
Changes in the expression of ion channels, connexins and Ca2+-handling proteins in the sino-atrial...
13
Experimental Physiology 426 Exp Physiol 96.4 pp 426–438 Research Paper Research Paper Changes in the expression of ion channels, connexins and Ca 2+ -handling proteins in the sino-atrial node during postnatal development Eman Abd Allah 1,2 , James O. Tellez 1 , Joseph Yanni 1 , Thomas Nelson 1 , Oliver Monfredi 1 , Mark R. Boyett 1 ∗ and Halina Dobrzynski 1 ∗ 1 Cardiovascular Medicine, Faculty of Medical and Human Sciences, University of Manchester, Core Technology Facility, 46 Grafton Street, Manchester M13 9NT, UK 2 Department of Physiology, Faculty of Medicine, Assiut University, Assiut, Egypt There are important postnatal changes in the sino-atrial node (SAN), the pacemaker of the heart. Compared with the neonate, the adult has a slower intrinsic heart rate and a longer SAN action potential. These changes may be due to differences in ion channel expression. Consequently, we investigated postnatal developmental changes in the expression of ion channels and Ca 2+ - handling proteins in the SAN to see whether this is indeed the case. Using quantitative PCR, in situ hybridization and immunohistochemistry, we investigated the expression of ion channels, Ca 2+ -handling proteins and connexins in the SAN from neonatal (2–7 days of age) and adult (∼6 months of age) New Zealand White rabbits. The spontaneous beating rate of adult SAN preparations was 21% slower than that of neonatal preparations. During postnatal development, quantitative PCR revealed a significant decline in the SAN of the following mRNAs: HCN4 (major isoform responsible for I f ), Na V 1.5 (responsible for I Na ), Ca V 1.3 (in part responsible for I Ca,L ) and NCX1 (responsible for inward I NaCa ). These declines could be responsible for the slowing of the pacemaker during postnatal development. There was a significant decline during development in mRNA for delayed rectifier K + channel subunits (K V 1.5, responsible for I K,ur , K V LQT1 and minK, responsible for I K,s , and ERG, responsible for I K,r ) and this could explain the prolongation of the action potential. In situ hybridization confirmed the changes observed by quantitative PCR. In addition, immunohistochemistry revealed hypertrophy of nodal cells during postnatal development. Moreover, there were complex changes in the expression of Ca 2+ - handling proteins with age. In summary, there are significant postnatal changes in the expression of ion channels and Ca 2+ -handling proteins in the SAN that could explain the established changes in heart rate and action potential duration that occur during normal development. (Received 15 October 2010; accepted after revision 24 January 2011; first published online 28 January 2011) Corresponding author H. Dobrzynski: Cardiovascular Medicine, Faculty of Medical and Human Sciences, University of Manchester, Core Technology Facility, 46 Grafton Street, Manchester M13 9NT, UK. Email: [email protected] It is well known that the heart rate of the neonate is higher than that of the adult in many species, including rabbits (Toda, 1980; Yang et al . 2006) rats (Huang et al . 2007), guinea-pigs (Jones et al . 2004) and humans (Huang et al . 2007). For example, in humans, the heart rate is ∗ M. R. Boyett and H. Dobrzynski are joint senior authors. ∼140 beats min −1 in the neonate and ∼80 beats min −1 in the adult (Huang et al . 2007). The sino-atrial (SAN), the primary pacemaker of the heart, initiates the heart beat (Satoh, 2003; Boyett et al . 2006). The action potential in the rabbit SAN shows many changes during postnatal development, including a reduction in spontaneous beating rate, an increase in action potential duration (APD), and an increasingly negative maximal diastolic DOI: 10.1113/expphysiol.2010.055780 C 2011 The Authors. Journal compilation C 2011 The Physiological Society ) at The University of Manchester Library on February 5, 2014 ep.physoc.org Downloaded from Exp Physiol (
-
Upload
eastanglia -
Category
Documents
-
view
3 -
download
0
Transcript of Changes in the expression of ion channels, connexins and Ca2+-handling proteins in the sino-atrial...

Exp
erim
enta
lPhy
siol
ogy
426 Exp Physiol 96.4 pp 426–438
Research PaperResearch Paper
Changes in the expression of ion channels, connexinsand Ca2+-handling proteins in the sino-atrial nodeduring postnatal development
Eman Abd Allah1,2, James O. Tellez1, Joseph Yanni1, Thomas Nelson1, Oliver Monfredi1, Mark R. Boyett1∗
and Halina Dobrzynski1∗
1Cardiovascular Medicine, Faculty of Medical and Human Sciences, University of Manchester, Core Technology Facility, 46 Grafton Street,Manchester M13 9NT, UK2Department of Physiology, Faculty of Medicine, Assiut University, Assiut, Egypt
There are important postnatal changes in the sino-atrial node (SAN), the pacemaker of the heart.Compared with the neonate, the adult has a slower intrinsic heart rate and a longer SAN actionpotential. These changes may be due to differences in ion channel expression. Consequently,we investigated postnatal developmental changes in the expression of ion channels and Ca2+-handling proteins in the SAN to see whether this is indeed the case. Using quantitative PCR, insitu hybridization and immunohistochemistry, we investigated the expression of ion channels,Ca2+-handling proteins and connexins in the SAN from neonatal (2–7 days of age) and adult(∼6 months of age) New Zealand White rabbits. The spontaneous beating rate of adult SANpreparations was 21% slower than that of neonatal preparations. During postnatal development,quantitative PCR revealed a significant decline in the SAN of the following mRNAs: HCN4(major isoform responsible for I f ), NaV1.5 (responsible for I Na), CaV1.3 (in part responsiblefor I Ca,L) and NCX1 (responsible for inward I NaCa). These declines could be responsible for theslowing of the pacemaker during postnatal development. There was a significant decline duringdevelopment in mRNA for delayed rectifier K+ channel subunits (KV1.5, responsible for I K,ur,KVLQT1 and minK, responsible for I K,s, and ERG, responsible for I K,r) and this could explainthe prolongation of the action potential. In situ hybridization confirmed the changes observedby quantitative PCR. In addition, immunohistochemistry revealed hypertrophy of nodal cellsduring postnatal development. Moreover, there were complex changes in the expression of Ca2+-handling proteins with age. In summary, there are significant postnatal changes in the expressionof ion channels and Ca2+-handling proteins in the SAN that could explain the established changesin heart rate and action potential duration that occur during normal development.
(Received 15 October 2010; accepted after revision 24 January 2011; first published online 28 January 2011)Corresponding author H. Dobrzynski: Cardiovascular Medicine, Faculty of Medical and Human Sciences, Universityof Manchester, Core Technology Facility, 46 Grafton Street, Manchester M13 9NT, UK.Email: [email protected]
It is well known that the heart rate of the neonate ishigher than that of the adult in many species, includingrabbits (Toda, 1980; Yang et al. 2006) rats (Huang et al.2007), guinea-pigs (Jones et al. 2004) and humans (Huanget al. 2007). For example, in humans, the heart rate is
∗M. R. Boyett and H. Dobrzynski are joint senior authors.
∼140 beats min−1 in the neonate and ∼80 beats min−1 inthe adult (Huang et al. 2007). The sino-atrial (SAN), theprimary pacemaker of the heart, initiates the heart beat(Satoh, 2003; Boyett et al. 2006). The action potentialin the rabbit SAN shows many changes during postnataldevelopment, including a reduction in spontaneousbeating rate, an increase in action potential duration(APD), and an increasingly negative maximal diastolic
DOI: 10.1113/expphysiol.2010.055780 C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (

Exp Physiol 96.4 pp 426–438 Ion channel expression in developing sino-atrial node 427
potential (Toda, 1980). Furthermore, there is an ∼35%decrease in the hyperpolarization-activated current (I f )density (Accili et al. 1997), a reduction in the Na+ current(INa; Baruscotti et al. 1996), and a ∼31% decrease inL-type Ca2+ current (ICa,L) density (Protas et al. 2001). Inaddition, during ageing (to senescence), the conductiontime from the SAN to its surroundings is increasedand the conduction velocity decreased in rabbits andguinea-pigs (Protas et al. 2001; Jones et al. 2004). Thesechanges in electrical activity in the SAN during postnataldevelopment are likely to be the result of changes in geneexpression. In this study, we investigated changes in theexpression of ion channels (determinants of pacemakeractivity and action potential duration), Ca2+-handlingproteins (also determinants of pacemaker activity) andconnexins (determinants of conduction).
Methods
Neonatal (2–7 days of age) and adult male New Zealandwhite rabbits (∼6 months of age) were used. QuantitativePCR (qPCR) was used to measure the relative abundanceof 27 transcripts in eight neonatal and eight adultrabbits. For qPCR, samples from the SAN centre,right atrium and left ventricular free wall were used(see Fig. S1). For the SAN centre, tissue samples weretaken from the anatomical position of the leadingpacemaker site (∼0.5 mm from the main branch of thecrista terminalis as determined by extracellular potentialrecordings, as described in the Supplemental Methods andshown in Fig. S1). RNA was extracted using a modifiedQiagen protocol, and 100 ng of total RNA was reversetranscribed; qPCR was performed using SYBR greentechnology. Experimental data points (in triplicate) wereanalysed using the ��Ct method (double standardizationmethod) to obtain a quantitative measurement of mRNAabundance. All qPCR data were normalized to 28S(a housekeeping gene). In our study, there were nostatistically significant changes in the expression of 28SmRNA during development (data not shown). Whenpossible, in situ hybridization (ISH) was used to confirmthe accuracy of our qPCR results. Selected correspondingproteins were investigated using immunohistochemistry.For ISH and immunohistochemistry, tissue sections fromthree neonatal and three adult rabbits were used. Furtherdetails can be found in the Supplemental Methods andTables S1–S3.
Results
Postnatal changes in heart weight and cell size
Body and heart weights were significantly greater inadult rabbits compared with the neonates, although theheart weight-to-body weight ratio was significantly less
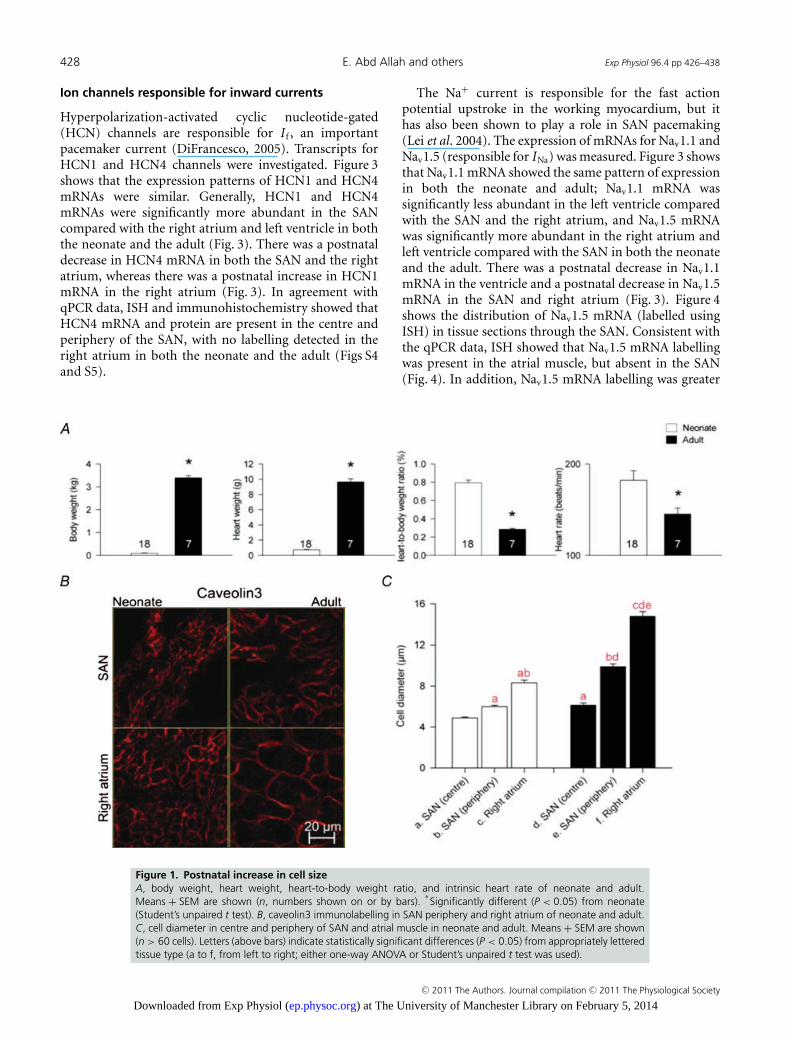
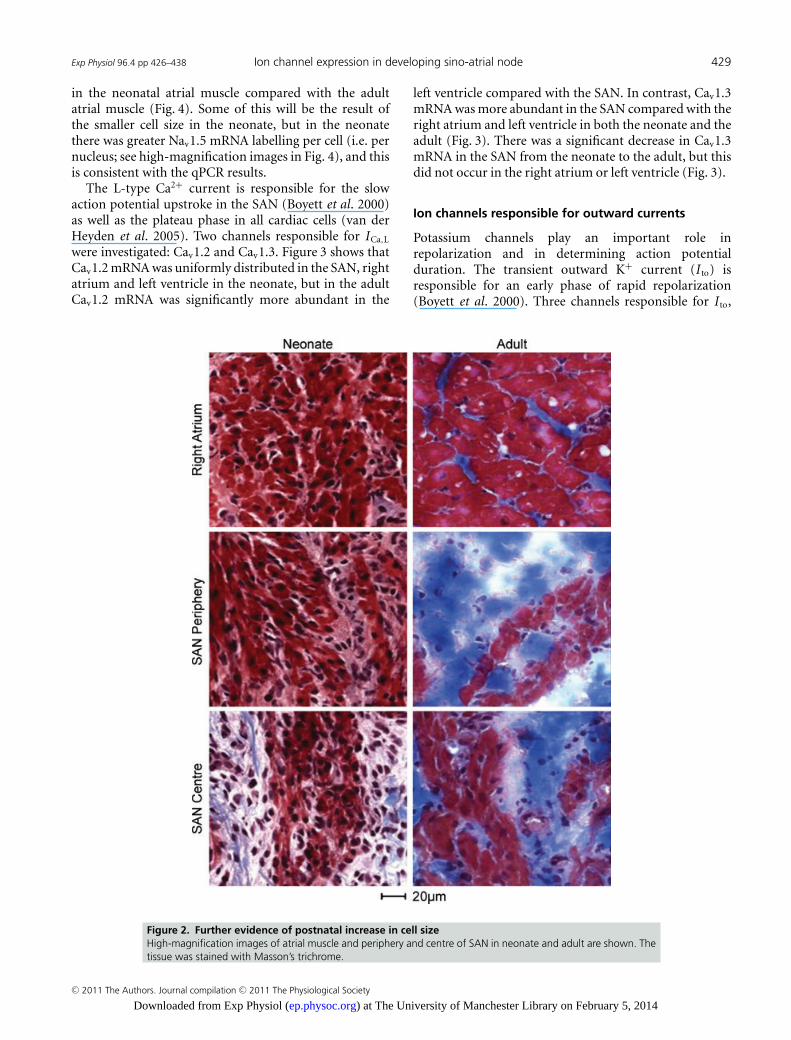
(Fig. 1A). To measure cell diameter, tissue sections wereimmunolabelled with either anti-caveolin3 or anti-NCX1antibodies. Figure 1B shows high-magnification imagesof caveolin3 labelling in the SAN as well as the atrialmuscle in the neonate and the adult. It shows that thenodal cells were modestly smaller than atrial cells in theneonate, but were substantially smaller in the adult. Thesehigh-magnification images were taken from the regionsindicated in Fig. S2. There was a prominent postnatalincrease in the cell diameter in all regions investigated.This is confirmed by Fig. 1C, which shows the mean celldiameter in the centre and periphery of the SAN and inatrial muscle. Figure 1C shows that cell diameter couldbe ranked as SANcentre < SAN periphery < atrial muscle,and from the neonate to the adult there was a small increasein cell diameter in the centre of the SAN (25%), a largerincrease in the periphery of the SAN (65%) and the largestincrease in the atrial muscle (78%). These results wereconfirmed by Masson’s trichrome staining (Fig. 2). One ofthe defining characteristics of the SAN is that the cells aresmaller than the surrounding atrial cells. Figure 1C showsthat this is primarily a characteristic of the adult.
Postnatal changes in ion channel expression
Figure 1A (right panel) shows that, as expected, thespontaneous beating rate of the isolated SAN (measureof intrinsic heart rate) was 21% slower in sevenpreparations from adult rabbits when compared witheighteen preparations from neonatal rabbits. To determinethe cause of the decrease in the intrinsic heart rate, tissuesamples (from SAN centre, atrial muscle and left ventricle;Fig. S1) were taken from eight neonates and eight adults,and qPCR was used to measure the abundance of differenttranscripts.
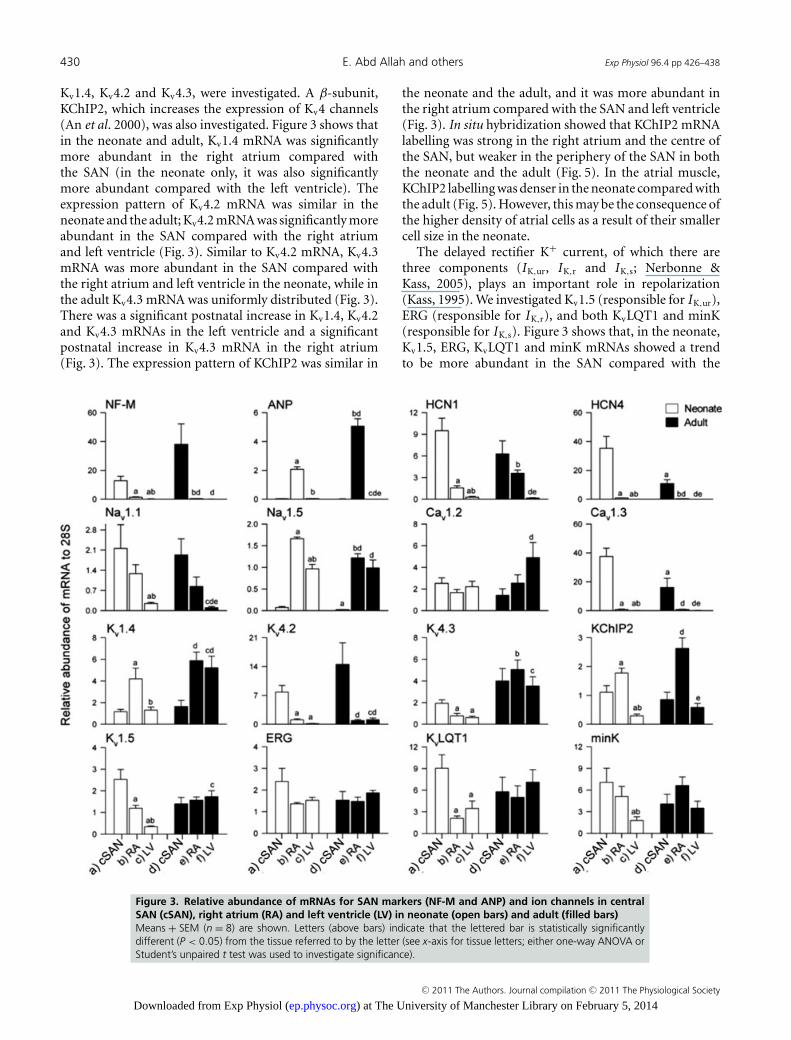
To test for atrial contamination of the SAN samples,neurofilament (NF-M) mRNA was used as a positivemarker of nodal tissue (ISH confirmed that NF-MmRNA was present in the SAN, but absent in the atrialmuscle in both neonate and adult; see Fig. S3). Sectionsfor investigation were cut perpendicular to the cristaterminalis. The centre of the SAN was located, as expected,in the thin intercaval region, whereas the periphery ofthe SAN (terminating in the right branch of the sino-atrial ring bundle, RSARB) was located next to and on thecrista terminalis (a thick bundle of atrial muscle). Atrialnatriuretic peptide (ANP) mRNA was used as a positivemarker of atrial muscle (Tellez et al. 2006). Figure 3 showsthat, in the neonate and adult, NF-M mRNA was abundantin the SAN and negligible in the right atrium and leftventricle, whereas ANP mRNA was abundant in the rightatrium and negligible in the SAN and left ventricle, thusconfirming that there was little or no cross-contaminationof the samples from neighbouring tissues.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
428 E. Abd Allah and others Exp Physiol 96.4 pp 426–438
Ion channels responsible for inward currents
Hyperpolarization-activated cyclic nucleotide-gated(HCN) channels are responsible for I f , an importantpacemaker current (DiFrancesco, 2005). Transcripts forHCN1 and HCN4 channels were investigated. Figure 3shows that the expression patterns of HCN1 and HCN4mRNAs were similar. Generally, HCN1 and HCN4mRNAs were significantly more abundant in the SANcompared with the right atrium and left ventricle in boththe neonate and the adult (Fig. 3). There was a postnataldecrease in HCN4 mRNA in both the SAN and the rightatrium, whereas there was a postnatal increase in HCN1mRNA in the right atrium (Fig. 3). In agreement withqPCR data, ISH and immunohistochemistry showed thatHCN4 mRNA and protein are present in the centre andperiphery of the SAN, with no labelling detected in theright atrium in both the neonate and the adult (Figs S4and S5).
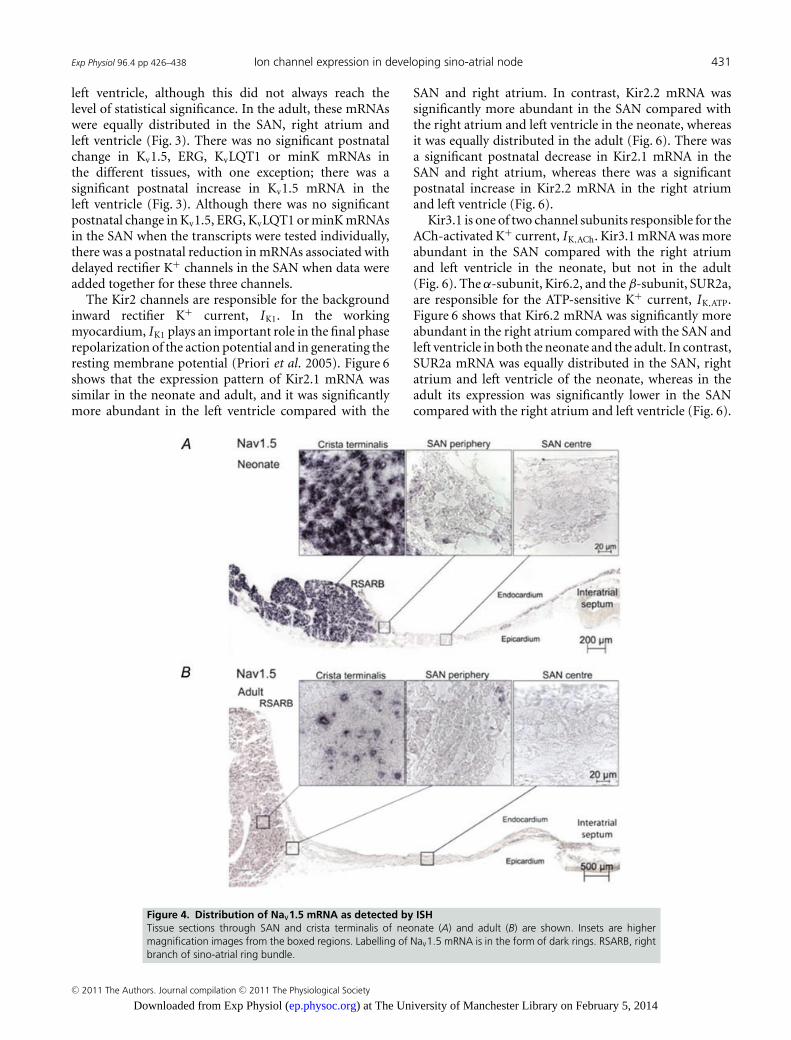
The Na+ current is responsible for the fast actionpotential upstroke in the working myocardium, but ithas also been shown to play a role in SAN pacemaking(Lei et al. 2004). The expression of mRNAs for Nav1.1 andNav1.5 (responsible for INa) was measured. Figure 3 showsthat Nav1.1 mRNA showed the same pattern of expressionin both the neonate and adult; Nav1.1 mRNA wassignificantly less abundant in the left ventricle comparedwith the SAN and the right atrium, and Nav1.5 mRNAwas significantly more abundant in the right atrium andleft ventricle compared with the SAN in both the neonateand the adult. There was a postnatal decrease in Nav1.1mRNA in the ventricle and a postnatal decrease in Nav1.5mRNA in the SAN and right atrium (Fig. 3). Figure 4shows the distribution of Nav1.5 mRNA (labelled usingISH) in tissue sections through the SAN. Consistent withthe qPCR data, ISH showed that Nav1.5 mRNA labellingwas present in the atrial muscle, but absent in the SAN(Fig. 4). In addition, Nav1.5 mRNA labelling was greater
Figure 1. Postnatal increase in cell sizeA, body weight, heart weight, heart-to-body weight ratio, and intrinsic heart rate of neonate and adult.Means + SEM are shown (n, numbers shown on or by bars).
∗Significantly different (P < 0.05) from neonate
(Student’s unpaired t test). B, caveolin3 immunolabelling in SAN periphery and right atrium of neonate and adult.C, cell diameter in centre and periphery of SAN and atrial muscle in neonate and adult. Means + SEM are shown(n > 60 cells). Letters (above bars) indicate statistically significant differences (P < 0.05) from appropriately letteredtissue type (a to f, from left to right; either one-way ANOVA or Student’s unpaired t test was used).
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
Exp Physiol 96.4 pp 426–438 Ion channel expression in developing sino-atrial node 429
in the neonatal atrial muscle compared with the adultatrial muscle (Fig. 4). Some of this will be the result ofthe smaller cell size in the neonate, but in the neonatethere was greater Nav1.5 mRNA labelling per cell (i.e. pernucleus; see high-magnification images in Fig. 4), and thisis consistent with the qPCR results.
The L-type Ca2+ current is responsible for the slowaction potential upstroke in the SAN (Boyett et al. 2000)as well as the plateau phase in all cardiac cells (van derHeyden et al. 2005). Two channels responsible for ICa,L
were investigated: Cav1.2 and Cav1.3. Figure 3 shows thatCav1.2 mRNA was uniformly distributed in the SAN, rightatrium and left ventricle in the neonate, but in the adultCav1.2 mRNA was significantly more abundant in the
left ventricle compared with the SAN. In contrast, Cav1.3mRNA was more abundant in the SAN compared with theright atrium and left ventricle in both the neonate and theadult (Fig. 3). There was a significant decrease in Cav1.3mRNA in the SAN from the neonate to the adult, but thisdid not occur in the right atrium or left ventricle (Fig. 3).
Ion channels responsible for outward currents
Potassium channels play an important role inrepolarization and in determining action potentialduration. The transient outward K+ current (I to) isresponsible for an early phase of rapid repolarization(Boyett et al. 2000). Three channels responsible for I to,
Figure 2. Further evidence of postnatal increase in cell sizeHigh-magnification images of atrial muscle and periphery and centre of SAN in neonate and adult are shown. Thetissue was stained with Masson’s trichrome.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
430 E. Abd Allah and others Exp Physiol 96.4 pp 426–438
Kv1.4, Kv4.2 and Kv4.3, were investigated. A β-subunit,KChIP2, which increases the expression of Kv4 channels(An et al. 2000), was also investigated. Figure 3 shows thatin the neonate and adult, Kv1.4 mRNA was significantlymore abundant in the right atrium compared withthe SAN (in the neonate only, it was also significantlymore abundant compared with the left ventricle). Theexpression pattern of Kv4.2 mRNA was similar in theneonate and the adult; Kv4.2 mRNA was significantly moreabundant in the SAN compared with the right atriumand left ventricle (Fig. 3). Similar to Kv4.2 mRNA, Kv4.3mRNA was more abundant in the SAN compared withthe right atrium and left ventricle in the neonate, while inthe adult Kv4.3 mRNA was uniformly distributed (Fig. 3).There was a significant postnatal increase in Kv1.4, Kv4.2and Kv4.3 mRNAs in the left ventricle and a significantpostnatal increase in Kv4.3 mRNA in the right atrium(Fig. 3). The expression pattern of KChIP2 was similar in
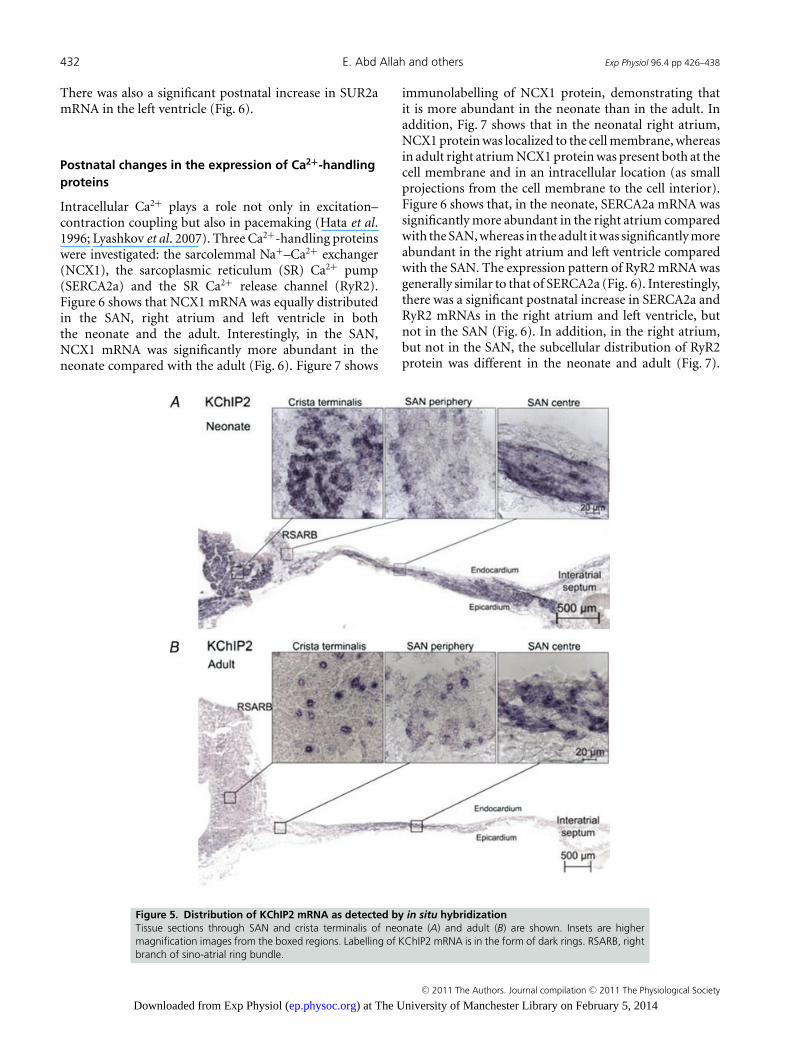
the neonate and the adult, and it was more abundant inthe right atrium compared with the SAN and left ventricle(Fig. 3). In situ hybridization showed that KChIP2 mRNAlabelling was strong in the right atrium and the centre ofthe SAN, but weaker in the periphery of the SAN in boththe neonate and the adult (Fig. 5). In the atrial muscle,KChIP2 labelling was denser in the neonate compared withthe adult (Fig. 5). However, this may be the consequence ofthe higher density of atrial cells as a result of their smallercell size in the neonate.
The delayed rectifier K+ current, of which there arethree components (IK,ur, IK,r and IK,s; Nerbonne &Kass, 2005), plays an important role in repolarization(Kass, 1995). We investigated Kv1.5 (responsible for IK,ur),ERG (responsible for IK,r), and both KvLQT1 and minK(responsible for IK,s). Figure 3 shows that, in the neonate,Kv1.5, ERG, KvLQT1 and minK mRNAs showed a trendto be more abundant in the SAN compared with the
Figure 3. Relative abundance of mRNAs for SAN markers (NF-M and ANP) and ion channels in centralSAN (cSAN), right atrium (RA) and left ventricle (LV) in neonate (open bars) and adult (filled bars)Means + SEM (n = 8) are shown. Letters (above bars) indicate that the lettered bar is statistically significantlydifferent (P < 0.05) from the tissue referred to by the letter (see x-axis for tissue letters; either one-way ANOVA orStudent’s unpaired t test was used to investigate significance).
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
Exp Physiol 96.4 pp 426–438 Ion channel expression in developing sino-atrial node 431
left ventricle, although this did not always reach thelevel of statistical significance. In the adult, these mRNAswere equally distributed in the SAN, right atrium andleft ventricle (Fig. 3). There was no significant postnatalchange in Kv1.5, ERG, KvLQT1 or minK mRNAs inthe different tissues, with one exception; there was asignificant postnatal increase in Kv1.5 mRNA in theleft ventricle (Fig. 3). Although there was no significantpostnatal change in Kv1.5, ERG, KvLQT1 or minK mRNAsin the SAN when the transcripts were tested individually,there was a postnatal reduction in mRNAs associated withdelayed rectifier K+ channels in the SAN when data wereadded together for these three channels.
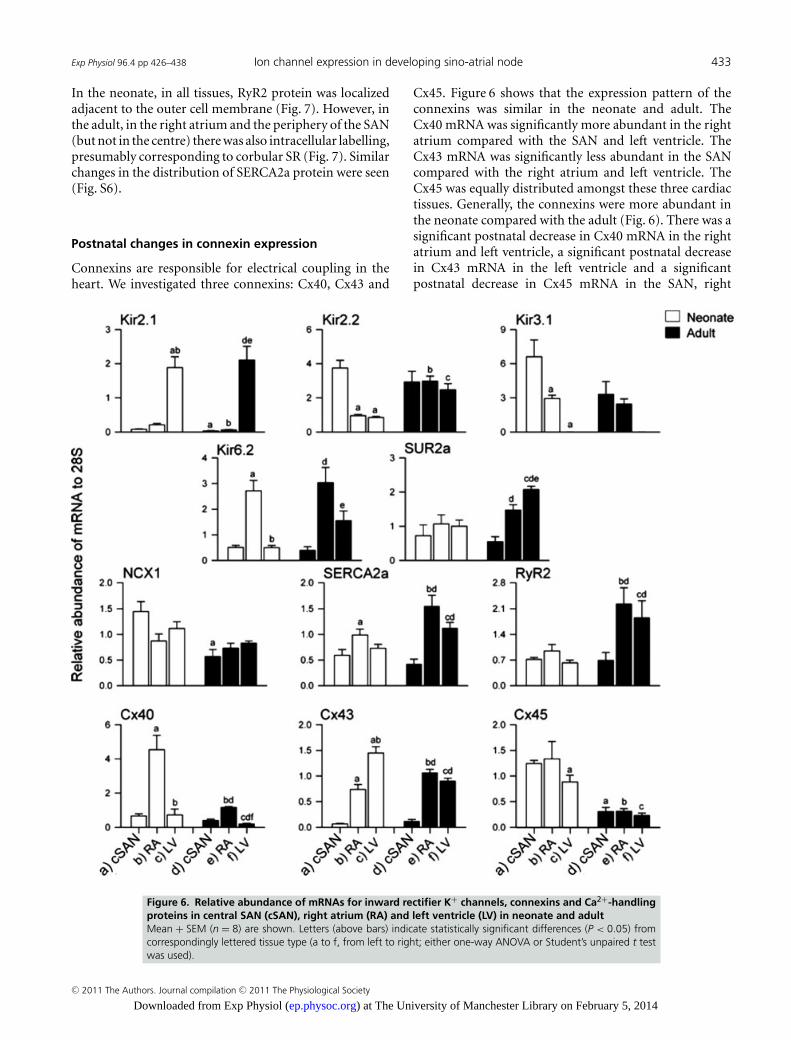
The Kir2 channels are responsible for the backgroundinward rectifier K+ current, IK1. In the workingmyocardium, IK1 plays an important role in the final phaserepolarization of the action potential and in generating theresting membrane potential (Priori et al. 2005). Figure 6shows that the expression pattern of Kir2.1 mRNA wassimilar in the neonate and adult, and it was significantlymore abundant in the left ventricle compared with the
SAN and right atrium. In contrast, Kir2.2 mRNA wassignificantly more abundant in the SAN compared withthe right atrium and left ventricle in the neonate, whereasit was equally distributed in the adult (Fig. 6). There wasa significant postnatal decrease in Kir2.1 mRNA in theSAN and right atrium, whereas there was a significantpostnatal increase in Kir2.2 mRNA in the right atriumand left ventricle (Fig. 6).
Kir3.1 is one of two channel subunits responsible for theACh-activated K+ current, IK,ACh. Kir3.1 mRNA was moreabundant in the SAN compared with the right atriumand left ventricle in the neonate, but not in the adult(Fig. 6). The α-subunit, Kir6.2, and the β-subunit, SUR2a,are responsible for the ATP-sensitive K+ current, IK,ATP.Figure 6 shows that Kir6.2 mRNA was significantly moreabundant in the right atrium compared with the SAN andleft ventricle in both the neonate and the adult. In contrast,SUR2a mRNA was equally distributed in the SAN, rightatrium and left ventricle of the neonate, whereas in theadult its expression was significantly lower in the SANcompared with the right atrium and left ventricle (Fig. 6).
Figure 4. Distribution of Nav1.5 mRNA as detected by ISHTissue sections through SAN and crista terminalis of neonate (A) and adult (B) are shown. Insets are highermagnification images from the boxed regions. Labelling of Nav1.5 mRNA is in the form of dark rings. RSARB, rightbranch of sino-atrial ring bundle.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
432 E. Abd Allah and others Exp Physiol 96.4 pp 426–438
There was also a significant postnatal increase in SUR2amRNA in the left ventricle (Fig. 6).
Postnatal changes in the expression of Ca2+-handlingproteins
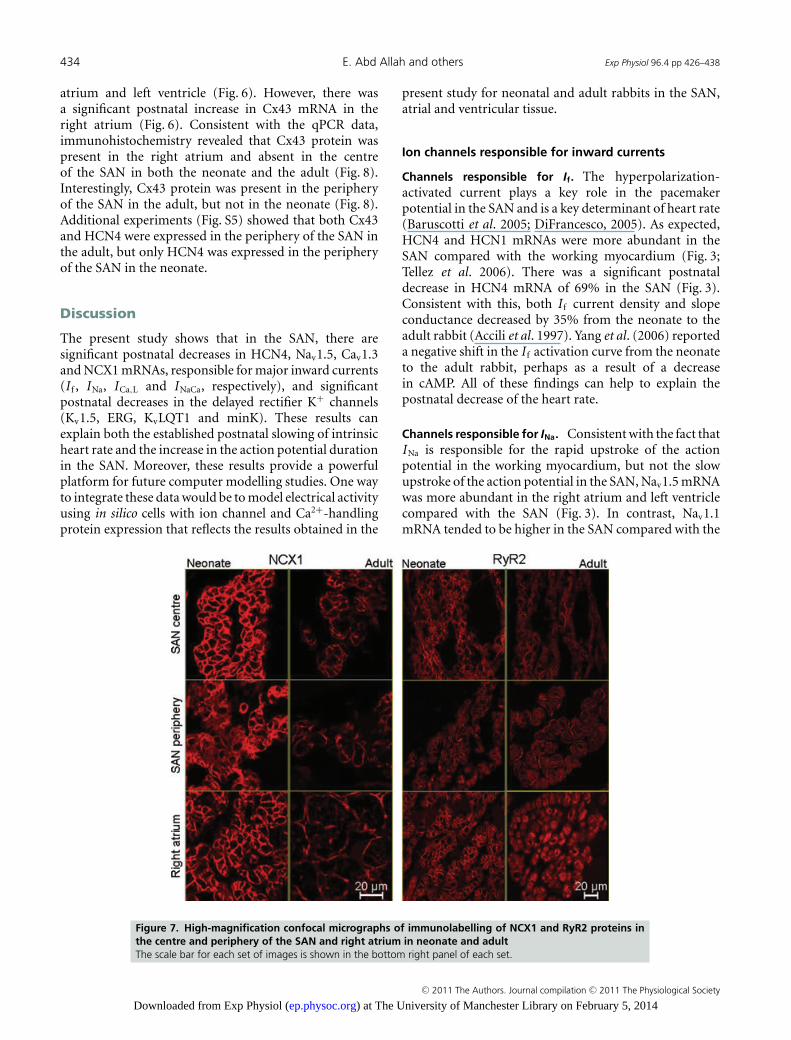
Intracellular Ca2+ plays a role not only in excitation–contraction coupling but also in pacemaking (Hata et al.1996; Lyashkov et al. 2007). Three Ca2+-handling proteinswere investigated: the sarcolemmal Na+–Ca2+ exchanger(NCX1), the sarcoplasmic reticulum (SR) Ca2+ pump(SERCA2a) and the SR Ca2+ release channel (RyR2).Figure 6 shows that NCX1 mRNA was equally distributedin the SAN, right atrium and left ventricle in boththe neonate and the adult. Interestingly, in the SAN,NCX1 mRNA was significantly more abundant in theneonate compared with the adult (Fig. 6). Figure 7 shows
immunolabelling of NCX1 protein, demonstrating thatit is more abundant in the neonate than in the adult. Inaddition, Fig. 7 shows that in the neonatal right atrium,NCX1 protein was localized to the cell membrane, whereasin adult right atrium NCX1 protein was present both at thecell membrane and in an intracellular location (as smallprojections from the cell membrane to the cell interior).Figure 6 shows that, in the neonate, SERCA2a mRNA wassignificantly more abundant in the right atrium comparedwith the SAN, whereas in the adult it was significantly moreabundant in the right atrium and left ventricle comparedwith the SAN. The expression pattern of RyR2 mRNA wasgenerally similar to that of SERCA2a (Fig. 6). Interestingly,there was a significant postnatal increase in SERCA2a andRyR2 mRNAs in the right atrium and left ventricle, butnot in the SAN (Fig. 6). In addition, in the right atrium,but not in the SAN, the subcellular distribution of RyR2protein was different in the neonate and adult (Fig. 7).
Figure 5. Distribution of KChIP2 mRNA as detected by in situ hybridizationTissue sections through SAN and crista terminalis of neonate (A) and adult (B) are shown. Insets are highermagnification images from the boxed regions. Labelling of KChIP2 mRNA is in the form of dark rings. RSARB, rightbranch of sino-atrial ring bundle.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
Exp Physiol 96.4 pp 426–438 Ion channel expression in developing sino-atrial node 433
In the neonate, in all tissues, RyR2 protein was localizedadjacent to the outer cell membrane (Fig. 7). However, inthe adult, in the right atrium and the periphery of the SAN(but not in the centre) there was also intracellular labelling,presumably corresponding to corbular SR (Fig. 7). Similarchanges in the distribution of SERCA2a protein were seen(Fig. S6).
Postnatal changes in connexin expression
Connexins are responsible for electrical coupling in theheart. We investigated three connexins: Cx40, Cx43 and
Cx45. Figure 6 shows that the expression pattern of theconnexins was similar in the neonate and adult. TheCx40 mRNA was significantly more abundant in the rightatrium compared with the SAN and left ventricle. TheCx43 mRNA was significantly less abundant in the SANcompared with the right atrium and left ventricle. TheCx45 was equally distributed amongst these three cardiactissues. Generally, the connexins were more abundant inthe neonate compared with the adult (Fig. 6). There was asignificant postnatal decrease in Cx40 mRNA in the rightatrium and left ventricle, a significant postnatal decreasein Cx43 mRNA in the left ventricle and a significantpostnatal decrease in Cx45 mRNA in the SAN, right
Figure 6. Relative abundance of mRNAs for inward rectifier K+ channels, connexins and Ca2+-handlingproteins in central SAN (cSAN), right atrium (RA) and left ventricle (LV) in neonate and adultMean + SEM (n = 8) are shown. Letters (above bars) indicate statistically significant differences (P < 0.05) fromcorrespondingly lettered tissue type (a to f, from left to right; either one-way ANOVA or Student’s unpaired t testwas used).
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
434 E. Abd Allah and others Exp Physiol 96.4 pp 426–438
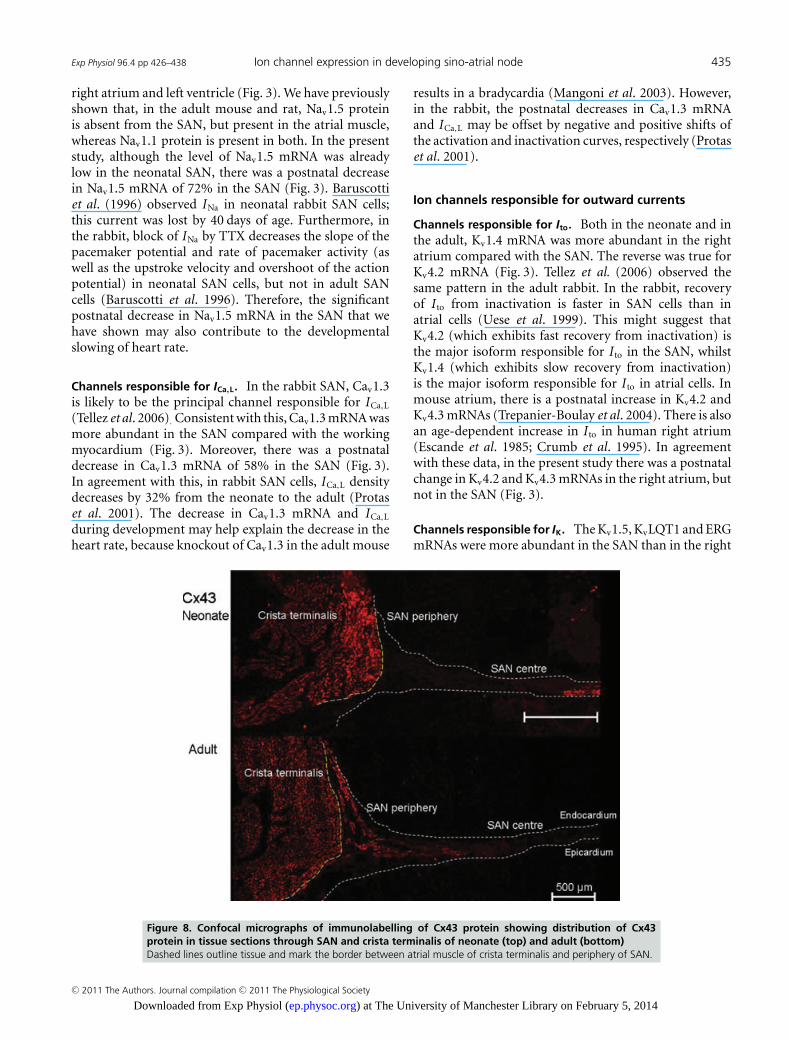
atrium and left ventricle (Fig. 6). However, there wasa significant postnatal increase in Cx43 mRNA in theright atrium (Fig. 6). Consistent with the qPCR data,immunohistochemistry revealed that Cx43 protein waspresent in the right atrium and absent in the centreof the SAN in both the neonate and the adult (Fig. 8).Interestingly, Cx43 protein was present in the peripheryof the SAN in the adult, but not in the neonate (Fig. 8).Additional experiments (Fig. S5) showed that both Cx43and HCN4 were expressed in the periphery of the SAN inthe adult, but only HCN4 was expressed in the peripheryof the SAN in the neonate.
Discussion
The present study shows that in the SAN, there aresignificant postnatal decreases in HCN4, Nav1.5, Cav1.3and NCX1 mRNAs, responsible for major inward currents(I f , INa, ICa,L and INaCa, respectively), and significantpostnatal decreases in the delayed rectifier K+ channels(Kv1.5, ERG, KvLQT1 and minK). These results canexplain both the established postnatal slowing of intrinsicheart rate and the increase in the action potential durationin the SAN. Moreover, these results provide a powerfulplatform for future computer modelling studies. One wayto integrate these data would be to model electrical activityusing in silico cells with ion channel and Ca2+-handlingprotein expression that reflects the results obtained in the
present study for neonatal and adult rabbits in the SAN,atrial and ventricular tissue.
Ion channels responsible for inward currents
Channels responsible for If. The hyperpolarization-activated current plays a key role in the pacemakerpotential in the SAN and is a key determinant of heart rate(Baruscotti et al. 2005; DiFrancesco, 2005). As expected,HCN4 and HCN1 mRNAs were more abundant in theSAN compared with the working myocardium (Fig. 3;Tellez et al. 2006). There was a significant postnataldecrease in HCN4 mRNA of 69% in the SAN (Fig. 3).Consistent with this, both I f current density and slopeconductance decreased by 35% from the neonate to theadult rabbit (Accili et al. 1997). Yang et al. (2006) reporteda negative shift in the I f activation curve from the neonateto the adult rabbit, perhaps as a result of a decreasein cAMP. All of these findings can help to explain thepostnatal decrease of the heart rate.
Channels responsible for INa. Consistent with the fact thatINa is responsible for the rapid upstroke of the actionpotential in the working myocardium, but not the slowupstroke of the action potential in the SAN, Nav1.5 mRNAwas more abundant in the right atrium and left ventriclecompared with the SAN (Fig. 3). In contrast, Nav1.1mRNA tended to be higher in the SAN compared with the
Figure 7. High-magnification confocal micrographs of immunolabelling of NCX1 and RyR2 proteins inthe centre and periphery of the SAN and right atrium in neonate and adultThe scale bar for each set of images is shown in the bottom right panel of each set.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (
Exp Physiol 96.4 pp 426–438 Ion channel expression in developing sino-atrial node 435
right atrium and left ventricle (Fig. 3). We have previouslyshown that, in the adult mouse and rat, Nav1.5 proteinis absent from the SAN, but present in the atrial muscle,whereas Nav1.1 protein is present in both. In the presentstudy, although the level of Nav1.5 mRNA was alreadylow in the neonatal SAN, there was a postnatal decreasein Nav1.5 mRNA of 72% in the SAN (Fig. 3). Baruscottiet al. (1996) observed INa in neonatal rabbit SAN cells;this current was lost by 40 days of age. Furthermore, inthe rabbit, block of INa by TTX decreases the slope of thepacemaker potential and rate of pacemaker activity (aswell as the upstroke velocity and overshoot of the actionpotential) in neonatal SAN cells, but not in adult SANcells (Baruscotti et al. 1996). Therefore, the significantpostnatal decrease in Nav1.5 mRNA in the SAN that wehave shown may also contribute to the developmentalslowing of heart rate.
Channels responsible for ICa,L. In the rabbit SAN, Cav1.3is likely to be the principal channel responsible for ICa,L
(Tellez et al. 2006). Consistent with this, Cav1.3 mRNA wasmore abundant in the SAN compared with the workingmyocardium (Fig. 3). Moreover, there was a postnataldecrease in Cav1.3 mRNA of 58% in the SAN (Fig. 3).In agreement with this, in rabbit SAN cells, ICa,L densitydecreases by 32% from the neonate to the adult (Protaset al. 2001). The decrease in Cav1.3 mRNA and ICa,L
during development may help explain the decrease in theheart rate, because knockout of Cav1.3 in the adult mouse
results in a bradycardia (Mangoni et al. 2003). However,in the rabbit, the postnatal decreases in Cav1.3 mRNAand ICa,L may be offset by negative and positive shifts ofthe activation and inactivation curves, respectively (Protaset al. 2001).
Ion channels responsible for outward currents
Channels responsible for Ito. Both in the neonate and inthe adult, Kv1.4 mRNA was more abundant in the rightatrium compared with the SAN. The reverse was true forKv4.2 mRNA (Fig. 3). Tellez et al. (2006) observed thesame pattern in the adult rabbit. In the rabbit, recoveryof I to from inactivation is faster in SAN cells than inatrial cells (Uese et al. 1999). This might suggest thatKv4.2 (which exhibits fast recovery from inactivation) isthe major isoform responsible for I to in the SAN, whilstKv1.4 (which exhibits slow recovery from inactivation)is the major isoform responsible for I to in atrial cells. Inmouse atrium, there is a postnatal increase in Kv4.2 andKv4.3 mRNAs (Trepanier-Boulay et al. 2004). There is alsoan age-dependent increase in I to in human right atrium(Escande et al. 1985; Crumb et al. 1995). In agreementwith these data, in the present study there was a postnatalchange in Kv4.2 and Kv4.3 mRNAs in the right atrium, butnot in the SAN (Fig. 3).
Channels responsible for IK. The Kv1.5, KvLQT1 and ERGmRNAs were more abundant in the SAN than in the right
Figure 8. Confocal micrographs of immunolabelling of Cx43 protein showing distribution of Cx43protein in tissue sections through SAN and crista terminalis of neonate (top) and adult (bottom)Dashed lines outline tissue and mark the border between atrial muscle of crista terminalis and periphery of SAN.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (

436 E. Abd Allah and others Exp Physiol 96.4 pp 426–438
atrium and left ventricle in the neonate and were uniformlydistributed in the adult (Fig. 3). We have previously shownthat, in the adult guinea-pig, Kv1.5 protein is equallydistributed in the SAN, right atrium and left ventricle(Dobrzynski et al. 2000). In the SAN, there was a postnataldecrease of the delayed rectifier K+ channels (Fig. 3),and this can explain the prolongation of the SAN actionpotential, as shown by Toda (1980).
Channels responsible for IK1. The background inwardrectifier K+ current, IK1, generates and stabilizes the restingmembrane potential in the atrium and ventricle, and IK1 islogically absent from the SAN (Dhamoon & Jalife, 2005).Consistent with this, Kir2.1 mRNA was less abundant inthe SAN than in the left ventricle in both the neonateand the adult (Fig. 6). However, curiously, in the neonateat least, Kir2.2 mRNA was more abundant in the SANcompared with the left ventricle (Fig. 6).
Ca2+-handling proteins
In the SAN, NCX1 was more abundant in the neonatecompared with the adult at both mRNA and proteinlevels (Figs 6 and 7). A similar postnatal decrease inNCX1 has been previously observed in rabbit ventricle(Artman et al. 1995; Chen et al. 1995; Dan et al. 2007),rabbit whole heart (Artman, 1992) and human ventricle(Qu et al. 2000). In addition, the distribution of NCX1protein was different in the neonate and the adult (Fig. 7).In the SAN, the postnatal decrease in NCX1 may befunctionally important, according to the ‘Ca2+ clock’hypothesis (according to this hypothesis, pacemaking isin part the result of spontaneous, rhythmic release ofCa2+ from the SR during late diastole and consequentactivation of inward Na+–Ca2+ exchange current, INaCa).The decrease in NCX1 can, therefore, help to explain thepostnatal decrease in the heart rate. We also observeda significant postnatal increase in SERCA2a and RyR2mRNA in the right atrium and left ventricle, but not in theSAN (Fig. 6). In agreement, it has previously been shownthat from embryo to adult, SERCA2a mRNA increases inrabbit ventricle (Arai et al. 1992) and human ventricle(Qu & Boutjdir, 2001). Likewise, in rabbit ventricularmyocytes, RyR2 mRNA has been previously reported toincrease from the embryo to the adult (Joyner & vanCapelle, 1986).
Connexins
The Cx40 and Cx43 mRNAs, but not Cx45 mRNA,were less abundant in the SAN compared with the rightatrium in both the neonate and the adult (Fig. 6). Tellezet al. (2006) observed the same pattern in the adult. Thedistribution of Cx43 protein is the same as its mRNA(Figs 6 and 8). The paucity of Cx43 and Cx40 in the
SAN is in agreement with theoretical considerations andmathematical modelling, which suggests that there is poorelectrical coupling in the SAN to protect the SAN from thehyperpolarizing effects of the surrounding atrial muscle(Joyner & van Capelle, 1986). In the adult rabbit, Cx43is present in the periphery but not the centre of the SAN(Dobrzynski et al. 2007). This was confirmed in the presentstudy (Fig. 8). It has been argued that the presence of Cx43in the periphery of the SAN is important for the SAN tobe able to drive the atrial muscle (Joyner & van Capelle,1986). It is, therefore, curious that Cx43 is absent from theperiphery of the SAN in the neonate (Fig. 8).
Conclusion
In conclusion, this is the first study to describe postnatalchanges in the SAN in the expression of mRNAs for a largenumber of ion channels as well as Ca2+-handling proteinsand connexins. The data show that there are significantpostnatal changes in ion channel expression in the SANthat can explain the physiologically observed changes inheart rate (HCN4, Nav1.5, Cav1.3 and NCX1) and actionpotential duration (Kv1.5, KvLQT1, minK and ERG).
References
Accili EA, Robinson RB & DiFrancesco D (1997). Propertiesand modulation of I f in newborn versus adult cardiac SAnode. Am J Physiol Heart Circ Physiol 272, H1549–H1552.
An WF, Bowlby MR, Betty M, Cao J, Ling HP, Mendoza G,Hinson JW, Mattsson KI, Strassle BW, Trimmer JS & RhodesKJ (2000). Modulation of A-type potassium channels by afamily of calcium sensors. Nature 403, 553–556.
Arai M, Otsu K, MacLennan DH & Periasamy M (1992).Regulation of sarcoplasmic reticulum gene expression duringcardiac and skeletal muscle development. Am J Physiol CellPhysiol 262, C614–C620.
Artman M (1992). Sarcolemmal Na+-Ca2+ exchange activityand exchanger immunoreactivity in developing rabbithearts. Am J Physiol Heart Circ Physiol 263, H1506–H1513.
Artman M, Ichikawa H, Avkiran M & Coetzee WA (1995).Na+/Ca2+ exchange current density in cardiac myocytesfrom rabbits and guinea pigs during postnatal development.Am J Physiol Heart Circ Physiol 268, H1714–H1722.
Baruscotti M, Bucchi A & DiFrancesco D (2005). Physiologyand pharmacology of the cardiac pacemaker (“funny”)current. Pharmacol Ther 107, 59–79.
Baruscotti M, DiFrancesco D & Robinson RB (1996). ATTX-sensitive inward sodium current contributes tospontaneous activity in newborn rabbit sino-atrial nodecells. J Physiol 492, 21–30.
Boyett MR, Honjo H & Kodama I (2000). The sinoatrial node,a heterogeneous pacemaker structure. Cardiovasc Res 47,658–687.
Boyett MR, Inada S, Yoo S, Li J, Liu J, Tellez J, Greener ID,Honjo H, Billeter R, Lei M, Zhang H, Efimov IR &Dobrzynski H (2006). Connexins in the sinoatrial andatrioventricular nodes. Adv Cardiol 42, 175–197.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (

Exp Physiol 96.4 pp 426–438 Ion channel expression in developing sino-atrial node 437
Chen F, Mottino G, Klitzner TS, Philipson KD & Frank JS(1995). Distribution of the Na+/Ca2+ exchange protein indeveloping rabbit myocytes. Am J Physiol Cell Physiol 268,C1126–C1132.
Crumb WJ, Pigott JD & Clarkson CW (1995). Comparison ofI to in young and adult human atrial myocytes: evidence fordevelopmental changes. Am J Physiol Heart Circ Physiol 268,H1335–H1342.
Dan P, Lin E, Huang J, Biln P & Tibbits GF (2007).Three-dimensional distribution of cardiac Na+-Ca2+exchanger and ryanodine receptor during development.Biophys J 93, 2504–2518.
Dhamoon AS & Jalife J (2005). The inward rectifier current(IK1) controls cardiac excitability and is involved inarrhythmogenesis. Heart Rhythm 2, 316–324.
DiFrancesco D (2005). Cardiac pacemaker I f current and itsinhibition by heart rate-reducing agents. Curr Med Res Opin21, 1115–1122.
Dobrzynski H, Boyett MR & Anderson RH (2007). Newinsights into pacemaker activity: promoting understandingof sick sinus syndrome. Circulation 115, 1921–1932.
Dobrzynski H, Rothery SM, Marples DD, Coppen SR, TakagishiY, Honjo H, Tamkun MM, Henderson Z, Kodama I, SeversNJ & Boyett MR (2000). Presence of the Kv1.5 K+ channel inthe sinoatrial node. J Histochem Cytochem 48, 769–780.
Escande D, Loisance D, Planche C & Coraboeuf E (1985).Age-related changes of action potential plateau shape inisolated human atrial fibers. Am J Physiol Heart Circ Physiol249, H843–H850.
Hata T, Noda T, Nishimura M & Watanabe Y (1996). The roleof Ca2+ release from sarcoplasmic reticulum in theregulation of sinoatrial node automaticity. Heart Vessels 11,234–241.
Huang X, Yang P, Du Y, Zhang J & Ma A (2007). Age-relateddown-regulation of HCN channels in rat sinoatrial node.Basic Res Cardiol 102, 429–435.
Jones SA, Lancaster MK & Boyett MR (2004). Ageing-relatedchanges of connexins and conduction within the sinoatrialnode. J Physiol 560, 429–437.
Joyner RW & van Capelle FJ (1986). Propagation throughelectrically coupled cells. How a small SA node drives a largeatrium. Biophys J 50, 1157–1164.
Kass RS (1995). Delayed potassium channels: cellular,molecular, and regulatory properties. In CardiacElectrophysiology: from Cell to Bedside, ed. Zipes D & Jalifa J,pp. 54–82. Saunders, Philadelphia, PA.
Lei M, Jones SA, Liu J, Lancaster MK, Fung SS, Dobrzynski H,Camelliti P, Maier SK, Noble D & Boyett MR (2004).Requirement of neuronal- and cardiac-type sodium channelsfor murine sinoatrial node pacemaking. J Physiol 559,835–848.
Lyashkov AE, Juhaszova M, Dobrzynski H, Vinogradova TM,Maltsev VA, Juhasz O, Spurgeon HA, Sollott SJ & Lakatta EG(2007). Calcium cycling protein density and functionalimportance to automaticity of isolated sinoatrial nodal cellsare independent of cell size. Circ Res 100, 1723–1731.
Mangoni ME, Couette B, Bourinet E, Platzer J, Reimer D,Striessnig J & Nargeot J (2003). Functional role of L-typeCav1.3 Ca2+ channels in cardiac pacemaker activity. ProcNatl Acad Sci USA 100, 5543–5548.
Nerbonne JM & Kass RS (2005). Molecular physiology ofcardiac repolarization. Physiol Rev 85, 1205–1253.
Priori SG, Pandit SV, Rivolta I, Berenfeld O, Ronchetti E,Dhamoon A, Napolitano C, Anumonwo J, di Barletta MR,Gudapakkam S, Bosi G, Stramba-Badiale M & Jalife J (2005).A novel form of short QT syndrome (SQT3) is caused by amutation in the KCNJ2 gene. Circ Res 96, 800–807.
Protas L, DiFrancesco D & Robinson RB (2001). L-type but notT-type calcium current changes during postnataldevelopment in rabbit sinoatrial node. Am J Physiol HeartCirc Physiol 281, H1252–H1259.
Qu Y & Boutjdir M (2001). Gene expression of SERCA2aand L- and T-type Ca channels during human heartdevelopment. Pediatr Res 50, 569–574.
Qu Y, Ghatpande A, el-Sherif N & Boutjdir M (2000). Geneexpression of Na+/Ca2+ exchanger during development inhuman heart. Cardiovasc Res 45, 866–873.
Satoh H (2003). Sino-atrial nodal cells of mammalian hearts:ionic currents and gene expression of pacemaker ionicchannels. J Smooth Muscle Res 39, 175–193.
Tellez JO, Dobrzynski H, Greener ID, Graham GM, Laing E,Honjo H, Hubbard SJ, Boyett MR, Billeter R (2006).Differential expression of ion channel transcripts in atrialmuscle and sinoatrial node in rabbit. Circ Res 99, 1384–1393.
Toda N (1980). Age-related changes in the transmembranepotential of isolated rabbit sino-atrial nodes and atria.Cardiovasc Res 14, 58–63.
Trepanier-Boulay V, Lupien MA, St-Michel C & Fiset C (2004).Postnatal development of atrial repolarization in the mouse.Cardiovasc Res 64, 84–93.
Uese K, Hagiwara N, Miyawaki T & Kasanuki H (1999).Properties of the transient outward current in rabbitsino-atrial node cells. J Mol Cell Cardiol 31, 1975–1984.
Van Der Heyden MA, Wijnhoven TJ & Opthof T (2005).Molecular aspects of adrenergic modulation of cardiacL-type Ca2+ channels. Cardiovasc Res 65, 28–39.
Yang ZF, Sun Y, Li CZ, Wang HW, Wang XJ, Zheng YQ, Liu &Liu YM (2006). Reduced sinoatrial cAMP content plays arole in postnatal heart rate slowing in the rabbit. Clin ExpPharmacol Physiol 33, 757–762.
Acknowledgements
This study was funded by the Ministry of Higher Educationof Egypt and the British Heart Foundation (grant no.RG/06005).
Supporting Information
Online supplementary material for this paper can beaccessed at Wiley Online Library:
Figure S1. A and B, typical neonatal (A) and adult (B)SAN preparations.Figure S2. Histological features of neonatal and adultrabbit SAN.Figure S3. NF-M mRNA as detected by ISH.Figure S4. HCN4 mRNA as detected by ISH.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (

438 E. Abd Allah and others Exp Physiol 96.4 pp 426–438
Figure S5. Double-labelling of Cx43 (a negative marker ofthe SAN; green) and HCN4 (a positive marker of the SAN;red) in tissue section through neonate and adult SAN andits surrounding atrial muscle.Figure S6. Immunolabelling of SERCA2a in the neonataland adult SAN centre and periphery and neonatal andadult atrium.Table S1. Primers used with ABI Prism 7900HT.Table S2. Primers used to generate DNA template forriboprobe synthesis.
Table S3. Summary of antibodies used inimmunohistochemistry.
As a service to our authors and readers, this journalprovides supporting information supplied by the authors.Such materials are peer-reviewed and may be re-organized for online delivery, but are not copy-edited ortypeset. Technical support issues arising from supportinginformation (other than missing files) should be addressedto the authors.
C© 2011 The Authors. Journal compilation C© 2011 The Physiological Society
) at The University of Manchester Library on February 5, 2014ep.physoc.orgDownloaded from Exp Physiol (




















