
Comparative study of shape, course, and disintegration of the rostral notochord in some vertebrates,...
22
Abstract The rostral part of the notochord reveals many pecularities compared with the trunk mesoderm. Further- more, its role in head formation and inductive processes in the head is not as well understood as the interaction of the trunk notochord with the spinal cord and somites. To interpret experimental and molecular biological examin- ations in the developing head region, exact knowledge about morphological features of the rostral notochord is fundamental. Here we show that the rostral notochord re- veals variations that depend on species and individual. We describe morphological characteristics of the rostral (head) notochord in human embryos (Carnegie stages X—XIV), which are shown in semithin sections and three-dimensional graphic reconstructions. Special atten- tion is paid to the relationship of the notochord with the prechordal mesoderm and the adenohypophysis. We pro- pose that in the human the rostral notochordal tip termi- nates at Rathke’s pouch, whereas in the chick prechordal mesoderm is found inbetween the notochordal tip and the anlage of the adenohypophysis. The behaviour of the notochord at the end of the embryonic period proper and early fetal time is shown in sagittal histological sections of 16 to 49 mm CRL human embryos. Position and dis- integration of the rostral notochord is also described in embryos of cat (8–25 mm), mouse (stage 21–24 accord- ing to Theiler) and chicken (stage 22–26 HH). A synop- sis reveals the different course of the notochord within, at the inner or outer side of the basioccipital cartilage. The course of rostral notochord is determined by its at- tachment points at the hypophysis, the pharynx or the footplate of the brain. In all species, it has an undulating course. Its rostral tip is highly coiled, and fragments or splinters are found within the anlage of the dorsum sell- ae. Thus, we have reasons to believe that the adenohy- pophysis is a hindrance for the rostrad elongation of the notochord. Variable adhesions between notochord and pharyngeal epithelium are considered to be responsible for invaginations of the pharyngeal wall forming bursae pharyngeae. In contrast to other authors, we observed in the mouse that rostrally the notochord bends ventrad and penetrates the chondrocranium at the level of the later synchondrosis basisphenoidale to build a bursa pharyng- ea. Finally, partial duplications of two human notochords are described. Key words Rostral notochord · Disintegration · Basichondrocranium · Human embryos · Vertebrate embryos Abbreviations in figures Ao Aorta · AH adenohypophysis · At atlas · BA basilar artery, · Bm buccopharyngeal membrane · BO basioccipital · BP bursa pharyngea · BS basisphenoidal · DA dens axis · DS dorsum sellae · E ectoderm · En endoderm · Fg foregut · Fgt foregut tip · H heart · Hy hypophysis · In infundibulum · M mandibula · N neural tube · NC notochord · NH neurohypophysis · Npu nucleus pulposus · NP neuropore · Nr neural ridge · Ph pharynx · PM prechordal mesoderm · RP Rathke’s pouch · SP Seessel’s pouch · V anastomosis of left and right Vena primaria capitis · Ve vertebra Introduction The notochord first appears during gastrulation as a cra- nial elongation of the primitive streak or Hensen‘s node (Spratt 1942; Rosenquist 1966; Nicolet 1970; Jurand 1974; O’Rahilly and Müller 1981; Bellairs 1986; Selleck and Stern 1991; Sausedo and Schönwolf 1993; Calata et al. 1996; Psychoyos and Stern 1996). In all vertebrates, it extends from the dorsal area of the hypophysis to the final extremity of the tail, forming a stiff rod that thus ensures a constant length of the body (Starck 1979). This supporting function of the notochord K. Barteczko · M. Jacob ( ✉ ) Abteilung für Anatomie und Embryologie, Institut für Anatomie, Ruhr-Universität Bochum, Universitätsstrasse 150, D-44780 Bochum, Germany e-mail: [email protected] Tel.: +49-234-700-5213, Fax: +49-234-709-4339 Anat Embryol (1999) 200:345–366 © Springer-Verlag 1999 ORIGINAL ARTICLE Klaus Barteczko · Monika Jacob Comparative study of shape, course, and disintegration of the rostral notochord in some vertebrates, especially humans Accepted: 27 April 1999
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Comparative study of shape, course, and disintegration of the rostral notochord in some vertebrates,...

Abstract The rostral part of the notochord reveals manypecularities compared with the trunk mesoderm. Further-more, its role in head formation and inductive processesin the head is not as well understood as the interaction ofthe trunk notochord with the spinal cord and somites. Tointerpret experimental and molecular biological examin-ations in the developing head region, exact knowledgeabout morphological features of the rostral notochord isfundamental. Here we show that the rostral notochord re-veals variations that depend on species and individual.We describe morphological characteristics of the rostral(head) notochord in human embryos (Carnegie stagesX—XIV), which are shown in semithin sections andthree-dimensional graphic reconstructions. Special atten-tion is paid to the relationship of the notochord with theprechordal mesoderm and the adenohypophysis. We pro-pose that in the human the rostral notochordal tip termi-nates at Rathke’s pouch, whereas in the chick prechordalmesoderm is found inbetween the notochordal tip andthe anlage of the adenohypophysis. The behaviour of thenotochord at the end of the embryonic period proper andearly fetal time is shown in sagittal histological sectionsof 16 to 49 mm CRL human embryos. Position and dis-integration of the rostral notochord is also described inembryos of cat (8–25 mm), mouse (stage 21–24 accord-ing to Theiler) and chicken (stage 22–26 HH). A synop-sis reveals the different course of the notochord within,at the inner or outer side of the basioccipital cartilage.The course of rostral notochord is determined by its at-tachment points at the hypophysis, the pharynx or thefootplate of the brain. In all species, it has an undulatingcourse. Its rostral tip is highly coiled, and fragments orsplinters are found within the anlage of the dorsum sell-ae. Thus, we have reasons to believe that the adenohy-pophysis is a hindrance for the rostrad elongation of the
notochord. Variable adhesions between notochord andpharyngeal epithelium are considered to be responsiblefor invaginations of the pharyngeal wall forming bursaepharyngeae. In contrast to other authors, we observed inthe mouse that rostrally the notochord bends ventrad andpenetrates the chondrocranium at the level of the latersynchondrosis basisphenoidale to build a bursa pharyng-ea. Finally, partial duplications of two human notochordsare described.
Key words Rostral notochord · Disintegration ·Basichondrocranium · Human embryos ·Vertebrate embryos
Abbreviations in figures Ao Aorta ·AH adenohypophysis · At atlas · BA basilar artery, · Bm buccopharyngeal membrane · BO basioccipital ·BP bursa pharyngea · BS basisphenoidal · DA dens axis ·DS dorsum sellae · E ectoderm · En endoderm ·Fg foregut · Fgt foregut tip · H heart · Hy hypophysis ·In infundibulum · M mandibula · N neural tube ·NC notochord · NH neurohypophysis · Npu nucleuspulposus · NP neuropore · Nr neural ridge · Ph pharynx ·PM prechordal mesoderm · RP Rathke’s pouch ·SP Seessel’s pouch · V anastomosis of left and right Venaprimaria capitis · Ve vertebra
Introduction
The notochord first appears during gastrulation as a cra-nial elongation of the primitive streak or Hensen‘s node(Spratt 1942; Rosenquist 1966; Nicolet 1970; Jurand1974; O’Rahilly and Müller 1981; Bellairs 1986; Selleckand Stern 1991; Sausedo and Schönwolf 1993; Calata etal. 1996; Psychoyos and Stern 1996).
In all vertebrates, it extends from the dorsal area ofthe hypophysis to the final extremity of the tail, forminga stiff rod that thus ensures a constant length of the body(Starck 1979). This supporting function of the notochord
K. Barteczko · M. Jacob (✉)Abteilung für Anatomie und Embryologie, Institut für Anatomie,Ruhr-Universität Bochum, Universitätsstrasse 150,D-44780 Bochum, Germanye-mail: [email protected].: +49-234-700-5213, Fax: +49-234-709-4339
Anat Embryol (1999) 200:345–366 © Springer-Verlag 1999
O R I G I N A L A RT I C L E
Klaus Barteczko · Monika Jacob
Comparative study of shape, course, and disintegrationof the rostral notochord in some vertebrates, especially humans
Accepted: 27 April 1999

is especially important for lower vertebrates. In Acrania,e.g. in the cephalochordate Amphioxus (Amphioxuslanzeolatus), which indeed lacks a hypophysis, the noto-chord extends to the tip of the rostrum (Theiler 1974).
In higher vertebrates, including man, the supportingfunction is taken over by the spinal column during onto-genesis. Only the cores of the intervertebral discs, thenuclei pulposi, are original remnants of the involutingnotochord and reminders of this transitory leading struc-ture (e.g. Christ and Wilting 1992).
Since the beginning of this century, the developmentof the notochord has been described by many scientists(e.g. von Bardeleben 1900; Bardeen 1919; Studnicka1926; Lehmann 1938; Jurand 1962; Bancroft and Bel-lairs 1976; Müller and O’Rahilly 1986, 1988; Sulik et al.1994; Ghanean et al. 1996).
The formation and composition of the notochordalsheath was particularly investigated by Mookerjee (1953),Carlson and Low (1971), Frederickson and Low (1971),Carlson (1973), Bancroft and Bellairs (1976), Carlson andKenney (1980), Götz et al. (1995). The phenomenon ofsegmental flexures of the notochord was discussed byBaur (1967), Verbout (1971), Adams et al. (1990).
However, the notochord does not only function astemporary axial skeleton but also operates as an inducerfor neural tube formation (this so-called primary induc-tion goes back to the classic experiments of Spemannand Mangold 1924) and in dorsoventral patterning of theneural tube. With regard to chick embryos, such an effectof the notochord on the differentiation of the floor platearea was demonstrated by van Straaten et al. (1988).Similarly, the notochord plays a crucial role in determingthe ventral compartment of the somite (reviewed byChrist et al. 1998), via the signal molecule sonic hedge-hog (SHH), which induces the expression of the Pax-1gene in the sclerotome preceding chondrogenesis of ver-tebrae (Brand-Saberi et al. 1993; Koseki et al. 1993;Ebensperger et al. 1995).
The development of the chondocranium and the for-mation of the skull has also been the subject of many in-vestigations and discussions (e.g. Levi 1900; Gaupp1901; Kernan 1916, Lewis 1920; Macklin 1921; de Beer1937; Doskocil and Jelinek 1976; Grube and Reinbach1976; Gans and Northcutt 1983; Hinrichsen 1990; Coulyet al. 1993; Le Douarin et al. 1993). Yet the organisationof the head and thus the skull is still not completely un-derstood and there even remain ambiguities about ametameric pattern in the head (for discussion see Jacobet al. 1984; Wachtler et al. 1986).
In Amphioxus, the notochord develops along thewhole neuraxis, whereas a prechordal ‘‘new head‘‘ hasbeen established during evolution, characterized by mes-enchyme deriving not only from paraxial mesoderm butfrom two other sources: neural crest and prechordal me-soderm (Gans and Northcutt 1983; Couly et al. 1993;Seifert 1995; Seifert et al. 1993a, b, 1995). The border ofthe skeleton, which derives from paraxial head meso-derm including occipital somites, is located at the transi-tion between the basipre- and basipostsphenoid bones
and is identical with the tip of the notochord (Le Douarinet al. 1993).
There is general agreement that the rostral part of thenotochord has other properties than the more posteriorpart, and after the works of Froriep (1882, 1894) specialattention has been paid to it.
The anteriormost notochord has been found to induce,e.g., expression of the homeobox gene engrailed 2,which defines the midbrain-hindbrain boundary (Hem-mati Brivalou et al. 1990). According to Darnell et al.(1992), however, notochordless chick embryos have anormal overall pattern of chick En-2 protein expression.Furthermore, Gardner and Barald (1992) found En-protein to appear in the “prechordal plate” and anteriornotochord only some time after its expression in themes-metencephalic neuroepithelium above. Rostralwardto this border (in contrast to the hindbrain and trunk)other than the Hox genes are expressed along the antero-posterior axis (for review see Kuratani et al. 1997).
Moreover, a special inductive influence of the rostraltip of the notochord on development and growth of the pi-tuitary was postulated by Eyal Gilady (1958). More re-cently Hermesz et al. (1996) found a homeobox gene Rpxas the earliest marker for the pituitary primordia, and thisgene is progressively activated in the prechordal plate, an-terior notochord and Rathke’s pouch. This implies that theprechordal region is also involved in inductive processesof the pituitary, which was already postulated by Ham-mond (1974) based on experimental studies of chicks.
Doskocil (1970a, b), on the other hand, emphasized thetopographical relationship of the notochord and individualparts of the hypophysis. The development of cranial flex-ure (Jacobson et al. 1979, 1982; Pikalow et al. 1994),which takes place in the mesencephalic region around therostral tip of the notochord, is in close relation with theformation of the adenohypophysis (Rathke’s pouch).
To interpret the results of experimental and molecularbiological investigations in the developing head regionexact knowledge about morphogenesis of the differentstructures is fundamental. In the case of the notochord,an evaluation of the significance of the notochord in themorphogenesis of the head may be a problem since thecourse and morphological features of the rostral noto-chord reveal many variations that depend on species andindividual, as will be shown in this paper. Furthermore,in the head region abnormal structures like chordomaand the bursa pharyngea (Slipka 1971, 1974) often ap-pear. We describe morphological features, the course(position) and disintegration of the rostral notochord inearly human embryos as well as in embryos of cat,mouse and chicken.
Materials and methods
The micrographs, graphic detail reconstructions and semi-schematic drawings, are based on light-microscopic investigationof more than 100 serially sectioned embryos of man, cat, mouseand chicken. As many as forty of there were human embryosstaged at the CC (Carnegie collection) staging system as intro-
346

duced by O’Rahilly and Müller (1987), which is based on hori-zons of Streeter. Thirty embryos with characteristic, species-spe-cific features of the rostral part of their notochord were selectedfor detailed analysis and are listed in Tables 1–4. Most of the em-bryos are from our own collection (Institute of Anatomy and Em-bryology, Ruhr University Bochum, established by Prof. K. Hin-richsen). Some of the human embryos (Table 1) were photo-graphed and examined microscopically by courtesy of Prof. G.Steding, who allowed access to his collection. The serially sec-tioned embryos were stained with hematoxylin and eosin, azan ortrichrome (Masson).
Reconstructions were made from the rostral part of the noto-chord of young human embryos at stages X CC (9 somites), XICC, XII CC (4.5 mm), XIII CC (5.5 mm) and XIV (6.8 mm). Forthe reconstructions, the required slices were directly projected on-to transparent paper, using a transparency, projecting apparatus.The series of tracings were arranged in blocks of ten slices. Theheight of the block was calculated from the product of slice thick-ness, number of slices and magnification. To ensure that theblocks are fitted to each other, prominent contours were used asfixation points. The blocks formed in this manner were arrangedor graduated side by side to form the definitive design.
Results
Morphological characteristics of the rostral (head)notochord in human embryos, Carnegie stages X–XIV
In the stage X embryo, the semicircular notochord, stillintracalated in the roof of the foregut (Fig. 1a 4, b), cra-nially changes into the broader notochordal plate with ir-regularly arranged cells (Fig. 1a 2, 3, b). The notochord-al plate is laterally delimited from the endoderm of theforegut. At the summit of the foregut, it passes over intoa mixed tissue in which distinction between mesodermaland endodermal cells is not possible. This tissue dispers-es at its cranialmost point but turns ventrally around thetip of the foregut down to the anlage of Rathke’s pouch.
A mid-sagittal section through the cranial part of amore advanced stage X embryo (Fig. 1d; 3,4 mm) showsa bent notochord separated from the foregut but withsome adhesion points. The situation is similar to the earlystage XI embryo shown in Fig. 2a–c. The rostral neuro-pore has not yet closed, as seen in the micrograph (Fig.
347
Table 1 Human embryos evaluated in this study listed by stage of development (Carnegie stage system after Streeter (1951), O’Rahillyand Müller (1987)
Figure Protocol Stage CC CRL Fixative Direction of Thickness Feature Collection/No. (mm) sectioning of sections Sponsor
1d 1954-04-09 X 3.4 – Sagittal 10 µm Prof. G. Steding a1a,b,c ME Esch X – – Horizontal 10 µm 9 somites Prof. G. Steding a2a,b,c 95-1-27 XI – – Sagittal Prof. G. Steding a3a–f ME 64 XII 4.5 Immers. GA Sagittal Semithin 2 pharyng.arches Own b4a–h ME 50 XII 4.5 GA/PA Sacch. Frontal Semithin 20–30 Own b5a,b ME 18 XIII 5.5 GA FA Para. Frontal Semithin Own b6a–c ME 52 XIV 6.8 J.F. Transverse Semithin Own b9a S. 403 XVIII 15 Formol Serra Frontal 8–10 µm Double notochord Own b8a,b S. 226 XVIII 16 Bouin Sagittal 10 µm Own b8c S. 388 XIX 19 Serra Sagittal 8 µm Own b8d S. 504 XXIII 26 Serra cold Sagittal 7 µm Own b8e, f S. 508 S. 542 Foetus 49 Serra Sagittal 8 µm Own b9b ‘‘Bodo” Foetus 50 Formol Transverse 7 µm Double notochord Own b
a Prof. Dr. med. G. Steding, Abt. f. Anatomie, Zentrum Anatomie des Fachbereichs Medizin, Georg-August-Universität, Kreuzbergring 36,D-37075 Göttingenb Sammlung der Abt. f. Anatomie u. Embryologie, Inst. f. Anatomie, Medizinische Fakultät der Ruhr-Universität, Universitätsstraße 150,D-44801 Bochum, Gründer der Sammlung: Prof. Dr. med. K. Hinrichsen (†1997)
Table 2 Laboratory animals used in this study. Embryos of cat (Felis domesticus; d.p.c. days post conception)
Figure Protocol Age a CRL Fixative Direction of Thickness of Collection/SponsorNo. d.p.c. (mm) sectioning sections
10 c S. 517 c 18 8 Serra Sagittal 10 µm Dr. H. Fromm b10 d,a S. 517 d 21 10 Serra Sagittal 10 µm Dr. H. Fromm b10 e S. 511 22 10.5 Serra Sagittal 10 µm Dr. H. Fromm b10 f S. 517 e 23 12 Serra Sagittal 10 µm Dr. H. Fromm b10 h S. 512 24 15 Serra Sagittal 10 µm Dr. H. Fromm b10 g S. 517 f 24 15 Serra Sagittal 10 µm Dr. H. Fromm b10 b,i S. 517 g 25 18 Serra Sagittal 10 µm Dr. H. Fromm b11 a,b,c S. 3 26 20 Serra Frontal 10 µm Dr. H. Fromm b10 j S. 517 h 28 25 Serra Sagittal 10 µm Dr. H. Fromm b
a After Rüsse I, Sinowatz F (1991) eds. Lehrbuch der Embryologie der Haustiere. Tab. 8.10, S. 202—203. Parey, Berlin Hamburgb Dr. med. vet. H. Fromm, D-58452 Witten

2a), but the oropharyngeal membrane has ruptured. Thepartial sagittal reconstruction (Fig. 2b) reveals a curvednotochord completely separated from the foregut endo-derm. It does not exactly follow the line of the foregut butis more undulated, with some protrusions. The medial-most block (the left in Fig. 2c) shows a close connectionbetween the rostral part of the notochord and the thinpharyngeal endoderm. At the summit of the foregut, theendoderm becomes thicker and curves ventrally and endsat the remnants of the oropharyngeal membrane. The ros-tral tip of the notochord continues with a loosely arrangedprechordal mesoderm that lies over the thickened foregutendoderm. Prechordal mesoderm cells have already mi-grated laterally, as shown in the serial units (Fig. 1c). Theprechordal mesoderm forms wing-like condensationssimilar to those described by Jacob et al. (1984) and Sei-fert et al. (1993b, 1995) in 2-day chick embryos.
In the stage XII embryo (Fig. 3a), both neuroporeshave closed, two pharyngeal arches can be distinguished,and the budding of limbs has started. The typical differ-entiation of notochord has occurred (Fig. 3b): enclosureby a basal lamina, vacuolization and translucent appear-ance of the cytoplasm. The closely adhering cells form acylindrical rod all over the trunk and neck region. Thecranialmost part, however, is formed irregularly: bendingventrally and broadening laterally. The reconstruction(Fig. 3g, h) and the corresponding histological sections(Fig. 3b–f) illustrate that the rostral notochord thus gets ahammer-head-like appearance. The bending of the noto-chord coincides with the cranial flexure and occurs at themesenchephalic level (Fig. 3g). Except for the close con-nections with the pharyngeal endoderm at some points inthe angle zone of the cranial flexure, it is separated fromthe endoderm by loosely arranged mesenchyme. Rem-nants of the oropharyngeal membrane form a very prom-inent ridge at the pharyngeal roof. Yet, a vein anastomo-sis (between Vv. capitis prim.) hinders adhesions of thenotochordal tip with this ridge (Fig. 3b–f) and, rostro-ventrally, in close relationship with this anastomosis anectodermal pouch appears adjacent to the ridge of the
pharyngeal roof, seemingly the first indication of the an-lage of the adenohypophysis.
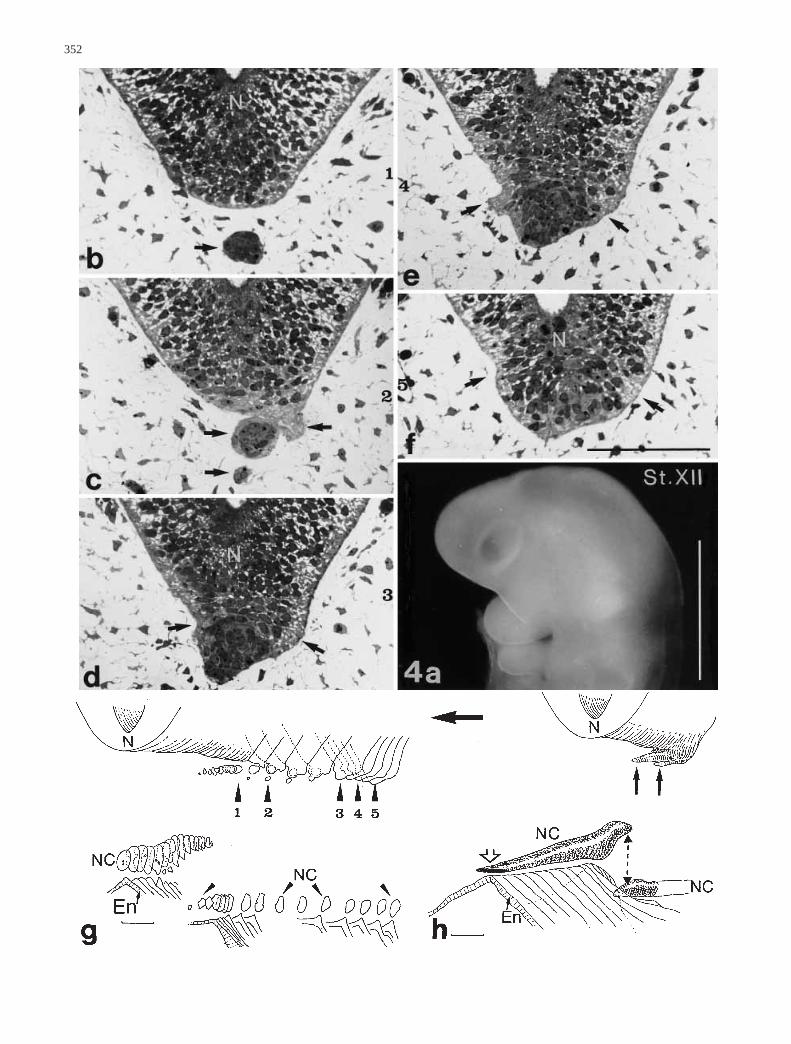
Another early stage XII CC embryo (Fig. 4a), with twodistinct pharyngeal arches reveals some pecularities of thenotochord and neural tube in the angle where the cervicalflexure develops. The rostral part of the notochord is stillin close relationship with the foregut endoderm, which hasformed a furrow in the midline where the notochord ad-heres (Fig. 4g, serial reconstruction; 4 h plastic recon-struction). There is a break in the notochord a short dis-tance more caudalwards, and this point coincides with aprotuberance extending rostroventrally from the floor ofthe neural tube (Fig. 4b–g). The subsequent part of the no-tochord is cast away and lies more ventrally, thus forminga depression in the dorsal pharynx wall. A horn-like protu-berance of the floor of the hindbrain comes out from afloorplate region like a prolongation of the rostral part ofthe notochord. It consists of irregularly arranged cells thatbulge the basal lamina and form a serrated outline of thispart of the neural tube (Fig. 4b–e). The cornu also con-tains cell detritus. These special configurations have notyet been described in human embryos. Considering thegraphic reconstruction, it seems that shearing forces haveworked in a similar way to geological faults.
In the stage XIII CC embryo (Fig. 5a), special atten-tion is paid to the relationship between notochord, brainanlage, Seessel’s and Rathke’s pouch.
The reconstruction (insert in Fig. 5a) is based on seri-al frontal sections and exhibits the irregular shape and
348
Fig. 1 a Detailed reconstruction of the human embryo Esch, stageX CC in four blocks, from cranial (1) to caudal (4). Note the tran-sition zone between neural ectoderm and superificial ectoderm(↔), the presumptive region of Rathke’s pouch and the roof of theforegut with the prechordal mesoderm (**). The rostral tip of thenotochordal plate (*) lies within the dorsal wall of the foregut. Bar0.1 mm. b, c Same embryo as in a. Transverse sections throughthe buccopharyngeal membrane (IIIII) and the notochordal plate(*) corresponding to 2 and 4 in a. Bar 0.01 mm. d Human embryo1954-04-09, late stage X CC. Sagittal section through the cranialnotochord tissue. Note the delicate connections (open arrow) withpharynx endoderm. Bar 0.1 mm
Table 4 Embryos of chicken (Gallus domesticus/White leghorn)
Figure Protocol Stage a Age (day) Fixative Direction of Thickness of CollectionHH sectioning sections
13 a, b N 2207 22 3.5 Bouin Sagittal 7 µm Own13 c N 2603 26 5 Bouin Sagittal 7 µm Own
a After Hamburger V and Hamilton HL (1951) A series of normal stages in the development of the chick embryo. J Morphol 88:49–92
Table 3 Embryos of mouse (Mus musculus)
Figure Protocoll-Nr. Stage a Age (day) Fixative Direction of Thickness of CollectionTheiler sectioning sections
12 c H 86/8 21 10–13 Serra Sagittal 10 µm Own12 a,b H 82/8 24 15–16 SerraGendre Sagittal 10 µm Own
a After Theiler K. (1972) ed. The house mouse. Berlin Heidelberg New York

349

350
Fig. 2a–c Human embryo 95-1-17, stage XI CC. SEM micro-graph of the head, ventral view. Note: the buccopharyngealmembrane (arrow) is already ruptured. b Detailed reconstructionof cranial notochord drawn after the sagittal sections shown in c.c Drawings depicting the position of prechordal mesoderm andnotochord in consecutive sagittal sections. Short arrows bucco-pharyngeal membrane. The big arrow shows ventral direction.Bar 0.1 mm
Fig. 3a–h Human embryo ME 64, stage XII CC. a Macrographof the whole embryo seen from the right side. Note the anlage ofheart and umbilical cord (arrows). Bar 1 mm. b–f Sagittal semi-thin sections through notochord cranial region, revealing tenderconnections (open arrow) with endoderm and with anastomosisbetween left and right Vena prim. cap. This remarkable anasto-mosis is situated between the notochord and a very prominentridge-shaped buccopharyngeal membrane remnant (*). Bar 0.1mm. g Detailed reconstruction of the notochord. ‘‘Hammerhead‘‘-like tip, remnant of buccopharyngeal membrane (*). Inset dia-gram (sagittal section) of whole head region. Bar 0.1 mm. h Dia-gram of the notochordal tip, cranial (c), frontal (f) and medial (m)view
▲

351
352

notochord, its penetration of the basioccipital cartilage andthe entrance into the basisphenoidal cartilage. However,they only pay little attention to the behaviour of the rostraltip of the notochord and they do not show the morphologyof this special part of the notochord. An identical course ofthe rostral notochord, as described by Müller andO’Rahilly (1980) in stage XXIII embryos, is also found inour stage XVIII (16 mm) embryo shown in Fig. 8a. Bypenetrating the chondrogenic anlage of the basioccipitale itturns ventrally from the apex of the dens axis and runswithin the ventral side of the developing perichondrium. Itlies parallel to the pharynx but is now separated from it byloose mesenchyme. At the anterior border of the basiocci-pital cartilage, it turns dorsally in a curved line pointed to-wards the anlage of the adenhypophysis but now separatedfrom it by the chondrogenic blastema of the basispenoidalskeleton. The higher magnification (Fig. 8b) reveals awell-preserved notochord with a hook-like extremity.
In a slightly older embryo (stage XIX, 19 mm), frag-ments of the notochord, which are linked at intervalswith the pharynx wall, can be seen in the midsagittal sec-tion (Fig. 8c).
At the end of the embryonic period (stage XXIII,26 mm: Fig. 8d), annularly arranged segments of the ros-tral tip of the notochord are found within the cartilage ofthe dorsum sellae.
More advanced regression of the anterior part of thenotochord is seen in the 49 mm fetus (Fig. 8e, f). A well-developed dorsum sellae separates degenerating fragmentsof the notochord from the pituitary gland. They lie withinthe fissure between basioccipital and basiphenoidal. Ven-trally, slivers of notochord can be seen to be in close con-tact with the pharyngeal wall. At each attachment point,the pharyngeal epithelium is thickened and/or reveals in-vaginations that may be considered as anlagen of the pha-ryngeal bursa according to Huber (1912), Marin-Padilla(1979), Müller and O’Rahilly (1980), Christ et al. (1988).
Fig. 9 shows another peculiarity of the notochord:doubling of the notochord is found in the cervico-occipi-tal transition zone of a stage XVIII embryo (Fig. 9a).The 50 mm fetus (Fig. 9b) exhibits duplication of the no-tochord within a vertebra.
The notochord in cat embryos of 8 to 25 mm
The strange position of the notochord within the base ofthe skull, observed in human embryos, is not a commonfeature in the vertebrates. In cats e.g. (Fig. 10a, 10 mm,10b, 18 mm), the notochord always runs straight aheadfrom the apex of the dens axis to the adenohypophysis. Itnever leaves its position dorsal to the skeleton anlageand lies in the perichondrium near the outer lamella ofthe dura mater. In the 10 mm embryo (Fig. 10a, d), anundulated course can be seen, whereas in the 18 mm em-bryo it lies a little ventrally to the perichondrium withinthe cartilage and keeps a more straight course.
Details of the behaviour of the rostral tip of the noto-chord are presented in Fig. 10c–j. In the youngest em-
353
coiled course of the rostral notochord. It lies over Sees-sel’s pouch and ends ventrally with a bifurcation at thebase of Rathke’s pouch. Whereas the notochord hascompletely lost its connection with the brain anlage, thebasement membrane at the rostral wall of the pouch is inintimate contact with the infundibular area of the dien-cephalon, as described by Müller and O’Rahilly (1988).
In the stage XIV embryo, the course and shape of therostral notochord do not change a lot (Fig. 6a–c). Again,the rostral tip is U-shaped or annulated and ends imme-diately at Rathke’s pouch. The undulated course of thenotochord is even more distinct. At the notochordal tip,short branches are directed towards the mesencephalonat an acute angle. Similar prolongations of the notochordpointing towards the apex of the mesencephalic flexureare also schematically shown in stage XII embryos byMüller and O’Rahilly (1987).
The semithin section of stage XII and XIV embryos(Figs. 5b, 6a, b) exhibit a well developed notochordalbasal lamina but not a notochordal sheath, which accord-ing to Shinohara and Tanaka (1988) appears in Stage XIVhuman embryos and may be detected by metachromacy.
Figure 7 summarizes the behaviour of the rostral noto-chord in stage XII–XIV embryos. Its extreme tip alwaysends near the remnants of the oropharyngeal membrane atthe dorsal wall of Rathke’s pouch. The outgrowing adeno-hypophysis more and more separates the tip of the noto-chord from the infundibular region of the forebrain, towhich it is closely related in the early stage. The noto-chord, which has a regular oval or round shape in the trunkregion, here reveals a very irregular form, with intumes-cences, constrictions and intersections. In correlation withthe formation of the cranial flexure, which according toMüller and O’Rahilly (1988) diminishes from an averageof 127° in stage X to 53° in stage XIII, the notochordal tip,at first bent at a right angle, acquires a more and morewavy, even a serrated course. Thus, during pronouncedventral flexion, the only fixed point seems to be the adhe-sion of the notochord to the dorsalmost part of the adeno-hypophyseal anlage. In humans, intimate contact of thehead notochord with the endoderm of the foregut persists,with the exception of the angular mesencephalic region.
The behaviour of the rostral notochord at the end of theembryonic period proper and in the early fetus
A careful analysis of the human chondrocranium at the endof the embryonic period was performed by Müller andO’Rahilly (1980), and we will refer to it for details of theskeleton. These scientists also describe the course of the
Fig. 4a–h Human embryo ME 50, stage XII CC. a Macrographfrom the left side. Bar 1 mm. b–f Frontal semithin sections of noto-chord dislocation and pointed neural tube outgrowth (arrows), bothin the same cranial region. Bar 0.1 mm. g Diagrams of sequence sec-tions, 1–5 corresponding to b–f. h Reconstruction of whole noto-chordal rostral tip (open arrow) and part of the neural tube with out-growth (black arrows) in the zone of dislocation (broken line witharrowheads). The big arrow points in ventral direction. Bar 0.1 mm

354
Fig. 5a, b Human embryo ME18, stage XIII CC. a Macro-graph of the right half of thehead region with projectiondrawing (inside) of Rathke’spouch, Seessel’s pouch, the no-tochord and the neural tube.Bar 1 mm. b Semithin trans-verse section through the rostralnotochord region. For total re-construction see inset above.Note the broad notochordal tiptouching Rathke’s pouch. Bar0.1 mm

355
Fig. 6a–c Human embryo ME 52, stage XIV CC. a, b Transversesemithin sections through splinters of the notochord and bifurcat-ing of the notochordal tip at Rathke’s pouch. Bar 0.01 mm. c
Block reconstruction diagram of the rostral notochord (see alsoFig. 7c). Bar 0.1 mm
Fig. 7a–c Three-dimensionaldetailed reconstructions of no-tochordal tip in three early hu-man embryos at stages XII CCto XIV CC to compare the dif-ferent shape of the rostral noto-chord in embryos of 4.5 mm(stage XII) with 6.8 mm (stageXIV) crown-rump length.a Embryo at stage XII CC.Sagittal views of the bisectednotochord. Remnant of bucco-pharyngeal membrane (arrow-heads). b Embryo at stage XIIICC, frontally sectioned. Noto-chord connecting Rathke’spouch. c Embryo at stage XIVCC. Note the splinters of thenotochord tip. Curved arrow tothe right: a segment with singleand carved fragments is shown.Asterisks Aa. carotis int. Longarrows point ventrally, shortarrows caudally. Bars 0.1 mm
▲

356

357
bryo (8 mm, Fig. 10c), a well-developed notochord at-taching to the anlage of adenohypophysis is shown. Dur-ing further development, the notochord meanders. Espe-cially, the extreme tip is highly coiled (Fig. 10e, g, h)and is separated from the adenohypophysis by con-densed mesenchyme between the adenohypophysis andthe dura mater. By rearing up, the notochord presses onthe dura mater, which lines the inner side of the skullskeleton (Fig. 10h, j). Fragmentation of the notochordaltip becomes obvious in the 18 mm embryo (Fig. 10i).These fragments are found between the dorsum sellaeand the mesenchyme condensation at the apex of thedorsum. Frontal serial sections of a 20 mm embryo re-veal the relationship between the notochordal tip and thismesenchymal condensation. More dorsally (Fig. 11a),the notochord is near the dura, then impresses the peri-chondrium (Fig. 11b) and losting volume, it ends withinthe dense mesenchyme (Fig. 11c) so that it will eventual-ly form the apex of the dorsum sellae or a ligament ad-hering to it.
Behavior of the rostral notochord in 13- to 16-day mouseembryos (stage 21–24 after Theiler)
With regard to this species, the course of the rostral partof the notochord exhibits another variation. As in cats, itlies at the inner (dorsal) side of the basioccipital cartilage(Fig. 12a). Rostrally, however, it turns towards the duralamella. After reaching it, the notochord bends ventrad,penetrates the skeleton of the skull base at the level ofthe later synchondrosis basisphenoidale (Fig. 12b). Itcomes in contact with the dorsal wall of the pharynx,there inducing the formation of a bursa pharyngea. Frag-ments of notochord are also found in the vicinity of thecapillary stem of the hypophysis (Fig. 12c).
The notochord in stage 22 HH to stage 26 HH chickembryos
In the chick embryo the notochord is much thicker thanin the other species described here. In contrast to them,the notochord runs in the middle of the basioccipitalecartilage.
In the youngest embryo (stage 22 HH), the skull skel-eton has not yet developed. The rostral notochord ischaracterized by large, vacuolar cells and is surroundedby a well-developed notochordal sheath (Fig. 13a). It iscontinuous with the prechordal mesoderm as analysed indetail by Seifert (1995), Seifert et al. (1993a, b, 1995).As already described by Doskocil (1970a), it is bent likea ‘‘walking-stick handle‘‘ and turns towards Rathke’spouch. There is no direct connection to the anlage of theadenohypophysis, only a junction formed by a strand ofmesenchymal cells.
Fig. 8 Sagittal histological sections through human embryos of16 mm (a, b), 19 mm (c), 26 mm (d) and 49 mm (e, f). Bars0.1 mm. a Embryo S. 226, stage XVIII CC. Note notochord coursein basioccipital and behaviour of its cranial tip (higher magnifica-tion in b) in the region of dorsum sellae. c Embryo S. 388, stageXIX CC. Notochord splinters near the pharyngeal roof in the mid-dle of basioccipital inducing a bursa pharyngea. d Embryo S. 504,stage XXIII CC. Common type of arrangement of notochord splin-ters in the dorsum sellae. e Fetus S. 542, 49 mm CRL. Involutingnotochord between basioccipital and dorsum sellae. Arrows spointto anlagen of bursae pharyngeae near notochord splinters. Highermagnification in f
Fig. 9a, b Two human embry-os with double notochord.a Embryo S. 403, stage XVIIICC. Micrograph of the cranialhead region near the rostral tipof the notochord (arrows dou-ble notochord). b Fetus ‘‘Bo-do‘‘, 50 mm CRL. Frontal his-tological section in the upperneck region. Distance of thecomplete duplication approxi-mately 70 µm (arrows noto-chord tissue with advanced in-volution, arrowheads noto-chord sheath, asterisks spacebetween notochord remnantsand sheath). Bars 0.1 mm

358

359
In the stage 26 HH embryo, the notochord is found inthe centrum of the precartilaginous anlage of the basioc-cipital, and it does not change this position up to its in-volution. In the middle of the basioccipital anlage, thismesenchymal condensation has separated the notochordfrom invaginations of the pharyngeal epithelium, seem-ingly a bursa pharyngea that has been induced by specialcontact with the notochord (Fig. 13b). At the extremetip, dissolution starts and fragments of notochord arefound within the mesenchymal condensation, whichmight indicate the formation of sella turcica (Fig. 13c).
Based on light- and electronmicroscopic investiga-tions on young chick embryos, the behaviour of the ros-tral notochord and subsequent prechordal mesoderm isschematically represented during the formation of Rath-ke’s pouch and the establishment of the cranial flexure(Fig. 14a, b). Between the vault of the pouch and the no-tochord, a tight connection always exists, mediated by
the prechordal mesoderm. While the pouch deepens andthe flexure increases, the ventral wall of the aden-ohypophysal anlage is in close contact with the foregut,as is the dorsal wall with the forebrain. The notochordand prechordal mesoderm become more and morecurved and lose their junction with the foregut. Onlysome extensions of the foregut epithelium remain of thisclose relation (Fig. 14b).
Synopsis of notochordal behaviour in four species:humans, cats, mice, chicken
For better comparison, the course and relationships ofthe rostral notochord are schematically represented inFig. 15a–d. The drawings reveal the different course ofthe notochord within or at the inner or outer side of thebasioccipital cartilage. Despite these differences, in allcases it ends near the tip of the adenohypophysis. In laterstages, however, the notochord has been separated fromthe hypophysis by the cartilaginous or mesenchymal an-lage of the dorsum sellae. The notochord always has anundulating course and is coiled at it extreme tip. Thus, itappears that the pitutiary or its cartilaginous capsule arean obstacle to the growth of the notochord. This barrierforces the notochord to deviate since it is encapsulatedby the stiff sheath. The border between ‘‘old”=chordaland ‘‘new”=prechordal head is assumed to be in the cen-trum of the sella turcica. A straightened notochord wouldend at the anterior border of the basipresphenoidal carti-lage, at least in humans (Fig. 16).
Discussion
The rostral (head) part of the notochord exhibits manypeculiarities compared with that in the trunk region.
First, in mammals (in contrast to birds) the notochordis temporarly integrated into the roof of the foregut as
Fig. 10a–j Sagittal histological sections through cat embryos of 8mm to 25 mm CRL. Note that in cat the notochord always runsunder the inner perichondrium of basioccipital. a, d Embryo S.517 d, 10 mm CRL with undulated notochord at the inner side ofbasioccipital (arrows). b i Embryo S. 517 g, 18 mm CRL. In b en-trance (1) and exit (2) of notochord under the perichondrium. Therostral notochord tip appears to be pressed against a mesenchymalcondensation (*) in a trigonum built of hypophysis, dura (open ar-rows) and dorsum sellae. c Embryo S. 517 c, 8 mm CRL. Noto-chord runs straight on and ends at adenohypophysis. e EmbryoS. 511, 10.5 mm CRL. Splintering of notochord. f Embryo S. 517e, 12 mm CRL. Splinter of the notochord rearing against outer du-ra lamina. g Embryo S. 517 f, 15 mm CRL, h Embryo S. 512,15 mm CRL. Both embryos reveal a rearing-up of the notochordtip before ending at the mesenchymal condensation. j EmbryoS. 517 h, 25 mm CRL. Notochord indenting on the dura. Bars0.1 mm
Fig.11a–c Frontal serial sections through cat embryo S. 3, 20 mmCRL. After pressing up the dura against the brain (a), the noto-chordal tip dives down into the mesenchymal condensation* (b, c).Bars 0.1 mm

360
notochordal plate (e.g. Jurand 174; Müller and O’Rahilly1983, 1985). This relationship to the pharynx epitheliumexhibits individual- and species-specific variations.
Second, whereas its origin from Hensen’s node (seeIntroduction) and its belonging to the mesoderm layer isgenerally accepted, the development at its extreme tip isfar less clear. Here the notochord, distinguishable byvacuolized cells and the notochordal sheath, is contin-ued with the prechordal mesoderm. The latter may notonly derive from Hensen’s node like the notochord butalso from the prechordal plate, an endodermal tissuewithin the roof of the foregut as described in chicken by
Seifert (1995) and Seifert et al. (1992a, b; 1993a, b;1995).
Third, the exact relationship of the rostral notochordand/or prechordal mesoderm to the adenohypophysis andtheir role in the formation of Rathke’s pouch and cranialflexure remains unsettled (see for discussion Doskocil1970a).
Finally, what is role of the notochord in the develop-ment of the base of the skull? Although the notochordhas been extensively examined for more than a hundredyears, details of the morphological behaviour of the ros-tral notochord remain nevertheless obscure. Its compli-cated morphology is hard to understand from a two-di-mensional view only. For that reason we here present forthe first time 3-dimensional reconstructions of the headnotochord during early stages of human development.
In the stage X CC human embryo, the notochord isstill intercalated within the roof of the foregut but re-mains always clearly delimited from it. Rostrally, how-ever, it is completely integrated into the thickened endo-derm of the foregut. According to the investigation of
Fig.12a–c Species-specific course and disintegration of the crani-al notochord in mouse embryos at 13 and 16 days gestation.a Embryo 82/8, 16 day. Total course of the notochord from densaxis. Note deviation of the notochord tip: after running in the di-rection of the mesencephalic flexure, it penetrates basioccipital toconnect pharynx epithelium (open arrow dura, asterisk dorsumsellae). Big arrow points in ventral direction. b Higher magnifica-tion of rectangle in a. c Embryo 86/8, 13 day. Notochord tip splin-ters near adenohypophysis. Bars 0.1 mm

361
Fig. 13 Sagittal sections through chick embryos at stage 22 HH(a, b) and 26 HH (c). a Rostral notochord with species-specificsolid shape bending against adenohypophysis. Note former con-nection with Rathke’s pouch (arrows). b Same embryo, middle
part of head notochord near pharynx epithelium. Splinter inducingthe anlage of bursa pharyngea. c Notochord tip and splinter nearadenohypophysis and infundibulum. Bars 0.1 mm
Fig. 14a, b Schematic dia-grams showing origin and de-velopment of Rathke’s pouchand its role in cranial flexure.Chick embryo stage 8 to. 14 HH(** buccopharyngeal mem-brane). a Note points of an-choring between the notochord,the prechordal mesoderm andRathke’s pouch during forwardshifting of forebrain as indicat-ed by arrows. Full circle tightconnection between prechordalmesoderm and Rathke’s pouch(hinge point). b Three-dimen-sional reconstruction of noto-chord cranial tip of stage 14HH chick embryo to show theconnection of notochord andRathke’s pouch by the medialpart of the prechordal meso-derm, thus the deepening ofRathke’s pouch is enabled.Wings of prechordal mesodermexpand laterally. Inset Diagramof a median section of stage14 HH chick embryo to showcranial flexure. Bar 0.5 mm

362
Seifert et al. (1993a, b) in chick, and Müller andO’Rahilly (1983) in human embryos, this intermingledtissue corresponds to the prechordal plate, which turnsventrally around the foregut to stop at the oropharyngealmembrane – an ovoid, mesoderm-free region that isfound dorsally to the anlage of the heart (It is notablethat in many textbooks of embryology the oropharyngealmembrane is called the prechordal plate!). From ‘‘our”endodermal prechordal plate cells may detach, contribut-ing to the head mesenchyme. Thus, the prechordal meso-derm in the human could derive, as in chicken (for dis-cussion see Seifert et al. 1993a, b), from the foregut andthe notochord.
At a slightly older stage, this prechordal mesodermdevelops as a strand of loosely arranged mesoderm thatinterconnects notochordal tip and prechordal plate. Theinitially axial prechordal mesoderm disperses laterallyand ventrally and forms wing-like mesenchymal conden-sations. The premandibular head cavities eventually ap-pear within these condensations (Adelmann 1922, 1926).Morphological and experimental studies in chick embry-os have shown that the external eye muscles derive fromthis tissue (Jacob et al. 1984; Wachtler et al. 1984, 1986).Later, Couly et al. (1992) proved that the muscles inner-vated by the oculomotorius nerve derive from the pre-chordal mesoderm.
We also found premandibular head cavities in humanembryos of stage XII and XIII, the youngest of 3–4 mmlength (not shown). This is in line with examinations ofGilbert (1957) who showed head cavities within preman-dibular condensations. These condensation are not ho-mologous with somites as discussed by Jacob et al.(1986) concerning chick embryos.
Fig. 15a–d Course of the notochord in basioccipital and its rela-tion to hypophysis, oropharynx roof and dura. A comparative syn-opsis from four species: man (a), cat (b), mouse (c), and chicken(d). Diagrams after original sagittal sections. Note the deviationsand splinters of notochords and anlagen of bursa pharyngea
Fig. 16 Schematic drawing of boundaries between ‘‘old” headand ‘‘new head” in the region of sella turcica. Median sectionview in consideration of findings after de Beer (1937), Gans andNorthcutt (1983), and Le Douarin et al. (1993); a old (chordal)head of mesodermal origin, b rostral part of basioccipital, c caudalpart of basispenoidal, d new (achordal) head of ectoderm (neural
crest) origin, e sella turcica, f basiprespenoidal, g basipostsphenoi-dal, h oblique midline of sella turcica, front of the old head, i dor-sum of sella turcica with proc. clinoideus post., j later synchron-drosis sphenooccipitalis, k notochord, pointed line supposed de-velopmental direction of the notochord after ‘‘straightening”, bigarrow points rostrally)

According to Müller and O’Rahilly (1994) the noto-chord completes development in stage XI—XII and thenotochordal sheath is present in stage XIII or stage XIVrostrally. During these stages, we found the rostral partof the notochord always attached to the dorsalmost wallof the adenohypophyseal anlage. In the chick embryo,we found Rathke’s pouch to develop at a slightly earlierstage (13—14 HH, ca. 20 somites) than in humans (stageXIII CC, ca. 30 somites) and prechordal mesoderm is in-between notochordal tip and the anlage of the adenohy-pophysis.
During the deepening of Rathke’s pouch, the head ofthe embryo bends more and more ventrally and thecranial flexure becomes conspicuous e.g. Müller andO’Rahilly (1988). According to Pikalow (1994), a pre-requisite for the cranial flexion is that the length of theforegut remains constant during the growth of the brainanlage. Because the ventral wall of the diencephalon isfirmly attached to Rathke’s pouch extending from the ec-toderm adjacent to the tip of the foregut, there is a linkbetween the neural tube and the pharynx. According toPikalow (1994), it is this link that causes the growingneural tube to bend around the foregut, thus forming thecranial flexure and the ectodermal infolding (Rathke’spouch). The role of the notochord is a passive one. How-ever, the attachment of its tip to the remnants of the oro-pharyngeal membrane, as found in stage XII CC humanembryos, seems to be the fixed point during this rotation(Frazer 1941; Doskocil 1970a). Since it is at points at-tached to the foregut, it has to follow the rotation duringcranial flexure. At first, it bends rectangularly, giving thetip a walking stick-like appearance.
Later on, the head notochord becomes undulated andbifurcated at its extreme tip since elongation of the ros-tral notochord, due partly to mitosis and mainly to vacu-olization, is obstructed by the adenohypophysis, and thenotochordal sheath prevents its thickening.
From stage XIII CC, clefts and fragmentation arefound at the rostral end of the notochord, perhaps due tothe forces that work during cranial flexure or as firstsigns of the involution of the notochord. With the elon-gation of Rathke’s pouch, the fixating-point of the noto-chordal tip shifts from the remnants of the oropharyngealmembrane and its transition into stomodeum ectoderm tothe dorsal wall of the adenohypophysis.
The partial duplication of the notochord seems to be arare anomaly. Johnston (1931) describes such a case inan 11 mm embryo. We can demonstrate two of theseanomalies, one in a stage XVIII CC and another in a40–50 mm embryo. Partial duplication may occur duringthe process of excalation of the notochord from the noto-chordal plate, which might be broader than usual. Sincethe two notochords are arranged dorso-ventrally, no con-sequences in the shaping of the foot-plate or the verte-brae could be observed.
The role of the notochord in inducing sclerotome dif-ferentiation and thus chondrogenesis of vertebrae isproven without doubt (for review see Christ and Wilting1992). Consequently, the notochord forms the central ax-
is of each vertebral body in the trunk region. However, itregresses during ossification and only remnants persist inthe nuclei pulposi of the intervertebral discs.
In the basichondrocranium, the situation is more com-plicated. The basioccipital, which is derived from thefirst five somites (for discussion see Wilting et al. 1995),and the bones of paraxial cephalic mesoderm origin, likethe sphenoid (Le Douarin et al. 1993). Although the ba-sioccipital is of somitic origin its segmental prepattern isnot visible in adults anymore. This may be due to theearly downregulation of the Pax-1 gene, which apparent-ly leads to the lack of intervertebral discs and thus fusionmight occur (Wilting et al. 1995). The mechanism of thisdownregulation is still unclear. Pax-1 is induced by noto-chord and floorplate signals (Ebensperger et al. 1995;Brand-Saberi et al. 1996), and in the trunk region bothtissues develop in close vicinity to each other, only sepa-rated by the developing cartilage of the vertebral bodies.In the head region, however, the notochord is not soclosely related to the footplate of the brain anlage sinceduring cranial flexion it moves away from the neuraltube, and involution starts earlier.
In this paper, we have compared the course of thehead notochord in four species and have found varia-tions depending on species and individual that mightnot be considered malformations. An analysis of theposition of the notochord in relation to the basioccipitaland basisphenoid gathered from the literature and ownobservations is given by Marin-Padilla (1979), reveal-ing the intracranial course of the notochord to be char-acteristic for each mammalian species. Yet, here we fo-cus on the ending of the rostral notochord. In the hu-man, the notochord turns ventrally after having left thedens axis and penetrates the basioccipital to adhere tothe pharyngeal epithelium at several points. Subse-quently, it bends dorsally, lying in the fissure betweenbasioccipital and sphenoid. It ends with a coiled tip inthe anlage of the dorsum sellae. When the regression ofthe notochord starts, fragments of the notochord appear.Such slivers of the notochord are usually found in thedorsum sellae and at the middle of the basioccipitallength close to the dorsal wall of the pharynx. The rela-tionship to the pharyngeal wall reminds one of thelong-lasting integration of the notochordal anlage (no-tochordal plate, Müller and O’Rahilly 1985) into theendoderm. This close connection to the endoderm mustbe the reason for the suboccipital position of the noto-chord in man. Variable adhesion between notochordand pharyngeal epithelium are considered to be respon-sible for invaginations of the pharynx wall, thus givingrise to bursae pharyngeae, as first described by Tour-neux and Tourneux in Corning (1925; for discussionsee Christ et al. 1988 and Marin-Padilla 1979). We alsofound thickenings and invaginations of the foregutwall, but in contrast to Corning, the notochord was nev-er interrupted at the tip of such a bursa pharyngea inour specimens.
The fragments of notochord in the sella turcica anlageare remnants from the fixing-points of the notochord at
363

the adenohypohyseal anlage or later at its cartilaginouscapsule. According to investigations of Kjaer et al.(1997) in 13–14-week-old human fetuses, about 30%had visible notochordal remnants in the cranial base.They concluded that the morphological appearance ofthe notochordal remnants in the dorsum sellae is clearlyassociated with the morphology of the sella turcica.Whereas we consider an undulated or coiled course ofthe notochordal tip as a common feature not only in hu-man embryos they connect a non-straight S- or Y-shapedcourse of persisting notochordal tissues in the dorsumsellae with pathological morphology. According to theirstudies, this region seems to be susceptible to malforma-tions. These observations point out the crucial role of thesphenoidal region in head development. However, in ouropinion a coiled course of the rostral notochord may notnecessarily be associated with malformations. Only per-sisting notochordal remnants could be indicators of mal-formations. Chordoma, which may develop from suchfragments, most commonly occur within the sella tur-cica.
All investigated species reveal the potencies for bur-sae pharyngeae. In mouse, they are situated more anteri-orly, since the notochord penetrates the base of the skulldorsoventrally at the level of the later synchondrosis ba-sisphenoidale after having been reared up dorsally at thedorsum sellae. Jurand’s (1974) careful study of the de-velopment of mouse notochord showed that in 10-day-old embryos the notochord was still connected with theforegut by two layers of the basal lamina that later disap-pear. In 14-day embryos, vacuolization begins and basi-occipital cartilage condensation appear. Although thisauthor showed the dorsal position of the notochord to theprecartilage the behaviour of the extreme tip of the noto-chord was not noticed. According to Jurand, the noto-chord is a straight rod rostrally and ends about 0.1 mmfrom Rathke’s pouch. The course through the fissura ba-sisphenoidale was also not observed by Marin-Padilla(1979). Therefore, he suggested the mouse represents theunique example in which the notochord remains dorsalto and outside of the entire chondrochranium. We see apersisting attachment of the notochordal tip to the endo-derm beside the anchoring of the rostral notochord with-in the dorsum sellae.
The course of the rostral notochord in cats is similarto that in mice. Leaving the dens axis, it again lies be-neath the dorsal perichondrium of the basioccipital an-lage, at some points bending dorsally, and bulges out thedura mater. Differing from its course in mice, it ends inthe region of the clivus separated from the adenohypoph-ysis by dense mesenchyme, seemingly the anlage of theprocessus clinoideus posterior. The coiled end splits upinto fragments.
In contrast to mammals, the notochord of chicks ismuch thicker and is found centrally in the basichondro-cranium, as in the vertebral anlagen. Our findings are inline with those of Doskocil (1970a), that the notochordaltip first touches the dorsal wall of Rathke’s pouch andthan is shifted dorsally by the growing infundibulum and
neurohypophysis. We also observed that the anterior endof the notochord is connected with a narrow strip ofcells. But in contrast to Doskocil, we do not assume thatthey belong to the notochordal sheath. Moreover, it isthose cells of the prechordal mesoderm, as described byJacob et al. (1984), which connect the lateral mesenchy-mal condensations of emigrated prechordal cells with thenotochordal tip. Views on the significance of the noto-chord for the formation of Rathke’s pouch are intensive-ly discussed by Doskocil (1970a and b).
Due to its thickness, the chick rostral notochord is notas coiled as in the other species. During cranial flexion,it bends semicircularly. In the middle of the basioccipi-tal, invaginations (bursae pharyngeae) of the pharyngealepithelium are found that are only separated from the no-tochord by the precartilaginous condensations. Thus itappears that they also develop in chicken at attachmentpoints of the notochord with the pharyngeal epithelium.
At later stages, when the precartilage anlage of thedorsum sellae develops, the rostral end of the notochordgets shifted away from the pituitary gland anlage. Its tipbecomes undulated and thickens slightly.
As was shown by Doskocil and Jelinek (1976), inchick embryos with experimental exencephalia, the noto-chord either comes into contact with the pharynx or withthe floor of the brain, but the relationship of the rostralend of the notochord to the pituitary anlage was alwaysthe same as in normal embryos. In this malformation, theanterior half of the skull is shortened and tilted upwardsas a response to the brain malformations. The course ofthe notochord is now similar to those in the other verte-brates we have described.
From these observations, we draw the conclusion thatthe attachment of the notochord to Rathke’s pouch, ei-ther directly or by the prechordal mesoderm, is a com-mon feature in all vertebrates and does not alter duringcranial flexion. The elongation of the notochord due tomitosis or vacuolization is obstructed by the developingpituitary gland. The notochord is thus forced to coil or toundulate. The precartilaginous condensations, which ap-parently develop under the influence of Pax-1 inducedby the notochord (Wilting et al. 1995), pushes the noto-chord away. Whereas the notochord induces chondrogen-esis, the morphological features of the skull depend onthe morphology of the brain. The course of the noto-chord is determined by its attachment points at the hy-pophysis, pharynx or footplate of the brain. Since theseattachments vary in individuals and species, the courseof the rostral notochord shows variations too. Persistingcontacts with the dorsal wall of the pharynx might in-duce the formation of bursae pharyngeae as described bySlipka (1971, 1974). The most rostral bursae, as in mice,might be confused with rudiments of Seessel’s pouch(see Snook 1934).
Acknowledgements We especially thank Prof. Dr. G. Steding(Göttingen) who kindly provided the micrographs Figs. 1b, c, d and2a. We are grateful to Antje Jaeger and Marion Köhn for expertphotographic work and to Kristina Klar for making the semithinsections.
364

365
References
Adams D, Keller R, Koehl MAR (1990) The mechanics of noto-chord elongation, straightening and stiffening in the embryo ofXenopus laevis. Development 110:115–130
Adelmann HB (1922) The significance of the prechordal plate: aninterpretative study. Am J Ant 31:55–101
Adelmann HB (1926) The development of the premandibular headcavities and the relations of the anterior end of the notochordin the chick and robin. J Morphol Physiol 42:371–439
Brancroft M, Bellairs R (1976) The development of the notochordin the chick embryo studied by scanning and transmissionelectron microscopy. J Embryo Exp Morphol 35:383–401
Bardeen CR (1910) Die Entwicklung des Skeletts und des Binde-gewebes. In: Keibel F, Mall FB (eds) Handbuch derEntwicklungsgeschichte des Menschen, Bd. 10, Hirzel, Leipzig,pp 296–456
Bardeleben K von (1900) Skelettsystem, Kap. III, Chorda andChordascheiden. In: Merkel F, Bonnet R (eds) Ergebnisse derAnatomie und Entwicklungsgeschichte Bergmann, Wiesbaden,Bd. X, pp 24–27
Baur R (1967) Über die segmentalen Wellen der Chorda dorsalisund ihre Bedeutung. Acta Anat 66:631–638
Beer GR de (1937) The development of the vertebrate skull. In:Hamilton WJ, Boyd JD, Mossman HW (eds) Human embryol-ogy, Williams & Wilkins, Baltimore, pp 546–547
Bellairs R (1986) The primitive streak. Anat Embryol 174:1–14Brand-Saberi B, Ebensperger C, Wilting J, Balling R, Christ B
(1993) The ventralizing effect of the notochord on somite dif-ferentiation in chick embryos. Anat Embryol 188:239–245
Brand-Sabri B, Wilting J, Ebensperger C, Christ B (1996) The for-mation of somite compartments in avian embryo. Int J DevBiol 40:411–420
Calata M, Teillet M-A, Robertis EM de, Le Douarin NM (1996) Aspinal cord fate map in the avian embryo: while regressing,Hensen’s node lays down the notochord and floor plate thusjoining the spinal cord lateral walls. Development122:2599–2610
Carlson EC (1973) Intercellular connective tissue fibrils in the no-tochordal epithelium of the early chick embryo. Am J Anat136:77–90
Carlson EC, Low F (1971) The effect of hydrocortisone on extra-cellular connective tissue fibrils in the early chick embryo.Am J Anat 130:331–346
Carlson EC, Kenney MC (1980) Surface ultrastructure of the iso-lated avian notochord in vitro: the effect of the perinotochord-al sheath. Anat Rec 197:257–276
Christ B, Wilting J (1992) From somites to vertebral column. AnnAnat 174:23–32
Christ B, Jacob HJ, Seifert R (1988) Über die Entwicklung derzerviko-okzipitalen Übergangsregion. In: Hohmann D, Kügel-gen B, Liebig K (eds) Neuroorthopädie 4. Springer, BerlinHeidelberg New York, pp 13–22
Christ B, Schmidt C; Huang R, Wilting J, Brand-Saberi B (1998)Segmentation of the vertebrate body. Anat Embryol 197:1–8
Corning HK (1925) ed. Entwicklung des Gehirnschädels und derSinneskapseln. In: Lehrbuch der Entwicklungsgeschichte desMenschen. Bergmann, München, pp 199–203
Couly GF, Coltey PK, Le Douarin NM (1992) The developmentalfate of the cephalic mesoderm in quail-chick chimeras. Devel-opment 114:1–15
Couly GF, Coltey PM, Le Douarin NM (1993) The triple origin ofskull in higher vertebrates: a study in quail-chick chimeras.Development 117:409–429
Darnell DK, Schoenwolf GC, Ordahl CP (1992) Changes in dor-soventral but not rostrocaudal regionalisation of the chick neu-ral tube in the absence of cranial notochord, as revealed by ex-pression of Engrailed-2. Dev Dyn 193:389–396
Doskocil M (1970a) Development of the chick hypophysis. ActaUniversitatis Carolinae, Medica Monographia XL, UniversitaKarlova, Praha
Doskocil M (1970b) The mutual morphogenetic influence of thestomodeal ceiling and of the brain. Folia Morphol (Prague)18:149–160
Doskocil M, Jelinek R (1976) Development and configuration ofthe base of skull and the notochord in chick embryos with ex-perimental exencephalia. Bull Czech Zool Sol 401–87
Ebensperger C, Wilting J, Brand-Saberi B, Mizutani Y, Christ BBalling R, Koseki H (1995) Pax-1, a regulator of sclerotomedevelopment is induced by notochord and floor plate signals inavian embryos. Anat Embryol 191:297–310
Eyal-Giladi H (1958) The notochord as inductor of the orohy-pophysis in urodeles (Pleurodeles Waltii). Proc K Med Wet(Amsterdam) 61:224–234
Frederickson RG, Low FN (1971) The fine structure of perinoto-chordral microfibrils in control and enzyme-treated chic em-bryos. Am J Anat 130:347–376
Frazer F (1941) ed. A manual of embryology. Williams & Wilkins,Baltimore
Froriep A (1882) Kopfteil der Chorda dorsalis bei menschlichenEmbryonen. Beiträge zur Anatomie und Embryologie als Fest-gabe für Jacob Henle, dargeboten von seinen Schülern. Bonn,pp 26–40
Froriep A (1894) Entwicklungsgeschichte des Kopfes. In: Merkel F,Bonnet R (eds) Ergebnisse der Anatomie und Entwicklungs-geschichte, Bd. III, Bergmann, Wiesbaden, pp 391–459
Gans G, Northcutt RG (1983) Neural crest and the origin of verte-brates: a new head. Science 220:268–274
Gardner CA, Barald KF (1992) Expression patterns of engrailed-like proteins in the chick embryo. Dev Dyn 193:370–388
Gaupp E (1901) Alte Probleme and neue Arbeiten über denWirbeltierschädel. In: Merkel F, Bonnet R (eds) Ergebnisse derAnatomie und Entwicklungsgeschichte, Bd. X. Bergmann, Wies-baden. pp 847–1001
Ghanem E, Cornelissen M, Thierens H, Ridder L de (1996) Mech-anism of age-dependent involution in embryonic chick noto-chords. Experientia 52:683–686
Gilbert PW (1957) The origin and the development of the humanextrinsic ocular muscles. Contrib Embryol 36:59–78
Götz W, Kasper M, Fischer G, Herken R (1995) Intermediate fila-ment typing of the human embryonic and fetal notochord. CellTissue Res 280:455–462
Grube D, Reinbach W (1976) Das Cranium eines menschlichen Em-bryo von 80 mm Sch.-St.-Länge. Anat Embryol 149:183–208
Hammond WS (1974) Early hypophysial development in the chickembryo. Am J Anat 141:303–316
Hemmati Brivanlou A, Steward RM, Harland RM (1990) Regionspecific neural induction of an engrailed protein by anteriornotochord in Xenopus. Science 250:800–802
Hermesz E, Mackem S, Mahon KA (1996) RPX: a novel anterior-restricted homeobox gene progressively activated in the pre-chordal plate, anterior neural plate and Rathke’s pouch of themouse embryo. Development 122:41–52
Hinrichsen K (1990) ed. Humanembryologie. Springer, BerlinHeidelberg New York
Huber GC (1912) On the relation of the chorda dorsalis to the an-lage of the pharyngeal bursa or median pharyngeal recess.Anat Rec 6:373–404
Jacob M, Jacob HJ, Wachtler F, Christ B (1984) Ontogeny of avi-an extrinsic ocular muscles. Cell Tissue Res 237:549–557
Jacob M, Wachtler F, Jacob HJ, Christ B (1986) On the problem ofmetamerism in the head mesenchyme of chick embryos.NATO ASI series. Series A, Life Sciences, 118:79–89
Jacobson A, Tam PPL (1982) Cephalic neurulation in the mouse em-bryo analyzed by SEM and morphometry. Anat Rec 203:375–396
Jacobson A, Miyamoto DM, Mai S-M (1979) Rathke’s pouchmorphogenesis in the chick embryo. J Exp Zool 207:351–366
Johnston TB (1931) Partial duplication of the notochord in a hu-man embryo of 11 mm greatest length. J Anat 66:48–49
Jurand A (1962) The development of the notochord in chick em-bryos. J Embryol Exp Morphol 10:603–621
Jurand A (1974) Some aspects of the development of the noto-chord in mouse embryos. J Embryol Exp Morphol 32:1–33

366
Kernan JD Jr (1916) The chondrocranium of a 20 mm human em-bryo. J Morphol 27:605–645
Kjaer I, Becktor KB, Nolting D, Fischer Hansen B (1997) The as-sociation between prenatal sella turcica morphology and noto-chordal remnants in the dorsum sellae. J Craniofac Genet DevBiol 17:105–111
Koseki H, Wallin J, Wilting J, Mitzutani Y, Kispert A, Ebensper-ger C, Herrmann BO, Christ B, Balling R (1993) A role forPax-1 as a mediator of notochord signals during the dorso-ventral specification of vertebrae. Development 119:649–660
Kuratani S, Matsuo I, Aizawa S (1997) Developmental patterningand evolution of the mammalian viscerocranium: genetic in-sights into comparative morphology. Dev Dyn 209:139–155
Le Douarin NM, Ziller C, Couly GF (1993) Patterning of neuralcrest derivatives in the avian embryo in vivo and in vitro stud-ies. Dev Biol 159:24–49
Lehmann FE (1938) Die morphologische Rekapitulation desGrundplans bei Wirbeltierembryonen und ihre ent-wicklungsphysiologische Bedeutung. Vierteljahresschrift derNaturforschenden Gesellschaft in Zürich, 83. Jahrg., Fest-schrift Karl Hescheler Nr. 30, pp 187–192
Levi G (1900) Beitrag zum Studium der Entwicklung des knorp-eligen Primordialcraniums des Menschen. Arch Mikrosc Anat55:341–414
Lewis WH (1920) The cartilaginous skull of a human embryotwenty one millimeters in length. Contrib Embryol 9:299–324
Macklin CC (1921) The skull of a human fetus of 43 mm greatestlength. Contrib Embryol 10:57–103
Marin-Padilla M (1979) Notochordal-basichondrocranium rela-tionships: abnormalities in experimental axial skeletal (dysra-phic) disorders. J Embryol Exp Morphol 53:15–38
Mookerjee S (1953) An experimental study of the development othe notochordal sheath. J Embyol Exp Morphol 1:411–416
Müller F, O’Rahilly R (1980) The human chondrocranium at theend of the embryonic period, proper, with particular referenceto the nervous system. Am J Anat 159:33–58
Müller F, O’Rahilly R (1983) The first appearence of the major divi-sions of the human brain at stage 9. Anat Embryol 168:419–432
Müller F, O’Rahilly R (1985) The first appearance of the neuraltube and optic primordium in the human embryo at stage 10.Anat Embryol 172:157–169
Müller F, O’Rahilly R (1986) The development of the humanbrain and the closure of the rostral neuropore at stage 11. AnatEmbryol 175:205–222
Müller F, O’Rahilly R (1987) The development of the humanbrain, the closure of the caudal neuropore, the beginning ofsecondary neurulation at stage 12. Anat Embryol 176:413–430
Müller F, O’Rahilly R (1988) The development of the human brainfrom closed neural tube at stage 13. Anat Embrol 177:203–224
Müller F, O’Rahilly R (1994) Occipitocervical segmentation instaged human embryos. J Anat 185:251–258
Nicolet G (1970) Analyse autoradiographique de la location desdifférentes ébauches présomptives dans la ligne primitive del’embryon de poulet. J Embryol Exp Morphol 23:79–108
O’Rahilly R, Müller F (1981) The first appearance of the humannervous system at stage 8.Anat Embryol 163:1–13
O’Rahilly R, Müller F (1987) Development stages in human embry-os, including revision of Streeters (Horizons) and a survey of theCarnegie-Collection. Carnegie Institute, Washington, Publ 637
Pikalow AS, Flynn ME, Searls RL (1994) Development of cranialflexure and Rathke’s pouch in the chick embryo. Anat Rec238:407–414
Psychoyos D, Stern CD (1996) Fates and migratory routes in primitivestreak cells in the chick embryo. Development 122:1523–1534
Rosenquist GC (1966) Autoradiographic study of labeled grafts inthe chick blastoderm. Development from primitive-streakstages to stage 12. Contrib Embryol 38:73–110
Sausedo RA Schoenwolf GC (1993) Cell behaviors underlying no-tochord formation and extension in avian embryos: quantita-tive and immunocytochemical studies. Anat Rec 237:58–70
Seifert R (1993a) Morphological and experimental studies of theaxial head region in avian embryos. Verh Anat Ges 88 [AnatAnz, Suppl 175]:322–323
Seifert R (1993b) The avian prechordal head region: a morpholog-ical study. J Anat 183:75–89
Seifert R (1995) Die prächordale Region des Vogelembryos undihre Bedeutung für die Entwicklung der vorderen Kopfregion.Inaugural Dissertation, Medical Faculty, Ruhr-UniversitätBochum
Seifert R, Jacob HJ, Jacob M (1992a) Differentiation capabilitiesof the avian prechordal head mesoderm. NATO ASI series A,Life Sciences 231:63–76
Seifert R, Jacob HJ, Jacob HJ (1992b) Mophologische Untersuc-hungen zum prächordalen Kopfmesoderm des Vogelembryos.Verh Anat Ges 87 [Anat Anz, Suppl 174]:92
Seifert R, Barteczko K, Hinrichsen KV (1995) Studies on originand fate of the prechordal head tissues of avian embryos. VerhAnat Ges 90 [Anat Anz, Suppl 177]:118
Selleck MAJ, Stern CD (1991) Fate mapping and cell lineage ana-lysis of Hensen’s node in the chick embryo. Development112:515–626
Shinohara H, Tanaka O (1988) Development of the notochord inhuman embryos: ultrastructural, histochemical and immuno-histochemical studies. Anat Rec 220:171–178
Slipka J (1971) Early development of the bursa pharyngea. FoliaMorphol (Prague) 20:138–140
Slipka J (1974) Relationship of the notochord to development ofthe skull basis. Folia Morphol (Prague) 22:244–246
Snook T (1934) The later development of the Bursa pharyngea:homo. Anat Rec 58:303–319
Spemann H, Mangold H (1924) Über Induktion von Embryonal-anlagen durch Implantation artfremder Organisatoren. Roux’sArch Dev Biol 100:599–638
Spratt NCT Jr (1942) Location of organspecific regions and theirrelationship to the development of the primitive streak in theearly chick blastoderm. J Exp Zool 89:69–101
Starck D (1979) ed. Vergleichende Anatomie der Wirbeltiere aufevolutionsbiologischer Grundlage. Chorda dorsalis (Noto-chord). Bd II. Springer, Berlin Heidelberg New York, pp35–43
Straaten HWM van, Hekking JWM, Wiertz-Hoessels EJLM,Thors F, Drukker J (1988) Effect of the notochord on the dif-ferentiation of a floor plate area in the neural tube of the chickembryo. Anat Embryol 177:317–324
Streeter GL (1951) Developmental horizons in human embryos.Description of age groups XIX, XX, XXI, XXII, and XXIII,being the fifth issue of a survey of the Carnegie Collection.Contrib Embryol Carnegie Inst Washington 34:165–196
Studnicka FK (1926) Untersuchungen am überlebenden Gewebeder Chorda dorsalis der Wirbeltiere. Z Zellforsch MikroskAnat 3:346–376
Sulik K, Dehart DB, Inagaki T, Carson JL, Vrablic T, Gesteland K,Schoenwolf GC (1994) Morphogenesis of the murine nodeand notochordal plate. Dev Dyn 201:260–278
Theiler K (1974) Phylogenetische Entwicklung des Achselskelets.In: Sonderdruck aus Handbuch der Medizinischen RadiologieEncyclopedia of medical Radiologie. Bd VI/I. Springer, BerlinHeidelberg New York, pp 1–14
Verbout AJ (1971) Die segmentalen Wellen der Chorda dorsalis.Ein intravitales oder ein postmortales Phänomen? Z Anat Ent-wicklungsgesch 133:172–183
Wachtler F, Jacob M (1986) Origin and development of the cranialskeletal muscles. Bibliotheca anatomica. Karger, Basel29:24–46
Wachtler F, Jacob HJ, Jacob M, Christ B (1984) The extrinsic ocu-lar muscles in birds are derived from the prechordal plate. Na-turwissenschaften 71:379–380
Wilting J, Ebensperger C, Müller TS, Koseki H, Wallin J, Christ B(1995) Pax-1 in the development of the cervico-occipital tran-sitional zone. Anat Embryol 192:221–227




















