
Correspondence to: Arjen Rinzema Lipids from yeasts and fungi: Tomorrow's source of biodiesel
13
Correspondence to: Arjen Rinzema, Bioprocess Engineering, Wageningen University, P.O. Box 8129, 6700 EV Wageningen, the Netherlands. E-mail: [email protected] © 2013 Society of Chemical Industry and John Wiley & Sons, Ltd Perspective Lipids from yeasts and fungi: Tomorrow’s source of biodiesel? Petra Meeuwse, Johan P.M. Sanders, Johannes Tramper, Arjen Rinzema, Wageningen University, the Netherlands Received October 31, 2013; revised February 26, 2013; accepted February 27, 2013 View online at Wiley Online Library (wileyonlinelibrary.com); DOI: 10.1002/bbb.1410; Biofuels, Bioprod. Bioref. (2013) Abstract: In the search for new transport fuels from renewable resources, biodiesel from microbial lip- ids comes into view. We have evaluated the lipid yield and energy use of a process for production of biodiesel from agricultural waste using lipid-accumulating yeast and fungi. We included different bio- reactors for submerged and solid-state fermentation in our evaluation. Using existing kinetic models, we predict lipid yields on substrate between 5% and 19% (w/w), depending on the culture system. According to the same models, improvement of the yield to 25–30% (w/w) is possible, for example by genetic modification of the micro-organisms. The net energy ratio of the non-optimized systems varies between 0.8 and 2.5 MJ produced per MJ used; energy use for pre-treatment and for oxygen transfer are most important. For the optimized systems, the net energy ratio increases to 2.9–5.5 MJ produced per MJ used, which can compete very well with other biofuels such as bioethanol or algal biodiesel. This shows that, although quite some work still has to be done, microbial lipids have the potential to be tomorrow’s source of biodiesel. © 2013 Society of Chemical Industry and John Wiley & Sons, Ltd Supporting information the appendices A–D as mentioned in the text can be found online. Keywords: biodiesel; energy yield; oleaginous fungi; microbial lipids; biomass Introduction L ipid accumulation in yeasts and fungi has been stud- ied for a long time because the lipids are rich in valu- able unsaturated fatty acids such as arachidonic acid and γ-linolenic acid. 1 Only recently, accumulated lipids are considered as a source of feedstock for biodiesel. 2–4 Especially the use of waste products from agriculture and agro-industry as substrate for lipid accumulating yeasts and fungi is studied extensively. 5–7 Unlike poly- unsaturated fatty acids, which are a specialty product with a high value, lipids for biodiesel production are a bulk product with a relatively low value and therefore have to be produced at low cost. Furthermore, energy input during production should be minimized to make the process effective from an energy point of view. Literature shows that the production costs can be reduced by using cheap substrates 5–7 and a cheaper process such as solid-state fermentation (SSF) instead of submerged fermentation (SmF). 7–11 Energy input during production, however, has not been considered before. In this paper, we estimate the energy input needed for large-scale production of lipids for biodiesel from organic waste material using lipid-accumulating yeast and fungi. Lipid yields in several cultivation systems are calculated with recently developed kinetic models for the filamentous fungus Umbelopsis isabellina; 12–14 two of these models can also describe data from yeasts, which shows their general
Transcript of Correspondence to: Arjen Rinzema Lipids from yeasts and fungi: Tomorrow's source of biodiesel

Correspondence to: Arjen Rinzema, Bioprocess Engineering, Wageningen University, P.O. Box 8129, 6700 EV Wageningen,
the Netherlands. E-mail: [email protected]
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd
Perspective
Lipids from yeasts and fungi: Tomorrow’s source of biodiesel?Petra Meeuwse, Johan P.M. Sanders, Johannes Tramper, Arjen Rinzema, Wageningen University, the Netherlands
Received October 31, 2013; revised February 26, 2013; accepted February 27, 2013View online at Wiley Online Library (wileyonlinelibrary.com); DOI: 10.1002/bbb.1410;Biofuels, Bioprod. Bioref. (2013)
Abstract: In the search for new transport fuels from renewable resources, biodiesel from microbial lip-ids comes into view. We have evaluated the lipid yield and energy use of a process for production of biodiesel from agricultural waste using lipid-accumulating yeast and fungi. We included different bio-reactors for submerged and solid-state fermentation in our evaluation. Using existing kinetic models, we predict lipid yields on substrate between 5% and 19% (w/w), depending on the culture system. According to the same models, improvement of the yield to 25–30% (w/w) is possible, for example by genetic modifi cation of the micro-organisms. The net energy ratio of the non-optimized systems varies between 0.8 and 2.5 MJ produced per MJ used; energy use for pre-treatment and for oxygen transfer are most important. For the optimized systems, the net energy ratio increases to 2.9–5.5 MJ produced per MJ used, which can compete very well with other biofuels such as bioethanol or algal biodiesel. This shows that, although quite some work still has to be done, microbial lipids have the potential to be tomorrow’s source of biodiesel. © 2013 Society of Chemical Industry and John Wiley & Sons, Ltd
Supporting information the appendices A–D as mentioned in the text can be found online.
Keywords: biodiesel; energy yield; oleaginous fungi; microbial lipids; biomass
Introduction
Lipid accumulation in yeasts and fungi has been stud-ied for a long time because the lipids are rich in valu-able unsaturated fatty acids such as arachidonic acid
and γ-linolenic acid.1 Only recently, accumulated lipids are considered as a source of feedstock for biodiesel.2–4 Especially the use of waste products from agriculture and agro-industry as substrate for lipid accumulating yeasts and fungi is studied extensively.5–7 Unlike poly-unsaturated fatty acids, which are a specialty product with a high value, lipids for biodiesel production are a bulk product with a relatively low value and therefore have to be produced at low cost. Furthermore, energy input during
production should be minimized to make the process eff ective from an energy point of view. Literature shows that the production costs can be reduced by using cheap substrates5–7 and a cheaper process such as solid-state fermentation (SSF) instead of submerged fermentation (SmF).7–11 Energy input during production, however, has not been considered before.
In this paper, we estimate the energy input needed for large-scale production of lipids for biodiesel from organic waste material using lipid-accumulating yeast and fungi. Lipid yields in several cultivation systems are calculated with recently developed kinetic models for the fi lamentous fungus Umbelopsis isabellina;12–14 two of these models can also describe data from yeasts, which shows their general

© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
P Meeuwse et al. Perspective: Lipids from yeasts and fungi
other C/N-ratios that can be obtained by either addition of N-source (for example NH3) or addition of agricultural waste with a higher C/N-ratio. Examples of such additions are (pre-treated) corn stover or wheat straw17 which we assume to have a C/N-ratio of 120 Cmol/Nmol.
In solid-state fermentation, the enzymatic hydrolysis takes place during fermentation. For submerged culture, simultaneous saccharifi cation and fermentation as well as saccharifi cation prior to the cultivation are possible. We assume that the hydrolysis has a yield of 100%, that no inhibiting compounds are formed during pre-treatment, and that the viscosity of the hydrolyzed substrate solution is equal to that of water. Th e substrate concentration in sub-merged fermentation is limited to 200 kg dry substrate/m3 because of the water content of 80% in the main substrate (sugarbeet pulp).
Micro-organisms
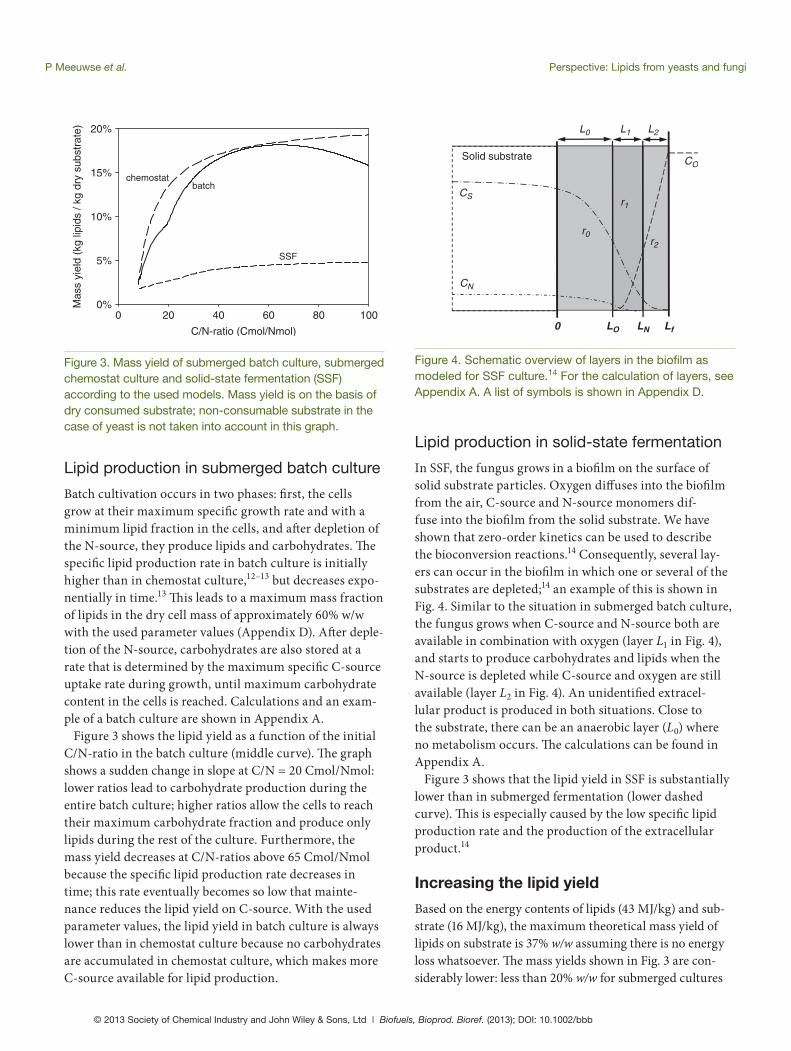
Figure 2 shows a simplifi ed scheme of a lipid-accumulating (oleaginous) organism.13 Th e organism uses an N-source and a C-source to produce cell mass, while C-source alone can also be used for lipid production, carbohydrate production and maintenance. In SSF, the production of a yet unidentifi ed extracellular product was also found and therefore included in the model (for SSF only). All of these processes also require oxygen. Th e accumulated lipids are a reserve component that can be consumed when the external C-source is exhausted.
applicability.13,15 Using basic engineering principles, we estimate the energy use during cultivation and part of the substrate pre-treatment and downstream processing steps.
The evaluated processes
Figure 1 gives an overview of (part of) the process for biodiesel production from a solid organic waste material. Included are a pre-treatment (substrate pre-treatment and sterilization), several bioreactors, a separation step and downstream processing to obtain biodiesel. We evaluated submerged fermentation and solid-state fermentation, each with its own advantages and disadvantages.
Substrate
As a model substrate, we used sugarbeet pulp, a cellulosic waste material from the European sugar industry. Th is material contains only a small amount of lignin, which makes it easily biodegradable.16 Although the composition of the material diff ers with harvest time and location, we assume that it contains 80% water and that the dry matter consists of one-third cellulose, one-third hemicellulose (mainly arabinan) and one-third pectin (Phyllis data-base17). To simplify calculations, we assume that it con-tains only three monomers: glucose (C6), arabinose (C5), and galacturonic acid (C6), which comprise up to 85% of total sugars present.18 Th e C/N-ratio of sugarbeet pulp is approximately 30 Cmol/Nmol. We also investigated
SterilizationPretreatmentSolid substrate
Enzymes
Water
Fungus
Yeast
Centrifugation
Extraction Refining
TransesterificationBiodiesel
Water with sugars
Biomass
STR
SSF
BCR
Pretreatment Fermentation Down stream processing
Figure 1. Schematic overview of process that is evaluated for lipid yield and energy use. STR = stirred tank reactor, BCR = bubble column reactor, SSF = solid state fermentation.

Perspective: Lipids from yeasts and fungi P Meeuwse et al.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
calculations, and to fi nd out what an improved micro-organism could achieve in the process.
Culture system design
Submerged and solid-state fermentation were evaluated. We considered submerged batch and chemostat cultures in a stirred-tank reactor and a bubble column. Submerged fermentation is widely used as a well-controlled system for cultivation. Solid-state fermentation is the culture of a micro-organism, usually a fi lamentous fungus, on a (wet) solid matrix without free-fl owing water. It is less com-monly used, but has several advantages:24–26 the process is cheaper, less power is needed for stirring and aeration, no centrifugation step is needed, and the amount of waste-water is reduced. We assume that an aerated packed-bed reactor with evaporative cooling and without mixing can be used.
Lipid production in submerged batch and chemostat cultures and in a model system for SSF (surface culture on carrageenan gel) were modeled before in our group.12–14 Th e metabolic processes included in these models are shown in Fig. 2. Th e kinetics of the models and the use of these kinetics in the diff erent production systems are shown in Appendix A.
Lipid production in submerged chemostat culture
In a chemostat culture, there is constant supply of both C-source and N-source. Growth, lipid produc-tion and maintenance occur simultaneously; we found no carbohydrate production in chemostat cultures with U. isabellina.12 When the N-source is limiting, the cell mass production rate depends on the N-source concentra-tion in the feed. Th e carbon source in the feed is used, in order of priority, for maintenance, cell mass production, and lipid production. Th erefore, the maximum specifi c lipid production rate is only reached when the C-source supply rate is suffi cient for all three processes. Our cal-culations show that the highest lipid fractions in the cells are reached at a low dilution rate combined with a high C/N-ratio; these calculations and an example of a chemo-stat culture are shown in Appendix A.
Figure 3 shows the lipid yield at diff erent C/N-ratios in the feed (upper dashed curve). Th is graph shows that the lipid yield increases with increasing C/N-ratio. At very high C/N-ratios, the shown curve reaches an asymptote of 20.8% w/w, but this coincides with unrealistically high lipid fractions (>90% w/w) in the cells. We therefore limited the C/N-ratio to 100 Cmol/Nmol in this graph and in our calculations.
We used a yeast and a fi lamentous fungus as lipid-accumulating organisms in our evaluation. Two main diff erences between yeast and fungus were considered: broth viscosity and substrate utilization capacity. A fi la-mentous fungus increases the viscosity of a submerged culture already at low dry weight concentrations, while this increase is much smaller for yeast. Th e viscosity is important for the energy needed for oxygen transfer in submerged cultures; for all cultures it was estimated using exponential functions of the cell mass concentrations as is shown in Appendix B.
As far as we know, there is no oleaginous yeast available that can convert all three monomers from sugarbeet pulp. Th e utilization of xylose has been reported for oleagi-nous yeasts19 as well as an oleaginous fungus,20 but for arabinose only a case of non-utilization is known.21 Th e utilization of pectin has been reported for fungi,22–23 but not for yeasts. Additionally, we have shown before that the oleaginous fungus Umbelopsis isabellina can con-vert sugarbeet pulp completely (Meeuwse, unpublished). Th erefore, we assume that the fungus can convert all mon-omers from sugarbeet pulp and the yeast can consume 50% (as an average between 33% for only glucose and 67% for glucose and either arabinose or galacturonic acid).
We used the same kinetic parameter values for both micro-organisms (Appendix D), based on experiments with the fi lamentous fungus U. isabellina in submerged batch culture.13 In this way, we take U. isabellina as a model organism, applying its biological parameters for both the fungus and the yeast in our calculations. As these parameter values infl uence the outcome of our evalua-tion, we carried out a sensitivity analysis to determine how important the used parameter values are in the
NH3
CH2O Glucose rS
rNCH1.77O0.55N0.17 Basic cell mass
CH1.95O0.11 Lipids
rX
rL
rCCO2
[H]
O2 H2OrO
rW
CH2O Carbohydrates
rIPCH4O
Extracellularproduct
rEP
Figure 2. Schematic overview of the modeled processes in the cell.13 Carbohydrate production (rIP) only occurs when no N-source is available, production of extracellular prod-uct (rEP) has only been observed in SSF. A list of symbols is shown in Appendix D.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
P Meeuwse et al. Perspective: Lipids from yeasts and fungi
Lipid production in solid-state fermentation
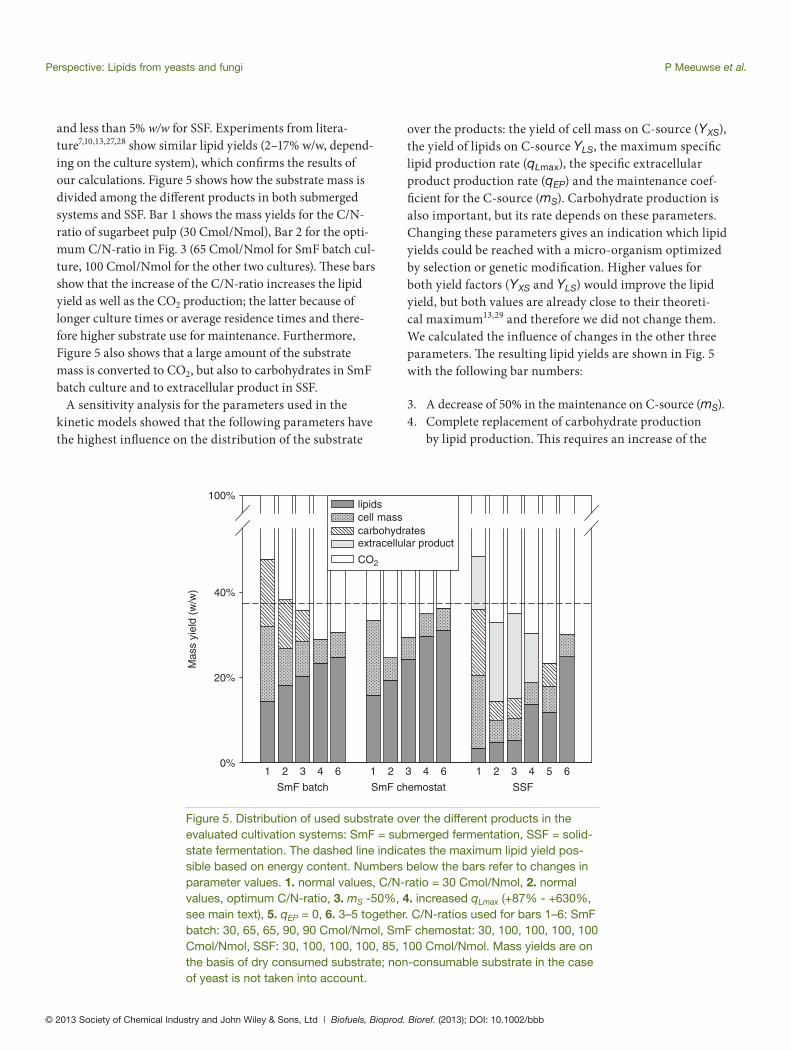
In SSF, the fungus grows in a biofi lm on the surface of solid substrate particles. Oxygen diff uses into the biofi lm from the air, C-source and N-source monomers dif-fuse into the biofi lm from the solid substrate. We have shown that zero-order kinetics can be used to describe the bioconversion reactions.14 Consequently, several lay-ers can occur in the biofi lm in which one or several of the substrates are depleted;14 an example of this is shown in Fig. 4. Similar to the situation in submerged batch culture, the fungus grows when C-source and N-source both are available in combination with oxygen (layer L1 in Fig. 4), and starts to produce carbohydrates and lipids when the N-source is depleted while C-source and oxygen are still available (layer L2 in Fig. 4). An unidentifi ed extracel-lular product is produced in both situations. Close to the substrate, there can be an anaerobic layer (L0) where no metabolism occurs. Th e calculations can be found in Appendix A.
Figure 3 shows that the lipid yield in SSF is substantially lower than in submerged fermentation (lower dashed curve). Th is is especially caused by the low specifi c lipid production rate and the production of the extracellular product.14
Increasing the lipid yield
Based on the energy contents of lipids (43 MJ/kg) and sub-strate (16 MJ/kg), the maximum theoretical mass yield of lipids on substrate is 37% w/w assuming there is no energy loss whatsoever. Th e mass yields shown in Fig. 3 are con-siderably lower: less than 20% w/w for submerged cultures
Lipid production in submerged batch culture
Batch cultivation occurs in two phases: fi rst, the cells grow at their maximum specifi c growth rate and with a minimum lipid fraction in the cells, and aft er depletion of the N-source, they produce lipids and carbohydrates. Th e specifi c lipid production rate in batch culture is initially higher than in chemostat culture,12–13 but decreases expo-nentially in time.13 Th is leads to a maximum mass fraction of lipids in the dry cell mass of approximately 60% w/w with the used parameter values (Appendix D). Aft er deple-tion of the N-source, carbohydrates are also stored at a rate that is determined by the maximum specifi c C-source uptake rate during growth, until maximum carbohydrate content in the cells is reached. Calculations and an exam-ple of a batch culture are shown in Appendix A.
Figure 3 shows the lipid yield as a function of the initial C/N-ratio in the batch culture (middle curve). Th e graph shows a sudden change in slope at C/N = 20 Cmol/Nmol: lower ratios lead to carbohydrate production during the entire batch culture; higher ratios allow the cells to reach their maximum carbohydrate fraction and produce only lipids during the rest of the culture. Furthermore, the mass yield decreases at C/N-ratios above 65 Cmol/Nmol because the specifi c lipid production rate decreases in time; this rate eventually becomes so low that mainte-nance reduces the lipid yield on C-source. With the used parameter values, the lipid yield in batch culture is always lower than in chemostat culture because no carbohydrates are accumulated in chemostat culture, which makes more C-source available for lipid production.
C/N-ratio (Cmol/Nmol)
0 20 40 60 80 100
Mas
s yi
eld
(kg
lipid
s / k
g dr
y su
bstr
ate)
0%
5%
10%
15%
20%
SSF
batchchemostat
Figure 3. Mass yield of submerged batch culture, submerged chemostat culture and solid-state fermentation (SSF) according to the used models. Mass yield is on the basis of dry consumed substrate; non-consumable substrate in the case of yeast is not taken into account in this graph.
0 LO LN Lf
CS
CN
CO
L0 L1 L2
r0
r1
r2
Solid substrate
Figure 4. Schematic overview of layers in the biofi lm as modeled for SSF culture.14 For the calculation of layers, see Appendix A. A list of symbols is shown in Appendix D.
Perspective: Lipids from yeasts and fungi P Meeuwse et al.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
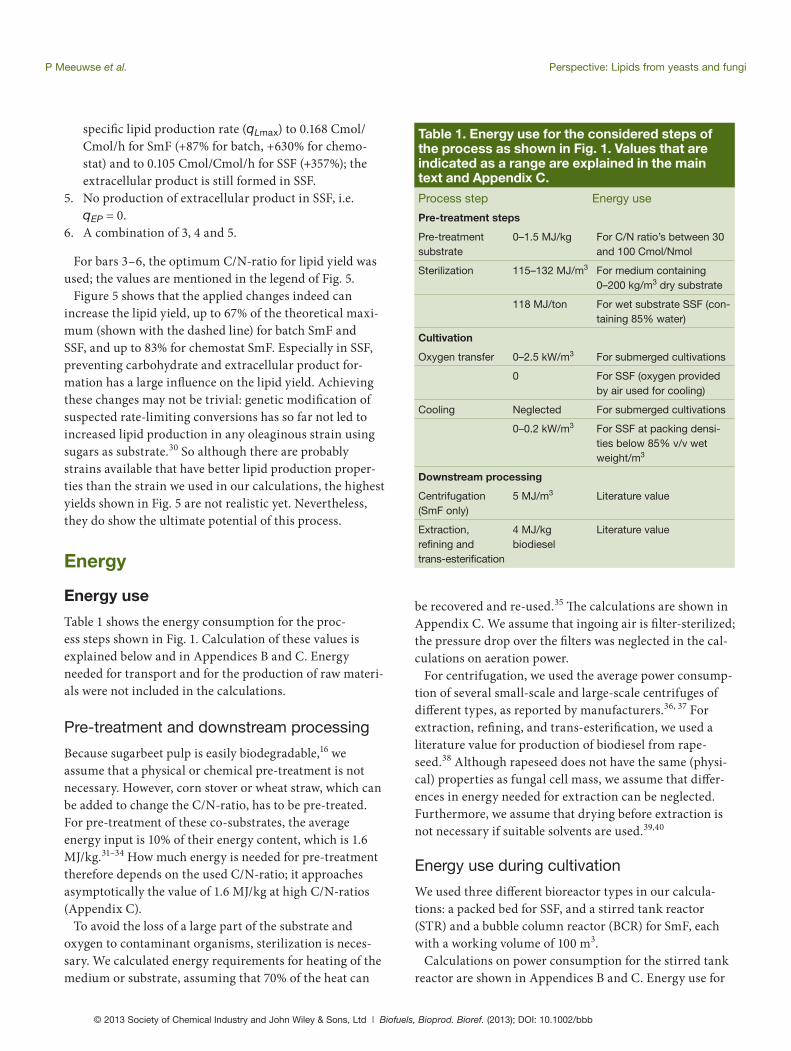
over the products: the yield of cell mass on C-source (YXS), the yield of lipids on C-source YLS, the maximum specifi c lipid production rate (qLmax), the specifi c extracellular product production rate (qEP) and the maintenance coef-fi cient for the C-source (mS). Carbohydrate production is also important, but its rate depends on these parameters. Changing these parameters gives an indication which lipid yields could be reached with a micro-organism optimized by selection or genetic modifi cation. Higher values for both yield factors (YXS and YLS) would improve the lipid yield, but both values are already close to their theoreti-cal maximum13,29 and therefore we did not change them. We calculated the infl uence of changes in the other three parameters. Th e resulting lipid yields are shown in Fig. 5 with the following bar numbers:
3. A decrease of 50% in the maintenance on C-source (mS).4. Complete replacement of carbohydrate production
by lipid production. Th is requires an increase of the
and less than 5% w/w for SSF. Experiments from litera-ture7,10,13,27,28 show similar lipid yields (2–17% w/w, depend-ing on the culture system), which confi rms the results of our calculations. Figure 5 shows how the substrate mass is divided among the diff erent products in both submerged systems and SSF. Bar 1 shows the mass yields for the C/N-ratio of sugarbeet pulp (30 Cmol/Nmol), Bar 2 for the opti-mum C/N-ratio in Fig. 3 (65 Cmol/Nmol for SmF batch cul-ture, 100 Cmol/Nmol for the other two cultures). Th ese bars show that the increase of the C/N-ratio increases the lipid yield as well as the CO2 production; the latter because of longer culture times or average residence times and there-fore higher substrate use for maintenance. Furthermore, Figure 5 also shows that a large amount of the substrate mass is converted to CO2, but also to carbohydrates in SmF batch culture and to extracellular product in SSF.
A sensitivity analysis for the parameters used in the kinetic models showed that the following parameters have the highest infl uence on the distribution of the substrate
Mas
s yi
eld
(w/w
)
0%
20%
40%
100%lipidscell masscarbohydratesextracellular product
CO2
1 2 3 4 6 1 2 4 6 1 2 3 6
SmF batch SmF chemostat SSF
3 4 5
Figure 5. Distribution of used substrate over the different products in the evaluated cultivation systems: SmF = submerged fermentation, SSF = solid-state fermentation. The dashed line indicates the maximum lipid yield pos-sible based on energy content. Numbers below the bars refer to changes in parameter values. 1. normal values, C/N-ratio = 30 Cmol/Nmol, 2. normal values, optimum C/N-ratio, 3. mS -50%, 4. increased qLmax (+87% - +630%, see main text), 5. qEP = 0, 6. 3–5 together. C/N-ratios used for bars 1–6: SmF batch: 30, 65, 65, 90, 90 Cmol/Nmol, SmF chemostat: 30, 100, 100, 100, 100 Cmol/Nmol, SSF: 30, 100, 100, 100, 85, 100 Cmol/Nmol. Mass yields are on the basis of dry consumed substrate; non-consumable substrate in the case of yeast is not taken into account.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
P Meeuwse et al. Perspective: Lipids from yeasts and fungi
be recovered and re-used.35 Th e calculations are shown in Appendix C. We assume that ingoing air is fi lter-sterilized; the pressure drop over the fi lters was neglected in the cal-culations on aeration power.
For centrifugation, we used the average power consump-tion of several small-scale and large-scale centrifuges of diff erent types, as reported by manufacturers.36, 37 For extraction, refi ning, and trans-esterifi cation, we used a literature value for production of biodiesel from rape-seed.38 Although rapeseed does not have the same (physi-cal) properties as fungal cell mass, we assume that diff er-ences in energy needed for extraction can be neglected. Furthermore, we assume that drying before extraction is not necessary if suitable solvents are used.39,40
Energy use during cultivation
We used three diff erent bioreactor types in our calcula-tions: a packed bed for SSF, and a stirred tank reactor (STR) and a bubble column reactor (BCR) for SmF, each with a working volume of 100 m3.
Calculations on power consumption for the stirred tank reactor are shown in Appendices B and C. Energy use for
specifi c lipid production rate (qLmax) to 0.168 Cmol/Cmol/h for SmF (+87% for batch, +630% for chemo-stat) and to 0.105 Cmol/Cmol/h for SSF (+357%); the extracellular product is still formed in SSF.
5. No production of extracellular product in SSF, i.e. qEP = 0.
6. A combination of 3, 4 and 5.
For bars 3–6, the optimum C/N-ratio for lipid yield was used; the values are mentioned in the legend of Fig. 5.
Figure 5 shows that the applied changes indeed can increase the lipid yield, up to 67% of the theoretical maxi-mum (shown with the dashed line) for batch SmF and SSF, and up to 83% for chemostat SmF. Especially in SSF, preventing carbohydrate and extracellular product for-mation has a large infl uence on the lipid yield. Achieving these changes may not be trivial: genetic modifi cation of suspected rate-limiting conversions has so far not led to increased lipid production in any oleaginous strain using sugars as substrate.30 So although there are probably strains available that have better lipid production proper-ties than the strain we used in our calculations, the highest yields shown in Fig. 5 are not realistic yet. Nevertheless, they do show the ultimate potential of this process.
Energy
Energy use
Table 1 shows the energy consumption for the proc-ess steps shown in Fig. 1. Calculation of these values is explained below and in Appendices B and C. Energy needed for transport and for the production of raw materi-als were not included in the calculations.
Pre-treatment and downstream processing
Because sugarbeet pulp is easily biodegradable,16 we assume that a physical or chemical pre-treatment is not necessary. However, corn stover or wheat straw, which can be added to change the C/N-ratio, has to be pre-treated. For pre-treatment of these co-substrates, the average energy input is 10% of their energy content, which is 1.6 MJ/kg.31–34 How much energy is needed for pre-treatment therefore depends on the used C/N-ratio; it approaches asymptotically the value of 1.6 MJ/kg at high C/N-ratios (Appendix C).
To avoid the loss of a large part of the substrate and oxygen to contaminant organisms, sterilization is neces-sary. We calculated energy requirements for heating of the medium or substrate, assuming that 70% of the heat can
Table 1. Energy use for the considered steps of the process as shown in Fig. 1. Values that are indicated as a range are explained in the main text and Appendix C.
Process step Energy use
Pre-treatment steps
Pre-treatment substrate
0–1.5 MJ/kg For C/N ratio’s between 30 and 100 Cmol/Nmol
Sterilization 115–132 MJ/m3 For medium containing 0–200 kg/m3 dry substrate
118 MJ/ton For wet substrate SSF (con-taining 85% water)
Cultivation
Oxygen transfer 0–2.5 kW/m3 For submerged cultivations
0 For SSF (oxygen provided by air used for cooling)
Cooling Neglected For submerged cultivations
0–0.2 kW/m3 For SSF at packing densi-ties below 85% v/v wet weight/m3
Downstream processing
Centrifugation (SmF only)
5 MJ/m3 Literature value
Extraction, refi ning and trans-esterifi cation
4 MJ/kg biodiesel
Literature value

Perspective: Lipids from yeasts and fungi P Meeuwse et al.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
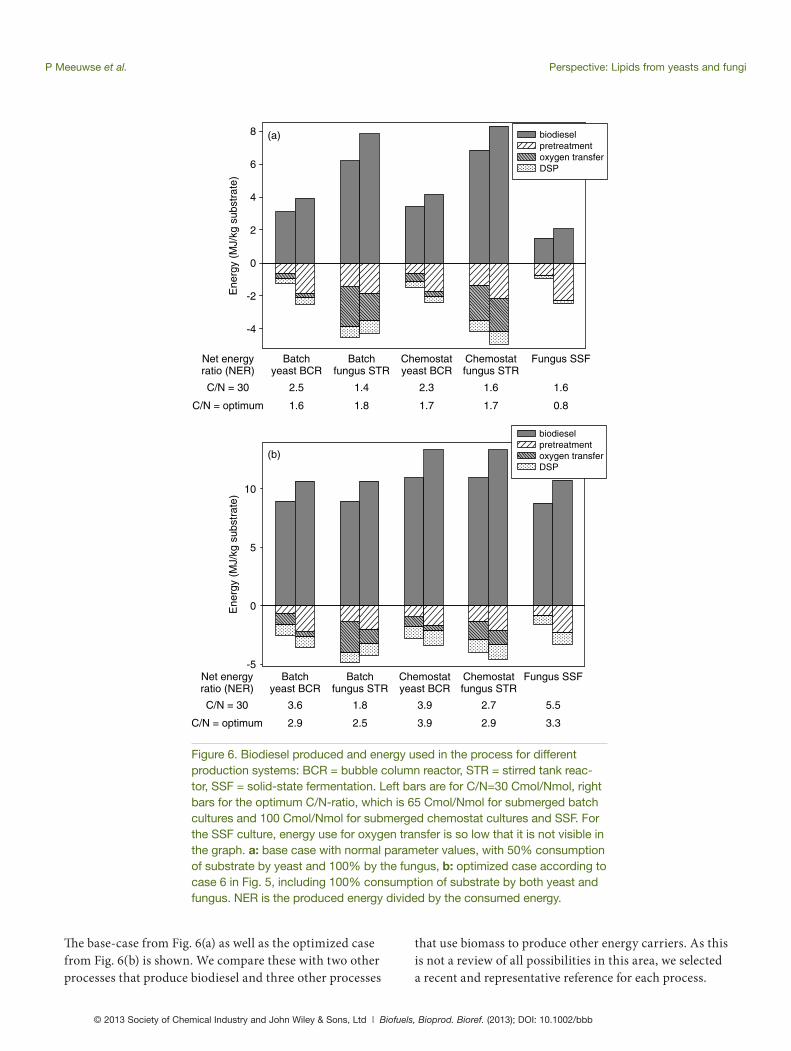
use for pre-treatment (Appendix C). Furthermore, the substrate concentration in the cultures also infl uences the energy use: at low substrate concentrations, energy use for sterilization of the complete medium (including water) in SmF is relatively high; at high substrate concentrations, the viscosity of the fungal cultures increases the energy use for oxygen transfer. Figure 6 shows the results for C/N = 30 Cmol/Nmol and for the optimum C/N-ratio, in all cases at the substrate concentration giving the lowest energy use.
Th e most important result shown in Fig. 6 is that in many cases the higher lipid yield at the optimum C/N-ratio cannot compensate for the higher energy use at this higher C/N-ratio. In these cases, the pre-treatment of the co-substrate costs more energy than is available from the extra biodiesel produced. Only for the submerged fungal cultures, a higher C/N-ratio is favorable. At a high C/N-ratio, more lipids are produced compared to biomass than at a low C/N-ratio. Th erefore, relatively less oxygen trans-fer is needed at a high C/N-ratio, because growth needs more oxygen than lipid production. Th is eff ect is strongest for submerged fungal culture because of the high viscosity, which increases energy use for oxygen transfer. Because of the high energy use for oxygen transfer in submerged fungal culture, the NER value for yeast cultures is higher, although the biodiesel yield is lower because of the 50% substrate utilization. In SSF with the normal fungal strain, the produced energy is low because of the low lipid yields (Fig. 3). Energy use is also low, but not low enough to give a high NER value. For the optimized strain, however, SSF has the highest NER value of all processes shown. Finally, Fig 6 shows that the diff erences between batch culture and chemostat culture are small.
In the NER calculation, energy used in the form of heat and in the form of electricity are added up without taking into account that electricity is oft en produced from heat with yield of approximately 40%. Th erefore, the NER value for systems which need a lot of energy for oxygen transfer (electricity for stirrers and air pumps) can be lower than shown in Fig. 6.
Comparison to other fuel production systems
Table 2 shows some examples of other systems that pro-duce energy carriers from renewable resources. From our own calculations, we added both submerged batch proc-esses (yeast BCR and fungus STR) and the SSF process. We used the values with the C/N-ratio that gave the highest NER: 30 Cmol/Nmol for yeast in SmF and fungus in SSF, and the optimum C/N-ratio for fungus in SmF.
(water) cooling can be neglected compared to stirring and aeration in submerged fermentation, in a stirred tank reac-tor as well as in a bubble column reactor (calculations not shown).
In a stirred tank reactor, viscosity only has an infl uence on oxygen transfer above a threshold of 50×10–3 Nms–2.41 Th is threshold is not reached in the yeast culture. In the fungal culture, it is reached at a cell mass concentration of 17 kg/m3; above this value, the oxygen transfer coeffi cient decreases rapidly.
In a bubble column reactor, oxygen transfer decreases very rapidly when the viscosity of the liquid increases. Th e power consumption for a bubble column is lower than for a stirred tank reactor for the same oxygen transfer rate, except for very high yeast concentrations and fungal cul-tures (Appendix C). Th ese high yeast concentrations are not reached because we limited the maximum substrate concentration in the reactor and the feed to 200 kg/m3. We therefore chose the bubble column reactor for all yeast cul-tures and the stirred tank reactor for all submerged fungal cultures.
In solid-state fermentation, the packed bed is aerated to remove heat and to supply oxygen. Literature42–44 shows that if the aeration rate is suffi cient for heat removal, it is also suffi cient for oxygen supply. Th erefore, the aeration rate is calculated from the heat production rate, which is directly proportional to the oxygen consumption rate. Th is is shown in Appendix B. Th e power consumption for aeration depends on the pressure drop over the packed bed. Th is pressure drop depends, amongst others, on the volume fraction of solid particles in the packed bed, which we assume to be constant during the culture. At a fraction of wet solid material below 85% v/v, the pressure drop is low (<0.05 bar); we therefore limited the wet substrate con-centration to 85% v/v. Th ese calculations are also shown in Appendices B and C.
Energy yields
Figure 6 shows the biodiesel produced and the energy used for all processes, both expressed in MJ per kg sub-strate, and it shows the ratio of both (the Net Energy Ratio, NER). Figure 6(a) shows results for the normal strain with parameter values shown in Appendix D. Figure 6(b) shows results for the optimized strains that were shown as case 6 in Fig. 5. In addition, we assumed that the substrate utili-zation is optimized to 100% for the yeast cultures.
Th e results shown in Fig. 6 have a complex back-ground. For example, biodiesel production depends on the C/N-ratio of the substrate as was shown in Fig. 3, and the C/N-ratio of the substrate also determines the energy
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
P Meeuwse et al. Perspective: Lipids from yeasts and fungi
that use biomass to produce other energy carriers. As this is not a review of all possibilities in this area, we selected a recent and representative reference for each process.
Th e base-case from Fig. 6(a) as well as the optimized case from Fig. 6(b) is shown. We compare these with two other processes that produce biodiesel and three other processes
Ene
rgy
(MJ/
kg s
ubst
rate
)
-4
-2
0
2
4
6
8 biodieselpretreatmentoxygen transferDSP
Net energyratio (NER)
Batchyeast BCR
Batchfungus STR
Chemostatyeast BCR
Chemostatfungus STR
Fungus SSF
C/N = 30 2.5 1.4 2.3 1.6 1.6
C/N = optimum 1.6 1.8 1.7 1.7
Ene
rgy
(MJ/
kg s
ubst
rate
)
-5
0
5
10
biodieselpretreatmentoxygen transferDSP
Net energyratio (NER)
Batchyeast BCR
Batchfungus STR
Chemostatyeast BCR
Chemostatfungus STR
Fungus SSF
C/N = 30 3.6 1.8 3.9 2.7 5.5
C/N = optimum 2.9 2.5 3.9 2.9 3.3
0.8
(b)
(a)
Figure 6. Biodiesel produced and energy used in the process for different production systems: BCR = bubble column reactor, STR = stirred tank reac-tor, SSF = solid-state fermentation. Left bars are for C/N=30 Cmol/Nmol, right bars for the optimum C/N-ratio, which is 65 Cmol/Nmol for submerged batch cultures and 100 Cmol/Nmol for submerged chemostat cultures and SSF. For the SSF culture, energy use for oxygen transfer is so low that it is not visible in the graph. a: base case with normal parameter values, with 50% consumption of substrate by yeast and 100% by the fungus, b: optimized case according to case 6 in Fig. 5, including 100% consumption of substrate by both yeast and fungus. NER is the produced energy divided by the consumed energy.
Perspective: Lipids from yeasts and fungi P Meeuwse et al.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
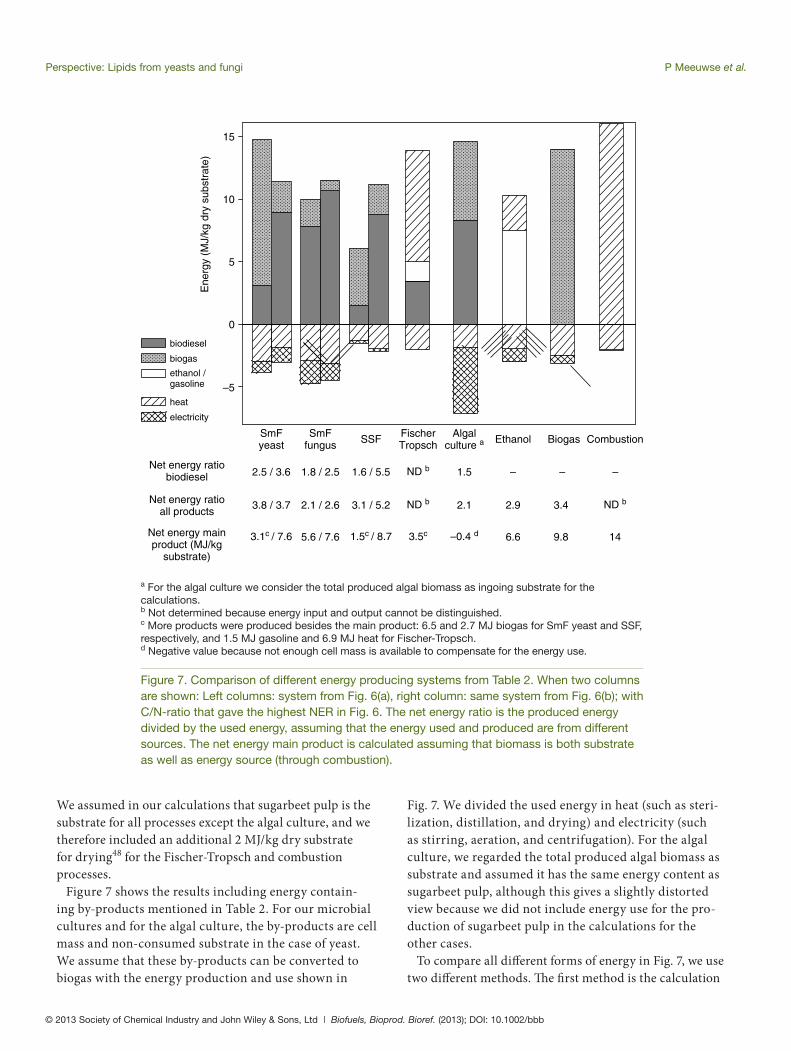
Fig. 7. We divided the used energy in heat (such as steri-lization, distillation, and drying) and electricity (such as stirring, aeration, and centrifugation). For the algal culture, we regarded the total produced algal biomass as substrate and assumed it has the same energy content as sugarbeet pulp, although this gives a slightly distorted view because we did not include energy use for the pro-duction of sugarbeet pulp in the calculations for the other cases.
To compare all diff erent forms of energy in Fig. 7, we use two diff erent methods. Th e fi rst method is the calculation
We assumed in our calculations that sugarbeet pulp is the substrate for all processes except the algal culture, and we therefore included an additional 2 MJ/kg dry substrate for drying48 for the Fischer-Tropsch and combustion processes.
Figure 7 shows the results including energy contain-ing by-products mentioned in Table 2. For our microbial cultures and for the algal culture, the by-products are cell mass and non-consumed substrate in the case of yeast. We assume that these by-products can be converted to biogas with the energy production and use shown in
Ene
rgy
(MJ/
kg d
ry s
ubst
rate
)
–5
0
5
10
15
biodiesel
biogas
ethanol /gasoline
heat
electricity
SmFyeast
SmFfungus SSF Fischer
TropschAlgal
culture a Ethanol Biogas Combustion
Net energy ratiobiodiesel 2.5 / 3.6 1.8 / 2.5 1.6 / 5.5 ND b 1.5 – ––
Net energy ratioall products
3.8 / 3.7 2.1 / 2.6 3.1 / 5.2 ND b 2.1 2.9 3.4 ND b
Net energy mainproduct (MJ/kg
substrate)
3.1c / 7.6 5.6 / 7.6 1.5c / 8.7 3.5c –0.4 d 6.6 9.8 14
a For the algal culture we consider the total produced algal biomass as ingoing substrate for the calculations.b Not determined because energy input and output cannot be distinguished.c More products were produced besides the main product: 6.5 and 2.7 MJ biogas for SmF yeast and SSF, respectively, and 1.5 MJ gasoline and 6.9 MJ heat for Fischer-Tropsch.d Negative value because not enough cell mass is available to compensate for the energy use.
Figure 7. Comparison of different energy producing systems from Table 2. When two columns are shown: Left columns: system from Fig. 6(a), right column: same system from Fig. 6(b); with C/N-ratio that gave the highest NER in Fig. 6. The net energy ratio is the produced energy divided by the used energy, assuming that the energy used and produced are from different sources. The net energy main product is calculated assuming that biomass is both substrate as well as energy source (through combustion).
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
P Meeuwse et al. Perspective: Lipids from yeasts and fungi
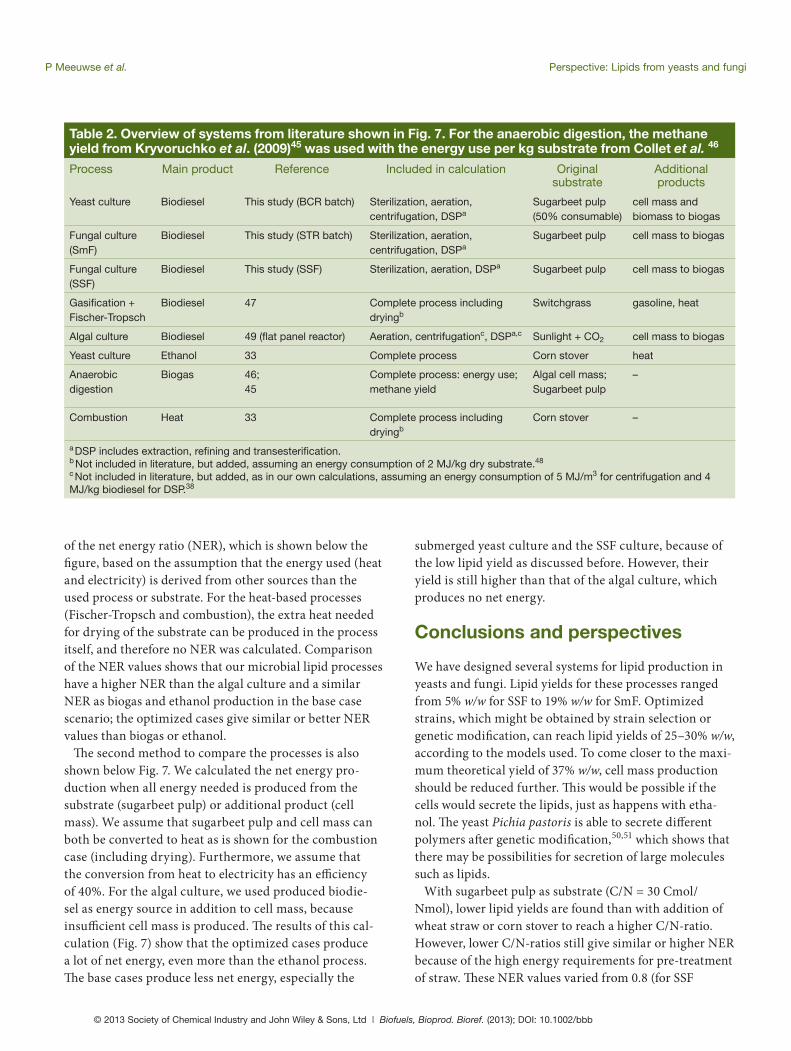
Table 2. Overview of systems from literature shown in Fig. 7. For the anaerobic digestion, the methane yield from Kryvoruchko et al. (2009)45 was used with the energy use per kg substrate from Collet et al. 46
Process Main product Reference Included in calculation Original substrate
Additional products
Yeast culture Biodiesel This study (BCR batch) Sterilization, aeration, centrifugation, DSPa
Sugarbeet pulp (50% consumable)
cell mass and biomass to biogas
Fungal culture (SmF)
Biodiesel This study (STR batch) Sterilization, aeration, centrifugation, DSPa
Sugarbeet pulp cell mass to biogas
Fungal culture (SSF)
Biodiesel This study (SSF) Sterilization, aeration, DSPa Sugarbeet pulp cell mass to biogas
Gasifi cation + Fischer-Tropsch
Biodiesel 47 Complete process including dryingb
Switchgrass gasoline, heat
Algal culture Biodiesel 49 (fl at panel reactor) Aeration, centrifugationc, DSPa,c Sunlight + CO2 cell mass to biogas
Yeast culture Ethanol 33 Complete process Corn stover heat
Anaerobic digestion
Biogas 46;45
Complete process: energy use;methane yield
Algal cell mass;Sugarbeet pulp
–
Combustion Heat 33 Complete process including dryingb
Corn stover –
a DSP includes extraction, refi ning and transesterifi cation.b Not included in literature, but added, assuming an energy consumption of 2 MJ/kg dry substrate.48
c Not included in literature, but added, as in our own calculations, assuming an energy consumption of 5 MJ/m3 for centrifugation and 4 MJ/kg biodiesel for DSP.38
of the net energy ratio (NER), which is shown below the fi gure, based on the assumption that the energy used (heat and electricity) is derived from other sources than the used process or substrate. For the heat-based processes (Fischer-Tropsch and combustion), the extra heat needed for drying of the substrate can be produced in the process itself, and therefore no NER was calculated. Comparison of the NER values shows that our microbial lipid processes have a higher NER than the algal culture and a similar NER as biogas and ethanol production in the base case scenario; the optimized cases give similar or better NER values than biogas or ethanol.
Th e second method to compare the processes is also shown below Fig. 7. We calculated the net energy pro-duction when all energy needed is produced from the substrate (sugarbeet pulp) or additional product (cell mass). We assume that sugarbeet pulp and cell mass can both be converted to heat as is shown for the combustion case (including drying). Furthermore, we assume that the conversion from heat to electricity has an effi ciency of 40%. For the algal culture, we used produced biodie-sel as energy source in addition to cell mass, because insuffi cient cell mass is produced. Th e results of this cal-culation (Fig. 7) show that the optimized cases produce a lot of net energy, even more than the ethanol process. Th e base cases produce less net energy, especially the
submerged yeast culture and the SSF culture, because of the low lipid yield as discussed before. However, their yield is still higher than that of the algal culture, which produces no net energy.
Conclusions and perspectives
We have designed several systems for lipid production in yeasts and fungi. Lipid yields for these processes ranged from 5% w/w for SSF to 19% w/w for SmF. Optimized strains, which might be obtained by strain selection or genetic modifi cation, can reach lipid yields of 25–30% w/w, according to the models used. To come closer to the maxi-mum theoretical yield of 37% w/w, cell mass production should be reduced further. Th is would be possible if the cells would secrete the lipids, just as happens with etha-nol. Th e yeast Pichia pastoris is able to secrete diff erent polymers aft er genetic modifi cation,50,51 which shows that there may be possibilities for secretion of large molecules such as lipids.
With sugarbeet pulp as substrate (C/N = 30 Cmol/Nmol), lower lipid yields are found than with addition of wheat straw or corn stover to reach a higher C/N-ratio. However, lower C/N-ratios still give similar or higher NER because of the high energy requirements for pre-treatment of straw. Th ese NER values varied from 0.8 (for SSF

Perspective: Lipids from yeasts and fungi P Meeuwse et al.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
because of the low lipid yield) to 2.5. For the optimized strains, the NER could be increased to 3–5.5; SSF has the highest NER due to the low energy costs of oxygen transfer in this system. Th ese values can compete very well with NER values for other biofuels such as algal biodiesel and bioethanol.
Th e NER value might be increased further by energy savings in the process. We used separate steps for pre-treatment and sterilization, while it is probably possible to combine these steps and save energy. In addition, the extraction and trans-esterifi cation step might be com-bined.52 Large energy savings could also be achieved if it were possible to produce lipids in an anaerobic culture.53 However, there are no reports of lipid accumulation in yeast and fungi under anaerobic conditions and among the lipid-accumulating bacteria there are no anaerobic species.54 Furthermore, intracellular lipid storage would be limited by the low cell yield of anaerobic cultures. Th erefore, anaerobic lipid production would only be pos-sible when the lipids are excreted.
A topic that has not been discussed in this review so far is the economics of this process. Submerged aerobic fermentation is known to be expensive. Rough estimates show that the costs for submerged cultivation only will be in the range of 200–1000 €/GJ biodiesel for the non-opti-mized cases. Th is is very high compared to the production costs for other biofuels: 7–20 €/GJ for biodiesel from dif-ferent oils,55 13–23 €/GJ for bioethanol from straw,56 and 75–450 €/GJ for biodiesel from algae.57 For the optimized submerged cultures, the cultivation costs are a factor 10 lower, but this is still quite high. For SSF, the production price is much lower: 50–100 €/GJ for the base case and a factor 10 lower for the optimized case, according to our estimates. Furthermore, SSF is a low-tech application, which produces less wastewater and can be used on a small scale at the source of biomass production, i.e. farms and agricultural processing plants. Th is reduces transport costs considerably, which gives SSF an extra advantage compared to the submerged systems.
Summarizing, the lipid yields found in the base-case sce-nario are quite low, especially in SSF, but we have shown that there is a potential for higher lipid yields. Th e energy use of the processes is quite high, but also here there is room for improvement. Microbial lipids have only been produced commercially for high value unsaturated fatty acids so far; process development for bulk production has yet to be started. Th is review shows that the potential lipid yield as well as the energy balance is promising enough to continue research on this subject. Th e high cultiva-tion costs of submerged aerobic fermentation might be a
serious problem, but the cultivation costs in SSF are much lower. With the improvement of lipid yield in SSF, micro-bial lipids produced in SSF cultures might be tomorrow’s source of biodiesel.
Acknowledgments
Th is work was fi nancially supported by the DEN program of SenterNovem under project number 2020-03-12-14-006.
References 1. Certik M and Shimizu S, Biosynthesis and regulation of micro-
bial polyunsaturated fatty acid production. J Biosci Bioeng 87:1–14 (1999).
2. Li Q, Du W and Liu DH, Perspectives of microbial oils for biodiesel production. Appl Microbiol Biot 80:749–756 (2008).
3. Feofi lova EP, Sergeeva YE and Ivashechkin AA, Biodiesel-fuel: Content, production, producers, contemporary biotechnology (Review). Appl Biochem Micro+ 46:369–378 (2009).
4. Meng X, Yang JM, Xu X, Zhang L, Nie QJ and Xian M, Biodiesel production from oleaginous microorganisms. Renew Energ 34:1–5 (2009).
5. Angerbauer C, Siebenhofer M, Mittelbach M and Guebitz GM, Conversion of sewage sludge into lipids by Lipomyces starkeyi for biodiesel production. Bioresource Technol 99:3051–3056 (2008).
6. Papanikolaou S and Aggelis G, Biotechnological valorization of biodiesel derived glycerol waste through production of sin-gle cell oil and citric acid by Yarrowia lipolytica. Lipid Technol 21:83–87 (2009).
7. Peng X and Chen H, Single cell oil production in solid-state fermentation by Microsphaeropsis sp. from steam-exploded wheat straw mixed with wheat bran. Bioresource Technol 99:3885–3889 (2008).
8. Jang HD, Lin YY and Yang SS, Polyunsaturated fatty acid pro-duction with Mortierella alpina by solid substrate fermentation. Bot Bull Acad Sinica 41:41–48 (2000).
9. Stredansky M, Conti E, Stredanska S and Zanetti F, gamma-Linolenic acid production with Thamnidium elegans by solid-state fermentation on apple pomace. Bioresource Technol 73:41–45 (2000).
10. Gema H, Kavadia A, Dimou D, Tsagou V, Komaitis M and Aggelis G, Production of gamma-linolenic acid by Cunninghamella echinulata cultivated on glucose and orange peel. Appl Microbiol Biot 58:303–307 (2002).
11. Fakas S, Makri A, Mavromati M, Tselepi M and Aggelis G, Fatty acid composition in lipid fractions lengthwise the mycelium of Mortierella isabellina and lipid production by solid state fer-mentation. Bioresource Technol 100:6118–6120 (2009).
12. Meeuwse P, Tramper J and Rinzema A, Modeling lipid accumulation in oleaginous fungi in chemostat cultures. I: Development and validation of a chemostat model for Umbelopsis isabellina. Bioprocess Biosyst Eng 34:939–949 (2011).
13. Meeuwse P, Akbari P, Tramper J and Rinzema A, Modeling growth, lipid accumulation and lipid turnover in submerged batch cultures of Umbelopsis isabellina. Bioprocess Biosyst Eng 35:591–603 (2012).

© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
P Meeuwse et al. Perspective: Lipids from yeasts and fungi
14. Meeuwse P, Klok AJ, Haemers S, Tramper J and Rinzema A, Growth and lipid production of Umbelopsis isabellina on a solid substrate - Mechanistic modeling and validation. Process Biochem 47:1228–1242 (2012).
15. Meeuwse P, Tramper J and Rinzema A, Modeling lipid accumulation in oleaginous fungi in chemostat cultures. II: Validation of the chemostat model using yeast culture data from literature. Bioprocess Biosyst Eng 34:951–961 (2011).
16. Spagnuolo M, Crecchio C, Pizzigallo MDR and Ruggiero P, Synergistic effects of cellulolytic and pectinolytic enzymes in degrading sugar beet pulp. Bioresource Technol 60:215–222 (1997).
17. http://www.ecn.nl/phyllis
18. Micard V, Renard GMGC and Thibault J-F, Enzymatic saccha-rifi cation of sugar-beet pulp. Enzyme Microb Tech 19:162–170 (1996).
19. He MX, Hu QC, Gou XH, Liu XY, Li Q, Pan K et al., Screening of oleaginous yeast with xylose assimilating capacity for lipid and bio-ethanol production. Afr J Biotechnol 9:8392–8397 (2010).
20. Fakas S, Papanikolaou S, Batsos A, Galiotou-Panayotou M, Mallouchos A and Aggelis G, Evaluating renewable car-bon sources as substrates for single cell oil production by Cunninghamella echinulata and Mortierella isabellina. Biomass Bioenerg 33:573–580 (2009).
21. Glatz BA, Hammond EG, Hsu KH, Baehman L, Bati N, Bednarski W et al., Production and modifi cation of fats and oils by yeast fermentation, in Biotechnology for the Oils and Fats Industry, ed by Ratledge C, Dawson PSS and Rattray J. American Oil Chemists’ Society, United states of America, pp. 163–176 (1984).
22. Papanikolaou S, Galiotou-Panayotou M, Fakas S, Komaitis M and Aggelis G, Lipid production by oleaginous Mucorales cultivated on renewable carbon sources. Eur J Lipid Sci Tech 109:1060–1070 (2007).
23. Richard P and Hilditch S, d-Galacturonic acid catabolism in microorganisms and its biotechnological relevance. Appl Microbiol Biot 82:597–604 (2009).
24. Pandey A, Solid-state fermentation. Biochem Eng J 13:81–84 (2003).
25. Holker U and Lenz J, Solid-state fermentation - are there any biotechnological advantages? Curr Opin Microbiol 8:301–306 (2005).
26. Bhargav S, Panda BP, Ali M and Javed S, Solid-state fermen-tation: An overview. Chem Biochem Eng Q 22:49–70 (2008).
27. Papanikolaou S, Komaitis M and Aggelis G, Single cell oil (SCO) production by Mortierella isabellina grown on high-sugar content media. Bioresource Technol 95:287–291 (2004).
28. Zhu LY, Zong MH and Wu H, Effi cient lipid production with Trichosporon fermentans and its use for biodiesel preparation. Bioresource Technol 99:7881–7885 (2008).
29. Ratledge C, Biochemistry, stoichiometry, substrates and economics, in Single Cell Oil, ed by Moreton RS. Longman Scientifi c & Technical, London, pp. 33–70 (1988).
30. Beopoulos A, Nicaud JM and Gaillardin C, An overview of lipid metabolism in yeasts and its impact on biotechnological proc-esses. Appl Microbiol Biot 90:1193–1206 (2011).
31. Aden A, Ruth M, Ibsen K, Jechura J, Neeves K, Sheehan J et al., Lignocellulosic biomass to ethanol process design and economics utilizing co-current dilute acid prehydrolysis an
enzymatic hydrolysis for corn stover, report no. NREL/TP-510-32438. National Renewable Energy Laboratory, Golden, CO (2002).
32. Cardona Alzate CA and Sánchez Toro OJ, Energy consumption analysis of integrated fl owsheets for production of fuel ethanol from lignocellulosic biomass. Energy 31:2447–2459 (2006).
33. Luo L, van der Voet E and Huppes G, An energy analysis of ethanol from cellulosic feedstock–Corn stover. Renew Sust Energ Rev 13:2003–2011 (2009).
34. Piccolo C and Bezzo F, A techno-economic comparison between two technologies for bioethanol production from lignocellulose. Biomass Bioenerg 33:478–491 (2009).
35. Soderberg AC, Fermentation design, in Fermentation and Biochemical Engineering Handbook, ed by Vogel HC and Todaro CL. Noyes Publications, Westwood, pp. 67–121 (1997).
36. www.uscentrifuge.com/decanter-centrifuges.htm
37. www.labocon.com/Centrifuge-7.html
38. Esteban B, Baquero G, Puig R, Riba JR and Rius A, Is it envi-ronmentally advantageous to use vegetable oil directly as biofuel instead of converting it to biodiesel? Biomass Bioenerg 35:1317–1328 (2010).
39. Mercer P and Amerta AE, Development in oil extraction from microalgae. Eur J Lipid Sci Tech 113:539–547 (2011).
40. Samori C, Barreiro DL, Vet R, Pazzolesi L, Brilman DWF, Galletti P et al., Effective lipid extraction from algae cultures using switchable solvents. Green Chem 15:353 (2013).
41. van’t Riet K and Tramper J, Basic Bioreactor Design. Marcel Dekker, Inc., New York (1991).
42. Gowthaman MK, Rao K, Ghildyal NP and Karanth NG, Gas concentration and temperature-gradients in a packed-bed solid-state fermenter. Biotechnol Adv 11:611–620 (1993).
43. Weber FJ, Tramper J and Rinzema A, A simplifi ed material and energy balance approach for process development and scale-up of Coniothyrium minitans conidia production by solid-state cultivation in a packed-bed reactor. Biotechnol Bioeng 65:447–458 (1999).
44. Weber FJ, Oostra J, Tramper J and Rinzema A, Validation of a model for process development and scale-up of packed-bed solid-state bioreactors. Biotechnol Bioeng 77:381–393 (2002).
45. Kryvoruchko V, Machmuller A, Bodiroza V, Amon B and Amon T, Anaerobic digestion of by-products of sugar beet and starch potato processing. Biomass Bioenerg 33:620–627 (2009).
46. Collet P, Hélias A, Lardon L, Ras M, Goy RA and Steyer JP, Life-cycle assessment of microalgae culture coupled to biogas production. Bioresource Technol 102:207–214 (2011).
47. Larson ED, Jin HM and Celik FE, Large-scale gasifi cation-based coproduction of fuels and electricity from switchgrass. Biofuel Bioprod Bioref 3:174–194 (2009).
48. Lorenz F and Voss C, Wie viel energie benötigt die verdamp-fungstrocknung? Zuckerind 132:766–769 (2007).
49. Jorquera O, Kiperstok A, Sales EA, Embirucu M and Ghirardi ML, Comparative energy life-cycle analyses of microalgal biomass production in open ponds and photobioreactors. Bioresource Technol 101:1406–1413 (2010).
50. Werten MWT, Wisselink WH, van den Bosch TJJ, de Bruin EC and de Wolf FA, Secreted production of a custom-designed, highly hydrophilic gelatin in Pichia pastoris. Protein Eng 14:447–454 (2001).
51. Schipperus R, Teeuwen RLM, Werten MWT, Eggink G and de Wolf FA, Secreted production of an elastin-like polypeptide by Pichia pastoris. Appl Microbiol Biot 85:293–301 (2009).

Perspective: Lipids from yeasts and fungi P Meeuwse et al.
© 2013 Society of Chemical Industry and John Wiley & Sons, Ltd | Biofuels, Bioprod. Bioref. (2013); DOI: 10.1002/bbb
52. Vicente G, Bautista LF, Gutierrez FJ, Rodriguez R, Martinez V, Rodriguez-Frometa RA et al., Direct transformation of fungal biomass from submerged cultures into biodiesel. Energ Fuel 24:3173–3178 (2010).
53. Weusthuis RA, Lamot I, van der Oost J and Sanders JPM, Microbial production of bulk chemicals: Development of anaerobic processes. Trends Biotechnol 29:153–158 (2011).
54. Alvarez HM and Steinbuchel A, Triacylglycerols in prokaryotic microorganisms. Appl Microbiol Biot 60:367–376 (2002).
55. Jegannathan KR, Eng-Seng C and Ravindra P, Economic assessment of biodiesel production: Comparison of alkali and biocatalyst processes. Renew Sust Energ Rev 15:745–751 (2011).
56. Gnansounou E and Dauriat A, Techno-economic analysis of lignocellulosic ethanol: A review. Bioresource Technol 101:4980–4991 (2010).
57. Norsker NH, Barbosa MJ, Vermue MH and Wijffels RH, Microalgal production - A close look at the economics. Biotechnol Adv 29:24–27 (2011).
Petra Meeuwse
Petra Meeuwse received her PhD in 2011 at Wageningen University for research on the production of fungal lipids. This work included experimental work in both submerged and solid-state fermentation, as well as kinetic modeling and process design. She received her Msc degree in 2004 at
Wageningen University in bioprocess engineering. She currently works as a scientist at Dyadic Netherlands.
Arjen Rinzema
Dr Arjen Rinzema is Assist-ant Professor in the Bioprocess Engineering Group at Wageningen University. His area of expertise is bioreactor modelling, with empha-sis on solid-substrate cultivation of fungi. Dr Rinzema received his MSc in Chemical Engineering in 1983 from the
University of Groningen and his PhD in Environmental Engineering in 1988 from the Wageningen University.
Johan Sanders
Johan Sanders is Professor at Wage-ningen University of the Biobased Commodity Chemicals chair with the focus of reducing CO2 production in a cost effective way using chemistry, fermentation and biorefineries at large as well as at small scale to enable optimal application of all plant com-
ponents. He holds a PhD in Molecular Biology from the University of Amsterdam for researching the physical map of yeast mitochondrial DNA. From 1977 to 1993 he worked at Gist Brocades, starting the Genetic Engineer-ing group and working on various projects in the field of enzyme research; he became Associate Director of Food Research. From 1993 to 2001 Sanders worked at AVEBE as R&D Director focusing on the enzymatic and genetic modification of starch.
Johannes Tramper
Dr Johannes Tramper is Professor of Bioprocess Technology at Wagenin-gen University. In addition to his main activities in teaching and research, he also spends much time and effort informing the wider public about mod-ern biotechnology. He has degrees from the Technical University of Delft
(Ir/MSc), Purdue University (MSc) and Wageningen University (PhD).




















