
Use of Stingless Bee Propolis and Geopropolis against ... - MDPI

G ModelA
Cpdo
AAa
Ab
c
d
e
a
ARRAA
KPMRMBF
ogm
0
ARTICLE IN PRESSNIFEE-13189; No. of Pages 10
Animal Feed Science and Technology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Animal Feed Science and Technology
journal homepage: www.elsevier.com/locate/anifeedsci
omparison of the in vitro efficiency of supplementary beeropolis extracts of different origin in enhancing the ruminalegradability of organic matter and mitigating the formationf methane
.S. Morsya, Y.A. Soltanb,∗, S.M.A. Sallamb, M. Kreuzerc, S.M. Alencard,
.L. Abdallae
Livestock Research Department, Arid Land Cultivation Research Institute, City of Scientific Research and Technological Applications,lexandria, EgyptAnimal Production Department, Faculty of Agriculture, Alexandria University, Alexandria, EgyptETH Zurich, Institute of Agricultural Sciences, Zurich, SwitzerlandDepartamento de Zootecnia, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, São Paulo, BrazilLaboratório de Nutric ão Animal, Centro de Energia Nuclear na Agricultura, Universidade de São Paulo, Piracicaba, São Paulo, Brazil
r t i c l e i n f o
rticle history:eceived 9 November 2013eceived in revised form 30 October 2014ccepted 1 November 2014vailable online xxx
eywords:ropolisonensin
uminal fermentationethane
atch cultureeed additive
a b s t r a c t
The current study was conducted to compare the effects of ethanolic extracts of Brazilianred propolis (BRP) and Egyptian brown propolis (EBP) characterized by different individ-ual compounds on the ruminal degradation of nutrients and methanogenesis. The utilityof propolis as a natural alternative to the ionophore antibiotic monensin, which was usedas a positive control, was also tested. The propolis extract was incubated with 0 (neg-ative control), 125, 250, or 500 �g per 500 mg of dietary dry matter. A semi-automaticin vitro gas production (GP) system was operated with three batches of rumen fluid inthree replicates per batch for each treatment. The major compounds of the BRP extractwere isoflavonoids and chalcones, which represented 0.43 of the total. Isoflavonoids andsaturated fatty acids made up 0.53 of the EBP extract. Extract of BRP25 �g enhanced the trueorganic matter degradability (TDOM) (P<0.05) compared with negative control. All doses ofBRP and EBP25 �g enhanced (P<0.05) TDOM compared with monensin while no differenceswere observed among the propolis extract treatments. Compared with the negative control,the treatment suppressing (P<0.05) methane (CH4) and total GP was monensin. The propo-lis extracts were only half or less than half as effective as monensin against CH4 production.Both the BRP and EBP extracts increased (P<0.05) the production of total and all major(acetate, propionate, butyrate) short-chain fatty acids (SCFA). Monensin enhanced (P<0.05)
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
only propionate production, clearly reduced (P<0.05) the acetate-to-propionate ratio com-pared with the negative control. Ruminal ammonia remained unaffected by the treatments,but the propolis extracts reduced (P<0.05) the protozoal count. This study demonstrated theefficiency of two types of propolis extracts, although they were less effective than monensin
Abbreviations: ADFom, ash-free acid detergent fiber; aNDFom, ash-free neutral detergent fiber treated with heat-stable �-amylase expressed exclusivef residual ash; BRP, Brazilian red propolis; CH4, methane; CP, crude protein; DM, dry matter; EE, ether extract; EBP, Egyptian brown propolis; GC–MS,as chromatography–mass spectrometry; GP, gas production; lignin(sa), lignin determined by solubilization of cellulose with sulfuric acid; OM, organicatter; PF, partitioning factor; SCFA, short-chain fatty acids; TDOM, truly degraded OM.∗ Corresponding author. Tel.: +20 1093589902; fax: +20 3 5922780.
E-mail address: uosra [email protected] (Y.A. Soltan).
http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004377-8401/© 2014 Elsevier B.V. All rights reserved.

G Model
ARTICLE IN PRESSANIFEE-13189; No. of Pages 102 A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx
in mitigating CH4 emissions. It also showed that propolis with very different composi-tions seemed to act in a similar way and that the efficiency of the two types of propolisin modifying the ruminal degradation of nutrients and mitigating methane formation wascomparable. The most active component(s) in propolis still have to be identified.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Ionophores are an especially important group of antibiotic growth promoters, which are widely used in ruminant hus-bandry as feed additives to improve energy and protein utilization and decrease methane (CH4) emissions from ruminants(Russell and Strobel, 1988; Wischer et al., 2013). However, there is a controversy about the use of such additives containingantibiotics in animal diets because of the risk of transferring residues into meat and milk and promoting the development ofresistant strains of bacteria. These concerns have promoted the search for alternative natural feed additives, such as propolis(Oeztuerk and Sagmanligil, 2009; Morsy et al., 2013).
Propolis is a mixture of a plant product and secretions by honeybees (Apis mellifera). Bees collect a resinous mixture frombuds of deciduous trees and from crevices in the bark of coniferous and deciduous trees. Beeswax and the secretions fromthe bees’ mandibular and hypopharyngeal glands are added to this mixture. Propolis is used by the bees to cover hive walls,fill cracks, and embalm invading insects that they are killed (Zhou et al., 2008). Propolis is known to be active against (grampositive) bacteria, viruses, fungi, oxidants, inflammation, tumors, and parasites, and it may act as an immunomodulator(Alencar et al., 2007; de Aguiar et al., 2013; Morsy et al., 2013). Propolis could also be of use as a natural feed alternative toionophores in ruminant diets, but the effects reported have been variable. For instance, Zawadzki et al. (2011) concludedthat brown propolis could be used as a substitute for monensin in diet supplementation, as it improved feed efficiency andbody weight gain in feedlot-finished bulls. Ítavo et al. (2011) reported similar findings in growth performance in a study onfeedlot lambs fed a diet supplemented with brown propolis, but they found that green propolis was not efficient.
The administration of Brazilian red propolis was beneficial for ewe’s health during the flushing period (Morsy et al., 2013)and Egyptian (brown) propolis improved semen quality (Hashem et al., 2013). In vitro, propolis was effective in decreasingthe formation of ammonia in the rumen (Oeztuerk et al., 2010) and inhibited hyperammonia-producing species of bacteria(de Aguiar et al., 2013). Broudiscou et al. (2000, 2002) reported that propolis seemed to have rather weak effects, determinedin vitro, on ruminal fermentation, other than the selective enhancement of propionate, and that it appeared to have no effecton methanogenesis (Broudiscou et al., 2000).
The biological activity of propolis may vary, together with its chemical composition, as it comprises numerous con-stituents. The chemical composition of propolis is intrinsically related to the floristic and ecological characteristics of theenvironment visited by the bees (Alencar et al., 2007). The method of propolis extraction may also affect its biological activ-ity. Due to the sticky nature of propolis, it cannot be used in its crude form and is therefore often purified by extractionwith solvents, which are intended to preserve the polyphenolic fractions (de Aguiar et al., 2013). Thus, the type and con-centration of the solvent represent another source of variation in the biological activity of the propolis extracts. Propolisexists in different colors (red, brown, yellow, and green), with the same colored samples from different countries apparentlyshowing a greater resemblance to each other than different colored samples from the one country (Alencar et al., 2007). Theaforementioned suggests that the influence of propolis extracts on ruminal fermentation may differ according to propolisorigin and extraction method. However, other than the study by Ítavo et al. (2011), the efficiency of different sources ofpropolis extracts in modifying ruminal fermentation has not been investigated. In most studies, the propolis material thatwas used originated exclusively from Brazil. Often, the color of the propolis is not even specified in publications.
In the present study, the following hypotheses were tested: (i) bee propolis can be used as an effective natural dietaryadditive in ruminants’ diets. (ii) The effect of bee propolis on ruminal fermentation, degradability, and CH4 production notonly depends on the amount of propolis added to the diet but also on the color of the propolis and the country of origin.These hypotheses were tested with in vitro batch cultures using two contrasting types of propolis collected from two differentcountries (Brazil and Egypt). To exclude any other source of variation, the crude propolis was extracted in the same way.The present study investigated the ruminal degradation of nutrients and the potential antimethanogenic activity of propolis,areas that have still been the focus of only few studies and where monensin is considered to be effective.
2. Material and methods
2.1. Origin, processing, and analysis of the batches of experimental propolis
Raw Brazilian red propolis (BRP) was collected from July 2010 to January 2011 from the mangrove region in Marechal
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
Deodoro, a city in the vicinity of Maceio, capital of Alagoas State, in Northeastern Brazil (9◦42′36′′S, 35◦53′42′′W), an areawith a wet tropical climate. Raw Egyptian brown propolis (EBP) was collected from October 2010 to March 2011 from anexperimental apiary in Alexandria Governorate, Northwestern Egypt (31◦11′53′′N, 29◦55′9′′E), a region characterized by adry subtropical climate. Both propolis types were collected manually from colonies of A. mellifera bees. Six replicates each

G ModelA
oe
tbpTad
oBc2mf(
2
pE
2
Pkn5sVsrB
2
wosc2
utAhicgo
2
mmfaru
ARTICLE IN PRESSNIFEE-13189; No. of Pages 10
A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx 3
f the two raw propolis types were cleaned, weighed, bottled in 10 g portions and stored at −18 ◦C until being subjected toxtraction and analysis (Morsy et al., 2013).
The same extraction method was applied to both raw propolis types. Ten grams each were treated with liquid nitrogen andhen ground to a fine powder and mixed with 100 ml of ethanol (700 ml/l). The mixture was then ultrasonically (ultrasonicath, Pulsatron KC3, Kerry, Guyson, UK) treated for 30 min. The ethanolic solution was subsequently filtered through a filteraper (Whatman no. 41). The extract was kept at −5 ◦C overnight and was filtered again to remove any remaining impurities.he supernatant was transferred to a rotary evaporator (RE301/601/801, Yamato Scientific America Inc., USA) and treatedt 42 ◦C for 30 min to remove the ethanol. The concentrated extract recovered in the volumetric flask was lyophilized for 3ays to obtain pure propolis extracts in a powdery form.
All extracts (six replicates of each type) were subjected separately to an in-depth compositional analysis at the Departmentf Agri-Food industry, Food and Nutrition, “Luiz de Queiroz” College of Agriculture, University of Sao Paulo (USP), Piracicaba,razil, after being methylated as described by Markham et al. (1996). The compositional analysis was performed with gashromatography–mass spectrometry (GC–MS) according to the method of Fernández et al. (2008). A Shimadzu Model GC-010 Series gas chromatograph, coupled with a Shimadzu series mass-selective detector quadrupole mass spectrometerodel GCMS-QP 2010 plus, was used (Shimadzu, Kyoto, Japan). The GC–MS peaks were identified by comparison with data
rom the literature and profiles from the libraries of Wiley 138 and the National Institute of Standards and TechnologyAlencar et al., 2007).
.2. In vitro experiment
The in vitro experiment was carried out at the Centre for Nuclear Energy in Agriculture (CENA, USP) Piracicaba, Brazil. Therotocol, which included rumen fluid sampling, was approved by the Internal Commission for Environmental and Ethics inxperimentation with Animals of CENA/USP (approval no. CIEEA/CENA 001/2011).
.2.1. Basal dietThe basal diet consisted, per kg of dry matter (DM), of 500 g of hay prepared from Bermuda grass (Cynodon dactylon (L.)
ers.; hybrid “Tifton-85”), 327 g of ground maize, 150 g of soybean meal, 10 g of limestone, and 13 g of mineral premix. Perg of DM, analyzed according to AOAC (1995) (ID number 930.15), this mixture contained 923 g of organic matter (OM; IDumber 942.05), 151 g of crude protein (CP; as 6.25 × N; ID number 954.01), 48 g of ether extract (EE; ID number 920.39),48 g of neutral detergent fiber (analyzed according to Van Soest et al. (1991), exclusive of residual ash and using heat-table �-amylase and sodium sulfite [aNDFom]), 256 g of acid detergent fiber (ADFom; determined following Goering andan Soest (1970), exclusive of residual ash), and 25.6 g of lignin (lignin(sa); determined by solubilization of cellulose withulfuric acid (720 ml/l) according to the method of Van Soest (1973)). This diet was formulated to meet or exceed NRC (2007)ecommendations for energy and protein for growing cattle. The diet was ground in a Wiley mill (Marconi, Piracicaba SP,razil) to pass a 1 mm screen.
.2.2. Experimental designStock solutions of monensin (M5273; Sigma–Aldrich Co., St. Louis, MO, USA; MW, 692.85) and the BRP and EBP extracts
ere prepared by diluting 15.6 mg of monensin and 12.5, 25, and 50 mg of the BRP or EBP extracts (the pooled of remaindersf the same 10 g portions of BRP or EBP that were analyzed in six replicates) in 1 ml of pure ethanol. These solutions weretored at −10 ◦C before supplementation. At zero time of the incubation, 10 �l of the stock solution were added to eachorresponding bottle to achieve, per 500 mg of basal diet DM, final concentrations of 156 �g of monensin, as well as 125,50, and 500 �g of either extracts of BRP or EBP.
A non-supplemented dietary treatment (negative control) completed the study, giving a total of eight treatments. Fornification of all the treatments, 10 �l of ethanol were also added to the blanks (without any other supplementation) ando the negative control. With this set-up, the effects could be related to the treatments, exclusive of the ethanol addition.ccording to Araujo et al. (2011) and Selje-Assmann et al. (2008), this amount of ethanol in 75 ml of buffered rumen fluidas no effect on the fermentation process. The ionophore monensin was selected for the positive control treatment because
t is one of the most common additives used to modulate ruminal fermentation and is therefore used very often as a positiveontrol in in vitro studies (e.g., Zawadzki et al., 2011). The dosage tested was previously found to decrease the production ofas, CH4, and acetate and to increase the synthesis of propionate while exerting minimal effects on the ruminal degradationf OM (Araujo et al., 2011).
.2.3. Experimental protocolSix adult rumen cannulated Santa Inês sheep (60 ± 2.3 kg body weight) grazing on tropical grass pasture and supple-
ented with ground maize and soybean meal (0.7 kg/100 kg live weight, 156 g crude protein/kg) with free access to aineral premix and fresh water were used as inoculum donors. Ruminal liquid and solid fractions were collected separately
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
rom each animal before morning feeding. A liquid fraction was obtained by using a stainless steel probe (2.5 mm screen)ttached to a syringe with a large capacity. The solid phase was collected through the cannula by hand from the dorsalumen sac and squeezed. The liquid and solid fractions were placed separately into prewarmed thermo containers (39 ◦C)nder anaerobic conditions and transported immediately to the laboratory. The two fractions were blended at 1:1 (v/v)

G Model
ARTICLE IN PRESSANIFEE-13189; No. of Pages 104 A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx
during 10 s, squeezed through three layers of cheesecloth, and maintained in a water bath (39 ◦C) under CO2 until startingthe inoculation (Bueno et al., 2005).
Each treatment was incubated in three inocula, each prepared from two animals. This was necessary due to the largeamount of inoculum needed and ensured that the natural variation among animals was maintained and, at the same time,the effects of unusual rumen inocula or rumen environments were reduced. When producing the experimental inocula, theruminal solid and liquid phases from each of the two animals were combined in equal portions. With each inoculum, sixbottles per treatment were prepared: three for determining truly degraded organic matter (TDOM) and the other three forassessing other fermentation traits. This resulted in a total of 18 bottles per treatment. The same procedure was applied tothe blanks (bottles without substrate but containing inoculum and medium) to correct the gas production (GP) from theinoculum and to an internal standard (Tifton-85 Cynodon sp. hay) to enable adjustments among the inocula. In total, thisresulted in 180 bottles. Biased incubation inocula (GP variations between the internal standards >0.1 from the average ofthe other inocula) were rejected and repeated again. In cases where the inocula results were within the accepted range, theinternal standard data were used to correct sensitivity changes induced by the inoculum. This was also done to avoid theeffects of unusual rumen inocula in the ruminal environment (Soltan et al., 2012).
The in vitro GP assay described by Theodorou et al. (1994) was adapted to a semi-automatic system (Bueno et al., 2005),equipped with a pressure transducer and a data logger (Pressure Press Data 800, LANA, CENA/USP, Piracicaba, Brazil). Thedried ground substrate diet (500 mg) was placed in 160-ml glass bottles and then incubated with 50 ml of incubation medium(Menke’s buffered medium) and 25 ml of the ruminal inoculum. The bottles were sealed immediately with 20 mm butyl sep-tum stoppers (Bellco Glass Inc, Vineland, NJ, USA), manually shaken, and incubated at 39 ◦C in a forced air oven (MarconiMA35, Piracicaba, SP, Brazil) for 24 h. When measuring the gas pressure and sampling gas for CH4 determination, 18 bottleswere put on each tray. All subsequently described procedures were accomplished in a laminar air flow chamber situateddirectly besides the oven, and all 180 bottles were treated within 10 min. These steps, together with the rather high temper-ature in the room, kept the bias low (e.g., bias caused by the gas composition of the headspace changing the carbon dioxidesolubility of the liquid due to cooling down of the samples). However, bias could not be totally excluded. The head space gaspressure (85 ml) was measured after 4, 8, 12, and 24 h. The gas volume produced (ml) was calculated as 7.365 × measuredpressure (psi) (Soltan et al., 2012). The net cumulative GP was obtained during 24 h of incubation.
For the CH4 analysis, 2.5 ml of gas was sampled at each measuring event using a 5 ml syringe (Becton Dickson IndústriaCirúrgica LTDA, Curitiba, PR, Brazil) and stored in a 10 ml vacuum tube. After each gas sampling, the bottles were vented torelease the gas pressure, shaken, and returned to the oven. The CH4 concentration was then determined on a gas chromato-graph (Model 2014, Shimadzu, Tokyo, Japan) equipped with a Shincarbon ST 100/120 micropacked column (1.5875 mm OD,1.0 mm ID, 1 m length; Ref. no 19809; Restek, Bellefonte, PA, USA). An external analytical calibration curve (0, 30, 60, 90,and 120 ml/l) was prepared with pure CH4 (White Martins PRAXAIR Gases industrial Inc., Osasco, SP, Brazil; 995 ml/l purity).The temperatures of the column, injector, and flame ionization detector were 60, 200, and 240 ◦C, respectively. Helium wasadded at 10 ml/min and used as a carrier gas. The amounts of CH4 produced were calculated according to the method ofLongo et al. (2006) as:
CH4 produced (ml) = (GP (ml) + headspace, 85 ml) × CH4 concentration (ml/ml). The net GP and CH4 were determined bycorrecting for the corresponding blank values.
All the bottles were placed on ice to terminate the incubation after 24 h. The TDOM was determined according to themethod of Blümmel and Becker (1997) by adding 70 ml of neutral detergent solution without heat-stable �-amylase toeach flask, followed by incubation at 105 ◦C for 3 h. The contents of the bottles were filtered in preweighed crucibles,washed with hot water and then acetone, and the residual DM and ash were determined. The partitioning factor (PF),an indicator of microbial efficiency, was calculated as the ratio of TDOM (mg) and gas volume (ml in 24 h) (Blümmelet al., 1997). The contents of the other three bottles were used for determining further fermentation variables. In theincubation liquid, the pH was measured using an epoxy body pH electrode (WD- 35801-00, Oakton) connected to a pHmeter (Ion 6, Acorn series, Oakton, USA). The net release values of NH3–N were measured according to the method ofPreston (1995) using the micro-Kjeldahl method. Short-chain fatty acids (SCFA) were determined according to the methodof Palmiquist and Conrad (1971) using a gas chromatograph (GC HP 5890 Series II/integrator HP 3396 Series II/automaticinjector HP 6890 Series, Agilent Technologies, Palo Alto, CA, USA). Butyric acid (MW = 116.16; Sigma Chemie GmbH, Stein-heim, Germany) was used as an internal standard, and a mixture of SCFA of known concentrations was used as theexternal standard for calibration of the integrator. For counting of ruminal protozoa by a microscope following the pro-cedure described by Dehority et al. (1983), 2 ml of rumen fluid were mixed with 2 ml of methyl green-formalin salinesolution.
2.3. Statistical analyses
The in vitro gas production was completed in one run for all eight treatments, using three inocula (true statistical repli-
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
cates), with each sample incubated in triplicate (analytical replicates) to obtain the most accurate estimate of a true replicate.The analytical replicates were averaged prior to statistical analysis with each inoculum being the statistical replicate, thusthe statistical number of replications of treatments (n = 3) are the true replications. The experimental unit was thereforethe mean of the three analytical replicates from one statistical replicate. The data were subjected to analysis of variance

ARTICLE IN PRESSG ModelANIFEE-13189; No. of Pages 10
A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx 5
Table 1Identity of constituents determined by gas chromatography/mass spectrometry in the ethanolic extracts of Brazilian red propolis ordered by proportionof total.
Peak Compounds tR (min)a Proportion of total area
1 Vestitol 38.37 0.24622 Benzenepropanoic acid, 3,5-bis[(trimethylsilyl)oxy]-, trimethylsilyl ester 38.24 0.16493 Medicarpin 37.55 0.13524 4,4′-Bis[(trimethylsilyl)ethynyl]-2,2′-bithiophene-5,5′-dicarbaldehyde 39.07 0.07075 Silane, trimethyl[5-methyl-2-(1-methylethyl)phenoxy]a 37.41 0.04976 Benzenepropanoic acid, 3,4-bis[(trimethylsilyl)oxy]-, trimethylsilyl estera 38.11 0.03937 Silane, trimethyl[1-[(trimethylsilyl)ethynyl]-2-naphthalenyl]a 44.91 0.03308 Silane, (9,19-cyclo-9.beta.-lanost-24-en-3.beta.-yloxy)trimethyla 48.11 0.03099 9h-Fluorene-4,5-diamine, N,N,N′ ,N′-tetramethyla 40.51 0.028810 Isoliquiritigenin 41.74 0.026411 Lup-20(29)-en-3-yl acetate 49.26 0.025412 Beta-amirin trimethylsilyl ether 47.57 0.023313 Hydrocinnamic acid, P-(trimethylsiloxy)-, trimethylsilyl estera 39.43 0.019614 Formononetin 40.86 0.018415 2,5-Tri-OH-phenylacetate 4tms 41.44 0.017716 3,4-Dihydroxy-9-methoxypterocarpan 39.68 0.015117 9,19-cyclolanostan-3-ol, 24-methylene-, (3.beta.)a 48.78 0.012118 4,4′ Bis[(trimethylsilyl)ethynyl]-2,2′-bithiophene-5,5′ dicarbaldehyde 36.49 0.008019 3,8-Dihydroxy-9-methoxypterocarpan (3-hydroxy-8,9-dimethoxypterocarpan) 40.13 0.007920 Silane, 9h-fluoren-9-ylidenebis(trimethyl)a 41.17 0.007821 Propanedioic acid, bis[(trimethylsilyl)oxy]-, bis(trimethylsilyl) estera 45.60 0.007822 2-Propenoic acid, 3-(3,4,5-trimethoxyphenyl)-, methyl estera 42.02 0.007023 Solanesol 49.88 0.0048
t
N
uc
3
3
empi3sh
3
tctco
(Hunbc
w
R (min)a = retention time, min.a Components were similarity more than 50% of the search results by comparison with data from literature and the profiles from the Wiley 138 andational Institute of Standards and Technology’98 (NIST’98).
sing the general linear model procedure (PROC GLM) procedure of SAS (2002). Differences between the treatments wereonsidered significant at P<0.05 using Tukey’s procedure for multiple comparisons among means.
. Results
.1. Compounds present in the propolis extracts of different origin
A number of specific constituents were identified from the chromatograms of the ethanolic propolis extracts of thexperimental types of propolis. Most abundant according to the relative percentages of the chromatogram areas of theajor compounds of BRP extract identified were isoflavonoids and chalcones, which represented almost 0.43 of total
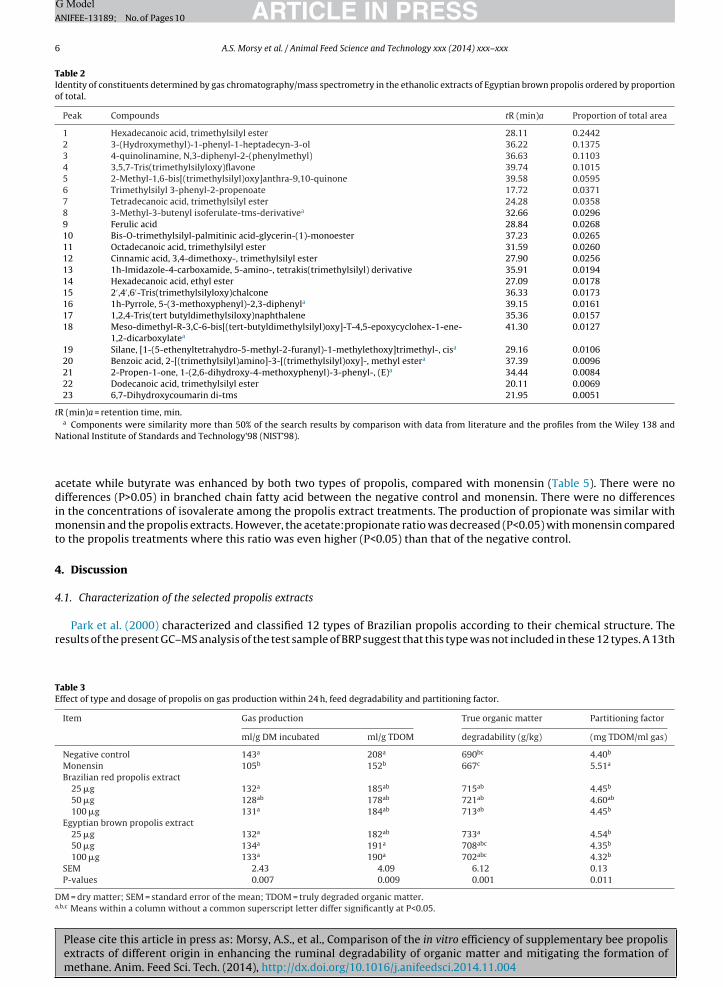
ropolis (Table 1). In detail, the proportions of total of the compounds identified were: vestitol (0.25), medicarpin (0.14),soliquiritigenin (0.025), and formononetin (0.018). Isoflavonoids were also the major components of the EBP extract, with-(hydroxymethyl)-1-phenyl-1-heptadecyn-3-ol, flavone and 4-quinolinamine, and n,3-diphenyl-2-(phenylmethyl) repre-enting 0.35 of the total (Table 2). Almost 0.25 of all the EBP compounds analyzed consisted of saturated fatty acids, especiallyexadecanoic acid and palmitic acid, which were not found in the BPR.
.2. In vitro effects of the propolis extracts on ruminal fermentation
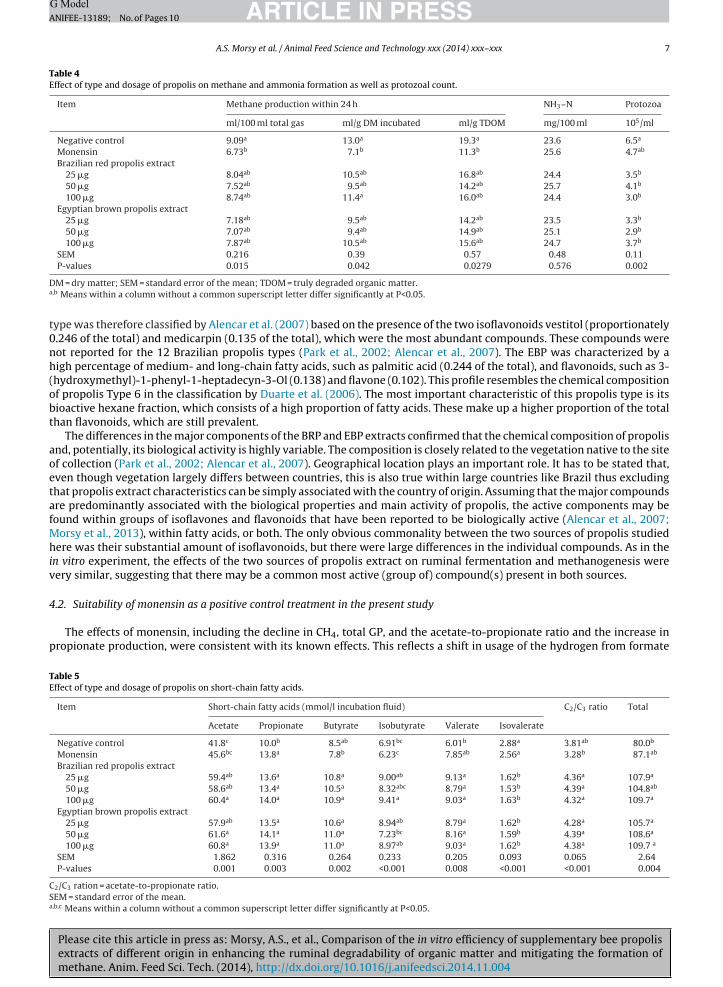
In the well-buffered batch system, the average pH was maintained at 6.70, without any significant differences betweenhe treatments. Both per unit of DM and TDOM, monensin resulted in the lower GP (P<0.05) compared with the negativeontrol (Table 3), whereas no differences were observed among the propolis treatments or the negative control. The EBP25 �greatment had the highest TDOM (P<0.05) compared with all other treatments, while all extracts of BRP enhanced the TDOMompared with the negative and positive control. The PF of the monensin was enhanced (P<0.05) compared to that of thether treatments.
A similar trend to that seen in GP was observed for CH4 given as ml/100 ml GP, as ml/g DM incubated, or as ml/g TDOMTable 4). Monensin inhibited (P<0.05) CH4 per unit of TDOM by 0.45 of the total compared with the negative control.owever, the effects of the treatments with the BRP50 �g and EBP25 �g extracts on CH4 production per unit of TDOM declined,p to 0.27 of the total compared with the negative control, but there were no significant differences observed even for theegative control or monensin. There were no significant differences in the NH3–N concentration of the incubation fluid
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
etween the treatments. Both types of propolis extracts decreased (P<0.05) the protozoal count compared with the negativeontrol, and the protozoal count was intermediate with monensin.
All the propolis treatments enhanced (P<0.05) the production of acetate, propionate, valerate and total SCFA comparedith the negative control. All EBP extracts and the highest dose of BRP (BRP100 �g) increased (P<0.05) the production of
ARTICLE IN PRESSG ModelANIFEE-13189; No. of Pages 10
6 A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx
Table 2Identity of constituents determined by gas chromatography/mass spectrometry in the ethanolic extracts of Egyptian brown propolis ordered by proportionof total.
Peak Compounds tR (min)a Proportion of total area
1 Hexadecanoic acid, trimethylsilyl ester 28.11 0.24422 3-(Hydroxymethyl)-1-phenyl-1-heptadecyn-3-ol 36.22 0.13753 4-quinolinamine, N,3-diphenyl-2-(phenylmethyl) 36.63 0.11034 3,5,7-Tris(trimethylsilyloxy)flavone 39.74 0.10155 2-Methyl-1,6-bis[(trimethylsilyl)oxy]anthra-9,10-quinone 39.58 0.05956 Trimethylsilyl 3-phenyl-2-propenoate 17.72 0.03717 Tetradecanoic acid, trimethylsilyl ester 24.28 0.03588 3-Methyl-3-butenyl isoferulate-tms-derivativea 32.66 0.02969 Ferulic acid 28.84 0.026810 Bis-O-trimethylsilyl-palmitinic acid-glycerin-(1)-monoester 37.23 0.026511 Octadecanoic acid, trimethylsilyl ester 31.59 0.026012 Cinnamic acid, 3,4-dimethoxy-, trimethylsilyl ester 27.90 0.025613 1h-Imidazole-4-carboxamide, 5-amino-, tetrakis(trimethylsilyl) derivative 35.91 0.019414 Hexadecanoic acid, ethyl ester 27.09 0.017815 2′ ,4′ ,6′-Tris(trimethylsilyloxy)chalcone 36.33 0.017316 1h-Pyrrole, 5-(3-methoxyphenyl)-2,3-diphenyla 39.15 0.016117 1,2,4-Tris(tert butyldimethylsiloxy)naphthalene 35.36 0.015718 Meso-dimethyl-R-3,C-6-bis[(tert-butyldimethylsilyl)oxy]-T-4,5-epoxycyclohex-1-ene-
1,2-dicarboxylatea41.30 0.0127
19 Silane, [1-(5-ethenyltetrahydro-5-methyl-2-furanyl)-1-methylethoxy]trimethyl-, cisa 29.16 0.010620 Benzoic acid, 2-[(trimethylsilyl)amino]-3-[(trimethylsilyl)oxy]-, methyl estera 37.39 0.009621 2-Propen-1-one, 1-(2,6-dihydroxy-4-methoxyphenyl)-3-phenyl-, (E)a 34.44 0.008422 Dodecanoic acid, trimethylsilyl ester 20.11 0.006923 6,7-Dihydroxycoumarin di-tms 21.95 0.0051
tR (min)a = retention time, min.a
Components were similarity more than 50% of the search results by comparison with data from literature and the profiles from the Wiley 138 andNational Institute of Standards and Technology’98 (NIST’98).
acetate while butyrate was enhanced by both two types of propolis, compared with monensin (Table 5). There were nodifferences (P>0.05) in branched chain fatty acid between the negative control and monensin. There were no differencesin the concentrations of isovalerate among the propolis extract treatments. The production of propionate was similar withmonensin and the propolis extracts. However, the acetate:propionate ratio was decreased (P<0.05) with monensin comparedto the propolis treatments where this ratio was even higher (P<0.05) than that of the negative control.
4. Discussion
4.1. Characterization of the selected propolis extracts
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
Park et al. (2000) characterized and classified 12 types of Brazilian propolis according to their chemical structure. Theresults of the present GC–MS analysis of the test sample of BRP suggest that this type was not included in these 12 types. A 13th
Table 3Effect of type and dosage of propolis on gas production within 24 h, feed degradability and partitioning factor.
Item Gas production True organic matter Partitioning factor
ml/g DM incubated ml/g TDOM degradability (g/kg) (mg TDOM/ml gas)
Negative control 143a 208a 690bc 4.40b
Monensin 105b 152b 667c 5.51a
Brazilian red propolis extract25 �g 132a 185ab 715ab 4.45b
50 �g 128ab 178ab 721ab 4.60ab
100 �g 131a 184ab 713ab 4.45b
Egyptian brown propolis extract25 �g 132a 182ab 733a 4.54b
50 �g 134a 191a 708abc 4.35b
100 �g 133a 190a 702abc 4.32b
SEM 2.43 4.09 6.12 0.13P-values 0.007 0.009 0.001 0.011
DM = dry matter; SEM = standard error of the mean; TDOM = truly degraded organic matter.a,b,c Means within a column without a common superscript letter differ significantly at P<0.05.
ARTICLE IN PRESSG ModelANIFEE-13189; No. of Pages 10
A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx 7
Table 4Effect of type and dosage of propolis on methane and ammonia formation as well as protozoal count.
Item Methane production within 24 h NH3–N Protozoa
ml/100 ml total gas ml/g DM incubated ml/g TDOM mg/100 ml 105/ml
Negative control 9.09a 13.0a 19.3a 23.6 6.5a
Monensin 6.73b 7.1b 11.3b 25.6 4.7ab
Brazilian red propolis extract25 �g 8.04ab 10.5ab 16.8ab 24.4 3.5b
50 �g 7.52ab 9.5ab 14.2ab 25.7 4.1b
100 �g 8.74ab 11.4a 16.0ab 24.4 3.0b
Egyptian brown propolis extract25 �g 7.18ab 9.5ab 14.2ab 23.5 3.3b
50 �g 7.07ab 9.4ab 14.9ab 25.1 2.9b
100 �g 7.87ab 10.5ab 15.6ab 24.7 3.7b
SEM 0.216 0.39 0.57 0.48 0.11
Da
t0nh(obt
aoetafMhiv
4
p
TE
CSa
P-values 0.015 0.042 0.0279 0.576 0.002
M = dry matter; SEM = standard error of the mean; TDOM = truly degraded organic matter.,b Means within a column without a common superscript letter differ significantly at P<0.05.
ype was therefore classified by Alencar et al. (2007) based on the presence of the two isoflavonoids vestitol (proportionately.246 of the total) and medicarpin (0.135 of the total), which were the most abundant compounds. These compounds wereot reported for the 12 Brazilian propolis types (Park et al., 2002; Alencar et al., 2007). The EBP was characterized by aigh percentage of medium- and long-chain fatty acids, such as palmitic acid (0.244 of the total), and flavonoids, such as 3-hydroxymethyl)-1-phenyl-1-heptadecyn-3-Ol (0.138) and flavone (0.102). This profile resembles the chemical compositionf propolis Type 6 in the classification by Duarte et al. (2006). The most important characteristic of this propolis type is itsioactive hexane fraction, which consists of a high proportion of fatty acids. These make up a higher proportion of the totalhan flavonoids, which are still prevalent.
The differences in the major components of the BRP and EBP extracts confirmed that the chemical composition of propolisnd, potentially, its biological activity is highly variable. The composition is closely related to the vegetation native to the sitef collection (Park et al., 2002; Alencar et al., 2007). Geographical location plays an important role. It has to be stated that,ven though vegetation largely differs between countries, this is also true within large countries like Brazil thus excludinghat propolis extract characteristics can be simply associated with the country of origin. Assuming that the major compoundsre predominantly associated with the biological properties and main activity of propolis, the active components may beound within groups of isoflavones and flavonoids that have been reported to be biologically active (Alencar et al., 2007;
orsy et al., 2013), within fatty acids, or both. The only obvious commonality between the two sources of propolis studiedere was their substantial amount of isoflavonoids, but there were large differences in the individual compounds. As in the
n vitro experiment, the effects of the two sources of propolis extract on ruminal fermentation and methanogenesis wereery similar, suggesting that there may be a common most active (group of) compound(s) present in both sources.
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
.2. Suitability of monensin as a positive control treatment in the present study
The effects of monensin, including the decline in CH4, total GP, and the acetate-to-propionate ratio and the increase inropionate production, were consistent with its known effects. This reflects a shift in usage of the hydrogen from formate
able 5ffect of type and dosage of propolis on short-chain fatty acids.
Item Short-chain fatty acids (mmol/l incubation fluid) C2/C3 ratio Total
Acetate Propionate Butyrate Isobutyrate Valerate Isovalerate
Negative control 41.8c 10.0b 8.5ab 6.91bc 6.01b 2.88a 3.81ab 80.0b
Monensin 45.6bc 13.8a 7.8b 6.23c 7.85ab 2.56a 3.28b 87.1ab
Brazilian red propolis extract25 �g 59.4ab 13.6a 10.8a 9.00ab 9.13a 1.62b 4.36a 107.9a
50 �g 58.6ab 13.4a 10.5a 8.32abc 8.79a 1.53b 4.39a 104.8ab
100 �g 60.4a 14.0a 10.9a 9.41a 9.03a 1.63b 4.32a 109.7a
Egyptian brown propolis extract25 �g 57.9ab 13.5a 10.6a 8.94ab 8.79a 1.62b 4.28a 105.7a
50 �g 61.6a 14.1a 11.0a 7.23bc 8.16a 1.59b 4.39a 108.6a
100 �g 60.8a 13.9a 11.0a 8.97ab 9.03a 1.62b 4.38a 109.7 a
SEM 1.862 0.316 0.264 0.233 0.205 0.093 0.065 2.64P-values 0.001 0.003 0.002 <0.001 0.008 <0.001 <0.001 0.004
2/C3 ration = acetate-to-propionate ratio.EM = standard error of the mean.,b,c Means within a column without a common superscript letter differ significantly at P<0.05.

G Model
ARTICLE IN PRESSANIFEE-13189; No. of Pages 108 A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx
and CH4 production to succinate or propionate production as alternate hydrogen-sink products in the mixed microbialpopulation (Russell and Strobel, 1988). The decrease in methanogenesis may be due to a shift in the bacterial populationfrom gram-positive to gram-negative organisms rather than a direct effect of the ionophores on methanogens. The effectsof monensin are due to its lipophilic properties, which cause a decrease in intracellular K+ and in intracellular pH and anincrease in intracellular Na+, which is detrimental to gram-positive bacteria. This group of ruminal bacteria produces moreammonia, hydrogen, and lactate than gram-negative bacteria (Russell and Strobel, 1988).
4.3. Effect of the different types of propolis extracts on the ruminal degradation of nutrients and methanogenesis
Although the effects of propolis were clearly less pronounced than those of monensin in the present study, propolis extractseemed to combat ruminal methanogenesis, thus likely adversely affecting Archaea. To the best of our knowledge, the onlystudy to describe the effects of propolis on CH4 (Broudiscou et al., 2000) did not indicate that it had an antimethanogeniceffect. In contrast to monensin, the propolis extracts also promoted the ruminal degradation of nutrients, as can be seenfrom the increase in the TDOM and SCFA and the decrease in the protozoal count. Among the flavonoids that were found inthe BRP extract, vestitol and chalcone (isoliquiritigenin) were described to be the most effective antioxidants in this noveltype of BPR and these compounds were found not to be synergistic in their activity for microbial control (Oldoni et al.,2011). The combination of flavonoids and saturated fatty acids found in EBP may also explain its effects. Zhou et al. (2013)demonstrated that certain fatty acids damage cells by increasing the cell membrane permeability and triggering K+ efflux.They also found that the cell membrane permeability and K+ efflux increased the most with saturated fatty acids that havea shorter chain length, such as lauric and myristic acid.
The decrease in the protozoal count caused by the addition of propolis in the present study is partially in agreement withthe study by Rispoli et al. (2009) who found the same in the rumen of buffaloes fed a diet consisting of maize silage andconcentrate (1:1) but not in cattle fed the same diet. An antiprotozoal effect would help to explain the reduction in CH4 emis-sions, as the inhibition of protozoa reduces the release of CH4, since protozoa are known to provide hydrogen as a substratefor the methanogens which are at least temporarily ecto- and endosymbiotically associated with them (Ushida and Jouany,1996). All levels of propolis extract enhanced concentrations of acetate, propionate, valerate and total SCFA. Concerning theseeffects, it seems that propolis caused a stimulation of rumen microorganisms responsible for other hydrogen-consumingpathways. At a closer look, the increase in total SCFA is mainly due to a massive increase in acetate production by 45%compared to control. Therefore, the change in total VFA might be attributed to the production of more acetate (the majorcontributor to VFA) at cost of CH4. This indicates that propolis, regardless of its type, has a different mode of action thanmonensin. However, it should be noted that Broudiscou et al. (2000) found that propolis selectively enhanced only themolar proportion of propionate. The large increase in the acetate-to-propionate ratio in all the propolis extract treatmentscompared with the monensin treatment, without a concomitant decline in SCFA concentrations, may reflect a differentmode of action of propolis: more extensive dietary fiber digestion. Accordingly, the propolis extract treatments may haveinfluenced fungi and cellulolytic bacteria, the two most important fiber-digesting groups. In the present study, the directeffects of propolis on the bacterial community were not assessed and have therefore be addressed experimentally in followup studies.
When compared to monensin, the propolis extracts improved the TDOM. This finding is in agreement with that of Costaet al. (2012) in a study of the nutrient digestibility of a diet supplemented with propolis in buffaloes. The degradation endproducts of the flavonoids found in both propolis types are known to be capable of influencing fiber degradability, as bacte-rial ring fission of aglycone flavonoids produces phenolic acids, such as 3,4-dihydroxyphenylacetic acid from isoquercitrinand quercetin or phenylacetic acid from naringenin (Winter et al., 1989). Some of these simple phenolic compounds mayinfluence the biosynthesis of aromatic amino acids, as both biosynthesis pathways are linked through cinnamic acid. Phenyl-propanoic acid and phenylacetic acid have been reported to enhance cellulose degradation and the growth of several strainsof Ruminococcus albus (Stack and Cotta, 1986).
A number of the major constituents of propolis (i.e., flavonoids, propanoic acid, and medicarpin) may act as antioxidants.However, information about the effects of such compounds on ruminal fermentation and microbial growth is very scarce(Cattani et al., 2012). Some natural and synthetic antioxidants were found to enhance the activity and growth of ruminalmicrobes and to promote the digestion and efficiency of diet utilization in vitro (Vázquez-Anón and Jenkins, 2007). As mostruminal microbes are predominantly strict anaerobes with less developed antioxidant capacity than facultative anaerobicand aerobic microbes, it is possible that the addition of antioxidants would lessen oxidative stress, thus promoting betterconditions for microbial growth (Cattani et al., 2012). Smith et al. (2002) observed that the addition of 50 mg/kg of ethoxyquinto the diet of dairy cows can improve milk yields, as well as DM digestibility. This finding suggests that ethoxyquin mayexert an antioxidant effect in ruminal fermentation. However, further investigations are needed to clarify this assumption.
Propolis had only weak effects, if any, on the PF. The PF is an indicator of the efficiency of microbial protein synthesis. Theweak effects in the present study are consistent with the relatively constant concentration of the incubation fluid ammonia,as this illustrates that the nitrogen used by microbes for their protein synthesis remained unchanged. In contrast to the
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
findings of the current study, Broudiscou et al. (2002), using a dual outflow fermenter, reported a slight increase in ruminalmicrobial outflow in response to propolis treatment.
A very puzzling finding of the present study was the lack of any dose–response relationship with both types of propolistested. Certain levels of BPR and EBP extracts (BRP50 �g and EBP25 �g) appeared to occasionally be more effective than others

G ModelA
war
5
(psofpb
C
A
fP
R
A
AA
B
B
B
B
B
C
C
d
D
D
F
G
H
Í
L
M
M
N
O
ARTICLE IN PRESSNIFEE-13189; No. of Pages 10
A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx 9
hen compared to those of the negative control or monensin or both. We have no explanation for this finding other thanssuming that the two propolis types were already sufficiently efficient at low levels to produce more or less a maximumesponse in ruminal fermentation.
. Conclusions
The current study demonstrated that the composition of the individual compounds of the two selected sources of propolisBRP and EBP) differed greatly. Nevertheless, there was a large similarity in the ruminal degradation of nutrients, GP, and CH4roduction in response to the BRP and EBP treatments. Additional source-specific variation was small. Although the findingsuggest that there could be an underlying basic effect of propolis, independent of the type or site of collection, existing reportsf the inefficiency of certain types of propolis question this assumption. In common with monensin, propolis reduced CH4ormation, but it was not as effective as monensin in reducing emissions. Further studies need to demonstrate the efficacy ofropolis in vivo. They also need to identify the active component(s) of propolis and subsequently analyze these in commercialatches to be able to predict their efficiency.
onflict of interest
There is no conflict of interest.
cknowledgments
This project was funded in part by the The World Academy of Sciences (TWAS), Trieste, Italy, and the National Councilor Scientific and Technological Development (CNPq) (3240188145) in Brazil. The assistance of Dr. Sônia Maria de Stefanoiedade, Exact Science Department of Agriculture College ESALQ/USP, Brazil, in the statistical evaluation is highly appreciated.
eferences
lencar, S.M., Oldoni, T.L.C., Castro, M.L., Cabral, I.S.R., Costa-Neto, C.M., Cury, J.A., Rosalen, P.L., Ikegaki, M., 2007. Chemical composition and biologicalactivity of a new type of Brazilian propolis: red propolis. J. Ethnopharmacol. 113, 278–283.
OAC, 1995. Official Methods of Analysis, 18th ed. Association of Analytical Chemists, Gaithersburg, MD, USA.raujo, R.C., Pires, A.V., Mouro, G.B., Abdalla, A.L., Sallam, S.M.A., 2011. Use of blanks to determine in vitro net gas and methane production when using
rumen fermentation modifiers. Anim. Feed Sci. Technol. 166, 155–162.lümmel, M., Becker, K., 1997. The degradability characteristics of fifty-four roughages and roughage neutral-detergent fibre as described by in vitro gas
production and their relationship to voluntary feed intake. Br. J. Nutr. 77, 757–786.lümmel, M., Steingass, H., Becker, K., 1997. The relationship between in vitro gas production, in vitro microbial biomass yield and 15N incorporations for
the prediction of voluntary feed intake of roughages. Br. J. Nutr. 77, 911–921.roudiscou, L.P., Papon, Y., Broudiscou, A.F., 2000. Effects of dry plant extracts on fermentation and methanogenesis in continuous culture of rumen microbes.
Anim. Feed Sci. Technol. 87, 263–277.roudiscou, L.P., Papon, Y., Broudiscou, A.F., 2002. Effects of dry plant extracts on feed degradastion and the production of rumen microbial biomass in a
dual outflow fermenter. Anim. Feed Sci. Technol. 101, 183–189.ueno, I.C.S., Filho, S.L.S., Gobbo, S.P., Louvandini, H., Vitti, D.M.S.S., Abdalla, A.L., 2005. Influence of inoculum source in a gas production method. Anim.
Feed Sci. Technol. 123, 95–105.attani, M., Franco, T., Lucia, B., Stefano, S., 2012. Synthetic and natural polyphenols with antioxidant properties stimulate rumen microbial growth in vitro.
Anim. Prod. Sci. 52, 44–50.osta, J.B.G., Zeoula, L.M., Franco, S.L., de Moura, L.P.P., Valero, M.V., Simioni, F.L., da Paula, E.M., Samensari, R.B., 2012. Effect of propolis product on digestibility
and ruminal parameters in buffaloes consuming a forage-based diet. Ital. J. Anim. Sci. 11 (4), http://dx.doi.org/10.4081/ijas.2012.e78, article e78.e Aguiar, S.C., Zeoula, L.M., Franco, S.L., Peres, L.P., Arcuri, P.B., Forano, E., 2013. Antimicrobial activity of Brazilian propolis extracts against rumen bacteria
in vitro. World J. Microbiol. Biotechnol. 29, 1951–1959.ehority, B.A., Damron, W.S., McLaren, J.B., 1983. Occurrence of the rumen ciliate Oligoisotricha bubali in domestic cattle (Bos taurus). Appl. Environ.
Microbiol. 45, 1394–1397.uarte, S., Pedro, L.R., Mitsue, F.H., Jaime, A.C., William, H.B., Marquis, R.E., Vera, L.G.R., Adilson, S., Masaharu, I., Hyun, K., 2006. The influence of a novel
propolis on mutans streptococci biofilms and caries development in rats. Arch. Oral Biol. 51, 15–22.ernández, M.C., Osmany, C.R., Istides, R.P., Rodny, M.O., Ingrid, R.N., Anna, L.P., Luca, R., 2008. GC-MS determination of isoflavonoids in seven red Cuban
propolis samples. J. Agric. Food Chem. 56, 9927–9932.oering, H.K., Van Soest, P.J., 1970. Forage fiber analysis (apparatus, reagents, procedures and some applications). In: Agric. Handbook No. 379. US Agricultural
Research Service, Washington (DC).ashem, N.M., Abd El-Hady, A., Hassan, O., 2013. Effect of vitamin E or propolis supplementatin on semen quality, oxidative status and hemato-biochemical
changes of rabbit bucks during hot season. Livest. Sci. 157, 520–526.tavo, C.C.B.F., Morais, M.G., Costa, C., Ítavo, L.C.V., Franco, G.L., Da Silva, J.A., Reis, F.A., 2011. Addition of propolis or monensin in the diet: behavior and
productivity of lambs in feedlot. Anim. Feed Sci. Technol. 165, 161–166.ongo, C., Bueno, I.C.S., Nozella, E.F., Goddoy, P.B., Cabral Filho, S.L.S., Abdalla, A.L., 2006. The influence of head-space and inoculum dilution on in vitro
ruminal methane measurements. In: Soliva, C.R., Takahashi, J., Kreuzer, M. (Eds.), Greenhouse Gases and Animal Agriculture: An Update. Int. Congr.Series No. 1293. Elsevier, The Netherlands, pp. 62–65.
arkham, K.R., Mitchel, K.A., Wilkins, A.L., Daldy, J.A., Lu, Y., 1996. HPLC and GC-MS identification of the major organic constituents in New Zealand propolis.Phytochem 42, 205–211.
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
orsy, A.S., Abdalla, A.L., Soltan, Y.A., Sallam, S.M.A., El-Azrak, K.M., Louvandini, H., Alencar, S.M., 2013. Effect of Brazilian red propolis administration onhematological, biochemical variables and parasitic response of Santa Inês ewes during and after flushing period. Trop. Anim. Health Prod. 45, 1609–1618.
ational Research Council, 2007. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids. National Academy of Sciences,Washington, DC, USA.
eztuerk, H., Sagmanligil, V., 2009. Role of live yeasts on rumen ecosystem. Deutsche Tieraerztl. Wochensschr. 116, 244–248.

G Model
ARTICLE IN PRESSANIFEE-13189; No. of Pages 1010 A.S. Morsy et al. / Animal Feed Science and Technology xxx (2014) xxx–xxx
Oeztuerk, H., Emre, B., Sagmanligil, V., Piskin, I., Fidanci, U.R., Pekcan, M., 2010. Effects of nisin and propolis on ruminal fermentation in vitro. J. Anim. Vet.Adv. 9, 2752–2758.
Oldoni, T.L.C., Cabral, I.S.R., Regitano d’Arce, M.A.B., Rosalen, P.L., Ikegaki, M., Andrea, M., Nascimento, A.M., Alencar, S.M., 2011. Isolation and analysis ofbioactive isoflavonoids and chalcone from a new type of Brazilian propolis. Separat. Purif. Technol. 77, 208–213.
Palmiquist, D., Conrad, H., 1971. Origin of plasma fatty acid in lactating dairy cows fed high fat diets. J. Dairy Sci. 54, 1025–1031.Park, Y.K., Alencar, S.M., Scamparini, A., Regina, P., Aguiar, C.L., 2002. Própolis produzida no sul do Brasil, Argentina e Uruguai: evidências fitoquímicas de
sua origem vegetal. Cienc. Rural 32 (6(nov./dez.)), 997–1003.Park, Y.K., Ikegaki, M., Alencar, S.M., Moura, F.F., 2000. Evaluation of Brazilian propolis by both physicochemical methods and biological activity. Honeybee
Sci. 21, 85–90.Preston, T.R., 1995. Biological and chemical analytical methods. In: Preston, T.R. (Ed.), Tropical Animal Feeding: A Manual for Research Workers. FAO, Rome,
pp. 191–264, Chap. 9.Rispoli, T.B., Rodrigues, I.L., Martins Neto, R.G., Kazama, R., Prado, O.P.P., Zeoula, L.M., Arcuri, P.B., 2009. Ruminal ciliate protozoa of cattle and buffalo fed
on diet supplemented with monensin or extracts from propolis. Pesquisa Agropecuária Brasileira, Brasília, DF 44, 92–97.Russell, J.B., Strobel, H.J., 1988. Effects of additives on in vitro ruminal fermentation: a comparison of monensin and bacitracin, another gram-positive
antibiotic. J. Anim. Sci. 66, 552–558.Selje-Assmann, N., Hoffmann, E.M., Becker, K., 2008. A batch incubation assay to screen plant samples and extracts for their ability to inhibit rumen protein
degradation. Anim. Feed Sci. Technol. 145, 302–318.Smith, J.L., Sheffield, L.G., Saylor, D., 2002. Impact of ethoxyquin on productivity of dairy cattle. J. Dairy Sci. 85 (Suppl. 1), 358.Soltan, Y.A., Morsy, A.S., Sallam, S.M.A., Louvandini, H., Abdalla, A.L., 2012. Comparative in vitro evaluation of forage legumes (prosopis, acacia, atriplex, and
leucaena) on ruminal fermentation and methanogenesis. J. Anim. Feed Sci. 21, 759–772.Stack, R.J., Cotta, M.A., 1986. Effect of 3-phenylpropanoic acid on growth of, and cellulose utilization by, cellulolytic ruminal bacteria. Appl. Environ. Microbiol.
52, 209–210.Statistical Analysis System, 2002. SAS PC Windows Version 9.2.0. SAS Institute Inc., Cary, NC, USA.Theodorou, M.K., Williams, B.A., Dhanoa, M.S., McAllan, A.B., France, J., 1994. A simple gas production method using a pressure transducer to determine the
fermentation kinetics of ruminant feeds. Anim. Feed Sci. Technol. 48, 185–197.Ushida, K., Jouany, J.P., 1996. Methane production associated with rumen-ciliated protozoa and its effect on protozoan activity. Lett. Appl. Microbiol. 23,
129–132.Van Soest, P.J., 1973. Collaborative study of acid detergent fibre and lignin. J. Assoc. Off. Anal. Chem. 56, 781–784.Van Soest, P.J., Robertson, J.B., Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal
nutrition. J. Dairy Sci. 74, 3583–3597.Vázquez-Anón, M., Jenkins, T., 2007. Effects of feeding oxidized fat with or without dietary antioxidants on nutrient digestibility, microbial nitrogen, and
fatty acid metabolism. J. Dairy Sci. 90, 4361–4367.Winter, J., Moore, L.H., Dowell Jr., V.R., Bokkenheuser, V.D., 1989. C-ring cleavage of flavonoids by human intestinal bacteria. Appl. Environ. Microbiol. 55,
1203–1208.Wischer, G., Boguhn, J., Steingaß, H., Schollenberger, M., Hartung, K., Rodehutscord, M., 2013. Effect of monensin on in vitro fermentation of silages and
microbial protein synthesis. Arch. Anim. Nutr. 67, 219–234.Zawadzki, F., Prado, I.N., Marques, J.A., Zeoula, L.M., Rotta, P.P., Sestari, B.B., Valero, M.V., Rivaroli, D.C., 2011. Sodium monensin or propolis extract in the
Please cite this article in press as: Morsy, A.S., et al., Comparison of the in vitro efficiency of supplementary bee propolisextracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation ofmethane. Anim. Feed Sci. Tech. (2014), http://dx.doi.org/10.1016/j.anifeedsci.2014.11.004
diets of feedlot-finished bulls: effects on animal performance and carcass characteristics. J. Anim. Feed Sci. 20, 16–25.Zhou, X., Meile, L., Kreuzer, M., Zeitz, J.O., 2013. The effect of saturated fatty acids on methanogenesis and cell viability of Methanobrevibacter ruminantium.
Archaea http://dx.doi.org/10.1155/2013/106916Zhou, J.H., Li, Y., Zhao, J., Xue, X.F., Wu, L.M., Chen, F., 2008. Geographical traceability of propolis by high-performance liquid-chromatography fingerprints.
Food Chem. 108, 749–759.
Copyright © 2022 FDOKUMEN




















