
A new settling velocity model to describe secondary sedimentation

FranceE. Kebreab, M. Schulin-Zeuthen, S. Lopez, J. Soler, R. S. Dias, C. F. M. de Lange and J.
of phosphorus utilization in growing pigsComparative evaluation of mathematical functions to describe growth and efficiency
published online June 12, 2007J ANIM SCI
http://jas.fass.org/content/early/2007/06/12/jas.2006-738.citationthe World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
by guest on May 18, 2011jas.fass.orgDownloaded from

Growth and phosphorus intake analysis in pigs1
2
Comparative evaluation of mathematical functions to describe 3
growth and efficiency of phosphorus utilization in growing pigs14
5
E. Kebreab,*†2 M. Schulin-Zeuthen,* S. Lopez,‡ J. Soler,§ R. S. Dias,* C. F. M. de6
Lange,* and J. France*7
8
*Centre for Nutrition Modelling, Department of Animal and Poultry Science, University of 9
Guelph, Guelph, N1G 2W1, Canada; †National Centre for Livestock and Environment, 10
Department of Animal Science, University of Manitoba, Winnipeg, Manitoba R3T 2N2, 11
Canada; ‡Departmento Produccion Animal, Universidad de León, 24071 Leon, Spain; §IRTA, 12
Centre de Control Porcí, 17121 Monells, Girona, Spain13
14
15
16
17
18
1This research was undertaken, in part, thanks to funding from the Canada Research Chairs19
Program20
Corresponding author: [email protected]
Page 1 of 31 Journal of Animal Science
Published Online First on June 12, 2007 as doi:10.2527/jas.2006-738 by guest on May 18, 2011jas.fass.orgDownloaded from

2
ABSTRACT: Success of pig production depends on maximizing return over feed costs and 22
addressing potential nutrient pollution to the environment. Mathematical modeling has been 23
used to describe many important aspects of inputs and outputs of pork production. This study 24
was undertaken to compare 4 mathematical functions for best fit in describing specific datasets 25
on pig growth, and to compare these 4 functions for describing P utilization for growth in a 26
separate experiment. Two datasets with growth data were used to conduct growth analysis and 27
another dataset was used for P efficiency analysis. All datasets were constructed from 28
independent trials that measured BW, age, and intake. Four growth functions representing 29
diminishing returns (monomolecular), sigmoidal with fixed point of inflection (Gompertz),30
and sigmoidal with variable point of inflection (Richards and von Bertalanffy) were used. 31
Meta-analysis of data was conducted to identify the most appropriate functions for growth and 32
P utilization. Based on Bayesian Information Criteria, the Richards equation described BW vs. 33
age data best. The additional parameter of Richards equation was necessary because the data 34
required a lower point of inflection (138 d) than the Gompertz (189 d) could accommodate,35
being fixed at 1/e times the final weight and was a limitation to accurate prediction. The 36
monomolecular equation was best at determining efficiencies of P utilization for weight gain37
compared to the sigmoidal functions. The parameter estimate for the rate constant in all 38
functions decreased as available P intake increases. Average efficiencies during different 39
stages of growth were calculated and offer insight into targeting stages where high feed40
(nutrient) input is required and when adjustments are needed to accommodate loss of 41
efficiency and reduction of potential pollution problems. It is recommended that the Richards 42
and monomolecular equations be included in future growth and nutrient efficiency analyses.43
Key words: functions, growth, phosphorus, pigs44
Page 2 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

3
INTRODUCTION45
Traditionally, growth functions have been used to describe increase in BW over time. 46
One of the outputs of generating growth curves is to have the ability to predict accurately the 47
time at which pigs will be ready for market. A useful growth function should describe data 48
well and contain biologically and physically meaningful parameters (France et al., 1996). 49
Protein accretion (Ferguson et al., 1994), BW (Bridges et al., 1992), and mineral deposition 50
(Mahan and Shields, 1998) are among attributes to be quantified using growth functions. In 51
swine nutrition, Schinckel and de Lange (1996) identified protein accretion, partitioning of 52
energy, maintenance requirement, and feed intake as primary requirements in developing 53
growth models. Growth functions can be grouped into 3 categories: those that represent 54
diminishing returns behavior (e.g., monomolecular), sigmoidal behavior with a fixed point of 55
inflection (e.g., Gompertz, logistic), and those encompassing sigmoidal behavior with a 56
flexible point of inflection (e.g., Richards, von Bertalanffy). The flexible functions are often 57
generalized models that encompass simpler models for particular values of certain additional 58
parameters. Although the Gompertz has been applied widely, other functions have been used 59
to describe growth in nonruminant animals (e.g., Darmani Kuhi et al., 2003a).60
Another application of growth curves is to relate BW to cumulative nutrient intake. It is 61
useful to examine intake in order to assess genetic improvements (Bermejo et al., 2003) or to 62
increase return over feed costs. Phosphorus is an essential nutrient that has received attention 63
for environmental reasons such as limited supply and pollution of ponds and streams (Schulin-64
Zeuthen et al., 2005). Therefore, optimizing P intake and understanding factors affecting P 65
utilization in pigs have environmental and economic benefits. Growth functions can be used to 66
determine efficiency of nutrient utilization (Darmani Kuhi et al., 2003b), which is the 67
Page 3 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

4
derivative of the relationship between BW and cumulative nutrient intake. The aim of this68
study was to evaluate 4 candidate mathematical functions for best fit in describing specific 69
datasets on pig growth, and to evaluate these 4 functions for describing P utilization for70
growth in a separate experiment.71
72
MATERIALS AND METHODS 73
Data Sources74
A total of 2,450 observations from 102 pigs originating from 3 independent datasets 75
(Table 1) was used in this study. All 3 datasets had data on individual pigs and were derived 76
from experiments conducted in Denmark, Spain, and Canada. Separate analyses were 77
undertaken to relate BW to time and BW to cumulative P intake. Datasets 1 and 2 were used 78
for growth analysis, and dataset 3 for P efficiency analysis.79
Growth analysis. Two datasets with a total of 1,120 observations from 48 pigs were used 80
to compare 4 equations to describe growth in pigs. Dataset 1 consisted of birth to maturity 81
records from 5 castrated, 5 male, and 5 female pigs. The pigs were crossbred ¼ Danish 82
Landrace, ¼ Yorkshire, and ½ Duroc (LYxD). Individual BW was recorded weekly and feed 83
intake twice a wk. All pigs received 7 different diets (based on barley and soybean) during 84
their lifespan in order to maximize growth based on Danish standards. The pigs were raised 85
between 1996 and 2001 (Schulin-Zeuthen, 2000; Schulin-Zeuthen and Danfær, 2001).86
Dataset 2 consisted of data collected from 33 Spanish grower-finisher pigs (25 to 140 87
kg) of 2 sexes (females and castrated males) with BW and feed intake recorded every third 88
week. The pigs were crossbred ¼ Large White, ¼ Landrace, and ½ Large White or Pietrain. 89
They received a grower and a finisher diet, based on corn (32.6 and 28.5 %), soybean meal 90
Page 4 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

5
(24.5 and 21 %), barley (10 and 15 %), and wheat (10 and 10 %) for grower and finisher diets, 91
respectively (Tibau et al., 2002; Jondreville et al., 2004).92
Phosphorus efficiency analysis. Dataset 3 with 1,330 observations was used to test the 93
fit of 4 functions to describe efficiency of P utilization (dW/dP) in growing pigs. An 94
experiment using 54 Yorkshire pigs (30 males and 24 females), with an initial BW of 25 kg95
conducted at the University of Guelph (Swidersky et al., 2003), was used to construct Dataset96
3. The pigs were randomly assigned to 1 of the 4 dietary P levels. The treatments were low, 97
medium, recommended, and high available P intakes corresponding to 52, 78, 105 and 130%,98
respectively, of the NRC (1998) requirements for grower and finisher pigs. Pigs had ad libitum 99
access to feed and feed intake was recorded automatically. For individual pigs housed in 100
groups, 2 diets were prepared for grower and finisher phases, based on corn (73.1 and 80%) 101
and soybean meal (24.0 and 17.3%) (Swidersky et al., 2003). The dataset was selected because 102
of the wide range of P intakes. The 4 functions can also describe Ca efficiency, as Ca was 103
varied in tandem with P because of their close association; however, diets were formulated to 104
be first-limiting in P.105
Mathematical Models106
Growth analysis. The growth equations chosen in the study represent diminishing 107
returns behavior (monomolecular), sigmoidal with a fixed point of inflection (Gompertz), and 108
sigmoidal with a variable point of inflection (von Bertalanffy and Richards) as described in 109
Chapter 5 of Thornley and France (2007).110
The monomolecular equation, the simplest equation used in this study, was used in the 111
following form with 3 parameters:112
W = Wf – (Wf – W0)e–kt,113
Page 5 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

6
where W is BW (kg) at age t (d), W0 is initial weight at birth (t = 0), Wf is asymptotic weight 114
reached as t � ∞, and k is a rate constant (d-1). The monomolecular equation describes the 115
progress of a simple, irreversible first-order reaction.116
The Gompertz equation can be derived by assuming substrate (feed) is non-limiting, the 117
quantity of growth machinery is proportional to W, and effectiveness of the growth machinery 118
decays exponentially with time according to a decay constant k (/d). The Gompertz equation 119
was used in the following form:120
))e1)(exp((ln0
0ktf
W
WWW −−= .121
Inflection in this sigmoidal growth function occurs at W = Wf /e. 122
In the von Bertalanffy equation, the assumptions are that the substrate is non-limiting123
and the growth process is the difference between anabolism and catabolism. The form of the 124
von Bertalanffy used in this study was:125
nktnnf
nf WWWW /1
0 ]e)([ −−−= , 126
where n (dimensionless, with bounds 0 < n ≤ 1/3) and k (/d) are parameters. Inflection occurs 127
at W = (1 – n)1/nWf .128
Richards equation is an empirical construct and therefore does not have the underlying 129
biological basis of the previous models. However, it belongs in the same group of classic 130
growth models and its flexibility, due to its shape parameter n (dimensionless), makes it a 131
generalized alternative to other equations (e.g., those with fixed inflection points such as the 132
Gompertz). The Richards equation was used in the following form:133
nktnnf
n
f
WWW
WWW
/100
0
)e)(( −−+= ,134
Page 6 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

7
where k (/d) is a positive constant, and n ≥ –1 in order to ensure physiological growth rates as 135
W � 0. Inflection occurs, provided n > –1, at W = Wf (1/(n + 1))1/n.136
Phosphorus efficiency analysis. Growth equations can also be used to obtain parameters 137
describing conversion of intake (e.g., feed, P, or other nutrient) to animal products such as 138
BW. Dataset 3 contains data with different levels of P and Ca intake but similar intake of other 139
nutrients, so the equations can be tested for their ability to determine values of efficiency of P 140
utilization. To describe efficiency, BW needs to be expressed in terms of intake. In this 141
analysis BW, W (kg), was regressed against cumulative intake of available P, P (g), and the 142
models described above, with age replaced by cumulative intake, were evaluated for their 143
ability to determine the efficiency of P utilization.144
The instantaneous efficiency of P utilization for gain, kP (kg BW/g P), is given by145
differentiating the W vs. P relationship: 146
P
WkP d
d= ,147
and the average efficiency of utilization of P for gain )( Pk between 2 P intake levels (P1 and 148
P2) is:149
12
d2
1
PP
Pk
k
P
P
P
P−
=∫
1212
2
1
2
1
dddd
PP
W
PP
PP
WW
W
P
P
−=
−=
∫∫)(
)(
12
12
PP
WW
−−
= .150
151
Statistical Procedures152
Growth analysis. The 4 models described were fitted to the 2 datasets combined using 153
the non-linear mixed procedure (PROC NLMIXED, Littell et al., 1996; SAS Inst., Inc., Cary, 154
NC). Mixed model analysis was chosen because the data were gathered from various studies;155
Page 7 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

8
therefore, it was necessary to consider analyzing not only fixed effects of the dependent 156
variable but also random effects (because the studies represent a random sample of a larger 157
population of studies). For this analysis, a nonlinear mixed procedure based on Craig and 158
Schinckel (2001) and Schinckel and Craig (2002) was employed. Forward selection was used 159
to find the optimum model with lowest value of Bayesian Information Criterion (BIC) 160
(Leonard and Hsu, 2001). Trial was coded as a random effect and between-pig variability was 161
modeled by introducing a parameter (c) to vary the mature BW which was also a random 162
effect (Craig and Schinckel, 2001). For example, for the monomolecular equation, the model 163
fitted was W = (c + µ)*(Wf – (Wf – W0)e–kt) where µ is the random effect of each pig. 164
Introducing sex as a fixed source of variation in the model did not improve the performance of 165
the models because most of the variability associated with differences in sex had been 166
explained by the between-pig variability parameter.167
Phosphorus efficiency analysis. Nonlinear mixed model analysis was also used in 168
determining efficiency of P utilization. First, the equations were fitted to the data and then 169
derivatives of the growth curves were calculated. In fitting the equations, fixed effects of 170
available P intake level and sex were considered. The NLMIXED procedure in SAS does not 171
support directly the concept of an independent discrete (class) variable, so the analysis was 172
performed using dummy variables to represent treatments, sex, or both. Each parameter was 173
considered individually at first and then a combination of parameters used. Model comparison 174
using an F-test showed that the rate parameter (k) was significantly affected by treatment (P < 175
0.01) and inclusion of other parameters did not improve the overall model performance. 176
Therefore, the model with separate k values but a common W0, Wf and n was fitted.177
Page 8 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

9
Distribution of random effects was assumed to be normal and the dual quasi-Newton 178
technique was used for optimization with adaptive Gaussian quadrature as the integration 179
method. Performance of the models was evaluated using significance level of the parameters 180
estimated, variance of error estimate, and its approximate SE. Comparison of models was 181
based on BIC which are model-order selection criteria centered on parsimony and imposing a 182
penalty on more complicated models for inclusion of additional parameters. Bayesian 183
Information Criterion combines the maximum likelihood (data fitting) and the choice of model 184
by penalizing the (log) maximum likelihood with a term related to model complexity as 185
follows:186
BIC = –2 log(Ĵ) + Klog (N),187
where Ĵ is the maximum likelihood, K is number of free parameters in the model and N is 188
sample size. A smaller numerical value of BIC indicates a better fit when comparing models.189
Both analyses were performed with the assumption that variance distribution for the 190
fixed factor was normal. Random effects were assumed to be normally distributed.191
192
RESULTS193
Growth Analysis194
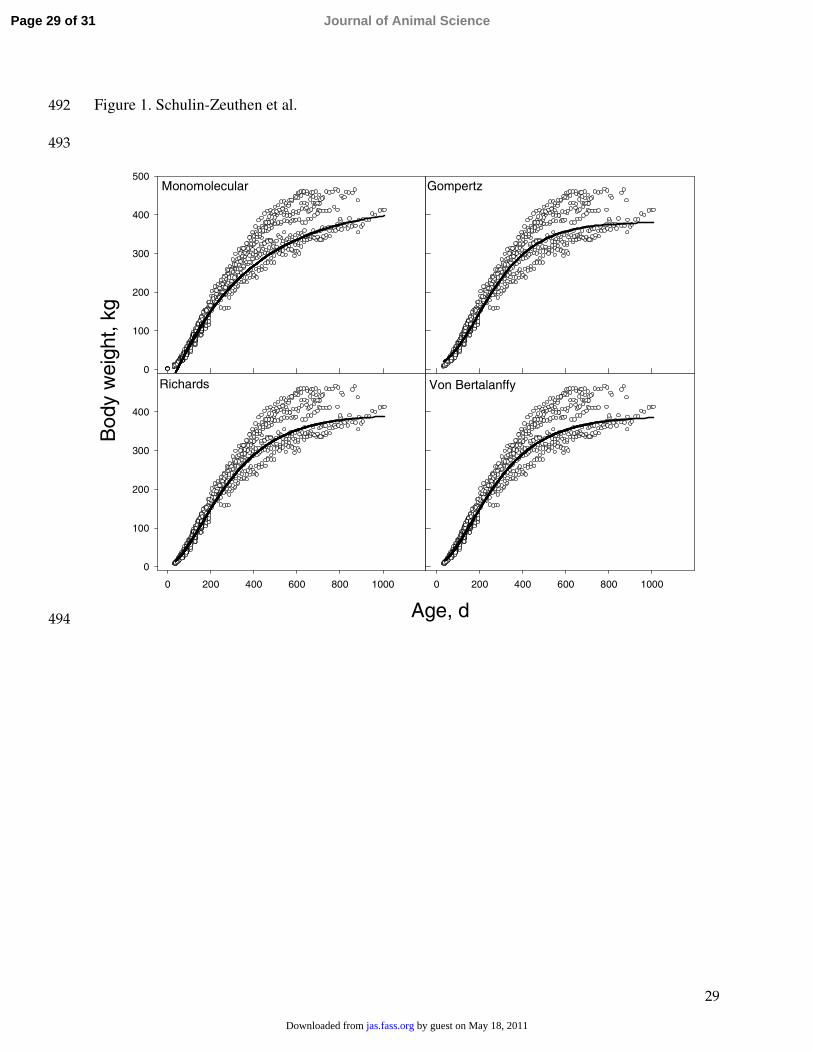
In general, all the equations fitted BW vs. age data well below approximately 350 kg 195
(Figure 1). The non-linear analysis of the pooled datasets showed that W0 estimates were close 196
to zero for all of the models compared. The estimate of W0 from the monomolecular equation197
was highest compared to the other equations. The von Bertalanffy equation estimated W0 to be 198
decreased compared to the other equations (Table 2). Estimates of Wf were within a 0.07-kg 199
range for the sigmoidal equations. More importantly, the random effect multiplier based on 200
Page 9 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

10
BW, (c), was also within a narrow range. Due to the introduction of a random effect of 201
between-pig variation, the parameter estimate of ‘mature’ pig weights (Wf) as described above 202
is not absolute. Therefore, to aid comparison between models, the estimated mature weight 203
was calculated by multiplying Wf and c (Table 2) with the monomolecular and Gompertz 204
equations showing highest and the lowest estimates, respectively. The rate constant parameter 205
estimates (k) ranged between 0.0028 and 0.0067, and were significantly different from zero for206
all models. Finally, the parameter n estimates were –0.40 and 0.33 for Richards and von207
Bertalanffy equations, respectively.208
Comparison of models based on BIC suggested an advantage in using a more complex 209
model (Richards) compared with simpler models such as the Gompertz or monomolecular. 210
The von Bertalanffy equation also appeared to improve predictions (Figure 1) compared with 211
the monomolecular equation. However, the addition of an extra parameter in the von 212
Bertalanffy equation did not result in significant improvement in goodness-of-fit with this 213
model compared with the Gompertz equation, as reflected by their similar BIC values. 214
Phosphorus Efficiency Analysis215
Cumulative available dietary P intake was calculated from feed intake measurements and 216
corresponding available P content of the diet. For a given weight, the amount of available P 217
consumed up to that particular time was entered in the database. 218
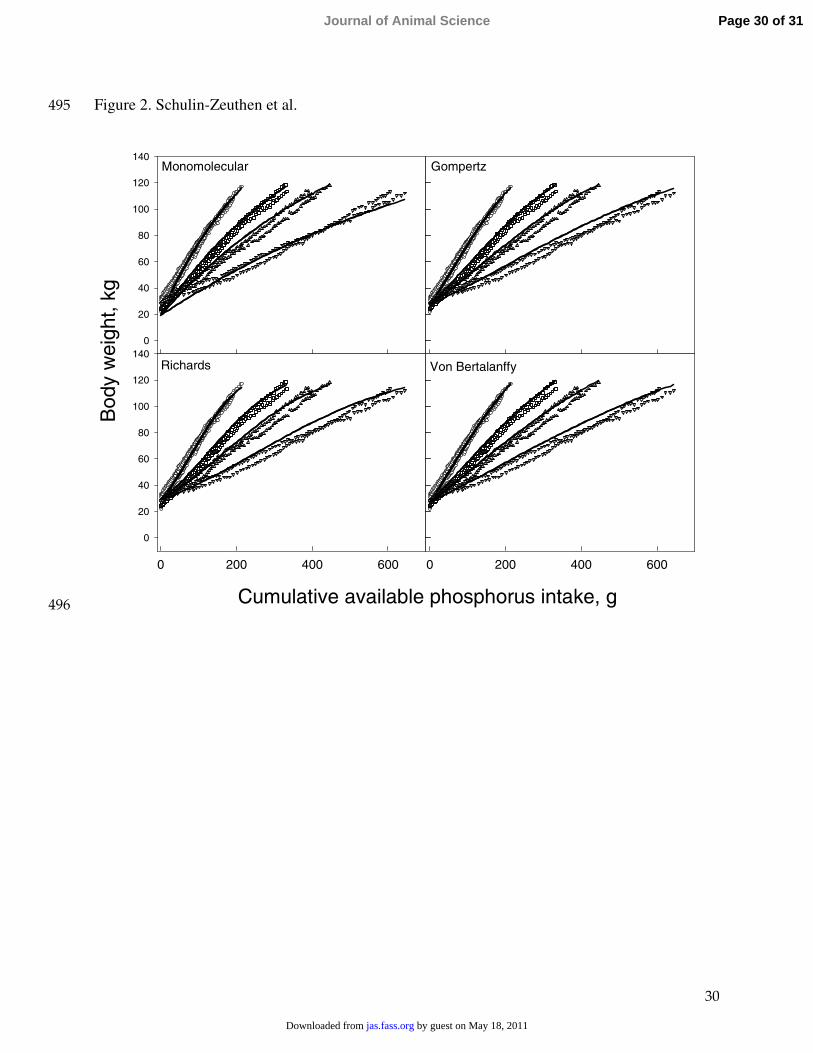
Fitting the equations to plots of BW vs. cumulative available dietary P intake (Figure 2) 219
showed that the parameter estimate for initial weight at zero P intake ranged from 19.2 for the 220
monomolecular equation to 28.7 kg for Richards equation (Table 3). Final weight estimates221
ranged from 131.9 (Richards) to 193.3 kg (monomolecular). All initial and final weight 222
estimates for all 4 models were significant parameters in the model. Rate constant estimates 223
Page 10 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

11
ranged from 0.0009 to 0.0082; n parameter estimates were –0.87 and 0.33 for the Richards and 224
von Bertalanffy equations, respectively.225
The model with separate rate constant values for each treatment category significantly 226
improved on the general model in all equations compared. However, comparisons of the 227
models using t-tests of parameter values failed to reject the null hypothesis that BW gain was 228
not significantly different between sexes (P > 0.05). Animals fed high levels of available P 229
showed the lowest estimates of rate constants, which increased as the levels of available P 230
decreased. In all models, the differences in the estimate of rate constants among dietary P 231
levels were significant (Table 3).232
Comparison of models based on BIC did not show any advantage in using a more 233
complex equation than the monomolecular (Table 3). The 3 sigmoidal equations did not do as 234
well as the monomolecular equation, but their BIC values were within a narrow range. The 235
Richards equation was marginally improved compared to the other sigmoidal equations and its 236
parameter estimates were close to those of the monomolecular equation, with a point of 237
inflection at a very small weight over the range of cumulative P intake. The Gompertz and von238
Bertalanffy equations were the least suitable for analyzing efficiency of P utilization.239
Calculations of maximum efficiency of P utilization showed wide variation between 240
models. Maximum efficiency was at a considerably lower weight for Richards (8.0 kg) than 241
for von Bertalanffy (56.5 kg) and Gompertz (57.9 kg) equations. The monomolecular equation 242
assumes the efficiency is continuously decreasing until an asymptote is reached, when 243
efficiency becomes zero.244
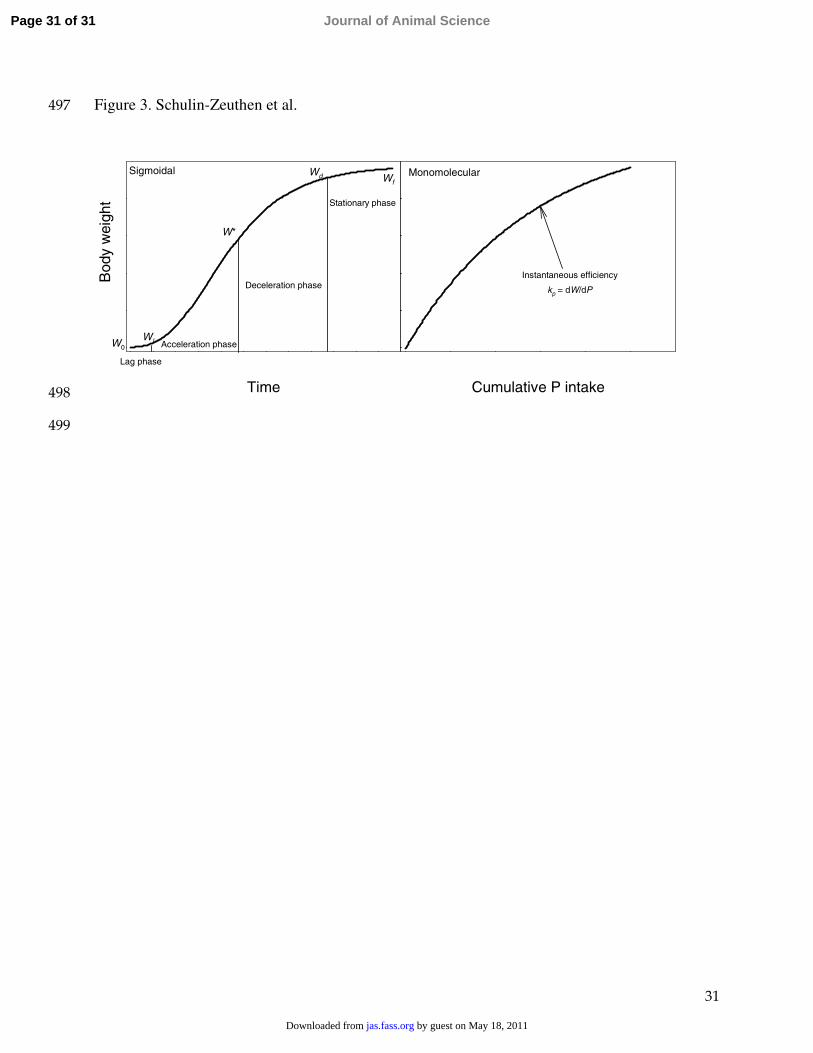
The growth curve was divided into 4 phases as shown in Figure 3. The ‘lag’ phase was 245
defined as time from the initial weight estimate until the animal attains 10% of its final weight, 246
Page 11 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

12
the ‘accelerating’ phase continues from the end of lag phase until maximum growth rate (point 247
of inflection) is achieved, then the animal goes into the ‘deceleration’ phase until it reaches248
90% of its mature weight, and finally into the ‘stationary’ phase which continues until the 249
approach of asymptotic final weight. Figure 3 also depicts graphically instantaneous efficiency 250
of P utilization taking the monomolecular equation as an example.251
Average efficiency of P utilization for gain in the stages described above is given in 252
Table 4. The calculations were made by taking into account the significant differences in the 253
rate constant. Averages from 25 to 100 kg, 25 to 50 kg, and 50 to 100 kg (weight intervals254
covering the growing-finishing periods) are also given, as these could have implications for 255
commercial production systems. As expected, the greatest average weight gain and efficiency 256
of P utilization for the monomolecular equation was during the first phase, while for the 257
Gompertz and von Bertalanffy equations, greatest weight gain was during the acceleration258
phase. For all the equations, P efficiency approaches zero in the stationary phase.259
260
DISCUSSION261
Weight gain has been described using growth equations, and in nonruminants, the 262
Gompertz has traditionally been the model of choice (Wellock et al., 2004). In this study, a 263
simpler model (monomolecular) and others without the limitation of a fixed point of inflection264
(which occurs at 1/e times the final weight in the Gompertz), such as Richards and von265
Bertalanffy equations, were evaluated. The analyses showed that, in general, all 4 growth 266
equations fitted the data reasonably well (Figures 1 and 2).267
Performance of the equations in describing the datasets was evaluated using BIC, which 268
suggested differences in model choice for growth in BW and efficiency of P utilization. A 269
Page 12 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

13
notable difference in parameter estimates in both sets of analyses was for W0. Estimates of W0270
were lower in the growth analysis because the data contained information on birth weights 271
while birth weight data were not available for the P efficiency analysis. The W0 estimates were 272
within the range of 0.042 and 9.5 kg in the first and second analyses, respectively. Wf273
estimates in growth analysis have to be looked at in conjunction with the c parameter, which 274
was within 18 kg for the sigmoidal curves. The introduction of random effect of mature weight 275
increased the goodness-of-fit (lower BIC value) compared to a generalized model with only 276
experiment as the random element in the model. It has also improved the approximate SE of 277
the parameter estimates.278
Comparison of BIC values for the growth analysis revealed that the Richards equation, 279
perhaps due to the broad limits on its additional parameter, was superior to the other 280
equations. As parameter n approaches particular values, the Richards equation encompasses 281
other simpler models, such as the monomolecular (n = –1), Gompertz (n = 0), and logistic (n = 282
1). Table 2 shows that n estimate was about halfway between the monomolecular and 283
Gompertz equations. The flexibility of the Richards equation increased the accuracy of 284
prediction, and the need for an additional parameter was justified because the shape of the 285
curve could not be described as well by either the monomolecular (because it is not a 286
sigmoidal equation) or the Gompertz (due to its fixed inflection point).287
Four phases were arbitrarily assigned to evaluate weight gain (Figure 3), and the average 288
efficiency of BW gain with time within the phases was calculated. The values based on289
generalized sigmoidal equations were close to each other but different from those based on the 290
monomolecular equation. The importance of this kind of calculation is to identify periods of 291
greatest weight gain so that inputs can be adjusted accordingly. More crucially, by recognizing 292
Page 13 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

14
the rate of decline in growth during the stationary phase, more informed decisions can be 293
made about the impact of varying slaughter weight on nutrient use and income over diet costs.294
From the Richards equation, it can be calculated that the pigs were expected to gain nearly 1 295
kg/d during the accelerating phase, which goes down by about 40% in the next phase; by the 296
time they reach the stationary phase, the animals were 97% less efficient in gaining BW. The 297
data also show that there was uniformity up to the end of the accelerating phase (d 300 of age);298
then, as animals matured, there were greater differences among animals, due to the weight 299
gains being mainly attributable to fat deposition.300
Apart from describing the pattern of growth, non-linear equations can be used to 301
investigate efficiency of utilization of individual nutrients for BW gain. Usually, mineral 302
requirements in pigs are estimated by empirical methods based on feeding a range of dietary 303
concentrations of each mineral element and examining the response in terms of certain304
performance (weight gain) or physiological (mineral balance) attributes to assess the most 305
adequate dietary concentration (i.e., the one resulting in maximum response). In general, as P 306
intake is increased, there is an increase in weight gain, feed intake, and feed efficiency (gain to 307
feed ratio); this relationship is curvilinear, such that maximum response is reached at a certain 308
level of P intake, which is variable depending upon animal characteristics (genotype, sex) and 309
the growing stage (Ketaren et al., 1993; Ekpe et al., 2002; Hastad et al., 2004). The response is 310
less pronounced as the animal grows, hence in animals of greater weight the level of P intake 311
required to reach the maximum response is lower (Ketaren et al., 1993; Hastad et al., 2004).312
From this approach, requirements have been mostly expressed as percentage of the mineral in 313
the diet (or alternatively per unit of energy in the diet) (ARC, 1981). However, in order to 314
assess optimal requirements for a given growth rate, it is important to express requirements of315
Page 14 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

15
P (and any other mineral) directly in relation to protein and fat gain, but this goal cannot be 316
achieved with the information currently available in the literature.317
Alternatively, the use of non-linear equations to describe the relationship between BW318
and cumulative available P intake can be used to estimate the efficiency of P utilization in 319
terms of weight gain per unit of available P intake at any stage of growth, provided that other 320
nutrients are not limiting expression of nutrient retention rates. Using this approach, 4 non-321
linear equations were examined. The Richards equation seemed to yield as good a fit as the322
Gompertz, given the almost identical BIC values. However, in this case, the estimate of 323
parameter n was close to –1 (Table 3), approaching diminishing returns behavior without a324
point of inflection, so that the simple monomolecular equation was adequate to describe the 325
relationship between BW and cumulative available P intake in pigs, which is confirmed by the 326
lowest BIC value of the model. Most of the rate parameter estimates of the Richards and 327
monomolecular equations were not significantly different from each other, which further 328
proves the flexibility of the Richards equation and the suitability of the monomolecular 329
equation to fit these profiles, without the need for an additional parameter. The same equation 330
(Richards) can be used for both growth analysis and efficiency of nutrient utilization, and can 331
yield different growth patterns for each of the 4 dietary P levels. Therefore, only 1 equation is 332
required for all levels to assess the optimum level.333
Calculations for efficiency of P utilization based on the best performing model 334
(monomolecular equation) showed that the average efficiency is greatest during the 2 initial 335
phases and slows down as the pig’s cumulative consumption of available P increases. By the 336
time the pig reaches the stationary phase, efficiency approaches zero. This trend is the same in 337
pigs fed low or high levels of P (Table 4). The steady decline in efficiency of P utilization 338
Page 15 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

16
observed as a pig grows agrees with the lower P requirements recommended (ARC, 1981; 339
NRC, 1998) for different growth phases (decreasing from weaning to 100 kg) and can be 340
attributed to a number of reasons. The amount of P required for maintenance is greater in341
larger pigs, and thus relatively less P is used for growth (ARC, 1981; NRC, 1998). Also, it is 342
important to examine variations in P deposition in the weight gain. Considering the principles 343
of skeletal tissue growth, there is a steady and progressive increase in the rate of P 344
accumulation per unit of fat-free BW from birth to slaughtering weight (Mahan and Shields, 345
1998). However, given the exponential increase in fat deposition in adipose tissue, the rate of 346
increment of P content in empty BW is inversely related to BW, as demonstrated by Mudd et 347
al. (1969) in trials using carcass analysis, reporting values of 8.1, 5.3, and 1.6 g P/kg weight 348
gain at 20, 50, and 90 kg of empty BW, respectively. Our analysis also shows that as BW 349
increases, the proportion of P retained decreases. The ARC (1981) suggested that the rate of 350
storage of Ca and P (g/kg weight gain) with growth could be represented by a linear fall 351
between birth and 50 kg BW (with a linear relationship between that rate and BW) and a 352
constant rate thereafter. Calculations of efficiency of P utilization as d(BW)/d(P intake) using 353
non-linear methods takes into account the relationship between BW and P as described by 354
different classical growth equations. It is noteworthy that the levels of Ca and P resulting in 355
maximum growth rate and efficiency of gain are not necessarily adequate for maximum 356
growth mineralization (ARC, 1981). To achieve maximum deposition of Ca and P, and 357
increase the bone mineral content, a greater mineral supply would be required (NRC, 1998; 358
Hastad et al., 2004). The approach suggested herein could be used not only to quantify the 359
trend in efficiency of P utilization throughout the lifespan of the animal, but also to examine 360
differences between genotypes with different lean growth rate (Bertram et al., 1994) or the 361
Page 16 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

17
effects of sex (comparison among gilts, barrows, or boars; Thomas and Kornegay, 1981; Epke 362
et al., 2002). This information could also be used to estimate P requirements in pigs at various 363
stages of the life cycle, adjusting P input levels during the finishing stages of pig production.364
Non-linear mixed analysis can give reliable parameter estimates and is a useful tool 365
when analyzing data from multiple studies. Although the Gompertz equation has been used 366
extensively, the Richards equation was superior in describing growth over time and it is 367
recommended for use in data analysis in nonruminant animals. The extra parameter in the 368
model improved the predictions and therefore was required as indicated by lower BIC values. 369
The non-linear relationship between BW and cumulative available P intake was best described 370
by the simpler monomolecular equation. Calculations of average or instantaneous efficiency of 371
P utilization can be used to predict the animals’ response to changes in intake and the potential 372
to reduce excess P excretion to the environment. It is recommended that future studies on 373
growth and nutrient intake analyses consider other models such as the monomolecular and 374
flexible equations such as the Richards as well as the Gompertz.375
Page 17 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

18
LITERATURE CITED376
377
Andersen P. E., and A. Just. 1983. Tabeller over Foderstoffers Sammensætning m.m. Kvæg. 378
Svin. Jordbrugsforlaget, DK, Copenhagen, Denmark.379
ARC. 1981. The Nutrient Requirements of Pigs. Commonwealth Agric. Bureaux, Slough, 380
U.K.381
Bermejo, J. L., R. Roehe, G. Rave, and E. Kalm. 2003. Comparison of linear and nonlinear 382
functions and covariance structures to estimate feed intake pattern in growing pigs. 383
Livest. Prod. Sci. 82:15-26.384
Bertram, M. J., T. S. Stahly, and R. C. Ewan. 1994. Impact of lean growth genotype and 385
dietary phosphorus regimen on rate and efficiency of growth and carcass characteristics 386
of pigs. J. Anim Sci. 72 (Suppl. 2):68 (Abstr.).387
Bridges, T. C., L. W. Turner, T. S. Stahly, J. L. Usry, and O. J. Loewer. 1992. Modeling the 388
physiological growth of swine part I: Model logic and growth concepts. Trans. ASAE389
35:1019-1028.390
Craig, B. A., and A. P. Schinckel. 2001. Nonlinear mixed effects model for swine growth. 391
Prof. Anim. Scientist 17:256-260.392
Darmani Kuhi, H., E. Kebreab, S. Lopez, and J. France. 2003a. An evaluation of different 393
growth functions for describing the profile of live weight with time (age) in meat and 394
egg strains of chicken. Poult. Sci. 82:1536-1543.395
Darmani Kuhi, H., E. Kebreab, S. Lopez, and J. France. 2003b. A comparative evaluation of 396
functions for the analysis of growth in male broilers. J. Agric. Sci. 140:451-459.397
Page 18 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

19
Ekpe, E. D., R. T. Zijlstra, and J. F. Patience. 2002. Digestible phosphorus requirement of 398
grower pigs. Canadian J. Anim. Sci. 82:541-549.399
Ferguson, N. S., R. M. Gous, and G. C. Emmans. 1994. Preferred components for the 400
construction of a new simulation model of growth, feed intake and nutrient requirements 401
of growing pigs. South African J. Anim. Sci. 24:10-17.402
France, J., J. Dijkstra, and M. S. Dhanoa. 1996. Growth functions and their application in 403
animal science. Ann. Zootech. 45:165-174.404
Hastad, C. W., S. S. Dritz, M. D. Tokach, R. D. Goodband, J. L. Nelssen, J. M. DeRouchey, 405
R. D. Boyd, and M. E. Johnston. 2004. Phosphorus requirements of growing-finishing 406
pigs reared in a commercial environment. J. Anim. Sci. 82:2945-2952.407
Jondreville, C., P.-S. Revy, J.-Y. Dourmad, Y. Nys, S. Hillion, F. Pontrucher, J. Gonzalez, J. 408
Soler, R. Lizardo, and J. Tibau. 2004. Influence of genotype and sex on minerals (Ca, P, 409
K, N, Mg, Fe, Zn, Cu) content and retention in the body of pigs from 25 to 135 kg of 410
body weight. J. Recherche Porcine 36:17-24.411
Ketaren, P. P., E. S. Batterham, E. White, D. J. Farrell, and B. K. Milthorpe. 1993. Phosphorus 412
studies in pigs .1. Available phosphorus requirements of grower finisher pigs. Br. J. 413
Nutr. 70:249-268.414
Leonard, T., and S. J. Hsu. 2001. Bayesian Methods: An Analysis for Statisticians and 415
Interdisciplinary Researchers. Cambridge Series in Statistical & Probabilistic 416
Mathematics. Cambridge Univ. Press, Cambridge U.K., 348 pp. 417
Littell, R. C., G. A. Milliken, W. W. Stroup, and R. D. Wolfinger. 1996. SAS® System for 418
Mixed Models. SAS Institute Inc., Cary, NC.419
Page 19 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

20
Mahan, D. C., and Jr. R. Shields. 1998. Macro- and micromineral composition of pigs from 420
birth to 145 kilograms of body weight. J. Anim. Sci. 76:506-512.421
Mudd, A. J., W. C. Smith, and D. G. 1969. Retention of certain minerals in pigs from birth to 422
90-kg live weight. J. Agric. Sci., Camb. 73: 181-188.423
NRC. 1998. Nutrient Requirements of Swine. 10th rev. ed. Natl. Acad. Press, Washington, DC.424
Schinckel, A. P., and C. F. de Lange. 1996. Characterization of growth parameters needed as 425
inputs for pig growth models. J. Anim. Sci. 74:2021-2036.426
Schinckel, A. P., and B. A. Craig. 2002. Evaluation of alternative nonlinear mixed effects 427
models of swine growth. Prof. Anim. Scientist 18:219-226.428
Schulin-Zeuthen, M. 2000. Capacity for retention of macro nutrients in growing pigs. M.Sc.429
Diss., Royal Veterinary and Agricultural Univ., Copenhagen, Denmark.430
Schulin-Zeuthen, M., and A. Danfær. 2001. Descriptions of the capacity of macro nutrient 431
retention in growing pigs. Page 7 in Proc. 52nd Ann. Meeting Europ. Assoc. Anim.432
Prod., Budapest, Hungary. 433
Schulin-Zeuthen, M., J. B. Lopes, E. Kebreab, D. M. S. S. Vitti, A. L. Abdalla, M. DeL. 434
Haddad, L. A. Crompton, and J. France. 2005. Effects of phosphorus intake on 435
phosphorus flow in growing pigs: Application and comparison of two models. J. Theor. 436
Biol. 236:115-125.437
Swidersky, A., J. Atkinson, and C. F. M. de Lange. 2003. Optimum dietary phosphorus levels 438
for grower-finisher pigs. Ontario Swine Research Review. Ontario Min. Agric. Fisheries, 439
Guelph, Canada.440
Page 20 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

21
Thomas, H. R., and E. T. Kornegay. 1981. Phosphorus in swine. I. Influence of dietary 441
calcium and phosphorus levels and growth rate on feedlot performance of barrows, gilts 442
and boars. J. Anim. Sci. 52:1041-1048.443
Thornley, J. H. M., and J. France. 2007. Mathematical Models in Agriculture. 2nd ed. CABI444
Int., Wallingford, U.K. 923 pp.445
Tibau, J., J. Gonzalez, J. Soler, M. Gispert, R. Lizardo, and J. Mourot. 2002. Influence du 446
poids à l'abattage du porc entre 25 et 140 kg de poids vif sur la composition chimique de 447
la carcasse: effets du génotype et du sexe. J. Recherche Porcine 34:121-127.448
Wellock, I. J., G. C. Emmans, and I. Kyriazakis. 2004. Describing and predicting potential 449
growth in the pig. Anim. Sci. 78:379-388.450
451
Page 21 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

22
Table 1. Data sources used in the study452
Dataset Source Number of pigs
Growth period (d)
Genotype Diet (% of as fed)1
1 Schulin-Zeuthen, (2000);
Schulin-Zeuthen and
Danfær (2001)
911 Birth to
1007
(Landrace × Yorkshire) ×Duroc
DM 89.9, dCP 14.3, NE 9.26, P 0.586, aP 0.378DM 89.6, dCP 13.1, NE 9.11, P 0.551, aP 0.317DM 88.9, dCP 11.9, NE 8.96, P 0.517, aP 0.256DM 88.2, dCP 12.7, NE 8.49, P 0.507, aP 0.241DM 89.0, dCP 11.2, NE 8.18, P 0.497, aP 0.244DM 88.8, dCP 10.7, NE 7.87, P 0.458, aP 0.224DM 88.4, dCP 9.56, NE 7.87, P 0.450, aP 0.224
2 Jondreville et al. (2004);
Tibau et al. (2002)
209 68 to 210 (Large White × Landrace) ×Pietrain or (Large White× Landrace) × Large White
DM 89.6, CP 20.0, NE 9.55, Ca 1.07, P 0.845, aP 0.406DM 87.3, CP 18.5, NE 9.21, Ca 1.02, P 0.817, aP 0.378
3 Swidersky et al. (2003) 1,330 70 to 155 Yorkshire DM 88.3, dCP 13.2, Ca 0.348, P 0.377, aP 0.122DM 87.2, dCP 13.5, Ca 0.521, P 0.436, aP 0.183DM 87.1, dCP 13.2, Ca 0.685, P 0.493, aP 0.240DM 88.9, dCP 13.1, Ca 0.858, P 0.553, aP 0.301
1CP = 6.25 × N (NRC, 1998;), dCP = digestible protein, NE (Net Energy, MJ/kg) = 7.72 Scandinavian Feed Units (Andersen and 453
Just, 1983), NE = 0.75 digestible energy (Andersen and Just, 1983), aP = available phosphorus.454
Page 22 of 31Journal of Animal Science
by guest on May 18, 2011
jas.fass.orgD
ownloaded from
23
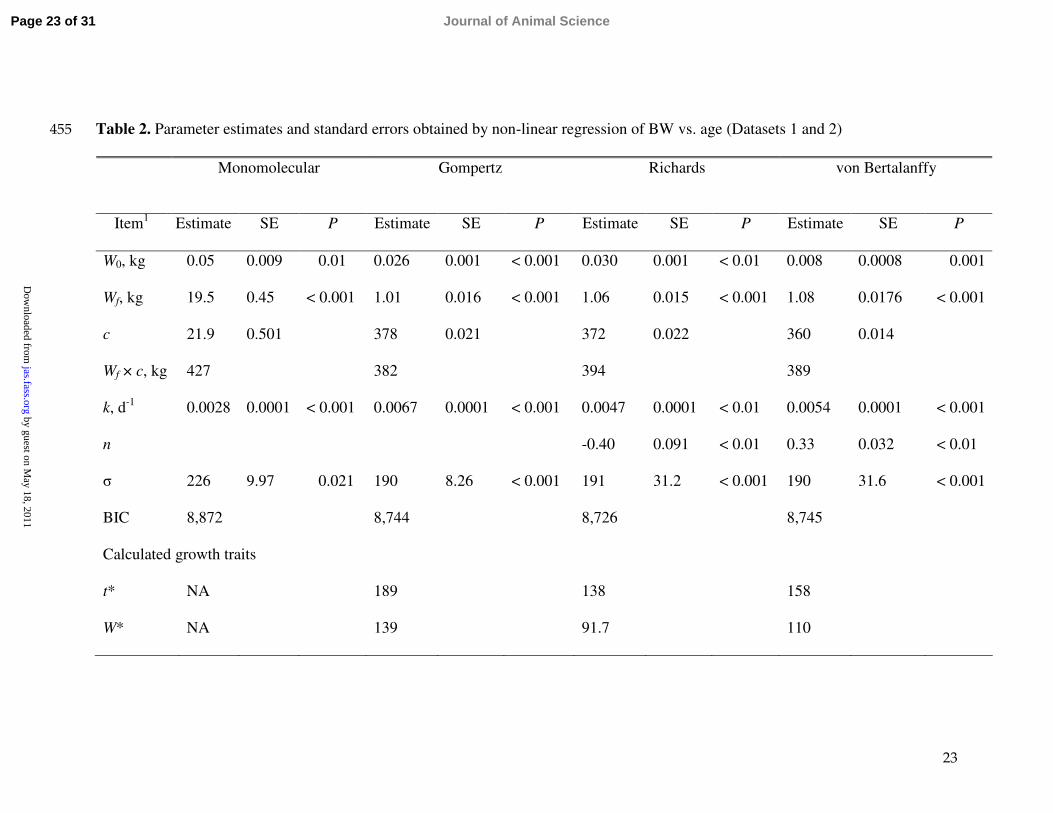
Table 2. Parameter estimates and standard errors obtained by non-linear regression of BW vs. age (Datasets 1 and 2)455
Monomolecular Gompertz Richards von Bertalanffy
Item1 Estimate SE P Estimate SE P Estimate SE P Estimate SE P
W0, kg 0.05 0.009 0.01 0.026 0.001 < 0.001 0.030 0.001 < 0.01 0.008 0.0008 0.001
Wf, kg 19.5 0.45 < 0.001 1.01 0.016 < 0.001 1.06 0.015 < 0.001 1.08 0.0176 < 0.001
c 21.9 0.501 378 0.021 372 0.022 360 0.014
Wf × c, kg 427 382 394 389
k, d-1 0.0028 0.0001 < 0.001 0.0067 0.0001 < 0.001 0.0047 0.0001 < 0.01 0.0054 0.0001 < 0.001
n -0.40 0.091 < 0.01 0.33 0.032 < 0.01
σ 226 9.97 0.021 190 8.26 < 0.001 191 31.2 < 0.001 190 31.6 < 0.001
BIC 8,872 8,744 8,726 8,745
Calculated growth traits
t* NA 189 138 158
W* NA 139 91.7 110
Page 23 of 31 Journal of Animal Science
by guest on May 18, 2011
jas.fass.orgD
ownloaded from

24
1W0 is initial weight at t = 0, Wf is asymptotic mature weight, c a parameter to estimate between-pig variability of mature body 456
weight, k is a rate constant, n is a shape parameter influencing point of inflection, σ is SE of model (= √variance (σ2) of error), BIC 457
(Bayesian Information Criterion) is a measure of regression fit, and t* and W* are age and weight at inflection point, respectively.458
459
Page 24 of 31Journal of Animal Science
by guest on May 18, 2011
jas.fass.orgD
ownloaded from
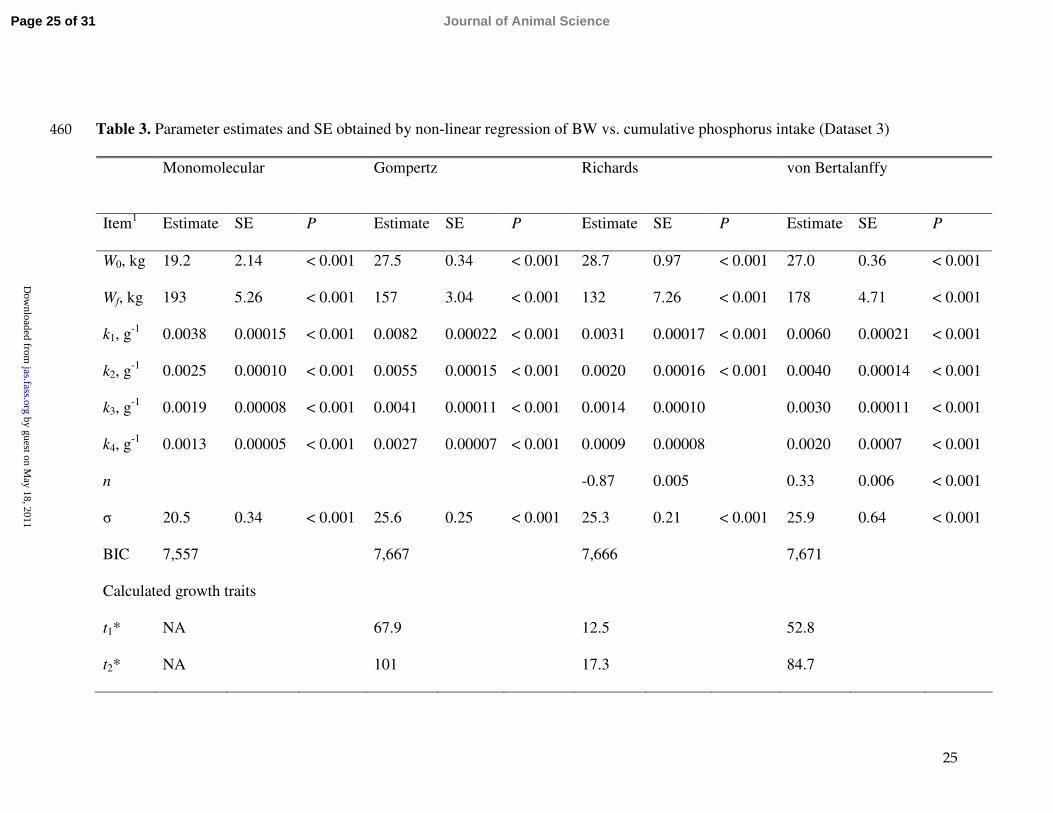
25
Table 3. Parameter estimates and SE obtained by non-linear regression of BW vs. cumulative phosphorus intake (Dataset 3)460
Monomolecular Gompertz Richards von Bertalanffy
Item1 Estimate SE P Estimate SE P Estimate SE P Estimate SE P
W0, kg 19.2 2.14 < 0.001 27.5 0.34 < 0.001 28.7 0.97 < 0.001 27.0 0.36 < 0.001
Wf, kg 193 5.26 < 0.001 157 3.04 < 0.001 132 7.26 < 0.001 178 4.71 < 0.001
k1, g-1 0.0038 0.00015 < 0.001 0.0082 0.00022 < 0.001 0.0031 0.00017 < 0.001 0.0060 0.00021 < 0.001
k2, g-1 0.0025 0.00010 < 0.001 0.0055 0.00015 < 0.001 0.0020 0.00016 < 0.001 0.0040 0.00014 < 0.001
k3, g-1 0.0019 0.00008 < 0.001 0.0041 0.00011 < 0.001 0.0014 0.00010 0.0030 0.00011 < 0.001
k4, g-1 0.0013 0.00005 < 0.001 0.0027 0.00007 < 0.001 0.0009 0.00008 0.0020 0.0007 < 0.001
n -0.87 0.005 0.33 0.006 < 0.001
σ 20.5 0.34 < 0.001 25.6 0.25 < 0.001 25.3 0.21 < 0.001 25.9 0.64 < 0.001
BIC 7,557 7,667 7,666 7,671
Calculated growth traits
t1* NA 67.9 12.5 52.8
t2* NA 101 17.3 84.7
Page 25 of 31 Journal of Animal Science
by guest on May 18, 2011
jas.fass.orgD
ownloaded from
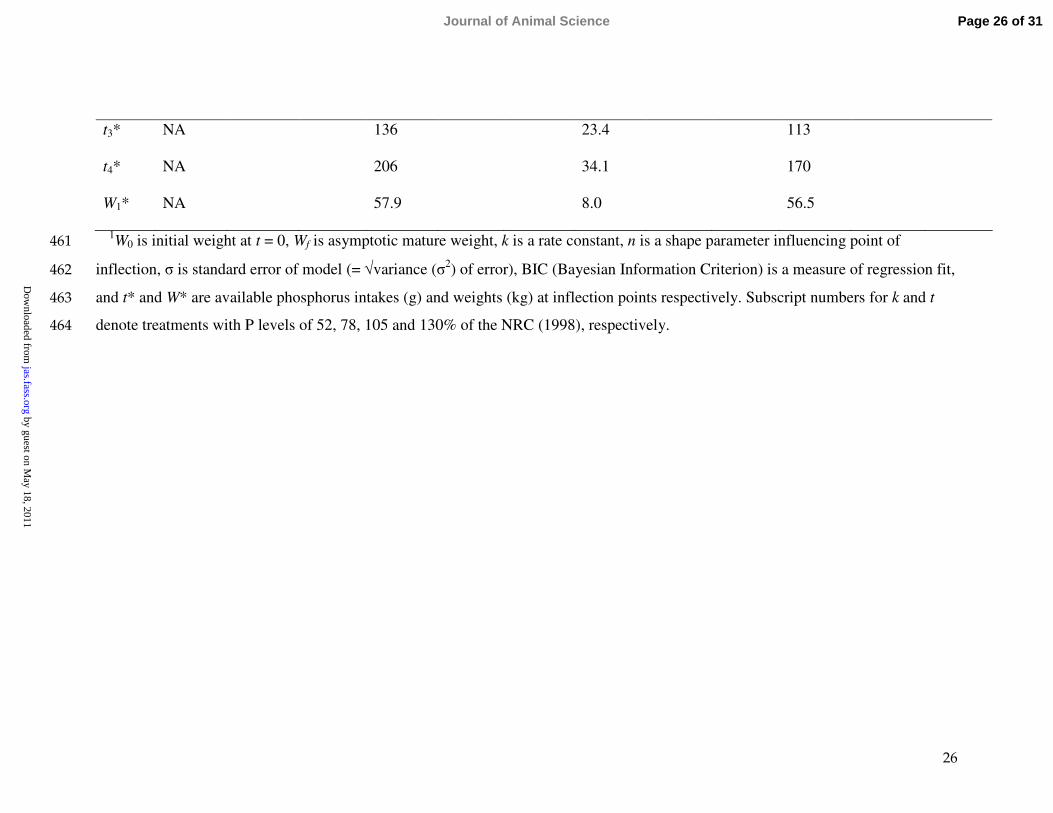
26
t3* NA 136 23.4 113
t4* NA 206 34.1 170
W1* NA 57.9 8.0 56.5
1W0 is initial weight at t = 0, Wf is asymptotic mature weight, k is a rate constant, n is a shape parameter influencing point of 461
inflection, σ is standard error of model (= √variance (σ2) of error), BIC (Bayesian Information Criterion) is a measure of regression fit, 462
and t* and W* are available phosphorus intakes (g) and weights (kg) at inflection points respectively. Subscript numbers for k and t463
denote treatments with P levels of 52, 78, 105 and 130% of the NRC (1998), respectively.464
Page 26 of 31Journal of Animal Science
by guest on May 18, 2011
jas.fass.orgD
ownloaded from
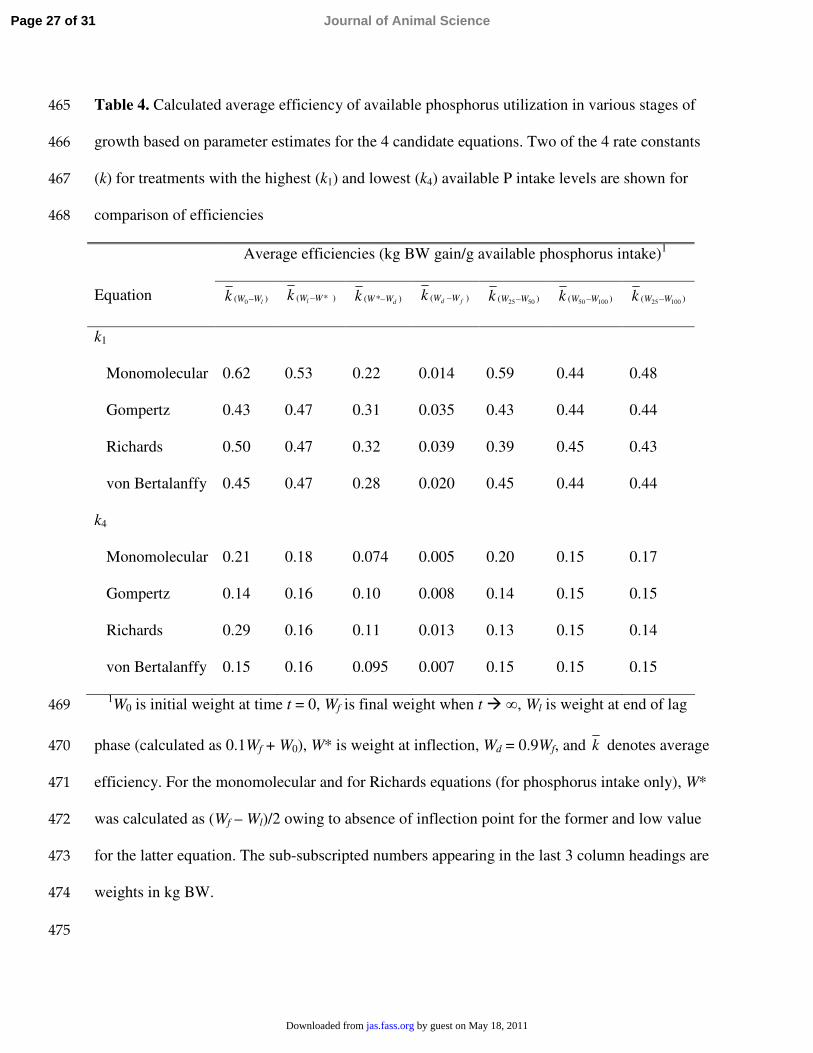
Table 4. Calculated average efficiency of available phosphorus utilization in various stages of 465
growth based on parameter estimates for the 4 candidate equations. Two of the 4 rate constants 466
(k) for treatments with the highest (k1) and lowest (k4) available P intake levels are shown for467
comparison of efficiencies468
Average efficiencies (kg BW gain/g available phosphorus intake)1
Equation )( 0 lWWk − )*( WWlk − )*( dWWk − )( fd WWk − )( 5025 WWk − )( 10050 WWk − )( 10025 WWk −
k1
Monomolecular 0.62 0.53 0.22 0.014 0.59 0.44 0.48
Gompertz 0.43 0.47 0.31 0.035 0.43 0.44 0.44
Richards 0.50 0.47 0.32 0.039 0.39 0.45 0.43
von Bertalanffy 0.45 0.47 0.28 0.020 0.45 0.44 0.44
k4
Monomolecular 0.21 0.18 0.074 0.005 0.20 0.15 0.17
Gompertz 0.14 0.16 0.10 0.008 0.14 0.15 0.15
Richards 0.29 0.16 0.11 0.013 0.13 0.15 0.14
von Bertalanffy 0.15 0.16 0.095 0.007 0.15 0.15 0.15
1W0 is initial weight at time t = 0, Wf is final weight when t � ∞, Wl is weight at end of lag 469
phase (calculated as 0.1Wf + W0), W* is weight at inflection, Wd = 0.9Wf, and k denotes average 470
efficiency. For the monomolecular and for Richards equations (for phosphorus intake only), W* 471
was calculated as (Wf – Wl)/2 owing to absence of inflection point for the former and low value 472
for the latter equation. The sub-subscripted numbers appearing in the last 3 column headings are 473
weights in kg BW.474
475
Page 27 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

28
Figure legends:476
477
Figure 1. Relationship between BW (kg) and age (d) in Datasets 1 and 2. Symbols represent 478
observed values and the lines are regression lines fitted using the different functions.479
480
Figure 2. Relationship between BW (kg) and cumulative available phosphorus intake (g) in 481
Dataset 3. Symbols represent observed values (circle, square, triangle, inverted triangle 482
correspond to treatments with available P intakes of 52, 78, 105 and 130% of the NRC (1998), 483
respectively) and the lines are regression lines fitted using the 4 candidate equations.484
485
Figure 3. Growth stages for a representative sigmoidal curve and graphical representation of 486
instantaneous efficiency for the monomolecular equation. W0 is initial weight at time t = 0, Wf is 487
final weight when t � ∞, Wl is weight at the end of lag phase (calculated as W0 + 0.1Wf), W* is 488
weight at inflection, Wd = 0.9Wf, and kP is instantaneous efficiency of phosphorus utilization for 489
growth.490
491
Page 28 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from
29
Figure 1. Schulin-Zeuthen et al.492
493
Monomolecular
Bod
y w
eigh
t, kg
0
100
200
300
400
500Gompertz
Richards
Age, d
0 200 400 600 800 1000
0
100
200
300
400
Von Bertalanffy
0 200 400 600 800 1000
494
Page 29 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from
30
Figure 2. Schulin-Zeuthen et al.495
Monomolecular
Bod
y w
eigh
t, kg
0
20
40
60
80
100
120
140Gompertz
Richards
0 200 400 600
0
20
40
60
80
100
120
140Von Bertalanffy
Cumulative available phosphorus intake, g
0 200 400 600
496
Page 30 of 31Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from
31
Figure 3. Schulin-Zeuthen et al.497
Sigmoidal
Time
Bod
y w
eigh
t
Monomolecular
Cumulative P intake
Instantaneous efficiency
kp = dW/dP
Stationary phase
Acceleration phase
Lag phase
Deceleration phase
Wf
W*
WlW0
Wd
498
499
Page 31 of 31 Journal of Animal Science
by guest on May 18, 2011jas.fass.orgDownloaded from

Citations
cleshttp://jas.fass.org/content/early/2007/06/12/jas.2006-738.citation#otherartiThis article has been cited by 1 HighWire-hosted articles:
by guest on May 18, 2011jas.fass.orgDownloaded from
Copyright © 2022 FDOKUMEN




















