
stability analysis of dump with admixture of fly-ash ... - CORE
Upload
independentCategory
view
0download
0
Characterization of Admixture in an Urban Sample from BuenosAires, Argentina, Using Uniparentally and Biparentally InheritedGenetic Markers
VERONICA L. MARTINEZ MARIGNAC,1,3 BERNARDO BERTONI,2 ESTEBAN J. PARRA,3
AND NESTOR O. BIANCHI1
Abstract In this study we analyzed a sample of the urban population ofLa Plata, Argentina, using 17 mtDNA haplogroups, the DYS199 Y-chromo-some polymorphism, and 5 autosomal population-associated alleles (PAAs).The contribution of native American maternal lineages to the population ofLa Plata was estimated as 45.6%, whereas the paternal contribution wasmuch lower (10.6%), clearly indicating directional mating. Regarding auto-somal evidence of admixture, the relative European, native American, andWest African genetic contributions to the gene pool of La Plata were esti-mated to be 67.55% (�2.7), 25.9% (�4.3), and 6.5% (�6.4), respectively.When admixture was calculated at the individual level, we found a low corre-lation between the ancestral contribution estimated with uniparental lineagesand autosomal markers. Most of the individuals from La Plata with a nativeAmerican mtDNA haplogroup or the DYS199*T native American alleleshow a genetic contribution at the autosomal level that can be traced primar-ily to Europe. The results of this study emphasize the need to use both uni-parentally and biparentally inherited genetic markers to understand thehistory of admixed populations.
The process of gene and cultural exchange in the Americas involved three mainpopulations: native Americans, Europeans, and West Africans. Gene introgres-sion from other populations (e.g., South Asia, East Asia, and the Middle East)has been much smaller and limited to speciÞc geographic regions. Throughoutthe Americas, historical differences in the pattern of settlement and migrationhave resulted in a wide range of parental genetic contributions to present ruraland urban populations (Sans 2000).
1Multidisciplinary Institute of Cell Biology, Department of Population Molecular Genetics, Calle 526 e/10y 11, C.C. 403, La Plata, C.P. 1900, Buenos Aires, Argentina.
2Department of Genetics, College of Medicine, Universidad de la Republica, Avda. Gral. Flores 2125, C.P.11800, Montevideo, Uruguay.
3Department of Anthropology, University of Toronto at Mississauga, 3359 Mississauga Rd. North, Room4019, South Building, Mississauga, ON L5L 1C6, Canada.
Human Biology, August 2004, v. 76, no. 4, pp. 543�557.Copyright � 2004 Wayne State University Press, Detroit, Michigan 48201-1309
KEY WORDS: Y CHROMOSOME, MITOCHONDRIAL DNA, mtDNA HAPLOGROUPS, AUTO-SOMES, PV92, WI1319, DR2DI, OCA2, SB19.3, DYS199, ADMIXTURE, POPULATION-ASSOCIATEDALLELES (PAAs), ARGENTINA, PARAGUAY, URUGUAY, SOUTH AMERICA.

544 / martinez marignac et al.
The presence of molecular markers of restricted geographic distribution inthe mitochondrial genome and the non-pseudo-autosomal region of the Y chro-mosome provides a powerful tool to explore the history of female and malemigrations. mtDNA and Y-chromosome-speciÞc sequences are haploid, do notrecombine, and are transmitted through the maternal and paternal lines, respec-tively. Given their mode of inheritance, mtDNA and Y-chromosome-speciÞc se-quences provide a useful but limited view of the past history of an individual orpopulation. The full perspective can be gained only by combining these uniparen-tal markers with autosomal markers, which provide the ��average�� history ofadmixture through the generations. A particularly useful set of autosomal mark-ers to study admixture are those markers showing large allele-frequency differ-ences (� values) between geographic populations (Shriver et al. 1997; Parra etal. 1998, 2001). These markers have been termed population-speciÞc alleles(PSAs) or population-associated alleles (PAAs) (Pfaff et al. 2001).
A good example of the application of uniparental and biparental markersto understand population history was described by Bravi et al. (1997) and Sanset al. (2002). These researchers used mtDNA, Y-chromosome, and autosomalpolymorphisms to study a relatively isolated Afro-Uruguayan population that,according to historical and sociocultural data, was assumed to have primarily anAfrican genetic contribution. In this population 52% of the mtDNA lineages wereof African ancestry, 29% of native American ancestry, and 19% of Europeanancestry, whereas the relative contributions observed for the Y-chromosome lin-eages were 30% African, 6% native American, and 64% European. The use ofblood groups and protein enzyme markers indicated a mixture composed of 47%,15%, and 38% African, Amerindian, and European autosomal genetic compo-nents, respectively. This Afro-Uruguayan example was chosen to illustrate threepoints: (1) Genetic mixture occurs in most human populations, even in seeminglyisolated ones; (2) admixture estimates provided by mtDNA, Y-chromosome, andautosomal markers may differ because each system evaluates a distinct path ofgene introgression; and (3) genetic data provide information that is critical tounderstanding the history of any given population, complementing and expand-ing the evidence that can be gathered from other sources.
Most urban populations in Argentina are assumed to have a predominantlyEuropean genetic component (mainly because of immigration from Spain andItaly and to a lesser extent from Germany and France) with low levels of Amerin-dian and African admixture (INDEC 1997; Sala et al. 1999; Goicoechea et al.2000). However, estimation of admixture using genetic markers is almost nonex-istent for Argentinean urban populations.
La Plata city, the capital of the province of Buenos Aires, is a city of750,000 inhabitants. The city was founded in 1882 in a region that had no Amer-indian population, and the early settlers were Europeans, mainly Italians, whocame to the city as workers in charge of building the ediÞces housing the politi-cal, cultural, and administrative branches of the provincial government (Bejarano1967; Barba 1983). At the end of the 19th century the population of La Plata
Admixture in Buenos Aires / 545
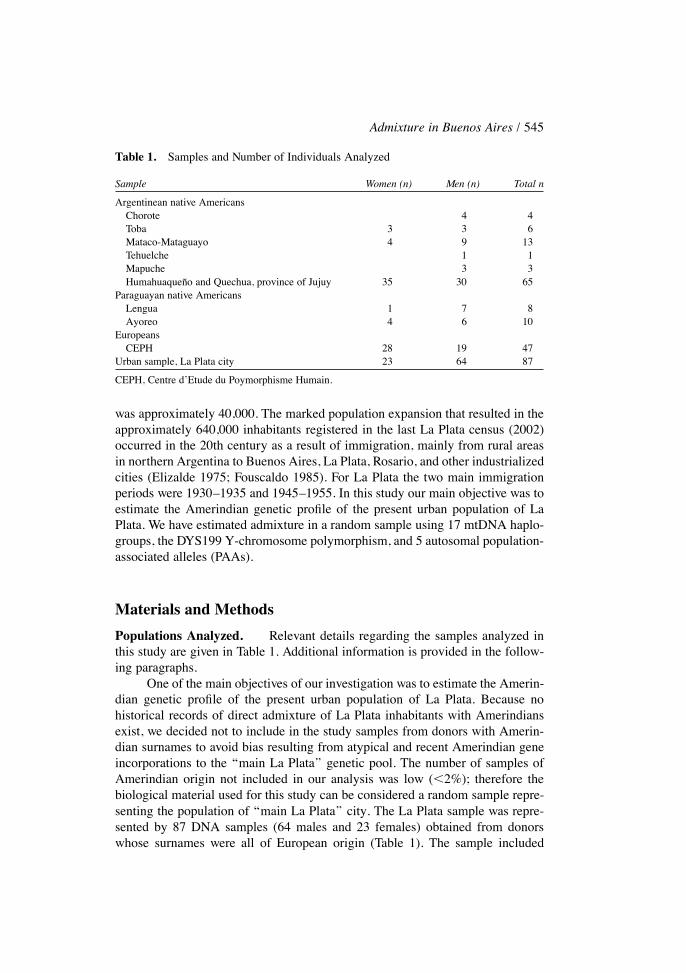
Table 1. Samples and Number of Individuals Analyzed
Sample Women (n) Men (n) Total n
Argentinean native AmericansChorote 4 4Toba 3 3 6Mataco-Mataguayo 4 9 13Tehuelche 1 1Mapuche 3 3Humahuaquen�o and Quechua, province of Jujuy 35 30 65
Paraguayan native AmericansLengua 1 7 8Ayoreo 4 6 10
EuropeansCEPH 28 19 47
Urban sample, La Plata city 23 64 87
CEPH, Centre d�Etude du Poymorphisme Humain.
was approximately 40,000. The marked population expansion that resulted in theapproximately 640,000 inhabitants registered in the last La Plata census (2002)occurred in the 20th century as a result of immigration, mainly from rural areasin northern Argentina to Buenos Aires, La Plata, Rosario, and other industrializedcities (Elizalde 1975; Fouscaldo 1985). For La Plata the two main immigrationperiods were 1930�1935 and 1945�1955. In this study our main objective was toestimate the Amerindian genetic proÞle of the present urban population of LaPlata. We have estimated admixture in a random sample using 17 mtDNA haplo-groups, the DYS199 Y-chromosome polymorphism, and 5 autosomal population-associated alleles (PAAs).
Materials and Methods
Populations Analyzed. Relevant details regarding the samples analyzed inthis study are given in Table 1. Additional information is provided in the follow-ing paragraphs.
One of the main objectives of our investigation was to estimate the Amerin-dian genetic proÞle of the present urban population of La Plata. Because nohistorical records of direct admixture of La Plata inhabitants with Amerindiansexist, we decided not to include in the study samples from donors with Amerin-dian surnames to avoid bias resulting from atypical and recent Amerindian geneincorporations to the ��main La Plata�� genetic pool. The number of samples ofAmerindian origin not included in our analysis was low (�2%); therefore thebiological material used for this study can be considered a random sample repre-senting the population of ��main La Plata�� city. The La Plata sample was repre-sented by 87 DNA samples (64 males and 23 females) obtained from donorswhose surnames were all of European origin (Table 1). The sample included

546 / martinez marignac et al.
individuals attending a public hospital and also staff and faculty from the Univer-sity of La Plata. At the time of collection, all the individuals were working andliving in the city of La Plata.
Our reference populations include the native American, European, andWest African populations used to obtain the PAA reference frequencies employedto estimate admixture. Native American DNA samples were obtained from sixArgentinean and two Paraguayan communities inhabiting geographic areas his-torically occupied by their ancestors. Donors were selected only if they had well-documented Amerindian genealogies and no family links with other donors inthe same population. The samples are composed of individuals from the follow-ing native American groups: (1) Humahuaquen�o and Quechua from the provinceof Jujuy in northwestern Argentina; (2) Tehuelche from Pampa de Chalõa andLoma Redonda, province of Neuquen; (3) Mapuche, from the province of RõoNegro; (4) Mataco-Mataguayo from the village of Santa Victoria, northwest ofthe province of Salta; (5) Chorote and (6) Toba from the village of Santa Victoria,in the northwest province of Salta; and (7) Ayoreo and (8) Lengua from southernParaguay.
Europeans were represented by DNA samples corresponding to 19 malesand 28 females that were part of CEPH genealogies of known European ancestry.In addition, we used previously published data on allele frequencies from Spainand West Africa to estimate the parental frequencies that were used in the admix-ture analysis (Parra et al. 1998). These data are available in the dbSNP databaseunder the submitter handle PSU_ANTH (available at http://www.ncbi.nlm.nih.gov/SNP/).
All La Plata donors gave informed consent for their participation in thisstudy. The native American and European samples analyzed in the laboratorywere submitted by various investigators who were in charge of obtaining in-formed consent from the donors. For the La Plata samples in which the surnameof the donors was made available to the research group, the correspondence be-tween each sample and the surname of the donor was maintained in anonymity.
Molecular Markers. The samples were screened for the following mtDNAhaplogroups: A1, A2, B, C1, C2, D1, and D2 (native American�speciÞc haplo-groups); H, I, J, K, T, U, V, W, and X (European-speciÞc haplogroups); andL1/L2 (African-speciÞc haplogroups). Information on the experimental conditionsused to characterize these haplogroups, including primer sequences and PCR con-ditions, can be found in Bailliet et al. (1994) and Torroni et al. (1994, 1996).
The samples were screened for the presence of a CVT transition in theDYS199 locus, a marker speciÞc to native American Y chromosomes, usuallyfound in 70�90% of South American native American populations (Underhill etal. 1996; Bianchi et al. 1998).
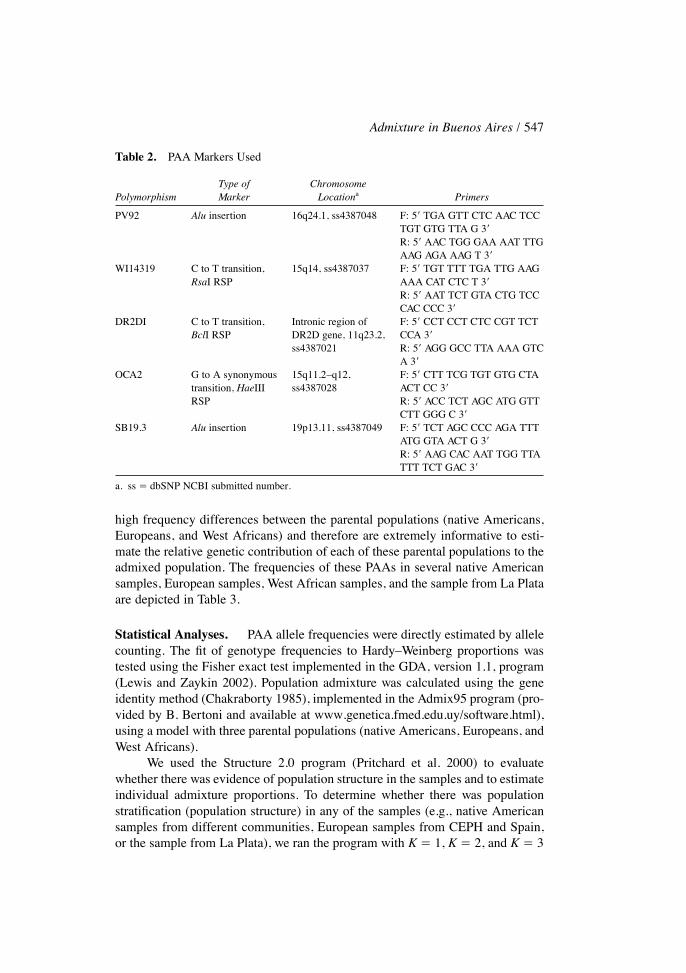
The samples were characterized for Þve PAA biallelic markers (PV92,WI1319, DR2DI, OCA2, and SB19.3). Further information on these polymor-phisms is provided in Table 2. These Þve PAAs were selected because they show
Admixture in Buenos Aires / 547
Table 2. PAA Markers Used
Type of ChromosomePolymorphism Marker Locationa Primers
PV92 Alu insertion 16q24.1, ss4387048 F: 5� TGA GTT CTC AAC TCCTGT GTG TTA G 3�R: 5� AAC TGG GAA AAT TTGAAG AGA AAG T 3�
WI14319 C to T transition, 15q14, ss4387037 F: 5� TGT TTT TGA TTG AAGRsaI RSP AAA CAT CTC T 3�
R: 5� AAT TCT GTA CTG TCCCAC CCC 3�
DR2DI C to T transition, Intronic region of F: 5� CCT CCT CTC CGT TCTBclI RSP DR2D gene, 11q23.2, CCA 3�
ss4387021 R: 5� AGG GCC TTA AAA GTCA 3�
OCA2 G to A synonymous 15q11.2�q12, F: 5� CTT TCG TGT GTG CTAtransition, HaeIII ss4387028 ACT CC 3�RSP R: 5� ACC TCT AGC ATG GTT
CTT GGG C 3�SB19.3 Alu insertion 19p13.11, ss4387049 F: 5� TCT AGC CCC AGA TTT
ATG GTA ACT G 3�R: 5� AAG CAC AAT TGG TTATTT TCT GAC 3�
a. ss � dbSNP NCBI submitted number.
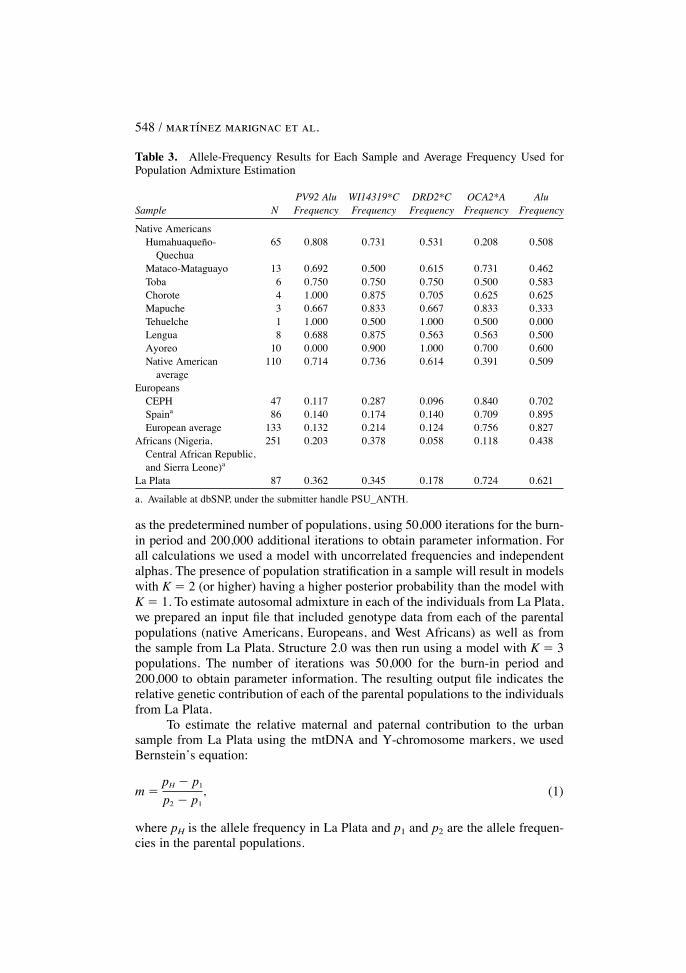
high frequency differences between the parental populations (native Americans,Europeans, and West Africans) and therefore are extremely informative to esti-mate the relative genetic contribution of each of these parental populations to theadmixed population. The frequencies of these PAAs in several native Americansamples, European samples, West African samples, and the sample from La Plataare depicted in Table 3.
Statistical Analyses. PAA allele frequencies were directly estimated by allelecounting. The Þt of genotype frequencies to Hardy�Weinberg proportions wastested using the Fisher exact test implemented in the GDA, version 1.1, program(Lewis and Zaykin 2002). Population admixture was calculated using the geneidentity method (Chakraborty 1985), implemented in the Admix95 program (pro-vided by B. Bertoni and available at www.genetica.fmed.edu.uy/software.html),using a model with three parental populations (native Americans, Europeans, andWest Africans).
We used the Structure 2.0 program (Pritchard et al. 2000) to evaluatewhether there was evidence of population structure in the samples and to estimateindividual admixture proportions. To determine whether there was populationstratiÞcation (population structure) in any of the samples (e.g., native Americansamples from different communities, European samples from CEPH and Spain,or the sample from La Plata), we ran the program with K � 1, K � 2, and K � 3
548 / martinez marignac et al.
Table 3. Allele-Frequency Results for Each Sample and Average Frequency Used forPopulation Admixture Estimation
PV92 Alu WI14319*C DRD2*C OCA2*A AluSample N Frequency Frequency Frequency Frequency Frequency
Native AmericansHumahuaquen�o- 65 0.808 0.731 0.531 0.208 0.508
QuechuaMataco-Mataguayo 13 0.692 0.500 0.615 0.731 0.462Toba 6 0.750 0.750 0.750 0.500 0.583Chorote 4 1.000 0.875 0.705 0.625 0.625Mapuche 3 0.667 0.833 0.667 0.833 0.333Tehuelche 1 1.000 0.500 1.000 0.500 0.000Lengua 8 0.688 0.875 0.563 0.563 0.500Ayoreo 10 0.000 0.900 1.000 0.700 0.600Native American 110 0.714 0.736 0.614 0.391 0.509
averageEuropeans
CEPH 47 0.117 0.287 0.096 0.840 0.702Spaina 86 0.140 0.174 0.140 0.709 0.895European average 133 0.132 0.214 0.124 0.756 0.827
Africans (Nigeria, 251 0.203 0.378 0.058 0.118 0.438Central African Republic,and Sierra Leone)a
La Plata 87 0.362 0.345 0.178 0.724 0.621
a. Available at dbSNP, under the submitter handle PSU_ANTH.
as the predetermined number of populations, using 50,000 iterations for the burn-in period and 200,000 additional iterations to obtain parameter information. Forall calculations we used a model with uncorrelated frequencies and independentalphas. The presence of population stratiÞcation in a sample will result in modelswith K � 2 (or higher) having a higher posterior probability than the model withK � 1. To estimate autosomal admixture in each of the individuals from La Plata,we prepared an input Þle that included genotype data from each of the parentalpopulations (native Americans, Europeans, and West Africans) as well as fromthe sample from La Plata. Structure 2.0 was then run using a model with K � 3populations. The number of iterations was 50,000 for the burn-in period and200,000 to obtain parameter information. The resulting output Þle indicates therelative genetic contribution of each of the parental populations to the individualsfrom La Plata.
To estimate the relative maternal and paternal contribution to the urbansample from La Plata using the mtDNA and Y-chromosome markers, we usedBernstein�s equation:
m �pH � p1
p2 � p1
, (1)
where pH is the allele frequency in La Plata and p1 and p2 are the allele frequen-cies in the parental populations.

Admixture in Buenos Aires / 549
Results
Parental Samples. Ninety-nine percent of the native American samples (109/110) had a native American mtDNA lineage, and 88% of the males (53/60)showed the DYS199*T native American allele. A combination of Y-chromosomeand mtDNA native American lineages was observed in 52 males (87%). AllCEPH samples showed European-speciÞc mtDNA lineages and showed theDYS199*C allele.
Table 3 shows the frequencies of the autosomal PAAs analyzed in thisstudy. We used the Fisher exact test to evaluate the goodness of Þt to Hardy�Weinberg proportions for each locus and over all loci (3,200 shufßing runs).Single-locus testing showed signiÞcant departures from Hardy�Weinberg propor-tions for PV92, DR2D, OCA2, and SB19.3 in the native American sample, forPV92 and DR2D in the CEPH samples, and for PV92, DR2DI, and SB19.3 lociin the sample from La Plata.
The genetic structure analyses using Structure 2.0 showed that there wasno evidence of the presence of a signiÞcant genetic structure in the Europeanparental sample composed of individuals from Spain and the CEPH. The modelwith the highest posterior probability was K � 1 in this sample. However, Struc-ture 2.0 indicated the presence of genetic structure in the native American group.The model with K � 2 had the highest posterior probability (ln P(D) � �668.4for K � 2; ln P(D) � �709.9 for K � 1). Of the two groups deÞned by Structure2.0, one group consisted of Ayoreos (10 subjects) and 18 additional individualsfrom several other aboriginal communities (11 Humahuaquen�os and Quechuas,1 Mapuche, 3 Mataco-Mataguayos, 2 Lenguas, and 1 Toba). The second groupincluded the remaining native American samples.
Urban Sample. As mentioned, before the characterization of admixture pro-portions in the sample from La Plata, we genotyped a set of mtDNA, Y-chromo-some, and autosomal PAA markers in several samples from European (CEPH)and native American groups (Omaguacas or Humahuaquen�os, Quechuas, Tehuel-ches, Mapuches, Mataco-Mataguayos, Chorotes, Tobas, Ayoreos, and Lenguas).We used these frequencies, in addition to published data for other European andAfrican samples, to estimate admixture contributions in the sample from LaPlata.
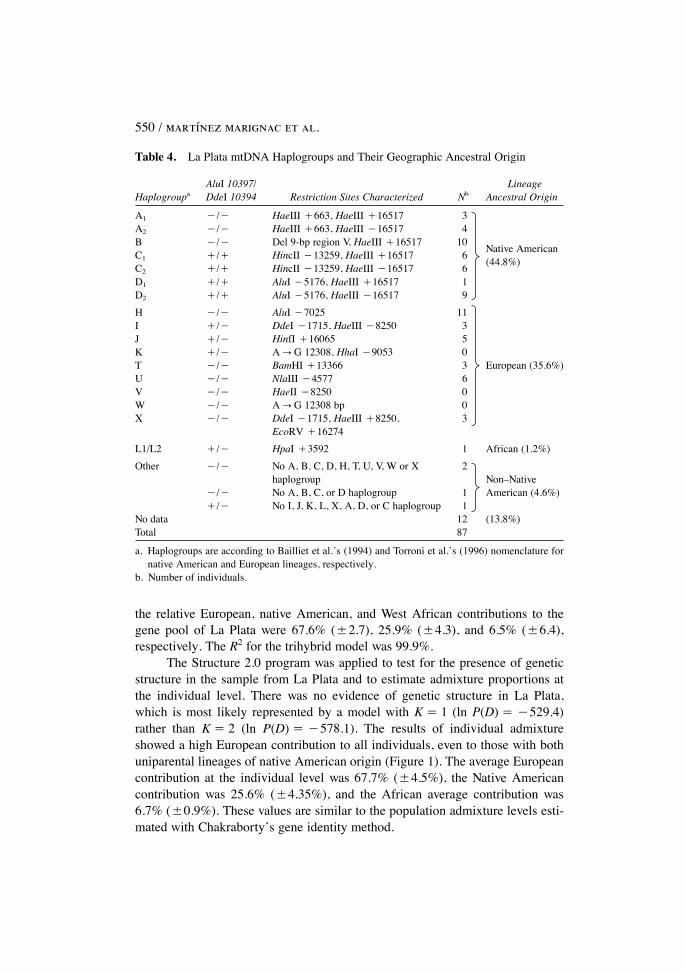
Table 4 displays the mitochondrial results for La Plata: 44.8 % (39/87) ofthe samples showed a native American maternal lineage, 41.4% (36/87) werenon�native American mtDNA haplogroups, and for the remaining 13.8% wecould not determine the lineage using the 17 mitochondrial markers analyzed inthis study. Regarding the Y chromosome, 9.4% of the samples (6 of the 64 males)had the DYS199*T allele. According to Bernstein�s formula, the contribution ofnative American maternal lineages to the gene pool of La Plata was estimated as45.2%, whereas the paternal contribution was much lower (10.6%).
The assessment of ethnic admixture using autosomal markers showed that
550 / martinez marignac et al.
Table 4. La Plata mtDNA Haplogroups and Their Geographic Ancestral Origin
AluI 10397/ LineageHaplogroupa DdeI 10394 Restriction Sites Characterized Nb Ancestral Origin
A1 �/� HaeIII �663, HaeIII �16517 3
Native American
A2 �/� HaeIII �663, HaeIII �16517 4
(44.8%)
B �/� Del 9-bp region V, HaeIII �16517 10C1 �/� HincII �13259, HaeIII �16517 6C2 �/� HincII �13259, HaeIII �16517 6D1 �/� AluI �5176, HaeIII �16517 1D2 �/� AluI �5176, HaeIII �16517 9
H �/� AluI �7025 11
European (35.6%)
I �/� DdeI �1715, HaeIII �8250 3J �/� HinfI �16065 5K �/� AVG 12308, HhaI �9053 0T �/� BamHI �13366 3U �/� NlaIII �4577 6V �/� HaeII �8250 0W �/� AVG 12308 bp 0X �/� DdeI �1715, HaeIII �8250, 3
EcoRV �16274
L1/L2 �/� HpaI �3592 1 African (1.2%)
Other �/� No A, B, C, D, H, T, U, V, W or X 2Non�NativehaplogroupAmerican (4.6%)�/� No A, B, C, or D haplogroup 1
�/� No I, J, K, L, X, A, D, or C haplogroup 1No data 12 (13.8%)Total 87
a. Haplogroups are according to Bailliet et al.�s (1994) and Torroni et al.�s (1996) nomenclature fornative American and European lineages, respectively.
b. Number of individuals.
the relative European, native American, and West African contributions to thegene pool of La Plata were 67.6% (�2.7), 25.9% (�4.3), and 6.5% (�6.4),respectively. The R2 for the trihybrid model was 99.9%.
The Structure 2.0 program was applied to test for the presence of geneticstructure in the sample from La Plata and to estimate admixture proportions atthe individual level. There was no evidence of genetic structure in La Plata,which is most likely represented by a model with K � 1 (ln P(D) � �529.4)rather than K � 2 (ln P(D) � �578.1). The results of individual admixtureshowed a high European contribution to all individuals, even to those with bothuniparental lineages of native American origin (Figure 1). The average Europeancontribution at the individual level was 67.7% (�4.5%), the Native Americancontribution was 25.6% (�4.35%), and the African average contribution was6.7% (�0.9%). These values are similar to the population admixture levels esti-mated with Chakraborty�s gene identity method.
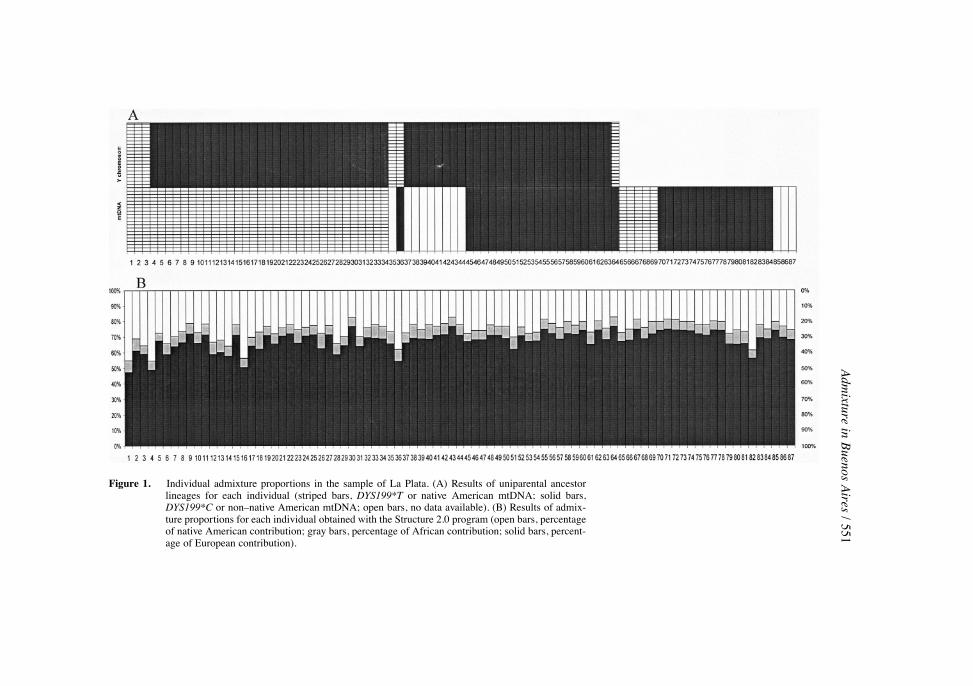
Adm
ixturein
Buenos
Aires
/551
Figure 1. Individual admixture proportions in the sample of La Plata. (A) Results of uniparental ancestorlineages for each individual (striped bars, DYS199*T or native American mtDNA; solid bars,DYS199*C or non�native American mtDNA; open bars, no data available). (B) Results of admix-ture proportions for each individual obtained with the Structure 2.0 program (open bars, percentageof native American contribution; gray bars, percentage of African contribution; solid bars, percent-age of European contribution).

552 / martinez marignac et al.
Comparison of Admixture Estimates Using Y-Chromosome-SpeciÞc, mtDNA,and PAA Systems in La Plata. Our results showed a lack of correlation be-tween mtDNA and Y-chromosome systems versus the PAA systems used to de-termine the ethnic genetic proÞle in the La Plata sample (Figure 1): Only 2 ofthe 42 individuals showing at least one Amerindian parental lineage also had apredominant Amerindian autosomal background, whereas in all other cases PAAmarkers indicated a prevalent European endowment. On the contrary, in the na-tive American sample, uniparental and autosomal systems coincided in showinga prevalent Amerindian genetic proÞle (Figure 2).
Discussion
The aim of this study was to characterize the parental genetic contributionsto an urban sample from Argentina through the analyses of uniparentally andbiparentally inherited genetic markers, both at the population and the individuallevel.
We used two sets of DNA samples representing well-deÞned European andnative American populations in order to test the efÞciency of the uniparental andbiparental markers employed in this study to estimate the genetic admixture inthe La Plata population. No evidence of genetic structure was observed in thesamples of European origin. Conversely, a marked structuring (K � 2) and asigniÞcant Hardy�Weinberg disequilibrium were detected in Amerindian popula-tions, very likely resulting from well-documented population reductions associ-ated with genetic drift and endogamy (Ubelaker 1992; Guerreiro et al. 1994;Trachtenberg et al. 1996; Zago et al. 1996; Tourret et al. 2000; Goicoechea et al.2001). The Structure 2.0 program showed a signiÞcant non-Amerindian geneticproÞle in only 3 of the 110 Amerindians included in this study (Figure 2). ThisÞnding further supports the assumption that isolation and genetic drift and notgene introgression are the major factors inducing the genetic structuring detectedin Amerindians (Cavalli-Sforza, et al. 1996; Salzano and Bortolini 2002). Thewide dispersion of allele frequencies detected in native American populations(Table 4) is in good agreement with this hypothesis.
Both mtDNA and Y-chromosome data show evidence of a native Americangenetic contribution to the urban sample from La Plata. However, the nativeAmerican paternal contribution was substantially lower (10.6%) than the nativeAmerican maternal contribution (45.2%). Clearly, there has been sex-biased geneßow in La Plata, in agreement with historical reports indicating that during colo-nial times, Spanish men embarking on the conquest of South America commonlypracticed polygamous unions with native American women (Herren 1992). Previ-ous studies have also shown directional mating in other South American popula-tions (Dipierri et al. 1998; Batista dos Santos et al. 1999; Rodriguez-DelÞn et al.2001). The autosomal markers showed an intermediate picture, with a high Euro-pean genetic contribution (68%) and native American and African estimates of

Adm
ixturein
Buenos
Aires
/553
Figure 2. Individual admixture proportions in the native American parental sample. (A) Results of uniparen-tal ancestor lineages for each individual (striped bars, DYS199*T or native American mtDNA; solidbars, DYS199*C or non�native American mtDNA; open bars, no data available). (B) Results ofadmixture proportions for each individual obtained with the Structure 2.0 program (open bars,percentage of native American contribution; gray bars, percentage of African contribution; solidbars, percentage of European contribution).

554 / martinez marignac et al.
26% and 6%, respectively. The PAA data in La Plata show concordance withprevious studies in urban regions from Argentina, where a high European ances-tral contribution was detected but relatively small native American and Africancontributions were also present (Martõnez Marignac et al. 1999; Avena et al.2001).
In La Plata the autosomal native American contribution is relatively low atthe individual level, even in those individuals showing native American maternalor paternal lineages. This Þnding indicates that the native American contributionto La Plata did not take place in recent times. The uniparental and biparentaladmixture data from the urban sample of La Plata can be interpreted to be aresult of admixture during colonial times or the Argentinean state conformation.It is important to note that the present population of La Plata can be traced backnot only to relatively recent immigrants from Europe but also to immigration ofpreviously admixed individuals from rural areas and neighboring countries. Inaddition, it is known that Africans also migrated to the region (Picotti 2001), andindeed a minor African genetic contribution was detected in the sample from LaPlata.
The data indicate that admixture studies with the goal of understanding ourpast should integrate the use of uniparental and biparental markers. Y-chromo-some-speciÞc markers and mtDNA can provide information about male and fe-male gene ßow patterns and are useful for detecting directional mating. Inaddition to the Y-chromosome and mtDNA evidence, the autosomal evidenceprovides a picture of the global history of admixture through the generations.The precision of the autosomal admixture estimates is dependent on the numberand ��s of the markers studied. Although the analysis relied on only Þve autoso-mal markers, these markers showed high frequency differences between nativeAmerican, European, and West African populations and were informative forestimating admixture proportions. In our judgment, admixture studies based onlyon uniparental markers must be interpreted with caution because of the potentialeffect of directional mating. In places such as La Plata, the results of mtDNA,Y-chromosome, and autosomal DNA can be substantially different. NativeAmerican contribution is highest at the level of mtDNA, intermediate at the au-tosomal level, and lowest at the level of the Y chromosome. In addition, most ofthe individuals from La Plata who showed a native American mtDNA haplogroupor the DYS199*T native American allele also showed a genetic contribution atthe autosomal level that can be traced primarily to Europe.
Finally, we would like to stress that the characterization of population andindividual ancestry is important, and not just from the historical and anthropolog-ical perspectives. This subject far exceeds its scientiÞc limits; several constitu-tions in South American countries have been recently reformed (e.g., inArgentina in 1994, in Colombia in 1991, and in Ecuador in 1998). The newconstitutions recognize the existence and rights of indigenous people, but cur-rently there is no reliable deÞnition of what ��indigenous�� means for legal mat-ters. The inclusion and recognition of rights to the ��indigenous people�� in the

Admixture in Buenos Aires / 555
Argentinean National Constitution has raised controversial interpretations, andsome populations have become interested in the characterization of their geneticproÞles because of the potential need to defend their own rights (Bianchi andMartõnez Marignac 2001; Bianchi et al. 2003). In this sense, it is important tomention that the history of populations and individuals is complex, and under-standing this history requires gathering evidence from a variety of sources, notjust genetics.
Received 22 July 2003; revision received 26 November 2003.
Literature Cited
Avena, S.A., S.A. Goicoechea, J.M. Dugoujon et al. 2001. Analisis antropogenetico de los aportesindõgena y africano en muestras hospitalarias de la ciudad de Buenos Aires. Rev. Arg. Antro-pol. Biol. 3(1):79�99.
Bailliet, G., F. Rothhammer, F.R. Carnese et al. 1994. Founder mitochondrial haplotypes in Amerin-dian populations. Am. J. Hum. Genet. 54:27�33.
Barba, F.E. 1983. Tiempo: El momento historico de la fundacion de La Plata. In La Plata ciudadnueva, ciudad antigua: Historia, forma y estructura de un espacio urbano singular, F. Teran,ed. La Plata, Argentina: Universidad Nacional de La Plata, Instituto de Estudios de Admin-istracion Local, 16�27.
Batista dos Santos, S.E., J.D. Rodrigues, A.K. Ribeiro-dos-Santos et al. 1999. Differential contribu-tion of indigenous men and women to the formation of an urban population in the Amazonregion as revealed by mtDNA and Y-DNA. Am. J. Phys. Anthropol. 109(2):175�180.
Bejarano, M. 1967. Inmigracion y estructura demograÞca de La Plata: 1884�1914. Bulletin 6, Direc-cion de Museos, Monumentos y Lugares Historicos. La Plata, Argentina: Provincia de BuenosAires, Ministerio de Educacion y Subsecretarõa de Cultura, 13�35.
Bianchi, N.O., C. Catanesi, G. Bailliet et al. 1998. Characterization of ancestral and derived Y-chromo-some haplotypes of New World Native populations. Am. J. Hum. Genet. 63:1,862�1,871.
Bianchi, N.O., and V.L. Martõnez Marignac. 2001. Aportes de la genetica y antropologõa moleculara los derechos indõgenas argentinos por la posesion de tierras. La Plata, Argentina: Institutode Estudios Judiciales de la Suprema Corte de Justicia, Provincia de Buenos Aires, ConsejoDepartamental La Plata, 59�93.
Bianchi, N.O., V.L. Martõnez Marignac, and R. Guanuco. 2003. IdentiÞcacion de Amerindios pormedio del analisis de and: Su aplicacion en los litigios por posesion de tierras, y otros temaslegales y eticos. Interciencia 28:1�8.
Bravi, C.M., M. Sans, G. Bailliet et al. 1997. Characterization of mitochondrial and Y-chromosomehaplotypes in an Uruguayan population of African ancestry. Hum. Biol. 69:641�652.
Cavalli-Sforza, L.L., P. Menozzi, and A. Piazza. 1996. The History and Geography of Human Genes.London: Princeton University Press.
Chakraborty, R. 1985. Gene identity in racial hybrids and estimation of admixture rates. In GeneticMicrodifferentiation in Man and Other Animals, J.V. Neel and Y. Ahuja, eds. New Delhi:Indian Anthropological Association, 171�180.
Dipierri, J.E., E. Alfaro, V.L. Martõnez Marignac et al. 1998. Paternal directional mating in twoAmerindian subpopulations located at different altitudes in northwestern Argentina. Hum.Biol. 70:1,001�1,010.
Elizalde, D. 1975. La migracion interna en la Argentina: 1960�1970. Buenos Aires, Argentina:INDEC.

556 / martinez marignac et al.
Fouscaldo, L. 1985. El proceso de constitucion del proletariado rural de origen indõgena en el Chaco.In Antropologõa, M. Lischetti, comp. Buenos Aires, Argentina: Eudeba, 337�359.
Goicoechea, A.S., S.A. Avena, J.M. Dugoujon et al. 2000. La presencia africana en poblacionesaborigenes y cosmopolitas de la Argentina. In Resumenes y Programa Sintetico: VI Congresode la Asociacion Latino Americana de Antropologõa Biologica. Montevideo, Uruguay: De-partamento de Publicaciones de la Facultad de Humanidades y Ciencias de la Educacion, 24.
Goicoechea, A.S., F.R. Carnese, C. Dejean et al. 2001. Genetic relationships between Amerindianpopulations of Argentina. Am. J. Phys. Anthropol. 115(2):133�143.
Guerreiro, J.F., E.J. dos Santos, and S.E. dos Santos. 1994. Effect of average heterozygosity on thegenetic distance of several Indian tribes from the Amazon region. Ann. Hum. Biol. 21(6):589�595.
Herren, R. 1992. La conquista erotica de las Indias. Buenos Aires, Argentina: Planeta.INDEC. 1997. La migracion internacional en la Argentina: sus caracterõsticas e impacto. Estudios
29. Buenos Aires, Argentina: INDEC.Lewis, P.O., and D. Zaykin. 2001. Genetic Data Analysis: Computer Program for the Analysis of
Allelic Data, Version 1.0 (d16c). Free program distributed by the authors; available at http://lewis.eeb.uconn.edu/lewishome/software.html
Martõnez Marignac, V.L., C.M. Bravi, H.B. Lahitte et al. 1999. Estudio del ADN mitocondrial de unamuestra de la ciudad de La Plata. Rev. Arg. Antropol. Biol. 2(1):281�300.
Parra, E.J., R.A. Kittles, G. Argyropoulos et al. 2001. Ancestral proportions and admixture dynamicsin geographically deÞned African Americans living in South Carolina. Am. J. Phys. Anthro-pol. 114(1):18�29.
Parra, E.J., A. Marcini, J. Akey et al. 1998. Estimating African American admixture proportions byuse of population-speciÞc alleles. Am. J. Hum. Genet. 63(6):1,839�1,851.
Pfaff, C.L., E.J. Parra, C. Bonilla et al. 2001. Population structure in admixed populations: Effect ofadmixture dynamics on the pattern of linkage disequilibrium. Am. J. Hum. Genet. 68(1):198�207.
Picotti, D.V. 2001. El negro en la Argentina: Presencia y negacion. Buenos Aires, Argentina: Edi-tores de America Latina.
Pritchard, J.K., M. Stephens, and P. Donnelly. 2000. Inference of population structure using multi-locus genotype data. Genetics 155(2):945�959.
Rodriguez-DelÞn, L.A., V.E. Rubin-de-Celis, and M.A. Zago. 2001. Genetic diversity in an Andeanpopulation from Peru and regional migration patterns of Amerindians in South America: Datafrom Y chromosome and mitochondrial DNA. Hum. Hered. 51(1�2):97�106.
Sala, A., G. Penacino, R. Carnese et al. 1999. Reference database of hypervariable genetic markersof Argentina: Application for molecular anthropology and forensic casework. Electrophoresis20(8):1,733�1,739.
Salzano, F.M., and M.C. Bortolini. 2002. The Evolution and Genetics of Latin American Populations.Cambridge: Cambridge University Press.
Sans, M. 2000. Admixture studies in Latin America: From the 20th to the 21st century. Hum. Biol.72(1):155�177.
Sans, M., T.A. Weimer, M.H. Franco et al. 2002. Unequal contributions of male and female genepools from parental populations in the African descendants of the city of Melo, Uruguay. Am.J. Phys. Anthropol. 118(1):33�44.
Shriver, M.D., M.W. Smith, L. Jin et al. 1997. Ethnic-afÞliation estimation by use of population-speciÞc DNA markers. Am. J. Hum. Genet. 60(4):957�964.
Torroni, A., Y.S. Chen, O. Semino et al. 1994. mtDNA and Y-chromosome polymorphisms in fourNative American populations from southern Mexico. Am. J. Hum. Genet. 54(2):303�318.
Torroni, A., K. Huoponen, P. Francalacci et al. 1996. ClassiÞcation of European mtDNAs from ananalysis of three European populations. Genetics 144:1,835�1,850.
Tourret, M.N., C.I. Catanesi, and L. Vidal-Rioja. 2000. Variability of the F13B locus in South Ameri-can populations. Hum. Biol. 72(4):707�714.

Admixture in Buenos Aires / 557
Trachtenberg, E.A., G. Keyeux, J.E. Bernal et al. 1996. Results of Expedicion Humana. I. Analysisof HLA class II (DRB1-DQA1-DPB1) alleles and DR-DQ haplotypes in nine Amerindianpopulations from Colombia. Tissue Antigens 48(3):174�181.
Ubelaker, D.H. 1992. Patterns of demographic change in the Americas. Hum. Biol. 64(3):361�379.Underhill, P.A., L. Jin, R. Zemans et al. 1996. A pre-Columbian Y-chromosome-speciÞc transition
and its implication for human evolutionary history. Proc. Natl. Acad. Sci. USA 93:196�200.Zago, M.A., W.A. Silva Jr., M.H. Tavella et al. 1996. Interpopulational and intrapopulational genetic
diversity of Amerindians as revealed by six variable number of tandem repeats. Hum. Hered.46(5):274�289.
Copyright © 2022 FDOKUMEN




















