
Condition factor and diet of the catfish Loricariichthys castaneus (Castelnau, 1855): gender...
10
This article was downloaded by: [CHRISTINA BRANCO] On: 04 March 2014, At: 15:54 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Studies on Neotropical Fauna and Environment Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/nnfe20 Condition factor and diet of the catfish Loricariichthys castaneus (Castelnau, 1855): gender differences in three tropical reservoirs Gustavo Luiz Ferreira a , Vanessa Guimarães Lopes b , José Henrique Cantarino Gomes c & Christina Wyss Castelo Branco a a Departamento de Zoologia, Instituto de Biociências, Universidade Federal do Estado do Rio de Janeiro, Rio de Janeiro, Brazil b Laboratório de Ecologia Teórica e Síntese, Instituto de Ciências Biológicas, Universidade Federal de Goiás, Brazil c Transpetro-Petrobras Transporte S.A., DGN/SMSOP; Rio de Janeiro, Brazil Published online: 27 Feb 2013. To cite this article: Gustavo Luiz Ferreira, Vanessa Guimarães Lopes, José Henrique Cantarino Gomes & Christina Wyss Castelo Branco (2013) Condition factor and diet of the catfish Loricariichthys castaneus (Castelnau, 1855): gender differences in three tropical reservoirs, Studies on Neotropical Fauna and Environment, 48:3, 190-198, DOI: 10.1080/01650521.2014.890849 To link to this article: http://dx.doi.org/10.1080/01650521.2014.890849 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Condition factor and diet of the catfish Loricariichthys castaneus (Castelnau, 1855): gender...

This article was downloaded by: [CHRISTINA BRANCO]On: 04 March 2014, At: 15:54Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Studies on Neotropical Fauna and EnvironmentPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/nnfe20
Condition factor and diet of the catfish Loricariichthyscastaneus (Castelnau, 1855): gender differences inthree tropical reservoirsGustavo Luiz Ferreiraa, Vanessa Guimarães Lopesb, José Henrique Cantarino Gomesc &Christina Wyss Castelo Brancoa
a Departamento de Zoologia, Instituto de Biociências, Universidade Federal do Estado doRio de Janeiro, Rio de Janeiro, Brazilb Laboratório de Ecologia Teórica e Síntese, Instituto de Ciências Biológicas, UniversidadeFederal de Goiás, Brazilc Transpetro-Petrobras Transporte S.A., DGN/SMSOP; Rio de Janeiro, BrazilPublished online: 27 Feb 2013.
To cite this article: Gustavo Luiz Ferreira, Vanessa Guimarães Lopes, José Henrique Cantarino Gomes & ChristinaWyss Castelo Branco (2013) Condition factor and diet of the catfish Loricariichthys castaneus (Castelnau, 1855):gender differences in three tropical reservoirs, Studies on Neotropical Fauna and Environment, 48:3, 190-198, DOI:10.1080/01650521.2014.890849
To link to this article: http://dx.doi.org/10.1080/01650521.2014.890849
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

ORIGINAL ARTICLE
Condition factor and diet of the catfish Loricariichthys castaneus (Castelnau, 1855): genderdifferences in three tropical reservoirs
Gustavo Luiz Ferreiraa, Vanessa Guimarães Lopesb, José Henrique Cantarino Gomesc &Christina Wyss Castelo Brancoa*aDepartamento de Zoologia, Instituto de Biociências, Universidade Federal do Estado do Rio de Janeiro, Rio de Janeiro, Brazil;bLaboratório de Ecologia Teórica e Síntese, Instituto de Ciências Biológicas, Universidade Federal de Goiás, Brazil; cTranspetro-Petrobras Transporte S.A., DGN/SMSOP; Rio de Janeiro, Brazil
(Received 22 August 2011; accepted 31 January 2014)
Condition factor and diet of male and female of Loricariichthys castaneus were studied in three tropical reservoirsin which it is the most abundant species of fish. The females had higher condition factors than males in all threeenvironments, but their diet differed from males only in the best-preserved ecosystem (Lajes Reservoir).Loricariichthys castaneus was considered as detritivorous-omnivorous. Starvation during breeding possibly con-tributes to a decrease of condition factor in males. By studying the gut content, sex-specific foraging and theinfluence of the diet importance in relation to the available food items in the three reservoirs were analyzed.
O fator de condição e a dieta de machos e fêmeas de Loricariichthys castaneus foram estudados em trêsreservatórios tropicais onde esta espécie é considerada a mais abundante. As fêmeas apresentaram maior fatorde condição nos três ambientes, mas tiveram dieta diferente da dos machos apenas no ecossistema maispreservado (Reservatório de Lajes). L. castaneus foi considerado detritívoro-onívoro. Escassez de alimentaçãoem machos durante a incubação dos ovos possivelmente contribui para o decréscimo do fator de condição.Através do estudo do conteúdo do trato digestório foram analisados tanto a alimentação específica de machos efêmeas como a influência da importância na dieta de itens alimentares disponíveis nos três reservatórios.
Keywords: Loricariichthys castaneus; sexual dimorphism; natural diet; habitat differences; detritivory; Brazil
Introduction
Brazil has one of the most developed systems ofhydroelectric plants, which are responsible forapproximately 76% of the electrical power distributedto more than 150 million people (Branco et al. 2009).Brazilian reservoirs are used for a wide variety ofpurposes, including the production of electric power,drinking water, recreation, fishing and agriculture,and all Brazilian river basins of major and mediumsize contain dams. The dams deeply influence thecomposition and structure of aquatic communities,creating great challenges, especially for fish popula-tions (Mérona et al. 2001; Lazzaro et al. 2003;Loureiro-Crippa & Hahn 2006; Delariva et al. 2007;Gubiani et al. 2007; Petesse et al. 2007; Luz-Agostinho et al. 2009).
The natural distribution of Loricariidae (armoredcatfish) encompasses the Neotropical region, reachingPanama and Costa Rica (Salvador-Jr et al. 2010).This family includes fish that live near the bottom,possessing sucker-shaped mouths. The speciesLoricariichthys castaneus (Castelnau 1855) is one ofthe most abundant species of the Loricariids in
freshwater environments of southeastern Brazil(Duarte et al. 2007). It is characterized by a sexualdimorphism associated with the male brood care.Males possess an elongated and flattened papilla ontheir lips to which the eggs adhere after fecundationand until the hatching of larvae (Taylor 1983; Reis &Pereira 2000). Recently, Gomes et al. (2011) foundlarvae-bearing males of L. castaneus in LajesReservoir, the first record of larvae transportationfor this species. The external morphological charac-teristics suggest a detritivorous diet but point to thepossibility of different types of feeding in males andfemales.
Despite L. castaneus being ubiquitous in theParaíba do Sul river system (Araújo et al. 2001),being constantly observed in the Lajes Reservoir(Araújo et al. 1998; Duarte & Araújo 2001; Gomeset al. 2011) and widely regarded as a detritivorousorganism (Kasper et al. 2009), there are no studiesdetailing the diet of this species. It is regarded domi-nant in the fish community of our study area, in thetropical reservoirs of Lajes, Santana and Vigário(Gomes et al. 2008). In Lajes predominance of
*Corresponding author. Email: [email protected]
Studies on Neotropical Fauna and Environment, 2013Vol. 48, No. 3, 190–198, http://dx.doi.org/10.1080/01650521.2014.890849
© 2014 Taylor & Francis
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014

females was observed by Araújo et al. (1998) andDuarte and Araújo (2001).
Due to the reported dominance of females and tothe modification of the male mouth adapted to thecare of eggs, we wanted to clarify sexual differences incondition factor and diet within a same reservoir.Condition indices are frequently used as indicatorsof the general well being of fish populations (Joneset al. 1999) and they are the result of several factorsthat include environmental features of the aquaticecosystem and aspects peculiar to the species, e.g.feeding and reproductive strategies (Sunardi et al.2007; Abujanra et al. 2009).
As the three reservoirs differ in limnological char-acteristics and levels of human impact, we assumedthere to be variable environmental conditions, in par-ticular in the available food, resulting in differences incondition factors. In order to clarify this, we analyzedthe gut content of the captured fishes.
Materials and methods
Study area
The three reservoirs are located within coordinates22°32′ S, 22°48′ S and 43°49′ W, 44°02′ W, approxi-mately 80 km from the city of Rio de Janeiro. Theyare used for the production of power, recreation andsport fishing, and also supply water for approximatelynine million people (Gomes et al. 2008).
The reservoir of Ribeirão das Lajes (LajesReservoir) is regarded as oligo-mesotrophic (Brancoet al. 2009) and is surrounded by remnants of theAtlantic Rainforest. The reservoirs of Vigário andSantana are considered eutrophic (Gomes et al.2008) having a high degree of human influence sincethey are surrounded by pasture and villages, respec-tively. Both present a massive colonization by aquaticplants, which are restrained by artificial barriers andperiodically removed by mechanical means.
Fish sampling
Fishes were caught in Lajes, Vigário and Santanareservoirs during the dry and wet seasons in theyears 2004 and 2005. The same standardized fishingeffort was applied in each reservoir, using 16 gill-nets3.5–53 m long, with heights between 1.15 and 3.8 mand with 1.5–5.5 cm knot-to-knot mesh-sizes. Gillnets were set towards dusk (proceedings commencedabout 16 h), and remained set for a period of 12hours, checked every four hours and being pulledfrom the water at dawn.
Voucher specimens were deposited in theCollection of the Museu Nacional (UFRJ), underthe number MNRJREG20050418.
The specimens were weighed (TW) and measuredfor evaluation of standard body length (SL) and totallength (TL). The identification of gender was ascer-tained through outer and inner morphological analy-sis. The digestive tract was measured (DL). Theintestinal quotient (IQ) was determined as the ratiobetween the length of the intestine and the total lengthof the individual (Barbieri et al. 1994).
The body condition was evaluated by the allo-metric condition factor (Ka), with the equation Ka= TW/SLb ×100 (Bagenal & Tesh 1978), where b wasobtained by the weight–length relationship lineariza-tion using the logarithmic transformation. Weight–length relationship was estimated according to theexpression TW = aSLb, where a is the intercept, andb the regression coefficient. The parameters a and bwere estimated after mass and length were log-trans-formed. Differences between biometric features ofgenders within reservoirs were estimated with thenon-parametric analysis of Kruskal–Wallis and theDunn test.
Fish diet analysis
The content of the first third of the digestive tract wasseparated and the content identified to the lowesttaxonomic level possible. The qualitative and quanti-tative analyses of the feeding items were performedwith a stereomicroscope and optic microscope withthe use of Sedgewick-Rafter chambers according tothe methodology in Branco et al. (1997). From theestimate of the frequency of occurrence (FO), relativeabundance (RA) and percentage by volume (%V) ofeach item, the Index of Relative Importance (IRI)(Pinkas et a1. 1971; Hyslop 1980) was applied, ascalculated by the formula: IRI = (RA + %V) × FO.
Analysis of similarity (ANOSIM, Clarke 1993)was applied to Bray–Curtis similarities using quanti-tative data (volume of food items) to determine ifgender led to changes in diet within each reservoirand also was performed within gender to determineif the reservoir led to changes in diet. ANOSIMemploys the R statistic to assess the existence of sig-nificant differences between groups (e.g. males andfemales) for a given factor; R ranges between –1 and+1, and 0 indicates a completely random grouping.The SIMPER (similarity percentage analysis) proce-dure was used to examine the contribution of eachfood item to the average dissimilarity between gendersin each reservoir and among the three reservoirs foreach gender. All analyses were done using the R
Studies on Neotropical Fauna and Environment 191
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014
program (R Development Core Team 2010) and theVegan package (Oksanen et al. 2009).
Results
Morphological characters
A total of 325 adult fishes were measured (136 fromLajes Reservoir, 132 from Vigário and 57 fromSantana). SL, DL, TW and Ka, but not IQ(Table 1) were significantly different between malesand females of the same reservoir (Kruskal–Wallis,p < 0.05). In all three reservoirs the females showed,on average, highest values of TW, SL, Ka, DL andIQ. Again, except for IQ, all morphological featureswere significantly different among the reservoirs. Thelowest values were found in the Vigário Reservoir andthe highest in Lajes (Dunn test, p < 0.05).
Fish diet
The analysis of the gut content of 128 individuals withfilled stomachs (52 from Lajes, 51 from Vigário and25 from Santana) resulted in the identification of 55items (Table 2). More females than males with filledstomachs were found in all three reservoirs (73% inLajes, 55% in Vigário and 64% in Santana). Datawere combined into nine large groups: algae, rotifers,microcrustaceans, testate amoebae, insects, otherinvertebrates, plant debris, sand and detritus.Detritus, sand, insects, plant debris, microcrustaceansand algae were found in the diet of fish from all threeenvironments (Table 3).
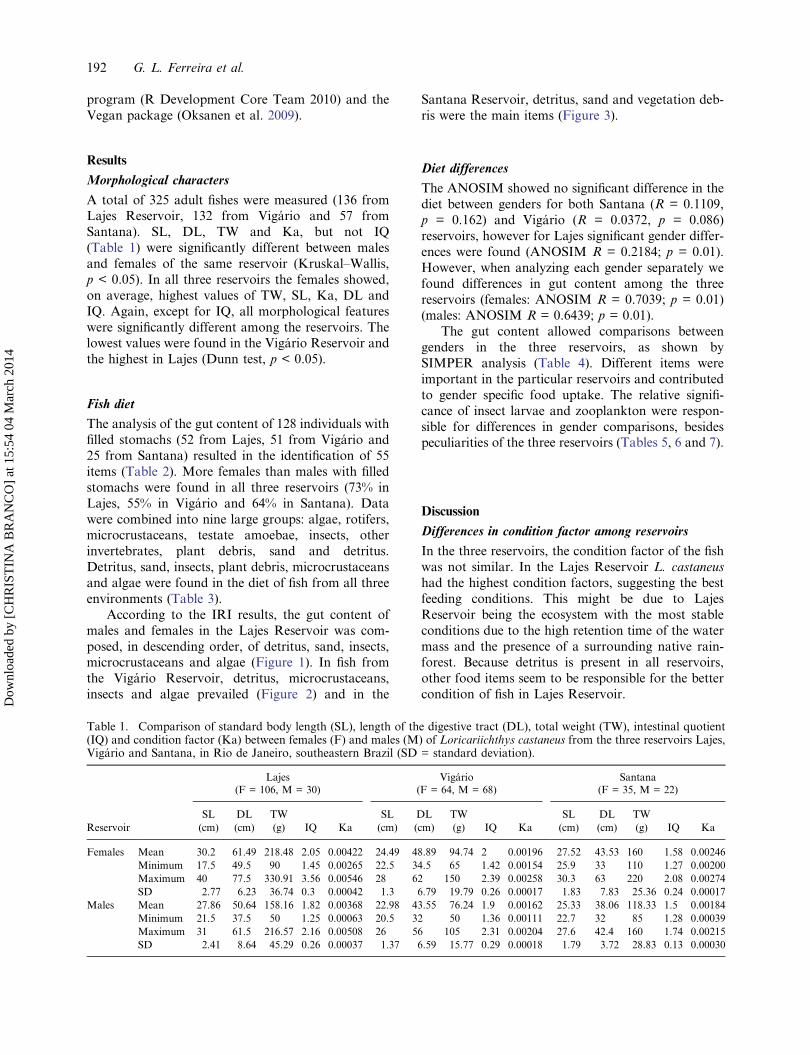
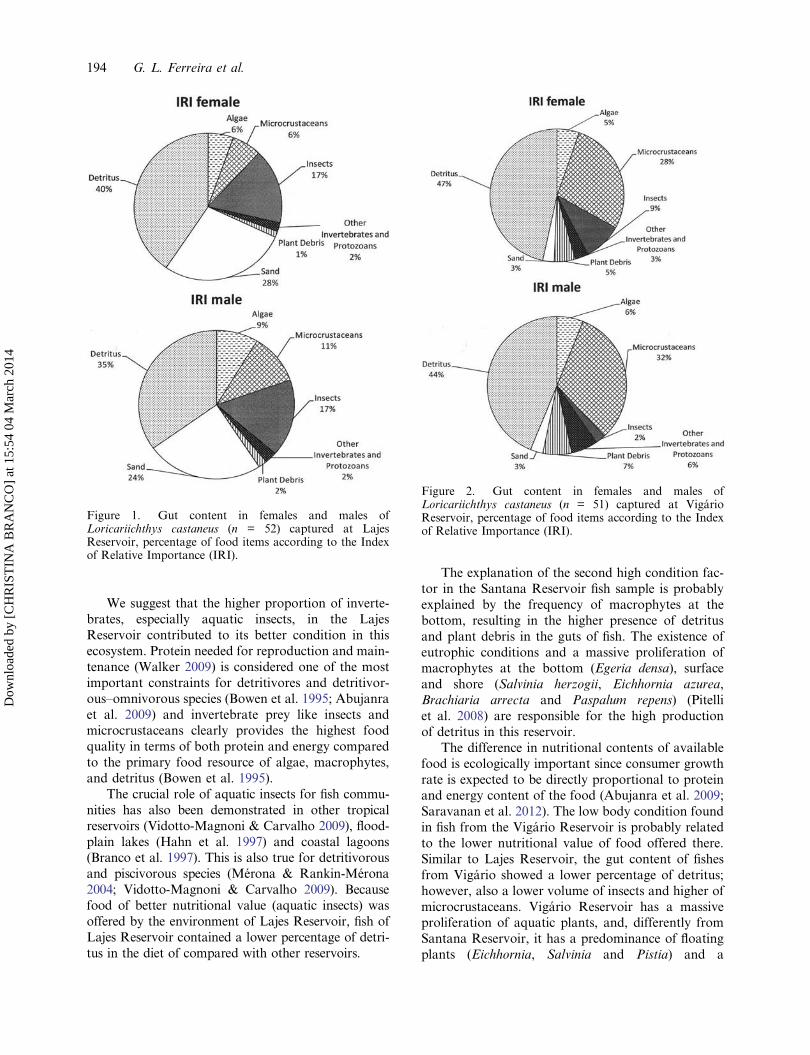
According to the IRI results, the gut content ofmales and females in the Lajes Reservoir was com-posed, in descending order, of detritus, sand, insects,microcrustaceans and algae (Figure 1). In fish fromthe Vigário Reservoir, detritus, microcrustaceans,insects and algae prevailed (Figure 2) and in the
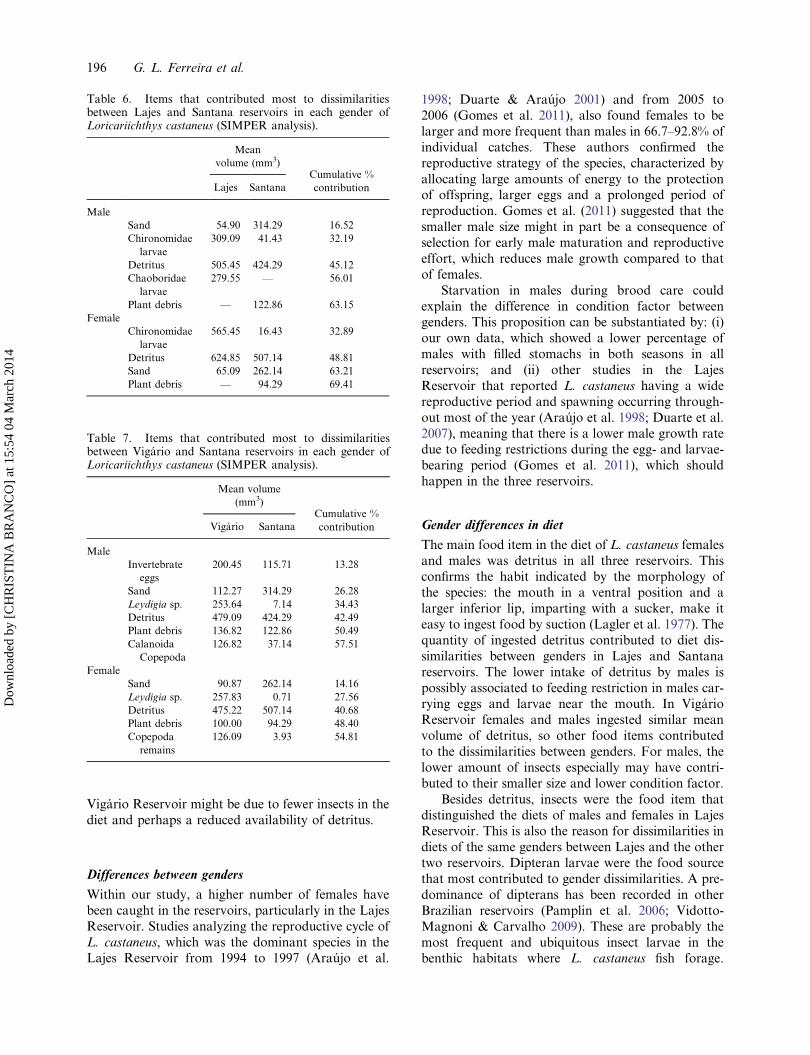
Santana Reservoir, detritus, sand and vegetation deb-ris were the main items (Figure 3).
Diet differences
The ANOSIM showed no significant difference in thediet between genders for both Santana (R = 0.1109,p = 0.162) and Vigário (R = 0.0372, p = 0.086)reservoirs, however for Lajes significant gender differ-ences were found (ANOSIM R = 0.2184; p = 0.01).However, when analyzing each gender separately wefound differences in gut content among the threereservoirs (females: ANOSIM R = 0.7039; p = 0.01)(males: ANOSIM R = 0.6439; p = 0.01).
The gut content allowed comparisons betweengenders in the three reservoirs, as shown bySIMPER analysis (Table 4). Different items wereimportant in the particular reservoirs and contributedto gender specific food uptake. The relative signifi-cance of insect larvae and zooplankton were respon-sible for differences in gender comparisons, besidespeculiarities of the three reservoirs (Tables 5, 6 and 7).
Discussion
Differences in condition factor among reservoirs
In the three reservoirs, the condition factor of the fishwas not similar. In the Lajes Reservoir L. castaneushad the highest condition factors, suggesting the bestfeeding conditions. This might be due to LajesReservoir being the ecosystem with the most stableconditions due to the high retention time of the watermass and the presence of a surrounding native rain-forest. Because detritus is present in all reservoirs,other food items seem to be responsible for the bettercondition of fish in Lajes Reservoir.
Table 1. Comparison of standard body length (SL), length of the digestive tract (DL), total weight (TW), intestinal quotient(IQ) and condition factor (Ka) between females (F) and males (M) of Loricariichthys castaneus from the three reservoirs Lajes,Vigário and Santana, in Rio de Janeiro, southeastern Brazil (SD = standard deviation).
Lajes Vigário Santana(F = 106, M = 30) (F = 64, M = 68) (F = 35, M = 22)
ReservoirSL(cm)
DL(cm)
TW(g) IQ Ka
SL(cm)
DL(cm)
TW(g) IQ Ka
SL(cm)
DL(cm)
TW(g) IQ Ka
Females Mean 30.2 61.49 218.48 2.05 0.00422 24.49 48.89 94.74 2 0.00196 27.52 43.53 160 1.58 0.00246Minimum 17.5 49.5 90 1.45 0.00265 22.5 34.5 65 1.42 0.00154 25.9 33 110 1.27 0.00200Maximum 40 77.5 330.91 3.56 0.00546 28 62 150 2.39 0.00258 30.3 63 220 2.08 0.00274SD 2.77 6.23 36.74 0.3 0.00042 1.3 6.79 19.79 0.26 0.00017 1.83 7.83 25.36 0.24 0.00017
Males Mean 27.86 50.64 158.16 1.82 0.00368 22.98 43.55 76.24 1.9 0.00162 25.33 38.06 118.33 1.5 0.00184Minimum 21.5 37.5 50 1.25 0.00063 20.5 32 50 1.36 0.00111 22.7 32 85 1.28 0.00039Maximum 31 61.5 216.57 2.16 0.00508 26 56 105 2.31 0.00204 27.6 42.4 160 1.74 0.00215SD 2.41 8.64 45.29 0.26 0.00037 1.37 6.59 15.77 0.29 0.00018 1.79 3.72 28.83 0.13 0.00030
192 G. L. Ferreira et al.
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014

Table 3. Frequency of occurrence (FO), relative abundance (RA), and Index of Relative Importance (IRI) of items in the dietof Loricariichthys castaneus in the three reservoirs.
Reservoirs
Lajes (n = 52) Vigário (n = 51) Santana (n = 25)
Items FO RA IRI FO RA IRI FO RA IRI
Algae 100 10.38 6.28 95.56 7.86 5.72 76.19 4.78 2.58Rotifers 13.64 0.06 — 24.44 0.2 0.03 — — —
Microcrustaceans 100 8.23 7.52 100 39.6 29.96 76.19 4.41 2.68Protozoans (Testacea) 70.45 2.64 1.09 77.78 1.62 4.21 23.81 0.23 0.03Insects 100 9.18 16.6 86.67 4.98 2.07 76.19 2.79 2.86Other invertebrates 88.64 1.9 0.98 91.11 7.97 4.06 42.86 4.92 1.24Plant debris 84.09 1.33 1.42 93.33 6.4 5.9 90.48 9.97 10.68Sand 100 51.18 26.91 97.78 5.5 2.83 100 26.85 14.54Detritus 100 15.1 39.19 100 25.88 45.22 100 46.06 65.39
Table 2. Comparison of the presence of main items in the stomach content of Loricariichtys castaneus collected in the threereservoirs, in Rio de Janeiro, southeastern Brazil.
Reservoirs
Food items Lajes (n = 52) Vigário (n = 51) Santana (n = 25)
AlgaeFilamentous algae X X XClosterium sp. X XDesmidium sp., Microcystis sp., Micrasterias sp. Spirulina sp. XPediastrum sp., Staurastrum sp., Volvox sp. XTabellaria sp. X XDiatoms X X X
RotifersKeratella americana, Lecane signifera XLecane lunaris XLecane sp. X XPolyarthra sp., Ptygura sp., Trichocerca sp., bdelloids X
MicrocrustaceansBosmina hagmanni, Bosminopsis sp. Simocephalus sp. XBosmina sp. X XCeriodaphnia cornuta, C. silvestrii, Daphnia gessneri, Daphnia sp. X XChydoridae, conchostracs, Diaphanosoma sp. XDiaphanosoma birgei XLeydigia sp., copepod nauplii, calanoid copepods, cyclopoid copepods X X X
OstracodsTestaceansArcella costata, Euglypha sp. XArcella sp., Difflugia sp. X XCentropyxis sp., Lesquereusia sp. Protocurcubitella sp. X
InsectsChironomid larvae, insect debris X X XDipteran larvae XChaoborid larvae X XHemipteran nymphs X
Other invertebratesNematods X XMollusk veligers XAcari X XInvertebrate debris X X X
Plant debris X X XSand grains X X XDetritus X X X
Studies on Neotropical Fauna and Environment 193
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014
We suggest that the higher proportion of inverte-brates, especially aquatic insects, in the LajesReservoir contributed to its better condition in thisecosystem. Protein needed for reproduction and main-tenance (Walker 2009) is considered one of the mostimportant constraints for detritivores and detritivor-ous–omnivorous species (Bowen et al. 1995; Abujanraet al. 2009) and invertebrate prey like insects andmicrocrustaceans clearly provides the highest foodquality in terms of both protein and energy comparedto the primary food resource of algae, macrophytes,and detritus (Bowen et al. 1995).
The crucial role of aquatic insects for fish commu-nities has also been demonstrated in other tropicalreservoirs (Vidotto-Magnoni & Carvalho 2009), flood-plain lakes (Hahn et al. 1997) and coastal lagoons(Branco et al. 1997). This is also true for detritivorousand piscivorous species (Mérona & Rankin-Mérona2004; Vidotto-Magnoni & Carvalho 2009). Becausefood of better nutritional value (aquatic insects) wasoffered by the environment of Lajes Reservoir, fish ofLajes Reservoir contained a lower percentage of detri-tus in the diet of compared with other reservoirs.
The explanation of the second high condition fac-tor in the Santana Reservoir fish sample is probablyexplained by the frequency of macrophytes at thebottom, resulting in the higher presence of detritusand plant debris in the guts of fish. The existence ofeutrophic conditions and a massive proliferation ofmacrophytes at the bottom (Egeria densa), surfaceand shore (Salvinia herzogii, Eichhornia azurea,Brachiaria arrecta and Paspalum repens) (Pitelliet al. 2008) are responsible for the high productionof detritus in this reservoir.
The difference in nutritional contents of availablefood is ecologically important since consumer growthrate is expected to be directly proportional to proteinand energy content of the food (Abujanra et al. 2009;Saravanan et al. 2012). The low body condition foundin fish from the Vigário Reservoir is probably relatedto the lower nutritional value of food offered there.Similar to Lajes Reservoir, the gut content of fishesfrom Vigário showed a lower percentage of detritus;however, also a lower volume of insects and higher ofmicrocrustaceans. Vigário Reservoir has a massiveproliferation of aquatic plants, and, differently fromSantana Reservoir, it has a predominance of floatingplants (Eichhornia, Salvinia and Pistia) and a
Figure 1. Gut content in females and males ofLoricariichthys castaneus (n = 52) captured at LajesReservoir, percentage of food items according to the Indexof Relative Importance (IRI).
Figure 2. Gut content in females and males ofLoricariichthys castaneus (n = 51) captured at VigárioReservoir, percentage of food items according to the Indexof Relative Importance (IRI).
194 G. L. Ferreira et al.
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014

prevalence of pastures in marginal areas. A low pro-duction of detritus from macrophytes and the lack ofaquatic vegetation on the shore probably contributed
to increased foraging on microcrustaceans by L. cas-taneus as an alternative source of protein and energy.The comparatively lower fish body condition in
Figure 3. Gut content in females and males ofLoricariichthys castaneus (n = 25) captured at SantanaReservoir, percentage of food items according to the Indexof Relative Importance (IRI).
Table 4. Items that contributed most to dissimilaritiesbetween genders in the gut contents of Loricariichthys cas-taneus in each reservoir (SIMPER analysis).
Reservoir
Mean volume (mm3)Cumulative %contributionMales Females
LajesChironomidae
larvae309.09 565.45 27.19
Detritus 505.45 624.85 45.85Chaoboridae
larvae279.55 71.21 61.49
Ostracoda 96.36 21.21 67.23Santana
Sand 314.29 262.14 28.16Detritus 424.29 507.14 46.35Plant debris 122.86 94.29 60.12
VigárioLeydigia sp. 253.64 257.83 15.14Detritus 479.09 475.22 24.79Invertebrate
eggs200.45 87.83 33.65
Copepodaremains
149.55 126.09 40.77
Plant debris 136.82 100.00 47.88Algae 80.00 73.04 54.01Calanoida
Copepoda126.82 42.17 59.76
CyclopoidaCopepoda
100.45 63.04 65.32
Sand 112.27 90.87 69.32
Table 5. Items that contributed most to dissimilarities between Lajes and Vigário reservoirs in eachgender of Loricariichthys castaneus (SIMPER analysis).
Mean volume (mm3)
Cumulative % contributionLajes Vigário
MaleChironomidae larvae 309.09 22.73 12.28Detritus 505.45 479.09 22.51Chaoboridae larvae 279.55 3.64 31.39Invertebrate eggs 13.36 200.45 39.14Leydigia sp. 0.00 253.64 45.13Plant debris 0.00 136.82 51.07Copepoda remains 2.00 149.55 56.56Calanoida Copepoda 10.91 126.82 61.17
FemaleChironomidae larvae 565.45 46.52 23.43Detritus 624.85 475.22 36.33Leydigia sp. 0.20 257.83 45.95Plant debris 0.00 100.00 51.17Chaoboridae larvae 71.21 53.91 56.24Copepoda remains 2.85 126.09 60.85
Studies on Neotropical Fauna and Environment 195
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014
Vigário Reservoir might be due to fewer insects in thediet and perhaps a reduced availability of detritus.
Differences between genders
Within our study, a higher number of females havebeen caught in the reservoirs, particularly in the LajesReservoir. Studies analyzing the reproductive cycle ofL. castaneus, which was the dominant species in theLajes Reservoir from 1994 to 1997 (Araújo et al.
1998; Duarte & Araújo 2001) and from 2005 to2006 (Gomes et al. 2011), also found females to belarger and more frequent than males in 66.7–92.8% ofindividual catches. These authors confirmed thereproductive strategy of the species, characterized byallocating large amounts of energy to the protectionof offspring, larger eggs and a prolonged period ofreproduction. Gomes et al. (2011) suggested that thesmaller male size might in part be a consequence ofselection for early male maturation and reproductiveeffort, which reduces male growth compared to thatof females.
Starvation in males during brood care couldexplain the difference in condition factor betweengenders. This proposition can be substantiated by: (i)our own data, which showed a lower percentage ofmales with filled stomachs in both seasons in allreservoirs; and (ii) other studies in the LajesReservoir that reported L. castaneus having a widereproductive period and spawning occurring through-out most of the year (Araújo et al. 1998; Duarte et al.2007), meaning that there is a lower male growth ratedue to feeding restrictions during the egg- and larvae-bearing period (Gomes et al. 2011), which shouldhappen in the three reservoirs.
Gender differences in diet
The main food item in the diet of L. castaneus femalesand males was detritus in all three reservoirs. Thisconfirms the habit indicated by the morphology ofthe species: the mouth in a ventral position and alarger inferior lip, imparting with a sucker, make iteasy to ingest food by suction (Lagler et al. 1977). Thequantity of ingested detritus contributed to diet dis-similarities between genders in Lajes and Santanareservoirs. The lower intake of detritus by males ispossibly associated to feeding restriction in males car-rying eggs and larvae near the mouth. In VigárioReservoir females and males ingested similar meanvolume of detritus, so other food items contributedto the dissimilarities between genders. For males, thelower amount of insects especially may have contri-buted to their smaller size and lower condition factor.
Besides detritus, insects were the food item thatdistinguished the diets of males and females in LajesReservoir. This is also the reason for dissimilarities indiets of the same genders between Lajes and the othertwo reservoirs. Dipteran larvae were the food sourcethat most contributed to gender dissimilarities. A pre-dominance of dipterans has been recorded in otherBrazilian reservoirs (Pamplin et al. 2006; Vidotto-Magnoni & Carvalho 2009). These are probably themost frequent and ubiquitous insect larvae in thebenthic habitats where L. castaneus fish forage.
Table 6. Items that contributed most to dissimilaritiesbetween Lajes and Santana reservoirs in each gender ofLoricariichthys castaneus (SIMPER analysis).
Meanvolume (mm3)
Cumulative %contributionLajes Santana
MaleSand 54.90 314.29 16.52Chironomidae
larvae309.09 41.43 32.19
Detritus 505.45 424.29 45.12Chaoboridae
larvae279.55 — 56.01
Plant debris — 122.86 63.15Female
Chironomidaelarvae
565.45 16.43 32.89
Detritus 624.85 507.14 48.81Sand 65.09 262.14 63.21Plant debris — 94.29 69.41
Table 7. Items that contributed most to dissimilaritiesbetween Vigário and Santana reservoirs in each gender ofLoricariichthys castaneus (SIMPER analysis).
Mean volume(mm3)
Cumulative %contributionVigário Santana
MaleInvertebrate
eggs200.45 115.71 13.28
Sand 112.27 314.29 26.28Leydigia sp. 253.64 7.14 34.43Detritus 479.09 424.29 42.49Plant debris 136.82 122.86 50.49Calanoida
Copepoda126.82 37.14 57.51
FemaleSand 90.87 262.14 14.16Leydigia sp. 257.83 0.71 27.56Detritus 475.22 507.14 40.68Plant debris 100.00 94.29 48.40Copepoda
remains126.09 3.93 54.81
196 G. L. Ferreira et al.
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014

Chironomids were found in the diet of L. castaneus inall three reservoirs and chaoborids mainly in LajesReservoir. The typical vertical movement of chaobor-ids through the water column (Keppeler & Hardy2004) should be facilitated in the deeper water columnof Lajes Reservoir. Since chironomids were moreabundant in females and chaoborids in males, wecan suppose slight differences in foraging sitesbetween genders in the Lajes Reservoir. Males carry-ing eggs might face difficulties in foraging on thebottom and therefore might switch to preying oninsects less restricted to benthic habitats.
In other species of Loricariinae similar dietaryitems were found (Bennemann et al. 2006). Our recordof planktonic microcrustaceans in the stomachs of bothgenders of L. castaneus and in all reservoirs is remark-able. Though adapted to feed on the bottom, the fishescaptured cladocerans, being evidently capable of feed-ing on plankton in the water column. However, themost recurrent zooplankton resource found wasLeydigia sp., a Chydoridae cladoceran that apparentlylives on the surface of mud (Kotov 2009). Therefore,cladocerans and copepods may not be just occasionalfood sources for Loricariichthys as reported byAlbrecht and Silveira (2001), and they contributed todietary dissimilarities between males and females andamong the three reservoirs.
In conclusion, despite the success of L. castaneusin the three reservoirs, differences among the threeenvironments influenced the species’ diets and corre-sponding fitness. Since the highest condition factorswere found in the Lajes Reservoir, better conditionsfor the species in a less eutrophicated environment areinferred. Breeding males with probably impaired feed-ing capacities presumably contributed to decreasedcondition factors of males. The ability of L castaneusto exploit detritus and invertebrates probably explainsits success in the three reservoirs.
AcknowledgmentsThe authors wish to thank A.C.I. Dias and L.C. Souza for assis-tance in the field and Light Energia S.A. for financial and logisticsupport in carrying out this study. We wish to thank to the anon-ymous referees for the critical review of the manuscript.
ReferencesAbujanraF,AgostinhoAA,HahnNS. 2009. Effects of the flood regime
on the body condition of fish of different trophic guilds in the UpperParaná River floodplain, Brazil. Braz J Biol. 69(2):469–479.
Albrecht MP, Silveira CM. 2001. Alimentação deLoricariichthys anus(Teleostei: Loricariidae) nas lagoas Marcelino e Peixoto, planíciecosteira do Rio Grande do Sul. Acta Limnol Bras. 13(2):79–85.
Araújo FG, Fichberg I, Duarte S. 1998. Ciclo reprodutivo deLoricariichthys spixii (Steindachner, 1882) (Pisces Loricariidae)
na represa de Ribeirão das Lajes, RJ. Acta Biol Leopold.20(1):309–318.
Araújo FG, Fichberg I, Pinto BCT, Peixoto MG. 2001. Variaçõesespaciais na assembléia de peixes no Rio Paraíba do Sul (BarraMansa, Barra do Piraí), Rio de Janeiro, Brasil. Rev Bras Zool.18(2):483–492.
Bagenal TB, Tesh FW. 1978. Age and growth. In: Bagenal TB,editor. Methods for assessment of fish production in fresh waters.Oxford (UK): Blackwell Scientific Publications; p. 101–136.
Barbieri G, Peret AC, Verani JR. 1994. Notas sobre a adaptação dotrato digestivo ao regime alimentar em espécies de peixes daregião de São Carlos (SP). I. Quociente intestinal. Rev BrasBiol. 54(1):63–69.
Bennemann ST, Casatti L, Oliveira DC. 2006. Alimentação depeixes: proposta para análise de itens registrados em conteúdosgástricos. Biota Neotrop. 6(2):1–8.
Bowen SH, Lutz EV, Ahlgren MO. 1995. Dietary protein andenergy as determinants of food quality: trophic strategies com-pared. Ecology. 76(3):899–907.
Branco CWC, Aguiaro T, Esteves FA, Caramaschi EP. 1997. Foodsources of the teleost Eucinostomus argenteus in two coastallagoons of Brazil. Stud Neotrop Fauna E. 32(1):33–40.
Branco CWC, Kozlowsky-Suzuki B, Sousa-Filho IF, GuarinoAWS, Rocha RJ. 2009. Impact of climate on the vertical watercolumn structure of Lajes Reservoir (Brazil): a tropical reservoircase study. Lakes R Res Manage. 14(3):175–191.
Clarke KR. 1993. Non-parametric multivariate analysis of changesin community structure. Aust J Ecol. 18(1):117–143.
Delariva RL, Hahn NS, Gomes LC. 2007. Diet of a catfish beforeand after damming of the Salto Caxias Reservoir, Iguaçu River.Braz Arch Biol Tec. 50(5):767–775.
Duarte S, Araújo FG. 2001. Abundância relativa e distribuição deLoricariichthys spixii no reservatório de Lajes, RJ. Rev BrasZool. 18(2):465–477.
Duarte S, Araújo FG, Sales A, Bazzoli N. 2007. Morphology ofgonads, maturity and spawning season of Loricariichthys spixii(Siluriformes, Loricariidae) in a subtropical reservoir. Braz ArchBiol Tec. 50(6):1019–1032.
Gomes ID, Araújo FG, Uehara W, Sales A. 2011. Reproductivebiology of the armoured catfish Loricariichthys castaneus(Castelnau, 1855) in Lajes Reservoir, southeastern Brazil.J Appl Ichthyol. 27(6):1322–1331.
Gomes JHC, Dias ACIM, Branco CWC. 2008. Fish assemblagecomposition in three reservoirs in the State of Rio de Janeiro.Acta Limnol Bras. 20(4):117–130.
Gubiani EA, Gomes LC, Agostinho AA, Okada EK. 2007.Persistence of fish population in the upper Paraná River: effectsof water regulation by dams. Ecol Freshw Fish. 16(2):191–197.
Hahn NS, Fugi R, Almeida VLL, Russo MR, Loureiro VE. 1997.Dieta e atividade alimentar de peixes do reservatório de Segredo. In:Agostinho AA, Gomes LC, editors. Reservatório de Segredo: basesecológicas para o manejo. Maringá (Brazil): Eduem; p. 141–162.
Hyslop EJ. 1980. Stomach contents analysis – a review of methodsand their application. J Fish Biol. 17(4):411–429.
Jones RE, Petrell RJ, Pauly D. 1999. Using modified length–weight relationship to assess the condition of fish. AquacultEng. 20(4):261–276.
Kasper D, Palermo EFA, Dias ACMI, Ferreira GL, Leitão RP,Branco CWC, Malm O. 2009. Mercury distribution in differenttissues and trophic levels of fish from a tropical reservoir, BrazNeotrop Icht. 7(4):751–758.
Keppeler EC, Hardy ER. 2004. Abundance and composition ofRotifera in an abandoned meander lake (Lago Amapa) in RioBranco, Acre, Brazil. Revista Brasileira de Zoologia.21(2):233–241.
Studies on Neotropical Fauna and Environment 197
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014

Kotov AA. 2009. A revision of Leydigia Kurz, 1875 (Anomopoda,Cladocera, Branchiopoda), and subgeneric differentiation withinthe genus. Zootaxa. 2082(1):1–84.
Lagler KF, Bardach JE, Miller RR, Passino DRM. 1977.Ichthyology. New York (NY); p. Wiley. 506.
Lazzaro X, Bouvy M, Ribeiro-Filho RA, Oliveira VS, Sales LT,Vasconcelos ARM, Mata MR. 2003. Do fish regulate phyto-plankton in shallow eutrophic Northeast Brazilian reservoirs?Freshw Biol. 48(4):649–668.
Loureiro-Crippa VE, Hahn NS. 2006. Use of food resources by thefish fauna of a small reservoir (rio Jordão, Brazil) before andshortly after its filling. Neotrop Icht. 4(3):357–362.
Luz-Agostinho KDG, Agostinho AA, Gomes LC, Júlio-Jr HF,Fugi R. 2009. Effects of flooding regime on the feeding activityand body condition of piscivorous fish in the Upper Paraná Riverfloodplain. Braz J Biol. 69(2):481–490.
Mérona B, Rankin-de-Mérona J. 2004. Food resource partitioningin a fish community of the central Amazon floodplain. NeotropIcht. 2(2):75–84.
Mérona B, Santos GM, Almeida RG. 2001. Short term effects ofTucuruí Dam (Amazonia, Brazil) on the trophic organization offish communities. Environ Biol Fish. 60(4):375–392.
Oksanen J, Kindt R, Legendre P, O’Hara B, Simpson GL, Solymos P,StevensMHH,WagnerH. 2009.Vegan: community ecologypackage.R package version 1.15–3 [computer program]; [cited 2009 Feb 15].Available from: http://CRAN.R-project.org/package=vegan
Pamplin PAZ, Almeida TCM, Rocha O. 2006. Composition anddistribution of benthic macroinvertebrates in AmericanaReservoir (SP, Brazil). Acta Limnol Bras. 18(2):121–132.
Petesse ML, Petrere Jr M, Spigolon RJ. 2007. The hydraulic man-agement of the Barra Bonita reservoir (SP, Brazil) as a factorinfluencing the temporal succession of its fish community. Braz JBiol. 67(3):433–445.
Pinkas L, Oliphant MS, Iverson ILK. 1971. Food habits ofAlbacore, Bluefin Tuna, and Bonito in California waters. CalifFish Game. 152(1):1–105.
Pitelli RLCM, Toffaneli CM, Vieira EA, Pitelli RA, Velini ED.2008. Dinâmica da comunidade de macrófitas aquáticas noreservatório de Santana, RJ. Pl Dan. 26(3):473–480.
R Development Core Team. 2010. [cited 2013 Sep 25]. R: A lan-guage and environment for statistical computing [Internet].Vienna: R Foundation for Statistical Computing. Availablefrom: http://www.R-project.org
Reis RE, Pereira EHL. 2000. Three species of the loricariid catfishgenus Loricariichthys (Teleostei: Siluriformes) from SouthernSouth America. Copeia. 2000(4):1029–1047.
Salvador-Jr LF, Salvador GN, Santos GB. 2010. Morphology of thedigestive tract and feeding habits of Loricaria lentiginosa Isbrücker,1979 in a Brazilian reservoir. Acta Zool. 90(2):101–109.
Saravanan S, Schrama JW, Figueiredo-Silva AC, Kaushik SJ,Verreth JAJ, Geurden I. 2012. Constraints on energy intake infish: the link between diet composition, energy metabolism, andenergy intake in rainbow trout. PLoS ONE. 7(4):e34743.
Sunardi, Asaeda T, Manatungue J, Takeshi F. 2007. The effects ofpredation risk and current velocity stress on growth, conditionand swimming energetics of Japanese minnow (Pseudorasboraparva). Ecol Res. 22(1):32–40.
Taylor JN. 1983. Field observations on the reproductive ecology ofthree species of armored catfishes (Loricariidae: Loricariinae) inParaguay. Copeia. 1983(1):257–259.
Vidotto-Magnoni AP, Carvalho ED. 2009. Aquatic insects as themain food resource of fish the community in a Neotropicalreservoir. Neotrop Icht. 7(4):701–708.
Walker I. 2009. Omnivory and resource-sharing in nutrient-deficientRio Negro waters: Stabilization of biodiversity? Acta Amaz. 39(3):617–626.
198 G. L. Ferreira et al.
Dow
nloa
ded
by [
CH
RIS
TIN
A B
RA
NC
O]
at 1
5:54
04
Mar
ch 2
014




















