
Polyamines Interact with Hydroxyl Radicals in Activating Ca2+ and K+ Transport across the Root...
14
Polyamines Interact with Hydroxyl Radicals in Activating Ca 2+ and K + Transport across the Root Epidermal Plasma Membranes 1[W] Isaac Zepeda-Jazo 2,3 , Ana Marı´a Velarde-Buendı´a 2 , Rene ´ Enrı´quez-Figueroa, Jayakumar Bose, Sergey Shabala, Jesu ´s Mun ˜ iz-Murguı ´a, and Igor I. Pottosin* Centro Universitario de Investigaciones Biome ´dicas, Universidad de Colima, 28045 Colima, Mexico (I.Z.-J., A.M.V.-B., R.E.-F., J.M.-M., I.I.P.); and School of Agricultural Science, University of Tasmania, Hobart, Tasmania 7001, Australia (J.B., S.S.) Reactive oxygen species (ROS) are integral components of the plant adaptive responses to environment. Importantly, ROS affect the intracellular Ca 2+ dynamics by activating a range of nonselective Ca 2+ -permeable channels in plasma membrane (PM). Using patch-clamp and noninvasive microelectrode ion flux measuring techniques, we have characterized ionic currents and net K + and Ca 2+ fluxes induced by hydroxyl radicals (OH d ) in pea (Pisum sativum) roots. OH d , but not hydrogen peroxide, activated a rapid Ca 2+ efflux and a more slowly developing net Ca 2+ influx concurrent with a net K + efflux. In isolated pro- toplasts, OH d evoked a nonselective current, with a time course and a steady-state magnitude similar to those for a K + efflux in intact roots. This current displayed a low ionic selectivity and was permeable to Ca 2+ . Active OH d -induced Ca 2+ efflux in roots was suppressed by the PM Ca 2+ pump inhibitors eosine yellow and erythrosine B. The cation channel blockers gadolinium, nifedipine, and verapamil and the anionic channel blockers 5-nitro-2(3-phenylpropylamino)-benzoate and niflumate inhibited OH d -induced ionic currents in root protoplasts and K + efflux and Ca 2+ influx in roots. Contrary to expectations, polyamines (PAs) did not inhibit the OH d -induced cation fluxes. The net OH d -induced Ca 2+ efflux was largely prolonged in the presence of spermine, and all PAs tested (spermine, spermidine, and putrescine) accelerated and augmented the OH d -induced net K + efflux from roots. The latter effect was also observed in patch-clamp experiments on root protoplasts. We conclude that PAs interact with ROS to alter intracellular Ca 2+ homeostasis by modulating both Ca 2+ influx and efflux transport systems at the root cell PM. Increased reactive oxygen species (ROS) production is a common denominator of plant adaptive responses to a large number of abiotic and biotic stresses (Mittler, 2002). ROS are produced by cell wall-associated perox- idases, apoplastic diamine and polyamine oxidases, plasma membrane (PM) NADPH oxidase, and oxidases and peroxidases in mitochondria, chloroplasts, and peroxisomes (Mahalingam and Fedoroff, 2003). A com- bination of increasing ROS production and limited energy resources to replenish the antioxidant activity results in ROS accumulation (Taylor et al., 2004). The most abundant types of ROS in plants are hydrogen peroxide (H 2 O 2 ) and the two free oxygen radicals, namely superoxide radical ( d O 2 2 ) and hydroxyl radical (OH d ), the latter being the most reactive with biomol- ecules and structures. The lifetime of the OH d is only 10 29 s, which implies that it acts within 1 nm from the point of its formation and does not cross the membrane (Mori and Schroeder, 2004). Generation of the OH d in roots, via the activity of intrinsic peroxidases, is a prerequisite for cell wall loosening and normal root growth (Schopfer et al., 2002; Liszkay et al., 2004). The extent of ROS accumulation is ultimately a determinant of whether ROS production is part of a signal mecha- nism (at low levels) or a harmful event (at high levels) for plants (Foyer and Noctor, 2005; Miller et al., 2010). Thus, stress-specific modulation of ROS production and scavenging is crucial. Up-regulation of ROS- responsive transcripts under osmotic stress was con- fined almost exclusively to shoots, whereas during salinity, these changes were observed almost exclu- sively in roots (Davletova et al., 2005a, 2005b; Miller et al., 2010). Ion channels have long been considered as potential ROS targets. In the PM, H 2 O 2 activates the hyper- polarization-activated nonselective Ca 2+ -permeable channels (Pei et al., 2000; Demidchik et al., 2007) and inhibits outward- and inward-rectifying potassium channels (Ko ¨hler et al., 2003). The induction of Ca 2+ influx by H 2 O 2 in guard cells mediates elicitor- or abscisic acid-induced stomatal closure (Lee et al., 1999; Pei et al., 2000; Schroeder et al., 2001). OH d has 1 This work was supported by Consejo Nacional de Ciencia y Tecnologı ´a (grant no. CB 82913 to I.I.P. and fellowships to I.Z.-J. and A.M.V.-B.), University of Tasmania Visiting Fellowship to I.I.P., and the Australian Research Council (grant no. DP1094663 to S.S.). 2 These authors contributed equally to the article. 3 Present address: Instituto de Biotecnologı ´a, Universidad Nacio- nal Auto ´noma de Me ´xico, Cuernavaca, Morelos 62210, Mexico. * Corresponding author; e-mail [email protected]. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Igor I. Pottosin ([email protected]). [W] The online version of this article contains Web-only data. www.plantphysiol.org/cgi/doi/10.1104/pp.111.179671 Plant Physiology Ò , December 2011, Vol. 157, pp. 2167–2180, www.plantphysiol.org Ó 2011 American Society of Plant Biologists. All Rights Reserved. 2167
Transcript of Polyamines Interact with Hydroxyl Radicals in Activating Ca2+ and K+ Transport across the Root...

Polyamines Interact with Hydroxyl Radicals inActivating Ca2+ and K+ Transport across the RootEpidermal Plasma Membranes1[W]
Isaac Zepeda-Jazo2,3, Ana Marıa Velarde-Buendıa2, Rene Enrıquez-Figueroa, Jayakumar Bose,Sergey Shabala, Jesus Muniz-Murguıa, and Igor I. Pottosin*
Centro Universitario de Investigaciones Biomedicas, Universidad de Colima, 28045 Colima, Mexico (I.Z.-J.,A.M.V.-B., R.E.-F., J.M.-M., I.I.P.); and School of Agricultural Science, University of Tasmania, Hobart,Tasmania 7001, Australia (J.B., S.S.)
Reactive oxygen species (ROS) are integral components of the plant adaptive responses to environment. Importantly, ROSaffect the intracellular Ca2+ dynamics by activating a range of nonselective Ca2+-permeable channels in plasma membrane(PM). Using patch-clamp and noninvasive microelectrode ion flux measuring techniques, we have characterized ionic currentsand net K+ and Ca2+ fluxes induced by hydroxyl radicals (OH
d
) in pea (Pisum sativum) roots. OHd
, but not hydrogen peroxide,activated a rapid Ca2+ efflux and a more slowly developing net Ca2+ influx concurrent with a net K+ efflux. In isolated pro-toplasts, OH
d
evoked a nonselective current, with a time course and a steady-state magnitude similar to those for a K+ efflux inintact roots. This current displayed a low ionic selectivity and was permeable to Ca2+. Active OH
d
-induced Ca2+ efflux in roots wassuppressed by the PMCa2+ pump inhibitors eosine yellow and erythrosine B. The cation channel blockers gadolinium, nifedipine,and verapamil and the anionic channel blockers 5-nitro-2(3-phenylpropylamino)-benzoate and niflumate inhibited OH
d
-inducedionic currents in root protoplasts and K+ efflux and Ca2+ influx in roots. Contrary to expectations, polyamines (PAs) did not inhibitthe OH
d
-induced cation fluxes. The net OHd
-induced Ca2+ efflux was largely prolonged in the presence of spermine, and all PAstested (spermine, spermidine, and putrescine) accelerated and augmented the OH
d
-induced net K+ efflux from roots. The lattereffect was also observed in patch-clamp experiments on root protoplasts. We conclude that PAs interact with ROS to alterintracellular Ca2+ homeostasis by modulating both Ca2+ influx and efflux transport systems at the root cell PM.
Increased reactive oxygen species (ROS) productionis a common denominator of plant adaptive responsesto a large number of abiotic and biotic stresses (Mittler,2002). ROS are produced by cell wall-associated perox-idases, apoplastic diamine and polyamine oxidases,plasmamembrane (PM)NADPH oxidase, and oxidasesand peroxidases in mitochondria, chloroplasts, andperoxisomes (Mahalingam and Fedoroff, 2003). A com-bination of increasing ROS production and limitedenergy resources to replenish the antioxidant activityresults in ROS accumulation (Taylor et al., 2004). Themost abundant types of ROS in plants are hydrogenperoxide (H2O2) and the two free oxygen radicals,namely superoxide radical (
d
O22) and hydroxyl radical
(OHd
), the latter being the most reactive with biomol-ecules and structures. The lifetime of the OH
d
is only1029 s, which implies that it acts within 1 nm from thepoint of its formation and does not cross the membrane(Mori and Schroeder, 2004). Generation of the OH
d
inroots, via the activity of intrinsic peroxidases, is aprerequisite for cell wall loosening and normal rootgrowth (Schopfer et al., 2002; Liszkay et al., 2004). Theextent of ROS accumulation is ultimately a determinantof whether ROS production is part of a signal mecha-nism (at low levels) or a harmful event (at high levels)for plants (Foyer and Noctor, 2005; Miller et al., 2010).Thus, stress-specific modulation of ROS productionand scavenging is crucial. Up-regulation of ROS-responsive transcripts under osmotic stress was con-fined almost exclusively to shoots, whereas duringsalinity, these changes were observed almost exclu-sively in roots (Davletova et al., 2005a, 2005b; Milleret al., 2010).
Ion channels have long been considered as potentialROS targets. In the PM, H2O2 activates the hyper-polarization-activated nonselective Ca2+-permeablechannels (Pei et al., 2000; Demidchik et al., 2007) andinhibits outward- and inward-rectifying potassiumchannels (Kohler et al., 2003). The induction of Ca2+
influx by H2O2 in guard cells mediates elicitor- orabscisic acid-induced stomatal closure (Lee et al.,1999; Pei et al., 2000; Schroeder et al., 2001). OH
d
has
1 This work was supported by Consejo Nacional de Ciencia yTecnologıa (grant no. CB 82913 to I.I.P. and fellowships to I.Z.-J. andA.M.V.-B.), University of Tasmania Visiting Fellowship to I.I.P., andthe Australian Research Council (grant no. DP1094663 to S.S.).
2 These authors contributed equally to the article.3 Present address: Instituto de Biotecnologıa, Universidad Nacio-
nal Autonoma de Mexico, Cuernavaca, Morelos 62210, Mexico.* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Igor I. Pottosin ([email protected]).
[W] The online version of this article contains Web-only data.www.plantphysiol.org/cgi/doi/10.1104/pp.111.179671
Plant Physiology�, December 2011, Vol. 157, pp. 2167–2180, www.plantphysiol.org � 2011 American Society of Plant Biologists. All Rights Reserved. 2167

activated a nonselective Ca2+-permeable current in themature zone of Arabidopsis (Arabidopsis thaliana)roots, whereas external H2O2 does not evoke anycurrent in this tissue (Demidchik et al., 2003, 2007).The OH
d
-induced current was implicated in elonga-tion via Ca2+ signaling (Foreman et al., 2003). Also, inArabidopsis roots, OH
d
was shown to activate theconstitutively expressed outward-rectifying K+ cur-rent, as part of a programmed cell death scenario(Demidchik et al., 2010). Therefore, it appears thatdifferent ROS may have rather different spectra ofphysiological activities and differ in their effects onthe PM ion channels.
The accumulation of the polyamines (PAs) putrescine(Put2+), spermidine (Spd3+), and spermine (Spm4+) isanother common component of stress responses ofplants and correlates with plant stress resistance(Bouchereau et al., 1999; Walters, 2003). The up- ordown-regulation of genes involved in the biosynthesisand degradation of PAswas reported tomodulate plantsensitivity to drought, salt, osmotic, oxidative, and coldstresses (Kusano et al., 2007a, 2007b; Rhee et al., 2007;Alcazar et al., 2010; Takahashi and Kakehi, 2010). Understress conditions, PAs can act either directly, as chemicalchaperones for DNA and other macromolecules, orindirectly, as positive regulators of stress responsegenes (Rhee et al., 2007). PAs normally reduce themembrane leakage induced by abiotic stresses in plants.In addition to unspecific increase of the membrane“rigidness,” due to the immobilization of negativelycharged phospholipids, more specific effects could bein work, such as activation of the antioxidant enzymesby PAs (Groppa and Benavides, 2008; Gill and Tuteja,2010). In plants, PAsmay play a dual role, as free radicalscavengers and antioxidant enzyme activators but alsoas a source of H2O2 (Takahashi and Kakehi, 2010). If thelatter mechanism prevails, instead of preventing it, PAscould increase the oxidative damage (Mohapatra et al.,2009, and refs. therein).
Among immediate molecular targets for PAs, ionchannels and receptors are receiving growing atten-tion. In animal cells, PAs block a variety of K+ andother cation-selective channels (Drouin and Hermann,1994; Ficker et al., 1994; Lopatin et al., 1994; Bahringet al., 1997; Lu and Ding, 1999). In plants, PAs inhibitPM Shaker-type K+ channels in guard, cortical, epi-dermal, and xylem parenchyma cells (Liu et al., 2000;Zhao et al., 2007), nonselective cation channels inmesophyll and root PM (Shabala et al., 2007; Zhaoet al., 2007), as well as nonselective cation fast andslow vacuolar channels (Bruggemann et al., 1998;Dobrovinskaya et al., 1999a, 1999b). PAs are the onlyorganic polycations that are present in sufficient quan-tities under stress to play the role of channels blockerswithout compromising cell metabolism (Alcazar et al.,2010). At the same time, PAs could act as cofactors inthe activation of PM H+ pumps (Reggiani et al., 1992;Liu et al., 2005b; Garufi et al., 2007).
If the blockage of PM K+ and nonselective cationchannels were the dominant effect of PAs, it would
assist the retention of intracellular K+ and the reduc-tion of Na+ influx under salt stress, thus amelioratingits detrimental effects on plant ionic homeostasis(Zepeda-Jazo et al., 2008). However, recent results byPandolfi et al. (2010) suggested that PAs, depending ongrowth conditions or particular root zone, could eithersuppress the salt-induced K+ efflux or stimulate it.Such stimulation may take place via cross talk betweenPAs and ROS, while the latter are produced in theapoplast via PA catabolization by amino oxidases, asalready mentioned. Indeed, it was shown recently thatPAs may be actively exported to the apoplast and beoxidized there, generating ROS; the latter will activateCa2+ entry across the PM. Such a pathway was shownto mediate abscisic acid-induced stomatal closure inVicia faba guard cells (An et al., 2008), the production ofvolatile terpenoids in lima bean (Phaseolus lunatus)leaves (Ozawa et al., 2009), and to control pollen tubegrowth (Wu et al., 2010). Importantly, salt stress alsoprovokes the exodus of PAs into the apoplast, with itsfurther oxidation and ROS production. Depending onthe conditions, this may result in either a toleranceresponse or lead to programmed cell death (Moschouet al., 2008a). A coproduction of PAs and ROS and theirinterplay under stresses, therefore, may influence PMion conductance in different ways, affecting in partic-ular Ca2+ signaling.
In this work, we have studied ROS-induced Ca2+
and K+ fluxes and currents in pea (Pisum sativum) rootsand tested the effects of PAs on their kinetics. Ourresults suggest a novel mechanism, where PAs act ascofactors in the ROS induction of PM Ca2+-permeablenonselective current and as inducers of the active Ca2+
efflux across the PM.
RESULTS
Kinetics and Pharmacology of the OHd
-Induced K+ andCa2+ Fluxes in Pea Roots
OHd
was generated by the application of copperascorbate (Cu/A; Biaglow et al., 1997; Halliwell andGutteridge, 1999). Briefly, being reduced by ascorbate,copper catalyzes a sequence of one-electron reduc-tion steps from molecular oxygen to
d
O22, from
d
O22 to
H2O2, and finally, from H2O2 to OHd
. The last step isanalogous to the classical Fenton reaction, where re-duced Fe2+ is normally used instead of Cu+ (Biaglowet al., 1997).
Net Ca2+ and K+ fluxes induced by 1 mM Cu/Awere measured by the noninvasive microelectrodeion flux measuring (MIFE) technique from pea rootepidermis (for details, see “Materials and Methods”).The concentration of 1 mM Cu/Awas selected here tomake our results comparable with previously pub-lished studies on roots (Demidchik et al., 2003, 2010;Cuin and Shabala, 2007). This treatment provoked along-lasting (more than 40-min) net K+ efflux frompea roots (Fig. 1A), while the Ca2+ flux showed a morecomplex kinetics, undergoing a switch from a rapidly
Zepeda-Jazo et al.
2168 Plant Physiol. Vol. 157, 2011

developed net Ca2+ efflux immediately after Cu/Aapplication to a net Ca2+ influx 10 to 15 min later (Fig.1B). The application of ascorbate (1 mM) alone did nothave any significant (P, 0.05) effect on net ion fluxes,whereas 1 mM Cu2+ evoked a relatively small K+ andCa2+ efflux (Fig. 1, C and D). As Cu2+ alone acts as asuperoxide radical scavenger (Schopfer et al., 2002),the observed flux stimulation may be related to OH
d
production, due to the reduction of Cu2+ added byintrinsic apoplastic ascorbate. Ca2+ and K+ flux re-sponses were observed also at lower Cu/A concen-trations (see below).The identity of ion flux components evoked by 1 mM
Cu/A was subjected to a pharmacological analysis.Pretreatment with the nonspecific cation channelblocker gadolinium (Gd3+; 0.1 mM) strongly dimin-ished the development of the OH
d
-induced K+ effluxand abolished Ca2+ influx, favoring net Ca2+ efflux(Fig. 1A). Eosine yellow (EY; 0.5 mM), a specific inhi-bitor of the PM Ca2+ pump (Romani et al., 2004;Beffagna et al., 2005), almost completely suppressedthe OH
d
-induced Ca2+ efflux (Fig. 1B) without a sig-nificant effect on the OH
d
-induced K+ efflux (Fig. 1A).Pretreatment with another fluorescein derivative,erythrosine B (0.5 mM), caused a decrease of the OH
d
-induced Ca2+ efflux by 77% (n = 10) without significanteffects on Ca2+ influx or K+ efflux (P , 0.05). Thus, atleast two transport systems appear to mediate theobserved Ca2+ fluxes. One of them is an active Ca2+
efflux system. This system is rapidly activated by OHd
and is sensitive to EY (Fig. 1B), suggesting the Ca2+-ATPase as a possible candidate (White and Broadley,2003). The second (slower) transport component ap-pears to be passive, it is sensitive to Gd3+, and it couldmediate both Ca2+ influx and K+ efflux (Fig. 1, A and B).
Slower (steady-state) Ca2+ and K+ flux componentswere subjected to a further analysis, where pharmaco-logical agents were added directly to the experimentalchamber after 30 min of OH
d
treatment. Figure 2 showsthat the application of 0.1 mM Gd3+, the Ca2+ channelblockers nifedipine and verapamil, which also blocknonselective cation channels activated by OH
d
(Demid-chik et al., 2003; Demidchik and Maathuis, 2007), and5-nitro-2(3-phenylpropylamino)-benzoic acid (NPPB)and niflumic acid, which block several anionic chan-nels in plants (Roberts, 2006), all cause significant (P ,0.05) inhibition of K+ and Ca2+ fluxes. The substantialinhibition of Ca2+ influx by Gd3+, nifedipine, andniflumate unmasked a continuing Ca2+ efflux (Fig.2B). Therefore, both OH
d
-induced Ca2+ efflux and influxappeared to occur at steady state, but at high (1 mM)Cu/A concentration, the influx dominates over theefflux, resulting in a net Ca2+ influx.
PAs Potentiate the OHd
-Induced Ion Fluxes in Pea Roots
In pea roots, the concentration of free PAs rangedfrom submillimolar (Spm4+ and Spd3+) to low milli-molar (up to 4 mM for Put2+; Shen and Galston, 1985).Therefore, we have tested the effects of Put2+, Spd3+, orSpm4+ at 1 mM concentration on the OH
d
-induced K+
and Ca2+ fluxes. Similar concentrations, justified bynatural PA contents, were tested previously againstPM channels in different plant tissues (Liu et al., 2000;Shabala et al., 2007; Zhao et al., 2007). Each of threePAs added in combination with 1 mM Cu/A aug-mented and accelerated the OH
d
-induced K+ efflux(Fig. 3). Simultaneous application of Cu/Awith either1 mM Put2+ or Spd3+ suppressed the Ca2+ influx andslightly diminished the peak Ca2+ efflux. At the same
Figure 1. Effects of Gd3+ and EY on the OHd
-induced K+ and Ca2+ fluxes in pea roots. OH
d
radicals were generated by mixing 1 mM CuCl2with 1 mM sodium ascorbate in the bath at thetimes indicated by arrows. A, K+ fluxes werestrongly suppressed in the presence of Gd3+ (0.1mM; triangles) but were insensitive to EY (0.5 mM;squares). B, Gd3+ at 0.1 mM suppressed the OH
d
-induced Ca2+ influx, whereas the presence of 0.5mM EY in the bath abolished the Ca2+ efflux. Thesign convention “efflux negative” applies to allMIFE measurements. Data are means 6 SE; n = 6,4, and 4 individual roots for control conditions(1 mM Cu/A only), Gd3+, and EY treatments,respectively. C and D, Control experiments,where 1 mM CuCl2 (Cu; white triangles) or sodiumascorbate (A; black circles) was applied sepa-rately. Data are means 6 SE; n = 3 or 4 individualroots, respectively.
Polyamines Potentiate OHd
-Induced Ion Fluxes
Plant Physiol. Vol. 157, 2011 2169

time, in the presence of Spm4+, the OHd
-induced Ca2+
efflux was greatly prolonged (Fig. 4). The effects ofPut2+ or Spm4+ alone and on the background of var-iable concentrations of Cu/A are described below.
Physiological Concentrations of Redox-Active TransientMetals Catalyze the Activation of K+ and Ca2+ Fluxes
The apoplastic space normally contains submillimo-lar concentrations of ascorbate (Pignocchi and Foyer,2003; Kukavica et al., 2009). In legume nodules, copperconcentration is 10 to 50 mM, whereas the concentrationof iron, another redox-active metal, is between 0.1 and0.7 mM (Becana and Klucas, 1992). Similar contents ofcatalytically active copper and iron, equivalent toapproximately 30 mM and 0.3 mM, were found in pearoot cell walls (Kukavica et al., 2009). The productionof OH
d
by the Cu/A mixture depends only weaklyon the ascorbate concentration, within a range of 0.1 to4.0 mM, but strongly on the copper concentration(Biaglow et al., 1997). In light of the above, we havetested the effects of different copper concentrations onthe induction of K+ and Ca2+ fluxes using a fixed (0.1mM) concentration of ascorbate. Results shown inFigure 5A indicate that at physiological concentrations(0.01–0.1 mM) of Cu2+, the Cu/A mixture evokeda relatively small K+ efflux (10–20 nmol m22 s21);this, however, was strongly (up to 10-fold) potentiated
in the presence of Spm4+ or Put2+. Spm4+ alone didnot provoke a significant K+ efflux (36 5 nmol m22 s21),whereas Put2+ induced a K+ efflux of 41 6 5 nmolm22 s21. However, even in the latter case, the K+ effluxproduced by a combination of 0.01 to 0.1 mM Cu2+ (+0.1mM ascorbate) and 1 mM Put2+ was twice as highcompared with the sum of fluxes produced by Cu/Aand Put2+ separately. The synergistic effect of PAs withROS, albeit less pronounced in relative terms, is con-served up to the highest (1 mM) Cu/A concentrationtested.
At the same time, Cu/A-induced Ca2+ efflux reachedits maximum level already at the copper concentrationof 0.1 mM (Fig. 5B). This implies a lower threshold forthe activation of the PMCa2+ efflux system as comparedwith the induction of passive cation permeability, me-diating both K+ efflux and Ca2+ influx. However, theaddition of Put2+ or Spm4+ alone also provoked Ca2+
Figure 2. Pharmacology of the steady-state OHd
-induced K+ and Ca2+
fluxes in pea roots. Drugs (0.1 mM Gd3+, nifedipine, verapamil, NPPB,or niflumic acid) were introduced to the bath 30 min after the inductionof the ion fluxes by 1 mM Cu/A. A, Net K+ fluxes. B, Net Ca2+ fluxes. Ionfluxes were averaged over 5-min periods immediately before (as acontrol) and after drug addition. Fluxes are plotted as means6 SE, withthe number of individual roots for each treatment indicated in paren-theses.
Figure 3. Natural PAs potentiate OHd
-induced K+ efflux in pea roots.Roots were treated either by 1 mM Cu/A alone (circles) or in acombination with 1 mM Put2+ (A), Spd3+ (B), or Spm4+ (C). Data aremeans 6 SE; n = 6 to 7 individual roots assayed for each treatment.
Zepeda-Jazo et al.
2170 Plant Physiol. Vol. 157, 2011
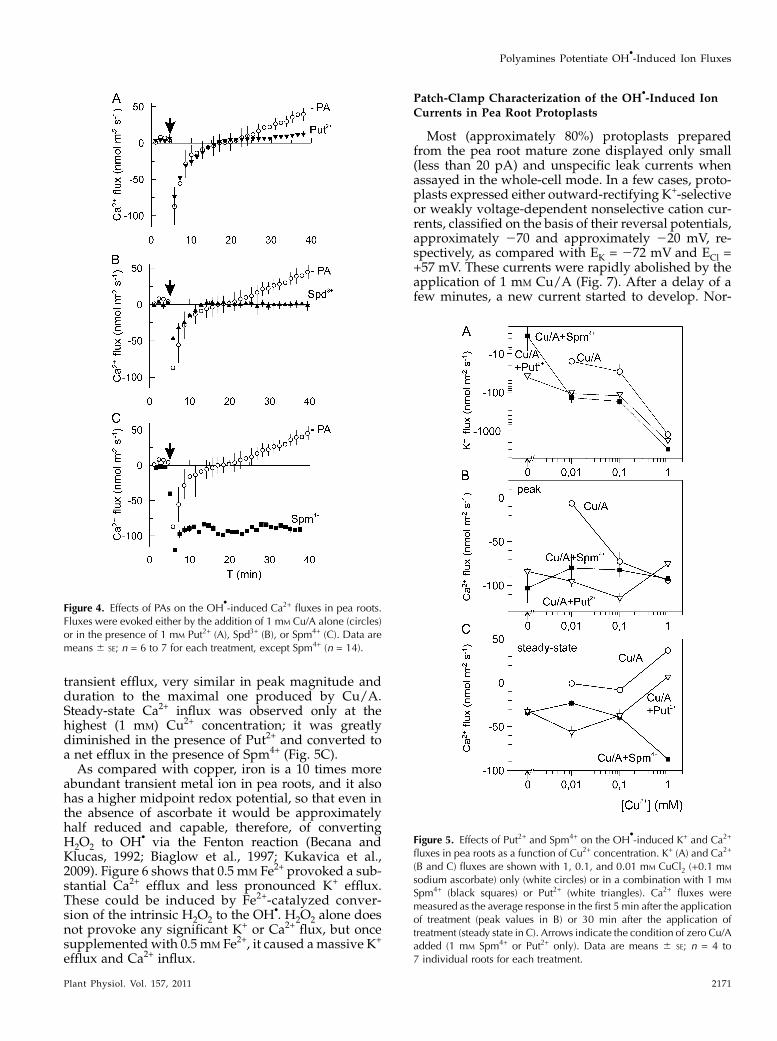
transient efflux, very similar in peak magnitude andduration to the maximal one produced by Cu/A.Steady-state Ca2+ influx was observed only at thehighest (1 mM) Cu2+ concentration; it was greatlydiminished in the presence of Put2+ and converted toa net efflux in the presence of Spm4+ (Fig. 5C).As compared with copper, iron is a 10 times more
abundant transient metal ion in pea roots, and it alsohas a higher midpoint redox potential, so that even inthe absence of ascorbate it would be approximatelyhalf reduced and capable, therefore, of convertingH2O2 to OH
d
via the Fenton reaction (Becana andKlucas, 1992; Biaglow et al., 1997; Kukavica et al.,2009). Figure 6 shows that 0.5 mM Fe2+ provoked a sub-stantial Ca2+ efflux and less pronounced K+ efflux.These could be induced by Fe2+-catalyzed conver-sion of the intrinsic H2O2 to the OH
d
. H2O2 alone doesnot provoke any significant K+ or Ca2+ flux, but oncesupplemented with 0.5 mM Fe2+, it caused amassive K+
efflux and Ca2+ influx.
Patch-Clamp Characterization of the OHd
-Induced Ion
Currents in Pea Root Protoplasts
Most (approximately 80%) protoplasts preparedfrom the pea root mature zone displayed only small(less than 20 pA) and unspecific leak currents whenassayed in the whole-cell mode. In a few cases, proto-plasts expressed either outward-rectifying K+-selectiveor weakly voltage-dependent nonselective cation cur-rents, classified on the basis of their reversal potentials,approximately 270 and approximately 220 mV, re-spectively, as compared with EK = 272 mV and ECl =+57 mV. These currents were rapidly abolished by theapplication of 1 mM Cu/A (Fig. 7). After a delay of afew minutes, a new current started to develop. Nor-
Figure 5. Effects of Put2+ and Spm4+ on the OHd
-induced K+ and Ca2+
fluxes in pea roots as a function of Cu2+ concentration. K+ (A) and Ca2+
(B and C) fluxes are shown with 1, 0.1, and 0.01 mM CuCl2 (+0.1 mM
sodium ascorbate) only (white circles) or in a combination with 1 mM
Spm4+ (black squares) or Put2+ (white triangles). Ca2+ fluxes weremeasured as the average response in the first 5 min after the applicationof treatment (peak values in B) or 30 min after the application oftreatment (steady state in C). Arrows indicate the condition of zero Cu/Aadded (1 mM Spm4+ or Put2+ only). Data are means 6 SE; n = 4 to7 individual roots for each treatment.
Figure 4. Effects of PAs on the OHd
-induced Ca2+ fluxes in pea roots.Fluxes were evoked either by the addition of 1 mM Cu/A alone (circles)or in the presence of 1 mM Put2+ (A), Spd3+ (B), or Spm4+ (C). Data aremeans 6 SE; n = 6 to 7 for each treatment, except Spm4+ (n = 14).
Polyamines Potentiate OHd
-Induced Ion Fluxes
Plant Physiol. Vol. 157, 2011 2171

mally, this current became apparent at about 10 minafter the application of Cu/A and reached a steady-state level after 30 to 40 min of incubation (Fig. 8A).The addition of 1 mM Put2+ or Spm4+ alone did notevoke any current (data not shown). When 1 mM Put2+
or Spm4+ was added simultaneously with 1 mM Cu/A,
the current that developed in a response to ROS grewup faster and reached a larger amplitude as comparedwith the application of 1 mM Cu/A alone (Fig. 8B;Supplemental Fig. S1). PAs (Put2+ and Spm4+) greatlyreduced the delay in the development of OH
d
-inducedcurrent and augmented two or four times, respectively,the maximal current magnitude (Fig. 8C).
To check whether the OHd
-induced current conductsCa2+, we increased the bath CaCl2 concentration to 20mM. If the current conducts cations (Ca2+ in this case)better than anions (Cl2), one should expect an increaseof the inward current due to the increased Ca2+ influx,without or with a little increase of the outward one. Onthe contrary, if the current is carried mainly by anions,an increased influx of Cl2 needs to be reflected by anincrease of the outward current. As can be seen fromthe data presented in Figure 9, A and B, it was theinward current that increased by up to 100% at 2160mV, and this could only be explained by the fact thatOH
d
-induced current mediates a substantial calciuminflux. However, the OH
d
-induced current could alsoconduct other cations, even as large as tetraethylam-monium (TEA+; Fig. 9B). It should be noted that theincrease of the external salt (Ca2+ or TEA+ plus Cl2)concentration had little effect on the reversal potentialof the OH
d
-induced current, which was close to zero inall cases. This behavior is consistent with a weakpreference between cations and anions for their en-trance into the pore from either membrane side, indi-cating a small difference in their relative permeability.Yet, cations appear to be conducted easier across themembrane (i.e. they display a higher absolute perme-ability), reflected by a larger increment of the respec-tive current, inward versus outward, upon theincrease of salt concentration in the bath (for the basesof selective permeability, see Hille, 2001). To verify thehypothesis of a dual cation and Cl2, permeability of
Figure 6. H2O2 alone is unable to induce K+ and Ca2+ fluxes in pearoots. K+ (A) and Ca2+ (B) fluxes were recorded after the application of5 mM H2O2 (white squares), 0.5 mM FeSO4 (white circles), or 5 mM
H2O2 after 0.5 mM FeSO4 (black circles). Data are means 6 SE; n = 5 to6 individual roots for each treatment.
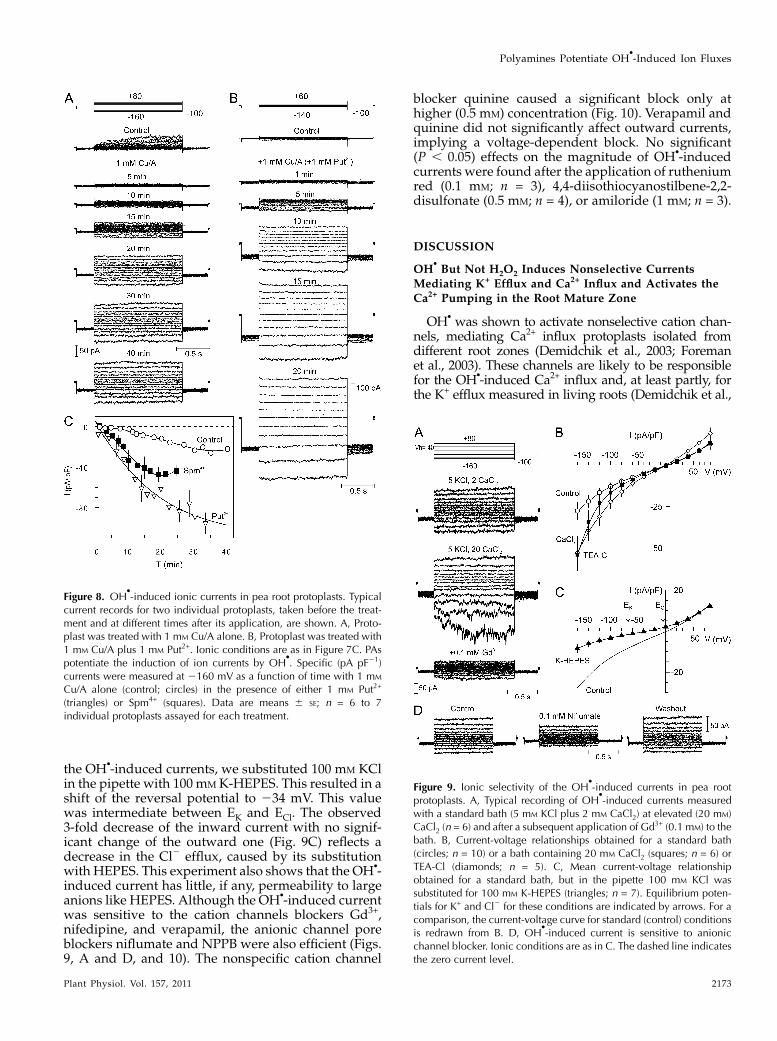
Figure 7. OHd
inhibits ionic currents constitutively expressed in pea root protoplasts. After the achievement and stabilization ofwhole-cell configuration, a control record of currents as a response to a standard voltage steps protocol (depicted at the top) wasundertaken. Original records from the two distinct protoplasts are presented. A, Protoplast expressing outward-rectifyingpotassium current. B, Another protoplast expressing a nonselective cation current. Bottom traces show currents recorded in thesame protoplast as above 3 min after the application of 1 mM Cu/A into the bath. Bath contained 5 mM and pipette contained 100mM KCl; for detailed solution compositions, see “Materials and Methods.” Dashed lines indicate zero current levels.
Zepeda-Jazo et al.
2172 Plant Physiol. Vol. 157, 2011
the OHd
-induced currents, we substituted 100 mM KClin the pipette with 100 mM K-HEPES. This resulted in ashift of the reversal potential to 234 mV. This valuewas intermediate between EK and ECl. The observed3-fold decrease of the inward current with no signif-icant change of the outward one (Fig. 9C) reflects adecrease in the Cl2 efflux, caused by its substitutionwith HEPES. This experiment also shows that the OH
d
-induced current has little, if any, permeability to largeanions like HEPES. Although the OH
d
-induced currentwas sensitive to the cation channels blockers Gd3+,nifedipine, and verapamil, the anionic channel poreblockers niflumate and NPPB were also efficient (Figs.9, A and D, and 10). The nonspecific cation channel
blocker quinine caused a significant block only athigher (0.5 mM) concentration (Fig. 10). Verapamil andquinine did not significantly affect outward currents,implying a voltage-dependent block. No significant(P , 0.05) effects on the magnitude of OH
d
-inducedcurrents were found after the application of rutheniumred (0.1 mM; n = 3), 4,4-diisothiocyanostilbene-2,2-disulfonate (0.5 mM; n = 4), or amiloride (1 mM; n = 3).
DISCUSSION
OHd
But Not H2O2 Induces Nonselective CurrentsMediating K+ Efflux and Ca2+ Influx and Activates theCa2+ Pumping in the Root Mature Zone
OHd
was shown to activate nonselective cation chan-nels, mediating Ca2+ influx protoplasts isolated fromdifferent root zones (Demidchik et al., 2003; Foremanet al., 2003). These channels are likely to be responsiblefor the OH
d
-induced Ca2+ influx and, at least partly, forthe K+ efflux measured in living roots (Demidchik et al.,
Figure 8. OHd
-induced ionic currents in pea root protoplasts. Typicalcurrent records for two individual protoplasts, taken before the treat-ment and at different times after its application, are shown. A, Proto-plast was treated with 1 mM Cu/A alone. B, Protoplast was treated with1 mM Cu/A plus 1 mM Put2+. Ionic conditions are as in Figure 7C. PAspotentiate the induction of ion currents by OH
d
. Specific (pA pF21)currents were measured at 2160 mV as a function of time with 1 mM
Cu/A alone (control; circles) in the presence of either 1 mM Put2+
(triangles) or Spm4+ (squares). Data are means 6 SE; n = 6 to 7individual protoplasts assayed for each treatment.
Figure 9. Ionic selectivity of the OHd
-induced currents in pea rootprotoplasts. A, Typical recording of OH
d
-induced currents measuredwith a standard bath (5 mM KCl plus 2 mM CaCl2) at elevated (20 mM)CaCl2 (n = 6) and after a subsequent application of Gd3+ (0.1 mM) to thebath. B, Current-voltage relationships obtained for a standard bath(circles; n = 10) or a bath containing 20 mM CaCl2 (squares; n = 6) orTEA-Cl (diamonds; n = 5). C, Mean current-voltage relationshipobtained for a standard bath, but in the pipette 100 mM KCl wassubstituted for 100 mM K-HEPES (triangles; n = 7). Equilibrium poten-tials for K+ and Cl2 for these conditions are indicated by arrows. For acomparison, the current-voltage curve for standard (control) conditionsis redrawn from B. D, OH
d
-induced current is sensitive to anionicchannel blocker. Ionic conditions are as in C. The dashed line indicatesthe zero current level.
Polyamines Potentiate OHd
-Induced Ion Fluxes
Plant Physiol. Vol. 157, 2011 2173

2003, 2010). To verify this relation, we have analyzedthe kinetics and pharmacology of OH
d
-induced Ca2+
and K+ fluxes in pea roots, applying a noninvasiveMIFE technique. The results presented in Figures1 and 2 suggest that there are at least two kineticcomponents of the OH
d
-induced fluxes: (1) a relativelyslow K+ efflux paralleled by approximately 20-foldsmaller Ca2+ influx; and (2) a rapidly developed Ca2+
efflux. This tentative component separation was fur-ther confirmed by a pharmacological analysis, as theactive Ca2+ efflux was sensitive to the Ca2+ pump-specific inhibitors EY (Fig. 1B) and erythrosine B.Fluorescein derivatives EY and erythrosine B alsoinhibit the PM H+ pump, but at 1,000-fold higherconcentrations (De Michelis et al., 1993). On the otherhand, both supposedly passive processes, K+ effluxand Ca2+ influx, were blocked by Gd3+, nifedipine,verapamil, niflumate, or NPPB (Fig. 2). We suggested,therefore, that both K+ efflux and Ca2+ influx aremediated by a nonselective passive conductance. Sev-eral lines of evidence are consistent with this sugges-tion. First, the kinetics of the OH
d
-induced currentmeasured in the whole-cell mode in patch experi-ments on individual root protoplasts was similar tothat for K+ efflux and Ca2+ influx in MIFE experimentson intact roots. Second, the steady-state magnitudeof the outward (mainly carried by K+) OH
d
-inducedcurrent, 12 pA pF21 at +80 mV, is equivalent to 1,200nmol m22 s21, fairly comparable to the OH
d
-inducedK+ efflux reported by the MIFE technique (Fig. 1).Third, a 10- fold increase of external Ca2+ provoked an
increase of the inward current by 50% to 100% (be-tween 2120 and 2160 mV; Fig. 9B). This implies that,initially, the Ca2+ fraction was about 5% to 10% of thetotal current, which is comparable to the relationbetween K+ and Ca2+ flux magnitudes in MIFE exper-iments (Fig. 1). Such a relation is to be expected for anonselective channel, whose relative conductance fordifferent ions is determined by the relation of theirconcentrations in experimental media, in this case,more than 1 order of magnitude higher for K+ ascompared with Ca2+. Finally, the inhibition pattern ofthe OH
d
-induced current in patch-clamp experiments(Fig. 10) was qualitatively similar to that obtained byMIFE for the K+ efflux in intact roots (Fig. 2A), withGd3+ being the most potent blocker. One may note thatpositively charged compounds, Gd3+ and verapamil,were more potent in MIFE experiments on intact rootsthan in patch-clamp experiments on isolated proto-plasts. A similar quantitative difference was reportedbefore by Demidchik and coworkers (2003). This dif-ference may be expected due to a lower ionic strengthin MIFE experiments, which tends to increase thenegative surface potential and the local concentrationof cations; surface potential also may be reduced dueto the removal of cell walls upon protoplast isolation.
In studies on plant cells, no selective blockers weredeveloped against any type of ion channel. Moreover,some blockers that were previously considered to berelatively selective (e.g. dihydropyridines such as ni-fedipine or phenylalkylamines such as verapamil,which block voltage-dependent Ca2+-selective chan-nels in animal cells) have different targets in plantcells. These block several types of nonselective Ca2+-permeable channels and even some outward-rectifyingK+ channels (Demidchik and Maathuis, 2007). There-fore, one may not rely on a single blocker and needs totest a variety of broad-spectrum inhibitors.
The effects of TEA+, Gd3+, and verapamil on theOH
d
-induced current reported here were qualitativelysimilar to those reported for the Arabidopsis rootmature epidermis (Demidchik et al., 2003), althoughwith somewhat lower affinity. An apparent lack ofcation/anion selectivity of the OH
d
-induced currentforced us to test additionally some anionic poreblockers; these were proved to be equally efficient(Figs. 2 and 10). Yet, a stilbene derivative, 4,4-diiso-thiocyanostilbene-2,2-disulfonate, irreversibly modi-fying and inhibiting some anion channels andtransporters, was inefficient in our case. No significantinhibition of OH
d
-induced current was found for eitherruthenium red (a known inhibitor of Ca2+ uniporterand a variety of Ca2+ and Ca2+-permeable channels) oramiloride (an inhibitor of cation transporters and somenonselective cation channels).
A poor ion selectivity of the OHd
-induced currentand its sensitivity to both cation and anion channelinhibitors (Figs. 9 and 10) seems surprising at firstglance. Yet, there are multiple reports on plant PMchannels with a low cation-to-anion selectivity (forreview, see Demidchik et al., 2002; for experimental
Figure 10. Pharmacology of the OHd
-induced ion currents in pea rootprotoplasts. Relative ion currents in the presence of Gd3+ (0.1 mM),nifedipine (0.1 mM), verapamil (0.1 mM), quinine (0.5 mM), NPPB (0.1mM), or niflumate (0.1 mM) in the bath are shown. Negative (black) andpositive (white) bars (means 6 SE; n = 3–8 protoplasts for eachtreatment) correspond to currents measured at 2160 mV and +80mV, respectively; control-specific currents (n = 25) at these potentialswere 229 6 1 and 12 6 1 pA pF21, respectively.
Zepeda-Jazo et al.
2174 Plant Physiol. Vol. 157, 2011

evidence, see Zepeda-Jazo et al., 2008). When it comesto OH
d
-induced currents in the root PM, it should benoted that they were never rigorously tested for cationover anion selectivity, as these currents were onlyassayed under pseudosymmetric ionic conditions, inthe absence of steep salt gradients across the mem-brane. Under these ionic conditions, reversal potentialvalues were close to zero, which was a compromisebetween equilibrium potential values for Cl2, mono-valent, and divalent cations (Foreman et al., 2003). Thisobservation might imply a significant anion permea-bility of the OH
d
-induced currents, not only a weakdiscrimination between different cations. Indeed, un-der the ionic conditions of the experiment presented inFigure 9C, the reversal potential of the whole-cellcurrent was substantially different from zero, butagain, it was in the middle between equilibrium po-tentials for K+ and Cl2. Besides, substitution of the 90%of intracellular Cl2 with a large nonpermeable anion(HEPES) unraveled a contribution of the Cl2 influx tothe inward current, which dramatically decreasedwithout a significant change of the outward one. Onthe contrary, the H2O2-activated Ca2+-permeable chan-nels in guard cells and the root elongation zonedisplay a somewhat better selectivity among cations(e.g. lacking permeability for TEA+) and a clear pref-erence of cation over Cl2 (Pei et al., 2000; Demidchiket al., 2007). As shown previously (Demidchik et al.,2007) and in this paper (Fig. 6), H2O2-activated chan-nels are not present in the mature root zone, at leastin the plant species studied. Thus, two different ROSspecies, OH
d
andH2O2, activate distinct Ca2+-permeable
channels in the root PM.
Physiological Concentrations of Copper and Iron CanCatalyze OH
d
Production, Which Induces a Substantial
K+ Leak
The ion fluxes and currents discussed so far wereinduced by high (1 mM) Cu/A concentration. Al-though even higher Cu/A concentrations were usedto demonstrate the role of OH
d
in cell wall looseningand stretching during the elongation process (Schopfer,2001) and equivalent concentrations were applied forthe activation of Ca2+-permeable channels in roots(Demidchik et al., 2003; Foreman et al., 2003), theeffects of lower Cu/A concentrations need to be stud-ied to reveal the thresholds for OH
d
-activated currents.Demidchik and coworkers (2010), using long-livingOH
d
-specific spin-trap 5,5-dimethyl-1-pyrroline-N-oxide, have shown that production of OH
d
by 1 mM
Cu/A in pure solution without plants was approxi-mately equivalent to that produced by intact Arabi-dopsis roots subjected to 100 mM NaCl. However, inthe presence of living roots naturally producing H2O2,the generation of OH
d
induced by 1 mM Cu/A in-creased severalfold and was equivalent to 3- or 5-foldof its production upon the application of 250 or 100 mM
NaCl, respectively. Extreme (several hundred micro-
moles) and lethal OHd
production occurs in illumi-nated photosynthetic tissues treated with herbicides(paraquat) or upon the inhibition of ascorbate perox-idase (Babbs et al., 1989; Burkhard and Heber, 1996).However, even a much lower physiological (10 mM)concentration of copper reduced by ascorbate is able togenerate 2 mmol of OH
d
within 10 min (Biaglow et al.,1997). In biological systems, not only total OH
d
gener-ation but also the location of the production site withrespect to the target molecules and scavenging mecha-nisms existing within this locality is important. Ourdata imply that at up to 0.1 mM Cu/A, only small K+
efflux resulted from the mature zone of pea roots(Fig. 5A). However, this efflux was strongly (up to1 order of magnitude) potentiated by PAs, reaching100 to 200 nmol m22 s21 (see discussion of the mecha-nism of PA action below). On the other hand, iron(another redox-active transient metal capable ofgenerating OH
d
via the Fenton reaction) is present atpea cell walls at 10-fold higher concentrations thancopper, mainly as a part of peroxidase reaction centers(Becana and Klucas, 1992; Kukavica et al., 2009).Moreover, on an equimolar iron basis, the peroxidaseiron is a 1 to 2 orders of magnitude better Fentonreaction catalyst as compared with the inorganic ironor Fe-EDTA (Chen and Schopfer, 1999). In our exper-iments, inorganic iron alone at 0.5 mM concentrationprovoked a relatively large (up to 100 nmol m22 s21) K+
efflux, which grew severalfold upon the application ofmillimolar H2O2 (Fig. 6A). For a comparison, K+ effluxfrom the root mature zone, induced by 50 to 100 mM
NaCl, was (in nmol m22 s21) 100 to 200 for Arabidop-sis, 100 for maize (Zea mays; Pandolfi et al., 2010), 100to 400 for barley (Hordeum vulgare) varieties differentin their salt tolerance (Chen et al., 2007), and about 50on average for different wheat (Triticum aestivum)cultivars (Cuin et al., 2008). Therefore, physiologicalconcentrations of copper (supplemented by PAs) andiron are capable of catalyzing OH
d
production suffi-cient to induce quite a significant K+ leak from pearoots persisting tens of minutes.
PAs Potentiate the OHd
-Induced PM
Passive Conductance
The kinetics of the OHd
-induced cation currents andK+ fluxes across the PM were further modulated byPAs. PAs are present at high, submillimolar to milli-molar, concentrations in pea roots. Spm4+ and Spd3+
are more abundant near root apices, whereas Put2+
is almost equally distributed between the root baseand apex (Shen and Galston, 1985). When it comes tothe known effects of PAs on passive membrane con-ductance, only the inhibitory effects, either direct orindirect, on K+-selective and nonselective cation chan-nels were documented for plants so far (Bruggemannet al., 1998; Dobrovinskaya et al., 1999a, 1999b; Liuet al., 2000; Shabala et al., 2007; Zhao et al., 2007). Thus,beneficial roles of PAs during stresses were discussed
Polyamines Potentiate OHd
-Induced Ion Fluxes
Plant Physiol. Vol. 157, 2011 2175

in this context. As an example, the PA contribution tosalt stress tolerance in plants was explained on thebasis of their inhibition of Na+ influx, reducing theconsequent membrane depolarization and K+ efflux(Shabala et al., 2007; Zhao et al., 2007; Zepeda-Jazoet al., 2008). However, our recent study (Pandolfi et al.,2010) showed that PAs, in particularly Spm4+, depend-ing on the growing condition and root zone, canreduce NaCl-induced K+ efflux, cause no change, oreven stimulate it. Clearly, for explanations of theeffects of externally applied PAs in vivo, one needsto consider not only the effects of PAs per se but also ofPA metabolization associated with ROS generation aswell as their roles in ROS scavenging as early effects.In particular, all natural PAs are powerful OH
d
scav-engers, at concentrations above 1.5 mM eliminatingvirtually all OH
d
produced by a mixture of 0.2 mM
H2O2 with 0.04 mM iron (Das and Misra, 2004). On theother hand, oxidation of Spm4+ and Spd3+ by poly-amine oxidase, and Put2+ by diamine oxidase, leads tothe formation of H2O2 (Moschou et al., 2008b), whichcould be further converted to OH
d
. External applica-tion of PAs to roots mimics their export to apoplast,which occurred in response to different environmen-tal cues, and is associated with a stimulation of Ca2+
influx by ROS, generated as a result of PA oxidation(An et al., 2008; Moschou et al., 2008a; Ozawa et al.,2009; Wu et al., 2010). In addition to the Ca2+ influx,overall PM leakage could increase, also leading to K+
loss. Thus, catabolization of PAs and ROS genera-tion may outweigh their roles as ROS scavengers,activators of the antioxidant enzymes, and micro-somal NADPH oxidase inhibitors (Papadakis andRoubelakis-Angelakis, 2005; Mohapatra et al., 2009;Takahashi and Kakehi, 2010). Increase of ROS pro-duction due to PA catabolization may indeed takeplace in intact pea roots. Indirect evidence for this isthat Put2+ alone provoked a much higher K+ efflux ascompared with Spm4+ (Fig. 5A), which may reflect amuch higher apoplast expression of diamine oxidaseas compared with polyamine oxidase, in the apoplastof dicots, particularly pea (Moschou et al., 2008b). Thecontribution of such a mechanism should be ruled out,however, in the case of isolated protoplasts, becauseapoplastic amine oxidases are washed out upon theprotoplast isolation procedure (Kaur-Sawhney et al.,1981). Besides, we have not observed any membranecurrent stimulation by treatment with PAs alone. Still,PAs may act as OH
d
scavengers, but this could onlypreclude or handicap the development of OH
d
-induced ionic currents. Therefore, exogenous PAscould stimulate the induction of ionic currents inprotoplasts by OH
d
only if PAs are acting as cofactorsin this process. To the best of our knowledge, thispossibility has not been considered in the literature sofar. However, it was reported that PAs may stabilizethe binding of other cofactors essential for the ionchannel activity, such as PIP2 for Kir and plant ShakerK+ channels (Liu et al., 2005a; Xie et al., 2005). Thus, theprecise biophysical mechanisms of this synergism
between PAs and ROS interaction warrant a separateinvestigation.
PAs and OHd
Activate Ca2+ Pumping in Intact Roots
Another intriguing finding of this study is the acti-vation of active Ca2+ efflux across the root epidermisby OH
d
and PAs (Figs. 1B, 4, and 5). This flux compo-nent has a substantially lower threshold for its activa-tion by OH
d
as compared with the K+ efflux (Fig. 5,A and B). Stimulation of the active EY-sensitive Ca2+
efflux across the PM by ROS was reported in a singlestudy on an Arabidopsis cell culture (Romani et al.,2004). It was shown that ROS generated by oligoga-lacturonide treatment assisted Ca2+-ATPase activationby calmodulin. However, in that paper, ROS inducednet Ca2+ influx first, and net Ca2+ efflux was measuredonly in 10 to 20 min; this is exactly the opposite ordercompared with the OH
d
-induced Ca2+ flux kinetics inour work (Figs. 1B and 4). On the other hand, there is alarge body of data on animal PM and sarcoplasmicreticulum Ca2+ pumps showing their inhibition byROS, which may originate from protein cross-linking,lipid peroxidation, and concurrent inhibition by oxi-dized forms of the calmodulin (for review, see Waring,2005). These effects develop slowly (hours), whichmay preclude their observation under the conditionsof our study.
At the same time, Put2+ and Spm4+ evoked Ca2+
efflux that was very similar to that induced by OHd
.One possibility is that this efflux was actually causedby ROS, generated during PA catabolization. Alterna-tively/additionally, PAs in principle can activate PMCa2+-ATPase in one of the ways they affect anotherP-type pump, H+-ATPase. These involve the PA-induced promotion of the interaction of 14-3-3 proteinswith the autoinhibitory domain at the H+-ATPase Cterminus (Garufi et al., 2007) and the activation of the H+
pump via a nitric oxide-dependent pathway (Tun et al.,2006; Arasimowicz-Jelonek et al., 2009; Zandonadi et al.,2010). Finally, if PAs activate the H+ pump, they couldcause the stimulation of Ca2+ pumping across the PMvia a coupled mechanism, as PM Ca2+-ATPase exportstwo Ca2+ ions in the exchange for the two imported H+
ions (Beffagna et al., 2005).
Implications of PA and OHd
Effects on Intracellular
Ca2+ Homeostasis
Irrespective of the precise mechanism of ROS andPA early effects on Ca2+ fluxes across the root epider-mis PM, some consequences for Ca2+ homeostasisand signaling could be drawn from our experimen-tal data. Moderate OH
d
production results in a tran-sient increase of Ca2+ pumping, outweighing theOH
d
-induced passive Ca2+ influx (Fig. 5, B and C).The resultant depletion of the intracellular Ca2+ poolshould reduce the Ca2+-dependent PM NADPH oxi-dase activity in a feedback manner (Takeda et al.,2008), thus diminishing ROS generation. Membrane-
Zepeda-Jazo et al.
2176 Plant Physiol. Vol. 157, 2011

bound NADPH oxidase is an electrogenic enzyme thattransports an electron from the interior to the exteriorof the cell upon oxygen reduction, and it is stimulatedby membrane hyperpolarization. The latter may begenerated by an increased PM pump activity. As anexample, activation of the H+-ATPase by fusicoccinresults in higher OH
d
production (Liszkay et al., 2004).However, activation of the nonselective ion conduc-tance by OH
d
(Fig. 9) would tend to depolarize themembrane, although opposed by enhanced H+ andCa2+ pumping as a response to the depolarization. ThePM H+ pump is also activated by hyperosmotic stressor salt. This increases cytosolic pH, which leads to alower synthesis of NADPH, thus decreasing its avail-ability for NADPH oxidase (Beffagna et al., 2005).And, as mentioned in a previous section, the H+ pumpmay be activated by PAs. As Ca2+-ATPase operates as a1:1 Ca2+:H+ exchanger, it might be of secondary rele-vance which enzyme, H+- or Ca2+-ATPase, is theprimary target for OH
d
/ PA, while the activity ofboth pumps will be mutually coupled via the H+
circuit. Thus, most likely, under conditions of moder-ate OH
d
production, the joint activity of H+ and Ca2+
pumps would tend to diminish ROS production bythe PM NADPH oxidase. Oscillations of intracellularH+ and Ca2+, in parallel with oscillations of ROSproduction levels, may also be expected under theseconditions.At strong oxidative stress, here experimentally pro-
voked by 1 mM Cu/A, in the absence of PAs, net Ca2+
influx was observed at steady state (after 30 minof incubation; Figs. 1B and 5C). However, on thebackground of PAs (Figs. 4 and 5C), it is either largelydiminished (Put2+) or reverted to an efflux (Spm4+).The direction of the net Ca2+ flux at any time is definedby the sign of the algebraic sum of Ca2+ efflux andinflux, which are generated by distinct PM transportsystems. As can be seen from Figures 3 and 8C, atlonger incubation times, Put2+ stimulates the OH
d
-induced passive K+ efflux (and, presumably, also theassociated Ca2+ influx) more than Spm4+ does. At thesame time, active Ca2+ efflux observed in the presenceof Put2+ and Spm4+ is approximately equal (Fig. 5B).Thus, at longer incubation times, one may expect arelatively higher net Ca2+ efflux in the presence ofSpm4+ as compared with Put2+, which is indeed thecase (Fig. 4). Thus, the ratio between different PAs isimportant to define the direction of a net Ca2+ fluxacross the PM. In the pea root mature zone, Put2+ isaccumulated at a higher concentration than the sumof Spd3+ and Spm4+ concentrations (Shen and Galston,1985), although in the apoplast of pea and other dicots,the activity of diamine oxidase dominates over theactivity of polyamine oxidase (Moschou et al., 2008b).PA concentration patterns are known to be not onlyspecies, tissue, and age specific but also stress specific(Alcazar et al., 2010). As an example, salt stress shiftsthe distribution in favor of higher PAs, Spm4+ andSpd3+, at the expense of Put2+, even though the totalPA concentration changes are not very dramatic. Yet,
plants opposing this tendency (i.e. maintaining ahigher Put2+/Spm4+ ratio) turn out to be more salttolerant and, importantly, to better accumulate cat-ions, including Na+, as required for the osmoticadjustment (Zapata et al., 2008). Among other expla-nations, this might be related to the fact that Put2+
could support a longer lasting potentiation of the OHd
-induced passive conductance (Fig. 3A) and could also,in contrast to Spm4+ and Spd3+, allow some net Ca2+
influx at steady state (Fig. 4).Summarizing, our study demonstrates that lower
OHd
concentrations induce Ca2+ pumping, while higherOH
d
levels activate also a passive Ca2+ uptake across theroot PM. The overall direction of the net Ca2+ flux isfurther modulated by natural PAs in a species-specificmanner. Thus, stress-induced changes in OH
d
produc-tion and in the levels of different PAs may be translatedinto intracellular Ca2+ changes.
MATERIALS AND METHODS
Plant Material
Pea (Pisum sativum ‘Greenfeast’) seeds were surface sterilized (full-strength
commercial bleach for 30 min) and thoroughly rinsed with distilled water.
Seeds were germinated in a dark growth cabinet at +24�C in two layers of wet
paper in petri dishes for 2 to 3 d. Uniformly germinated seedlings were
selected and transferred to a bubbled hydroponic culture unit comprising a
3-L plastic container over which seedlings were suspended on a plastic grid so
that their roots were almost completely immersed in the growth solution (0.5
mMKCl and 0.1 mM CaCl2). Aeration was provided by one aquarium air pump
via flexible plastic tubing. Seedlings were grown under constant (+24�C)conditions in a lighted growth cabinet until 5 d old. Roots of 8 to 10 cm long
were used for current and flux measurements.
Measurements of Ion Fluxes
Net K+ and Ca2+ fluxes were measured using the noninvasive MIFE
technique (University of Tasmania innovation). The principles of the MIFE
measurements, microelectrode fabrication, and calibration are available in
previous publications (Shabala et al., 2001, 2007), and the theory of the MIFE
measurements is available elsewhere (Newman, 2001; Shabala et al., 2006a).
During experiments, pea seedlings were placed in 30-mL measuring cham-
bers. Their roots were immobilized in a horizontal position as described
elsewhere (Cuin and Shabala, 2005) and preincubated in a new solution
containing 0.5 mM KCl, 0.1 mM CaCl2, 5 mM MES, and 2 mM Tris base, pH 6.0,
for 1 h. Normally, two ion-selective microelectrodes, one for K+ and another
for Ca2+, were used in the same experiment. During measurements, electrodes
were moved between positions M1 and M2, 50 and 150 mm from the root
surface, respectively, in the mature zone (approximately 15–20 mm from the
tip). OHd
was generated by the addition of CuCl2 (0.01–1 mM) plus sodium
ascorbate (0.1–1 mM) into the chamber. PAs (Spm4+, Spd3+, and Put2+) were
added simultaneously with Cu/A mixture, or individually. Net K+ and Ca2+
fluxes were measured for over 40 min after the treatment. Different blockers
were added 10 min before the Cu/A treatment (EY, erythrosine B, Gd3+) or 30
min later (Gd3+, nifedipine, NPPB, niflumate) as indicated.
Isolation of Pea Protoplasts from the Root Mature Zone
The isolation of root protoplasts was developed based on the previously
described protocols used for mesophyll protoplasts (Demidchik and Tester,
2002; Shabala et al., 2006b) using the same modifications as described by Chen
et al. (2007). Exodermal cylinders of root segments of pea seedlings, after
mechanical separation of steles, were placed into 3 mL of the enzyme solution
containing 2% (w/v) cellulase (Yakult Honsha), 1.2% (w/v) cellulysin
(Calbiochem), 0.1% (w/v) pectolyase, 0.1% (w/v) bovine serum albumin, 10
mM KCl, 10 mM CaCl2, and 2 mM MgCl2, pH 5.7, adjusted with 2 mM MES, and
Polyamines Potentiate OHd
-Induced Ion Fluxes
Plant Physiol. Vol. 157, 2011 2177

osmolality was adjusted to 750 mosmol with sorbitol. After 40 min of
incubation in the enzyme solution (in the dark at 30�C; agitated on a 90-rpm
rotary shaker), root segments were transferred to the so-called wash solution
(as above, minus enzymes) and thoroughly washed for another 2 min.
Segments were then transferred into the measuring chamber filled with a
solution containing 5 mM KCl, 2 mM CaCl2, 0.5 mM MgCl2, and 2 mM MES-
KOH, pH 5.7, with osmolality of 650 mosmol adjusted by D-sorbitol. By gently
shaking, protoplasts were released into the measuring chamber used for
patch-clamp experiments (see next section). Once the protoplasts were re-
leased, we washed them with EDTA-bath solution (the same as release-bath
solution plus 5 mM EDTA) to clean the cell membrane surface and then
washed them again with release-bath solution alone. The origin of released
protoplasts was verified by size distribution, as within a mature root zone the
average cortical cell is about twice as large in diameter as an epidermal one
(Rost et al., 1988). The mean diameter for protoplasts used in patch-clamp
experiments was 11.5 mm, which coincided with a position of a peak
corresponding to a fraction of the smallest protoplasts in our preparation
(mean diameter of 11.2 mm, close to the value reported for pea protoplasts
from the root epidermis); due to some overlapping of peaks corresponding to
different protoplast fractions (Supplemental Fig. S2), one may not exclude
some contribution of cortical protoplasts into those studied by patch clamp,
with a probability of more than 60% that the patched protoplasts originated
from the epidermis.
Patch-Clamp Experiments
Measurements on individual pea root protoplasts were made using an
Axopatch 200 patch-clamp amplifier (Axon Instruments) in the conventional
whole-cell configuration as described by Shabala et al. (2006b). The basic
pipette solution contained (in mM) 100 KCl, 3 MgCl2, 0.8 CaCl2, 2 K2EGTA, and
5 HEPES-KOH, pH 7.4, osmolality of 650 mosmol adjusted with D-sorbitol.
Membrane potentials were clamped at2100 mV throughout the experiments,
and voltage pulses were applied in 20-mV steps, from 2140 or 2160 mV to
+40 or +80 mV. A typical access resistance was between 12 and 30
MV, compensated by 60% to 70% using the Axopatch 200 compensation
circuit, and the whole-cell capacitance was in the range between 2 and 6 pF. To
stimulate the production of OHd
, 1 mM CuCl2 and sodium ascorbate were
mixed directly in the experimental chamber. PAs (Spm4+ or Put2+) were added
directly into the bath solution to give a final concentration of 1 mM. In
pharmacological assays, the inhibitors Gd3+ (0.1 mM), nifedipine (0.1 mM),
verapamil (0.1 mM), quinine (0.5 mM), amiloride (1 mM), ruthenium red (0.1
mM), and NPPB and niflumate (both 0.1 mM) were added directly into the
bath. In selectivity measurements, external solutions with 20 mM CaCl2 or
TEA-Cl were applied by bath perfusion. Anion (Cl2) permeability was
verified by substitution of 100 mM Cl2 in the patch pipette with 100 mM
K-HEPES (100 mM KOH plus 244 mM HEPES, pH 7.4). In the latter case, the
necessary correction for liquid junction potential between the pipette and the
bath was made using the JPCalc program by P.H. Barry (University of New
South Wales). All chemicals were of analytical grade, purchased from Sigma.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Development of the OHd
-induced current is
accelerated in the presence of spermine.
Supplemental Figure S2. Smallest, mostly of epidermal origin, protoplasts
are selected for patch-clamp assays.
Received May 6, 2011; accepted October 3, 2011; published October 6, 2011.
LITERATURE CITED
Alcazar R, Altabella T, Marco F, Bortolotti C, Reymond M, Koncz C,
Carrasco P, Tiburcio AF (2010) Polyamines: molecules with regulatory
functions in plant abiotic stress tolerance. Planta 231: 1237–1249
An ZF, Jing W, Liu YL, Zhang WH (2008) Hydrogen peroxide generated by
copper amine oxidase is involved in abscisic acid-induced stomatal
closure in Vicia faba. J Exp Bot 59: 815–825
Arasimowicz-Jelonek M, Floryszak-Wieczorek J, Kubis J (2009) Interac-
tion between polyamine and nitric oxide signaling in adaptive re-
sponses to drought in cucumber. J Plant Growth Regul 28: 177–186
Babbs CF, Pham JA, Coolbaugh RC (1989) Lethal hydroxyl radical pro-
duction in paraquat-treated plants. Plant Physiol 90: 1267–1270
Bahring R, Bowie D, Benveniste M, Mayer ML (1997) Permeation and
block of rat GluR6 glutamate receptor channels by internal and external
polyamines. J Physiol 502: 575–589
Becana M, Klucas RV (1992) Transition metals in legume root nodules:
iron-dependent free radical production increases during nodule senes-
cence. Proc Natl Acad Sci USA 89: 8958–8962
Beffagna N, Buffoli B, Busi C (2005) Modulation of reactive oxygen species
production during osmotic stress in Arabidopsis thaliana cultured cells:
involvement of the plasma membrane Ca2+-ATPase and H+-ATPase.
Plant Cell Physiol 46: 1326–1339
Biaglow JE, Manevich Y, Uckun F, Held KD (1997) Quantitation of
hydroxyl radicals produced by radiation and copper-linked oxidation
of ascorbate by 2-deoxy-D-ribose method. Free Radic Biol Med 22:
1129–1138
Bouchereau A, Aziz A, Lahrer F, Martin-Tanguy J (1999) Polyamines and
environmental challenges: recent development. Plant Sci 140: 103–125
Bruggemann L, Pottosin I, Schonknecht G (1998) Cytoplasmic polyamines
block the fast-activating vacuolar cation channel. Plant J 16: 101–105
Burkhard J, Heber U (1996) Photoproduction and detoxification of hy-
droxyl radicals in chloroplasts and leaves and relation to photoactiva-
tion of photosystem I and II. Plant Cell Physiol 37: 629–635
Chen SX, Schopfer P (1999) Hydroxyl-radical production in physiological
reactions: a novel function of peroxidase. Eur J Biochem 260: 726–735
Chen Z, Pottosin II, Cuin TA, Fuglsang AT, Tester M, Jha D, Zepeda-Jazo
I, Zhou M, Palmgren MG, Newman IA, et al (2007) Root plasma
membrane transporters controlling K+/Na+ homeostasis in salt-stressed
barley. Plant Physiol 145: 1714–1725
Cuin TA, Betts SA, Chalmandrier R, Shabala S (2008) A root’s ability to
retain K+ correlates with salt tolerance in wheat. J Exp Bot 59: 2697–2706
Cuin TA, Shabala S (2005) Exogenously supplied compatible solutes
rapidly ameliorate NaCl-induced potassium efflux from barley roots.
Plant Cell Physiol 46: 1924–1933
Cuin TA, Shabala S (2007) Compatible solutes reduce ROS-induced
potassium efflux in Arabidopsis roots. Plant Cell Environ 30: 875–885
Das KC, Misra HP (2004) Hydroxyl radical scavenging and singlet oxygen
quenching properties of polyamines. Mol Cell Biochem 262: 127–133
Davletova S, Rizhsky L, Liang H, Shengqiang Z, Oliver DJ, Coutu J,
Shulaev V, Schlauch K, Mittler R (2005a) Cytosolic ascorbate peroxi-
dase 1 is a central component of the reactive oxygen gene network of
Arabidopsis. Plant Cell 17: 268–281
Davletova S, Schlauch K, Coutu J, Mittler R (2005b) The zinc-finger
protein Zat12 plays a central role in reactive oxygen and abiotic stress
signaling in Arabidopsis. Plant Physiol 139: 847–856
De Michelis MI, Carnelli A, Rasi-Caldogno F (1993) The Ca-pump of the
plasma membrane of Arabidopsis thaliana: characteristics and sensitivity
to fluorescein derivatives. Bot Acta 106: 20–25
Demidchik V, Cuin TA, Svistunenko D, Smith SJ, Miller AJ, Shabala S,
Sokolik A, Yurin V (2010) Arabidopsis root K+-efflux conductance
activated by hydroxyl radicals: single-channel properties, genetic basis
and involvement in stress-induced cell death. J Cell Sci 123: 1468–1479
Demidchik V, Davenport RJ, Tester M (2002) Nonselective cation channels
in plants. Annu Rev Plant Biol 53: 67–107
Demidchik V, Maathuis FJM (2007) Physiological roles of nonselective
cation channels in plants: from salt stress to signalling and develop-
ment. New Phytol 175: 387–404
Demidchik V, Shabala SN, Coutts KB, Tester MA, Davies JM (2003) Free
oxygen radicals regulate plasma membrane Ca2+- and K+-permeable
channels in plant root cells. J Cell Sci 116: 81–88
Demidchik V, Shabala SN, Davies JM (2007) Spatial variation in H2O2
response of Arabidopsis thaliana root epidermal Ca2+ flux and plasma
membrane Ca2+ channels. Plant J 49: 377–386
Demidchik V, Tester MA (2002) Sodium fluxes through nonselective cation
channels in the plasma membrane of protoplasts from Arabidopsis
roots. Plant Physiol 128: 379–387
Dobrovinskaya OR, Muniz J, Pottosin II (1999a) Asymmetric block of the
plant vacuolar Ca(2+)-permeable channel by organic cations. Eur Bio-
phys J 28: 552–563
Dobrovinskaya OR, Muniz J, Pottosin II (1999b) Inhibition of vacuolar ion
channels by polyamines. J Membr Biol 167: 127–140
Drouin H, Hermann A (1994) Intracellular action of spermine on neuronal
Ca2+ and K+ currents. Eur J Neurosci 6: 412–419
Zepeda-Jazo et al.
2178 Plant Physiol. Vol. 157, 2011

Ficker E, Taglialatela M, Wible BA, Henley CM, Brown AM (1994)
Spermine and spermidine as gating molecules for inward rectifier K+
channels. Science 266: 1068–1072
Foreman J, Demidchik V, Bothwell JH, Mylona P, MiedemaH, Torres MA,
Linstead P, Costa S, Brownlee C, Jones JDG, et al (2003) Reactive
oxygen species produced by NADPH oxidase regulate plant cell
growth. Nature 422: 442–446
Foyer CH, Noctor G (2005) Redox homeostasis and antioxidant signaling: a
metabolic interface between stress perception and physiological re-
sponses. Plant Cell 17: 1866–1875
Garufi A, Visconti S, Camoni L, Aducci P (2007) Polyamines as physio-
logical regulators of 14-3-3 interaction with the plant plasma membrane
H+-ATPase. Plant Cell Physiol 48: 434–440
Gill SS, Tuteja N (2010) Polyamines and abiotic stress tolerance in plants.
Plant Signal Behav 5: 26–33
Groppa MD, Benavides MP (2008) Polyamines and abiotic stress: recent
advances. Amino Acids 34: 35–45
Halliwell B, Gutteridge JMC (1999) Free Radicals in Biology and Medi-
cine, Ed 3. Oxford University Press, Oxford
Hille B (2001) Ion Channels of Excitable Membranes, Ed 3. Sinauer
Associates, Sunderland, MA
Kaur-Sawhney R, Flores HE, Galston AW (1981) Polyamine oxidase in oat
leaves: a cell wall-localized enzyme. Plant Physiol 68: 494–498
Kohler B, Hills A, Blatt MR (2003) Control of guard cell ion channels by
hydrogen peroxide and abscisic acid indicates their action through
alternate signaling pathways. Plant Physiol 131: 385–388
Kukavica B, Mojovic M, Vuccinic Z, Maksimovic V, Takahama U,
Jovanovic SV (2009) Generation of hydroxyl radical in isolated pea
root cell wall, and the role of cell wall-bound peroxidase, Mn-SOD and
phenolics in their production. Plant Cell Physiol 50: 304–317
Kusano T, Yamaguchi K, Berberich T, Takahashi Y (2007a) Advances in
polyamine research in 2007. J Plant Res 120: 345–350
Kusano T, Yamaguchi K, Berberich T, Takahashi Y (2007b) The polyamine
spermine rescues Arabidopsis from salinity and drought stresses. Plant
Signal Behav 2: 251–252
Lee S, Choi H, Suh S, Doo IS, Oh KY, Choi EJ, Schroeder Taylor AT, Low
PS, Lee Y (1999) Oligogalacturonic acid and chitosan reduce stomatal
aperture by inducing the evolution of reactive oxygen species from
guard cells of tomato and Commelina communis. Plant Physiol 121:
147–152
Liszkay A, van der Zalm E, Schopfer P (2004) Production of reactive
oxygen intermediates (O2.-, H2O2, and
.OH) by maize roots and their role
in wall loosening and elongation growth. Plant Physiol 136: 3114–3123
Liu HP, Yu BJ, Zhang WH, Liu YL (2005b) Effect of osmotic stress on the
activity of H+-ATPase and the levels of covalently and noncovalently
conjugated polyamines in plasma membrane preparation from wheat
seedling roots. Plant Sci 168: 1599–1607
Liu K, Fu H, Bei Q, Luan S (2000) Inward potassium channel in guard cells
as a target for polyamine regulation of stomatal movements. Plant
Physiol 124: 1315–1326
Liu K, Li LG, Luan S (2005a) An essential function of phosphatidylinositol
phosphates in activation of plant shaker-type K+ channels. Plant J 42:
433–443
Lopatin AN, Makhina EN, Nichols CG (1994) Potassium channel block by
cytoplasmic polyamines as the mechanism of intrinsic rectification.
Nature 372: 366–369
Lu Z, Ding L (1999) Blockade of a retinal cGMP-gated channel by polya-
mines. J Gen Physiol 113: 35–43
Mahalingam R, Fedoroff N (2003) Stress response, cell death and signal-
ing: the many faces of ROS. Physiol Plant 119: 56–68
Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R (2010) Reactive oxygen
species homeostasis and signalling during drought and salinity stresses.
Plant Cell Environ 33: 453–467
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends
Plant Sci 7: 405–410
Mohapatra S, Minocha R, Long S, Minocha SC (2009) Putrescine over-
production negatively impacts the oxidative state of poplar cells in
culture. Plant Physiol Biochem 47: 262–271
Mori IC, Schroeder JI (2004) Reactive oxygen species activation of plant
Ca2+ channels: a signaling mechanism in polar growth, hormone trans-
duction, stress signaling, and hypothetically mechanotransduction.
Plant Physiol 135: 702–708
Moschou PN, Paschalidis KA, Delis ID, Andriopoulou AH, Lagiotis GD,
Yakoumakis DI, Roubelakis-Angelakis KA (2008a) Spermidine exodus
and oxidation in the apoplast induced by abiotic stress is responsible for
H2O2 signatures that direct tolerance responses in tobacco. Plant Cell 20:
1708–1724
Moschou PN, Paschalidis KA, Roubelakis-Angelakis KA (2008b)
Plant polyamine catabolism: the state of the art. Plant Signal Behav 3:
1061–1066
Newman IA (2001) Ion transport in roots: measurement of fluxes using ion-
selective microelectrodes to characterize transporter function. Plant
Cell Environ 24: 1–14
Ozawa R, Bertea CM, Foti M, Narayana R, Arimura GI, Muroi A,
Horiuchi JI, Nishioka T, Maffei ME, Takabayashi J (2009) Exogenous
polyamines elicit herbivore-induced volatiles in lima bean leaves:
involvement of calcium, H2O2 and jasmonic acid. Plant Cell Physiol
50: 2183–2199
Pandolfi C, Pottosin I, Cuin T, Mancuso S, Shabala S (2010) Specificity of
polyamine effects on NaCl-induced ion flux kinetics and salt stress
amelioration in plants. Plant Cell Physiol 51: 422–434
Papadakis AK, Roubelakis-Angelakis KA (2005) Polyamines inhibit
NADPH oxidase-mediated superoxide generation and putrescine pre-
vents programmed cell death induced by polyamine oxidase-generated
hydrogen peroxide. Planta 220: 826–837
Pei ZM, Murata Y, Benning G, Thomine S, Klusener B, Allen GJ, Grill E,
Schroeder JI (2000) Calcium channels activated by hydrogen peroxide
mediate abscisic acid signalling in guard cells. Nature 406: 731–734
Pignocchi C, Foyer CH (2003) Apoplastic ascorbate metabolism and its role
in the regulation of cell signalling. Curr Opin Plant Biol 6: 379–389
Reggiani R, Milano I, Zaina S, Bertani A (1992) Plasmalemma ATPase in
rice coleoptiles: stimulation by putrescine and polyamines. Phytochem-
istry 31: 417–419
Rhee HJ, Kim EJ, Lee JK (2007) Physiological polyamines: simple primor-
dial stress molecules. J Cell Mol Med 11: 685–703
Roberts SK (2006) Plasma membrane anion channels in higher plants and
their putative functions in roots. New Phytol 169: 647–666
Romani G, Bonza MC, Filippini I, Cerana M, Beffagna N, DeMichelis MI
(2004) Involvement of the plasma membrane Ca2+-ATPase in the short-
term response of Arabidopsis thaliana cultured cells to oligogalactur-
onides. Plant Biol (Stuttg) 6: 192–200
Rost TL, Jones TJ, Falk RH (1988) Distribution and relationship of cell
division and maturation events in Pisum sativum (Fabaceae) seedling
roots. Am J Bot 75: 1571–1583
Schopfer P (2001) Hydroxyl radical-induced cell-wall loosening in vitro
and in vivo: implications for the control of elongation growth. Plant J 28:
679–688
Schopfer P, Liszkay A, Bechtold M, Frahry G, Wagner A (2002) Evidence
that hydroxyl radicals mediate auxin-induced extension growth. Planta
214: 821–828
Schroeder JI, Kwak JM, Allen GJ (2001) Guard cell abscisic acid signalling
and engineering drought hardiness in plants. Nature 410: 327–330
Shabala L, Ross T, McMeekin T, Shabala S (2006a) Non-invasive micro-
electrode ion flux measurements to study adaptive responses of micro-
organisms to the environment. FEMS Microbiol Rev 30: 472–486
Shabala L, Ross T, Newman I, McMeekin T, Shabala S (2001) Measure-
ments of net fluxes and extracellular changes of H+, Ca2+, K+, and NH4+
in Escherichia coli using ion-selective microelectrodes. J Microbiol
Methods 46: 119–129
Shabala S, Cuin TA, Pottosin I (2007) Polyamines prevent NaCl-induced
K+ efflux from pea mesophyll by blocking non-selective cation channels.
FEBS Lett 581: 1993–1999
Shabala S, Demidchik V, Shabala L, Cuin TA, Smith SJ, Miller AJ, Davies
JM, Newman IA (2006b) Extracellular Ca2+ ameliorates NaCl-induced
K+ loss from Arabidopsis root and leaf cells by controlling plasma
membrane K+-permeable channels. Plant Physiol 141: 1653–1665
Shen HJ, Galston AW (1985) Correlations between polyamine ratios and
growth patterns in seedling roots. Plant Growth Regul 3: 353–363
Takahashi T, Kakehi JI (2010) Polyamines: ubiquitous polycations with
unique roles in growth and stress responses. Ann Bot (Lond) 105: 1–6
Takeda S, Gapper C, Kaya H, Bell E, Kuchitsu K, Dolan L (2008) Local
positive feedback regulation determines cell shape in root hair cells.
Science 319: 1241–1244
Taylor NL, Day DA, Millar AH (2004) Targets of stress-induced oxidative
damage in plant mitochondria and their impact on cell carbon/nitrogen
metabolism. J Exp Bot 55: 1–10
Polyamines Potentiate OHd
-Induced Ion Fluxes
Plant Physiol. Vol. 157, 2011 2179

Tun NN, Santa-Catarina C, Begum T, Silveira V, Handro W, Floh EI,
Scherer GF (2006) Polyamines induce rapid biosynthesis of nitric oxide
(NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol 47: 346–354
Walters DR (2003) Resistance to plant pathogens: possible roles for free
polyamines and polyamine catabolism. New Phytol 159: 109–115
Waring P (2005) Redox active calcium ion channels and cell death. Arch
Biochem Biophys 434: 33–42
White PJ, Broadley MR (2003) Calcium in plants. Ann Bot (Lond) 92:
487–511
Wu J, Shang Z, Wu J, Jiang X, Moschou PN, SunW, Roubelakis-Angelakis
KA, Zhang S (2010) Spermidine oxidase-derived H2O2 regulates pollen
plasma membrane hyperpolarization-activated Ca(2+)-permeable chan-
nels and pollen tube growth. Plant J 63: 1042–1053
Xie LH, John SA, Ribalet B, Weiss JN (2005) Long polyamines act as
cofactors in PIP2 activation of inward rectifier potassium (Kir2.1)
channels. J Gen Physiol 126: 541–549
Zandonadi DB, Santos MP, Dobbss LB, Olivares FL, Canellas LP, Binzel
ML, Okorokova-Facanha AL, Facanha AR (2010) Nitric oxide mediates
humic acids-induced root development and plasma membrane H+-
ATPase activation. Planta 231: 1025–1036
Zapata PJ, Serrano M, Pretil MT, Botella MA (2008) Changes in free
polyamine concentration induced by salt stress in seedlings of different
species. Plant Growth Regul 56: 167–177
Zepeda-Jazo I, Shabala S, Chen Z, Pottosin II (2008) Na-K transport in
roots under salt stress. Plant Signal Behav 3: 401–403
Zhao F, Song CP, He J, Zhu H (2007) Polyamines improve K+/Na+
homeostasis in barley seedlings by regulating root ion channel activi-
ties. Plant Physiol 145: 1061–1072
Zepeda-Jazo et al.
2180 Plant Physiol. Vol. 157, 2011




















