
Deletion and interallelic complementation analysis of Steel mutant mice
Upload
independentCategory
view
3download
0
Cloning of a breast cancer homozygous deletion junction narrows the regionof search for a 3p21.3 tumor suppressor gene
Yoshitaka Sekido1,2, Mohsen Ahmadian1,7, Ignacio I Wistuba1, Farida Latif3,8, Scott Bader1,9,Ming-Hui Wei3,4, Fuh-Mei Duh3,4, Adi F Gazdar1,5, Michael I Lerman3 and John D Minna1,2,6
1Hamon Center for Therapeutic Oncology Research, Departments of 2Internal Medicine, 5Pathology and 6Pharmacology,University of Texas Southwestern Medical Center, Dallas, Texas 75235; 3Laboratory of Immunobiology and 4BiologicalCarcinogenesis and Development Program, SAIC Frederick, National Cancer Institute-Frederick Cancer Research andDevelopment Center, Frederick, Maryland 21702, USA
Chromosome 3p abnormalities and allele loss arefrequent in lung and breast cancers, and several lungcancer cell lines exhibit homozygous deletions of 3pindicating potential sites of tumor suppressor genes atregions 3p21.3, 3p14.2 and 3p12. We have identi®ed andcharacterized a new 3p21.3 homozygous deletion in abreast cancer cell line and the primary tumor thatoverlaps those previously described in small cell lungcancer (SCLC). This homozygous deletion is approxi-mately 220 kb in length and represents a somaticallyacquired change in the primary breast cancer. Cloningand sequencing of the breakpoint demonstrated that thisresulted from an interstitial deletion and preciselypinpoints this deletion within the three SCLC homo-zygous deletions previously reported. This deletionsigni®cantly narrows the minimum common deletedregion to 120 kb and is distinct from the previouslyreported region that suppresses tumor formation of themurine A9 ®brosarcoma cells. These ®ndings suggestthat a common homozygous deletion region on 3p21.3 isimportant in both lung and breast cancers. It is likelythat this very well characterized region either containsone tumor suppressor gene common to both tumor typesor two closely linked tumor suppressor genes speci®c foreach tumor.
Keywords: breast cancer; chromosome 3; homozygousdeletion; somatic mutation; breakpoint cloning; lungcancer
Introduction
Deletions and allele loss frequently a�ect chromosomeregion 3p in both SCLC and non-small cell lung cancer(NSCLC), implying the existence of a tumor suppres-sor gene (TSG) or genes in this region. Cytogeneticevidence ®rst showed a consistent deletion on the shortarm of chromosome 3 and loss of heterozygosity(LOH) was subsequently con®rmed by restrictionfragment length polymorphism (RFLP) analysis andallelotyping by microsatellite markers (Kok et al., 1987;
Naylor et al., 1987; Whang-Peng et al., 1982). Theseanalyses detected the deletion in nearly all SCLCs andabout three quarters of NSCLC, although the identityof such a gene(s) remains elusive despite intensiveinvestigation. So far, several distinct 3p regions havebeen identi®ed as showing frequent allele loss: 3p25-26,3p21.3-22, 3p14, and 3p12 (Hibi et al., 1992).Furthermore, homozygous deletions have been
found in the 3p21.3 region in three SCLC cell lines,NCI-H740, NCI-H1450 and GLC20 (Daly et al., 1993;Kok et al., 1994), in the 3p21.3-22 region in ®ve SCLCcell lines including ACC-LC-5 (Yamakawa et al.,1993), in the 3p14.2 FHIT/FRA3B region in six lungcancer cell lines (Fong et al., 1997; Ong et al., 1997)and in the 3p12 region in one SCLC cell line, U2020(Drabkin et al., 1992; Latif et al., 1992). Thesedeletions when originally identi®ed were large withestimated sizes of 1500 kb for NCI-H740, 800 kb forACC-LC-5 (Murata et al., 1994) and about 8 Mb forU2020. This implied a requirement of further massivee�orts for positional cloning of TSGs in these regions.Therefore, new homozygous deletions or rearrange-ments have been sought to narrow down these targetregions. In addition, homozygous deletions of 3p21.3were also found in three uncultured lung tumors by¯uorescence in situ hybridization analysis (Todd et al.,1997), indicating that the deletion is not an event ofcell culture but appears critical for primary cancerdevelopment.We have studied in detail several candidate 3p
TSGs. One candidate gene in the 3p25 region, the vonHippel-Lindau (VHL) tumor suppressor gene, is onlyrarely mutated in lung cancer (Sekido et al., 1994).Another candidate gene FHIT, which spans theFRA3B fragile site at 3p14.2, is associated withhomozygous deletions in lung cancer and withcancer-related FHIT cDNA splicing aberrations(Fong et al., 1997; Sozzi et al., 1996). However, innearly all cancers a wild-type FHIT transcript is alsoexpressed, making this gene di�erent from otherknown TSGs (Fong et al., 1997). A reasonableinterpretation of FHIT abnormalities is that theyrepresent alterations in the FRA3B fragile site(Boldog et al., 1997; Fong et al., 1997). The H740homozygous deletion region at 3p21.3 has also beenextensively studied by us and others, includingconstruction of a cosmid contig covering 700 kb (Weiet al., 1996), identi®cation of new genes including 3pK(Sithanandam et al., 1996), SKMc15 (Latif et al.,1997), Semaphorin A(V) (proposed symbol SEMAA)(Sekido et al., 1996), and Semaphorin IV (also calledSema III/F and proposed symbol SEMAK) (Roche et
Correspondence: JD MinnaCurrent addresses: 7KLA-Corp. San Jose, CA8Department of Paediatrics and Child Health, Clinical Genetics Unit,University of Birmingham, UK9Department of Pathology, The University of Edinburgh MedicalSchool, UKReceived 21 November 1997; revised 21 January 1998; accepted 22January 1998
Oncogene (1998) 16, 3151 ± 3157 1998 Stockton Press All rights reserved 0950 ± 9232/98 $12.00
http://www.stockton-press.co.uk/onc
al., 1996; Sekido et al., 1996; Xiang et al., 1996), andmutational analysis of these genes. However, thesegenes have been shown not to have a signi®cantnumber of mutations in lung cancer although somegenes such as Semaphorin A(V) seemed to be down-regulated dramatically in SCLC, which may indicatethat epigenetic changes are involved in inactivation ofthe gene (Sekido et al., 1996).Deletions of 3p have been demonstrated in breast
cancer by cytogenetic and LOH studies, indicatingseveral regions (3p24-26, 3p21.3, and 3p13-14.3)involved in breast cancer (Chen et al., 1994; Devileeet al., 1989; Pandis et al., 1995; Pandis et al., 1993;Sato et al., 1991). LOH at 3p21.3 was initially shownin seven of the 17 informative cases of 28 primarytumors with the D3F15S2 (previously calledDNF15S2) probe (Devilee et al., 1989). A homo-zygous deletion was identi®ed in a primary breastcancer with the D3S2 probe which maps to 3p14.3-21.1(Buchhagen et al., 1994). The FHIT gene was alsodemonstrated to be abnormal in breast cancer(Ahmadian et al., 1997; Hayashi et al., 1997; Negriniet al., 1996). In addition, LOH was detected inmorphologically normal lobules adjacent to breastcancers and the most frequent aberration was atchromosome region 3p22-25 (Deng et al., 1996). Thisevidence encouraged us to screen breast cancers fordeletions in the 3p21.3 region, leading us to identifyand characterize a new homozygous deletion in abreast cancer cell line recently started by us.
Results
Detection of a homozygous deletion in a breast cancercell line
Based on the evidence that there are three SCLCoverlapping homozygous deletions on 3p21.3 and thatbreast cancers also show frequent LOH at the 3p21.3locus, we performed studies to detect genomic DNAabnormalities in 26 breast cancer cell lines. SelectedPCR primers, localized to the H740 deletion region,were tested for detection of homozygous deletions, andSouthern blot analyses were performed to survey theH740 homozygous deletion region to con®rm PCRdata as well as to independently detect abnormalities.One cell line among 26 breast cancers, HCC1500,
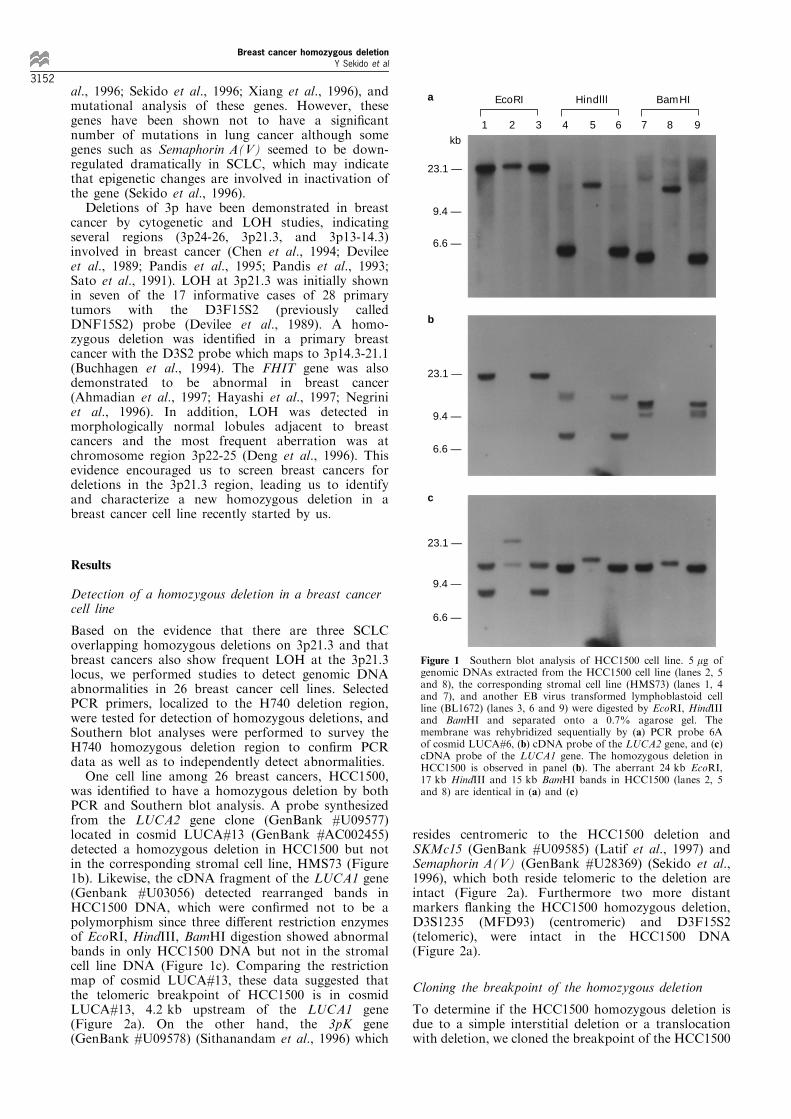
was identi®ed to have a homozygous deletion by bothPCR and Southern blot analysis. A probe synthesizedfrom the LUCA2 gene clone (GenBank #U09577)located in cosmid LUCA#13 (GenBank #AC002455)detected a homozygous deletion in HCC1500 but notin the corresponding stromal cell line, HMS73 (Figure1b). Likewise, the cDNA fragment of the LUCA1 gene(Genbank #U03056) detected rearranged bands inHCC1500 DNA, which were con®rmed not to be apolymorphism since three di�erent restriction enzymesof EcoRI, HindIII, BamHI digestion showed abnormalbands in only HCC1500 DNA but not in the stromalcell line DNA (Figure 1c). Comparing the restrictionmap of cosmid LUCA#13, these data suggested thatthe telomeric breakpoint of HCC1500 is in cosmidLUCA#13, 4.2 kb upstream of the LUCA1 gene(Figure 2a). On the other hand, the 3pK gene(GenBank #U09578) (Sithanandam et al., 1996) which
resides centromeric to the HCC1500 deletion andSKMc15 (GenBank #U09585) (Latif et al., 1997) andSemaphorin A(V) (GenBank #U28369) (Sekido et al.,1996), which both reside telomeric to the deletion areintact (Figure 2a). Furthermore two more distantmarkers ¯anking the HCC1500 homozygous deletion,D3S1235 (MFD93) (centromeric) and D3F15S2(telomeric), were intact in the HCC1500 DNA(Figure 2a).
Cloning the breakpoint of the homozygous deletion
To determine if the HCC1500 homozygous deletion isdue to a simple interstitial deletion or a translocationwith deletion, we cloned the breakpoint of the HCC1500
23.1 —
23.1 —
23.1 —
9.4 —
9.4 —
9.4 —
6.6 —
6.6 —
6.6 —
1 2 3 4 5 6 7 8 9
EcoRI Hindlll BamHI
kb
a
b
c
Figure 1 Southern blot analysis of HCC1500 cell line. 5 mg ofgenomic DNAs extracted from the HCC1500 cell line (lanes 2, 5and 8), the corresponding stromal cell line (HMS73) (lanes 1, 4and 7), and another EB virus transformed lymphoblastoid cellline (BL1672) (lanes 3, 6 and 9) were digested by EcoRI, HindIIIand BamHI and separated onto a 0.7% agarose gel. Themembrane was rehybridized sequentially by (a) PCR probe 6Aof cosmid LUCA#6, (b) cDNA probe of the LUCA2 gene, and (c)cDNA probe of the LUCA1 gene. The homozygous deletion inHCC1500 is observed in panel (b). The aberrant 24 kb EcoRI,17 kb HindIII and 15 kb BamHI bands in HCC1500 (lanes 2, 5and 8) are identical in (a) and (c)
Breast cancer homozygous deletionY Sekido et al
3152
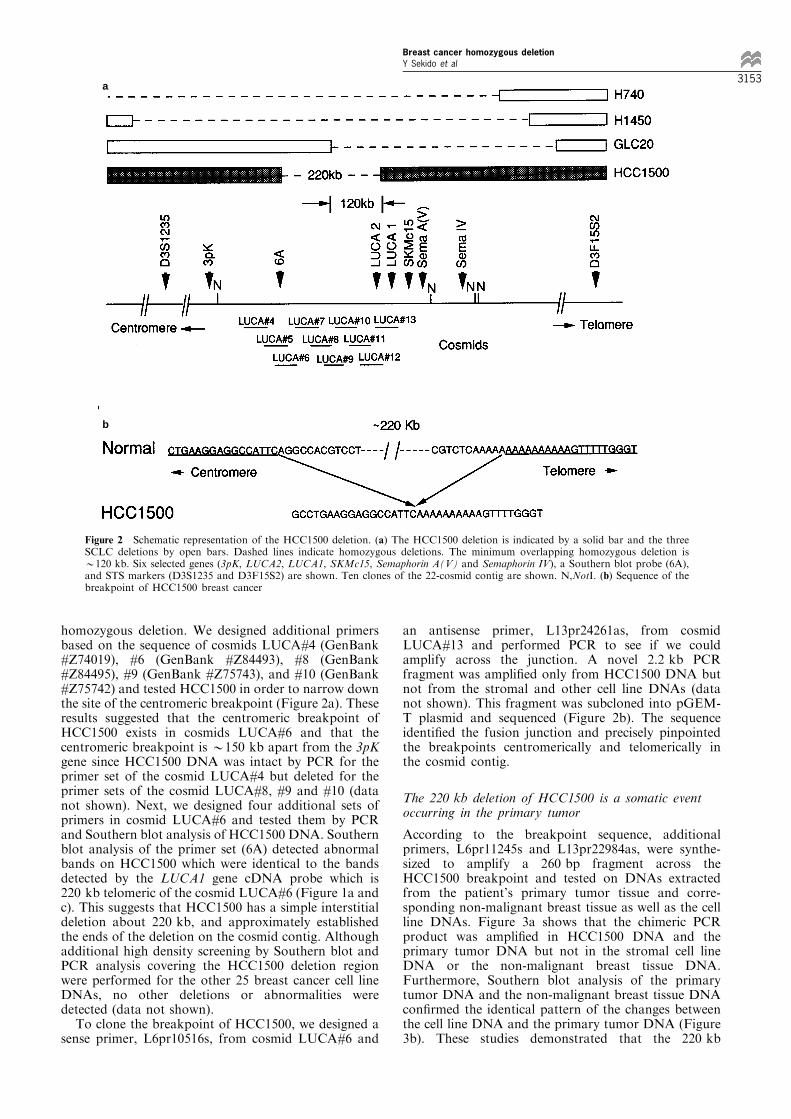
homozygous deletion. We designed additional primersbased on the sequence of cosmids LUCA#4 (GenBank#Z74019), #6 (GenBank #Z84493), #8 (GenBank#Z84495), #9 (GenBank #Z75743), and #10 (GenBank#Z75742) and tested HCC1500 in order to narrow downthe site of the centromeric breakpoint (Figure 2a). Theseresults suggested that the centromeric breakpoint ofHCC1500 exists in cosmids LUCA#6 and that thecentromeric breakpoint is *150 kb apart from the 3pKgene since HCC1500 DNA was intact by PCR for theprimer set of the cosmid LUCA#4 but deleted for theprimer sets of the cosmid LUCA#8, #9 and #10 (datanot shown). Next, we designed four additional sets ofprimers in cosmid LUCA#6 and tested them by PCRand Southern blot analysis of HCC1500 DNA. Southernblot analysis of the primer set (6A) detected abnormalbands on HCC1500 which were identical to the bandsdetected by the LUCA1 gene cDNA probe which is220 kb telomeric of the cosmid LUCA#6 (Figure 1a andc). This suggests that HCC1500 has a simple interstitialdeletion about 220 kb, and approximately establishedthe ends of the deletion on the cosmid contig. Althoughadditional high density screening by Southern blot andPCR analysis covering the HCC1500 deletion regionwere performed for the other 25 breast cancer cell lineDNAs, no other deletions or abnormalities weredetected (data not shown).To clone the breakpoint of HCC1500, we designed a
sense primer, L6pr10516s, from cosmid LUCA#6 and
an antisense primer, L13pr24261as, from cosmidLUCA#13 and performed PCR to see if we couldamplify across the junction. A novel 2.2 kb PCRfragment was ampli®ed only from HCC1500 DNA butnot from the stromal and other cell line DNAs (datanot shown). This fragment was subcloned into pGEM-T plasmid and sequenced (Figure 2b). The sequenceidenti®ed the fusion junction and precisely pinpointedthe breakpoints centromerically and telomerically inthe cosmid contig.
The 220 kb deletion of HCC1500 is a somatic eventoccurring in the primary tumor
According to the breakpoint sequence, additionalprimers, L6pr11245s and L13pr22984as, were synthe-sized to amplify a 260 bp fragment across theHCC1500 breakpoint and tested on DNAs extractedfrom the patient's primary tumor tissue and corre-sponding non-malignant breast tissue as well as the cellline DNAs. Figure 3a shows that the chimeric PCRproduct was ampli®ed in HCC1500 DNA and theprimary tumor DNA but not in the stromal cell lineDNA or the non-malignant breast tissue DNA.Furthermore, Southern blot analysis of the primarytumor DNA and the non-malignant breast tissue DNAcon®rmed the identical pattern of the changes betweenthe cell line DNA and the primary tumor DNA (Figure3b). These studies demonstrated that the 220 kb
a
b
Figure 2 Schematic representation of the HCC1500 deletion. (a) The HCC1500 deletion is indicated by a solid bar and the threeSCLC deletions by open bars. Dashed lines indicate homozygous deletions. The minimum overlapping homozygous deletion is*120 kb. Six selected genes (3pK, LUCA2, LUCA1, SKMc15, Semaphorin A(V) and Semaphorin IV), a Southern blot probe (6A),and STS markers (D3S1235 and D3F15S2) are shown. Ten clones of the 22-cosmid contig are shown. N,NotI. (b) Sequence of thebreakpoint of HCC1500 breast cancer
Breast cancer homozygous deletionY Sekido et al
3153

deletion represents a somatic alteration acquired in theprimary tumor.The primary tumor specimen used to prepare bulk
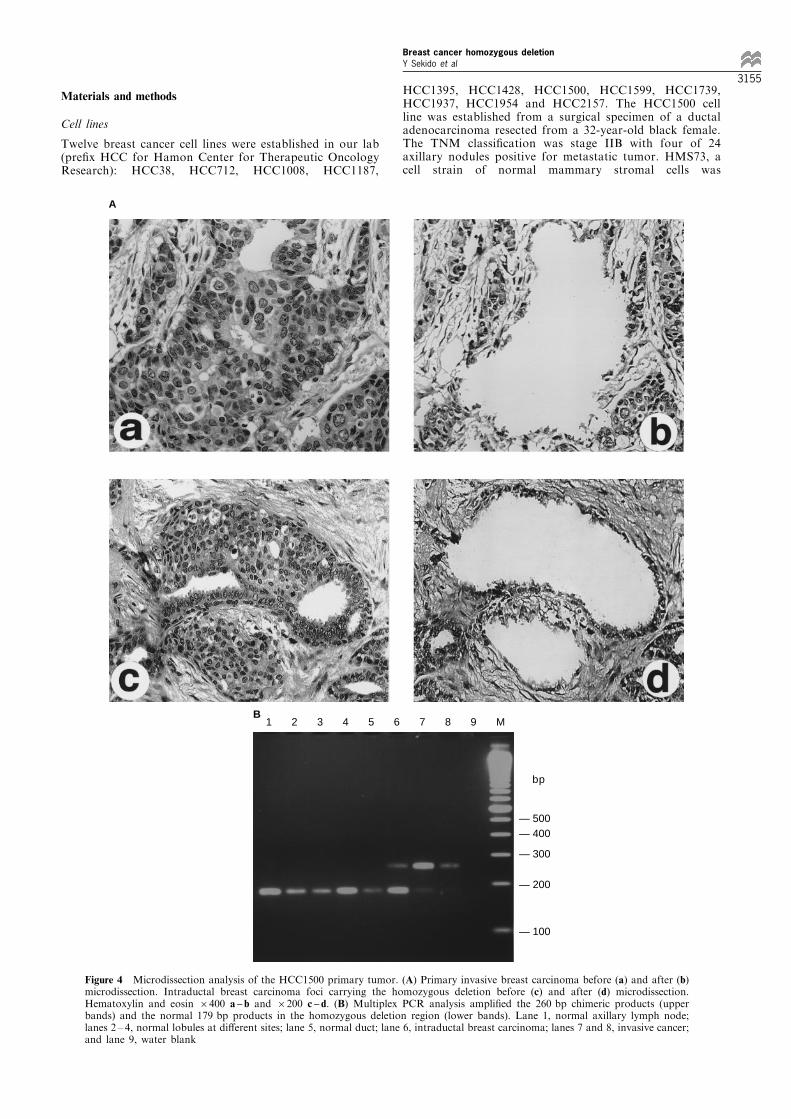
DNA was contaminated with normal cells. Thus theSouthern blot gave bands hybridizing to both thenormal germline fragments as well as the rearrangednovel fragments (Figure 3b). To determine if only themutant or both mutant and wild-type allele were inthe tumor cells, we employed microdissectiontechnique followed by PCR assay to avoid contam-ination with normal cells. Precisely microdissectedspecimens were obtained from the para�n-embeddedprimary invasive breast carcinoma and accompanyingintraductal carcinoma, histologically normal lobulesand ductal breast epithelia (Figure 4A). Theintraductal carcinoma cells were contiguous tohistologically normal ductal cells and contaminationin the microdissection could not be avoided (Figure4A, c and d). Multiplex PCR was performed with aprimer set of L6pr11245s and L13pr22984as amplify-ing the chimeric 260 bp fragment and a primer set ofL12pr35s and L12pr36as amplifying a control normal179 bp fragment which resides in the 220 kbhomozygous deletion region. The results showedthat the 260 bp chimeric band was detected in theinvasive cancer, while the wild-type band was weakor absent, indicating loss of the wild-type allele(Figure 4B lanes 7 and 8). Of interest, both the260 bp chimeric band and the 179 bp normal banddetecting the deletion were ampli®ed in the intraduc-tal breast carcinoma (Figure 4B lane 6). This couldbe explained either by contamination with normalductal cells or by retention of the wild-type allele in
the intraductal component. Several samples ofadjacent histologically normal cells showed only thenormal band, which again con®rmed that theinterstitial deletion was tumor-speci®c.
Discussion
The present data show that one breast cancer cell line,HCC1500, and its primary tumor have a 220 kbhomozygous deletion overlapping the three commonhomozygous deletions in SCLC cell lines, NCI-H740,NCI-H1450 and GLC20, which themselves have anestimated common region around 370 kb (Kok et al.,1994; Wei et al., 1996). The homozygous deletion ofHCC1500 has now narrowed the common region ofdeletion to 120 kb, which suggests that lung and breastcancers might share the same TSG for the 3p21.3locus. Alternatively, it is possible that there are twoclosely linked TSGs, one for lung cancer and the otherfor breast cancer. While the three homozygousdeletions in SCLC were demonstrated only in theestablished tumor cell lines, the 220 kb deletion ofHCC1500 provides clear evidence that this change wasacquired somatically and existed in the primary tumor.These results, along with those of Todd et al. (1997),provide strong support that this region is critical fortumor development.Cloning and sequencing of the breakpoint demon-
strated that this resulted from an interstitial deletion.We performed a BLAST search with the fusionjunction sequence but no high quality matches werefound. It will be interesting to compare other fusionjunctions from cancer related interstitial deletions ortranslocations to see if there is a common theme. Thiscould provide a clue to the enzymatic mechanismgenerating these common changes found in cancer.The telomeric deletion of HCC1500 is 4.2 kb
upstream of the LUCA1 gene, which potentiallycould involve 5' regulatory sequence of LUCA1. Itappears to exclude other candidate TSGs in thecontig such as SKMc15, Semaphorin A(V), GNAI2,GNAT1 and Semaphorin IV (also called SemaIII/F)residing in the telomeric region of the H740homozygous deletion. This region is also contiguousto, but quite distinct from that identi®ed by atransfection assay using a P1 clone into murine A9cells (Todd et al., 1996). This region is much furthertelomeric from the breakpoint of the HCC1500homozygous deletion (Todd et al., 1996). However,this P1 clone did suppress tumorigenicity, albeit inmouse cells, providing functional evidence of a TSGin this distal region. It will be necessary to performsimilar functional tests in the HCC1500 deletionregion. The centromeric breakpoint of the HCC1500deletion was about 150 kb centromeric from the 3pKgene, which was found not to have mutations(Sithanandam et al., 1996). In the 120 kb commondeletion region, we have identi®ed eight distinct genesso far including LUCA1, LUCA2, fus1, 123F2, BLU2,101F6, PL6 (Wei et al., 1996) and Gene 26, all ofwhich now require detailed mutational analysis inbreast and other cancers as well as lung cancers. It isof course possible that more than one TSG exists inthis region, one of which is important for breast andthe other for lung cancer.
1 2 3 4 5 6 M
bp
— 500— 400
— 300
— 200
— 100
No
rmal
Tis
sue
Pri
mar
y Tu
mo
r
— 23.1
— 9.4
— 6.6
— 4.4
— 2.3— 2.0
a bkb
Figure 3 Identi®cation of the 220 kb deletion in the HCC1500primary tumor. (a) PCR analysis of genomic DNAs using aprimer set to cross the HCC1500 breakpoint. A 260 bp abnormalband is ampli®ed for HCC1500 and the primary tumor DNAs butnot in the corresponding stromal cell lines (HMS73), the non-malignant breast tissue or another cell line DNAs (BL1672). Lane1, HMS73; lane 2, HCC1500; lane 3, BL1672; lane 4, non-malignant breast tissue of the HCC1500 patient; lane 5, primarytumor of the HCC1500 patient; and lane 6, water blank. (b)Southern blot analysis of the normal breast tissue and theprimary tumor DNAs of HCC1500. 5 mg of genomic DNAs weredigested by BamHI and the membrane was hybridized by PCRprobe 6A. An arrow indicates the abnormal 15 kb band which isthe same as the one of lane 8 on Figure 1a
Breast cancer homozygous deletionY Sekido et al
3154
Materials and methods
Cell lines
Twelve breast cancer cell lines were established in our lab(pre®x HCC for Hamon Center for Therapeutic OncologyResearch): HCC38, HCC712, HCC1008, HCC1187,
HCC1395, HCC1428, HCC1500, HCC1599, HCC1739,HCC1937, HCC1954 and HCC2157. The HCC1500 cellline was established from a surgical specimen of a ductaladenocarcinoma resected from a 32-year-old black female.The TNM classi®cation was stage IIB with four of 24axillary nodules positive for metastatic tumor. HMS73, acell strain of normal mammary stromal cells was
— 500— 400
— 300
— 200
— 100
1 2 3 4 5 6 7 8 9 M
bp
B
Figure 4 Microdissection analysis of the HCC1500 primary tumor. (A) Primary invasive breast carcinoma before (a) and after (b)microdissection. Intraductal breast carcinoma foci carrying the homozygous deletion before (c) and after (d) microdissection.Hematoxylin and eosin 6400 a ± b and 6200 c ± d. (B) Multiplex PCR analysis ampli®ed the 260 bp chimeric products (upperbands) and the normal 179 bp products in the homozygous deletion region (lower bands). Lane 1, normal axillary lymph node;lanes 2 ± 4, normal lobules at di�erent sites; lane 5, normal duct; lane 6, intraductal breast carcinoma; lanes 7 and 8, invasive cancer;and lane 9, water blank
A
Breast cancer homozygous deletionY Sekido et al
3155

established from a non-malignant portion of the mastect-omy specimen, and was used as the source of constitutionalDNA.
Fourteen breast cancer cell lines were obtained from theAmerican Type Culture Collection (Rockville, MD); HTB19(BT-20), HTB20 (BT-474), HTB22 (MCF-7), HTB23 (MDA-MB-131-VI), HTB24 (MDA-MB-157), HTB25 (MDA-MB-157-VII), HTB26 (MDA-MB-231), HTB27 (MDA-MB-361),HTB121 (BT-483), HTB129 (MDA-MB-435S), HTB130(MDA-MB-436), HTB131 (MDA-MB-453), HTB132(MDA-MB-468), and HTB133 (T-47D).
Southern blot analysis
High molecular weight DNAs were extracted by standardmethods (Sambrook et al., 1989). Five mg of DNAs,digested with appropriate restriction endonucleases (Gib-co BRL), were electrophoresed in 0.7% agarose gels,transferred to Hybond N+ membranes (Amersham),hybridized with 32P-labeled probes synthesized withrandom primers and subjected to autoradiography afterwashing according to the company protocol (Amersham).Reprobing of the same membranes was performed afterstripping of previous probes.
PCR analysis
Primers were designed either from sequences of cDNAclones we isolated in the 3p21.3 homozygous deletionregion or from genomic sequences of the 700 kb cosmidand P1 clone contig (Wei et al., 1996). This contig is beingsequenced jointly by The Genome Sequence Center atWashington University, St Louis and The Sanger Centre,Hinxton, UK, and most of the sequences have beendeposited in GenBank. Primers were synthesized based onthe sequences of the cosmids and the key primer sets aregiven below while the rest are available upon request. Theprimer set (6A) for Southern blot probe (Figure 1a)designed from cosmid LUCA#6 is L6pr11s, 5'-AAGGA-CAACTTTGTCACCAG-3' and L6pr12as, 5'-CAAGGA-GAGATACAAAGACTC-3'. The primer set for cloning
the 2.2 kb fragment of the HCC1500 breakpoint isL6pr10516s, 5'-AGGTGGAGATTCTGCAGAAG-3' fromcosmid LUCA#6 and L13pr24261as, 5'-AGCAGTCTA-CAGCCTGAGAC-3' from cosmid LUCA#13. The primerset for demonstrating the 260 bp novel fusion junctionfragment of the HCC1500 breakpoint is L6pr11245s, 5'-TCTCAGCACTTGAGGCATAG-3' from cosmidLUCA#6, and L13pr22984as, 5'-TGACAGAGCAAGAC-TCCATCTG-3' from cosmid LUCA#13. The primer setfrom cosmid LUCA#12 for detecting the normal 179 bpfragment in the homozygous deletion region for micro-dissected samples is L12pr35s, 5'-CTGTCCTTTACCC-TACCCTT-3', and L12pr36as, 5'-AGGAGTGAGAAGA-GGTATGAA-3'.
Some PCR products were further puri®ed and used asprobes for Southern blot analysis. The 2.2 kb PCR productcovering the breakpoint of the interstitial deletion ofHCC1500 was subcloned into pGEM-T vector (Pharmacia),pHC1500-2.2, and bidirectionally sequenced by automatedcycle sequencing (ABI).
Microdissection and DNA extraction
Microdissection and DNA extraction were performed frommicroslides as previously described (Hung et al., 1995).Precisely identi®ed areas of the para�n-embedded primaryinvasive carcinoma and accompanying intraductal carcino-ma, histologically normal breast lobules and ductal breastepithelia were microdissected under microscopic visualiza-tion. Lymphocytes from a normal axillary lymph nodewere also obtained. After DNA extraction, 5 ml of thedigested samples, containing DNA from at least 100 cells,were used for each multiplex PCR.
AcknowledgementsWe would like to acknowledge Dr JW Shay for providingthe HMS73 cell line. This work was supported by Susan GKomen Breast Cancer Foundation, National CancerInstitute Grant R01 CA71618, and Texas AdvancedResearch Program Grant ARP003660-080.
References
Ahmadian M, Wistuba II, Fong KM, Behrens C, KodagodaDR, Saboorian MH, Shay J, Tomlinson GE, Blum J,Minna JD and Gazdar AF. (1997). Cancer Res., 57, 3664 ±3668.
Boldog F, Gemmill RM, West J, Robinson M, Robinson L,Li E, Roche J, Todd S, Waggoner B, Lundstrom R,Jacobson J, Mullokandov MR, Klinger H and DrabkinHA. (1997). Hum. Mol. Genet., 6, 193 ± 203.
Buchhagen DL, Qiu L and Etkind P. (1994). Int. J. Cancer,57, 473 ± 479.
Chen LC, Matsumura K, Deng G, Kurisu W, Ljung BM,Lerman MI, Waldman FM and Smith HS. (1994). CancerRes., 54, 3021 ± 3024.
Daly MC, Xiang R-H, Buchhagen D, Hensel CH, GarciaDK, Killary AM, Minna JD and Naylor SL. (1993).Oncogene, 8, 1721 ± 1729.
Deng G, Lu Y, Zlotnikov G, Thor AD and Smith HS. (1996).Science, 274, 2057 ± 2059.
Devilee P, van den Broek M, Kuipers-Dijkshoorn N, KolluirR, Khan PM, Pearson PL and Cornelisse CJ. (1989).Genomics, 5, 554 ± 560.
Drabkin HA, Mendez MJ, Rabbitts PH, Varkony T, Bergh J,Schlessinger J, Erickson P and Gemmill RM. (1992).Genes Chrom. Cancer, 5, 67 ± 74.
Fong KM, Biesterveld EJ, Virmani A, Wistuba I, Sekido Y,Bader SA, Ahmadian M, Ong ST, Rassool FV, Zimmer-man PV, Giaccone G, Gazdar AF and Minna JD. (1997).Cancer Res., 57, 2256 ± 2267.
Hayashi S, Tanimoto K, Hajiro-Nakanishi K, Tsuchiya E,Kurosumi M, Higashi Y, Imai K, Suga K and Nakachi K.(1997). Cancer Res., 57, 1981 ± 1985.
Hibi K, Takahashi T, Yamakawa K, Ueda R, Sekido Y,Ariyoshi Y, Suyama M, Takagi H, Nakamura Y andTakahashi T. (1992). Oncogene, 7, 445 ± 449.
Hung J, Kishimoto Y, Sugio K, Virmani A, McIntire DD,Minna JD and Gazdar AF. (1995). JAMA, 273, 558 ± 563.
Kok K, Osinga J, Carritt B, Davis MB, van der Hout AH,van der Veen AY, Landsvater RM, de Leij LFMH,Berendsen HH, Postmus PE, Poppema S and BuysCHCM. (1987). Nature, 330, 578 ± 581.
Kok K, van der Berg A, Veldhuis PMJF, van der Veen AY,Franke M, Schoenmakers EFPM, Hulsbeek MMF, vander Hout AH, de Leij L, van de Ven W and Buys CHCM.(1994). Cancer Res., 54, 4183 ± 4187.
Latif F, Duh F-M, Bader S, Sekido Y, Li H, Geil L, Zbar B,Minna JD and Lerman MI. (1997). Human Genetics, 99,334 ± 341.
Breast cancer homozygous deletionY Sekido et al
3156

Latif F, Tory K, Modi WS, Graziano SL, Gamble G,Douglas J, Heppell-Parton AC, Rabbitts PH, Zbar B andLerman MI. (1992). Genes Chrom. Cancer, 5, 119 ± 127.
Murata Y, Tamari M, Takahashi T, Horio Y, Hibi K,Yokoyama S, Inazawa J, Yamakawa K, Ogawa A,Takahashi T and Nakamura Y. (1994). Hum. Mol.Genet., 3, 1341 ± 1344.
Naylor SL, Johnson BE, Minna JD and Sakaguchi AY.(1987). Nature, 329, 451 ± 454.
Negrini M, Monaco C, Vorechovsky I, Ohta M, Druck T,Ba�a R, Huebner K and Croce CM. (1996). Cancer Res.,56, 3173 ± 3179.
Ong ST, Fong KM, Bader SA, Minna JD, Lebeau MM,Mckeithan TW and Rassool FV. (1997). Genes Chrom.Cancer, 20, 16 ± 23.
Pandis N, Jin Y, Gorunova L, Petersson C, Bardi G, Idvall I,Johansson B, Ingvar C, Mandahl N, Mitelman F andHeum S. (1995). Genes Chrom. Cancer, 12, 173 ± 185.
Pandis N, Jin Y, Limon J, Bardi G, Idvall I, Mandahl N,Mitelman F and Heim S. (1993). Genes Chrom. Cancer, 6,151 ± 155.
Roche J, Boldog F, Robinson M, Varella-Garcia M,Swanton M, Waggoner B, Fishel R, Franklin W, GemmillR and Drabkin H. (1996). Oncogene, 12, 1289 ± 1297.
Sambrook J, Fritsch EF and Maniatis T. (1989). MolecularCloning: A laboratory Manual, Second edition, Cold SpringHarbor Laboratory Press: New York.
Sato T, Akiyama F, Sakamoto G, Kasumi F and NakamuraY. (1991). Cancer Res., 51, 5794 ± 5799.
Sekido Y, Bader S, Latif F, Chen J-Y, Duh F-M, Wei M-H,Albanesi JP, Lee C-C, Lerman MI and Minna JD. (1996).Proc. Natl. Acad. Sci. USA, 93, 4120 ± 4125.
Sekido Y, Bader S, Latif F, Gnarra JR, Gazdar AF, LinehanWM, Zbar B, Lerman MI and Minna JD. (1994).Oncogene, 9, 1599 ± 1604.
Sithanandam G, Latif F, Duh F-M, Bernal R, Smola U, LiH, Kuzmin I, Wixler V, Geil L, Shrestha S, Lloyd PA,Bader S, Sekido Y, Tartof KD, Kashuba VI, ZabarovskyER, Dean M, Klein G, Lerman MI, Minna JD, Rapp URand Allikmets R. (1996). Mol. Cell. Biol., 16, 868 ± 876.
Sozzi G, Veronese ML, Negrini M, Ba�a R, Cotticelli MG,Inoue H, Tornielli S, Pilotti S, De Gregorio L, PastorinoU, Pierotti MA, Ohta M, Huebner K and Croce CM.(1996). Cell, 85, 17 ± 26.
Todd MC, Xiang R-H, Garcia DK, Kerbacher KE, MooreSL, Hensel CH, Liu P, Siciliano MJ, Kok K, van den BergA, Veldhuis P, Buys CHCM, Killary AM and Naylor SL.(1996). Oncogene, 13, 2387 ± 2396.
Todd S, Franklin WA, Varella-Garcia M, Kennedy T,Hilliker CEJ, Hahner L, Anderson M, Wiest JS, DrabkinHA and Gemmill RM. (1997). Cancer Res., 57, 1344 ±1352.
Wei MH, Latif F, Bader S, Kashuba V, Chen JY, Duh FM,Sekido Y, Lee CC, Geil L, Kuzmin I, Zabarovsky E, KleinG, Zbar B, Minna JD and Lerman MI. (1996). CancerRes., 56, 1487 ± 1492.
Whang-Peng J, Kao-Shan CS, Lee EC, Bunn PA, CarneyDN, Gazdar AF and Minna JD. (1982). Science, 215,181 ± 182.
Xiang RH, Hensel CH, Garcia DK, Carlson HC, Kok K,Daly MC, Kerbacher K, van den Berg A, Veldhuis P, BuysCHCM and Naylor SL. (1996). Genomics, 32, 39 ± 48.
Yamakawa K, Takahashi T, Horio Y, Murata Y, TakahashiE, Hibi K, Yokoyama S, Ueda R, Takahashi T andNakamura Y. (1993). Oncogene, 8, 327 ± 330.
Breast cancer homozygous deletionY Sekido et al
3157

Copyright © 2022 FDOKUMEN




















