
The effects of a synthetic curcuminoid analogue,...
9
Immunopharmacology and Inflammation The effects of a synthetic curcuminoid analogue, 2,6-bis-(4-hydroxyl-3-methoxybenzylidine)cyclohexanone on proinflammatory signaling pathways and CLP-induced lethal sepsis in mice Chau Ling Tham a , Kok Wai Lam b , Revathee Rajajendram a , Yoke Kqueen Cheah a , Mohd Roslan Sulaiman a,b , Nordin H. Lajis b,c , Min Kyu Kim b,d, ⁎, Daud A. Israf a,b, ⁎ a Department of Biomedical Science, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia b Laboratory of Natural Products, Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia c Department of Chemistry, Faculty of Science, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia d National Research Foundation of Korea, 137-748 Seoul, Republic of Korea abstract article info Article history: Received 28 July 2010 Received in revised form 12 October 2010 Accepted 31 October 2010 Available online 30 November 2010 Keywords: BHMC Curcumin MAPK NF-κB Monocyte Sepsis We previously showed that 2,6-bis-(4-hydroxyl-3-methoxybenzylidine)cyclohexanone (BHMC), suppressed the synthesis of various proinflammatory mediators. In this study we explain the mechanism of action of BHMC in lipopolysaccharide (LPS)-induced U937 monocytes and further show that BHMC prevents lethality of CLP-induced sepsis. BHMC showed dose-dependent inhibitory effects on p38, JNK and ERK 1/2 activity as determined by inhibition of phosphorylation of downstream transcription factors ATF-2, c-Jun and Elk-1 respectively. Inhibition of these transcription factors subsequently caused total abolishment of AP-1–DNA binding. BHMC inhibited p65 NF-κB nuclear translocation and DNA binding of p65 NF-κB only at the highest concentration used (12.5 μM) but failed to alter phosphorylation of JNK, ERK1/2 and STAT-1. Since the inhibition of p38 activity was more pronounced we evaluated the possibility that BHMC may bind to p38. Molecular docking experiments confirmed that BHMC fits well in the highly conserved hydrophobic pocket of p38 MAP kinase. We also show that BHMC was able to improve survival from lethal sepsis in a murine caecal-ligation and puncture (CLP) model. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Curcumin (1,7-bis-(4-hydroxy-3-methoxyphenyl)-1,6-hepa- diene,3,5,-dione) is one of the major components in the rhizome of turmeric and has been used over centuries in traditional remedies for the treatment of a variety of diseases, including inflammatory conditions (Ammon and Wahl, 1991). The ability of a single molecule like curcumin to exhibit a wide range of pharmacological effects against a variety of diseases remains an enigma under intense scrutiny (Alpers, 2006). Numerous molecular targets for curcumin have been identified over the years. Curcumin has been shown to down-regulate various transcription factors, such as nuclear factor (NF)-κB, activator protein (AP)-1, signal transducers and activators of transcription protein (STAT), specificity protein (SP)-1, protein kinases, growth factor receptors, chemokines, and inflammatory mediators such as interleukin (IL)-8, monocyte inflammatory protein (MIP)-1α, monocyte chemotactic protein (MCP)-1 (Abe et al., 1999), tumour necrosis factor (TNF)-α, IL-1, IL-6, cyclooxygenase (COX-2), inducible nitric oxide synthase (iNOS), and 5-lipoxygenase (5-LOX) (Alpers, 2006). It has been reported that curcumin blocks both the expression and signaling of proinflammatory cytokines through suppression of NF-κB nuclear translocation. Inhibition of NF-κB nuclear translocation is achieved by inhibition of IκBα phosphorylation via an inhibitory effect of curcumin on the IκB kinase (IKK) complex (Aggarwal, 2006; Bharti et al., 2003). It has been shown that the anti-inflammatory effects of curcumin are also mediated through its ability to down-regulate the activation of STAT1, STAT3, JAK1 and JAK 2 in microglia activated with gangliosides, LPS or IFN-γ (Kim et al., 2003). In inflammatory bowel disease, the signaling pathway leading to MAP kinase p38 activation was interrupted by curcumin (Salh et al., 2003). Curcumin also inhibits c-Fos transcription factor activation by down-regulation of extracellular-signal-related-kinase (ERK) and Jun N-terminal kinase (JNK) (Squires et al., 2003). The direct interaction of curcumin with AP-1 binding to its DNA binding motif results in the suppression of AP-1 as well as the inhibition on c-Jun and c-Fos, which are major components of the AP-1 assembly (Park et al., 1998). Previous studies have proven that the presence of bis-α, β-unsaturated β-diketone, two methoxy groups, two phenolic hydroxyl groups and two double-conjugated bonds to be critical in the anti-inflammatory activities assigned to curcumin (Sandur et al., 2007). BHMC was synthesized based European Journal of Pharmacology 652 (2011) 136–144 ⁎ Corresponding authors. Laboratory of Natural Products, Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia. Fax: + 60 3 8947 2337. E-mail addresses: [email protected] (M.K. Kim), [email protected] (D.A. Israf). 0014-2999/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2010.10.092 Contents lists available at ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar
Transcript of The effects of a synthetic curcuminoid analogue,...

Immunopharmacology and Inflammation
The effects of a synthetic curcuminoid analogue,2,6-bis-(4-hydroxyl-3-methoxybenzylidine)cyclohexanone on proinflammatorysignaling pathways and CLP-induced lethal sepsis in mice
Chau Ling Tham a, Kok Wai Lam b, Revathee Rajajendram a, Yoke Kqueen Cheah a, Mohd Roslan Sulaiman a,b,Nordin H. Lajis b,c, Min Kyu Kim b,d,⁎, Daud A. Israf a,b,⁎a Department of Biomedical Science, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysiab Laboratory of Natural Products, Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysiac Department of Chemistry, Faculty of Science, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysiad National Research Foundation of Korea, 137-748 Seoul, Republic of Korea
a b s t r a c ta r t i c l e i n f o
Article history:Received 28 July 2010Received in revised form 12 October 2010Accepted 31 October 2010Available online 30 November 2010
Keywords:BHMCCurcuminMAPKNF-κBMonocyteSepsis
We previously showed that 2,6-bis-(4-hydroxyl-3-methoxybenzylidine)cyclohexanone (BHMC), suppressedthe synthesis of various proinflammatory mediators. In this study we explain the mechanism of action ofBHMC in lipopolysaccharide (LPS)-induced U937 monocytes and further show that BHMC prevents lethalityof CLP-induced sepsis. BHMC showed dose-dependent inhibitory effects on p38, JNK and ERK 1/2 activity asdetermined by inhibition of phosphorylation of downstream transcription factors ATF-2, c-Jun and Elk-1respectively. Inhibition of these transcription factors subsequently caused total abolishment of AP-1–DNAbinding. BHMC inhibited p65 NF-κB nuclear translocation and DNA binding of p65 NF-κB only at the highestconcentration used (12.5 μM) but failed to alter phosphorylation of JNK, ERK1/2 and STAT-1. Since theinhibition of p38 activity was more pronounced we evaluated the possibility that BHMC may bind to p38.Molecular docking experiments confirmed that BHMC fits well in the highly conserved hydrophobic pocket ofp38 MAP kinase. We also show that BHMC was able to improve survival from lethal sepsis in a murinecaecal-ligation and puncture (CLP) model.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Curcumin (1,7-bis-(4-hydroxy-3-methoxyphenyl)-1,6-hepa-diene,3,5,-dione) is one of the major components in the rhizome ofturmeric and has been used over centuries in traditional remedies forthe treatment of a variety of diseases, including inflammatoryconditions (Ammon and Wahl, 1991). The ability of a single moleculelike curcumin to exhibit a wide range of pharmacological effectsagainst a variety of diseases remains an enigma under intense scrutiny(Alpers, 2006). Numerous molecular targets for curcumin have beenidentified over the years. Curcumin has been shown to down-regulatevarious transcription factors, such as nuclear factor (NF)-κB, activatorprotein (AP)-1, signal transducers and activators of transcription protein(STAT), specificityprotein(SP)-1,proteinkinases, growth factor receptors,chemokines, and inflammatory mediators such as interleukin (IL)-8,monocyte inflammatory protein (MIP)-1α, monocyte chemotacticprotein (MCP)-1 (Abe et al., 1999), tumour necrosis factor (TNF)-α,
IL-1, IL-6, cyclooxygenase (COX-2), induciblenitric oxide synthase (iNOS),and 5-lipoxygenase (5-LOX) (Alpers, 2006). It has been reported thatcurcumin blocks both the expression and signaling of proinflammatorycytokines through suppression of NF-κB nuclear translocation. Inhibitionof NF-κB nuclear translocation is achieved by inhibition of IκBαphosphorylation via an inhibitory effect of curcumin on the IκB kinase(IKK) complex (Aggarwal, 2006; Bharti et al., 2003). It has been shownthat the anti-inflammatory effects of curcumin are alsomediated throughits ability to down-regulate the activation of STAT1, STAT3, JAK1 and JAK2inmicroglia activatedwith gangliosides, LPS or IFN-γ (Kimet al., 2003). Ininflammatoryboweldisease, the signalingpathway leading toMAPkinasep38 activation was interrupted by curcumin (Salh et al., 2003). Curcuminalso inhibits c-Fos transcription factor activation by down-regulation ofextracellular-signal-related-kinase (ERK)and JunN-terminal kinase (JNK)(Squires et al., 2003). Thedirect interactionof curcuminwithAP-1bindingto its DNA binding motif results in the suppression of AP-1 as well as theinhibition on c-Jun and c-Fos, which are major components of the AP-1assembly (Park et al., 1998).
Previous studies haveproven that thepresence of bis-α,β-unsaturatedβ-diketone, twomethoxy groups, two phenolic hydroxyl groups and twodouble-conjugated bonds to be critical in the anti-inflammatory activitiesassigned to curcumin (Sandur et al., 2007). BHMCwas synthesized based
European Journal of Pharmacology 652 (2011) 136–144
⁎ Corresponding authors. Laboratory of Natural Products, Institute of Bioscience,Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia. Fax: +60 3 8947 2337.
E-mail addresses: [email protected] (M.K. Kim), [email protected](D.A. Israf).
0014-2999/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2010.10.092
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar

on the structure of curcumin by eliminating the unstable β-diketonemoiety and modifying it into conjugated double bonds while preservingthe phenolic OH group (Fig. 1). Based on our previous studies, we foundthat in comparison to curcumin, BHMC is more selective in thesuppression of various inflammatory mediators, such as monocytechemotactic protein (MCP)-1, inducible nitric oxide synthase (iNOS),IL-10, TNF-α, and IL-6. It alsodiffers fromcurcuminas it doesnot affect thesynthesis of prostaglandin E2 (PGE2), IL-1β, and IL-8 (Tham et al., 2010).We believe that differences in structural and functional characteristics ofBHMC may be due to more selective inhibitory effects uponproinflammatory signaling pathways. In this study, the effects of BHMCon proinflammatory signaling pathways were explored to provide moreaccurate information regarding the precise target molecule involved.Furthermore, a preliminary attempt was made to study the dose-dependent effect of BHMC in terms of protection from lethal sepsis in amurine caecal-ligation puncture model of severe sepsis.
2. Materials and methods
2.1. Reagents
Antibiotics (5000 U/ml penicillin and 5000 μg/ml streptomycin),Amphotericin B (250 μg/ml), Fetal bovine serum (FBS) andRoswell ParkMemorial Institute medium (RPMI)-1640 were purchased fromHyclone (Utah, USA). p44/42 MAP kinase (#9820), SAPK/JNK (#9810),p38 MAP kinase (#9800) assay kits, rabbit polyclonal antibody specificfor SAPK/JNK, p-SAPK/JNK, STAT1, p-STAT1, and rabbit anti-p-IκBαmonoclonal antibody were purchased from Cell Signaling Technology(Danvers, MA, USA). Rabbit polyclonal antibody specific for p38, p-p38,ERK 1, ERK2, p-ERK 1/2, IκBα, NFκB p65, TFIIB, HRP-conjugated donkeyanti-rabbit Ig-G, HRP-conjugated mouse anti-β-actin, NF-κB and AP-1oligonucleotideswere purchased from Santa Cruz Biotechnology (SantaCruz, CA, USA). All other reagents for Western blot analysis, bovineserum albumin (BSA) and dimethyl sulfoxide (DMSO) were purchasedfrom Amresco (Solon, Ohio). The polyvinylidene fluoride (PVDF)membrane was purchased from Milipore (Bredford, MA). Super SignalWest Femto Maximum Sensitivity Substrate, Bicinchoninic acid (BCA)protein determination kit (CatNo. 23225), Lightshift ChemiluminescentEMSA kit (Cat No. 20148), Biodyne precut nylonmembranes, and Biotin3′ End DNA Labeling Kit (Cat No. 89818) were purchased from Pierce(Rockford, IL). Phorbol-12-Myristate-13-Acetate (PMA), dexametha-sone, E. coli lipopolysaccharide (LPS) 055:B5 and curcumin werepurchased from Sigma Chemical co. (St Louis, MO). 2′-Amino-3′methoxyflavone (PD98059), 4-(4-Fluorophenyl)-2-(4-methylsulfinylphenyl)-5-(4-pyridyl) 1H-imidazole (SB203580), Anthra(1,9-cd)pyra-zol-6(2H)-one (SP600125), phosphatase inhibitor cocktail set III (CatNo. 524627) were purchased from Calbiohem (San Diego, CA). Proteinladder was purchased from Fermentas (Glen Burnie, MD). Benzonase
nuclease and NucBuster Protein Extraction Kit (Cat No. 71183-3) werepurchased fromNovagen (SanDiego, CA). BHMCwas synthesized in theLaboratory of Natural Products from 4-hydroxy-3-methoxybenzalde-hyde (vanillin or vanillaldehyde) and cyclohexanone based on thepublished procedure (Tham et al., 2010). For in vitro studies, BHMC,curcumin (Cur), dexamethasone (Dex), SB203580, PD98056 andSP600125 were dissolved in 100% DMSO as a stock and diluted toappropriate concentrations for assay as described in Materials andmethods and Results sections. The final concentration of DMSO wasmaintained at 0.1%.
2.2. Cell culture
U937 cells were purchased from the American Type CultureCollection (ATCC). Cells were cultured in RPMI-1640 supplementedwith 10% FBS, 4.5 g/l glucose, sodium pyruvate (1 mmol/l), L-glutamine(2 mmol/l), amphotericin B, streptomycin (50 μg/ml) and penicillin(50 U/ml). Cells were split when cell confluency reached 80–90%.
2.3. Phenotypic differentiation of U937 monocyte into macrophage
U937 cells at logarithmic phase of growthwereharvested, adjusted to2!106 cells/ml in the presence of 10 nM PMA, and incubated at 37 °C in5% CO2 incubator for 24 h to differentiate into adherent macrophages.After 24 h incubation, cells were washed twice with phosphate bufferedsaline (PBS) to remove non-adherent cells. Cell counts were performedeach time prior to and following the cell differentiation process to ensureconsistency. The variation in the differentiation process was minimalwithmore than 90% of differentiated cells remaining attached and viable.Remaining adherent cells were quiesced in fresh RPMI medium in theabsence of PMA for 24 h before stimulation with 10 μg/ml of LPS.
2.4. Preparation of whole cell lysates
Media was discarded following treatments and cells were rinsedtwice with ice-cold PBS. The cells were lysed in 500 μl of sample lysisbuffer (125 mM, 4% SDS, 20% glycerol, 0.004% bromophenol blue)with phosphatase inhibitor cocktail (Calbiochem, CA). Benzonasenuclease (25 U/ml) was added to digest nucleic acid. Followingincubation at room temperature for 15 min, the cells were scraped outand boiled at 100 °C for 10 min to denature protein and unfold DNA.Lysates were cooled in ice and centrifuged at 25,150!g, 4 °C, for15 min. The supernatant was collected and protein content wasmeasured using the BCA assay kit (Pierce, USA). The lysates werestored at !80 °C prior to analysis.
2.5. Preparation of cytosolic and nuclear extracts
This procedure was carried out by following Protocol # 71183-3 ofthe NucBuster Protein Extraction Kit (Novagen, CA). Treated U937cells were collected and rinsed centrifugally with ice-cold PBS(80 mM Na2HPO4, 100 mM NaH2PO4, pH 7.5). The cell pellet wasresuspended in NucBuster Reagent 1 (supplied with kit) and vortexedat high speed twice on ice. The cell suspensionwas then centrifuged at16,000!g, 4 °C for 5 min, and the supernatant was stored as thecytosolic fraction. The remaining pellet containing nuclei was lysedwith NucBuster Reagent 2 containing protease inhibitor cocktail andDTT (supplied with kit) followed by high speed vortexing on ice. Thesuspension was then centrifuged at 16,000!g, 4 °C for 5 min and thesupernatant (nuclear extract) collected was stored at !80 °C prior toanalysis. Protein content was quantified with the BCA assay kit(Pierce, USA).
Fig. 1. Chemical structure of (A) BHMC [2,6-bis-(4-hydroxyl-3-methoxybenzylidine)cyclohexanone] and (B) curcumin [1,7-bis-(4-hydroxy-3-methoxyphenyl)-1,6-hepa-diene,3,5,-dione].
137C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144

2.6. Western blot analysis
Differentiated U937 cells (2!106 cells/ml) were induced with10 μg/ml LPS as described earlier and treatedwith various concentrationsof BHMC for 30 min or 1 h. For the detection of p65NF-κB, IκBα, p-IκBα,p38, p-p38, STAT, and p-STAT, the cells were treated for 1 h. As for JNK,p-JNK, ERK and p-ERK, the cells were only treated for 30 min. Whole celllysates were used to analyze IκBα, p-I-κBα, p38, p-p38, JNK, p-JNK, ERK,p-ERK, STAT, p-STAT proteins, while both cytosolic and nuclear extractswere used for p65NF-κB analysis. Equal amounts of protein (20–50 μg)were electrophoresed on a 7%, 10% and 12% SDS-polyacrylamide geldepending on the proteinmolecular weight. Proteins were blotted onto aPVDFmembranewithaTrans-Blot SDSemiDryTransferCell (BioRad, CA).The membrane was blocked with 5–7% BSA in Tris buffered saline(TBS)-Tween 20 (0.05%) for 1 h and incubated overnight at 4 °C withrabbit polyclonal antibody specific for p38 (1:500), p-p38 (1:500), JNK(1:500), p-JNK (1:500), ERK1 (1:1000), ERK 2 (1:1000), p-ERK (1:500),STAT (1:500), p-STAT (1:500), p65NF-κB (1:500), IκBα (1:500) andp-IκBα (1:1000) in TBS-Tween containing 5% BSA. After washing threetimes with TBS-Tween, the membrane was hybridized with HRP-conjugated donkey anti-rabbit secondary antibody (1:2500–5000) for2 h and washed three times with TBS-Tween. The same membrane wasstripped and re-probed with HRP-conjugated mouse anti-mouse β-actin(1:10,000) or rabbit anti-TFIIB polyclonal antibody (1:1000). Themembrane was incubated with Super Signal West Femto MaximumSensitivity Substrate Reagent (Pierce, Rockford, IL) and viewed underchemiluminescence by a CCD camera imaging system (Vilber Lourmet,Marne-la-Vallee, France). Band intensities were quantified by BioProfil-Bio-1D software and normalized by comparison to β-actin or TFIIB.
2.7. Immunoprecipitation and in vitro kinase activity assays
This procedure was carried out by following Protocol # 9800, 9810and 9820 by Cell Signaling Technology (Danvers, MA, USA). U937macrophages (2!106 cells/ml) were induced with 10 μg/ml LPS asdescribed earlier and treated with various concentrations of BHMC.For the detection of p38MAP kinase activity, the cells were treated for1 h. As for SAPK/JNK and p44/42 MAP kinase activity, the cells wereonly treated for 30 min. Cells were rapidly washed with PBS, thenlysed with ice-cold lysis buffer (supplied with kit) and the p38 MAPK,SAPK/JNK, and p44/42 MAPK proteins immunoprecipitated. 200 μg oftotal cell extract was incubated overnight at 4 °C with 20 μl ofimmobilized antibody bead slurry. The beads were then washed threetimes with lysis buffer and incubated for 30 min at 30 °C in 50 μl ofkinase buffer (supplied with kit) containing 200 μM of ATP and kinasesubstrate. The reaction was terminated by the addition of samplebuffer and subjected to 12% SDS-PAGE. The membrane was incubatedwith Super Signal West Femto Maximum Sensitivity SubstrateReagent (Pierce, Rockford, IL). Bands of phosphorylated ATF-2,Elk-1, and c-Jun were then visualized under chemiluminescence by aCCD camera imaging system (Vilber Lourmet, Marne-la-Vallee, France).
2.8. Electrophoretic mobility shift assay (EMSA)
Control cells or cells treated with various concentrations of BHMCwere treated with 10 μg/ml LPS for 1 h, then nuclear extracts wereprepared as described previously by usingNucBuster Protein ExtractionKit. Oligonucleotides corresponding to thedownstreamNF-κB (5′-AGTTGAGGGGACTTTCCCAGGC-3′) and AP-1 (5′-CGCTTGATGACTCAGCCGGAA-3′) binding sequences were synthesized, annealed and end-labeledwith Biotin 3′ End DNA Labeling Kit. EMSA was performed as describedin the protocol of Lighshift Chemiluminescent EMSA Kit. Briefly, 20 μgnuclear extracts prepared as indicated above were incubated with100 fmol of oligonucleotides for 10 min at room temperature andseparated on a 5% native polyacrylamide gel. The specificity of bindingwas examined by competition with unlabeled oligonucleotides. The
nylon membrane was visualized, and chemiluminescent bands werequantified by a CCD camera imaging system (Vilber Lourmet,Marne-la-Vallee, France) using BioProfil-Bio-1D software.
2.9. Molecular docking experiments
The binding pattern of the BHMCmolecule in the active site of p38αMAP kinasewas studied using the protocol of AUTODOCK 4.2. The BHMCligand was built with hydrogen atoms and optimized throughMMFF94sforce field calculation. The crystal structure of the p38:SB203580complex (1A9U) was obtained from the protein data bank (Wanget al., 1997). Solventmoleculeswere deletedprior to the additionof polarhydrogen atoms and Kollman united atomic charges assigned. The gridpoints were set as 40!40!40 with the spacing value at 0.375 on thecatalytic site of the 1A9Uwhile the grid centerwas placed at 3.708, 15.85,and 28.973. The validity of the docking was justified from the redockingof the crystal ligand into the active site. Water molecules were removed.Ligand docking was performed using the AUTODOCK 4.2 LamarckianGenetic Algorithm (LGA). The free binding energieswere generated fromthe empirical terms multiplied by coefficients determined by linearregression analysis of complexes with known 3D-structures and knownbinding free energies (Morris et al., 1998). Other docking parameterswere in default mode during the course of docking.
2.10. Animals
Eight week-old male ICR mice were housed in a temperature-controlled roomwith food and water provided ad libitum and were keptunder a 12-h light/dark cycle diurnal cycle. The experiments describedbelowwere performed in accordancewith the current guidelines for thecare of laboratory animals and the ethical guidelines for animalexperimentation as approved by the Animal Experimentation EthicsCommittee, Faculty of Medicine and Health Sciences, Universiti PutraMalaysia.
2.11. Treatment groups and surgery
Two experiments were conducted; first of which evaluated severaldoses of BHMC todetermine the optimal dose for protection fromdeath.The second experiment compared the optimal dose of BHMC againsttwo doses of curcumin. Mice were randomly divided into groups of 12animals each. Sepsis was induced via the caecal ligature puncture (CLP)procedure as previously described (Rittirsch et al., 2009). Sham controlswere subjected to surgery involving caecal manipulation withoutligation and puncture. All other groups received treatment 1 h prior toCLP surgery. A vehicle control groupwas treatedwith an intraperitonealdose (10 ml/kg) of vehicle (5% ethanol, 5% Tween-20 and 90% distilledwater). Mice in other groups received BHMC (0.5, 1, 2 and 10 mg/kg)and curcumin (1 and 10mg/kg) dissolved in the vehicle via theintraperitoneal route. After 1 h post-treatment, the mice wereanaesthetized with an intraperitoneal injection of avertin (10 mg/kg)followed by the CLP surgical procedure. Following surgery mice weremonitored and death recorded every 6 h for 7 days.
2.12. Statistical analysis
Data is presented as means±S.E.M of at least three separateexperiments. Significant differences between treatments weredetermined by one-way analysis of variance (ANOVA) followed by posthoc comparisonusingDunnett test. TheChi-square exact test and Logranktest were used to analyze the differences in mouse mortality amonggroups. P-values of less than 0.05 were considered significant.
138 C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144
3. Results
3.1. BHMC mildly inhibits LPS-stimulated I!B" phosphorylation
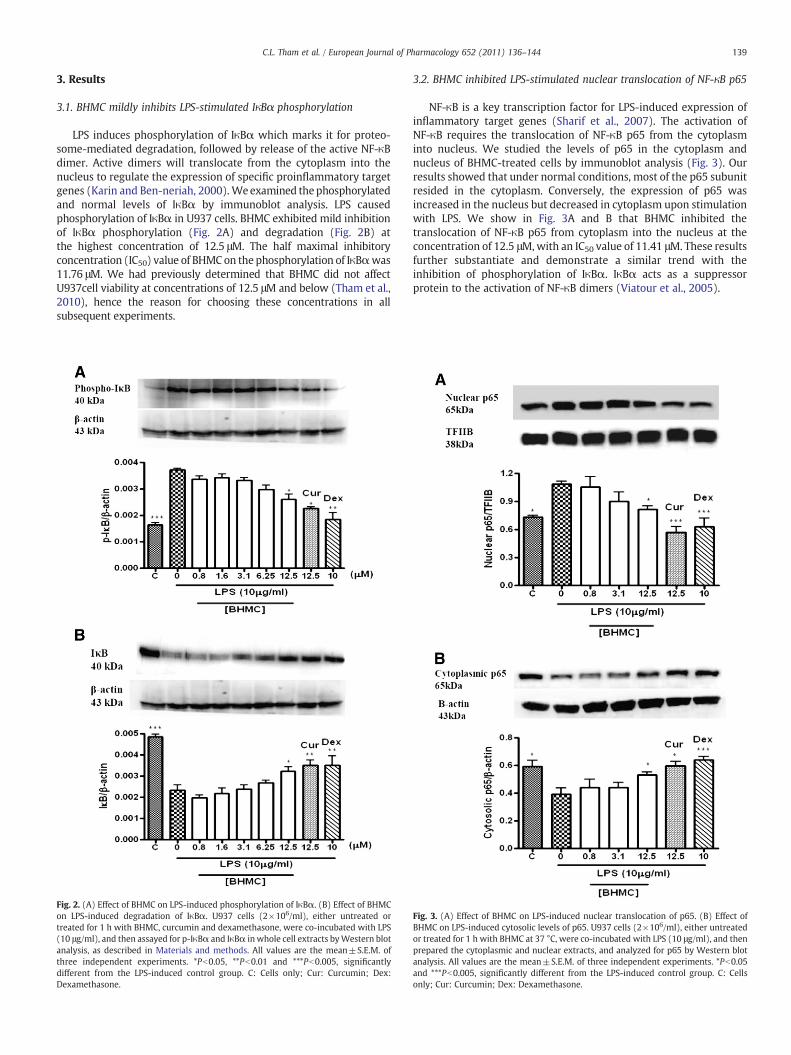
LPS induces phosphorylation of IκBα which marks it for proteo-some-mediated degradation, followed by release of the active NF-κBdimer. Active dimers will translocate from the cytoplasm into thenucleus to regulate the expression of specific proinflammatory targetgenes (Karin and Ben-neriah, 2000). Weexamined thephosphorylatedand normal levels of IκBα by immunoblot analysis. LPS causedphosphorylation of IκBα in U937 cells. BHMC exhibited mild inhibitionof IκBα phosphorylation (Fig. 2A) and degradation (Fig. 2B) atthe highest concentration of 12.5 μM. The half maximal inhibitoryconcentration (IC50) value of BHMC on the phosphorylation of IκBαwas11.76 μM. We had previously determined that BHMC did not affectU937cell viability at concentrations of 12.5 μM and below (Tham et al.,2010), hence the reason for choosing these concentrations in allsubsequent experiments.
3.2. BHMC inhibited LPS-stimulated nuclear translocation of NF-!B p65
NF-κB is a key transcription factor for LPS-induced expression ofinflammatory target genes (Sharif et al., 2007). The activation ofNF-κB requires the translocation of NF-κB p65 from the cytoplasminto nucleus. We studied the levels of p65 in the cytoplasm andnucleus of BHMC-treated cells by immunoblot analysis (Fig. 3). Ourresults showed that under normal conditions, most of the p65 subunitresided in the cytoplasm. Conversely, the expression of p65 wasincreased in the nucleus but decreased in cytoplasm upon stimulationwith LPS. We show in Fig. 3A and B that BHMC inhibited thetranslocation of NF-κB p65 from cytoplasm into the nucleus at theconcentration of 12.5 μM,with an IC50 value of 11.41 μM. These resultsfurther substantiate and demonstrate a similar trend with theinhibition of phosphorylation of IκBα. IκBα acts as a suppressorprotein to the activation of NF-κB dimers (Viatour et al., 2005).
Fig. 2. (A) Effect of BHMC on LPS-induced phosphorylation of IκBα. (B) Effect of BHMCon LPS-induced degradation of IκBα. U937 cells (2!106/ml), either untreated ortreated for 1 h with BHMC, curcumin and dexamethasone, were co-incubated with LPS(10 μg/ml), and then assayed for p-IκBα and IκBα in whole cell extracts byWestern blotanalysis, as described in Materials and methods. All values are the mean±S.E.M. ofthree independent experiments. *Pb0.05, **Pb0.01 and ***Pb0.005, significantlydifferent from the LPS-induced control group. C: Cells only; Cur: Curcumin; Dex:Dexamethasone.
Fig. 3. (A) Effect of BHMC on LPS-induced nuclear translocation of p65. (B) Effect ofBHMC on LPS-induced cytosolic levels of p65. U937 cells (2!106/ml), either untreatedor treated for 1 h with BHMC at 37 °C, were co-incubated with LPS (10 μg/ml), and thenprepared the cytoplasmic and nuclear extracts, and analyzed for p65 by Western blotanalysis. All values are the mean±S.E.M. of three independent experiments. *Pb0.05and ***Pb0.005, significantly different from the LPS-induced control group. C: Cellsonly; Cur: Curcumin; Dex: Dexamethasone.
139C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144
3.3. BHMC does not affect phosphorylation of MAPKs
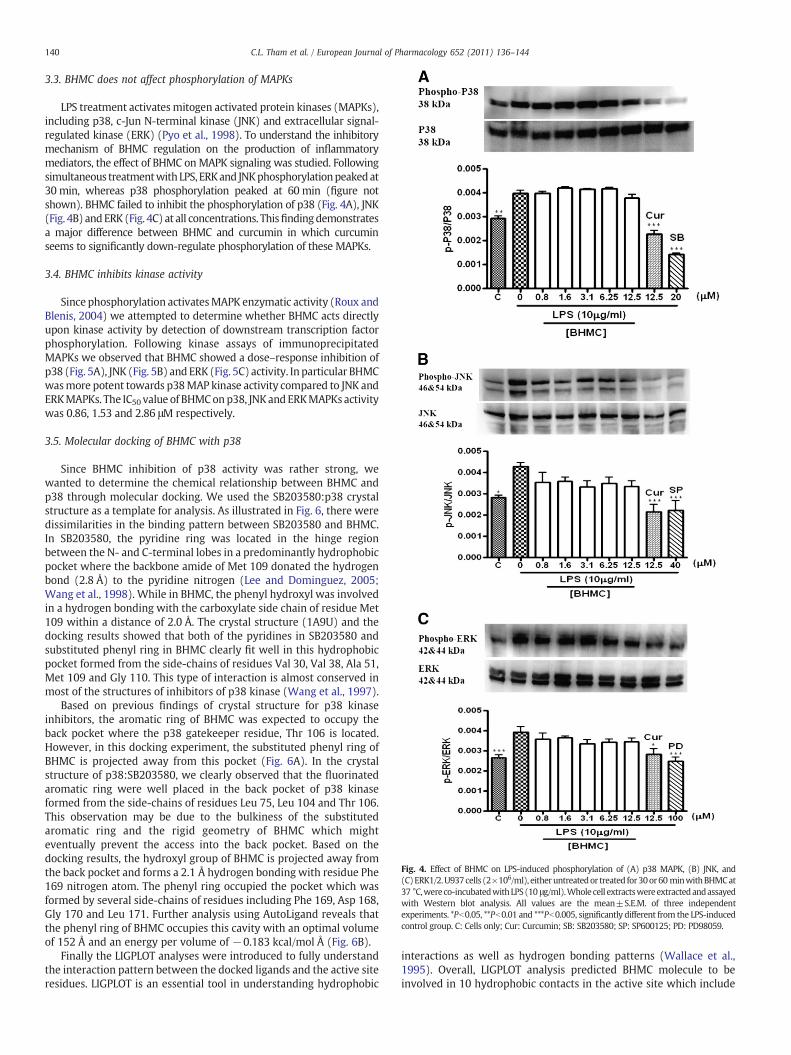
LPS treatment activatesmitogen activated protein kinases (MAPKs),including p38, c-Jun N-terminal kinase (JNK) and extracellular signal-regulated kinase (ERK) (Pyo et al., 1998). To understand the inhibitorymechanism of BHMC regulation on the production of inflammatorymediators, the effect of BHMC onMAPK signaling was studied. Followingsimultaneous treatmentwithLPS, ERKand JNKphosphorylationpeakedat30 min, whereas p38 phosphorylation peaked at 60 min (figure notshown). BHMC failed to inhibit the phosphorylation of p38 (Fig. 4A), JNK(Fig. 4B) andERK (Fig. 4C) at all concentrations. Thisfindingdemonstratesa major difference between BHMC and curcumin in which curcuminseems to significantly down-regulate phosphorylation of these MAPKs.
3.4. BHMC inhibits kinase activity
Since phosphorylation activatesMAPK enzymatic activity (Roux andBlenis, 2004) we attempted to determine whether BHMC acts directlyupon kinase activity by detection of downstream transcription factorphosphorylation. Following kinase assays of immunoprecipitatedMAPKs we observed that BHMC showed a dose–response inhibition ofp38 (Fig. 5A), JNK (Fig. 5B) andERK (Fig. 5C) activity. In particular BHMCwasmore potent towards p38MAP kinase activity compared to JNK andERKMAPKs. The IC50 value of BHMConp38, JNKandERKMAPKsactivitywas 0.86, 1.53 and 2.86 μM respectively.
3.5. Molecular docking of BHMC with p38
Since BHMC inhibition of p38 activity was rather strong, wewanted to determine the chemical relationship between BHMC andp38 through molecular docking. We used the SB203580:p38 crystalstructure as a template for analysis. As illustrated in Fig. 6, there weredissimilarities in the binding pattern between SB203580 and BHMC.In SB203580, the pyridine ring was located in the hinge regionbetween the N- and C-terminal lobes in a predominantly hydrophobicpocket where the backbone amide of Met 109 donated the hydrogenbond (2.8 Å) to the pyridine nitrogen (Lee and Dominguez, 2005;Wang et al., 1998). While in BHMC, the phenyl hydroxyl was involvedin a hydrogen bonding with the carboxylate side chain of residue Met109 within a distance of 2.0 Å. The crystal structure (1A9U) and thedocking results showed that both of the pyridines in SB203580 andsubstituted phenyl ring in BHMC clearly fit well in this hydrophobicpocket formed from the side-chains of residues Val 30, Val 38, Ala 51,Met 109 and Gly 110. This type of interaction is almost conserved inmost of the structures of inhibitors of p38 kinase (Wang et al., 1997).
Based on previous findings of crystal structure for p38 kinaseinhibitors, the aromatic ring of BHMC was expected to occupy theback pocket where the p38 gatekeeper residue, Thr 106 is located.However, in this docking experiment, the substituted phenyl ring ofBHMC is projected away from this pocket (Fig. 6A). In the crystalstructure of p38:SB203580, we clearly observed that the fluorinatedaromatic ring were well placed in the back pocket of p38 kinaseformed from the side-chains of residues Leu 75, Leu 104 and Thr 106.This observation may be due to the bulkiness of the substitutedaromatic ring and the rigid geometry of BHMC which mighteventually prevent the access into the back pocket. Based on thedocking results, the hydroxyl group of BHMC is projected away fromthe back pocket and forms a 2.1 Å hydrogen bonding with residue Phe169 nitrogen atom. The phenyl ring occupied the pocket which wasformed by several side-chains of residues including Phe 169, Asp 168,Gly 170 and Leu 171. Further analysis using AutoLigand reveals thatthe phenyl ring of BHMC occupies this cavity with an optimal volumeof 152 Å and an energy per volume of !0.183 kcal/mol Å (Fig. 6B).
Finally the LIGPLOT analyses were introduced to fully understandthe interaction pattern between the docked ligands and the active siteresidues. LIGPLOT is an essential tool in understanding hydrophobic
interactions as well as hydrogen bonding patterns (Wallace et al.,1995). Overall, LIGPLOT analysis predicted BHMC molecule to beinvolved in 10 hydrophobic contacts in the active site which include
Fig. 4. Effect of BHMC on LPS-induced phosphorylation of (A) p38 MAPK, (B) JNK, and(C)ERK1/2.U937cells (2!106/ml), eitheruntreatedor treated for30or60minwithBHMCat37 °C,wereco-incubatedwithLPS(10 μg/ml).Wholecell extractswereextractedandassayedwith Western blot analysis. All values are the mean±S.E.M. of three independentexperiments. *Pb0.05, **Pb0.01 and ***Pb0.005, significantly different from the LPS-inducedcontrol group. C: Cells only; Cur: Curcumin; SB: SB203580; SP: SP600125; PD: PD98059.
140 C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144

interactions with residues Leu 108, Val 30, Ala 51, Val 38, Ile 84, Asp168, Leu 167 and Leu 75 (Fig. 6C). Interestingly, this analysis alsoshows that the cyclohexanone moiety of BHMC formed threehydrophobic contacts with nearby residues. However, the presenceof the ketone group did not reveal any hydrogen bonding interactionwithin the active site.
Fig. 5. Effect of BHMC on LPS-induced MAP kinase activity. (A) p38 MAP kinase activitywas determined by immunoprecipitation studies using an ATF-2 fusion protein assubstrate. (B) SAPK/JNK kinase activity was determined by immunoprecipitation studies.SAPK phosphorylated the c-Jun substrate upon addition of ATP. (C) p44/42 MAP kinaseactivity was determined by immunoprecipitation studies using an Elk-1 fusion protein assubstrate. Result shown is a representative blot from three independent experiments. Allvalues are the mean±S.E.M. of three independent experiments. *Pb0.05, **Pb0.01 and***Pb0.005, significantly different from the LPS-induced control group. C: Cells only; Cur:Curcumin; SB: SB203580; SP: SP600125; PD: PD98059. Fig. 6. (A) Bindingmode of BHMC into the binding site of p38MAP kinase (PDB code 1A9U).
Residues involved in the clashes and contacts with BHMC in the binding site with estimatedfree binding energy of !6.96 kcal/mol. Both the hydroxyl group of BHMC was involved inhydrogen bonding with residues MET 109(2.086 Å) and PHE 169(2.137 Å) respectively. Oneof the phenyl groups was clearly seen occupying the hinge region while the other ring wasfilling in another cavity at the back of the ATP-site. (B) Interatomic contacts (yellow) of theligand with the residues in active site based on Van der Waals radii generated by Chimerasoftware. (C) The 2D representation docking result of compound with the residues in theactive site of p38 MAP kinase (PDB code 1A9U) showed by LIGPLOT. The hydrogen bondinginteraction andhydrophobic contactswere represented by the greendashed lines and the arcwith spokes radiating towards the ligand atoms they contact respectively.
141C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144
3.6. BHMC inhibited NF-!B and AP-1 DNA binding activity
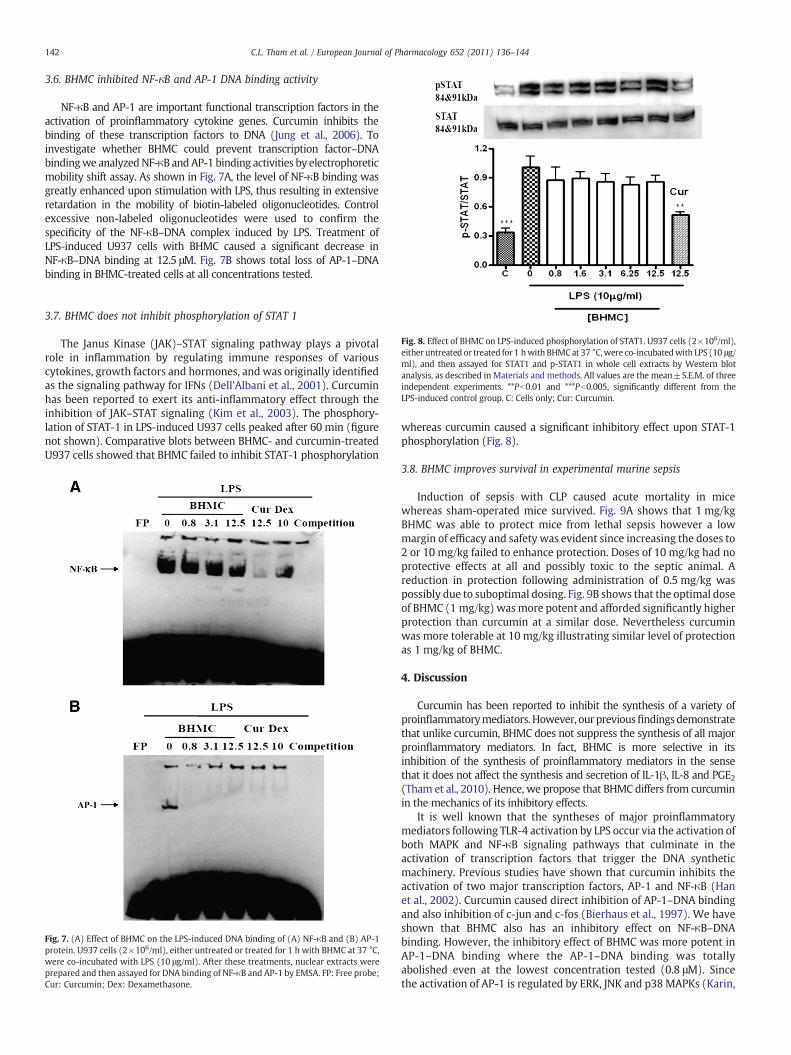
NF-κB and AP-1 are important functional transcription factors in theactivation of proinflammatory cytokine genes. Curcumin inhibits thebinding of these transcription factors to DNA (Jung et al., 2006). Toinvestigate whether BHMC could prevent transcription factor–DNAbindingwe analyzedNF-κB andAP-1 binding activities by electrophoreticmobility shift assay. As shown in Fig. 7A, the level of NF-κB binding wasgreatly enhanced upon stimulation with LPS, thus resulting in extensiveretardation in the mobility of biotin-labeled oligonucleotides. Controlexcessive non-labeled oligonucleotides were used to confirm thespecificity of the NF-κB–DNA complex induced by LPS. Treatment ofLPS-induced U937 cells with BHMC caused a significant decrease inNF-κB–DNA binding at 12.5 μM. Fig. 7B shows total loss of AP-1–DNAbinding in BHMC-treated cells at all concentrations tested.
3.7. BHMC does not inhibit phosphorylation of STAT 1
The Janus Kinase (JAK)–STAT signaling pathway plays a pivotalrole in inflammation by regulating immune responses of variouscytokines, growth factors and hormones, and was originally identifiedas the signaling pathway for IFNs (Dell'Albani et al., 2001). Curcuminhas been reported to exert its anti-inflammatory effect through theinhibition of JAK–STAT signaling (Kim et al., 2003). The phosphory-lation of STAT-1 in LPS-induced U937 cells peaked after 60 min (figurenot shown). Comparative blots between BHMC- and curcumin-treatedU937 cells showed that BHMC failed to inhibit STAT-1 phosphorylation
whereas curcumin caused a significant inhibitory effect upon STAT-1phosphorylation (Fig. 8).
3.8. BHMC improves survival in experimental murine sepsis
Induction of sepsis with CLP caused acute mortality in micewhereas sham-operated mice survived. Fig. 9A shows that 1 mg/kgBHMC was able to protect mice from lethal sepsis however a lowmargin of efficacy and safety was evident since increasing the doses to2 or 10 mg/kg failed to enhance protection. Doses of 10 mg/kg had noprotective effects at all and possibly toxic to the septic animal. Areduction in protection following administration of 0.5 mg/kg waspossibly due to suboptimal dosing. Fig. 9B shows that the optimal doseof BHMC (1 mg/kg) wasmore potent and afforded significantly higherprotection than curcumin at a similar dose. Nevertheless curcuminwas more tolerable at 10 mg/kg illustrating similar level of protectionas 1 mg/kg of BHMC.
4. Discussion
Curcumin has been reported to inhibit the synthesis of a variety ofproinflammatorymediators.However, ourpreviousfindingsdemonstratethat unlike curcumin, BHMC does not suppress the synthesis of all majorproinflammatory mediators. In fact, BHMC is more selective in itsinhibition of the synthesis of proinflammatory mediators in the sensethat it does not affect the synthesis and secretion of IL-1β, IL-8 and PGE2(Tham et al., 2010). Hence, we propose that BHMC differs from curcuminin the mechanics of its inhibitory effects.
It is well known that the syntheses of major proinflammatorymediators following TLR-4 activation by LPS occur via the activation ofboth MAPK and NF-κB signaling pathways that culminate in theactivation of transcription factors that trigger the DNA syntheticmachinery. Previous studies have shown that curcumin inhibits theactivation of two major transcription factors, AP-1 and NF-κB (Hanet al., 2002). Curcumin caused direct inhibition of AP-1–DNA bindingand also inhibition of c-jun and c-fos (Bierhaus et al., 1997). We haveshown that BHMC also has an inhibitory effect on NF-κB–DNAbinding. However, the inhibitory effect of BHMC was more potent inAP-1–DNA binding where the AP-1–DNA binding was totallyabolished even at the lowest concentration tested (0.8 μM). Sincethe activation of AP-1 is regulated by ERK, JNK and p38 MAPKs (Karin,
Fig. 7. (A) Effect of BHMC on the LPS-induced DNA binding of (A) NF-κB and (B) AP-1protein. U937 cells (2!106/ml), either untreated or treated for 1 h with BHMC at 37 °C,were co-incubated with LPS (10 μg/ml). After these treatments, nuclear extracts wereprepared and then assayed for DNA binding of NF-κB and AP-1 by EMSA. FP: Free probe;Cur: Curcumin; Dex: Dexamethasone.
Fig. 8. Effect of BHMC on LPS-induced phosphorylation of STAT1. U937 cells (2!106/ml),either untreated or treated for 1 hwith BHMCat 37 °C,were co-incubatedwith LPS (10 μg/ml), and then assayed for STAT1 and p-STAT1 in whole cell extracts by Western blotanalysis, as described in Materials and methods. All values are the mean±S.E.M. of threeindependent experiments. **Pb0.01 and ***Pb0.005, significantly different from theLPS-induced control group. C: Cells only; Cur: Curcumin.
142 C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144

1995), we suggest that the loss of AP-1–DNA binding of BHMC-treatedU937 is possibly due to the inhibitory effects upon MAPK activation,especially the particularly potent effect of BHMC upon p38 activity.
Curcumin has been shown to target phosphorylation of JNK, p38(Kim et al., 2006) and ERK (Cho et al., 2007) and upstream kinases(Chen and Tan, 1998). A major departure in the mechanism of actionof BHMC from that of curcumin is the specific effect upon p38, JNK,and ERK1/2 enzymatic activity without any inhibitory effect uponphosphorylation. We suggest that BHMC does not affect upstreamkinases such as MEK and the chemical modification of curcumin hasintroduced a degree of selectivity in which it directly targetsenzymatic activation especially on p38. Since BHMC demonstrated apotent effect on p38 activation we attempted to explain the molecularinteraction between these two molecules through docking studies.Our findings were supported by the data that showed the potentialbinding mode of BHMC in the p38 active site as illustrated in Fig. 6.Once p38 is activated by MAPKK, conformational changes occurfollowed by the binding of ATP to the ATP pocket which in turnrenders the kinase to phosphorylate and activate its substrate such asthe transcription factor ATF-2. The docking results showed that BHMCpotentially competes with ATP from binding to the ATP pocket in p38.Instead of occupying the back pocket of p38where the p38 gatekeeperresides, Thr 106 is located at the other phenyl ring of BHMC anddeviating far from this position due to its bulkiness and rigidgeometry. However, the binding pattern showed that BHMC clearlyfits well in the hydrophobic pocket consisting of residue Val 30, Val 38,Ala 51, Met 109 and Gly 110.We speculate a similar binding pattern inJNK and ERK1/2 since the ATP-pocket is highly conserved amongkinases. The relevant residues Met and Lys are generally conservedwhile the gatekeeper differs within these enzymes to contribute totheir substrate specificity (Peifer et al., 2007). Most synthetic drugs
that target p38 contain pyridinyl and pyrimidinyl moieties, which areinvolved in a direct competition with ATP binding. However, some ofthese inhibitors have failed in clinical trials due to target selectivityrelated toxicity and off-target effects (Angell et al., 2008).
Activation of the p38 signaling pathway plays a significant role inthe development of immune suppression seen in the septic host. It hasbeen previously reported that an inhibitor of p38 markedly improvessurvival rates when administered after the initial proinflammatorystage of polymicrobial sepsis (Song et al., 2001). Curcumin has beenshown to reduce muscle proteolysis in a rat CLP model throughinhibition of p38 and NF-κB activation (Poylin et al., 2008).Furthermore a recent study reported that mice injected with100 mg/kg of curcumin prior to LPS/GalN challenge exhibited asurvival rate of 65% in LPS/GalN-induced lethal sepsis (Yun et al.,2010). Since the major effect of BHMC is targeted towards inhibitionof p38, we compared the ability of this compound and curcumin atequivalent doses in their preventive effects upon lethality in a CLPmodel. Clearly the modification of curcumin into BHMC retained itsability to prevent death due to a severe systemic inflammatorysyndrome. Interestingly the protection afforded by 1 mg/kg was aseffective as that of curcumin at 10 mg/kg. However our preliminaryexperiments suggest that BHMC has a narrow margin of safety in theseptic host. It was evident that a 10 mg/kg dose had no significantprotective effect and this suggests toxicity. Our unpublished data onthe 24 h acute toxicity of BHMC showed that mice did not show anysigns of toxicity with doses up to 1 g/kg. Therefore we are of theopinion that BHMC could be toxic once normal metabolic processesare compromised by the severe renal and liver damage in the septicmouse. Whether the protective effect of BHMC is directly related top38 alone remains to be elucidated, however it is extremely possiblethat the enhanced inhibition of p38 activity may play a major role andtherefore BHMC is of considerable interest in the prophylaxis andtherapeutics of experimental sepsis.
In conclusion, the generation of BHMC through elimination ofunstable β-diketone moieties and modification into conjugateddouble bonds while preserving the phenolic OH group of thecurcumin molecule has induced a certain degree of selectivity in itstarget molecules, with p38 being the most significantly affectedthrough inactivation via competitive binding to the ATP pocket. BHMCis also potent in preventing death following lethal sepsis and this ispossibly due to its enhanced potency towards p38 inhibition that mayblock the synthesis of several proinflammatory mediators responsiblefor the severe systemic inflammatory syndrome observed in sepsis.Further work is needed to explain the prophylactic and therapeuticrole of BHMC in septic hosts.
Acknowledgements
We thank Norazren Ismail and Zulkhairi Zainol for excellenttechnical assistance and Ms Azra Elia Zamri for proofreading. Thisinvestigation was financially supported by the Fundamental ResearchGrant Scheme (FRGS 04-11-09-704FR/F1) and the Research UniversityGrant Scheme (RUGS 04-02-10-0922RU), Universiti Putra Malaysia.Chau Ling Tham, KokWai Lam and Revathee Rajajendramare recipientsof a UPM graduate research fellowship scheme.
References
Abe, Y., Hashimoto, S., Horie, T., 1999. Curcumin inhibition of inflammatory cytokineproduction by human peripheral blood monocytes and alveolar macrophages.Pharmacol. Res. 39, 41–47.
Aggarwal, S., 2006. Curcumin (diferuloylmethane) down-regulates expression of cellproliferation and anti-apoptotic andmetastatic gene products through suppressionof IκBα kinase and Akt activation. Mol. Pharmacol. 69, 195–206.
Alpers, D.H., 2006. The potential use of curcumin in management of chronic diseases:too good to be true? Curr. Opin. Gastroen. 24, 173–175.
Ammon, H.P., Wahl, M.A., 1991. Pharmacology of Curcuma longa. Planta Med. 57, 1–7.
Fig. 9. (A) Dose–response effects of BHMC on the survival rate in murine CLP-inducedsepsis. (B) Effect of curcumin and BHMCon the survival rate inmurine CLP-induced sepsis.Mice were randomly divided into groups of 12 animals each. Treatments wereadministered intraperitoneally 1 h prior to surgery. Survival was monitored over 7 days.*Pb0.05, **Pb0.01 and ***Pb0.005, significantly different from the vehicle control group.#Pb0.05, significantly different from the BHMC 1 mg/kg treatment group. Cur: Curcumin.
143C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144

Angell, R.M., Bamborough, P., Cleasby, A., Cockerill, S.G., Jones, K.L., Mooney, C.J.,Somers, D.O., Walker, A.L., 2008. Biphenyl amide p38 kinase inhibitor 1: discoveryand binding mode. Bioorg. Med. Chem. Lett. 18, 318–323.
Bharti, A.C., Donato, S., Singh, S., Aggarwal, B.B., 2003. Curcumin (diferuloylmethane)down-regulates the constitutive activation of nuclear factor-kappa B and IkappaBalpha kinase in human multiple myeloma cells, leading to suppression ofproliferation and induction of apoptosis. Blood 101, 1053–1062.
Bierhaus, A., Zhang, Y., Quehenberger, P., Luther, T., Haase, M., Muller, M., Mackman, N.,Ziegler, R., Nawroth, P.P., 1997. The dietary pigment curcumin reduces endothelialtissue factor gene expression by inhibiting binding of AP-1 to the DNA andactivation of NF-kappaB. Thromb. Heamostasis 77, 772–782.
Chen, Y.R., Tan, T.H., 1998. Inhibition of the c-Jun-N-terminal kinase (JNK) signallingpathway by curcumin. Oncogene 17, 173–178.
Cho, J.W., Lee, K.S., Kim, C.W., 2007. Curcumin attenuates the expression of IL-1β, IL-6,and TNF-α as well as cyclin E in TNF-α-treated HaCaT cells; NF-κB and MAPKs aspotential upstream targets. Int. J. Mol. Med. 19, 469–474.
Dell'Albani, P., Santangelo, R., Torrisi, L., Nicoletti, V.G., Vellis, J., Giuffrida Stella, A.M.,2001. JAK/STAT signaling pathway mediates cytokine induced iNOS expression inprimary astroglial cell cultures. J. Neurosci. Res. 65, 417.
Han, S.S., Keum, Y.S., Seo, H.J., Surh, Y.J., 2002. Curcumin suppresses activation of NF-κBand AP-1 induced by phorbol ester in cultured human promyelocytic leukemiacells. J. Biochem. Mol. Biol. 35, 337–342.
Jung, K.K., Lee, H.S., Cho, J.Y., Shin, W.C., Rhee, M.H., Kim, T.G., Kang, J.H., Kim, S.H., Hong,S., Kang, S.Y., 2006. The inhibitory effect of curcumin on nitric oxide productionfrom lipopolysaccharide-activated primary microglia. Life Sci. 79, 2022–2031.
Karin, M., 1995. The regulation of AP-1 activity by mitogen-activated protein kinases. J.Biol. Chem. 270, 16483–16486.
Karin, M., Ben-neriah, Y., 2000. Phosphorylation meets ubiquitination: the control ofNF-κB activity. Annu. Rev. Immunol. 18, 621–663.
Kim, H.Y., Park, E.J., Joe, E., Jou, I., 2003. Curcumin suppresses Janus Kinase–STATinflammatory signaling through activation of Src homology 2 domain-containingtyrosine phosphatase 2 in brain microglia. J. Immunol. 171, 6072–6079.
Kim, Y.S., Ahn, Y.K., Hong, M.H., Joo, S.Y., Kim, K.H., Sohn, I.S., Park, H.W., Hong, Y.J., Kim,J.H., Kim, W., Jeong, M.H., Cho, J.G., Park, J.C., Kang, J.C., 2006. Curcumin attenuatesNF-κB, c-jun N-terminal kinase and p38 in tumor necrosis factor-α-stimulatedendothelial cells. Korean Circ. J. 36, 482–489.
Lee, M., Dominguez, C., 2005. MAP kinase p38 inhibitors: clinical results and an intimatelook at their interactions with p38α protein. Curr. Med. Chem. 12, 2979–2994.
Morris, G.M., Goodsell, D.S., Halliday, R.S., Huey, R., Hart, W.E., Belew, R.K., Olson, A.J.,1998. Automated docking using a Lamarckian Genetic Algorithm and empiricalbinding free energy function. J. Comput. Chem. 19, 1639–1662.
Park, S., Lee, D.K., Yang, C.H., 1998. Inhibition of fos–jun–DNA complex formation bydihydroguaiaretic acid and in vitro cytotoxic effects on cancer cells. Cancer Lett.127, 23–28.
Peifer, C., Kinkel, K., Abadleh, M., Schollmeyer, D., Laufer, S.J., 2007. From five- to six-membered rings: 3, 4-diarylquinolinone as lead for novel p38MAP kinaseinhibitors. J. Med. Chem. 50, 1213–1221.
Poylin, V., Fareed, M.U., O'Neal, P., Alamdari, N., Reilly, N., Menconi, M., Hasselgren, P.O.,2008. The NF-κB inhibitor curcumin blocks sepsis-induced muscle proteolysis.Mediat. Inflamm. 317851, 1–13.
Pyo, H., Jou, I., Jung, S., Hong, S., Joe, E., 1998. Mitogen-activated protein kinases activatedby lipopolysaccharide and amyloid in cultured rat microglia. Neuro. Rep. 9, 871–874.
Rittirsch, D., Huber-Lang, M.S., Flierl, M.A., Ward, P.A., 2009. Immunodesign ofexperimental sepsis by cecal ligation and puncture. Nat. Protoc. 4, 31–36.
Roux, P.P., Blenis, J., 2004. ERK andp38MAPK-activatedprotein kinases: a family of proteinkinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 68, 320–344.
Salh, A., Assi, K., Templeman, V., Parhar, K., Owen, D., Gomez-Munoz, A., Jacobson, K.,2003. Curcumin attenuates DNB-induced murine colitis. Am. J. Physiol. Gastr. L.285, 235–243.
Sandur, S.K., Pandey, M.K., Sung, B., Ahn, K.S., Murakami, A., Sethi, G., Limtrakul, P.,Badmaev, V., Aggarwal, B.B., 2007. Curcumin, demethoxycurcumin, bisdemethox-ycurcumin, tetrahydrocurcumin and turmerones differentially regulate anti-inflammatory and anti-proliferative responses through a ROS-dependent mecha-nism. Carcinogenesis 28, 1765–1773.
Sharif, O., Bolshakov, V.N., Raines, S., Newham, P., Perkins, N.D., 2007. Transcriptionalprofiling of the LPS induced NF-κB response in macrophages. BMC Immunol. 8, 1.
Song, G.Y., Chung, C.S., Chaudry, I.H., Ayala, A., 2001. MAPK p38 antagonism as a novelmethod of inhibiting lymphoid immune suppression in polymicrobial sepsis. Am. J.Physiol. Cell Physiol. 281, 662–669.
Squires, M.S., Hudson, E.A., Howells, L., Sale, S., Houghton, C.E., Jones, J.L., 2003.Relevance of mitogen activated protein kinase (MAPK) and phosphotidylinositol-3-kinase/protein kinase B (PI3K/PKB) pathways to induction of apoptosis bycurcumin in breast cells. Biochem. Pharmacol. 65, 361–376.
Tham, C.L., Liew, C.Y., Lam, K.W., Mohamad, A.S., Kim, M.K., Cheah, Y.K., Zakaria, Z.A.,Sulaiman,M.R., Lajis, N.H., Israf,D.A., 2010. A synthetic curcuminoid derivative inhibitsnitric oxide and proinflammatory cytokine synthesis. Eur. J. Pharm. 628, 247–254.
Viatour, P., Merville, M.P., Bours, V., Chariot, A., 2005. Phosphorylation of NF-κB and IκBproteins: implications in cancer and inflammation. Trends Biochem. Sci. 30, 43–52.
Wallace, A.C., Laskowski, R.A., Thornton, J.M., 1995. LIGPLOT: a program to generateschematic diagrams of protein–ligand interactions. Protein Eng. 8, 127–134.
Wang, Z., Harkins, P.C., Ulevitch, R.J., Han, J., Cobb, M.H., Goldsmith, E.J., 1997. Thestructure of mitogen-activated protein kinase p38 at 2.1-A resolution. Proc. NatlAcad. Sci. USA 94, 2327–2332.
Wang, B., Canagarajah, J., Boehm, J., Kassisa, S., Cobb, M., Young, P., Abdel-Meguid, S.,Adams, J., Goldsmith, E., 1998. Structural basis of inhibitor selectivity in MAPkinases. Structure 6, 1117–1128.
Yun, S.S., Kim, S.P., Kang, M.Y., Nam, S.H., 2010. Inhibitory effect of curcumin on liverinjury in a murine model of endotoxemic shock. Biotechnol. Lett. 32, 209–214.
144 C.L. Tham et al. / European Journal of Pharmacology 652 (2011) 136–144




















