
Clp ATPases and ClpP proteolytic complexes regulate vital biological processes in low GC,...
11
MicroReview Clp ATPases and ClpP proteolytic complexes regulate vital biological processes in low GC, Gram-positive bacteria Dorte Frees, 1 Kirsi Savijoki, 2 Pekka Varmanen 3 and Hanne Ingmer 1 * 1 Department of Veterinary Pathobiology, Faculty of Life Sciences, University of Copenhagen, Stigbøjlen 4, DK-1870 Frederiksberg C, Denmark. 2 Institute of Biotechnology and 3 Department of Basic Veterinary Sciences, University of Helsinki, Finland. Summary Clp proteolytic complexes consisting of a proteolytic core flanked by Clp ATPases are widely conserved in bacteria, and their biological roles have received considerable interest. In particular, mutants in the clp genes in the low-GC-content Gram-positive phyla Bacillales and Lactobacillales display a diverse range of phenotypic changes including general stress sensitivity, aberrant cell morphology, failure to initiate developmental programs, and for patho- gens, severely attenuated virulence. Extensive research dedicated to unravelling the molecular mechanisms underlying these complex phenotypes has led to fascinating new insights that will be covered by this review. First, Clp ATPases and ClpP- containing proteolytic complexes play indispensable roles in cellular protein quality control systems by refolding or degrading damaged proteins in both stressed and non-stressed cells. Secondly, ClpP proteases and the chaperone activity of Clp ATPases are important for controlling stability and activity of central transcriptional regulators, thereby exerting tremendous impact on cell physiology. Targets include major stress regulators like Spx (oxidative stress), the antisigma factor RsiW (alkaline stress) and HdiR (DNA damage) in addition to regulators of developmental programs like ComK (competence development), s H and Sda (sporulation). Thus, Clp proteins are central in co-ordinating developmental decisions and stress response in low GC Gram- positive bacteria. The bipartite ClpP proteolytic complexes and the Clp chaperones The highly conserved ATP-dependent ClpP proteases are two-component proteases consisting of separately encoded ATPase and peptidase subunits (Fig. 1A). Exten- sive structural analysis of the ClpP proteases from Escherichia coli has formed the paradigm for the way we understand the function of these proteins (reviewed by Sauer et al., 2004). The central proteolytic core consists of 14 ClpP serine peptidase subunits stacked in two hep- tameric rings, forming an internal chamber in which the active sites are sequestered from the cytoplasm (Wang et al., 1997). Narrow axial pores that exclude native pro- teins and all but the smallest peptides control access to this proteolytic chamber. To gain proteolytic activity, the ClpP multimer associates with one or two hexameric rings of Clp ATPases, forming the ClpP-containing proteolytic complex (designated the ClpP protease). In association with the proteolytic core, the Clp ATPases are responsible for the recognition, unfolding and translocation of sub- strates into the ClpP degradation chamber (Zolkiewski, 2006). This overall structure and organization bears resemblance to the 26S proteasome of eukaryotic cells (Kessel et al., 1995). The Clp ATPases constitute a family of closely related proteins that carry one or two nucleotide binding domains typical of the AAA+ superfamily of ATPases (Schirmer et al., 1996; Neuwald et al., 1999). Clp ATPases are divided into subfamilies on the basis of the presence of specific signature sequences and the number and spacing of the nucleotide binding sites (Fig. 1B). A role in protein folding, activation, or disaggregation seems to be shared by all Clp ATPases (Zolkiewski, 2006). However, only a subgroup of the Clp ATPases can interact with ClpP, a property that is associated with the presence of a ClpP recognition tripeptide found in ClpA, ClpC and ClpE Accepted 5 January, 2007. *For correspondence. E-mail hi@life. kudk; Tel. (+45) 3528 2773; Fax (+45) 3528 2755. Molecular Microbiology (2007) 63(5), 1285–1295 doi:10.1111/j.1365-2958.2007.05598.x © 2007 The Authors Journal compilation © 2007 Blackwell Publishing Ltd
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Clp ATPases and ClpP proteolytic complexes regulate vital biological processes in low GC,...

MicroReview
Clp ATPases and ClpP proteolytic complexes regulatevital biological processes in low GC, Gram-positivebacteria
Dorte Frees,1 Kirsi Savijoki,2 Pekka Varmanen3
and Hanne Ingmer1*1Department of Veterinary Pathobiology, Faculty of LifeSciences, University of Copenhagen, Stigbøjlen 4,DK-1870 Frederiksberg C, Denmark.2Institute of Biotechnology and 3Department of BasicVeterinary Sciences, University of Helsinki, Finland.
Summary
Clp proteolytic complexes consisting of a proteolyticcore flanked by Clp ATPases are widely conservedin bacteria, and their biological roles have receivedconsiderable interest. In particular, mutants in theclp genes in the low-GC-content Gram-positive phylaBacillales and Lactobacillales display a diverserange of phenotypic changes including generalstress sensitivity, aberrant cell morphology, failureto initiate developmental programs, and for patho-gens, severely attenuated virulence. Extensiveresearch dedicated to unravelling the molecularmechanisms underlying these complex phenotypeshas led to fascinating new insights that will becovered by this review. First, Clp ATPases and ClpP-containing proteolytic complexes play indispensableroles in cellular protein quality control systems byrefolding or degrading damaged proteins in bothstressed and non-stressed cells. Secondly, ClpPproteases and the chaperone activity of Clp ATPasesare important for controlling stability and activity ofcentral transcriptional regulators, thereby exertingtremendous impact on cell physiology. Targetsinclude major stress regulators like Spx (oxidativestress), the antisigma factor RsiW (alkaline stress)and HdiR (DNA damage) in addition to regulators ofdevelopmental programs like ComK (competencedevelopment), sH and Sda (sporulation). Thus, Clpproteins are central in co-ordinating developmental
decisions and stress response in low GC Gram-positive bacteria.
The bipartite ClpP proteolytic complexes andthe Clp chaperones
The highly conserved ATP-dependent ClpP proteases aretwo-component proteases consisting of separatelyencoded ATPase and peptidase subunits (Fig. 1A). Exten-sive structural analysis of the ClpP proteases fromEscherichia coli has formed the paradigm for the way weunderstand the function of these proteins (reviewed bySauer et al., 2004). The central proteolytic core consistsof 14 ClpP serine peptidase subunits stacked in two hep-tameric rings, forming an internal chamber in which theactive sites are sequestered from the cytoplasm (Wanget al., 1997). Narrow axial pores that exclude native pro-teins and all but the smallest peptides control access tothis proteolytic chamber. To gain proteolytic activity, theClpP multimer associates with one or two hexameric ringsof Clp ATPases, forming the ClpP-containing proteolyticcomplex (designated the ClpP protease). In associationwith the proteolytic core, the Clp ATPases are responsiblefor the recognition, unfolding and translocation of sub-strates into the ClpP degradation chamber (Zolkiewski,2006). This overall structure and organization bearsresemblance to the 26S proteasome of eukaryotic cells(Kessel et al., 1995).
The Clp ATPases constitute a family of closely relatedproteins that carry one or two nucleotide binding domainstypical of the AAA+ superfamily of ATPases (Schirmeret al., 1996; Neuwald et al., 1999). Clp ATPases aredivided into subfamilies on the basis of the presence ofspecific signature sequences and the number andspacing of the nucleotide binding sites (Fig. 1B). A role inprotein folding, activation, or disaggregation seems to beshared by all Clp ATPases (Zolkiewski, 2006). However,only a subgroup of the Clp ATPases can interact withClpP, a property that is associated with the presence of aClpP recognition tripeptide found in ClpA, ClpC and ClpE
Accepted 5 January, 2007. *For correspondence. E-mail [email protected]; Tel. (+45) 3528 2773; Fax (+45) 3528 2755.
Molecular Microbiology (2007) 63(5), 1285–1295 doi:10.1111/j.1365-2958.2007.05598.x
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
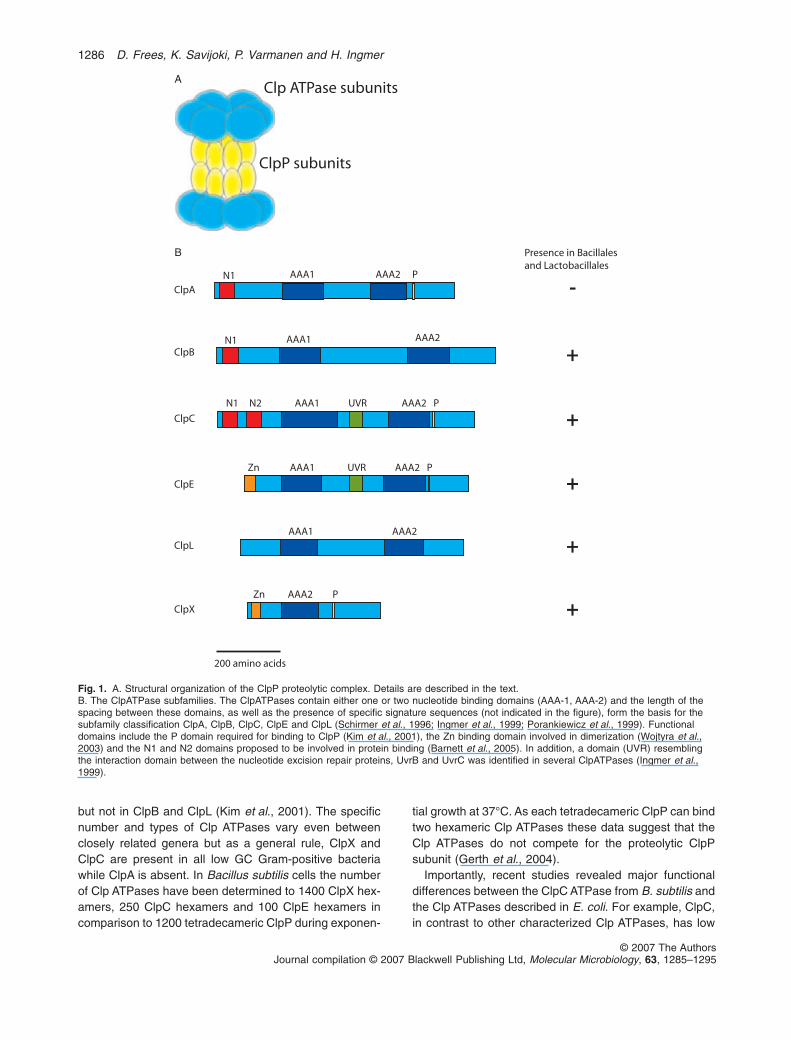
but not in ClpB and ClpL (Kim et al., 2001). The specificnumber and types of Clp ATPases vary even betweenclosely related genera but as a general rule, ClpX andClpC are present in all low GC Gram-positive bacteriawhile ClpA is absent. In Bacillus subtilis cells the numberof Clp ATPases have been determined to 1400 ClpX hex-amers, 250 ClpC hexamers and 100 ClpE hexamers incomparison to 1200 tetradecameric ClpP during exponen-
tial growth at 37°C. As each tetradecameric ClpP can bindtwo hexameric Clp ATPases these data suggest that theClp ATPases do not compete for the proteolytic ClpPsubunit (Gerth et al., 2004).
Importantly, recent studies revealed major functionaldifferences between the ClpC ATPase from B. subtilis andthe Clp ATPases described in E. coli. For example, ClpC,in contrast to other characterized Clp ATPases, has low
Clp ATPase subunitsA
B
ClpP subunits
N1 N2 AAA1 AAA2
ClpC
ClpE
Zn
UVR
ClpL
P
P
PAAA1 AAA2UVR
Zn
AAA1 AAA2
AAA2
ClpX
ClpBN1
ClpAN1
AAA1 AAA2
AAA1 AAA2
Presence in Bacillales and Lactobacillales
-
+
+
+
+
+
P
200 amino acids
Fig. 1. A. Structural organization of the ClpP proteolytic complex. Details are described in the text.B. The ClpATPase subfamilies. The ClpATPases contain either one or two nucleotide binding domains (AAA-1, AAA-2) and the length of thespacing between these domains, as well as the presence of specific signature sequences (not indicated in the figure), form the basis for thesubfamily classification ClpA, ClpB, ClpC, ClpE and ClpL (Schirmer et al., 1996; Ingmer et al., 1999; Porankiewicz et al., 1999). Functionaldomains include the P domain required for binding to ClpP (Kim et al., 2001), the Zn binding domain involved in dimerization (Wojtyra et al.,2003) and the N1 and N2 domains proposed to be involved in protein binding (Barnett et al., 2005). In addition, a domain (UVR) resemblingthe interaction domain between the nucleotide excision repair proteins, UvrB and UvrC was identified in several ClpATPases (Ingmer et al.,1999).
1286 D. Frees, K. Savijoki, P. Varmanen and H. Ingmer
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

intrinsic ATPase activity and depends on cofactors to gainchaperone activity (Schlothauer et al., 2003; Kirsteinet al., 2006). MecA, the best-characterized ClpC cofactor,facilitates ClpC oligomerization into a complex containingboth ClpC and MecA (Kirstein et al., 2006). Formation ofthis complex is a prerequisite for all ClpC mediated activi-ties, including ATPase activity, substrate recognition andinteraction with ClpP (Kirstein et al., 2006). In E. coliadaptor proteins are required only to modulate substraterecognition, not for basic functions of their ClpATPasepartner (see review by Dougan et al., 2002). Based onthis important difference we propose to use the termcofactor instead of adaptor protein for MecA and relatedproteins.
Regulation of ClpP and the Clp ATPases
Production of ClpP and most Clp ATPases is stronglyincreased in response to heat-shock and other stressconditions. In low GC Gram-positive bacteria, transcrip-tion control of the clp genes is primarily mediated bybinding of the CtsR repressor to a heptanucleotide repeatA/GGTCAAA/T located in the sA promoter region of targetgenes (Derré et al., 1999; Nair et al., 2000). However, insome organisms the HrcA repressor as well as the alter-native sigma factor, sB are also involved as summarized inFig. 2 (Krüger and Hecker, 1998; Derré et al., 1999; 2000;Varmanen et al., 2000; Chastanet et al., 2001; Chastanetand Msadek, 2003; Grandvalet et al., 2005).
The central question of how the repressor activity ofCtsR is relieved in response to heat shock has beenaddressed in B. subtilis, where detailed in vitro studieshave demonstrated that DNA binding by CtsR is modu-lated by a complex titrating mechanism involving McsB,McsA and ClpC, all encoded by the ctsR operon (Krügeret al., 2001; Kirstein et al., 2005). According to the presentmodel illustrated in Fig. 3, McsA, McsB and ClpC form aternary complex in which the kinase activity of McsB isinhibited during non-stress conditions (Kirstein andTurgay, 2005; Kirstein et al., 2005). Upon protein denatur-ation, a hallmark of most stress conditions and notablyof heat stress, ClpC is titrated away from the ternarycomplex (Kirstein and Turgay, 2005). In the absence ofClpC, the kinase activity of McsB is stimulated by McsAand the resulting phosphorylation of McsB enhances itsaffinity for CtsR, leading to the formation of a new ternarycomplex consisting of phosphorylated McsA, McsB andCtsR. The formation of this complex and the phosphory-lation of CtsR result in sequestration of CtsR from itstarget promoters, leading to derepression of the CtsRregulon. Interestingly, the phosphorylation of CtsR alsoseems to be implicated in tagging it for degradation byClpCP and ClpEP during stress conditions (Kirstein et al.,2005; Miethke et al., 2006). CtsR is part of the CtsR
regulon and its synthesis is greatly stimulated in responseto stress. Therefore, rapid inactivation of newly synthe-sized CtsR by phosphorylation and subsequent degrada-tion is probably required to sustain derepression of theCtsR regulon until the problems with non-native proteinsare overcome (Kirstein and Turgay, 2005). Notably, clpXappears not to be part of the CtsR regulon in any organ-ism examined so far. Based on this observation and thelack of heat induction of ClpX we speculate that theprimary cellular role of ClpX is not linked to handlingstress-damaged proteins in low GC Gram-positive bacte-ria (Frees et al., 2003; Gerth et al., 2004).
Clp proteins are indispensable in protein qualitycontrol during both stress and non-stressconditions
The implication of Clp proteins in protein quality controlin the stressed cell
Unfolding and subsequent aggregation of denatured pro-teins is the hallmark of heat shock and most other stressconditions and ClpP proteases clearly play an essentialrole in removing heat-damaged proteins from the low GCGram-positive bacteria. This notion was first demonstratedindirectly by showing that bacterial mutants lacking ClpP
Bacillales Lactobacillales
S. salivariusS.thermophilus
clpP/C/B/E/L
clpP/E/C/B
groESLdnaK
clpPgroESLdnaK
clpP/E/LgroESLdnaK
clpP/C/E
groESLdnaK
clpP/C/B
groESLdnaK
clpP/C/LgroESLdnaK
Lactobacillusgasseri
LactococcusStreptococcus
Oenococcusoeni
S. aureus
BacillusListeria
clpL
σΒ
Fig. 2. Regulation of clp gene expression by CtsR, HrcA and sB.In low GC firmicutes the composition of CtsR (boxed) and HrcA(circle) and overlapping CtsR and HrcA (dotted) regulons showconsiderable variation. In Bacilli the CtsR and HrcA regulons aredistinct, whereas the HrcA regulon is embedded within the CtsRregulon in staphylococci. Alternative sigma factor sB plays a minorrole (dashed line, B) in expression of clpP and clpC in B. subtilisand L. monocytogenes, whereas clpL of S. aureus is solelytranscribed by sB. In Lactobacillales the CtsR and HrcA regulonscoexist and partially overlap in several members ofStreptococcaceae. The clp gene regulation in the other members ofthis order are predominantly controlled by either CtsR or HrcA.(Model adapted from Chastanet and Msadek, 2003.)
Clp ATPases and ClpP proteolytic complexes 1287
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

were restricted for growth at high temperatures in a widerange of bacteria including B. subtilis, Lactococcus lactis,Listeria monocytogenes, Staphylococcus aureus andStreptococci (Msadek et al., 1998; Frees and Ingmer,1999; Gaillot et al., 2000; Lemos and Burne, 2002; Rob-ertson et al., 2002, Frees et al., 2003; Nair et al., 2003).Directly, it was observed that inactivation of ClpPdecreased in vivo degradation of non-native model sub-strates (puromycyl-peptides) to less than 30% of the wild-type level in L. lactis, L. monocytogenes and in B. subtilis(Frees and Ingmer, 1999; Gaillot et al., 2000; Krüger et al.,2000). To recognize and degrade non-native proteins,ClpP must associate with a ClpATPase partner and currentknowledge supports that ClpC is the principal ClpATPaseresponsible for recruiting ClpP to degrade non-native pro-teins in B. subtilis and S. aureus (Krüger et al., 2000; Freeset al., 2004; Kock et al., 2004a). Similarly, ClpE appears toplay a role in protein quality control during heat stresswhereas the function of ClpX varies between organisms(Nair et al., 1999; Chastanet et al., 2001; Miethke et al.,2006). For example a B. subtilis clpX mutant is heat sen-sitive (Gerth et al., 1998) while deletion of clpX improvedheat tolerance of S. aureus (Frees et al., 2003; 2004).ClpX is essential in lactococci and streptococci but themolecular basis behind this observation is unknown(Robertson et al., 2003; K. Savijoki, unpubl. results).
Finally, ClpB is important for growth at very hightemperatures as well as for induced thermotolerance inL. monocytogenes and S. aureus (Chastanet et al., 2004;Frees et al., 2004). The absence of a ClpP interactiondomain in ClpB suggests that ClpB primarily functions asa chaperone and that the unique role of ClpB in solubiliz-ing protein aggregates described in E. coli is conserved inGram-positive bacteria (Mogk et al., 1999). Interestingly,ClpL also lacks the ClpP interaction domain and the phe-notypic resemblance between clpL and clpB mutants in
S. aureus indicates that ClpL and ClpB may have a similarfunction (Frees et al., 2004).
Protein quality control in the non-stressed cell
Measurements of overall protein degradation rates inB. subtilis growing under non-stress conditions revealedthat cellular proteins were largely stable during exponen-tial growth but overall protein degradation occurred at arelatively high rate upon entry into stationary phase (Kocket al., 2004a). Importantly, this growth phase-dependentbreakdown of cellular proteins did not occur in cellslacking ClpP, suggesting that ClpP is the major determi-nant of bulk protein turnover in B. subtilis even undernon-stress conditions. The importance of ClpP for proteinquality control in non-stressed cells was further empha-sized by the finding that the absence of ClpP caused20–30% of newly synthesized proteins to aggregate(Kock et al., 2004a). In accordance, electron-dense par-ticles representing insoluble protein aggregates wereobserved in non-stressed Bacillus cells lacking ClpP activ-ity (Krüger et al., 2000). An example of a conservedprokaryotic ClpP-dependent protein quality mechanismoperating in the non-stressed cell is the cotranslationallytagging of proteins synthesized from damaged mRNAs byan 11 amino acid peptide (AANDENYALAA) that willdestine the tagged proteins for degradation by the ClpPproteases (Keiler et al., 1996; Wiegert and Schumann,2001). Hence, cotranslationally tagged proteins may rep-resent one group of newly synthesized proteins that endup in aggregates in cells devoid of ClpP.
ClpP proteolytic complexes control specific stressresponses by regulated proteolysis
Recent data emphasize that ClpP mediated proteolysisalso plays a more specific role in cellular stress responses
McsA
CtsR
Heat shock gene
McsA
McsB
CtsR
P
CtsR
Unfoldedprotein
McsAMcsB
P
P
McsB
CtsR
Heat shock gene Heat shock geneekpression
P
ClpC/ClpE
Heat shock geneekpression
ClpP
ClpC
ClpC
Protein unfolding stress
Co-factor?
P
Fig. 3. In response to stress the repressor activity of CtsR is inhibited by McsA/McsB-mediated phosphorylation and subsequent degradationby ClpCP. Details of the model are described in the text. Long arrow indicates high level gene expression, broken arrow indicates low levelgene expression and P indicates phosphorylation.
1288 D. Frees, K. Savijoki, P. Varmanen and H. Ingmer
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295
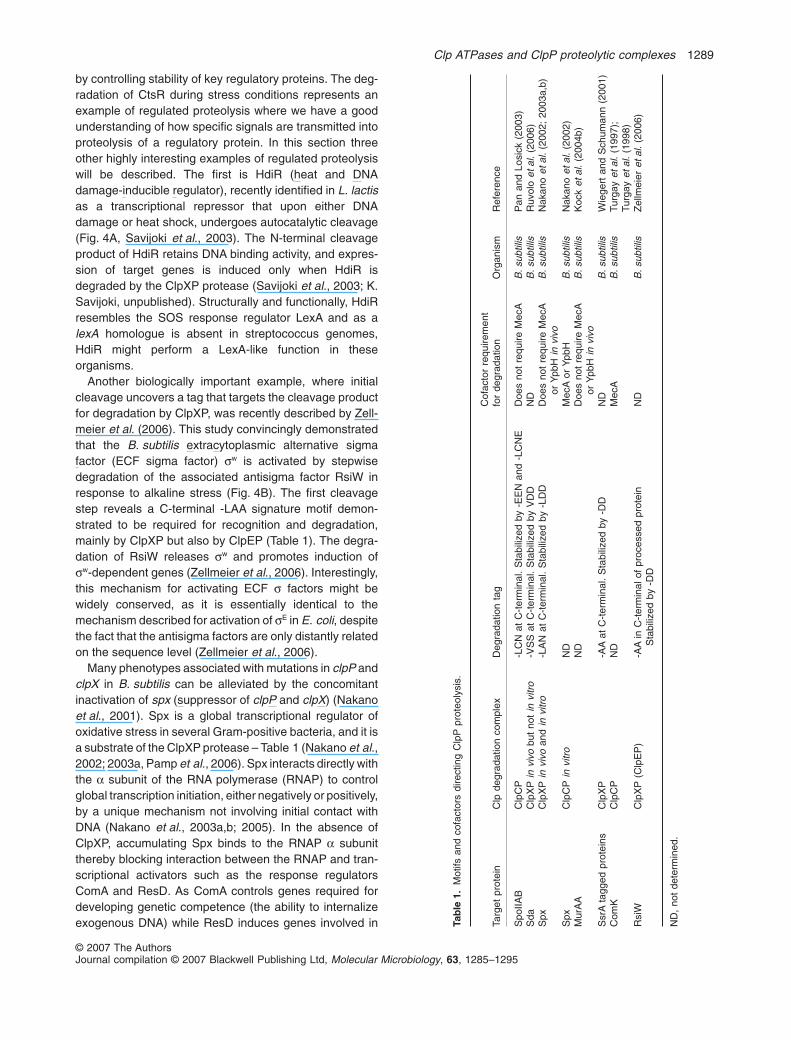
by controlling stability of key regulatory proteins. The deg-radation of CtsR during stress conditions represents anexample of regulated proteolysis where we have a goodunderstanding of how specific signals are transmitted intoproteolysis of a regulatory protein. In this section threeother highly interesting examples of regulated proteolysiswill be described. The first is HdiR (heat and DNAdamage-inducible regulator), recently identified in L. lactisas a transcriptional repressor that upon either DNAdamage or heat shock, undergoes autocatalytic cleavage(Fig. 4A, Savijoki et al., 2003). The N-terminal cleavageproduct of HdiR retains DNA binding activity, and expres-sion of target genes is induced only when HdiR isdegraded by the ClpXP protease (Savijoki et al., 2003; K.Savijoki, unpublished). Structurally and functionally, HdiRresembles the SOS response regulator LexA and as alexA homologue is absent in streptococcus genomes,HdiR might perform a LexA-like function in theseorganisms.
Another biologically important example, where initialcleavage uncovers a tag that targets the cleavage productfor degradation by ClpXP, was recently described by Zell-meier et al. (2006). This study convincingly demonstratedthat the B. subtilis extracytoplasmic alternative sigmafactor (ECF sigma factor) sw is activated by stepwisedegradation of the associated antisigma factor RsiW inresponse to alkaline stress (Fig. 4B). The first cleavagestep reveals a C-terminal -LAA signature motif demon-strated to be required for recognition and degradation,mainly by ClpXP but also by ClpEP (Table 1). The degra-dation of RsiW releases sw and promotes induction ofsw-dependent genes (Zellmeier et al., 2006). Interestingly,this mechanism for activating ECF s factors might bewidely conserved, as it is essentially identical to themechanism described for activation of sE in E. coli, despitethe fact that the antisigma factors are only distantly relatedon the sequence level (Zellmeier et al., 2006).
Many phenotypes associated with mutations in clpP andclpX in B. subtilis can be alleviated by the concomitantinactivation of spx (suppressor of clpP and clpX) (Nakanoet al., 2001). Spx is a global transcriptional regulator ofoxidative stress in several Gram-positive bacteria, and it isa substrate of the ClpXP protease – Table 1 (Nakano et al.,2002; 2003a, Pamp et al., 2006). Spx interacts directly withthe a subunit of the RNA polymerase (RNAP) to controlglobal transcription initiation, either negatively or positively,by a unique mechanism not involving initial contact withDNA (Nakano et al., 2003a,b; 2005). In the absence ofClpXP, accumulating Spx binds to the RNAP a subunitthereby blocking interaction between the RNAP and tran-scriptional activators such as the response regulatorsComA and ResD. As ComA controls genes required fordeveloping genetic competence (the ability to internalizeexogenous DNA) while ResD induces genes involved in Ta
ble
1.M
otifs
and
cofa
ctor
sdi
rect
ing
Clp
Ppr
oteo
lysi
s.
Targ
etpr
otei
nC
lpde
grad
atio
nco
mpl
exD
egra
datio
nta
gC
ofac
tor
requ
irem
ent
for
degr
adat
ion
Org
anis
mR
efer
ence
Spo
IIAB
Clp
CP
-LC
Nat
C-t
erm
inal
.S
tabi
lized
by-E
EN
and
-LC
NE
Doe
sno
tre
quire
Mec
AB
.sub
tilis
Pan
and
Losi
ck(2
003)
Sda
Clp
XP
invi
vobu
tno
tin
vitr
o-V
SS
atC
-ter
min
al.
Sta
biliz
edby
VD
DN
DB
.sub
tilis
Ruv
olo
etal
.(2
006)
Spx
Clp
XP
invi
voan
din
vitr
o-L
AN
atC
-ter
min
al.
Sta
biliz
edby
-LD
DD
oes
not
requ
ireM
ecA
orY
pbH
invi
voB
.sub
tilis
Nak
ano
etal
.(2
002;
2003
a,b)
Spx
Clp
CP
invi
tro
ND
Mec
Aor
Ypb
HB
.sub
tilis
Nak
ano
etal
.(2
002)
Mur
AA
ND
Doe
sno
tre
quire
Mec
Aor
Ypb
Hin
vivo
B.s
ubtil
isK
ock
etal
.(2
004b
)
Ssr
Ata
gged
prot
eins
Clp
XP
-AA
atC
-ter
min
al.
Sta
biliz
edby
-DD
ND
B.s
ubtil
isW
iege
rtan
dS
chum
ann
(200
1)C
omK
Clp
CP
ND
Mec
AB
.sub
tilis
Turg
ayet
al.
(199
7);
Turg
ayet
al.
(199
8)R
siW
Clp
XP
(Clp
EP
)-A
Ain
C-t
erm
inal
ofpr
oces
sed
prot
ein
Sta
biliz
edby
-DD
ND
B.s
ubtil
isZ
ellm
eier
etal
.(2
006)
ND
,no
tde
term
ined
.
Clp ATPases and ClpP proteolytic complexes 1289
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

HdiR hdiR hdiRHdiR-N
hdiR geneexpression
HdiR-NA
σW
ECF regulated gene
σW
σW promoter
ECF regulated geneσW promoter
Expression of ECF regulated gene
σW
B
Alkaline stress
DNA damage or heat
ClpX
ClpP
hdiR promoter
RsiWRsiW-N
RsiW-N
ClpX
ClpP
SpoIIABσF
Sporulation gene
σF
SpoIIAB
SpoIIAA
SpoIIAB
σF promoter
Sporulation geneσF promoter
Sporulation geneexpression
σF
C
Sporulationsignal ClpC
ClpP
D
MecA
Competence gene
ComK
ClpC
Competence gene
MecA
ComS
ClpC
ComK
Competencesignal
Competence geneexpression
ComS
MecA
ComK
ClpC
ClpP
ClpC
ClpP
ComK
Fig. 4. Examples of mechanisms underlying regulated proteolysis by ClpP proteases.A. Degradation of the transcription factor HdiR by ClpXP in response to heat stress or DNA damage.B. Regulated proteolysis of the antisigma factor RsiW by ClpXP/ClpEP in response to alkaline stress.C. Regulated proteolysis of the competence regulator, ComK, by ClpCP.D. Degradation of the antisigma factor SpoIIAB by ClpCP during sporulation. See text for details of the depicted models.
1290 D. Frees, K. Savijoki, P. Varmanen and H. Ingmer
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

adaptation to oxygen limitation, this explains why clpP andclpX mutants are defective in developing genetic compe-tence and in growth under oxygen restricted conditions inB. subtilis (Nakano et al., 2003a; Zuber, 2004).
Homologues of Spx are highly conserved among lowGC Gram-positive bacteria (Zuber, 2004). Interestingly,the spx homologue trmA was previously identified as thesite of mutations that could alleviate the heat-sensitivephenotype of clpP and recA mutant in L. lactis (Duwatet al., 1999; Frees et al., 2001). L. lactis encodes 7 Spxparalogues and we hypothesize that these proteins rep-resent RNAP binding global regulators whose activity iscontrolled by ClpP mediated proteolysis. The signals thatgovern ClpXP regulated proteolysis of Spx have not yetbeen identified.
Finally, a recently published DNA-microarray studycomparing overall transcription in wild-type S. aureus andits clpP mutant revealed that ClpP influences expressionof multiple central regulons, including the PerR, Fur, MntRand LexA regulons (Michel et al., 2006). Thus, the numberof regulators whose activity is controlled by ClpP medi-ated proteolysis might be significantly underestimated atpresent.
Clp proteins in cellular differentiation
Clp proteins controlling developmental programs
In response to starvation for essential nutrients B. subtilisinitiates several developmental programs including sporu-lation and competence development. The ClpXP medi-ated control of the cellular level of Spx represents anexample of how ClpXP regulates development of compe-tence, however, Clp protein control multiple key switchesin these programs. Central to competence development isthe transcriptional activator, ComK that is required forexpression of genes encoding the DNA uptake apparatus.Importantly, ComK also positively regulates its ownexpression and therefore only small fluctuations in thelevel of ComK can initiate a positive feed-back loopleading to the development of competence as recentlydiscussed by Dubnau and Losick (2006). Accordingly, pro-duction of ComK is carefully controlled at multiple levelsand any ComK molecules that escape this tight controlare via binding to MecA directed to ClpCP for degradation(Fig. 4C, Msadek et al., 1994; Kong and Dubnau, 1994;Turgay et al., 1997; 1998). The ComK pathway, and con-sequently competence, is induced when synthesis ofComS is initiated as a result of signalling by a quorum-sensing two component regulatory pathway. Subse-quently, ComS displaces ComK from the MecA-ClpCternary complex and the released ComK then activatestranscription of its own gene, thereby initiating a burst ofcompetence gene expression (Turgay et al., 1997).
In response to more severe starvation B. subtilis ini-tiates sporulation. Timely and spatial expression of tran-scription factors is essential for successful sporulationand both chaperone and proteolytic activities of the Clpproteins are required at multiple levels. ClpX is required atthe onset of sporulation to activate the alternative sigmafactor sH (Spo0H) and once it has completed its tasks sH
is removed in a process involving ClpC, most likely as partof a ClpCP complex (Nanamiya et al., 1998; Liu et al.,1999; Liu and Zuber, 2000). During sporulation, the activ-ity of the sporulation sigma factor sF is confined to theforespore by ClpCP (Pan et al., 2001). At the transitionfrom the vegetative cell to the forespore sF is activated bybeing released from the anti-sigma factor SpoIIAB in aprocess involving the anti-antisigma factor SpoIIAA(Fig. 4D). Dissociation of sF from SpoIIAB uncovers aClpC recognition motif leading to degradation by ClpCP(Pan et al., 2001; Table 1).
Finally, ClpP affects the co-ordination of DNA replicationand initiation of sporulation by degrading Sda, a smallprotein whose synthesis is induced in response to pertur-bations in chromosome replication or elongation. Sdaspecifically inhibits the histidine kinases required to ini-tiate spore development, and recovery from this staterequires the removal of Sda by the ClpXP protease(Table 1, Ruvolo et al., 2006).
Clp proteins affecting cell division and transitionbetween growth phases
Inactivation of Clp proteins often leads to aberrant mor-phology such as filamentation, impaired cell septation andseparation, abnormal cell wall structure and irregular celldivision (Gerth et al., 1998; Msadek et al., 1998; Nairet al., 1999; 2003). Collectively, these data suggest thatClp ATPases and ClpP proteolytic complexes are requiredfor basic biological processes fundamental to cell integrity.Recent reports have shed some light on the processesthat might underlie these observations. As describedabove, massive ClpP-dependent degradation of proteinsproduced during exponential growth occurs in the transi-tion to stationary phase in B. subtilis (Kock et al., 2004a).One of the ClpCP-degraded substrates is MurAA(Table 1), which catalyses carboxyvinyl transfer fromphosphoenolpyruvate to UDP-N-acetyglucosamine, thefirst committed step in peptidoglycan biosynthesis (Kocket al., 2004b). Notably, cells overproducing MurAA displaythe same filamentous growth as clpP mutant cells, sug-gesting that this clpP phenotype is at least in part be dueto failure to degrade MurAA (Kock et al., 2004b). Interest-ingly, an independent role of ClpX in Bacillus cell divisionwas recently demonstrated by showing that ClpX func-tions as an inhibitor of the cell division protein FtsZ, pos-sibly by hindering FtsZ polymerization that is required for
Clp ATPases and ClpP proteolytic complexes 1291
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

septum formation by sequestering unassembled FtsZprotein (Weart et al., 2005).
Clp proteins affecting virulence of pathogens
Intriguingly, inactivation of ClpP or Clp ATPases signifi-cantly reduces virulence of the important pathogens,S. aureus, L. monocytogenes and Streptococcus pneu-moniae (Gaillot et al., 2000; Frees et al., 2003; Ibrahimet al., 2005; reviewed by Butler et al., 2006).
The entry of bacterial pathogens into host organismsconstitutes a dramatic environmental change and theapparently conserved role of Clp proteins in virulencecould be a consequence of their involvement in handlingstress imposed damage of bacterial proteins. As anexample, S. aureus cells lacking the ClpB chaperone areunable to replicate intracellularly in bovine cells (Freeset al., 2004). As ClpB is needed for growth only underconditions generating massive protein aggregation, anddoes not affect production of any known virulencefactors, this result supports the idea that the Clp proteinquality control systems are required in the host.However, Clp chaperones and proteases also contributeto virulence by controlling synthesis of major virulencefactors. For example, ClpC is required for transcription ofinlA, inlB and actA encoding proteins important for hostcell invasion of L. monocytogenes; while the absence ofClpP reduces activity, but not production of listeriolysinO, a major virulence factor implicated in phagosomelysis (Nair et al., 2000; Gaillot et al., 2000; 2001). Simi-larly, S. aureus ClpXP is required for transcription of anumber of important virulence genes encoding haemol-ysins, extracellular proteases surface adhesins andothers (Frees et al., 2003; 2005a; Michel et al., 2006).Interestingly, the regulatory activity of ClpXP seems tobe tightly linked to the quorum sensing Agr two-component system, the best-characterized virulenceregulator in this organism (reviewed by Novick, 2003).Induction of the agr operon in response to cell densityleads to the production of the 514 nt small regulatoryRNA, RNAIII, that is the actual effecter of virulence generegulation. How RNAIII regulates the transcription ofmore than 50 genes remains puzzling but recently it wasshown that RNAIII inhibits translation of rot encoding atranscriptional regulator of virulence genes (Geisingeret al., 2006). We have proposed that RNAIII may addi-tionally bind directly to regulatory proteins thereby induc-ing a conformational change that destine the proteins fordegradation by ClpXP (Frees et al., 2005a). Noteworthy,ClpX independent of ClpP is essential for transcription ofspa, encoding the important Staphylococcal surfaceprotein, Protein A (Frees et al., 2003; 2005a). At presentwe believe that ClpX is required either for expression,folding or dimerization of Rot that is a transcriptional
activator of spa transcription (Frees et al., 2005a;Oscarsson et al., 2005).
The conserved role of Clp proteins in virulence mayalso be exploited therapeutically. Interestingly, S. aureusClpP was recently shown to be the target of a newpromising class of antibiotics, the acyldepsipeptides(Brotz-Oesterhelt et al., 2005). This compound apparentlyenables ClpP to degrade proteins in the absence of anassociating Clp ATPase, leading to uncontrolled proteoly-sis that eventually kills the cells.
Concluding remarks and future perspectives
Bacteria can adapt to a wide range of growth conditions.Traditionally, studies have focused on changes in tran-scription, but it has become clear that regulated proteoly-sis represent an equally important aspect of growthadaptation. In E. coli four classes of ATP-dependent pro-teases [ClpAP/XP, ClpYQ (HslVU), Lon and FtsH] haveoverlapping substrate specificities, perhaps explainingwhy inactivation of individual proteases causes onlymodest phenotypic changes under the conditions tested(reviewed by Gottesman, 2003). In striking contrast thewide range of phenotypes conferred by inactivation ofClpP in the low GC Gram-positive bacteria suggest thatthe ClpP proteolytic proteases are the major proteasesboth for eliminating misfolded proteins and for controllingthe activity of central regulatory proteins in theseorganisms. In streptococci homologues of Lon and ClpYQare missing whereas in S. aureus ClpYQ is required forgrowth at very high temperature, suggesting that ClpYQmediated proteolysis becomes essential only when heat-induced protein denaturation exceeds the capacity of theClpP complex (Frees et al., 2005b). Thus, the ATP-dependent proteases appear to be dedicated to special-ized tasks in Gram-positive bacteria. Additional importantmechanistic differences to the E. coli paradigm was dem-onstrated by the finding that ClpC in contrast to othercharacterized ClpATPases has low intrinsic ATPase activ-ity and depends on cofactors for all basic activities. Onemajor task of future studies will be to understand the roleof cofactors in relation to selection of substrates. A break-through in understanding the general rules governing sub-strate recognition by the ClpXP protease was recentlyobtained in E. coli, where specifically one large class ofsubstrate proteins carry the C-terminal -LAA signaturethat is part of the SsrA tag (Flynn et al., 2003). Similarmotifs have been shown to be required for ClpP-dependent proteolysis in Gram-positive bacteria suggest-ing that recognition of substrate proteins follow thegeneral rules established in E. coli (Table 1).
A very interesting aspect of regulated proteolysis is theconditional degradation that occurs only in response tospecific cellular signals. Examples of factors modulating
1292 D. Frees, K. Savijoki, P. Varmanen and H. Ingmer
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

degradation are shown in Figs 3 and 4. One challenge ofthe future will be to unravel the complex networks control-ling regulated proteolysis. In the case of competencedevelopment proteolysis is regulated in response to envi-ronmental signals perceived and transmitted via two-component signal transduction systems. Alternatively,cellular processes such as DNA damage-repair may com-municate with ClpATPases via the UVR domain (Fig. 1B),which resembles the UvrB–UvrC interaction domain. Thiscommunication event is functionally similar to the interac-tion that takes place between the DNA damage repairproteins and the 26S proteasome in eukaryotes (Swederand Madura, 2002). Studies aimed at elucidating themechanisms underlying regulated proteolysis, and spe-cifically the ways external signals are communicated tothe proteases, will be central for future work in this field.
Acknowledgements
We are thankful for the support of the Danish ResearchCouncil to D. Frees and for the support of the Academy ofFinland to P. Varmanen (project no. 211165). We also grate-fully acknowledge the helpful comments provided by M.T.Cohn, L. Jelsbak, L. E. Thomsen and M. Bojesen.
References
Barnett, M.E., Nagy, M., Kedzierska, S., and Zolkiewski, M.(2005) The amino-terminal domain of ClpB supportsbinding to strongly aggregated proteins. J Biol Chem 280:34940–34945.
Brotz-Oesterhelt, H., Beyer, D., Kroll, H.P., Endermann, R.,Ladel, C., Schroeder, W., et al. (2005) Dysregulation ofbacterial proteolytic machinery by a new class ofantibiotics. Nat Med 11: 1082–1087.
Butler, S.M., Festa, R.A., Pearce, M.J., Darwin, K., and H.(2006) Self-compartmentalized bacterial proteases andpathogenesis. Mol Microbiol 60: 553–562.
Chastanet, A., and Msadek, T. (2003) ClpP of Streptococcussalivarius is a novel member of the dually regulated classof stress response genes in Gram-positive bacteria.J Bacteriol 185: 683–687.
Chastanet, A., Prudhomme, M., Claverys, J.P., and Msadek,T. (2001) Regulation of Streptococcus pneumoniae clpgenes and their role in competence development andstress survival. J Bacteriol 183: 7295–7307.
Chastanet, A., Derre, I., Nair, S., and Msadek, T. (2004) clpB,a novel member of the Listeria monocytogenes CtsRregulon, is involved in virulence but not in general stresstolerance. J Bacteriol 186: 1165–1174.
Derré, I., Rapoport, G., and Msadek, T. (1999) CtsR, a novelregulator of stress and heat shock response, controls clpand molecular chaperone gene expression in Gram-positive bacteria. Mol Microbiol 31: 117–131.
Derré, I., Rapoport, G., and Msadek, T. (2000) The CtsRregulator of stress response is active as a dimer and spe-cifically degraded in vivo at 37 degrees C. Mol Microbiol38: 335–347.
Dougan, D.A., Mogk, A., Zeth, K., Turgay, K., and Bukau, B.(2002) AAA+ proteins and substrate recognition, it alldepends on their partner in crime. FEBS Lett 529: 6–10.
Dubnau, D., and Losick, R. (2006) Bistability in bacteria. MolMicrobiol 61: 564–572.
Duwat, P., Ehrlich, S.D., and Gruss, A. (1999) Effects ofmetabolic flux on stress response pathways in Lactococ-cus lactis. Mol Microbiol 31: 845–858.
Flynn, J.M., Neher, S.B., Kim, Y.I., Sauer, R.T., and Baker,T.A. (2003) Proteomic discovery of cellular substratesof the ClpXP protease reveals five classes of ClpX-recognition signals. Mol Cell 11: 671–683.
Frees, D., and Ingmer, H. (1999) ClpP participates in thedegradation of misfolded protein in Lactococcus lactis. MolMicrobiol 31: 79–87.
Frees, D., Varmanen, P., and Ingmer, H. (2001) Inactivationof a gene that is highly conserved in Gram-positive bacteriastimulates degradation of non-native proteins and con-comitantly increases stress tolerance in Lactococcus lactis.Mol Microbiol 41: 93–103.
Frees, D., Qazi, S.N., Hill, P.J., and Ingmer, H. (2003) Alter-native roles of ClpX and ClpP in Staphylococcus aureusstress tolerance and virulence. Mol Microbiol 48: 1565–1578.
Frees, D., Chastanet, A., Qazi, S., Sorensen, K., Hill, P.,Msadek, T., and Ingmer, H. (2004) Clp ATPases arerequired for stress tolerance, intracellular replication andbiofilm formation in Staphylococcus aureus. Mol Microbiol54: 1445–1462.
Frees, D., Sorensen, K., and Ingmer, H. (2005a) Global viru-lence regulation in Staphylococcus aureus: pinpointing theroles of ClpP and ClpX in the sar/agr regulatory network.Infect Immun 73: 8100–8108.
Frees, D., Thomsen, L.E., and Ingmer, H. (2005b) Staphylo-coccus aureus ClpYQ plays a minor role in stress survival.Arch Microbiol 183: 286–291.
Gaillot, O., Pellegrini, E., Bregenholt, S., Nair, S., andBerche, P. (2000) The ClpP serine protease is essential forthe intracellular parasitism and virulence of Listeriamonocytogenes. Mol Microbiol 35: 1286–1294.
Gaillot, O., Bregenholt, S., Jaubert, F., Di Santo, J.P., andBerche, P. (2001) Stress-induced ClpP serine protease ofListeria monocytogenes is essential for induction of listeri-olysin O-dependent protective immunity. Infect Immun 69:4938–4943.
Geisinger, E., Adhikari, R.P., Jin, R., Ross, H.F., and Novick,R.P. (2006) Inhibition of rot translation by RNAIII, a keyfeature of agr function. Mol Microbiol 61: 1038–1048.
Gerth, U., Krüger, E., Derre, I., Msadek, T., and Hecker, M.(1998) Stress induction of the Bacillus subtilis clpP geneencoding a homologue of the proteolytic component of theClp protease and the involvement of ClpP and ClpX instress tolerance. Mol Microbiol 28: 787–802.
Gerth, U., Kirstein, J., Mostertz, J., Waldminghaus, T.,Miethke, M., Kock, H., and Hecker, M. (2004) Fine-tuningin regulation of Clp protein content in Bacillus subtilis.J Bacteriol 186: 179–191.
Gottesman, S. (2003) Proteolysis in bacterial regulatorycircuits. Annu Rev Cell Dev Biol 19: 565–587.
Grandvalet, C., Coucheney, F., Beltramo, C., and Guzzo, J.(2005) CtsR is the master regulator of stress response
Clp ATPases and ClpP proteolytic complexes 1293
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

gene expression in Oenococcus oeni. J Bacteriol 187:5614–5623.
Ibrahim, Y.M., Kerr, A.R., Silva, N.A., and Mitchell, T.J.(2005) Contribution of the ATP-dependent protease ClpCPto the autolysis and virulence of Streptococcuspneumoniae. Infect Immun 73: 730–740.
Ingmer, H., Vogensen, F.K., Hammer, K., and Kilstrup, M.(1999) Disruption and analysis of the clpB, clpC, and clpEgenes in Lactococcus lactis: ClpE, a new Clp family inGram-positive bacteria. J Bacteriol 181: 2075–2083.
Keiler, K.C., Waller, P.R., and Sauer, R.T. (1996) Role of apeptide tagging system in degradation of proteins synthe-sized from damaged messenger RNA. Science 271: 990–993.
Kessel, M., Maurizi, M.R., Kim, B., Kocsis, E., Trus, B.L.,Singh, S.K., and Steven, A.C. (1995) Homology in struc-tural organization between E. coli ClpAP protease and theeukaryotic 26 S proteasome. J Mol Biol 250: 587–594.
Kim, Y.I., Levchenko, I., Fraczkowska, K., Woodruff, R.V.,Sauer, R.T., and Baker, T.A. (2001) Molecular determi-nants of complex formation between Clp/Hsp100 ATPasesand the ClpP peptidase. Nat Struct Biol 8: 230–233.
Kirstein, J., and Turgay, K. (2005) A new tyrosine phospho-rylation mechanism involved in signal transduction in Bacil-lus subtilis. J Mol Microbiol Biotechnol 9: 182–188.
Kirstein, J., Zuhlke, D., Gerth, U., Turgay, K., and Hecker, M.(2005) A tyrosine kinase and its activator control the activityof the CtsR heat shock repressor in B. subtilis. EMBO J 24:3435–3445.
Kirstein, J., Schlothauer, T., Dougan, D.A., Lilie, H., Tischen-dorf, G., Mogk, A., et al. (2006) Adaptor protein controlledoligomerization activates the AAA+ protein ClpC. EMBO J25: 1481–1491.
Kock, H., Gerth, U., and Hecker, M. (2004a) The ClpP pep-tidase is the major determinant of bulk protein turnover inBacillus subtilis. J Bacteriol 186: 5856–5864.
Kock, H., Gerth, U., and Hecker, M. (2004b) MurAA, catalys-ing the first committed step in peptidoglycan biosynthesis,is a target of Clp-dependent proteolysis in Bacillus subtilis.Mol Microbiol 51: 1087–1102.
Kong, L., and Dubnau, D. (1994) Regulation of competence-specific gene expression by Mec-mediated protein–proteininteraction in Bacillus subtilis. Proc Natl Acad Sci USA 91:5793–5797.
Krüger, E., and Hecker, M. (1998) The first gene of theBacillus subtilis clpC operon, ctsR, encodes a negativeregulator of its own operon and other class III heat shockgenes. J Bacteriol 180: 6681–6688.
Krüger, E., Witt, E., Ohlmeier, S., Hanschke, R., and Hecker,M. (2000) The clp proteases of Bacillus subtilis are directlyinvolved in degradation of misfolded proteins. J Bacteriol182: 3259–3265.
Krüger, E., Zuhlke, D., Witt, E., Ludwig, H., and Hecker, M.(2001) Clp-mediated proteolysis in Gram-positive bacteriais autoregulated by the stability of a repressor. EMBO J 20:852–863.
Lemos, J.A., and Burne, R.A. (2002) Regulation and Physi-ological Significance of ClpC and ClpP in Streptococcusmutans. J Bacteriol 184: 6357–6366.
Liu, J., and Zuber, P. (2000) The ClpX protein of Bacillussubtilis indirectly influences RNA polymerase holoenzyme
composition and directly stimulates sigma-dependenttranscription. Mol Microbiol 37: 885–897.
Liu, J., Cosby, W.M., and Zuber, P. (1999) Role of lon andClpX in the post-translational regulation of a sigma subunitof RNA polymerase required for cellular differentiation inBacillus subtilis. Mol Microbiol 33: 415–428.
Michel, A., Agerer, F., Hauck, C.R., Herrmann, M., Ullrich, J.,Hacker, J., and Ohlsen, K. (2006) Global regulatory impactof ClpP protease of Staphylococcus aureus on regulonsinvolved in virulence, oxidative stress response, autolysis,and DNA repair. J Bacteriol 188: 5783–5796.
Miethke, M., Hecker, M., and Gerth, U. (2006) Involvement ofBacillus subtilis ClpE in CtsR degradation and proteinquality control. J Bacteriol 188: 4610–4619.
Mogk, A., Tomoyasu, T., Goloubinoff, P., Rudiger, S., Roder,D., Langen, H., and Bukau, B. (1999) Identification of ther-molabile Escherichia coli proteins: prevention and rever-sion of aggregation by DnaK and ClpB. EMBO J 18: 6934–6949.
Msadek, T., Kunst, F., and Rapoport, G. (1994) MecB ofBacillus subtilis, a member of the ClpC ATPase family, is apleiotropic regulator controlling competence gene expres-sion and growth at high temperature. Proc Natl Acad SciUSA 91: 5788–5792.
Msadek, T., Dartois, V., Kunst, F., Herbaud, M.L., Denizot, F.,and Rapoport, G. (1998) ClpP of Bacillus subtilis isrequired for competence development, motility, degrada-tive enzyme synthesis, growth at high temperature andsporulation. Mol Microbiol 27: 899–914.
Nair, S., Frehel, C., Nguyen, L., Escuyer, V., and Berche, P.(1999) ClpE, a novel member of the HSP100 family, isinvolved in cell division and virulence of Listeriamonocytogenes. Mol Microbiol 31: 185–196.
Nair, S., Derre, I., Msadek, T., Gaillot, O., and Berche, P.(2000) CtsR controls class III heat shock gene expressionin the human pathogen Listeria monocytogenes. MolMicrobiol 35: 800–811.
Nair, S., Poyart, C., Beretti, J.L., Veiga-Fernandes, H.,Berche, P., and Trieu-Cuot, P. (2003) Role of the Strepto-coccus agalactiae ClpP serine protease in heat-inducedstress defence and growth arrest. Microbiology 149: 407–417.
Nakano, M.M., Hajarizadeh, F., Zhu, Y., and Zuber, P. (2001)Loss-of-function mutations in yjbD result in ClpX- andClpP-independent competence development of Bacillussubtilis. Mol Microbiol 42: 383–394.
Nakano, S., Zheng, G., Nakano, M.M., and Zuber, P. (2002)Multiple pathways of Spx (YjbD) proteolysis in Bacillussubtilis. J Bacteriol 184: 3664–3670.
Nakano, S., Kuster-Schock, E., Grossman, A.D., and Zuber,P. (2003a) Spx-dependent global transcriptional control isinduced by thiol-specific oxidative stress in Bacillus subtilis.Proc Natl Acad Sci USA 10: 13603–13608.
Nakano, S., Nakano, M.M., Zhang, Y., Leelakriangsak, M.,and Zuber, P. (2003b) A regulatory protein that interfereswith activator-stimulated transcription in bacteria. Proc NatlAcad Sci USA 100: 4233–4238.
Nakano, S., Erwin, K.N., Ralle, M., and Zuber, P. (2005)Redox-sensitive transcriptional control by a thiol/disulphideswitch in the global regulator, Spx. Mol Microbiol 55: 498–510.
1294 D. Frees, K. Savijoki, P. Varmanen and H. Ingmer
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295

Nanamiya, H., Ohashi, Y., Asai, K., Moriya, S., Ogasawara,N., Fujita, M., et al. (1998) ClpC regulates the fate of asporulation initiation sigma factor, sigmaH protein. Bacillussubtilis at elevated temperatures. Mol Microbiol 29: 505–513.
Neuwald, A.F., Aravind, L., Spouge, J.L., and Koonin, E.V.(1999) AAA+: a class of chaperone-like ATPases associ-ated with the assembly, operation, and disassembly ofprotein complexes. Genome Res 9: 27–43.
Novick, R.P. (2003) Autoinduction and signal transduction inthe regulation of staphylococcal virulence. Mol Microbiol48: 1429–1449.
Oscarsson, J., Harlos, C., and Arvidson, S. (2005) Regula-tory role of proteins binding to the spa (protein A) and sarS(staphylococcal accessory regulator) promoter regions inStaphylococcus aureus NTCC 8325-4. Int J Med Microbiol295: 253–266.
Pamp, S.J., Frees, D., Engelmann, S., Hecker, M., andIngmer, H. (2006) Spx is a global effector impacting stresstolerance and biofilm formation in Staphylococcus aureus.J Bacteriol 188: 4861–4870.
Pan, Q., and Losick, R. (2003) Unique degradation signal forClpCP in Bacillus subtilis. J Bacteriol 185: 5275–5278.
Pan, Q., Garsin, D.A., and Losick, R. (2001) Self-reinforcingactivation of a cell-specific transcription factor by proteoly-sis of an anti-sigma factor in B. subtilis. Mol Cell 8: 873–883.
Porankiewicz, J., Wang, J., and Clarke, A.K. (1999) Newinsights into the ATP-dependent Clp protease: Escherichiacoli and beyond. Mol Microbiol 32: 449–458.
Robertson, G.T., Ng, W.L., Foley, J., Gilmour, R., andWinkler, M.E. (2002) Global transcriptional analysis of clpPmutations of type 2 Streptococcus pneumoniae and theireffects on physiology and virulence. J Bacteriol 184: 3508–3520.
Robertson, G.T., Ng, W.L., Gilmour, R., and Winkler, M.E.(2003) Essentiality of clpX, but not clpP, clpL, clpC, or clpE.Streptococcus pneumoniae R6. J Bacteriol 185: 2961–2966.
Ruvolo, M.V., Mach, K.E., and Burkholder, W.F. (2006) Pro-teolysis of the replication checkpoint protein Sda is neces-sary for the efficient initiation of sporulation after transientreplication stress in Bacillus subtilis. Mol Microbiol 60:1490–1508.
Sauer, R.T., Bolon, D.N., Burton, B.M., Burton, R.E., Flynn,J.M., Grant, R.A., et al. (2004) Sculpting the proteome withAAA (+) proteases and disassembly machines. Cell 119:9–18.
Savijoki, K., Ingmer, H., Frees, D., Vogensen, F.K., Palva, A.,and Varmanen, P. (2003) Heat and DNA damage inductionof the LexA-like regulator HdiR from Lactococcus lactis ismediated by RecA and ClpP. Mol Microbiol 50: 609–621.
Schirmer, E.C., Glover, J.R., Singer, M.A., and Lindquist, S.(1996) HSP100/Clp proteins: a common mechanismexplains diverse functions. Trends Biochem Sci 21: 289–296.
Schlothauer, T., Mogk, A., Dougan, D.A., Bukau, B., andTurgay, K. (2003) MecA, an adaptor protein necessary forClpC chaperone activity. Proc Natl Acad Sci USA 100:2306–2311.
Sweder, K., and Madura, K. (2002) Regulation of repair bythe 26S proteasome. J Biomed Biotechnol 2: 94–105.
Turgay, K., Hamoen, L.W., Venema, G., and Dubnau, D.(1997) Biochemical characterization of a molecular switchinvolving the heat shock protein ClpC, which controls theactivity of ComK, the competence transcription factor ofBacillus subtilis. Genes Dev 11: 119–128.
Turgay, K., Hahn, J., Burghoorn, J., and Dubnau, D. (1998)Competence in Bacillus subtilis is controlled by regulatedproteolysis of a transcription factor. EMBO J 17: 6730–6738.
Varmanen, P., Ingmer, H., and Vogensen, F.K. (2000) ctsR ofLactococcus lactis encodes a negative regulator of clpgene expression. Microbiology 146: 1447–1455.
Wang, J., Hartling, J.A., and Flanagan, J.M. (1997) The struc-ture of ClpP at 2.3 Å resolution suggests a model forATP-dependent proteolysis. Cell 91: 447–456.
Weart, R.B., Nakano, S., Lane, B.E., Zuber, P., and Levin,P.A. (2005) The ClpX chaperone modulates assembly ofthe tubulin-like protein FtsZ. Mol Microbiol 57: 238–249.
Wiegert, T., and Schumann, W. (2001) SsrA-mediatedtagging in Bacillus subtilis. J Bacteriol 183: 3885–3889.
Wojtyra, U.A., Thibault, G., Tuite, A., and Houry, W.A. (2003)The N-terminal zinc binding domain of ClpX is a dimeriza-tion domain that modulates the chaperone function. J BiolChem 278: 48981–48990.
Zellmeier, S., Schumann, W., and Wiegert, T. (2006) Involve-ment of Clp protease activity in modulating the Bacillussubtilis sigma stress response. Mol Microbiol 61: 1569–1582.
Zolkiewski, M. (2006) A camel passes through the eye of aneedle: protein unfolding activity of Clp ATPases. MolMicrobiol 61: 1094–1100.
Zuber, P. (2004) Spx–RNA polymerase interaction and globaltranscriptional control during oxidative stress. J Bacteriol186: 1911–1918.
Clp ATPases and ClpP proteolytic complexes 1295
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 63, 1285–1295




















