
Levels of organochlorine compounds in spotted dolphins from the Coiba archipelago, Panama

J. DulcÏ ic á M. Kraljevic á B. Grbec á A. Pallaoro
Composition and temporal ¯uctuations of inshore juvenile ®shpopulations in the Kornati Archipelago, eastern middle Adriatic
Received: 23 March 1997 /Accepted: 11 April 1997
Abstract Temporal ¯uctuations of species compositionand abundance of ®sh juveniles in the National ParkKornati were studied over 12 months from January toDecember 1992. A total of 40 168 individuals, repre-senting 24 families and 69 species, were collected using a50 m long beach seine. The community was dominatednumerically by a few species: Atherina hepsetus (45.90%),Sarpa salpa (21.21%), Diplodus vulgaris (10.49%), andSymphodus ocellatus (6.98%), constituting over 84.58%of the total catch. Similarity between patterns in theabundance of the 17 most common species were exam-ined using correlation matrix-based principal compo-nent analysis. Results were compared to the abioticfactors leading to the conclusion that only a low amountof variation in the abundance ®eld can be explained bytemperature, salinity and dissolved oxygen.
Introduction
Numerous studies have been published on ®sh popula-tions in lagoons, estuaries and coastal regions in manyparts of the world. Most of the studies are on seasonalvariations in the number of individuals, species diversity,and the in¯uence of abiotic factors on abundance andspecies performance (Haedrich and Haedrich 1974; Allenand Horn 1975; Allen 1982; Allen et al. 1983; Beckley1984; Reina-Hervas and Serrano 1987; Tzeng and Wang1992; Whit®eld 1994; Laegdsgaard and Johnson 1995).However, most of the published material on ®shes in theeastern Adriatic is restricted to taxonomy, general ®sheryaspects, and, to a lesser extent, to the biology of variousspecies (Morovic 1960, 1962, 1965; GrubisÏ ic 1974; Jardas1980 a,b, 1982, 1986, 1996; Jardas and Pallaoro 1989;
DulcÏ ic 1993a, 1995; DulcÏ ic and Cetinic 1993; DulcÏ ic andKraljevic 1995; Kraljevic et al. 1995, 1996; Kraljevic andDulcÏ ic 1996. There have only been a few papers on ju-venile ®sh populations in the eastern Adriatic. Some re-search has dealt with the temporal distribution of youngmugilids in the coastal area of the eastern middleAdriatic (Katavic 1980; Jug-Dujakovic 1988) and somewith feeding of striped sea bream, Lithognathus mo-rmyrus (Froglia 1977; Jardas 1985), and annular bream,Diplodus annularis (Jardas et al. 1986). Previous prelim-inary investigations on juvenile ®sh populations inKornati Archipelago are limited and incomplete, con-cerned just with occurrence and quantity of species(Kraljevic and Jug-Dujakovic 1987; Kraljevic andPallaoro 1991).
The present study provides the ®rst data on thetemporal ¯uctuations in juvenile ®sh composition, di-versity indices and ®sh species association in the coves ofNational Park Kornati (eastern middle Adriatic). Therelative importance of temperature, salinity and dis-solved oxygen on biotic factors was examined usingmultivariate techniques.
Materials and methods
Study area
The Republic of Croatia with a surface area of 88 438 km2 (land,sea and islands) has seven national parks and numerous otherprotected areas. The Kornati Islands lie along the central part ofthe Croatian coast, between Zadar and SÏ ibenik. The Kornati Ar-chipelago covers an area of approximately 224 km2 between Pro-versa (NW) and Samogradska vrata (SE) (Fig. 1) and has 91islands, islets and reefs. The Kornati Islands have the most irreg-ular coastlines of any islands in the Mediterranean. Human activ-ities in the area include ®shing, the traditional occupation ofKornati inhabitants and inhabitants from the island Murter, andtourism (MihelcÏ ic 1997).
Sampling techniques
Sampling was conducted on a monthly basis from January toDecember 1992 from ®ve coves: SÏ ipnata, Lojena, Studena, Lavsa
Marine Biology (1997) 129: 267±277 Ó Springer-Verlag 1997
Communicated by O. Kinne, Oldendorf/Luhe
J. DulcÏ ic á M. Kraljevic á B. Grbec á A. PallaoroInstitute of Oceanography and Fisheries,SÏ etalisÏ te Ivana MesÏ trovic a 63, P.O.B. 500,21000 Split, Croatia

and ZÏ akan (Fig. 1). Fish samples were collected using a 50 m longbeach seine. Net depth at the beginning of wings was 30 cm and250 cm at the central part together with the sac. Outer wings wereof 8 mm mesh size and central sac of 4 mm. The net was alwayshauled from the entrance to the cove (max. 7 m depth) to its innerend. Collected organisms were sorted and preserved in 4% formalin(pH from 8.5 to 9.0). The ®sh species were identi®ed according toSÏ oljan (1975) and Jardas (1996). The juveniles of the species weretaken as specimens with already formed scales, and were taken assuch until the moment of ®rst sexual maturity (Katavic 1984). Thesampling area was characteristically hard, sandy, sandy-clay andsandy-mud laterally overgrown by meadows of Posidonia oceanicaor Cymodocea nodosa. Physical±chemical parameters were mea-sured by classical methods in every cove before net hauling. Tem-perature was measured with a mercury thermometer, salinity with alaboratory inductive salinometer, and dissolved oxygen (DO) withWinkler's method.
Data analysis
Temporal ¯uctuations in community structure, abiotic factors andinteractions between these variables were analysed using multi-variate techniques: multiple linear regression and principal com-ponent analysis (Preisendorfer 1982). Community structure wasspeci®ed by species richness (D), diversity (H¢ ) and evenness (J¢ )using formulae proposed by Margalef (1968), Shannon and Weaver(1949) and Pielou (1966), respectively. The degree of relationshipexisting between these variables and abiotic factors (temperature,salinity and dissolved oxygen) was determined by R2, e.g. the co-e�cient of multiple determination.
Abundance in the species compositions was subject to thecorrelation matrix-based principal component analysis (PCA)(Preisendorfer 1982) to examine a minimal number of groups(clusters) the variability of which describes the maximum amountof the total variability in the ®sh community. The PCA was per-formed on the correlation matrix of standardised variables (zeromean and unit standard deviation). We used Bartlett's test (Fulgosi1988) for testing the signi®cance of the correlation matrix. The
signi®cance of extracted principal components (eigenvalues) weretested with Rule N (Overland and Preisendorfer 1982), usingMonte Carlo simulation of the random matrix of the same size asthe original data matrix. To enhance the interpretation of thecomponents, Varimax orthogonal rotation was applied (Richman1986).
The relationship of species composition between the ®ve sta-tions was compared using Spearman's rank correlation test on thebasis of the yearly catch in terms of number of each individualspecies. For comparing abiotic factors between ®ve sampling lo-cations, an ANOVA model was used.
Results
Temporal ¯uctuations of abiotic factors
Analysis of variance revealed signi®cant correlationsbetween temperature (ANOVA: p < 0.01; df = 3.65;F = 0.14), salinity (F = 2.51) and dissolved oxygen(F = 1.09) among the ®ve sampling stations. Meanmonthly variation in sea temperature, salinity and dis-solved oxygen (DO) averaged over the ®ve coves arepresented in Fig. 2. Mean monthly temperature rangedfrom 11.1 °C (SD = 0.98) in February to 26.2 °C(SD = 1.64) in August. Mean monthly salinity values,however, were more stable (ranging from 37.46 � 1.68to 38.52 � 0.51 psu, with the exception of Novemberwhen the salinity declined sharply to 35.86 � 4.40 psu).Oxygen concentration level ¯uctuated between 6.07 mll)1 (SD = 0.94) and 8.17 ml l)1 (SD = 0.88). While seawater temperature shows typically Adriatic seasonalcycles, seasonal cycles in salinity are not evident, mainlybecause the sampling stations are situated in the coastal
Fig. 1 Locations of the sam-pling stations (d) in the KornatiArchipelago
268

area which is strongly in¯uenced by precipitation andevaporation.
Temporal ¯uctuations in ®sh compositionsand abundance
Spearman's rank correlation coe�cients of speciescomposition between the ®ve stations were signi®cant atp = 0.01, indicating that the rank of species composi-tion was highly correlated with sampling location. So,data for all coves were combined. Sixty-nine species,representing 24 families, were caught between Januaryand December 1992 (Table 1), the highest number wascollected in June (41) and the lowest during the winterperiod from December (23) to March (22). The numberof species captured increased in late spring and earlysummer (May to June) and also in autumn (Septemberto November), and decreased in winter (December toMarch).
A total of 40 168 ®shes were collected: the highestcollection was in April (8958), and the lowest in theperiod from June (1108) to September (1078). Atherinahepsetus (45.895%), Sarpa salpa (21.206%), Diplodusvulgaris (10.488%) and Symphodus ocellatus (6.983%)comprised 84.572% of the total sample (Table 1).A. hepsetus was the dominant species in the collection,and had the highest abundance in April. S. salpa andD. vulgaris had the highest abundance from January toJune, while the abundance of S. ocellatus was highest inthe period from August to December. The remainingspecies (65) comprised from 2.338 to 0.003% of the to-tal catch. The overall value of richness D was 6.42,ranging from 2.35 in March to 5.71 in June. H¢ values¯uctuated from 1.12 in April to 2.79 in June, with anoverall value of 2.94. J¢ ranged from 0.77 to 0.32. Meanmonthly variations in these biotic factors are presentedin Fig. 3.
Multivariate analysis of relationship between bioticand abiotic factors
The relationship between abiotic factors such as tem-perature, salinity and dissolved oxygen (included inmodel as independent variables) and the biotic factors(number of species, number of individuals, diversity andabundances: dependent variables) was analysed usingmultiple linear regression models. The signi®cance levelfor the variables included in the model was set atp < 0.05. Results indicated that variations in abioticfactors could predict more than 85% of monthly varia-tions in number of species mainly because of the inter-action e�ect of temperature and salinity. Variation innumber of individuals showed no signi®cant correlationswith independent variables. Results also suggest thatabout 78% of the variability associated with monthly¯uctuations in diversity could be predicted by the tem-perature and oxygen concentration levels. These resultsindicated that even if there is a temperature ``control-ling'' monthly ¯uctuations of number of species anddiversity, derivation of a non-zero association betweenthese biotic factors and sea water temperature does notnecessarily imply a casual relationship. In temperate seassuch as the Adriatic, most species have internalised theirbiological cycles according to seasonal patterns. Theyspawn or settle in a quite short and well-de®ned periodof the year. So, abundances of many species were posi-tively or negatively correlated with sea water tempera-ture (Table 2), because they settle during the hot or coldseason. This is a reason why multilinear correlationcoe�cients between the number of individuals and watertemperature are nonsigni®cant. Based on the multiplelinear regression model it was found that six of 20common species positively (Coris julis and Mullussurmuletus) or negatively (Diplodus puntazzo, Diplodusvulgaris, Sarpa salpa and Oedalechilus labeo) correlatedwith temperature, indicating that timing for settling ofthe individual species is di�erent. Only one species (Liza
Fig. 2 Monthly variations in the mean sea water temperature (°C),salinity (psu) and dissolved oxygen (ml l)1) in the Kornati Archipel-ago between January and December 1992
269
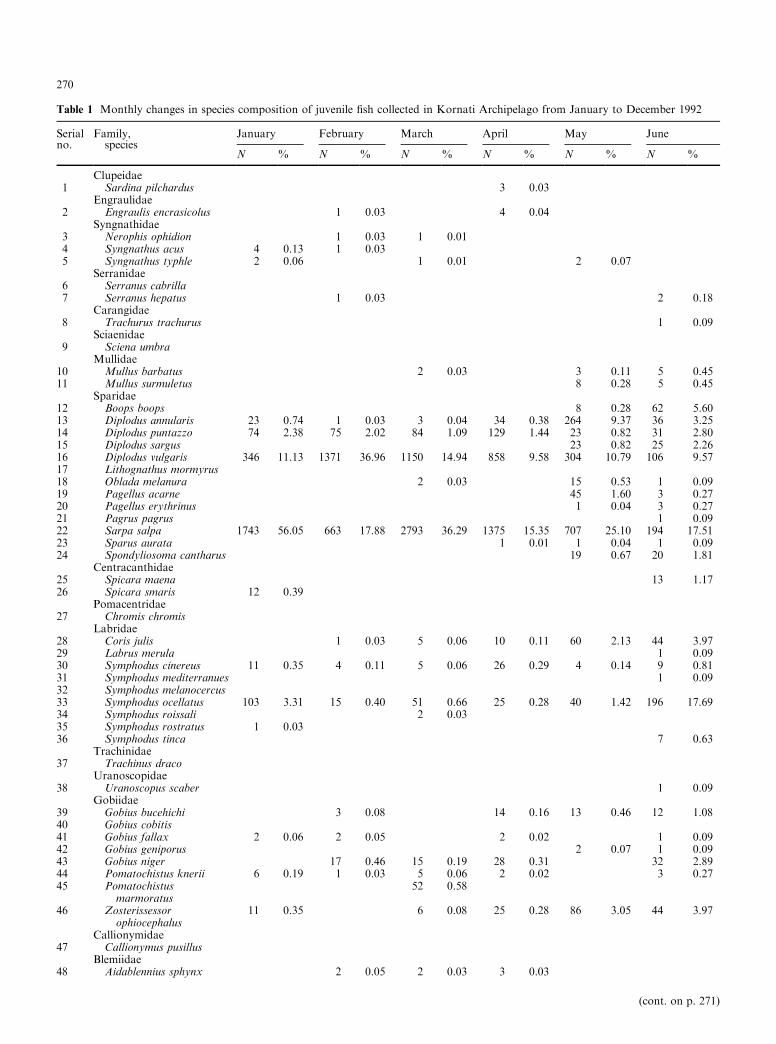
Table 1 Monthly changes in species composition of juvenile ®sh collected in Kornati Archipelago from January to December 1992
Serial Family, January February March April May Juneno. species
N % N % N % N % N % N %
Clupeidae1 Sardina pilchardus 3 0.03
Engraulidae2 Engraulis encrasicolus 1 0.03 4 0.04
Syngnathidae3 Nerophis ophidion 1 0.03 1 0.014 Syngnathus acus 4 0.13 1 0.035 Syngnathus typhle 2 0.06 1 0.01 2 0.07
Serranidae6 Serranus cabrilla7 Serranus hepatus 1 0.03 2 0.18
Carangidae8 Trachurus trachurus 1 0.09
Sciaenidae9 Sciena umbra
Mullidae10 Mullus barbatus 2 0.03 3 0.11 5 0.4511 Mullus surmuletus 8 0.28 5 0.45
Sparidae12 Boops boops 8 0.28 62 5.6013 Diplodus annularis 23 0.74 1 0.03 3 0.04 34 0.38 264 9.37 36 3.2514 Diplodus puntazzo 74 2.38 75 2.02 84 1.09 129 1.44 23 0.82 31 2.8015 Diplodus sargus 23 0.82 25 2.2616 Diplodus vulgaris 346 11.13 1371 36.96 1150 14.94 858 9.58 304 10.79 106 9.5717 Lithognathus mormyrus18 Oblada melanura 2 0.03 15 0.53 1 0.0919 Pagellus acarne 45 1.60 3 0.2720 Pagellus erythrinus 1 0.04 3 0.2721 Pagrus pagrus 1 0.0922 Sarpa salpa 1743 56.05 663 17.88 2793 36.29 1375 15.35 707 25.10 194 17.5123 Sparus aurata 1 0.01 1 0.04 1 0.0924 Spondyliosoma cantharus 19 0.67 20 1.81
Centracanthidae25 Spicara maena 13 1.1726 Spicara smaris 12 0.39
Pomacentridae27 Chromis chromis
Labridae28 Coris julis 1 0.03 5 0.06 10 0.11 60 2.13 44 3.9729 Labrus merula 1 0.0930 Symphodus cinereus 11 0.35 4 0.11 5 0.06 26 0.29 4 0.14 9 0.8131 Symphodus mediterranues 1 0.0932 Symphodus melanocercus33 Symphodus ocellatus 103 3.31 15 0.40 51 0.66 25 0.28 40 1.42 196 17.6934 Symphodus roissali 2 0.0335 Symphodus rostratus 1 0.0336 Symphodus tinca 7 0.63
Trachinidae37 Trachinus draco
Uranoscopidae38 Uranoscopus scaber 1 0.09
Gobiidae39 Gobius bucehichi 3 0.08 14 0.16 13 0.46 12 1.0840 Gobius cobitis41 Gobius fallax 2 0.06 2 0.05 2 0.02 1 0.0942 Gobius geniporus 2 0.07 1 0.0943 Gobius niger 17 0.46 15 0.19 28 0.31 32 2.8944 Pomatochistus knerii 6 0.19 1 0.03 5 0.06 2 0.02 3 0.2745 Pomatochistus
marmoratus52 0.58
46 Zosterissessorophiocephalus
Callionymidae
11 0.35 6 0.08 25 0.28 86 3.05 44 3.97
47 Callionymus pusillusBlemiidae
48 Aidablennius sphynx 2 0.05 2 0.03 3 0.03
(cont. on p. 271)
270
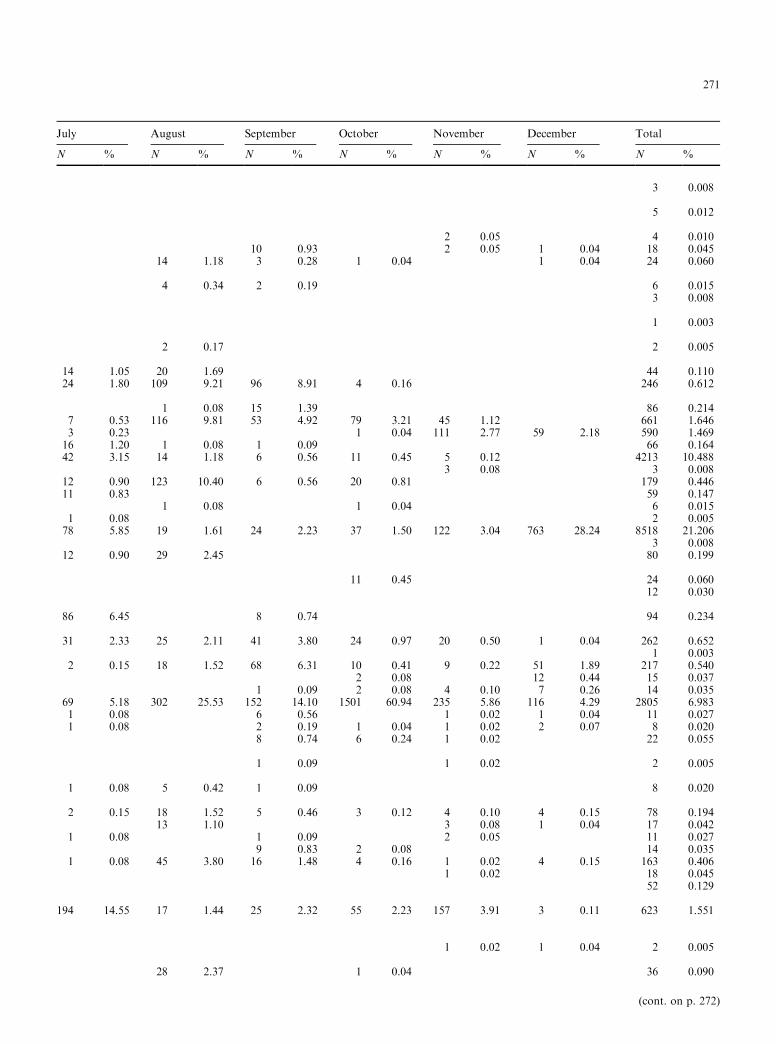
July August September October November December Total
N % N % N % N % N % N % N %
3 0.008
5 0.012
2 0.05 4 0.01010 0.93 2 0.05 1 0.04 18 0.045
14 1.18 3 0.28 1 0.04 1 0.04 24 0.060
4 0.34 2 0.19 6 0.0153 0.008
1 0.003
2 0.17 2 0.005
14 1.05 20 1.69 44 0.11024 1.80 109 9.21 96 8.91 4 0.16 246 0.612
1 0.08 15 1.39 86 0.2147 0.53 116 9.81 53 4.92 79 3.21 45 1.12 661 1.6463 0.23 1 0.04 111 2.77 59 2.18 590 1.46916 1.20 1 0.08 1 0.09 66 0.16442 3.15 14 1.18 6 0.56 11 0.45 5 0.12 4213 10.488
3 0.08 3 0.00812 0.90 123 10.40 6 0.56 20 0.81 179 0.44611 0.83 59 0.147
1 0.08 1 0.04 6 0.0151 0.08 2 0.00578 5.85 19 1.61 24 2.23 37 1.50 122 3.04 763 28.24 8518 21.206
3 0.00812 0.90 29 2.45 80 0.199
11 0.45 24 0.06012 0.030
86 6.45 8 0.74 94 0.234
31 2.33 25 2.11 41 3.80 24 0.97 20 0.50 1 0.04 262 0.6521 0.003
2 0.15 18 1.52 68 6.31 10 0.41 9 0.22 51 1.89 217 0.5402 0.08 12 0.44 15 0.037
1 0.09 2 0.08 4 0.10 7 0.26 14 0.03569 5.18 302 25.53 152 14.10 1501 60.94 235 5.86 116 4.29 2805 6.9831 0.08 6 0.56 1 0.02 1 0.04 11 0.0271 0.08 2 0.19 1 0.04 1 0.02 2 0.07 8 0.020
8 0.74 6 0.24 1 0.02 22 0.055
1 0.09 1 0.02 2 0.005
1 0.08 5 0.42 1 0.09 8 0.020
2 0.15 18 1.52 5 0.46 3 0.12 4 0.10 4 0.15 78 0.19413 1.10 3 0.08 1 0.04 17 0.042
1 0.08 1 0.09 2 0.05 11 0.0279 0.83 2 0.08 14 0.035
1 0.08 45 3.80 16 1.48 4 0.16 1 0.02 4 0.15 163 0.4061 0.02 18 0.045
52 0.129
194 14.55 17 1.44 25 2.32 55 2.23 157 3.91 3 0.11 623 1.551
1 0.02 1 0.04 2 0.005
28 2.37 1 0.04 36 0.090
(cont. on p. 272)
271

aurata) was negatively correlated with salinity, while theother species' correlations were insigni®cant. This indi-cates that most species could tolerate wide variation ofsalinity. Only two species (Symphodus ocellatus andBoops boops) were signi®cantly correlated with dissolvedoxygen (Table 2).
To consider which species, or group of species, con-tributes signi®cantly to the community structure, prin-cipal component analyses were performed. According tononsigni®cance of the correlation matrix (Bartlett's test)for all determined species (20), PC analysis was done forthe 17 most frequent species in the area of investigation(Liza saliens, Chromis chromis and Boops boops wereexcluded). Correlation matrix (for 17 species) was sig-ni®cant for p = 0.05. The ®rst ®ve principal compo-nents together describe 79.03% of the total abundancevariance: PC1 contributes with 33.86%, PC2 with18.15%, and PC3 with 11.02%. The remaining two ex-tracted principal components contribute with 8.57 and7.42% to the total variability in the abundance ®eld.When using only percentages of total variances it isdi�cult to choose the number of PCs with signi®cantmeanings. Therefore, Rule N was applied. The ®rst threePCs are signi®cant because the eigenvalues of the real
data matrix are smaller than those of the random datamatrix. On the basis of the distribution of rotatedloadings it was possible to distinguish ®ve groups(Fig. 4), but only the ®rst three signi®cantly contributewith di�erent amounts to the total variability in speciesabundance. PC1 has the highest loadings for Atherinahepsetus, Diplodus puntazzo and Chelon labrosus. PC2
has highest loadings for Mullus surmuletus, Oblada me-lanura, Liza ramada and Gobius niger. In the case of PC3,four species were grouped: Coris julis, Diplodus annul-aris, Atherina boyeri and Symphodus ocellatus.
Scores of the ®rst three principal components werecompared with sea water temperature, salinity and DOshowing that only a low variability in abundances can beexplained by these abiotic factors. Signi®cant linearcorrelation was found between temperature and PC2
scores only ( p < 0.10; r = 0.54).
Discussion
The Adriatic Sea is an oligotrophic sea (Zore-Armandaet al. 1987). It is relatively rich in ichthyofauna and ac-cording to Jardas (1996) has 407 ®sh species and sub-
Table 1 (cont.)
Serial Family, January February March April May Juneno. species
N % N % N % N % N % N %
49 Lipophrys dalmatinus 2 0.03 1 0.0450 Lipophrys pavo 2 0.02 5 0.18 8 0.7251 Lipophrys trigloides 2 0.0652 Parablennius incognitus 1 0.03 2 0.07 1 0.0953 Parablennius
sanguinolentus2 0.02 2 0.18
54 Parablenniustentacularis
1 0.03 1 0.01 2 0.07
Sphyraenidae55 Sphyraena sphyraena
Mugilidae56 Chelon labrosus 21 0.68 4 0.11 13 0.17 40 0.45 5 0.4557 Liza aurata 94 3.02 179 4.83 34 0.44 20 0.22 4 0.14 61 5.5158. Liza ramada 1 0.03 23 2.0859 Liza saliens 11 0.35 20 0.54 12 0.4360 Oedalechilus labeo 75 2.41 186 5.01 112 1.45 189 2.11 42 1.49 65 5.87
Atherinidae61 Atherina boyeri 157 4.2362 Atherina hepsetus 564 18.14 1003 27.04 3407 44.27 6110 68.21 1120 39.76 77 6.95
Scorpaenidae63 Scorpaena porcus 2 0.06 1 0.01 1 0.01 1 0.04 2 0.18
Triglidae64 Trigla lucerna
Bothidae65 Arnoglossus lanterna 1 0.0966 Arnoglossus thori67 Bothus podas podas 1 0.01 2 0.18
Soleidae68 Solea impar 1 0.01
Gobiesocidae69 Diplecogaster bimaculata 1 0.03
Total no. of individuals 3110 3709 7696 8958 2817 1108Total no. of species 23 23 22 27 29 41
272

species. According to the results of this study, 69 species,either permanently or temporarily, occupy small covesof Kornati Islands. This number represents 17.0% ofAdriatic ichthyofauna.
Juveniles of four species (Atherina hepsetus, 45.90%;Sarpa salpa, 21.21%; Diplodus vulgaris, 10.49% andSymphodus ocellatus, 6.98%) were dominant, forming84.58% of the total. The dominant species of the ®shcommunity in this study area belonged to the productiveand low trophic level species with high ecological e�-ciency. The ®sh entering the coves of Kornati Archi-pelago were mainly in the juvenile stage (as a startingpoint for the ®sh to be considered juvenile total length of1.5 cm was taken, and ®sh were considered juvenilesuntil the point of their ®rst sexual maturity) of devel-opment, and a small number of adults (from their ®rstsexual maturity onwards) were found. This di�erence inabundance can be explained by the fact that the inshorearea is closely related to the presence of certain coastalspecies and of the larvae and juvenile stages of more orless neritic species, which in many cases go through theircomplete biological cycle in this zone. These facts alsoindicate that the study sites probably play an importantrole as a nursery ground for the ®sh during the inshore±
o�shore migration of their early life history. Such areasalso provided suitable food, shelter, and a reduction ofpredation. The nursery function of estuaries and inshorewaters has been well-documented all over the world(Blaber and Blaber 1980; Lenanton 1982; Robertson andDuke 1987; Blaber and Milton 1990). Kraljevic andPallaoro (1991) observed that juveniles of S. ocellatus(35.42%) and A. hepsetus (19.26%) were dominant inthe same area, but only for 3 months during sampling(May, July and October), which is in agreement with ourresults. According to our investigations two families,Atherinidae (46.29%) and Sparidae (36.02%), domi-nated, while Labridae (8.35%), Mugilidae (5.12%) andGobiidae (2.43%) were less abundant and all otherfamilies (19) were rare and occasional (<1.00%). Similarresults were obtained by Kraljevic and Pallaoro (1991),who found that Atherinidae comprised 19.26%, Spar-idae 24.79%, and Labridae 37.88% of the sample, whileMugilidae (4.88%) and Gobiidae (7.60%) were lessabundant.
A high abundance of di�erent species was observedduring two periods in the year: May to June and Sep-tember to November, being characterised by mediantemperatures in comparison to the hot summer and cold
July August September October November December Total
N % N % N % N % N % N % N %
2 0.15 1 0.02 2 0.07 8 0.02018 1.35 18 1.67 8 0.32 2 0.05 61 0.15210 0.75 8 0.20 20 0.050
2 0.08 6 0.0155 0.38 5 0.46 2 0.05 16 0.040
1 0.08 19 1.61 5 0.46 2 0.08 3 0.08 34 0.085
2 0.19 2 0.005
5 0.38 18 1.52 8 0.74 13 0.32 4 0.15 131 0.3265 0.42 24 2.23 215 5.36 36 1.33 672 1.67377 6.51 68 6.31 3 0.08 12 0.44 184 0.45834 2.87 44 4.08 8 0.30 129 0.321
19 1.43 52 4.40 14 1.30 44 1.79 30 0.75 111 4.11 939 2.338
2 0.17 159 0.396661 49.59 50 4.23 316 29.31 627 25.46 2998 74.74 1502 55.59 18435 45.895
1 0.08 1 0.08 8 0.74 3 0.12 3 0.08 23 0.057
1 0.08 1 0.003
1 0.02 1 0.0033 0.28 1 0.04 5 0.012
2 0.15 3 0.28 1 0.02 9 0.022
1 0.003
1 0.003
1333 1183 1078 2463 4011 2702 4016832 32 38 28 35 23 69
273

winter. GrubisÏ ic (1982) and Jardas (1996) indicated thatspring and summer were the spawning periods for mostcommon species in the eastern Adriatic. So, we supposerecruitment timing of these species could be the reasonfor a high abundance during the mentioned periods. Ahigh abundance in autumn was due to the increase ofAtherina hepsetus, Diplodus puntazzo, Oedalechilus labeoand Symphodus ocellatus juveniles, and this could becorrelated with the spawning periods (Jardas 1996) andduration of embryonic development of this species(DulcÏ ic 1993b). Ecological separation of the dominantspecies by recruitment timing resulted in the fact thatspecies did not compete one with the other for the sameniche (Tzeng and Wang 1992). Temporal staggering ofrecruitment may be a mechanism for reducing possibleinterspeci®c competition, as has been observed in other®sh communities (Doherty 1991). Segregation of somesparid species, for example, was seasonal in KornatiArchipelago since they recruit at di�erent times of theyear (Diplodus sargus, Pagellus acarne and Spondylio-soma cantharus in spring; Oblada melanura duringsummer; Diplodus puntazzo at the beginning of autumn;Sarpa salpa during autumn; Diplodus vulgaris at thebeginning of winter), with each species occupying zonespreviously occupied by another species some timebefore.
The Kornati Archipelago ®sh assemblage exhibited agreat range for the values of D (2.35 to 5.71), H¢ (1.12 to2.79), and J¢ (0.32 to 0.77). These values could becomparable with values obtained in studies performed insimilar areas, bays and estuaries (Table 3). This rela-tively wide range of values re¯ects a di�erential utilisa-tion of Kornati Archipelago habitats by various ®shspecies, which is similar to the ®ndings of Allen (1982) inupper Newport Bay, California. McErlean et al. (1973)considered the wide range of richness values as an in-
Fig. 3 Monthly variations in the number of individuals (N ), numberof species (S ), richness, diversity and evenness in the KornatiArchipelago between January and December 1992
Table 2 Coe�cient of multiple determination (R2) identifying the association according to rank of the 20 most dominant species,(abbreviations in parentheses) with temperature, salinity and dissolved oxygen (DO)
Rank no. Serial no. Species Temperature Salinity DO R2 p
1 62 Atherina hepsetus (AH) 0.33 0.342 22 Sarpa salpa (SS) )0.73 0.55 0.083 16 Diplodus vulgaris (DV) )0.72 0.55 0.084 33 Symphodus ocellatus (SO) )0.64 0.45 0.175 60 Oedalechilus labeo (OL) )0.78 0.64 0.046 57 Liza aurata (LA) )0.70 0.64 0.047 13 Diplodus annularis (DA) 0.19 0.628 46 Zoosterisessor ophiocephalus (ZO) 0.51 0.119 14 Diplodus puntazzo (DP) )0.74 0.65 0.0310 28 Coris julis (CJ) +0.72 0.55 0.0811 11 Mullus surmuletus (MS) +0.65 0.48 0.1412 30 Symphodus cinereus (SC) 0.03 0.9713 58 Liza ramada (LR) 0.39 0.2414 18 Oblada melanura (OM) 0.44 0.1715 43 Gobius niger (GN) 0.25 0.4816 61 Atherina boyeri (AB) 0.22 0.5417 56 Chelon labrosus (CL) 0.06 0.9118 59 Liza saliens (LS) 0.12 0.7819 27 Chromis chromis (CC) 0.22 0.5520 12 Boops boops (BB) +0.69 0.54 0.09
274

dicator of the nursery function of a speci®c area. So, wecan suppose that the overall high richness values of ®shassemblages in Kornati Archipelago underline the im-portance of the area as a nursery ground for several localspecies.
The highest number of juvenile individuals in KornatiArchipelago was collected during the period fromMarch to April, and the lowest from June to September.Morovic (1960) and Kraljevic and Pallaoro (1991) sug-gested that ®ngerlings of many species migrate to thedeeper waters of the Kornati Archipelago (>7 m) in thesummer and winter (extremely high and low tempera-tures in the shallow coves). Allen and Horn (1975) foundthat temperature was the main factor a�ecting the ®shpopulation in the Colorado Lagoon. Allen (1982) ob-served that temperature and salinity accounted for 83%of the variation in abundance of individual ®sh inNewport Bay. The high positive correlation of temper-ature to number of species played an important role in®sh migration to and from Kohr-al-Zubair in thenorthwestern Arabian Gulf (Ali and Hussain 1990). Thenumber of species, abundance, and indices of speciesrichness and diversity of the community of ®sh larvaeand juveniles in the Tanshui River estuary were posi-tively correlated with temperature and salinity (Tzengand Wang 1992). The same authors observed that dis-solved oxygen, pH, depth, transparency and current
velocity are also positively or negatively correlated withthe number and abundance of ®sh species (larvae andjuveniles) in the Tanshui River estuary (two speciescorrelated with DO, ®ve species correlated with depth).
It is di�cult to explain species arrangement accordingto PC loadings. However, species in the case of PC1 arespecies (Atherina hepsetus, Diplodus puntazzo, and Che-lon labrosus) which were generally abundant and hadtwo peaks in their abundances during the year (Apriland November). The same conclusion could be drawnfor species in the case of PC2, with a peak in August andpartial peak in September (Mullus surmuletus, Obladamelanura, Liza ramada and Gobius niger). PC3 hashighest loadings for Coris julis, Diplodus annularis, At-herina boyeri, and Symphodus ocellatus, which are ben-thic species associated with Posidonia oceanica beds andcharacterised by a very similar reproduction time (Jar-das 1996).
Substrate type (Carr 1991; Levin 1991) and depth(Garcia-Rubies 1995; Garcia-Rubies and Macpherson1995) are two of the main factors a�ecting ®sh recruit-ment in the shallow waters, and a�ect the abundance,mortality and growth rate of recruits (Connell and Jones1991; Macpherson 1994). It is apparent from the fore-going discussion that a number of parameters in¯uencethe di�erential distribution of juveniles of the speciesstudied. The relative importance of each factor di�ersaccording to species. Perhaps the only commondenominator is the preference of juveniles for relativelyshallow water. There may be several di�erent explana-tions for the shallow distribution of recruits. Di�erentauthors have reported that prey abundance (Jones 1986;Forrester 1990), predation (Doherty and Sale 1985), andthe presence of adult conspeci®cs (Victor 1986; Jones1987) may exert a positive or negative in¯uence on thedistribution and abundance of juveniles. The distribu-tion in shallow waters could be also interpreted as asheltering behaviour from possible predators, becausepredator manoeuvrability may be hampered in shallowwater. It is also possible that sampling design (our choicein number of surveys, speci®c locations, type of net)could partially in¯uence the results.
Preliminary results of the present study provide abasis for establishing the temporal and spatial patterns
Fig. 4 Scatter plot of the loadings along PC1 versus PC2 of 17 mostcommon species. Letter coding see Table 2
Table 3 Diversity index values from the literature
Area D H¢ J¢ Source
Upper Galvestone Bay, Texas 0.11±2.01 Bechtel and Copeland (1970)Colorado Lagoon, California 0.5±1.73 0.03±1.11 0.01±0.57 Allen and Horn (1975)Lower Medway Estuary, Kent 1.5±3.4 0.25±1.89 0.10±0.96 van den Broek (1979)Upper Newport Bay, California 0.42±1.76 Allen (1982)Iriven Bay, west coast of Scotland 1.5±3.08 1.17±1.97 0.57±0.77 Nash and Gibson (1982)Lynn of Lorn, west coast of Scotland 1.4±3.34 1.23±1.95 0.50±0.72 Nash and Gibson (1982)Cabrillo beach area, Los Angeles 0.50±2.50 Allen et al. (1983)Malaga Bay, Spain 1.17±3.00 0.27±0.65 Reina-Hervas and Serrano (1987)Kohr-al-Zubair, northwestern Arabian Gulf 1.17±3.47 1.19±2.36 0.57±0.80 Ali and Hussain (1990)Island el Hierro, Canary Islands 0.57±1.39 Bortone et al. (1991)Four Islands, Canarian Archipelago 0.27±0.78 Falco n et al. (1996)Kornati Archipelago, eastern Adriatic 2.35±5.71 1.12±2.79 0.32±0.77 Present study
275

of recruitment in various ®sh species. Following thisstudy, we recommend long-term research to establishwhether the correlation between biotic factors andtemperature really exists. This should be done aftereliminating seasonal patterns from both data sets. Fur-ther work is needed to elucidate which additional aspectsmay in¯uence the distribution and abundance of juve-niles in the small coves of Kornati Archipelago.
Acknowledgements We dedicate this paper to Mr. Zvonko Mod-rusÏ an and Mr. Miran TurcÏ inov, who supported our work at theNational Park Kornati and died tragically in a car accident on28 March 1997. We are very thankful to R. C uzela Papata,E. KulusÏ ic , D. Dobric , K. KulusÏ ic M. RamesÏ a, N. Stipica, andCÏ . SkracÏ ic (sta� of National Park Kornati) for their great help incollecting the material. Also, we would like to thank the sta� of thechemical laboratory of the Institute of Oceanography and Fisheriesfor their help in chemical analysis and to N. Skakelja for technicalpreparation of the manuscript. The authors express their gratitudeto the Ministry of Science, Technology and Informatics of theRepublic of Croatia for their ®nancial support.
References
Ali TS, Hussain NA (1990) Composition and seasonal ¯uctuationsof intertidal ®sh assemblage in Kohr-al-Zubair, northwesternArabian Gulf. J appl Ichthyol 1: 24±36
Allen LG (1982) Seasonal abundance, composition and produc-tivity of ®sh assemblage in upper Newport Bay, California. FishBull US 80: 769±790
Allen LG, Horn MHK (1975) Abundance, diversity and season-ality of ®shes in Colorado Lagoon, Alamitos Bay, California.Estuar cstl mar Sci 3: 371±380
Allen LG, Horn MHK, Edmonds FA, Usui CA (1983) Structureand seasonal dynamics of ®sh assemblage in the Cabrillo Beacharea of Los Angeles Harbor, California. Bull Sth Calif Acad Sci82: 47±70
Bechtel TJ, Copeland BJ (1970) Fish species diversity indices asindicators of pollution in Galveston Bay, Texas. Contrib marSci Univ Tex 15: 103±132
Beckley LE (1984) The ichtyofauna of the Sundays Estuary, SouthAfrica, with particular reference to the juvenile marine com-ponent. Estuaries 3: 248±258
Blaber SJM, Blaber TG (1980) Factors a�ecting the distribution ofjuvenile estuarine and inshore ®sh. J Fish Biol 17: 143±162
Blaber SJM, Milton DA (1990) Species composition, communitystructure and zoogeography of ®shes of mangrove estuaries inthe Solomon Islands. Mar Biol 105: 259±267
Bortone SA, van Tassel J, Brito A, Falco n JM, Bundrick CM(1991) A visual assessment of the inshore ®shes and ®sheryresources o� E1 Hierro, Canary Islands: a baseline survey.Scientia mar 3: 529±541
Carr MH (1991) Habitat selection and recruitment of an assem-blage of temperate zone reef ®shes. J exp mar Biol Ecol 146:113±137
Connell SD, Jones GP (1991) The in¯uence of habitat complexityon postrecruitment processes in a temperate reef ®sh popula-tion. J exp mar Biol Ecol 151: 271±294
Doherty PJ (1991) Spatial and temperature patterns in recruitment.In: Sale PF (ed) The ecology of ®shes on coral reefs. AcademicPress, London, pp 261±292
Doherty PJ, Sale PF (1985) Predation on juvenile coral reef ®shes:an exclusion experiment. Coral Reefs 4: 225±234
DulcÏ ic J (1993a) Stock assessment of the Adriatic anchovy (En-graulis encrasicolus L.) using egg surveys. Acta Ichthyol Pisca 1:69±76
DulcÏ ic J (1993b) IstrazÏ ivanje sastava i bronjnosti ihtioplanktona usrednjem Jadranu. Morsko ribarstvo 3: 73±79
DulcÏ ic J (1995) Estimation of age and growth of sardine, Sardinapilchardus (Walbaum, 1792), larvae by reading daily otolithincrements. Fish Res 22: 265±277
DulcÏ ic J, Cetinic P (1993) Estimate of spawning biomass of an-chovy, Engraulis encrasicolus, in the eastern part of Adriaticfrom 1989 to 1990 by means of egg surveys. Acta Ichthyol Pisca2: 67±76
DulcÏ ic J, Kraljevic M (1995) Age, growth and mortality of dam-sel®sh (Chromis chromis L.) in the eastern middle Adriatic. FishRes 22: 255±264
Falco n JM, Bortone SA, Brito A, Bundrick CM (1996) Structure ofand relationships within and between the littoral, rock-substrate®sh communities o� four islands in the Canarian Archipelago.Mar Biol 125: 215±231
Forrester GE (1990) Factors in¯uencing the juvenile demographyof a coral reef ®sh population. Ecology 71: 1666±1681
Froglia C (1977) Feeding of Lithognathus mormyrus (L.) in centralAdriatic Sea (Pisces: Sparidae). Rapp P-v Re un Commn intExplor scient Mer Me diterr 24: 95±97
Fulgosi A (1988) Faktorska analiza. SÏ kolska knjiga, ZagrebGarcia-Rubies A (1995) Poblacions de peixos litorals sobre substrat
rocos. Factors que in¯uexen la seva distribucio. Ph.D. Thesis,University of Barcelona, Barcelona
Garcia-Rubies A, Macpherson E (1995) Substrate use and tem-poral pattern of recruitment in juvenile ®shes of the Mediter-ranean littoral. Mar Biol 124: 35±42
GrubisÏ ic F (1974) Znakovi osiromasÏ enja Jadrana-posljedica pre-intenzivnog ribolova. Acta adriat 5: 97±117
GrubisÏ ic F (1982) Ribe, rakovi i sÏ koljke Jadrana. ITRO Naprijed,Zagreb and GRO Liburnija, Rijeka
Haedrich RL, Haedrich SO (1974) A seasonal survey of the ®shesin the Mystic River, a polluted estuary in downtown Boston,Massachusetts. Estuar cstl mar Sci 2: 59±73
Jardas I (1980a) Fishery resources and Yugoslav ®sheries in thecoastal area of the Adriatic Sea. FAO Fish Rep 239: 53±59
Jardas I (1980b) Obalna naselja riba i njihova eksploatacija. Mo-rsko ribarstvo 4: 153±157
Jardas I (1982) Utjecaj intenzivnog ribolova na naselja riba obal-nog istocÏ nog Jadrana. Ichthyologia 1: 21±39
Jardas I (1985) The feeding of juvenile striped sea bream, Lit-hognathus mormyrus (L., 1758) (Pisces: Sparidae). Rapp P-vRe un Commn int Explor scient Mer Me diterr 29: 107±108
Jardas I (1986) Trammel bottom set catches along the easternAdriatic coast (1971±1984). FAO Fish Rep 345: 189±199
Jardas I (1996) Jadranska ihtiofauna. SÏ kolska knjiga, ZagrebJardas I, Antolic B, Skaramuca B, Bender A, Jug-Dujakovic J
(1986) Preliminary report of feeding of juvenile annular bream,Diplodus annularis (L., 1758) (Pisces: Sparidae). Rapp P-v Re unCommn int Explor scient Mer Me diterr 30: p 229
Jardas I, Pallaoro A (1989) Neki pokazatelji opadanja biolosÏ kogbogatstva priobalnog podrucÏ ja Jadrana (1960±1988). Pogledi 4:159±176
Jones GP (1986) Food availability a�ects growth in a coral reef®sh. Oecologia 70: 136±139
Jones GP (1987) Some interactions between residents and recruitsin two coral reef ®shes. J exp mar Biol Ecol 114: 169±182
Jug-Dujakovic J (1988) Prilog poznavanju ekologije mladi go-spodarski interesantnih vrsta riba u sÏ ibenskom priobalju. M.Sc.Thesis, University of Zagreb, Zagreb (mimeo)
Katavic I (1980) Temporal distribution of young mugilids (Mug-ilidae) in the coastal waters of the central eastern Adriatic. Actaadriat 2: 137±150
Katavic I (1984) Inducirano mrijesÏ c enje i uzgoj ranih razvojnihstadija lubina Dicentrarchus labrax (Linnaeus, 1758) i komarcÏ e,Sparus aurata (Linnaeus, 1758). Ph.D. Thesis, University Za-greb, Zagreb
Kraljevic M, DulcÏ ic J (1996) Age, growth and mortality of thegolden grey mullet Liza aurata (Risso, 1810) in the easternAdriatic. Arch Fish mar Res 44: 69±80
Kraljevic M, DulcÏ ic J, Cetinic P, Pallaoro A (1996) Age, growthand mortality of the striped sea bream, Lithognathus mormyrusL., in the northern Adriatic. Fish Res 28: 361±370
276

Kraljevic M, DulcÏ ic J, Pallaoro A, Cetinic P, Jug-Dujakovic J(1995) Sexual maturation, age and growth of striped sea bream,Lithognathus mormyrus L., on the eastern coast of the AdriaticSea. J appl Ichthyol 11: 1±8
Kraljevic M, Jug-Dujakovic J (1987) Preliminary ecological andbiological studies of juvenile ®sh species of commercial interestin the National Park ``Kornati''. FAO Fish Rep 394: 291±299
Kraljevic M, Pallaoro A (1991) Ihtiocenoze plitkih uvala nacio-nalnog parka ``Kornati''. Morsko ribarstvo 3: 81±90
Laegdsgaard P, Johnson CR (1995) Mangrove habitats as nurser-ies: unique assemblages of juvenile ®sh in subtropical man-groves in eastern Australia. Mar Ecol Prog Ser 126: 67±81
Lenanton RCJ (1982) Alternative non-estuarine nursery habitatsfor some commercially and recreationally important ®sh speciesof south-western Australia. Aust J mar Freshwat Res 33: 881±900
Levin PS (1991) E�ects of microhabitat on recruitment variation ina Gulf of Marine reef ®sh. Mar Ecol Prog Ser 75: 183±189
Macpherson E (1994) Substrate utilization in a Mediterranean lit-toral ®sh community. Mar Ecol Prog Ser 114: 211±218
Margalef R (1968) Perspectives in ecology theory. University ofChicago Press, Chicago
McErlean AJ, O'Connor SG, Mihursky JA, Gibson CI (1973)Abundance, diversity and seasonal patterns of estuarine ®shpopulations. Estuar cstl mar Sci 1: 19±36
MihelcÏ ic V (1997) National Park ``Kornati'' ± Croatia. Europarc,Federation of Nature and National Parks of Europe, Grafenau,Germany
Morovic D (1960) Mugilids of the Adriatic with a special review ofMugil cephalus, L. and Mugil chelo Cuv. Ph.D. Thesis, Uni-versity of Zagreb, Zagreb (mimeo)
Morovic D (1962) Prilog poznavanju lova migavicom u podrucÏ juDugog otoka 1953±1957. BiljesÏ ke Instituta za oceanogra®ju iribarstvo, Split
Morovic D (1965) Le probleme de la peche cotiere d'apres uneanalyse des captures au tramail. Proc gen ®sh Coun Mediterr40: 351±360
Nash RDM, Gibson RN (1982) Seasonal ¯uctuations and com-positions of two populations of small demersal ®shes on thewest coast of Scotland. Estuar cstl Shelf Sci 15: 485±495
Overland JE, Preisendorfer RW (1982) A signi®cance test forprincipal component applied to a cyclone climatology. MonWeath Rev 101: 1±4
Pielou EC (1966) Shannon's formula as measure of species diver-sity: its use and misuse. Am Nat 100: 463±465
Preisendorfer RV (1982) Principal component analysis in meter-orology and oceanography. Elsevier, Amsterdam
Reina-Hervas JA, Serrano P (1987) Structural and seasonal vari-ations of inshore ®sh populations in Ma laga Bay, southeasternSpain. Mar Biol 95: 501±508
Richman MB (1986) Rotation of principal component. J Climatol3: 293±335
Robertson AI, Duke NC (1987) Mangroves as nursery sites:comparisons of the abundance and species composition of ®shand crustaceans in mangroves and other nearshore habitats intropical Australia. Mar Biol 96: 193±205
Shannon CE, Weaver C (1949) The mathematical theory of com-munication. University of Illinois Press, Urbana
SÏ oljan T (1975) I pesci dell'Adriatico. O�cine gra®ce di Verona,Verona
Tzeng WN, Wang YT (1992) Structure, composition and seasonaldynamics of the larval and juvenile ®sh community in themangrove estuary of Tanshui River, Taiwan. Mar Biol 113:481±490
van den Broek WLF (1979) Seasonal survey of ®sh populations inthe lower Medway Estuary, Kent, based on power stationscreen samples. Estuar cstl mar Sci 9: 1±15
Victor BC (1986) Larval settlement and juvenile mortality in arecruitment-limited coral reef ®sh population. Ecol Monogr 56:145±160
Whit®eld AK (1994) Abundance of larval and 0+ juvenile marine®shes in the lower reaches of three southern African estuarieswith di�ering freshwater inputs. Mar Ecol Prog Ser 105: 257±267
Zore-Armanda M, Morovic M, Stojanoski L, Vukadin I (1987) Dali je u toku eutro®kacija otvorenog Jadranskog mora? Pomorskizbornik 1: 627±634
277
Copyright © 2022 FDOKUMEN




















