
Comparative study on Lewis lung tumour lines with ‘low’ and ‘high’ metastatic capacity III....
11
CLIN~ EXPL. METASTASIS, 1989, VOL~ 7, NO. 6, 659-669 Comparative study on Lewis lung tumour lines with 'low' and 'high' metastatic capacity III. Glycosaminoglycan synthesis, transport and degradation in cell lines G. POGANYt, E. MOCZAR;~, A. JENEYw J. TIMARw F. TIMARw K. DITROIt, and K. LAPISw t Joint Research Organization of the Hungarian Academy of Sciences and Semmelweis Medical University, Department of Molecular Pathology, Ull6i. fit 26, H-1085 Budapest, Hungary Laboratoire de Biochimie du Tissu Conjonctif, CNRS, Facult~ de M~decine, Universit6 de Paris, 8 rue du General Sarrail, 94010 Cr~teil, France w First Institute of Pathology and Experimental Cancer Research, Semmelweis Medical University, Ull6i fit 26, H-1085 Budapest, Hungary (Received 25 July 1988; accepted 20 March 1989) The glycosaminoglycans (GAGs) of low (LM) and highly metastatic (HM) cell lines of the Lewis lung tumour (3LL) were compared using [3H]glucosamine labelling techniques. The GAGs isolated from nuclei, cytoplasm, pericellular fractions and medium were analysed by cellulose acetate electrophoresis and by digestion with specific enzymes, and the following conclusions were drawn. 1. Increased cellular uptake and incorporation of [3H]glucosamine into glycoconju- gates of the cytoplasm was a typical feature of the highly metastatic cell line after a 48-h labelling. However, there was no elevated radioactivity in glycolipids. 2. Radioactivity of the purified GAGs was two and three times higher in nuclear and cytoplasmic fractions of HM cells than in those of LM cells. There was much less difference between the two cell lines in the pericellular fractions. 3. A definite change from chondroitin sulphate to dermatan sulphate dominancy was re- corded in each GAG fraction. Higher heparan sulphate labelling was observed in the cytoplasmic and periceltular GAGs of HM cultures. 4. In the post-labelling period about three times more GAG was present in the extracellular compart- ment of the HM cultures compared with the LM cultures. 5. In the LM cultures the total GAG-associated radioactivity decreased by 73 per cent in the 48-h chase period whereas in the HM cultures it decreased by only 30 per cent. This indicates a higher rate of GAG degradation in the LM cultures. Abbreviations: GAG, glycosaminoglycan; 3LL, Lewis lung tumour; HM, highly metastatic; LM, low metastatic; FCS, foetal calf serum; PBS, phosphate- buffered saline; HP, heparin; HS, heparan sulphate; HA, hyaluronic acid; CS, chondroitin sulphate; DS, dermatan sulphate; PCA, perchloric acid; TCA, trichloroacetic acid; CPC, cetylpyridinium chloride; E, extracellular; P, peri- cellular; C, cellular; CY, cytoplasmic; N, nuclear compartment; ECM, extra- cellular matrix. Introduction The composition of the cell surface and of the microenvironment plays an important role in contacts between various type of cells, such as malignant and normal types [34]. Thus proteoglycans and their sugar moieties, the glycosamino- glycans, are critical elements of both the pericellular compartment and the extra- 0262-0898/89 $3,00 1989 Taylor & Francis Ltd.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Comparative study on Lewis lung tumour lines with ‘low’ and ‘high’ metastatic capacity III....

CLIN~ EXPL. METASTASIS, 1989, VOL~ 7, NO. 6, 659-669
Comparative study on Lewis lung tumour lines with 'low' and 'high' metastatic capacity III. Glycosaminoglycan synthesis, transport and degradation in cel l lines
G. P O G A N Y t , E. MOCZAR;~ , A. JE N E Y w J. T I M A R w F. T I M A R w K. D I T R O I t , and K. L A P I S w
t Joint Research Organization of the Hungarian Academy of Sciences and Semmelweis Medical University, Department of Molecular Pathology, Ull6i. fit 26, H-1085 Budapest, Hungary
Laboratoire de Biochimie du Tissu Conjonctif, CNRS, Facult~ de M~decine, Universit6 de Paris, 8 rue du General Sarrail, 94010 Cr~teil, France w First Institute of Pathology and Experimental Cancer Research, Semmelweis Medical University, Ull6i fit 26, H-1085 Budapest, Hungary
(Received 25 July 1988; accepted 20 March 1989)
The glycosaminoglycans (GAGs) of low (LM) and highly metastatic (HM) cell lines of the Lewis lung tumour (3LL) were compared using [3H]glucosamine labelling techniques. The GAGs isolated from nuclei, cytoplasm, pericellular fractions and medium were analysed by cellulose acetate electrophoresis and by digestion with specific enzymes, and the following conclusions were drawn. 1. Increased cellular uptake and incorporation of [3H]glucosamine into glycoconju- gates of the cytoplasm was a typical feature of the highly metastatic cell line after a 48-h labelling. However, there was no elevated radioactivity in glycolipids. 2. Radioactivity of the purified GAGs was two and three times higher in nuclear and cytoplasmic fractions of H M cells than in those of LM cells. There was much less difference between the two cell lines in the pericellular fractions. 3. A definite change from chondroitin sulphate to dermatan sulphate dominancy was re- corded in each GAG fraction. Higher heparan sulphate labelling was observed in the cytoplasmic and periceltular GAGs of H M cultures. 4. In the post-labelling period about three times more G A G was present in the extracellular compart- ment of the H M cultures compared with the LM cultures. 5. In the L M cultures the total GAG-associated radioactivity decreased by 73 per cent in the 48-h chase period whereas in the H M cultures it decreased by only 30 per cent. This indicates a higher rate of G A G degradation in the LM cultures.
Abbreviations: GAG, glycosaminoglycan; 3LL, Lewis lung tumour; HM, highly metastatic; LM, low metastatic; FCS, foetal calf serum; PBS, phosphate- buffered saline; HP, heparin; HS, heparan sulphate; HA, hyaluronic acid; CS, chondroitin sulphate; DS, dermatan sulphate; PCA, perchloric acid; TCA, trichloroacetic acid; CPC, cetylpyridinium chloride; E, extracellular; P, peri- cellular; C, cellular; CY, cytoplasmic; N, nuclear compartment; ECM, extra- cellular matrix.
Introduction T h e c o m p o s i t i o n of the cell surface and of the m i c r o e n v i r o n m e n t p lays an
i m p o r t a n t role in contac t s be tween var ious type of cells, such as m a l i g n a n t and n o r m a l types [34]. T h u s p ro t eog lycans and the i r sugar moie t ies , the g l y c o s a m i n o - g lycans , are cr i t ica l e l emen t s of b o t h the pe r i ce l lu l a r c o m p a r t m e n t and the ex t r a -
0262-0898/89 $3,00�9 1989 Taylor & Francis Ltd.

660 G. Pogg~ny et al.
cellular matrix and deserve special attention in the elucidation of the molecular mechanism implicated in the metastatic process of the tumour cells [6, 7, 24].
In previous studies, having compared the in vivo liver-selected Lewis lung tumour cells with the parent cells [21, 22], we found a remarkable change in the G A G pattern. The results indicated that the increased G A G biosynthesis and elevated HS/CS ratio may be related to the liver-colonizing potential of these tumour cells [14, 31]. However, the actual amount of GAGs present is determined not only by the synthesis but also by the rate of degradation. Further, the transport of these molecules between the intracellular and extracellular compartments must be taken into account. Therefore, the purpose of this work was to determine the distribution of GAGs in the nuclear, cytoplasmic, pericellular and extracellular fractions after a 48-h labelling and their subsequent changes during the post- labelling incubation period.
Materials and m e t h o d s
Materials RPMI-1640 medium and FCS were obtained from Flow Laboratories (U.K.).
Gentamycin sulphate and Pronase E from Streptomyces griseus (E.C. 3.4.24.4) were purchased from Serva (F.R.G.). [3H]Glucosamine [specific activity 7'4 • 10 l~ Bq/mmol (2 Ci/mmol)] was from Amersham (U.K.). The G A G standards (CS type A, B and C; HP-grade I; HS; HA-grade I) and the specific enzymes for digestions, hyaluronidase from Streptomyces hyalurolyticus type IX (E.C. 4.2.2.1) and chondroitinase ABC from Proteus vulgaris (E.C. 4.2.2.4), were from Sigma (U.S.A.). Hyaluronidase from bull testes (E.C. 3.2.1.35) was purchased from Reanal (Hungary). All of the chemicals used were of analytical-reagent grade.
Cell lines and cultures The LM cell line [derived from Lewis lung adenocarcinoma (3LL)] and H M cell
line (derived from the highly metastatic 3LL tumour [21]) were described previously [14] and maintained by serial passage in RPMI-1640 medium containing 2 mg/ml gentamycin sulphate, 10 per cent heat-inactivated FCS and cultured in a humidified, 7"5 per cent CO 2 atmosphere at 37~ For the radioactive labelling of the glycoconjugates, 1"5 x 106 cells were incubated in 2 5 - c m 2 Greiner culture flasks in the presence of 3"7 x t05 Bq/ml (10~tCi/ml) [3H]glucosamine for 48 h in 8ml of medium. During this period the number of cells of both L M and H M cultures nearly doubled. At the end of the labelling period, half of the cultures were processed and the others were incubated further without radioactive glucosamine for 0"5, 5, 24 and 48 h in the presence of 10-5 M non-radioactive glucosamine. The size of inoculum for cell culturing was selected in a way allowing the study of the biosynthesis of GAGs in growing cultures, whereas secretion and degradation of GAGs was followed in non-growing cultures. (In this post-labelling chase period, there was no change in the number of cells.) The liver-metastatic capacity of tumour cells was continuously tested by using the spleen-liver model described previously [21].
Collection of various glycoconjugate fractions The extracellular glycoconjugates (E) were isolated from culture medium
separated by centrifugation (500g, 5 min) from the cells. The cells were washed with

Lewis lung tumor lines 661
calcium- and magnesium-free PBS and were digested in 3 ml of PBS containing 0"08 per cent trypsin and 0"02 per cent E D T A for 30 rain at 4~ Tryps in was then inactivated with 40 #1 of FCS. After this mild trypsination, the viability of the cells was higher than the 80 per cent, assessed by the T r y p a n blue dye exclusion test. The samples were centrifuged at 125g for 20 rain and the supernatants containing the pericellular glycoconjugates (P) were collected. The cells were washed with PBS and suspended in hypotonic buffer (10raM NaC1, 3 r a m MgCI2, 1 0 r a m Tr is -HC1, p H 7"4) for 1 h at 4~ The swollen cells were disrupted in a P T F E homogenizer, then centrifuged (500g, 15 min). The sediment was washed first with the above buffer containing 0"25 M sucrose, then in the presence of 0"5 per cent Tr i ton X-100. The supernatants were collected as cytoplasmic fractions (CY) and the sedimented nuclei (N) suspended in 0"05 M Tr i s -HC1 buffer (pH 8). The puri ty of the nuclei was checked under a light microscope using haematoxylin stain. All samples were stored at - 20~
Separation of the cellular glycoconjugates To determine the distribution of radioactivity among the various cellular
fractions, sequential extractions were per formed by applying cold 0"5 M PCA, ethanol-diethylether (3 : 1, v/v) and 0"2 M N a O H . T h e pool of low molecular weight compounds was obtained from the cold 0'5 M PCA extract after precipitating the protein-free glycosaminoglycans, glycopeptides and polysaccharides (glycoconju- gate 1 fractions) with ethanol at 4~ Glycopipids were recovered f rom the cold PCA precipitates by extraction with ethanol-diethylether (3:1 , v/v), whereas gly- coproteins and proteoglycans were solubilized by 0"2 M N a O H (glycoconjugate 2 fractions).
Study of the glycosaminoglycans Aliquots of cellular and extracellular fractions were digested with pronase [17].
The fl-elimination was carried out [2] with 0'5 M N a O H for 3 h at 4~ to remove core peptides. After neutralization with concentrated acetic acid, the samples were adjusted to 10 per cent T C A at 4~ The mixture was centrifuged at 500g for 25 min and the supernatant was dialysed against several changes of distilled water at 4~ dried and dissolved in 200 #1 of distilled water. After each purification step, the effect of the t reatment was monitored by precipitation (aliquots of 50#1) on Wha tman 3 M M filter-paper with 1 per cent (w/v) CPC [33]. T o estimate the composit ion of GAG, aliquots of the samples were dissolved after CPC precipitation and the purified glycosaminoglycans were digested alternatively by chondroitinase ABC [25], Streptomyces hyaluronidase [1,20], testicular hyaluronidase [25] or treated with nitrous acid [29]. The undigested G A G was precipitated with ethanol [23]. T h e radioactivity in supernatants containing the released disaccharides was taken as the labelling of the appropriate type of G A G . The calculation of the radioactivity of each G A G component was performed principally according to the method described by Angello et al. [1], completed with the determination of dermatan and chondroit in sulphate. To calculate the radioactivity of chondroit in sulphate, the count of the radioactivity of the disaccharides released after the digestion by Streptomyces hyaluronidase was subtracted f rom that obtained by testicular hyaluronidase digestion. The radioactivity of dermatan sulphate was calculated by subtracting the radioactivity of chondroitin sulphate (calculated as described above) f rom the radioactivity of the disaccharides obtained by chondroitinase ABC. The specificity

662 G. Pogdtny et al.
of the enzymes was controlled using G A G standards and cellulose acetate electrophoresis. T h e electrophoretic separation of the isolated G A G s was performed according to Newton et al. [18] on cellulose acetate, but using 0"1 M calcium acetate- acetic acid buffer (pH 5'8). Samples of 2 #1 of G A G s were subjected to electro- phoresis for 1"4 h, then stained with a solution of 0"1 per cent Alcian blue, 0"025 M sodium acetate and 0"05 M MgCI 2 in 50 per cent ethanol (pH 5"8). The spots corresponding to the separated molecules were cut out and subjected to liquid scintillation counting.
The radioactivity of each fraction was determined in toluene-based scintillating cocktails as reported previously [31] by applying a Beckman LS 100C spectrometer. The nucleic acid content of the cells was determined by measuring A260 in the hot PCA fractions according to Scott et al. [27]. Statistical calculations were performed according to Student ' s t-test.
R e s u l t s The established cell lines ( L M and H M ) isolated f rom the original tumours
retained their altered metastatic capacity (table 1). Table 2 shows the incorporation of [3H]glucosamine into the various cytoplasmic glycoconjugate fractions of the cell lines after 48-h labelling. The incorporation was higher in the H M than in the L M cells, with the exception of the glycolipids. These differences may be caused by the significantly enlarged precursor pool of H M cells. The greatest differences between the labelling of the two lines were found in the glycoconjugate 1 fractions of the cytoplasms. These fractions were characterized by their solubility in cold perchloric acid and by their insolubility in ethanol (i.e. protein-free GAGs , oligosaccharides and glycopeptides). The labelling of total glycoconjugates was similar in the nuclei of the L M and H M cells, in contrast to their cytoplasms. Considering the distribution of [3H]glucosamine among the various cytoplasmic fractions, an increased labelling was apparent in the glycoconjugate 1 fraction of H M cells compared with L M cells (78 per cent of total label). The labelling of the other glycoconjugate fractions decreased at the same t ime in the H M cells, related to the total radioactivity.
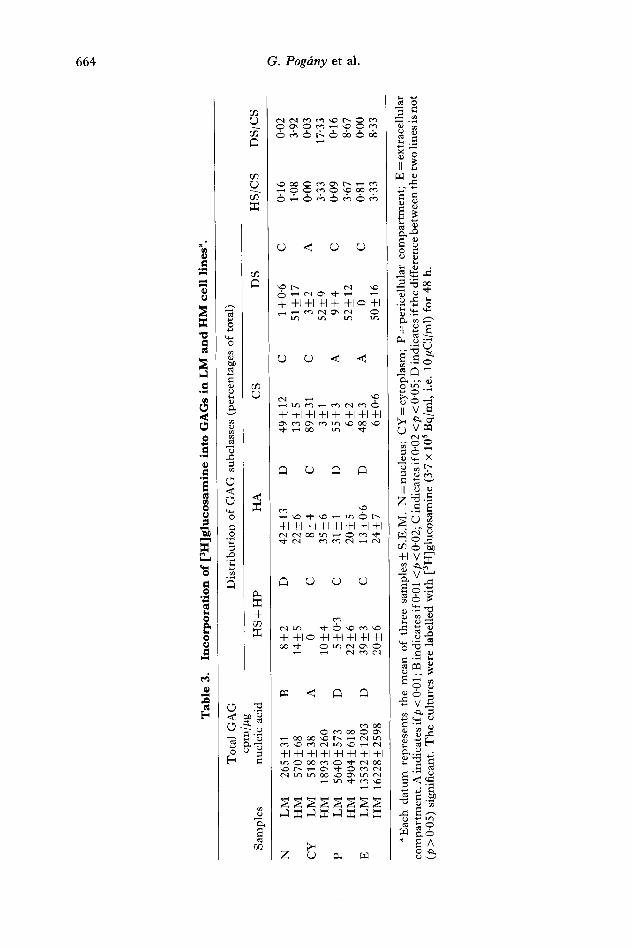
Measurements on purified G A G provided data for the higher [3H]glucosamine labelling of the G A G molecules in the cytoplasmic and nuclear fractions of the H M cells compared with the L M cells (table 3). However , there was no significant difference between the two cell lines in the level of labelling of the pericellular and extracellular fractions (see total G A G values). This implies that the distribution of the total cellular G A G shows a different pattern in the two cell lines: the ratio of labelling between the nuclear, cytoplasmic and pericellular fractions was 1 : 2 : 21 and 1 : 3 : 9 in the L M and in the H M cells, respectively. In the L M cells only 12 per cent of the total [3H]glucosamine associated with G A G was recovered in the nuclear and cytoplasmic fractions. This value was 33 per cent in the H M cells.
The composit ion of the G A G s showed differences between the cellular and extracellular fractions and between the L M and the H M Lewis lung cell lines (table 3). In the cytoplasms of the L M cells G A G was represented almost entirely by chondroitin sulphate. In the nuclear and pericellular fractions there was also substantial labelling of hyaluronate. T h e most remarkable feature of the G A G composit ion in the H M cells was the dominance of dermatan sulphate, presumably at the expense of chondroitin sulphate. Increased heparan sulphate labelling of the cytoplasmic and pericellular fractions was also observed.

Tab
le 1
. G
row
th a
nd
met
asta
tic
cap
acit
y to
th
e li
ver
of L
M a
nd
HM
cel
ls"
.
No
. o
f
Cel
l in
tras
ple
nic
ally
ty
pe
ino
cula
ted
cel
ls
Pri
mar
y t
um
ou
r
Inci
den
ce
Mea
n w
eigh
t +-
S.E
.M.
(g)
Liv
er m
etas
tasi
s
Inci
den
ce
Ind
ivid
ual
nu
mb
ers
Mea
n +
- S.E
.M.
LM
10
6 6/
6 1.
56 +
- 0.4
4
HM
1
0 6
9/
9 1"
68 +
- 0"3
0
4/6
76,
60,
50,
12,
0, 0
3
2"8
+-1
3"4
9/9
460,
430
, 20
3, 1
43,
172-
1 +
-55-
2 13
1, 8
6, 4
2, 4
0, 1
4
"Ev
alu
atio
n o
n t
he 1
6th
po
st-i
no
cula
tio
n d
ay.
Eac
h d
atu
m r
epre
sent
s m
ean
+ S
.E.M
.
Tab
le 2
. D
istr
ibu
tion
of
[3H
]glu
cosa
min
e la
bel
in
th
e ce
llu
lar
frac
tion
s of
LM
an
d H
M c
ells
in
vit
ro".
Poo
l G
lyco
con
jug
ate
1 G
lyco
lip
ids
Gly
coco
nju
gat
e 2
To
tal
Cel
l li
nes
cpm
~o
cp
m
~ cp
m
~ cp
m
~ cp
m
N
LM
2
70
+4
9
+0
'1
15
21
+6
2
52+
_2
252+
-4
9_+
0'1
888+
-24
30
_+
0"8
29
31+
-94
100_
+3
HM
25
3+-6
8+
-0"2
18
72+
-83
60+
-3
25
9+
16
8+
-0-5
74
0+-6
5 24
+-2
"1
3124
+-1
69
100+
-5
CY
L
M
7937
_+60
6 2
0•
2386
3+-6
28
62+
-2
1601
+-1
21
4+-0
.3
5451
_+18
1 14
+-0
.5
3885
2+-1
536
100_
__4
HM
28
395+
-137
15
+-0
'1
14
91
25
+1
03
10
78
+-5
16
48+
-41
1_+
0'1
1240
4+-7
78
6+-0
.4
19
15
72
+1
12
65
10
0+-6
a T
he
valu
es li
sted
are
ave
rage
s __
_ S.E
.M.
fi'o
m th
ree
diff
eren
t cu
ltu
re d
ishe
s. D
ata
are
in c
pm
/pg
nu
clei
c ac
id. P
ool =
col
d P
CA
ext
ract
aft
er r
emov
al
of g
lyco
conj
ugat
e 1
frac
tion
wit
h et
han
ol
prec
ipit
atio
n; G
lyco
nco
nju
gat
e 1
=al
coh
oli
c pr
ecip
itat
e of
col
d P
CA
-so
lub
le f
ract
ion;
Gly
coli
pid
s =
alc
oh
ol-
die
thy
l et
her
extr
act
of P
CA
pre
cipi
tate
; gl
ycoc
onju
gate
2 =
res
idua
l m
ater
ial
diss
olvi
ng i
n N
aOH
; N
= n
ucl
eus;
CY
= c
ytop
lasm
. T
he
cult
ure
s w
ere
labe
lled
wit
h [
3H]g
luco
sam
ine
(3'7
x 1
0 s B
q/m
l, i
.e,
10/~
Ci/
ml)
for
48
h.
GO
4~
Tab
le 3
. In
corp
orat
ion
of
[3H
]glu
cosa
min
e in
to G
AG
s in
LM
an
d H
M c
ell
lin
es a.
Sam
ple
s
N
LM
To
tal
GA
G
Dis
trib
uti
on
of
GA
G s
ubcl
asse
s (p
erce
ntag
es o
f to
tal)
cp
m//
~g
nu
clei
c ac
id
HS
+ H
P
HA
C
S
DS
H
S/C
S
DS
/CS
0-02
CY
P E
26
5+
31
B
8
+2
D
4
2+
13
D
4
9+
12
C
1+
-0"6
C
0"
16
HM
5
70
+6
8
14
+5
2
2+
6
13
+5
51
+-1
7 1"
08
3-92
L
M
51
8+
38
A
0
C
8+
4
C
89
+3
1
C
3+
2
A
0-00
0"
03
HM
18
93+
-260
1
0+
4
35
+6
3
+1
52
_+9
3-33
17
'33
LM
56
40__
_573
D
5_
__0"
3 C
3
1+
1
D
55-t
-3
A
9-t-
4 C
0-
09
0"16
H
M
4904
___6
18
22+
_6
20+
-5
6+-2
52
_+12
3"
67
8"67
L
M
1353
2+-1
203
D
39+
-3
C
13_+
0-6
D
48-t
-3
A
0 C
0-
81
0-00
H
M
16
22
8+
25
98
20
+-6
24
+-7
6_
+0"6
50
+-1
6 3-
33
8-33
g
a E
ach
dat
um
re
pre
sen
ts
the
mea
n
of t
hre
e sa
mpl
es__
+ S
.E.M
. N
= n
ucle
us;
CY
= c
yto
pla
sm;
P =
per
icel
lula
r co
mp
artm
ent;
E
= e
xtr
acel
lula
r co
mp
artm
ent.
A i
ndic
ates
ifp
< 0
"01
; B i
ndic
ates
if 0
-01
<p
< 0
-02;
C i
ndic
ates
if 0
"02
<p
< 0
"05;
D i
ndic
ates
if t
he
diff
eren
ce b
etw
een
the
tw
o li
nes
is n
ot
(p >
0-0
5) s
igni
fica
nt.
Th
e cu
ltu
res
wer
e la
bell
ed w
ith
[3
H]g
luco
sam
ine
(3"7
x 1
05 B
q/m
l, i
.e.
10 k
tCi/
ml)
for
48
h.

Lewis lung tumor lines 665
Most of the synthesized G A G was excreted f rom the cells and appeared in the extracellular compar tment (table 3). Therefore, the dynamics of G A G transport were moni tored in a pulse chase experiment, i.e. we measured the translocation of the prelabelled G A G molecules f rom the cellular to the pericellular and extracellular compar tments in the presence of unlabelled glucosamine. As our previous studies had revealed that these cells achieved their dynamic equil ibrium in 48 h, we applied a 48-h labelling period.
Table 4 shows that the radioactivity of cellular G A G did not decrease in the L M cells in the first 24 h of the post-labelling incubation period. In contrast, nearly a 50 per cent decrease occurred in the H M cells during the same period. However , in the pericellular fraction the decrease in G A G labelling was more rapid (84 per cent in 5 h), especially in the L M cells. These observations suggest that the bulk of increasing extracellular GAG-associa ted radioactivity originates f rom the peri- cellular compar tment . Considering the similarly high elimination rate of G A G - associated radioactivity from the cellular +pericellular compar tment of both H M and L M cells, it was an unexpected observation that about three times less G A G was recovered in the extracellular compar tment of the L M cells than in the H M cultures. Admittedly, this was due to the differences in the G A G degradation between the two cell cultures. The data for total GAG-associa ted radioactivity in the culture (cellular + pericellular + extracellular) indicate that about 70 per cent of the G A G is degraded in 48 h in the L M cultures, but only 30 per cent in the H M cells.
D i s c u s s i o n Glycosaminoglycans are endowed with regulatory functions, both on the cell
surface and in the nucleus, owing to the variability of their chemical structures, their biological half-lives and their easy movements among various cellular compar t - ments. Thus proteoglycans are good candidates for biopolymers, contr ibuting not only to the formation of the cells's phenotype but also to the modification of gene function. In our previous studies on low and high metastatic variants of Lewis lung tumours (3LL, 3 L L - H H ) , an increased biosynthesis of G A G was observed in the highly metastatic variant [14, 31]. Further , an elevated amount of sialic acid on the cell surface, the appearance of transplantation antigens, fibronectin expression and a decrease in terminal Ga l /GalNAc content was observed in the pr imary cultures of the highly metastatic Lewis lung tumour ( 3 L L - H H ) . This was accompanied by enhanced metastasis formation [15]. These properties were implicated in the increased in vitro invasiveness and in altered in vivo heterotypic interactions of the H M cells [22]. These results indicated that the G A G metabol ism is presumably related to the increased metastatic capacity of the H M cells.
Increased cellular uptake and incorporation of [3H]glucosamine into glycoconju- gates of the cytoplasm (except glycolipids) was a typical feature of the highly metastatic cell line after a 48-h labelling (table 2). It was similar to the uptake previously observed in cells isolated f rom in vivo maintained tumour lines [31]. The higher [3H]glucosamine labelling in the cytoplasm and nucleus may be due to the enlarged precursor pools. The increase of the pools may be correlated with the facilitated t ransport of small molecules through the H M cell membranes .
The radioactivity of the purified G A G s was two and three times higher in nuclear and cytoplasm fractions, respectively, of H M cells than in those of L M cells (table 3). There was much less difference between the two cell lines in the pericellular

o~
Tab
le 4
. C
ha
ng
es o
f G
AG
rad
ioac
tivi
ty i
n t
he
cell
ula
r, p
eric
ellu
lar
and
ex
tra
cell
ula
r fr
acti
ons
of L
M a
nd
HM
cu
ltu
res
du
rin
g t
he
pos
t-
lab
elli
ng
(ch
ase)
in
cub
ati
on
tim
e R
.
Tim
e (h
)
0 0"
5 5'
0 24
"0
48"0
cpm
/#g
cp
m/#
g
cpm
/#g
cp
m/#
g
cpm
/#g
n
ucl
eic
acid
~o
nu
clei
c ac
id
~ n
ucl
eic
acid
~
nu
clei
c ac
id
~o
nu
clei
c ac
id
C
LM
7
83
+6
9
100_
+9
671_
+36
86
_+5
938_
+79
1
20
+1
0
805_
+61
1
03
__
_8
33
3-+
35
43_+
5 H
M
2463
-+32
8 10
0-+
13
2108
_+12
86
-+0-
5 22
51-+
237
91_+
10
1408
_+12
0 57
_+5
740_
+35
30
_+1
P L
M
5640
_+57
3 1
00
_+
10
1867
_+46
3
3_
__
0.8
88
5-t-
103
I6__
_2
91
2+
6
16
_+
0-1
30
9_+
35
6-t-
0.6
HM
49
04_+
618
100_
+13
21
21_+
225
43
+5
8
02
+2
3
16
+0
-5
76
5+
52
1
6+
1
724_
+10
4 15
-t-2
E
L
M
0 0
333-
+43
3
0+
4
651_
+87
58
-+8
12
26
+7
5
110_
+7
1106
_+15
6 10
0_+
14
HM
0
0 22
02_+
289
60_+
8 24
24+
__17
9 66
_+5
2931
-+32
9 8
0+
9
3652
_+22
5 10
0_+
6 T
L
M
6423
_+64
2 1
00
_+
10
2871
_+12
5 45
_+2
2474
_+26
9 39
_+4
2943
_+14
2 46
_+2
1748
_+22
6 27
_+4
HM
7
36
7+
94
6
100_
+13
64
31_+
526
87-+
7 54
77_+
439
74-+
6 51
04_+
501
69
+7
51
16_+
364
70_+
5
R T
he
GA
Gs
wer
e m
easu
red
in
tri
plic
ate
for
each
dis
h fr
om
tw
o di
ffer
ent
exp
erim
ents
. E
ach
dat
um
rep
rese
nts
mea
n +
S.E
.M.
Th
e cu
ltu
res
wer
e la
bell
ed w
ith
[3H
]glu
cosa
min
e (3
-7 x
105
Bq/
ml,
i.e.
10
#C
i/m
l) f
or 4
8 h.
Th
e la
bell
ed c
ells
wer
e w
ashe
d an
d in
cub
ated
for
a fu
rth
er 4
8 h
in th
e pr
esen
ce o
f 10
- 5
M n
on
-rad
ioac
tiv
e gl
ucos
amin
e. C
= c
ellu
lar
com
par
tmen
t; P
= p
eric
ellu
lar
com
par
tmen
t; E
= e
xtra
cell
ular
co
mp
artm
ent;
T =
tot
al (
C +
P +
E)
GA
G-a
sso
ciat
ed r
adio
acti
vity
.
g

Lewis lung tumor lines 667
fractions. The changes in the distribution pat tern of G A G s in the various cellular fractions and the changes in the amount of G A G s accumulated in the extracellular compar tment indicate a considerable shift in G A G metabolism. This shift is accompanied by the expression of the enhanced metastatic capacity of the H M cells. The association of HS and DS to the high metastatic capacity of the L L T cells is in accord with the observations indicating that HS plays an important role in the cell to cell and cell to extracellular matrix (ECM) interactions [30, 32]. I t was concluded that HS makes the at tachment of the tumour cells to host cell membranes possible, which is a crucial early step in the invasive process [5, 6, 24]. The H S proteoglycans can specifically interact with extracellular matrix constituents such as fibronectin, laminin and collagens [4, 8], so if the amount of the HS increases in the membrane , the probabil i ty of interactions between cells and extracellular matr ix (ECM) may also be facilitated. Our previous studies showed that the affinity of the 3 L L - H H cells to fibronectin and collagen type I - I I I was higher both in vitro and in vivo [15, 22]. Dermatan sulphate may also support cell adhesion and a t tachment [10, 11].
I t has been shown that sulphated G A G s were able to influence the replication and transcription of D N A by interacting with histones or by modulat ing the action of nuclear enzymes [9, 12, 13]. In addition, heparin binding to growth factors shows that it may also regulate the mechanism of cell proliferation [16, 28]. Hence the changes in the G A G levels in the nuclei of the H M cells may also have significance in the shift f rom a low to a high metastatic phenotype.
The results of our pulse chase exper iment showed that more G A G was released into the culture medium of the H M cells, originating mainly f rom the pericellular compar tment . This finding is consistent with reports that an increased shedding of surface membrane vesicles was implicated in the molecular process of the metastasis of rat m a m m a r y carcinoma and lymphomas (Eb, ESb) [3, 19, 26, 34].
The H M cells degraded more G A G in their intra- or extracellular compar tments than did the L M cells. This difference may be associated with the shift in the dominance of chondroitin sulphate to dermatan sulphate. This G A G may be less susceptible to enzymatic degradation. T h e data presented above suggest that the accumulation of HS and DS in the H M cells and their increased shedding may be related to the highly metastatic phenotype of 3 L L cells and that these alterations may contribute to the formation of an ' invasion enhancing' microenvironment .
Acknowledgements We are grateful to Ms. J. Olfih for excellent technical assistance. This work was
supported by grants f rom the Hungar ian Ministry of Heal th (Drug action on metastatic formation of human and murine tumours; p rog ramme leader Prof. K. Lapis) and by the Hungarian Academy of Sciences (grant No. 85/1-084 to Prof. K. Lapis).
References [1] ANGELLO, J. C., DANIELSON, K. G., ANDERSON, L. W., and HosIcK, H. L., 1982,
Glycosaminoglycan synthesis by subpopulations of epithelial cells from a mammary adenocarcinoma. Cancer Research, 42, 2207-2210.
[2] CONSTANTOPOULOS, G., McCoMB, R. D., and DEKABAN, A. S., 1976, Neurochemistry of the mucopolysaccharidoses: brain glycosaminoglycans in normal and four types of mucopolysaccharidoses. Journal of Neurochemistry, 26, 901-908.

668 G. Pog~ny et al.
[3] DOLJANSKI, F., 1982, Cell surface shedding. The Glycoconjugates, Vol. 4, edited by M. I. Horowitz (New York: Academic Press), pp. 155-187.
[4] FRANSSON, L., CARLSTEDT, I., and COSTER, L., and MALMSTROM, A., 1986, The functions of the heparan sulphate proteoglycans. Functions of the Proteoglycans, Ciba Foundation Symposium 124, edited by D. Evered and J. Whelan (Chichester: Wiley), pp. 125-142.
[5] GALLAGHER, J. T., LYON, M., and STEWARD, W. P., 1986, Structure and function of heparan sulphate proteoglycans. Biochemical Journal, 263, 313-325.
[6] H66K, M., KJELLt~N, L., JOHANSSON, S., and ROBINSON, J., 1984, Cell surface glycosaminoglycans. Annual Reviews of Biochemistry, 53, 847-869.
[7] Iozzo, R. V., 1985, Biology of disease. Proteoglycans: structure, function and role in neoplasia. Laboratory Investigation, 53, 373-396.
[8] IRIMURA, T., HESTER, J. E., YAMORI, T., BELLONI, P. N., FIDLER, I. J., OTA, D. M., and NICOLSON, G. L., 1987, Adhesive properties of human colon carcinoma cells established from primary tumor and metastases. Proceedings of 78th Annual Meeting of AACR, 28, 73.
[9] ISHII, K., KATASE, A., ANDOH, T., and SENO, N., 1982, Inhibition oftopoisomerase I by heparin. Biochemical and Biophysical Research Communications, 104, 541-547.
[10] KITTLICK, P. D., 1985, Glycosaminoglycans. Recent biochemical results in the fields of growth and inflammation. Experimental Pathology, Supplement 10, edited by F. Block, H. David, D. De Paola, C. C. Harriss, R. Hembry, B. A. Lapin, K. Lapis, U. Mohr, Sh. Takayama, G. Waldmann and H. Wrba, (Jena: VEB Gustav Fischer Verlag), pp. 39~5.
[11] KJELLI~N, L., OLDBERG, A., and H66K, M., 1980, Cell surface heparan sulphate. Journal of Biological Chemistry, 255, 10407-10413.
[12] KRAEMER, R. J., and COFFEY, D. S., 1970, The interaction of natural and synthetic polyanions with mammalian nuclei. I. DNA synthesis. Biochimica et Biophysiea Acta, 224, 553-567.
[13] KRAEMER, R. J., and COFFEY, D. S., 1970, The interaction of natural and synthetic polyanions with mammalian nuclei. I I. Nuclear swelling. Biochimica et Biophysica Acta, 224, 713-717.
[14] LAPIS, K., TIMAR, J., PAL, K., JENEY, A., TIMAR, F., and KOPPER, L., 1987, Membrane properties of Lewis lung tumor cells with low and high metastatic capacity. Antimetasta- tic effect of a glycosaminoglycan biosynthesis blocking agent 5-hexyl-2'-deoxyuridine (HUdR). Cancer Biology and Therapeutics. Proceedings of the 1st Annual H. Lee Moffitt International Symposium, edited by J. G. Cory and A. Szentivfinyi (New York: Plenum Press), pp. 79-94.
[15] LAPIS, K., TIM.~R, J., TIM.~R, F., PAL, K., and KOPPER, L., 1985, Differences in cell surface characteristics of poorly and highly metastatic Lewis lung tumour variants. Biochemistry and Molecular Genetics of Cancer Metastasis. Proceedings of the N I H Symposium, Bethesda, USA, edited by K. Lapis, L. A. Liotta and A. S. Rabson (Boston: Martinus Nijhoff), pp. 225-235.
[16] MACIAG, T., MEHLMAN, T., FRIESEL, R., and SCHREIBER, A. B., 1984, Heparin binds endothelial cell growth factor, the principal endothelial cell mitogen in bovine brain. Science, 225, 932-935.
[17] MOCZAR, E., LAURENT, M., and COURTOIS, Y., 1981, Effect of retinal growth factor and sulfated glycosaminoglycans of bovine lens epithelial cells. Biochimica et Biophysica Acta, 675, 132-139.
[18] NEWTON, D. J., SCOTT, J. E., and WHITEMAN, P., 1974, The estimation of acid glycosaminoglycan-Alcian Blue complexes eluted from electrophoretic strrips. Analyt- ical Biochemistry, 62, 268-273.
[19] NICOLSON, G. L., 1984, Cell surface molecules and tumour metastasis. Regulation of metastasic phenotypic diversity. Experimental Cell Research, 150, 3-22.
[20] OHYA, T., and KANEKO, Y., 1970, Novel hyaluronidase from Streptomyces. Biochimica et Biophysica Acta, 198, 607-609.
[21] PAL, K., KOPPER, L., and LAPtS, K., 1983, Increased metastatic capacity of Lewis lung tumour cells by in vivo selection procedure. Invasion and Metastasis, 3, 174-182.
[22] P~.L, K., KOPPER, L., TIMAR, J., RAJNAI, J., and LAPIS, K., 1985, Comparative study on Lewis lung tumour lines with low and high metastatic capacity. I. Growth rate, morphology and resistance to host defence. Invasion and Metastasis, 5, 159-169.

Lewis lung tumor lines 669
[23] RODI~N, L., BAKER, J. L., CIFONELLI, J. A., and MATHEWS, M. B., 1974, Isolation and characterisation of connective tissue polysaccharides. Methods in Enzymology, 28, 73- 140.
[24] ROLLINS, B. J., CATHCART, M. K., and CULP, L. A., 1982, Fibronectin-proteoglycan binding as the molecular basis for fibroblast adhesion to extracellular matrix. The Glycoeonjugates, Vol. 3, edited by M. I. Horowitz (New York: Academic Press), pp. 289- 329.
[25] SAITO, H., YAMAGATA, T., and SUZUKI, S., 1969, Enzymatic assay of chondroitin sulphates. Journal of Biological Chemistry, 243, 1536-1542.
[26] SCHIRRMACHER, V., and BARZ, D., 1986, Characterization of cellular and extracellular plasma membrane vesicles from a low metastatic lymphoma (Eb) and its high metastatic variant (ESb): inhibitory capacity in cell-cell interaction systems. Biochimica et Biophysica Acta, 860, 236-242.
[27] SCOTT, F. J., FRaCCASTORO, A. P., and TAFT, E. B., 1956, Studies in histochemistry: I. Determination of nucleic acids in microgram amounts of tissue. Journal of Histochemis- try and Cytochemistry, 4, 1-10.
[28] SHING, Y., FOLKMAN, J., SULLIVAN, R., BUTTEREIELO, C., MURRAY, J., and KLAGSBRUN, M., 1984, I-teparin affinity: purification of a tumor-derived capillary endothelial cell growth factor. Science, 223, 1296-1299.
[29] SHIVLEY, J. E., and CONRAO, H. E., 1976, Formation of anhydrosugars in the depolymerisation of heparan. Biochemistry, 15, 3932-3942.
[30] STECK, P. A., CHEONG, P. H., NAKAJIMA, M., LUNG, W. K. A., MOSER, R. P., and NICOLSON, G. L., 1987, Altered expression of glycosaminoglycans in metastatic 13762NF rat mammary adenocarcinoma cells. Biochemistry, 26, 1020-1028.
[31] TIMJ~R, J., MOCZAR, E., TIMER, F., P~L, K., KOPPER, L., LAPIS, K., and JENEY, A., 1987, Comparative study on Lewis tumour lines with low and high metastatic capacity. II. Cytochemical and biochemical evidence for differences in glycosaminoglycans. Clinical and Experimental Metastasis, 5, 79-87.
[32] TURLEu E. A., and TRETIAK, M., 1985, Glycosaminoglycan production by murine memaloma variants in vivo and in vitro. Cancer Research, 45, 5098-5105.
[33] WASTESON, A., UTHNE, K., and WESTERMARK, B., 1973, A novel assay for the biosynthesis of sulphated polysaccharide and its application to studies on the effects of Somatomedin on cultured cells. Biochemical Journal, 136, 1069-1074.
[34] YOGEESWARAN, G., 1983, Cell surface glycolipids and glycoproteins in malignant transformation. Advances in Cancer Research, 38, 289-350.


















![C.S. Lewis. Adaptacje życia i twórczości [C.S. Lewis: adaptations of life and work]](https://static.fdokumen.com/doc/165x107/631b31c4665120b3330b4035/cs-lewis-adaptacje-zycia-i-tworczosci-cs-lewis-adaptations-of-life-and.jpg)

