
Chondroitin sulfate proteoglycans in the developing central nervous system. I. Cellular sites of...
14
THE JOURNAL OF COMPARATIVE NEUROLOGY 355515-628 (1995) Chondroitin Sulfate Proteoglycans in the Developing Cerebral Cortex: The Distribution of Neurocan Distinguishes Forming Afferent and Efferent Axonal Pathways BRAD MILLER, ALLAN M. SHEPPARD, ALMA R. BICKNESE, AND ALAN L. PEARLMAN Departments of Cell Biology (B.M., A.M.S., A.L.P.), Neurology (A.R.B.,A.L.P.), and Pediatrics (A.R.B.),Washington University School of Medicine, St. Louis, Missouri 63110 ABSTRACT The first thalamocortical axons to arrive in the developing cerebral cortex traverse a pathway that is separate from the adjacent intracortical pathway for early efferents, suggesting that different molecular signals guide their growth. We previously demonstrated that the intracortical pathway for thalamic axons is centered on the subplate (Bicknese et al. [19941 J. Neurosci. 14:3500-3510), which is rich in chondroitin sulfate proteoglycans (CSPGs; Sheppard et al. [1991] J. Neurosci. 11:3928-39421, whereas efferent axons cross the subplate to exit in a zone containing much less CSPG. To define the molecular composition of the subplate further, we used antibodies against CSPG core proteins and chondroitin sulfate disaccharides in an immunohistochemical analysis of their distribution in the developing neocortex of the rat. Immunolabeling for neurocan, a central nervous system-specific CSPG (Rauch et al. [19921 J. Biol. Chem. 267: 19537-19547), and for chondroitin 6-sulfate and unsulfated chondroitin becomes prominent in the subplate before the arrival of thalamic afferents. Immunolabeling is initially sparse in the cortical plate but appears later in maturing cortical layers. A postnatal decline in immunolabeling occurs uniformly for most proteoglycans, but, in the somatosensory cortex, labeling for neurocan, phosphacan, and chondroitin 4- and 6-sulfate declines in the centers of the whisker barrels before the walls. In contrast to neurocan, immunolabeling for other proteoglycans is either uniformly distributed (syndecan-1, N-syndecan, 5F3, phosphacan, chondroitin 4-sulfate), restricted to axons (PGMl), distributed exclusively on nonneuronal elements (2D6, NG2, and CD44), or undetectable (9.2.27, aggrecan, decorin). Thus, neurocan is a candidate molecule for delineating the intracortical pathway of thalamocortical axons and distinguishing it from that of cortical efferents. o 1995 Wiley-Liss, Inc. Indexing terms: development, extracellular matrix, glycosaminoglycan The paths taken by the first axons to enter and leave the embryonic cerebral cortex are adjacent but distinct (McCo- nnell et al., 1989; De Carlos and O'Leary, 1992; Erzurumlu Accepted October 13, 1994, and Jhaveri, 1992; Miller et a1.9 1993; Bicknese et al.9 l994), Brad ill^^,^ present address is ~ i v i ~ i ~ ~ of Experimental ~~~~~l~~ and indicating that the ~Okcular Signals that guide these two sets of axons also differ. In prior studies, we have demon- strated that the intracortical trajectory of the first axons to (Bicknese et al., 1994), a layer rich in chondroitin sulfate Neurological Surgery, Washington University School of Medicine, St. Louis, Allan M. Sheppard's present address is Division of Respiratory and MO 63110. arrive from the thalamus is centered on the subplate Critical Care, Department of Medicine, Washington University School of Alma R. Bicknese's present address is Department of Neurology, State Address reprint requests to Alan L. Pearlman, Department of Cell Biology-Box 8228, Washington University School of Medicine, 660 South Euclid Avenue, St. Louis, MO 63110. Medicine, St. Mo 63110. proteoglYcans (CSPGs; Sheppard et al., lggl; Bicknese et University ofNew York, Stony Brook, Ny 11794, 1994) and Other extracellular matrix (ECM) cornPo- nents (Derer and Nakanishi, 1983; DelPech and DelPech, 1984; Stewart and Pearlman, 1987; Chun and Shatz, 1988; o 1995 WILEY-LISS, INC.
Transcript of Chondroitin sulfate proteoglycans in the developing central nervous system. I. Cellular sites of...

THE JOURNAL OF COMPARATIVE NEUROLOGY 355515-628 (1995)
Chondroitin Sulfate Proteoglycans in the Developing Cerebral Cortex: The
Distribution of Neurocan Distinguishes Forming Afferent and Efferent Axonal
Pathways
BRAD MILLER, ALLAN M. SHEPPARD, ALMA R. BICKNESE, AND ALAN L. PEARLMAN
Departments of Cell Biology (B.M., A.M.S., A.L.P.), Neurology (A.R.B., A.L.P.), and Pediatrics (A.R.B.), Washington University School of Medicine, St. Louis, Missouri 631 10
ABSTRACT The first thalamocortical axons to arrive in the developing cerebral cortex traverse a
pathway that is separate from the adjacent intracortical pathway for early efferents, suggesting that different molecular signals guide their growth. We previously demonstrated that the intracortical pathway for thalamic axons is centered on the subplate (Bicknese et al. [19941 J. Neurosci. 14:3500-3510), which is rich in chondroitin sulfate proteoglycans (CSPGs; Sheppard et al. [1991] J. Neurosci. 11:3928-39421, whereas efferent axons cross the subplate to exit in a zone containing much less CSPG. To define the molecular composition of the subplate further, we used antibodies against CSPG core proteins and chondroitin sulfate disaccharides in an immunohistochemical analysis of their distribution in the developing neocortex of the rat. Immunolabeling for neurocan, a central nervous system-specific CSPG (Rauch et al. [19921 J. Biol. Chem. 267: 19537-19547), and for chondroitin 6-sulfate and unsulfated chondroitin becomes prominent in the subplate before the arrival of thalamic afferents. Immunolabeling is initially sparse in the cortical plate but appears later in maturing cortical layers. A postnatal decline in immunolabeling occurs uniformly for most proteoglycans, but, in the somatosensory cortex, labeling for neurocan, phosphacan, and chondroitin 4- and 6-sulfate declines in the centers of the whisker barrels before the walls. In contrast to neurocan, immunolabeling for other proteoglycans is either uniformly distributed (syndecan-1, N-syndecan, 5F3, phosphacan, chondroitin 4-sulfate), restricted to axons (PGMl), distributed exclusively on nonneuronal elements (2D6, NG2, and CD44), or undetectable (9.2.27, aggrecan, decorin). Thus, neurocan is a candidate molecule for delineating the intracortical pathway of thalamocortical axons and distinguishing it from that of cortical efferents. o 1995 Wiley-Liss, Inc.
Indexing terms: development, extracellular matrix, glycosaminoglycan
The paths taken by the first axons to enter and leave the embryonic cerebral cortex are adjacent but distinct (McCo- nnell et al., 1989; De Carlos and O'Leary, 1992; Erzurumlu Accepted October 13, 1994, and Jhaveri, 1992; Miller et a1.9 1993; Bicknese et al.9 l994), Brad ill^^,^ present address is ~ i v i ~ i ~ ~ of Experimental ~~~~~l~~ and indicating that the ~ O k c u l a r Signals that guide these two sets of axons also differ. In prior studies, we have demon- strated that the intracortical trajectory of the first axons to
(Bicknese et al., 1994), a layer rich in chondroitin sulfate
Neurological Surgery, Washington University School of Medicine, St. Louis,
Allan M. Sheppard's present address is Division of Respiratory and MO 63110.
arrive from the thalamus is centered on the subplate Critical Care, Department of Medicine, Washington University School of
Alma R. Bicknese's present address is Department of Neurology, State
Address reprint requests to Alan L. Pearlman, Department of Cell Biology-Box 8228, Washington University School of Medicine, 660 South Euclid Avenue, St. Louis, MO 63110.
Medicine, St. Mo 63110.
proteoglYcans (CSPGs; Sheppard et al., lggl; Bicknese et University ofNew York, Stony Brook, Ny 11794, 1994) and Other extracellular matrix (ECM) cornPo- nents (Derer and Nakanishi, 1983; DelPech and DelPech, 1984; Stewart and Pearlman, 1987; Chun and Shatz, 1988;
o 1995 WILEY-LISS, INC.

616 B. MILLER ET AL.
Sheppard et al., 1991). In contrast, efferent cortical plate axons cross the CSPG-rich subplate and then turn abruptly to follow a trajectory below that of the thalamic afferents, which contain far less CSPG (Bicknese et al., 1994). These relationships suggest that CSPGs and other ECM compo- nents may be parts of the molecular substrate that distin- guishes between afferent and efferent pathways in the developing cortex.
A variety of experimental studies indicate that CSPGs have a role in defining axonal pathways, but the evidence regarding the nature of that role is conflicting (Letourneau et al., 1994). CSPGs are associated with structures that are inhibitory boundaries for axonal growth in the peripheral nervous system (Fichard, 1991; Oakley and Tosney, 19911, and certain CSPGs inhibit neurite Outgrowth in vitro (Snow et al., 1990; McKeon et al., 1991; Bovolenta et al., 1993). In contrast, other studies have shown that CSPGs promote neurite outgrowth in tissue culture (Iijima et al., 1991; Lafont et al., 19921, and our analysis of the relation- ship of axonal trajectories to the distribution of CSPGs indicates that they are not inhibitory in the developing cortex (Bicknese et al., 1994).
CSPGs, like all proteoglycans, have two major structural components, a core protein and one or several covalently linked glycosaminoglycan side chains (Esko, 1991; Jackson et al., 1991; Kjellen and Lindahl, 1991). Although CSPGs are classified on the basis of the repeating disaccharides that make up the side chains, the number, length, and sulfation pattern of these chains vary considerably from one core protein to another (Gallagher, 1989). In some instances, it is apparently the glycosaminoglycan side chains that affect neurite outgrowth (Snow et al., 1990; Fichard, 1991; McKeon et al., 1991; Oakley and Tosney, 1991; Lafont et al., 1992; Bovolenta et al., 19931, whereas, in other cases, the core protein has the active role (Iijima et al., 1991; Lafont et al., 1992).
The structural diversity of proteoglycans and their ability to interact with adhesion molecules and other matrix components (Gallagher, 1989; Ruoslahti, 1989; Rauch et al., 1992; Grumet et al., 1993; Stipp et al., 1994) make them particularly interesting candidate molecules for roles in axon guidance. In prior studies defining the distribution of CSPGs in the developing nervous system, we and others (Sheppard et al., 1991; Snow et al., 1991; Brittis et al., 1992; Pindzola et al., 1993; Bicknese et al., 1994) have taken advantage of a monoclonal antibody that recognizes anti- genic features common to many chondroitin sulfate side chains (Avnur and Geiger, 1984; Sorrel1 et al., 1993). However, there are a variety of CSPGs present in the developing brain (Oohira et al., 1988; Gowda et al., 1989; Herndon and Lander, 1990; Zaremba et al., 1990; Rauch et al., 1991, 1992); an antibody directed against features common to chondroitin sulfate side chains would not be expected to distinguish among them. In the present study, we sought to achieve greater molecular specificity in identi- fying the CSPGs present during major cortical developmen- tal events such as the establishment of axonal pathways. We used a panel of antibodies directed against recently identified core proteins or particular sulfation forms of glycosaminoglycan side chains in an immunohistochemical analysis of their spatial and temporal distributions during neocortical development in the rat. A preliminary report of this work has been presented elsewhere (Miller et al., 1992).
MATERIALS AND METHODS Experimental animals
Prior studies from this laboratory (Sheppard et al., 1991; Bicknese et al., 1994) have examined the distribution of CSPGs in the developing neocortex of the mouse with an antibody that recognizes chondroitin sulfate glycosaminogly- can chains (CS-56; Sigma, St. Louis, MO; Avnur and Geiger, 1984). Preliminary immunolabeling with several of the monoclonal antibodies to core proteins used in the current study indicated that they do not immunolabel sections of cortex from the mouse, but they do recognize rat. We, therefore, carried out this analysis in the rat, because both the major steps in cortical development and the distribution of immunolabeling with the CS-56 anti- body are quite similar in rat and mouse. We examined the cerebral cortex of a total of 67 Sprague-Dawley rats (Sasco, Inc., Omaha, NE) at 5 embryonic (E13-16, E18) and 7 postnatal ages (PO, P4, P7, P10, P14, P21, and adult). The day of conception, as defined by the appearance of a vaginal plug after an overnight mating, was designated EO.
Immunohistochemistry Timed-pregnant females were killed by C02 inhalation,
followed by cervical dislocation; embryonic rats were re- moved via caesarian section. Early fetal animals (E13-15) were fixed by immersion (4% paraformaldehyde in phos- phate-buffered saline; PBS). The brain was removed from later embryos (El6 and E18) and then immersed in fixative. Young postnatal (PO-3) animals were anesthetized by hypothermia and perfused briefly with PBS followed by fixative. Older postnatal animals (P4+) and adults were anesthetized with chloral hydrate (0.1 ccil0 g, i.p.) and then perfused briefly with PBS followed by fixative. The brains were postfixed for 2-18 hours, washed in PBS for 30 minutes, immersed in 30% sucrose overnight (4"C), embed- ded in M1 (Lipshaw, Pittsburgh, PA), then frozen in liquid nitrogen. In some cases, brains were embedded and frozen without fixation.
Coronal and sagittal sections were cut on a cryostat at 18 ym and mounted on glass slides. Sections were hydrated with PBS for 5-10 minutes, blocked in 2% fish gelatin (Sigma) in PBS for 30-60 minutes, then incubated with the primary antibody in 2% fish gelatin at 4°C overnight or for 1-2 hours at room temperature. After six washes with PBS, the sections were incubated for 1 hour at room temperature with a fluorescein isothiocyanate (F1TC)- or tetramethyl rhodamine isothiocyanate (TR1TC)-conjugated secondary antibody (goat anti-mouse or goat anti-rabbit, Jackson Immunoresearch, West Grove, PA; or rabbit anti-rat, Cap- pel, Durham, NC) diluted in 2% fish gelatin. Adjacent sections were stained with hematoxylin. Selected immuno- labeled sections were counterstained with bisbenzimide (Sigma) for cytoarchitectonic identification. Immunola- beled sections were washed with PBS, then coverslipped with 0.01% p-phenylenediamine (Sigma) in glycerin (Johnson and Nogueira-Araujo, 1981) or Vectashield (Vec- tor Labs, Burlingame, CA) and sealed with nail polish.
Some antibodies required permeabilization of the tissue with methanol or Triton X-100 for effective immunolabel- ing. Sections were either immersed in methanol (-2O"C, 10 minutes) and air dried prior to hydration in PBS or 0.2% Triton X-100 (Sigma) was included in all incubations and 0.1 M Tris acetate buffer (pH 7.3) was used in place of PBS. Sections were examined and photographed with an epifluo-

CSPGS IN THE NEOCORTICAL SUBPLATE 617
rescence microscope (Nikon). Control experiments included 1) incubation with the diluent (2% fish gelatin in PBS) without the primary antibody, 2) incubation with a mono- clonal antibody (MEL 14) to a lymphocyte surface molecule (Gellatin et al., 1983), and 3) incubation with normal rabbit serum in place of the primary antiserum.
Antibodies We used monoclonal antibodies and polyclonal antisera
(Table 1) directed against several identified CSPG core proteins isolated from the nervous system and other sites. In addition, monoclonal antibodies against unsulfated chon- droitin and the 4- and 6-sulfated forms were employed to characterize better the CSPGs present during neocortical development. The CS-56 antibody (Avnur and Geiger, 1984) was purchased from Sigma, and the antibody to unsulfated chondroitin (0-CS, antibody 1B5) was pur- chased from ICN (Costa Mesa, CAI. Other antibodies were generously provided by the following investigators (for references, see Table 1): 1D1 (neurocan), 3F8 (phospha- can), 5F3, and 3H7, Renee K. Margolis (State University of New York Health Science Center, Brooklyn, NY); 281-2 (Syndecan-1) and MEL 14, Kevin Hooper (Harvard Medical School, Boston, MA); PGM1, Jean-Paul Zanetta (Centre de Neurochemie du CNRS, Strasbourg, France); chondroitin 4-sulfate (4-CS; antibody 2B6) and 6-sulfate (6-CS; anti- body 3B3), Bruce Caterson (University of North Carolina, Chapel Hill, NC); 2E9 and 5 C l l (Aggrecan), Thomas F. Kresina (Brown University, Providence, RI); NG2, William B. Stallcup (La Jolla Cancer Research Foundation, La Jolla, CA); 2D6, John R. Couchman (University of Alabama at Birmingham, Birmingham, AL); 9.2.27, Ralph A. Reisfeld (Scripps Research Institute, La Jolla, CA); IM7.8.1 (CD44), Robert Hyman (Salk Institute, San Diego, CAI; KM 201 (CD44), Paul Kincade (Oklahoma Medical Research Foun-
dation, Oklahoma City, OK); N-syndecan, David J. Carey (Geisinger Clinic, Danville, PA); and Decorin, Hans Kresse (University of Munster, Munster, Federal Republic of Germany).
Chondroitinase ABC treatment Chondroitinase ABC (chABC; Sigma; 0.25 units/ml 0.1
M Tris-acetate buffer, pH 7.3) was applied to a series of sections for 30-60 minutes at 37°C prior to application of primary antibodies to remove the major portion of chondroi- tin sulfate and dermatan sulfate glycosaminoglycan chains. Elimination of immunolabeling with a particular antibody by chABC indicates that the antibody recognizes an epitope that includes the glycosaminoglycan chain (e.g., CS-56; Table 1). Epitopes on the core protein are unaffected by chABC treatment (e.g., neurocan; Table 1). The terminal disaccharides recognized by the antibodies to unsulfated and 4- and 6-sulfated chondroitin require chABC treatment (Caterson et al., 1990), but the antibody to chondroitin 4-sulfate (2B6) also recognizes a cytoplasmic component without chABC treatment (see Results).
RESULTS The distribution of immunolabeling for various CSPG
forms will be described at several points in neocortical development, with emphasis on early stages. The initial stage in neuronal migration and layer formation occurs when the first postmitotic neurons migrate out of the ventricular zone to form the preplate (Rickmann et al., 1977; also called the primordial plexiform layer, Marin- Padilla, 1971). Arriving cortical plate cells then split the preplate into the marginal zone above and the subplate below (Marin-Padilla, 1971; Raedler and Raedler, 1978;
TABLE 1. Monoclonal Antibodies and Polyclonal Antisera Directed Against Several Identified CSPG Core Proteins Isolated From the Nervous System and Other Sites
Effect of Antibody Antigen CoreiGAG Antigen Source Host' Dilution ch.4BC References
Laminar in early cortex ID1 Neurocan Core Rat brain Mouse 1:200 No effect Rauch et al., 1991,1992 CS-56 Chondroitin sulfate GAG Chick gizzard Mouse M O O Eliminated Avnur and Geiger, 1984 1B5 0-Sulfate GAG Rat chondrosarcoma Mouse 1:250 Required Caterson et al., 1985 2B6 4-Sulfate GAG Bovine nasal cartilage Mouse 1:1,000 Enhanced Catersonet d., 1981 3B3 6-Sulfate GAG Rat chondrosarcoma Mouse 1:500 Required Caterson et d., 1985
Diffuse in early cortex 3F8 Phosphacan Core Rat brain Mouse 1 : l O O No effect Rauch et d ,1991; Maurel et al.,
5F3 - Core Rat brain Mouse 1:20 No effect Rausch et d., 1991 281-2 Syndecan-l Core Mouse mammary gland Rat 1:lOO No effect Solursh et d., 1990 N-Syndecan Syndecan-3 Core Recombinant fragment Rabbit 1:50 No effect Carey et al., 1992
1994
Labels axon PGMl - Core Rat brain Rabbit 1:500 No effect Normand et d., 1988
Labels non-neuronal elements IM 7.8.1 CD44 Core Mouse hematopoietic cells Rat 1:l No effect Trowbridge et al., 1982 KM 201 CD44 Core Mouse bone marrow Rat 1: l No effect Miyake et al., 1990 NG2 NG2 Core Rat brain Mouse 1:1,000 No effect Levine and Card, 1987 2D6 - Core Rat placenta Mouse 1 : l O O No effect McCarthy et al., 1989 3H7 - Core Rat brain Mouse 1:lOO No effect Rauch et al., 1991
No immunolabe12 9.2.27 - Core Human melanoma Mouse - - Schrappe et al.. 1991 2E9 and5C11 Aggrecan (AIDlD1) Core Rabbit cartilage Mouse - - Kresina and Malemud, 1986 Decorin Decorin Core Human fibroblast Rabbit - - Glossl et d., 1984
'Antibodies derived from mouse or rat are monoclonal; antisera derived from rabbit are polyclonal. *Antibodies were used in a wide range of dilutions from 1:lO to 1:500 on h e d and unfixed tissue of various ages with or without chondroitinase ARC.

618 B. MILLER ET AL.
Luskin and Shatz, 1985; Bayer and Altman, 1991; Wood et al., 1992). Immunolabeling for chondroitin sulfates is very prominent in the preplate and its derivatives, the subplate and marginal zone (Sheppard et al., 1991; Bicknese et al., 1994). Axons of preplate cells traverse the CSPG-rich preplate obliquely to enter the forming intermediate zone, which contains much less CSPG (Bicknese et al., 1994). Efferent axons of cortical plate neurons cross the CSPG- rich subplate directly, then turn abruptly to extend through the intermediate zone (Bicknese et al., 1994). In contrast, growing axons of afferent thalamic projections travel di- rectly within the subplate (Catalano et al., 1991; Erzu- rumlu and Jhaveri, 1992; Kageyama and Robertson, 1993; Miller et al., 1993; Bicknese et al., 1994): double labeling for CSPGs and thalamic afferents demonstrates that the intra- cortical trajectory of the afferents is centered on this CSPG-rich region (Bicknese et al., 1994).
As the cortex matures, postmitotic cells continue to migrate from the ventricular zone to the top of the cortical plate, while older cells at the bottom of the cortical plate differentiate into mature neurons (Caviness, 1982; Bayer and Altman, 1991). Coincident with the maturation of the lower cortical plate, thalamic axons grow into the differenti- ating layers (Catalano et al., 1991; Kageyama and Robert- son, 1993; Miller et al., 1993; Bicknese et al., 1994). At approximately the same time, immunolabeling for CSPGs appears in these maturing layers and gradually extends upward as the lower aspect of the cortical plate differenti- ates (Bicknese et al., 1994; see below). This deep-to- superficial maturation continues until all layers of the neocortex are distinct, which occurs at approximately the end of the first postnatal week in the rat (Kageyama and Robertson, 1993).
We have grouped our findings into five sections based on the distribution of immunolabeling during cortical develop- ment: 1) comparison of the distribution of the central nervous system (CNS)-specific proteoglycan neurocan (Rauch et al., 1991, 1992) with that of chondroitin sulfate glycosaminoglycan chains identified with the CS-56 anti- body; neurocan is the only CSPG core protein in our panel that we found to have a distinct laminar distribution during early stages and to be particularly prominent in the sub- plate; 2) distribution of chondroitin disaccharides that are either unsulfated or sulfated in the 4 or 6 position of N-acetylgalactosamine in the oligosaccharide stub that remains after c U B C digestion (Caterson et al., 1990); 3) proteoglycan core proteins that do not have a laminar pattern but, instead, are widely distributed in early stages; 4) CSPGs that are associated with nonneuronal structures such as blood vessels or with axons in late development; and 5 ) distribution of CSPGs in the barrel field of the postnatal somatosensory cortex, where it has been previously demon- strated that tenascin and the cytotactin binding proteogly- can are more prominent in the barrel walls than in the centers of the barrels (Crossin et al., 1989; Steindler et al., 1989; Jhaveri et al., 1991; Sheppard et al., 1991). Although it is not possible to determine the subcellular localization of proteoglycans with certainty using light microscopic immu- nolabeling, we have attempted to identify labeling of the cytoplasm (see, e.g., Fig. 2B) and to distinguish it from labeling that is membrane-associated or extracellular (see, e.g., Fig. 2A). For convenience, the latter will be grouped together and referred to as peri- or extracellular.
Immunolabeling for chondroitin sulfate glycosaminoglycan chains and neurocan is prominent in the subplate in early cortical
development Neurocan, one of the first proteoglycans of brain to be
cloned and sequenced (Rauch et al., 19921, is identified by the 1D1 antibody (Rauch et al., 19911, which we used in our analysis of its distribution during cortical development. For comparison, we include a description of immunolabeling for chondroitin sulfate side chains with the CS-56 antibody (Avnur and Geiger, 1984), which was used in prior studies in this laboratory to establish the presence of immunolabel- ing for CSPGs in the developing cortex (Sheppard et al., 1991) and the relationship of growing efferent and afferent axons to these proteoglycans (Bicknese et al., 1994).
Prior to the formation of the preplate, there is scattered immunolabeling for chondroitin sulfate chains with CS-56 in the ventricular zone that is somewhat more prominent near the ventricle (not illustrated). As the first postmitotic neurons form the preplate, immunolabeling becomes quite prominent in that layer and decreases in the ventricular zone (Fig. 1A; see also Sheppard et al., 1991; Bicknese et al., 1994). Immunolabeling is also present in association with blood vessels (not illustrated) and is very intense in the pia-arachnoid. Throughout the period when immunolabel- ing for CS-56 is present, it appears to be extracellular. Immunolabeling for neurocan with 1D1 is sparse and homogeneous in the ventricular zone prior to preplate formation (not illustrated). Labeling for neurocan increases slightly in the preplate as it forms (Fig. 1B) but is delayed in comparison with CS-56 labeling. Neurocan is also present in association with blood vessels and in the pia-arachnoid but is much less prominent in the latter than the CS-56 antigen. Most immunolabeling with 1D1 appears to be peri- or extracellular, but intracellular labeling is also occasion- ally evident.
The arrival of cortical plate neurons divides the preplate into the marginal zone above and the subplate below; immunolabeling with CS-56 is also divided, resulting in intense labeling in the marginal zone and subplate (Fig. 1C). Very little CS-56 immunolabelingis present among the cells of the cortical plate. There is moderate labeling in the upper intermediate zone, whereas labeling in the ventricu- lar zone is nearly undetectable. Immunolabeling for neuro- can increases in the subplate as it is formed and is, therefore, prominent prior to the arrival of the first cortical axons that takes place well after subplate formation (De Carlos and O’Leary, 1992; Erzurumlu and Jhaveri, 1992; Miller et al., 1993; Bicknese et al., 1994). As with the immunolabeling with CS-56, it is also present at reduced levels in the intermediate zone and nearly absent from the cortical plate and ventricular zone (Fig. 1D). There is a further increase in the immunolabeling for neurocan in the subplate as the cortical plate becomes thicker and begins to differentiate at its lower border (Fig. 1F). In contrast, the intensity of labeling for CS-56 declines somewhat in the subplate (Fig. 1E). Immunolabeling with CS-56 in the pia is greatly reduced at this stage and remains quite low through- out the remainder of cortical development (Fig. lE,G).
Immunolabeling with both CS-56 and 1D1 becomes evident throughout the cortical layers as they differentiate from below upwards in the early postnatal period. The cell-dense cortical plate remains distinct, with very little immunolabel with either antibody (Fig. lG,H). Immunola-

Fig. 1. Distribution of chondroitin sulfate side chains and neurocan during neocortical development. Immunofluorescent labeling of chon- droitin sulfate glycosaminoglycan side chains (mab CS-56; top) and the neurocan core protein (mab 1D1; bottom) in adjacent coronal sections of neocortex. A,B: With formation of the preplate (PPZ stage; E14), CS-56 labels the preplate and pia-arachnoid (PA) intensely, and there is punctate labeling in the ventricular zone (VZ). Immunolabeling for neurocan is more prominent in the preplate than the ventricular zone, but this difference is delayed relative to the CS-56 antigen. Labeling for neurocan is also associated with the pia-arachnoid and blood vessels (arrow) but is much less prominent than CS-56 labeling. C,D: Cortical plate formation (CP stage early; El61 divides the preplate into the marginal zone (MZ), above, and the subplate (SP), below. Strong immunolabeling with CS-56 continues to be associated with preplate cells in their new locations; neurocan labeling has increased substan-
tially in the subplate. Less prominent immunolabeling for both the CS-56 antigen and neurocan is evident in the upper intermediate zone (IZ); very little is present in the cortical plate or ventricular zone. E,F: The cortical plate has begun to differentiate at its lower border (CP stage late; E18). CS-56 labeling has decreased in the lower subplate, whereas neurocan labeling has increased throughout the subplate. Labeling for the CS-56 antigen has declined substantially in the pia-arachnoid and remains low throughout the remainder of develop- ment. G,H: During early postnatal development of cortex (Ctx stage early; P4), labeling for both the CS-56 antigen and neurocan is evident throughout the differentiating cortical layers 5 and 6 and is sparse in the thin remnant of cell-dense cortical plate. Scale bars = 50 pm in A (also applies to B), 100 +m in C (also applies to C-F), 100 K r n in G (also applies to HI.

620 B. MILLER ET AL.
beling in the emerging cortex is relatively homogeneous, without a dense band of labeling in the subplate (Fig. lG,H). Immunolabeling for both the CS-56 antigen and neurocan declines in the centers of the whisker barrels in the parietal somatosensory cortex by P7, but the decline in CS-56 labeling proceeds more rapidly than the decline in neurocan (see below). Immunolabeling for both continues to be present in the remainder of the cortex through P10, when a decrease in labeling begins that is nearly complete by P14. Immunolabeling for the CS-56 antigen is undetect- able in the adult, whereas labeling for neurocan is evident intracellularly in some cells throughout the cortex and the white matter. Labeled cells are quite small (approximately half the diameter of granule cells in layer IV), suggesting that they are glia rather than neurons (not illustrated).
Thus, although there are many similarities in the distri- bution of the CS-56 antigen and neurocan, there are also several differences. Immunolabeling for both is evident in the preplate, but CS-56 is much more prominent. Neurocan becomes prominent in the layers derived from the preplate, the marginal zone, and, especially, the subplate shortly after these layers are formed by the intervention of the cortical plate. Labeling for CS-56 is very intense in the pia-arachnoid in the early stages, then declines, whereas neurocan is never prominent in the pia-arachnoid. 1D1 immunolabels a population of small cells in the cortex and the white matter of the adult rat that are probably glia and that are not labeled by CS-56. These observations suggest that neurocan makes up some, but not all, of the CSPGs identified by CS-56 and that neurocan may also be present without the CS-56 antigen.
Distribution of chondroitin and the 4- and &sulfated isomers of chondroitin sulfates
Differing isomers of CS glycosaminoglycans result from ester sulfation at distinct sites in the repeating disaccharide units. Chondroitinase ABC digestion removes the majority of the glycosaminoglycan chain, leaving oligosaccharide stubs that are either unsulfated or sulfated in the 4 or 6 position of N-acetylgalactosamine (Caterson et al., 1990). To determine the distribution of three of these isomers in the developing cortex, we used monoclonal antibodies that recognize distinct epitopes in the CS oligosaccharide stub that results from chABC digestion.
Monoclonal antibody 3B3 rec- ognizes epitopes that contain a terminal nonreducing hex- uronic acid adjacent to a 6-sulfated N-acetylgalactosamine (Caterson et al., 1990). The distribution of 6-sulfated CS isomers (6-CS) in the developing cerebral cortex is initially quite similar to that of the CS-56 antigen, including a predominantly extracellular distribution. For example, dur- ing early cortical plate formation (E15), there is intense labeling for 6-CS in the subplate and pia (Fig. 2A), as there is with CS-56 (Fig. 1C). The pattern of labeling for 6-CS begins to differ from that for CS-56 after the first postnatal week, when immunolabeling for CS-56 rapidly diminishes, whereas labeling for 6-CS persists until 2 weeks after birth. In addition, although the CS-56 antibody does not label adult brain, pericellular immunolabeling for 6-CS is appar- ent in association with a small subset of cortical neurons that are also labeled by the antibody to nonsulfated isomers (see below; see also Bertolotto et al., 1991). Thus, the spatial and temporal patterns of immunolabeling for 6-sul- fated CS isomers are similar, but not identical, to those of the CS-56 antibody.
6-Sulfated disaccharides.
4-Sulfated disaccharides. Monoclonal antibody 2B6 identifies an epitope that includes a terminal hexuronate residue adjacent to a 4-sulfated N-acetylgalactosamine (Ca- terson et al., 1990). The 4-sulfated antigen (4-CS) is prevalent early in brain development and is more broadly distributed than the CS-56 antigen or the 6-CS and unsul- fated isomers. Immunolabeling is both cytoplasmic and pericellular (Fig. 2B). Cells in the upper ventricular zone are more intensely labeled than those in the lower ventricu- lar zone (Fig. 2B). Immunolabeling is less prominent in the preplate than in the ventricular zone (not illustrated) but becomes prominent in the subplate as the cortical plate forms (Fig. 2B). Some cells in the cortical plate also show evidence of cytoplasmic labeling (Fig. 2B). Pericellular labeling requires chABC pretreatment, whereas cytoplas- mic labeling i s evident without chABC. This result differs from results of a prior study (Bertolotto et al., 1990) that found no cytoplasmic immunolabeling with or without chABC. Differences in tissue fixation between the two studies probably account for the differences in immunolabel- ing.
Immunolabeling for 4-CS is diffuse throughout the neuro- pi1 of the cortex as it matures. Labeling occurs in all layers of the developing cortex but is most intense in the differen- tiated region just below the cell-dense cortical plate. By P10, immunolabel for 4-CS is homogeneous in all layers of the cortex except for a reduction of label in the centers of the barrels of the somatosensory cortex (see below). In adults, cytoplasmic immunolabeling is evident in most cells, and the neuropil is diffusely labeled (see also Bertolotto et al., 1990).
An epitope that includes an unsulfated N-acetylgalactosamine adjacent to an unsatu- rated terminal hexuronate residue is recognized by the 1B5 antibody (Caterson et al., 1990). Immunolabeling for this antigen (0-CS), which appears to be primarily extracellular, first becomes evident in the preplate, predominantly in its upper aspect (not illustrated). Labeling for 0-CS is not evident in the ventricular zone. When the cortical plate forms, immunolabeling for 0-CS is very precisely localized to the subplate and marginal zone (Fig. 2C), with only traces of labeling in the cortical plate and upper intermedi- ate zone. The blood vessels and pia are not labeled at any stage.
As with the other isomers, immunolabeling for 0-CS involves the developing cortex as it matures. Labeling gradually diminishes postnatally; by P 14, the immunolabel- ing of the neuropil is almost gone, but pericellular labeling of a small population of cells with neuronal morphology becomes evident at approximately this stage and persists in the adult. Double labeling with immunoglobulin-type- specific second antibodies demonstrates that the same cells are labeled by the antibodies to both 6-CS and 0-CS (see also Bertolotto et al., 1991).
Unsulfated chondroitin.
Immunolabeling for several proteoglycan core proteins is nonlaminar
Several of the antibodies against core proteins of CSPGs in the panel that we tested labeled developing cortex diffusely with little or no indication of a laminar distribu- tion.
The 5F3 CSPG (350-450 kD core protein) was first identified in the rat cerebellum: its distribution overlaps that of neurocan, although it labels Purkinje cells and some astrocytes in addition (Rauch et al., 1991). Immunolabeling
5F3.

CSPGS IN THE NEOCORTICAL SUBPLATE 621
Fig. 2. Distribution of unsulfated and 4- and 6-sulfated isomers of chondroitin sulfates. Immunofluorescent labeling of coronal sections of cortex in the cortical plate stage (E15). A Labeling for 6-sulfated chondroitin disaccharides (mab 3B3) after chondroitinase ABC treat- ment is dense in the subplate (SP) and marginal zone (Mi2 and moderate in the intermediate zone (IZ). B: Labeling for 4-sulfated
isomers (mab 2B6) is more broadly distributed, with some concentra- tion in the subplate and upper ventricular zone. C: The distribution of labeling for unsulfated chondroitin disaccharides (mab 1B5) is almost completely limited to the subplate and marginal zone. Scale bar = 50bm.
for 5F3 in the developing neocortex of the rat is evenly distributed throughout all layers (Fig. 3A). It is primarily intracellular and is most intense at E13-15. At late fetal ages (ElB), immunolabeling begins to diminish in the lower, more mature layers and continues to diminish postnatally until it is nearly undetectable at P7-10. Moder- ate immunoreactivity for 5F3 returns in the adult cortex, where it is homogeneously distributed in the neuropil. 5F3 labels the pia-arachnoid but not the cortical blood vessels.
Syndecan-1 is a cell-surface proteoglycan containing both chondroitin sulfate and heparan sulfate glycosaminoglycans (Brauker et al., 1991; Cizmeci-Smith et al., 1992) that has previously been found in the adult CNS with antibody 281-2 (Corless et al., 1992). We extend these results by demonstrating that immunolabeling for synde- can-1 is present in the neocortical wall throughout pre- and postnatal development. Dense extracellular immunolabel- ing is present in all layers in early stages of development (Fig. 3B). Immunolabeling is also evident along axons in the intermediate zone and the internal capsule. Although there is some reduction in immunolabeling in the ventricular zone near the time of birth (PO), immunolabeling remains relatively high for the first week postnatally, then declines substantially, and is barely detectable by P21. In the adult brain, immunolabeling for syndecan-1 increases slightly. Immunolabeling i s evident in association with the pia- arachnoid, but not the blood vessels, at all ages.
N-syndecan (syndecan-3) is a transmem- brane heparan sulfate proteoglycan. Unlike syndecan, it
Syndecan-1.
N-syndecan.
contains no chondroitin sulfate glycosaminoglycans. It is included in this study because it is a member of the syndecan family that is present in large amounts in the nervous system (Carey et al., 1992). The spatial and temporal distributions of immunostaining for N-syndecan are similar to those of syndecan-1. Pericellular immunola- beling is uniformly distributed in all laminae (Fig. 3C) and is also associated with axons in the intermediate zone. Labeling remains prominent in the first postnatal week, decreases considerably by P10, and is faint but distinct in adult cortex. The pia-arachnoid, but not the blood vessels, is labeled at all ages.
Phosphacan. The 3F8 antibody recognizes a large CSPG core protein first identified in the cerebellum of the rat (Rauch et al., 1991). The recently determined primary sequence of the 3F8 proteoglycan, now called phosphacan, indicates that it is an extracellular variant of a receptor- type protein tyrosine phosphatase (Maurel et al., 1994). In the neocortex, immunolabeling for phosphacan is mini- mally present in the ventricular zone; it increases with formation of the preplate and cortical plate. During most stages of neocortical development, phosphacan immunola- beling appears to be primarily extracellular and is homoge- neously distributed in all cortical layers (Fig. 3D), with minor exceptions. There is slightly less labeling in the cortical plate when it first forms (E15; Fig. 3D); by E18, labeling has increased in the cortical plate and has de- creased somewhat in the subplate and intermediate zone. Immunolabeling for phosphacan decreases first in the

622 B. MILLER ET AL.
Fig. 3. Nonlaminar distribution of proteoglycan core proteins. Immunofluorescent laheling of coronal sections of cortex at the cortical plate stage (E15-16) labeled with antibodies to the core proteins of two chondroitin sulfate proteoglycans, 5F3 (A) and phosphacan (D); a heparan sulfate proteoglycan, N-syndecan (C); and syndecan-1 (B), which contains both chondroitin sulfate and heparan sulfate. Labeling
centers of presumptive barrels of somatosensory cortex (P7; see below). In adults, granular phosphacan labeling is evident in the cortical neuropil and subcortical white matter, and there is some labeling of blood vessels. We found no immunolabeling of the pia-arachnoid with 3F8 at any age.
Immunolabeling of nonneuronal and primarily axonal CSPGs
Three of the antibodies in the panel label only nonneuro- nal structures, and one labels axons in major white matter tracts.
206. The 2D6 antibody recognizes a CSPG core protein (160 kD) associated with basement membranes (McCarthy et al., 1989; Couchman et al., 1990). We found immunolabel- ing for this CSPG only on blood vessels (Fig. 4A) in the cortex from E l 3 through adult stages. The 2D6 antigen is sensitive to paraformaldehyde fixation; labeling is best on unfixed tissue.
This CSPG has previously been demonstrated on the cell membrane of presumptive 0-2A glial precursor cells and protoplasmic astrocytes in the cerebellum and optic nerve (Levine and Card, 1987; Stallcup and Beasley, 1987; Levine et al., 1993). From E l 6 through adult stages, the NG2 monoclonal antibody mixture labels small cells (Fig. 4B) in neocortex that appear to be nonneuronal by morpho- logical criteria and are similar in appearance to the NG2- positive cells that have been identified as 02-A precursors and protoplasmic astrocytes (Levine and Card, 1987). In addition, from the earliest stages of neocortical develop- ment to approximately P10, the NG2 monoclonal antibody
NG2.
is relatively uniform for all of these, with the exception of phosphacan, which is slightly less intense in the cortical plate a t this age but increases later (see text). The size differences between panels reflect slight differences in the cortical location selected for illustration: the distribution of a given proteoglycan does not vary with cortical location at this stage. Abbreviations as in Figures 1 and 2. Scale bar = 50 pm.
mixture labels blood vessels in the pia and cortex (Fig. 4B) but does not do so after P10.
CD44 is a cell-surface proteoglycan of hematopoi- etic and other cell types (Trowbridge et al., 1982). At least one form of CD44 is thought to be involved in cell motility (Faassen et al., 1992; Thomas et al., 1992). We used two different antibodies to CD44 (IM 7.8.1 and KM 201); neither labels embryonic (EX) or early postnatal (PO) cortex, but both label cells in adult subcortical white matter with the morphology of astrocytes (Fig. 4C). Some of these cells have processes that appear to contact blood vessels. Both antibodies label the same population of cells by morphological criteria, and both require unfixed tissue.
Immunolabeling for this large CSPG has been found on a subset of axons in the brain of the adult rat (Normand et al., 1988). We find that the PGMl antibody labels axons in the developing brain as well but does not label cell bodies. Many axons are labeled as early as E14, and they are prominent in the internal capsule by El6 (Fig. 4D). Most of the labeled axons in the neocortex appear to be of thalamic origin, because they are primarily in the subplate and upper intermediate zone, although some labeled axons that are probably cortical efferents are also evident in the lower intermediate zone. The antibody also labels axons in several other locations including the optic tract, corticospinal tract, and hippocampus.
The proteoglycans (61-105 kD core glycoproteins) recognized by the 3H7 antibody were initially studied in rat cerebellum (Rauch et al., 1991). In the postnatal and adult cerebellum, 3H7 labels most layers (Rauch et al., 19911, but the distribution of this antigen in the cerebral cortex is
CD44.
PGMZ.
3H7.
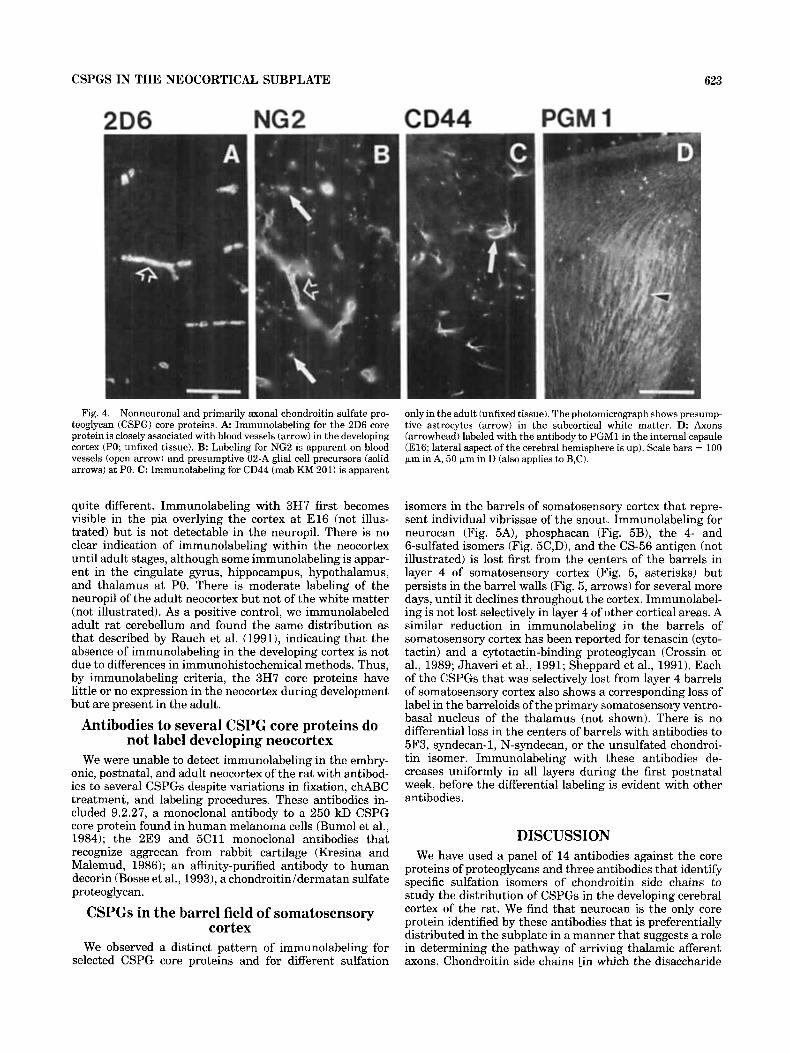
CSPGS IN THE NEOCORTICAL SUBPLATE 623
Fig. 4. Nonneuronal and primarily axonal chondroitin sulfate pro- teoglycan (CSPG) core proteins. A Immunolabeling for the 2D6 core protein is closely associated with blood vessels (arrow) in the developing cortex (PO; unfixed tissue). B: Labeling for NG2 is apparent on blood vessels (open arrow) and presumptive 02-A glial cell precursors (solid arrows) at PO. C: Immunolabeling for CD44 (mab KM 201) is apparent
only in the adult (unfixed tissue). The photomicrograph shows presump- tive astrocytes (arrow) in the subcortical white matter. D: Axons (arrowhead) labeled with the antibody to PGMl in the internal capsule (E16; lateral aspect of the cerebral hemisphere is up). Scale bars = 100 IJ-m in A, 50 IJ-m in D (also applies to B,C).
quite different. Immunolabeling with 3H7 first becomes visible in the pia overlying the cortex a t E l 6 (not illus- trated) but is not detectable in the neuropil. There is no clear indication of immunolabeling within the neocortex until adult stages, although some immunolabeling is appar- ent in the cingulate gyms, hippocampus, hypothalamus, and thalamus at PO. There is moderate labeling of the neuropil of the adult neocortex but not of the white matter (not illustrated). As a positive control, we immunolabeled adult rat cerebellum and found the same distribution as that described by Rauch et al. (1991), indicating that the absence of immunolabeling in the developing cortex is not due to differences in immunohistochemical methods. Thus, by immunolabeling criteria, the 3H7 core proteins have little or no expression in the neocortex during development but are present in the adult.
Antibodies to several CSPG core proteins do not label developing neocortex
We were unable to detect immunolabeling in the embry- onic, postnatal, and adult neocortex of the rat with antibod- ies to several CSPGs despite variations in fixation, chABC treatment, and labeling procedures. These antibodies in- cluded 9.2.27, a monoclonal antibody to a 250 kD CSPG core protein found in human melanoma cells (Bumol et al., 1984); the 2E9 and 5 C l l monoclonal antibodies that recognize aggrecan from rabbit cartilage (Kresina and Malemud, 1986); an affinity-purified antibody to human decorin (Bosse et al., 19931, a chondroitiddermatan sulfate proteoglycan .
isomers in the barrels of somatosensory cortex that repre- sent individual vibrissae of the snout. Immunolabeling for neurocan (Fig. 5A), phosphacan (Fig. 5B), the 4- and 6-sulfated isomers (Fig. 5C,D), and the CS-56 antigen (not illustrated) is lost first from the centers of the barrels in layer 4 of somatosensory cortex (Fig. 5, asterisks) but persists in the barrel walls (Fig. 5, arrows) for several more days, until it declines throughout the cortex. Immunolabel- ing is not lost selectively in layer 4 of other cortical areas. A similar reduction in immunolabeling in the barrels of somatosensory cortex has been reported for tenascin (cyto- tactin) and a cytotactin-binding proteoglycan (Crossin et al., 1989; Jhaveri et al., 1991; Sheppard et al., 1991). Each of the CSPGs that was selectively lost from layer 4 barrels of somatosensory cortex also shows a corresponding loss of label in the barreloids of the primary somatosensory ventro- basal nucleus of the thalamus (not shown). There is no differential loss in the centers of barrels with antibodies to 5F3, syndecan-1, N-syndecan, or the unsulfated chondroi- tin isomer. Immunolabeling with these antibodies de- creases uniformly in all layers during the first postnatal week, before the differential labeling is evident with other antibodies.
DISCUSSION We have used a panel of 14 antibodies against the core
proteins of proteoglycans and three antibodies that identify specific sulfation isomers of chondroitin side chains to study the distribution of CSPGs in the developing cerebral
CSPQ in the barrel field of somatosensory cortex
cortex of the rat. We find that neurocan is the i n l y core protein identified by these antibodies that is preferentially distributed in the subdate in a manner that suggests a role
We observed a distinct pattern of immunolabeling for selected CSPG core proteins and for different sulfation
in determining the pathway of arriving t h a l a z c afferent axons. Chondroitin side chains [in which the disaccharide

624 B. MILLER ET AL.
Fig. 5 . Immunolabeling for specific CSPGs declines first in the centers of vibrissal barrels in somatosensory cortex. Immunofluorescent labeling of coronal sections of somatosensory cortex P7-10. Immunolabeling for neurocan (A), phosphacan (B), 6-CS ( C ) , and 4-CS (D) is reduced in the barrel hollows (asterisks) o f somatosensory cortex before it declines in the barrel walls (arrows). Scale bar = 100 bm.
that remains after chABC digestion is either unsulfated or chondroitins initially, but immunolabeling for all three sulfated in the 6 position of the N-acetylgalactosamine becomes evident as cortical neurons mature. Three chondroi- residue (Caterson et al., 1990)] also have an initially tin sulfate or chondroitiniheparan sulfate proteoglycans restricted distribution. Immature cortical plate neurons do (syndecan-1, 5F3, and phosphacan), one heparan sulfate not express neurocan or the 6-sulfated and unsulfated proteoglycan (N-syndecan), and 4-sulfated disaccharide

CSPGS IN THE NEOCORTICAL SUBPLATE
stubs on CSPG core proteins are diffusely distributed in early cortical development and, thus, are not likely to play a role in defining early axonal tracts. The vibrissal barrel field of somatosensory cortex is unique among cortical areas in its pattern of CSPG expression. Immunolabeling for neuro- can, phosphacan, and the 4- and 6-sulfated isomers of chondroitin sulfate is lost first from the centers of barrels and, later, from the barrel walls, when labeling is also decreasing in other cortical areas.
Neurocan and the intracortical pathway for thalamic afferent axons
The temporal and spatial distributions of neurocan sug- gest that it is a major component of the CSPGs we previously identified in the subplate trajectory of thalamo- cortical axons (Sheppard et al., 1991; Bicknese et al., 1994). Neurocan becomes prominent in the subplate, as it is delineated by the insertion of the cortical plate into the preplate and, thus, before thalamic axons approach the cortical wall (Bicknese et al., 1991, 1994; Catalan0 et al., 1991; Miller et al., 1993). Immunolabeling for neurocan remains high in the subplate during the period when thalamic axons are extending along it. Neurocan has re- cently been shown to have an identical distribution when labeled with a different monoclonal antibody (Oohira et al., 1994). Neurocan is, therefore, a candidate molecule for delineating the intracortical pathway of thalamocortical axons and distinguishing it from the subjacent pathway for cortical efferent axons.
The complete coding sequence for neurocan has recently been defined (Rauch et al., 1992); the deduced amino acid sequence strengthens the prospect that it has a role in cell-cell or cell-substrate interactions. In the 168 kD, full- length form of the neurocan core protein that is prominent in early postnatal brain, the N-terminal portion has a hyaluronic acid-binding region, an immunoglobulin-like domain, and an RGDS sequence; the C-terminal portion has approximately 60% identity with the C-termini of versican and aggrecan, including two epidermal growth factor-like domains, a lectin-like domain, and a complement regulatory protein-like region (Rauch et al., 1992). The N-terminal half of neurocan is lost by proteolytic processing to form the 68 kD adult form of the core protein (Rauch et al., 1992). Both the full-length and the truncated forms are recognized by the 1D1 antibody. We find that immunolabel- ing for neurocan is predominantly peri- or extracellular and is associated with neurons in the embryonic and early postnatal periods, whereas, in the adult, it appears to be intracellular and is associated with small cells in the cortex and white matter that are probably glia. In the early postnatal cerebellum, electron microscopic immunolabeling with 1D1 is predominantly extracellular, whereas it is intracellular in the adult cerebellum (Rauch et al., 1991). These observations raise the interesting prospect that the full-length, early postnatal form of the core protein is predominantly peri- or extracellular and neuronal, whereas the C-terminal half found in adult brain is predominantly intracellular and glial (Rauch et al., 1991). However, be- cause the 1D1 antibody recognizes both the early postnatal and the adult forms of the neurocan core protein (Rauch et al., 19921, confirmation of this prospect will require antibod- ies that recognize noncorresponding parts of the two forms.
Neurocan binds directly to neurons in assays carried out in vitro: it inhibits the homophilic binding of neurons mediated by N-CAM or Ng-CAM, inhibits the binding of
625
neurons to Ng-CAM-coated substrates, and inhibits neurite outgrowth on Ng-CAM substrates (Grumet et al., 1993; Friedlander et al., 1994). However, neurocan added to in vitro assays may cause these effects by interfering in neuronal interactions with Ng-CAM substrates rather than through an independent inhibitory or repellent mechanism. The former interpretation is more consistent with the presence of neurocan in zones of active axonal extension in both the cerebellum (Rauch et al., 1991) and the cerebral cortex (present study), which indicates that neurocan does not inhibit axonal growth in these locations. The promi- nence of neurocan in the subplate suggests that it is part of a complex ECM matrix (for review, see Sheppard et al., 1991) that may define the pathway for thalamocortical axons, perhaps through the specific interactions with neu- ral cell adhesion molecules that have been demonstrated (Grumet et al., 1993; Friedlander et al., 1994).
Distribution of unsulfated and 4- and 6-sulfated chondroitin disaccharides
We have observed that immunoreactivity for the unsul- fated chondroitin disaccharides that remain on the core protein after chABC digestion is prominent as a band in the subplate soon after it is established. Neurocan, which is also prominent in the subplate at this stage (E16), does not contain detectable unsulfated disaccharides when analyzed in early postnatal (P7) brain (Rauch et al., 1991). On average, full-length neurocan has three chondroitin sulfate chains; approximately 80% of the disaccharides are 4-sul- fated, and 20% are 6-sulfated (Rauch et al., 1991). Thus, our observation that immunoreactivity for unsulfated chon- droitin stubs is prominent in the subplate suggests that core proteins other than neurocan may also be present in the subplate in a restricted fashion or that neurocan in embryonic stages differs from that present later.
The relative composition of the different sulfation forms of chondroitins in the brain changes with development. Both unsulfated and 6-sulfated forms decrease substan- tially after the early postnatal period; 4-sulfated forms become dominant in the adult (Flaccus et al., 1991; Matsui et al., 1993). The transient nature of unsulfated and 6-sulfated forms suggests that they have a special function during early brain development. In addition, the changing composition of chondroitin sulfate forms may reflect devel- opmental changes in the proteoglycan core protein. This has been demonstrated in the case of neurocan, which has 4-sulfated and 6-sulfated glycosaminoglycans on the large, early postnatal core protein, whereas the smaller adult form has a single, 32 kD chondroitin 4-sulfate side chain (Rauch et al., 1991, 1992). Although 6-sulfated isomers have been associated with structures that serve as barriers to axonal extension in the peripheral nervous system (Oakley and Tosney, 1991) and in glial scars after injury in the CNS (McKeon et al., 19911, they are prominent in the subplate pathway of thalamocortical axons (present study) and in the cerebellar white matter (Flaccus et al., 1991) and, thus, are not likely to be inhibitory in either of these sites.
The distributions of immunolabeling with the 1D1 anti- body for neurocan, the 3B3 antibody for 6-sulfated terminal disaccharide stubs, and the CS-56 antibody are similar in many respects and are different from the immunolabeling with antibodies to other core proteins or sulfation forms. Although the epitope recognized by the CS-56 antibody has not been identified with certainty, the antibody is relatively

626 B. MILLER ET AL.
specific for chondroitin sulfates (Avnur and Geiger, 1984) in the interior of glycosaminoglycan chains that have not been treated with chABC (Sorrell et al., 1993). The similar- ity in distribution of immunolabeling with 1D1 and CS-56 suggests that CS-56 is recognizing the glycosaminoglycan chains of neurocan, but this does not exclude the prospect that it is also recognizing chains on other core proteins. The similarity in distribution of immunolabeling with CS-56 and the 3B3 antibody against 6-sulfated terminal disaccha- ride stubs suggests that CS-56 may be recognizing 6-sul- fated disaccharides. Caution is warranted in this conclu- sion, however, because 3B3 recognizes only a 6-sulfated disaccharide unit close to the core protein, and the chondroi- tin sulfate side chains are probably not uniform throughout their length (Lander, 1993; Sorrell et al., 1993).
Functional role of proteoglycans in cortical development
Although the developing neocortex contains a rich vari- ety of CSPGs and other proteoglycans (Margolis et al., 1975; Oohira et al., 1988; Herndon and Lander, 1990; Zaremba et al., 1990; Rauch et al., 1991, 19921, the differential expression of neurocan in the newly formed subplate and marginal zones suggests a functional relation- ship between it and specific developmental processes taking place in these layers. Because the distribution of neurocan (and, probably, other as yet unidentified CSPGs) is associ- ated with the pathway for thalamic afTerents but not that for cortical efferents (Bicknese et al., 1994; present study), we have placed emphasis on the prospect that neurocan in the subplate may be involved in defining the pathway for thalamic afferents. However, neurocan could also play a role in the other important developmental events that involve preplatei subplate cells (reviewed in Allendoerfer and Shatz, 19941, including the formation of synapses with early thalamic axons (Friauf et al., 1990), the later invasion of cortex by thalamic axons (Ghosh et al., 19901, and the reorganization of cortical connections that is responsible for the formation of ocular dominance columns (Ghosh and Shatz, 1992,1993). In addition, the subsequent developmen- tally regulated expression of neurocan and other CSPGs in all cortical layers as well as the variety of functions that have been ascribed to proteoglycans indicate that their role is not limited to pathway definition.
CSPGs have been associated with the promotion or inhibition of a number of developmental processes that involve cell migration (Perris and Johansson, 1987; Morriss- Kay and Tuckett, 1989; Hoffman et al., 1990; Lane and Solurish, 1991) and cell adhesion (Solursh et al., 1990; Jackson et al., 1991). Proteoglycans also play a role in ECM formation by binding other ECM components, including fibronectin, which is prominent in the subplate (Stewart and Pearlman, 1987; Chun and Shatz, 1988; Sheppard et al., 1991). In addition, glycosaminoglycan side chains of heparan sulfate proteoglycans and the core proteins of other proteoglycans bind and sequester a variety of growth factors, and the epidermal growth factor-like repeats pre- sent in several core proteins may also have a role in modulating the metabolic activity and differentiation of cells (reviewed in Ruoslahti, 1989; Jackson et al., 1991; Kjellen and Lindahl, 1991; Hardingham and Fosang, 1992).
I t is of note that immunolabeling for neurocan (Oohira et al., 1994; present study) and phosphacan declines in the centers of the vibrissal barrels in laver 4 of somatosensow
of cortex. This change in distribution is similar to that previously described for tenascin (cytotactin) and a cytotac- tin-binding proteoglycan (Crossin et al., 1989; Steindler et al., 1989; Sheppard et al., 1991). Both neurocan and phosphacan bind with high affinity to tenascin (Grumet et al., 1994). Because the decline in labeling for these mol- ecules in the centers of barrels occurs after the thalamic afferents have formed distinct vibrissa-related axonal clus- ters (Jhaveri et al., 1991; for review, see Steindler, 1993), it cannot be directly responsible for the axonal patterning as earlier studies had suggested (Steindler et al., 1990). In- stead, afferents have a primary role in barrel formation (Woolsey, 1990; Schlaggar and O’Leary, 1991; Senft and Woolsey, 1991), and the loss of ECM molecules appears to be a secondary event (Jhaveri et al., 1991). Thus, although the maturational signal is not known, the expression of neurocan, phosphacan, and several other proteoglycans is down-regulated throughout the cortex as maturation pro- ceeds; the centers of vibrissal barrels are of particular interest, because they are precocious in this regard.
ACKNOWLEDGMENTS We thank Carol Boyd, Joelle Kaliki, and Judy Speck for
excellent technical assistance and Drs. Janice Brunstrom, Richard Margolis, and RenCe Margolis for many helpful discussions during the course of this work. We are grateful to the many colleagues who generously provided antibodies used in this study. Their names and affiliations are provided in Materials and Methods. This work was supported by a research grant to A.L.P. (EY00621; NEI) and fellowship support to B.M. (NIH training grant EY07057; NEI), A.R.B. (NIH training grant NS07027; NINDS), and A.M.S. (McDonnell Centers for Higher Brain Function and Cellu- lar and Molecular Neurobiology, Washington University).
LITERATURE CITED Allendoerfer, K.L., and C.J. Shatz (1994) The subplate, a transient cortical
structure: Its role in the development of connections between thalamus and cortex. Annu. Rev. Neurosci 17:185-218.
Avnur, Z., and B. Geiger (1984) Immunocytochemical localization of native chondroitin-sulfate in tissues and cultured cells using specific monoclo- nal antibody. Cell 38:811-822.
Bayer, S.A., and J. Altman (1991) Neocortical Development. New York: Raven Press.
Bertolotto, A,, G. Rocca, and D. Schiffer (1990) Chondroitin 4-sulfate proteoglycan forms an extracellular network in human and rat central nervous system. J. Neurol. Sci. 100:113-123.
Bertolotto, A,, G. Rocca, G. Canavese, A. Migheli, and D. Schiffer (1991) Chondroitin sulfate proteoglycan surrounds a subset of human and rat CNS neurons. J. Neurosci. Res. 29:225-234.
Bicknese, A.R., A.M. Sheppard, D.D.M. O’Leary, and A.L. Pearlman (1991) Thalamocortical axons preferentially extend along a chondroitin sulfate proteoglycan-enriched pathway coincident with the neocortical subplate and distinct from the efferent path. SOC. Neurosci. Abstr. 17:764.
Bicknese, A.R., A.M. Sheppard, D.D.M. O’Leary, and A.L. Pearlman (1994) Thalamocortical axons preferentially extend along a chondroitin sulfate proteog1ycan.enriched pathway coincident with the neocortical subplate and distinct from the efferent path. J. Neurosci. 14:3500-3510.
Bosse, A,, K. Schwarz, E. Vollmer, and H. Kresse (1993) Divergent and co-localization of the two small proteoglycans decorin and proteoglycan- 100 in human skeletal tissues and tumors. J. Histochem. Cytochem. 41:13-19.
Bovolenta, P., F. Wandosell, and M. Nieto-Sampedro (1993) Characteriza- tion of a neurite outgrowth inhibitor expressed after CNS injury. Eur. J. Neurosci. 5454465.
Brauker. J.H.. M.S. Trautman. and M. Bernfield (1991) Svndecan. a cell surface proteoglycan, exhibits a molecular polymorphism during lung
cortex before it declines in the barrel walls or in other parts development. Dev. KOI. 147:285-292.

CSPGS IN THE NEOCORTICAL SUBPLATE 62 7
Brittis, P.A., D.R. Canning, and J. Silver (1992) Chondroitin sulfate as a regulator of neuronal patterning in the retina. Science 255333-736.
Bumol, T.F., L.E. Walker, and R.A. Reisfeld (1984) Biosynthetic studies of proteoglycans in human melanoma cells with a monoclonal antibody to a core glycoprotein of chondroitin sulfate proteoglycans. J. Biol. Chem. 259: 12733-12741.
Carey, D.J., D.M. Evans, R.C. Stahl, V.K. Asundi, K.J. Conner, P. Garbes, and G. Cizmeci-Smith (1992) Molecular cloning and characterization of N-syndecan, a novel transmembrane heparan sulfate proteoglycan. J. Cell Biol. 117t191-201.
Catalano, S.M., R.T. Robertson, and H.P. Killackey (1991) Early ingrowth of thalamocortical afferents to the neocortex of the prenatal rat. Proc. Natl. Acad. Sci. USA 88t2999-3003.
Caterson, B., J.R. Baker, J.E. Christner, J.F. Kearney, and R.C. Stohrer (1981) The characterization of clonal antibodies directed against bovine nasal cartilage proteoglycan and link protein. In: Monoclonal antibodies and T-cell hybridomas. Eds. G.U. Hammerling, U. Hammerling and J.F. Kearney. ElsevieriNorth-Holland Biomedical Press, Amsterdam, pp. 259-267.
Caterson, B., J.E. Christner, J.R. Baker, and J.R. Couchman (1985) Production and characterization of monoclonal antibodies directed against connective tissue proteoglycans. FASEB J., 44.386-393.
Caterson, B., J. Griffen, F. Mahmoodian, and J.M. Sorrel1 (1990) Monoclonal antibodies against chondroitin sulfphate isomers: Their use as probes for investigating proteoglycan metabolism. Biochem. SOC. Trans. 18r820- 823.
Caviness, V.S., Jr. (1982) Neocortical histogenesis in normal and reeler mice: A developmental study based upon PHIthymidine autoradiography. Dev. Brain Res. 4t293-302.
Chun, J.J.M., and C.J. Shatz (1988) A fibronectin-like molecule is present in the developing cat cerebral cortex and is correlated with subplate neurons. J. Cell Biol. 106t857-872.
Cizmeci-Smith, G., V. Asundi, R.C. Stahl, L.J. Teichman, M. Chernousov, K. Cowan, and D.J. Carey (1992) Regulated expression of syndecan in vascular smooth muscle cells and cloning of rat syndecan core protein cDNA. J. Biol. Chem. 267r15729-15736.
Corless, C.L., A. Mendoza, T. Collins, and J. Lawler (1992) Colocalization of thrombospondin and syndecan during murine development. Dev. Dy- nam. 193:346-358.
Couchman, J.R., J.L. King, and K.J. McCarthy (1990) Distribution of two basement membrane proteoglycans through hair follicle development and the hair growth cycle in the rat. J. Invest. Dermatol. 94165-70.
Crossin, K.L., S. Hoffman, S.-S. Tan, and G.M. Edelman (1989) Cytotactin and its proteoglycan ligand mark structural and functional boundaries in somatosensory cortex of the early postnatal mouse. Dev. Biol. 136:381- 391.
De Carlos, J.A., and D.D.M. O’Leary (1992) Growth and targeting of subplate axons and establishment of major cortical pathways. J. Neuro- sci. 12:1194-1211.
Delpech, A., and B. Delpech (1984) Expression of hyaluronic acid-binding glycoprotein, hyaluronectin, in the developing rat embryo. Dev. Biol. 101:391400.
Derer, P., and S. Nakanishi (1983) Extracellular matrix distribution during neocortical wall ontogenesis in “normal” and “reeler” mice. J. Hirn- forsch. 24t209-224.
Erzurumlu, R.S., and S. Jhaveri (1992) Emergence of connectivity in the embryonic rat parietal cortex. Cereb. Cortex2:33&352.
Esko, J.D. (1991) Genetic analysis of proteoglycan structure, function and metabolism. Curr. Opin. Cell Biol. 3.805-816.
Faassen, A.E., J.A. Schrager, D.J. Klein, T.R. Oegema, J.R. Couchman, and J.B. McCarthy (1992) A cell surface chondroitin sulfate proteoglycan, immunologically related to CD44, is involved in type I collagen-mediated melanoma cell motility and invasion. J. Cell Biol. 116t521-531.
Fichard, A,, J.-M. Verna, J. Olivares, and R. Saxod (1991) Involvement of a chondroitin sulfate proteoglycan in the avoidance of chick epidermis by dorsal root ganglia fibers: A study using B-D-xyloside. Dev. Biol. 148:l-9.
Flaccus, A., A. Janetzko, H. Tekotte, R.K. Margolis, and R.U. Margolis (1991) Immunocytochemical localization of chondroitin and chondroitin 4- and 6-sulfates in developing rat cerebellum. J. Neurochem. 56t1608- 1615.
Friauf, E., S.K. McConnell, and C.J. Shatz (1990) Functional synaptic circuits in the subplate during fetal and early postnatal development of cat visual cortex. J. Neurosci. 10r2601-2613.
Friedlander, D.R., P. Milev, L. Karthikeyan, R.K. Margolis, R.U. Margolis, and M. Grumet (1994) The neuronal chondroitin sulfate proteoglycan neurocan binds to the neural cell adhesion molecules Ng-CAMILIINILE
and N-CAM, and inhibits neuronal adhesion and neurite outgrowth. J. Cell Biol. 125r669-680.
Gallagher, J.T. (1989) The extended family of proteoglycans: Social residents of the pericellular zone. Cum. Opin. Cell Biol. lr1201-1218.
Gellatin, W.M., I.L. Weissman, and E.C. Butcher (1983) A cell-surface molecule involved in organ-specific homing of lymphocytes. Nature 304t30-34.
Ghosh, A., and C.J. Shatz (1992) Involvement of subplate neurons in the formation of ocular dominance columns. Science 255:1441-1443.
Ghosh, A,, and C.J. Shatz (1993) A role for subplate neurons in the patterning of connections from thalamus to neocortex. Development 11 7:1031-1047.
Ghosh, A,, A. Antonini, S.K. McConnell, and C.J. Shatz (1990) Requirement for subplate neurons in the formation of thalamocortical connections. Nature 347; 179-181.
Glossl, J., M. Beck, and H. Kresse (1984) Biosynthesis of proteodermatan sulfate in cultured human fibroblasts. J. Biol. Chem. 259t14144-14150.
Gowda, D.C., R.U. Margolis, and R.K. Margolis (1989) Presence of HNK-1 epitope on poly(N-acetyllactosaminyl) oligosaccharides and identifica- tion of multiple core proteins in the chondroitin sulfate proteoglycans of brain. Biochemistry 28:44684474.
Grumet, M., A. Flaccus, and R.U. Margolis (1993) Functional characteriza- tion of chondroitin sulfate proteoglycans of brain: Interactions with neurons and neural cell adhesion molecules. J. Cell Biol. 120:815-824.
Grumet, M., P. Milev, T. Sakurai, L. Karthikeyan, M. Bourdon, R.K. Margolis, and R.U. Margolis (1994) Interactions with tenascin and differential effects on cell adhesion of neurocan and phosphacan, two major chondroitin sulfate proteoglycans of nervous tissue. J. Biol. Chem. 269: 12 142-12 146.
Hardingham, T.E., and A.J. Fosang (1992) Proteoglycans: Many forms and many functions. FASEB J. 6:861-870.
Herndon, M.E., and A.D. Lander (1990) A diverse set of developmentally regulated proteoglycans is expressed in the rat central nervous system. Neuron 4r949-961.
Hoffman, S., K.L. Crossin, F.S. Jones, D.R. Friedlander, and G.M. Edelman (1990) Cytotactin and cytotactin-binding proteoglycan. Ann. NY Acad. Sci. 580,288-301.
Iijima, N., A. Oohira, T. Mori, K. Kitabatake, and S. Kohsaka (1991) Core protein of chondroitin sulfate proteoglycan promotes neurite outgrowth from cultured neocortical neurons. J. Neurochem. 56:706-708.
Jackson, R.L., S.J. Busch, and A.D. Cardin (1991) Glycosaminoglycans: Molecular properties, protein interactions, and role in physiological processes. Physiol. Rev. 71t481-539.
Jhaveri, S., R.S. Erzurumlu, and K. Crossin (1991) Barrel construction in rodent neocortex: Role of thalamic afferents vs. extracellular matrix molecules. Proc. Nati. Acad. Sci. USA 88t4489-4493.
Johnson, G.D., and G.M. Nogueira-Araujo (1981) A simple method of reducing the fading of immunofluorescence during microscopy. J. Immu- nol. Methods 43:349-350.
Kageyama, G.H., and R.T. Robertson (1993) Development of geniculocorti- cal projections to visual cortex in rat: Evidence for early ingrowth and synaptogenesis. J. Comp. Neurol. 335: 123-148.
Kjellen, L., and U. Lindahl (1991) Proteoglycans: Structures and interac- tions. Annu. Rev. Biochem. 60r443-475.
Kresina, T.F., and C.J. Malemud (1986) Murine monoclonal antibodies recognizing rabbit proteoglycans. Collagen Rel. Res. 6: 15-39.
Lafont, F., M. Rouget, A. Triller, A. Prochiantz, and A. Rousselet (1992) In vitro control of neuronal polarity by glycosaminoglycans. Development 114: 17-29.
Lander, A.D. (1993) Proteoglycans. In T. Kreis and R. Vale (eds): Guidebook to the Extracellular Matrix and Adhesion Proteins. Oxford: Oxford University Press, pp. 12-1.6.
Lane, M.C., and M. Solurish (1991) Primary mesenchyme cell migration requires a chondroitin sulfateidermatin sulfate proteoglycan. Dev. Biol. 143t389-397.
Letourneau, P.C., M.L. Condic, and D.M. Snow (1994) Interactions of developing neurons with the extracellular matrix. J. Neurosci. 14:915- 928.
Levine, J.M., and J.P. Card (1987) Light and electron microscopic localiza- tion of a cell surface antigen (NG2) in the rat cerebellum: Association with smooth protoplasmic astrocytes. J. Neurosci. 72711-2720.
Levine, J.M., F. Stincone, andY.3. Lee (1993) Development and differentia- tion of glial precursor cells in the rat cerebellum. Glia 7r307-321.

628 B. MILLER ET AL.
Luskin, M.B., and C.J. Shatz (1985) Studies of the earliest generated cells of the cat’s visual cortex: Congeneration of the cells of the marginal zone and subplate. J. Neurosci. 5t1062-1075.
Margolis, R.U., R.K. Margolis, L.B. Chang, and C. Preti (1975) Gly- cosaminoglycans of brain during development. Biochemistry 14:85-88.
Marin-Padilla, M. (1971) Early prenatal ontogenesis of the cerebral cortex (neocortex) of the cat (Felis domestics). A Golgi study. I. The primordial neocortical organization. Z. Anat. Entwickl. Gesch. 134:117-145.
Matsui, F., A. Oohira, R. Shoji, Y. Kariya, and K. Yoshida (1993) Biochemical comparison of brain glycosaminoglycans hetween normal and reeler mutant mice. Neurosci. Res. 16:287-292.
Maurel, P., U. Rauch, M. Flad, R.K. Margolis, and R.U. Margolis (1994) Phosphacan, a chondroitin sulfate proteoglycan of brain that interacts with neurons and neural cell adhesion molecules, is an extracellular variant of a receptor-type protein tyrosine phosphatase. Proc. Natl. Acad. Sci. USA91t2512-2516.
McCarthy, K.J., M.A. Accavitti, and J.R. Couchman (1989) Immunological characterization of a basement membrane-specific chondroitin sulfate proteoglycan. J. Cell Biol. 109r3187-3198.
McConnell, S.K., A. Ghosh, and C.J. Shatz (1989) Subplate neurons pioneer the first axon pathway from the cerebral cortex. Science 245978-982.
McKeon, R.J., R.C. Schreiber, J.S. Rudge, and J. Silver (1991) Reduction of neurite outgrowth in a model of glial scarring following CNS injury is correlated with the expression of inhibitory molecules on reactive astrocytes. J. Neurosci. llt3398-3411.
Miller, B., A.M. Sheppard, and A.L. Pearlman (1992) Expression of two chondroitin sulfate proteoglycan core proteins in the subplate pathway of early cortical afferents. SOC. Neurosci. Abstr. 181778.
Miller, B.M., L. Chou, and B.L. Finlay (1993) The early development of thalamocortical and corticofugal projections. J. Comp Neurol. 335116- 41.
Miyake, K., C.B. Underhill, J. Lesley, and P.W. Kincade (1990) Hyaluronate can function as a cell adhesion molecule and CD44 participates in hyaluronate recognition. J. Exp. Med., 17269-75.
Morriss-Kay, G., and F. Tuckett (1989) Immunohistochemical localisation of chondroitin sulphate proteoglycans and the effect of chondroitinase ABC in 9- to 11-day rat embryos. Development 106:787-798.
Normand, G., S. Kuchler, A. Meyer, G. Vincendon, and J.-P. Zanetta (1988) Isolation and immunohistochemical localization of a chondroitin sulfate proteoglycan from adult rat brain. J. Neurochem. 51:665-676.
Oakley, R.A., and K.W. Tosney (1991) Peanut agglutinin and chondroitin-6- sulfate are molecular markers for tissues that act as barriers to axon advance in the avian embryo. Dev. Biol. I47:187-206.
Oohira, A., F. Matsui, M. Matsuda, Y. Takida, and Y. Kuboki (1988) Occurrence of three distinct molecular species of chondroitin sulfate proteoglycan in the developing rat brain. J. Biol. Chem. 263: 10240- 10246.
Oohira, A,, F. Matsui, E. Watanabe, Y. Kushima, and N. Maeda (1994) Developmentally regulated expression of a brain specific species of chondroitin sulfate proteoglycan, neurocan, identified with a monoclonal antibody IG2 in the rat cerebrum. Neuroscience 60,145-157.
Perris, R., and S. Johansson (1987) Amphibian neural crest cell migration on purified extracellular matrix components: A chondroit,in sulfate proteo- glycan inhibits locomotion on fibronectin substrates. J. Cell Biol. 105: 2511-2521.
Pindzola, R.R., C. Doller, and J. Silver (1993) Putative inhibitory extracellu- lar matrix molecules a t the dorsal root entry zone of the spinal cord during development and after root and sciatic nerve lesions. Dev. Biol. 156t3448.
Raedler, E., and A. Raedler (1978) Autoradiographic study of early neurogen- esis in rat neocortex. Anat. Embryol. 154.267-284.
Rauch, U., P. Gao, A. Janetzko, A. Flaccus, L. Hilgenberg, H. Tekotte, R.K. Margolis, and R.U. Margolis (1991) Isolation and characterization of developmentally regulated chondroitin sulfate and chondroitinikeratin sulfate proteoglycans of brain identified with monoclonal antibodies. J. Biol. Chem. 266: 14785-1480 1.
Rauch, U., L. Karthikeyan, P. Maurel, R.U. Margolis, and R.K. Margolis (1992) Cloning and primary structure of neurocan, a developmentally
regulated, aggregating chondroitin sulfate proteoglycan of brain. 3. Biol. Chem. 267t19536-19547.
Rickmann, M., B.M. Chronwall, and J.R. Wolff (1977) On thedevelopment of non-pyramidal neurons and axons outside the cortical plate: The early marginal zone as a pallial anlage. Anat. Emhryol. 151285-307.
Ruoslabti, E. (1989) Proteoglycans in cell regulation. J. Biol. Chem. 264: 13369-13372.
Schlaggar, B.L., and D.D.M. O’Leary (1991) Potential of visual cortex to develop an array of functional units unique to somatosensory cortex. Science 25211556-1560.
Schrappe, M., F.G. Klier, R.C. Spiro, T.A. Waltz, R.A. Reisfeld, and C.L. Gladson (1991) Correlation of chondroitin sulfate proteoglycan expres- sion on proliferating brain capillary endothelial cells with the malignant phenotype of astroglial cells. Cancer Res. 51:49864993.
Senft, S.L., and T.A. Woolsey (1991) Growth of thalamic afferents into mouse barrel cortex. Cereb. Cortex 1:308-335.
Sheppard, A.M., S.K. Hamilton, and A.L. Pearlman (1991) Changes in the distribution of extracellular matrix components accompany early morpho- genetic events of mammalian cortical development. J. Neurosci. 11:3928- 3942.
Snow, D.M., V. Lemmon, D.A. Carrino, A.I. Caplan, and J. Silver 11990) Sulfated proteoglycans in astroglial barriers inhibit neurite outgrowth in vitro. Exp. Neurol. 109:lll-130.
Snow, D.M., M. Watanahe, P.C. Letourneau, and J. Silver (1991) A chondroitin sulfate proteoglycan may influence the direction of retinal ganglion cell outgrowth. Development I13:1473-1485.
Solursh, M., R.S. Reiter, K.L. Jensen, M. Kato, and M. Bernfield (1990) Transient expression of a cell surface heparan sulfate proteoglycan (syndecan) during limb development. Dev. Biol. 140.83-92.
Sorrell, J.M., D.A. Carrino, and A.I. Caplan (1993) Structural domains in chondroitin sulfate identified hy anti-chondroitin sulfate monoclonal antibodies. Immunosequencing of chondroitin sulfates. Matrix 13:351- 361.
Stallcup, W.B., and L. Beasley (1987) Bipotential glial precursor cells of the optic nerve express the NG2 proteoglycan. J. Neurosci. 72737-2744.
Steindler, D.A. (1993) Glial boundaries in the developing nervous system. Annu. Rev. Neurosci. 16:445-470.
Steindler, D.A., N.G.F. Cooper, A. Faissner, and M. Schachner (1989) Boundaries defined by adhesion molecules during development of the cerebral cortex: The Jlitenascin glycoprotein in the mouse somatosen- sory cortical barrel field. Dev. Biol. 131.243-260.
Steindier, D.A., A. Faissner, and M. Schachner (1990) Brain “cordones”: Transient boundaries of glia and adhesion molecules that define develop- ing functional units. Commun. Dev. Neurohiol. 1129-60.
Stewart, G.R., and A.L. Pearlman (1987) Fibronectin-like immunoreactivity in the developing cerebral cortex. J. Neurosci. 7:3325-3333.
Stipp, C.S., E.D. Litwack, and A.D. Lander (1994) Cerebroglycan: An integral membrane heparan sulfate proteoglycan that is unique to the developing nervous system and expressed specifically during neuronal differentiation. J. Cell Biol. 1241149-160.
Thomas, L., H.R. Byers, J. Vink, and I. Stamenkovic (1992) CD44H regulates tumor cell migration on hyaluronate-coated substrate. J. Cell Biol. 118:971-977.
Trowbridge, I.S., J. Lesley, R. Schulte, R. Hyman, and J. Trotter (1982) Biochemical characterization and cellular distribution of a polymorphic, murine cell-surface glycoprotein expressed on lymphoid tissues. Immuno- genetics 15299-312.
Wood, J.G., S. Martin, and D.J. Price (1992) Evidence that the earliest generated cells of the murine cerebral cortex form a transient population in the subplate and marginal zone. Dev. Brain Res. 66t137-140.
Woolsey, T.A. (1990) Peripheral alteration and somatosensory development. In J.R. Coleman (ed): Development of Sensory Systems in Mammals. New York: John Wiley and Sons, pp, 461-516.
Zaremba, S., J.R. Naegele, C.J. Barnstable, and S. Hockfield (1990) Neuronal subsets express multiple high-molecular-weight cell-surface glycoconju- gates defined hy monoclonal antibodies Cat-301 and VCl.1. J. Neurosci. 10:2985-2995.




















