
Oxygen radicals and tissue damage
222
-?-K oxY6Eil RßD|CÊLS RllD T|SSUE DfiÌlÊfiE: T]IE EFFECT OF OXYGEil RRDICRLS O]I T]IE STPUCTURE ffr{D sYilTHEStS 0F }tYRLUR0lllC RCID by JULIÊN DAVID llcNElL H.8.8.5., F,tl.Íl.C.P. Ê Thesls Submitted for the Degree of Doctor of Philorophy in The Unirersity of Êdelaida. Department of Pathology The llnlrersity of Adelalde December, 1986. :.), \ \..1? 1 i\'*i.:... ,! t,d \ ,.\u^o
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of Oxygen radicals and tissue damage

-?-K
oxY6Eil RßD|CÊLS RllD T|SSUE DfiÌlÊfiE:
T]IE EFFECT OF OXYGEil RRDICRLS O]I T]IE STPUCTURE
ffr{D sYilTHEStS 0F }tYRLUR0lllC RCID
by
JULIÊN DAVID llcNElL H.8.8.5., F,tl.Íl.C.P.
Ê Thesls Submitted for the Degree of Doctor of Philorophy
in The Unirersity of Êdelaida.
Department of PathologyThe llnlrersity of Adelalde
December, 1986.
:.), \ \..1? 1i\'*i.:... ,! t,d\,.\u^o

TâBLE OF COTTE;Í5
DECtâRTTIOT
ACßTOTIEDGIIEXTS
5U1,IHâNT
GrosfìâRr
it
iv
vil
I I flTRoDllcTlox AtD lErl Elf¡:-- GaËtal lntroductlon 2'É.
oxuqtl ntdtctlt trú Jol nt Blolog y 4
l. lntroductlon?. Daflnltlonr tú clPmlttrg
t. 9uProxldc tnlonb. HlJdrryn Pcrod&c. Hgdroxgl rtdlæld. Slftgltt oxlsôn
3. Posslblciouræïú oøoryn radlcalt in thc lnlltrrd Jolntt. Tlp r¿lææ sl wW¡ radle¡lr from ætivtttd ptnqocgtts
b. pæsiblc ¡¡¡pn iÁttæl productlon dw to lln grærtnsl rúwl¿¡rts tld trtnsltlonmattl¡ rrithln t]æ lnfltm.d Joint
c. Could cgnovlal iæhæmit lcáO to o(Wn rdictl Prsd'ßllon1¡rtJ3.?4. Protactlvt mæhnitm¡
t. SuProxlü dltmuttætb, Gluttthtonc/gluttthiona Wrwl&rcc. Cattlæed. tllçr dB.lc¡wtc. fl¡<lûnt &lerrrs vlthf n thc Jolnt
5. Ttrgatt for oxg rdiæl tttækô. Protcintb. ilrælcic æld cnd DllA
c. l"tcmbrtna llÍrldtd. Extræcllultr mæromolæulcs otlçr thtn hpluronic æidÊ. Ccllultr targett
| ) Rd ælltÐ 0tlpr ælls
C. Hgtluronic Acidl. lntroductlon2. Chtmlctl rtrrcturc5, Hytluronlc æld ln qæow tolutlon--tho formatlon ú t ¡¡rtrß4. lñpllcttlontlor tymvltl lluld furction
ü. Lubrlætlonb. Yltcælcrtlc aroPrllstc. A codult for nutrlantr
5. Eiotqnthcalr6. Turmvcr7 . Hgtl uronic æid-æll I ntcrætiont8. Sgnovltl fluld hPluronlc acld
2?

D. St¡llæ on (hrgcn Rtdlæl lnducod Hpluronlc Acld hpolUrprlzrtlon 42l. lnltltl tttllæ2. Hlttlon lnduccd hpluronlc æld dcgrtûtlonÍ. fhldatlon- rcductloñ dcpol Umarlætlon (0RD ræctlon)4. Emgmatiællg producad oxpn radlælr tld lnhlbltorr5. lmprtaru of mrltl clrlatorr tnd cnttlnlltmmtorg drqc6. Effæt of suproxl&dhmutæc?. DcArdttlon of hrpluronlc æid bg ncutrophlb8. l'lolæular vcight ch¡rp¿s9. Conclusion
il. TltE DEpotTHEnratTt0x p[oDucTs 0F HTAtuRoilc Æ¡D âFTER EXP0!¡UnE T0 0DrR lf,TITNOFlntrøuctlon 52B. I'lrtarials tld t4cthoú 55
l. Hgaluronlc æld Purification2. 0xggrn radlctl çncrdlonä. Farrour lon tutocldttionb. xÛl¡þt,c. Stlmuldad trlUrprphonrrlær lcucocqtor
!, Vlrcomctrg4. Êal chromatogrtphu5. Uronlc ætd tnal¡pit6. Amlgtictl ultræcntrlfugttfon
C. Ræultc 57l. Visco¡ltu2, Ì4olccultr cxclurion çl chromctogrtphg
t. Scphtdcx f¡- 100b. $ptnræc 48c. *glnrw 2B
3. Amlgtictl ullrlrn¡ûriitrytionD. Discusrion 67
?2E. Summarg
ilt. GHAIGES tt TltE ]to[EcutâP tfEtGHT SPECTRâ 0F STf,fHlAt rtulD Hrâtumtlc tclDSEET IT ITTLAHHâTOPT.NITT D|5EâI¡Eå. lntroductlon 75B. llttaritls and mctfre& 77
l. SgnMtl fluidZ. Hgaluronic æid reptrttion5. Eclluloæ tcctata trl clcctrophoræit4. Gcl chromatogrcphg5. Uronic æid tmlUsis6. 0xgçn radictl çncrationRæultsDiæuæionSummrg
IY. I,IEASUREI'IE]IT OF REDIICIlG E]ID GROUPÍ¡ GEIEPATED 8I EXPfT!¡UTE OF HYAI.URO]IIC
ACID TO OXTGE]I RâDICâU¡A. lntroduction 93B. Hdcrltls and mattpdr 94
l. 0xggcn rdlctl çmrttionË. Fcrrous lon ¡utoxlútlonb. xDlJ#
c.D,
E.
82889l

2. Erugmtlc dcgrútlon of hçluronlc ældÊ. Tætlcultrhpluronlfutb. Strcptomgæs hgtluronldæt Ûlçttlon
5. Gcl chromttognPhg4. Colorlmatrlc $ttlp
Ë. Uronic ældb. Ptrk-Johmn c$tgc. l'lorpn-flæn ruætlon
5. C$nlü lcbtlllm6. Atccnding ptpr chrorutagruPhU
C. RtsultsD. DlæussionE. Sumrurg
T. DOES ÍITTGET NTDICâI EXPÍËURE âITER THE âBILITT OF HTâLUROTIC âCID TO
l'l0DULtTE I{åER0PHåGE Fc REGEPT0R BltDltG?A. lntroduction I 08B. lttcritls and methods 109
l. Adhcrcnt monæUtæ?. Fc rcccptor æsag
t. Praptrttion of antiMt¡ cottcd red blæd ællsb. Fc rcccPtor asstgc. Sttining tnd æuntiq
3. Hgtluronic æid prcparttiont. 0xr¡gcn radictl cxposed htpluronic æidb. Enzgmc cxPoscd hgaluronic æid
C. Rrsultr lllD. Discussion ll4E. Summtrg ll?
TI. ETPOSURE OF CUTTURED STMTIåI GETLS TO ONTGET RTDIGâI FTIHE!ì. ETMTüßIGITTâTD EFTEGT OlI IITAIUMXIG TCID PRODIETIO]IA. lntrodrætlon I 19
B. Matarlals and l'tthdr t l9l. S¡novltl æll culturt?. Chromium rularstæu¡3. fhgçn rtülctl çncntion4. Hgtluronic æid Prodtrtion5. Ctllulæa tætttu çl tlætrophor¿sis
C. Ræults I e5l. N0/HX llductd cgtotoxlcitg2. Effcct of oxgçn raüictl cxpoturc on lH production
D. Discussion I 5eE. Summaru I 36
il t.GEIERAI- DISCUSSIoX
APPEXDICESI Dctail of thc Colorimetric æsôUlll Titsue Culture l.1eúia
lll Publicctionr
9?t04t06
137
t49152r53
t?0BIBTIOGRAPHT

I
DECLRRRTIOII
Thls thesls contalnr no materlal uhlch hac bcÉî accepted or
submltted for the auard of any other degnee or dlploma ln any
lJnlrerslty. Furthumore, to the best of my knouledge and bellef, thls
thesls contalns no matenlal prerlously publlshed on r¡rltten by another
pÊnson, except uhere due referÊnce ls made ln the text.
The r*¡ork descrlbed hereln has been the subJect oî the follorrrlng
publlcatlons:-
HcÌ{ElL, J.D., tdlEEKltl, 0.1d., EETT9, ld.l'|. and CLELÊÌ{D, L.E. (1985)
Depoþerlzatlon products of hyaluronlc acid after Êxpoiureto oxygen derlved free radlcals.Ênnals of the Rheumatlc Dlseases 11, ?80-?Eg.
McllElL, J.D,, lrJlEEKll{, 0.hJ., CLELÊIiD, L.6. and 7ERN0i{-R0BERT3, E.
( 1986). The generation of reduclng ends by exposure ofhyaluronlc acid to oxygen derired free radicals.Êgents and Êctlons Supplements 18, 95- lg l.
and abstract¡:-tlcNElL, J.0., lrjlEEKll{, 0,lrJ., CLELÊHD, L.E. and EETTS' hj.ll. ( lgBl)
Ênalysls of hyaluronlc acld degradatlon products afterexposurË to an oxygen derlved free radical flux.Connective Tissue Research I l, 25A.
Hct{ElL, J.D., kJlEEKllt, 0.kJ., ELELAiID, L.E. and EETT3, [d,H. (1983)Hechanlsms of hyaluronlc acid degradatløt by oxygen derlvedfree radlcals. Aust. N. Z. J. Hed. 13, 2l l.
tlcHElL, J.D., CLELÊND, L.E. and EETTS, lrJ.ll. (1983)The cytotoxlclty of oxygen derlrred frce radlcal¡.Êust. ll, Z, J. Hed. l\ 225.
HcllElL, J.0., [ÂllEEKllJ, O.bJ., and ELELÊIJD, L.E. (1981)Ênalyrls of the molecular uelghts of hyaluronlc acld fromlnflamed and non-lnflamed synovlal flulds.Êust, ì1. Z. J. Hed. l{ (suppl, l'), 313.
llcNElL, J.0., lrJlEEKltl, 0.UJ., and CLELÊHD, L.6, (tgS5)The cytotoxic effect oî oxygen dertved lree radlcals on borinesynovial cells in culture. Aust. 11. Z. J. 357.
Slgned:J. D. HcHeil
December, l9EE

1i.
ßCKll0l¡lLEDGllEllTs
The urork degcrlbed ln thlr thesls uras :upported by grantt from
the Natlonal llealth and lledlcal Research Councll of Rustralla and
from an E. R. Dau¡es fellor¡shlp from the Royal Adelalde llospltal.
I hereby ulsh to thank my supÊrylsors Dr. Ole lrliebktn, Dr. Leslie
Cleland and Professor Barrle Pernon-Roberts. Dr. Ole lrjlebkln has
helped me r¡lth day to day teachhngr guldance, enthuslasm and
frlendshlp. Dr. Leslie Eleland has lnfluenced me touards a càreer
lnrolrlng laboratory resÊàrch and has provlded support, raluable
coun¡el and frlendshlp. Professor Earrle /ernon-Roberts has prorided
orzerall superrlslon and advlce and has provlded all the facilltles of hls
department for thls uork. I also r¡lsh to thank Proîessor J. L.
Skosey rr.rho provlded valuabla supervision and edltorlal as¡lstance
durlng the r¡rltlng up phare.
0ther people uho hare helped mË ärÊ Dr. ld. llenry Eettc urho
prorlded teachlng help and as¡lstance r¡lth the chemlstry of oxygen
radicals, Dr. Stephen Mllazzo r¡ho provlded rraluable interect, support
and cltnlcal teaching, llrr. S. C. lrJlabkln rr.rho provlded agglstance in
tcchnical matters, l1?s. Eenevieve LaPinska r¡ho provided uilling help
ln typlng thls manuscript, I1n. Dayid llaynes for help r¡ith the
macrophage Fc receptor binding àssåpr and Hr. Dale Carille r¡ho
contributed of his photographic talents. I haye also had raluable
discussions r¡ith Dr. lllchael lrJhltehouse for r¡hlch I am grateful.
I am also indebted to the South Êustrallan lleat Corporation
for thelr glft of bortne hocks and Pharmacla, South Seas, for thelr
glft of llealon.

111.
I r¡ould llke to make a speclal trlbute to my late father and my
mother, both of uhonr hare provlded me' orer many y¿a?\ r¡rlth an
lnexhaustlble supply of encouragËmÊ-nt and support. Ftnalþ' I t¡ould
llke to thank my ullfe, Jane, r.rho has patlently cndured so much so
that I could undertake thls proJect.

iv.
SU}IIIRRY
ln order to examlnc the role of oxygÉTr radlcalc ln lnflarnmatory
Jotnt damage, a study of oxygen radlcal lnduced hyaluronlc acld
depolymerlzatlon uas undertaken. Hyaluronlc acld u,às uscd because
It ls a maJor extracellular compon"nt of s¡moylal fluld and the
s¡morlal membrane and a prlme secretory product of s¡rnorlal cells.
Êlso lt ls kroum to be sensltlve to oxygen radlcal inducad
depol¡merlzatlon and ls altered ln lnflammatory Jolnt dlsease.
The methods of approach used uære:
a. Ênalysls of the products of oxygen radlcal lnduced hyaluronlc
acld depolymerlztlon h rltro to determlne changÊ ln molecular
r.r.lelght, generatlon of reduclng c,nds and, to a tlmlted extent, change
ln blologlcal propertle¡.
b. Synorlal fluld hyaluronlc acld from lnflamed synovlal flulds rr.ras
isolated and the spectrum of molecular rrrrlghtg idËïtfled in order to
make comparisons urlth those obtalned E yJl¡4.
c. The effect of an oxygen radlcal flux on s¡norlal cells ln
culture uas determlncd and the effect on hyaluronlc acid produced
by thesc cells r¡as examlned.
Ênaþsls by gel chrornatography of oxygen radical induced
hyaluronlc acld depolymerlzation products revealed polydlsperslty tn
sizÊ. The smallest molecutes detected had a molecular rlelght of lla,¡¡hlch ulas not reduced further by exposure to a second oxygen
radical flux. Conslstentþ a relatlyely rapld transition uJàs seÊn from
large to small materlal suggestlng àn ondered element to the
breakdoum process.
Ênaþrls by analytlcal ultrccentrlfugatlon conflrmed the
decrease ln slze of hyaluronlc acld due to Ëxpoiure to oxygen
radlcals; houerer, thls procedure reyealed that those ramples of

v
louest vlscoslty dld not exhlblt the louest sedlmentatlon values, ¡
posslbþ reflectlng oxygsn radlcal lnduced repoþerlzatlon. '^v ") "'"r; ")
Oxygen radlcal lnduced hyaluronlc acld depoþcrlzatlon lg
Ëccompânlcd by the generatlon of rcduclng cndr ât mcåtured by
both colorlmctrlc assayg and cyanlde labellng, There ls, houerÊr' no
detectable lncreage ln llorgan-Elson reactlvlty thur lmplylng that
the reduclng ends released ärË prÊlent on the D-glucurøtlc acld
corriFonÊnt of the hyaluronlc acld product. Thls ls therefors evldence
that lt ls the Þ | -3 glycosldlc llnkage that ls surceptlble to clearage
by oxygur radlcal¡.
Fractlonatlon oî s¡novlal fluld hyaluronlc acid from patlents
r¡.rlth both o¡teoarthrltls and lnflanmatory arthropàthlcs revealed
uronlc acld contalnlng materlal that u.rag retarded on Sepharose '18-
EL chromatography rn contrast to non-arthrltlc cadaver subJects in
r¡hlch cäsÊ all s¡novlal fluld hyaluronlc acld uas excluded on
Sepharose 'iE-CL. ln the patlent group no materlal u,lar present
belou a molecular r.uelght, of 2 x l0a and no correlatlong could be
made betuccn the amount of uronate retarded and cllnlcal lndlces of
inflarnmatory actlvlty. Extracted cadaver synovial fluld hyaluronlc
acld ¡¡as susceptlble to o,(ygcfi radlcal lnduced depoþarlzation þrlLÊg.
0xygen radlcal exposed hyaluronlc acld had the rêmc effect on
Fc receptor blndtng a5 non-oxygen radlcal exposed hyaluronlc acid of
the same molecular r.uelght. Thus, at least ln thls context, no
alteratlon tn the propertlec of hyaluronlc acld due to oxygcn radlcal
ÊxposurÊ uas dc'rnonstrable beyond that due to decreaslng molecular
u.relght.
Êppllcatlon of än oxygÊn radlcal flux, produced by the actlon of
xanthlne oxldasc on hypoxanthlne, ls cytotoxlc to cultured borine

vf
t!,norlàl cells. Êdmlnlstratlon of catalase, but not supenoxlda Lqa(L-
dlrmutase, ls protectlye thus lndlcatlng HzO¿ ls the agent nÊc"ssäry i.;';'" ,
for toxlclty. Sublethal oxygen radlcal fluxes r¡¡lll result ln tha
cessatlon of hyaluronlc acld s¡rnthesls before the cessatlon of
s¡.rnthesls of other glucosamlne-contalnlng products. No lor¡
molecular r.lelght hyaluronlc acld pnoducts could be detected ln the
supÊrnatants of oxygen radlcal exposed cells. These flndlngs mply
that the lor¡ered molecular r.lelght hyaluronlc acld seen ln
lnflammatory Jolnt flulds ls dut to extracellular depoþerlzation.
These results prorlde furthen erldence for the role of oxygËn
radlcals as agents of hyaluronic acld depol!¡rnerlzation þ ¿Ug.

vi1.GLOSSARY
âDP - adenoslne dlphosphateÊïP - adenosine trlphosphateEPS - bathophenanthrollne sulphonateConA- concanavallnÊCTÊE - cetyl trimethyl ammonlum bromideCSb - the f¡fth component of complement (actlrated)DETÊPÊC - diethylenetrlamine pentaacetic acidEDTÊ - ethylamlnedlamlne tetnaacetic acidFe (ll) - ferrous lonFe (lll) - ferric lonFllLP - formyl methlonyl leucyl phrnylalanine6ÊG - glycosaminoglycanGSI'I - reduced glutathioneIìSSE - oxidized glutathionellÊ - hyaluronlc acidl'lBZ - peroxyl radicalKay - partltlon coefflcient descrlbed by the fdrmula:-
Kay= Pe-Po
Vt-VoHsD- molecular ureight mËäsurÊd by sadimeryrtation-dlffu:ion
t{ÊD - nicotine adenine dinucleotldeNÊDPll - nicotlne adenine dinucleotide phosphateNr - relative vlscosltyNrp - specific vlscoslty0RD - oxygeî radical depolymerlzationO2- - superoxide anion
0ll. - hydroxyl radicalOCL-- hypochlorous lonùBZ- delta singlet oxygen
ZEZ - sigma singlet oxygËn
Pl'lÊ - phytohaemagglutininPllÊ - phorbol myristate acetatePPD - purified protein derirzatirzeRPlll - tissue culture medium [lhJÊKÊTÊ and 6RÊCE, 1961]500 - superoxide dismutaseUDP-tìlcÊ - uridinedephospho-D-glucuronic acidUDP-GlcNÊc-uridinedephorpho-N-acatyglucosamineUV - ultrarioletVe - elutlon volumeVo- vold volumeVt- total rolumeXOlllK - xanthine oxldase/hypoxanthineXO/X - xanthlne oxldaselxanthine

1
C]IfiPTER I
ßTRoDUCTloll RllD RElrlEtd

¡¡
R. 6E]IERRL I]ITRODUCTIO]I
Dlarthrodal Jolnts àre speclallzed connectlve tlssue structuras
that haye dereloped to allou moblllty for an organlsm r¡lth a rlgld
skeleton. The surfaces of the artlculatfng bones àrê corered r¡lth
hyallne cartllage and the bones are unlted both by a fìbrous capsule
and by llgaments. ln addltlon, ovÊrlyins muscles and tendons con-
trlbute to the stablllty of the Jotnt. The lnner surface of the Jolnt
capsule ls llned by thc rascular s¡rnorlal membrane, thus formlng the
Jolnt spac? r¡hlch ls totalþ enclosed by synorlal membrane and
artlcular cartllage. The JoÌnt spàcË contalns a small amount of ¡otnt
fluld r¡hlch ls broardly slmllar ln composltlon to blood plasma to r¡hich
has been added hyaluronlc acld IHcEÊRTY, l9E5]. llyaluronlc acld ls a
secretory product of the synorial linlng cell. lt imparts to s¡rnorial
fluld a dlstlnctlrre high rlscosity and contrlbutes to tha biologlcal
functions of s¡rnorlal fluid. '
Joint tnflammation results in characterlstlc changes in all of the
above-mentloned anatomlcal structures, leading to damage to the
rarlous components of the Joint and ultlmateþ to Jotnt deformity
and loss of Jotnt functlon. Earþ tnflammatory changes are seen ln
the s¡rnoylal mambrana and are rÊry soon reflected ln the s¡rnorial
fluld, uhich becomes both lncreased ln volume and ls often much less
r¡lscous than normal fluld.
A r¡lde varlety of effector agents havs beer examlned to
determlne the blochemlcal mechanlgm¡ lnvolv¿d ln the loss of
structural lntegrlty of Joint tlssues. Those examined most closely
havc been ËnzymÊ systemsr prlnclpally acld proteares and neutral
proteases IEÊRRETT and 9AKLÊTVÊLÊ, l9E5l. lt ha¡ become clear from
these studles that ulthln the tnflanrmatory proccli there arÊ a
multltude of effector systems (including effectors, their precursors,

3
actlråtorr ând äntagonlsts) that can contrlbute to lnflanrmatory
Jolnt destructlon.
One such system that has come to llght ln the last l3 years
lnvolves the productlon and release of orqgen radlcals by actlrated
inflarnmatory cells. Êlthough the exlstÊnce of oxygen radlcals in
chemical systems has been appreciated for ove? 100 years, until
recently they urÊrË thought to be too actlve to play a rolc in
biologlcal processes. lt is nou clear that stlmulated phagocytes, tn
partlcular poþmorphonuclear leukocytes, undergo a "respiratory
burst" r¡hich ls characterized by lncreased oxygen uptake,
chemiluminescence, lncreased hexosË monophosphate shunt actlvlty
and production of the superoxldÊ anlmr. Superoxide anions dlsmutate
either spontaneousþ or catalysed by th¿ enzpe superoxide
dismutase to form hydrogen peroxlde. Superoxide anlons may react
uith hydrogen peroxlde to produce slnglet oxygen and the hydroxyl
rad¡cal. These radlcals ârÊ iËcreted lnto the phagosonre and are
utlllzed tn mlcroblcldal kllllng. llydrogen peroxlde is a substrate for
phagosomal myeloperoxidase r¡hich produces hypochlorlte ions, r¡hich
also hare potent oxldant actirlty. 0xygen nadicals also escape into
tha extracellular space t.lhere, in the absrnce of complete sca-
renglngr adjacent host components may be exposed to their effects.
The intra-articular space of inflamed joints prorides an important
site to examlne the effects of oxygen radicals fon reasons outlined
orlginally by tlcE0RD I l9?{]. Flrstþ, lerels of endogenous scapengers
of oxygen radlcals ln s¡,movial fluid are lou. Superoxide dlsmutase,
catalase and the glutathione/glutathione peroxidase system are
predomlnantly intracellular and thelr lerels in extracellular fluids in
general, and synorlal fluld ln pärtlcular, a?e ;,cveral orders of mag-
nltude less than lntracellular levels [l'fcCORD, 1971i ELÊKE Êt Al.,

4
lgE lJ. Secondly, hyaluronic acld hag beÊt shoum to be surceptlble to
the effects of oxygen radlcals ln ways that parallel the changer reÊ'Tì
tn lnflammatory Joint dlsea:e, that ls, lou.rered vlscoslty and a de-
crÊâse ln the areragÊ molecular u.relght.
ln order to elucldate the role of oxygÊn radlcals ln lnflam-
matory Jolnt dlsease, I harre choscn to examtne in detail the effect
of oxygen radicals on the structure, propertle¡ and cellular pro-
ductlon of hyaluronlc acld þ tzltro, and to determlne r¡hether any of
these changes can be correlated ulth the changer found ln fnflamcd
s¡,morlal fluld hyaluronlc acld. The results of thege studleg form the
basl¡ of thls theslg,
E. 0xYf¡Elt RADTCßLs ßltD J0tltT 8t0L06Y
l. lntroductlon.
0xygen has been present ln slgnlficant quantltles in the earth's
atmosphere for an estimated 2.5 billion yearl, its appearâncË cor-
responds to the ËmËrgÉnce of the flrst photosynthetlc orgánlsms
IBERKNER and HÊRSI.|ALL, l9E'f]. Since that time the atmospheric
oxygËn concentration has gradually increased and lifa forms haye
dereloped that are dependent on aeroblc metabolism. Concurnantþ
there has bcen an erolutionary pressurÊ for these organlsms toderelop effectlve defenses agalnst the toxlc lntermedlates that are
formed during the reductlon of oxygen to uatar TFRANK and Ì1ÊSSÊR0,
l980l. Host of the oxygen consumÊd by resplnlng cells ls accounted
for by the mltochondrlal cytochrome C oxldase system [ÊNTON|N| Êt
d., | 9?0]. Thls system effects the tetrayalent reductlon of oxygen
to r¡ater r¡rlthout the nrleasc of eithar superoxlde or peroxlde
IFRlD0PlCll, lg?BJ. llor¡eyen thæe are blological processÊs thatproduca superoxlde anlon, and effectlve cellulan antloxldant defense
mechanlsms hare ¿volved to protect the cell from these reduced

Ð
forms of oxygen ISIES and CÊDE|'|AS, l985l. lronlcalþ, lt r.uas the
elucldatlon of the functlon of one of these antloxidant enz¡nner,
superoxlde dlsmutase [llcCORD and FRlD0PlCtl, l96El, that led to aulde appreclatlon of the role of partlally reduced forms of oxygen (as
oxygÊn radlcals) ln blology and thelr posslble pathoge¡etlc role ln a
number of dlsearË procËlrËt.
2.
Ê frer radlcal mày bc dcfined as any ipcclel capable of lnde-
pendcnt cxlstencc that has one or morÊ unpalred clectrong
II'IÊLLIIJELL and GUTTERIDGE, l9E5J. ln general, the preic'rìcË of an
unpalred electron accountg for thc high reactlvity of radicals. As
stated, thls deftnitlon lncludes the hydrogen atom and most
transltlon metals. Radlcals may be formed f a covalent bond,
conslstlng of tuo shared electrons, ls spllt such that one electron
stays r¡ith each atom (homolytlc fission). This may be considered an
"lnltlatlon" reactlon. lndirldual radlcals mày react u.lith non-radlcals
r¡lth the formatlon of another radlcal specles (propagatlon reactlons)
or tr¡o radlcals mäy react together r¡ith the fonmation of a non-
radlcal product (tarmlnatlon reaction).
The ground state oxygèn molecule ltself mày be consldered a
dl-radlcal slnce lt has turo unpalred electrong each located ln a
different r antibondlng orbltal. Eoth of thcse electrons hare the
râmê spln quantum number. Thus, rl oxygen atte'rnpts to oxidize
another atom or molecule by acceptlng a palr of electrons from it,both neu electrons must be of parallel spin so as to enter the
racancies tn the unpaired ø orbitals [HÊLLltdELL and EUTTERIDGE,
198{a]. Thls restrictlon on oxldatlons oî oxygur dictates thato,rygen ulll tend to accept electroni onÊ at a tlme, 1.e., uÍtlvaleryrt

6
reductlon, räthËr than to be dlrectly reduced to tlater by
tetravalent reductlon.
e- e- Ê- e-02>
+211+
The unlralent reductlon of oxygen leads to the productlon of three
maJor oxygÊn-centred radlcal lntermedlates r¡hlch are dlscussed
belor*l:
a. fuFtr¡¡ldlÊulctrhJhen oxygÊn accepts a slngle electron lt ls converted to the
superoxlde anion (Br-) or its protonated form, the hydr operoxyl
radical (ll0z.). ln aqueous solution at ä pll ol 1.Ê the turo forms are
present in a ratlo of l:1 and at physiological pll oriy l% is present in
the pronated form IFEE and I/ÊLEHTII{E, 197?]. Superoxide aníon
hor¡erer disappears from aqueous solution via a dismutation
reactlon, uhlch proceeds.
H0r' * Oz + ll+ -----) llr!, + E"
This reaction may occur :pontaneousllt in r¡hich ca:,e it uill occur
most rapidly at an acidic pll (due to the higher concentratlon of
l'102.). lt may also be catalyzed by the enz¡tme superoxide dismutase
[llcC0RD and FRlDlVlEll, lgEE], u¡hich is activ¿ over a r¡ride pll range.
b. Hyircgen Peroxide
Êdditlon of tuo electrons to molecular oxygen r.rllll lead tothe formatlon of the paroxide ion (0.2-). Tha pK¡ of the reactlon
describlng the equlllbrlum betr¡een 0.2; and llr0, is so high that atphysiological pH rzlrtually all ls present as llr0r. Sereral ?nzymesr
e.9., glucose oxldase, ârË capable of the direct dir¿alent reduction of
0, independent of the dismutation reaction [FRlD0PlCll, l9?E].
?'t

7
G.@Ê mcchanlsm r.,.lhereby a three electron reductlon product of
oxygÊn could be formed from H"E" uas flrst propored by IIABER and
hJElSS [193{]. Houerer the rate conctant for thls reactlon ln
âquÊoui solutlon has been ¡horrrn to be vlrtually ze?o [llÊLLIUJELL,
tgB l]. Thls ls not the caiÊ hor¡erer lf traces of a transltlon metal
âre prÊsent, in r¡hich case a Fenton reaction ls possible, i.e.,
I Fe(lll) + Az- Fe(ll) + gz
2 Fe(ll) , HzOz >
Nett ReactionFe++ soln
0i * ltrg z -----> 0z + 0l-1. + 0l'l -catalyst
5 lron catalyzedHaber hJei¡c reaction
Sincc the sum of reactions 1 & 2 is the ¡ame ai the l'laber lrjeisE
reaction this reactlon is al¡o knor*m as the lron-catalyzed l'laber
l¡Jeiss reactlon. Thls reaction r¡lll be promoted if the iron ls äppro-
priately chelated, Ë.9., by EDTÊ []1cCORD and DÊY, l9?E; EETT!ì and
ELELÊND, l382l. 0n the other hand chelators uith à very strong
affinity for Fe(lll) do not allou Fe to shuttla betr¡een Fe(lll) and
Fa(ll) and therefore inh¡bit the reaction [6UTTER|D6E, RlCllH0llD and
I'|ÊLL||¡JELL, l9?91. Physiological chelators such as ÊTP or ÊDP can act
as iron chelators and support the superoxide-driven hydroxyl radical
production IFLOYD, I gE3] at physiological iron concentrations Ín the
absence of EDTÊ IFLITTER, R0hJLEY and l-lÊLLIIJELL, l9E3]. The
hydroxyl radical is extremely reactive and u¡ill react r.rith any
molecule uithin the immediate vlclnity of lts productlon in contrast
to 0r- and llr0, r.,.rhich are relatlvely stable and can dtÍîuse away
from thelr site of productlon IÊNEAR and NETÊ, 196?J.

I
d. Slnglat 0xFgen
Slnglet o,rygcn ls a hlghþ reactlve form of oxygen that ts
formed uhcn molecular oxygÉn absorbg sufflclÊît cne?gy to cause a
shlft of one of lts tr.rlo unpalrcd electrons of slmllar spln to an orbltal
porltlon of hlgher lin¿?gy, Thlg ls agsoclated r¡lth the lnrzerglon of spln
of one of the electrong IKASHA and ERABHÊH, 19?91. lf the excltcd
electrons form a palr then lt ls deslgnated lAgO, (therefore, by
deflnltlon not a radlcal) and lf the electront are not ln the Jäme
orbltal 3Ig0r. Slnglet oxygÊTr ha¡ been reported to be produced är âresult of the non-catalyzed dlsmutatlon reactlør and durlng the
Haber lrJelss reactlon [KELL066 and FRlDOVlEll, l9?5J. ln addltlon, lthas been reported to be produccd by the myeloperoxlde-Hr0r-hallde
system of neutrophlls [R0SEtl and KLEBAT{0FF, l9??1, uhere tt ls
bellered to be produced by the lnteractlon of the hypochlorlte lon and
H.o.'
3. Posslble Source¡ of 0xpgËn Radlcals urlthln an lnflamed Jolnt
ln addltlon to the general exposure to oÅygen radlcalc that any
cell exlgt¡ng ln Ên oxygÉTr contalnlng enrlronment must endure, (for
rerleu¡s see; llÊLLlldELL and IìUTTERIDGE, l9E5; FRlDOTlCll, l9?E), cells
and extra-cellular componentg of thç lnflamed Jolnt ärË Êxposed to
oxygÊn radlcals at å result of lnflanrmatlon. Êg a baslc for dlcusslon of
these erents, lt is approprlate to conslder brlefly lromË lnflanmatory
mechanlsms that ulll lead to .Jolnt lnflanrmatlon.
ln some sltuatlons the lncltlng agent ls obvlous, îor example, a
bacterlum ln the case of scptlc arthrltls or ân lrnmune complex ln
assoclatlon r¡ith ä strum sickrress reactlon. llorË conrmonþ, horr.rerer,
the nature of the tncltlng agent ls unknourn. Nerertheless lt ls likely
that the tratn of events slmllar to that outllned by ZVAIFLER I l9?5]
for rhcumatold arthrltls ls set ln motlon. lle proposed that the

9
lncltlng antlgen becomes locallzed ln the artlcular carlty and
stlmulates the productlon of antlbody. These combtnt tn the s¡rnorlal
fluld or the synorlal membrane and actlvate the complement
pathuay thus generatlng chemotactlc agents, partlcularþ CSb.
Pol¡nnorphonuclear neutrophlls r¡lll then accumulate, partlcularly
r¡lthln the s¡morlal fluld, and along r¡lth type Ê s¡rnoriocytes, r.uill
ingcst the fmmunË complexes. ât the time that thls theory uJàs
flrst proposed lt r¡as bellered that lt r¡as the hydrolytlc Ênzym?s
that ur?rÊ released from þsosomes durlng the act of phagocytosls
that accounted for the pnollferatlve and destructlve changes
characterlstlc of rheumatold arthrltls [ldElSSllANNr ZURIER and
H0FFSTEIN, t9?2], lt has nou becomê âpparent that oxygÊn radicals
måy also be rehasrd at thls stage [l¡JElSSllÊNN el il., 19?91 and may
contrlbute slgnlflcantly tn the genesls of these changes,
a. @en radicals from actiyated phagggl¿l!å.
Phagocytes become actlyated on contact r¡ith particulate
stlmull, such as fmmune complexÊs or bacterla that have been
opsonlzed (1.e. coated r¡lth host-derlred proteins) [R00T and
HETCÊLF, l9??]. Several soluble factors may also lead to phagocyte
actlyatlon. Êctlration tnvolves the "sr¡itchlng on" of a number of
metabollc processes, the end result belng that the partlcle ls
engulfed lnto an organelle called a phagosome. Eranules present in
the cytoplasm of the phagocyte fuse r¡lth the phagosome and
dlscharge thelr contrnts lnto lt (degranulatlon) to effect bacterial
kllllng and/or degradatlon of the lngested materlal. BÊB|0R, KIPNES
and CÊRIIUTTE I l9?5] urÊrË the first to shor¡ that lncluded ln the
procËss of actlyatlon ls the nelease of superoxlde anion. This ¡s
formed às ä componËnt of the "resplratory burst" of thr neutrophll
r¡hlch lnvolres :

r0
l) increased O" uptake l¡hlch ls lndependent of mltochondrtal
resplratlon ISEÊRRA and KÊRi|OPSKY, 19591.
2) lncreased glucose oxldatlsn vla the hexorÊ monopho:phate
shunt r¡lth the resultlng productlon of i{ÊDPH IIYER, |SLA]I and
ouÊsTEL, t9EIl.
3) Êctlratlon of an llADPll oxldasc IEÊEl0R, l9?EJ, uhlch csnverts
molecular oxygÊn to superoxldc anlon according to the reacflon ¡
2llADPtl +?Oz---) ZHÊDP+ *2E;+ jtzl
Thls tnz¡mc lg sltuated on the lnner rurface of the plasma
membrane and accepts t{ÊDPll from the cell cytoplasm; lt also accepts
oxygËn and releätÊs ruperoxlde anion from the outer ¡urface of the
plasma membranr [BÊE|0R Êt Ê!., l9E I ]. This mechanicm Érrsures thatsuperoxlde anion r¡ill be generated and deposited on the in¡ide of the
phagosome as the plasma membrane inrzaglnates durlng the cellular
ingestion of a partlcle. Houerer it is clear that not all the
superoxlde produced ln this uay ls contalned by the phagosomÊ
[lrlEl35t1ÊllN Êt å1., l9?gl, posslbþ because the enzyme ls acUvated
before the phagosorrc has been sealed, but alco becauge ënz1yt"r,C úr
contlguous årcås of mcrynbränê ls also actlrated. Also, r¡lthln the
Jolnt' lt lr clear that lrnmuna complexes mäy adhere to gtructural
componÊnts, Ç.9., collagen, leadlng to a sltuatlon of 'Trustrated
phagocytosls" r,.rhereby phagosomal cørtentr, lncludlng superoxlde
anlon, cän Êrcäpe freely lnto the extracellular mllleu ÍZVAIîLER,
lg?51. l¡Je hare recently obtained somÊ evidence that this may
occur ulth particulate stimull in the prÊsencÊ of hlghly polymerized
hyaluronlc acid [t1cHElL, Cl-l0lrJ and SKOSEY, l9E6J.
The ability of an organic system to be able to generate as
hlghþ reactlve an agent as the hydroxyl radical r¡as flrst lnferred by
EEÊUCHAHP and FRIDO]/lCll IlS?0], uho exposed the thtoether

11
methlonal to the xanthlne oxldase/xanthlne oxygÈn radlcal
generatlng rystem and detected ethylene. Slnce thls could not be
accompllshed by the actlon of elther superoxlde anlon or hydrogen
peroxlde alone, they postulated that hydroxyl nadlcals urÉrË
produced. tlydroxyl radlcals hare been detected ln suspenslons of
actlvated phagocytes, also by the pnoductlon of ethylene from
mcthlonal ITÊUBER and EÊB|0R, l9??]. Tha most lilreþ sequence of
events ls that superoxlda anions produced by the phagocytes
undergo dlsmutatlon to form Hr0, and, ln the presÊnc? of lron or
another transltion metal às â cataþst, hydroxyl radicals are formed
by the iron-catalyzed Haber l,rJelss reactlon.
Eridence for the extracellular generatlon of thls agent comes
from the experlments of SÊL|N and HcCORD [13?5] uho shor'.red that
the auto-inactlvatlon of stlmulated phagocytes can be averted by
addltion of superoxide dlsmutace, catalase or Ol1. scavångers to the
suspenslon medlum. Thls suggests that the componcnts necellaîy
for hydroxyl radlcal productlon are released lflto the extracellular
mllleu by actlrzated phagocytes r¡here they can medlate cytotoxlc
and other effectc.
Ehronlc granulomatous dl¡ease uhich lg due to a non-functlonal
NÊDPll oxldase, provldes lnslghts lnto the lmportance of orlygen
radlcals ln mlcrobial kllling. At least tuo lnherlted forms of chronic
granulomatous disease exist. Eiochemlcalþ, the tuo forms of the
disease demonstrate an inability of neutrophils to effect a
resplratory burst. Thls manifests clinlcally as persistent infections
rr¡ith sereral non-peroxlde producing bacteria, typlcally staphlococcus
aurÊus. lrJlthout the production of superoxlde anions, hydrogen
peroxlde ls not arallable to the phagocyte and both hydroxyl radlcal
productlon and hypochlorous lon productlon rla the myeloperoxldase

LZ
Ên abnormallty ln lron utlllzatlon has long been appreclated
ln actlye rheumatold arthrltlg. Thls manlfests cllnlcally ar anaemla,
r¡hlch ls assoclated r¡lth a lor¡ sËrum lron and à concomltant raised
sËrum ferrltln INASC|OL| and ELÊCKEURN, l9B5]. The mechanlsm
äppËårr to be related to an lnadequate release of lron from
retlculoendothellal cells IBENNETT, l9??].
NUIRDEH and SENÊT0R [1966] have demonstrated that iron
deposlts rrËrË a constant feature of the histopatholog¡l of the
rheumatold synovlum. Electron mlcroscopy of the s¡morial
membranes of a number of thelr patients shor¡ed that the iron r¡as
present both as ferrltin and haemosiderin IHUIRDEN, 1966].
Speculatlon exlsts as to urhether the accumulation of thls lron is due
to local factors, l.e. repeated eplsodes of mlcrobleeding consequent
upon an abnormal microvasculature, or u.lhether there is a
generallzed dlsturbance of lron storage due to chronic systmúc
dlsease [0GlLPlE-llÊRRlS and F0RI{ÊS|ER, 1 g80l.
Êftcr abrorptlon from the gut, lron, ac Fc(lll), ls transported
in the plasma bound avldly to the camler protein transferrin.
Lactoferrln, prerent ln sereral body fluids, ls a protein similar to
tranferrln ln that tr¡o moles of Fe(lll) are bound per mole of protein.
Transferrin blnds to receptors on the cell surface and can br
internallzed. lrJhen the cytoplasmic yacuole so formed becomes
acidified, the lron ls released and may become bound to yanlous
cellular constltuents such as cltrate, ÊTP and 6TP forming a small
pool of non protein-bound iron. lrJ¡thin the cell, iron is stored in the
protein ferritin, uhlch comprlses a high molecular ueight protein shell
surrounding an hydrated Fe(lll) oxlde-phosphate complex. lron
enters femltln as Fe(ll) becomlng oxldlzed by the proteln to Fc(lll)
and deposlted tn the lnterlor; conversÊly, ln order to be removed,

t3
Fc(lll) naeds to be reduced to Fe(ll), r¡hlch ls believed to be
accompllshed by blologlcal reducfng agents such .ts àscorbate,
cystetne or reduced flavlns. Lysosomal degradation of ferritin leaves
the tnsoluble product, hacmosld?rtn. FerrltÌn has a hlgh storage
capaclty for lron and ls normally only I /5 saturated.
The nett effect of thls system ls to keep the intracellular pool
of non proteln-bound lron lou. IIALLIUJELL and 6UTTERID6E Il9S{]
hare suggested that thls may be requlred in order to restrlct the
ablllty of non proteln-bound lron, (chelated ulith, say, ÃfP, 6TP or
citrate) from reacting r¡ith HrO" tn a Fenton reactlon thus leading to
the productlon of hydroxyl radicals. EUTTERID0E, RO|.dLEY and
llALLllrJELL [l9BlJ hare developed an ä5iåy for unbound iron salts,
r¡hich utlllzes the requirement of Eleomycln for unbound iron as a
catalyst in Bleomycln-induced DNÊ degradation. Their results indicate
no such "catalytic" iron in serum (consistent r¡ith the iron being
tightly bound by transferrin); houerer, a lerel of 2.8 t 1.2 ¡tmol/L
non-protein bound iron ¡alts in rheumatoid synovial fluid. These
data shou that fî llzÛz becomes arailable t¡ithin the rheumatold
Joint, then a Fenton reactlon ls possible IIìUTTERIDBE, R0ITJLEY and
HÊLLIhJELL, t 9ËZl.
Other erldence for the role of lron in joint inflanrmation is less
dlrect. The adminlstratlon of iron to a rheumatoid patient, partic-
ularly by the parenteral route ls often associated r¡ith àn exacer-
batlon of artlcular Ínflanrmation IREDDY and LElrIlS, 1969J.
Conrersely, ELÊKE gt ê1. t l983(b)l hav¿ clalmed that administration of
a chelatlng agent to gutnea plgs uith antigen-induced chronic
arthritis is associated uith a transient exacerbation of their disease
but ls follor¡ed by . long term lmprorement. The samË group IELÊKE

L4
!.1 d., l9E{l claim å corrÇlatlon betucÊn ltlrìovlal lrsn deposlts and
outcome of dlsaase ln a group of patlents urlth rheumatold arthrltls.
Studlas of the pathologl of lnon orerload s¡rndromes hare
lndlcated radlcal-medlated mechanlsms IY0UNG and âISEN, lgBZ ¡ ]IEYS
and D0RÌ1RIiDY, l9E l]. ln addltlon, ln the treatment of these
condltlons, ascorbate admlnistration r¡lll enhance the effect of the
chelating agent desfenrloxamine (consistent r¡lth its ability to
retrlere Fe(lll) from lts stonage sltes). lf glren alone, hourerer,
ascorbate ulll hare a deleterlous, sometfmes fatal, effect [NlENllUlS,
l98l J. This has been linked to its abllity to interact r¡ith iron to
produce hydroxyl radicals, leading to increased lipid peroxidation.
Finallyr þ rltro eridenct shor¡s that unsaturated lactoferrln r¡ill
lnhlblt hydroxyl radlcal formatlon IEUTTERIDGE É-êl.r l9Ell. llowerer,
lf fully saturated, lactoferrin r¡ill enhancc hydroxyl radlcal production
and lipid peroxidation IÊHERUSO and J0HN50i{, l9E l]. ln surnmary,
tnflamed synorlum ls lron laden and thls ls assoclated ulth an
lncrease ln "blologlcalþ arallable" ¡ron that could lnteract ulth HrÛ,
lÐ vlvo to produce hydroxyl radicals vla a Fenton reactlon.
c. Could stmorial ischaÉmia lead to oxrrggn radical production in
vivo?
Thls thlrd mechanlsm of oxygen radlcal productlon ls much more
speculatire and is reviewed as an arÊa u¡arranting further inrzesti-
gatlon.
Xanthine oxldase has been widely used g vltro to produce a
flux of oxygen radicals because it r¡lll react r¡ith molecular oxygen
and one of a rarlety of substrates (e.g. hypoxanthfne) to produce 0..and llr0r. Houeyer lt äppËärs that þ yiyo "xanthine oxidase" is à
xanthtnc dehydrogÊnàsË tuhlch transfers electnons to NAD+ rathen
than o¡rygËn. Attack by proteoþtlc enz4¿i or oxldation of romÊ

I5
thlol groups durlng purlflcatlon ls bellered to account for ltsconverslon to an oxldase IHALLIh,ELL and GUTTERIDGE, lgESl. Durlng
lschaemla xanthlne dehydrogÊnàsË ls also converted to an oxldase,
prÇsumably also by proteolytlc attack. llypoxla algo results ln the
accumulatlon of one of lts gubstrater, hypoxanthlne, fronr the
degradatlon of ÊTP ln hypoxlc cells. PÊRKS gt gl. [l9Bl] and others
hare suggcsted that thls may account for tlssue damage occurring
durlng reperfuslon of lschaemlc tlsgueg. As olgen suppþ ls
restorcd to hypoxlc tlssue ruperoxldc anlon may be produced by the
damaged Ënzt¡mÊ.
GRÊI|BER, RUTILI and llcC0RD Il9El] shor¡cd that reperfuslon
tnJury of cat small lntestlna, a tlssue rlch fn thls enzyme, ls
dacreased lf superoxlde dlsmutase ls added to the reperfuslon fluld.
Thls mechanlsm has dlnect releyance ln sltuations of coronary
lschaemla IBÊRDNER t$-91., l9B5l and stroke [FLÊlll1 g!j|., l9?E].
Desplte lncreased blood flou to the s¡moylum äs à result of
lnflammatlon, therc exlsts a state of relatlre hypoxla r¡lthln the
s¡rnorrlal fluld and presumabþ also r¡lthln the s¡norium ILUND-OLESEN,
l9?0¡ TREUIIAFT and l1cCÊRW, l9? I ]. ln patients ulth seyÊrr actiye
rheumatold arthrltls a synorlal fluld p0. as lou as I mm llg has been
rcported IFÊLCI'IUK, 60ETZL and KULKÊ, l9?0]. The mlcroyascular
leslons of rheumatold arthrltls assoclated t¡lth lncreased demand of
lnflamed s¡ttrovlal tls¡ue for oxygc.n and glucose may lead to areas of
unstable oxygÊn supply r¡hlch mlght ln turn lead to areas of
reperfurlon lnJury. lndced focal lnfarctlon and necroslg harc becn
ured to explaln the appearance of s¡novlal fluld detrltus knor*m as
"rlcÊ bodles" and also the cllnlcal flndlng of a lou tcmperaturc effu-
slon fluld found ln sevare rheumatold arthrltls hac becn attrlbuted
to an lschaemlc synovlum [tlßRRlg, l9B5l.

t6
lnltlal studles on the dlstrlbutlon of xanthlne oxldase hare
lndlcated that lt ls extremely wldely dlstrlbuted betueen specler
lRlCHEgf and ITJESTERFIELD, 1951, ÊL-KllÊLlDl and C||AGLRSSlAtl, 19651.
It ls not, hotlerer found in the parenchyma of many human tiscuec
(apart from llrer and small lntestlne) but ls prese'nt ln human
endothellal cells. Llttle ls knoun of the xanthlne oxldage content of
s¡rnorlal tlssues.
The potential relerance of thls mechanlsm uarrants further
study, since the concept of enztmatlc denaturatlon of an enzqe
due to hypoxlc or proteolyllc damage may apply to other
flarroproteln dehydrogenases, e.g. aldehyde dehydrogenaie, and could
lead to oxidase actlvity uith superoxlde production [LÊH05, personal
communicatlon].
1.
Êppreclation that biologlcal systems cån generate highly
reactlve partlal reductlon products of oxygên has largeþ come
about by the demonstratlon of the cellular defense mechanlsms thatprotect agalnst them. The prlmary defcnse ts provlded by " systenr
of enztrmes. Thr superoxlde dlcmutases ulll ellminate superoxlde
anlon by catalyzfîg lts csnverslon to hydrogrï pËroxlde, I'lydrogen
peroxlde ls remored by catalase r¡hlch lt r¡lll csnvert to uater plus
oxygÊn and by peroxldaces h,hlch also reduce hydrogen peroxide to
uater using a rarlety ol reductants arailable to the cell. Efflcient
remoral of one of the first tr¡o fntermediates of oxygen reduction
prerents the formation of the thlrd, the hydnoxyl radlcal, rrlhich is so
reactire that enzymatlc scayenging uould be tmpossible.
a. fupæ,ËidsiiÐs!.êEe¡.The exlstencÊ of haemocupreln, ä coppËr containlng protetn
componËnt of borlne blood r¡lth a molecular r,uelght of 5,1,000 uac

l?
dlscorered ln l93E by tlÊNN and KElLlN. tloueyer, lt urar not unfll 30
yÊärt later that l'lcC0RD and FRIDOVIEII Il98El ldentlfled lts functton.
They demonstrated that haemocupreln, nou Jrnourn ås superoxrde
dismutase, rrräs able to cataþtlcalþ remove rruperoxlde anlon. i{oother substrate has been ldentlfled for thls enzqe uhtch ls ublq-
ultous ämong aeroblc organlsms. Three dlstlnct classes of superoxide
dismutase, each r¡lth a dlfferent metal at thelr acüye site, haye
no¡¡ been rÊcognlzed [FRlD0PlEll, l9?5]. Enz]¡mes r¡tth iron or
mangänËse at their actlre sltes äpË gËnÊrally chanactenistlc for
prokaryocytes and mltochondrla. They share amino acld sequ?nce
homologlÊs suggestlng a common erolutlonany orlgin. The coppen
contalnlng Ënzymr studlad by llcCord and Frldorlch has been shor¡n
to contaln zlnc as tr.rell, although thls ls belleyed to fulfìll a stnuctural
role and ls not essentlal fon enz¡rmatlc actlrlty [EEEÌ1, RIEHÊRDS0N
and RÊJÊGOPÊLÊN, l9??1. lts amino acid sequËncË is qurte unnelated
to that of the othen tr¡o anzymes and lt is found in the cytosol of
eukaryotlc cells IKEELE, Hcc0RD and FRlD0Plcll, l9?0]. The copper
contalnlng enzyme is lnhibited by cyanide but ls stable to ethanol
and chloroform uhereas ¡¡lth the mangånese and iron containing
enzr¡mes the rererse is the case IFRlDOPlCtl, 1982].
The mechanlsm of enzqe cataþsed dismutation of superoxide
anion aPPeårs to lnvolve alternate reduction and oxldatlon of the
transltlon metal lon at the actlve Elte [RgTlLl0, BRAY and FIELDEN,
l9?21. The three dlmenslonal structure of the proteln subunits of
bovine coPPËr containlng supËroxlde dlsmutase suggests a positirely
charged track that leads to the actlye site and a slmllar mechanlsm
ls llkely for the mängänËse and lron contalnlng enzqÊs [llÊLLll¡JELL
and GUTTERIDEE, 1985J.

I8
The superoxlde dlsmutasÊs arÊ essentialþ lntracellular Ënzqes,
Lerels found ln extracellular flulds haye been ve?y lor¡ [l{cC0RD, l9?{].
llore recentþ a hlgh molecular r.r.relght form of copper contatntng
superoxlde dlsmutase has been neported II1ÊRKLUND, l gEZ]. Thls form
ls present ln the extracellular tpacÊr predomlnantþ of the lung. lt'sexact role ls yet to be elucldated.
hb. BlutaÏone f[ìlutathlone oeroxldase).
Glutathlone peroxldase detoxlfles hydrogen peroxlde through
thc oxldatlon of reduced glutathlonc accordlng to the reactlon¡
ZGSII * HzEz >
glutathloneperoxidase
The oxidlzed glutathione is then reduced by ^ second cnzqeglutathione reductase uslng l{ÊDPll (generated by the hexose mono-
phosphate shunt) as the reduclng agent. 0ther peroxides can act
as substrates for glutathlone peroxldase e.!.r lipid hydroperoxides
r¡hich r¡lll be metabollzed to hydroxy fatty aclds.
ElutathlonÊ pËroxldase (l1lÁl 85,000) lt a selÉrrlum dependcryrt
enz¡,rmÉ found ln the cytosol. lt ls effectlve at lou concentratlons of
hydrogen peroxlde and thereforu probably constltutes the cell'c flrstllne of defense agalnst thls agent. Furthcr crldence of thlc comËt
from the fact that glutathione peroxidase deficient indiriduals sulfer
a dlsease slmllar to chronlc granulomatous disease [llOLt1ES Êt Al.,
ts?01.
c. Eatalase.
Catalase catalyze¡ the dlyalent reductlon of hydrogen
peroxide to uater accordlng to the reaction:
211282 >

t9
It hac à, very hlgh P¡¡¡¡ for hydrogen peroxlde l.e. reactlon
rates approach the dfffusion controlled limit and thu¡ lt is very
difficult to saturäte. tln the other hand, houever, it is not efficic'nt
at remorzlng lorrr concentratlons of hydrogÊn peroxHe [SCl'lONBAUll and
CllÊNCE, l9?61. llost purlfied catalacec consist of four protein
subunits, each of r¡hich has a molecular u.leight of 60,000 and
contains a haem group bound to its active slte [PÊlNSllTElN BÈ 3!.,
l98 ll. Catalases are intracellular enz],l¡"n.as found in most aerobic
cells and ln animals, particularly in liver and red blood cells.
Êcatalasemic indiriduals are relatively symptom free INÊTSUDÊ et ê!.,
l9?61, thus implying that catalase is not the cell's primrary defence
against oxldant stress. Nerertheless it rr.rill complement the
glutathione/glutathione peroxidase system particularly at high levels
of hydrogen peroxide.
d.@.Ê number of other mechanisms exist uhich appear to
functlon, at least in part, as antioxidants. Eaeruloplasmin, r¡hich is
present in serum, has the ability to act as aî oxygen radical
scâvenger. 60LD5TE!N et ¡!. [19?9] have claimed that in combination
uith copper ions it has the ability to act as a superoxide dismutase,
houeyer this has been disputed [EÊtlHlSTER gt ê!., lg80l. Êscorbic
acld may act as a reductant, in r¡hlch case lt forms the semi-
dehydroascorbate radical uhich is relatiyely unreactiv¿ and usually
undengoes disproportionation to ascorbate and dehydroascorbate.
ln this manner ascorbate may react urlth superoxide anlon actíng to
detoxlfy it and its subsequent more reactire radical products.
Êscorbate is found in high concentratlons in the lens of the eye,
r¡hich has lou superoxide di¡mutase levels. lts function there is
belierzed to be an , antioxldant. Êlso uric acid, at lerels found in

20
the blood (0.12-0.'15 nrmol/l), can act âs ä porr.rerful scarenger of
stnglet oxygËn and of hydroxyl radlcals [Êl'1ES Êt å1., lgBl].
e..llcCORD t l9?{l flrst reported that boylne s¡rnorial fluld con-
tains a barely detectable amount of endogÊnous catalasr (1.e., 0.05
¡rgm/ml or less) and rery lor¡ concentratlons of endogenous
superoxlde dlsmutase (l ¡.rgm/ml). These leyels are at least Z-5
orders of magnltude less than the lntracellular concentrations of
these enzymÊi. He also shouJed that the activity of the superoxlde
dlsmutase present could be inhibited by cyanide, thus suggesting
that it t¡.ras the copper containing enzqe. The endogenous
superoxide dismutase presert ln fresh borine synovial fluid in Hc0ord's
experiments uas enough to effect only partlal protection of synovial
fluld hyaluronlc acid from än exogenously applied srrygen radical flux.
The magnitude of the flux that he applled uas equizalent to thatproduced by 5xl0ó actirated neutrophlls/ml, E yJgg. Sqne
inflammatory synovial flulds contaln a number äs much as tuentyfold
greater than thls. Extracellular flulds other than synovial fluid,
includlng human iÊrum, human cerebro-spinal fluid and borine
äquËous humour, have simllar lor¡ larels of both ënzqes. ELAKE Etå!., Il9El] examined the sgmovial ffuid from eight human patients ¡¡ith
rheumatoid arthritis and found no detectable superoxide dismutase
actituity ås åssessed by abillty to inhibit reduction of nitroblue
tetrazolium. They also found lor¡ catalase actirity (0-55 units
catalase/ml). l6ÊRl et al. [1982], using inhibition of reduction of
ferrlcytochrome C, reported that lerels of superoxide dlsmutase in
the s¡rnorial fluid of patients uith rheumatoid arthniüs r¡as
lncreased compared to that found ln ostaoarthritls. Î{erertheless
the absolute lerels that they report Ðte r,¿yeral orders of

zt
mâgnltude less than lntracellular lerels [FRlD0flCH, l9?5J. ï¡novlal
fluld ceruloplasmln lerels urere found to be lncreased ln rheumatold
arthrltlc ISCUDDER Êt Al., l9?El. llouerer, the results reported by
ELÊKE Êt gl. I l9E I I ¡¡ould suggest thls ls not associated r¡lth slgnlf-
lcant (i.e., detectable) superoxlde dlsmutase actirlty. Total
ascorbate lerels ln rheumatold s¡morlal fluld ur?re found by BLAKE ç¡!
¡!. I l9E l] to be at the lor¡ end of the normal ràng". Reduced
ascorbate hJâs not measured.
5. Targets for Oxy Radical Êttack
A large numbcr of biologlcal moleculer have been reported to be
denatured ln somÊ way by rcactlon¡ r¡lth oxygen radlcals. lt ls my
lntentlon to drau attentlon to tho¡e morÊ relevant, to inflanrmatory
Jolnt dlgeare,
a. Ec,glElE¡,
Holecules that ärË uniaturated or contain a sulphur molety
ârÊ susceptlble to free radlcal attack [PRY0R, l9?6]. Thrse include
the unsaturated amino acids, tryptophan, tyroslne, phenylalanine,
and histidine, and sulphur containing amlno acids, methionine and
cysteine. Thus, enzryes r¡hich depend on these amino acids for their
reactlvlty arÊ llkely to bc lnactirated by oxygÉTr radicals [Lltt and
ÊRI15TR0N6, l9?E; EUCIIÊNÊI{ and ARHSTRONIì, l9?El. Tu¡o examples of
this are, flrstlyr the fnactivation of serum cr I -pnoteinase inhibitor
upon ?xposurË to an oxygen radlcal flux generated by stlmulated
neutrophlls [EARP and JÊNOFF, l9?9]. lnactlyation is due to oxidation
to a sulphoxlde of the methlonlne resldue at the actlre sitc of thls
ËnzlrmÊ tnhlbltor U0l'll-130H and TRÊ/15, lg?g]. This has been proposed
äs å mechanl¡m of lung damage ln cigarette smoke-lnduced
emphysema, slnce clgarette smoke ls a potent source oî oxygen
radlcals ICÊRP and JÊtl0FF, l9?EJ. Secondly, JÊ51Ì{ [l9EZJ has

z2
rcportcd the generatlon of lgG aggrËgäter, u.rlth the propertles of
irnmune complexes, upon expoture of lgG to the myeloperoxlde-ll"O"-
hallde system. Further uork on thls phenonrenffi by tlULLlNÊX and
NULLIi{ÊX tl9E5] ha¡ shoum the formatlon of dltyrosine crosslinks (1.e.,
lnterproteln dlsulphlde bond formatlon). Thls could represent an
antlgen-lndependent means of formatlon of "lmmune complexes", r¡hich
could lead to the production of rheumatold factor.
b'@'Ê considerable literature exlsts r¡hich deccrlbes cell mutatlon
and death from ionizing radiation [I1YERS, l9E0J. These effects are
primrarily due to free radical reactlonc r'rlth DHÊ. A number of
enzlrmatlc radical generating systems hare beer tested, including
the X0/llX system [ERÊlrJll and FRlDOVlEll, l9Ell, that hare shou¡n
DNÊ strand scission. llydroxyl radical scärÊngers have been
protectlre, thus lmpllcating this agent.
E. Membrane lipids.
The unsaturated bonds of membrane cholesterol and îattyacids can readily react r¡lth free radicals leading to lipid penoxidation
IFRÊNKEL, l9Ë0]. Êfter lnltlatlon r¡lth an âppropriateþ raactlre
agent, e.g, the hydroxyl radlcal, thls process ulll become
autocatalytlc, yielding a variety of products including lipld peroxides,
llpid alcohols, aldehydes. Ên increase in the senum and synorrial fluid
content of llpid peroxidation products ln actire rheumatoid arthritis
has been reported ILUNEC É å]., l9Ell. Llpid peroxidation has been
proposed as ä cytotoxic erent Jn a number of conditions. llou.lerer,
as HÊLLlhlELL and EUTTERIDGE t1981(b)l have pointed out, lipid
peroxldatlon is a common pathr.uay in the dissolution of a cell and the
presence of llpld peroxldatlon products per se, does not Ìmpllcate llpld
peroxldation as the primany cytotoxic eyent.

23
d. Extracallular mac olecules other than hr.raluronlc acld.
The depol¡¿merlzatlon of several mäcromolecules of blologlcal
lmportance by rxporunË to a rarlety of oxygen radlcal generaÜng
systems has been demonstnated. Perlodate oxldation, for example,
ulll lnduca a loss of vlscoslty ln solutlons of proteoglycans and
glycosaminoglycans ISCOTT, TIEhJELL and SÊJDERA, l9?u], alginate
IPÊINTER and LARSEN, l9?01 and methylcellulose [5C0TT and TlGtrlELL,
l9?51. 5c0TT and PÊ6E-T||0HÊS [19?6] have presented evidencr for
the productlon of hydroxyl radlcals tn àquÊous periodate solutlons
and ln general the depolymerlzatlons descrlbed could be inhibited by
the hydroxyl radlcal scayenger, propan-l-ol.
Ê decrease in riscoslty in solutions of lsolated proteoglycan
subunit from borine nasal cartllage occurs upon exposure to the
xO/l'lx onygen radlcal generatlng system itr ritro IBREENUJÊLD, l'1BY
and LÊZÊRUS, l9?61, EÊRTOLD, hJIEEKIN and TH0NÊRD I l9E{(a)] haye
demonstrated a decneasa ln hydrodynamlc size of a glngiral
proteoglycân upon exposur? to oxygen radlcals. These authors
suggesttd that glycosaminoglycan side chains urËre cleayed from the
corÊ protein.
GREENITJÊLD and l10Y Ilg?gl have ¡hor¡n that soluble collagen
after Êxposure to an oxygen radical flux (generated by the X0/l'lx)
does not gel normally l¡hen heated. These authors and
PENKÊTÊSUERAIIAI|lÊtl and J0SEPll [13777 have postulated that this-
effect ls due to cleavage of the telopeptides of the non-helical
region. ll0NEOlSSE É e!. [19e5; l9B{], hor¡eyer, hare reported thatexPosure of soluble collagen to the X0/llX oxygen radical generating
system leads to multlple clearages of the collagen mrcrofibrrls
lncludlng the trlple hellcal reglon. lt ls not äpparent from these

24
studles, houerer, hour susceptlble fully aggregôted collagen flbres
are to oxygÊn radlcals.
e.@' l) Red cells.
Both stlmulated phagocytes [5lNCl'l0ldlTZ and gPILEERE,
l9?9; trJElSS, l9E0l and non-cellular onygên radtcal generailng
systcms IKELL0GG and FRlDOylCH l9??] ârË cäpable of medtatÍng red
blood cell lysls. The blochemlcal mechanlsms leadlng to lysls appear tobe complex and not nccessarlly appllcable to other cell types, The
fir¡t conslderatlon ls the presencÊ of oxyhaemoglobin r¡lthln the red
crll. Thls Is bellerzed to be prcsent ä! iupÊroxy-fercihaeme compound
IC0LLHAN É å1., l9?6] uhlch may autoxldtz e provtding a conflnuous
sourcÊ of superoxlde anlon r¡lthin the cell, uhlch ls ellmhated by
endogenous supÊroxlde dlsmutase unless this autoxidaflon rs
accelerated, e.9., by haemoþtlc drugs such as phenylhydrazfne or
menadlone. Reaction of superoxide anlon l¡lth haemoglobin leads totha formatlon of haemoglobin breakdor*¡n products (including
methaemoglobh) rr.rhich bind to the red cell membrane and cause
lncreased osmotic fragillty of the cell IIìOLDEERE and STERi1, 197?7.
LYNCH and FRlD0rlcll [19?s] hare demonstrated the exrstence of
anlonlc membrane channels ulthln the erythrocyte membrane by
l¡hlch superoxlde anlon can galn free acces¡ to the cell interlor and
thus react ulth oxyhaemoglobin. ln addiflon, [rlEl53 [lgE0] has
postulated that phagocytË genËrated hydrogËn peroxlde, uhlch can
dlffuse freely acro55 the cell membrane, cän react ulth methacryno-
globin to fonm a cytotoxlc complex capable of haemolysrng the
erythrocyte. Thls has stemmed from experiments that hare shou¡n
accelerated phagocyte-lnduced red cell lysls ulth exogerìous
superoxide dlsmutase, or inhlbltion of endogenous cataläse or

z5
glutathlonc peroxldase and protectlon r¡lth exogÉ.nous catalase
[lrJElSS, l9E0l. 0ther uorkers have demonstrated damage to
membrane llplds and protelns u.¡lth the use of red cell ghosts, r.uhere
the effects of oxyhaemoglobin have been elimlnated. Thus, prlmary
llpld peroxldatlon ls an alternatlre mechanlsm fon red cell þsls
[KELL066 and FRlD0PlCll, l9??; FLYNN Êtå!., l9E3; DRS and NÊlR, l9E0;
R05EÌi, EÊREER and RÊUCKI1ÊN, l9E5l.
2) 0ther cells
6lren the nonspeclflc reactirzity orlygÊn radlcals, lt ls not
surprlsing that enz',matic and chemical superoxide generating
systems have been shoun to be cytotoxic to cells in culture. Eells
lines studled hare lncluded human gllal cells IDEL HÊESTR0 .Êt ê1.,
19801, human lyrnphocytes [HcNElL, CLELÊND and EETÏS, l9B5l, lung
cells [F0X and ÊUTOR, l9?8] and human f¡broblasts [5lt1ON,5E0EGlH and
PÊTTERS0N, l9E I l. Êlso, sublethal exposure to oxygeî radicals causes
a r¡ide rängÊ of alteratlons to cell function. For example, KRÊUT and
SÊ60NE Il9El] hare demonstrated that exposurË of þphocytecultures to the X0/X system reduces the ability of these cells to
form rosettes ulth sheep red blood cells, thus indicatlng cell
membrane receptor damage. EÊTES, LOUITHER and I.|ÊNDLEY I l9E{]
hare demonstrated that cultured borine artlcular chondrocytes
respond to exposure to the X0/llX oxygen radical generating system
r¿rlth reduced proteogþcan and proteln synthesls that pensists fon
at least 5 days follouÍng exposurÊ. ln general these effects haye
been completely prerentable by catalase IEÊTES, L0lrJTtlER and
|.|ÊNDLEY, l9E{; KRÊLT and SÊBONE, l9E2¡ Sll10N, SCOGG|N and
PÊTTERS0N, l9E I ¡ DEL Ì1ÊESTRO E! E!., l9E0l, thus implicatlng hydnogen
peroxlde äs å key lntermedlate. lt ls appropnlate to note that fn
these cases the oxygen radical generating system is applied

z6
externally to the cell and the non-charged nature of hydrogen
penoxlde may allor¡ lt to penetrate hydrophoblc reglons of the cell
membrane. lndeed the cellular alteratlons and cytotoxlc ereîts may
depend upon the slte of secondany oxyg?n radlcal generatlon, lnduced
by hydrogen pËroxlde. The factors llkely to be inyolyed here are
complex, Ê.9.r ät/äilabllity of transltllon metals and/or chelator,
cellular antioxldant defence rtatus and target susceptlbllity.
Systems utilizing neutrophlls as cellulan sourcÊs of oxygen
radlcals hare also been described. Êutocytotoxicity of stimulated
neutrophils can be inhibited by superoxide dismutase [5ÊLlN and
P1cCORD, I g?sJ. This effect and the presencÊ of the myeloperoxide-
Hr0.-hallde system create difficulties tn interpreting studies of the
effects of oxygen radicals generated by stimulated neutrophils.
hJElSS É ê1. Il9ElJ demonstrated that granulocyte-induced injury
to cultured endothelial cells, as measured by ='Cr release, brås
lnhlbited by catalase but not by superoxlde dlsmutàse. Êdditional
myetoperoxidase dld not enhance slCr release and myeloperoxidase
deficlent neutrophils shor¡ed slmilar effects to normal neutrophils.
Thls suggests a myeloperoxldase tndependent, hydnogen peroxide
medlated cytotoxlclty.
There are, hourey¿rt tlssue systems uhere superoxide
dismutase äppcars protecllve. Superoxide dismutase r.uill protect
against the increased microrasculature permeability observed in the
hamster cheek pouch u¡hen lt is perfused r¡ith XE/X IDEL NÊESTR0 Êtê1., l980l. lt uill also ameliorate reperfusion injury Ín the cat small
intestlne IPÊRI(S Êt ê1., 19Ê21. Clearly, as the systems studied
become more complex, it becomes more dlfficult to fnterpret the
results of scarenger studles ln a r.lay that leads to a clear
understanding of the important inltial cellular events leading to

z7
toxlclty and to detenmtne the radlcals lnrolrad.
It also appcêrt that oxygÉTt radlcal dependent cytotoxlclty
mäy medlate such lrnmunologlcal procËJr as neutrophll-rnedlated
antlbody-dependent cytotoxlclty IE0RREGAÊRD and KRAGBALLE, l9E0J
and posslbly natural klllcr cell cytotoxlclty [SUltlÊtfTtllRÊÌ1 gt tl.,198{] although the latter contentlon ls far frorn establlshed.
c. HYRLUR0ÎilC RC|D
l. ln$sd$s:llsn
The term "synovla" uJas colned by Paracelcu¡ to dcscrlbe Jolntfluld because he llkened lts vlscous nature to that of an egg
IR0DNÊN, EENEDEK and PÊÌIETTA, l96El. The component nesponsible
for the hlgh rlscoslty of s¡.rnorlal fluld ls hyaluronlc acld. Thls
u.rldespread polysaccharlde ls classlfled as a glycosamtnogþcan
although màny of lts characterlstics mank lt as belng unlque. ln
the follotrlng pàràgràphs I shall neylerr¡ the physlology of thls
màcromolecula, ln partlcular, lts structure, llkeþ functions,
blos¡,tnthasls, turnorer and lts lntaractlons r¡¡lth cells partlcularþ ln
relatlon to lts ppÊsËncê ln the synorlal fluid of dlathrodal jotnts.
Z. Chemlcal Structure
flyaluronlc acld uac dlscovered by lleyer and Palmer ln 193{ ln
borlne vltreous humour. lt hag since becn lsolated lrs¡¡t numeroug
tlssue¡ lncludlng human rltreous, skln, umblllcal cord, blood vessel
u.ralls, and ls also prescnt ln lot¡ concentratlon ln the serum. ln-
deed lt ls a ublqultout component of extracellular matrlces. lnltlal
che.rnlcal studles by IIEYER and PÊL!'1ER [ | 938] ¡houed that lt uac a
large molecular urelght, non-rulphated, acldlc polysaccharlde. ltconglstg of equlmolar amounts of D-glucuronlc acld and l'{-acctyl-
glucosamhe (2-acetamldo-2-deoxy-D-glucose) both ln the
pyrannose form [|'1EYER, 19,1?J.

z8
Studles by JEANLOZ and FORCIIELLI Il95ll, more reccntþ cørflrmed
by LOllBÊS and HEYER Il98ll, demonstrated that the polymer l¡
unbranched and that the tuo constltuent sugari occur alternately
all the r.uay along the molecule (Ftgure l-l). The nature of the
glycosldlc llnkages urepa shoun, r¡lth the use of speclflc
hyaluronldases, to be alternatmg $ | -5 and D | -{ glycosldlc llnkages.
Eoth testlcular and bacterlal hyaluronldases cleave at the D | -{llnkages and leech hyaluronldase cleaves at the Fl-5 ltnkage
ILÊURENI, lg?0]. llyaluronic acld pol¡mers shor¡ conslderable
yarlatlon ln length (1.e., polydispersltg) and therefore total molecular
rrelght, but apart from thls no chemlcal dlfferences have been
demonstrated ämong hyaluronlc aclds from varlous blologlcal sources
including bacteria IBR|iIÊCOHEE and IdEBEER' lgE'îJ. The lsolation
mcthods that have bec'n used to separate hyaluronic acld harze
produced a product that contalns rarlable amounts of protefn.
Using mlld lsolatlon technlqucs 0fì5T0H and STÊt{lER [1950] separated
a hyaluronate-proteln conrplex from ox Jqtrvlal fluld that contalned
?5-3O% protein. Because thls sedlmcnted at a sfngle peak on
ultracentrlfugatlør they suggested that comblnatlon ulth protein
mäy ba an tntegral part of the structure oî hyaluronlc acld and
nËcetiary for somÊ of lts functlons. The flndlng of a rarlatlon ln the
amount of proteln bound r¡lth change ln pll has led CUgfÊlH [l955l
and othcrs to suggest that the bulk of thlg lnteractlon ls lonlc.
Êlternatlvely, houerer, proteln may bc adsorbed (trapped) lnto
mlcelles of the randomþ colled hyaluronlc acld molecules [0GST0Ì{ and
STÊN|ER, l95ll. Using morc rlgorous technlques hyaluronlc acld can
be lsolated r¡lth as llttle as 8.35% proteln [SlrJÊNl{, l9EEJ. lrJhether
thls proteln ls coralentþ bound l¡ unclear [LÊUREfff, l9?0; lllKUlll-
TÊKAEÊK| and P00LE, lgEl; TSIBÊÌ103, yYNl0S and KALPÊX|S, l9E6l.

Flgure l-l STRUCTURE 0F HYALUR0NIC ACID
The structure of hgoìuronic ocid showing the two
component sugars D-glucuronic ocid ond N-ocetglglucosomine
joined ot the centre bg o ßl-3 glgcosldic link0ge. At the
opposite end of each sugar Ëre the components of 0 ßl-4glgcosidic linkage. These two sugars alternote along the
whole length of this unbranched molecule,

cH2oHcooHo
HHH
\
:
.3¡
NH H3
o
HOHH

80
lndeed rcmoral of proteltt by trypsln dlgestlon does not affect the
tntrtnslc rlscoslty of hyaluronlc acld solutlons IEÊLÊZS and SUIiDELÊD'
lg5gl,
3. Ëyaluronlc Êcld fn Êou
Hyaluronlc acld has a hlgh afflnlty for uater. hlhen hydrated lt
sr¡ells to occupy a volume onÊ thousand tlme¡ greater than lts
anhydrous yolume [065T0H and STÊlllER, l95lJ. Thus a tingle molecule
of sod¡um hyaluronate (t1lrJ = 5 x l0É) r¡lll olcupy a spheroidal domaln
r¡lth a diameter of 0.5 !r or onË gram dissolrred Tn physlologlcal ¡aline
r.lill completely occupy ã liters of solrent. ThereforÊ, irr
concentratlons abore 0.35 mg/ml there r'rlll be overlapping of the
molecular domains of macromolecular hyaluronate and the formatlon
of a polymer matrlx [EÊLÊ25, l9?4J. Ê coarse neturork of branching
ftbrtllar strands has been vlsuallzed by llÊDLER Êt å!. lJgAZl using
electron mlcroscopy of freeze etch replicas of hyaluronate solutions.
lnitlal studles based on vlscotltyr sedtmentatlon and llght
scattering data tuggerted that the molecule adopts a random coll
conformation ln solution [OttsT0]l and 5TÊl{lER, l95l; LÊUREHT' 1955;
ELUNEERIì and 0tì5T0lt' 195?J. Hore recent studies uslng xray
crystallography have demonstrated that tndlvidual chains of
hyaluronlc acld adopt a stable left handed hellx - most conrmonþ
r¡ith 5 disaccharide units per complete helical turn IhJIHTER' Sl'1lT]l
and ÊRNOTT, l9?5J. The chains tend to be fully extended and are
htghly stable regandless of degree of hydration or lonic strength ln
the preparatlons used for crystallography and therefore likeþ to be
found as such in free solution. ln addltion, structural considerations
uslng the fact that the pyrannose rlngs of hyaluronic acid are both
ln the chalr form, lndlcate that lntramolecular hydrogen bondlng
äcross each glycosldlc ltnkage cân occur [ÊTK|NS, HEADER and SCOTT'

8r
l9E0¡ UJINTER, SHlTTl and ARll0TT, lg?51. Erldence for a secondary
structure stablllzed fn this manner has been found by SCOTT Êt À1.
t t Sg ll ustng ll'l n.m.r., although that study uas done using dimethyl
sulphoxide rather than r¡¡ater as the solvent. ln additlon,
lntermolecular lnteractlon betr¡een hyaluronlc acld molecules
medlated by r.rater molecules, formfng bridges betueen hyaluronate
catlon chalns, has also been postulated [SHEEllÊtl, ÊTKlllS and
NIEDUSZYNSKI, l9?51. These tnterchaÍn lnteractions, although
translent, r.,.rould tend to lead to a degree of order through chaln
packlng ulthtn the matrix, albelt temporarlly. Ê full dlscussion of
these polnts ls prorlded by l-lÊDLER and NÊPIER [19??].
{' lua. L$.ÞrLç^illlsn
Ên appreciatlon of all the functlons of hyaluronlc acld in
synorrial fluld is far from complete. The rlscous nature that hy^-
luronate lmparts to synovlal fluld has led to the suggestion that itacts äs a cartllage-cartilage lubricant, llouerer, RÊDll{, St¡JÊl{tJ and
IJEISSER [19?0] clearly demonstrated that the cartilage lubricating
actlrzity of r¡hole synovial fluid ls present in the protein fraction and
is undimlnlshed by hyalurmridase treatment. SldÊlttt and couorkers
hare isolated tr.rro high molecular r,reight glycoproteins (lubricin I and
2) r¡hlch account for thls property l5tdAl{l-l and RÊDl11, 1912i StÂlÊtll-l gt
ê!., l9E l; ShJÊNN É ê1., lg8{]. l{evertheless, a role for hyaluronic
acld in prorldlng lubrlcation betr¡een the soft tissues of the joint
has also been postulated. þ vltro eridence for this has been
prorzlded by RÊDlltl 9È å!., [19?l] r.,.rho shor¡ed that, coîìpared urith
buffer, the coefflclent of frlction betueen a flat surface and an
approprlately loaded sectlon of synovlal tlssue uas decreased
equiralently by synovlal fluld and a solutlon of hyaluronic acid alone.

82
b.@FrrlErl'lyaluronlc acld lmparts to s¡novlal fluld lts characterlstlc
riscoelastlc propertles. The rheologlcal beharrior of s¡novlal fluld and
of proteln-free sodlum hyaluronate solutions are rirtually identical
lElEBS, IIERR|LL and SlllTll,l96El and thlc beharlor may be considered
as haylng a rlscous component (d¡mamic loss module) and an elastic
component (dynamic storage module). These parameters can be
meåsured orer a nànge of strain frequencies corresponding to
rr.ralklng and runnlng. Predominantþ viscous beharior ulll result in
energy being dlssipated to surrounding tissues as heat and elastic
beharior results in energy belng stored in the deformatlon of the
hyaluronic acld molecules lfor full discussion, see EALÊZS, l9?{]. hlhen
these modull are measured n synovlal fluids from normal (young) knee
joints an elastoriscous transformation ls observed, 1.e., a
tranEformation from predominantly viscous behavior (at lorr.t straln
frequencies) to predominantly elastic beharior (at strain frequencies
corrËspondlng to a îast rr.ralk). This conrersion to elactic beharior is
less marked ln synovlal fluids from older subjects and uas not
obserred at all n synovlal fluids from patients uith osteoarthritis
and inflammatory joint disease evën at strain frequencies
corrÊspondlng to running [EÊLÊ23, l9?{]. Elastic beharior mày serve
the important function of protecting synovial membrane and
articular cartilage from mechanlcal shock and deformation (i.e.,
acting as ä shock absorber).
c. Ê Conduit for l{utrients
The second main function of synovial fluid ls that of nutri-tion. lndeed the nutrients required by the surface layers of carti-
lage are derlred from the synovlal vasculature and therefore must
pass through the synorlal fluld hyaluronate matrix. The posslble

88
influence of hyaluronate on the transfer of nutrientc has been the
subJect of con¡lderable lnrzestlgatlon. For example, llÊDLER I l9E I ]
claimed that transport of gluco¡e through an hyaluronate matrlx is
factlltated (1.e. lt occurs fa¡ter than through uater alone). This
flndlng has, hot.rÊyer, been dlsputed [NoRToN' UREÊN and Ì1ÊRoUDÊS'
lgBzl. The synoylal membrane has a rlch blood supply IR0PES and
BÊUER, lg55l and the caplltarics hare a fenestrated structure ISUTER
and HÊJN0, l9E4l. Thus, although s¡novlal linlng cells have lntercel-
lular Junctlons [tìR0Tll, t9?5], they do not form a discrete layer
IEHÊDIÊLLY, l9?EJ. The result ls that, ln certaln areas, the "ground
substancË" of the synoyial membrane has free àccess to the joint
carlty. These morphological features suggest that the extracellulan
matrlx of the synorial membrane plays an lmportant nole in
regulating the outflor¡ of fluid and soluble blood comPonents fnom the
capillaries of the synorium. lndirect eridence r¡hich supports this
concept u¡as obtained by NETTELELÊDT, SUNDELÊD and J0N550N [1965]'
uho shoued that sedimentation of plasma through a solution of
hyaluronlc acld produced an ultrafiltrate u.¡ith composition similar to
that found in normal s¡novial fluid.
5. Eios,unthesis
In recent years our understanding of the biosynthesis of hyal-
uronic acid has increased conslderably. PREIIH [1983 and l9E'l] and
PlllL|PS0N and SCHIdÊgtZ Llgïl) have shor*m that hyaluronate
synthetase is situated on the interior surface of the plasma
membrane and that as the molecule ls synthesized it extends out
lnto the extracellular späce. lt ls syntheslzed at the reducing end
by alternate transfer of the substrates UDP-GlcNÊc and UDP-G|cA to
the hyaluronate chain. The substrates can initiate chain formation
and no protein primers are rËquired. This is in contrast to the

84
synthËsis of other màmmällan glycosamlnoglycäns r¡hlch ane
syntheslzed at the 6olgl apparatus and undergo post-s¡rnthetlc
modlflcatlon. The mechanlsm of chatn tenmlnatlon for hyaluronic acld
synthesls remalns unclear.
6. Turnorer
Lrlhllst lt has been establlshed for many yeart that hyaluronic
acld ls syntheslzed ln the tissues r¡hene lt ls found I6R0SSFIELD eÈ
ê!., l95EI, knouledgÊ concÊrnlng lts turnorer has developed only
recently r¡lth the arallablllty of radiolabelled high molecular ueight
hyaluronlc acid produced in cell culture IEÊXTER, FRÊSER and CLÊRR15,
19?31. The lnltlal study in this ärea uas from ÊiTT0HÊS, FRÊSER and
I'IUIRDEN [19?3], r¡ho follor¡ed the fate of radiolabelled high molecular
u.relght hyaluronic acid after tnjectlon into rabbit knees. They
detected radloactirzity fn the synorlum, cartllage, negional lymph
nodes and blood plasma. âppearance r¡rithin regional lymph nodes r.,.ras
obserred wlthln l5 minutes of injection. Hyaluronic acid entens the
genenal clrculatlon most probably fnom the l¡mph ILAURENT and
LÊURENT, l9E I l. The normal serum concentration is of the order of
200 ng/ml ILÊURENT and TEN6ELÊ0, l9B0] and labelled hyaluronic acid
is cleared very rapidly from the blood by the liyen r¡here it is taken
up by the endothelial cells IERIKSSON Ê!å!., l9E5] by. receptor
mediated mechanism [SHEDSR0D et g]., l9E{]. There it is degraded
rapidly and completely IFRÊSER G! è!., l9E I ] in lysosomes [SHEDSROD
g! al., l984l. The major products of hyalunonic acid catabolism by
endothelial cell in culture àre acetate and lactate [S|'1EDSR0D, l9E{],
although ln the presence of hepatocytes and in ¡¡ ¡liyo expeniments,
these are further metabolised to C0, and u¡aten.

85
?. Hpaluronlc acld-Cell lnteractlons
lnltlal demonstratlon of an hyaluronlc acld-cell lnteractlon
related to the demonstratlon that a rarlety of cell types hare a
perlcellular coat of hyaluronate [60LDEERG and T00LE, l9E{] r¡hich
has the ablllty to exclude partlcles. lt has bee,n suggested that this
cell surface hyaluronate may be important in such diverse cellular
propertles as äggregätion, resistance to viral infection and
tumourlgeneslg. lJlEEKll''{ and llUlR [19?5] demonstrated thathyaluronlc acld uas able to lnhlblt the synthesis of proteoglycans by
chondnocytes ln culture and therefore may have a regulatory role in
this chondrocyte functlon. The nature of the lnteraction is very
speciflc slnce no inhibition u.raE seerì r¡ith the other
glycosaminoglycans or índeed r+rlth chondroltln, the desulphated form
of chondroitln sulphate, ulhlch dflfers chemically from hyaluronic acid
only in the orlentation of the hydroxyl group at E4 on the N-
acetylglucoEamine.
Further eridence suggesting that hyaluronic acid influerrces
cellular actirzity was advanced by T00LE ll98?l, r¡ho demonstrated
that during chick embryo derrelopment, cell morement takes place
ulthln extnacellular matrlces rich in hyaluronate. For example,
inraslon of the chick embryo cornea by mesenchymal cells colncldes
r¡lth lncreased hydration and suellÍng of the acellular stroma atr¡hich stage a maJor component of the extracellular matrix is
hyaluronate. Subsequenlly, however, migration ceases, the stroma
loses r;later, hyaluronldase actlvlty increases and hyaluronate
content of the extracellular stroma falls. lt ls at thls stage thatcellular differentlation starts. Toole suggested that hyaluronate
probably lnhlblts cell-cell lnteractlons, thereby permlttlng migration
and prollferation but delaylng the onset of diffenentiation until a

86
conrêct sequencÊ of tlssue organlzatlon could ensue. UNDERIIILL and
T00LE I l9?91 hare demonstrated a cell receptor for hyaluronate, tn
slmlan rlrus transformed 5T5 cells. Thls neceptor has a relailreþhlgh afflnlty. They càlculated that approxtmateþ 5 x l0! molecules
could blnd to each cell and that hyaluronlc acld r.uas pnobabþ bound
at sereral sltes.
The functlons that these receptorr ärÊ llkely to subserve àre)
flrstlyr the attachment of the penlcellular hyalunonate layen.
Secondly, thay are bellered to medlate the endocytosls of
hyaluronate before degradation by lysosomal hyaluronldases. Th.y
are also llkely to modulate metabollc functlons r¡ithin the cell, for
exampfe, the aforementioned lnhlbltlon of chondrocyte proteoglycan
synthesis.
llyaluronate has also been shourn to affect a number of inflam-
matory cell functions. Studies in this area have generally been con-
ducted at hyaluronate concentratlons similar to those found in the
extracellular spacË, i.Ë., l-1 mglml. Houerer more recently
lmportant obserratlons hare beer made using concentratlons llkely
to be found ln plasma 1 l,é., 200 ng/ml.
ERÊNDT I lg?1] reported that in solutlons of isola ted s¡novial
fluld hyaluronate, the lngestlon of monosodlum urate crystals by
perlpheral blood polymorphonuclear leukocytes uas inhlbited propor-
tlonal to hyaluronate concentratlon ove? the range 8,2 - l,O mglml
This effect uas diminishe d by decreasing the molecular uleight of the
hyaluronic acid to belou 106 uith mild hyaluronidase treatment.Houerer at a molecular uleight ó¡
= x 105, he noted an enhanced
uptake of crystals. F0RRESTER and EÊLÊZS [1980] have extended
these studles. Uslng ellclted mouse perltoneal macrophage
phagocytosis of latex spheres, they demonstrated that vrscous

Ðz
solutlons of hlgh molecular r.lelght hyalurontc actd (.1.6 x lgi -2.8 x
106) caused dose-dcpendent lnhlbltlon of phagocytosls orcr the
concentratlons 0.1 - ?.5 mg1ñ but, agaln, at the molecular rleight
of 9 x l0a, a paradoxlcal stlmulatlon of phago cytotls uJät teËn.
The lnhlbltlon of phagocytosls uas not present ff the cells r¡ere
pre-lncubated uith hyaluronic acid r¡hlch r¡as then removed by
r.'.lashing. Nor ulas it setn lf other large polyanions such as heparin
or chondroitln sulphate uJere substituted for the hyaluronic acid,
thus lndlcatlng that lt r¡as not a charge effect, Thry postulated
that the lnhibitory affect r¡as due to molecular lnteraction betueen
large coils of poþmeric polysaccharide prerenting the attachment
phase of phagocytosls.
Neutrophlls hare also been studled ln this regard by FORREIIER
and LdILKINSON IlgSl] r*rho reported that hyaluronic acid, ln the jame
concentratlon ränge, lnhlblted both dlnected and random locomotion
of neutrophils tn a dose-dependent and molecular-uelght dependent
manner. lnhlbltlon uJås more pronounced u.rlth higher molecular
r.uelght chemoattractant¡ such as casein, than lor small
chemoattractant peptides such as FHLP. ln addlticlrt, hyaluronic acid
inhibited the binding of the chemotactlc factor, denatured human
serum albumln, to the neutrophll surface. Thls effect úJas îeversible.
It appears, therefore, that at these concentrations the physical
effects such as molecular exclusion provide the best explanation for
the results obtained.
EÊLÊZS and DÊRZYIiKlElÅJICZ Í,19731 have examined the effects of
hyaluronlc acld on rarlous lymphocyte properties. They shor.,.red, using
a slmple migratlon inhibition assay, that concentrations of hyaluronic
acld slmllar to those found tn s¡novtal fluld tnhrbrted lymphocyte
moblllty. Ês rrlell, a number of other lymphocyte functions are

38
depressed, lncludlng the rÊsponsÊ to mltogens such as PllÊ'
l¡mphocyte dependent cytotoxlclty and experimental graft vÊrsus
host dlsease. Êgaln, each of these actlyltles ls inhlblted most
effectlrely by hlgh molecular t lelght hyaluronlc acld and uras
proportlonal to the concentratlon of hyaluronic acid.' They found
that the effect of hyaluronic acid uas cimilar to that observed by
decreaslng the cell denslty. For example, hyaluronlc acid utas more
effectlrze ln suppresslng ¡tlmulatlon of l¡mphocytes using pokerr.reed
mitogen and PPD, r¡hlch don't cautÊ signlflcant leukoagglutination'
than lt r¡as In suppresslng PllÊ lnduced stlmulatíon, urhere
leukoagglutination is ¡een. 0ther mänoÊurrcs that increase cell
dcnsity also tended to decrea¡e the effecllveness of hyaluronic acid
in lnhibltlng the stimulatlon of these cells. Other parallels betr¡een
the effect of hyaluronic acld and decreasing cell-cell distance urerË
seen such as delayed onset of DNÊ synthesis, delayed appearance of
blasts and the effects of both urere additire. They felt that in this
sltuation, hyaluronic acid may act to decrease cell-cell interaction,
although the possibility of interference r¡ith the dÌffusabillty of an
intercellular messenger such as intenleukin-Z could be an alternate
explanation.
I'IÊKÊNSSOH, llÊLLEREl,l and 1/Etl6E t 19801 have reported that lflou concentnations of hyaluronic acld (betr¡een 5-508 ¡.rgm/llten)
uJere lncubated r¡lth r¡hole blood, a stïmulation of vanious neutrophil
functlons uJås sËËn lncludlng phagocytosls, adherence, random
mlgratlon and chemllumlnescence. Ê simllar effect ls seen after a
subcutaneous inJectlon of hyaluronlc acid. They hav¿ subseque'ntly
determined that the serum cofactor necessa?y to see this
stlmulatlon ls flbronecttn [|iÊKÊNSS0N and PENGE, 1985]. Thls effect is
apparently specific for neutrophils since ÄnnSfnSSlÊDES and

89
ROBERTSON I l9B{] hare shou¡n that at slmllar lou concentratlons of
hyaluronlc acid, mltogen lnduced l¡trnphocyte stimulatlon ls lnhlblted,
although these authors separated thelr mononuclear cell fractlon
before lncubatlon r¡lth hyaluronlc acld thus not allol¡lng the
hyaluronic acld to interact ulth ierum flbronectin.
ln summaryr therefore, there arË ä number of lmport ant ways
in r¡hlch hyaluronate ls llkely to lnteract r¡lth cells. Êt concentra-
tions llkely to be found in extracellular matrixes (up to 2 mg/ml)
undegraded hyaluronate causes inhibitlon of a number of inflam-
matory cell functions including random and directed locomotion,
phagocytosls and lymphocyte stimulatlon. The llkeþ cause of theEe
effects is the entanglement of hyaluronlc acid molecules forming a
matrix. This ulll act to increase cell-cell distance and uill alter the
abllity of ranious factors to interact uith cells. Êt much lower
concentratlons, analogous to those found in plasma, however,
hyafuronlc acid, in concert ulth flbronectln, appears to cause
neutrophil stlmulatlon or priming.
E. Su¡s riêlljUslhyslur oniejgid.-Hyaluronic acid lsolated from any glven iource is polydisperse
and substantlal difference¡ ln the arerage molecular rrleight of
hyaluronic acld occur in fluids and tissues of the same species
ICLELÊND,lg?0]. Studies of the molecular ueight of nonmal synorrial
fluid hyaluronic acid are r¡ell represented by EÊLÊZS g!, al. [ 196?] r¡ho
studied pooled fluids from 268 normal adult male yolunteers. Using
analytical ultracentrifugation, lntrlnsic rziscoslty meàsurements and
light scatteringr they estimated an arerage molecular ueight of 6.5-
10.9 x 106 r¡hich is essentially fn agreement r¡ith others u.rho haye
looked at synorlal fluld obtalned from fresh cadayers UO|iNSTON, 1955;
DÊl-lL €t ê1., l985l.

40
RÊ6AN änd |-1EYER [ 1919] urere the flrst to report a decreace tn
the arerage molecular size of synorlal fluld hyaluronlc acid from
patlents ulth rheumatoid arthrltls. lt has been long appneciated
cllnlcally that r¡hole s¡rnorrial fluld from patients r¡tith lnflammatory
arthropathles has a decreased rlscosity. llouerer the hyaluronate
concentratlon is also decreased (see Table l) thus no inference
about the slze of the hyaluronic acld present can be made from this
obserratlon alone. RÊGÊN and IIEYER determlned a quoflent (log
tziscoslty/hyaluronlc acid concentnation) r¡hich uas linear oyer the
concentrations measured and uas dependent upon the degree of
polymerization. Thus rhey urere able to demonstrate that the
degree of polymerization uras decreased in a series of patients urith
rheumatoid arthritis. 0ther workers have used other techniques
including intrinsic rriscosity, anaþtical ultracentnifugation and light
scatterlng to confirm this obserration. There has, horr.reyerr been
some raniation in the estimates ol average molecular r.ueight forlnflamed synorzial fluid hyaluronic acid. These studies åre summarized
in Table l. lnitial reports suggested that the decreåse in arerage
molecular ueight of hyaluronic acid cornelated uith the degree of
lnflammatlon present [RÊ6ÊN and HEYER, l g.lg]. Thls, houerer, has
not been confirmed by other authors ISUNDELAD, lgSS¡ DÊl'lL É al.,
1ggsJ.
Êlso apparent from these studies uJas an increase in the Foly-disperslty of the hyalurontc acid present. lt is only tn relatively
recent years that reports have appeared that shor¡
chromatographic profiles of sSnovial fluid hyaluronic acid. BALAZS,
ERILLER and DENLINGER I l98l] fracflonated synovial fluid from
patlents I¡lth arthrltls on glycerallzed controlled pone glass columns.
They shoued the presence of a considerable amount of material in

Table l-l ÊPERÊEE H0LECULÊR hJElGl-lT ESTIÌ1ÊT|ONS 0F NORHÊL ÊND PÊT|-|OL0G¡CÊL SYllOt/lÊL FLUID
HORHÊLSYTIOYIÊL FLUID
HÊ Conc.
PÊTHOLOGICÊLSY]IOYIÊL FLUID
HÊ Conc Hâ l',ll¡J
0.08-0.850 decr.eased**
l.?3x106
l.?x106
1.2-1.5x106
dccr.ea¡ed**
1.8-2.15x106
1.2-2.2x1O6
5.2-6.8x106(avc=4.8)
Reference
I RÊGÊII & T4EYER [I94S]
2 SUHDBLÊD Irss3]
3 FESSLER, OGSTON & 5TÊTIIER II95{I
+ JoHHSToH 1955
5 BÊRKER et al. [1985]
6 CAST0R [rs66l
? BÊLÊZS et al. [196?]
I DÊHL et El. [19S5]
l'lethod'*mg/ml
HÊ HUJ
mg/ml
v
lv.
ll,trÊU
ÊU
TC
tv
ly, Êu, Ls
GC
r.36
?.9?
r .+- I 0.0
2.gr -3.5t
t.Ê1-?.25
r.45-5.t 2
2.85xt 06
I -,tx I 0õ
8.+-21xl0Ë
2.?5-{.0x10Ê
6.5-l tt.9xl06
6.5-?.6x106(are=?.0)
0.5-5
t.g?-8.0
1.05-1.,+5
<l.g
0.t ?-1.52
*V = viscosity; l!¡ = ¡ntrlnsic riscosity; TC = turb¡d¡ty cu-lve; ÊU = analytical ultracentrifugation; LS = light scattering;GC = gel chromatography;
'r* These methods shour decreased parameters, correlating to molecular r,reight, in the diseased fluids.rÈ)J

42
the molecular ueight fractlons 0.5 x l0l - 1.5 x 106. ln addlilon they
shoued at least trr.ro characterlstlc proflles of uronate-containlng
materlal, one unlmodal ulth sker,,.rlng into the louer molecular u.reight
fractlons and the other blmodal ulth the presÊnc; of a lot¡er molec-
ular r¡elght peak. BJELLE, ÊNDERSS0N and 6RÊNÊTH I l9E2] also
demonstrated conslderable heterogenelty ln molecular uelght profiler
of l? patlents r¡ith rarlou¡ form¡ of lnflanrmatory arthropathy.
Hore recently DÊllL Êt Ê1. I l9E5] utlllzlng a speclflc radloass ay lorhyaluronlc acld and therefore dolng away r¡lth the necesslty îor
potentlally degra datlve lsolation procedures obtained similar results.
These latter studies also failed to find a corelation betueen degree
of lnflammatlon and the amount of small moleculan r.reight hyaluronic
acid present.
D. STUDIES OII OXYGEI{ RRDICÊL I]IDUCED ]IYffLUROI{IC RCIDDEPOLY]îER I ZRTI OI{
l. lnltial Studies
Shortly after the enzqatlc depolqerization of hyaluronlc acid
r¡as desmlbed [CHÊl¡l and DUTI'||E, 1939; tlEYER, DUE0S and SllYTll,
193?l a number of reports appeared descrlblng the non-ÊnzWatic
degradatlon of hyaluronlc acld containfng substaÍrces. R0EEtrfS0H,
ROPES and BÊUER I lg39] described a spontaneour loss of viscosity n
fresh cattle ritreous upon standing. Because this tis¡ue has a high
ascorblc acid content they reasoned that this may be the "mucoþtic
agent" and subsequently shoued that both s¡novlal fluid hyaluronic
acid and extracted hyaluronic acid lost viscosity upon addition of
ascorbic acid in the presence of phosphate buffer [R0EERI50]1, ROPES
and EÊUER, 1939; l91l l. These uorkers urere quick to appreciate
that the process uJas irrerersible. lt r¡as soon shoum that a

43
numbËr of other polysaccharldes includlng pectln and starch could be
depol¡,rmerized in this fashlon [R0BERIS0N, R0PES and EÊUER, I g4l J and
that other organic reductants such ös pyrogallol and hydrogen
sulphide could replace ascorblc acid. HcCLEAIJ and IIALE I l91l]reported that ascorblc acid-induced hyaluronic acid depolymerization
uas inhibited by catala¡e, r.,.lhlch r.las important evidence thathydrogen peroxide played an intermedlate role.
The next maJor step in the elucidation of the mechanism of this
mÊans of depolymerizatlon Lras taken by SKÊllSE and SU|.iDELÊD [ 1913]
urho shor^led that ascorbic acid-lnduced depolymerization of
hyaluronlc acid dld not occur tn an anaeroblc enrlronment, but
proceeded as soon as ot(,ygen L'.,a5 added. They also reported thatthe addltlon of coppËr greatly enhancrd the effect. ln 1919 JEi|SEH
reported that ferrous ions alone or femic ions in the presence of a
reducing agent (hydrazine sulphate) could degrade hyaluronic acid.
There r+,as considerable debate in these early papers as to the role
of hydrogen penoxide and ascorblc acld. Indeed part of this confusion
has subsequently been resolred because ule nouJ realize that the
phosphate buffers used in these experiments contain iron in sufficient
quantity to cataþze a Fenton reactlon [h,0ll6 É ê!., lgBl].
Ê major concern at this tlme r¡as the unexplained degradation
of hyaluronic acid associated l¡ith various extraction procedures and
"presenratires". Êysteine, f or example, [¡Jas used to actirate papain
r¡hlch r¡as utilized in the extraction of hyaluronlc acid from various
organic sources and merthiolate LJas used as a preservatlve. Eoth
could act as reductants IPIEHÊN and RlZVl, 1959; 065T0H and
Sl-lERt1AN, 19591. Perhaps of more interest uas the depolymertzatisn
seen upon lyophlllzatlon. This uas noted by P|6HÊH and RIZVI [1S59]
to be accelerated by the presence of phosphate ioni. Subsequent

44
urork by LTJEDLOCK Êt El. I l9E5] has shor,m that the þophlllzatlon
procedure is assoclated r¡ith the appeärancÊ of radicals as detected
by electron spln reronänce spectroscopy.
Z. Radiatlon lnduced llyaluronlc Êcid Degradatlon
RÊ6ÊN gt è1. I l9{?] urere the first to report that the vlscosity
of both human synorzial fluids and solutions of hyaluronic acid urerÊ
dlmlnlshed after exposure to x-lrradlatlon. EÊLÊZS and LÊURENT
I l95l] demonstrated a slmllar effect sËcn upon Êxposure to IJV llght.
Ntlther UP nor x-lmadlatlon r¡ere assoclatcd r¡lth any change ln the
polyanlonlc charactrr of hyaluronic acid, thus indicatlng that the
carboxyllc acld groups uJËrË not affected.
EÊLÊZS Êt d. [ 1959] reported that the molecular ueight of a
hyaluronlc acid preparation, calculated from sedimentation and
diffusion constants, decreased from 80,000 to l9r7Ûg after
inradiation r¡ith U/ light, l-le has al:o reported the appearance of a
dialyzable fraction after x-imadiation suggesting a soîìeurhat smaller
end product. SUNDELÊO and EÊLÊ25 Ll96El have suggested however
that the rlscosity lor.lering effect of lor¡ doses of x-irradiation may
not be solely due to depolymerizatlon of the macrofiiolecule but also
mäy be due to changes ln shape, deformablllty or lnternal structure.
EÊLÊZS Ê! ê!. [1959] hare reported a decreas¿ n totalhexosamine and ln total glucuronic acid of hyaluronic acid exposed to
0.5-4 x l0ó rads, thus suggesting a degree of destruction of these
sugârs due to lnradiation uith electrons. Reducing substances,
meäsured by the femicyanide method, lncreased after UV ircadiation.
Degradation of hyaluronic acid by x-irradiation could be
prerented by the addition of sodlum thiosulphate, an effect belteved
to be medlated by the scavenging of hydroxyl radicals,[ERIHKHÊH glê!., I 96l l. BÊLÊZS et al. I l9E?] reported the presence of transient

45
radlcal intermedlate¡ after the pulse radlolysls of âqueous solutlon¡
of hyaluronlc acld as detected by electron pärämagnetlc rÊsonancÉ.
They postulated that the radlcal lntermedlates obs¿rv¿d could be
best explalned by cleavage of the gþcosldic linkage and alro hydrogen
abstractlon from the E, carbon of the glucuronic acld moiety r¡ith
subsequent resonänce stablllzatlon rllth C..
5. pelt¿Esrizê3lgL(ORD Reaction).
This term t¡as formulated by PlGtlÊN's group uho publtshed a
number of papers on the non-enzt¡matic depoþerizatlsn of
hyaluronic acid ln the 1960's. These uorkers uJere among the flrst topostulate that ascorbate-induced hyaluronic acid depolymerizatton
r¡as due to an oxygen radical reaction, Seyerd lines of evidence led
to thls concluslon. Flrstly, they clted the fact that x-irradiation of
aqueous solutions of hyaluronic acid leads to depolymerizallon due to
the generation of radicals (the hydroxyl radical is generated along
r¡lth hydrated electrons). The requirement for oxygeryr and the
presencÊ of a reducing agent suggested that oxygeî radlcals may be
found r¡ith the ORD reaction. Secondly a number of hydroxyl radical
scarengers including ethanol, toluene and D-glucose kJere inhibitors of
the ORD reaction. Finalþ electron spin resonance studies
demonstrated the presÉncÊ of free radicals upon the oxidation of
ascorblc acid in the presÊnce of molecular oxygen ILAEERÊRÊNTZ,
1gE.f l.
llÊRRl5, HERP and PlGllÊH [19?1] demonstrated that Fe(lI)-EDTÊ
complex could be regenerated from Fe(llI)-EDTÊ at a platinum
electrode (cathode) and thls uould depolymerize hyaluronic acid and
alginic acid. This prorzides further support for cycllc reduction r¡ith
subsequent autoxldation of Fe(ll) hyaluronic acld depolymerization
induced by ascorbate and other reductänts.

46
A. EnZy¡gtlgÀlly Produced 0xp$en Radicals and lnhtbltors
HcCORD I l9?{] uas the flrst to repont the effect of an
enzymatlc oxygen radlcal generatlng system (X0/llx) on slrrìortal flutd
and hyaluronlc acld solutlons r¡hlch demonstrated loss of riscoslty.
Uslng thls system he shor¡ed that both supcroxlde dlsmutasa and
catalast prerentad tha loss of rlscoslty at catalytlc concentratlons
(superoxlde dlsmutase I ¡.rg/ml; catalase 0.05 ¡rg/ml). lle reasoned
that slnce both supenoxlde dlsmutase and catalase alone inhiblt thls
reactlon thcn nelther the superoxlde anlon nor hydrogen peroxldà
could be the depoþmerlzlng agent but must be present
slmultaneously for the process to occur. The impllcation being thatin this sytem the actual depolymerizing species is generated
secondarily by a reactlon betureen superoxide anion and hydrogen
peroxide. The most likely candidate being the hydroxyl radical
generated by a l-laber-l,rJeiss reaction (see page 7), llannitol, a
scärengen of the hydroxyl radical, u.ras also found to prevent loss of
rlscosity in hls system. llÊLLIITJELL [19?E] conflrmed these findings
t¡ith X0/llx uslng the hyaluronlc acid solutlons and de'rnonstrated the
dependence of the observed reductlon ln vlscoslty upon the prerence
of lron salts, albelt tn trace quantlties.
5. l¡nportance of Hetal C fllnflanrmatorr.r Drugs
l'lÊLLlt¡JELL [19?E] and subsequently EETT9 and CLELÊND I lgBZ]
hare shoun that the r.lay the metal lons are chelated is of vitalimportance in both the X0IHX induced depolymerization and other
systems dependent on the oxldaflon of fercous ions (e.g., ascorbic
acid induced hyaluronic acid depolryerizatlon in phosphate buffer). lfthe chelator binds the Fe(lll) tightly (as uith the chelators Eps,
DETÊPÊC or desfemloxamfne) then thls milltates âgainst the reduction
of Fe(lll) and reductant-driren iron-dependent hydroxyl radical

4r
production i¡ inhibited. 0n the other hand iron chelators r,.rhich allor¡
iron to shuttle betr¡een Fe(lll) and Fe(ll) by transfer of electrons
r¡ill facllltate the neaction. The concentration of metal and chelator
are of importance in detenmining this effect fon example in the XO/l-lX
system. EETTS and ELELÊND I l9B2] hare shor¡n that the
antirheumatic agent D-penicillamine can act as a chelator and u.¡ill
inhlbit hyaluronic depolymerization in concentrations greater than
5m11. CIn the other hand in the ferrous ion autoxidation syslem atconcentrations of less than 5ml1 D-penicillamine is stimulatory but athlgher concentrations is inhibitory. Other antirheumatic drugs
lncludlng salicylate, chloroquine, gold sodlum aunothiomalate and
indomethacin r¡ere in inhibltory ln both systems.
6.EflÈçl-sf¡s@l'lydroxyl radlcal productlon appeärs to proceed by dlfferent
pathurays in the ,19ll1>1 system and ln systems dependent on the
autoxidatlon of femou¡ ions. The clearect evldence for this ís thatX0/l'lX induced hyaluronic acid depolymerization is inhibitable by
superoxlde dismutase but ferrous ion autoxidation and ascorbic acid
induced hyaluronic acid depolymerizatisrt are not [HOFHAltlN and
SC|-|NUT, 19Ê01. This is also the case uith ascorbate índuced Dl{Ê
depolymerization [|.40R6ÊN, C0NE and ELIìERT, l9?E]. IJIFfIERE0URI'|E
[19?9] has suggested that iron-EDTÊ catalyzed hydroxyl radical
production from hydrogen peroxide and ascorbate could proceed by ^
mechanism largely independent of superoxide, uhereas the reduction
of Fe(lll) to Fe(ll) m the XU/l'lX system is effectedby superoxide
anion [8Efi5 and CLELÊI|D lïEZl. 0ther dlfferences exist betr¡een
these tr.rro oxygen radlcal generatfng systems for example X0 can
effect the dlrzalent reductlon of oxygen as r¡ell as the unirzalent
reductlon and the relatlrc amount of superoxide anion and hydrogeïr

48
peroxide producÊd directly are dependent on pll, pO" and substrate
concentration IFRlDOl/lCl-|, I 9?CI].
?. Deqnadation of Hualuronic Êcid bu Neutroohils
----GREENhJÊLD and t10Y I l9E0] reported that phorbol myristate
acetate (Pl'1Ê) stimulated neutrophils induced a progressive decrease
in riscosity that r¡as abolished by addition of superoxide dismutase
on mannltol. These cells do not contain hyaluronidase ISODER, l9?0].
Funther uork from this group IBREENIIÊLD and H0ÊK, lg84l has
demonstrated that Fe(ll) or Fe(lll) are required, and thatstimulants other than PIlÊ (¡ncluding zymosän, Eon Ê or Fl"lLP) nequine
added hydrogen peroxide. They also demonstrated that neutnophil
lysates do not lnduce loss of rriscosity such as r¡ould be expected if
thls r¡ere due to a simple enzyme effect.
E. llolecular lrjeight Changg
PlGtlÊN and RIZPI [1959] did not detect dialysable material
aften treatment of hyaluronic acid r¡ith ranious oxidation-reduction
systems yet they did, horr.lÊve?, detect a broadening of the
hyaluronic acid peak on ultracentrifugal and electrophoretic analysis,
similar to mild treatment r¡ith hyaluronidase. These results indicate
that a true depolymerization occurs. This conclusion is alEo in
keeping uith the obserration that the changes in intrin¡ic viscosity
urere irrerersible [Pl6PlÊN, RIZll and l-l0LLEY, 1961]. EREEI{IIJÊLD and
l10Y I l9E0] uJere the first to publlsh gel chromatograms of hyalurontc
acid expoEed to any of these axyg¿n radical generating systems.
They shoued that their lnitial ppËparätion of (human umbilical cord)
hyaluronic acid rr.ras large enough to be excluded from Sepharos e 2B-
cL. Êfter elrposure to X0/llX or to Pl'lÊ stimulated neutrophils,
materlal had a reduced hydrodynamlc slze being retarded
throughout the column. Ho such information uas available until this

49
current study [tlcNElL Ê! ê!., 19ts5] for the ferrous ion autoxidation
system. EREEN|dÊLD and t10Y I I gEO] shou¡ed the presÊnce of uronic
acid containing material rzirtually to the /1 on Sepharose ZB-EL lKav
. = 0.8) corresponding to a molecular r.leight of approximatdy ?-3 x
l0s. This uas achiered using a level of super oxide anion generation
that they had calculated could be attalned r¡ithin an inflamed joínt.
They reported that part of their reactlon product became dtalyzable
but the llmiting pore size of their dlalysls tubing u¡as not given.
Studles of the generation of reducing endE from hyaluronic acSd
after exPosurÊ to oxygen radicals have provided conflicting results.
Early studies r¡ith ascorbate induced hyaluronic acid
depolymerization suggested that reducing ends kJÊre not generated
lPlcLEÊN and llÊLE, 13411. Houerer, ELELÊI|D Ë! e!. [ 1969]
subsequently detected reducing ends using sodium borohydride
labelling ln hyaluronic acid depolymerlzed by ascorbic acid. GREEHITJÊLD
and l"l0Y [1980] dld not detect reducing ends an X1/llX exposed
hyaluronic acid.
9. Eonclusion
llost inrzestlgatlons into the oïygen radical induced hyaluronic
acld depolymerizatlon systems have used change in viscosity as an
index of depolymerization -and have centered upon mechanisms of
oxygen radical production. Relatirely little information exist s as tothe molecular r.rlelght changes and conflicting information exists as tothe generation of reduclng ends ln the reaction product.
The molecular r.leight of hyaluronic acid is however altered ín
inflamed synoruial fluid hyaluronic acid. ln this investigation I have
therefore posed the follouing questions:
l. hJhat are the moleculan u.reight changes induced ln
solutions of hyaluronic acid by jn ritro oxygen nadical generating

50
systËms that resemble those in the inflamed joint? (i.e., femous ion
autoxidation, X0/HX, activated neutrophils)
2. l,rJhat are the molecular rrleights of hyaluronic acid
extracted fnom lnflamed Joints? Could any neduction in size be as a
result of oxygen radical generating systems?
5. Êre reducing ends generated by exposure of hyaluronic
acld to oxygen radicals?
1. ls the biologlcal activlty of hyaluronic acid altered if it is
depolymerized by hyaluronidase or an oxygen radical system?
5. hlhat happens to synovial cells r¡hen they are exposed tooxygen radicals and in particular to the hyaluronic acid that they
produce?

5t
C]IRPTER I I
T}IE DEPOLYIIERIZfiTIOÌI PRODUCTS OF }IYÊLURO]IIC RCID RFTER
EXP05URE T0 oDFR !Ë ytTRo

t2
R. II{TRODUETIO]I
lluch information about the molecular r.leight changes in
hyaluronic acld exposed to oxygen radical generating systems har¡e
been inferred from changes in intrinsic riscosity. Ên exception to this
is represented by the tr,.ro Sephanose ZE-CL chromatograms
published by IìREENIJÊLD and l10Y I l9B0]. Thelr data cleanly
demonstrate an incnease in polydispersity but they do not shor¡
sequential changes, nor do they demonstrate the effect of a
maximal oxygen radical flux. Sereral studies [Pl6l1ÊN and RlZPl, lg53;
EREENhJÊLD and 110Y, lgÊ01 suggest that products as ¡mall as ffono-
and disaccharides are not obtained follor.rring ü1ygen radical induced
depolymerization. Ho¡¡eyer the size of the smallest products
obtainable has not been established.
This inrestigation uas therefore undertaken to define by gel
chromatography:-
l. the sequence of molecular r¡eight changes seen after exposure
of hyaluronic acid to increasing levels of oxygen radical flux.
?,. to determine the molecular r.leight of the smallest degradation
products thus obtained.
3. to determine the effect of re-exposure to a second oxygen
radlcal flux.
1. to determine changes in an umbilical cord hyaluronate
preparation uith a characterized starting molecular r.rreight.
ln onder to obtain a r¡ide range of relative hydrodynamrc size
resolutions three gels rr.rere used:- Sepharose ZE-EL (included rolume
for polysaccharides 105 - 28x10õ), sepharose flB-EL (3x1oa - Í¡x1o6)
and Sephadex Gt00 (103 - 105) [pttRRtlÊctÊ - PR0DUET lF{F0Rt1ÊTt0]-tl.

53
Three oxygËn radical generating systems, each of possible
biological relerance uJere studied i.e. the ferrous ion autoxidation
system, the X0/HÅ system and PF1Ê stimulated neutrophils. The
generation of 0ll' is thought to proceed by different mechanisms in
each of these systems, as discuEsed in Chapter l.
llolecular rleight r¡as al¡o determined by analytical ultra-centrifugation as än independent confirmation of the results of gel
chromatography in the ferrous ion autoxidation system.
ln addition, a purified sample of hyaluronate was prepared from
the commercially arzailable human umbilical hyaluronic acid in order tobetter reflect the molecular r.leight spectrum of normal synovial flu¡d
hyaluronic acid.
E. HÊTERtÊLS Êt{D HETH0DS
Hyaluronic acid (grade lll from human umbilical cordJ and xanthine
oxidase (grade lll fnom buttermilk) r,.lere obtained from the Sigma
Ehemical Eo., 5t. Louis, l1o. Sepharose EL-28 and EL-18, Sephadex E-
100 and Sephacryl 5-100 brÊre obtained from Pharmacia, Upp:ala,
Sr¡eden. Êll other chemicals brere of the highest purity available
from Sigma or ÊJax Chemicals, Sydney, Êustralia.
I . H,ygI¿ronls--ËEld_ps¡flsÊSjsÐ
l-lyaluronic acid r¡as further purified by pasøage through a
Sephacryl 5-100 preparatire column (rrolume ZEE mls) using 0.5 l-1
sodium acetate as the buffer. Fractions r¡ith a Kar betueen 0.1
and [1.25 ulere pooled, This represented a hydrodynamic size of
betueen ?x1O6 and 5x l0s. Protein contamination uaE reduced toless than l% wlv by predigestion uith pronase, a procedure shoun
by EÊRT0LD, l,',llEEKlN and Tl-l0NÊRD [1981(b)] to have no detectable
effect on the hydrodynamic slze of hyaluronic acid.

54
2. !¡¡¿gen radical Benenation
a. Ferrous lon autoxidatlon
The reaction mixture comprised:
(l) hyaluronic acid, I mg/ml
(Z) potassium phosphate buffer, 50 mH, pll ?.4 .v
(3) ferrous sulphate at concentrations rrarying betr¡een 5
¡.r11 and 1000 ¡lt1 .-.,,.. ,o û_,,.
(1) EDTÊ in a molar ratio to ferrous ions of l:1. ./r,ì,:r,ir.:i.i .
Stock solutlons of hyaluronic acid r¡ere prepared at an initial
concentration of 4 mg/ml ln sterile distilled r¡råtep. This slightly
cloudy solution r¡as cleared by passäge thnough a l.? l.r Hillipure
filter and r¡as then stored either fnozen or at {oC until nequired.
Prlor to use, a solution of Z mg/ml hyaluronic acid in 50 ml1
phosphate buffeF urås prepaned by the addttion of an equal rolume
of 100 mll potassium phosphate buffen to the inttial 4 mg/ml
hyaluronlc acld stock solution. Stock solutions of l0 mÌl EDTÊ urËre
prepared in phosphate buffer and stock solutions sf l0 mt1 and 1 mt1
fenrous sulphate uJerÊ prepared in u.rater, The reaction rolume in
each case uJas l.? ml. Under these conditions and in the presence of
atmospheric oxygen, autoxidation of ferrous ions pnoceeds r¡ith the
production of the hydroxyl radlcal (OH.J, the agent neponted to be
directly effecting hyaluronic acid depolymerization [llÊLLlldELL, lgTE; 1,..i.r,Ì'
EETTS and ELELÊND, l9Ë81. -The neaction proceeds to completion in
less than 5 minutes IEETTS and ÊLELÊND, lgB2].
b' X.g/l-lx
For the enzymatic system the reaction mixture comp risedz
(l) hyalurontc acid, I mg/ml
(2) potasElum phosphate buffer, 50 mH, pl1 7.i

55
(3) l'lx, E mll
(1) X0 at concentrations varylng betu.reen 5xl0-3 and I lJlml
The action of X0 on HX under aerobic conditions j¡ vitro leads
to the production of superoxide anion and hydrtrgtrt peroxide in
addition to xanthine and subsequently uric acid, Trace quantities of
lron, present ln the phosphate buffer used, allor¡ further reactions
leading to the production of hydroxyl radlcals IBETTS and CLELÊi{D,
l9EZl. The reaction uaE allor¡ed to proceed to completion (greater
than tr.renty minutes) [gETfS and CLELÊND, l9E2].
c. Stimulated poll,mor@For the Pl1Ê Etimulated polymorphonuclear leucocyte system
the reaction mixture comprised:
(l) hyaluronic acid, I mg/ml
(Z) Dulbecco's phosphate buffered saline, pl'l ?.{
(3) EDTÊ, 60 ¡M('î) ferric chlonide, t 0 ¡.rH
(5) phorbol myristate acetate (Pt1Ê), 200 ng/ml
Elood. r¡as obtained by rrenipuncture from healthy labunatory
personnel and r¡as anticoagulated r¡ith EDTÊ. Polymorphonuclean
leucocytËs uJere separated by lor.'.r speed centrifugation oyer Ficoll-
llypaque ln a single step procedure as outltned by FERRÊNTE and
Tl'ltlNG I lg?E]. The cells trene then r¡ashed three times in Dulbecco's
phosphate buffered saline. lmmediately prior to the experiment,
riability uräs ässessed by tnypan blue exclusion. 0nly cell
preparations exhlbiting greater than 9596 exclusion uJere used. Pl'1Ê,
200 ng/ml, r.,las used as the stimulant and superoxide production r¡as
confirmed by obserring the neduction of nitroblue tetnazolium [SEGÊL,
l9?'11. Graded fluxes Lrere obtained by varytng the number of cells

ã6
added to the reaction mixture betr¡een 0.Exl06/ml and l2xl06/ml.
The cells bJere incubated at 37oE for 3 hrs before the cells ¡¡ere
sedimented by centrifugation for l0 minutes at 1000 g. The
vlscosity of the supernatant r¡as then measured.
5. l/iscometrl¡-
The riscosities of solutions of undegraded and degraded hyalur-
onate urere meäsured simpþ and nepnoducibly by using a I ml
tuberculin syringe and a ?0 gauge needle at ZoC according to the
method of EEfiS and ELELÊND I l gBZ]. Relatir¡e riscosities rrene
calculated by dirziding the time taken for 0.Ë ml of test soution todrain from the syrlnge by the time taken for the same volume of
buffer to drain. Specific viscosity (llsp) uas then calculated by
subtracting I from all the relatire rlscosities (1.e., subtracting the
contribution of the buffer). Changes in Nsp were expressed a¡ the
percentage ratio of degraded material to undegraded material. The
t¡iscosities of solutions of oxygen radical exposed material r¡.rere
determined at 2 hrs for solution: exposed to the ferrous ion
autoxidation or XO/l'lX systems, and at 3 hrs for solutions exposed
to the cellular system.
4. 6el chromatograp_hy-
Êliquots of O.25 ml of the reaction products urere fractionated
on Sepharose 2Ê or 4E and Sephadex6-100 (30 cm x 0.? cm). Frac-
tions of 0.? ml I¡ere eluted at ? ml/hr r¡ith 0.5 l"l sodium acetate atpll 5.8. Characterizatlon of the columns uas determined r¡ith
Dextran Blue, S3s-sulphate and glucuronic acid. ln addition, ín order
to rerify the resolution of these small columns a series of standards
uJas run through both large (i.e., 1.5 x 90 cm) and ¡mall columns and
the K¿y's of the profile peaks compared.

67
5. Uronic acid anal,usis
Uronic acid uas estimated by the meta-hydroxydiphenyl method
of ELUIIENKRÊ]-|TZ and ÊSB0E-|-|ÊNSEH I tg?3] (see Êpp endix I ) or amodlfication of the automated method of R0SENTI-IÊL, BENTLEY and
ÊLBIN I Ig?EJ.
E. Ênalytical ultracentrifugation
The sedimentation coefficients of intact and depolymerized
hyaluronlc acld samples rJepe determined on a Beckman llodel E
analytical ultracentrifuge. Samples (0.5 ml in 58 mt1 phosphate
buffer) urÊre centrifuged at 59?Ê0 r.p.m. (Zïgrg1} g åt the cell
centre) in a l2 mm single ¡ector ccll r¡ith plane quartz uindor¡s in an
ÊnD rotor at lÊoÊ for up to ? hrs, Photographr of Schlieren
patterns uJere taken at E minute interrals and images measured on
a Nikon profile analyser r¡ith a magnification cmEtant of 2.5, The
arerage molecular r.r.reights of the peak material kJere calculated by
neference to standard preparations of hyaluronic acid kindly provided
by 0r. 11. llather¡s of Ehicago and to the ralu es derived from the
data of ELELÊND and kJÊNÊ [ 19?0] as r¡ell as a standard preparation
of l-lealon analysed by BÊRT0LD, lrlElEKlN and TI'|ONÊRD Ilg8l(b)].C. RESULTS
l. Piscositu
Ê substantial decrease in llsp r¡as obs erved rJpon exposure of
hyaluronic acid to oxyge'n radicals uhether generated by the
autoxidation of femous ions, enzymailcally r¡ith >18 /t1x or by
incubation r¡ith Pl1Ê activated polymorphonuclear leucocytes (Table
?-1). The decrËases in viscosity seen uJere proportional to the
intensities of the fluxes generated r¡ithin each system. Follorr;ing
exPosure to fluxes of maxlmal lntenslty the vlscosity of hyaluronlc

58
Table 2-l 1055 0F lzlSCOSlTY INDUCED EY EÊCl'l 0XYRÊD|CÊL EENERÊTINE
5Y5TET1
Autoxidrtion of ¡(0/Hlt fictirrtrd nrutrophibfrrnou¡ lon¡
Fr lonconc. (pl"lì
H¡P(r)
t{0(U/ml)
Crll conc.(cdl,¡lml)
Ìlrp$t
lkp(ß,
r00
65
60
56
6?
65
98
0
25
50
100
e00
500
r00
{?3lt9lt1
0
0.05
0.t
t.0
t00
2Í22
t2
0
0.6 x 106
6 x l0ól? x 106
te x toG (+soD)
12 x l0ô (no PMfi)
tu x t0õ ({oc)
ln thc fmnou¡ ion ¡utoxidrtion hyeluronic acld, I rng/ml, r¡Jrs rxposrd to fennous ionsin thc pFÇsenct of 50 mH potassium phosphetc buffer, pH ?.{. EDTA was pnesent in arrtio to fcrrsuc ions of l:1. In the X0/lll{ systcm, hyaluronic rcid, I mg/ml, r,.ras
exposod to ll0 in thr prtsêncc of potessium buffcr, pH ?.{, and llX, 6 ml'l. ln thecollulan syrtcm hy*luronic ecid, I mg/rnl bns rxpo¡rd to cclls ln thc presence ofDulbçcco'¡ phosphetr buffercd srlinc to uhich hrd brrn rddrd FrClg l0 ¡rM, EDTÊ 60
¡M end PHÊ 200 ng/ml. SOD r¡rs u¡rd rt e concrntnrtion of 60 units/ml.

59
acid approached that of the buffer alone. The decrease in Nsp using
the cellular system hras partly inhibited by superoxide dismuta¡e
thus implying that superoxide ions and/or derived oxygen radicals
uJeFe, at least in part, responsible.
2. llolecular exclusion gel chromatograp[y-
a. Ss,Fh.e.Cs¡-È.!00
Samples of hyaluronic acid r¡hich had been exposed to
different oxygen radical genenating systeTns urÊre applied to
Sephadex 6- 100 columns. fln elution r¡ith 0.5 N sodium acetate,
hyaluronlc acid from preFarations r¡ith Hsp's of lO% of control, or
greater, urere largely excluded from the gel, t¡hile a major proportion
of the hyaluronic acid from lor¡er viscosity samples had a hydro-
dynamic size sufficient to be retarded by this gel. The smallest
products uJere seen at a Kay of 0.66 uhich corresponded to a
molecular r.leight of 104 (Fag. Z-ll.Ê further experlment uas performed to determine r¡hether the
Êxposure of hyaluronic acld to tr.ro sequential otlygln radical fluxes
r¡ould reduce the hydrodynamic size yet further. llyaluranic acid
r¡as first exposed to the XE/11>1 system and an aliquot r¡as then
chromatographed (Fig. Z-?). The remainder r¡as then exposed to the
ferrous ion autoxldation system using I ml1 ferrous sulphate, After
this double exposure to an ffiygsîr radical flux no material r¡as
obserred past a Rav of 0.68 on Sephadex E- 100 although the overall
change in shape of the proflle seeî on Sepharose 4E r¡ith acharacteristic shift tor¡ards the total column volu'me (V¡) tndicated
that some further depolymerization of larger molecules had
occurred.

FIEURE Z- I SEPHÊDEX Iì- I OO CHROIIÊTOERÊPHY OF HYHLURONIE ACID
EXPOSED TO TTIE FERROUS ION ÊUTOXIDÊTION SYSTEÌ1
l'lyaluronic acid, I mg/ml uràs exposed to fernous ions ata concÉrìtratlon of 50 ¡rt1 (Nsp = ?B%) and I mH (Nsp = 6g¡
in the prËsÊnce of 50 ml"l potassium phosphate buffer' pH
?.4. EDTÊ uJäs ppÊsant in a ratio to ferrous lons of l: l.

Nsp IOO5
1
3
2
I
o
5
4
3
2
t
o
5
I
3
2
I
20 25 30
20 25 30
ro
ro
t5
r5
5
ôtIox
-E
oE
zIl-gÊ,]-zll¡(Jzouô-U
IzoÈÊ,3
Ntp 78
5
Ntp I
or0
Vo5 3025
Vr20t5

FIEURE Z-2 |'|YÊLUR0NIE ÊClD EXP0SED T0 Tl¡J0 SEOUENTIÊL 0XYBEN RÊD|CÊL
FLUXES.
llyaluronic acld urat "xposed
to 0.5 unitc/ml X0 and 6 mH
l'lx in 50 mH phosphate buffer, pl'l ?.4. Ên aliquot r¡ar then
fractlonated on SapharotÊ 4E and anothcr uas frac-
tionated on Sephadex 6- 100. These profllas are rhorgn ln
the middle panel. I'lyaluronic acld contalning fractlonl rJËrÊ
then poolcd, dlalyzed agalnst urater, thctt dried in Z.e¡¡¡g
ov?? phosphorus perrtoxide beforÊ ÊxPoiure to the ferrouc
ion autoxidation using a concÇntration of ferroug ions of I
mll. Profllec obtained after fractionation of this material
on the ramÊ gels is shoun in the bottom panel.

SEPHAROSE 1B
UNE)OOSED HA
ro 15 20
HA EXPOSED TO xO/H
r0 l5 20 25 3025 30 5
x
25
THEN
30
SEPHADEX GIOO
ôaI
9x
çË
5
1
3
2
I
o
5
1
3
2
I
o5
E
zIÉ,
zt¡¡UzoUô6
IzoolD
5
I
3
2
'|
0
5
1
3
2
5
5
r0 15
HA EXPOSED TO
l0
vor5
3020 510 152025
TO FERROUS ION AUTOXIDATION
l0 t5 20
vo
30
30
5
1
3
2
I
0
5
I
3
2
xo/nx
V¡
25
V1
52520
FRACTION

6Z
b. EPierus-1EFractionation of hyaluronic acid (from human umbilical cord,
Sigma type lll) on Sephacryl s-400 provided a nominally high
moleculan rrreight starting materiall however there r¡as still same
retarded material present on Sepharose 4E chrom atography, Using
this material, a series of chromatographic profile5 LJas obtained for
graded fluxes generated by the ferrous ion autoxidation, the HOlt¡lÅ
and the stimulated polymorphonuclear leucocyte systems. They
appear as Fig. ?-3.
The effect of an increasing anygen radical flux on these high
molecular uleight hyaluronlc acld preparations in each of the axygen
radical generating systemi, rrras to sh¡ft the amount of uronic acid
containing material from the rzoid rolume (V6) to the íncluded rolume,
i.e., t0 the night. Êny shift of the chromatographic profile to the
right uJas accompanied by a decrease in Nsp in each of the
generating systems. The greatest decrease in viscosity was ¡een in
the ferrous ion autoxidation system. l-lere an Nsp of 5% of control
r¡as obtained and at this r¡isc osity no uronic acid containing material
uJas monitored at Vs. Ê significant amount of material uJas' however
retarded by the gel (i.e., Kav = 8.17 corresponding to a molecular
rrrelght of ?xl0s). Êlso, r¡ith an Nsp as lou as 3?% of control, uronic
acid containing material remained present at Vs ndicating the
presence of material r¡ith a hydrodynamic size of .l06 still preserrt in
this sample.
llyaluronic acid exposed to the stimulat ed polymorphonuclear
leucocytÊs remained predominantly excluded by the gel orer the
rånge of viscosities achiered, however there wal an increa¡e in the
amount of material seen in the included rolume and in the samples
r¡ith Nsps of 639' and 56?ã of contnol, a small amount of material r¡as

FIEURE Z-5 SEPHÊROSE '{B CIIROIIÊTOBRÊPTIY OF I'IYÊLURONIE REID EXPOSED
TO EÊEII OF TTIE OXYEEN RÊDIEÊL BENERÊTIIIE SYSTEI'IS
ln the ferrous ion autoxidation system hyaluronic acid, I
mg/ml, bJas Êxposed to ferrous ions at concentrations of
100 ¡.rH (Nrp = 5l%l and I mH (l{sp = 5%) 'n the prese'nce of
50 ml-l potassium phorphata buffer, pH ?.4. EDTÊ r¡a¡
prescnt in a ratio to ferroug ions of l:1.
ln the X0/llX system, hyaluronic acid, t mg/ml, r.tas
expored to X0 at cørcertrations of 0.1 unit/ml (Nsp = 32%)
and I unit/ml (Ì{ry = ?E%J in the prÊiÊncÊ of 6 mH llX and
50 mll potassium phosphate buffer, pll ?,1.
ln the cellular system hyaluronic acid, I mg/ml, rr.ras
exposed to concentrations of polymorphonuclear leukocytes
of 0.6 x l0ô cells/ml (Hsp = El%J and l? x IEG cellslml (Nsp =
íEZ.J in the prÊrfllce of Dulbecco's phosphate buffered saline
to r¡hich had been added FeClS l0 ¡rÌ1, EDTÊ 60 ¡.rl't and PHÊ
200 ng/ml.

IERROUS ION AUTOXIDATION
Nsp IOO
ro r5 20 25 30 5
Nsp 5l
r5 20 25 30
Nsp 5
r0vo
t5 20
HYPOXANTHINE STIMUIATED POTYMORPHONUCTEARTEUCOCYTES
Nsp 100XANTHINE OXIDASE
Nsp lO0
Nsp 32
lo l5
Nsp 20
ôt'gx
E
oE
zIÉz¡¡¡uzoue|.)
9zoÉ.l
5
1
3
2
I
0
5
a
3
2
I
0
5
a
3
2
I
o
5
I
3
2
I
o
5
1
3
2
t
o
15 20 25 30r0 t5 20 2s 30 5 to
Nsp ó3
Nsp 5ó
5
5
1
3
2
o 20 25 30 lo5 t5
ì5 20
20 25 305lo5
5
1
3
2
I
0
5
I
3
2
I
0
5
4
3
2
I
0 25Vt
ì0Vo
530
53025vr
5t0\b
15 20 25Vt
30
FRACTION NUMBER

64
present at a Kar = 0.6Ê r¡hich r¡as not present in the starting
material.
c, &PhaCeåe 2E
Ê large series of hyaluronic acid ¡amples rrler e exPc.sed to
the ferrous ion autoxidation system in order to resolve changes in
the molecular ueight spectra induced by graded oxygen radical
fluxes. Small increments in fernous ion concentnation (and thenefore
oxygen radical flux) uJere used and chromatographic profiles r¡ere
obtained on Sepharose ?Ê. Ê representatire sample of these is
shor¡n in Fig. 2-4. Examination of these profiles indicated that thene
r¡Jere tr.lo maJor peaks, onÊ tor¡ard V s and the other aPProaching
V¡. Ês degnadation Progressed there rJas a rapid transition of
material from the ]/s to the 1/1 rather than a ProgrÊs¡ion acroíí
the profile. For example in the fractionation of the sample r¡ith an
Nsp of Ê7% of control the maj ority of uronic acld containing material
r¡as excluded r¡hilst a profile obtained for a sample r¡ith an l'lsp of 8%
of control shor¡s a peak of uronic acid containing material
approachlng P1. ln fact, betr¡een the relatively intact hyaluronic
acid and the lor¡ motecular r.r.leight degradation product, intermediate
sized material does not appear as a discrete peak. lrJhen the area
under the curre in the first, middle and final thirds of chromato-
graphic profiles from Sepharo se 2E separation are plotted against %
Nsp (Fig. Z-5), the area in the middle third does not increase above
33Z. of the total area.
Using a high molecutar r.leight fraction of hyaluronic acid,
obtained from a prepar ative Sephacryl 5-400 column and exposing it
to the autoxidation system a similar computation r¡as applied to the
first, middle and final thirds of a series of Sepharose 1Ê

FIGURE 2-{ Ê SELECTION OF SEPHÊROSE 2B EHROIIÊTOERÊPIIY PROFILES
ÊFTER EXPOSURE OF I.IYÊLURONIC ÊEID TO THE FERROUS ION
ÊUTOXIDÊTIBN SYSTEH U5IN6 5}1ÊLL INERET,IE].ÍTS IN FERROUS ION
EONCENTRÊTION
The experimerrtal condltiont hrÊrÊ the same at outlined in
Flg. l. The concentrations of ferrous ions used ¡n the
profiler shoun urÊrÊ Z0 ¡.rH (Nsp = A7%1r 50 pH (t{sp = 73%'1,,
100 ¡rl1 (Nsp = 3g%), 200 ¡rM (Nsp = 237<-) and 1000 ¡-rH (Nrp =
Bß).

TtxExE
a
3
2
1
3
2
5
a
2
2
I
o
5
1
3
2
I
o
o
6
3
NrP
N ¡p87
Nrp 73
N rp39
Np23
Nrp 0
-oÉ,Þ-¡¡¡2oIJ
-ê|J{I2oc,I
5
1
to
Y6
5
2
020 a,
FRACI|Ol{v1

FrfìuRE 2-5 ÊREÊ UI-IDER THE EURI/E ÊS Ê FUNCTI0N 0F 9ã Nsp FOR
IIYÊLURONIE AEID EXPOSED TB TtIE FERROUS ION AITTOXIDÊTION
SYSTEH ÊND THEN FRÊCTIBNÊTED ON SEPI.IÊROSE 2E.CL
A large saries of hyaluronic acid samples uert ÊxPosÊd to
thc ferrous lon autoxldatlon system and then fractlonated
on Sepharose 28. ln each profila obtalncd, thc årËå undar
tha curve ln each thlnd uJât mËåsurtd. This flgure sho¡¡s
thosa mÊäsurÊments plottad ås à functlon of ß Nsp.
The area for the flrst thlrd of each pnofile (r""r)therefonÊ rËprÊsents hlgh molecular uelght material. The
arÊa under the curve for the mlddle third of each
profile (r......r) represents middle molecular u.reight material
and the last third of each proflle (o"""'o ) rÊPrÊsents small
molecular r.r.laight material.

roo
Agoo-
.C(Joooooooo
oñv
t¡¡
É,fIllrIt-et¡¡ôzÐ
¡¡¡É
q
80
70
óo
50
&
30
20
t0
t
...
aj
E
.o*, *o.acr,.^fWEIGHT MATERIAI
iAIDDTE MOTECUTAR
WEIGHT MATERIAT
¡o..
¡. 4
tr
I 1+:.j
A
I{IGH MOI.ECUTAR
aMAlERtAl
1.......0 30 20 t0 o100 90 80 70 óo 50 10
SPECIFIC vlScOSlTY (Ntp Z )

67
fractionationg. Êgain no more than 13% of the uronic acid appeared
in the transition portion of the profiles (Fig. Z-Ê).
5. Ênalptical ultracentrifugation
Sedimentation of hyaluronic acid and its oxygen radical induced
degradation products uas achieved on the analytical
ultracentrifuge. The reciprocals of the sedimentation coefficients (5o-¡) and the extrapolations to derived M5g rralues [ÊLELÊl'lD and l,rJÊl-lG,
l9?0; EÊRTOLD, I,rJIEEKIN and TH0NÊRD, 1981(b)l are shor¡n in Tabte Z-2,
Thnee samples of hyaluronic acid hJere used, each of a diÍferent
starting molecular r.leight. They were all exposed to the ferous ion
autoxidation system. Êgain a louer molecular r.leight limit of
approximately lBa uas obtalned. The calculated moiecular uleightralues (t1sD) folloued a predictable overall drop r¡ith decreased
rziscosity. Hor¡erer in Experiment I the sample exposed to ?E yl1
ferrous ions (Nsp = E7% of control) had an I'l5g of 60,000, u.rhereas ín
the sample exposed to 100 ¡.rl'1 ferrous ions (Nsp = 3g% of control)the l15g ralue rr.ras Ê8,000. Similarly in ExperimEnt 5, u.rhere more
samples uJere analyzed, the lor¡est H5g rralue calculated r¡as 9,000 in
the sample that had been exposed to 125 ¡tP1 ferrous ions, r¡hereas
samples exposed to 150 and 2OO ¡tYl ferrous ions shor¡ed t15g yalues
to 39'000 and 't2'000 respectively prlor to a further drop occurring.
D. Dt5CU55tOil
The commercial hyaluronic acid used in this study uJas supplied
as a lyophilysate and Sepharose 4E chromatography shous this tobe polydlsperse as regards hydrodynamic size. Attempts to obtain ahigh molecular ueight hyaluronic acid preparation by fractionaflon on
Sephacryl 5-400 chromatography r¡ith subsequent dialysis and
lyophlllzatlon of the fraction¡ reduced the amount of material

FIGURE 2-6 AREÊ UNDER TtlE CURPE ÊS R FUNCTION 0F ß Nsp F0R PURIFIED
IIYÊLUROT{IC ÊCID EXPOSED TO TTIE FERROUS ION ÊLTTOXIDÊTION
5Y5TET1 TIIEN FRHCTIONÊTED ON SEPHÊROSE {B-EL

PERCENTAGE
OF
PROFILE
70
30
ro
óoLOW MOTECUTAR
WEIGHÍ MATÊRIAL
MIDDTE MOTECUTARWEIGHT MAIERIATA
.^
HIGH MOTECUIAR
WEIGHÍ MAÍERIAT
0 roo 90 80 70 óo 50 10 30 20 loNtP
0

69
Teblc 2-2 MOLECULÊR IJEIGHT ESTIMÊT|0]{ 0F HYÊLUP0HIC fiClD E1{P05ED T0 THE
FERROUS IOÌI ÊUTOIIIDÊTION SYSÍEM USITIG THE AIIÊLYTICAL ULTRÊCEI.ÍTRIFUÊE
E*P
?r cFFotJt ton
concrntretionmicnomoler
Sprcificvi=cosil,yHrp (f,)
Rrciprocel of thrccdirnçfitrtion
corff¡c¡rnt tt z¿noconcrntration (l lïo't)
C¡lculrtrdmoleculerrlcight(F1sD)
0t02g
100
020
t00I 000
t00918?ãg
100E9+tt0
r00E6?2565216352+t5
0.0600.0150.1858.118
0.29?0.50e0.+ee0.585
0.{280.{l ?0.1950.5180.89?t.t060.Ët00.5890.90Ê
l,+50,808g+,00060,u00EE,gOE
?
3
e5e,0ooe15.00095,000,15,0Ð0
02550?5
r00125t50200100
9e,00098,00065,00058.00015,0009,000
59,000,+2,000
15,000
For rach cxprnimcnt, thr rcection concçntmtion of hyelunonic rcid r¡¡s I mg/rnl,,EDTH wrs prusent in r netio to fcrnou¡ ions of l:l and the buffcn used r¡as 50 mMphosphate buffcn. Thr srdlmentation coefficicnts breFe then determined on concen-tnetions of hyeluronic ecid of I gm/ml, 0.5 mg/ml and 0.25 mg/ml, although et theletter concentnation in somG of the moFc dcpolymenl=ed samples no pcak r,.ras
ldentlfled rnd thrnrforc no rrlur fon thr srdlmrntrtion constent could be uscd.Sincr thr hyrlunonic rcld concrntretlon¡ u¡rd r,¡rnc nrletirrly lou.r, e linr¡nnrgnrssion of thr reciprocel of thr ¡rdimrntetion coefficient could br used todr[rrminr So'l [BÊnruUD, UJIEBKIII rnd Ttl0NÊRD, lg8{].

?0
retarded on Sepharose 4E chromatography but did not eliminate itentirely. This preparation uräs performed before the publication of
the r¡ork of hIEDL0CK et al. I I gE3]. They shor¡ed that lyophyltzation
' ls accompanled by radical generation and depolymerization.
Nerzertheless a large amount of material in the commencial pnoduut
that uas rrtarded by Sephanose {E-CL r¡as able to be eliminated.
Êll subsequent prËpåråtlons uJere made r¡lthout lyophylization.
The controllcd exposune of hyaluronlc acld to oxygen nadicals ls
åccompanlad by a predicted decreå,se in riscoslty and the
appËärance of a polydlspense population of breakdor¡n pnoducts. This
phenomenon is noted r¡hether the nadicals ane genenated by the
autoxldatlon of fernous ions, the action of xanthine oxidase on
hypoxanthine or by stimulated peripheral blood polymorphonuclear
leucocytes. EREENhJÊLD and HOY I l9E0] hare reported tr.ro
Sepharose ZB-EL chromatognaphic profiles r¡hich adequately
demonstrate ä genenalized reduction of hydrodynamic size of
hyaluronic acid exposed to the XO/l'lX system and to PHÊ stimulated
neutrophils. lrJe hare endearouned to define the pnogpess of oxygen
radlcal lnduced degradation by lncreasing the numben of profiles
analyzed. 0f the oxygen radical produclng systems studied, the
ferrous ion autoxidation system could ba most neadily manipulated
to proride a comprehensire serias of dagraded hyaluronic acid
samples. Less extensive studies using the X0/l-lX system produced
congruent findlngs compatible r¡ith a similar fonm of hydroxyl nadlcal
attack on hyalunonic acld in both systems. The nelatirely modest
degradatlon achiered by Pl'1Ê stimulated polymorphonculear leuco-
cytes could be partly explained by: (a) lessen fluxes of superoxide

7L
(not quantitated) and/or (b) less uniform production of oxygen
nadicals by the cellular system.
The effects of oxygen radicals on hyaluronic acid observed Ey
, concurrently monitoring several parameters (i.e., viscostty, change in
hydrodynamic profile sn several gels and l15g) indicated that change
in riscosity is a sensltive indlcator of early damage to hyaluronic
acid. For example, a sample r¡ith an Hsp = 3E% of control (Fig. 2-3)'still contains a signfficant amount of material that is excluded on
Sepharose 4E chromatography. Ên explanation of this may be
erident from the concept expounded by BfrLAZï Ug?4l and others
that hyaluronic acid in solution forms an interlocking molecular
matrix. lf sereral large chains of hyaluronic acid in this molecular
matrix are depolymerized, the matrix r¡ould become less ordered,
despite the persistence of some intact molecules. lndeed
fractionation of hyaluronic acid on Sepharose 1B after expasure to a
mild oxygen radical flux shor¡ed some excluded material, rep?e:,enting
the intact chains together r¡ith sorre material in the included volume
rÊpresÊnting the depolymerized chains. Yet as a r¡hole the ¡olution
r¡as dramatically altered in vtscosity, ln turn theEe changes are
likely to resr-tlt in alterations in physiological properties,
ln the pnesent study the smallest degradation products obtained
had a hydrodynamic size of the order of l0a. This lor¡er limit r¡as
not shifted by subsequent exposure to a second orlygln radical flux.
Ênalysis of the large Eeries of chnomatographic profiles
obtained r¡ith the ferrous ion autoxidatlon system suggested thatthe depolymerization of hyaluronic acid h radicals may be
at least partly ondered uith rapid progression from lange to small

72
material, such ås måy occun uith a chain nraction of clearrage
erents.
The obserred increase in sedimentation coefficient of louer vis-
cosity hyaluronic acid samples on analytical ultracentrifugation may
reflect some form of molecular interaction (aggregation) or
conformatlonal change due to crossllnklng. BALÊZS g! al. [ 196?] hare
postulated that, during pulse nadiolysis of hyalunonic acid, in addition
to direct oxidatire clearrage of the glycosidic linkage betr¡een
glucunonlc acid and N-acetylglucosamlne, uronate fnee radlcals may
be formrd by ellmlnatlon of the hydnogen at C5. This nadlcal r¡ould
then be nesonancÊ stabilized by lnteraction r¡ith the canboxyl group.
Thesr radicals could interact to form cross-llnked hyalunonate
degradation products. lndeed tha identiflcation of these branched
products in synorial fluid could be used to more directly implicate
radical-induced damage to macromolecules in inflamed joints,
Eertainly such structural alterations could have important conse-
quencÊs in terms of the altered physiological properties of the
hyaluronate matrix.
E. SUllllÊRY
Preparatlre chromatographic fractions of human umbilical cord
hyaluronic acid of a molecular r.r.reight of l0ô uJerÊ subjected to
graded oxygÊn radical fluxes produced by (a) the autoxidation of
ferrous ions, (b) the action of xanthine oxldase on hypoxanthÈe and
(c) by perlpheral blood polymorphonuclear leucocytes r¡hich had been
stimulated by PllÊ. Ênalysls by gel chromatography of the products
obtained r¡ith each of the oxygÊn nadlcal generating systems
rerealed polydispersity in size. The smallest molecules detected had
a molecular r.,.relght of 104. This limiting slze r¡as not reduced further

?8
by exposure to a second oxygen radical flux. The relatir¡e
proportions of large, medium and small degradation products urere
establiEhed for rarious lerels of oxygen radical flux. Consistently a
relatlr¿ely rapid transition from large to small material uJas seËìì on
Sepharose ZB chromatognaphy suggesting ån ordered element to the
breakdoun process, Êlthough the decrease ln molecular r.leight,
after oxygen nadical exposunÊ, uJas conflrmed by anaþtical
ultracentrifugation, thls pnocedune nerealed that samples of lor¡er
rziscosity did not necÊssanily exhibit lor¡er sedlmentation ralues,
posslbly reflecting oxygen radical-induced repolymerization. Ey
altering slze and possibly the conformational characteristics of
hyaluronic acid, oxygen radlcal exposure may be expected to alter
its biological propertles.

74
cllÊPTER lllCllRllGES lH II0LECULÊR hlElEllT SPECTRß OF SYll0V,ÃL FLUID
TIYÊLURO]IIC RCID SEEII I]I II{FLÊH}IßTORY JOI]IT DISERSE

76
R. lltrRoDucil0l{.
The changes in several parameters of synovial fluid hyaluronic
acid in resPonse to inflammation have been r¡ell char acterized.
These include changes in concentration, average molecular ueight
and ¡ntrinsic t¡iscosity and are summarized in Table 1-l' page 11'
llouerer, sparse and conflicting lnformation exists concerning the
effect of inflammation upon the polydispersity of molecular r.leights
of hyaluronic acld fnom inflamed synovlal fluids. One means of
lmptlcaflng or discounting the possibillty of oxygen radical induced
hyaluronic acid depolymerization ln inflamed joints is compari=on of
the moleculan r.rleight profiler of hyaluronic acid isolated from
lnflammatory synovlal effuslons to profilei leeî after exposure of
hyaluronic acid to radlcal fluxeE 'E yJ3,. The study descrihed in
Chapter ll prorzided a large range of profiles de+nonstrating the
changes noted in the molecular r.leight of hyaluronic acid as the
size of the oxygen radical flux uas increased. llowever no Euitable
data obtained from inflamed and non-inflamed synovial fluid
hyaluronic acid rJas arailable in the literature to enable a ralid
comparison.
CREENITJÊLD [ 19Ê0] has reported in abstract that in 2l patients
r¡ith rheumatotd arthrltls none of the synovial fluid hyalunonic acid
r¡as of sufflciently small hydrodynamic size to be retarded on
Sepharose ZB-EL chromatography. ln anothen study by tsÊLÊZS'
ERILLER and DENLINEER ItSel] r¡ho analysed molecular r.r.reight
profiles of synorial fluid hyaluronic acid fnom l5 horses r¡ith
traumatic arthritls and Z0 humans r¡.¡ith osteoanthritis' a lange
amount of hyalunonic acid r¡as identified r¡ithin the included
rolume of glycer allzed controlled pore glass columns calibrated
betrr.reen Ëx l0E and 5x 104. Their re¡ults therefore indicate the
presence of material r¡hich t¡ould be included upon Sepharose 2Ê
and Sepharose {E-ÊL chromatography. llore recently EJELLE,

z6
ÊNDERSSON and GRÊNÊTl'l t l3S2l have reported gel profiles of l?
"arthritic" patients, predominantly rheumatoid arthnitis, using a
composite gel column packed r¿¡ith consecutirre portions of
Sephanose ZE, Sepharose EE and agarose 0.58, cross-linked r*¡ith
dirzinyl-sulphone. This composite gel required the tneatment of
their samples r¡ith 2.5 11 sodium hydroxid. IPH < 12) and thein
column r¡as eluted using a buffer r¡ith a pl-l I l.E. This strong
alkaline treatment leayes this technique open to cniticism because
of the possibility of alkaline hydrolysis of the hyalunonic acid.
Houerer the authors claim this r¡ould har¡e been minimal due to
the napidity of analysis (< l2 hrs)' yet no internal standard r¡as
shoun to demonstrate this. Nerertheless they shor¡ed a r¡ide
rzariety of molecular ueight distributions uith material still
present at a Rav corresponding to a molecular r.leigh t of ? x l}aon their column. ¡
None of the above studies have included a control group of
"normal" fluids. The results of these latter tuo studies
suggested that a portion of hyaluronic acid from inflamed synorrial
flulds r¡oulå elute ¡¡ithin the included r¡olume aften fractionation
on Sepharose 4E-CL. The follotling study r¡as therefore
undertaken r¡ith the follor.ling aims:-
l. To determine the molecular r.leight profiles of synorial fluid
hyaluronic acid from a series of patients r¡ith inflammatory
arthropathies.
Z. To determine similar molecular ueight profiles for a grouP
of non-inflamed synorial fluids.
3. To determine r¡hether any correlation exiEts betueen the
amount of material retarded by these gels and clinical
parametens of inflammation (erythrocyte sedimentation rate and
synoruial fluid cell count).

77
4. To determine r¡hether extracted synovial fluid hyaluronic
acid r¡as susceptible to än i¡1 ritro oxygen radical flux in the same
uräy äs commencially arrailable human umbilical hyaluronic acid used
in the prerious study.
Non-lnflamed synovial fluid samples urere taken from fresh
cadarers. ln order to be included in the studlt there r¡as to be
no history of sepsis, malignancy or anthnopathy. ln addition thene
uas to be no mäcroscopic eridÊnce of osteoarthritis or othen
arthropathy at autopsy.
E. ]IETIIfIDS
Sepharose 4E-CL, and Sephadex Ê 100 LrÊre obtaÍned from
Pharmacia, Uppsala, Su.reden. Eellogel cellulo¡e acetate membraneE
rrlere obtained from Chemetron, Hllan, ltaly. Êlcian blue r¡as
obtained from Difco, Sumey, UK. Glycosaminoglycan standards
uJere a gift from Dr. H.E. llatther¡s, Ehicago. Êll other chemicals
LrerË of the highest punlty ar¡ailable from Sigma Chemical Eompany,
5t. Louis, l"lissouri or Êjax Ehemicals, Sydneyr Êustnalia.
l. Sunorial Fluid
Details of patients are given in Table 5- 1. The erythrocyte
sedlmentation rate r¡as determlned by the lrJestergren method
and synorlal fluld uhlte cells r¡ere counted using a
haemocytometen. Êfter aseptic asplration of the knee joint in
each case the synorial fluid uas placed into a sterile tube
(r¡ithout anti-coagulant) and diluted r¡ith an equål rrolume of
normal sallne. Details of the cadaver subJects are giren in Table
5-2. Synorrial fluid from these cadarers uas obtained r¡ithin E
hours of death by closed irrigation of the joint r¡ith l0 ml of
normal saline, after r¡hich the joint rra5 opened to establish the
lack of macroscopic evidence of joint pathology. The cellular
content of the fluids thus obtained uJas removed by

7E
Table 5-l CLll''llEÊL DETÊ|LS FR0ll PÊTlEffTS ldlTll ÊRTtlROPÊTllY
Petient Sex Ag.
(yru)
Duration ofdl¡tr,tt(yrù
Synovial lluidwhltr crll count
(ctll¡tr¡ni)
% oî totaluronic ¡cidrtt*drd by9rpherorr
{B-CL
E5R
(mm/hr)
Srnopositivr nhruTnetoid enthnitisPrF65leP2F?15P5MÊ615P{ M 45 llP5 F +'l e5PÊF?g16P?M60lePEF5218Pg 1.1 6? 21Pto F {E 19PilF56l+
Srnonrgetivc nhrumatoid crthrltisPre F 68 16PIs F Ê? IgPt{ F ?{ llPlS F eS Zmo.PI6MBtl¿
P¡orietic rrthnitisPI? H 81
Pr8 ll +5
Rcltrr'¡ syndromePlg 1''1 lg Zmo.
0stroarthritisP21 F
F22 F
Ênkylosing spondylitis uith pæiphcral arthropathyPzg F ?5 25 15 2,809
t09r08t05685039322120l9t2
8t?05tt5E
NÊ*Nâ
900Nâ
+,50031,900
t00NÊ
NR
{,500Nñ
20e9l6t0ete?t2t{tst{t9
I{II
+00500+00{00600
100
e?2ersI
r5
ll
35
t7
3l
62
I+
8,290I,7EÍ)
11Ê
ilÊ5g?9
2525
23
t3l+
tJlâ . not erailablo
6

z9
Table 5-Z DETÊ|LS FR0l'1 Tl'lE EÊDÊlzER SUEJECTS
Subject Sex Ag" Êutopsy diagnosis ß Haterial retardedby Srpharose 4B-CL
?
??
M
lt"l
F
M
cl
ce
c5
c{
55
5?
58
Myocardirl inferction
Myocerdial infanction
Rupturrd aonta
Sub¡rechnoid h¡cmomhage
t0
l0
l1

EO
centrifugation at lUB0 x g îor 15 min. The supernatant uas
collected and frozen until assayed.
2. Hualuronic acid seoaration
Synoruial fluid hyaluronic acid r¡as isolated by precipitation
r¡ith cetyl trimethyl ammonium brom¡de (ETÊE) according to the
method of SEOfi [ 1968]. Ê l% solution of CTÊE in 0.3 l'1 sodium
chloride r¡as added to synovid fluid in 0.3 l'1 sodium chloride in a
ratio of 5: l. This uas then centrifuged for 30 min at ll,OOO x g
to remove àny precipitate. lrlater uas then added dropuise to
the synor¡ial fluid: CTÊE mixture until the sodium chloride
concentration approached 0.0?5 H at uhich stage a heavy
precipitate formed. This contained the hyaluronic acid-ÊTÊB
complexes. The sodlum chloride concentration r¡a¡ then decreased
to 0.04? 11 to ensure that all the hyaluronic acid present r¡a¡ in
the pnecipitate. This uas then centrifuged (5,000 x g f or l0 min)
before remoral of the supernatant (3?). The precipitate r.las
then redissolrzed in 1 11 sodium chloride at r¡hich stage the
hyaluronic acid ETÊE complexes dissociatÊ. This solution r¡as then
neprecipitated r¡.rith 4 r¡olumes of ethanol at SoC and
recentrifuged. This procedure kras repeated twice, after r¡¡hich
the final pnecipitate uas redissolved in r¡ater (there being enough
salt present in the pellet to allou this to occur). lt r¡as then
ethanol precipitated, cettrifuged and dried Jn vacuo aveî
phosphorus pentoxide. The removal of the protein from the
synorial fluids rJas monitored by opttcal scanning of the
supernatants betr¡een 190-5?0 nm on a Perkin-Elmer
spectrophotometer. lt r¡as determined that material thatabsorbed at 280 nm hJäs removed u.rith supernatant 52 and the
final pnoduct had negligible absorption at this r¡ar¡elength.

8r
5. Eellulose acetate gel electrophgEEig
ln order to demontrate that the glycosaminoglycan in the
final product rr.ras hyaluronic acid, cellulose acetate gel
electnophoresis rr.ras penformed acconding to the method of
EÊRT0LD, hJIEEKIN and Tl-l0NÊRD [1382]. Ê characteristic "fingened"
Êlcian blue stalning band on electrophoresis connesponding to the
migration of a standard sample of hyaluronic acld r.rlas nequired
before the sample r'.las further analyzed.
4. Eel chromatogxgphy-
Êliquots oî 8,2 ml of a ¡olution of purified synovial fluid
hyaluronic acid (l mg/ml in buffer) rr.rere eluted on Sepharose 1Ê-
EL columns (30 x 0.? cm) at 7 ml/hr uslng 0.5 11 ¡odium acetate,
pll 5.8, as the buffer. Fractions of [J.? ml uere collected. The
columns rr.Jere characte rized using Dextran blue, 3J5-sulphate and
glucuronic acid. Retarded rolume material uas defined as uronic
acid containing material beyond fraction l2 (voad rolume atfractlon t 0). Êreas under the curre for each profile LJere
measured on a J'ler¡lett-Packard 9E?Ê digitizer.
5. Uronic acid anal,usis
Uronic acid r¡as determined by the meta-hydroxy diphenyl
method of ELUI1ENKRÊNTZ and ÊSBOE-HÊNSEN I l9?5] oF a
modification of the automated method of ROSEMI'IÊL, EENTLEY and
ÊLEtN I le?E].
E. Qðyl|en nadical qeneration-¡-E¡The ferrous ion autoxida tion systËm kJas used as.outlín ed in
Ehapter ll. Ê stock solutlon of 2 mg/ml r¡as made from the
extracted desiccated non-arthritlc cadaren synorial fluid
hyaluronlc acld ln potasslum phosphate buffer, 50 mll, pll ?.4. The
reaction mlxture consisted of hyaluronic acid, I mg/ml, buffen,
fernous sulphate at concentrations of 0, 50 and 1000 ¡.rÌ1 and EDTÊ

82
bJås prËsent in ä molar rätio to ferrous ionE of l: l. Viscasity was
measured after 2 hr as described in Ëhapter l.
E. RESULTS
Fractionation r¡ith subsequent analysts of rau ;,ynovial fluid
resulted in the prËsencÊ of another material or materials thateluted at least partly r¡lthin the lncluded volume and r¡hich
lnterfered r¡ith the uronlc acid assay used. Using the CTÊE
precipitation method, as outlined abore, no other
glycosaminoglycans urere detected by Êlcian blue staining of the
electrophoretically separated samples (Fig. 3- l) and no
interferencË occurred r¡ith the uronic acld a:.:,ay. lt r¡as alsg
established that degradation of contnol samples of human umbilical
cord hyaluronic acid did not occup during the separation
pnocedure.
Fractionatlon on Sepharose 4E-CL of synovial fluid hyaluronic
acld from control subjects demonstrated that the uronlc actd
containing material eluted at the V6 of the colurrn. Peaks ul,lre
either symmetrical (Flg. 3-?, Ã) or Lrere dlghtly skewed towardr
the included rolume. The mean % of total uronic acid in the
retarded material r¡as 10.3 (50 = 2.374, 5EP1 = 1.1%) [Fig. 3-3].
For the patients r¡ith inflammatory arthropathies the mean 96
of uronate in the included r¿olume after fractionation on
Sepharose {E-EL uas lE.0 (50 = ?.0, 5El'1 = 1.6) [Fig. 5-5]. This
r.ras significantly different. ln most casËs this r¡as pnesent ås a
sker.l.ring of the excluded rolume material peak (Fig. 5-?, E),
although in some cases there rJas a separate peak of netanded
unonic acld-containing matenial (F¡g. 5-Zr E). The Kar¿'s of these
peaks r¡arled betr¡een 0.2? and 0.?5, cornesponding to molecular
rrreights betr¡een 9x l0s and Zx l0{. No material r¡as seen r¡ith amoleculan r+reight lor¡er than ZxlE+. This finding rJas confirme d by

\
FIGURE 5- I ELECTROPI.IORESIS OF STÊNDÊRD 6LYEB5ÊIIINO6LYEÊN5 ÊND
EXTRÊETED PÊTIENT ÊND CTNTROL SÊHPLES.
5 microliters of each sample (l mg/ml) r,.ras applied to a
t.5 cm line on cellulose acetate strips and electnophoresed
at 5B r¡olts /cm in 0.? M calcium acetate. The strips urene
stained r¡ith 0.29 Êlcian blue.
Top panel: Standards
!r. l-lyaluronic acid
b. Ehondroitin 4 sulphate
c. Chondroitin E sulphate
d. Hixture of each of the aborre
Bottom panel: Sample patients and controls
Ê. l-lyalunonic acid standard
f. c2
9. Pt+
h. Ptg
i. P5

*ve
I trËI
dcbo
-ve
+llft {-+l e) a
hIre

FIÍìURE 5-Z CI.IROHÊTTERÊPI-IIC PROFILES OF EXTRÊETED SYNOI/IÊL FLUID
I.IYÊLURONIC ÊCID ÊFTER FRÊCTIONÊTION ON SEPI.IRROSE 4B-EL.
(lÊ) Control subject,
( I B) seronegatlrre rheumatoid arthritis,( lC) seroposltlre rheumatoid arthritis,( lD) psorlatic anthritis,
( lE) Re¡ter's syndrome,
( I F) patiant rejected as a contnol r¡ho died aften a
prolonged illness that included a lymphoma and a
mediastinal abscÊss complicated by an episode of
septicaemia.

voI
100
80
60
40
20
100
BO
60
40
20
100
80
60
40
20
Fig. 1Ac2
Fig. 18
Vt
I
?eoü,obo5z9l-ÊÞzl¡¡ozooeo9zoÊÐ
0
Fis. 1C
Fig. 1EPt9
Fig. 1F
P14
0
0
P3 c5
5 10 15 20 250 5 10 15 20 25 30
FRACTION NUMBER
Fis. 1DPr8
V
ItVe
I

FIBURE 5-5 PEREENTRGES OF IlÊTERIÊL RilÊRDED EY SEPIiÊROSE 4B-CL
IN EÊDÊPER SUBJECTS ÊND INFLÊHTIHTÍIRY ÊRTTIROPÊÏTIY
PÊTIENTS.

50
PERCENTMATERIALRETARDED
BY
40
SEPHAROSE 48
30
20
ro
oo
a
oa
oaooOo
.3.ooooo
0
ooaa
CADAVERS INFLAMMATORYARTHROPATHY
PATIENTS

86
fractionation on Sephadex 6100 as all material analysed from the
inflammatory arthropathy and control groupi uas excluded from
this gel. Fractionation of several samples an Sepharose ?ts-EL
(data not shor¡n) resulted in similar profiles and these r¡ere not
altered by fractionation under di=sociative conditions using 111
guanidine HEL as the elution buffer, Smaller molecular r.leight
material r¡a¡ found in all vaüeties of inflammatory arthropathy
¡tudied r¡hich included sero-positir¡e and sero-negatirre nheumatoid
arthritis (Fig. 5-2, E & E), psoriatic arthritis (Fig. 5-2, D), Reiten's
syndrome (Fig. 5-2, E) and ankylosing spondylitis r¡ith periphenal
arthropathy, Êlso in tr*ro patients r¡.rith osteoanthritis retanded
material ¡Jas present, although these patients uere not
othenr¡ise included in the analysis.
There uJäs no significant cornelation betuJeen the percentage of
uronate retarded on Sepharose {ts-CL from the synor¡ial fluids of
patients r¡ith inflammatory anthropathies and (a) the
erythrocyte sedimentation rate, (b) the synorrial fluid r¡hite cell
count, (c) age, or (d) duration of disease.
Extracted synovial fluid hyaluronic acid from cadaver subject
C I rJas exposed to the ferrous ion autoxidation system using
ferrous ion concentrations of 50 and 1000 ul"l. The results are
ilfustrated as Fig. 3-1. There is migration of the uronic acid
containing peak across the partially included volume in a manner
identical to that seen uith human umbilical cord hyaluronic acid.
No uronic acid-containing matenial from either the cadayer
subjects or the inflammatory arthropathy patients r¡as retarded
by Sephadex G100.

FIEURE 5-{ SEPHÊROSE .îB.EL ÊND SEPI.IÊDEX Iì TOO CI-IRBNÊTTIGRÊPI.IY OF
SYNBHIÊL FLUID I.IYÊLURIINIE ÊEID EXPOSED TO T}IE FERROUS
ION ÊUTOXIDATIBN SYSTEI.1.
Extracted synorrial fluid hyaluronic acid fnom contnol
subject ES uas exposed to the femous ion autoxidation
system at fernous ion concentrations of 50 Ft1 (E Nsp =
?6) and 1000 FH (96 Nsp = 9) under conditions outlined in
the text.

SEPHAROSE 4B-CL
l0
SEPHADEX G1OO
Nsp 100
=b-¡-
zIl-É,t-zt¡¡ozooeo
IzoÉ,f
Nsp 7ó
Nsp 9
5lzvt15 20 ,ì 2s 30 s
vr FRAcnoN
0
vot5 tol
vol5 20 30

E8
D. Dl5CU55l0l{
The initial studie¡ in this investigation using untreated
synorzial fluid highlight potential pitfalls in the use of colorimetric
assays for uronic acid upon untreated samples from human tiEsue
and body fluids. The detection of a substance that is retarded
on Sepharose 4E-CL and produces an interfering colour reaction in
the uronic acid assay used makes necessary a purification
procedure to isolate the hyaluronic acid before chrom atography
and uronic acid analysis. DÊllL et ê!. I I gS5] urere able to
circumrent this problem by using a specific radioa=say (,using
radiolabeled link protein). Ênother factor dictating the nece ssity
for a separation procedure is the fact that chondroitín sulphate
has been shor¡n to be present in normal synovial fluid IS|LPÊNÊTNÊ,
DUI{STONE and 065T0I{, lgE?l and to be present in elevated
concentrations in pathologlcal fluids ISEPPÊLÊ *. d., l97?l, This
mäy be cartilage derived or a secretory product of synovial cells
lt1ÊR5l'l e.[ ê1., l3?9; SÊÊRN| É ê!., 1gÊ0l. The fact that after
extraction this glycosaminoglycan could not be demonstrated by
Êlcian blue staining after electrophoretic separation indicates
that chondroitin sulphate is unlikely to account for any of the
uronic acid containing material retarded by Sepharose íÊ-EL.
The selection criteria for donor patients (i.e. lack of infectisn or
cancer in the prÊ-morbid illness) r.r.rere imposed due to the
possibility oî colonization of the synorzial fluid and/or synovial
membrane uith bacteria or metaEtatic carcinom a cells, both of
r¡hich may be sources of exogenous hyaluronic acid [5T00LNlLLEE
and DORFI1ÊN, l9?0; KltlÊTÊ É al., lg8'fl. These criteria excluded
one autopsy subject as a non-arthrltic control. This subject (Fig.
3-?., F) demonstrated a large amount of .hyaluronic acid (a3%J in
the included rolume on Sepharose 4E-CL chromat ography. ln this

89
case no orert sync//ial inflammation uas present at autop=y and
her synorial fluid uras analyzed before the full clinical details rrere
arzailable. Those clinical details revealed lymphoma, mediastinal
abscess and an episode of septicaemia. She r¡as not used as a
control in this study due to the possibility of bactenially-deriyed
hyaluronic acid. l'lot.lerer, another explanation for the lange
amount of lor¡ molecular r,r.reight hyaluronic acid in hen cäse is the
effect of a prolonged and serere illness r¡ith its associated
metabolic derangements upon synovtal cell hyaluronate production
(see Ehapter /l). This is an area that uarrants furtherinrestigation.
Êadarers cannot be considered an ideal so,Jrce of control
synorzial flu¡d due to the possibility of post-mortem changes. ltfJas, hor¡erer, considered unethical to expose rolunteers (if they
could be found!) to the risks of arthrocentesis, hourerer small.
Ner¡ertheless, our results shor¡.1 that the molecular rrleight of
synorzial fluid hyaluronate fnom non-arthritic subjects, rr.rho died
suddenly, uras uniformly high, r¡ith mainly symmetnical peaks at thePs on Sepharose 4E-EL. No mope than l{16 of the uronic acid
containing material r¡as netarded by Sepharose +E-CL. Thus
post-mortem depolymerization does not appear to hare been a
factor. DÊl'lL et ê!. t lSË5] hare independently reached the same
conclusion.
llolecular rr.reight profiles on Sepharose íE-CL demonstrate
considenable rariation in the shape. Ê striking feature, however,
uas that the predominant amount of uronic acid-containing
material uas excluded by Sepharose 4E-CL i.e, it represented a
hydnodynamic size of I x 106 or greater. Samples run on
Sepharose ZE-CL, ruhich can char acterize molecule s rp to a higher
limiting molecular r,leight, índicated that much of the material has

90
a hydrodynamic size as high as l0?. Therefore, in inflamed synovial
fluid a significant proportion of the hyaluronic acid present is still
of a high molecular u.reight and r¡ell able to interact to form a
polymer netr¡ork at concentrations normally found r¡ithin inflamed
joints, i.e., I mg/ml [EÊLÊ25, 1971].
The amount of material included on Sepha?ose íB-ÊL nanged
bet¡¡een E9ã and 35?6 of the total and the smallest material
present had a K¿y of 0.?5 comesponding to a molecular ueight of
? x l0'+, Thls ls r¡lthln the neglon r¡here hyalunonic acld is found
to be stimulatony of macrophage phagocytosis IF0RRESTER and
EÊLÊ25, lgË01.
There rJas no correlatlon betr¡een the relatíve amount of
smaller moleculan rr.leight hyaluronic acid and clinical indices of
serzerity of joint inflammation including erythrocyte ¡edimentatisn
nate and synorial fluid cell count
Extracted cadarer synovial fluid hyaluronic acid uJas susceptible
to oxygen radical induced depolymerization to the same degree a=
human umbilical cord hyaluronic acid. Oxygen radical exposure
results in depolymerization products that fall r¡ithin the saTfie
molecular ueight range as sÊen in extracted inflamed =ynovial fluid
hyaluronic acld. Ê mild atrygen radical flux ([Fe ++l = 1L]fi applied
to extracted cadarer synovial fluid hyaluronic acid produced a
Sepharose 4E-CL profile similar ín appearance to many extracted
lnfl¿med synorial fluld hyaluronic acid's. ThiE finding supports the
hypothesls that otlygsît radlcal depolymerization can cäu5e a
decrease in arerage molecular ueight (and increased
polydispersity) of ínflamed synovial fluid hyaluronic acid E vivç).

9t
E. SU]1]1ÊRY
llyaluronic acid uas isolated from the synovial fluid sf ?E
patients r¡ith inflammatory arthropathies, Z osteoarthritíc
patlents and 4 cadarer subjects. Fractionation of hyaluronic acid
from inflammatory arthropathy and osteoarthritis patients on
Sepharose 'fE-EL revealed that uronic acid containing materiat
r¡as retarded (ave. 17.3%). Synovial fluid hyaluronic acid from
cadarer subjects r¡ho died suddenly r¡as excluded from Sepharose
{E-CL (i.e., molecular ueight >5x 106). ln most patients retarded
unonate uJas present as a sker.ling of the excluded volu'ne peak
torr.rards the included rzolume, hou.lever in some patients Emall
discrete peaks uJÊre present. No material r¡as present belou a
molecular r.leight of Z x l0a and most material present had a
molecular r+leight greater than l0õ, 1.e., r¡lthin the region thathyaluronic acid r¡ill form a polymer netuork at concentrations
found in the inflamed joint. ln patlents r¡ith inflammatony
arthropathies, there uJas no significant correlation betr¡-leen the
percent of uronate retarded on Sepharose 4E-CL and (a) the
patient's erythnocyte sedimentation rate, (b) the synorrial fluid
r¡hite cell count, (c) age, on (d) disease duration. Extracted
synorzial fluid hyaluronic acid hJas susceptible to oxygen radical
induced depolymerization'n vttro to the säme degree as human
umbilical cord hyaluronic acid.

92
C]IRPTER I1'
IIERSURE}IEIIT OF REDUCI]IG EIID SROUPS GEIIERRTED EY
EXPOSURE OF ]IYRLUPOIIIC RCID TO OXYGEII RRD¡CRLS

98
Ê. iltTRoDUCTl0ll
Enzymatlc depol¡,rmerization of hyaluronic acid ls accompanied by
the release of reducing ends irrespective of the linkage cleaved.
Testiculan hyaluronidase (EC 5.2.1.35) and hyaluronate lyase from
Streptomyces hyaluronolyticus (EE 1.Z.Z.l) are both endoenztmes
that can clearze hyaluronlc acld at the þ1-1 glycosldlc llnkage
lll0UEK and PEÊRCE, 195?; LIHKER, NEYER and t'l0FFl1ÊH' lgsÊl. ln
the lattËr cårc ân unläturated uronide lr formcd at thc non-
reducing end. Thi¡ form of clcaragc r.rill"learc l1-acetylglucosamine
at the reducing end and can thus be meaEurcd by an increase in
Horgan-Elson reactivity, 0n the other hand leech hyaluronidase
(hyaluronic acid-endo-p-glucuronidase), also an endoenzyme,
clearrs at the F | -3 glycosidic linkage, thus leavng D-glucuronic
acid at the reduclng end IYUKI and FlSl-lMÊN, l9E3J. No increase in
llorgan-Elson reactirzity can be detected follor.uhg this form of
depolymerization, houerer an increase in total reducing ends r¡ill
be detected by less reEtricted a#ays of reducing ends' such as
that of PÊRK and J0lltl50N I tg49].
It iE not clear from the llterature if reduclng ends are
detected after depol¡rmerlzation of hyaluronic acid induced by
oxygËn radlcals. tlcELEÊN and llÊLE I l9+0] dld not dctect reducing
ends after ascorbata lnduced depolymenization of hyaluronic acid
and morr racently EREENhJÊLD and l10Y I l9B0] dld not detect
reducing ends after Ë,(poture of hyaluronic acld to the XCI/HX
oxygËn radical generatlng system. llor¡erer CLELÊiID qË åL t lgEgl
urerË able to dcmonstrate that the depolymerization of
hyaluronlc acld exposed to ascorbic acid at pll 5.5 is accompanied
by the release of reduclng ends as demonstrable by reaction r¡ith
sodium borohydride. The resolution of this discrepancy is

94
of importäncË in the further clucidation of thc mechanirm of
oxygËn radical induced dcpolymerization.
The aim of this inrzestigation r¡as to determine r¡¡hether
reducing ends brerÊ detectable in samples of hyaluronic acid thathad been exposed to oxygen radicals generated either by the
autoxldatlon of femous ions or by the actlon of XE upon l'lX. The
assays for neducing sugars uJere the ferricyanide assay of Park
and Johnson and ta?-cyanide labelling of reducing ends, both of
r¡hich r¡ill detect N-acetylglucosamine and glucuroflic acid at the
reducing end and the Horgan-Elson ä,ssäpr l¡hich is specific for N-
acetylglucosamine at the reducing end in this system.
E, }IET]IODS
l. gry
a'
l'lyaluronlc acid uJäs e¡(posed to ferroug ionE in the
prËsence of 50 ml1 phosphate buffer pll ?,1 under conditions
identical to thoEe outlined ln Ehapter ll. Fcrrous ion
concentrations betr.¡een 50- 1000 ul1 uJere used.
b, x0/Ëf-lyaluronic acld uras exposed to the XEll'lK system under
conditions similar to those outlined in Chapter ll, except that in
this series of experiments a fixed concentnation of X0 of 5 mU/ml
r¡as usÊd. The magnitude of the flux ¡¡as varied by altering the
inltial concentratlon of l'lX betueen l0- 1000 ull and the reaction
¡¡as allor¿.led to go to completion (i.e. I hr).
Z. En¿y:nggg-dËgr a dation of h,r.r alur onic acid
a.
Eorlne testlcular hyaluronldase (Slgma Typ, lV-1, EC,
3.?.1.35) r¡as added to hyaluronic acid, I tg/ml, in 50 mll
phosphate buffer, pll 7.1, and incubated at 3?oE at e'nzyme

e5
concÊntratlons betl¡een l0 and 500 ug/ml for 2 lts. The reactlon
r¡as stopped by changing the pl'l to I r¡ith the addition of Na0l'l
and placing the dlgestion tube on lce.
b. Stfs,plegylefiyalurÉnldase d¡ gestion.
Hyalunonate lyase from Streptomyc,æ hyalurolytlcus (EE
1.?.?.1) r¡as obtalned from Selkagaku Kogo Co., Ltd., Tokyo, Japan.
Solutlons of I mg/ml hyaluronlc acld ln 50 ml{ phosphate buffer' PH
8.0, u.rare lncubatcd uith a concËntratlon of hyaluroflate lyaæ of
l0 TRU/ml at E0oE for tlmes betueen 5 and 120 mlnutes. The
reactlon rräi ågäln stopped by raising the pll of the solution to B
r¡ith NaOH and placing the sample on icÊ.
5. Esl-hæ¡nÊlsgcaphy-
The arerage molecular r.leights of the hyaluronic acid
samples urere determined by gel permeation chromatography,
Êliquots of 0.2 ml of a I mg/ml solution of hyalursnic acid t¡¡ere
applied to Sepharose 'IE-CL or Sephadex 6100 (Pharmacia, Uppsala'
Sr¡eden) using perspelr columns (50 x 0.? cm). The columns rrÊre
eluted at ? ml/hr uslng 0.5 11 sodlum acetate, pll 5.8, as the
elutlon buffer and fractlons of 0.? ml r¡ere collected. The colurnns
rrêFÊ callbratcd ulth Dextran bluc, [353]-gulphate and D-
glucuronic acld. Ka,r's uJÊrË mÊàsurÊd and conrented to molecular
r.letghts by refarÊncË to data glrran by Flshar I l9B0] for dextPàns.
't'@Prlor to analy¡il thc samples urÊPÉ cxhaustlvely dialyzed
agalnst uJatËr.
a. !.@ig-ê.g!.d
Uronic acids urere determined by the metahydroxydiphenyl
method of ELUIIENKRÊNTZ and ASE0E-|'|ÊNSEN I l9?5].

96
b. Park-Johnson assal¿
A modlflcatlør of the ferlcyanlde method of Park and
Jolrnson as outlined by ÊSlllrJELL IlSS?l uag used. This assay is a
ve?y sensltlre mËêiurË of rcduclng rubstances. The modlfication
applied urâs èr outlined by llOUEK and PEÊRCE [lg5?] and entails
ensuring that thc carbonata-cyanlde reagent ha¡ a pll no higher
than 9. Êt a morË alkallne ptl, hydrolysls of the hyaluronic acld
occurs laadlng to falseþ eleyated ralues of thc reducing ends
obtained [R0ElN30N and tl0PlrJO0D, lg?31.
G. l'lorgan-Elson reactlon
The Rrissig modlflcation IREl33ltì, STR0NIIIGER and LEL0lR,
19551 of the llorgan Elson reactlon uag used. Thls assay ls
spcclflc because ln order to be porltlrc, ln addltlon to a frea
reduclng end, lt requlres the presÊncÊ of an acylamldo group and
a frce hydroxyl at the 'lth carbon of the rugar molecule.
ThereforË, ln thc case of hyalursnlc acld, lt r¡ill only be posltirze
r¡hen l{-acetylglucoramtne ig at the reduclng end.
5. Ê'¿¡niCcJsÞellhg
The method usad b,ä5 crsÉrrtlalþ that outllned by SlrJÊtlH Et
e!. If 9821. Degraded samples oî hyaluronlc acld r¡ere dlalyzed
agalnst 1000 volumes of E.? l'1 l{arE0r/ÌlallC0, buffer, pll 9.1. The
uronlc acld concertratlon of the solutlon uas then determlned
and an equlmolar conccntratlsn ol KtlCn (gO y}l.ml) was
incubated uith the mlxture for 3E hrs al ?2o8. Then unlabelled
KCÌ1 r¡as added to achleve a concentratlon four tlmes that of the
uronlc acld concetrtratlon. Êfter a furthrr l2 hour¡ the reaction
uras terminated by the addltlon of acetlc acid to glve a pll of 4.5.
The resultant sample uas thçrr fractlonated ør a Sapharose 48-
CL column ( I x E0 cnr) at l0 ml/hr urlng 0,5 H sodium acetate, pH
5.6, as the elution buffen. I ml fractlons uJËre collectad and

l
1
I
The reason that there is a discrepency betr¡een thedegr-e of Par*-Jofrnson reactlvlty for a gtvenmolecular r,reight rrcm¡in¡ unclear, although it maybe related to the inherent inaccuracy in using thebro¡d pcak, obtsined aftcr gcl chromatographl, lom"¡3un molecul¡n r,rcight,

97
ästayËd for ursnlc acid crtttrrt and radloactlvlty kJaJ counted
uslng a Beckman llquid sclntlllation counter.
E. Êscendlng Paper Êhromatograohp.
tlyaluronlc acld sampler tabelted uith '4C-CH urÉrÊ hydrolysed by
incubation at l00oÊ r¡lth Ën Êqual rolume of 4Ì{ llCl for .lE hrs
under nltrogen. MonoiaccharldÊr uJerÊ lsolatad å! a slngle peak
after fractlonatlon on Elogel P2. ÊscËîdlng chronratography oî
matcrlal from the monosaccharide peak rlas pcrformed on
hJhatman lJo I chronratography päpêr. Th¿ solve¡t uged bra¡
ethyl acetate I pyr6ne / r¡ater in a ratlo ol 12z3z1, Samples of
D-glucuronlc acld, l{-acetylglucoramlne and glucuronlc acid lactone
urÊr? labelled r¡ith I{C-CN as deccribed prevlously and uerÊ run
concurrcntly. Êfter lE hrs thc paper uJat dried and cut lnto lcm
strips and the radloactlvlly counted.
C. RESULTS
The rcsults of the colorhnetric assays are ¡houm in Flgure l- l,llyaluronlc acld degraded by either tasflcular hyaluronidase or
hyaluronate lyase from Streptomyces hyaluronoþticus shor¡s an
increase ln Parl<-Jolrnswr raactirlty. Thls ls closeþ reflectrd by a
slmilar lncrease ln l'lorgan-Elson rcactirlty. Thls ls in keeping ulith
the kno¡rrn actlon of thesa Ênzlrmer r¡hlch are both endocnz!¡m-es
that claaye haxosaminldlc llnkages thus learfng N-
acetylglucosamlnr at the reduclng cnd ILAUREM, l9?0]. ln both
oxygrtr radlcal generatlng systcms, hourerer, there is a marked
dlrergancc betr¡ecn the reduclng end to uronic acld raüo using _
the tr¡o assåys. Thene is a rise ln neducing ends uith decreasing
molacular r.lelght as mêåsurcd by the Park-Johnron assayr but
thls ls not mlrrored by a rlsa ln reducfng ends as mÊåsured by the
Relssig modlfication of the llorgan-Elson âssày.

FIGURE I- I ËENERÊTION OF REDUEINË ENDS HFTER DEPOLYI'IERIZRTION OF
HYÊLURONIC AEID EY EXPOSURE TO ENZYIIÊTIE RND OXYGEN
RHDIEÊL GENERÊTINE SYSTEIIS.
Reducing ends urere detected by the Park-Johnson assay
as modified by l-lOUCK and PEÊREE I lg5?], t ) and the
modification of the llorgan-Elson assay acconding to RElSSlE,
STRoIItNEER and LEL0IR ItgS?], (r-).Each point represents the mean of triplicate
determinations of the reducing end assay used, expressed
as å ratio of the mean of tniplicate determinations of the
uronic acid ralue of the sample.
The rar¡ data are shor¡n in Table 4- l.

STREPTOMYCES HYALURONIDASE TESTTCULAR HYALURONI0.15
o.Fl!(Ú 0. l0ú
.Jo 0.05
0.15
10 10 6
FERROUS ION
€
o.r{Éotr
!É
tr¡
è0c
.r.loЀc,É
ro5 to4
AUTOXIDATION
ro7 106 105 ro4
XANTHINE OXIDASE/HYPOXANTHINE
0. 10
0.05
10 1067 ro5 ro4 to7Molecular Weight
106 ro5 ro4

99
TRELE {- I COTIPÊRISON OF REDUÊINE EI*ID TO UROTIIS ÊgID RÊTIOS Ê5
DETERTIINED BY TTIE PÊRK-JOIINSIIN ÊSSÊY ÊND HOREÊ¡I-EL5OI'.I Ê55ÊY
Perk-Johncon UÊ natioHIJ x 5D
Morgan-Elson UA ratio1'4lJ x 5D
Fenrous ion autoxidationFc(ll) ¡M
I25I5
E
I23
1
5
6
0t0
100200500I 000
5xl.5x
g6góg6g5glga,lx
0.00r 60.00r 80.00830.01510.05560.015 r
0.0036g.alzE0.01Ê60.03820.055,1E.OS12
0.00060.0r 250.0190O.E?ZEo.l2180.t ?90
0.00810.00940.0{050.05130.t 1680.1 8t 5
0.0002û.00010.001 r0.0018E,ÊE+Z0.0080
E.EAZ10.008?0.00310.00220.00?g0.0040
0.00050.00100.00+00,00550.00t 10.0106
0.00110.00r 40.000Ê0.00180.00500.0060-
5xl.5x
lx(r(
1x1x
3xl 0ól.5xt 06
1x106lxlOj1x103SxlO+
2x13x[
00
0.sat20.00280,00130.01 t 5
0.00160.00150.008s0.0t 060.0 r 190.0159
t.001?0.026a.g?g+0.r 0200. r 5290.2325
0.00?50.029?0.12100.19630.3030.356
0,00020.00050.00100.001 I
0.0003
0.00030,0005Ð.00020.0010
0.00t 89.00?20.01820.0501E.O?92E.EBl?
0.000r8.00s20,0072a.a2350.005?0.0325
g6gÉ
6693gJga
lx(xlx
ilypoxanthinr/llenthine oxidrsrHx ¡rMl0
z2E3 t001 2905 5006 t000
3xl 0ó5xl 0ölxl06'lxl05lxl0s5xl 0a
2x1063x1053xl0alxl0a9x1038x103
2x1063x1059xl 0+5xl0aflxl0+Zxl0a
Testlcular HyalunonidaseConc(u9/ml)
102 t0550+ 1005 2006 500
g6g5gaga
93g3
gÉgsgag+g+ga
5x.5x9x8x
2x3x9x5x1x¿x
Strcptococcal Hyeluronid¡¡cTime(mins)
05
t530EO
tzE

r00
The rise in Park-Jol'rnson acllvlty ln hyaluronlc acld exposcd tothe ferrous ion autoxldatlon system uJèr further tested by
measurlng the Park-Johnron to uronic acld ratlo beforc and afterextenslra dlaþsls agalnst uJätÊr. Slnce alt the componcnt¡ of thl¡autoxldation systÊm årr of lor¡ molecular ueight they uould not
ba ratained by the dlaþsls bag (molecular r.relght cut off =
10,000). l{o changc in the Park-Johnson to uronlc acld ratlo uas
rcÉrr after this manoÉurrÊ (data not shoum).
Since lt has beer suggested that oxygcn radlcal cxposurÊ
mlght caulc port-clearägÊ modlflcatlon of the reductng c'nd
[tìREE]llÂlÊLD and ilOY, l980l, thls posslbllity uas tested. Ê rample
of hyaluronlc acld r¡a¡ exposed to a llrnited dlgestion uithhyaluronate lyase and uas then retrleyed by ethanol
preclpltatlon X3 and uag thcn drled þ yacqo oyer phosphorus
pentoxlda. Thls r¡as thcn expoted to the famous autoxldation
system (FIgure 1-21. Thc llorgan-Elson to uronlc acid raflo of
enz¡rmatlcally degraded hyaluronlc acld krêt unchanged by
ÊxposurÊ to the oxygÉrt radlcal gcneratlng rystcm, Thls
damonstrateg that post-cleavag¿ modflcatlon cannot account forthe dlrergs'ncÊ betr¡cerr Parle-Johngon and llorgan-Elssn
rcactlritlcs noted fn Flgure {-1.Flgura 'l-3 shor¡s the proflle, after fractlonatlør ør SepharosË
{B-CL of hyaluronic acld samples that hare bec,n reductirrely
labelled ¡¡ith l4c-cN. Labelllng utth l'c-cN occurr urth hyaluronic
acld that has been expored to both hyaluronldase digestion as
r¡ell as both the fcmous ion autoitdaflon and the X0ltlX oxygcn
radlcal ganaratlng systems. The proflle for laC mirrors the uronic
acid profih until P¡ rr.las reached r¡here fnee |{C-CN elutes. The
rate of lncrea¡e ln tha ratio of radloactlrity to uronlc acid
beturen fractlons Z0 - ZB r¡as drtcrmlned for each proflle by

FIEURE t-Z EFFECT OF ÊN IIXYBEN RÊDICRL FLUX ON TIIE I'IORIìÊN.ELSON
REÊCTIYIW OF ÊN ENZYHÊTIEÊLLY DEERÊDED SÊ]-IPLE OF
HYÊLURONIC RCID.
Ê seria¡ of samples of hyaluronic acid urere digested
r¡ith hyaluronate lyase from Streptomycei
hyaluronolyticus for t0 min under the conditlons outlined
in the text. The hyaluronic acid u.¡as then retrieved by
precipitation x3 r¡ith ethanol, drled in ZSC¡¡C ove?
phosphorus pentoxide and made up rrlith the appropriate
components of the ferrous ion autoxidation systc'rn at
ferrous ion concentrations of 100' 200 and 1000 ul1
respectively,

E-0-
E
I
o.r{
\¡JÉ(tloútn
F{€
f4 "-{o
ci <(ü@o¡{ 'r-loÉ=ot{
0. 15
0. 10
0.05
10 106Molecular
hyal uroni daseoxy radical
104105
I'lei ght

FtGuRE {-s sEpHÊRosE +E-cL cHRoHÊToGRÊpHv 0F I{c-cvR¡uDE LÊEELLED
IIYÊLURONIC ACID DEBRÊDÊTION PRBDUCTS.
The uronic acid value is closely reflected by the lerel of
I {E-cyanide until Pt is neached r¡hen unbound llC-C¡t
elutes.

lF{É
o0
50€o
o.r{É
I 10
00
10
50
10
50
0
STREPTOMYCES HYALURONIDASE
FERROUS ION AUTOXIDATION
XANTHINE OXI DAS N /HYPOX ANTHI NE
vo vt
5000
2 500
5000
2500
0
5000
ã0)ÈH.o0)ocr
(-f
tFÚ
3uroEH
I2 500
0
10 20
Fraction Number
30 35

FIGURE I-{ ÊSCENDINIì PÊPER EIIRBIIÊTOERÊPHY OF I'1ONI]SÊCEI'IÊRIDES
ISoLÊTED ÊFTER ÊEtD HYDRoLYSTS 0F l{C-CVR¡IIDE LÊBELLED
I.IYÊLURBNIC ÊCID DEGRÊDÊTION PRODUCTS.
The top panel shot¡s the results obtained after initial
depolymerization r¡ith the X0/HX system. The bottom
panel shot¡s the results obtained after initial digestion
r¡ith hyaluronidase from streptomyces hyaluronidasa. A
l{C-.yanide labelled N-acetylglucosamine standard
mlgrated to l? cm, l4C-cy"ttide labelled D-glucuronic acid
and D-glucuronolactone standards migrated to bet¡¡een
5 and ? cms.

120
100
80
60
40
20
ASCENDING PAPER CHROMATOGRAPHY
XANTH I NE OX I DASE/HYPOXANTH I NE
STREPTOCOCCAL HYALURONI DASEDPM/CM
5000
4000
3000
2000
1 000
5 10
DI STANCE ( C}4 )
20

t04
llnear rcgrcssion anaþsls. The slopËs årË all comparable
(hyaluronldase = lg.E, Fe2+ autoxldatlon = l{.0 and X0/llX = 19.6).
Slnce the only aldehyde that thc enzr¡rne uill rclease is at the
reduclng end, thls flndlng suggerts that aldehyde or ketone
formatlon ln thc onygÉn radlcal systems does not occur
slgnlflcantþ other than from the reduclng end crcated at the
clearage slte (1.c. cach clcaragc rlte = I aldehydc Aroup created).
Furthcr efforts uJGrË undertaken to ldcntlfy the rugar tor¡htch |{C-CN tr btndtng by hydrolystng the rtC-CN labellcd
degradatlon products. llonosaccharldes uJÇrË isolated by
fractlonatlon on Elogel PZ and urÊrê separated by ascandtng päpêr
chromatography. Flgure {-.t shot¡s that for hyaluronldase
dcgraded hyaluronlc acld the label mlgratas r¡lth a similarly
labelled l{-acetylglucosamlne standard. Thls ls ln keeping r.ulth the
knoum actlon of thls Ênz!¡mÊ. l¡Jlth hyaluronic acld exposed to the
X0/tlX system r{E-CÌ{ does not mlgrate ulth the N-
acatylglucosamine standard but ls spread oy?p the areas of
mlgratlon of the lrE-CN labellad D-glucurontc actd and a stmllarly
labelled glucuronlc acld lactone standard.
D" Dt3CU35l0il
The "reduclng Érrd" of a rugar molecule ls due to the præencÊ
of a frae or potcntlally free aldehyde group [LEHNIilGER, l9B3l,
The structurc of glx carbon tugarr ls usualþ deplcted as a ring
structura, hourercr r¡lth reductng rugær a small portiør ls
present in rolutlon al än open rlng structurç uith an aldehyde
group centered on the I tt carbsr. The tuo formg are fn an
equilibrlum that rtrongþ îavours thc closed ring form.
The colorlmetrlc assay of Park and Johnson rellas on the
ablllty of the aldehyde group to act ôs â reduclng agcrrt ¡¡ith ametal ¡on (Fe3*). Cyanldc r¡ill atso rcact u¡ith thc free aldehyde

r0õ
group of a sugar in a reaction first described by FISCIIER [1890]
and knoun ås cyanohydrin synthesis. lnltially utilized as a mêanj
of adding an extra carbon to a sugar molecule, this reaction r¡as
subsequently dereloped by H0YER and ISEELL Ilg5E] as means of
reductlr¡e labelllng of sugars. lt is clear from our rÊsults r¡ith
both of these assay;, that free aldehyde groups (i.e. reducing
sugars) arË prÊsent in oxygÉrr radical dcpolymcrtzed hyaluronic
acld"
The flndlng that one docs not sec Horgan-Elson reactlvity
after dcpolymcrizatlon of hyaluronic acid by the tr¡o olrygm
radlcal grnenating systems tested indlcateg that uhrn
depolymerization occurr, thç rcducing rugar gÉ'nÊrated ig not
centered on the l"l-acetylglucosamlne. Thls i¡ contrary to the
findlngs of ELELÊiiD Èt gl. I lgEg], r,.rho reported that for hyaluronic
acld depolymerlzed by an arcorbate drlren systen, thc $l-4hexosaminidlc llnkage rrras prefererttially cleaved (thus learzing l{-
acetylglucosamlne at the reducing eîd). ln Chapter l, I hare
draum attentlon to the similaritics betuJeen the ascorbate
syste'rn and the femous ion autoxidatlon system (ascorbate
reduces Fe3* to Fez+J. l'louerer, it may bc the dffferencer in
these systems that axplaln the dlvergent flndtngs c.f. the
posslbillty of a mopÊ direct reaction bet¡¡een ascorbate or its
oxidlzed products and hyaluronic acld.
0ur flndings apÊ consistent r¡lth the hypothecis that olrygerr
radicals clearc at thc glucoraminidic linkage thus learing D-
glucuronic acid at the rcducing etd. lndccd EALAZS gt tl. [ 198?l
hare postulatcd that it ir the this glycosidlc linkage that ig
cleared during pulsc radioylsis of aquÊous solutlon¡ of hyaluronic
acld. Eoth pulse radiolysis and the tuo o,(ygen radical generating

t06
tystËmr used in tha prescnt study are bellered to càusÊ
hyaluronlc acld dcpolymrrizatlon by OHt productlon.
It is al¡o posslbla to interpret from the colorimetnlc data thatclearrage mäy occup by lysls of the carbon rlng of elther sugar
uith the creatlon of a terminal aldehydc or ketone gnoupr both of
r¡hlch mäy also react r¡ith cyanida. Hor¡eyer this ls less llkely
than F | -3 glucosaminidlc llnkage ruptura because although the
laballed monosaccharides do not clearly separate on ascanding
paper chromatography solely as labelled D-glucuronic acid,
probably due to lactone formation duning hydroþsis, it is clean
that a considerable portion ls labelled D-glucuronic acid. Should
clearage occur by lysls of a sugar ringr than no such labelled
sugår should be present"
The ldentlflcatlon of reductng sugars by radloactire labelling and
subsrquent separatlon måy p?ov¿ to be a sultably úscfmlnatlng
method to determlne r¡hether hyaluronlc acid has been exposed to
oxyg"n radlcals !n EiEg. ,, "
,.-ï-rr"l,"';, ''E. SU]1HfiRY
Êfter depol¡rmerlzatlon of hyaluronlc acid Ey expoture to the
ferrous ion autoxldation syrtem and to X0/l'lX therc ic an
increase in reducfng cnd actlrlty ar mÊåiured by the Park-
Johnson assåy and by reductirue labelling ulth radiolabclled
cyanide. Therc is houJerer no increa¡e in llorgan-El¡on reactivityt
nor lg there any change ln l"lorgan-Elson reactirzity n hyaluronic
acid pre-exposed to hyaluronidase and subsequently exposed an
oxygËn radical generating system. These flndings uJÊre consistent
uith our hypothesis that oxygen radlcal lnduced depolymerization
of hyaluronic acid occurs by preferentlal clearage of the Þ 1-5
glucuronidlc glycosidlc llnkage thus learzlng D-glucuronic acid at thc
reducing end.

r0?
C]IRPTEN Y
DOES OXYEE]I RRDICRL EXPOSURE RLTER T]IE REILITY OF
llYRLUR0lllC RCID T0 ]{0DULÊTE }î0ll0cYTE Fc RECEPT0R
EHDHE?

t08
R. I]ITRODUCTIOil
The indicatlon that ÊxporurÊ to oxygcn derlved free radlcalg
mlght lead to clcaragc. of speciflc Aþcostdtc llnkager and posstbly
other altaratlont along the lcrrgth of the hyaluronic acid molccule
leads to the questlon:- Do the chemical requclae frsrn exposurt tooxygrrt frec radlcals lead to an alteratlon ln biologlcal or chemical
propertles apart from those rerultlng frorn decreased molecular
trlalght?
The obserratlon of F0RREÍTER and EÊLÊZ,ï l.lg80l thathyaluronlc acld r¡ould lnhlb¡t the phagocytosis of latex beads
demonstrated a speclflc cellular effect that ls of conslderablc
potcntlal blologlcal releyance, partlcularþ ln relaflon to
phagocytosls occurrlng ulthln an lnflamcd Jolnt. Thcy obserred
that thc phagocytorlr of latex bcads by eltcttcd rat perrtoneal
macrophågËs r¡as lnhlbltcd by hlgh molccular r.lclght (1.c. llhl = 1,8
x l0r - ?.f, x l0É), hyaluronlc acld at concrntraflons betueen 0, 1-
0.2 mg/ml. Paradoxtcalþ, hor¡erer hyaluronlc acid of molecular
r.rreight 9.0 x 101 had a gtimutatory effect at concerrtration¡
betr¡een 0.2 -0.5 mg/ml" Hyaluronic acld of thig ¡lze can be
expectcd folloulng depolymerlzatton r¡tth oxygrrt radtcals (see
Chapter ll) and ls found ln thc synovtal fluld of a considerablc
proportlon of patlerrts r¡¡lth infla¡matory arthropathies (see
Chaptcr ttl),
lrJe thercfore sought to verlîy the flndtngs of Fomcstar and
Ealazs by utlllzlng the Fc receptor blndlng assay. There urerÊ
several rÊasons r.rlhy thls assày uas used. Ftrst;ly the Fc receptor
medlates both blndlng and lnterrrallzatlon of l96 coated partlcles
and lgfì aggregates. lt ls a r¡cll characterlzcd membrane
rcceptor [ERlFFlll, ElAl{c0 and 9ILI/ERSTEII{, l9?5J. Fhalþ, the
paramatêr mÊåsurcd, l.e. Fc receptor binding is more dlrectþ

r0g
linked to the phagocyto¡ls of immune conrplexeg and ls thuE ls
morË clearþ llnked to the pathogcneslg of lnflanrmatory Jointdlseasc than the phagocytosls of latex baads.
E. NET}IODs
l" Adherent monocptas
l0 ml samplcs of yenous blood, antlcoagulated ulth llthlum
heparhate, urcre obtalned from healthy Red Cross blood donorg.
These bJGpÉ thcryr dlluted ¡¡lth Ën Êqual rolumc of l'lank's balanced
salt solutlon (833), placed ln a 50 ml slllcontzcd glass centrtfuge
tube and underlayed r¡lth ?.5 ml Flcoll Paque (Pharmacta, South
Seas, Sydncy). The tubes ur?rê thcn centrffuged at 1009 for 35
min at poom tanperaturc. The mononuclcar cell fraction uas
then asplratad fronr the Flcoll Paqua-Hank's 833 tnterfacc. Th.y
brÊrÊ then rrrashed three tlmes ln llank's 853. The flnal call ptlleturär r?suspendad ¡n RPHI-|E{0 medlum supplementad r¡tth l09E
fetal calf serum (FCS) and penlclllln/streptomyctn. They kr?rÊ thcn
transfemed to Z{-r¡ell tlssue culture plates ( l E mm dlam) Costar
(Cambrldge, llass, U.S.Ê). Êt the bottom of each r¡ell had baen
placed a small clrcular glars coversllp that had prertously beeryr
r¡ashcd ln l0% rulphurlc acld folloucd by 7O% ethanol.
âpproxlmataþ { xlOË cells ucrc added to each uell. The celb uJËrÊ
then lncubatad for I hour at S?oC tn a 5% C0, incubator, l{on-
adherent cells uJÊre then r¡ashed off r¡ith l{ank's BSS.
2. Fc receptor assap-
a. Preparatlon of anttbod blood cells
Ê mlxturÊ r¡ra! made conslgtfng of 50 ¡rl packed r¡a¡hed
red blood cells, 200 pl antl-D tËnum (Commonrr¡ealth Serum
Laboratories, Sydnay, Australia) and 2 ml Hank'¡ 835. This mixture
r.r.ras placed in a S?oC tncubator for I hour. Durlng the incubation,
the tube r¡as invertad ¿vety 20 minutes. The red cells r¡ere then

rt0brËthÊd three tlmeg in gterlle normal sallnc and flnalþ resurpended
ln 25 ml RPÌ1|.
b. Fc recaFtor_ansa}¿
500 ul of thc anttbody coated erythrocytes Lrere added
to aach r¡ashed mwtocyte monolayer. Thls uas then incubatc d forI hour after uhlch each uell r¡as r¡ashed three ttme¡ r¡ith llank's
ESS to rÊmova non-bound erythrocytes. Thc cells LJæÊ flxed by
tha addltion of 39ã gluteraldchyde in llank'¡ 833 for 30 min atroom temparature (or 12 hrs at 4oE).
c.@The corcrsllps r¡JcFË stainçd ulth Elemsa staln. Êfter Etl
seconds the corerrllps uJÊrÊ uaghed ln uatcr and allor¡ed tostand ln dlstlllcd llz0 for 50 mln, the uatcr being changed êvery
It mlnutes. Th.y LrÊrÇ drlcd and mountcd. The mlcrorcoplc
appeåpånce of these prËpËrètlons shouJcd rsadlly ldcnflflable
adherent monocytrs to r¡hlch urcrÊ attached rad blood celb,
eithar touchfng or partlalþ or completaþ lngested. Sinca the erd
point in thls inrzestigation r¡as rcceptor attaclrmrnt, no attemptuas made to dlstlngulsh attaclrmrnt fronr conrplete phagocytosls.
For each corersllp flra hlgh pouJÇr flelds brcre counted or cnough
to guarantce at least 100 mørocyles and thc comespondlng
number of erythrocytes. Thlr uas thrn ÊxprÊæed as a bindlng
ratlo. The results for cach Àstay uJÊrê exprÊJsed as the Blndlng
lndax.
EFtdlng lndex = ratlo REC per mmoc¡,¡te uith ïAratlo REC per monocyte r¡ithout I'lA
3,
a.
Purlflad hlgh molecular ludght hyalurontc acid (Healon: J"lhJ =
>2x l0t) r¡as tha gift of Pharmacia, south Seas, sydney, Êustralia,

rtlThis r¡as exposed to the fcrrous ion autoxrdation orlygen radlcal
gancratfng system under condltlon¡ ldentical to thosc outllned fn
Chapten ll. Êrerage molecular rletghts of the depol¡rmerized
samples urËrÊ determlned by fractionaflng allquots upon
Sepharose {E-CL columns ( l0 x 0,5 cm) and the K6y of the peak
uronlc acld raluÊ rrtâl mcasured. The correrponding molecular
uleight r¡as determlned by referÊncÊ to data for poþsaccharldrs
glrcrr by FISHER [19?0]" The ferrous lon concentraüon and the
corrËsPonding åvÇrâgc molecular weight of the product are shourn
ln Table 5- l.
b. Enzl¡me exoosed hpaluronlc acld
Ê preparation of llcalon uJa! ênposed to testicular
hyaluronidase (Sqma Typ, lV-ï, EE 3.2.1.35), 0.1 mg/ml for S mins
at EO.C and the reactlen u¡as stopped by decreastng the pll to8.0 and dropplng the temperature to .loC. Thls r¡as then
fractlonated on ü Sephacryl 51000 column. Poolad fractions uJêre
dlaþzrd extenslveþ agalnst uater, ethanol prectpttated and then
drlad h g¡Sl¡C. The average molecular rrrclghts of pooled fraction
uscd uas detcrmlned by fractlonatlon on à Sepharosa {E-EL
column as dascrlbed preriousþ. Charactarlsflcs of these
prËparètlsns ara glran ln Table 5- l. ln thls âssày onþ ont
hyaluronlc acld concentratlon of 0.5 mg/ml uas used.
C, RESULTS
Using undegraded high molecular rrralght hyaluronic acid there
l¡rËs å dose dependent lnhib¡tlon of Fc receptor blndlng orer the
concerrtration rângÊ 0.2 - I mg/ml (Fig. 5-l).Êt a concentratlon of 0.3 mg/ml there urar a biphasic effect
upon Fc neceptor blndfng dependent upon molccular rrleight.
lnhlbltlsn of Fc receptor blndlng h,ar iËÉrt r¡ith prcparatlons r¡lth
an äreràge molecular ¡¡elght of I x l0ó and abo v¿ and stimulaflon

rl2
Table 5- | P1BLECULÊR hJEIGHTS 0F THE I'|YÊLURON|E ÊClD PREPÊRÊT|0l-15 USED
Itl TllE Fc RECEPTOR EIND|NB Ê55ÊYs
0xygen nedical depolymerized JIA
Fcnr^ou¡ ion Êrcragc l"lLJ
concentration(micromol¡r) (xl O6)
Enzymc degradcd HÊ
Poohd Êrcragc MLJ
frectiong(xt O'6)
050
r00500
r 000
505
2
t.t.0.
?1.51.00.50.2
HEÊLOf'|
11- 2E5r- 80101-il0

Frf¡uRE 5- | EFEEff OF TTIE CONCENTRÊTION OF H FIIÍìII ¡TOLEEULÊR
l¡JEIEFfT |'|YÊLUR0NIC ÊElD PREPÊRÊT|0N (HEÊLON -PIIRRÌ1ÊC|Ê) UP0N Fc RECEPTOR BINDINE.
Each data polnt = mÊân (t 3.D.) of trlplicate
datcrmlnatlons.
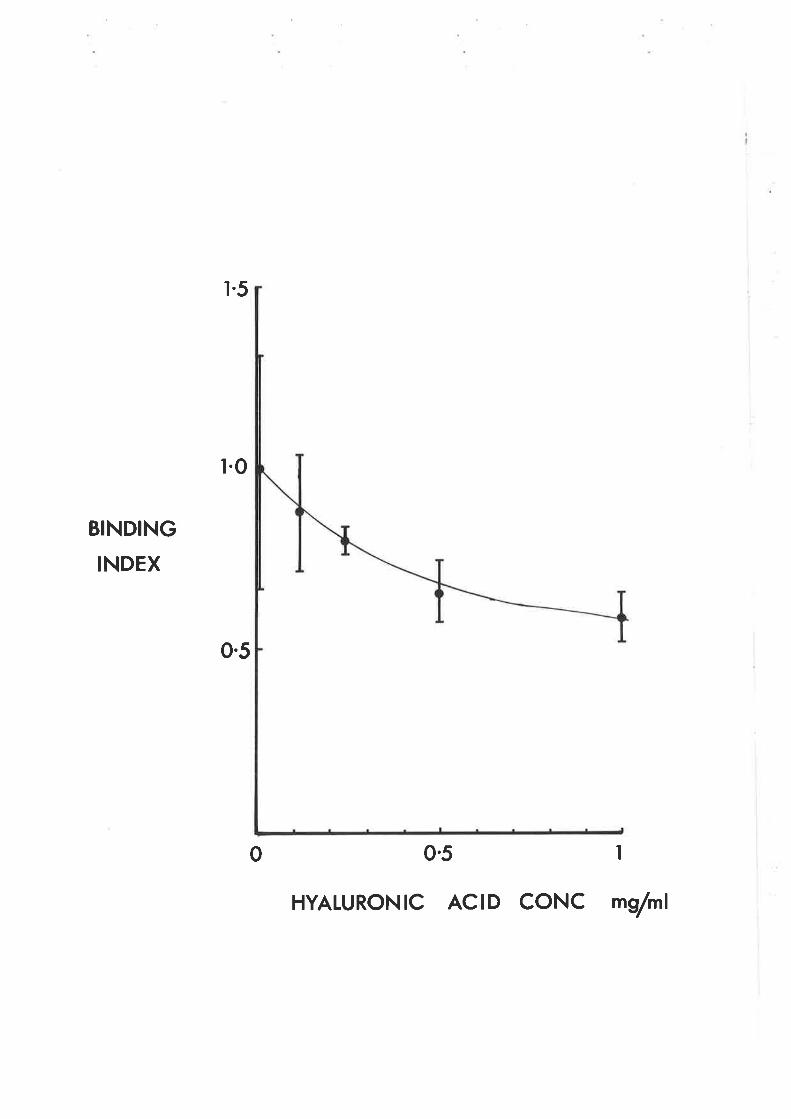
BINDING
INDEX
r.5
r.0
0.5
0 0.5
HYALURONIC ACID CONC ns/nl
I

rl4of Fc recaptor blndlng uJär tÊÉrì ln preparaflons ulth Ën ärÊragÊ
molecular uletght of 5x 105 and belor¡ (Flg. S-z). Thls effect ts rËen
t¡lth both o¡rygÉTt radlcal exposed and enzyme dtgestcd hyaluronic
acld"
D" DtSCUSSt0il
The Fc receptor blndlng àssày rystcm used ln this studydfrTered fnom the latex phagocytosts âsray studted by F0RREIIER
and EÊLÊZS [1980] becausa a specrftc receptor for the uptake olpoþstyrene latex rphercs has not bcen demonstrated. Latex ls
consldered to be crrgulfad ln a non-spËclflc manner porstbþ rclatedto lts hydrophoblclty [FR0ilnfìEfIT, I g?11, The rcsults of thts rtudylndlcate that recrptor medrated brndrng ts tnhtbtted by htgh
molecular rrlelght hyaluronrc acrd (>lxl0ê), t{ouercr at a lou
concertratlon (1.e., 0.3nrg/ml) of lor¡¡ molacular ualght hyaluronlc
acld, a paradoxlcal silmulaflon of Fc rcceptor bindlng lr erident.Thls l¡ ln accord ulth the flndlngs of Fomcster and Ealaz¡ forlatex phagocytosls, houevcr the molecular r.leight at r¡hich thisstimulatlon flrst becomes errdent rs sqnreu.rhat higher in our
system (i.e. 5x105 companed ulth gxl0{).
Depol¡rnrarlzatlon of hyaluronlc acld by both enz4aflc dtgerflon
and oxygan radlcal exposupÊ resulted in hyalurørlc acld
dagradatlon products that urcrÊ lnhlbltory ãt htgh hyaluronic acld
molecular rrrelght yet caused paradoxlcal gtimulafiør at louered
molecular rlelghts. Oxygen radlcal depol¡,rmerlzed hyaluronlc acld
caused a greatar dagree of sttmulaflon of blndlng lndcx but thlg
dld not raach statlsflcal slgnlflcancÊ.
The hyaluronlc acld concerrtraflon of 0.r mglml uas choser-r
after consldaraflon of Fomester and Earaz's data to glre maxlmal
¡tlmulatlon at the decraased molccular tr,leight and it ls partinent

Ftt¡uRE 5-Z EFFEET 0F ENZYHRïC (ao....t) ÊND 0XYEEN RRDIEIIL
DEP0LYI.'IERIZED (- ) |'|YÊLUR0NIE ÊClD UPON Fc RECEPT0R
BINDINE ÊT H CONCENTRRTION OF 0.5 IIE/T11.
Each data point = mean t S.D. of triplicate detenminations.

BINDING
INDEX
r.5
r.o
t.0
MOLECULAR WEIGHT
r.5 2-Oo.2 0.5
x loó

rr6to nota that at thig concentratlon a matrlx of hyaluronlc acid is
unllkaþ to form aspeclalþ at louertd molacular r¡eights.
Clcarly mor? extenslyc studles äpê requlred to probe the
mechanlsms lnvolved ln both the lnhlbltory and the stimulatory
lffccts dcmonstratad. llouevar lt is pantlnent to conslder sareral
rcctnt studles, ln conslderlng the lnhlbltory effect, Forrcster and
Ealazs concluded that alectrostaflc repulslon (latex, lgG, and
hyaluronlc acld are electronegaüre) ls unllkeþ to haye an effect
since sulphated gþcosamlnogþcans r¡hlch arÊ more electronegatire
are t¡lthout effect. Stmilarþ an hyaluronic acid tnduced membrane
pertubation ls unllkaþ since other membrane modtfying agents
r.,crÇ r¡lthout cffect. Th.y favorrd a starlc excluslon of latex
from the area of the cell surface thus decreaslng the proxinnity of
call and latcx. Êlternataly they speculated that latax beads and
lndard lgtì and complement coatad parUdes are hydrophoblc thus
maklng lt easlcr for the cell to effect phagocytosts. tlyaluronic
acld, betng hydrophillc r¡¡ould dacreasa this phagocytic adrantage.
Eoth of thcse explanatlons could appþ to our àssày. Stmilarþ,
starlc excluslon effects have been lmpltcatad ln the lnhlbitory
affect of hyaluronic acld on mttogen induced lyrnphocyte
stimulation.

rr?
E. SUtltlÊRY'
l'ligh molecular r.reight hyaluronic acid (l'lealon-Pharmacia) caused
a dose dependent inhibitlon of macrophage Fc receptor binding
betr¡een the concentrations of E,Z - I mg/ml. Êt a concentration
of 0.5 mg/ml both oìlyg?n radical depolymerized and enzymatically
degraded hyaluronic acid cäule an inhibition of Fc receptor binding
at molecular r¡eights of lxl06 and 1.5 x 106. Jlowever r¡hen the
molecular rrleight r¡a¡ reduced to 2 x 10s and 5 x I g= by axygen
radical degradation, ¡timulation of Fc receptor binding occurred.
Stimulation uas al¡o ¡ean r¡ith enzymatically degraded hyaluronic
acid r¡ith a molccular r.rleight of 2.5 x l[Js.

rt8
C}IRPTER YI
EXPOSURE OF CULTURED SYÎIOYIRL CELIS TO OXTGE]I RßDICRL
FLUXES. CYTOTOXICITY .R]ID EFFECT OII IIYRLURO]IIC RCID
PR0DUST|oil

119
R. iltrRoDUCTt0H
Tha studtes descrlbcd so far hare explored tha posslblllty that
oxygÊn radlcals, generatcd ln the lnflamed Jolnt, can act to
depolyrnerlze fully s¡.mtheslzed hyaluronlc acld. Houerer' the
flndlng of hyaluronlc acld of decreased arerage molecular u.leight ln
inflamed s¡morlal flulds may also be cxplalned by the synthasls of a
lcss pol¡merlzed product. The rynorlal cell rÊprÇlctlts a potcntlal
target for damagc by oxidants releascd by leucocytss as part of
the lnflarnrnratory procÊts. lt is porsible that a synovlocyte
damaged by oxldants may contlnue to producË a smaller molacular
rlelght hyaluronlc acld product.
The aim of thls expertmcnt uas to dctermlne the extent of
thc susceptlbltlty of synovlal cells to an cnzymatlcalþ gcnerated
oxygÉrt radlcal flux þ rltro. and to detarmlne the molecular rleight
charactælstlcs of the hyaluronlc acld produccd by synovlocytes
glrcrr a sub-lcthal expoturË to an oxygërl radical flux.
E" HETHoDS
l.@Eoylne s¡rnorzlal cellg rrJcre obtained by trypsln larage of borlne
metacarpophalangaal Jolnts accordlng to a modlflcatlon of the
method of FRASER and tlcCALL [1965l, Fre¡h bavtng^ hocks uJÊrÊ Ë
gift fronr the South Êustrallan lleat Corporation. trllthln { hours
of slaughter they urËra u¡ashed and the skln uas dlssectcd frea
from the mctacarpophalangeal joint. Ên area of ¡ubcutaneou¡
tissue brts rurabbed ulth 7E% alcohol and an I E gaugc needle
lnserted. ThË metcarpophalangeal Jolnt uas laraged u.lith tuo 50
ml rolumes of normal sallne to remoyÊ l',norlal fluld and slnorlal
fluld cells. {0 ml of 0.25 trypsln (tyPe lll from boytne Pancrtâs'
Slgma Chemlcal to., St. Louls, Hlssourl) in Êa++ and llg++ free
phosphate buffcred saline (Dulbecco's formula) uas then inJected.

r?'0
Êpproxtmateþ 25 ml uas therr uithdraunr lnto the syrtnge and the
remaindar uJas left h sltu for 15 mlnutes. The Jolnt rr.ras gcrrtly
malragÊd initially to ensure that the trypsln solution ent+ed
rêcassËs in the synorial lpacÊ. The jolnt r¡as then rer¡ashed r.¡¡ith
the conrplete rolume of the trypsln solutlon u¡hlch r¡as r¡lthdrarr¡n
and cantrlfugad at 1000 x g.
The cell pellet obtalned r¡as then rËruspênded in lo tlssue
culture medlum (see Êppendlx ll) supplcmented r¡ith 20fi foetal
calf serum and seedad lnto a plastlc 25 cm2 tlssue culture flask
(tìrand lsland Elologlcal Ëompanlr 6rand lsland, ller¡ York). The cells
urerÊ thcn grourn at 5?oC ¡n 5% carbon dloxlde. Êftar Z days
scatterçd adherent calls could be seÊn u.¡lth charactarlstic sharp
cytoplasmlc processes" The cells brËre then r¡ashed tr¡ice ulth
Dulbecco's phosphata buffered sallne (PEg) and the I o tlssue
culture madlum replaced. Thc medlum uag changed ln thls mannËr
tr¡lce ucekþ. Tha prhnary culture urär maintained until the cells
had rcached confluÉncÊ (usually about l0- l{ days). Th.y urerÊ
then subcultured by uashlng tr¡lce in Dulbecco's PES and then
incubatlng ät 5?oC for 5 mln in 2-5 ml of O.25% trypsfn ¡n PES.
lrJhen lt u¡¡as obsenved that the cells had separatcd, l0 ml oî 2o
tlgsua culture medlum (see Appendlx ll), supplementad r¡¡th l0%
fetal calf sarum uJâr added. Thls ¡¡as than dlvided into tr¡o on
thrce allquots r¡hlch urêrÊ then added to fresh tlssue culture
flasks. Êll subsequcnt cultures urerÊ grourn ln Zo tlssue culture
medlum unlesg otherr¡lre rpecifled. Cells LJerË harrestad for the
chromium release arrËy betu¡cerr the Zttd and ?th subcultures. ln
thls r.r.lay a relatlveþ hornogÉ'nout populatlon of synoviacytss r¡¡ith
a flbroblast-llke åppËaräncË urÇre obtalncd IFRÊSER Êt å1., l9?31,

t2l
Z.
S¡morlal cells r¡crr haryestad for tht chromlum relcase
assåy by subculturlng ln the mânnÊr dascrlbed abovc. l0l calls (ln
1.5 ml tlssue culture medlum) þrËrÊ then added to aach urell of a
2{ r¡cll tlsrue culture plate (Eostar, lE nrm dlametcr u,relb,
Cambrldge, Hass.). Thegc urêrÊ thcn lncubated at 3?oC rî 5% Egz
for tuo days r¡hsn a confluerrt layer of cells could be seÉî at the
bottom of each u¡ell. Each ¡¡cll uas then u,¡ashed tu¡lce r¡lth
Dulbecco's PBS. Carc uas takcn durlng ulashlng to pofnt the tip of
the pasteur plpette at a rlngle polnt on the circumferencÊ of the
uall ln order to dlsturb the cell laycr as little as posrlble. 0ne-
tenth mllllllter of trBr (speclflc actltzlty = 50 ¡.rCllml, Amersham,
Êustralla) r¡as added to each r¡cll and lncubated for I hour at5?oC. Thc cell laycrs bJËrË thcî uashrd three tlmes ln Dulbecco'¡
PBS and the reagcnts for cach cxpcrlmcnt uerc addcd ln a total
rolume of I ml. Expcrlrn+nts uJerc performcd ln trlpllcata u¡lth
+ve controls (20?ã acctlc acld) and -rc controls (buffer) ln each
platc.
Êftar lncubatlon for the approprlatc tlme, the cell layerr
urÊrÊ lnspected to datcrmlne that they uJcrr stlll adherÉnt. Then
0.5 ml of thc supÊrîËtant (3) uas arplrated and placed in a poþ-
carbonate tube for countfng. The remalnder (R) uag then
asplrated and the cell laysrs uJcrÊ remored by frypslnlzatlon x2
r¡ith 0.5% trypsln ln PEg and a flnal r¡ash ulth uater tohypotonically lysc any rasldual cells. These urashtngs rdcrÊ then
pooled, the volumes equallzcd and the tuber uJerÊ counted on a
Roche Eammamatlc gamma countar.
Perce¡tagÇ llCr rclearÊ urâr obtalned by the formula
ß 3t Cr releasc = ?3 X 100
R+3

LZ2
Baclrgroun d %3lÊr releate k as determlned fronr the -ve control
and r¡as subtracted from the gros s % 3lCr release. The mean
and standard derzlatlon of trlpllcate determlnatlsnr uJÊre then
calculated.
3. gry
The oxygcn radlcal geteratlng systsm used t¡¡as the XOlllX
system as prerlously outllned (sae Ehapter ll). Protcoþtlc
contamlnatlon of the xanthlne oxldase prÊprratlons uJËrÊ
decrcased accordlng to the method of IìREENhJALD and NOY I 19?91
by Scphadcx [ì100 chrornatography, Bccauge all thc proteare
actlrlty uas not conrpletely rsmovcd by thls method and lndced
afforts to complrtely rernorÇ all protea¡e actlvlty wlll rcsult ln
loss of xanthlne oxldare actlrlty IBHTES, L0trfT?lER and llÊtJDLEY,
l9B'tI, the axpcrlmcrrts urrrÊ camled out ln the presÉncê of l%foetal calf sarum ln order to ¡uamp rerldual protc ase actlvlt,y
(ree rasults sactlon).
{"Ë
llyaluronic acld productlon uräs assesged urlng synovial cells
grouJn to confluÊïrcË ln 25cmz tlssue culture ftagks. After uashing
r¡lth Dulbecco's P85, 5 ml¡ llam's F l0 tlssue culture medlum
contalnlng 50 ¡rfi 3H-gluco¡amlne hydrochlorlde wac added. Thls
r¡as then lncubatad for 'lE hours before the supernatant uas
collccted aftcr gentla urashlng of thr cell laycr t¡lth th;rupernËtant.
Tha collacted rupËrîatant uas therr cmrcerrtrated ln a
Mllllpore 25 mm stlrred cell orer a Pclllcon F[-EC flltcr.
Hpproxlmateþ 2.5 ml of concentratad superîatant ua¡ thcn
appllad to a Sapharose'iE-CL colunrn (100 x I cm) and eluted r¡lth
'l 11 guanldlnlum chlorlde. Slnce the peak of radloactlvity identifled
as hyaluronic acld dld not changc slgnlflcantþ under assoclative or

rz3
dissoclatirc condltlons rubsequent clutlons u¡ere performed urlng
0.5 H NaÊc pll 5.8. Fractlons of I ml uere collected. 5 ,¡l of cach
fraction urÊpÊ thc.n added to 2 ml scintillation fluid and counted in
a Eeckrnann llquld scfntlllatlon countËr.
5. Cellulosc acctate gel electrophoreír
The hyalurwrlc acld containing pcaks after fractløration
brÊrÊ ldentlflcd by applylng 5 ¡rl of the peak fractlon to cellulose
acetate strips. Elactrophoresls uJät conducted a¡ descrlbed tn
Chapter lll. Êfter electrophorerls, standard hyalursnlc acid
samples bJere read by Êlclan blue staining t¡hereas radlolabelled
samples uJerÊ rcad by cuttlng the cellulosc acctate strlp nto ?
mm sectlons fronr the potnt of orlgln. Each strlp r¡as addcd to acountlng ylal ulth 2 ml of llquld sclntlllatlon fluld and counted.
C."RESULTs
l. f,I/HX induccd c].rtotoxlclt¡¿
The lnltlal part of thls experlment ua¡ to detcrmlne the
leyels of oxygcn radlcal flux that produced a cytotoxlc effect
uslng s¡morlal calls as targets. Flgure 6- I shor¡s thr resultg of a
tlma coupre experlmcnt ln ¡¡hlch I cmz uells of confluent
monolayar¡ of syrnavlocylss uJÊrÊ expored to l0 mU/ml xanthine
oxldase ln the prËsrncÇ of 2 mll hypoxanthlne. Êfter E hrs a
cytotoxic efract bJät mênfestcd by the releagc of stCr into the
medlum. Subsequmtly all measurements in the stf;r release assay
uJerÊ made at 12 hrs.
Eecause xanthlne oxldase prËparatlørs contaln tomÊ
proteoþtic actirltyr ln splte of chromatographlc puriflcation,
foetal calf serum uJas added to the mlxture to suamp this
actlrlty. Flgure E-Z shor¡s the results of an expertment ustng
progrËrrlvely lncreaslng ümounts of foetal calf serum up to 50ß.
Êbore l0% foetal calf sÊrum, the F s'Erreleasa is brought back

FIÍ¡URE 6-I KINETICS OF SICr RELEÊSE FROI1 OXYBEN RÊDIEÊL EXPOSED ÊND
UNEXPOSED SYNOI/ I OCYTES.
1
- ): X0 = l0 mU/ml, llx = Z ml.l
( .."".... )¡ llx (2 mll) alone"
(Data points rÊprÊsÊnt mean I 5.0.)
FIÍiURE 6-2 EFFEET OF FOETÊL CÊLF SERUI'I ETNEENTRÊTION ON XO/IIX
INDUEED SICT RELEHSE.
Hatchad bars: X0 = 5 mU/ml, HX = Z mH
Elank bars: flX onþ

r00
PERCENT
80
CHROMIUM
ó0
RELEASE40
TIME COURSE FOR CHROMIUM RETEASE AFTER EXPOSURE
TO AN OXYRADICAL FIUX
20
.¡.
o
r00
PERCENT
80
CHROMIUM
óo
RELEASE40
20
2 4 8
T¡ME
l0
HOURS
E FFECT OF FOETAT CAIF SERUM CONCENTRATION
oN xo/nx INDUcED cHRoMluM RELEASE
hotched bor: xo hx
blonk: control
6
Nofcs I%fcs 2%1cs 5%Ícs lO%fcs 50%fcs

rz6
to control lerels, using an xanthine oxldarË concËntration of
SrnU/ml. Subsequently l% foetd calf sÊrum r¡as added to the
reactlon mlxture r¡hen the cells urcrc exposed to the X0/HX
systenr.
Figure E-3 shor¡s thc affact of superoxlde dlsmutasË on XO/HX
lnduccd slCr releasc. Êt a superoxlde dlsmutasa concentratlon of
?5 U/ml, no protcctlrc effcct uJar rËGn ove? a xanthlne oxldage
concentratlon rangË oJ 5 to 100 mU/ml. Thcse l¿v¿ls of ruperoxtda
dlsmutasc hare becn rhoum to be protectlv¿ ln the XO/tlX induced
hyaluronlc acld degradatlon rystrm and X0/acetaldchydc lnduced
erythrocyte lysls (KELL0CB and FRlD0ylCll, lg11'), Catalasc, on the
other hand uras protcctlra at a concartratlon of 50 U/ml (Figure
B-'l)" Ilannltol had a mlnor protectlve eflect at lor¡ xanthlne
oxldase concrntratlons (Flgure 6-5).
Eased on the lndlcation from the aboye expertments thathydrogen paroxide r¡¡as the agent necËrsapy for demonstration of
a cytotoxlc effcct, the effect of glucose/glucose oxidasÊ r¡Jas
examlned. This É'nzlrmt producas the dlralent reduction of oxygen
to I'1r0, ulthout productlon of the suparoxlde anlon [N|L350N, PICK
and ERÊY, l3E9l. Ê cytotoxlc affact r¡as raadlþ damonstrated ata glucosa oxldasË concentratlons of 50 - 500 mU/ml. Thls r¡as
conrpletely lnhlblted by the addltlon of catalase at aconcentratlon of 50 U/ml (Ftgura E-6).
2. Effact of oxpgen radlc
Predr¡rllcD
Tha next phase ln thls serlcs of exper¡nients uag todetrrmlne the effect of an Ëxog€'nousþ applled oxygen radlcal flux
upon hyaluronlc acld productlon by sytavlal celb ln culture. Ì1on-
dlseased boyine synovlal cells, ag used in the cytotoxlclty studies,
uJerÊ incubated rrlith 3H-glucosamine hydrochloride. lnltlal

FIEURE E-3 EFFEET OF SUPEROXIDE DISHUTÊSE ON XO/I.IX INDUEED 5IËT
RELEÊSE.*
500 concentration = ?5 units/ml
FIfìURE E-{ EFFECT OF EÊTÊLÊSE OH XO/IIX INDUCED 5IC' RELEASE.,I
Eatalase concentration = 50 units/ml
FIfìURE 6.5 EFFEET OF NÊNNITOL ON XO/HX INDUEED 5IE' RELEÊsE.*
* The error bars rÉprÊtÊnt the mÊan t 5.0. of triplicate
determination¡. Th¿ % 5l Cr relaase in the +re controls in
these experiments = 91.2 ! 1.1.

100
PERCENT80
CHROMIUM
ó0
REIEASE
10
EFFECT OF SOD ON XØX INDUCED CHROMIUM RELEASE
soDNO SOD
05 20 50
20
100
PERCENT
80
CHROMIUM
óo
RELEASE10
20
XANTHINE OXIDASE CONC
,( lO-3 units/ml
EFFECT OF CATALASE ON XqhX TNDUCED CHROMIUM RELEASE
ro0
r0005 20
CATATASE
50
XANTHINE OXIDASE CONC
, lO-3 units/ml
CH RETEASE
NO MANNIÍOI
2OmM MANNIIOL
IOOmM MANNIIOL
EFFECT OF MANNTTOT ON XqfiX lNpUCEpr00
PERCENT
80
CHROMIUM
ó0
RETEASE10
5
XANTHINE OXIDASE
. lO-3 units/ml
20
coNc.
20

1
I
FtfìuRE 6-6 EFFEET 0F IìLUEosE/BLUcoSE oxtDÊsE 0N Slcr RELEÊSE.
[ìlucose oxidase = 50-500 mU/ml
[ìlucose = 8"5 mH
Catalase = 50 units/ml

100
PERCE NT
80
CHROMIUM
óo
RELE AS E
40
EFFEC T OF
ON
GLU COSE/GLUCOSE OXIDASE
CHROMIUM RELEASE
Il
20CATALAS E
GLUCOSE OXIDASE
unity'ml
50510

Peak Ê is also completeþ susceptible to digestionr,tlth hyaluronate -lyase from streptomyceshyelur.onolyticus (d¡t¡ not ¡hor^m).

rz8
ÊxpËrlmÉrlts, shorrm ln Flgurc E-? and E-8, rr.lere performed toldentlfy hyaluronlc acld tn thc call culture mpdtum. Thc
suPËrnätants fronr tr¡ò 25 cmz tl¡ruc culture flasks contalnlng
confluent synorlocytes brËrÊ pooled after a .18 hr laballing period.
Êftar concentratlon, these uJÊrÊ fracflonated on ä Sepharose 4E-
ÊL column under dlssoclaflre condlflons (i.e" { Ì1 guanidtnium
chloride)' The chromatggraphtc proftle demonstrated 2 peaks oflH-glucosamlne containlng matarial. Tha higher molecular peak
(peal A) elutad niar Po and a louer molecular rreight peak (peak
E) cluted in the lncluded rolume. Electrophoreüc separaüon of
material from each peak uJàs pËrformad on cellulosa acetate
strlps, (Flgure E-B), and hyalurontc actd r¡as ldenttfled tn paak Ê
but not peak E"
Eorlne s¡rnorlal cells thtrefore produced rslalclvely small
quantltlas of a hlgh molccular rlelght hyaluronlc acld that elutcr
at the Po of a Sepharo!Ç {E-CL colunn undcr dlsrociaflre
condltlons.
l¡Je next exposed confluÉît monolaye?s of sgtnovlocytes h Zs
cmz tlssuc culture flasks to the xEltlx axygeî radical generaüng
rystem. The xanthlne oxldasÊ concentratlon used uas 5 mU/ml ln
the preseïcÊ of 2 ml-l hypoxanthlne. Thts resutcá rn conrplete
abolitlon of both peaks and all the radtoactivity eluted near V¡
(Flgure 6-9).
Subsequently confluent monolayerr uJerÊ exposed to a xanthine
oxldase concerrtratlon of I mU/ml. Flgure 6-10 ghours the rcsults
one such expcrlment. Exposure to xanthine oxldagç alsne shok s
the prescncÊ of tr¡o peaks, a¡ ls rcÉrì ulth unËxpoicd cclls,
houerer ln the prËtcncË of 2 ml1 hypoxanthlne the htgher
molecular r.rlelght peak dlrappearr. The proflles rhoun ln Flgure E-
l0 dúrcr from those shoum in Ftgure 6-? in that the cell products

FIT¡URE E-? SEPI.IÊRI]SE {B-CL EI.IRONÊTOERRPI.IY IlF T}IE SUPERNÊTANTs OF
Stt-ctucûsÊÌ1rNE LÊEELLED EoprNE syNoptÊL EELLS.
Borine synorrial cells, grouJn to confluence in tu.ro ?5 cmZ
. tissue culture flasks. Th.y uJene then cultured in routine
tissue culture medium (see Êppendix lll) containing 50 ¡.rCi
SH-glu.osamine for 2{ hrs. The supernatants uJere then
pooled and concantratad, and the concentnate applied to aSepharose 4E-CL column r¡hich r¡as eluted r¡ith { H guanidine
chloride. Tr.,-ro peaks of glucosamine containing material are
apparent, a high molecular rrreight peak (Ê) and a lou.ren
molecular r.rraight paak (B).
FIEURE 6-fl CELLULOSE ÊEETÊTE GEL ELEETROPIIORESIS OF T1ÊTERIÊL FROTI
PEÊKS Ê ÊND E"
5 ¡.rl of fraction 2E 19- <) and 5 pl from fraction 5E (o"<1
from the material demsnstrated in Figure E-7 were
electrophoresed under conditions outlined in Ëhapter lll. Ê
standard of hyaluronic acid r¡as run concurnently and stained
r¡ith Êlcian Elue. The area of Êlcian Blue staining is indicated
at the bottom of the figure. Radioactirre hyaluronic acid r¡as
identified in peak A only.

3x- GTUCOSAMINE INCORPORATION BY UNEXPOSED SYNOVIAL CELLS
DPM 15,000
50¡l
10,000
5,000
SEPHAROSE 48 - CL CHROMATOGRAPHY
Vg
tPeak A
Peak B
a
0 10 20 30 40 50 60 70
CELLULOSE ACETATE CEL ELECTROPHORESIS OF HATERIALFROT PEAKS A&B FROT FIGURE I
DPil 3002mm
r-< Fraction 20 (F¡gue 1)
O€ Fract¡on 56 (FigurelJ
200
100
12 14 16 18 20
vt
t
B0 90 100
FRACTION NUiIBER
oI
I
I
I
I
I
I
I
I
II
I
o24TILLIMETERS FROT T}IE ORIGIN +VE
lAREA OF ALCIAN ELUE SIA¡IIIIIGFOR AI{ HYALUßOIIIC ACID STAÍ'IDARD
0
-VE

FI6URE 6-9 SEPIIÊROsE .18-CL EIiROHÊTOIìRÊPI'IY OF SYNOPIÊL CELL
SUPERNÊTÊNT:' LÊBELLED ÊFTER EXPOSURE TT] TTIE XÊNT}IINE
OXIDÊSE/HYPOXÊNfiINE OXYBEN RÊDICÊL IìENERÊTING SYSTEM.
Eovine s¡morial cells, gpouJn to confluÊncÊ in tr¡o ?5 cm2
tlssuc culture flasks uJÊFÊ axposrd to xanthlna oxidase 5
mU/ml X0 and hypoxanthinc 2 mll in Dulbecco's PES for 2 hours.
Thry uJÊrÊ then l¡aghed and cultured in routlne tissue culture
medium containing 50 ¡.rEi 5n-glucogamlne for 24 hrs. The
supÊrîätantg uJÊpÊ thcn pooled and concmtrated, and the
concentrate appliad to a Sepha?ol,e'{E-EL colurnn uhich r¡as
eluted r¡ith 4 l'1 guanidine chloride.

3H - GLUCOSAUINE INCORFORATION BY SYNOVIAL CELLS EXPOSED TO5 mU/ml XANTHINE OXIDASE
DPM 3OOO SEPI-IAROSE 48 - CL CHROMATOGRAPHY50¡¡l
Vg
IYt
I
2000
1000
0 10 20 30 40 50 60 70 80 90 100 110
FRACTION NUMBER

FIGURE E-IO SEPIIÊROSE +E-EL EI.IROI'IRTOIìRÊPI'IY FROH SINÍìLE E5 CHZ
TISSUE CULTURE FLÊSKS CONTAININE CONFLUENT SYNOTIÊL
CELLS ÊFTER EXPOSURE TO XÊNT}IINE OXIDÊSE RLIINE ÊND
XÊNTTI I NE OX I DÊSE /IIYPOXHNTTII NE.
The panel shor¡s results from a tissue cultura flask thatrdas Êxposed to I mU/ml X0 in Dulbecco's PE5 before a 21 hour
labelllng period uith 3tl-glu.osamhe as outlined in the legend
to Fig. 6-8. The bottom panel shous the ruper'natant fronr a
flask exposed to I mU/ml X0 in the prÊrencÊ of 2 ml"î tlX. The
columns urÊrË eluted r¡ith 0.5 11 NaÊc' pll 5.6. The l/s r¡as at
fraction Z I and /1 at fraction 93.
Hyaluronic acid uas not identifled by cellulose acetate gel
electrophoresis in peak E ln either profllc.

3H - GLucosAMtNE rNcoRpoRATroN By syNovtAl cELLsEXPOSED TO 1 mU /mI XANTHINE OXIDASE
-
DPM1o ¡l
300
250'
200
150
100
50
0
400
350
300
2æ
200
150
100
50
0
XO ALONE
XO/HX
10 20 30 40 50 60 70 80 90 100
FRACTION NUMBER

r32
fractionated arc the contents of one 25 ct¡tz tiscue culture flask
that has been labelled for 21 hrs. ln addition, fractlonation uras
performed under associatlre conditions (1.e. 0.51'{ l'la Ac, pll 5.8).
Thls has resulted ln moremcnt of peak E touards Vo suggetting
that the componmts of peak E hare the ablllty to âggrÊgate and
that thls ls not af?cctad by exposurc of the cell to the full X0/HX
system ln thc mËînËr dcgcrlbed. Efforts to ldcntíy lche Prclc-ncË
of hyaluronlc acld, by gd elactrophorcslr, ln thc rlnglc peak of thc
proflla sÊÉ'n tn the X0/tlX cxposed cell rupernatantl u,crÊ
unsuccÊrsful (data not shor.m).
The flnal serles of experlnrtnts, shorrnr ln Flgure E- I I ' LJÊre
deslgned to examlne thc effect of superoxlde dlgmuta¡e and
catalase in thls systrm" The crrdltlons urÊrÇ identical to those
descrlbed above. Suproxlde dlsmutase added at a concsntration
of 100 U/ml had no ef?ect, houerer ulth the addltion of 100 U/ml
catalase, tulo peaks uJGrÊ agaln erzldcnt after fractlonation.
D. DlSCUSSloll
The cytotoxlclty studles descrlbcd abore demonstrate that
catalase protects adhercnl s¡novlocytes from the cfïects of an
çrtraccllular oxygcn radlcal generatlng rysttm. Superoxlde
dlsmutasc r¡lll renrorc 02-r to a dcgrec that ulll lnhlblt a Fenton
rcactlon and thus hydroxyl radlcal productlon. Thc flnd¡ng that
superoxlde dlsmutasc uag not effcctlve ln thls tystsm uould
suggest that hydroxyl radlcal, generated extracellularly, uas not
nÊcÊrtäFy îor the lethal ererrt to occur. The fact that mannltol,
å icèrÊrrger of the hydroxyl radlcal, u.ras minimalþ effectlre r¡ould
support this ffndlng. The toxicity oî the glucoselglucose oxidase
system and lts ablatlon by catalase, conflrms the ablllty of
hydrogen peroxlder !ËhËpâted extracellularly, to act as a
cytotoxlc agent.

FIEURE E-I I EFFEET OF SUPEROXIDE DIST1IJTÊSE ÊND EÊTÊLÊSE ON XÊNTHINE
OXIDÊSE/I.IYPIIXÊNTTIINE INDUCED SUPPRESSION OF HYÊLURONIC
ÊEID PRODUETION BY CULTURED EOPINE SYNOPIAL CELLS.
This figure shous a series of profiles, each rcprÇsenting the
radloactlvll.y aften fractlonation of the concentrated
rupÊfiìatants of ringle 25 cnß tlg¡uc culture flaskr. The
elutlon buffer r¡ag 0.5 11 NaHc, pl'l 5.8.
The top panel rÇprÊrentg the cmrtrol l.e. unÊxposed cells.
Thc gecond shor¡r expolure to tha X0/l'lX system (X0, I mU/ml;
I'lX, 2 mH).
The third panel ¡hous the effect of the addltiør of 100 U/ml
500.
The fourth panel shouc the effect of the additim of 100 U/ml
catalase"
The tr¡o primary peaks as illustrated in Fig. 6-? are
apparerrt. Exposure to the X0/l'lX system result¡ in the
dlsappearance of the highar molecular r.ueight llÊ containing
peak uhlch is partialþ rertored by catala¡e but not by S0D.

500
DPM 050 rul
CONTROL
VO400t
300
200
100
500 XO/HX
400
300
200
100
0
400
200
500 SOD100 /ml
300
100
500 CATALASE
400 100 /ml
300
200
100
30 40 50
FRACTION NUMBER
0
0 20

r34
llydrogen peroxide ls a small and unchargcd molçculc and
rdatlrely stabla r¡hen cornparËd to thc hydroxyl radical
I'lydrogen peroxide may therefore be exptcted to be abla todiffuse auJày from lts extracallular slte of producflør and crors
the cell membrane. Thi¡ r¡lll raise intra-cellular hydrogcrr peroxide
lerzels and lf enough hydrogen peroxlde ls generatad, the lntra-cellular antloxldant defensÊs urould ba oyeruhelmed" I'lydrogen
peroxide uould then be frea, not only to madlate intnacellular
damage pËr lrÊr but also to fnteract l¡lth other radlcal specles
that arÊ gËnÊrated as by-products of ccll metabolism.
Suparoxidc anlsn for example, is genaratad by a number of
lntraccllular procersÊs. lf endogcnously produced superoxide anion
ls not adequateþ scaranged ln a sltuatlon of oxldatlre strass, ltcould lnteract '¡lth hydrogen peroxlde to lcad to hydroxyl radlcal
productlsn rla a llabtr-hJelss reactlsn. The prurc'nce of
tntracellular lrwr rr¡ould cåtalyse thls rcactlon (1.e. al a Feryttsn
reactlon). Erldence that supports thls mechanlsm has recerrtþ
been produced by SCI'IRÊUFSTHTTER gÈ el. [ | 985] uho hare recently
documented the depletlon of lntracellular l1ÊD and glutathione in a
cell linc exposed to hydrogÉn pcroxide. Thls lndicateg a depletion
of the capaclty of the glutathlone/glutathlone peroxidase system
to act as an oxldant defence mechanlsm.
The addition of ruperoxlde dismutase and catalase to tha
extracellular medlum ls unllkely to alter lntracellular superoxlde
dlsmutasc or catalage lcrels due to the size of thesc molecules.
llannltol, hoularer ls small and unchargcd and therefore likeþ tocrors the cell menrbrane. lt¡ mlnor protectlve ¿ff¿ct mËy be due
to hydroxyl radlcal rcar+rrglng at lntracallular sltcg. Also houeve?
its mlnor protactlve ;ffect-, tËÉrt at high conccntratlont, may be
slo

135
due to non-tpËciflc scayenging since so-called oxygÊn radlcal
tcêrffigers trÊ notorlousþ non-speciflc.
Chrsrnlum release ls a manlfastatlon of cell þsls slnce
lntracellular chromlum ls bound to protetns that ulll onþ be
released r¡lth loss of ccll mambrana lntegrlty [R0tlAl, l9E9]. Êell
menrbranÊr år? partlcularþ susceptlbla to the deleterlous efrects
of oxldants through thc procËss of llpld peroxidatlon. Nelthcr of
thesc obseryatlons ln themsalyes indlcate ¡¡hather mambrane
damage ls a I o or 2o erent in oxldant inducad cell damage.
llouever tha sansltlrlty of hyaluronic acld production to lor¡ lerels
of X0/HX r¡ould suggest call membrana damage uràs ån earþ
eve,nt ln oxldant damage" PREHI1 tlgB{l and PtllLlPSON and
SCtlURRfZ I l98ll hare demonstratad that hyaluronlc acld
synthatase is situatad on the lntarnal surface of the call
membrane and nerr.lly syntheslzed hyaluronlc acld ls secrated
dlrectly Ínto the extracellular spacÊ uhereas other
gþcosamlnoglycans arÊ s¡mtheslzcd ln the cytoplasm. Thercfore
the obsarratlon that ËxpoturÊ of s¡novlocytes to I mU xanthine
oxldase ln thc prËsÉ.ncÊ of hypoxanthlne rcsult¡ ln the lo¡s of
hyaluronata productlon, but not the productlon of other Il-
acetylglucosaminc contalnlng products, ruggcstg earþ membrane
damage at a stage r¡hctr cytoplasmlc conrpmeîtg are still
functlonal.
PUflRl0 Êt il. t l gEZl hare reported that synoviocytes groun
from patiants ulth rhcumatoid arthritis ulll produce lou molecular
uelght hyalunonlc acid uhen establlshed in culture. Synthesis of
such a product fi vlvo lg an alternatlre explanatlon for the flndfng
of decreased arerägÊ molecular rrlelght hyaluronlc acid found tn
lnflammatory arthropathles. 0ur flndlng that expoture tosublathal lerals of X0/HX rçsults in cessation of hyaluronic acid

r36
productlon rËthÊr than s¡mtheslg of a smalltr product uould
suggest that oxldant Ê,(posurÊ can be dlscounted a¡ a cause for
such an alteratlon ln synthesir. lt ls of intercst to note that
EATES, L0lrJTtlER and l'lÊllDLEY [19E5] hare also reported thatchondrocytes cxposad to sublethal lerels of oxygan radlcals uill
decrease the amount of protcogþcan produced, but r¡lll not
cffcct the glze of the proteogþcan produced by these calls.
E" SU]I}IRRY
Eorlnc s¡novlocyta¡ ln culture êrÊ tusceptlble to oxygen
radlcalg pnoduccd by X0/llX at mcäturcd by'lCr releage.
Êdnrlnlstratlon of catalasa uJat protactlre u¡herËêl luPËroxide
dlsmutasê uJât tneffectlvc and mannltol had a minlmal efl'ect. This
lndlcatr¡ that hydrogcn perorlde ls thc ag+nt nccattËPy for
toxlclty. Sublethal oxygcn radlcal fluxes ulll result ln ccssatlon of
hyaluronlc acld synthesls evsn I¡hct the r¡mthsglr of othcr lll-
glucosamlne contalnlng products contlnucg. l{o lor¡ molecular
r.rleight hyaluronic acld products could bc detectad in the
superîatants of oxygen radlcal cxposcd cells.

r3?
C}IRPTER YI I
Dtscu35toil

t3E
fi. lü vlTRo oxYtEil RÊDtcRL HDUCED HTRLUR0ilTC RC|D
DEPOLYIIER I ZÊTI O]I.
l. llolecular r¡clght studlas.
Tha shnple chemlcal structure of hyaluronlc acld belles lts
complax phyrlco-chemlcal propartles and blologlcal roles. Thasa
propartlÊ! ËrÊ duc to lts macromolecular structure ¡¡hlch
depends upon both lnter- and lntra- chaln interactlon¡ and r¡hich
lcad to thc formatlon of a matrlx ln âqucoul solutltrtg of
sufflclant contentratlon. lt ls also âppârËnt from thc data
presented in this thesis, that a consideration of macromolecular
structure is necersåry bafore an hypothasls of hyaluronic acid
degradation by oxygÉï radlcals can be formulated.
The findlngs that Émarge from the studles dme ør the
molacular r.rreight changes after oxygÉyr radical lnduced
depolymerization and r¡hich must be reconcllcd lnto ruch an
hypothesls are:
l. The smallast dapol¡merlzatlon product lr 104 molccular
r.lclght.
Z. Thera ls a rapld progragslon from large molccular ulclght
hyaluronlc acld to the gmallegt rlze dcgradatlon product.
3. Ê dagrce of rcpol¡merlzatlon of degradatlon products ls
probable.
The flrst flndfng ls the most clear cut and may be accounted
for by a number of depol,,¡merlzation mechanlsms, Ìlost obriou¡
are those mechanlsms that require that there be a site along
the hyaluronic acid molecule that ir particularþ susceptiblc toclearage by oxygen radicab.
Perhaps the most llkely propcrty of hyaluronic acid that¡¡¡ould lead to a serles of susceptible sites along its length is its

t89
tcrtlary structurc ln aquËoul solutlon. Ês outlfncd in Chapter I'
hyaluronate, in aqueou¡ solutlon, ls bellerued to adopt random coll
best characterlzed alt a left-handed hellx. The random nature of
the colling of the hyaluronate molecule ls one feature that may
lead to GxporurË (or increased accc¡gabttity) of a glycosidlc lÍnkage
ts the surroundlng cnrzironm?nt. This in turn could make such a
linkage susceptlble to oxldatirze damagc" Êlternately structural
cEnstralnts may be imposed upon the tertlary structura of a
hyaluronic acld molccule in solutlon by lts interactlon ulth other
hyaluronate molcculcr ln the rämË rolutlon. Structural studics
IhJ|NTER, Sl'llTTl and ÊRi{0TT, l9?5J hdlcate a degrec of stffneæ of
the hyaluronata molacule ln aqueous solution, clearþ then the
formatlon of a matrlx ln s.olutlon r¡ith so mäny "ttiff' moleculas
r¡ould cåurË lrregularitles,fcrtiary structura to occuP. l,rlhat is
claar fnom our studles ls that these susceptible sites must be a
conslderable distäncÊ apart to account for the smallest moleculan
r.lelght degradàtion products harlng.a molecular rr¡cight of 101
(i.e. approximateþ 50 saccharldes)" Thls hypothesis predlcts that
an hyaluronate chain of lesser length is not long enough to adopt
the tertlary structure that leads to the formatløt of a
susceptlble slte.
The chclatlon of lron ls clearly a critlcal factor in all the
oxygÉî radlcal reactlon¡ utlllzed in this studyt and manl E vlvo
reactlons lnrolrlng radlcals (gee Chapter l). lt may be that by
rlrtue of lts tartlary gtructurc that hyaluronlc acid can chelate
lror or another metal lør ln such a r¡Jäy that the gcneratlon of
tha hydroxyl radlcal occurs ln close proxlmlty to the hyaluronate
chaln. lt ls claar nouJ from a numbar of studles that hydroxyl
radlcal productlon ls nÊcÊssäry for hyaluronic acid

r40
depolymerlzatlon [EffT:t and CLELHND' I gEZ; HBFl"lÊNl{ ÊND SCHHUT'
l9E0l. The extremely high rcactirzlty of the hydroxyl radical
dlctates that lt must bc generated close to its target ¡ite since
It so lndlsrfminately neactire [FEE and PÊLEt{Tlt'lE' 197?1" This
sltuatlon r¡ould thereforË rÊPrcleltt a form of "site-specific
Fenton reaction" i.Ê. the site of chelation of a metal ion capable
of catalysing a Fenton neaction determines r¡hene hydroxyl radical
damage occurs. ln tha case of hyaluronic acld this r¡ould be close
to tha Fl-S glycosidic linkage as d¡scussed ln Ehaptar lP. This
hypothesls t¡ould thus neconcile the high and non-speciflc
reacür¡ity of the hydroxyl radlcal u¡lth a specific alycosidlc bond
clearage"
Hrn lmpllcation of theEe "susceptlble site" hypothesËs ls that
they may prorzide an explanatlon for the second statcd flnding of
the molecular r.lelght studles. Depolymerization proceedlng from
"susceptlble sltc" to "lulceptlble sitÊ", ln an Íneruitable chaln
reaction r¡ould regult in the obscrration of a rapid progresssion
of largc molecular r.leight hyalunonic acid to tha smallest size
degradation product r¡ith a ralatire paucity of intermediate sized
materlal.
0ther hypothetÊl arË porslbla. The gencration of lor¡er
molecular r.r,relght hyaluronic acld may interferc uith further
orlygÉTt rad¡cal production. This may occur lf the small molecular
rrletght products act as hlghly efflclent oxygÉTt radlcal lcarengers.
Alternately hyaluronic acid depolymerlzation products rnay
interfere uith chelator-rnetal bindingr thus prerenting further
hydroxyl radical generatlon. The chelator may be EDTÊ' or' äs
suggested prerlously, hyaluronlc acld ltself.

r4l
Finally the ruggestion that axygcn radical damaged
hyaluronlc acld may repolymerlze suggests a furthcr mcchanism
that r¡ould llmlt the slze of the degradatlon product to 104 . Ên
equlllbruim may be reached r¡ith depolymerizatlen predominating
at large molecular rrreights and repolymerization predominating at
small molecular rlalghts.
Ê number of there hypothese¡ are testable and lead to tha
folloulng speciflc questions being posed.
l. To r¡hat extent and hor¡ tlghtly does hyaluronic acid bind
(labelled) lron on other metal ions?
2. Can hyalunonic acld degnadatlon products interfcra r¡ith
chalator-metal bindlng?
3. Êre hyaluronic acid degradation productg morê efflcient
o¡(ygÉrr radlcal scavÉngÊrt on a mole for mole basls that
undepolymerlzed hyaluronate?
4. Can crolt llnked or branchcd hyaluronlc acid be found in
the depoll,merlzation products of oxygen nadlcal exposed
hyaluronic acid?
SAT0 and llll.dÊ (l9ES) hare addressed the question of the
nadical scarenglng capaclty of hyalunonlc acid (yas) and its
constlturnt sugars, D-glucuronic acld (No) and N-
acetylglucosamlne (yss¡. llyaluronlc acld dcAradatlør products (i.e.
of about 104 mohcular r.rlelght or gmaller ollgosaccharldcr)
houevçr harç not barn axamlned.
lrJa hara looked for crosslinked or "brânchad" hyalunonic acid
in oxygen radlcal exposed hyaluronlc acld. Thls r¡as dont by gel
chromatography of oxygan radlcal exposed hyaluronic acid that
uas axhaustlrely digested r¡ith streptomycÊs hyalunonidase.
llo¡¡ever our abillty to nesolve the oligosaccharides uas not

r42
sufficlent to allor¡ a concluslon. Ner¡ar methods using
polyacrylamide gel electrophonesis to separate hyaluronate
oligosaccharides may be helpful in this negand [l-lHllPS0N and
tìÊLLÊ61'lER, l9E5l. The findlng of a product, chanactenistically
altered by oxygen radical exposure r¡ould prorride a means to
establlsh unequirocally r¡hether lnflamed synorrial fluid hyalunonic
acid is depol¡,merized by oxygen nadical exposune in rivo.
Z. Reducino Ends.
-
The findlng in our study that reducing ends are released
during the oxygen radical induced depolymerization of hyaluronic
acid is clear cut since it has been obserred by both colorimetric
and radiolabeling techniques. Thls finding is in contrast tu some
othen early reponts IHcELEÊN and llÊLE, lg4tl; EREENITIÊLD and
H0Y, tg80l. Houerer ELELÊN0 et ê1, I lgEgl, using the aEcorbic acid
induced drpolymenization of hyaluronic acid r¡ene able to detect
the release of reducing ends uslng sodium bonohydnide labelling.
l.rJe dlffer fnom these authons hor¡erer in finding that it is the F l-5 glucuronidic linkage that is preferentially cleaved nather than
the F l-{ hexosaminidic linkage. l,rJe har¡e been careful not to let
clearage by the alkaline conditions required during both the
colorimetnlc and radiolabelling assays lead to misleading nesults.
ShJÊNN É êt t I 96Êl hare determined that hyaluronic acid is not
depolymerized unden the conditions used in oun study for cyanide
labelling. Ê similar cniterion r¡as also established for the
colorimetric assay of Park and Johnson (1.e. pl'l not greater than
9.0). This is of importance since the F l-3 glycostdic linkage of
hyaluronic acid is relatlvely labile in alkaline csndltions [R0EIN50H
and l'l0Pl,rJ00D, l9?31.
Tr¡o other llner of evidence r¡ould also support clcarage atthe F l-5 glycosidlc linkage. The first comÊs fnom the uork of

r43
BÊLÊZS et êt I lgE?] urho reported the electron spin resonance
spectroscopy findings of hyaluronic acid in aqueous solution
exposed to pulse radiolysis. Pulse radiolysis in aqueous solutions"
leads to the generation of the hydroxyl radical as r¡ell as the
aquÊous electron and superoxide anion. Their findings uJere best
explalned by the formatlon of tuo translent lntermedlates thatcould be produced by eithen the hydroxyl radlcal (relevant to our
oxygÉTr radical ge.nerating systems) or the aqueous electron (not
relerant). The first transient intermediate is produced by
remoral of an electron fnom the oxygen of the F l-5 glycosidic
llnkage. This r¡ould react furthen girring rise to clearage at this
linkage. The second intermediate, seen at high doses of nadiation,
inrolves labilization of the hydrogen on E5 of D-glucuronic acid.
This could be stabillzed by a Fesonance lnteraction uith the
caboxyllc acld group of this sugar. This latten intermediate måy
hare significance in any rÊpolymerization neaction seen as à result
of interaction of hyaluronic acid ¡¡ith the hydroxyl radical.
Secondly our results allor¡,an explanation of the findings of
EREENUJÊLD and t10Y I l9?9], r,.rho reported that hyaluronic acid
exposed to oxygen radicals is mone susceptible to dlgestion by the
neutrophil lysosomal enzyme, N-acetylglucosaminidase. This
enzyme uill remore N-acetylglucosamine fnom the non-neducing
end of hyaluronic acid ILINKER, |'1EYER and l.¡JElSSt1Ê¡lì 1955].
Therefore if oxygen radical depolymerization of hyaluronic acid
representE a situation r¡here only the F 1-ã glycosldic línkage ís
cleaved, then all cleavages r¡ill leare H-acetylglucosamine at the
non-reducing end. This uill allol¡ maximal subEtrate for this
enzymÊ and thus a maximal amount of N-acetylglucosamine r¡íll be
released upon digestion r¡ith this enzyme.

L44
E. STUDIES OF sY]IOV'ÃL FLUID }IYÊLUROIIIC RCID ISOLßTED
FRO}I IIIFLR]IED JOI]ÍTS. RRE OXYf¡E]I RRDICßLs RESPO]ISIELE?
Ê large number of studles hare reported the changes in
synovlal fluid that occur as a result of tnflanrmation. Êlthough a
numbcr have presentad dlrzergtnt results for the molecular
r.r.relght changcs lcËn ln synorrlal fluld hyaluronlc acld (see BÊLÊZS
ç3 åL I l9E?], DÊl'lL Êt åL I I 985]), a number of characteristlc
changes arË agrÊed upon. These arc:
" incretsed synovlal fluid rolume
. decreased synorlal fluid hyaluronic acld concentration (and
hence the clinical finding of a decreased viscosity of rau
synorzlal fluid).
. lncneased "total amount" of hyaluronlc acld presetrt ln the
synorzlal fluld as ê result of the lncreased rolume.
" decrças¿d avsragc molecular r.lelght of the hyaluronic acid
presant.
Thc molecular rlelght spectra of lnflamm atory synovlal fluid
hyaluronlc acld prescnted in thls thesis årË conilstent ulth other
studles demon¡tratlng that the decreasÇ rn average molccular
rrrelght, sÊÉrr during fnflarnmation ls smalf but dcfinlte. ln inflamed
synovial flulds, mort synovlal fluid hyaluronic acld rerflains excluded
on Sepharose 'tE suggesting a ve?y hlgh molecular ureight"
l{erertheless, ulth inflarnmatlon a greater proportlon elutes in the
fncluded rolume and material can be indentifled doum to amolecular uatght of 2x l0{. ttf addit¡onal intarest are the shapes
of the proflles. Host lncluded rolume materlal ls seen ås a sker.rllng
of the excluded voluma peak into the lncluded volume houever a
number of our lnflamed flulds hare ä sÉparäte pcak in the includad
rolume on Saphaross 18. Thls l¡ould be pradlcted îrsr¡ our flndlngs

r4õ
in the þ ritro studies that degradation procecds rapidly to the
smaflest degradation product tÊËn (¡.e. molecular r.leight of lElJ.
This second peak may rÊprËrênt small molecular ulelght breakdol^m
productsr prÊseît in sufflcient quantity before thay can be
cleared from the Jolnt spacË.
l¡Je, llke others ISUI{DELÊD, 1953; DÊl'lL Et åLlgA5] could not
flnd any rlgnlflcant corrclatlon bctLlËêt1 thc decr?atÊ in areragc
molccular uclght and othcr lndlces of lnflanrmation (crythrocyte
sedlmantation rate, haemoglobln, synorrial fluld r¡hlte cell counts)"
Eoth ¡idcs of this attempted correlation hou¡ever, represent
impreclse mËärurÊr" Systemlc mÇasurÊs of inflarnmation
(erythrocyte sedimentation rate and fall in haemoglobh) may be
at rariance r¡ith the dagree of synovltls 'nvolvlng the joint
sampled. Local meaiurcs (synovial fluid cell count) are abo
lmpreclse e.g. some ve?y lnflamed Joints hare no effuslon at all"
0ther factors, such as the productlon and clearance of
rarious slzcs of hyaluronic acid from tha synovial fluld are goÍng
to effect the molecular r.r;eight proflle of s¡novial fluid hyaluronic
acid sampled.at any particular time. l'lyaluronic acld is being
samplcd at one point in a dynamlc procËst depcndtnt upon the
ra-tes of both s¡,rnthesis of hyaluronlc acld and its clearancË from
the joint rpäcË. The peturbatlons of thcse latter Processes by
inflanrmatory change are not knoum, indecd llttle ig knot fi of
hyaluronlc acid turnorÉr. Studies by ÊNIOI.|AS' FRÊSER and
HUIRDEN [19?2] hare shor,rn the very rapid äPPearance ((15
mlnutes) of labelled hyaluronic acid in the draining lWph node of
the rabblt knee after intra-articular lnjection.' Thls suggests a
rapld rate of turnorer. Êny molecular r.r.relght change produced by
a depolymerizatlon procÊrs !tr vivo is depcndent in part upon the

r46
"length of time hyaluronic acid is exposed to the depolyrnerzlng
agent, Ê teleological explanation for a high rate of hyaluronic acid
turnorer is that this brill limit the extent of deterioration of the
synovial fluid hyaluronate matrix due to a depolymerization process
and thus r¡ill preserre its biological functions,
C" EFFECT OF OXYÍ¡EH RRDICRLS OIt SYIIOYIOCYTES ffI{D T}IEIR
PRODUCTIOII OF TIYffLUROIIIC RCID.
Finally there remaing the proposition that hyaluronic acid is
not depolymerized n vivo - that there are enough oxygen radical
scarenging mechanisms arzailable in the synovial fluid to prevent
post- synt h et ic de polymerization. This hypo th esi¡ s u g ge s ts th at
inflammatory damage to type 1l sgnovtocytes causes them to
synthesize hyaluronic acid r¡ith a decreased average molecular
r,.reight. ln support of this hypothesis PUORIO É 4 tl gÊ21 have
published the profiles of hyaluronic acid produced in culture by
synoviocytes obtained by explant culture of rheumatoid and normal
synovtum. They reported that synaviocytes obtained from inflamed
rheumatoid and joints inflamed for other neasons produced
hyaluronic acid of a lor¡er molecular r¡eight than synovtocytes
obtained from normal or non-inflamed synovium. The Sepharo:e 2B
profiles for the products of inflamed synoviocytes shobJ minimal or
no material in the excluded volume and the peak of the included
rolume materiEl has a Kar of approximately B.?5 rorresponding to a
moleculan r.reight of 105. Simply stated this material is too small to
compapÊ r¡ith hyaluronic acid extracted fnom inflamed synorial
fluids. Ê number of factors r¡ill affect N-acetylglucosamine
inconponation into hyaluronic acid under cultune conditions. These
include the grou.rth phase of the cells and the glucose
concentnation. Êlso it may be preferable in this circumstance to
considen orgån culture as suggested by HYERS I l9E5]. The findings

L 47
of PUORl0 et al. [1981] therefore are more likely to indlcate thatthe synorriocytes fnom inflamed joints äne moFe susceptible to in
ritro lnfluences that cÊusÊ an altenation in hyalunonic acid
synthesis nathen than pnorring that a smallen size hyaluronic acid
molecule is produced by inflamed synoriocytes h 4!4,.
Ue appnoached thls question by asking uhether cultuned
synorrial cells could be altered by exposure to oxygen radicals in
such a rJay as to produce a louer molecular rrreight hyalunonic acid"
ln short u¡e found that cells exposed to sublethal doses of oxygen
radicals shut off thein productlon of hyaluronic acid nather than
produced a smaller product. l'lyaluronic acid production bJas more
sensitlre to oxygen radlcal damage than other cellular synthetic
functions. This is in keeping u.rith the r¡ork of BÊTES gE gt t tg8ll
r¡ho found that upon elrposupe of chondnocytes to a sublethal
oxygen radlcal flux the nates of synthesls decreased but the si¡e of
the product r¡as unaltened. Clearly r¡e hare only tested one of a
possibly very large number of factors acting on the synoriocyte at
an inflammatory site and r¡hich includes other neutnophil secnetony
products, T cell and macrophage products, cytokines and so on. ltis possible that one or more of these could lead to an alteration in
the size of the hyaluronate produced. Êlso more understanding of
the mechanisms of lnitiation and cesEation of hyaluronate chain
spnthesis are nequined before this question can be completely
resolred. llyaluronate synthetase, by rirtue of its close
association r¡ith the cell membrane is likely to be susceptible to
membrane disruption. [Jne of these is the production of oxygen
radicals due to the propensity of membranes to undergo lipid
peroxldation. Ê logical extenslon to oun studies is therefore toexamine other membrane damaging agents for their effect on
hyaluronate synthetase function and also to examine the cell

14E
supernatants for the aPPearance of lipid peroxidation products
particularly ln relation to the cessation of hyaluronic synthesis.
D. COHCLUDIHf¡ REHRRKS.
The effect of oxidants generated r¡ithin a biologic al system
hare been seen as an interplay betr¡een the reactivity of the
oxldant generated and the defence or scarenger systerns arzailable
to protect against that oxidant. lt has emerged frsn the r¡ork in
thts thesis that a third important factor in this lnterplay is target
susceptibility, The evidence in our gtudies on isolated hyaluronic
acid has pointed tor¡ards a site specific susceptibtlity to
depolymerization. Specifically þl-3 glycosidic linkages spaced at
sites along the molecule are susceptible. Equally r¡ith synoviocytes
in monolayer culture the loss of cell rziability and cessation of
hyaluronate synthetase function are mediated by hydrogen
peroxide, underlining the impontance of access to a target site
r¡ithin the cell.
Ê study of the effects of oxidants r¡ithin the context of joint
inflammatlon has been undertaken by examining in detail an
important component of joint physiology, namely hyaluronic acid.
Such a reductionlst tactlc has been used since the number of
effector agents, modffying factors and structures at risk r¡ithin
the inflamed join t arelÐnumerous to study effectively. Yet thiE
approach has p?oveî useful in exposing target sltes susceptibile to
oxidant damage
Ên ultimate aim of research in the field of rheumatology is
prerrention, by therapeutic interrention, of joint deformity u.rought
by the lnflammatory Pnocess. Ê clear appreciation of the specific
tyFe and extent of this damage that can be attr'ibuted to
oxidants is required so that appropriate therapeutic steps can be
taken to prerzent this form of joint damage from occunning.

r49
ÊPPE]IDIX I
uRoÌilc fictD Ê55RY
The technique desribed by HLUiIEÌ{KRÊNTZ and Ê5BUE-|'|ÊNSEI{
[19?51 uag used.
Ê" REÊGENTS
l. Heta-hl¡droxydiph
Ên 0" 15% solution of mcta-hydroxydlphenyl (Eastman Kodak
Eo., Rocherter, New York) ¡n O.5% t{a0ll urar prÊpared and ¡tored
in the refrigerator r¡¡ith an aluminum foil corering. This solution
uJas relnade erery three monthE.
Z. il;FL+/tetraborate solution
Ên 0.0125 n golution of sodium tetraborate in concentrated
sulphuric acid.
Ë. METHOD
0.2 ml of unknoun and standards containing from ?.5 - 100
ug/ml D-glucuronic are added to 15 xl00 mm test tubes. 1.2 ml
of the sulphuric acid/tetraborate solution is then added and the
tubes are refrigerated on crushed ice. The tubes are then
¡haken in a Portex mixer and heated in a boiling r.r.later bath for 3
minutes. Êfter coollng on crushed ica for l0- 15 mlnutes, 20 ¡.rl of
tha m-hydroxydiphenyl reagent is added. The tubes are shaken
and the color alloued to derelop fon 5 minutes. Êbsorbance
mÊåturãïents are tht¡ read at 520 nll on a Perkin-Elmer (LS
1500) spectrophotometer. Ileasurements on sämples fnom
fractions after gel chnomatography are done in duplicate, all
other determinations are in triplicate.

t50
FERRICYßI{IDE IIETTIOD OF PRRK RIID JO}I]IsO]I
Thic technlqut l¡ a modlflcation of the àsray orignally described
by PÊRK and J0l'll'150H [ 19111 and sub¡equently modified by H0UEK
and PEÊRCE [ 195?l îor use r¡ith hyaluronic acid.
Ê. E!.Agg¡lå Three reagents arÊ prepared I
I ) Femic,uanide solution. 0.5 gt of potassium ferricyanide per
liter; stored in a bror*m bottle"
Z) Earbonate-cpanide ¡olution; 5.5 gm of sodium carbonate +
0"ES gm of potassium per liter of solution. ln our experÍments the
pll of this reagent r.rras adjusted to a pl'l of 9.0 [ll0UCK and
PEÊRCE, tg5?1"
5) Femic ion solution; 1.5 gm of ferric antmøtium sulphata + I
gm of Duponol (sodium monolauryl rulphate, Du Pont lnc.,
l,rlilmington, Delar¡are) in t liter of 0.05N ¡ulfuric acid.
E. Hathod
0.5 ml of the unknoun¡ and standards contaÍning I -ZE ug/ml
glucuronic acid are added to 15 x 100 mm PyrËx test tubes. 0.5 ml
potassium fcrricyanida and 0.5 ml carbonate-cyanide solutions are
then added. The mixtune is then shaken on a Pontex mixen and
heated in a boiling r.r.rater bath for 15 minutes. The tubes ane
then cooled by immersion in a r¡ater bath at room temperature
for a further 15 minutes. 2.5 mls of the ferric ion solution are
then added and the mixture again shaken on a Pontex mixÊr. The
colour reaction is then read at 690 nPl on a Perkin Etmer (LS 1500)
spectrophotometer. Êll measurements are done in triplicate.

r5t
Ìl0Rf¡Rl{ - Elsoll REHCTI0I{
Thls technique ig thc modificatlon of tha Horgan-Elson reaction
. according to RElSSltì, STR0|'4|NGER and LEL0IR [1955],
Ê. BEÊEESIE
l " ; Hn [J.E l'l potatsium
tetraborate solution ig madc and the pll adjustÊd to 9.1 ¡¡ith
potassium hydroxide.
Z. P - dimeth,ulam¡nob Dl'1ÊE)-el¡¡lieg; l0 9m Dt1ÊE
(Eastman Kodak, Rochester, Fleu York.) is dissolved in 100 ml
reagent grade glacial acetic acid l¡ith lZ.5% (rrolume per rrolume)
l0 N hydrochloric acid. This reagemt can be stored at ?oE for up
to a month in a dark bottle. Shortly bcfore uge the golution ig
diluted ¡¡ith I rolumes of neagant grade acetic acid.
E. TIËIEED,
0"ZS ml roluma¡ of unknorrms and standards contalníng 2.5 -50
ug/ml N-acatylglucosamine are added to 15 x 100 mm PyrÊx test
tubes" 50 ul of potassium tetraborate solution is then added.
The tube! ârÊ heated in a boiling ulater bath îar 3 minutes. Than
they are cooled in tap urater and 1.5 Dl'1ÊE ¡olution i¡ added and
the tubes are shakerr on a Vortex mixer. The tubes are then
heated in a r¡ater bath at 3E - 38oE for 2O minutes. They are
then read at SES nH on a Perkin Elmer (LS 1500)
spectrophotomcter. Êll measurements are done im triplicate.

r52
RPPEI{DIX II
T|55UE CULTURE 50LUTl0l{5
Ê. PRI]1RÊRY CELL CULTURE IIEDIUI'I"
- llam's F l0 medium [HÊH' l9E3]
- Z0t6 foetal caff serum.
(heat inactivated by incubating ät 56oE for 50 mins)
- gentamycin 0.2 mg/ml.
- L-glutamine 5 mH added fresh.
E. ROUTINE SYNOI/IÊL EELL CULTURE IIEDIUII.
- llam's F l0 medium.
l0% foetal calf serum. (heat inactirated)
- gentamycin 0.2 mg/ml.
- L-glutamine 5 mll added fresh.
E. DULEEEEO'S PI'IOSPI.IRTE BUFFERED 5ÊLINE.
NaEl 8.0 g/l KEI 0.2 g/l
CaCl2 - 0.1 g/l HgCl2.6ll2B - 0.1 g/l
Na2HP0'1.2H20 l.l5 g/l KH2P04 0.? g/l

McNeil, J. D., Wiebkin, O. W., Betts, W. H., & Cleland, L. G. (1985).
Depolymerisation products of hyaluronic acid after exposure to oxygen-derived free
radicals. Annals of the Rheumatic Diseases, 44(11), 780-789.
NOTE:
This publication is included in the print copy
of the thesis held in the University of Adelaide Library.
It is also available online to authorised users at:
http://dx.doi.org/10.1136/ard.44.11.780

McNeil, J. D., Wiebkin, O. W., Cleland, L. G., & Vernon-Roberts, B. (1986). The
generation of reducing ends by exposure of hyaluronic acid to oxygen derived free
radicals. Agents and actions. Supplements, 18, 95-101.
NOTE:
This publication is included in the print copy
of the thesis held in the University of Adelaide Library.

r?0
EIELIOÍ¡RRP}IY
Êtenrcr, J.11., PlcERctxy, C.Ê. and llunst, J.K. (l9E l)" Eiological
reactir¡lty of hypochlorous acid: impllcations for microbicidal
mechanisms of leucocyte myeloperoxidase" Proc. Natl. Êcad.
Sci.USÊ ?8, 210-e 14.
Êl-Kr¡uor, U"Ê.3. and CrnalßssroN, T.ll. ( 1965). The specles dlstribution
of xanthine oxidase. Eiochem. J. 9?, 3lE-521.
Êmanuso, D.R. and Jottttsolt, R.E. ( l9E l). Lactoferrin enhances hydroxyl
radlcal productlon by human neutnophils, neutrophil
particulate fractlons and an enzymatic generatlng system.
J. Ëlln. lnvest. 6?, 352-380.
Êmes, 8.N., EßTHcñRT, R.r Scrutens, E. and llocnsrutn, P. ( | 9E I ). Uric acid
prorldes an antioxidant defense in humans against oxidant
and nadical-caused aging and cancer: ä hypothesis. Proc.
Natl. Êcad. Sci. USH ?8, 6ESE-6862.
ÊnnsrRsstRoEs, T. and RoeeRTsoN, lrJ. ( l9E{). l.lodulation of mitogen-
dependent lymphocyte stimulatlon by hyaluronic acid. J.
Rheumatol. I l, ?29-?54.
ÊneRn, 11. and NFrñ, P" ( | 96?). Ê compilation of specific bimolecular
rate constants for the reactions of hydrated electrons,
hydrogen atoms and hydroxyl radicals r¡lth inorganic and
organlc compounds in aqueous solution. lnt. J. Êppl. Rad.
lsotopes lE,495-5U5.
ÊtnonRs, K.l'|., Fnf,sEp, J.R.E. and Hu¡noEN, K.D. ( l9?3). Distribution of
biologlcally labelled radioactlve hyaluronic acid injected into
joints. Ênn. Rheum. Dis. 32, 103- I I l.
Êtnonrnr, E., Bnunopr, l{., Gnegnuooo, C. and J'lnlmsrnom, E.E. ( 1970).
Catalytic mechanism of cytochrome oxidas¿. Nature (London)
228, 93ã-937.

r71
Êsnulrtt, 6. ( lg5?)" Colorimatric analysis of sugårs. llethods
Enzymol. 5, ?3- l29.
Êtxrtts, E"D.T., He Roen, D" and Scon, J.E. ( lgËB). llodel for hyaluronic
acid tncorporating four intramolecular hydrogen bonds. lnt.
J" Elol. Hacromol. 2, 5l B-519.
Eaeton, 8.L., RosEN, R.E., Hcllunnrcr, EJ., Penns, N.Ê. and BRBIoR, 8.11.
( l98 l). Êrrangement of respiratory burst oxidase in the
plasma membrane of the neutrophll. J. Cl¡n. lnrest. 6?,
r ?2{- I ?28.
EReton, E.N, (19?E). 0xygen dependent microbial klllingby
phagocytes" l{err.r England J. Hed. 298, 859-6ã81721-'?23.
BReton, 8.H., KtRttEs, R"5. and ÊRnnurE, J,T. ( l9?5), Biological defense
mechanisms. The productlon by leucocytes of superoxide, a
potent bactericldal agent. J. Êlln" lnrest. 52, ?{ l-?4'1.
ERtnzs, E.Ê. ( l9?'l). The physlcal propertles of s¡rnorrial fluid and the
speclal role of hyaluronlc acld. ln: Dlsorders of the Knee.
Helfer Ê. (Edltor) pp Ê3-?5. J.E. Lipplncott, Philadelphia.
ERrnzs, E.Ê. and DRnzvrr¡¡blcz, Z, (.19?3). The effect oÍ hyaluronic
acld on flbroblasts, mononuclear phagocytes and
l¡mphocytes. ln: Blology of the Fibroblast. Kulonen, E. and
PlkkaralnËn, J. (Ed¡tons) pp 25?-25?. Êcademlc Pness, London.
Entnzs, E.Ê. and Lnunttn, T.E. ( lg5 l )" Tha viscosity function of
hyaluronlc acid ås a polyelectrolyte. J" Polymer Sci. 6' 665-
EEE.
ERtRzs, E.Ê. and Suttoltno, L. ( 1959). Vlscoslty of hyaluronic acld
solutions contafning proteins. Êcta Soc. HÞd. Upsalien. 54,
r 5?- r 46.

L7 2
ERtRzs, E.Ê., BntLLEn, S.l. and DgtttH6Ep, J.L. (1381). Na-hyaluronatc
molacular size rariations in equine and human arthritlc
synorrlal flulds and the effcct on phagocytlc cells. Semin.
Êrthritis Rheum. I l(suppl. l), l4l- 145.
ERtRzs, E.Ê., LRuRENT, T"C., HorrlE, Ê.F. and Hnnen, L" (1959). lnnadiation
of muco-polysaccharides ulth ultraviolet llght and electpons.
Radlat. Res. I I, l {9- 16'1.
ERtnzs, E.Ê., DfiutEs, J.P", Ptltttes, 6.0. and Youne, 11.0. ( 196?).
Translent fntermediates in the radiolysls of hyaluronic acid"
Radlat. Res. 5l, 245-245.
Eatnzs, E.Ê., hIRTsoN, D, Durr, l.F. and RosemRn, 5. (l9E?). l'lyalunonlc
acld ln synorrlal fluld. l. Holecular paramcters of hyaluronic
acld ln normal and arthrltls fluids" Êrthritls Rhaum. l0' 35?-
5?6.
Ennnlsmn, J.V,, BRrrttrten, lJ.l1., H|LL, ll.Ê.0", llÊttooo, J.F., hllLtsottr R.L. and
hlolrgnoer, 8.5. ( l9E0). Does ceruloplasmin dismutase
superoxlde? FEES Lett. I lE, 12?-129.
ERnrEn, 5.Ê., EfryyuK, J.5., Enlmnco¡,leE, J.5., HAt.lrltts, C.F. and STRCEY, l'1.
( 1965). Fhgerprinting the hyaluronlc acid component of
normal and pathological synovlal flulds. Clin. Êhim. Êcta E,
902-989.
ERntRno, P., l'loutroFF, Ê.8. and l'lnr¡gnmf,H, D. ( lgEZl. Electronmtcroscopy
of the human synorrlal membranc. J. Eell Elol. I'1, 20?-220"
ERnnm, Ê.J. and SnrlmvRlR, J. ( l9Ë5). Protelnases in Jolnt dlseage.
ln: Textbook of Rheumatology, Znd Ed. Kelley' Ld.N., llanris,
E.0., Ruddy, S. and Sledgê, 8.8., (Edltors) pp lEZ- 192. hJ.E.
Saundrns Eompanyr Phlladalphla.
BRntoto, P.H., lrltEextn, 0.1¡J. and TxonRno, J.E. ( lgEZ). Holaculan r.lelght
estlmatlon of suJfated glycosamfnogþcans in human gingirae.
Eonn. Tlss. Res. 9, 165- l?2.

r7 3
Enntoto, P.l*|., lrJteertr, O.[rJ. and Tror'rnno, J.Ê. ( l9f.1a). The effect of
o,(ygËn derired frce radlcals on glngirzal proteoglycans and
hyaluronlc acld. J. Periodont. Res. 19, 390-400,
ERmoto, P.l*|., l,rJttertn, 0.trJ. and TttonRno, J.E. ( lgB4b). Eeharlor of
hyaluronlc acld from glnglral epithelium and conncctlre tissue
on thç analytlcal ultracentrlfuge. CoÍln. ïr!, Res. 12r 25?-
2Ê4.
ERtEs, EJ., LounxEn, D.Ê. and llnnoLEy, C,J. ( l9B4). Exygen fnee-
radlcals medlatc an lnhibitlon of pnoteoglycan synthesis in
culturcd artlcular cartilage" Élnn. Rheum. Dis. 45, {62-469.
BRxnn, E., FRESER, J.R.E. and l'lnnusr 6.5. ( l9?5). Fractionation and
recovÇry of secretions of synorzial cells synthesized in culture
¡¡lth radioactire precursors. Ênn" Rheum. Dis. 52, 35-{0.
BgRucttRnp, E. and Fntoourctt, l. (19?0). Ê mechanlsm fon the
production of ethylene from methional: the generation of
hydroxyl radlcal by xanthine oxidasa. J. Elol. Ehem. 245,
.lEl l -+E{8.
Eeem, K.11., R¡crnnosoÌr, D.C. and Ra.rn60pf,Lnn, R.V, (13771. lletal Eltes of
coppËp-zlnc superoxide dlsrflutast. Eiochc'rnistry 18, 1950-
I gEE.
Etrnm, R.l'î. ( 197?). llaematologlcal changes ln rheumatoid diseasc.
Elln. Rheumàt. Dls. 5, {55-{65.
BEnrnen, L.E. and Hnnsrntt, L.E. ( l9E+). The hlstory of oxygenic
concentratlon in the earth's atmosphare. Faraday Discuss.
Ehem. Soc. 5?, I 12.
Ems, lrJ.lJ. and EtetRro, L.6. ( lgÊZ). Effect of metal chelators and
antiinflammatory drugs on the degradation of hyaluronic
acld. Êrthritis Rheum. 25, l'lE9- 1'l?8.

L74
Elette, Ê., ÊNoEnsson, T. and GnnnfrTr, K. ( lg82). I'lolecular r.leight
distribution of hyaluronic acld of human synorial fluid in
rheumatic diseases. Scand. J. Rheumatol. l?, ISS- lSE.
EtRre, 0.R., llRLL, N.D., Tneey, D.Ê., llRLLil¡JELL, B. and Gunen¡oee, J.l1.E.
( l9Ë l). Protection against superoxide and hydnogen peroxide
in s¡rnorlal fluid fnom rheumatoid patients" Clfn" scl" 61, 485-
{Ë6.
EtRre, 0.R., ELßÌ{il, É1., ERcott, P.Ê., Ffinn, ll., 6ulrERtD6E, J"H.E. and
HRU¡r¡eu, E" ( 1985a). Femoxidase and ascorbate oxidaEe
actitzitles of ceruloplasmin ln synovial fluid frsm rheumatoid
patients. Clln. Scl. 84, 551-553.
BtRt<t, 0.R., llfrLL, ll.D., ERcott, P.4., DtEppE, P.Ê., l-lnll¡l¡et¡., B. and
[ìunrn¡oee, J.Fl.C. ( lg8rb). Effect of a specific tron chelating
agent on animal models of inflammation. Ênn, Rheum. Dis. 12,
Eg-95.
EtRre, D.R., BßLLfiGHEn, PJ., P0TÍER, 4.R., Egtt, l1J. and EficoN, p.Ê.
( l9E4). The effect of synovtal inon on the progresEion of
rheumatoid disease. Êrthritis Rheum. 27, lgí-581.Etumeenc, 8.5, and 0esror, Ê.8. ( 195?). The effect of proteolyflc
Ënzymes on the hyaluronic acld complex of ox synorrial fluid.
Biochem. J. 66, 5'12-5+8.
EtumsnxnnnTz, N. and Êsaoe-llRnsEn, G. ( l9?5). Ner¡ method for
quantitatire detenmlnatlon of uronic acids. Ênalyt. Eiochem.
54' 'l8'î-'lBg'Eonngc¡Rno, N. and KnResarre, K. ( l9E0). Role of oxygert in anfibody
depandent cytotoxicity medlated by monocytes and
nautrophils. J. Cl¡n. lnrcst. 66, E?6-685.
Ennnot, K.D. ( l9?.1). The effect of synorzlal hyaluronate on the
ingestion of monosodium urate crystals by leucocytes. Clin.
Ehim. Êcta 55, 30?-315.

rz5
Ënnun, K. and Fntooutcx, l. ( l9B l). DNÊ strand scisslon by enz¡,rmlcalty
generated oxygen radicals. Êrch. Eiochem. Elophys. 206,
{ 1.1-419.
En¡mRcot"tgE, J.5" and [rJeeeEn, J.11. ( l96rî). I'lyaluronic acid. ln:
l"lucopolysaccharldeg: chemical structure, dlstribuflon and
lsolatlonn Ehap. 3, pp ,{3-83. Elserler, Êmsterdam.
BntnrmRn, R., Lfim8Enrg, H"8., lrJRoetn J. fitt0 Zuloeueto, J. ( | gE l).
Ëontributions to the study of immediate and early x-îay
reactlons l¡lth regard to chemoprotectlon. l. Further
analysis of the immediate drop in inJection pressure on
moderate irradiation" lnt. J. Radiat. Elol. 5, ZE5-210.
Eucrnnnn, J.D. and Ênmyrnone, D.Í1. (19?E)" The radtolysis of
glyceraldehyde-5-phosphate dehydrogÊnä,se. lnt. J. Radiat.
Blol. 53, 409 labstl.
CRnl, ll. and JRnorr, Ê. ( l9?E). Posstble machanismt of emphysema in
smokers. þ ritno suppression of serum elastase-inhibitory
capaclty by fresh cigarette smoke and lts pnerention by
antloxidants" tlnnu. Rer. Resp. Dis. I lE, 6l?-621.
ERRo, H. and JRnorr, H. ( l9?9). þ r¡ltno supprÈssion of serum
elastase inhlbitory capacity by neactlre oxygen species
gencrated by phagocytosing poþmorphonuclean leukocytes.
J. Clln. lnyest. 65, ?95-?9?.
Efiston, E.hJ., Pntncr, R.K. and HnzEtrott, HJ. ( l96E). l'lyalunonic acid in
human synorial effusions: a sensitire indlcaton of altered
connectlve tlsstte cell functlør durlng lnflammation. Êrthrltis
Rheum. g, ?Ë5-?9+.
EttRltt, E. and Dutnle, E.5. ( 1959). Ê mucolytic enzyme in testio
extracts. Natuna 144, 9??-9?8.

r?6
Etetnno, R.L. ( lg?0)" Holecular r.leight dlstribution in hyaluronic
acid. ln: Chemistry and Holecular Biology of the lntercellular
Ilatrix. Ealazs, E.Ê., (Editon) pp. ?55-?{2. Êcademic" Pness,
London and Ner¡ York.
EtetRno, R.L. and lrJnne, J.L. (19?B). lonic polysaccharides. lll. Dilute
solution pnoperties of hyaluronic acid fractlons. Eiopolymers
g, ?99-ElB.
EtrtRro, R.L., STo0LMILLER, Ê.C., R00EH, L, and LnunEHT, T.C. ( 1963).
Partial characterlzatlon of rcaction products formed by the
degradatlon of hyaluronic acid r¡ith ascorbic acid. Elochim.
Blophys. Êcta 192, 585-59{.
Cottt"tRtt, J.P., EEfiuHfrH, J"1., llRteenn, T.R. and Susucx, K.S. (19?Ê).
Hature of E" and C0 bfnding to metalloporphyrins and hc'rne
proteins" Proc. l{atl. Êcad. Scl. U5Ê 13r,3333-3337.
CumRtr, C.C. ( 1955). The nature of the protein in the hyaluronic
complex of borine synovial fluid. Eiochem. J. 61, EÊE-E9E.
DRttt, L., DfiHL, 1.11.5., Engsrno¡r-Launert, Ê. and 6nRrRTr, K. ( lgBS).
Concentration and molecular r.leight of sodium hyaluronate ín
synorial fluid from patients r¡ith rheumatoid arthritis and
other arthropathies. Ênn. Rheum. Dls. 11, B|7-BZ?,
DRs, 3.K. and NRtn, R.t. ( l9A0). Superoxide dismutase, glutathione
peroxidase, catalase and lipid peroxidation of normal and
slckled erythrocytes. Erit" J. Haematöt 11, A?-92.
DglssenoTx, Ê. and Dounce, Ê.L. ('19?0). Eatalasez physlcal and
chemical properties, m;chanisms of catalysls and physiological
role. Physiol. Rer. 50, 319-5?5.
DeL llnesrn0, R.F., T}lRtt, H.H., EJoRK, J,, PtRnren, 11. and ÊnFons, K.E.
( l9E0). Free radicals as medlators of tlssue inJury. Êcta
Physiol. Scand. suppl. 492, {3-5?.

L77
Enrrssott, S., FRf,sEß, J.tl.E., LRunent, T.8., PERToFT, 11. and 3meo8R00, E.
( l9E5). Endothellal cells ârË â slte of uptake and
degradation of hyaluronic acld in the llren. Exp. Eell Res.
111, 223-224.
FRtcttut<, K.11., EoETaL, EJ, and Kulrf,, J.P. (19?0). Rerpiratary gaset
of synorrlal flulds. Êmep. J. Hed. '13, 225-U 51.
FEE, J.Ê. and PnleNTHE, J"3. ( lg??). Êhemlcal and physlcal properties
of superoxlde. ln: Superoxide and Superoxide Dismutases.
lllchelson, É1.H., llcEord, J. and Fridorich, l. (Editors) pp l9-80.
Êcademlc Press, London.
Fennntm, Ê. and Tnone, Y.H. ( I g?B). Ê napld one-step procedure for
purlficatlon of mononuclear and polymonphonuclear
leucocytes from human blood using a modlflcation of the
Hypaque-Flcoll technlque. J" lmmunol. llethods 21, 589.
Ftssten, J.ll., 0csroÌ{f Ê.8. and SrnntEnr J.E. ( 195'{). Some propenties of
human and other s¡rnorlal fluids. Elochtm. J. 58, 658-680.
Flscuen, E. ( lE90). Synthesan in der zuckcrgruppÊ. Eerichte der
dautcshen chemischen geselschaft. 25' 2l l{-21+ l.
Ftscmen, L. ( l9E0). Eel flltratlon chromatography. ln: Laboratory
Techniquas in Eiochemistry and Holecular Eiolog¡1. lrlork, T.S.
and Eurdon, R.ll. (Eeneral Edltors). Elserrien/North llolland
Elomedlcal Press, Êmstendam, Nerl Yonk, and 0xfond.
Ftnmm, E.5., DEM0pouL0st ll.E., SEL|EMRNr fí.L., Posen, R.6. and RRNsotoFr, J.
( I g?E). Fnee radicals ln cerebral ischaemla. Stroke 9, '1'{5-
44?.
Ftrrun, hl., RouLEy, D.Ê. and Hnl¡.tt'rELL, E. ( l9Ë5). Superoxide-
dependent formation of hydnoxyl radlcals in the presence of
lron salts. hJhat is the physiological lron chelator? FEBS
Latt. l5A, 510-31?.

t78
FtoYo, R.Ê. ( lgES). Direct damonstratlon that fernous ion
complexes of di- and triphosphate nucleotides catalyze
hydroxyl fnee nadical formation fnom hydrogen peroxide.
Êrch, Eiochem. Blophys. 225, 263-2?0.
FLynn, J.P., ÊLLEN, D.hl.r Jottttsott, 6J. and hlxre, J.6. ( lgES). 0xidant
damage of the llpids and protelns of the erythrocyte
membranes ln unstable hemoglobin dlsease. Eridence for the
role of llpld penoxidatlon. J. Elln. lnrest. ? l, lZ 15- l?25.
Fomr, C.3. (lg?9). Detectlon of slnglet olrygÉ'n in complex systems:
a critique. lnz Exygen: Eiochemical and Clinical Êspects.
Caughey, hJ.S" (Ed¡tor) p 805" Êcademic Press, l{eu York.
Fonnesren, J.7. and BntRzs, E"Ê. (1980)" lnhlbitlon of phagocytosis by
high molecular rrelght hyaluronatÊ. lmmunologg {0' 435-4{6"
Fonnesnn, J.P. and lrl¡t-xtxsolt, P.C. ( lgB I ). lnhibitlon of lucocyte
locomotion by hyaluronlc acld. J. Eell Sci. 48, 515-551.
FoN, Ê.1d. and Êuron, Ê.P. ( l9?E). guperoxide anlon mediated toxicity
in isolated lung cell preparatlons. flnn. Rer. Resp. -Dls. I 11,
535? [abst].
FnRnr,"L. and J{RssRgo, D. ( lg8tl). Exygen toxlcity. Élmer. J. lled. 89,
I r?-r26.
FnRrrtt, E.N. (19Ë0). Llpld oxidatlon. Progr, Lipld Rcs. 19, l-Z?.
Fnnsen, J.R.E., Etannts, BJ. and ERrtel, E. ( l9?g). Pattarns of induced
rarlatlon in the morpholo1!, hyaluronlc acld secretion and
lysosomal erzyme actlvlty of cultured human synovial cells.
Ênn. Rheum. Dii. 38, ?A7-291.
Fnnsen, J.R.E. and llcERtt , J.F, ( l9E5). Culture of synovlal cells iÐ
Eil¡e. Ênn. Rheum. Dis. 2'1, 551-559.

1?9
FnRsen, J.R.E., LRunem, T.8., PEmoFT, 11. and Enxrep, E. ( l9E l), Plagma
clcarance, tlrrua dlstributlon and metabollgm of hyaluronic
acid lnjected intrarenously in the nabbit. Eiochem. J. 200,
.115-{2+.
Fntooutcr, l" (19?0). 0uantltatlrç aspects of thc productlon of
superoxlde anlon radlcal by mllk xanthine oxidase. J. Elol.
Chem. ?15, 1853-'105?.
Fuooulctt, l. ( l9?5), Superoxlde dlgmutasçs. Annu, Rer. Elochem. 44,
l.î?- 159.
Fntooulctt, l. (19?E). 0xygen radicals, hydrogen peroxlde and oxygen
toxiclty. ln: Free radicals in biology, Vol. l. Pryor, Lil.H.
(Ed¡tor) pp 239-217. Êcadcmlc Press, lJer¡ York.
Fntooulctt, l. (lg?E). ThÇ biology of oxygm radlcals. Sclc,nce 201,
8?5-Ë80.
Fntooutctt, l. (19ÊZ). Supcroxlde dlgmutage ln blology and mcdlc¡ne.
ln; Pathology of 0xygen. Êutor, Ê, (Edltor) pp l - 19.
Êcademlc Prcsr, Hcr¡ York.
Fnor¡Rr¡or, 11.P. ( l9??). Êdsorptlon of human erythrocyte surf ace
glycoprotein on to polystyrene latex rpheres. J. Lab. Elin.
lled. 98, 321.
ERnonsn, T.J., SrEurßFT, J.R., ERsRte, Ê.5., DobIHEy, J.11. and CtlnmBEns, D.E.
( l9A5). Reduction of myocardial ischaemic injury r¡ith
oxygen-derived free radlcal scarengers. Surgery 31, 123-
1?7.
EnnotRtty, F.N. ( l9?E). Fine structure of joints. ln: The Joints and
Synorzlal Fluld, Pol. l. (sokoloff, L., Editon) pp 105- l?6.
Êcademlc Pness, Ner¡ Yonk.
6roas, D.Ê., HERRILL, E.hl. and Smm¡ K.Ê. ( l gEE). Rheology of
hyaluronic aÊld. Blopolymers Ë, ???-?9 t.

t80
Botoeene, E. and Srunn, A. ( l9??). Thc role of thc ruperoxide anlon
ås å toxic species ln the enythrocyte. Êrch. Biochem"
Elophys. l?8, 2l E-225.
Gotoeenc, R.L. and Toote, E.P. ( l9E4). Hyaluronate coat formation
and cell spreading ln rat flbrosarcoma cells. E*p. Ëell Res.
l5l, z5B-zâ3.
Gotomern, 1.H., KfipLfir, H.8., Eoeusolt, H.S" and ldg¡ssrtRrtt, E. (19?9).
Ceruloplasmin: a scarÊnger of superoxide anion radicalE. J.
Elol. Ehem. 231, 1E1O-1O13"
6nRncrn, 0.N., RuïLt, 6. and HcCono, J. ( I gB I ). Superoxide radicals in
feline intestlnal ischaemia. Gastroenterology E I, 22-29.
Gnee¡rt¡Rro, R.Ê" ( l9Ê I ). The role of oxygen-derived fnee radicals
(0DFR) in connectlve tissut degradation" lll. Studles on
hyaluronic acld depolymerlzatlon in lnflamed synorrial fluids.
Semin. Êrthnitis Rheum. I l(suppl. l), 9?.
Eng¡nl¡Rt o, R.Ê. and Honx, S.Ê. ( l9E4). Degradatlon of hyaluronlc acid
by neutrophils. lnflammation Research Êssoclation.
Therapeutic control of lnflanmatory di¡eages. ll. Êbst.
papers, p.21.
EneentJnlo, R.É1. and HoY, h,,hJ. (l9?9). lnhibition of collagen gelation
by the actlon of the superoxide radlcal. Arthnitls Rheum. 22,
25 I -259.
GnrenuRlo, R.Ê. and l-10Y, hJ.kI. ( 1980). Effect of orygen-derived free
radlcals on hyalunonic acid. Êrthritls Rheum. U5, '155-465.
Gn¡gnurRr-o, R.Fl., l1oy, lrl.lrl. and LRzRnus, D. ( l9?6). Degradation of
cartilage proteoglycans and collagen by supenoxide radlcal
[Êbstract]. Êrthritls Rheum. 19, ?99.

t8l
6ntrnn, F.11., BtRnco, E" and Srt pgnsrulr, S.E. ( l9?5). Characterization of
the macrophage receptor for complemert and demonEtration
of its functlonal independence from the receptor for the Fc
portlon of immunoglobulln B. J. Exp. Med. 111, 1269-12?7.
EnossrsLo, ll" (1958). Studies on production of hyaluronic acid in
tissue culture: the presence of hyaluronidase in embryo
extract. Exp. Cell Res. 14, 213-218.
6non, l'f.P. ( I g?5). Cellular contacts in tha synovlal membrane of
the cat and rabbit: an ultnastructural study. Eell Tlssue
RËs. 16'1, 525-5{ l.
Burrgnoc¡, J.fí.E", Rrcrmoro, R. and llRl¡.n¡ell, E. ( l9?9). lnhlbltion of
iron-catalyzed formatlon of hydroxyl radicals from
superoxide and of lipid peroxidatiøt by desfercio,ramine.
Eiochem. J. lE4, 469-1?2.
[ìurrenroee, J.11.8., Routey, D.Ê. and I'lRlt ¡l¡ell, E. ( l9E I ). Superoxide-
dependent formation of hydroxyl-radicals in the presence of
lron salts. Blochem. J. 199, 283-265.
6un¡n¡oee, J.l'î.C., Routey, D.Ê. and HRt-r"n¡¡r-t, B. ( l gEZ). Superoxide-
dependent fonmatlon of hydroxyl radlcals and lipid
peroxdation in the presence of iron salts. Detection of
"catalytic" lron and antloxidant actlvity in extracellular
flulds. Eiochem. J. ?OÊ, 805-609.
[ìungn¡oa¡, J.Ì1.C., Pmuneon, 5.K., SEEf,L, Ê.h1. and HnlltLrELL, E. ( l98l ).
lnhibition of lipid peroxidatlon by thc lron bindlng protein
lactoferrin. Elochem. J. l99, Z5g-ZÊ1.
llReen, F. and l,rlutss, J. ( | 934). The catalytlc decompoEition of
hydrogen peroxide by iron salts. Proc. Royal Soc. London,
Ser. Ê l+7,332-351.
llRoten, N.l'!. (19Êl). Enhanced dlffusionity of glucose in a matrix of
hyaluronic acid. J. Eiol. Chem. 255, 5552-5555.

182
llRoten, N.l{. and NRlttn, H.Ê. ( l9??). Structure of hyaluronlc acid in
synorzlal flu¡d and lts lnfluence on the mopÊmÊnt of solutes.
Semin. Êrth. Rheum. ?, l4l- 152.
HRotrn, N.J1., DounmRsrruH, R.R., Nenmur, H.P. and lrJttttRms, L.D" ( ßAZ),
Ultrastructure of a hyaluronic acid matrix" Proc. Ì{atl. Ãcad.
Sci. U5Ê ?9, 50?-509.
llRrnnssott, L. and Punce, P. ( l9Ê5). The combined action of
hyaluronic acid and fibronectfn stimulates neutrophil
migration. J. lmmunol. 135, 2?55-U?59.
HnrRnssotr, L., llÊL[6pEN, R. and Psner, P. ( | 980). Regulation of
granulocyte function by hyaluronic acid. þ r¡itro and þ riroeffacts on phagocytosls, locomotion and metabolism. J. Cl¡n.
lnrçst. 86, 298-505.
HRH, R. ( tgES). Ên lmprored nutnlent solution fon dlploid chlnese
hamsten and human cell llnts. Exptl. Cell Res. 2S, 515-526.
HRt"usoN, I and 6Rt tRengn, J. ( lgES). Separatlon of nadiolabelled
glycosamtnoglycan ollgosaccharldes by polyacnylamide gel
electrophoresis. Biochem. J. 2?l, E9?-?05.
f'lRul¡¡lt , E. ( l9?E). Superoxide-dapendent fomnation of hydroxyl
radicals in the prÊience of iron salts. FEES Lett. gE, Z3B-
21?,.
f'lnLL¡ueLL, B. (l98l). Free radicals, oìrygen toxiclty and aging. ln:
Êge Pigments. Sobel, R.5. (Editor) pp l-EZ, ElsevlerlNorth
Holland Elomedical Press, Êmsterdam.
l'lRttn¡Etr., E. and EunenloeE, J.J"l.C. ( I gË4a). Oxygen toxicityr oxygen
radlcals, transitlon metals and disease. Eiochem. J. 213' I -14.
llntttuletr, E. and Gurren¡oc¡, J.l'l.C. ( l9Ê4b). Llpid peroxidatlon, ot4ygen
nadlcals, call damage and antloxldant tharapy. Lancet
i:1596- 159?.

183
Hnlu¡¡gtt, E. and Eunen¡gee, J.ll.E. ( l9E5)" Free Radicals in Ëlology
and lledlcine. Elanendon Pness, Oxfond.
l'lnnnts, E.D, ( l9Ë5). Pathogenesis of rheumatold arthnitis. ln:
Textbook of Rheumatologyr ãnd Ed. Kelley, hJ.¡|., llarrls, E.0.,
Ruddy, 5. and Sledge, C.E. (Editors) pp EË6-9 t5. ld.E.
Saunders Eompany' Phllädelphla.
llRnnts, 11.J., HEnP, Ê. and PlemRn, hJ. ( 19? I ). Depolymerlzation of
polysaccharldas thnough the gcneratlon of free nadlcals at aplatlnum surface: a norel procedure for the controlled
productlon of fnee-radlcEl oxidatlons. Êrch. E¡ochem. Biophys.
l{2,615-62U.
llRnntsott, J.E. and Scruttz, J. ( | 9?Ë). Studlcs on the chlorinating
actlrlty of myeloperoxldasa. J. Elol. Cham. 251' l3? l - 15?{.
lleys, Ê.0. and DonmRnoY, T.L. ( l9E I ). Llpld peroxidatlon ln lron-
orerloaded spleens. Clln. Sci. 60' 295-301.
Hormnnn, ll. and Scrmut, 0. ( 1980). The lnabillty of superoxide
dismutase to inhiblt the depolymerizatlon of hyaluronic acid
by ferrous ions and ascorbate. [ìnaefes Êrch. Kl¡n.
0phthalmol. 211, I I I - I Ê5.
l{otmes, 8., Pf,nK, 8.H., l'lfiLlsrfr, S.E.r 0ul, P.G., Netsott, D.L" and 6000, R.Ê.
( I g?0). Chronlc granulomatous dlscage ln fc'males: a
deflclcncy af laucocytc alutathione peroxldase. l{crrl Engl. J.
Hed. 283, Zl?-Z?1,
lloucr, J.E. and PeRncl, R.ll. ( 195?). The mcchanlsm of hyaluronida¡e
actlon. Biochlm. Elophys. Êcta 25' 555-58?.
l6Rnr, T., Kf,HEDR, H., l'lontucttt,5. and 0no, S. (lgBU). Ê rtmarkable in-
crËase of superoxlde dlsmutase actlvlty in synovial fluid of
patlents uith rheumatoid arthritls. Clln. 0rthop. Rel. Res.
1ã?, ZB?-2Ë?.

IE4
IutrRtn, 5. and EnRce, J.T. (1981) Eultiratlon l¡ vitro of myelobla¡tE
from human leukaemia. N. Y" J, Jlad. 64' 22?9-2282.
lyER, 6.Y.N., lsLRH, H.F. and 0unsrct, J.ll. (lgEl). Elochemical aspects
of phagocytosis. Nature (London) 192' 535-541.
JRsttt, t1.E. (l9EZ). Eeneratlon of lgG aggregates by the
myeloperoxldase-hydrogen pcroxldc systcrn. Êrthritls Rheum.
Z5(suppl), 53 [abst].
JeRrtOZ, R.bJ" and Foncrettt, E" ( l95l). Studles on hyaluronic acld and
related substances. lP. Periodate oxidation. J. Eiol. Ehem.
190, 55?-5'+6.
Jrnsen, C.E" ( 1949). The influence of metalllc lons on the vlscosity of
hyaluronic acid solutions. Êcta Chem. Scand" 5' 5E{-588.
Jonnson, D. and TnRuts, J. ( l9?9)" The oxldatire tnactlratlon of
human a- | -protetnase lnhlbltor. J. Elol. Chtm. Z3+, 1DZ2-
{0?Ë,
Jornsror, J.P. (1955). Ê comparlson of the propcrtl¿s of hyaluronic
acid from normal and pathologlcal human synovial fluids.
Elocham. J. 59, EZE:635.
Jonnsnn, J.P. ( 1955). Tha vlscoslty of normal and pathologlcal
human synorial fluids. Eiochem. J. 59, 635-85?.
Ktete, 8.8., lfcEoRD, J.H. and FnlOolrlcn, l. (t9?0). Superoxide-dlsmutase
from Escherlchia Eoll B. Ê ner¡ manganese-containing
enzyme. J. Elol. Chem. 2{5' 6l ?6-E I E l.
K¡tloee, E.lJ. and Fuooutcr, l. (19?5). Superoxlde, hydrogen peroxide
and singlet oxygen in lipid peroxidation by a xanthine oxidase
system. J. Biol. Chem. 250' 8A lz-Bïn.Kettoee, E.hJ. and Fntooulcr, l. ( 19??). Llposome oxidation and
erythrocyte lysls by enzymlcally generated superoxlde and
hydrogen peroxlde. J. Elol. Ehem. 232, E72l-Ê728.

185
KtttRtR, K., lloNMfi, Y., 0KfiyßMñ, H., 0cunt, K., llozumr, Ì1. and Suzurt, S.
( l9Ë'{). lncreased synthesis of hyaluronic acld by mouse
mammåry carcinoma cell rarlants uith high metastatic
potentlal. Eancer Res. 45, 1311-1354.
KLssRHorr, S.J. (l9E?). lodinatlon of bacterla: a bactericidal
mechanlsm. J. Exp. Hed. l?6, t0E3- l0?8.
Kt¡sRnorr, 5J. ( l9A0). 0xygcn metabollsm and the toxlc properties
of phagotytes. Ann" lntern" HÊd. 93, {Ë0-4Ê9.
Klesnnorr, 5J. and Rosel{, H. ( l9?E). Ethylene formatlon by
pol¡rmorphonuclear elucocytes" Role of myeloperoxidase. J.
Exp" I1ed. l1A, 498-506.
KnRtn, E.ll. and SReore, Ê.L. ( lg8 I ). The effect of oxldant injury on
the lymphocyte membrane and functions. J. Lab" Elin. Hed.
98, 69?-?05.
LReencnRnfz, E. ( l9E't). Free radlcals tn the auto-oxldatlon of
ascorbic acid. Êcta Chem. Scand. lïr SEZ.
Lnunent, T.E. ( 1955)¡ Studles on hyaluronic acld tn the ritreous
body. J. Biol Ehem. 216, 265-2? l.
LRunert, T.C. ( l9?0). Structure of hyaluronic acld. ln: Chemtstry
and llolecular Elology of the lntercellular l'latrlx. Vol. ll,Glycosamlnoglycans and Proteoglycans. Ealazs, E. (Edltor)
pp ?03-?4?. Êcademic Presl, London and l{eu York"
Leunem, U.E.E. and LRUnEHT, T.C. ( lg8 l). 0n the orlgin of hyaluronate
in the blood. Elochemistry Internatlonal 2, 195- 199.
LRunem, U.8.6. and TeneBLRo, Ê. ( lgË0). Determinatlon of hyaluronate
ln biological samples by a speclflc radloassay technique.
Ênalyt. Eiochem. 109, 586-394.
Lenntrern, Ê.L. (19?5). Blochemistry'. The llolecular Easis of Ëell
Structure and Function, Znd Ed. lrJorth Publishers, lnc.,
Ner¡ York.

r86
LtN, lrl.5. and ÊnusrRoNc, D.Ê. ( lg?E). Effects of supenoxide
dlsmutasa, dithiothreltol and formate lon on the lnactiyatlon
of papaln by the hydroxyl and superoxide radicals in aerated
solutions. lnt. J. Radiat. Eiol. 55, 251-23{.
LrnxEn, Ê., HEyEn, K. and Horrunn, D. ( l95E). The production of
unsaturated uronldes by bacterlal hyaluronldases. J. Elol.
Ehem. Zl9, 15-25.
Lrnren, Ê., l.lEyEn, K. and l,rJerssmRn E. ( 1955). Enzryatlc formation of
monosaccharides from hyaluronate. J. Biol. Chem. Zl3,237-
Z,IE.
Lonc, C.Ê. and Etetsrt, E.l'lJ. ( l9Ë0). Rate of reaction of superoxide
radlcal '"ulth
chlorlde-contalnlng specles. J. Phys. Ehem. E{,
555"
LottcRs, H.0. and Heyen, K. (l3A 1)" Sequential hydrolysls oî
hyalunonata by p-glucuronldase and p-N-acetyl
hexosamlnldase. Blochem. J. l9?, 2?5-242.
Luno-0lesEN, K. ( l9?0). 0xygen tenslon in synorial fluids. Êrthritis
Rheum. 15, ?69-??6.
Lunec, J., HfittoRRNr S.P.r lrJrre, Ê.8. and Donmnnoy, T.L. ( l9E I ). Fnee
radlcal oxidation (peroxidation) products Ìn serum and
synorzial fluld in rheumatoid arthrltis" J. Rheumatol. E, 253-
215.
Lyttctt, R.E. and Fntooutcr, l. ( t 9?E). Effects of superoxide on the
erythrocyta membrane. J. Eiol. Ehem. 255, I ESE- I845.
l'lRnn, T. and Kelutt, D, ( l33E). llaemocuprefn and hepatocuprlen,
coppÊr-proteln compounds oî blood and llrer ln mammals.
Proc. Roy. 3oc., Ser.E:Eiol. Scl. l2E, 305-315.
HRnrtuno, 3.L. ( lgEZ). lluman coppËr-contalnlng superoxlde
dismutase of hlgh molecular rrlelght, Proc' llatl. Êcad. Sci.
u3Ê ?9, ?811-?83Ê.

187
J'1Rnsn, J,M., hltEsxnt, O.hJ., 6RtE, 5., l"lutR, I',l. and Hntnt, R.N. ( l9?g).
Synthesis of sulphated proteoglycans by nheumatoid and
normal synorrial tissue in cultune. Ênn. Rheum. Dis. 58, 166-
r ?0,
llnscrorr, E.Ê. and Elncreunn, E.L. ( l gES). l'{utrltlon and rheumatic
dlseases. ln: Textbook of Rheumätology, ?nd ed. Kelley, lÁ|.H.'
llarrls, E.D., Ruddy, S" and SledgË, 8.8., (Edltors) pp 552-560.
lrl.E" Saundars, Phlladelphla.
HRtsuoR, 1., 0Kn, T., TfiNl6ucHl, N., FunuYRmn, H., KRoRt'tñ, 5., ÊnRsllme, S. and
Hmsuynno, T. ( l9?E). Leucocyte glutathione penoxidase
deflciency in a male patient r¡ith chronic gnanulomatous
dlsease. J. Pedlat. 88, 5A I -583.
llcCRmv, D.J. ( ISES)" S¡morrlal fluld. ln: Êrthrltls and Êllled
Eondltlons, lOth Ed. l'tcEarty, DJ. (Edltor) pp 5a-75. Lea &
Febiger, Philadelphia.
llcCleRn, D. and llRte, C.hJ. ( l9'î0). lluclnase and tlssue Permeability.
Natune l't5, 86?-Ë88.
PlcClenn, D. and llRte, C.ld. ( l9{ I ). Studles on diffusing factons: the
hyaluronldase actirlty of testlcular extracts, bactrial
culture flltrates and other agents that increasa tissue
permaablllty. Blochem. J. 55, l59- I85.
HcEono, J.H. ( l9?4). Free radlcals and inflammatlon: protection of
synorzlal fluld by suparoxide dlsmutase. Science lE5' 529-55t.
ilcEono, J.J"l. and DfiY, E.D. ( l9?E). Superoxlde dependent production
of hydroxyl radlcal catalyzed by iron-EDTÊ complex. FEBS
Lett.86, 133-11?.
f'lcEono, J.11. and Fuooutcr, l.: The reduction of cytochrome C by milk
xanthlne oxidase. J. Blol. Ehem. l9EB; Z'13: 5?55-5?68.

I8E
l'lcNelu, J.D., EHotrI, D.Ë. and SrosEy, J.L. (lgEE). Effect of a high
molacular rrreight hyaluronic acid prËparation on the
stimulation of polymorphonuclear leucocytes. Fed. Proc. {5,
658 [abst].
llcNen, J.0., CLELfiHo, L.G. and E¡ns, hJ.l{. (lg8g). The cytotoxicity of
oxygËn derired fnee radlcals. Êust. N.Z. J" l'led. 15, ZZs
Iabst].
HcNen, J.D., hlrEaKrt, 0.hJ., Ems, hJ.l1. and EuetRno, L.g. ( 19ES).
Depolymerization products of hyaluronic acid after eilposure
to oxygen-derlred free radicals. Ênn. Rheum. Dis. 1+, ?tE-
?89.
lfeao, J.F" ( lg?E). Free radlcal mechanlsms of llpid damage and
consÊquÊrrceg for cellular membraneg. ln: Free Radlcab in
Elology, Vol, l. Pryor, hJ.Ê. (Edltor) pp 5l-EE. Êcademic
Press, Ner¡ York.
lfeven, K. ( | 94?). The biological significance of hyaluronic acid and
hyaluronldase. Physiol. Rev" 2?, 335-359.
Heven, K. and PRtmen, J,hJ. (1934). The polysaccharide of ritreous
humor. J. Biol. Chem. I B?, 829-85'1.
Hrven, K. and Pntmtn, J.lrJ. (1958). tln glycoproteins. ll. The polysac-
charldes of ritneous humon and of umbilical cord. J" Biol.
Chem. I l{, EÊ9-?05.
llEyEn, K., DuBos, R. and St,tyttt, E.N. ( 193?). The hydrolysis of the
polysaccharide acids of vltreous humon of umbillcal cord and
of streptococcus by the autolytic enzyme of pneumococcus.
J. Elol. Chem. I tE, ? l-?8.
Hrxult¡-TnKfrGßKr, Y. and Tools, E.P. ( l9Ë I ). llyaluronate-proteln
complex of rous rarcoma rlrus-transformed chlck embryo
flbroblasts. J. Eiol. Chem. 258, 8'{85-E'tEg.

t8g
Honeotsse, J.8., BRfiouET, P. and BoneL, J.P. (lgBl). 0xygen-free
radlcals as mediators of collage breakage. Êgents Êctions
15, 49-50.
llotmorsse, J.Ê., ERf,ouET, P., Rnnooux, Ê. and Eonet, J.P. ( l9E5). Non-
enzymatlc degradation of acld-soluble calf skln collagen by
superoxide lon: protectlre effect by flavnoids. Elochem,
Pharmac ol. 32, 53-58.
lloneRn, Ê.R., EoNE, R.L. and Eleenr, T.J'|. ( l9?E). The mechanism of DHÊ
strand breakage by ritamln C and superoxide and the
protective roles of catalase and superoxide dismutase.
Nucleic Êcid Res. 3, 1 159- I I {9.
lloven, J"D. and lseett, 11.3. ( I gSE). Determination of reducing sugars
and reduclng end gpoups in polysaccharides by neaction r¡ith
carbon- l {-labelled cyanlde. Ênalyt. Ehem. 50, l9?5- l9?8.
l"lutnotn, K.D. ( 1966). Ferritin in synorlal calls in patients r¡ith
rheumatoid arthrltls. Ênn. Rheum. Dls. 25, 3E?-401.
Hutnoen, K.D. and Senatonr 6.E. (1968). lron ln the s¡,rnovlal membnane
ln rheumatold arthrltls and othan Joint diseases. Ênn.
Rheum. Dis. 2?, 3E-{8.
Hutttnnx, F. and HutttttRx, 6.L. ( l9Ë5). Tyroslne-tyrosine Eross links in
lgG. Generation by peroxidation and occurnÊnce in RÊ synorrial
fluids" Êrthritis Rheum. ZE(suppl), S lE [abst.].
llyens, L.S. (1980). Free nadical damage of nucleic acids-and their
components: the dlrect absorption of enËrglr. ln: Free
Radicals in Eiology. Pryor, hJ.Ê. (Ed¡tor) pp 95- l l'1. Êcademic
Press, Ner¡ Yonk.
Nm¡le¡-Ror, E., Sunoetno, L. and Jonsson, E. ( 1963). Permeability of the
synorzial membrane to proteins. Êcta Rheum. Scand. 9, 28-
52.

r90
Nrentuls, Ê.hJ. ( l9E I ). l/ltamin E and iron. Nerrr England J. Hed. 504,
t?0-t?r.
I'lltssott, R., PtcK, F.J1. and Bnny, R.C. ( l969). EPR studies on reduction
of oxygen to superoxide by some biochemical system.
Elochim. Eiophys. Êcta 132, l45- l'16.
Nonton, J., UnBfiH, J. and HnnouoRs, Ê. ( | 9BZ). Ê failure to obserre
enhanced diffuslrzlty of glucose in a matrix of hyaluronic acid.
J. B¡ol. Ehem. 25?, 14151- l'{ 135.
0ol¡.yle-Hnnnts, D.J" and FonnfistEn, V.L. (1980). ï¡novial iron depoEition
in osteoarthritls and rheumatoid arthnitis. J. Rheumatol. ?,
30-56"
Ocsron, 3. and SrenmRr, T.F, ( 1959). Degradation of the hyaluronic
acid complax of s¡morzial fluid by protealytic Ënzqet and by
EDTÊ. Eiochem. J. 72' 301-505.
0esron, Ê.8. and Stnnttn, J.E. ( 1950). 0n the state of hyaluronic acid
in s¡rnorrial fluid. Biochem. J. {8, 56'f-5?6.
0cstott, Ê.6. and StRuen, J.E. ( l95lJ. The dimensions of the particle
of hyaluronic acid complex in synorial fluid. Eiochem. J. 49,
5Ê5-590.
Ptnr, J.T. and Jottttsott, ll.J. ( l9'19). Ê submlcrodeterminatlon of
glucose. J. E¡ol, Ehem. I E l, 149- l5l.
PRnrs, D.Ê., BULKLEY, 6.8., Gnnncen, D.ll., HñMILT0N' 5.R. and llcC0nD, J.11.
(l9EZ). lschemlc injury in the cat small lntestine. Role of
superoxide radlcal¡. Ba¡tnoenterology 82' 9- 15.
PrRn¡lnClR-PnOOUCT lnfonmmOn. Eel Filtration - Theory and Practice.
Pharmacia: Laboratory Separation Division, Plscatar.layr Netl
Jersey, U.S.Ê.
Pxtueson, L.ll. and Scru.lRmz, H.E. ( l9B't). Subcellular locallzation of
hyaluronate synthetase in ollgodendroglioma cellc. J. Eiol.
Ehem. ZSg, 5011-5023.

191
Pr6mRn, lrJ. and Rtzut, 5. ( 1959). llyaluronlc acld and the ORD
neactlon. Elochem. Eiophys. Res. Eommun. l, 59-{2.
PleNnn, l,rJ., Rtzut, 5. and Hortty, ll.L. ( l3E l). Depolymenization of
hyaluronic acid by the ORD reaction, Êrthritis Rheum. 4,
u +0-252"
Pnexm, P. ( l3A3). Synthesls of hyaluronate In differerntlated
tcratocarcinoma cells: mechanism of chain gror.r.lth. Eiochem.
J. ?l l, l9l-198.
Pnenm, P. ( | gB{). llyaluronate is synthesized at plasma
membranes. Eiochem. J. 220, 59?-800.
Pnvon, lrJ.Ê. ( l9?E). The role of free radical reactions in biological
systems. ln: Free Radlcals in Eiology, Pol. l. Pnyor, l,rJ.Ê.
(Edltor) pp l-49. Êcademlc Press, Ner¡ Yorì<.
Raottt, E.L., StIRNH, D.Ê. and lrJ¡¡ssEn, P.Ê. ( l9?0). Separation of a
hyaluronate-free lubnicatlng fraction from synorial fluid.
Natune 228, 5??-5?8.
RRottt, E.L., PÊuL, 1.L., SURtttt, D.Ê. and ScxorrsrngDnr E.S. (19?l).
Lubrication of synorial membrane. Ênn. Rheum. Dis. 50, 5e2-
. 3U5"
RReRn, C. and Heyen, K. (1949)" The hyaluronic acid of synovial fluid
in rheumatoid arthrltls. J. Elhl. lnregt. 28, 5Ë-59.
RRcRtt, C., DoHtRu,8.P., Coss, J.Ê. and 6nuetr, Ê.F. (131?r. Effects of x-
ray irradiation on viscosity of synovial fluid. Proc. Soc. Exp.
Eiol. Hed. 68, l?E-l??,
Rtooy, P.S. and Leulls, H.. ( l gEg). The adrerse effect of intr aveno.Js
irõn dextran in rheumatoid arthritls. Êrthritis Rheum. lZ,
454-45?.
R¡HNcnoNR, 5., SrEsJo, E.K. and S¡,lnr, 0.5. ( l9Ë0). Rererslble lschaerynia
of the braln: blochemical factors influencfng restitution.
Êcta Physiol. Scand. {9Z(suppl), 155- l'10.

r92
Retssre, J.L., STR0M¡N6ER' J.L. and Lttotn, L.P. ( 1955)" Ê modlfled
colorlmatric mathod for the estimatlon of N-acetylamino
sugars. J. Eiol. Ehem. Z l?' 959-986.
Rtcnent, D.Ê. and lrJesr¡nrtelû, l,lJ.U. ( tgS l). Xanthine oxldases in
different species. Proc. Soc. Exp. E¡ol. Hed. ?Ê,252-251.
RoseffisoN, hJ.E., Rotes, H.hJ. and Eauen, hl. ( 1953). Enzymatlc
hydrolysls of mucln by a bacterlal enzyme and by t¡itamin E.
Êmer, J. Physlol" l2E, 609 labst.l
RoggntTsoÌ{, hJ.E., Role s, l1.lJ. and ERuen, Ld. ( | 9'10). Huclnage: a
bactçrial enzyms ¡¡hlch hydrolysês synÊvial fluid mucln and
other muclns. J. Blol. Chem. l3,3' ?E|-Z?E.
Rogeprson, hJ.B., Roles, l1.hl, and"Enuen, hJ. (19'îl)' Thc degradation of
mucins and polysaccharides by ascorbic acld and hydrogen
paroxida. Elochem. J. 55' 9CI5-908.
Roelnson, H.E. and Hottloou, J.J. (l9?5). The alkallne clearage and
borohydnlde reductlon of cantllaga proteogþcen. Biochem. J.
155, ,{5?-+?0.
RoonRn, 6.P., EENEDEK, T.6. and Pnnetrf,, hl.E. (1386). The early history
of synorzial fuoint) fluid. Ênn. Intern. lled. 65' E? I -E{2.
RonRt, P.Ì1. ( lgEg). The elutlon of t'Cr from labellcd leucocytes - a
nËuJ ässây. Blood 53' 408-{ 15.
Root, R.K. and lletcRtF, J.Ê. ( l9??). H"O, release from human
. granulocytrr durfng phagocytoslg: rclatlonghlp to rup+oxldc
anlon formatlon and cellular metabollsm of llr9rt studles r¡lth
normal and cytochalasln E trrattd crlls. J. Clln. lnrest. 80,
r 286- I 2?9.
Rores, lf.hl. and Enuen, U. ( 1953). Synovial Fluid Changes in Joint
Dlsease. llarrard Unlrerslty Press, Eambrldge, HÊ,

r9g
Roser, E.H. and KLegnnorr, 5.J. ( lg??). Formation of singlet otrygen
by thc myeloperoxldase-medlated antlmlcroblal system. J.
Blol. Chem. 252, {805-'1810.
Rosttt, 6.J",l., BRneen, 11.J. and RRucxmnn, E.J. ( l9E5). Disnuptlon of
crythrocyte membranal onganizatlon by superoxide. J. Eiol.
Ehem. ?58, ZZ?5-2ZZË.
Rosenrrat, H.8., EEnnev, J.P. and Êtetn, E.E. ( 19'lE). Ê sensitlre auto-
mated procedure for the detenmination of glucuronic acid.
Conn. Tiss. Res. 4, 155- lE t"
Roltto, 8., Ënfiy, R.E. and Flrtoen, E.H. ( l9?2). Ê pulse radiolysis
study of superoxide dismutase. Eiochim. Eiophys. Êcta 268'
EB5.EBg.
SRRnnt, ll., TrMMr, H., [/u0nt0, E. and PerrnHEH, R" ( l9E0). Distribution of
glycosamtnoglycans in nheumatold cultures and effects of
cortlsol on lt. Scand. J. Rheumatol. 9, I l- 16.
Sncrs, T., l{0L00tJ ) C.F., CnRooocr, P.R., EobJERsr T.L. and JRcoB, l'1.S.
( t 9?E). 0xygelr radlcals mediate endothellal cell damage by
complement-Etimulated granulocytesz an in vltro model of
immune vascular damage. J. Elin. lnrest. E l, 1 lE I - I lE?.
Snun, J1.L. and llcEono, J.H. ( l9?5). Protection of phagocytosing
leukocytes by suparoxide dismutase. J. Clin. lnrest" 56,
t5l9-1525.
SRto, l. and NlttR, Y. The oxygen radical scarenging actlrzlty of
hyaluronic acid and its comPonÊnt sugars, in press.
SeRnnn, Ê.J. and KRnnousrv, 11.L. ( 1959). The biochemical basis of
phagocy-tosls. l. Hetaabolic changes during the lngestion of
particles by polymorphonuclear leucocytes. J. Biol. Ehem'
231, I 355- l1EZ.

194
Scronenum, G.R. and ExnncE, E. ( l9?E). Êatalase. ln: The Enzymei,
5rd Ed. Eoyer, P.D. (Edltor) Pol. 15, pp 365-459. Êcademic
Press, Ner¡ York.
ScrnRursrRTTER, 1.U., lilNsHfirr¡, 0.8., l'lystoP, P.Ê., SPnßGE, R.6. and CocrRRNE,
Ë.6. ( l9E5). Elutathlone cycle actlrrlty and pyrldine nucleotide
lerels ln oxldant-induced inJury of cells. J. Elln. lnvest, ?Ê,
I l5l-l t39.
Scor, J.E. ( 1980). Allphatlc ammonlum salts ln the a;.say of acidlc
polysaccharldes from tlssucg. Methods Elochern. Ãnalysis Ê,
I 45- r 9?.
Scorr, J.E. and Pnag-Tto¡1fis, D.P.P. ( l9?E). Spectrofluorimetric
detectlon and measurement of hydroxyl radicalc in periodate
solutlon. Earbohydrata Ras, 52, Z l4-? I E.
Scotr, J.E. and Ttcuett, HJ. ( lg?3). Periodate-induced riscosity
decreases in aquÊous solutions of acetal and ether linked
polymers. Êarbohydrate Res. 28, 53-59.
Scorf, J.E., TTcLIELL, ll.J. and Sn.lo¡nfi, UJ. (1372). Loss of basophilic
(sulphated) matenial from sections of cartilage treated r¡ith
periodate solutlon. l'listochemlcal J. 4, t55- 16?.
Scon, J.E., HEÊTLEv, F., Hoogcnorr, D, and 0tRuesen, ll. ( | 9E l).
Secondary structures of hyaluronate and chondroitin
sulphate. a lH n.m.r. study of NH slgnals in dimÇthylsulphoside
solutlons. Elochem. J. 199' 829-432.
Scuooen, P.R., ÊL-Ttmtmt, D., HcHunnfiY, lrJ., hlnm, A,8.,2008, E.E. and
Donmanov, T.L; ( l9?A). Serum coppÊr and rçlated rarlables in
rheumatoid arthrltls. Ênn. Rheum. Dis. 31, E?-79.
5e ent, Ê.lrJ. ( l9?'{). Nltroblue-tetrazolium tests. Lancet il, IUA-
1252.

r95
SerRRtR, P.0., KfinKKRTNENT J., LEHToNEN, Ê. and llRxtsñRfi, P. ( lg?Z).
Ehondroitln sulphate in normal and rheumatoid synorzial fluid.
Elln. Ehim. Êcta 58, 549-555.
SreexRn, J.K., ÊTKrNs, E.D.T. and Nl¡ouszYNsKl' T.A. ( l9?5). X-ray
diffraction of polysaccharldes. Tr¡o-dimensional packing
schemes for threefold hyalunonic chalns. J. Hol. Elol. 3l ' 155-
I 83.
SrEs, ll. and Eaoenfis, E. ( t 983). Biological basis of detoxlcation of
oxygen free radlcals. ln: Biological basis of detoxication.
Caldrr.rell, J. and Jakoby, hJ.E. (Edltors) pp lBl-Zl l. Êcademic
Press, San Dlego.
SnpnnRrnR, P", DuNsToHE, J.R. and 0egfor, Ê.8. ( l9E?). Fractionation of
a hyaluronic acld prÉparätion in a dansity gradlent: the
lsolation and ldentlflcatlon of a chondroitln sulphate.
Blochem. J. l0+, {0{-409.
SrmcrotJrrz, L. and Selteeno, l. (19?9). Evidence for the role of
superoxide radicals in neutrophil-medlated cytotoxicity.
lmmunologg 5?, 301-509.
Srt-lon, R.ll., ScoGGtH, E.l.l. and Pnrrglsot{, B. ( l9E l). llydrogn peroxlde
cäuses fatal inJury to human fibroblasts exposed to oxygen
radlcals. J. Biol. Chem. 256' ? lE l-? lEË.
SrRnse, E. and SunoetRo, L. ( l9'{5). Oxidatlre bneakdor¡n of
hyaluronlc and chondroitln sulphurlc acld. Êcta Physiol. Scand.
E,5?-51.
Smeosnoo, 8., PEnroFT, l''l., Enlrssott, 5., FRfrsEn, J.R.E. and LnunENTr T.C.
( l9E'1). gtudles þ ritro on the uptake and degradatlon of
sodlum hyaluronate in rat lirer endothellal cells. Elochem. J.
2?1, El?-Ê?E.

196
Sooen, P.0., NoRD, C.E., LuttoBuRo, 6. and K,leutmRn, 0. ( l9?0J.
lnrzestigations on lysozgffie, protease and hyalunonidase
actirzity in extracts from human leucocytes. Êcta Ehem.
Scand.2{, l29-158.
Sroou¡,tu.gn, Ê.C. and DonrMfiN, Ê. ( l9?0). The bios¡mthesis of
hyaluronlc acld ln group Ê streptococcl. ln: Chemlstny and
Holecular Elology of the lntercellular Hatrlx, Pol. 2. Ealazs'
E.Ê. (Edltor) pp 7A3-791. Êcademlc Press, Het¡ York.
SunoatRo, L. ( lg53). Studles on hyaluronic acid in synovial fluids.
Êcta Soc. Hed. Upsalla 58, I 15-256.
SunoeuRo, L. and ERtRzs, E.Ê. ( lgËE). Ehemical and physical changes of
glycosamlnoglycans and glycoprotelns caused by oxidation-
rcducction systems and radlatlon. ln: The Amino Sugars, Pol.
28. l'letabolism and lntenactions. Balazs, E.fl. and Jeanloz,
R.hl. (Edltors) pp 229-250. Êcademic Press, Neu York and
London.
Swen, E.R. and lln.lno, 6. ( l9E'l). Ultrastructure of the joint capsule
in the rat: presence of t¡¡o kinds of capillaries. Nature 202,
920-92 l.
SunRnrHnßN, 1"1., Sotot'tolt, 5.0., hJtLltfiMs, P.5., RuElN, 4.L., N0u06n00sKYr Ê.
and SrenzEL, K.11. (198.1). liydroxyl radlcal s:avËngers inhibit
human natural killer cell actirzity. lïature 381r 21ã-??A.
Sl¡RNn¡ D,Ê. (1968). Studles on hyaluronic acid. ll. The protein
component(s) of rooster comb hyaluronic acid. Eiochim.
Biophys. Êcta 160, 96- 105.
SrrtRnx, D.Ê. and Rfioltt, E.L. ( 197?). The molecular basis of articular
lubrication. l. Purification and properties of a lubricating
fractlon from borine synovial fluid. J. Eiol. Ëhem. Z1?, AEE9-
B0?3.

I9?
SuRnn, D.Ê., I'|EN0nEH, R.8,, Rnotn, E.L., SoTMfiN, 3.L. and Duon, E.A. ( l9E l).
The lubrlcating actirlty of synovial fluid glycoproteins.
Êrthrltis Rhaum. 21, 22-38.
SttRnn, D.Ê., SrLuER, F.ll., Stott,tRlt, S.L. and Henmnnn, l'1. (lgE?).
Heasurements of reduclng end groups on borzine ritreous-
humour hyaluronic acid by reaction r¡ith l"E7-cyanide.
Eiochem. J. ?87, '{09-{ l'1.
St*lRtttt, D.Ê., ELocH, KJ., Sutnoett, D. and Srone, E. ( l9E{). The
lubricatfng actlvity of human synorial flulds. Êrthritls
Rheum. 2?, 552-558.
TRuaen, Ê.1. and ERelon, B.l'1. (19??). Eridence lor hydroxyl radlcal
productlon by human neutrophlls. J. Elln. lnvest. 60' 5?{-
5?9.
Toote, E.P. ( I gEZ)" Derzalopmental nolc of hyaluronätê. Conn. Tlss.
Res. 10,95-100.
TnuexRrt, P.5. and HcCRnrv, D. ( 19? I ). Synorial fluid pll, lactatc,
oxygen and carbon dioxlde: partlal pressure n various joint
dlseases. Êrthritls Rheum. 14, 4?5-443.
TsrcRltos, C.P., l/yntos, D.l'|. and KRt-pf,Nts, D.L. ( l9EE). Rooster comb
hyaluronate-protein a non-coralently llnked complex.
B¡ochem. J. 235, l17-123.
Unoennttt, E.E. and Toote, E.P. ( l9?9). Einding of hyaluronate to the
surface of cultured cells. J. Eell BIol. 82, {?5-48'1.
PR¡nsnnrn, 8.K., lteur-ÊonHyfrN, hJ.R., EnnYnln, P., PnGtN, Ê.Ê. and Gntetnro,
Ê"1. (19Ël). Three dlmanslonal structure of the enzyme
catalase. Natune 295, 'l I l-'t 12.
PgnrntnsuonftMfrNrñN, K. and JosERtt, K.T. ( I g??). Êctlon of slnglet
oxygÊn on collagen. lnd. J. Elochem. Elophys. l{' 2l?-2?0.

198
Puonro, E., EINoLR, s., HfrKKfiRfitNEn, s. and pe nntnen, R. ( lg8z). synthesis
of underpolymerized hyaluronlc acid by fibnoblasts culturedfrom rheumatoid and non-rheumatoid synorritis. Rheumatol.
lnt. 2, g?- l0Z.
l,rJeotocr, D.J,, PHtLLtpg, 8.0., Dautes, Ê., EoRHf,LLy, J. and l¡Jytt-JoHEs, E,
( l9E3). Depolyrmertzaflon of godtum hyaluronatc durrng
freeze drying" lnt. J. Blol. Hacromol. E, I EE- lËÊ.
lrlelss, S.J. ( lg80)" The role of superoxidc ln the de¡truction ofeythrocyte targets by human neutrophils. J. Eiol. Ehem.
255, 99 t?-ggt?.
hlulss, 5.J., RusrfiGt, P.K. and Logu6Lt0, Ê.F. ( l g?E). Human granulocyte
generation of hydroxyl radical. J. Exp. Hed. r4?, gr6-sag.
l,rJetss, sJ., YouN6, J., Loeuetto, 4.F., sLruKfr, Ê. and N¡HSH, N.F. ( lgE I ).
Role of hydrogen peroxide in neutrophil-mediated destructionof cultured endothelial cells. J. clln. Inrest. 68, ? l4-?z l.
hJerssmRrn, 8., ZuRrER, R.E. and llorrsTurN, s. ( lg?z). Leucocyte
pnoteases and the lmmunological release of þsosomalenzymes. Élmep. J. Pathol. 6Ë, 539-560.
l,rJelss¡lnnn, G., KoRcHfrK, l1.l't., Punez, ri.D., 3M0LEN, J,E., fìotosrurn, 1.11. and
llorrsntn, s.T. ( l9?9). LeukocytÊr ä5 secreiory organs of
inflammatlon. Êdr. lnflanrmation Res. l, g1-l lZ,
|rJrerrtn, 0.1¡,l. and Hurp, ll. ( lg?s). lnfluence of the cells on thepericellular ertvironment. The effect of hyaluronic acid on
proteoglylan synthesis and secreilon by chondrocytes ofadult cartilage. Phil. Trans. Royal soc. Lond. E zllr zgS-zgl,
hJrmen, hl.l., SMFH, PJ, and Ên¡rolT, s. (lg?s). llyaluronic acid:
structure of a fulþ extended 5-fold helical sodium salt and
comparison uith the less extended l-fold helical forms. J.
llol. Elol. 99, 219-255.

r9g
lrJrnrensounnE, E.E. ( lg?9). Comparison of suparoxlde and other
reduclng agents in the biological productlon of hydnoxyl
radicals. Elochem. J. I E2' EZ5-628.
hlOfe, S.F., HfiLLtbJELL, 8., Rlcttt.,tOno, R. and SfOlrlRoNECKr lrJ.R. (l3E l). The
role of superoxide and hydroxyl radicals in the degnadation
of hyalunonlc acld tnduced by metal Ions and by asconblc acid.
J. lnorganic Blochem. I {' lZ?- 154.
YoUne, 5.P. and Êtsen, P. ( | $EZ). The llrzer and lron. ln: The Lirzer:
Elology and Pathobiology. Êriäs, 1., Popper, ll.r Schacter, D.
and Shafrltz, D.Ê. (Edltors) pp 393-40'1. Raren Press, f'ler.,.l
York.
Yurcr, l'1. and Frsnman, hJ.l1. ( lSEg). Purlflcatlon and charactcrlzatlon
of lcach hyaluronlc acld-endo-p-glucuronldagc. J. Elol. Chem.
238, lA?1-18?9.
ZuRlrten, N. ( l9?5). The lntmunopathology of joint lnflammatlon in
rheumatoid arthrltiE. Êdr. lmmunol. l6' 285-338.

P. ?, r"cact¡on (3)
P. 10, reectlon llne I
P. 18, line ?
P. I g, linc 28
P. 139, linc I {
P. ll0, Table 5-l
P. l{1, line 2
P. I {8, line 22
P. 90, lnc 2l ([Fee+] = 50 ¡rL) should read ([Fe?+l = 50 ¡rl'l)
P. g?, |:me 29, Insert aftcr'llorgan-Elson assay'The reason that ther.e is a discnepency betr¡een thedegree of Park-Johnson reactlvlty for a glvenmolecular r.reight r.emains unclear, although it meybe related to the inherent inaccuracy in using thebroad pcak, obtained after gcl chromatography, tom?¡rur1r molcculan urcight.
P. 128, linc 15, Inscrt after 'but not Peak B'Peek Ê is also completeþ susceptible to digestionr.rlth hyaluronate lyase fnom streptomyceshyeluronolyticus (d¡ta not shor,rn).
ERRâTA SHEEÏ
0a- + Ha0a 0a + 0H.+ 0H-
zltÊDPH + 20¿ zltÊDP+ + 202 + HaO
'Glutatione' should r-ed'Glutathione'
'to bc ån an antioxidant' ¡hould re¡d 'to be ¡nantioxidant'
inscrt 'in' after 'irægularities'
headtng factor ls 'x l0-6'
'r',epolymenized' shou|d read'nepolymerize'
'so' should rced 'too'




















