
Biofuels (alcohols and biodiesel) applications as fuels for ...
Upload
independentCategory
view
3download
0
Characterization of PartiallyFolded Intermediates ofPapain in Presence ofCationic, Anionic, and NonionicDetergents at Low pH
Aabgeena Naeem
Sadaf Fatima
Rizwan Hasan KhanInterdisciplinary
Biotechnology Unit,Aligarh Muslim University,
Aligarh-202002, India
Received 22 September 2005;revised 17 March 2006;accepted 23 March 2006
Published online 5 April 2006 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/bip.20520
Abstract: A systematic investigation of the effects of detergents [Sodium dodecyl sulphate (SDS),hexa decyltrimethyl ammonium bromide (CTAB) and Tween-20] on the structure of acid-unfoldedpapain (EC.3.4.22.2) was made using circular dichroism (CD), intrinsic tryptophan fluorescence,and 1-anilino 8-sulfonic acid (ANS) binding. At pH 2, papain exhibits a substantial amount of sec-ondary structure and is relatively less denatured compared with 6 M GdnHCl (guanidine hydro-chloride) but loses the persistent tertiary contacts of the native state. Addition of detergents causedan induction of a-helical structure as evident from the increase in the mean residue ellipticity valueat 208 and 222 nm. Near-UV CD spectra also showed the regain of native-like spectral features inthe presence of 8 mM SDS and 3.5 mM CTAB. Induction of structure in acid-unfolded papain wasgreater in the presence SDS followed by CTAB and Tween-20. Intrinsic tryptophan fluorescencestudies indicate the change in the environment of tryptophan residues upon addition of detergents toacid-unfolded papain. Addition of 8 mM SDS resulted in the loss of ANS binding sites exhibited by adecrease in ANS fluorescence intensity, suggesting the burial of hydrophobic patches. MaximumANS binding was obtained in the presence of 0.1 mM Tween-20 followed by CTAB, indicating a com-pact ‘‘molten-globule’’–like conformation with enhanced exposure of hydrophobic surface area. Acid-unfolded papain in the presence of detergents showed the partial recovery of enzymatic activity. Theseresults suggest that papain at low pH and in the presence of SDS exists in a partially folded state char-acterized by native-like secondary structure and tertiary folds. While in the presence of Tween, acid-unfolded papain exists as a compact intermediate with molten-globule–like characteristics, viz.enhanced hydrophobic surface area and retention of secondary structure. While in the presence ofCTAB it exists as a compact intermediate with regain of native-like secondary and partial tertiarystructure as well as high ANS binding with the partially recovered enzymatic activity, i.e., a moltenglobule state with tertiary folds. # 2006 Wiley Periodicals, Inc. Biopolymers 83: 1–10, 2006
This article was originally published online as an accepted preprint. The ‘‘Published Online’’ datecorresponds to the preprint version. You can request a copy of the preprint by emailing theBiopolymers editorial office at [email protected]
Keywords: anion-induced state; circular dichroism; molten globule; papain; partially foldedintermediate state
INTRODUCTION
To understand the factors governing the formation of
three-dimensional protein structures, it is important
to elucidate the hierarchy of interactions that stabilize
the native and molten globule states. A molten globule
exists as an intermediate between native and denatured
states, is defined as a compact conformation with a
Correspondence to: R. H. Khan; e-mail: [email protected] or [email protected]
Biopolymers, Vol. 83, 1–10 (2006)
# 2006 Wiley Periodicals, Inc.
1

comparable amount of native-like secondary structure
but a high enhancement in the intramolecular motion,
i.e., a largely disordered tertiary structure.1 A large
barrier to crack the protein-folding code is the high
cooperativity of folding reactions at equilibrium. A
solvent perturbation study is a good approach to evalu-
ate the stabilizing force of a protein structure. The
interesting cosolvents in this context can be either
polyols and sugars or alcohol and surfactants since
they have potential protecting effects on various types
of protein denaturation.2,3
One of the main objectives in the solution of the
‘‘protein-folding problem’’ is to characterize these par-
tially folded stable intermediate states.4,5 Several stud-
ies have shown that the amount of secondary structure
and the compactness of the intermediate state formed
in the folding pathway of protein are not necessarily
close to those of the native state, but vary greatly
depending on the protein species.6,7 This suggests the
presence of various intermediate states, from one close
to the fully unfolded state to one close to the native
state, depending upon the protein and experimental
conditions. There has been considerable interest in
whether a new type of intermediate state, which may
be located between the native state and the molten
globule state or the molten globule state and the
unfolded state, exists.4,8 Characterization of the transi-
tion state of proteins, which is believed to be located
between the molten globule state and the native state,
is an important topic in protein folding.7,9–11 A number
of equilibrium intermediates have been detected and
characterized. The high cooperativity and complexity
of the protein-folding process makes the characteriza-
tion of conformational transitions and equilibrium in-
termediate states hard to achieve; different methods
are employed for the characterization of folding inter-
mediates and for understanding the different factors
leading to the formation and stabilization of these
intermediates. There is evidence that supports the idea
that the molten globule may also possess well-defined
tertiary contacts.12–15
Surfactants have a polar head and a nonpolar tail;
the latter causes a hydrophobic effect, which arises pri-
marily from the strong attractive forces between water
molecules themselves. The forces involved between
protein and surfactants are both electrostatic and hydro-
phobic. Surfactants are unique among protein denatur-
ants; even as low as 1 mM SDS, for instance, can often
alter protein conformation. The addition of surfactant
can either increase or decrease the helical conforma-
tion. This is unlike the unfolding of most proteins in 5–
6 M guanidine hydrochloride or 8–10 M urea. Deter-
gent extraction of integral proteins from the membranes
often results in disruption of protein subunits or even
converts � forms into helices.16,17
Papain, one of the sulfhydryl proteases from the la-
tex of Carica papaya, is well studied because of the
molecular structure and many industrial applications.
The interesting feature of this protein is its molecular
structure, which is divided essentially in the form of
two distinct lobes separated by a cleft. The structural
stability of each lobe is at least partly due to the hydro-
phobic core to which many side chains contribute. It is
suggested that the folding of each lobe is independent
of the other and this feature of the papain may indicate
its binuclear nature. The folding of the polypeptide
chain is seen to be different in the two lobes.18,19 Here
we report the different equilibrium intermediates of pa-
pain in the presence of different detergents. The
approach taken was to obtain different intermediates
starting from a common point, i.e., the acid-unfolded
state to detergent-induced states.
MATERIALS AND METHODS
Materials
Papain (EC.3.4.22.2) from the dried latex of papaya was
purchased from Sigma (St. Louis, MO, USA) and the pu-
rity was checked by SDS–PAGE. Sodium dodecyl sulphate
(SDS), hexa decyltrimethyl ammonium bromide (CTAB),
and Tween (Sigma) were used without any further purifica-
tion. All other chemicals used in this study were of analyti-
cal grade.
Methods
All of the measurements were carried out at room tempera-
ture. Typically, papain stock solution (5 mg/mL) was pre-
pared in and dialyzed against 20 mM Gly HCl buffer,
pH 2.0. The concentration of native papain in 20 mM phos-
phate buffer was determined from the extinction coefficient
of 25 M–1cm–1 at 280 nm, pH 7. For studying the effect of
detergents on acidic pH, the unfolded protein was incu-
bated at room temperature for 30 min to carry out spectro-
scopic measurements.
Circular Dichroism Measurements. Circular dichroism
(CD) was measured with a JASCO J 715 spectropolarim-
eter (Tokyo, Japan) calibrated with ammonium D-10
camphorsulfonate. Cell path lengths of 0.1 and 1 cm were
used for scanning between 250–200 and 300–250 nm,
respectively. Protein concentrations of the samples were
typically 20 �M for far-UV CD and 50 �M for near-UV
CD studies. The results were expressed as the mean residue
ellipticity (MRE in deg cm2/dmol), which is defined as
MRE ¼ � obs ðmdegÞ=ð10� n� Cp� 1Þ
where � obs is the observed ellipticity in degrees, Cp is the
molar fraction, and l is the length of the light path in cm.
The �-helical content of papain was calculated from the
2 Naeem, Fatima, and Khan
Biopolymers DOI 10.1002/bip
MRE value at 222 nm using the following equation as
described by Chen et al.20:
% �� helix ¼ MRE222 � 2340=30300� 100
Fluorescence Measurements. Fluorescence spectra were
recorded with a Shimadzu RF 540 spectrofluorophotometer
(Japan) in a 10-mm path-length quartz cell. Samples con-
taining different concentrations of detergents were equili-
brated at room temperature for 30 min before recording for
tryptophan fluorescence measurements. The excitation
wavelength was 280 nm and the emission was recorded
from 300 to 400 nm. The final protein concentration was
10 �M for all experiments.
ANS-Fluorescence Measurements. 8-Anilino-1-naphtha-
lene-sulfonic acid (ANS) binding was measured by fluores-
cence emission with excitation at 380 nm and emission
was recorded from 400 to 600 nm. Typically, ANS concen-
tration was 100 molar excess of protein concentration and
protein concentration was in the vicinity of 10 �M.
Percent Residual Activity. In the presence of detergents the
% residual activity was calculated relative to native papain as
100% from the hydrolysis of casein at pH 7.0 and 378C.
Caseinolytic Activity Measurement. Proteolytic activity
measurements were made using 2% casein as substrate and a
stock enzyme concentration of 0.1 mg/mL. Casein (0.25 mL)
was treated with enzyme solution (0.3 mL) in 0.1 M sodium
phosphate buffer, pH 7.0 at 378C for 15 min, and the reaction
was stopped by the addition of 5% TCA (0.25 mL). The pre-
cipitated material, from each reaction mixture, was removed
by centrifugation and the TCA soluble fraction was assayed
by the method of Lowry et al.21 An appropriate blank solu-
tion was also prepared and absorbance values at 700 nm were
measured with reference to the blank. The absorbance values
obtained could be correlated with the amount of product
formed. The results are expressed in terms of % residual ac-
tivity compared with native.
Data Analysis
The folding curves for the D$ N transition were normalized to
the apparent fraction folded, fN, using the following equation22:
fN ¼ ðY � YDÞ=ðYN � YDÞwhere Y is the observed variable parameter and YN and YDare the values of the variable characteristics of the folded
and unfolded conformations. The difference in free energy
between the folded and the unfolded states, DG was calcu-
lated by the following equations:
�G ¼ �RT lnK ¼ �RT ln ½ð1� fDÞ=fD�¼ �RT ln ½fN=1� fN�:
fN ¼ 1� fD or fN þ fD ¼ 1
where fN is the fraction folded, fD is the fraction unfolded, Kis the equilibrium constant, R is the gas constant (1.987 cal/
deg/mol), and T is the absolute temperature (298 K).
RESULTS AND DISCUSSION
CD Measurements
The effect of detergents on acid-unfolded papain was
studied by far UV and near UV circular dichroism.
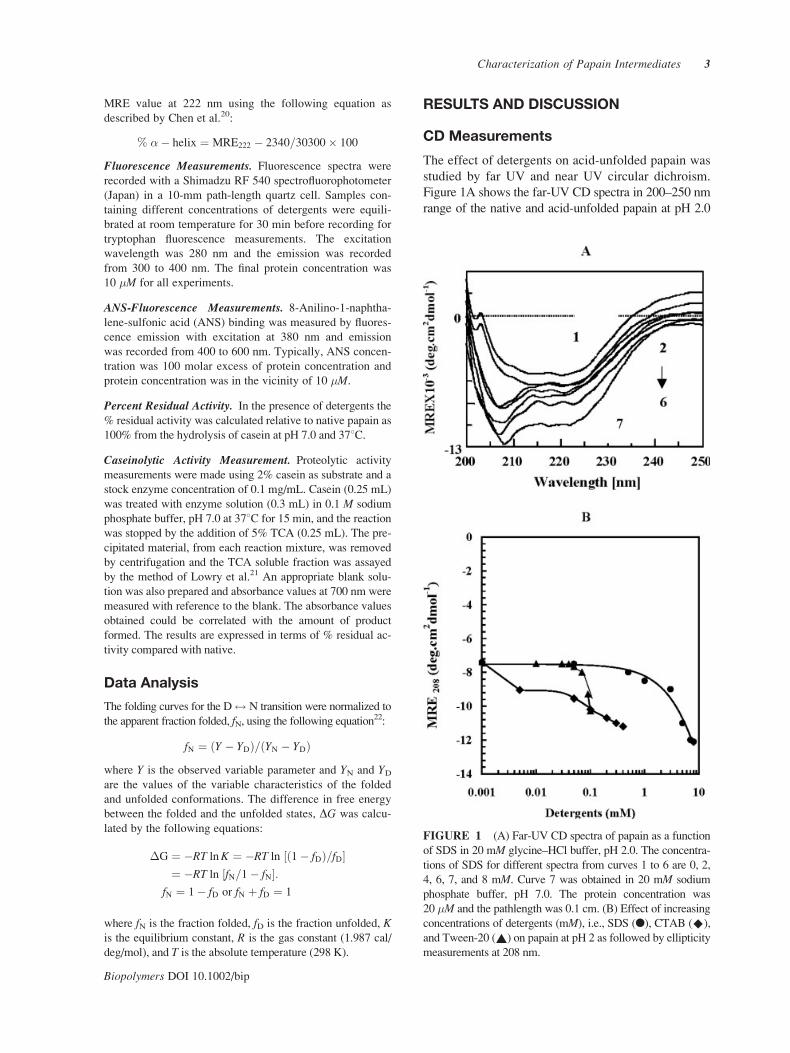
Figure 1A shows the far-UV CD spectra in 200–250 nm
range of the native and acid-unfolded papain at pH 2.0
FIGURE 1 (A) Far-UV CD spectra of papain as a function
of SDS in 20 mM glycine–HCl buffer, pH 2.0. The concentra-
tions of SDS for different spectra from curves 1 to 6 are 0, 2,
4, 6, 7, and 8 mM. Curve 7 was obtained in 20 mM sodium
phosphate buffer, pH 7.0. The protein concentration was
20 �M and the pathlength was 0.1 cm. (B) Effect of increasing
concentrations of detergents (mM), i.e., SDS (l), CTAB (^),
and Tween-20 (~) on papain at pH 2 as followed by ellipticity
measurements at 208 nm.
Characterization of Papain Intermediates 3
Biopolymers DOI 10.1002/bip
both in the absence and presence of various concen-
trations of SDS (0–8 mM). Curve 1 represents the
far-UV CD spectra of acid-unfolded papain, whereas
the far-UV CD spectrum of native papain is repre-
sented by curve 7. The spectra of acid-unfolded pro-
tein obtained in the presence of different concentra-
tions (2–8 mM) of SDS are shown in curves 2 and 6.
All of the spectra were characterized by the presence
of minima at 208 and 222 nm, the characteristic fea-
tures of helical secondary structure. As can be seen
from the figure, papain in acidic conditions retains
some amount of secondary structure. Addition of SDS
on acid-unfolded papain up to 8.0 mM led to an in-
crease in helical content as evidenced by an increase in
negative MRE value at 208 and 222 nm, hence ap-
proaches the native value. The 12-carbon hydrophobic
chain of SDS creates a nonpolar environment that de-
creases the polarity of the solvent at pH 2 causing an
increase in hydrophobicity and also interacting with
the hydrophobic cluster of the protein. The addition
of sulphate anion to papain leads to electrostatic inter-
actions with the positively charged centers on the pro-
tein (binding of SO42– anion), so as to effectively shield
the repulsive forces, which favor unfolding, and con-
sequently the intrinsic hydrophobic interactions of
protein in an aqueous environment manifest them-
selves.11 This results in protein folding, for the protein
to adopt a compact structure with increased hydropho-
bic surface area. Further increase in the SDS concen-
tration beyond that which maximum ellipticity was
observed leads to the transition of the SDS monomer
to the formation of micelles. The critical micellar con-
centration of SDS in water is reported to be 8.3 mM,
whereas it further increases with increase in dielectric
constant of the solvent.23 Hiramatsu and Yang16 have
reported a similar study on cytochrome c.SDS induces the anion transition in acid-unfolded
papain. The helical contents calculated from the MRE
value at 222 nm by the method of Chen et al.20 for the
native state at pH 7.0, the acid-unfolded state at pH 2.0,
and the acid-unfolded state at pH 2.0 in the presence of
8 mM SDS were 29, 16.7, and 28%, respectively.
The effect of detergents, i.e., cationic (CTAB),
anionic (SDS), and nonionic (Tween-20) on the acid-
unfolded state of papain at pH 2.0 was studied by
measuring MRE at 208 nm (Figure 1B). All three
detergents used were found to induce helical struc-
ture on acid-unfolded papain. However, SDS was
found to be more effective than CTAB or Tween 20,
with maximum ellipticity being observed at 8 mM.
Further increases in detergent concentration led to
protein precipitation. Detergents form micelles at a
particular concentration called the critical micelle
concentration. It has been observed that, at concen-
trations beyond their ‘‘CMC’’ values, detergents in-
duce precipitation in proteins. Detergent-induced tran-
sition of acid-unfolded papain as observed in this study
can be explained on the basis of addition of deter-
gent, known to stabilize the helical structure of a pro-
tein. A summary of respective maximum MREs at
208 and 222 nm of detergents, their Cm values (the
concentration of detergent at which half of the struc-
ture is induced), and their �-helix is given in Table I.
Figure 2A shows the far UV CD spectra in the
200–250 nm range of native papain (curve 6), acid-
unfolded papain, and in the presence of various
detergents at their respective maximum concentrations.
As can be seen from the figure, papain in acidic condi-
tions (curve 2) retains some amount of secondary
structure compared with 6M GdnHCl (curve 1). Maxi-
mum induction of structure at 208 nm is seen in 8 mMSDS (curve 5) followed by 3.5 mM CTAB (curve 4)
and 0.1 mM Tween-20 (curve 3). Hence SDS is found
to be most effective in inducing secondary structure
almost approaching native (curve 6), among the above-
mentioned detergents.
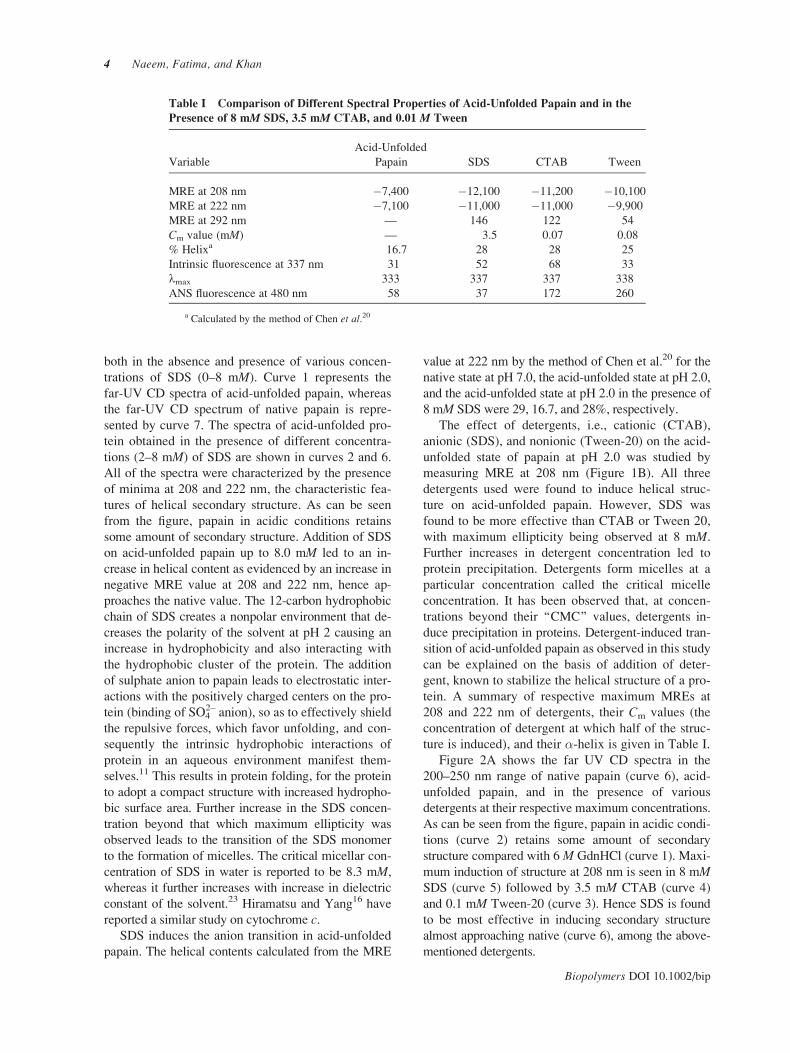
Table I Comparison of Different Spectral Properties of Acid-Unfolded Papain and in the
Presence of 8 mM SDS, 3.5 mM CTAB, and 0.01 M Tween
Variable
Acid-Unfolded
Papain SDS CTAB Tween
MRE at 208 nm �7,400 �12,100 �11,200 �10,100
MRE at 222 nm �7,100 �11,000 �11,000 �9,900
MRE at 292 nm — 146 122 54
Cm value (mM) — 3.5 0.07 0.08
% Helixa 16.7 28 28 25
Intrinsic fluorescence at 337 nm 31 52 68 33
lmax 333 337 337 338
ANS fluorescence at 480 nm 58 37 172 260
a Calculated by the method of Chen et al.20
4 Naeem, Fatima, and Khan
Biopolymers DOI 10.1002/bip

Figure 2B shows near-UV CD spectra in the 300–
250 nm range of native papain (curve 5), acid-
unfolded papain (curve 1), and acid-unfolded papain
in the presence of 0.1 mM Tween-20 (curve 2),
3.5 mM CTAB (curve 3), and 8 mM SDS (curve 4).
As can be seen from the figure, native papain is char-
acterized by the presence of three prominent peaks at
292, 280, and 269 nm. At pH 2.0, the protein com-
pletely lost its tertiary structure with the disappear-
ance of all of the prominent peaks. In the presence of
8 mM SDS, all three peaks reappear but the MRE
value does not approach the native value. Whereas in
the case of 3.5 mM CTAB, the only peak reappear-
ance was at 292 nm. In the presence of 0.1 mMTween protein there was a peak at around 272 nm in
comparison to acid-unfolded papain, in which the
spectrum was almost zero. Thus, acid-unfolded pa-
pain in the presence of 8 mM SDS and 3.5 mMCTAB retains its partial tertiary structure.
Intrinsic Fluorescence
The intrinsic fluorescence maximum (lmax) is an
excellent parameter to monitor the polarity of the
tryptophan environment in the protein and is sensi-
tive to the protein conformation.24 Contribution of
detergents alone to the emission spectra was taken
into account and we report here the subtracted fluo-
rescence spectra. To rule out aggregation of protein
and hence light scattering, the samples were read at
350 nm and the negligible absorbance was observed.
Intrinsic tryptophan fluorescence studies were per-
formed to monitor the modification of the microen-
vironment of tryptophan in the presence of deter-
gents (SDS, Tween-20, and CTAB). The emission
maximum of the native protein was obtained at 337 nm
(curve 2) and a red shift of 13 nm was observed in
the completely unfolded state in 6 M Gdn HCl (curve
1) as all the five tryptophans are exposed to the sol-
vent. A decrease in the fluorescence intensity was
observed at pH 2 (curve 6) with a blue shift of
17 nm, i.e., lmax at 333 nm with a 30% decrease in
fluorescence signal relative to 6 M Gdn HCl. This ob-
servation suggests that the papain molecule at pH 2.0
acquires a conformation different from native and
denatured states. This blue-shift fluorescence of pa-
pain at pH 2.0 can be attributed to the conformational
changes in the vicinity of surface-exposed trypto-
phans, presumably because of internalization in a
more hydrophobic environment. A similar blue-
shifted fluorescence has been reported earlier for
glucose isomerase,25 bovine growth hormone,26 and
interferon-�.27 An alternative possibility is that sur-
face-exposed tryptophans are quenched differently
from the internal tryptophans at low pH without inter-
nalizing them, leading to a greater proportion of the
remaining signal coming from the more buried tryp-
tophans that are located in the interior of the native
protein. Upon addition of 8 mM SDS, the internalized
trytophans in the acid-unfolded state (curve 6) get
partially exposed from a nonpolar to a polar environ-
ment as indicated by a red shift of 4 nm and an
increase in fluorescence intensity (curve 3). This indi-
cates that tryptophan residues of the acid-unfolded
FIGURE 2 (A) Far-UV CD spectra of acid-unfolded pa-
pain (curve 2) in the presence of detergents: 8 mM SDS
(curve 5), 3.5 mM CTAB (curve 4), and 0.1 mM Tween-20
(curve 3) at their respective maximium concentrations in
20 mM glycine–HCl buffer, pH 2.0. Curve 1 represents Gdn–
HCl–denatured papain. Curve 6 represents papain at pH 7.
The protein concentration was 20 �M and the pathlength was
0.1 cm. (B) Near-UV CD spectra of native papain (curve 5),
acid-unfolded papain at 8 mM SDS (curve 4), 3.5 mM CTAB
(curve 3), and 0.1 mM Tween-20 (curve 2). Curve 1 repre-
sents papain at pH 2. The protein concentration was 50 �Mand the pathlength was 1 cm.
Characterization of Papain Intermediates 5
Biopolymers DOI 10.1002/bip

state in the presence of SDS acquire a state that
resembles more closely native protein. Similar spectra
were obtained in 3.5 mM CTAB (curve 4) and
0.1 mM Tween (curve 5) but the effect was less pro-
nounced in CTAB and the least in Tween. This result
is consistent with our CD results, in which the sulfate
ions present in the electronegative series were found
to be effective in inducing secondary and partial terti-
ary structure. The electronegative series given by
Goto and collaborators28–31 is as follows:
FeðCNÞ3�6 >SO2�4 >CCl3COO�>SCN�>ClO�4
>I�>NO�3 >CF3COO
� >Br� >Cl�
Figure 3B depicts the fluorescence intensity at 337 nm
as a function of detergent concentration (mM). On
increasing the concentration of detergents there was
increase in fluorescence intensity. More enhancement
was found in case of SDS followed by CTAB and
Tween-20.
As papain consists of more basic amino acids, sul-
phate ions, being negatively charged, interacted with
the positive charge on the protein more effectively
than cationic (CTAB) or nonionic (Tween 20) deter-
gents. As the concentration of SDS (0–8 mM) is
increased, it acts as source of sulphate anion, which
binds positively charged groups of the protein result-
ing in screening of the repulsive effect, thus over-
coming charge–charge repulsion, causing a compact
structure. Alternatively, the electrostatic effects of
the formation of ionic couples at the protein surface
due to specific sulfate anion on the conformation of
acid-unfolded papain can be partially interpreted
within the framework of Debye–Huckel theory with
simple electrostatic consideration. The sulphate
anion exerts an ‘‘ionic atmosphere’’ effect around the
protein. The counterion presence around the charge
groups will weaken the repulsion, thus permitting the
other forces favoring folding to become relatively
strengthened and resulting in folding of the protein.
Thus, SDS is found to be effective in inducing sec-
ondary structure (as observed by far-UV CD) and
partial tertiary structure (as observed by near-UV CD
and tryptophan fluorescence).
ANS Fluorescence
The intermediate state induced by detergents is fur-
ther confirmed by extrinsic fluorescence studies. Since
ANS is known to bind to hydrophobic patches on pro-
FIGURE 3 (A) Tryptophan fluorescence emission spectra
of papain in the native state in 20 mM sodium phosphate
buffer, pH 7.0 (curve 2); acid-unfolded state in 20 mM gly-
cine–HCl buffer, pH 2.0 (curve 6); in the presence of 8 mMSDS (curve 3) at pH 2.0; in the presence of 3.5 mM CTAB
(curve 4) at pH 2.0; and in the presence of 0.1 mM Tween-20
(curve 5) at pH 2.0; curve 1 represents 6 M Gdn–HCl–dena-
tured state. The protein concentration was 10 �M. (B) Effect
of increasing concentrations of detergents (mM), i.e., SDS
(l) CTAB (^), and Tween-20 (~) on papain at pH 2 as fol-
lowed by fluorescence intensity at 337 nm.
6 Naeem, Fatima, and Khan
Biopolymers DOI 10.1002/bip
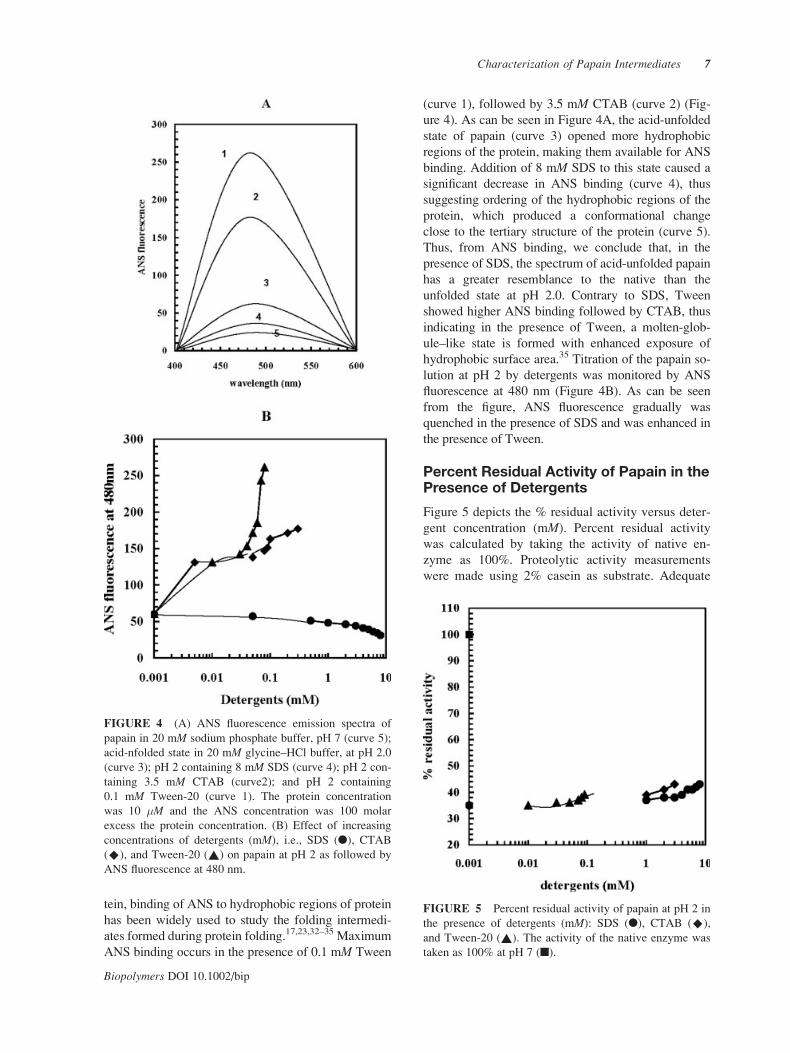
tein, binding of ANS to hydrophobic regions of protein
has been widely used to study the folding intermedi-
ates formed during protein folding.17,23,32–35 Maximum
ANS binding occurs in the presence of 0.1 mM Tween
(curve 1), followed by 3.5 mM CTAB (curve 2) (Fig-
ure 4). As can be seen in Figure 4A, the acid-unfolded
state of papain (curve 3) opened more hydrophobic
regions of the protein, making them available for ANS
binding. Addition of 8 mM SDS to this state caused a
significant decrease in ANS binding (curve 4), thus
suggesting ordering of the hydrophobic regions of the
protein, which produced a conformational change
close to the tertiary structure of the protein (curve 5).
Thus, from ANS binding, we conclude that, in the
presence of SDS, the spectrum of acid-unfolded papain
has a greater resemblance to the native than the
unfolded state at pH 2.0. Contrary to SDS, Tween
showed higher ANS binding followed by CTAB, thus
indicating in the presence of Tween, a molten-glob-
ule–like state is formed with enhanced exposure of
hydrophobic surface area.35 Titration of the papain so-
lution at pH 2 by detergents was monitored by ANS
fluorescence at 480 nm (Figure 4B). As can be seen
from the figure, ANS fluorescence gradually was
quenched in the presence of SDS and was enhanced in
the presence of Tween.
Percent Residual Activity of Papain in thePresence of Detergents
Figure 5 depicts the % residual activity versus deter-
gent concentration (mM). Percent residual activity
was calculated by taking the activity of native en-
zyme as 100%. Proteolytic activity measurements
were made using 2% casein as substrate. Adequate
FIGURE 4 (A) ANS fluorescence emission spectra of
papain in 20 mM sodium phosphate buffer, pH 7 (curve 5);
acid-nfolded state in 20 mM glycine–HCl buffer, at pH 2.0
(curve 3); pH 2 containing 8 mM SDS (curve 4); pH 2 con-
taining 3.5 mM CTAB (curve2); and pH 2 containing
0.1 mM Tween-20 (curve 1). The protein concentration
was 10 �M and the ANS concentration was 100 molar
excess the protein concentration. (B) Effect of increasing
concentrations of detergents (mM), i.e., SDS (l), CTAB
(^), and Tween-20 (~) on papain at pH 2 as followed by
ANS fluorescence at 480 nm.
FIGURE 5 Percent residual activity of papain at pH 2 in
the presence of detergents (mM): SDS (l), CTAB (^),
and Tween-20 (~). The activity of the native enzyme was
taken as 100% at pH 7 (n).
Characterization of Papain Intermediates 7
Biopolymers DOI 10.1002/bip

controls of substrate casein in the presence of deter-
gents were taken to rule out its chaperone activity
and changes induced by casein on acid-unfolded pa-
pain or being itself denatured by detergents. The pre-
cipitated material was removed and the TCA soluble
fraction was assayed by the method of Lowry et al.21
At pH 2, in the absence of detergents, the activity of
papain was reduced to 35%. While in the presence of
detergents, there was partial recovery of enzymatic
activity of acid-unfolded papain. The % recovery in
enzyme activity, calculated in the presence of 8 mMSDS, 3.5 mM CTAB, and 0.1 mM Tween-20, was
43, 41, and 39%, respectively. No distinct difference
was observed in the activity on acid-unfolded papain
among the detergents. This result is consistent with
near- and far-UV CD spectra, as acid-unfolded papain
in the presence of 8 mM SDS and 3.5 mM regains the
tertiary structure and almost native-like secondary
structure (Figure 2). The gain of signal in the near-UV
CD region with low activity may be attributed to the
initial occurrence of nonnative structures and subse-
quent reorganization toward the native one.
Thermodynamic Parameter Derivedfrom the Detergent-DependentTransition Curve
The fraction of protein-folded at pH 2 in the pres-
ence of detergents is shown in Figure 6. fN of papain
at pH 2 in the presence of detergents was calculated
by taking papain at pH 7 as the native value. In the
presence of SDS and CTAB at pH 2, no intermediate
state could be observed, as the curves were sharp,
suggesting a single-step two-state transition. No dis-
tinct transition could be seen in the Tween-20 curve
(fluorescence intensity at 337 nm). However, the tran-
sition curve of the ellipticity change at 208 nm shows a
single-step, two-state transition. Nonoverlap of the two
transitions suggests a noncooperative transition. The
calculated free energy of folding, DG, in the presence
of detergents is shown in Table II.36
CONCLUSION
Detergents have gained an important status in protein
chemistry and are applied in the biosciences and in
many (bio)technological processes owing to the fact
that detergents mimic the native, hydrophobic envi-
ronment of the phospholipid bilayer in vivo. Keeping
in view the wide applicability of detergents in mem-
brane protein solubilization, electrophoresis, etc., it
becomes imperative to develop a better understand-
ing of the effects of detergents on proteins. Acid-
unfolded papain in the presence of different deter-
gents attains different conformations, making avail-
able different types of information for the usage of
protein purification. The primary role of detergents
in protein purification is to affect solubilization. The
nondenaturing property is the most difficult issue to
address in the selection of a detergent. A precise def-
inition of denaturation is usually given in terms of
effects on secondary and tertiary structure. It is there-
fore essential to recognize the detergents that will ac-
complish the task of maintaining solubility while not
significantly compromising the native structure and
consequently the biological function of the protein.
Earlier scattering and hydrodynamic data were mainly
used to probe the structural changes in protein–deter-
gent complexes. However, more recently, spectro-
scopic approaches have gained more attention. Oneda
FIGURE 6 Fraction folded versus detergent: fraction
folded at acid-unfolded papain in the presence of SDS (l,
*), CTAB (^, ^), and Tween (~, ~) by the change in
MRE at 208 nm and fluorescence intensity at 337 nm.
Filled symbols represent the changes monitored by MRE
208 nm and open symbols represent changes monitored by
fluorescence intensity at 337 nm.
Table II DG Values for the Detergent-Induced
Transition of Acid-Denatured Papain Studied by MRE
Measurements at 208 nm and Fluorescence Intensity
Measurements at 337 nm
Probe
DGH2O
(kcal/mol)
SDS
DGH2O
(kcal/mol)
CTAB
DGH2O
(kcal/mol)
Tween-20
MRE208 �1.39 �1.85 �1.82
Fluorescence337 �1.23 �0.42 �0.014
8 Naeem, Fatima, and Khan
Biopolymers DOI 10.1002/bip

and Inouye37 have used Brij-35 to refold and recover
the recombinant human matrix metalloproteinase 7
(matrilysin) from inclusion bodies expressed by Esche-richia coli. Cardamone et al.38 have used CTAC (cetyl
tri methyl ammonium chloride), a cationic detergent
for spectroscopic and equilibrium binding analysis
using a recombinant porcine growth hormone. SDS at
a low surfactant/protein ratio acts as a structure-stabi-
lizing additive on BSA. A similar study shows that in-
teraction of HSA with surfactant becomes stronger
with increasing alkyl chain length.39 SDS is used to
solubilzed interferon and interleukin 2 (IL-2) from in-
clusion bodies. In our study, all three detergents were
able to induce �-helix in acid-unfolded papain, as
evident from the induction of MRE values at 208 and
222 nm. The anion-induced state at low pH in the pres-
ence of SDS showed a diminished ANS binding com-
pared with acid-unfolded papain or that of the Tween-
induced state. Moreover, in presence of SDS there was
gain of signal in the near-UV (aromatic) region of the
CD spectrum, suggesting an ordering of the tertiary
structure. Also, in the presence of SDS, acid-unfolded
papain recovers partial activity followed by CTAB and
Tween-20. While in the case of Tween, an intermedi-
ate state with native-like secondary structure, loss of
tertiary structure and high ANS binding was obtained.
This state can be characterized as a molten-globule–
like state. In the case of CTAB, the intermediate state
obtained can also be characterized as molten globule,
due to high ANS binding and a native-like secondary
structure that also retains native-like tertiary contacts.
These species may hence be described as compact
molten-globule–like intermediates, as similar proper-
ties have been observed for the molten globule states
of a number of proteins. Molten globules are partially
structured a protein-folding intermediate that adopt a
native-like overall backbone topology in the absence
of extensive detectable tertiary interactions. It is im-
portant to determine the extent of the specific tertiary
structure present in molten globules and to understand
the role of specific side-chain packing in stabilizing
and specifying the molten globule structure.40 Moreover,
the A-state of equine ferricytochrome c has native-likecompactness, native-like helix content, native helix–
helix interactions, and one native heme ligand.41–43
Tertiary A state has been reported for �-lactalbumin,14
RNase H,44 myoglobin,45 and ubiquitin.46 A summary
of observations on acid-unfolded papain in presence of
detergents at pH 2 is presented in Table I. The effect of
detergents on proteins depends on the nature of both
the detergent and the protein component; in addition,
the environmental conditions are of importance. In our
studies, SDS and CTAB proved to be the better deter-
gents, as the information obtained from experiments
shows that protein regains secondary as well as tertiary
structures. Nevertheless, there are still a lot of promis-
ing possibilities of protein–detergent interaction that
need to be explored for solubilization of inclusion
bodies and membrane protein purification.
The authors are very thankful for the facilities obtained at
AMU Aligarh. The authors are also thankful to DST–FIST
for financial support. A. N. is the recipient of CSIR-SRF
and S.F. is the recipient of CSIR-JRF.
REFERENCES
1. Ptitsyn, O. B. Adv Protein Chem 1995, 47, 83–87.
2. Jacnicke, R. Biochemistry 1991, 30, 3147–3161.
3. Naeem, A.; Khan, R. H. Int J Biochem Cell Biol.
2004, 36, 2281–2292.
4. Ptitsyn, O. B. Trends Biochem Sci 1995, 20, 376–379.
5. Barrick, D.; Hughson, F. M.; Baldwin, R. L.; J Mol
Biol 1994, 237, 588–601.
6. Ballery, N.; Desmadril, M.; Minard, P.; Yon, J. M.
Biochemistry 1993, 32, 708–714.
7. Kuwajima, K. Proteins 1989, 6, 87–103.
8. Matousschek, A.; Serrano, L.; Meiering, E. M.; By-
croft, M.; Ferscht, A. R. J Mol Biol 1992, 224, 837–
845.
9. Dryden, D.; Weir, M. P. Biochim Biophys Acta 1991,
1078, 94–100.
10. Schulman, B. A.; Kim, P. S.; Dobson, C. M.; Redfield, C.
Nat Struct Biol 1997, 4, 630–634.
11. Naeem, A.; Akram, M.; Khan, R. H. J Protein Chem
2004, 23, 185–195.
12. Naeem, A.; Khan, A.; Khan, R. H. Biochem Biophys
Res Commun 2005, 331, 1284–1294.
13. Song, J.; Bai, P.; Luo, L.; Peng, Z. Y. Protein Sci
2001, 10, 53–62.
14. Wu, L. C.; Kim, P. S. J Mol Biol 1998, 280, 175–182.
15. Shortle, D.; Ackerman M. S. Science 2001, 293, 487–
489.
16. Hiramatsu, K.; Yang, J. T. Biochim Biophys Acta
1983, 743, 106–114.
17. Das, T. P.; Mazumdar, S.; Mitra, S. Eur J Biochem
1998, 254, 662–670.
18. Edwin, F.; Jagannadham, M. V. Biochem Biophys Res
Commun 1998, 252, 654–660.
19. Naeem, A.; Khan, K. A.; Khan, R. H. Arch Biochem
Biophys 2004, 432, 79–87.
20. Chen, Y. H.; Yang, J. T.; Martinez, H. M. Biochemistry
1972, 114, 120–413.
21. Lowry, D. H.; Rosebrough, N. J.; Farr, A. L.; Randal,
R. J. J. Biol Chem 1951, 193, 265–275.
22. Tanford, C. Adv Protein Chem 1968, 23, 122–282.
23. Neugebauer, J. M. Methods Enzymol 1990, 182, 239–
253.
24. Stryer L.J Mol Biol 1965, 13, 482–495.
25. Pawar, S. A.; Deshpande V. V. Eur J Biochem 2000,
267, 6331–6338.
Characterization of Papain Intermediates 9
Biopolymers DOI 10.1002/bip

26. Holzman, T. E.; Dougherty, J. J.; Brems, D. N.; Mackenzie
N. E. Biochemistry 1990, 29, 1255–1261.
27. Nandi, P. K. Int J Biol Macromol 1998, 22, 23–31.
28. Goto, Y.; Takahashi, N.; Fink, A. L. Biochemistry
1990, 29, 3480–3488.
29. Goto, Y.; Nishikiori, S. J Mol Biol 1991, 222, 679–686.
30. Goto, Y.; Calciano, L. J.; Fink A. L. Proc Natl Acad
Sci USA 1990, 87, 573–577.
31. Fink, A. L.; Calciano, C. T.; Goto, Y.; Kurotsu, T.;
Palleros, D. R. Biochemistry 1994, 33, 12504–12511.
32. Matulis, D.; Lovrien, R. Biophys J 1998, 74, 422–429.
33. Stryer, L. Science 1968, 162, 526–540.
34. Matulis, D.; Baumann, C. G.; Bloomfield, U. A.; Lov-
rien, R. E. Biopolymers 1999, 49, 451–458.
35. Semisotnov, G. V.; Rodionava, N. A.; Razgulyaev, O. I.;
Uversky, V. N.; Gripas, A. F.; Gilmanshin R. I. Biopoly-
mers 1991, 31, 119–128.
36. Pace, C. N.; Shaw, K. L. Proteins 2000, 4, 1–7.
37. Oneda, H.; Inouye, K. J Biochem (Tokyo) 1999, 126,
905–911.
38. Cardamone, M.; Puri, N. K.; Sawyer, W. H.; Capon, R. J.;
Brandon, M. R. Biochim Biophys Acta 1994, 1206, 71–
82.
39. Gharibi, H.; Javadian, S.; Hashemianzadeh, M. Colloids
Surf A Physochem Eng Aspects 2004, 232, 77–86.
40. Arai, M.; Kuwajima, K. Adv Protein Chem 2000, 53,
209–282.
41. Hamada, D.; Kuroda, Y.; Kataoka, M.; Aimoto, S.;
Yoshimura, T.; Goto, Y. J Mol Biol 1996, 256, 172–
186.
42. Kataoka, M.; Hagihara, Y.; Mihara, K.; Goto, Y. J Mol
Biol 1993, 229, 591–596.
43. Colon, W.; Roder H.Nat Struct Biol 1996, 3, 1019–
1025.
44. Raschke, T. M.; Marqusee, S. Nat Struct Biol 1997, 4,
298–304.
45. Kay, M. S.; Baldwin R. L. Nat Struct Biol 1996, 3,
439–445.
46. Khorasanizadeh, S.; Peters, I. D.; Roder, H. Nat Struct
Biol 1996, 3, 193–205.
Reviewing Editor: Yuji Kobayashi
10 Naeem, Fatima, and Khan
Biopolymers DOI 10.1002/bip
Copyright © 2022 FDOKUMEN




















