
Comparative analysis of Late Jurassic sauropod trackways from the Jura Mountains (NW Switzerland)...
26
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Belvedere, Matteo] On: 28 May 2010 Access details: Access Details: [subscription number 922595506] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Historical Biology Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713717695 Comparative analysis of Late Jurassic sauropod trackways from the Jura Mountains (NW Switzerland) and the central High Atlas Mountains (Morocco): implications for sauropod ichnotaxonomy Daniel Marty a ; Matteo Belvedere b ; Christian A. Meyer c ; Paolo Mietto b ; Géraldine Paratte a ; Christel Lovis a ; Basil Thüring c a Office de la culture, Section d'archéologie et paléontologie, Porrentruy 2, Switzerland b Dipartimento di Geoscienze, Università degli Studi di Padova, Padova, Italy c Naturhistorisches Museum Basel, Basel, Switzerland First published on: 19 April 2010 To cite this Article Marty, Daniel , Belvedere, Matteo , Meyer, Christian A. , Mietto, Paolo , Paratte, Géraldine , Lovis, Christel and Thüring, Basil(2010) 'Comparative analysis of Late Jurassic sauropod trackways from the Jura Mountains (NW Switzerland) and the central High Atlas Mountains (Morocco): implications for sauropod ichnotaxonomy', Historical Biology, 22: 1, 109 — 133, First published on: 19 April 2010 (iFirst) To link to this Article: DOI: 10.1080/08912960903503345 URL: http://dx.doi.org/10.1080/08912960903503345 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Comparative analysis of Late Jurassic sauropod trackways from the Jura Mountains (NW Switzerland)...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Belvedere, Matteo]On: 28 May 2010Access details: Access Details: [subscription number 922595506]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Historical BiologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713717695
Comparative analysis of Late Jurassic sauropod trackways from the JuraMountains (NW Switzerland) and the central High Atlas Mountains(Morocco): implications for sauropod ichnotaxonomyDaniel Martya; Matteo Belvedereb; Christian A. Meyerc; Paolo Miettob; Géraldine Parattea; ChristelLovisa; Basil Thüringc
a Office de la culture, Section d'archéologie et paléontologie, Porrentruy 2, Switzerland b Dipartimentodi Geoscienze, Università degli Studi di Padova, Padova, Italy c Naturhistorisches Museum Basel, Basel,Switzerland
First published on: 19 April 2010
To cite this Article Marty, Daniel , Belvedere, Matteo , Meyer, Christian A. , Mietto, Paolo , Paratte, Géraldine , Lovis,Christel and Thüring, Basil(2010) 'Comparative analysis of Late Jurassic sauropod trackways from the Jura Mountains(NW Switzerland) and the central High Atlas Mountains (Morocco): implications for sauropod ichnotaxonomy',Historical Biology, 22: 1, 109 — 133, First published on: 19 April 2010 (iFirst)To link to this Article: DOI: 10.1080/08912960903503345URL: http://dx.doi.org/10.1080/08912960903503345
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Comparative analysis of Late Jurassic sauropod trackways from the Jura Mountains(NW Switzerland) and the central High Atlas Mountains (Morocco): implications forsauropod ichnotaxonomy
Daniel Martya*, Matteo Belvedereb, Christian A. Meyerc, Paolo Miettob, Geraldine Parattea, Christel Lovisa and
Basil Thuringc
aOffice de la culture, Section d’archeologie et paleontologie, Paleontologie A16, Hotel des Halles, P.O. Box 64, 2900 Porrentruy 2,Switzerland; bDipartimento di Geoscienze, Universita degli Studi di Padova, Via Giotto 1, 35137 Padova, Italy; cNaturhistorischesMuseum Basel, Augustinergasse 2, 4000 Basel, Switzerland
(Received 2 October 2009; final version received 22 November 2009)
Late Jurassic sauropod trackways from the Jura Mountains (NW Switzerland) and the central High Atlas Mountains(Morocco) are described and compared. Emphasis is put on track preservation and trackway configuration. The trackwaysare similar with respect to preservation and the pes and manus track outlines, but they show a large range of trackwayconfiguration. Only one of the trackways reveals digit and claw impressions, and thus differences in trackway gauge and theposition of pes and manus tracks are the most explicit characters for their distinction. The Late Jurassic to Early Cretaceousichnotaxa Brontopodus, Parabrontopodus and Breviparopus are reviewed and a differential diagnosis is given for thetrackways studied. The reference trackway of Breviparopus corresponds to one of the studied trackways of Morocco.Parabrontopodus and Breviparopus are considered to be both valid ichnotaxa, even though we recommend the latter to beformally erected based on better-preserved tracks than those currently exposed. The analysed trackways and ichnotaxasuggest that trackway configuration, notably trackway gauge (width), is not decisively influenced by extrinsic factors such asontogenetic stage, locomotion speed and substrate properties. However, it cannot be excluded that it is related to otherfactors such as individual behaviour or even sexual dimorphism.
Keywords: Sauropod footprint; trackway configuration, trackway gauge; Late Jurassic; Switzerland; Morocco
Introduction
The present paper describes in detail Late Jurassic
sauropod trackways from the Jura Mountains (NW
Switzerland) and the central High Atlas Mountains
(Morocco), with particular emphasis put on track
preservation, track morphology and trackway configur-
ation, notably trackway gauge (width). The major issues
are (1) to compare the trackways, (2) to explain similarities
and differences between the trackways (e.g. substrate
properties, trackmakers and behaviour) and (3) to evaluate
their ichnotaxonomical assignation. We also comment on
the ichnotaxon Breviparopus Dutuit and Ouazzou (1980)
since one of the analysed trackways from Morocco (i.e.
trackway Deio-D) is the reference trackway for this
ichnotaxon. The term ‘reference’ is used instead of ‘type’
throughout this paper, because this ichnotaxon is not
formally emended (see below).
In the description of dinosaur ichnotaxa, trackway
configuration is traditionally used. This is also the case for
sauropod trackways, and a good example is Brontopodus
birdi, which is based on both track morphology and
trackway parameters (Farlow et al. 1989). More
particularly, Farlow (1992) proposed that sauropod track-
ways could be classified according to trackway width. He
introduced the term trackway gauge stressing that
sauropod trackways can generally be described as narrow
gauge when pes tracks are ‘close to or even intersecting the
trackway midline’ or wide gauge when they are ‘well away
from the trackway midline’. Because not all trackways
could unambiguously be classified into one or the other
category, Lockley et al. (1994b) and Meyer et al. (1994)
proposed an intermediate category (i.e. medium gauge),
but they also stated that a formal classification of sauropod
trackways on gauge alone was premature and that there is
‘a need to carefully describe well-preserved trackways and
refine sauropod ichnotaxonomy . . . ’. Nonetheless, track-
way gauge is commonly used for the classification of
sauropod trackways and the identification of their track-
makers (e.g. Lockley et al. 1994a, 1994b, 2002a, 2002b;
Moratalla et al. 1994; Lockley and Hunt 1995; Dalla
Vecchia and Tarlao 2000; Lockley and Meyer 2000; Day
et al. 2002, 2004; Marty et al. 2003; Moreno and Benton
2005; Gonzalez Riga and Calvo 2009 and dos Santos et al.
ISSN 0891-2963 print/ISSN 1029-2381 online
q 2010 Taylor & Francis
DOI: 10.1080/08912960903503345
http://www.informaworld.com
*Corresponding author. Email: [email protected]
Historical Biology
Vol. 22, Nos. 1–3, March–June–September 2010, 109–133
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

2009) and also for ichnofacies (e.g. Lockley et al. 1994c;
Mannion 2008) or herding behaviour (Day et al. 2002).
When Farlow (1992) introduced the trackway gauge
concept, he stressed that trackway configuration including
gauge may also be related to substrate properties and
locomotion (speed), in a way that the trackmakers of the
wide-gauge trackways may have angled their legs inward
with respect to the sagittal plane, and if so that trackway
gauge would have little systematic value. This was also
discussed by Wilson and Carrano (1999), but they
concluded that ‘it is unlikely that any single sauropod
was able to produce both wide- and narrow-gauge
trackways’, because they assumed that skeletal mor-
phology is responsible for trackway gauge and that the
wide stance was a synapomorphy of the titanosaurids.
More recently, Wright (2005) has suggested that trackway
gauge also seems to be influenced by other characteristics
such as locomotor style and ontogeny. Carpenter (2009)
suggested that trackway gauge could also be related to the
degree of lateral motion of the trackmaker, i.e. wide-gauge
trackways were produced by trackmakers that had less
lateral motion than the makers of the narrow-gauge ones.
However, despite the importance of trackway gauge in
the classification of sauropod trackways and the proposed
link with the anatomy of the trackmakers, a quantification
for trackway gauge was provided only recently by
Romano et al. (2007) introducing the trackway ratio
(TR) and by Marty (2008) introducing a ratio between the
width of the angulation pattern and the corresponding
track length (i.e. the [WAP/PL] ratio for pes tracks).
An important part of this work is the characterisation
of trackway gauge, and therefore, the two trackway gauge
ratios mentioned above are compared. This is also
presented for three sauropod ichnotaxa Breviparopus,
Brontopodus and Parabrontopodus, contributing towards
their better characterisation. Finally, we test whether
sauropod trackway gauge is influenced by parameters such
as substrate consistency, behaviour (locomotion speed)
and ontogenic stage (pes length).
General setting
NW Switzerland
The four studied trackways were documented on four
different tracksites near Porrentruy, Ajoie district, Canton
Jura (Figure 1(D) and (E)): Chevenez–Combe Ronde
(CHE-CRO), Courtedoux–Pommerat (CTD-PMM), Cour-
tedoux–Sur Combe Ronde (CTD-SCR) and Courtedoux–
Tchafoue (CTD-TCH). In this tracksite naming system, the
community is indicated first followed by the name of the
tracksite. The excavations of the tracksites started in 2002
and are carried out by a paleontological survey project (the
Paleontologie A16) on or close to the future course of the
Figure 1. Geographical setting of Europe and North Africa (C) exhibiting the location of the studied tracksites in the central High AtlasMountains, Morocco (A and B), and in the Jura Mountains, NW Switzerland (D and E).
D. Marty et al.110
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Swiss federal highway A16, also named Transjurane (e.g.
Marty et al. 2004, 2007). By the end of 2009, over 50
ichnoassemblages (i.e. an association of true tracks found
on a single paleosurface) revealing over 8000 tracks,
including 234 trackways of tridactyl dinosaurs (mainly
attributed to theropods) and 177 sauropod trackways, have
been excavated and documented.
The sauropod trackways occur in three track-bearing
laminite intervals (named lower, intermediate and upper
levels) interbedded between shallow marine carbonate
platform sediments (Marty 2008). These laminites were
deposited in inter- to supratidal paleoenvironments, which
is indicated by macroscopic (stromatolitic lamination,
desiccation cracks, wave ripples and invertebrate burrows)
and microscopic (e.g. cryptmicrobial lamination, fenestrae
and brecciation) sedimentological features (Marty 2008;
Marty and Pacton 2009) (Figure 2(C) and (D)). Each
laminite interval bears several distinct track-bearing
levels, whereas the intermediate levels with a thickness
of 1 m and at least 15 track-bearing levels are the track-
richest interval (Marty et al. 2007). In total over 30 distinct
levels, some of which (notably the main track levels at the
base of each laminite interval) can be correlated between
the different tracksites, have been identified so far. The
sequence with the three track-bearing intervals is part of
the Reuchenette Formation (Thalmann 1966; Gygi 2000),
and it can be precisely dated with ammonites to the
Tethyan Divisum to Acanthicum ammonite zones, i.e. late
Early to early Late Kimmeridgian (Marty et al. 2003; Jank
et al. 2006a, 2006b).
During the Late Kimmeridgian, the localities were
located at the northern margin of the oceanic Ligurian
Tethys as a large, structurally complex carbonate platform
(e.g. Thierry et al. 2000a, 2000b and Stampfli and Borel
2002). This platform was at a paleolatitude of around 308N,
at the threshold between the Paris Basin to the northwest
and the Tethys Ocean to the south (Figure 3) and, thus,
influenced by both the Tethyan and Boreal realms (e.g.
Ziegler 1988; Thierry 2000a, 2000b and Jank et al. 2006a).
During the Kimmeridgian, the climate of the Jura carbonate
platform was subtropical and is generally considered semi-
arid to arid (e.g. Hallam 1984, 1985; Frakes et al. 1992;
Ross et al. 1992; Weissert and Mohr 1996 and Bertling and
Insalaco 1998).
Morocco
The analysed trackways are from two different tracksites
located in the western part of the Iouaridene Valley near
the village of Taghbalout, i.e. in the central High Atlas
Mountains, around 15 km east of the town of Demnat and
120 km east of Marrakech (Figure 1(A) and (B)). Known
since 1930s (Plateau et al. 1937), the Iouaridene tracksites
became important with the work of Dutuit and Ouazzou
(1980) because of their description of a narrow-gauge
trackway with very large tracks, which they named
Breviparopus taghbaloutensis. Later, Ishigaki (1985a,
Figure 2. Typical macroscopic and microscopic features of the studied tracksites in Morocco (A and B) and NW Switzerland (C and D).(A) Cross section of the track RP3 of the Deio Lav-A trackway. The dashed line indicates the track outline. Note the absence ofundertracks indicating that the subjacent sediment was already consolidated at the time of track formation. (B) Thin section with planarcryptmicrobial lamination and laminoid fenestrae of level 3 near Taghbalout. (C) The lower track-bearing laminites at the CHE-CROtracksite. The laminite interval is capped by a shallow marine, massif limestone bed. Hammer (33 cm in length) for scale is put on themain track level (level 500). (D) Thin-section CRO004-132a of layer 545 from the CHE-CRO tracksite exhibiting crinkled cryptmicrobiallamination, characterised by an alternation of clotted micrite and irregular seams enriched in organic matter.
Historical Biology 111
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

1985b, 1985c, 1986, 1989) published several papers on
some of the sauropod tracksites of the Iouaridene Valley,
but only recently more complete studies have been
achieved by Nouri (2007) and Belvedere (2008). The latter
reported around 800 tridactyl tracks, mostly attributed to
theropods, 200 sauropod tracks and 2 tracks attributed to
the stegosaur ichnogenus Deltapodus (Belvedere and
Mietto 2010). Consequently, sauropod tracks represent
about 20% of the studied tracks of the Iouaridene
ichnosite, even though it has to be considered that the very
long Breviparopus reference trackway (i.e. Deio-D)
accounts for a large part of the sauropod tracks.
All tracksites of the Iouaridene basin are located in the
lower member (sensu Charriere et al. 2005) of the
Iouaridene Formation (Jenny et al. 1981), in which
Belvedere (2008) identified 21 track-bearing levels.
The sequence with the track-bearing levels is composed
of a cyclic alternation of siliciclastic mudstones and
carbonate-cemented mudstones to very fine sandstones
topped by mud cracks sometimes associated with
symmetrical ripple marks and often bearing dinosaur
tracks. Fluvial channels, with fining upward sequences,
climbing ripples and herringbone cross-stratification, are
present at the top of the sequence with the track-bearing
levels. Dinosaur tracks are never deeper than 15 cm, even
for the largest sauropod tracks, indicating a coherent
and/or well-laminated (e.g. due to microbial mats)
sediment. In thin sections, the carbonate-cemented levels
often exhibit a pronounced microbial lamination with
laminoid fenestrae and birds eyes (Figure 2(B)). The
presence of evaporitic minerals, concertina-like veins and
pseudo-anticlines with evaporitic infillings suggest an arid
to semi-arid environment (Belvedere 2008).
The track-bearing sequence has long been attributed to
the Middle Jurassic (e.g. Jenny et al. 1981; Jenny and
Jossen 1982 and Jenny 1985). However, Charriere et al.
(2005) identified charophytes (Prochara kimmeridgiensis,
Dictyoclavator ramalhoi) that indicate a Late Jurassic
Figure 3. Paleogeographical setting of the studied localities during the Late Jurassic. The central High Atlas Mountains (Morocco) wereformerly located at the north-eastern part of the Sahara Craton (1), and the Jura Mountains (NW Switzerland) on the Jura carbonateplatform (2) between the Massif Central to the southwest and the Rhenish Massif to the north. Redrawn after Thierry et al. (2000a, 2000b).The paleogeographical position of Corsica and Sardinia is interpreted differently in other reconstructions (e.g. Stampfli et al. 2002). (1)Exposed land; (2) hypersaline; (3) aeolian, fluviatile, lacustrine and fluvio-lacustrine; (4) shallow-water environments with fluctuatingsalinities; (5) coastal marine and shallow marine (terrigenous); (6) shallow marine (carbonate); (7) deeper carbonates and (hemi)pelagicoozes; (8) deep marine; (9) deep oceanic basins (mid ocean ridge) and (10) major faults.
D. Marty et al.112
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

(Oxfordian to Kimmeridgian) age to the track-bearing
sequence, i.e. the upper part of the lower member of the
Iouaridene Formation. This is further supported by the
occurrence of a Barremian lacustro-marine ostracod
assemblage (Globator trochiliscoides, Atopochara trivol-
vis triquetra) in the upper member of the Iouaridene
Formation (Charriere et al. 2005).
During the Late Jurassic, the domain of today’s central
High Atlas Mountains was located at the northeastern part
of the Sahara Craton at a paleolatitude of about 208 N
(Figure 3), and continental red beds (couches rouges) were
deposited, whereas in the western area of the High Atlas, a
more coastal-marine deposition prevailed due to the
opening of the central Atlantic (e.g. Ziegler 1990;
Charriere et al. 2005 and Haddoumi et al. 2009).
Methods and terminology
Only quadrupedal trackways or pes-dominated trackways
(i.e. trackways with some but not all manus tracks missing
due to overprinting) with tracks considered as true tracks
(for definition see below) are studied. For all trackways,
track and trackway data were systematically measured
(Figure 4) following commonly used ichnological
terminology (e.g. Leonardi 1987 and Thulborn 1990),
and the mean parameters are given in Table 1. Tracks
directed outward with respect to the line connecting it with
the consecutive track (the stride) have an outward
(positive) rotation and those directed inward an inward
(negative) rotation. For the quantification of sauropod
trackway gauge, two quantitative ratios are used: the TR
and the ratio between the width of the angulation pattern
and the corresponding track length.
Trackways are illustrated by outline drawings exhibit-
ing the distinct and essential characters of the tracks
(Figure 5) and photographs (Figure 6). In the trackway
outline drawings, the internal track outline marks the
actual imprint of the foot and defines the track dimensions,
whereas the external track outline and the external limit of
the displacement rim define the extramorphological
features of the tracks (Figure 5).
Preservation terminology
True track
Lockley (1991) called the track emplaced on the actual
tracked surface true track, and the term true track is used
here in this sense. The track bottom or true track sensu
Figure 4. Schematic sauropod trackway showing the essentialpes and manus track and trackway parameters measured: PL, peslength; PW, pes width; a, pes rotation; LPP, left pes pace length;RPP, right pes pace length; PS, pes stride length; g, pes paceangulation; WAP, width of the pes angulation pattern; ML, manus
length; MW, manus width; b, manus rotation; LMP, left manuspace length; RMP, right manus pace length; MS, manus stridelength; d, manus pace angulation; WAM, width of the manusangulation pattern; SW, side width; OW, overall width; LP, leftpes; RP, right pes; LM, left manus and RM, right manus track.
R
Historical Biology 113
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

stricto may, under appropriate substrate conditions and
if the foot is placed in an ideal way, reveal anatomical
details of the trackmakers’ foot. Such tracks were also
named elite tracks, which have the connotation of well
preserved, visually clear true tracks that are not
distorted (e.g. Lockley and Hunt 1995 and Lockley and
Meyer 2000).
Underprint
A special case of a true track, which is produced when the
foot penetrates or breaks through the uppermost or through
several layers of sediment and leaves the true track on
sediment below the initially exposed sediment surface
(Marty et al. 2009). If in this case the sediment is split open
at successively deeper layers, the overall track will be
found to be less and less complete (Thulborn 1990), while
the track bottom (or the true track sensu stricto) may still
reveal anatomical details of the foot.
Undertrack
A track that is formed in (bio-) laminated and plastic
substrate when the foot does not penetrate the sediment
but compresses it in a way that it creates a miniature
stratigraphic sequence or stack of transmitted prints
(stack of casts and moulds) (Thulborn 1990;
Lockley 1991).
Trackway ratio
The TR is defined as the ratio between the side width (SW)
and the overall width (OW) of a trackway (Figure 4),
expressed as a percentage (Romano et al. 2007):
TR ¼ ðSW=OWÞ £ 100%:
Both measurements (SW and OW) are taken
perpendicular to the trackway axis of the trackway: as
such SW may not correspond to the true width of the
track, which is measured at right angles to the length of
the track. Only in cases where the track is oriented
parallel to the long axis of the trackway the SW
represent the true track width. The higher the TR, the
narrower the trackway gauge is; the provisional
boundary between wide gauge and medium gauge is at
35% and that between medium gauge and narrow gauge
is at 50% (Romano et al. 2007).
The TR is calculated for both pes (pes TR, PTR)
and manus (manus TR, MTR) tracks. Only the SW
and OW of the Moroccan trackways were measured in
the field. For the studied Swiss trackways, SW and OW
were measured on magnifications of outline drawings,
and for the ichnotaxa from magnifications of published
outline drawings or the TR was taken from Romano et al.
(2007) as indicated in Table 2.
Ratio between the width of the angulation pattern and thecorresponding track length
Defined for both pes and manus tracks as (Marty 2008):
(1) width of the pes angulation pattern/pes length
(i.e. [WAP/PL] ratio)
(2) width of the manus angulation pattern/manus width
(i.e. [WAM/MW] ratio).
In the case of manus tracks, the manus width is used
instead of its length because manus tracks are often
incomplete due to overprinting by the subsequent pes or
due to a more digitigrade impression of the manus. Thus,
manus length is subjected to a high variability, and width
better represents the size of the manus than the length. If
the [WAP/PL] ratio equals one (i.e. PL ¼ WAP), the pes
tracks are likely to touch the trackway midline. If the ratio
is smaller than one, tracks intersect the trackway midline,
which corresponds to the definition of narrow gauge
(Farlow 1992). Accordingly, a value of 1.0 separates
narrow-gauge from medium-gauge trackways, whereas the
value 1.2 is arbitrarily fixed between medium-gauge and
wide-gauge trackways, and trackways with a value higher
than 2.0 are considered as very wide gauge (Marty 2008).
For the studied trackways, all measurements (Table 1)
were made at the outcrops, and for the ichnotaxa included
in the discussion (Table 2), they were taken from
published data or measured on magnifications of published
outline drawings.
Ratio between the widths of the pes and manusangulation patterns
This ratio is introduced here and is defined as
(1) width of the pes angulation pattern/width of the manus
angulation pattern (i.e. [WAP/WAM] ratio).
This ratio is used to characterise whether manus tracks
are closer or farther away from the trackway midline than the
pes tracks. If this ratio equals one, pes and manus tracks are
located in a line at the same distance from the trackway
midline; if the ratio is smaller than one, manus tracks are
located farther away from the trackway midline than pes
tracks and if the ratio is higher than one, the manus tracks are
located closer to the trackway midline than the pes tracks.
Heteropody
Heteropody is defined as the difference in area (total track
area) between the pes and manus tracks in a given trackway
of a quadrupedal animal (Lockley et al. 1994b). Lockley
et al. (1994b) noted that differences in heteropody could be
used to distinguish ichnotaxa, even if heteropody is not
considered as a character in the phylogenetic taxonomy of
sauropods (e.g. Salgado et al. 1997; Wilson and Sereno
1998; Wilson and Carrano 1999 and Wilson 2002, 2005).
D. Marty et al.114
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Detailed calculations of total track areas and of the
heteropody ratio are not in the scope of this work, because
manus tracks are often incomplete due to overprinting or a
digitigrade stance, and their surface area is thus subjected
to an important variation and difficult to calculate.
Therefore, heteropody values should be considered only
as a gross approximation. The ratio between the indices of
pes and manus track size may also be used as an
approximate estimation of heteropody, and this ratio is
indicated in Table 1. The higher this ratio, the smaller the
manus with respect to the pes track is. The definition of the
index of track size follows Thulborn (1990, p. 234), and it
is calculated with the mean pes and manus lengths and
widths, respectively.
Locomotion speed
Calculation of locomotion speed (Tables 1 and 2) derives
from the relationship obtained by Alexander (1976). Because
of several shortcomings of this empiric relationship due to
the estimation of hip height based on tracks and the a priori
unknown precise trackmaker (e.g. Coombs 1978 and
Rainforth and Manzella 2007), as well as the unknown
precise relationship between relative stride length (S/h) and
the Froude number (speed2/leg length £ gravitation con-
stant) for dinosaurs (Alexander 2006), speed calculations are
considered as rough approximations only (Hutchinson et al.
2005; Alexander 2006). Nonetheless, Alexander’s (1976)
method is at least informative providing an estimation for the
magnitude of the locomotion speed of a dinosaur and, more
importantly, for the relative speed of a given sample of
trackways. Locomotion speed is used here in this way to test
whether it has an influence on trackway configuration,
notably trackway gauge.
Description of studied trackways
NW Switzerland
Four typical trackways from four different ichnoassem-
blages and from all three track-bearing laminite intervals
are described. The trackways of NW Switzerland are
either protected in situ (CHE-CRO-500-S10, CTD-SCR-
1000-S10, CTD-PMM-1505-S1) or the original trackways
were extracted and displaced (CTD-TCH-1055-S4) or
casted (CTD-SCR-1000-S10, CTD-PMM-1505-S1) and
are now housed in the collection of the Natural History
Museum of Porrentruy (Musee jurassien des sciences
naturelles MJSN).
Trackway CHE-CRO-500-S10
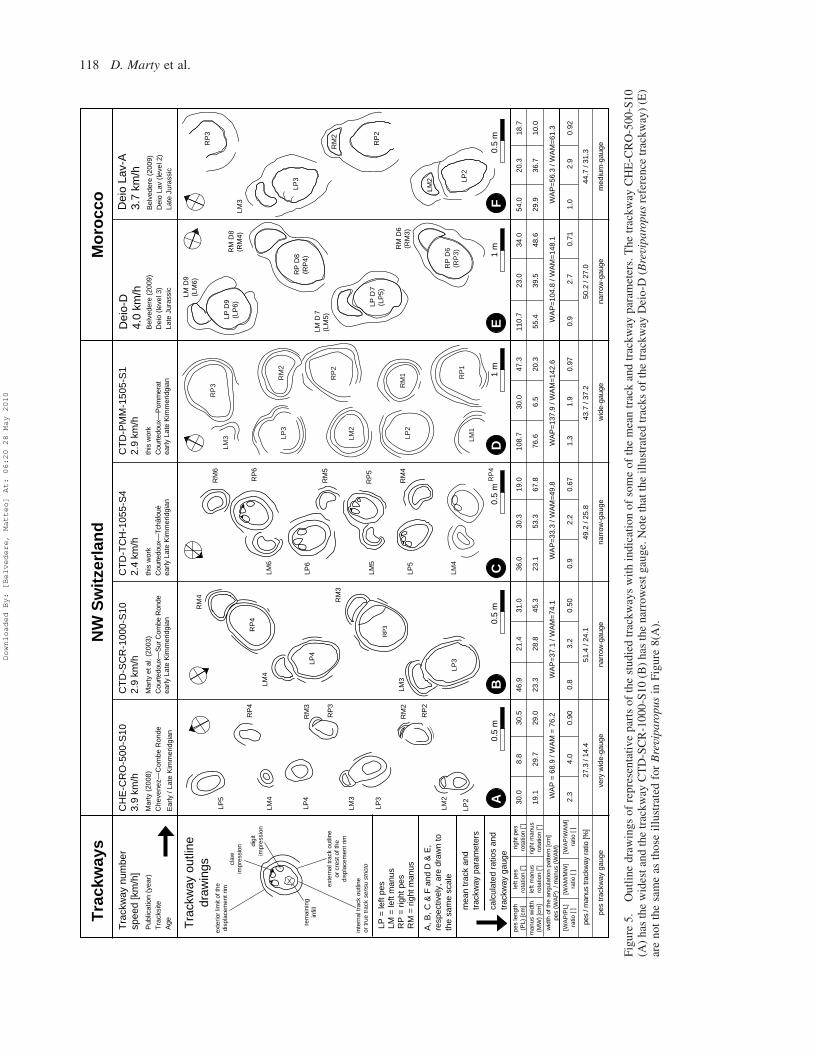
This straight, 8.5 m long trackway (Figures 5(A) and 6(A))
was described by Marty (2008) who, based on its very
wide gauge and rather small heteropody, tentatively
assigned it to the ichnotaxon Brontopodus.
The trackway is characterised by a very wide gauge,
a pes-dominated pattern (some manus tracks are
completely overprinted), an irregular configuration
(variable position of manus tracks with respect to the
pes tracks) and a relatively small heteropody coefficient
(Figure 5(A)). Manus tracks, undeformed by the pes, are
in front of the pes tracks with their centre located
slightly to the inside of the prolongation of the pes’ long
axis. Pes and manus tracks have a similar high mean
outward rotation.
Pes tracks are longer (30.0 cm mean length) than wide
(19.0 cm mean width) and have a subcircular to oval,
slightly elongated shape (Figure 6(C)).
Manus tracks are wider (19.1 cm mean width) than long
(9.6 cm mean length) and their shapes vary, due to different
degrees of overprinting by the subsequent pes and inclination
towards the anterior part of the tracks, from semicircular
to crescent shaped. Well-preserved and undeformed
manus tracks (e.g. LM4, Figure 6(B)) are semicircular
and lack evidence for digit and claw impressions.
Trackway CTD-SCR-1000-S10
This straight, 19 m long trackway (Figures 5(B) and 6(D))
was first described by Marty et al. (2003) and assigned to
the ichnotaxon Parabrontopodus. All tracks are rather
shallow and they are partially surrounded by small, and
narrow displacement rims.
This quadrupedal trackway is characterised by a
narrow gauge (Figure 6(D)) and a heteropody ratio of
about 1:3 for virtually undeformed manus tracks. The
centres of the manus tracks are placed about on the
prolongation of the pes’ long axis, and hence clearly
farther away from the trackway midline than the centres of
the pes tracks. Both pes and manus tracks are rotated
outwards with a higher outward rotation of the manus than
for the pes, and the trackway is slightly asymmetric such
as the right pes and manus tracks have a higher mean
outward rotation than the left ones.
The pes tracks have an oval shape and are longer
(46.9 cm mean length) than wide (35.8 cm mean width),
with the greatest width located in the anterior part of the
pes’ long axis but not so far away from its midpoint
(Figure 6(E) and (F)). Some of the pes tracks show up to
two very shallow and round digit impressions but without
evidence for claw impressions.
Virtually undeformed manus tracks (e.g. RM4) are
semicircular, convex forward, wider (23.3 cm mean width)
than long (12.3 cm mean length), and they exhibit neither
digit nor claw impressions (Figure 6(F)).
Trackway CTD-TCH-1055-S4
This straight, 14 m long trackway (Figures 5(C) and 6(G))
is one of the trackways with the best-preserved pes tracks
Historical Biology 115
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Table 1. Mean pes and manus track and trackway parameters and calculated gauge ratios for pes and manus tracks.
Country NW Switzerland Morocco
Tracksite CHE-CRO CTD-SCR CTD-TCH CTD-PMM Deio Deio Lav
Trackway number S10 S10 S4 S1 Deio-D Lav-A
Level 500 1000 1055 1505 3 2
Number of pes tracks 12 20 16 9 28 6Number of manus tracks 11 13 19 7 26 5Pattern (q, quadrupedal, p–d, pes dominated) p–d to q p–d to q q q q qTrackway length (m) 8.5 19 14 12 86 5Track orientation (8) 25 225 140 310 260 335
Track parametersPes tracksLength (PL) (cm) 30.0 46.9 36.0 108.7 110.7 54.0Width (cm) 19.0 35.8 27.1 92.1 88.5 40.5Index of track size (IPS)* (cm) 23.9 41.0 31.2 100.1 99.0 46.8Depth (cm) 4.3 3.3 2.6 3.0 13.0 9.2Left rotation (8) 8.8 21.4 30.3 30.0 23.0 20.3Right rotation (8) 30.5 31.0 19.0 47.3 34.0 18.7
Manus tracksLength (cm) 9.6 12.3 13.3 63.2 29.7 14.5Width (MW) (cm) 19.1 23.3 23.1 76.6 55.4 29.9Index of track size (IMS)* (cm) 13.5 16.9 17.5 69.6 40.6 20.8Depth (cm) 3.3 2.6 2.6 3.6 4.9 6.3Left rotation (8) 29.7 28.8 53.3 6.5 39.5 36.7Right rotation (8) 29.0 45.3 67.8 20.3 48.6 10.0
Trackway parametersPes tracksStride (cm) 137.0 159.2 117.0 284.3 350.5 203.8Width a. pattern (WAP) (cm) 68.9 37.1 33.3 137.9 104.8 56.3Left pace (cm) 98.0 90.2 68.0 201.8 207.8 109.2Right pace (cm) 91.7 90.2 68.4 191.8 195.3 120.5Pace angulation (8) 87.7 128.6 118.0 90.9 116.6 116.3
Manus tracksStride (cm) 141.3 144.7 121.7 287.0 354.9 202.0Width a. pattern (WAM) (cm) 76.2 74.1 49.8 142.6 148.1 61.3Left pace (cm) 107.0 105.8 75.4 201.0 233.8 108.6Right pace (cm) 112.0 80.3 80.3 182.0 231.7 123.9Pace angulation (8) 83.0 98.1 98.1 94.0 100.7 117.7Gleno-acetabular distance (cm) 98.3 104.0 104.0 261.3 216.0 126.7
Trackway gaugePes tracksPes trackway ratio (PTR) (%) 27.3 51.4 49.2 43.7 50.2 44.7[WAP/PL] ratio 2.3 0.8 0.9 1.3 0.9 1.0[WAP/IPS] ratio 2.9 0.9 1.1 1.4 1.1 1.2
Manus tracksManus trackway ratio (MTR) (%) 14.4 24.1 25.8 37.2 27.0 31.3[WAM/MW] ratio 4.0 3.2 2.2 1.9 2.7 2.1[WAM/IMS] ratio 5.6 4.4 4.8 2.0 3.7 2.9[PTR/MTR] ratio 1.9 2.1 1.9 1.2 1.9 1.4[WAP/WAM] ratio 0.90 0.50 0.67 0.97 0.71 0.92
[IPS/IMS] ratio (heteropody approximation) 1.8 2.4 1.8 1.4 2.4 2.2Pes trackway gauge Very wide Narrow Narrow Wide Narrow MediumLocomotion speed (km/h) 3.9 2.9 2.4 2.9 4.0 3.7
Notes: For details on the calculation of locomotion speed refer to text. The trackway pattern terminology follows Marty et al. (2006) and Marty (2008, p. 38, figure 2.16).Asterisks mark parameters, which were not measured in the field but calculated based on other parameters.
D. Marty et al.116
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

(digit and claw impressions clearly discernible) of the
Ajoie ichnoassemblages. The tracks are rather shallow
with small displacement rims, and they were probably left
under ideal substrate conditions such as a thin layer of
plastic carbonate mud or microbial mats. After track
formation a network of mud cracks with a diameter of 10–
20 cm was formed.
This quadrupedal trackway has a very narrow gauge
and a regular configuration, with manus located almost as
close to the trackway midline as pes tracks (Figure 6(G)).
Both pes and manus tracks are rotated outwards and the
rotation is much higher for the manus than for the pes
tracks, reaching up to 1078 (LM12). In particular, some
manus tracks are located well in front of the pes tracks,
closer to the following opposite pes track than to the
preceding pes track from the same side (e.g. LM5 closer to
RP5 than to LP5; Figure 5(C)).
Pes tracks have a subcircular to oval shape and are
rounded posteriorly and longer (36.0 cm mean length) than
wide (27.1 cm mean width) with the greatest width located
in the anterior part of the pes’ long axis but not so far away
from its midpoint. The best-preserved tracks show three
digits (digits I–III; decreasing in size) and two claw
impressions (Figure 6(H)–(J)). The latter are triangular in
shape, laterally directed and located on the tip of the first
and second digit impressions (Figure 6(J)).
Manus tracks are never overprinted, semicircular to
crescent shaped and convex forward, wider (23.1 cm mean
width) than long (13.3 cm mean length) and have a track
bottom inclined towards the anterior part of the track
(Figure 6(H)–(J)). Well-preserved manus tracks (e.g.
LM5, Figure 6(H)) lack, in contrast to the pes tracks,
impressions of digits and claws.
Trackway CTD-PMM-1505-S1
This straight, 12 m long trackway (Figures 5(D) and 6(K))
is located on the level 1505, about 5 cm above the main
track level (level 1500) of the upper levels, and to date it is
the only trackway excavated at this site. The trackway was
most probably left in a thin layer of moist carbonate mud
or microbial mat, superimposed on a hardened layer
because the tracks are – despite their large size – not very
deep. The soft sediment was squeezed outwards into the
displacement rims and the foot was put on the hardened
layer explaining the lack of anatomical details. These
tracks are considered as underprints and this is also
confirmed by the absence of tracks on the overlying
(removed) levels. After track formation, a network of mud
cracks with a diameter of 10–20 cm was formed.
This quadrupedal trackway has a regular configuration
with manus tracks located as close, or exceptionally closer,
to the trackway midline than the pes tracks (Figure 5(D)).
It has a wide gauge and the mean widths of the pes and
manus angulation patterns are almost equal. Pes and
manus tracks are always rotated outwards, manus having a
smaller rotation than pes tracks, and the right pes and
manus tracks showing a higher rotation than the left ones.
Well-preserved pes tracks are longer (108.7 cm mean
length) than wide (92.1 cm mean width), with the greatest
width located in the anterior part of the pes’ long axis, and
they do not exhibit digit and claw impressions. They are
bell shaped or slightly triangular and pointed posteriorly
(Figure 6(L) and (M)).
Manus tracks are never overprinted, semicircular to
oval and convex forward, wider (76.6 cm mean width)
than long (63.2 cm mean length) and they exhibit neither
digit nor claw impressions (Figure 6(N)).
Morocco
Two trackways named ‘Deio Lav-A’ and ‘Deio-D’ by
Belvedere (2008) are described, where ‘Deio’ is the
abbreviation for Demnat Iouaridene and ‘Lav’ for lavatory,
because this tracksite is located close to a small runnel,
where local women wash their cloths (Belvedere 2008). Both
trackways are located on the northern side of the Iouaridene
valley close to the village of Taghbalout (Figure 1(A))
and are easily accessible in the field; however, they have
been subjected to erosion and disintegration for several
decades. Over the past few years the GeoParc M’Goun, the
Association pour la protection du PatrimoineGeologique du
Maroc, and another local association, the Association des
Enseignants des Sciences de la Vie et de la Terre (AESVT),
undertook efforts to protect the localities in situ.
Trackway Deio Lav-A
This trackway is located on top of level 2 (Belvedere
2008), and it corresponds to the leftmost trackway
illustrated in the outline drawing of Ishigaki (1986,
figure 4) and to the trackway ‘1Ta1’ of Nouri (2007,
figures 214 and 215, ‘premier gisement du Taghbalout’).
The tracks are most probably true tracks because they are
well defined, quite deep, surrounded by large and well-
defined displacement rims, and because of the absence of
undertracks in the cross section of the right pes track at the
end of the trackway (Figure 2(A)). The latter fact also
suggests that the layer, in which the trackway was made
was a moist to water-unsaturated mud superimposed on an
already hardened or even lithified layer. After track
formation, a network of mud cracks with a diameter of
10–20 cm was formed.
The trackway Deio Lav-A is about 5 m long and
straight (Figures 5(F) and 6(O)), and it is the best-
preserved and leftmost trackway of at least five parallel
trackways left by medium-sized sauropods (Belvedere
2008). It is a quadrupedal trackway composed of five pes-
manus couples and an incomplete right pes track at the end
of the trackway. Manus tracks are overprinted to different
Historical Biology 117
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010
LP2
LM2
RP
2
RM
2
LP3
LM3
RP
3
LP4
RP
4
LM4
RM
4
LP3
RP
3
LM3
RM
3
LM1
RP
1
RM
1 LP
2
LM2
RP
2
RM
2 LP
3
LM3
RP
3
LM6
RP
6
RM
6
LP6
RM
4LP
5
LM5
RP
5
RM
5
LM4
RP
4
CH
E-C
RO
-500
-S10
3.9
km/h
Mar
ty (
2008
)C
heve
nez—
Com
be R
onde
Ear
ly /
Late
Kim
mer
idgi
an
CT
D-S
CR
-100
0-S
102.
9 km
/h
Mar
ty e
t al.
(200
3)C
ourte
doux
—S
ur C
ombe
Ron
deea
rly L
ate
Kim
mer
idgi
an
NW
Sw
itze
rlan
dM
oro
cco
Dei
o La
v-A
3.7
km/h
Bel
vede
re (
2009
)D
eio
Lav
(leve
l 2)
Late
Jur
assi
c
Dei
o-D
4.0
km/h
Bel
vede
re (
2009
)D
eio
(leve
l 3)
Late
Jur
assi
c
CT
D-T
CH
-105
5-S
42.
4 km
/hth
is w
ork
Cou
rtedo
ux—
Tch
âfou
èea
rly L
ate
Kim
mer
idgi
an
CT
D-P
MM
-150
5-S
12.
9 km
/hth
is w
ork
Cou
rtedo
ux—
Pom
mer
atea
rly L
ate
Kim
mer
idgi
an
Tra
ckw
ay n
umbe
rsp
eed
[km
/h]
Pub
licat
ion
(yea
r)T
rack
site
AgeT
rack
way
s
0.5
m0.
5 m
0.5
m0.
5 m
1 m
1 m
Tra
ckw
ay o
utlin
edr
awin
gs
A, B
, C &
F a
nd D
& E
,re
spec
tivel
y, a
re d
raw
n to
the
sam
e sc
ale
exte
rior
limit
of th
edi
spla
cem
ent r
im
exte
rnal
trac
k ou
tline
or c
rest
of t
hedi
spla
cem
ent r
im
inte
rnal
trac
k ou
tline
or tr
ue tr
ack
sens
u st
ricto
LP =
left
pes
LM =
left
man
usR
P =
rig
ht p
esR
M =
rig
ht m
anus
mea
n tr
ack
and
trac
kway
par
amet
ers
calc
ulat
ed r
atio
s an
dtr
ackw
ay g
auge
RP
2
RM
2
RP
3
RP
4
RM
3
LM2
LP3
LM3
LP4
LM4
LP5
LP2
D
C
B
A
E
F
pes
leng
th(P
L) [c
m]
left
pes
rota
tion
[˚]
right
pes
rota
tion
[˚]
man
us w
idth
(MW
) [c
m]
left
man
usro
tatio
n [˚
]rig
ht m
anus
rota
tion
[˚]
wid
th o
f the
ang
ulat
ion
patte
rn [c
m]
pes
(WA
P)
/ m
anus
(W
AM
)
pes
/ man
us tr
ackw
ay ra
tio [%
]
[WA
P/P
L]ra
tio [
][W
AM
/MW
]ra
tio [
]
30.0
8.
830
.5
19.1
29
.729
.0
46.9
21
.431
.0
23.3
28
.845
.3
36.0
30
.319
.0
23.1
53
.367
.8
108.
7 30
.0
47.3
76.6
6.
5 20
.3
110.
7 23
.034
.0
55.4
39
.548
.6
54.0
20
.318
.7
29.9
36
.710
.0
WA
P =
68.
9 / W
AM
= 7
6.2
WA
P=
37.1
/ W
AM
=74
.1W
AP
=33
.3 /
WA
M=
49.8
WA
P=
137.
9 / W
AM
=14
2.6
WA
P=
104.
8 / W
AM
=14
8.1
WA
P=
56.3
/ W
AM
=61
.3
27.3
/ 14
.451
.4 /
24.1
49.2
/ 25
.843
.7 /
37.2
50.2
/ 27
.044
.7 /
31.3
2.3
4.0
0.90
0.8
3.2
0.50
0.9
2.2
0.67
1.3
0.9
1.0
2.9
0.92
1.9
0.97
2.7
0.71
digi
tim
pres
sion
claw
impr
essi
on
rem
aini
ngin
fill
[WA
P/W
AM
]ra
tio [
]
pes
trac
kway
gau
geve
ry w
ide-
gaug
ena
rrow
-gau
gena
rrow
-gau
gew
ide-
gaug
ena
rrow
-gau
gem
ediu
m-g
auge
LP D
9(L
P6)
RM
D6
(RM
3)
LM D
7(L
M5)
RP
D8
(RP
4)
RM
D8
(RM
4)
RP
D6
(RP
3)
LM D
9(L
M6)
LP D
7(L
P5)
Fig
ure
5.
Ou
tlin
ed
raw
ing
so
fre
pre
sen
tati
ve
par
tso
fth
est
ud
ied
trac
kw
ays
wit
hin
dic
atio
no
fso
me
of
the
mea
ntr
ack
and
trac
kw
ayp
aram
eter
s.T
he
trac
kw
ayC
HE
-CR
O-5
00
-S1
0(A
)h
asth
ew
ides
tan
dth
etr
ack
way
CT
D-S
CR
-10
00
-S1
0(B
)h
asth
en
arro
wes
tg
aug
e.N
ote
that
the
illu
stra
ted
trac
ks
of
the
trac
kw
ayD
eio
-D(Breviparopusreference
trackway)(E)
arenotthesameas
those
illustratedforBreviparopusin
Figure
8(A
).
D. Marty et al.118
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Figure 6. Photographs of representative segments and tracks of the studied trackways from the Jura Mountains, NW Switzerland (A–N)and from the central High Atlas Mountains, Morocco (O–T). (A–C) CHE-CRO-500-S10. (D–F) CTD-SCR-1000-S10. (G–J) CTD-TCH-1055-S4. (K–N) CTD-PMM-1505-S10. (O–Q) Deio Lav-A. The arrow in Q points to the small lateral indentation at the interiorside in the middle of the manus track. (R–T) Deio-D (Breviparopus reference trackway).
Historical Biology 119
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

degrees, but one (LM2, Figure 6(Q)) is complete. The
heteropody ratio varies between 1:3 for the more
complete, semicircular manus tracks and 1:4 for the
incomplete, crescent-shaped manus tracks. The trackway
falls into the medium-gauge field and is clearly wider than
the trackway Deio-D. Pes and manus tracks are outward
rotated and manus tracks are generally located slightly to
the inside of the prolongation of the pes’ long axis. This is
also indicated by a similar width of the angulation pattern
for pes and manus tracks.
The pes tracks are sub-elliptical to bell shaped and are
less triangular in outline than those of the trackway Deio-
D. Pes tracks are longer (54.0 cm mean length) than wide
(40.5 cm mean width), with the greatest width located
between the middle and anterior part of the pes’ long axis
(Figure 6(P)). Notably the two left pes tracks LP1 and
LP2 exhibit several small depressions in their anterior
part, and these were interpreted as three to four digit
impressions separated by interdigital displacement rims
by Ishigaki (1986, figure 4) and Nouri (2007, figure 214).
Nonetheless, these impressions cannot clearly be ident-
ified as digit impressions because the track bottom is
generally very irregular. This can be explained, for
instance, by the adherence of the sediment to the foot
during withdrawal or due to a partial collapse of the track
interior. Displacement rims are large and well defined and
are best developed in the anterolateral exterior margin of
the tracks, where they cut across the displacement rims of
the manus tracks. Displacement rims are shallower at the
interior part and very shallow or absent in the rear part of
the pes tracks.
Manus tracks are crescent shaped to semicircular,
always wider (29.9 cm mean length) than long (14.5 cm
mean length), without digit and claw impressions (Figure
6(Q)). Manus tracks are generally placed on, inside and
outside the prolongation of the pes’ long axis and are
deformed by the subsequent pes track to different degrees
(Figure 6(P)). Contrary to the trackway Deio-D, manus
tracks of Lav-A are clearly deeper than the pes tracks and
are surrounded by well-defined, larger displacement rims
cut by those of the pes. The semicircular manus tracks are
suggested to have been left when the trackmaker did not
overprint it with the pes and when it put its manus in a less
digitigrade way resulting in a better impression of the shape
of the trackmaker’s manus. Such well-defined manus
tracks exhibit a small lateral indentation at the interior side
in the middle of the track (e.g. LM2, Figure 6(Q)), which is
probably a characteristic of the manus.
Trackway Deio-D
This trackway (trackway ‘4Am1’ of the ‘quatrieme
gisement d’Aıt Mimoun’ of Nouri 2007) is the reference
trackway of B. taghbaloutensis, first described by Dutuit
and Ouazzou (1980) and later illustrated by Ishigaki (1989,
figure 9.5). The trackway lies on the main track level
(named level 3), and all tracks of level 3 are interpreted as
true tracks. Undertracks are visible on a mud-cracked level
about 10 cm below level 3, where the true tracks of level 3
are already eroded. This suggests that the sediment was
moist and well laminated when the tracks were left,
possibly due to the former presence of microbial mats.
After track formation, a network of mud cracks with a
diameter of 5–10 cm was formed.
The trackway Deio-D (Figures 5(E) and 6(R)) is a
narrow-gauge, quadrupedal trackway composed of 33
tracks, and it is about 86 m long, even though there is a
22 m long gap between the tracks D/20 and D/21 due to the
erosion of the track level. After this gap the preservation is
poor, and one right pes–manus couple and a pes are
missing between the prints D/22 and D/23, while two other
pes–manus couples are missing between D/29 and D/30.
Trackway direction changes twice along the trackway
from 2628 to 2508 and then back to 2698, while the stride
length remains almost constant. The pes rotation is
outward and different between left and right tracks: the
average outward rotation for the left pes tracks is 158,
while for the right ones it is 308, without notable changes
in rotation close to the turns. The position of the manus is
almost always slightly more external with respect to the
prolongation of the pes’ long axis, also indicated by a
larger width of the angulation pattern of the manus. Manus
tracks also have an outward rotation, which is slightly
higher than for the pes tracks. Pes tracks are clearly deeper
(up to twice as deep) than the manus tracks, this was also
observed by Dutuit and Ouazzou (1980). Because all
manus tracks are at least partially overprinted, heteropody
varies from 1:3 to 1:4.5 depending on the degree of
overprinting. Dutuit and Ouazzou (1980) calculated a
heteropody ratio of 1:3.6, and Nouri (2007) of 1:4.5.
However, for undeformed manus tracks, heteropody is
suggested to be rather in the order of 1:3.
Pes tracks are longer (110.7 cm mean length) than wide
(88.5 cm mean width), the shape varies from elliptical to
bell shaped, but it is often slightly triangular (i.e. pointed
posteriorly). The maximum width of the track is generally
in the anterior part of the pes’ long axis, whereas in the bell
shaped to triangular tracks it is positioned more anteriorly.
Displacement rims are always present and are more
developed on the anterior and on the external sides of the
tracks, while they are smaller or sometimes even absent in
the posterior (rear) part of the pes tracks (Figure 6(S) and
(T)). Dutuit and Ouazzou (1980) stated that the pes tracks
have at least four digit impressions marked by three
interdigital displacement rims and accordingly to their
Figure 1, there are at least three claw impressions present.
Also, in the outline drawing of four pes–manus couples
illustrated in Ishigaki (1989, figure 9.5) (Figure 8(A)), up
to four digit impressions and up to three claw impressions
are drawn. On the other hand, Meyer and Monbaron
D. Marty et al.120
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

(2002), who re-examined the trackway, stated that ‘neither
manus nor pes imprints show traces of claws’. However,
only the track Deio-D/28, also illustrated by Ishigaki
(1989, figure 9.5, second right), exhibits three faint
depressions in the anteriormost part of the track, which
could be interpreted as digit impressions, and the claw
impressions figured by Dutuit and Ouazzou (1980) and
Ishigaki (1989) are reinterpreted here as desiccation cracks
dissecting the footprint outline. Thus, in their present
preservation state (in May 2009), the evidence for digit or
claw impressions is weak, and it is suggested that this is
not an unequivocal characteristic of the trackway Deio-D.
None of the manus tracks is completely unaffected by
the subsequent pes, and their shape varies from
semicircular to crescent shaped depending on the degree
of overprinting by the subsequent pes track (Figure 6(T)).
In one case, the pes (D/16) almost completely overprints
the manus track. Manus tracks are always wider (55.4 cm
mean width) than long (29.7 cm mean length). No digit or
claw impressions are present on any of the manus tracks
and this is in accordance with the description of Dutuit and
Ouazzou (1980). Displacement rims are well developed all
around the tracks and are cut by the displacement rims of
the subsequent pes.
Comparison of studied trackways
Basically, the trackways from NW Switzerland and
Morocco are similar and with the exception of the
trackway CTD-TCH-1055-S4 (Figure 5(C)) they all lack
anatomical details such as digit and claw impressions.
Thus, the most important differences that can be observed
regarding track preservation are overall pes and manus
track outlines (from oval to bell shaped), trackway gauge
(from very narrow to very wide gauge), the position of pes
and manus tracks with respect to the trackway midline
(manus as close to midline as pes up to manus clearly
farther away from midline as pes tracks), the degree of pes
and manus outward rotation and heteropody.
Track preservation
Despite the different sedimentological settings of the
tracksites in Morocco (siliciclastic) and NW Switzerland
(carbonate), the studied trackways are similar from a
preservational point of view. All trackways have relatively
shallow tracks, which are generally surrounded by well-
defined displacement rims. This suggests that the substrate
in both paleoenvironments was well consolidated possibly
due to the presence of microbial mats (Marty et al. 2009), or
that it was rapidly consolidating due to early diagenetic
processes, which may have been induced by microbial mats
(e.g. Chafetz and Buczynski 1992 and Dupraz and Visscher
2005). Under such circumstances, even very large and
heavy trackmakers (e.g. sauropods with a pes length of
.1 m) are unlikely to leave tracks deeper than 0.2 m.
Some of the studied tracksites are characterised by the
presence of true tracks with underlying shallow under-
tracks, others by true tracks without undertracks and at
some by the presence of underprints. Shallow undertracks
indicate biolaminated and plastic sediment, strongly
suggesting the former presence of microbial mats.
Underprints and/or the absence of undertracks indicate a
moist layer, which was squeezed outwards on an already
consolidated layer by the pressure of the foot forming
displacement rims (Marty et al. 2009). This case is well
expressed in the cross section of a track of the Moroccan
Deio Lav-A trackway (Figure 2(A)), where the underlying
clays were clearly not affected by the impact of the foot,
and thus was already consolidated at the time of track
formation. Deio Lav-A has the deepest tracks with the
largest displacement rims of all trackways studied. When
the sediment was completely squeezed outwards, the
actual imprint of the foot may have also been left on a
level, which was situated within the sediment prior to track
formation. Such tracks are called underprints (sensuMarty
et al. 2009), and this scenario is expressed by the trackway
CTD-PMM-1505-S1 and explains why this trackway does
not show anatomical details of the feet of its trackmaker.
With the exception of some tracks of the trackway
CTD-TCH-1055-S4, none of the trackways has tracks with
substantial anatomical details of the foot. This is either
related to unsuitable substrate properties (e.g. compo-
sition, grain size, moisture content and the presence or
absence of microbial mats), covering of the foot with
adhering sediment, unsuitable behaviour of the trackmaker
(e.g. moving or dragging of feet during track formation), to
overgrowth and modification by microbial mats forming
overtracks and resulting in amalgamated track infills, or to
a combination of the above factors. In the case of the
Moroccan trackways, this may further be related to recent
weathering due to their long exposure, and this may
occasionally have modified the track outlines.
Because the preservational setting of all studied
trackways is similar in as far that the sediment was not
susceptible for deformation to greater depths (even though
it cannot be excluded that it was slippery in some cases),
differences in substrate properties are not very likely to be
the reason for the observed variations in trackway
configuration and gauge described below.
Pes and manus track morphology
The overall pes track outline varies from oval and rounded
posteriorly (CTD-SCR-1000-S10) to bell shaped and
pointed posteriorly (CTD-PMM-1505-S1, Deio-D). Pes
tracks are always longer than wide, and the maximum
width is located in the anterior half of the track with
respect to the midpoint of the pes’ long axis. The position
Historical Biology 121
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

of its intersection with the pes’ long axis (i.e. the reference
point) is generally closer to the midpoint of the pes’ long
axis, even though in the two trackways with the largest
tracks (CTD-PMM-1505-S1 and Deio-D) it is closer to the
anterior end of the pes’ long axis. Some pes tracks of
the trackway CTD-TCH-1055-S4 have impressions of the
digits I–III and of two strongly outward rotated claws on
digits I and II.
Manus tracks are always wider than long, the overall
outline of undeformed and well-preserved manus tracks
varying from crescent shaped (CTD-SCR-1000-S10,
CTD-PMM-1505-S1) to semicircular (CHE-CRO-500-
S10, CTD-TCH-1055-S4, Deio Lav-A) and digit and
claw impressions are never discernible. Even along
single trackways, the shape of manus tracks is subjected
to strong variation, possibly related to a more or less
digitigrade posture of the manus depending on the
behaviour of the trackmaker. The best-defined manus
tracks are those of trackways CTD-TCH-1055-S4 and
Deio Lav-A, both being strongly inclined towards the
anterior part of the track and showing some evidence for
an anterior crescent-shaped groove. This possibly
indicates that digits II–IV were bound together in a
pad and that the manus was put in a pronounced
digitigrade way. The depth of pes and manus tracks is
always in the same order; only the trackway Deio-D has
slightly deeper pes than manus tracks.
Trackway configuration
The most obvious differences in trackway configuration
are the degree of overprinting of manus by the subsequent
pes tracks, the degree of outward rotation of pes and manus
tracks, heteropody, trackway gauge and the position of pes
and manus tracks with respect to each other and the
trackway midline.
Apart from CTD-TCH-1055-S4, where the manus
tracks are well in front of the pes tracks and closer to the
subsequent pes track of the opposite side, manus tracks are
located close to the subsequent pes tracks. Partial
overprinting of the manus by the subsequent pes occurs
(at least sometimes) in the trackways CHE-CRO-500-S10,
CTD-SCR-1000-S10, Deio-D and Deio Lav-A. However,
there is no evidence to suggest that this is related to
trackway gauge or locomotion speed, even though it is
generally assumed that overprinting in trackways of
quadrupedal tetrapods rather occurs at higher speeds (e.g.
Peabody 1959 and Haubold 1971). Nonetheless, Marty
(2008) observed on a trackway sample of the CHE-CRO
tracksite that overprinting tends to be associated with
lower locomotion speeds.
Pes and manus tracks are always rotated outward and
with the exception of CTD-PMM-1505-S1, manus tracks
generally have a higher outward rotation. Pes outward
rotation is in the order of 20–308 and manus outward
rotation in the order of 30–708, even though important
differences between left and right tracks are common (e.g.
CTD-PMM-1505-S1, Deio Lav-A). Manus outward
rotation may also be extremely pronounced with up to
1078 in CTD-TCH-1055-S4.
Heteropody, as expressed by the ratio between the
indices of pes and manus track size, is highest for CTD-
SCR-1000-S10 and Deio-D (2.4), followed by Deio Lav-A
(2.2). The trackways CHE-CRO-500-S10, CTD-TCH-
1055-S4 and notably CTD-PMM-1505-S1 (1.4) have
ratios smaller than 2.0.
Figure 7(A) shows that the [PTR] ratio and the
[WAP/PL] ratio give similar results for the gauge of the
trackways analysed, and this was also obtained by Marty
(2008, p. 163, figure 5.38) for other trackways from NW
Switzerland and selected ichnotaxa and other published
trackways. Gauge varies from narrow (CTD-SCR-1000-
S10, CTD-TCH-1055-S4, Deio-D) over medium (Deio
Lav-A) to wide (CTD-PMM-1505-S1) and very wide
(CHE-CRO-500-S10) (Figure 7(A) and (B), Table 1). The
trackway Deio Lav-A falls into the medium-gauge field
with a [WAP/PL] ratio slightly higher than 1.0 and a
PTR clearly smaller than 50%. The trackway CTD-SCR-
1000-S10 with a [WAP/PL] ratio of 0.8 and a PTR of 51.4
is the narrowest, and the trackway CHE-CRO-500-S10
with a [WAP/PL] ratio of 2.3 and a PTR of 27.3 is the
widest one. The [WAP/PL] ratio poorly correlates with
both mean pes length (Figure 7(C)) and locomotion speed
(Figure 7(D)) suggesting that trackmaker size and
differences in speed do not significantly contribute to the
observed differences in gauge. In Figure 7(B), the
[WAM/WM] ratio shows the variation in manus trackway
gauge. Manus trackway gauge is higher in the trackway
CHE-CRO-500-S10 because this trackway is much wider
than the other trackways, and it is comparatively high in
the narrow-gauge trackways CTD-SCR-1000-S10 and
Deio-D because of their pronounced exterior manus
position.
For all studied trackways, the ratio between the
widths of the pes and manus angulation pattern (i.e. the
[WAP/WAM] ratio) is smaller than 1.0 (Figure 7(B), (E)
and (F)) and thus manus tracks are located farther away
from the trackway midline than pes tracks. The trackway
outline drawings shown in Figure 5 and the distribution
of the trackways shown in Figure 7(E) and (F) further
suggest that the narrower trackways (CTD-SCR-S10,
CTD-TCH-1055-S4, Deio-D) tend to have manus tracks
located farther away from the trackway midline than the
wider trackways (Deio Lav-A, CTD-PMM-1505-S1,
CHE-CRO-500-S10), and that this configuration is
independent of trackmaker size because the mean pes
length correlates poorly with the [WAP/WAM] ratio
(Figure 7(E)). However, it has to be considered that the
good positive correlation in Figure 7(F) between the
[WAP/PL] ratio and the [WAP/WAM] ratio may be
D. Marty et al.122
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

related to the fact that both ratios have WAP in the
numerator. The poor correlation in Figure 7(C)–(E), on
the other hand, may be related to the fact that the
trackways were left by different sauropod trackmaker
species. This is very likely the case for the trackways of
NW Switzerland when compared to those of Morocco,
and it is also possible that the trackways of NW
Switzerland were left by different sauropods.
Summary
The trackways from NW Switzerland and Morocco wereleft by sauropods ranging in size from small to very large.The pes and manus tracks of all studied trackways have asimilar morphology (overall track outline), notably alsothe narrow- and very wide-gauge trackways within theNW Switzerland trackway samples. Pes tracks are alwayslonger than wide, more or less oval to bell shaped and
A B
C D
E F
PBro
Bro
Bro
Bro
PBro
Bre
Deio-D
Deio Lav-A
Bro
PBro
Deio Lav-A
Deio Lav-A
CTD-PMM-1505-S1
PB
roDeio Lav-A
narrow medium wide
wid
em
ediu
mna
rrow
0.8
30
120
110
100
90
80
70
Mea
n pe
s le
ngth
[cm
]
Loco
mot
ion
spee
d [k
m/h
]60
50
40
30
1.2
1.1
1
0.9
0.8
0.7
0.6
0.5
40
50
1 1.2 1.4 1.6
[WAP/PL] ratio
[WAP/PL] ratio
[WA
P/W
AM
] rat
io
1.2
1.1
1
0.9
0.8
0.7
0.6
0.5
[WA
P/W
AM
] rat
io
Pes
trac
kway
rat
io [%
]
1.8 2 2.2 2.4
0.8
30 40 50 60 70 80Mean pes lenght [cm]
90 100 110 120
1 1.2 1.4 1.6 1.8 2 2.2 2.4
[WAP/PL] ratio
0.8 1 1.2 1.4 1.6 1.8 2 2.2 2.4
[WAP/PL] ratio0.8 1 1.2 1.4 1.6 1.8 2 2.2 2.4
very wide
CT
D-T
CH
-105
5-S
4CTD-PMM-1505-S1
CTD-PMM-1505-S1
CTD-PMM-1505-S1
Dei
o-D
& B
re
Dei
o-D
& B
reD
eio-
DB
reP
Bre
N = 9r = –0.97r = –0.92
N = 9r = –0.26r = –0.10
N = 9r = 0.56r = 0.78
N = 9r = –0.18r = –0.33
N = 9r = 0.16r = 0.07
CHE-CRO-500-S10
4CTD-S
CR-1000-S10
Parabrontopodus
Deio-DBrevip
aropus
CTD-TCH-1055-S
4
Deio Lav-A
CTD-PMM-1505-S
1
Brontopodus
CHE-CRO-500-S
10
3.5
3
2.5
Tra
ckw
ay g
auge
rat
ios
2
1.5
1
0.5
7
6
5
4
3
2
1
[WAP/PL] ratio [WAM/MW] ratio [WAP/WAW] ratio
Dei
o-D
CT
D-S
CR
-100
0-S
10
CT
D-T
CH
-105
5-S
4
CH
E-C
RO
-500
-S10
CT
D-S
CR
-100
0-S
10
CT
D-T
CH
-105
5-S
4
CHE-CRO-500-S10
CT
D-S
CR
-100
0-S
10
CT
D-T
CH
-105
5-S
4
Deio Lav-A
Bro
CTD-PMM-1505-S1
CHE-CRO-500-S10
CT
D-S
CR
-100
0-S
10
CHE-CRO-500-S10
CTD-TCH-1055-S4
CTD-SCR-1000-S10
man
us c
lose
rto
mid
line
pes
clos
er to
mid
line
Bre
Figure 7. Plots exhibiting the differences in trackway gauge and in the position of pes and manus tracks with respect to the trackwaymidline among the studied trackways and for the three sauropod ichnotaxa included in the discussion. For the data used, refer to Table 1(studied trackways) and Table 2 (sauropod ichnotaxa). (A) Plot of the [WAP/PL] ratio against the pes TR. (B) Variation of the [WAP/PL]ratio, the [WAM/MW] ratio and the [WAP/WAM] ratio. (C) Plot of the [WAP/PL] ratio against the mean pes length. (D) Plot of the[WAP/PL] ratio against locomotion speed. (E) Plot of the mean pes length against the [WAP/WAM] ratio. (F) Plot of the [WAP/PL] ratioagainst the [WAP/WAM] ratio. In (A) the boundaries between the different gauge types with respect to the PTR ratio follow Romano et al.(2007), and in (A), (C), (D) and (F) the gauge boundaries with respect to the [WAP/PL] ratio follow Marty (2008). Bro is Brontopodus;PBro, Parabrontopodus and Bre, Breviparopus.
Historical Biology 123
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

occasionally slightly triangular (pointed posteriorly). The
latter pes tracks may be related to a slight sliding of the pes
on the surface while the track was left, or in the case of the
Moroccan trackway Deio-D also to recent weathering or to
a combination of both. A sliding or moving of the pes
during track formation may also explain the absence of pes
digit and claw impressions in some of the trackways.
Manus tracks, if undeformed by the subsequent pes, are
always wider than long, semicircular or slightly crescent
shaped, and without evidence for a claw impression on
digit I. This reflects the shape of the feet to a certain
degree, but the form of the tracks is also influenced to a
certain degree by the behaviour of the trackmaker and/or
the substrate properties (e.g. slippery, sediment backflow
into track). Manus tracks are often incomplete impressions
because the manus was put in a more pronounced
digitigrade way, resulting in the lack of the impression of
the rear part of the manus (e.g. Marty 2008). Such manus
tracks are inclined towards the anterior part of the track
(Figure 6(H)–(J)).
Apart from the wide range of trackway gauge, they are
also characterised by a similar general trackway configur-
ation with pes and manus tracks rotated outwards (where
manus tracks generally have a higher outward rotation than
pes tracks), more or less pronounced heteropody and
centres of manus being placed farther away from the
trackway midline than those of the pes tracks.
The trackways CTD-SCR-1000-S10 and Deio-D are
similar because they have a narrow gauge, manus tracks
located on the prolongation of the pes’ long axis and farther
away from the trackway midline than the pes and a
pronounced heteropody. CTD-TCH-1055-S4 is also
narrow gauge but it differs because of a smaller heteropody
coefficient, manus tracks located well in front of the pes
tracks and to the inside of the prolongation of the pes’ long
axis, a very high manus outward rotation and because of the
presence of digit and claw impressions in the pes track.
Deio Lav-A is different because it is a medium gauge, has
manus tracks located not much farther away from trackway
midline than pes tracks and relatively deep tracks.
CHE-CRO-500-S10 and CTD-PMM-1505-S1 are
similar because they have a wide gauge, a less
pronounced heteropody and semicircular manus tracks
located almost as close to trackway midline as pes
tracks. These characteristics are different from the other
trackways analysed. Nonetheless, regarding the overall
pes and manus track outline, even these two trackways
are similar to the other ones, notably the bell-shaped pes
tracks of CTD-PMM-1505-S1 are similar to the ones of
Deio-D.
Concluding this section, more trackways (including
wide-gauge ones) with digit and claw impressions need to
be examined for a more accurate distinction of the
trackways from NW Switzerland and Morocco, and also
for a distinction within the trackway samples of NW
Switzerland (notably between the narrow- and wide-gauge
trackways) and of Morocco.
Discussion
The aim of this discussion is to briefly review important
Late Jurassic to Early Cretaceous sauropod ichnotaxa, to
provide a differential diagnosis (based on track mor-
phology and trackway configuration) for the studied
trackways from NW Switzerland and Morocco, and to
evaluate whether they can be assigned ichnotaxonomically
and whether possible trackmakers can be identified.
Review of Late Jurassic to Early Cretaceous sauropodichnotaxa
Sauropod ichnotaxa were reviewed by Farlow et al.
(1989), Farlow (1992), Lockley et al. (1994b), Meyer et al.
(1994), Dalla Vecchia and Tarlao (2000), Avanzini et al.
(2003), Dalla Vecchia (2005) and Wright (2005), and
Table 2. Data source of sauropod ichnotaxa used for the plots of Figure 7.
Ichnotaxon B. taghbaloutensis P. mcintoshi B. birdi
References (a) Ishigaki (1989),(b) Belvedere (2008)
(c) Lockley et al. (1994),(d) Lockley et al. (1986)
(e) Farlow et al. (1989),(f) Romano et al. (2007)
Outline drawing in Figure 8 after (a, figure 9.5) (c, figure 7b) (e, figure 42.3)PL (cm) 110.7 (b) 60 (d) 86.5 (e)PTR (%) 50.2 (b) 51.90 36.04 (f)[WAP/PL] ratio 0.90 0.87 1.46
Wap measured on outline drawingPL after (d, figure 9.5) (b) (d, figure 7b) (d) (f, figure 3a) (e)
[WAM/MW] ratio 2.20 2.03 2.32WAM measured on outline drawingMW after (a, figure 9.5) (b) (d, figure 7b) (d) (f, figure 3a) (e)
[WAP/WAM] ratio 0.66 0.81 1.13Trackway gauge Narrow Narrow WideLocomotion speed (km/h) 4.00 (b) 6.80 (d) 1.21 (this work)
D. Marty et al.124
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

currently about a dozen of ichnotaxa are attributed to
sauropods. Of these, only a few are considered as valid by
the majority of the authors cited above, and in this
discussion, the ichnotaxa Breviparopus and Parabronto-
podus from the Late Jurassic and Brontopodus from the
Early Cretaceous are considered.
Breviparopus Dutuit and Ouazzou, 1980
The reference trackway of Breviparopus corresponds to
the trackway Deio-D of this study. According to the
detailed descriptions provided by Dutuit and Ouazzou
(1980), it is characterised by:
(1) A clearly narrow-gauge trackway with pes tracks
intersecting the trackway midline (l’axe de la piste
passe par la partie posterieure des empreintes des
pattes posterieures).
(2) Manus tracks located further away from the trackway
midline than pes tracks (l’empreinte du membre
anterieur est d’avantage ecartee de l’axe de la
marche que celle du membre posterieur).
(3) Manus tracks located in front of pes tracks and being
(slightly) overprinted by the pes tracks (l’empreinte
laissee par le membre posterieur suit immediatement
celle du membre anterieur, le bourrelet anterieur de
la premiere et le bourrelet posterieur de la seconde se
confondant).
(4) Manus tracks with a slightly semicircular shape
without evidence for digit impressions (l’empreinte
de patte anterieure est sensiblement en demi-cercle
. . . Aucun doigt n’est visible).
(5) Pes tracks with a slightly oval shape with at least four
digit impressions separated by three small interdigital
displacement rims, and evidence for claw marks
within the overall displacement rim (l’empreinte de
patte posterieure s’inscrit dans une figure geome-
trique intermediaire entre le triangle a sommet
posterieur et petit cote anterieur et l’oval deforme
s’effilant en arriere . . . on remarque aussi la presence
d’encoches sur le bourrelet anterieur de l’empreinte
posterieure, bourrelet qui temoigne ainsi de l’exist-
ence de griffes . . . au moins quatre doigts porteurs a
sa patte posterieure. On constate en effet la presence
de trois bourrelets interdigitaux).
(6) A pronounced heteropody in the order of 1:3.6
(La surface d’empreinte posterieure est donc 3.6
fois plus grande que celle de l’empreinte
anterieure).
We agree with this general description, but we could
not observe the described digit and claw impressions. The
trackway in the current preservation state – contra the
published outline drawings of Dutuit and Ouazzou (1980)
and Ishigaki (1989), illustrated in our Figure 8(A) – does
not show any evident digit and claw impressions, and this
was already noted by Meyer and Monbaron (2002). The
outline drawing in Ishigaki (1989) illustrates well the gross
outline of the Breviparopus pes tracks, but the illustration
of claws is misleading. In our opinion, digit and claw
impressions are also not clearly visible on the original
photos of the trackway published by Dutuit and Ouazzou
(1980). Furthermore, the Deio-D trackway is slightly
asymmetric with left manus tracks being located closer to
the trackway midline than the right ones and having a
smaller outward rotation than the right ones (Figures 5(E),
6(R) and 8(A)). At least, some of the left manus tracks are
located as close to the trackway midline as the
Figure 8. Outline drawings (drawn to scale) of Late Jurassic toEarly Cretaceous sauropod ichnotaxa as they were originallypublished. (A) Because Dutuit and Ouazzou (1980), whodescribed B. taghbaloutensis, only illustrated selected tracksbut not a trackway segment, here the outline drawing of Ishigaki(1989, figure 9.5, p. 85) is shown. These tracks correspond to thetracks D/25 to D/28 of Belvedere (2008). Compare the outlinedrawing of B. taghbaloutensis with our interpretation shown inFigure 5(E) and the photographs in Figure 6(R)–(T), even if theyare not the same tracks. (B) Outline drawing of the holotypetrackway CU-MWC 190.5, from Lockley et al. (1986; figure 7b,p. 1170). (C) Bird’s outline drawing (without the tridactyl tracks)of the slab excavated for the American Museum of NaturalHistory, from Farlow et al. (1989; figure 42.3, p. 374).
Historical Biology 125
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

corresponding subsequent pes tracks, contrary to Dutuit
and Ouazzou (1980), who stated that manus tracks are
located further away from the trackway midline than pes
(point 2 above).
A pes TR larger than 50% and a [WAP/PL] ratio
clearly smaller than 1.0 confirm that Breviparopus has,
despite a comparatively pronounced outward pes rotation,
a narrow gauge (Figure 7(A) and (B)), even if our PTR
(50.2%) is lower than that (51.48 and 53.54%) calculated
by Romano et al. (2007) based on the outline drawings of
Ishigaki (1989) and Thulborn (1990), respectively. This is
due to the fact that these published outline drawings refer
to the best-preserved and narrower trackway segment (i.e.
tracks Deio-D/25 to Deio-D/28), while our PTR is
calculated based on the whole trackway. Breviparopus is
further characterised by a low [WAP/WAM] ratio
indicating that the manus tracks are located clearly farther
away from the trackway midline than the pes tracks.
Dutuit and Ouazzou (1980) introduced the name
B. taghbaloutensis stating that they use it for description
only and without any taxonomical value. Nonetheless, this
ichnotaxon is generally considered as valid and was used
by Farlow (1992) as the ‘type’ example of a narrow-gauge
trackway. However, we suggest that Breviparopus needs to
be formally erected based on the reference trackway.
Because this trackway may have suffered from erosion
during the last 30 years, the description (notably the
presence and position of digit and/or claw impressions)
has to be verified and/or emended (cf. Sarjeant 1989;
Billon-Bruyat and Mazin 2003) based on new, better-
preserved tracks that could possibly be found by further
excavating at the beginning of the Deio-D trackway. The
type trackway should then be protected in situ and a cast of
at least one well-preserved pes–manus couple should be
made and housed in an official institute or museum.
Otherwise, the tracks risk rapid disintegration, continu-
ously lowering their ichnotaxonomic value.
Parabrontopodus Lockley, Farlow and Meyer 1994
This ichnotaxon is based on a trackway from the
Purgatoire Valley Dinosaur tracksite, USA (Unit B of
the Morrison Formation, Late Jurassic), described as:
Narrow sauropod trackway of medium to large size(footprint length about 50–90 cm), characterized by nospace between trackway midline and inside margin of pestracks. Pes footprint longer than wide with long axisrotated outward. Pes claw impressions, corresponding todigits I, II and III show strong outward rotation. Manustracks semicircular and small in comparison with pestracks (i.e. pronounced heteropody) (Lockley et al. 1994b)(Figure 8(B)).
Lockley et al. (1994b) further noted that Parabrontopodus
has a heteropody coefficient of about 1:4 or 1:5.
Even though Wright (2005) stated ‘Lockley et al.
(1994b) did not note how Parabrontopodus could be
differentiated from Breviparopus’ and that for this reason
‘Parabrontopodus may be a junior synonym of Brevipar-
opus’, Parabrontopodus is considered as valid here, also
because Breviparopus was never formally erected. A pes
TR larger than 50% and a [WAP/PL] ratio clearly smaller
than 1.0 confirm that Parabrontopodus is characterised by
a narrow gauge, in about the same order as Breviparopus
(Figure 7(A) and (B)).
The Parabrontopodus type tracks from the Purgatoire
Valley are found in high-energy oolitic shoreface
limestones with interbedded lacustrine shales (Lockley
et al. 1986; Prince and Lockley 1989), and judging from
illustrations in Lockley et al. (1986, 1994b), the holotype
tracks are relatively deep and do not have displacement
rims because they were left in a thick layer of water-
unsaturated carbonate mud with a high plasticity, and this
may be the reason why the claw impressions are quite well
defined.
Brontopodus Farlow, Pittman and Hawthorne 1989
This ichnotaxon was defined based on a trackway from the
Paluxy River Valley near Glen Rose, Texas (Trinity
Group, Aptian–Albian, Early Cretaceous), based on both
track morphology and trackway parameters (Figure 8(C)).
Farlow et al. (1989) stated that it is unknown
which features are diagnostic at the ichnogenus level
and provided the following detailed diagnosis for
Brontopodus:
Sauropod ichnites of small to large size, known pesfootprint length ranging 50 to over 100 cm. Manusfootprint length and width about the same in well-preserved tracks; manus tracks clawless, somewhat U-shaped, with digit impressions I and IV slightlyseparated from the impression of the conjoined digitsII–IV. Pes tracks longer than broad, with large, laterallydirected claw marks at digits I–III (diminishing in sizefrom I to III), a small claw, nail, or callosity mark atdigit IV, and a small callosity or pad mark at digit V;digit marks IV and V only seen in well-preservedfootprints. Manus tracks often (usually?) rotated outwardwith respect to direction of travel. Manus track medial toa line through pes track long axis, such that manus trackcentres are somewhat closer to the trackway midlinethan pes track centres. Trackway broad, with left andright manus and pes footprints often well away from thetrackway midline; trackway width roughly 1–1.5 timespes track length. Outer limits of trackway defined by pestracks. Manus-pes distance 0.5–1.2 times pes footprintlength. Stride length roughly 2–5 times pes track length.Step angle generally 1008–1208. Glenoacetabular lengthc. 3–4 times pes track length. Tail drag marks rare orabsent.
The B. birdi trackways of the Paluxy River are very well
preserved, and this has to be considered when comparing
these tracks with other less well-preserved tracks. Also, it
has to be born in mind that the Brontopodus tracks from
the Paluxy River Valley are very deep without displace-
D. Marty et al.126
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

ment rims and with a well impressed foot morphology
including the claws because they were left in a nearshore
carbonate setting (e.g. Farlow 1987 and Pittman 1989,
1990, 1992) possibly in a thick layer of water-unsaturated
carbonate mud with a high plasticity (personal observation
DM). Walking in such deep sediment certainly had an
impact on the trackmaker’s behaviour, and the pronounced
claw impressions may also be related to a functional
reason, e.g. they may have improved the animal’s footing
providing a ‘snowshoe’ effect (Gallup 1989), or as
suggested by Pittman and Gillette (1989) that ‘ . . . on
firmer substrates the claws were “wrapped” around the
lateral margin of the foot, and on softer substrates the
claws were often extended forward, probably to provide
better traction’.
The pes TR in the type trackway (Farlow et al. 1989)
varies from c. 32.5% to nearly 38% and characterises
Brontopodus as medium gauge to wide gauge (Romano
et al. 2007, p. 261 and figure 15A), and the [WAP/PL] ratio
as wide gauge, respectively (Figure 7(A) and (B); Table 2).
Brontopodus is further characterised by a [WAP/WAM]
ratio higher than one indicating that the manus tracks are
located closer to the trackway midline than the pes tracks,
and this is also indicated by a comparatively small
[WAM/WM] ratio (Figure 7(B)). Brontopodus has a
heteropody of about 1:3 (Lockley et al. 1994b).
Differential diagnosis
Regarding Breviparopus
The Moroccan trackway Deio-D is the reference trackway
of Breviparopus, and it is similar to the Swiss trackway
CTD-SCR-1000-S10 because of its narrow gauge,
pronounced heteropody and outward rotated manus tracks
located on the prolongation of the pes’ long axis and
clearly farther away from the trackway midline than the
pes, expressed by the lowest [WAP/WAM] ratio of all
trackways (Figure 7(B)). Nonetheless, CTD-SCR-1000-
S10 has much smaller pes tracks with a more oval overall
track outline than Breviparopus (Deio-D), which has large
(.1.0m length) pes tracks with a bell-shaped, slightly
triangular outline. This particular pes track morphology,
on the other hand, is also characteristic for CTD-PMM-
1505-S1, with pes tracks equally longer than 1.0m.
However, the latter trackway has a wide gauge, a less
pronounced heteropody and manus tracks located
much closer to the trackway midline than in Breviparopus
(Deio-D). Breviparopus (Deio-D) differs from Parabron-
topodus and Brontopodus because of the bell-shaped pes
track outline, the absence of claw marks on the pes tracks,
a higher pes and manus outward rotation and manus tracks
located on the prolongation of the pes’ long axis and
farther away from the trackway midline. It further differs
from Brontopodus because of its narrower gauge, a
semicircular manus shape and a higher heteropody.
Regarding Parabrontopodus
Marty et al. (2003) assigned the narrow-gauge trackway
CTD-SCR-1000-S10 to the ichnotaxon Parabrontopodus,
mainly based on the typical trackway configuration
characterised by a pronounced narrow gauge and
heteropody. However, heteropody is less pronounced in
CTD-SCR-1000-S10, and the manus tracks are located
farther away from the trackway midline than in
Parabrontopodus. Belvedere et al. (2007) suggested that
the medium-gauge trackway Deio Lav-A from Morocco is
more similar to Parabrotopodus than Breviparopus,
because manus tracks are semicircular, pes and manus
tracks have a lower outward rotation and manus tracks are
located closer to the trackway midline than in Brevipar-
opus. The third narrow-gauge trackway CTD-TCH-1055-
S4 analysed here is also similar to Parabrontopodus,
notably because it has evident impressions of the first three
digits and the first two claws, the latter being strongly
rotated outwards as in Parabrontopodus. On the other
hand, heteropody is decidedly smaller in CTD-TCH-1055-
S4 and it further has an exceptional high manus outward
rotation and a particular manus position well in front of the
pes tracks and never deformed by the subsequent pes,
when compared with the holotype trackway of Para-
brontopodus. Parabrontopodus differs from Breviparopus
(Deio-D) because of the oval-shaped pes tracks with claw
impressions and manus tracks with a smaller outward
rotation and being located closer to the trackway midline.
It differs from Brontopodus because of its narrower gauge,
a semicircular manus shape and a higher heteropody.
Regarding Brontopodus
Marty (2008) tentatively assigned the very wide-gauge
trackway CHE-CRO-500-S10 to the ichnotaxon Bronto-
podus mainly because of its very wide gauge. This is
supported by manus tracks located almost as close to the
trackway midline as pes tracks. Nonetheless, the
assignation is tentative at best because CHE-CRO-500-
S10 does not exhibit digit and claw impressions due to the
poor preservation of the pes tracks, and Brontopodus is less
wide gauge and has manus tracks located closer to the
trackway midline than pes tracks. CTD-PMM-1505-S1 is
similar to Brontopodus because of a wide gauge, small
heteropody, semicircular to subcircular manus track
outline and manus tracks located almost as close to
trackway midline as pes tracks. This trackway compares
favourably with Brontopodus, but it cannot be assigned
with certainty because of the absence of digit and claw
impressions. Finally, the trackways CTD-SCR-1000-S10,
Deio-D and Deio Lav-A are different from Brontopodus
because of their narrow gauge and manus tracks located
farther away from trackway midline than pes tracks. CTD-
TCH-1055-S4 differs because of the absence of small digit
and clawmarks at digits IVand V, even though Farlow et al.
Historical Biology 127
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

(1989) noticed that in Brontopodus, these are only seen in
well-preserved footprints. Brontopodus differs from
Breviparopus (Deio-D) because of pes tracks with claw
impressions and manus tracks with a smaller outward
rotation. It differs from Breviparopus and Parabrontopo-
dus because of its wider gauge, U-shaped manus tracks, a
smaller heteropody and manus tracks located somewhat
closer to the trackway midline than pes tracks.
Summary
Wright (2005) recommended that sauropod trackways
should be classified primarily on pes and manus
morphology and only secondarily on trackway configur-
ation (trackway gauge). However, with the exception of
CTD-TCH-1055-S4, all studied trackways lack anatom-
ical details and for this reason an ichnotaxonomical
assignation based on footprint morphology is not
possible for these trackways. Based on trackway
configuration (notably gauge and for some of the
trackways heteropody and the position of pes and manus
tracks with respect to trackway midline), the studied
trackways could be assigned equally between the
narrow-gauge ichnotaxa Breviparopus (Deio-D, CTD-
SCR-1000-S10) and Parabrontopodus (CTD-TCH-1055-
S4, Deio Lav-A) and the wide-gauge ichnotaxon
Brontopodus (CHE-CRO-500-S10, CTD-PMM-1505-
S1). In trackway CTD-TCH-1055-S4, the number and
outward rotation of digits and claws is furthermore
similar to Parabrontopodus, even though trackway
configuration, heteropody and preservation (track
depth) are clearly different from the type specimen of
Parabrontopodus (CU-MWC 190.5; Lockley et al. 1986,
figure 7B; 1994b, figures 3 and 4). However, some
trackways from the Parabrontopodus-type locality (e.g.
CU MWC 190.3; Lockley et al. 1986, figures 7A and
9A) also show considerable changes in trackway
configuration (e.g. manus well in front of pes tracks,
high manus outward rotation) with regard to the
holotype. This can also be observed within the Swiss
narrow-gauge trackways CTD-SCR-1000-S10, CTD-
TCH-1055-S4 and CTD-SCR-1000-S4 [described by
Marty et al. (2003)] of the intermediate track-bearing
levels, which exhibit very different trackway configur-
ations, notably regarding the outward rotation and
position of pes and manus tracks. This evidence suggests
that this variability in trackway configuration may be
related to the behaviour of the trackmaker rather than to
different ichnotaxa.
Identification of possible trackmakers
A precise trackmaker identification is outside the scope of
this paper, because this is generally not possible unless the
animal is preserved at the end of its fossil trackway. The
purpose here is to provide a gross identification based on
track morphology, trackway configuration and on a short
review of skeletal material found in relative close
proximity to the tracksite localities, even though it has to
be considered that skeletal remains of the trackmakers may
not as yet have been found.
Based on track morphology
The typical pes and manus overall track morphology and
trackway configuration of all studied trackways readily
identify them as trackways left by sauropods. In the Late
Jurassic, the vast majority of quadrupedal dinosaur
trackways are attributed to sauropods, even though they
could have also been left by ornithischian dinosaurs such
as basal ceratopsians, ankylosaurs or most likely
stegosaurs as indicated by the skeletal record (Currie and
Padian 1997; Meyer and Hunt 1998; Weishampel et al.
2004). Thus, poorly preserved ornithischian trackways
could possibly be misinterpreted as the trackways of
smaller sauropods. Ankylosaur tracks are known from the
Cretaceous only (review in McCrea et al. 2001), even
though an ankylosaur trackway from Dorset (England)
may be of Late Jurassic age (Wright 1996; Ensom 2002).
Late Jurassic stegosaur tracks assigned to the ichnotaxon
Deltapodus are described from the Middle Jurassic of
England (Whyte and Romano 2001; Romano and Whyte
2003; Whyte et al. 2007), the Morrison Formation
(Lockley and Hunt 1998; Milan and Chiappe 2009),
from Spain (Cobos et al. 2008; Lockley et al. 2008), from
Portugal (Mateus and Milan, Forthcoming 2009), from
Poland (Gierlinski and Sabath 2002) and also from the
Iouaridene Formation, Morocco (Belvedere and Mietto
2010), and they all clearly differ from the herein described
sauropod tracks because of a particular pes morphology
characterised by a triangular outline with three short,
blunt, rounded and forward facing digit impressions and a
broad massive heel area.
When compared with predicted pes track mor-
phologies of Wright (2005) – which she based on pes
synapomorphies inferred from phylogenetic studies
(Upchurch 1998; Wilson and Sereno 1998) – the three
digits and laterally directed two claws in the pes tracks of
CTD-TCH-1055-S4 suggest a diplodocid sauropod as
possible trackmaker. On the other hand, CTD-TCH-1055-
S4 does not show any evidence for a claw impression on
digit I of the manus, which is in contradiction with the
predicted manus track morphology for diplodocid
sauropods of Wright (2005). However, this could also be
related to a functional reason (eventually related to
substrate properties) – even though the pollex claw may
have possessed a limited range of movement (e.g.
Upchurch 1994) – and/or because the rear part of the
manus tracks of CTD-TCH-1055-S4 was not impressed
due to a pronounced digitigrade posture of the manus.
D. Marty et al.128
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Based on trackway gauge
Lockley (1999) and Wilson and Carrano (1999) suggested
that more derived sauropods such as the titanosaurs were
the trackmakers of wide-gauge trackways while narrow-
gauge trackways were attributed to more ‘basal’ sauropods,
basal macronarians or diplodocoidea sensu Wilson and
Sereno (1998). Accordingly, and based on trackway gauge,
the studied trackways could, apart from the medium-gauge
trackwayDeioLav-A, easily be attributed tomore basal and
more derived sauropods, respectively. This would indicate
the presence of both groups of sauropods in very close
temporal relationship (and even on single paleosurfaces) on
the Jura carbonate platform, and of more ‘basal’ sauropods
only in the northeastern part of the Sahara Craton during the
Late Jurassic.
On the other hand, wide-gauge trackways could have
occurred more than once in sauropod evolution as the
result of the anteriorisation of the centre of mass
(Henderson 2006) and the overall morphology (Lockley
et al. 2002a), and trackway gauge could also be
influenced by the degree of lateral motion of the
trackmaker (Carpenter 2009). Lockley et al. (2002a),
based on trackway evidence from the Late Cretaceous of
Bolivia, suggested that titanosaurids may have shown
changes from narrow gauge to wide gauge during their
ontogenetic growth. Farlow (pers. comm. 2004 in Wright
2005) noticed that some Brontopodus trackways of the
Paluxy River have a narrow gauge along some trackway
segments, and a change from wide to narrow gauge
along a single trackway was also described by Leonardi
and Avanzini (1994) from the Early Jurassic of northern
Italy. Based on wide-gauge trackways (ichnotaxon
Polyonyx gomesi) from Portugal, which are characterised
by a small heteropody, pes tracks with four claw marks
(claws I–II anteriorly oriented), and notably manus
tracks with large digit I mark and a large triangular claw
mark I posteriorly oriented, dos Santos et al. (2009)
recently argued that at least one basal eusauropod was
able to produce wide-gauge trackways by the Middle
Jurassic. For this reason, they concluded that wide-gauge
trackways are not exclusive to titanosauriform
sauropods.
Based on the skeletal record
For the Late Jurassic, the presence of both groups of
sauropods on the Jura carbonate platform is also supported
by the skeletal record. Cetiosauriscus greppini from the
Kimmeridgian of Moutier, NW Switzerland (von Huene
1922, 1927; Meyer and Thuring 2003; Schwarz et al.
2007), currently considered a nomen dubium (Glut 1997),
is provisionally regarded as Diplodocoidea incertae sedis
(Upchurch et al. 2004), but its phylogeny is currently
under re-evaluation by one of the authors (Meyer).
Bothriospondylus madagascariensis from the Oxfordian
of Damparis, France (Buffetaut 1988), was re-examined
by Moine (1999) and assigned to Brachiosaurus byWilson
(2002, 2005).
From the northeastern part of the Sahara Craton, basal,
vulcanodontid sauropods (Tazoudasaurus naimi) are
known from the Early Jurassic (Allain et al. 2004) and
diplodocid sauropods (Rebbachisaurus garasbae) from the
Early Cretaceous (e.g. de Lapparent 1960 and Monbaron
1978). The presence of more derived sauropods is also
suggested by the Late Jurassic skeletal record of Africa,
notably the well-known Brachiosauridae remains from the
Late Jurassic of Tendaguru, Tanzania (e.g. Russell et al.
1980; Bonaparte et al. 2000 and Schwarz and Fritsch
2006) such as Brachiosaurus brancai recently identified as
Giraffatitan brancai (Taylor 2009). Furthermore, a
brachiosaurid-like sauropod (Atlasaurus imelakei) is
described from the central High Atlas Mountains
(Monbaron et al. 1999), and the bone-bearing layer –
initially thought to be Middle Jurassic in age (Monbaron
et al. 1999) – may actually be of Late Jurassic age (cf.
Charriere et al. 2005).
Concluding remarks and outlook
Sauropod trackways from the Late Jurassic of NW
Switzerland and the central High Atlas Mountains in
Morocco are similar from a preservational point of view
and with respect to the overall pes and manus track outline,
but they show a wide range of trackway configuration and
trackway gauge.
Based on trackway gauge solely, the studied trackways
can either be attributed to one of the narrow-gauge
ichnotaxa Breviparopus or Parabrontopodus, or to the
wide-gauge ichnotaxon Brontopodus. However, taking into
account the absence of digit and/or claw impressions in all
but one of the trackways, and the variable differences in
trackway configuration, an unambiguous ichnotaxonomi-
cal assignation of the trackways studied is not possible. The
Moroccan trackway Deio-D is the reference trackway of
Breviparopus, which we consider a valid ichnotaxon, but
which is in need of a formal and emended re-description
based on better preserved tracks with evident digit and/or
claw impressions. The latter condition is crucial for a clear
distinction between Breviparopus and Parabrontopodus.
The reference trackway (Deio-D) of Breviparopus is
characterised by more bell-shaped to triangular pes tracks
and by manus tracks located clearly farther away from the
trackway midline than in Parabrontopodus. The ichno-
taxon Parabrontopodus is also considered valid (and not a
junior synonym of Breviparopus) and it is characterised by
an oval pes track outline, pes claw impressions on digits I–
III and manus tracks located closer to the trackway midline
than in Breviparopus.
For the trackways and three ichnotaxa analysed, the
narrower trackways tend to have manus tracks located
Historical Biology 129
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

farther away from the trackway midline than the wider
ones. This evidence may indicate that the association of
narrow-gauge trackways with manus tracks located farther
away from the trackway midline and wide-gauge track-
ways with manus tracks located closer to the trackway
midline could be an expression of the general posture of
the trackmakers, and this could eventually be useful in the
classification of sauropod trackways.
No clear relationships between trackway gauge
(expressed by the [WAP/PL] ratio) and trackmaker size
(expressed by pes length), locomotion speed or substrate
properties were found for the trackways studied, but gauge
may be influenced by other factors than those analysed,
such as a particular individual behaviour (different degrees
of lateral bending) or sexual dimorphism.
The degree of pes and manus outward rotation, and of
manus overprinting and position in front of the pes is
interpreted as being related to the general locomotion
capabilities of the trackmakers (individual walking style,
locomotion speed, etc.), whereas heteropody is difficult to
calculate due to incomplete manus tracks.
The lack of digit and claw impressions in most of the
trackways studied makes it difficult to more closely
identify the sauropod trackmakers. At least, the three digits
and laterally directed two claws in the pes tracks of the
trackway CTD-TCH-1055-S4 suggest a diplodocid
sauropod as a possible trackmaker for this trackway.
In order to better understand the variability of sauropod
trackway configuration in general and within trackways left
by a specific group (e.g. diplodocids, brachiosaurids and
titanosauriforms) of sauropods, many more trackways need
to be analysed and compared in a similar way. This could be
helped by an online trackway database including pictures,
and if available three-dimensional pictures based on laser
scanning and/or photogrammetry. Then, trackway data
could also be analysed in a statistically more sophisticated
way than done in this work. This may help to better
understand the general locomotion capabilities of sauropods,
and to determine which parameters of trackway configur-
ation are to what degree useful in their classification because
they are an expression of the general posture of the
trackmakers rather than they are influenced by extrinsic
factors. However, even if trackway configuration may – to a
certain degree – be useful in the classification of sauropod
trackways, we agree that ichnotaxa should only be erected
based on well-preserved true tracks with impressions of
substantial anatomical details of the feet such as digit and/or
claw impressions.
Acknowledgements
A part of this research was carried out during the PhD theses ofDM and MB and therefore the Swiss National ScienceFoundation (grant no. 20-109214.05) and the Universita degliStudi di Padova, respectively, are acknowledged. We thank allthe authorities of the Demnat region, from the community chief
to the governor of the Azilal Province, for permissions for thefieldwork. We also warmly thank the AESVT of Demnat,especially M. Chaouki and M. Essoufi, and to SimohamedBenhimou from Taghbalout, for support with the logistics andcontact with the local authorities and people. N.-E. Jalil(University Cadi Ayyad, Marrakech) is thanked for theorganisation of the field trips, the relationships with theM’Goun Geopark direction and the invitation to contribute tothis special volume. S. Ishigaki (Hayashibara Museum of NaturalSciences, Okayama) is thanked for kindly sharing his hugeamount of unpublished sketches of the Iouaridene tracksites.G. Dyke and N.-E. Jalil are thanked for editorial work and foraccepting us to contribute to this special volume on north Africanvertebrate paleontology.
References
Alexander R McN. 1976. Estimates of speeds of dinosaurs. Nature.261:129–130.
Alexander R McN. 2006. Dinosaur biomechanics. Proc R Soc Lond, SerB. 273:1849–1855.
Allain R, Aquesbi N, Dejax J, Meyer CA, Monbaron M, Montenat C,Richir P, Rochdy M, Russell D, Taquet P. 2004. A basal sauropoddinosaur from the Early Jurassic of Morocco. C R Palevol.3:199–208.
Avanzini M, Leonardi G, Mietto P. 2003. Lavinipes cheminii ichnogen.,ichnosp. nov., a possible sauropodomorph track from the LowerJurassic of the Italian Alps. Ichnos. 10:179–193.
Belvedere M. 2008. Ichnological researches on the Upper Jurassicdinosaur tracks in the Iouaridene area (Demnat, central High-Atlas,Morocco) [dissertation]. [Padova (Italy)]: Universita degli Studi diPadova.
Belvedere M, Mietto P. 2010. First evidence of stegosaurian Deltapodusfootprints in North Africa (Iouaridene Formation, Upper Jurassic,Morocco). Palaeontology. 53:233–240.
Belvedere M, Mietto P, Mehdi M. 2007. Dinosaur tracks from the UpperJurassic Iouaridene Formation (Demnat, Morocco). Geoitalia 2007,Rimini, Italy. Epitome. 2:306.
Bertling M, Insalaco E. 1998. Late Jurassic coral/microbial reefs from thenorthern Paris Basin – Facies, palaeoecology and palaeobiogeo-graphy. Palaeogeogr Palaeoclimatol Palaeoecol. 139:139–175.
Billon-Bruyat J-P, Mazin J-M. 2003. The systematic problem of tetrapodichnotaxa: the case study of Pteraichnus Stokes, 1957 (Pterosauria,Pterodactyloidea). In: Buffetaut E, Mazin J-M, editors. Evolutionand palaeobiology of pterosaurs. London: Geol Soc of London SpecPubl. 217: 315–324.
Bonaparte JF, Heinrich WD, Wild R. 2000. Review of Janenschia Wildwith the description of a new sauropod from the Tendaguru beds ofTanzania and a discussion on the systematic value of procoelouscaudal vertebrae in the Sauropoda. Palaeontographica. 256:25–76.
Buffetaut E. 1988. Les restes de dinosauriens de l’Oxfordien superieur deDamparis (Jura): preuves d’emersion sur place. Rev Paleobiol.7:301–306.
Carpenter K. 2009. Role of lateral body bending in crocodylian trackmaking. Ichnos. 16:202–207.
Chafetz HS, Buczynski C. 1992. Bacterially induced lithification ofmicrobial mats. Palaios. 7:277–293.
Charriere A, Haddoumi H, Mojon P-O. 2005. Decouverte de Jurassiquesuperieur et d’un niveau marin du Barremien dans les «couchesrouges» continentales du Haut Atlas central marocain: implicationspaleogeographiques et structurales. C R Palevol. 4:385–394.
Cobos A, Royo-Torres R, Alcala L, Luque L, Aberasturi A. 2008. Nuevosdatos de las icnitas de dinosaurios en la Formacion Villard elArzobispo (Teurel). In: Ruiz-Omenaca JI, Pinuela L, Garcıa-RamosJC, editors. Colunga (Spain): Libro de resumenes. XXIV Jornadas dela Sociedad Espanola de Paleontologıa, Museo del Jurasico deAsturias (MUJA). p. 25–26.
Coombs WP Jr. 1978. Theoretical aspects of cursorial adaptions indinosaurs. Q Rev Bio. 53:393–418.
D. Marty et al.130
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Currie PJ, Padian K. 1997. Encyclopedia of dinosaurs. San Diego (CA):Academic Press.
Dalla Vecchia FM. 2005. Between Gondwana and Laurasia: Cretaceoussauropods in an intraoceanic carbonate platform. In: Tidwell V,Carpenter K, editors. Thunder-lizards. Bloomington (IN): IndianaUniversity Press. p. 395–429.
Dalla Vecchia FM, Tarlao A. 2000. New dinosaur track sites in the Albian(Early Cretaceous) of the Istrian peninsula (Croatia). Part II –Paleontology. In: Dalla Vecchia FM, Tarlao G, Tunis G, Venturini S,editors. New dinosaur track sites in the Albian (Early Cretaceous) ofthe Istrian peninsula (Croatia). Padova (Italy): Memorie di ScienzeGeologiche di Padova 52. p. 227–292.
Day JJ, Norman DB, Gale AS, Upchurch P, Powell HP. 2004. A MiddleJurassic dinosaur trackway site from Oxfordshire, UK. Palaeonto-logy. 47:319–348.
Day JJ, Norman DB, Upchurch P, Gale AS, Powell HP. 2002. Sauropodtrackways, evolution, and behavior. Science. 296:1659.
de Lapparent AF. 1960. Les dinosauriens du “Continental intercalaire” duSahara central. Mem Soc Geol France. p. 88A.
dos Santos VF, Moratalla JJ, Royo-Torres R. 2009. New sauropodtrackways from the Middle Jurassic of Portugal. Acta Palaeontol Pol.54:409–422.
Dupraz C, Visscher PT. 2005. Microbial lithification in marinestromatolites and hypersaline mats. Trends Microbiol. 13:429–438.
Dutuit JM, Ouazzou A. 1980. Decouverte d’une piste de dinosauresauropode sur le site d’empreintes de Demnat (Haut-Atlasmarocain). Mem Soc Geol France N S. 139:95–102.
Ensom PC. 2002. Vertebrate trace fossils in the Purbeck Limestone groupof Southern England. In: Milner AR, Batten DJ, editors. Life andenvironment in Purbeck times. Oxford (UK): The PalaeontologicalAssociation. Special Papers in Palaeontology 68. p. 203–220.
Farlow JO. 1987. A guide to Lower Cretaceous dinosaur footprints andtracksites of the Paluxy River Valley, Sommervell County, Texas.Geological Society of America, South-Central Section, 21st AnnualMeeting, Field Trip Guide-Book, Waco, Texas.
Farlow JO. 1992. Sauropod tracks and trackmakers: integrating theichnological and skeletal record. Zubia. 10:89–138.
Farlow JO, Pittman JG, Hawthorne JM. 1989. Brontopodus birdi, LowerCretaceous sauropod footprints from the US Gulf Coastal plain. In:Gillette DD, Lockley GM, editors. Dinosaur tracks and traces.Cambridge: Cambridge University Press. p. 371–394.
Frakes LA, Francis JE, Sykes RM. 1992. Climate modes of thePhanerozoic. Cambridge: Cambridge University Press.
Gallup MR. 1989. Functional morphology of the hindfoot of the Texassauropod Pleurocoelus sp. indet. In: Farlow JO, editor. Paleobiologyof the dinosaurs. Geol Soc Am Spec Pap. 238:71–74.
Gierlinski G, Sabath K. 2002. A probable stegosaurian track from theLate Jurassic of Poland. Acta Palaeontol Pol. 47:561–564.
Glut DF. 1997. Dinosaurs: the encyclopedia. Jefferson (NC): McFarland& Company Inc.
Gonzalez Riga BJ, Calvo JO. 2009. A new wide-gauge sauropod track sitefrom the Late Cretaceous of Mendoza, Neuquen Basin, Argentina.Palaeontology. 52:631–640.
Gygi RA. 2000. Annotated index of lithostratigraphic units currently usedin the Upper Jurassic of Northern Switzerland. Eclogae geol Helv.93:125–146.
Haddoumi H, Charriere A, Andreu B, Mojon P-O. 2009. Les depotscontinentaux du Jurassique moyen au Cretace inferieur dans le HautAtlas oriental (Maroc): paleoenvironnements successifs et significa-tion paleogeographique. Carnets de Geologie. CG2008-A06:1–29.
Hallam A. 1984. Continental humid and arid zones during theJurassic and Cretaceous. Palaeogeogr Palaeoclimatol Palaeoecol.47:195–223.
Hallam A. 1985. A review of mesozoic climates. J Geol Soc (London).142:433–445.
Haubold H. 1971. Ichnia Amphibiorum et Reptiliorum fossilium. In:Kuhn O, editor. Encyclopedia of paleoherpetology. Handbuch derPalaoherpetologie. Part 18. Stuttgart (Germany): Fischer Verlag.
Henderson DM. 2006. Burly gaits: centers of mass, stability and thetrackways of sauropod dinosaurs. J Vertebr Paleontol. 26:907–921.
Hutchinson JR, Anderson FC, Blemker SS, Delp SL. 2005. Analysis ofhindlimb muscle moment arms in Tyrannosaurus rex using a three-
dimensional musculoskeletal computer model: implications forstance, gait, and speed. Paleobiology. 31:676–701.
Ishigaki S. 1985a. Dinosaur footprints from Atlas Mountains. Nat Study.31(10):5–8 [In Japanese].
Ishigaki S. 1985b. Dinosaur footprints from Atlas Mountains. Nat Study.31(12):5–7 [In Japanese].
Ishigaki S. 1985c. Compte rendu d’activite 1984. Unpublished ReportRabat (Morocco): Ministere de l’Energie et des Mines.
Ishigaki S. 1986. Dinosaur footprints from Atlas Mountains. Nat Study.32(1):6–9 [In Japanese].
Ishigaki S. 1989. Footprints of swimming Sauropods from Morocco. In:Gillette DD, Lockley MG, editors. Dinosaur tracks and traces.Cambridge: Cambridge University Press. p. 83–86.
Jank M, Meyer CA, Wetzel A. 2006a. Late Oxfordian to LateKimmeridgian carbonate deposits of NW Switzerland (Swiss Jura):Stratigraphical and palaeogeographical implications in the transitionarea between the Paris Basin and the Tethys. Sediment Geol.138:237–263.
Jank M, Wetzel A, Meyer CA. 2006b. A calibrated composite section forthe Late Jurassic Reuchenette Formation in northwestern Switzer-land (?Oxfordian, Kimmeridgian sensu gallico, Ajoie-Region).Eclogae geol Helv. 99:175–191.
Jenny J. 1985. Carte geologique du Maroc a 1:100’000, feuille Azilal.Notes Mem Serv geol Maroc 399 p.
Jenny J, Jossen J-A. 1982. Decouverte d’empreintes de pas dedinosauriens dans le Jurassique inferieur (Pliensbachien) du Haut-Atlas central Maroc. C R Acad Sci (Paris). 294:223–226.
Jenny J, Le Marrec A, Monbaron M. 1981. Les couches rouges duJurassique moyen du Haut Atlas central (Maroc): correlationslithostratigraphiques, elements de datations et cadre tectono-sedimentaire. Bull Soc geol Fr (7). XXIII(6):627–639.
Leonardi G. 1987. Glossary and manual of tetrapod footprintpalaeoichnology. Brasılia (Brasil): Publicacao do DepartementoNacional da Producao Mineral Brasil.
Leonardi G, Avanzini M. 1994. Dinosauri in Italia. Le Scienze, Quaderni.76:69–81.
Lockley MG. 1991. Tracking dinosaurs: a new look at an ancient world.Cambridge: Cambridge University Press.
Lockley MG. 1999. The eternal trail: a tracker looks at evolution.Cambridge (MA): Perseus Books.
Lockley MG, Farlow JO, Meyer CA. 1994b. Brontopodus andParabrontopodus ichnogen. nov. and the significance of wide- andnarrow-gauge sauropod trackways. Gaia. 10:135–146.
Lockley MG, Garcia-Ramos JC, Pinuela L, Avanzini M. 2008. A reviewof vertebrate track assemblages from the Late Jurassic of Asturias,Spain with comparative notes on coeval ichnofaunas from thewestern USA: implications for faunal diversity in siliciclastic faciesassemblages. Oryctos. 8:53–70.
Lockley MG, Houck KJ, Prince NK. 1986. North America’s largesttrackway site: implications for Morrison Formation paleoecology.Bull Geol Soc Am. 97:1163–1176.
Lockley MG, Hunt AP. 1995. Dinosaur tracks and other fossil footprintsof the western United States. New York (NY): Columbia UniversityPress.
Lockley MG, Hunt AP. 1998. A probable stegosaur track from theMorrison Formation of Utah. In: Carpenter K, Chure D, Kirkland J,editors. The Upper Jurassic Morrison Formation: an interdisciplinarystudy. Modern Geology. 23:331–342.
Lockley MG, Hunt AP, Meyer CA. 1994c. Vertebrate tracks and theichnofacies: implications for paleoecology and palichnostratigraphy.In: Donovan SK, editor. The palaeobiology of trace fossils.Chichester: Wiley. p. 241–268.
Lockley MG, Meyer CA. 2000. Dinosaur tracks and other fossil footprintsof Europe. New York (NY): Columbia University Press.
Lockley MG, Santos VF dos, Meyer CA, Hunt A, editors. 1994a. Aspectsof sauropod paleobiology. Gaia 10.
Lockley MG, Schulp AS, Meyer CA, Leonardi G, Kerumba Mamami D.2002a. Titanosaurid trackways from the Upper Cretaceous ofBolivia: evidence for large manus, wide-gauge locomotion andgregarious behaviour. Cretaceous Res. 23:383–400.
Lockley MG, Wright JL, White D, Jianjun L, Lu F, Hong L, MatsukawaM. 2002b. The first sauropod trackways from China. Cretaceous Res.23:363–381.
Historical Biology 131
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Mannion PD. 2008. Environmental associations of sauropod dinosaursand their bearing on the early Late Cretaceous ‘sauropod hiatus’.J Vertebr Paleontol. 28 (Suppl. 3):111.
Marty D. 2008. Sedimentology, taphonomy, and ichnology of LateJurassic dinosaur tracks from the Jura carbonate platform(Chevenez–Combe Ronde tracksite, NW Switzerland): insightsinto the tidal-flat palaeoenvironment and dinosaur diversity,locomotion, and palaeoecology [dissertation]. [Fribourg (Switzer-land)]: University of Fribourg. GeoFocus 21.
Marty D, Ayer J, Becker D, Berger J-P, Billon-Bruyat J-P, Braillard L,Hug WA, Meyer CA. 2007. Late Jurassic dinosaur tracksites of theTransjurane highway (Canton Jura, NW Switzerland): overview andmeasures for their protection and valorization. Bull Appl Geol.12:75–89.
Marty D, Cavin L, Hug WA, Jordan P, Lockley MG, Meyer CA. 2004.The protection, conservation and sustainable use of the Courtedouxdinosaur tracksite, Canton Jura, Switzerland. Rev Paleobiol. VolSpec 9:39–49.
Marty D, Cavin L, Hug WA, Meyer CA, Lockley MG, Iberg A. 2003.Preliminary report on the Courtedoux dinosaur tracksite from theKimmeridgian of Switzerland. Ichnos. 10:209–219.
Marty D, Meyer CA, Billon-Bruyat J-P. 2006. Sauropod trackwaypatterns expression of special behaviour related to substrateconsistency? An example from the Late Jurassic of northwesternSwitzerland. Hantkeniana. 5:38–41.
Marty D, Pacton M. 2009. Formation and preservation of Late Jurassicdinosaur track-bearing tidal-flat laminites (Canton Jura, NWSwitzerland) through microbial mats. In: Billon-Bruyat J-P, MartyD, Costeur L, Meyer CA, Thuring B, editors. 5th InternationalSymposium on Lithographic Limestone and Plattenkalk, Abstractsand Field Guides. Societe jurassienne d’emulation, actes 2009 bis.p. 56–58.
Marty D, Strasser A, Meyer CA. 2009. Formation and taphonomy ofhuman footprints in microbial mats of present-day tidal-flatenvironments: implications for the study of fossil footprints. Ichnos.16:127–142.
Mateus O, Milan J. Forthcoming 2009. A diverse Upper Jurassic dinosaurichnofauna from central-west Portugal. Lethaia. 13 pp.
McCrea RT, Lockley MG, Meyer CA. 2001. Global distribution ofpurported ankylosaur track occurrence. In: Carpenter K, editor. Thearmored dinosaurs. Bloomington (IN): Indiana University Press.p. 413–454.
Meyer CA, Hunt AP. 1998. The first stegosaurian dinosaur (Ornithischia:Thyreophora) from the Late Jurassic of Switzerland. N Jb GeolPalaont. 1998(3):141–145.
Meyer CA, Lockley MG, Robinson JW, dos Santos VF. 1994.A comparison of well-preserved sauropod tracks from the LateJurassic of Portugal and the Western United States: evidence andimplications. Gaia. 10:57–64.
Meyer CA, Monbaron M. 2002. Middle Jurassic dinosaur tracks fromMorocco – Facts and fiction. Paper presented at 7th EuropeanWorkshop on Vertebrate Palaeontology; 2002 Jul 2–7; Sibiu,Romania.
Meyer CA, Thuring B. 2003. Dinosaurs of Switzerland. C R Palevol.2:103–117.
Milan J, Chiappe LM. 2009. First American record of the Jurassicichnospecies Deltapodus brodricki and a review of the fossil recordof Stegosaurian footprints. J Geol. 117:343–348.
Moine O. 1999. Datation, condition de depot et position phylogenetiquede ‘Bothriospondylus madagascariensis’ (Damparis, Jura, France).[Master’s Thesis]. [Lyon (France)]: Ecole Normale Superieure 8:77–106.
Monbaron M. 1978. Nouveaux ossements de Dinosauriens de grandetaille dans le bassin jurassico-cretace de Taguelft (Atlas de Beni-Mellal, Maroc). C R Acad Sci Paris serie D. 287:1277–1279.
Monbaron M, Russell DA, Taquet P. 1999. Atlasaurus imelaki n.g., n. sp.,a brachiosaurid-like sauropod from the Middle Jurassic of Morocco.C R Acad Sci France. 329:519–526.
Moratalla JJ, Garcia-Mondejar J, dos Santos VF, Lockley MG, Sanz JL,Jimenez S. 1994. Sauropod trackways from the Lower Cretaceous ofSpain. Gaia. 10:75–84.
Moreno K, Benton MJ. 2005. Occurrence of sauropod dinosaur tracks inthe Upper Jurassic of Chile (redescription of Iguanodonichnusfrenki). J South Am Earth Sci. 20:253–257.
Nouri J. 2007. La paleoichnologie des empreintes de pas dinosauriens,imprimees dans les couches du Jurassique du Haut-Atlas Central duMaroc [dissertation]. [Rabat (Morocco)]: Universite Mohammed V,Faculte des Sciences, AGDAL.
Peabody FE. 1959. Trackways of living and fossil salamanders. UnivCalif Publ Zool. 63:1–72.
Pittman JG. 1989. Stratigraphy, lithology, depositional environment, andtrack type of dinosaur track-bearing beds of the Gulf Coastal Plain.In: Gillette DD, Lockley MG, editors. Dinosaur tracks and traces.Cambridge: Cambridge University Press. p. 135–153.
Pittman JG. 1990. Dinosaur tracks and trackbeds in the middle part of theGlen Rose Formation, Western Gulf Basin, USA. In: Bergman GR,Pittman JG, editors. Nearshore clastic-carbonate facies and dinosaurtrackways in the Glen Rose Formation (Lower Cretaceous) ofCentral Texas. Geological Society of America, 1990 AnnualMeeting, Field Trip Guidebook 8:47–83.
Pittman JG. 1992. Stratigraphy and vertebrate ichnology of the Glen RoseFormation, Western Gulf Basin, USA [dissertation]. [Austin (TX)]:University of Texas at Austin.
Pittman JG, Gillette DD. 1989. A new sauropod dinosaur tracksite inLower Cretaceous beds of Arkansas, USA. In: Gillette DD, LockleyMG, editors. Dinosaur tracks and traces. Cambridge: CambridgeUniversity Press. p. 313–332.
Plateau H, German G, Roch E. 1937. Sur la presence d’empreintes deDinosauriens dans la region de Demnat (Maroc). C R Somm Soc geolFrance. 16:241–242.
Prince NK, Lockley MG. 1989. The sedimentology of the Purgatoiretracksite region, Morrison Formation of southeastern Colorado. In:Gillette DD, Lockley GM, editors. Dinosaur tracks and traces.Cambridge: Cambridge University Press. p. 155–164.
Rainforth E, Manzella M. 2007. Estimating speeds of dinosaurs fromtrackways: a re-evaluation of assumptions. In: Rainforth E, editor.Contributions to the paleontology of New Jersey (II) – Field guideand proceedings. 2007 Oct 12–13; Geological Association of NewJersey XXIV Annual Conference and field trip, East StroudsburgUniversity; Pennsylvania, USA. p. 41–48.
Romano M, Whyte MA. 2003. Jurassic dinosaur tracks and trackways ofthe Cleveland Basin, Yorkshire: preservation, diversity anddistribution. P Yorks Geol Soc. 54:185–215.
Romano M, Whyte MA, Jackson SJ. 2007. Trackway ratio: a new look attrackway gauge in the analysis of quadrupedal dinosaur trackwaysand its implications for ichnotaxonomy. Ichnos. 14:257–270.
Ross CA, Moore GT, Hayashida DN. 1992. Late Jurassic paleoclimatesimulation–paleoecological implications for ammonoid provinci-ality. Palaios. 7:487–507.
Russell D, Beland P, McIntosh JS. 1980. Paleoecology of the dinosaurs ofTendaguru (Tanzania). Mem Soc Geol France NS. 139:169–175.
Salgado L, Coria RA, Calvo JO. 1997. Evolution of titanosauriddinosaurs, I: Phylogenetic analysis based on the postcranialevidence. Ameghiniana. 34:3–32.
Sarjeant WAS. 1989. ‘Ten paleoichnological commandments’: astandardized procedure for the description of fossil vertebratefootprints. In: Gillette DD, Lockley MG, editors. Dinosaur tracks andtraces. Cambridge: Cambridge University Press. p. 369–370.
Schwarz D, Fritsch G. 2006. Pneumatic structures in the cervicalvertebrae of the Late Jurassic Tendaguru sauropods Brachiosaurusbrancai and Dicraeosaurus. Eclogae geol Helv. 99:65–78.
Schwarz D, Wings O, Meyer CA. 2007. Super sizing the giants: firstcartilage preservation at a sauropod dinosaur limb joint. J Geol Soc(London). 164:61–65.
Stampfli GM, Borel GD. 2002. A plate tectonic model for the Palaeozoicand Mesozoic constrained by dynamic plate boundaries and restoredsynthetic oceanic isochrones. Earth Planet Sci Lett. 196:17–33.
Stampfli GM, Borel GD, Marchant R, Mosar J. 2002. Western Alpsgeological constraints on western Tethyan reconstructions. In:Rosenbaum G, Lister GS, editors. Reconstruction of the evolution ofthe Alpine–Himalayan Orogen. J Virtual Explorer 8:77–106.
Taylor MP. 2009. A re-evalutation of Brachiosaurus altithorax Riggs1903 (Dinosauria, Sauropoda) and its generic separation from
D. Marty et al.132
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010

Giraffatitan brancai (Janensch 1914). J Vertebr Paleontol.29:787–806.
Thalmann HK. 1966. Zur Stratigraphie des oberen Malm im sudlichenBerner und Solothurner Jura [dissertation]. [Bern (Switzerland)]:University of Bern.
Thierry J. 2000a. Early Kimmeridgian. In: Dercourt J, Gaetani M,Vrielvynck B, Barrier E, Biju-Duval B, Brunet MF, Cadet JP,Crasquin S, Sandulescu M, editors. Atlas Peri-Tethys Palaeogeo-graphical maps – Explanatory Notes. Paris: CCGM/CGMN(Commission for the Geological Map of the World). p. 85–97.
Thierry J. 2000b. Early Tithonian. In: Dercourt J, Gaetani M, VrielvynckB, Barrier E, Biju-Duval B, Brunet MF, Cadet JP, Crasquin S,Sandulescu M, editors. Atlas Peri-Tethys Palaeogeographical maps– Explanatory Notes. Paris. p. 99–110.
Thierry J, Abbate E, Alekseev AS, Ait-Ouali R, Ait-Salem H, Bouaziz S,Canerot J, Georgiev G, Guiraud R, Hirsch F, et al. 2000a. Map 10:Early Kimmeridgian (146–144 Ma). In: Dercourt J, Gaetani M,Vrielvynck B, Barrier E, Biju-Duval B, Brunet MF, Cadet JP,Crasquin S, Sandulescu M, editors. Paris (France): Atlas Peri-TethysPalaeogeographical maps.
Thierry J, Barrier E, Abbate E, Alekseev AS, Ait-Ouali R, Ait-Salem H,Bouaziz S, Canerot J, Georgiev G, Guiraud R, et al. 2000b. Map 11:Early Tithonian (141–139 Ma). In: Dercourt J, Gaetani M,Vrielvynck B, Barrier E, Biju-Duval B, Brunet MF, Cadet JP,Crasquin S, Sandulescu M, editors. Paris (France): Atlas Peri-TethysPalaeogeographical maps.
Thulborn T. 1990. Dinosaur tracks. London: Chapman & Hall.Upchurch P. 1994. Manus claw function in sauropod dinosaurs. Gaia.
10:161–172.Upchurch P. 1998. The phylogenetic relationships of sauropod dinosaurs.
Zool J Linn Soc. 124:43–103.Upchurch P, Barrett PM, Dodson P. 2004. Sauropoda. In: Weishampel
DB, Dodson P, Osmolska H, editors. The dinosauria. 2nd ed.Berkeley and Los Angeles (CA): University of California Press.p. 259–324.
von Huene F. 1922. Uber einen Sauropoden im oberen Malm des BernerJura. Eclogae geol Helv. 17:80–94.
von Huene F. 1927. Sichtung der Grundlagen der jetzigen Kenntnis derSauropoden. Eclogae geol Helv. 20:444–470.
Weishampel DB, Dodson P, Osmolska H. 2004. The dinosauria. 2nd ed.Berkeley and Los Angeles (CA): University of California Press.
Weissert H, Mohr H. 1996. Late Jurassic climate and its impact on carboncycling. Palaeogeogr Palaeoclimatol Palaeoecol. 122:27–43.
Whyte MA, Romano M. 2001. Probable stegosaurian dinosaur tracksfrom the Saltwick Formation (Middle Jurassic) of Yorkshire,England. P Geologist Assoc. 112:45–54.
Whyte MA, Romano M, Elvidge DJ. 2007. Reconstruction of MiddleJurassic dinosaur-dominated communities from the vertebrateichnofauna of the Cleveland Basin of Yorkshire, UK. Ichnos.14:117–129.
Wilson JA. 2002. Sauropod dinosaur phylogeny: critics and cladisticanalysis. Zool J Linn Soc. 136:217–276.
Wilson JA. 2005. Overview of sauropod phylogeny and evolution. In:Curry Rogers KA, Wilson JA, editors. The sauropods: evolution andpaleobiology. Berkeley and Los Angeles (CA): University ofCalifornia Press. p. 15–49.
Wilson JA, Carrano MT. 1999. Titanosaurs and the origin of ‘wide-gauge’ trackways: a biomechanical and systematic perspective onsauropod locomotion. Paleobiology. 25:252–267.
Wilson JA, Sereno PC. 1998. Early evolution and higher-level phylogenyof sauropod dinosaurs. J Vertebr Paleontol Mem. 5:198 p.
Wright JL. 1996. Fossil terrestrial trackways: function, taphonomy andpaleoecological significance [dissertation]. [Bristol]: University ofBristol.
Wright JL. 2005. Steps in understanding sauropod biology – Theimportance of sauropod tracks. In: Curry Rogers KA, Wilson JA,editors. The sauropods: evolution and paleobiology. Berkeley andLos Angeles (CA): University of California Press. p. 252–280.
Ziegler PA. 1988. Evolution of the Arctic – North Atlantic and theWestern Tethys. AAPG Memoir. 43:68 p.
Ziegler PA. 1990. Geological atlas of Western and Central Europe.2nd ed. London: Shell International Petroleum Mijnbouw andGeological Society.
Historical Biology 133
Downloaded By: [Belvedere, Matteo] At: 06:20 28 May 2010




















