
NPHS2 p.V290M mutation in late-onset steroid-resistant nephrotic syndrome

Aquaculture, 61 (1987) 59-74 Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands
59
Changes in Steroid Concentrations During Sexual Ontogenesis in Tilapia
S. R0THBARD2, B. MOAV’ and Z. YARON’
‘Department of Zoology, The George S. Wise Faculty of Life Sciences, Tel Aviv University, Tel Aviv 69978 (Israel) 2Fish Breeding Centre, Gun Shmuel38810 (Israel)
(Accepted 27 June 1986)
ABSTRACT
Rothbard, S., Moav, B. and Yaron, Z., 1987. Changes in steroid concentrations during sexual ontogenesis in tilapia. Aquaculture, 61: 59-74.
Testosterone, estiadiol and 1 1-ketotestosterone were determined by radioimmunoassay in whole body extracts of tilapia at various stages of ontogenesis: before, during and after sexual differen- tiation. The study was conducted on Tilapia nilotica ( = Greochrom~ niloticus), T. mossam~ica ( = 0. rnossarn~~~} and T. hornorum in a series of ~tr~peci~c and interspecific spawn com- binations. The con~ntrations of testosterone and estradiof measured in homogenates prepared from groups of individuals just after fertilization were relatively high (c. 1 ng/~divid~) and declined to undetectable levels in 3-4-week-old fry. In older fry (4-6 weeks), the concentrations of both steroids started to increase; however, the testosterone increase was tenfold that of estra- diol. In 6-8-week-old fry measured individually, the testosterone concentrations in intraspecific spawns showed a clear tendency toward two peaks of distribution. This bimodality in testosterone concentrations probably reflects the presence of males and females with different testosterone contents in the population. In interspecific crosses characterized by 100% male progeny, the dis- tribution pattern of testosterone was unimodal, which would be expected in a monosexual popu- lation. E&radio1 and Il-ketotestosterone concentrations in the young differentiated fry at this age were low in all cases. It is suggested, therefore, that, of the steroids examined in tilapia at the age of 6-8 weeks, only testosterone reflects the processes of sex diffe~ntiation.
INTRODUCTION
The puzzle of sex determination and sex differentiation in tilapia has intrigued both fish culturists and scientists since Hickling crossed two species of tilapia and unintentionally obtained monosex populations of males (Hick- ling, 1960). Based on chromosomal sex determination mechanisms, several models have been proposed in an attempt to explain the “abnormal” sex ratios obtained by crossing various tilapia species (Hickling, 1960; Chen, 1969; Jal- abert et al., 1971; Avtalion and Hammerman, 1978; Hammerman and Avtalion,
0044-8486/87/$03.50 0 1987 Elsevier Science Publishers B.V.

60
1979; Lovshin, 1982). Although results of some crosses fit the suggested chro- mosomal model, others imply that sex determination in tilapias involves sys- tems of a more complicated nature ( Wohlfarth and Hulata, 1983; Majumdar and McAndrew, 1983). Deviations from the expected 1:l sex ratio were also reported in a series of single pair intraspecific spawnings of Tilupia nilotica and of T. aurea (Shelton et al., 1983).
The fact that the phenotypic sex in fish can be manipulated by external intervention indicates that, whatever the mechanism of chromosomal sex determination may be, the subsequent process of sex ~fferentiation involves ad~tional mechanisms. These encompass certain sex steroids and also a cell surface component, the H-Y antigen found in individuals carrying the heter- ogametic sex (Zaborski, 1982; Kallman, 1984). Sex inversion can be attained by the administration of synthetic steroids during the undifferentiated stage of the young fish (e.g., Clemens and Inslee, 1968; Miiller, 1969; Guerrero, 1982; Goudie et al., 1983; Rothbard et al., 1983). This may imply that steroids play an important role in sex differentiation in fish (see the reviews by Hunter and Donaldson, 1983, and by Reinboth, 1983).
The present work was undertaken in order to clarify whether natural sex differentiation in tilapia is, indeed, associated with specific changes in the con- ~entrations of endogenous steroids such as testosterone, ll-ketotestosterone and estradiol.
MATERIALS AND METHODS
Fish stocks
Three stocks of tilapias were used for single pair spawnings, which were either interspecific or intraspecific:
1. T. nilotica (Ghana strain), introduced from Ghana by the Gan-Shmuel Fish Breeding Centre, which now serves as the major maternal parent in com- mercial crosses with T. uureu in Israel.
2. T. mossambicu is the most widely distributed tilapia in Africa and the Far East ( Wohlfarth and Hulata, 1983). The stock used for the present investi- gation was introduced from Natal, South Africa (Pike, 1983) by the Israeli Agricultural Research Organization (I.A.R.O. ) Fish and Aquaculture Research Station, Dor.
3. T. hornorum originated from Zanzibar; its crosses with T. mossambica yield all-male populations (Hickling, 1960; Chen, 1969). The Israeli stock of T. hornorum originates from the Ivory Coast ( Obi and Shelton, 1983) and was introduced into Israel from Brazil by the I.A.R.O. Fish and Aquaculture Research Station, Dor.
The stocks were m~nt~ned under controlled conditions at the Gan-Shmuel

61
TABLE 1
Mating combinations performed in the present study
Combination Female Male Expected sex
of progeny*
No. of
replicates
Intraspecific T. nilotica T. nilotica Females and males 2
7’. mossambica T. mossambica 2 T. hornorum T. hornorum 1
Interspecific T. niloticn T. hornorum Males only 2
T. mossambica T. homorum 3
*According TV Hickting (1960), Pruginin ( 1968), Chen (1969), Lovshin et al. (1977) and Hufata et al. (1981).
Fish Breeding Centre and the LA.R.0. Volcani Centre and Fish and Aquacul- ture Research Station, Dor.
Spawning combinations and production of single-pair spawns
The interspecific and intraspecific spawning combinations used are pre- sented in Table 1. The fish were maintained either in 500-I glass aquaria or in 600-I plastic tanks, in “families” consisting of one male and 7-10 females f Rothbard and Pruginin, 1975; Rothbard, 1979), At 3-4 days after spawning, the embryos were washed out from the mothers’ mouths and placed in a closed- system incubator to complete their development (Rothbard and Hulata, 1980). After yolk-sac absorption, each batch of fry (9-10 mm total length) was trans- ferred to 50-l aquaria or 100-l plastic containers connected to the hatchery water system, maintained at 24-28” C. They were fed trout starter (45% pro- tein, “Silver Cup”) ad libitum.
Steroid determinations in batches of specimens early in ontogenesis (one day to seven weeks)
In order to determine the concentrations of testosterone and estradiol in non-differentiated fish, batches of embryos, larvae or fry obtained from intra- specific single pair matings of 2’. niloticu were maintained in 30-l aquaria at 24-28°C. Every 3-4 days, samples consisting of 50 or more fish were randomly collected, homogenized in 3% trichloroacetic acid (TCA) and stored for fur- ther study at - 18°C. Sampling was initiated with l-day-old embryos and con- tinued until l’/2-2 weeks after yolk-sac absorption ( 21/2-3 weeks after spawning).
A further group of 2’. nilotica fry, the progeny of a single pair, was maintained for 4 weeks and then sampled every 3-4 days until the 7th week. On each occasion, a batch of at least 10 fish was sampled and homogenized as above.
The homogenates were extracted with diethyl ether, and the testosterone

and estradiol were determined by radioimmunoassay (RIA) according to Yaron et al. (1977). A monoclonal anti-testosterone antibody was donated by Dr. Naomi Moav (“Interyeda”, Ness Ziona, Israel). The antibody had less than 0.1% cross-reaction with either androsterone, dehy~oepi~n~osterone, 11-j?= hy~ox~estosterone, proges~rone~ ~~-ketotestos~rone, ~7-~~2~-~-~hy- foxy-4-pre~en-3-one or estradiol. The sensitivity of the standard curve was 9 pgj’tube (Yaron and Levavi-Zermonsky, 1986). The anti-estradiol serum was developed in our laboratory; its specificity is described by Bogomolnaya and Yaron (1984).
Eggs were obtained by stripping a ripe T. nilotica as described by Rothbard and Pruginin (1975). Testosterone and estradiol were determined in the unfer- tilized and washed eggs.
Batches of fry obtained from intraspecific and interspecific single pair mat- ings (as shown in Table 1) were maintained at 24-28” C for 6--12 weeks in 56 1 aquaria, Under similar conditions, most species of mouth-bro~ng tilapias will reach sexual ~fferentiation (Eckstein and Spira, 1965; Nakamura and Takahashi, 1973). At least 60 fish were randomly sampled from each batch and were individually homogenized in LO-l.5 ml TCA (3% ) and stored at - 18°C. The extraction and assays for testosterone and estradiol were as described above.
The distribution characteristics of testosterone concentration in the various combinations were analysed by compnting G, and Gz statistics according to Sokal and Ruhlf (1981) .
In order to compare the cuncentrationa of testosterone and fl-ketotestos- terone (ll-KT) in male tilapia fry, a batch of 75 fish, which were the progeny of a single pair interspecific cross between T. nilotica (female) and T. horno- rum (male), was grown to the age of 12 weeks. The fish were then individually homogenized in 3% TCA. The steroids were extracted from the homogenate of each fish and assayed as described above. Forty of the extract samples were each divided into two aliquots. In one, testosterone was determined as above, while the other was used for the determination of If-KT. The anti~dy used in this assay was a gift from Dr. D.E. Kime f Sheffield) and the isotopic stan- dard was synthesized from 3H-cortisol (The Radiochemical Centre, Amer- sham) according to Kime and Hyder (1983). The unlabelled standard was purchased from Ikapharm (Israel). Testosterone was measured in 40 of the fish while ll-KT was measured in all 75 specimens.
Larvae and fry of T. niloticu were collected at 5-day intervals for 12 weeks and immersed in Bouin’s fluid after incision of the abdomen. Transverse par-

63
1.0
0.5
at
r m z
2 0.05
-0 ._ 0
G
;;
i I I I 1 I i
t I
o-+ E,
1 -T
I
----kY- 1 weeks
Fii, 1. Testosterone (T) and estradiol (Ey ) concentrations in 3 batches of TiEapia ni&icu embryos, larvae and fry during the first 3 weeks after spawning; each point represents a sample of 50-57 individuals.
affin sections, 10 pm thick, were prepared from the middle third of the fish and stained with haematoxylin and eosin or with Mallory-Azan.
RESULTS
Steroid concentrations in batches of T. nilotica during the first seven weeks after s~awni~
Eggs stripped from the ripe female contained high concentrations of both testosterone and estradiol (c. 3 ng/egg ) .
In 3 different spawns of 2’. nilotica, the concentration of these steroids in the whole body extracts of developing fish decreased sharply and reached very low values 3 weeks after spawning: < 0.04 ng for testosterone and < 0.012 ng for estradiol (Fig. 1).
In another batch of 7’. nilotica grown until the 7th week, both steroids reached the lowest concentration during the 6th week and then began to increase. Dur- ing the following week, testosterone increased tenfold compared with estradiol, reaching 1 ng/in~vidual (Fig. 2 ) .
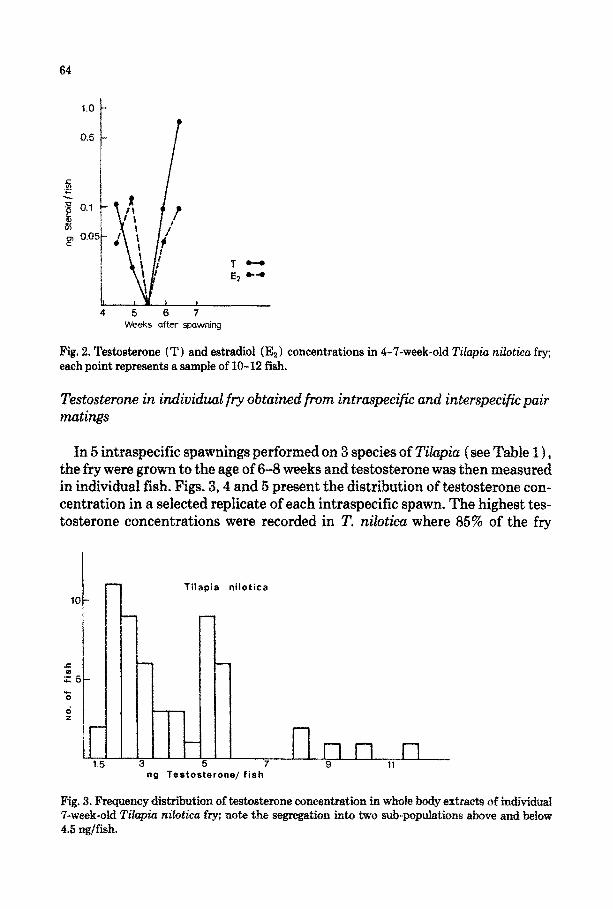
64
Weeks after spawning
Fig. 2. Testosterone (T) and estradiol (E,) concentrations in 4-‘7-week-old Tilapia nilotica fry; each point represents a sample of 10-12 fish.
Testosterone in individual fiy obtained from i~~~~ecif~c and interspecific pair matings
In 5 i~t~~~~~~~~ ~~~w~i~~ performed on 3 species of Ti~u~i~ (see Table I f t the fry were grown to the age of 6-8 weeks and testosterone was then measured in individual fish. Figs. $4 and 5 present the distribution of testosterone con- centration in a selected replicate of each intraspecific spawn. The highest tes- tosterone concentrations were recorded in 2’. nilotica where 85% of the fry
Tilapia nilotica
ng Testosterone/ fish
Fig. 3. Frequency d~t~b~~ion of testosterone concentration in whoie body extracts of indj~idual ?-week-old T&z+ n&&a fry; note the segregation into two sub-populations above and below 4.5 ng/fish.

65
~ J OEiCl2 _
ng 1estos
T. mossambica
Q6 08 Q85 ._. . ronej tisn
Fig. 4. Frequency distribution of testosterone concentration in wbob body extracts of individual 7-week-old Tag ~ssamb~a fry; note the segregation into two sub-~p~atio~ above and below 0.2 ngffish.
showed concentrations varying between 1.5 and 9.7 ng/fish. Testosterone val- ues clustered around two distinct peaks (Fig. 3 ) . In 2’. mossambica fry, testos- terone concentrations reached only 0.85 ng/f%h (Fig. 4). The lowest mean
Tilapia hornorum
0.1 0.2 ng Testosterone/ fish
Fig. 5. Frequency ~st~bution of testosterone concentration in whoie body extracts of individ~ &&week-old Taco ~r~~~~rn fry; note the segregation into two sub-~pulations above and below 0.12 ngffish.
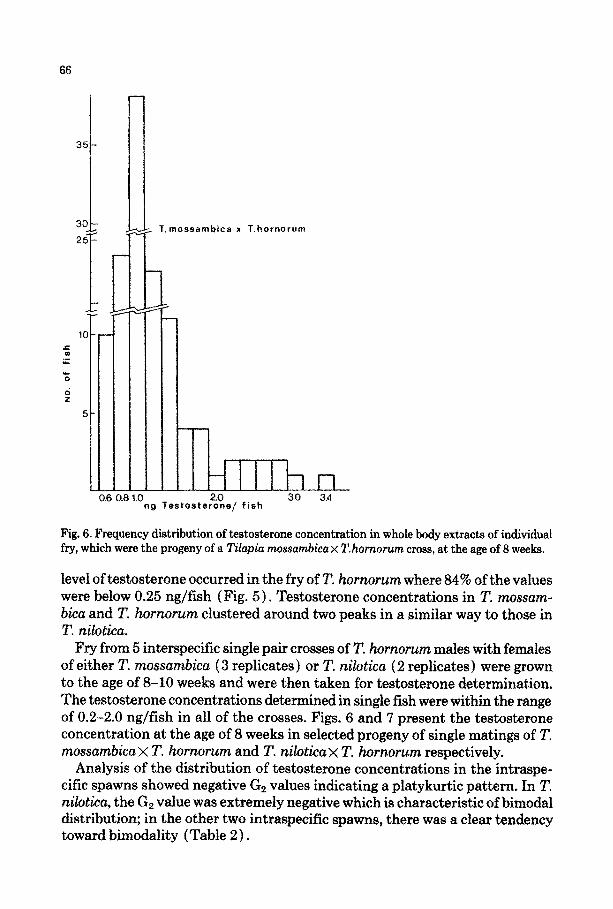
66
3!
31
2!
11 _c m E
b
z I
T. mossambica x T.hornorum
ix p Testosterone/ fish
Fig. 6. Frequency distribution of testosterone concentration in whole body extracts of individual fry, which were the progeny of a Tilapia mossambica x T. hornorum cross, at the age of 8 weeks.
level of testosterone occurred in the fry of 2’. ~r~ru~ where 84% of the values were below 0.25 ng/fish (Fig. 5). Testosterone concentrations in T. 1)2ossczn- bica and T. ~or~$u~ chxstered around two peaks in a similar way to those in T. ~~~~~U.
Fry from 5 interspecifEc single pair crosses of T. hornorum males with females of either T. ~~s~~~cu ( 3 replicates ) or T. nilotica (2 replicates) were grown to the age of 8-10 weeks and were then taken for ~stosterone determination. The testosterone concentrations determined in single fish were within the range of 0.2-2.0 ng/fish in all of the crosses. Figs. 6 and 7 present the testosterone concentration at the age of 8 weeks in selected progeny of single matings of T. ~oss~~bica x T. hor~oru~ and T. ni~ticu x T. ~rnoru~ respectively.
Analysis of the ~stribution of testosterone concentrations in the intraspe- cific spawns showed negative G, values indicating a platykurtic pattern In T. ~i~t~~, the Gz value was extremely negative which is characteristic of bimodal ~stribution; in the other two intrasp~i~c spawns, there was a clear tendency toward bimodality (Table 2 1.

67
n T. nilotica x T. hornorum
It- 0 1.0 2.0 3.0
ng Testosterone/ fish
Fig. 7. Frequency distribution of testosterone concentration in whole body extracts of individual fry, which were the progeny of a Tilapia nilotica x T. hornorum cross, at the age of 8 weeks.
In contrast, the ~stribution analysis of testosterone concentrations in the interspecific spawns producing all male progeny were unimodal and leptokur- tic as indicated by the positive G, values (Table 2). The G, values of all of the spawns differed from zero indicating an asymmetric distribution.
TABLE 2
Analysis of skewness and kurtosis (G, and G, statistics) of testosterone concentrations in the groups presented in Figs. 3-7 (a bimodal distribution is characterized by a large and negative G,)
Spawn N G,iSE G,kSE Shape of distribution
T. ni~otica 55 0.250 z!z 0.322 - 1.500 2 0.634 Platy~~i~ and skewed to the right’
T. mossambica 125 - 0.054 & 0.258 -0.899 rt 0.511 Platykurtic and skewed to the left’
T. hornorum 61 0.146 zk 0.306 -0.753 It 0.604 Platykurtic and skewed to the right’
T. mossambica 87 1.979+0.217 4.298 t 0.430 Leptokurtic and skewed X to the right T. hornorum T. nilotica 56 1.75620.319 3.933 It 0.628 Leptokurtic and skewed X to the right T. hornorum
‘The five extremely skewed data points in Figs. 3 and 4 were grouped in the last class. ‘The last seven extreme date points in Fig. 5 were omitted.

68
30
20
T.mossambica x T. hornorum
.~ .c - .I? lo-
o
d +
n n 0.1 0.2 0.3 0.4 0.5 0.6 07
ng Estradiol/ fish
Fig. 8. Frequency distribution of estradiol concentration in whole body extracts of individual fry, which were the progeny of a Tilapia mossambica x T. hornorum cross, at the age of 8 weeks.
~str~io~ and 11-~etotestosteron~ co~entrutio~
The concentration of estradiol in all of the spawns, whether intraspecific or interspecific, rarely exceeded 0.4 ng/individual and in most fish it was less than 0.1 ng/individual. Fig. 8 represents the distribution of estradiol in one of the progeny at the age of 8 weeks.
The androgen 11-ketotestosterone was only measured in the progeny of a cross between T. nilotica (female) and T. hornorum (male) at the age of 12 weeks. In most individuals the steroid was below 0.05 ng/fish. The mean ( -t SEM) concentration of ll-KT was 7.39 + 1.01 pg/fish (n = 75), compared with a mean ( +-SEM) testosterone concentration of 195.3sf:‘75.8 ngjifish in the same individuals ( n = 40 ) .
Histo~og~cul e~uminution of the developing gonads
T. niloticu at the age of 6 or 12 weeks showed either typical ovaries or undif- ferentiated gonads, probably presumptive testes. The ovary of a 34-mm, 12- week-old fish had a cross-section of 200 x 186 pm and already contained oocy- tes at the perinucleolar stage, surrounded by follicular elements. In addition, the ovaries contained smaller gonocytes and a distinct ovarian cavity (Fig. 9). At this age, the presumptive testes of a 33-mm fish were much smaller than the ovaries (116 x 60 ,um in cross-section). They consisted of clusters of germ cells, undifferentia~d mesodermal cells and blood capillaries (Fig. 10). Pri- mortal germ cells occurred around and near the un~fferentiated gonad and a

Fig. 9. Ovary of a 12-week-old ~i~~io ~i~otic~ fry, 34 mm in total length, showing oocytes at the perinucleolar stage (0) and their foliicukr envelope (F f , peripheral nucleoli ( N) and undiffer- entiated germ cells (G); stained with Mallory-Azan; scale: 40 pm.
Fig. 10. Undifferentiated gonad of a 12-week-old Tilapia nilotica fry (presumptive male), 33 mm in total length, showing undifferentiated mesodermal cells (R) , primordial germ cells (P) , dif- ferentiating germ cells (G) , capillaries (C) and a slit probably representing the future efferent duct (D) ; stained with haematoxyhn and eosin; scale: 40 pm.
typical slit could be seen, probably representing the future efferent duct (Nak- amura and Takahashi, 1985).
DISCUSSION
In the present work, an attempt was made to clarify whether testosterone and estradiol are involved in the normal process of sex differentiation in tila- pias. In most animals, a similar study would have been hindered by the diffi- culty in distinguishing between the sexes. Advantage was taken of the fact that hybridization between certain tilapia species results in all-male progeny, which enables prediction of the sex of all individuals.
The results show that, in T. nilotica, eggs stripped from the ovaries contained high concentrations of both testosterone and estradiol. Since estradiol has also been found in a total ovarian extract of 2’. aurea (91 t 7 ng/kg, Katz et al., 1971) , it is assumed that these steroids accumulate in the oocytes prior to and during vitellogenesis. In spite of the variability, in all of the batches the con- centrations of these steroids decreased sharply during embryonic and larval development, reaching very low values within 3-5 weeks (Figs. l-2). In the weeks following, testosterone concentrations increased again (Fig. 2) reaching 1.5-13.0 ng/fish at 7 weeks (Fig. 3). Apparently, this increase reflects self- production of this androgen in the fry.
The results of testosterone determinations in whole body extracts of 7-week- old T. nilotica, consisting of both females and prospective males, showed a clear bimodal distribution. In the other intraspecific spawns ( 7’. ~0s~~~~~~~ and T.

70
~r~rum) only a tendency toward bimodal distribution could be shown. It should be emphasized in this context that the absolute testosterone concen- tration in these species was considerably lower than that in 2’. nilotica (Figs. 3-5). In interspecific crosses expected to yield all-male progeny, the distribu- tion of testosterone concentrations in fry of the same age was unimodal,
Ovarian differentiation precedes that of the testis in 2’. aurea (Eckstein and Spira, 1965)) I: zillii (Yoshikawa and Oguri, 1978)) 2’. mossambica (Naka- mura and Takabashi, 1985) and also in ~e~i~~~c~~o~~ muZtico2or (Mtiller, 1969). The present work shows that females of ‘I’. ~i~ot~cu are also morphol- ogically differentiated and already show typical follicular elements at the age of 6 weeks. However, in other specimens, although they had a slit in the gonad (possibly a future efferent duct), endocrine components such as Leydig or Sertoli cells could not be identified even at the age of 12 weeks (Figs. 9-10).
The estradiol concentration was very low or undectable in the fry of all of the progeny tested at the age of 6-8 weeks (see Fig. 8 ) . Also the concentrations of ll-KT in whole body extracts of 2’. niZotica~ 7’. hornorum at the age of 12 weeks were extremely low compared with those of testosterone in the same individuals.
The results of the present study do not allow the precise determination of whether the sub-population clustered around the high testosterone values con- sists of males or females. The suggestion that the sub-population with a high testosterone level consists of males disregards the fact that, in the tilapias examined, female sex differentiation precedes that of the males. By 6-12 weeks after fertilization, the ovaries are already morphologically differentiated and possess typical oocytes (Fig. 9) and also follicular elements to which steroi- dogenic capacity is. attributed (Nagahama, 1983). At this age, the gonads of the would-be males have not yet attained the morphological characteristics of the testis (Fig. 10). Moreover, testosterone concentrations in all-male hybrids are clustered around a level closer to that of the “low testosterone” than that of “high testosterone” sub-population in intraspecific spawns (compare Figs. 3 and 7). This would indicate that the sub-population exhibiting high testos- terone levels in intraspeci~c spawns could well be the females which already have differentiated ovaries.
A bimodal distribution of another androgen was also found in immature Oncorhynchus kisutch, in which females show significantly lower levels than males ( Le Bail et al., 1983 ) . However, the differences between the sexes in the salmon were reflected in the plasma level of ll-KT. This androgen was found to be very low or undetectable in the tilapia fry examined in the present study although high levels have been found in mature male and female tilapias ( Katz and Eckstein, 1974; Kime and Hyder, 1983). It would appear, therefore, that ll-KT, although a functioning androgen in the adult stage of tilapia, is not involved in the physiology of the early gonads.
Mas~ulinization can be achieved in tilapia by androgen treatment at an early

71
critical age. Apparently, during this process, part of the population genetically determined to produce females is inverted into phenotypic males. The long list of steroids that can induce sex inversion in fish (Yamazaki, 1983; Hunter and Donaldson, 1983) may confirm the theory established by Yamamoto (1969) that sex inducers (“androinducers” and “gynoinducers”) are the main acti- vators in sex differentiation. However, this view is not supported by the elec- tron microscopy studies of Oryzias Zutipes demonstrating that sex differentiation of germ cells occurs prior to that of the steroid-secreting cells ( Satoh, 1974). In salmonids, enzymes capable of producing estrogens could be found only at an advanced age. At an earlier age the only androgens produced in vitro were androstenedione and ll-~-hy~oxyan~ostenedione. Several progestins were also produced by the undifferentiated gonad at a very early stage. Morphol- ogically-typical steroidogenic cells could be found only after sex differentia- tion; hence, the site of steroid production in the undifferentiated trout gonad is still uncertain (Van den Hurk et al., 1982). These results emphasize the gradual appearance of steroidogenic enzymes during larval development, and are in agreement with the present work where neither ll-KT nor estradiol could be found up to the age of 12 weeks.
The effect of androgen treatment on sex inversion is maximally exerted dur- ing a critical period of sex ~fferentiation, which occurs in tilapia fry between 2. and 6 weeks after spawning (Guerrero, 1982; Rothbard et al., 1983). Little is known, however, about which developmental events are susceptible to androgen induction or whether androgens play any role at all during this crit- ical period in the normal course of events. Several modes of action have been suggested for androgen-induced sex differences in the adult vertebrate nervous system (Arnold and Gorski, 1984). It has been suggested that androgens con- trol sex differences in neuron numbers in certain brain nuclei by preventing the normal cell death (Nordeen et al., 1985).
The steroid concentrations in developing tilapia, as determined in the pres- ent study, may be divided into three phases. The first phase (up to 3-4 weeks) is characterized by a continuous decrease in testosterone and estradiol stored in the egg. In the second phase (4-6 weeks), endogenous steroids are very lo& or undetectable. Sex inversion by androgen treatment is effective during a crit- ical period between 2-6 weeks when endogenous testosterone concentrations are relatively low. In the third phase (6-12 weeks), endogenous estradiol remains low while testosterone increases gradually, presumably only in females, reflecting the steroidogenic activity in the differentiating ovary.
It is speculated that the steroids stored within the egg do not allow differ- entiation into either sex during the initial period. Later, when the stored ste- roids are exhausted, female gonads begin to differentiate. Exogenous androgens may postpone this process by some unknown mechanism ( s) resulting in mas- culinization. Based on our results, it would appear that the increased testos-

72
terone production in 6-&week-old tilapia fry is a result, rather than the cause, of sexual differentiation.
ACKNOWLEDGEMENT
We thank Dr. D.E. Kime (Sheffield, England) for the anti-11-KT serum and Dr. Naomi Moav (“Interyeda”, Israel) for the testosterone monoclonal antibodies used in this research. We also thank Dr. G. Hulata (I.A.R.O. Vol- cani Center, Israel) for some of the tilapia groups examined in the study and Mrs. Hassia Rubin-Kedem for her assistance with the steroid determinations. Thanks are due to Professor L. Fishelson (Tel Aviv University), Dr. G. Hulata and Dr. G. Wolhfarth (I.A.R.O., Dor Station, Israel) and Professor R. Rein- both (Mainz) for their comments on the manuscript.
REFERENCES
Arnold, A.P. and Gorski, R.A., 1984. Gonadal steroid induction of structural sex differences in the central nervous system. Annu. Rev. Neurosci., 7: 413-442.
Avtalion, R.R. and Hammerman, I.S., 1978. Sex determination in Surotherodon (Tilapia). I. Introduction to a theory of autosomal influence. Bamidgeh, 30: 110-115.
Bogomolnaya, A. and Yaron, Z., 1984. Stimulation in vitro of e&radio1 secretion by the ovary of a cichlid fish, Sarotherodon aureus. Gen. Comp. Endocrinol., 53: 187-196.
Chen, F.Y., 1969. Preliminary studies on the sex-determining mechanism of Tilapiu mossambica Peters and T. hornorum Trewavas. Verh. Int. Ver. Limnol., 17: 719-724.
Clemens, H.P. and Inslee, T., 1968. The production of unisexual broods by Til~ia mossumbica sex-reversed with methyl-~s~s~~ne. Trans. Am. Fish. Sot., 97: 18-21.
E&stein, B. and Spira, M., 1965. Effect of sex hormones on gonadal differentiation in a cichlid, Tilapia aurea. Bioi. Bull., 129: 482-489.
Goudie, C.A., Redner, B.D., Simco, B.A. and Davis, K.B., 1983. Feminization of channel catfish by oral administration of steroid sex hormones. Trans. Am. Fish. Sot., 112: 670-672.
Guerrero III, R.D., 1982. Control of tilapia reproduction. In: R.S.V. Pullin and R.H. Lowe- McConnell (Editors 1, The Biology and Culture of Tilapias, ICLARM Conf. Proc., 7. ICLARM, Manila, pp. 309-316.
Hammerman, IS. and Avtahon, R.R., 1979. Sex determination in Sarotherodon (Tikzpia). II. The sex ratio as a tool for the destination of genotype - a model of autosomal and gonosomal intluence. Theor. Appl. Genet., 55: 177-18’7.
Hickling, C.F., 1960. The Malacca tilapia hybrids. J. Genet., 57: l-10. Hulata, G., Rothbard, S. and Wohlfarth, G., 1981. Genetic approach to the production of all-male
progeny of Tilapia. European M&culture Sot. Spec. Publ., 6: 181-190. Hunter, G.A. and Donaldson, E.M., 1983. Hormonal sex control and its application to fish culture.
In: W.S. Hoar, D.J. Randall and E.M. Donaldson (Editors), Fish Physiology, Vol. 9B. Aca- demic Press, New York, pp. 223-303.
Jalabert, B., Kammacher, P. and Lessent, P., 1971. D~~rminisme du sexe chez ies hybrides entre Tilapia ~croch~r et Tilapia niloticu. Etude de la sex-ratio dans les recroisements des hybrides de premiere generation par les especes pare&es. Ann. Biol. Anim. Biochim. Biophys., 11: 155-165.

73
Kallman, K.D., 1984. A new look at sex determination inpoeciliid fishes. In: B.J. Turner (Editor), Evolutionary Genetics of Fishes. Plenum, New York, pp. 95-171.
Katz, Y. and Eckstein, B., 1974. Changes in steroid concentration in blood of female Z’ilapia aurea (Teleoetei, Cichlidae) during initiation of spawning. Endocrinology, 35: 963-967.
Katz, Y., Eckstein, B., Ikan, R. and Gottlieb, R., 1971. Estrone and estradiol-17p in the ovaries of Tilapia aurea (Teleostei, Cichlidae) . Comp. Biochem. Physiol., 40B: 1006-1009.
Kime, DE. and Hyder, M., 1983. The effect of temperature and gonadotropin on testicular ste- roidogenesis in Sarotherodon ( Tilapia) mossambicus in vitro. Gen. Comp. Endocrinol., 50: 105-115.
Le Bail, P.-Y., Fostier, A. and Marcuzzi, O., 1983. Limites et amdliorations du sexage des salmo- nides par dosage de la 11-c~to~stos~rone circulante. Can. J. Zool., 61: 457-460.
Lovshin, L.L., 1982. Tilapia hybridi~tion. In: R.S.V. Pullin and R.H. owe-McConnell (Edi- tars) , The Biology and Culture of Tilapias. ICLARM Conf. Proc., 7. ICLARM, Manila, pp. 279-308.
Lovshin, L.L., Da Silva, A.B. and Fernandez, J.A., 1977. The intensive culture of all male hybrid of Tilapia hornorum (male) x T. nilotica (female) in Northeast Brazil. FAO Fish. Rep., 159: 162-179 (In Spanish with English abstract).
Majumdar, K.C. and McAndrew, B.J., 1983. Sex ratios from interspecific crosses within the tila- pias. In: L. Fishelson and Z. Yaron (Editors), Proceedings of the International Symposium on Tilapia in Aquaculture. Tel Aviv University, Tel Aviv, pp.261-269.
Miiller, R., 1969. Die Einwirkung von Sexsualhormonen auf die Geschlechtsdifferenzierung von Hemihaplochromis multicolor (Hilgendorf) - (Cichlidae). Zool. Jahrb. Physiol., 74: 519-562.
Nagahama, Y., 1988. The functional morphology of teleost gonads. In: W.S. Hoar, D.J. Rendall and E.M. Donaldson (Editors), Fish Physiology, Vol. 9A. Academic Press, New York, pp. 223-275.
Nakamura, M. and Takahashi, H., 1973. Gonadal sex differentiation in Tilapia mossambica with special regard to the time of estrogen treatment effective in inducing complete feminization of genetic males. Bull. Fat. Fish. Hokkaido Univ., 24 l-18.
Nakamura, M. and Takahashi, H., 1985. Sex control in cultured tilapia (Tilupia mossambica) and salmon (Oncorhynchus masou) . In: B. Lofts and W.N. Holmes (Editors), Current Trends in Comparative Endocrinology. Hong Kong University Press, Hong Kong, pp. 1255-1260.
Nordeen, E.J., Nordeen, K-W., Sengelaub, D.R. and Arnold, A.P., 1985. Androgens prevent nor- mally occurring cell death in a sexually dimorphic spinal nucleus. Science, 229: 671-673.
Obi, A. and Shelton, W.L., 1983. Androgen and estrogen sex reversal in Tilapia hornorum. In: L. Fisheison and Z. Yaron (Editors), Proceedings of the International Symposium on Tilapia in Aquaculture. Tel Aviv University, Tel Aviv, pp. 165-173.
Pike, T., 1988. Oreoch~rn~s mossambic~ in Nat& Natural distribution and growth studies. In: L. Fishelson and Z. Yaron (Editors), Proceedings of the In~rnation~ Symposium on Tilapia in Aquaculture. Tel Aviv University, Tel Aviv, pp. 33-47.
Pruginin, Y., 1968. The culture of carp and tilapia hybrids in Uganda. FAO Fish. Rep., 44: 223-229. Reinboth, R., 1983. The peculiarities of gonad transformation in teleosts. Differentiation Suppl.,
23: 82-86. Rothbard, S., 1979. Observations on the reproductive behaviour of Tilapia zillii and several Sar-
otherodon spp. under aquarium conditions. Bamidgeh, 81: 35-43. Rothbard, S. and Hulata, G., 1980. Closed-system incubator for cichlid eggs. Prog. Fish Cult., 42:
203-204. Rothbard, S. and Pruginin, Y., 1975. Induced spawning and artificial incubation of Tilapia. Aqua-
culture, 5: 315-321. Rothbard, S., Solnik, E., Shabbath, S., Amado, R. and Grabie, I., 1983. The technology of mass
production of hormon~y sex-inversed all-male tilapias. In: L. Fishelson and 2. Yaron (Edi-

74
tars) , Proceedings of the Inflations S~~sium on Tilapia in Aqua~ultu~. Tel Aviv Uni- versity, Tel Aviv, pp. 425-434.
Satoh, N., 1974. An ultrastructural study of sex differentiation in the teleost Oryzios latipes. J. Embryol. Exp. Morphol., 32: 192-215.
Shelton, W.L., Meriwether, F.H., Semmens, K.J. and Calhoun, W.E., 1983. Progeny sex ratios from intraspecific pair spawnings of Tilapia aurea and T. nilotica. In: L. Fishelson and Z. Yaron (Editors), Proceedings of the International Symposium on Tilapia in Aquaculture. Tel Aviv University, Tel Aviv, pp. 270-286.
Sokal, R.R. and Rohlf, F.J., 1981. Biometry. Second edition. W.H. Freeman, San Francisco, CA,
859 PP. Van den Hurk, R., Lambert, J.G.D. and Peute, J., 1982. Steroidogenesis in the gonads of rainbow
trout fry (Salmo gaird~ri) before and after the onset of gonadal sex diffe~ntiation. Reprod. Nutr. Dt?v., 22: 413-425.
Wohlf~h, G.W. and Hula@ G., 1983. Applied genetics of tilapias. Second edition. ICLARM Stud. Rev., 6. ICLARM, Manila, 26 pp.
Yamamoto, T., 1969. Sex differentiation. In: W.S. Hoar and D.J. Randall (Editors), Fish Phys- iology, Vol. 3. Academic Press, New York, pp. 117-175.
Yaron, 2. and Levavi-Zermonsky, B., 1986. Fluctuations in gonadotropin and ovarian steroids during the annual cycle and spawning of the common carp. Fish Physiology and Biochemistry, 2: 15-86.
Yaron, Z., Terkatin-Shimony, A., Shaham, Y. and Salzer, H., 1977. Occurrence and biological activity of estradiol-17/3in the intact and ovariectomixed Tilapia aurea (Cichlidae, Teleostei) . Gen. Comp. Endocrinol., 33: 45-52.
Yoshikawa, H. and Oguri, M., 1978. Sex differentiation in a cichlid, Tilapia G&i. Bull. Jpn. Sot. Sci. Fish., 44: 313-318.
Zaborski, P., 1982. Expression of H-Y antigen in non~m~m~i~ vertebrates and its reIation to sex differentiation. In: C.J.J. Richter and H.J.Th. Goes (Editors), Proceedings of the Inter- national Symposium on Reproductive Physiology of Fish. PUDOC, Wageningen, pp. 64-68.
Copyright © 2022 FDOKUMEN




















