
Coordination Between Fission Yeast Glucan Formation and Growth Requires a Sphingolipase Activity
15
Copyright 2001 by the Genetics Society of America Coordination Between Fission Yeast Glucan Formation and Growth Requires a Sphingolipase Activity Anna Feoktistova,* Paula Magnelli, † Claudia Abeijon, † Pilar Perez, ‡ Robert L. Lester, § Robert C. Dickson, § and Kathleen L. Gould* , ** *Howard Hughes Medical Institute and **Department of Cell Biology, Vanderbilt University School of Medicine, Nashville, Tennessee 37232, † Department of Molecular and Cell Biology, Boston University-Goldman School of Dental Medicine, Boston, Massachusetts 02118, ‡ Instituto de Microbiologia Bioquimica, CSIC, Universidad de Salamanca, Edificio Departmental, 37007 Salamanca, Spain and § Department of Biochemistry and the Lucille P. Markey Cancer Center, University of Kentucky College of Medicine, Lexington, Kentucky 40536-0096 Manuscript received April 3, 2001 Accepted for publication May 3, 2001 ABSTRACT css1 mutants display a novel defect in Schizosaccharomyces pombe cell wall formation. The mutant cells are temperature-sensitive and accumulate large deposits of material that stain with calcofluor and aniline blue in their periplasmic space. Biochemical analyses of this material indicate that it consists of - and -glucans in the same ratio as found in cell walls of wild-type S. pombe. Strikingly, the glucan deposits in css1 mutant cells do not affect their overall morphology. The cells remain rod shaped, and the thickness of their walls is unaltered. Css1p is an essential protein related to mammalian neutral sphingomyelinase and is responsible for the inositolphosphosphingolipid-phospholipase C activity observed in S. pombe membranes. Further- more, expression of css1 can compensate for loss of ISC1, the enzyme responsible for this activity in Saccharomyces cerevisiae membranes. Css1p localizes to the entire plasma membrane and secretory pathway; a C-terminal fragment of Css1p, predicted to encode a single membrane-spanning segment, is sufficient to direct membrane localization of the heterologous protein, GFP. Our results predict the existence of an enzyme(s) or process(es) essential for the coordination of S. pombe cell wall formation and division that is, in turn, regulated by a sphingolipid metabolite. S CHIZOSACCHAROMYCES pombe cells, like other tomannans; alkali-soluble -(1,3)-, -(1,6)-, and -(1,3)- linked glucans; and an alkali-insoluble -(1,3)-linked yeast, are surrounded by a rigid cell wall that pro- vides mechanical strength and protection from environ- glucan with the -(1,3)-linked glucans forming the ma- jority of cell wall polysaccharide (Bush et al. 1974; Man- mental stresses (Cabib et al. 1997). The wall is required for the formation and maintenance of cell shape and ners and Meyer 1977). These components are arranged in distinct layers that can be visualized by electron mi- is remodeled continuously during the cell cycle to allow cell expansion. Cell wall remodeling also occurs in re- croscopy (Kopecka et al. 1995; Osumi et al. 1998). There is an outer shell comprised of glycoproteins, a middle sponse to extracellular cues that trigger shape changes, such as nutrient deprivation and exposure to mating layer of amorphous glucan material, and an inner layer of densely woven glucan fibrils. Several S. pombe galacto- factors. During vegetative growth, S. pombe cells are rod shaped and grow by tip elongation, first at the old end syltransferases, Golgi enzymes required for the produc- tion of cell wall glycoproteins, have been identified and and then at both ends. Subsequently, cell wall deposi- tion occurs during septation and the two new ends are characterized (Chappell and Warren 1989; Chappell et al. 1994; Yoko-o et al. 1998). Several enzymes also thus sealed off (Mitchison and Nurse 1985). Patches of F-actin are concentrated at these areas of growth, have been identified that are required for the synthesis of - and -glucans. There are at least five -glucan and it is a widely held view that they direct wall formation to these sites (Marks et al. 1987; Nurse 1994). Electron synthases of which one, termed Mok1p/Ags1p, has been characterized (Hochstenbach et al. 1998; Katayama micrographic studies of regenerating S. pombe proto- plasts have substantiated this view by demonstrating an et al. 1999). There is also a family of -glucan synthases intimate association between F-actin structures and sites in S. pombe. One of these, Cps1p, is critical for the forma- of new cell wall formation (Osumi et al. 1998). tion of the septum (Ishiguro et al. 1997; Le Goff et al. The carbohydrate composition of the S. pombe cell 1999; Liu et al. 1999). Another, Bgs2p, is essential for wall has been known for some time to consist of -galac- maturation of the spore wall (Liu et al. 2000; Martin et al. 2000). Mok1p/Ags1p and Cps1p are essential for cell wall integrity and viability of vegetatively growing Corresponding author: Kathleen L. Gould, HHMI and Department cells. In their absence, the cell wall becomes less well of Cell Biology, B2309 MCN, Vanderbilt University School of Medi- organized, thicker, and more sensitive to degradation, cine, 1161 21st Ave. S., Nashville, TN 37232. E-mail: [email protected] and the cells become rounded, swollen, and eventually Genetics 158: 1397–1411 (August 2001)
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Coordination Between Fission Yeast Glucan Formation and Growth Requires a Sphingolipase Activity

Copyright 2001 by the Genetics Society of America
Coordination Between Fission Yeast Glucan Formation and GrowthRequires a Sphingolipase Activity
Anna Feoktistova,* Paula Magnelli,† Claudia Abeijon,† Pilar Perez,‡
Robert L. Lester,§ Robert C. Dickson,§ and Kathleen L. Gould*,**
*Howard Hughes Medical Institute and **Department of Cell Biology, Vanderbilt University School of Medicine, Nashville, Tennessee 37232,†Department of Molecular and Cell Biology, Boston University-Goldman School of Dental Medicine, Boston, Massachusetts 02118, ‡Instituto
de Microbiologia Bioquimica, CSIC, Universidad de Salamanca, Edificio Departmental, 37007 Salamanca, Spain and §Department ofBiochemistry and the Lucille P. Markey Cancer Center, University of Kentucky College of Medicine, Lexington, Kentucky 40536-0096
Manuscript received April 3, 2001Accepted for publication May 3, 2001
ABSTRACTcss1 mutants display a novel defect in Schizosaccharomyces pombe cell wall formation. The mutant cells are
temperature-sensitive and accumulate large deposits of material that stain with calcofluor and aniline bluein their periplasmic space. Biochemical analyses of this material indicate that it consists of �- and �-glucansin the same ratio as found in cell walls of wild-type S. pombe. Strikingly, the glucan deposits in css1 mutantcells do not affect their overall morphology. The cells remain rod shaped, and the thickness of their wallsis unaltered. Css1p is an essential protein related to mammalian neutral sphingomyelinase and is responsiblefor the inositolphosphosphingolipid-phospholipase C activity observed in S. pombe membranes. Further-more, expression of css1� can compensate for loss of ISC1, the enzyme responsible for this activity inSaccharomyces cerevisiae membranes. Css1p localizes to the entire plasma membrane and secretory pathway;a C-terminal fragment of Css1p, predicted to encode a single membrane-spanning segment, is sufficientto direct membrane localization of the heterologous protein, GFP. Our results predict the existence ofan enzyme(s) or process(es) essential for the coordination of S. pombe cell wall formation and divisionthat is, in turn, regulated by a sphingolipid metabolite.
SCHIZOSACCHAROMYCES pombe cells, like other tomannans; alkali-soluble �-(1,3)-, �-(1,6)-, and �-(1,3)-linked glucans; and an alkali-insoluble �-(1,3)-linkedyeast, are surrounded by a rigid cell wall that pro-
vides mechanical strength and protection from environ- glucan with the �-(1,3)-linked glucans forming the ma-jority of cell wall polysaccharide (Bush et al. 1974; Man-mental stresses (Cabib et al. 1997). The wall is required
for the formation and maintenance of cell shape and ners and Meyer 1977). These components are arrangedin distinct layers that can be visualized by electron mi-is remodeled continuously during the cell cycle to allow
cell expansion. Cell wall remodeling also occurs in re- croscopy (Kopecka et al. 1995; Osumi et al. 1998). Thereis an outer shell comprised of glycoproteins, a middlesponse to extracellular cues that trigger shape changes,
such as nutrient deprivation and exposure to mating layer of amorphous glucan material, and an inner layerof densely woven glucan fibrils. Several S. pombe galacto-factors. During vegetative growth, S. pombe cells are rod
shaped and grow by tip elongation, first at the old end syltransferases, Golgi enzymes required for the produc-tion of cell wall glycoproteins, have been identified andand then at both ends. Subsequently, cell wall deposi-
tion occurs during septation and the two new ends are characterized (Chappell and Warren 1989; Chappellet al. 1994; Yoko-o et al. 1998). Several enzymes alsothus sealed off (Mitchison and Nurse 1985). Patches
of F-actin are concentrated at these areas of growth, have been identified that are required for the synthesisof �- and �-glucans. There are at least five �-glucanand it is a widely held view that they direct wall formation
to these sites (Marks et al. 1987; Nurse 1994). Electron synthases of which one, termed Mok1p/Ags1p, has beencharacterized (Hochstenbach et al. 1998; Katayamamicrographic studies of regenerating S. pombe proto-
plasts have substantiated this view by demonstrating an et al. 1999). There is also a family of �-glucan synthasesintimate association between F-actin structures and sites in S. pombe. One of these, Cps1p, is critical for the forma-of new cell wall formation (Osumi et al. 1998). tion of the septum (Ishiguro et al. 1997; Le Goff et al.
The carbohydrate composition of the S. pombe cell 1999; Liu et al. 1999). Another, Bgs2p, is essential forwall has been known for some time to consist of �-galac- maturation of the spore wall (Liu et al. 2000; Martin
et al. 2000). Mok1p/Ags1p and Cps1p are essential forcell wall integrity and viability of vegetatively growing
Corresponding author: Kathleen L. Gould, HHMI and Department cells. In their absence, the cell wall becomes less wellof Cell Biology, B2309 MCN, Vanderbilt University School of Medi-
organized, thicker, and more sensitive to degradation,cine, 1161 21st Ave. S., Nashville, TN 37232.E-mail: [email protected] and the cells become rounded, swollen, and eventually
Genetics 158: 1397–1411 (August 2001)

1398 A. Feoktistova et al.
for dominance, the css1-3 strain was crossed to a mei1-102 strainlyse unless provided with osmostabilizing medium. The(mei1-102 mutants will mate with h� partners, but the resultingactivities of the glucan synthases are regulated in a com-diploids will not sporulate) and stable diploids were obtained
plex manner by the activities of Rho family GTPases using complementation of nutritional markers.and protein kinase C’s (reviewed in Arellano et al. The parental S. cerevisiae strain was diploid JK9-3d (homozy-
gous for leu2-3, 112 ura3-52 rme1 trp1 his4) and MATa/MAT�1999). Under- or overproduction of these regulators(Heitman et al. 1991). RCD319 is a MATa haploid derivativedramatically affects S. pombe morphology and cell wallof JK9-3d carrying the yer019-�1::Kan allele, which has basesintegrity. Indeed, in such studies, considerable progress�4 to �967 replaced by a lox-kanamycin-lox cassette (Guldener
was made in identifying and understanding the signal- et al. 1996). YPD (Sherman 1991) and S-leu (Skrzypek et al.ing pathways governing the synthesis of cell wall compo- 1998) media were used to culture S. cerevisiae cells from which
membranes were made. S. cerevisiae cells were grown in PYEDnents. In contrast, virtually nothing is understood in S.medium (Skrzypek et al. 1998) to radiolabel sphingolipids.pombe about how the various components of the wall
Plasmids and molecular biological techniques: All plasmidare organized into a functional unit subsequent to theirmanipulations and bacterial transformations were by standard
synthesis. techniques (Sambrook et al. 1989). Essential features of plas-Here, we present the isolation and characterization mid construction are described. All sequencing of plasmid
DNA was performed using Thermo Sequenase (Amershamof css1 mutants that display a novel defect in cell wallLife Sciences, Cleveland, OH) according to manufacturer’sformation. In these mutants, a large excess of �- andinstructions. Yeast genomic DNA was isolated as described
�-glucans accumulates in the periplasmic space without (Moreno et al. 1991; Hoffman 1993).being incorporated into the cell wall and without affect- For expression of css1� under control of the nmt1 promoter,ing the rod shape of the cells. We present evidence that a NdeI site was introduced at the initiating methionine codon
by PCR amplification of the open reading frame (ORF) withCss1p, related to human neutral sphingomyelinase, isPfu polymerase. A BamHI site was also introduced just 3� ofan essential membrane protein that functions as an ino-the termination codon. The PCR fragment was subcloned intositolphosphosphingolipid-phospholipase C in S. pombe. pZERO (Invitrogen, San Diego). The css1� ORF was then
Css1p can also substitute for loss of this enzyme activity removed with XhoI and BamHI and subcloned into the pREPXseries of fission yeast expression plasmids (Basi et al. 1993;in Saccharomyces cerevisiae encoded by ISC1 (Wells et al.Forsburg 1993; Maundrell 1993). For expression in S. cere-1998; Sawai et al. 2000). Many agonists and stresses ac-visiae, the same fragment was subcloned into the expressiontivate mammalian sphingomyelinases with subsequentvector pRS425GAL1 (Mumberg et al. 1994). By dropping out
hydrolysis of sphingomyelin in the plasma membrane to the internal NdeI fragment of pKG1560 (pREP1GFPcss1), thegenerate phosphocholine and ceramide, an intracellu- plasmid pKG1845 was created that fused green fluorescent
protein (GFP) to Css1p residues 332–424. GFP fusions tolar signaling molecule (Kolesnick and Kronke 1998;amino acids 332–389 and 332–663 of Css1p were made byKronke 1999; Levade and Jaffrezou 1999). The agonistsamplification of css1� sequences with Pfu polymerase usingand stresses include cytokines, interleukin 1� and Fas oligonucleotides 5�-CGTCTTCATATGAGACTTCG-3� and ei-
ligands, chemotherapeutic drugs, environmental stresses, ther 5�-CTAGGATCCTTATCTACCAAATAGCAATCC-3� orand injury or infections. Ceramide is thought to regulate 5�-CTAGGATCCTTACTTTAACCAAGCGGG-3�, respectively.
The PCR products were digested with NdeI and BamHI andsignaling pathways that control growth arrest, apoptosis,subcloned into pREP1GFP (pKG1538) to generate pKG1528senescence, and immune responses in mammals. Pro-and pKG1974. A GFP fusion protein to Css1p residues 1–331cesses regulated by sphingolipases have not been identi- was created by subcloning the css1� NdeI fragment into
fied in fungi (Dickson 1998). Our results suggest that pKG1538. A GFP fusion protein with Css1p residues 1–389Css1p hydrolyzes complex sphingolipids to yield cera- was created by inserting the css1� NdeI fragment (encoding
residues 1–331) into the unique NdeI site of pKG1528 to createmide, which regulates a pathway or process essentialpKG2138.for the coordination between cell wall formation and
Genomic cloning and DNA sequencing: The css1-3 straingrowth in S. pombe. This is the first example of a sphingol- was transformed with a S. pombe genomic library constructedipase that is essential for fungal cell growth. in the plasmid vector pUR19 (Barbet et al. 1992), and three
Ura� colonies that could grow at 36� were isolated. PlasmidDNAs were recovered from these colonies and a comparison ofthe genomic DNA inserts by restriction mapping and SouthernMATERIALS AND METHODSblotting revealed that they contained overlapping DNA se-
Yeast strains, methods, and media: S. pombe strains used in quences. The region of these clones responsible for comple-this study are listed in Table 1. They were grown in yeast mentation of css1-3 was narrowed by constructing subclonesextract medium (YE) or minimal medium with appropriate and testing them for their ability to rescue. The DNA se-supplements (Moreno et al. 1991). Crosses were performed quences of the ends of the smallest rescuing fragment (MluI-on glutamate medium (minimal medium lacking ammonium MluI) were then determined and compared to sequenceschloride and containing 0.01 m glutamate, pH 5.6). Random within the databases. At one end, the DNA sequence matchedspore and tetrad analyses were performed as described a terminal segment of cosmid c32F12 in the S. pombe genome(Moreno et al. 1991). Transformations were performed by sequencing project database. The other end did not match aelectroporation (Prentice 1992). For regulated expression sequence within this database but did match a region adjacentof css1� by the nmt1 promoter (Maundrell 1993), cells were to the orp2� gene (Leatherwood et al. 1996). However, theregrown either in minimal media lacking thiamine to allow was an �800-bp gap between the ends of these DNA fragments.expression or with the addition of 5 �m thiamine to repress The DNA sequence of this gap region was then determinedexpression. Double mutant strains were constructed and iden- on both strands using custom-synthesized oligonucleotides
that allowed the identification of a single open reading frame.tified by tetrad analysis or by random spore analysis. To test

1399Essential S. pombe Sphingolipase
TABLE 1
S. pombe strains used in this study
Strain Genotype Source
KGY28 h� 972 wild type Our stockKGY246 h� leu1-32 ura4-D18 ade6-M210 Our stockKGY1145 h� css1-3 leu1-32 ura4-D18 ade6-M210 This studyKGY1205 h� css1-5 ura4-D18 leu1-32 ade6-M210 This studyKGY2037 mei1-102/mei1� css1-3/css1� lys1-3/lys1� leu1-32/leu1� ura4-D18/ura4� ade6-M210/ade6� This studyKGY2351 h� css1::css1-myc/Kanr ura4-D18 leu1-32 ade6-M210 This studyKGY2378 h� css1-2 ura4-D18 leu1-32 ade6-M210 This studyKGY2380 h� css1-4 ura4-D18 leu1-32 ade6-M210 This studyKGY2482 h� css1-2/css1::ura4� leu1-32/leu1-32 ura4-D18/ura4-D18 ade6-M210/ade6-M216 This studyKGY2550 h� css1-2 This study
For integration mapping, the genomic fragment shown in �-(1,3)-linked glucan (Smith and McCully 1978). For immu-nostaining, cells were fixed with a mixture of formaldehydeFigure 4A was subcloned into pJK210, an integrating vector
that contains the ura4� selectable marker (Keeney and Boeke and glutaraldehyde (Balasubramanian et al. 1997) andstained with 9E10 monoclonal antibody followed by Alexa-1994). This construct was linearized with NdeI and integrated
into the genome of the css1-3 ura4-D18 strain by homologous conjugated goat anti-mouse IgG. GFP-Css1p fusion proteinswere visualized in live cells.recombination. In outcrosses of the integrant strain to wild
type, the ura4� marker segregated with a css1� phenotype in Analysis of cell wall polysaccharides: Two slightly differentprocedures were used to analyze the cell wall polysaccharide14 out of 14 tetrads analyzed.
For sequencing of css1 mutant alleles, genomic DNA was composition. Both are based on the fact that the metabolicconversion of uniformly labeled [14C]glucose to labeled man-prepared from the relevant strains and the css1 coding region
was amplified by PCR with the following oligonucleotides: nose and galactose occurs rapidly, leading to a homogeneouslabeling of every cell wall component. In the first procedure,css1-F6 (5�-TTGACGGCAATAACTCCG-3�) and css1-R6 (5�-TA
AATTTATGATTTAGC-3�). The PCR products were sequenced labeling and fractionation of cell wall polysaccharides wasdone as described (Arellano et al. 1997) and is presented indirectly using the thermosequenase cycle sequencing kit
(Amersham) and customized oligonucleotides. Figure 3A. Briefly, exponentially growing cultures of S. pombe,incubated at 25�, were supplemented with [U-14C]glucoseGene deletion: To delete css1� from the S. pombe genome, a
HindIII fragment containing the majority of the open reading (1 �Ci/ml) and incubated for an additional 4 hr at differenttemperatures (25�, 28�, 32�, and 37�). Cells were harvestedframe was replaced with the ura4� gene (see Figure 4). A
linear fragment from this construct including css1� 5� and 3� and total glucose incorporation was monitored by measuringthe radioactivity of the TCA insoluble material. Mechanicalflanking sequences was then transformed into a diploid strain
with the relevant genotype of css1�/css1-2. Stable Ura� trans- breakage of cells was done using prechilled glass beads addedto the cells, and lysis was achieved in a Fast-Prep system FP120formants were selected at 25� and then screened for tempera-
ture-sensitive growth. Temperature-sensitive diploids that (BIO 101, Vista, CA; Savant, Farmingdale, NY), using two15-sec intervals at 5.5 speed. Cell walls were pelleted at 1000 were presumably css1::ura4�/css1-2 were screened by PCR and
Southern blotting for the appropriate gene replacement. g for 5 min and washed with 5% NaCl three times and with1 mm EDTA three times. Aliquots (100 �l) of the total wallEpitope tagging of css1�: A genomic version of css1 encoding
13 copies of the myc epitope fused to the C terminus of were incubated with 100 units of Zymolyase 100T (SeikagakuAmerica, Ijamsville, MD) or Quantazyme (Quantum Biotech-Css1p was generated by the method described in Bahler et
al. (1998). Oligonucleotide primers were used to amplify the nologies, Blaine, WA) for 36 hr at 30�. Aliquots without enzymewere included as control. The samples were centrifuged andmyc13-kan cassette (Bahler et al. 1998), and the PCR frag-
ment was transformed into diploid cells (KGY1218). The cells the supernatant and washed pellet were counted separately.The supernatant from the Zymolyase 100T reaction was con-were allowed to recover for 12 hr on YE plates and then replica
plated onto YE plates containing 100 �g/ml G418 (Geneticin; sidered to be �-glucan plus galactomannan and the pellet tobe �-glucan. The supernatant from the Quantazyme reactionsGIBCO BRL, Grand Island, NY). G418-resistant colonies were
screened for homologous recombinants by PCR; the presence was considered to be �-glucan and the pellet to be �-glucanplus galactomannan.of the epitope was confirmed by immunoblotting with the
9E10 antibody. Sporulation and tetrad analysis showed 2:2 In the second procedure (Figure 3B and Table 2), wild-type and css1-2 cells were grown at 23� in 75 ml of YE 1%segregation of the G418 resistance, confirming that the hap-
loid strain, css1::css1-myc, was viable. glucose, 1 �Ci/ml d-[14C(U)]glucose (300Ci/mmol; Ameri-can Radiochemical Corporation, St. Louis) until an OD600 ofFluorescence microscopy: All fluorescence microscopy was
performed using a Zeiss axioscope and the appropriate set of 0.3. Cultures were either harvested or shifted to 36� for 12 hrbefore harvesting. Cells were washed in PBS-10 mm sodiumfilters. Calcofluor was used to detect cell wall material in live
or formaldehyde-fixed cells, DNA was visualized using 4�,6- azide, resuspended in PBS containing 1 mm phenylmethylsul-fonyl fluoride (PMSF), and subjected to mechanical breakagediamidino-2-phenylindole (DAPI), and actin was visualized
using rhodamine-conjugated phalloidin as described (Bala- with glass beads until 95% of the cells had lysed (microscopicobservation). The glass beads were allowed to settle and thesubramanian et al. 1997). The cell wall was also visualized by
staining formaldehyde-fixed cells with 0.5 mg/ml methyl blue recovered homogenate, together with three washes, was centri-fuged at 1600 g for 10 min to pellet the cell wall. The(M-6900; Sigma, St. Louis) in PBS (Kippert and Lloyd 1995).
Methyl blue contains aniline blue that stains specifically supernatant representing the total intracellular material was

1400 A. Feoktistova et al.
then separated into a 100,000-g pellet (intracellular organ- tube (Kimble no. 73785, 15 ml, screw thread, Teflon-linedcap) under a stream of N2. The dried lipids were suspendedelles) and cytosol. The total radioactivity incorporated into
every fraction was determined after total acid hydrolysis by in 270 �l of buffer [final concentrations: 250 mm 2-(N-Mor-pholino)ethanesulfonic acid monohydrate (pH 6.0), 0.6%liquid scintillation counting. In wild-type cells at both tempera-
tures, as well as the css1-2 mutant at 23�, 15–20% of the total n-octyl-�-glucoside, 5 mm MgCl2] and treated for 10 min in asonic water bath. Membrane protein (30 �l, 75–300 �g) wasradioactivity incorporated was found in the cell wall fraction.
In the css1-2 mutant grown at 36�, 50–55% of the total radioac- added and the reaction was incubated with intermittent shak-ing for up to 2 hr at 30�. The reaction was stopped by addingtivity incorporated was found in the cell wall fraction. The
wall fractions were boiled for 5 min, washed 10 times with 0.9 ml of 2.27% sodium dodecylsulfate and extracted twicewith an equal volume of methyl-tert-butylether. The upperPBS/1 mm PMSF, and frozen at �70�.
Cell walls were fractionated according to Manners and layer was transferred to a glass tube and extracted with 1 mlof water. The upper organic layer containing the [3H]cera-Meyer (1977) with minor modifications. Briefly, the walls
were extracted three times with 3% NaOH for 60 min at 75� mide was transferred to a glass scintillation vial, taken to dry-ness under a stream of N2, resuspended in 4.5 ml of Ultima-to obtain, after centrifugation, an alkali-soluble and an alkali-
insoluble fraction. The alkali-insoluble fraction contained Gold scintillation fluid (Packard, Meriden, CT), sonicated ina water bath for 10 min, and counted in a liquid scintillationabout one-half of the �-1,3 glucan, and the rest was in the
alkali-soluble fraction. To quantify the �-1,3 glucan, both frac- counter.Hydrolysis of radiolabeled S. pombe mannose-inositol-phos-tions were independently treated with �-1,3 glucanase (Zymo-
lyase 100T; Seikagaku America). This enzyme is not recombi- phoceramide (MIPC) was determined by drying 1.5 104 cpmof [3H]inositol-MIPC or 4000 cpm of [3H]dihydrosphingosine-nant but we determined that it was completely devoid of �-1,6
glucanase activity by its inability to hydrolyze purified �-1,6 MIPC in a glass screw-capped tube under a stream of N2,followed by suspension in reaction buffer (final concentra-glucan (pustulan). The digestion products were separated by
P4 (Bio-Rad, Richmond, CA) chromatography and character- tions: 25 mm potassium phosphate, pH 7.0, 2.5 mm dithiothrei-tol, 5 mm MgCl2, 0.6% n-octyl-�-glucoside). The sample wasized by thin-layer chromatography in Silica Gel G (solvent
system: butanol/ethanol/H2O; 5:3:2. The amount of laminari- sonicated as described above and warmed to 30�, and thereaction was started by adding 150 �g of membranes (totaloligosaccharides (�-1,3-linked glucose oligosaccharides) de-
tected served to quantify the �-1,3 glucan. The �-1,3 glucan volume of 150 �l). Reactions were incubated for up to 2 hrand stopped by addition of 15 �l of 0.5 m disodium ethylenedi-was obtained in the void volume of the P4 chromatography
of �-1,3 glucanase-treated alkalki-soluble fraction. The galacto- amine acetate, pH 7.1. Samples containing [3H]inositol-MIPCwere heated in a boiling water bath for 2 min followed bymannan was obtained from the alkali-soluble fraction by selec-
tive precipitation (Fehling’s reagent). Monosaccharide com- 2-min treatment in a sonic water bath. Reactions containing[3H]dihydrosphingosine-MIPC were vortexed with organic sol-position indicated the presence of mannose, galactose, and
�5% glucose. vent (5 volumes of sample:16 volumes chloroform:16 volumesmethanol) and centrifuged to pellet denatured proteins. InCell fractionation and immunoblotting: Protein lysates were
prepared by glass bead disruption of S. pombe cells in one of both cases 5–50 �l were chromatographed on a 15 15-cmpiece of silica gel-impregnated Whatman SG-81 paper treatedthree buffers: TEG (Katayama et al. 1999), NP-40, or SDS-
lysis (Gould et al. 1991). In the latter buffer, protein lysates with ethylenediamine tetraacetate (Steiner and Lester1972). The chromatogram was developed in chloroform/were heated immediately following lysis at 96� and diluted
with NP-40 buffer. Following removal by filtration of glass methanol/4.2 m ammonia (9:7:2, v/v/v), dried, and each lanewas cut into 2-cm wide by 1-cm tall pieces. The pieces werebeads, soluble and insoluble fractions of each lysate were sepa-
rated by microcentrifugation at 14,000 rpm for 20 min at 4�. placed in 4.5 ml of UltimaGold scintillation fluid and soni-cated for 3 min before counting in a liquid scintillationTo each supernatant fraction was added 1⁄5 volume of 5 SDS
gel sample buffer. Each pellet was resuspended in a volume counter. The background control sample lacked membranes.Membranes were prepared from 50–100 A600 units of S.of 1 SDS gel sample buffer equivalent to that of the total
supernatant fractions. Both fractions were then heated to 96� cerevisiae cells grown to an A600 of 3–6 in YPD medium at 30�.The cells were centrifuged at room temperature, washed twicefor 5 min. Equal volumes of soluble and insoluble fractions
were then resolved by 6–20% SDS-PAGE, transferred to poly- with distilled water, and resuspended in 1 ml of ice-cold extrac-tion buffer (50 mm potassium phosphate, pH 7.0, 5 mm dithi-vinylidene fluoride membrane, and Css1p-myc was visualized
by enhanced chemiluminescence (ECL) subsequent to incu- othreitol, 1 mm phenylmethylsulfonyl fluoride). One-half vol-ume of glass beads (0.5-mm diameter) was added, and thebation with the 9E10 antibody (1 �g/ml) and peroxidase-
conjugated goat anti-mouse IgG (1:50,000; Jackson Labora- sample was chilled on ice and then vortexed eight times for30 sec with a 30-sec chilling period between vortexing. Thetories, West Grove, PA).
Electron microscopy: For electron microscopy, 106 css1-2 sample was centrifuged for 10 min at 4000 g in a SorvallSS-34 rotor at 4�. The resulting supernatant fluid was centri-cells were fixed after a 6-hr shift to 36� in 1.5% potassium
permanganate. They were then treated with 2% glutaralde- fuged for 1 hr at 100,000 g in a Beckman TLA100 rotor at 4�.The resulting membrane pellet was suspended in extractionhyde and osmium tetroxide, block stained with uranyl acetate,
and dehydrated in ethanol series. Cells were embedded in buffer by 20 up-and-down strokes in a glass tissue homoge-nizer. The centrifugation and suspension steps were repeatedspurr in BEEM capsules, which were polymerized under vac-
uum at 65�. Sections were cut to 75 �m with a Leica Ultracut, and the final membrane pellet was suspended in extractionbuffer containing 30% glycerol. Aliquots of 50 �l were frozenpicked onto grids, stained with uranyl acetate and lead citrate,
and imaged on a Philips 300 electron microscope at 60 kV. and stored at �80�. For enzyme assay, the frozen sample wasthawed on ice and used immediately. Membranes were pre-Phospholipase C assays: The phospholipase assays were per-
formed as described (Wells et al. 1998) with minor modifica- pared in the same way from S. pombe cells grown at 25� inyeast extract medium to an A600 of 0.5 (before the temperaturetions. For each reaction 4 104 cpm of [3H]dihydrosphingo-
sine-labeled inositolphosphoceramide (IPC-3), which has a shift) and switched to 36� for 2 hr of incubation. Proteinconcentration was determined by using the Bio-Rad DC pro-ceramide composed of phytosphingosine and �-OH-C26 fatty
acid (Lester and Dickson 1993), and 10 nmol of nonradioac- tein assay kit with bovine serum albumin as the standard.Radiolabeled sphingolipids: S. cerevisiae sphingolipids weretive IPC-3 (both from S. cerevisiae) were dried in a conical glass

1401Essential S. pombe Sphingolipase
labeled with [3H]inositol and purified using a published proce- time at the nonpermissive temperature by increasingdure (Wells et al. 1998). S. pombe sphingolipids were labeled amounts of “cell wall” material (Figure 1, D–G). Thewith [3H]inositol or [3H]dihydrosphingosine in a similar man-
relative percentages of cells containing material at onener except that the cells were grown in yeast extract medium.end of the cell, both ends of the cells, or in the middleThe [3H]inositol-labeled S. pombe MIPC was purified by acid
precipitation. Following extraction of lipids from cells using of the cells were quantified in two alleles (Figure 1, Hsolvent B (Wells et al. 1998), 37.5 �l of Glacial acetic acid and I). The localization of this material suggested thatwas added per milliliter of sample and stored overnight at 4�. the onset of this phenotype was independent of cellThe sample was centrifuged at 10,000 rpm in a Sorvall SS-34
cycle position.rotor for 15 min at 4�. The resulting precipitate was resus-Physiological analysis of css1 mutants: To determinepended in 0.5 ml of solvent C (chloroform/methanol/water
16:16:5, v/v/v) by warming to 60� and treating for 5 min in how long it took css1 mutant cells to arrest division,a sonic water bath. The sample was dried under a stream cell numbers of wild-type and css1-2 mutant cells wereof N2, resuspended in 0.5 ml of monomethylamine reagent quantified after shift to the nonpermissive temperature.(Clarke and Dawson 1981), and incubated 30 min at 50�
Whereas wild-type cells continued to grow exponen-to deacylate glycerolipids. After evaporation to dryness andtially, css1-2 cells divided at most once (Figure 2A). Dur-resuspension in solvent C, the radiopurity of the MIPC was
verified by chromatography on a Whatman HP-K thin layer ing the same experiment, a portion of css1-2 and wild-plate (200 �m, 10 20 cm) using chloroform/methanol/ type cells was released from 36� to 25� to determine4.2 m ammonia (9:7:2, v/v/v) as solvent. Radioactivity was whether the division arrest of css1-2 cells was reversible.located by using a BioScan apparatus. For [3H]inositol-sphin-
Unlike wild-type cells, css1-2 cells lost viability rapidlygolipids �90% of the counts were in MIPC, 5% were in IPC,upon incubation at 36�, indicating that their arrest isand �5% were in two small broad bands of unknown composi-
tion near the origin. The same procedures were used to purify terminal (Figure 2B).[3H]dihydrosphingosine-labeled MIPC, except that further Although our initial analysis from asynchronous cul-purification was achieved by chromatography on a Whatman tures indicated that there was no specific time in theHP-K TLC plate. The band of MIPC on the TLC was located
cell cycle when cell wall material was deposited in css1(BioScan apparatus), scraped from the plate, loaded into amutants, we wanted to determine whether materialglass column (6 ml no. 504394; Supelco, Bellefonte, PA), and
eluted with solvent C. Radioactive fractions (0.5 ml) were would be deposited preferentially at one site if the cellspooled, dried, and resuspended in 0.5 ml of solvent C. were synchronized prior to inactivation of Css1-2p. To
examine this, css1-2 cells in the G2 phase of the cellcycle were selected by elutriation and were shifted to
RESULTS36�. We observed that when the septation index beganto rise, 50–60% of the cells failed to proceed beyond thatPhenotype of css1 mutants: In a visual screen for tem-
perature-sensitive lethal mutants (Balasubramanian et point (Figure 2C) and accumulated calcofluor-stainablematerial in the medial region (Figure 2D). The remain-al. 1998), we identified four mutants in which the accu-
mulation of the nonvital dye, phloxin B, was uneven der of the cells underwent septation and cell divisionnormally and then accumulated material at the old endwithin the cells. On phloxin B-containing plates, por-
tions of these mutant cells remained white while other of the cell where growth had resumed (Figure 2D).These observations indicate that the material is likelyparts were pink. Pairwise crosses between these mutants
indicated that all contained mutations within the same, to be deposited at the site of active growth when Css1-2p is inactivated.or closely linked, gene, which we termed css1� (see
below). The css1 mutations were recessive; diploids with css1 mutants accumulate �- and �-glucans: In an effortto determine the identity of the accumulated material,the relevant genotype css1-3/css1� and css1-2/css1� grew
well at 36� (data not shown). We chose two mutants, we stained css1 mutant cells with aniline blue. This fluo-rochrome was reported to bind preferentially to �-(1,3)css1-2 and css1-3, for further phenotypic analyses.
To address why the mutant cells stained unevenly with glucans (Smith and McCully 1978) and to stain spe-cifically the cell wall and septum of S. pombe (Kippertphloxin B, they were grown in liquid culture, shifted to
36� for various periods of time, and stained with cal- and Lloyd 1995). The material in css1 mutants stainedwell with aniline blue (Figure 2E), indicating that at leastcofluor to visualize cell wall material (Figure 1, B and
C). By 6 hr, it was clear that the mutant cells were a portion of it was comprised of �-glucan. Treatment ofeither whole cells or isolated walls with recombinantaccumulating material that stained intensely with cal-
cofluor and by 24 hr at nonpermissive temperature, the �-1,3 glucanase (Quantazyme) completely inhibited ani-line blue binding (data not shown).cells were nearly filled with this material (Figure 1C).
Because the cell wall, particularly that within the S. pombe To identify the material more precisely, cell wall con-stituents were isolated and characterized after growingseptum, is normally the only structure to stain intensely
with calcofluor (Figure 1A), the mutant was designated the cells in the presence of [U-14C]glucose. Incorpora-tion of radioactive glucose into the cell wall of css1-2css1� for can’t stop synthesizing cell wall. When cells
were stained with phalloidin to visualize F-actin and cells was similar to that of wild type at 25�, slightly higherat 28�, and considerably higher than in the wild type atDAPI to visualize DNA, it became clear that the cyto-
plasm of css1 mutant cells was being compressed over 32� (from 32 to 45% of total glucose incorporated) or

1402 A. Feoktistova et al.
Figure 1.—Phenotype of thecss1 mutants. css1-2 and css1-3 cellswere grown at 25� to midlog phasein YE medium, shifted to 36� forthe indicated number of hours,and fixed with formaldehyde.Wild type grown at 25� (A) andcss1-2 cells (B and C) were thenstained with calcofluor, and css1-3cells were permeabilized andstained with phalloidin (D and F)and DAPI (E and G). Arrows indi-cate the regions accumulating thecalcofluor-staining material. Thepercentage of css1-3 (H) and css1-2(I) cells accumulating calcofluor-stainable material at one end,both ends, in the middle, or bothat the tips and in the middle wasquantified.
36� (from 33 to 53%; Figure 3A). A dramatic six- to sured in vitro, was not increased (data not shown), itis likely that feedback regulation to prevent cell wallsevenfold increase in the amount of �-1,3 and �-1,3
glucan was detected in css1-2 mutants grown for 12 hr formation in the absence of further division is lost incss1 mutants. In this scenario, the glucan synthases wouldat 36� (Figure 3B and Table 2). The ratio between �/�-
glucan was the same as that found in wild-type S. pombe continue to produce cell wall material that is not needed.Gene cloning: To explore the molecular basis for thecell walls, indicating a simultaneous increase in both
glucan polymers (Figure 3 and Table 2). On the other css1� phenotype, as well as to determine the phenotypeof a css1 null allele, we cloned the css1� gene by comple-hand the amount of galactomannan was not signifi-
cantly affected, leading to a drastic alteration in the mentation of the css1-3 mutant phenotype. Sequenceanalysis of the smallest rescuing fragment of the clonedglucan to galactomannan ratio. At 23� both wildtype
and css1-2 mutants exhibited 2–3 times more glucan DNAs (MluI-MluI in Figure 4A) revealed the presenceof a single ORF that encoded a predicted protein prod-than galactomannan. At 36� we detected almost 10 times
more glucan than galactomannan in the css1-2 mutant uct of 424 amino acids and 48 kD (Figure 4B). A data-base search with the predicted amino acid sequence of(Table 2). These changes are solely due to the css1
mutation because the temperature shift per se did not Css1p revealed a hypothetical protein of S. cerevisiaeencoded by ORF YER019w (now termed ISC1), whichcause major alterations in the cell wall composition of
wild-type S. pombe (Figure 3 and Table 2). These results was 37% identical to Css1p (Figure 4B). These yeastproteins shared significant sequence similarity, espe-suggest that css1 mutants, when grown at the restrictive
temperature, have lost the ability to properly coordinate cially within their N-terminal 160 amino acids, to mam-malian neutral sphingomyelinases (Tomiuk et al. 1998).glucan polymer synthesis with cell growth. Because the
specific activity of the (1,3)�-d-glucan synthase, mea- Like the mammalian enzymes, Css1p and Isc1p lack a

1403Essential S. pombe Sphingolipase
Figure 2.—Physiological analysis ofcss1 mutant cells. Wild-type and css1-2cells were grown at 25� and then shiftedto 36� for the indicated number of hours.(A) Cell number was determined in trip-licate. (B) To determine the percentageof survival, samples were taken at eachtime point after shift to 36�, cell numberwas determined, and the same numberof cells was plated in triplicate. The num-ber of colonies to form at 25� after 5days was then quantified. (C and D)css1-2 cells were synchronized in earlyG2 phase by centrifugal elutriation andshifted to 36� for the indicated numberof hours. The septation index was deter-mined (C) and the positions where cal-cofluor-stainable material was depositedand accumulated were also determined(D). Abnormal deposits of material wereobserved in the middle of the cells andalso at old tips. (E) The calcofluor-stain-able material also binds the �-glucan-specific dye aniline blue. Wild-type cellswere grown at 25� and css1-2 cells weregrown at 25� to midlog phase in YE me-dium, shifted to 36� for 12 hr, and thenstained with aniline blue.
signal sequence at their N terminus, and a hydropathy copy suppressor was confirmed by integration mapping.In addition, the css1 mutant alleles were sequenced andplot suggests the presence of two adjacent membrane-
spanning domains at the Css1p C terminus (Figure 4C). found to contain single point mutations that resultedin single amino acid substitutions. In the css1-2 allele,The mammalian enzymes were found to be similar to
a large family of Mg2�-dependent phosphodiesterases glycine 18 was altered to arginine; in the css1-3 allele,alanine 291 was altered to valine; in the css1-4 al-(Tomiuk et al. 1998). In fact, subsequent mutagenesis
of certain conserved residues predicted to be involved lele, arginine 255 was altered to histidine; in the css1-5allele, glycine 15 was altered to glutamic acid (Figurein catalysis abrogates function of the mammalian en-
zyme in vitro (Rodrigues-Lima et al. 2000; Tomiuk et al. 4B). Thus, we concluded that the cloned DNA repre-sented the css1� gene.2000). Css1p contains these glutamic acid and histidine
residues, which are predicted to be involved in substrate To determine whether css1� was an essential gene,one copy of the gene was replaced with the ura4� genebinding and catalysis, respectively, of this family of en-
zymes (Figure 4B). in a diploid with the relevant genotype css1� ura4-D18/css1-2 ura4-D18 by the one-step gene deletion method.That the rescuing gene encoded css1� and not a high

1404 A. Feoktistova et al.
ization of Css1p, the chromosomal css1� locus was modi-fied to encode a protein with 13 copies of the mycepitope at its C terminus. Since the haploid strain pro-ducing Css1p-myc grew with morphology and kineticsindistinguishable from a wild-type strain, the tagged ver-sion of Css1p was functional. Because Css1p is predictedto contain two transmembrane domains, we testedwhether Css1p was a membrane-associated protein.Css1p-myc and wild-type cells were lysed in buffers con-taining no detergent, 1% NP-40, or 1% SDS, and solubleand insoluble fractions were separated by centrifuga-tion. Immunoblot analysis of the fractions indicated thatdetergent was required to release significant amounts ofCss1p into the soluble pool (Figure 4D). It also indicatedthat Css1p released into the supernatant was readilydegraded. Indeed, in NP-40 buffer, no soluble full-length protein was detected (Figure 4D, lane 3).
To determine the intracellular localization of Css1p,asynchronously growing css1myc cells were stained withantibodies to the myc epitope (9E10; Figure 5, A andB). Specific staining outlined the cell and nuclear pe-ripheries. Css1p-myc was also detected in specklesthroughout the cytoplasm and was excluded from nu-clei. These staining patterns are consistent with the lo-calization of Css1p to membrane structures, especiallythe plasma membrane, and to the secretory pathway.To confirm this localization pattern, the css1� ORF wasappended in frame to the 3� end of the gene encodingGFP and expressed in wild-type cells under control ofthe thiamine-repressible nmt1 promoter. Like Css1p-myc, GFP-Css1p was detected primarily around the en-tire cell periphery and in structures resembling vesiclesthroughout the cytoplasm (Figure 5C). Weak stainingaround the nuclei could be observed in some cells.
Figure 3.—Composition of the cell wall in the css1 mutant. To determine whether the predicted transmembrane(A) The relative levels of [14C]glucose radioactivity incorpo- domains were required for the observed localizationrated into each cell wall polysaccharide in a 4-hr labeling are
pattern, we removed from the GFP fusion the C-terminalshown for the wild-type (972h�) and css1-2 (KGY2550) strainssequences encoding the last 92 amino acids that aregrown at different temperatures. Values are the means of five
independent experiments with duplicated samples. Standard predicted to contain the transmembrane domains. Thedeviations for the total carbohydrate values are shown. (B) resultant protein, GFP-Css1�Cp, was present through-The levels of radioactivity from [14C]glucose incorporated into out the cytoplasm and excluded from the nucleus. Un-cell wall polysaccharides from wild-type and css1-2 mutants
like the full-length protein, it was not detected at the cellare shown. Values per milligram of wet weight are meansperiphery, at the division plane, or around the nucleusof duplicate cultures. Measurements of individual cell wall
fractions were done in triplicate for each culture. Cells were (Figure 5D). Overproduced GFP-Css1�Cp also ap-grown for 10 hr at 23� in YE, 1% glucose (1 �Ci [U-14C]glu- peared to form aggregates in some cells. To determinecose/ml), and then harvested or shifted to 36� for 12 hr before whether the last 92 amino acids (332–424) of Css1pharvesting. Cell walls were isolated and fractionated as de-
were sufficient for membrane localization, they werescribed in materials and methods. Exact values and stan-fused to GFP. GFP-Css1p332-424 was localized to thedard deviations are shown in Table 2.cell periphery and to speckles; its staining around thenuclear periphery was much more intense than thatobserved with the full-length protein (Figure 5E). ToUra4� temperature-sensitive diploids that had the cor-
rect gene replacement, as determined by PCR, were determine whether the two putative transmembrane do-mains of Css1p were sufficient for membrane localiza-sporulated. In each tetrad, only two Ura� temperature-
sensitive colonies formed (data not shown), indicating tion, Css1p residues 332–389 were fused to GFP. Again,we observed localization of this GFP fusion protein tothat css1� was indeed an essential gene. In contrast, ISC1
is not essential for S. cerevisiae viability. the nuclear and cell peripheries (Figure 5F). Finally,we fused amino acids 332–363 encoding just the firstIntracellular localization of Css1p: To study the local-

1405Essential S. pombe Sphingolipase
TABLE 2
Incorporation of radioactivity from [14C]glucose into cell wall polysaccharides in counts per minute per wet weight
�-/ GlucanStrain Temp. Galactomannan 36�/23� �-1,3-Glucan 36�/23� �-1,3-Glucan 36�/23� �-Glucan g-mannan
css1-2 23� 6,814 � 698 1 2,761 � 486 1 12,891 � 1,160 1 4.7 2.3css1-2 36� 10,509 � 1,001 1.5 20,717 � 574 7.5 79,923 � 3,048 6.2 3.8 9.6WT 23� 6,412 � 668 1 3,641 � 473 1 15,618 � 1,975 1 4.2 3.0WT 36� 6,263 � 579 0.98 4,892 � 181 1.3 15,461 � 1,056 0.99 3.1 3.2
The numbers represent the average � standard deviation of duplicate cultures. Measurements of individual cell wall fractionswere done in triplicate for each culture. The combined recovery of radioactivity in the different fractions was at least 80% ofthe total label incorporated into cell wall. These results are representative of those obtained in three independent experiments.
transmembrane domain to GFP and found that this to form inositolphosphosphingolipids (Dickson andLester 1999). We had previously determined that S.fusion protein localized to the plasma membrane and
apparently to membranes surrounding the nucleus (Fig- pombe contains primarily MIPC and a small amount ofIPC (G. B. Wells and R. L. Lester, unpublished re-ure 5G). We conclude from these data that the first
transmembrane domain of Css1p is necessary and suffi- sults). One hypothesis that would explain why css1-2cells stop growing at 36� is that they have an abnormalcient for its membrane localization. Consistent with a
putative function at the membrane, full-length GFP- amount of sphingolipids due to loss of Css1p activity.We examined this hypothesis by measuring the accumu-Css1p but not GFP-Css1�Cp was able to rescue growth
of css1-2 (data not shown). GFP-Css1p1-389, which lacks lation of radioactive sphingolipids following a shift ofcells to the restrictive temperature. At the time of shift-only sequences C-terminal to the transmembrane do-
mains, was able to localize correctly to membranes (Fig- ing log phase cells from 25� to 36�, [3H]inositol wasadded to the culture, which was incubated for 4 hr.ure 5H) but was unable to rescue growth of css1-2 (data
not shown). This observation suggests that this C-termi- Lipids were then extracted and analyzed. We found thatwild-type and mutant css1-2 cells contained about thenal extension of Css1p provides an essential function
unrelated to membrane targeting. same amount of radiolabeled MIPC per A600 unit of cells(data not shown). These data indicate that Css1p is notcss1 mutant cells maintained a normal rod shape de-
spite the accumulation of glucans. This is dissimilar to regulating sphingolipid synthesis or accumulation.We next determined if Css1p could hydrolyze IPC.the situation where loss of function or overproduction
of many genes involved in cell wall biosynthesis causes To assay for such activity, the css1� gene was overex-pressed in S. cerevisiae strain RCD318 using the galactose-changes in cell and cell wall morphology (reviewed in
Arellano et al. 1999). Thus, we wanted to determine inducible GAL1 promoter. The S. cerevisiae homolog ofcss1, ISC1, is deleted in strain RCD318 and the strainwhether excess glucans were being deposited in the
periplasmic space or in the cytoplasm. For this purpose, lacks the phospholipase C activity present in wild-typeS. cerevisiae cells that hydrolyzes inositolphosphosphin-we utilized the GFP-Css1p332-424 construct to visualize
the position of the plasma membrane relative to the golipids (Wells et al. 1998). For example, membranesprepared from strain RCD318 transformed with andeposits. When GFP-Css1p332-424 was expressed in css1-2
cells at the nonpermissive temperature, the plasma mem- empty vector (RCD318-GAL1) do not hydrolyze [3H]IPC-3whereas membranes prepared from wild-type JK9-3d S.brane was clearly no longer adjacent to the cell wall but
separated from it by the accumulated material (Figure cerevisiae cells do (Figure 7A). We found that mem-branes from the Css1p overexpressing strain (RCD-5I). This situation was also observed by electron micros-
copy of css1-2 mutant cells (Figure 6). The glucans accu- 318GAL1css1) hydrolyzed IPC-3 as well as wild-typeS. cerevisiae membranes (Figure 7A). These data stronglymulated between the cell wall and the plasma mem-
brane and did not affect the integrity or appearance suggest that Css1p is able to hydrolyze inositolphosphos-phingolipids.of the cell wall. These results are consistent with our
observation that css1-2 cells are no more resistant or The results shown in Figure 7A predict that mem-branes prepared from wild-type S. pombe cells shouldsensitive to cell wall degrading enzymes than wild-type
cells (data not shown). contain an activity that hydrolyzes IPC-3. This predictionwas verified using membranes prepared from cellsCss1p is a sphingolipase: Css1p shows greatest se-
quence similarity to mammalian neutral sphingomy- grown at 25� and from cells shifted to 36� for 2 hr priorto membrane isolation (Figure 7B). We expected thatelinases (Rodrigues-Lima et al. 2000). Fungi do not
contain sphingomyelin; rather they often contain sphin- membranes prepared from css1-2 cells grown at the re-strictive temperature (2 hr at 36�) should lack this en-golipids that have inositol phosphate linked to ceramide

1406 A. Feoktistova et al.
Figure 4.—Cloning of css1�. (A) Genomic organization of css1�,constructs tested for css1-3 rescue, and deletion strategy. The ORFis indicated by the solid box with the direction of transcriptionindicated by the arrow. X, XbaI; M, MluI; H, HindIII; A, Acc651. (B)Amino acid sequence alignment of Css1p with S. cerevisiae Isc1p.Identical amino acids are indicated by vertical lines. The glutamicacid and histidine residues predicted to be involved in substratebinding and catalysis, respectively, of this family of enzymes areindicated with an arrowhead and a box. The residues mutated inthe css1 mutant strains are indicated with asterisks. (C) A hydropathyplot of Css1p with the two potential transmembrane domains nearthe C terminus. (D) Css1p is a membrane-associated protein. css1myc(lanes 1, 3, 5, 7, 9, and 11) and wild-type (lanes 2, 4, 6, 8, 10, and12) cells were lysed with no detergent in TEG buffer (lanes 1, 2, 7,and 8), 1% NP-40 in NP-40 buffer (lanes 3, 4, 9, and 10), or in 1%SDS lysis buffer, and boiled immediately (lanes 5, 6, 11, and 12).The lysates were fractionated into soluble (lanes 1–6) and insoluble(lanes 7–12) fractions. Css1p-myc was detected by immunoblottingwith 9E10.
zyme activity and this is what we found (Figure 7B). ity in S. pombe cells behaves like a phospholipase typeC enzyme. Cleavage of MIPC by a phospholipase C-likeUnexpectedly, membranes prepared from css1-2 grown
at the permissive temperature (25�) also lacked enzyme enzyme should yield ceramide and MIP as the reactionproducts. For these experiments we used MIPC pre-activity (Figure 7B). Lack of activity in cells grown at
the permissive temperature probably reflects a variant pared from wild-type cells radiolabeled with [3H]inositolto label the polar head group or radiolabeled withprotein that is inactivated during membrane prepara-
tion or during the enzyme assay, which contains a low [3H]dihydrosphingosine to label ceramide. The radiola-beled product of the reactions was analyzed by paperconcentration of detergent. Whatever the reason, it is
clear from these data that css1-2 cells lack the IPC-3 chromatography using a solvent system in which MIPremained at the origin while ceramide moved with thehydrolase activity present in wild-type cells. These data
also establish that css1 encodes the only sphingolipid solvent front (Wells et al. 1998). Cleavage of [3H-inosi-tol]MIPC gave radiolabeled MIP as the product (Figurehydrolytic activity present in S. pombe cells under our
assay conditions. 7C) and cleavage of [3H-dihydrosphingosine]MIPC gaveradiolabeled ceramide as the product (Figure 7D).We next verified that the sphingolipid hydrolytic activ-

1407Essential S. pombe Sphingolipase
Figure 5.—Css1p is local-ized to membranes. css1myccells were grown at 32� andstained with (A) 9E10 and (B)DAPI. Wild-type cells express-ing (C) GFP-Css1p, (D) GFP-Css1�Cp, (E) GFP-Css1p332-424, (F) GFP-Css1p332-389, (G)GFP-Css1p332-363, and (H)GFP-Css1p1-389 were grown tomidlog phase in the absence ofthiamine for 18 hr. Images oflive cells were captured. (I)css1-2 cells expressing GFP-Css1p332-424 were grown tomidlog phase at 25� in the ab-sence of thiamine for 14 hr andshifted to 36� for 6 hr. Imagesof live cells were captured.
Thus, the sphingolipid hydrolase in wild-type S. pombe turnover and the activity of enzymes important for coor-dinating glucan synthesis with cell growth.cells behaves like a phospholipase C.
Using two different polysaccharide labeling and analy-sis strategies, the same conclusion was reached regard-ing increased levels of both �- and �-glucans, but notDISCUSSIONgalactomannans, in css1 mutant cells. This result is con-
In this study, we presented the characterization of sistent with the accumulated material staining with bothcss1 mutants that display a novel defect in cell wall me- calcofluor and aniline blue, dyes known to bind glucantabolism. css1 mutants arrest division quickly but con- polymers. Because there are so many genes encodingtinue to accumulate large amounts of both �- and �-glu- �- and �-glucan synthases in S. pombe cells, it was notcans in their periplasmic space. Because this material is possible to prevent glucan deposition by making appro-not incorporated into the cell wall, it does not affect priate mutant strains. For example, both css1mok1 andtheir rod shape but does cause compaction of the cell’s css1cps1 double mutant strains still accumulated largecytoplasm. The growth arrest and unusual glucan depo- amounts of calcofluor-staining material (data notsition observed in css1 mutants appear to be irreversible shown). This is not surprising given that in the absencelethal events. css1 mutants are defective in sphingophos- of a single glucan synthase gene, many others are pre-pholipid-phospholipase C activity and, thus, their phe- sumably still functional. Typically, there is strict coordi-
nation between the synthesis of cell wall componentsnotype points to a connection between sphingolipid
1408 A. Feoktistova et al.
the dramatic polymer accumulation, indicating that thesynchrony between �- and �-glucan synthases is not al-tered in css1 mutant cells. Since the other polysaccharidecomponent of the cell wall, galactomannan, did notaccumulate in the css1 mutant, there appears to be aspecific defect in the feedback control of �- and �-glucansynthases. Although their activity does not appear to beincreased in css1 mutants, as determined by measuringtotal �-glucan synthase activity in vitro (data not shown),it appears that the normal feedback control of theseenzymes is not operative in css1 mutants. It may bepossible to discover the biochemical nature of thesecontrol mechanisms by further analysis of css1-inter-acting genes.
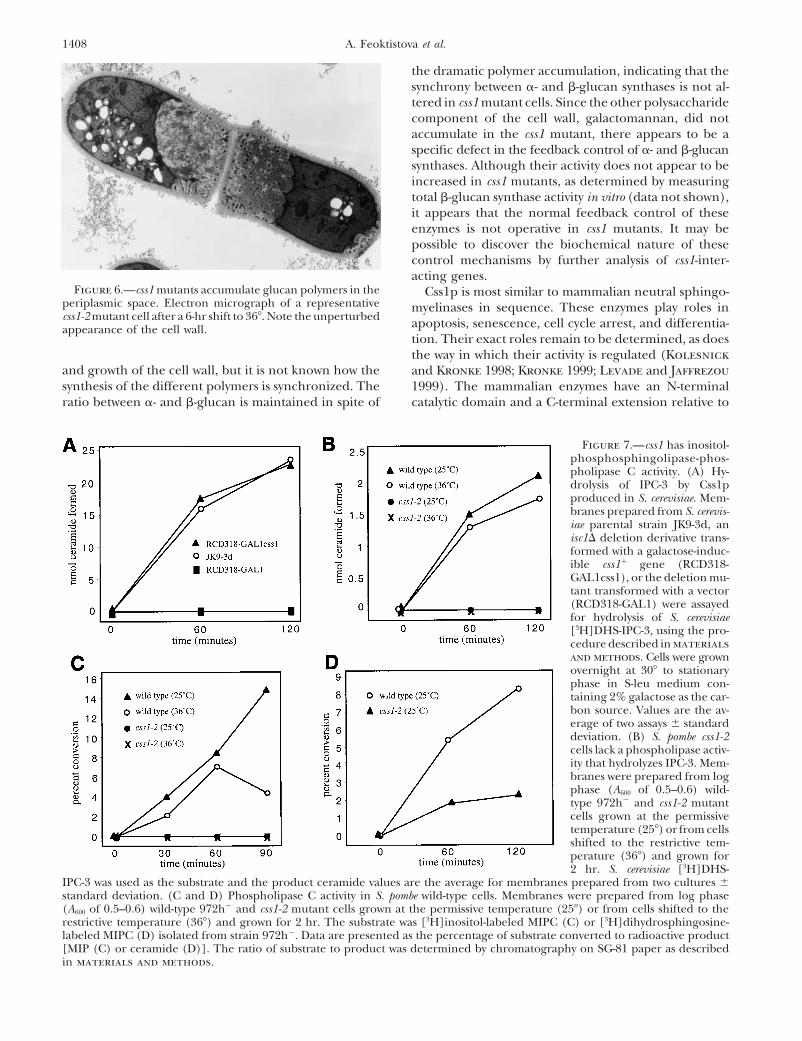
Figure 6.—css1 mutants accumulate glucan polymers in the Css1p is most similar to mammalian neutral sphingo-periplasmic space. Electron micrograph of a representative myelinases in sequence. These enzymes play roles incss1-2 mutant cell after a 6-hr shift to 36�. Note the unperturbed apoptosis, senescence, cell cycle arrest, and differentia-appearance of the cell wall.
tion. Their exact roles remain to be determined, as doesthe way in which their activity is regulated (Kolesnick
and growth of the cell wall, but it is not known how the and Kronke 1998; Kronke 1999; Levade and Jaffrezousynthesis of the different polymers is synchronized. The 1999). The mammalian enzymes have an N-terminal
catalytic domain and a C-terminal extension relative toratio between �- and �-glucan is maintained in spite of
Figure 7.—css1 has inositol-phosphosphingolipase-phos-pholipase C activity. (A) Hy-drolysis of IPC-3 by Css1pproduced in S. cerevisiae. Mem-branes prepared from S. cerevis-iae parental strain JK9-3d, anisc1� deletion derivative trans-formed with a galactose-induc-ible css1� gene (RCD318-GAL1css1), or the deletion mu-tant transformed with a vector(RCD318-GAL1) were assayedfor hydrolysis of S. cerevisiae[3H]DHS-IPC-3, using the pro-cedure described in materialsand methods. Cells were grownovernight at 30� to stationaryphase in S-leu medium con-taining 2% galactose as the car-bon source. Values are the av-erage of two assays � standarddeviation. (B) S. pombe css1-2cells lack a phospholipase activ-ity that hydrolyzes IPC-3. Mem-branes were prepared from logphase (A600 of 0.5–0.6) wild-type 972h� and css1-2 mutantcells grown at the permissivetemperature (25�) or from cellsshifted to the restrictive tem-perature (36�) and grown for2 hr. S. cerevisiae [3H]DHS-
IPC-3 was used as the substrate and the product ceramide values are the average for membranes prepared from two cultures �standard deviation. (C and D) Phospholipase C activity in S. pombe wild-type cells. Membranes were prepared from log phase(A600 of 0.5–0.6) wild-type 972h� and css1-2 mutant cells grown at the permissive temperature (25�) or from cells shifted to therestrictive temperature (36�) and grown for 2 hr. The substrate was [3H]inositol-labeled MIPC (C) or [3H]dihydrosphingosine-labeled MIPC (D) isolated from strain 972h�. Data are presented as the percentage of substrate converted to radioactive product[MIP (C) or ceramide (D)]. The ratio of substrate to product was determined by chromatography on SG-81 paper as describedin materials and methods.

1409Essential S. pombe Sphingolipase
through the secretory pathway (reviewed in Rapoportet al. 1996). Our results suggest that, once directed tothe ER by the transmembrane domain, other portionsof Css1p facilitate its transport through the secretorypathway since GFP fusions to the C-terminal domain ofCss1p were much more concentrated in juxta-nuclearregions than was full-length GFP-Css1p.
S. pombe cells make IPC and MIPC (Dickson andLester 1999) but the presence of other sphingolipids,such as sphingomyelin, has not been reported. Weshowed that Css1p function is clearly necessary for theyeast cells’ ability to hydrolyze IPC and MIPC and, atFigure 8.—Models of Css1p function. Three types of sub-least for MIPC, ceramide is generated in vitro (see Figurestrates that Css1p might act on along with the products of the
reactions are shown. Our data support model A, where the 8 for model of Css1p function). We also find that Css1psubstrates would be IPC and MIPC, but do not exclude models and its S. cerevisiae homolog, Isc1p, are responsible forB or C. Cellular processes or enzymes that could be targets this type of enzymatic activity in yeast membrane prepa-for the hydrolysis products are discussed in the text.
rations.The mammalian homolog, nSMaseI, is also able to
function as a major sphingolipase of membrane prepa-bacterial sphingomyelinases that contains two putativetransmembrane domains (Rodrigues-Lima et al. 2000). rations in vitro (Fensome et al. 2000; Tomiuk et al. 2000).
Unlike the situation in yeast membranes, however, thereCss1p is organized similarly. The mutations in the fourcss1 mutant alleles that we isolated map to sequences appear to be additional isozymes since immunodeple-
tion of nSMaseI does not remove all of the neutralencoding the putative catalytic domain. However, weshowed that the region C-terminal to the transmem- sphingomyelinase activity from such membrane prepa-
rations. Interestingly, nSMaseI produced in bacteria orbrane domains and the transmembrane domains arealso essential for its function. Why sequences C-terminal insect cells possesses sphingolipase activity comparable
to its activity in membrane preparations, arguing thatto the transmembrane domains are essential is notknown. We showed that they are not required for proper this class of enzyme does not require other subunits for
its activity (Fensome et al. 2000). While S. cerevisiae ap-Css1p localization; they might influence the activity ofthe catalytic domain. Given that Css1p behaves as an pear not to require sphingolipid hydrolysis since they
tolerate deletion of the css1 homolog ISC1, S. pombeintegral membrane protein and is localized to vesiclesand the plasma membrane, it is not surprising that the cells cannot survive without css1. We did not detect a
significant increase in the amount of S. pombe sphingoli-transmembrane domains are essential for its function.Most likely Css1p must be anchored in membranes to pids in css1 mutant cells (data not shown) and it may
be that there is reduced synthesis of sphingolipids infunction correctly. The closest known relative of Css1pamong the mammalian neutral sphingomyelinases, response to a drop in their hydrolysis.
Since there is no detectable buildup of sphingolipids,nSMase1, is thought to be localized predominantly toendoplasmic reticulum (ER) membranes (Rodrigues- the phenotype caused by loss of css1 function seems
more likely due to a decrease in a product of sphingoli-Lima et al. 2000; Tomiuk et al. 2000) although sphingo-myelinase activity is also abundant in brain plasma mem- pid hydrolysis, i.e., ceramide and MIP or IP (Figure 8,
model A). The role of ceramide in mammalian cells isbrane preparations (Tomiuk et al. 2000). Thus, whetherCss1p plays a role in the ER and secretory apparatus as of great interest because its level increases in response
to extracellular ligands and stresses, and it is thoughtwell as at the plasma membrane remains to be deter-mined. to act as a second messenger (reviewed in Dickson
1998). In S. cerevisiae, there is limited evidence thatAn unusual feature of Css1p is its membrane-targetingdomain. Although predicted to contain two membrane- sphingolipids or their metabolites serve as second mes-
sengers except perhaps during stress responses (reviewedspanning domains likely to form a hairpin loop throughthe membrane, the first membrane-spanning domain in Dickson and Lester 1999). The data presented here
suggest that Css1p hydrolyzes sphingolipids to generateof Css1p was sufficient to direct GFP to S. pombe mem-branes. Interestingly, a GFP fusion to the first transmem- ceramide, which acts as a signal to regulate cell wall
formation (Figure 8, model A).brane domain of mammalian nSNase1 also resulted inthe targeting of GFP to the ER of mammalian cells Besides signaling, other functions for ceramides have
been identified in S. cerevisiae. There is compelling evi-(Rodrigues-Lima et al. 2000). While unusual, such amembrane insertion strategy is not unique. Several pro- dence that ceramide enhances the transport of glycosyl-
phosphatidylinositol (GPI)-anchored proteins throughteins, including synaptobrevin, are known to insertthemselves into ER membranes through C-terminal the secretory apparatus (Horvath et al. 1994; Skrzypek
et al. 1997) and can be the lipid moiety of lipid-linkedmembrane-spanning domains and to be transported

1410 A. Feoktistova et al.
Fensome, A. C., F. Rodrigues-Lima, M. Josephs, H. F. Paterson andproteins that are major constituents of the cell wallM. Katan, 2000 A neutral magnesium-dependent sphingomye-
(Conzelmann et al. 1992; reviewed in Dickson 1998). It linase isoform associated with intracellular membranes and re-versibly inhibited by reactive oxygen species. J. Biol. Chem. 275:is possible, therefore, that loss of Css1p function causes a1128–1136.reduction in an unknown ceramide-anchored cell wall
Forsburg, S. L., 1993 Comparison of Schizosaccharomyces pombeprotein(s) whose function is to coordinate glucan syn- expression systems. Nucleic Acids Res. 21: 2955–2956.
Gould, K. L., S. Moreno, D. J. Owen, S. Sazer and P. Nurse, 1991thesis and incorporation into the wall with cell growthPhosphorylation at Thr167 is required for Schizosaccharomyces(Figure 8, model B). Finally, our data do not exclude thepombe p34cdc2 function. EMBO J. 10: 3297–3309.
possibility that Css1p acts on other lipid phosphodiesters Guldener, U., S. Heck, T. Fielder, J. Beinhauer and J. H. Hege-mann, 1996 A new efficient gene disruption cassette for re-lacking ceramide (Figure 8, model C). Because S. pombepeated use in budding yeast. Nucleic Acids Res. 24: 2519–2524.Css1p has an essential function, further analysis of css1-
Heitman, J., N. R. Movva, P. C. Hiestand and M. N. Hall, 1991interacting genes will provide a unique opportunity to FK 506-binding protein proline rotamase is a target for the immu-
nosuppressive agent FK 506 in Saccharomyces cerevisiae. Proc. Natl.probe the role of ceramides and other sphingolipidAcad. Sci. USA 88: 1948–1952.metabolites in yeast cell function.
Hochstenbach, F., F. M. Klis, H. van den Ende, E. van Donselaar,P. J. Peters et al., 1998 Identification of a putative alpha-glucanWe thank Dan McCollum and Mohan Balasubramanian for css1synthase essential for cell wall construction and morphogenesismutant isolation. This work was supported by National Institutes ofin fission yeast. Proc. Natl. Acad. Sci. USA 95: 9161–9166.Health (NIH) grant GM 59773 to C.A., NIH grant GM41302 to R.C.D,
Hoffman, C. S., 1993 Preparation of Yeast DNA, pp. 13.11.11–grant IFD97-1570-C02-01 from the Comision Interministerial de Cien-13.11.14 in Current Protocols in Molecular Biology, edited by F. A.cia y Technologia, Spain to P.P., and by the Howard Hughes MedicalAusubel, R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman
Institute of which K.L.G. is an associate investigator. et al. John Wiley & Sons, New York.Horvath, A., C. Sutterlin, U. Manning-Krieg, N. R. Movva and
H. Riezman, 1994 Ceramide synthesis enhances transport ofGPI-anchored proteins to the Golgi apparatus in yeast. EMBO J.LITERATURE CITED13: 3687–3695.
Ishiguro, J., A. Saitou, A. Duran and J. C. Ribas, 1997 cps1�, aArellano, M., A. Duran and P. Perez, 1997 Localisation of theSchizosaccharomyces pombe rho1p GTPase and its involvement in the Schizosaccharomyces pombe gene homolog of Saccharomyces cerevisiae
FKS genes whose mutation confers hypersensitivity to cyclosporinorganisation of the actin cytoskeleton. J. Cell Sci. 110: 2547–2555.Arellano, M., P. M. Coll and P. Perez, 1999 RHO GTPases in the A and papulacandin B. J. Bacteriol. 179: 7653–7662.
Katayama, S., D. Hirata, M. Arellano, P. Perez and T. Toda,control of cell morphology, cell polarity, and actin localizationin fission yeast. Microsc. Res. Tech. 47: 51–60. 1999 Fission yeast �-glucan synthase Mok1 requires the actin
cytoskeleton to localize the sites of growth and plays an essentialBahler, J., J. Q. Wu, M. S. Longtine, N. G. Shah, A. McKenzie, 3rdet al., 1998 Heterologous modules for efficient and versatile role in cell morphogenesis downstream of protein kinase C func-
tion. J. Cell Biol. 144: 1173–1186.PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14:943–951. Keeney, J. B., and J. D. Boeke, 1994 Efficient targeted integration
at leu1-32 and ura4-294 in Schizosaccharomyces pombe. Genetics 136:Balasubramanian, M. K., D. McCollum and K. L. Gould, 1997Cytokinesis in fission yeast Schizosaccharomyces pombe. Methods En- 849–856.
Kippert, F., and D. Lloyd, 1995 The aniline blue fluorochromezymol. 283: 494–506.Balasubramanian, M. K., D. McCollum, L. Chang, K. C. Wong, specifically stains the septum of both live and fixed Schizosaccharo-
myces pombe cells. FEMS Microbiol. Lett. 132: 215–219.N. I. Naqvi et al., 1998 Isolation and characterization of newfission yeast cytokinesis mutants. Genetics 149: 1265–1275. Kolesnick, R. N., and M. Kronke 1998 Regulation of ceramide
production and apoptosis. Annu. Rev. Physiol. 60: 643–665.Barbet, N., W. J. Muriel and A. M. Carr, 1992 Versatile shuttlevectors and genomic libraries for use with Schizosaccharomyces Kopecka, M., G. H. Fleet and H. J. Phaff, 1995 Ultrastructure of
the cell wall of Schizosaccharomyces pombe following treatment withpombe. Gene 114: 59–66.Basi, G., E. Schmid and K. Maundrell, 1993 TATA box mutations various glucanases. J. Struct. Biol. 114: 140–152.
Kronke, M., 1999 Involvement of sphingomyelinases in TNF signal-in the Schizosaccharomyces pombe nmt1 promoter affect transcrip-tion efficiency but not the transcription start point or thiamine ing pathways. Chem. Phys. Lipids 102: 157–166.
Le Goff, X., A. Woollard and V. Simanis, 1999 Analysis of therepressibility. Gene 123: 131–136.Bush, D. A., M. Horisberger, I. Horman and P. Wursch, 1974 The cps1 gene provides evidence for a septation checkpoint in Schizo-
saccharomyces pombe. Mol. Gen. Genet. 262: 163–172.wall structure of Schizosaccharomyces pombe. J. Gen. Microbiol. 81:199–206. Leatherwood, J., A. Lopez-Girona and P. Russell, 1996 Interac-
tion of Cdc2 and Cdc18 with a fission yeast ORC2-like protein.Cabib, E., T. Drgon, J. Drgonova, R. A. Ford and R. Kollar, 1997The yeast cell wall, a dynamic structure engaged in growth and Nature 379: 360–363.
Lester, R. L., and R. C. Dickson, 1993 Sphingolipids with inositol-morphogenesis. Biochem. Soc. Trans. 25: 200–204.Chappell, T. G., and G. Warren, 1989 A galactosyltransferase from phosphate-containing head groups. Adv. Lipid Res. 26: 253–274.
Levade, T., and J. P. Jaffrezou, 1999 Signalling sphingomyelinases:the fission yeast Schizosaccharomyces pombe. J. Cell Biol. 109: 2693–2702. which, where, how and why? Biochim. Biophys. Acta 1438: 1–17.
Liu, J., H. Wang, D. McCollum and M. K. Balasubramanian, 1999Chappell, T. G., M. A. Hajibagheri, K. Ayscough, M. Pierce and G.Warren, 1994 Localization of an alpha 1,2 galactosyltransferase Drc1p/Cps1p, a 1,3-beta-glucan synthase subunit, is essential for
division septum assembly in Schizosaccharomyces pombe. Geneticsactivity to the Golgi apparatus of Schizosaccharomyces pombe. Mol.Biol. Cell 5: 519–528. 153: 1193–1203.
Liu, J., X. Tang, H. Wang and M. Balasubramanian, 2000 Bgs2p,Clarke, N. G., and R. M. Dawson, 1981 Alkaline O leads to N-trans-acylation. A new method for the quantitative deacylation of phos- a 1,3-beta-glucan synthase subunit, is essential for maturation
of ascospore wall in Schizosaccharomyces pombe. FEBS Lett. 478:pholipids. Biochem. J. 195: 301–306.Conzelmann, A., A. Puoti, R. L. Lester and C. Desponds, 1992 105–108.
Manners, D. J., and M. T. Meyer, 1977 The molecular structuresTwo different types of lipid moieties are present in glycophospho-inositol-anchored membrane proteins of Saccharomyces cerevisiae. of some glucans from the cell walls of Schizosaccharomyces pombe.
Carbohydr. Res. 57: 189–203.EMBO J. 11: 457–466.Dickson, R. C., 1998 Sphingolipid functions in Saccharomyces cerevis- Marks, J., I. M. Hagan and J. S. Hyams, 1987 Spacial association
of F-actin with growth polarity and septation in the fission yeastiae : comparison to mammals. Annu. Rev. Biochem. 67: 27–48.Dickson, R. C., and R. L. Lester, 1999 Yeast sphingolipids. Biochim. Schizosaccharomyces pombe. Spec. Publ. Soc. Gen. Microbiol. 23:
119–135.Biophys. Acta 1426: 347–357.

1411Essential S. pombe Sphingolipase
Martin, V., J. C. Ribas, E. Carnero, A. Duran and Y. Sanchez, 2000 Identification of ISC1 (YER019w) as inositol phospho-sphingolipid phospholipase C in Saccharomyces cerevisiae. J. Biol.2000 bgs2�, a sporulation-specific glucan synthase homologueChem. 275: 39793–39798.is required for proper ascospore wall maturation in fission yeast.
Sherman, F., 1991 Getting started with yeast. Methods Enzymol.Mol. Microbiol. 38: 308–321.194: 3–21.Maundrell, K., 1993 Thiamine-repressible expression vectors pREP
Skrzypek, M., R. L. Lester and R. C. Dickson, 1997 Suppressorand pRIP for fission yeast. Gene 123: 127–130.gene analysis reveals an essential role for sphingolipids in trans-Mitchison, J. M., and P. Nurse, 1985 Growth in cell length in theport of glycosylphosphatidylinositol-anchored proteins in Saccha-fission yeast Schizosaccharomyces pombe. J. Cell Sci. 75: 357–376.romyces cerevisiae. J. Bacteriol. 179: 1513–1520.Moreno, S., A. Klar and P. Nurse, 1991 Molecular genetic analysis
Skrzypek, M. S., M. M. Nagiec, R. L. Lester and R. C. Dickson,of fission yeast Schizosaccharomyces pombe. Methods Enzymol. 194:1998 Inhibition of amino acid transport by sphingoid long795–823.chain bases in Saccharomyces cerevisiae. J. Biol. Chem. 273: 2829–Mumberg, D., R. Muller and M. Funk, 1994 Regulatable promoters2834.of Saccharomyces cerevisiae: comparison of transcriptional activity
Smith, M. M., and M. E. McCully, 1978 A critical evaluation ofand their use for heterologous expression. Nucleic Acids Res.the specificity of aniline blue induced fluorescence. Protoplasma22: 5767–5768.95: 229–254.Nurse, P., 1994 Fission yeast morphogenesis—posing the problems.
Steiner, S., and R. L. Lester, 1972 Studies on the diversity ofMol. Biol. Cell 5: 613–616.inositol-containing yeast phospholipids: incorporation of 2-deoxy-Osumi, M., M. Sato, S. A. Ishijima, M. Konomi, T. Takagi et al., 1998glucose into lipid. J. Bacteriol. 109: 81–88.Dynamics of cell wall formation in fission yeast, Schizosaccharomyces
Tomiuk, S., K. Hofmann, M. Nix, M. Zumbansen and W. Stoffel,pombe. Fungal Genet. Biol. 24: 178–206.1998 Cloned mammalian neutral sphingomyelinase: functionsPrentice, H. L., 1992 High efficiency transformation of Schizosac-in sphingolipid signaling? Proc. Natl. Acad. Sci. USA 95: 3638–charomyces pombe by electroporation. Nucleic Acids Res. 20: 621. 3643.Rapoport, T. A., B. Jungnickel and U. Kutay, 1996 Protein trans- Tomiuk, S., M. Zumbansen and W. Stoffel, 2000 Characterizationport across the eukaryotic endoplasmic reticulum and bacterial and subcellular localization of murine and human magnesium-
inner membranes. Annu. Rev. Biochem. 65: 271–303. dependent neutral sphingomyelinase. J. Biol. Chem. 275: 5710–Rodrigues-Lima, F., A. C. Fensome, M. Josephs, J. Evans, R. J. Veld- 5717.
man et al., 2000 Structural requirements for catalysis and mem- Wells, G. B., R. C. Dickson and R. L. Lester, 1998 Heat-inducedbrane targeting of mammalian enzymes with neutral sphingomye- elevation of ceramide in Saccharomyces cerevisiae via de novo synthe-linase and lysophospholipid phospholipase C activities. Analysis sis. J. Biol. Chem. 273: 7235–7243.by chemical modification and site-directed mutagenesis. J. Biol. Yoko-o, T., S. K. Roy and Y. Jigami, 1998 Differences in in vivoChem. 275: 28316–28325. acceptor specificity of two galactosyltransferases, the gmh3� and
Sambrook, J., E. F. Fritsch and T. Maniatis, 1989 Molecular Clon- gma12� gene products from Schizosaccharomyces pombe. Eur. J.ing: A Laboratory Manual, Ed. 2. Cold Spring Harbor Laboratory Biochem. 257: 630–637.Press, Cold Spring Harbor, NY.
Sawai, H., Y. Okamoto, C. Luberto, C. Mao, A. Bielawska et al., Communicating editor: P. Russell




















