
Changes in microbial biomass and activity in relation to shifting cultivation and horticultural...
10
Changes in microbial biomass and activity in relation to shifting cultivation and horticultural practices in subtropical evergreen forest ecosystem of north-east India Vanlalhruaii Ralte, H.N. Pandey *, S.K. Barik, R.S. Tripathi, Swapna D. Prabhu Plant Ecology Laboratory, Department of Botany, North-Eastern Hill University, Shillong 793 022, Meghalaya, India Received 17 September 2004; accepted 1 April 2005 Available online 10 May 2005 Abstract The present study conducted in the Nokrek biosphere reserve (BR) of Meghalaya in north-east India aimed at analyzing the impact of human activities such as shifting agriculture (‘Jhum’) and horticultural practices on temporal and spatial changes in microbial biomass and dehydrogenase and urease activities in soil. Microbial biomass-C (MBC) and -N(MBN) as well as dehydrogenase and urease activities were significantly (P < 0.01) greater in the soils of the undisturbed forest ecosystem in core zone of the BR than the soils under various land use practices in the buffer zone of the BR. The microbial biomass-N in the surface soil layer (0–10 cm) was maximum (285.4 μg g –1 ) in the undisturbed core zone and minimum (75.1 μg g −1 ) in the 1-year-old jhum fallow, which was subjected to intense human activities. Similarly, dehydrogenase (TPF released = 0.70 μg g −1 24 h −1 ) and urease activities (NH 4 released = 31.56 μg 100 g −1 6h −1 ) were maximum in the undisturbed core zone and minimum (TPF released = 0.37 μg g −1 24 h −1 and NH 4 released = 10.78 μg 100 g −1 6h −1 , respectively) in the 1-year-old jhum fallow. Thus, human activities in the buffer zone of the BR were responsible for significant reduction in both microbial biomass-N and enzyme activities. Both these properties showed recovery during regrowth of vegetation on jhum fallows. Microbial biomass-N and enzyme activities declined significantly (P < 0.01) with increasing soil depth at all the sites. Seasonal variations in both microbial biomass-N and enzyme activities at all sites were also significant (P < 0.01) with peak in autumn and trough in winter. © 2005 Elsevier SAS. All rights reserved. Keywords: Microbial biomass; Dehydrogenase; Urease; Human activities; Soil physico-chemical properties 1. Introduction The soil microbial biomass constitutes a transformation matrix for all natural organic materials in the soil and acts as a labile reservoir of plant available nutrients (Jenkinson and Ladd, 1981). Since it constitutes a significant part of the poten- tially mineralizable N and serves both as the transformation agent and source-sink of N (Bonde et al., 1998), it plays an important role in N cycling due to rapid turnover rate. Garcìa- Gil et al. (2000) concluded that microbial biomass is a much more sensitive indicator of changing soil conditions than the total organic matter content. In recent years, studies on micro- bial biomass and their activity have engaged the attention of many researchers. However, most of these studies are con- fined to agricultural soils (e.g. Grayston et al., 2001) and in forest ecosystem such studies are rather limited. Microbio- logical indicators have been used by research groups in numer- ous studies of soil restoration in forest ecosystem (e.g. Cara- vaca et al., 2002, 2003). Soil enzymes are essential for catalyzing reactions neces- sary for organic matter decomposition (Ajwa et al., 1999) and their activities are strongly influenced by organic matter content of the soil (Klose and Tabatabai, 2000). The enzyme activities have often been used as indices of microbial activ- ity and soil fertility (Kennedy and Papendick, 1995). Human activities that minimize the organic matter content of the soil may reduce enzyme activities and could alter the availability of nutrients for plant uptake (Dick et al., 1998). Therefore, the study of soil microbial biomass and their potential activ- ity is important for understanding early changes in biological quality of soil following changes in the land management * Corresponding author. Tel.: +91 364 272 2213 (O), 255 0084 (R); fax: +91 364 272 1000/272 2000. E-mail address: [email protected] (H.N. Pandey). Acta Oecologica 28 (2005) 163–172 www.elsevier.com/locate/actoec 1146-609X/$ - see front matter © 2005 Elsevier SAS. All rights reserved. doi:10.1016/j.actao.2005.04.001
Transcript of Changes in microbial biomass and activity in relation to shifting cultivation and horticultural...

Changes in microbial biomass and activity in relation to shiftingcultivation and horticultural practices in subtropical evergreen forest
ecosystem of north-east India
Vanlalhruaii Ralte, H.N. Pandey *, S.K. Barik, R.S. Tripathi, Swapna D. Prabhu
Plant Ecology Laboratory, Department of Botany, North-Eastern Hill University, Shillong 793 022, Meghalaya, India
Received 17 September 2004; accepted 1 April 2005
Available online 10 May 2005
Abstract
The present study conducted in the Nokrek biosphere reserve (BR) of Meghalaya in north-east India aimed at analyzing the impact ofhuman activities such as shifting agriculture (‘Jhum’) and horticultural practices on temporal and spatial changes in microbial biomass anddehydrogenase and urease activities in soil. Microbial biomass-C (MBC) and -N(MBN) as well as dehydrogenase and urease activities weresignificantly (P < 0.01) greater in the soils of the undisturbed forest ecosystem in core zone of the BR than the soils under various land usepractices in the buffer zone of the BR. The microbial biomass-N in the surface soil layer (0–10 cm) was maximum (285.4 µg g–1) in theundisturbed core zone and minimum (75.1 µg g−1) in the 1-year-old jhum fallow, which was subjected to intense human activities. Similarly,dehydrogenase (TPF released = 0.70 µg g−1 24 h−1) and urease activities (NH4 released = 31.56 µg 100 g−1 6 h−1) were maximum in theundisturbed core zone and minimum (TPF released = 0.37 µg g−1 24 h−1 and NH4 released = 10.78 µg 100 g−1 6 h−1, respectively) in the1-year-old jhum fallow. Thus, human activities in the buffer zone of the BR were responsible for significant reduction in both microbialbiomass-N and enzyme activities. Both these properties showed recovery during regrowth of vegetation on jhum fallows. Microbial biomass-Nand enzyme activities declined significantly (P < 0.01) with increasing soil depth at all the sites. Seasonal variations in both microbial biomass-Nand enzyme activities at all sites were also significant (P < 0.01) with peak in autumn and trough in winter.© 2005 Elsevier SAS. All rights reserved.
Keywords: Microbial biomass; Dehydrogenase; Urease; Human activities; Soil physico-chemical properties
1. Introduction
The soil microbial biomass constitutes a transformationmatrix for all natural organic materials in the soil and acts asa labile reservoir of plant available nutrients (Jenkinson andLadd, 1981). Since it constitutes a significant part of the poten-tially mineralizable N and serves both as the transformationagent and source-sink of N (Bonde et al., 1998), it plays animportant role in N cycling due to rapid turnover rate. Garcìa-Gil et al. (2000) concluded that microbial biomass is a muchmore sensitive indicator of changing soil conditions than thetotal organic matter content. In recent years, studies on micro-bial biomass and their activity have engaged the attention ofmany researchers. However, most of these studies are con-
fined to agricultural soils (e.g. Grayston et al., 2001) and inforest ecosystem such studies are rather limited. Microbio-logical indicators have been used by research groups in numer-ous studies of soil restoration in forest ecosystem (e.g. Cara-vaca et al., 2002, 2003).
Soil enzymes are essential for catalyzing reactions neces-sary for organic matter decomposition (Ajwa et al., 1999)and their activities are strongly influenced by organic mattercontent of the soil (Klose and Tabatabai, 2000). The enzymeactivities have often been used as indices of microbial activ-ity and soil fertility (Kennedy and Papendick, 1995). Humanactivities that minimize the organic matter content of the soilmay reduce enzyme activities and could alter the availabilityof nutrients for plant uptake (Dick et al., 1998). Therefore,the study of soil microbial biomass and their potential activ-ity is important for understanding early changes in biologicalquality of soil following changes in the land management
* Corresponding author. Tel.: +91 364 272 2213 (O), 255 0084 (R); fax:+91 364 272 1000/272 2000.
E-mail address: [email protected] (H.N. Pandey).
Acta Oecologica 28 (2005) 163–172
www.elsevier.com/locate/actoec
1146-609X/$ - see front matter © 2005 Elsevier SAS. All rights reserved.doi:10.1016/j.actao.2005.04.001

(Palma et al., 2000). The objective of the present study was toanalyze the effect of various human activities on soil biologi-cal properties of forest ecosystem in Nokrek biosphere reserve(BR), north-east India. The study sites that were exposed todifferent degrees of human activities, such as ‘Jhum’ (shift-ing cultivation or slash and burn agriculture) fallows of dif-ferent ages undergoing natural recovery, tea gardens andorange orchards recently raised in the buffer zone as well asthe undisturbed primary forest in the core zone. In order toachieve the aforesaid objective, the soil microbial biomass-Nand biomass-C, and soil enzyme (urease and dehydrogenase)activities were measured in the undisturbed forest and above-mentioned communities that varied in the degree of distur-bance and intensity of human intervention.
2. Materials and methods
2.1. Study site
The study was carried out in the Nokrek BR (latitude25°20′N–25°29′N; longitude 90°13′E–90°35′E) situated inthe western part of the state of Meghalaya, north-east India.The altitude of the BR ranged between 200 and 1412 m (a.s.l.).The BR has an area of 820 km2, of which 47.48 km2 is cov-ered by undisturbed primary subtropical evergreen forest. Thisportion is designated as the Nokrek National Park and con-stitutes the core zone of the BR spreading in east–west direc-tion. The northern aspect of National Park is comparativelyof gentle slope while the southern flank consists of hills withvery steep to moderate slopes.
About 129 villages of the Garo tribe with a total popula-tion of 39,432 are located in the buffer zone. Shifting agri-culture is extensively practiced by these people in the entirebuffer zone. This agricultural practice involves slashing orcomplete clearing of the vegetation and burning of dried slashduring dry winter, followed by pure and mixed cropping for1–2 years depending upon the availability of land in a par-ticular village. Thereafter the field is abandoned for naturalrecovery of soil fertility. Since shifting agriculture is causinglarge-scale degradation of land in the buffer zone of the BR,the government agencies have encouraged the establishmentof tea gardens and orange orchards during the past 5 years,with twin objective of improving the economic condition ofthe people and checking further land degradation in the BR.In order to study the impact of these activities on soil, fourabandoned jhum fields with 1, 3–4, 6–8 and 10–12 years oldvegetational regrowth, tea gardens and orange orchards in thebuffer zone and a primary forest stand in the undisturbed corezone were selected. For each of the four jhum fields, threereplicate sites were identified for the sampling purpose, whileonly two replicate sites were available for tea garden andorange orchard in the BR. The site characteristics of the sam-pling locations are summarized in Table 1.
The subtropical semi-evergreen forest in the core zone isdominated by tree species like, Calophyllum polyanthum,
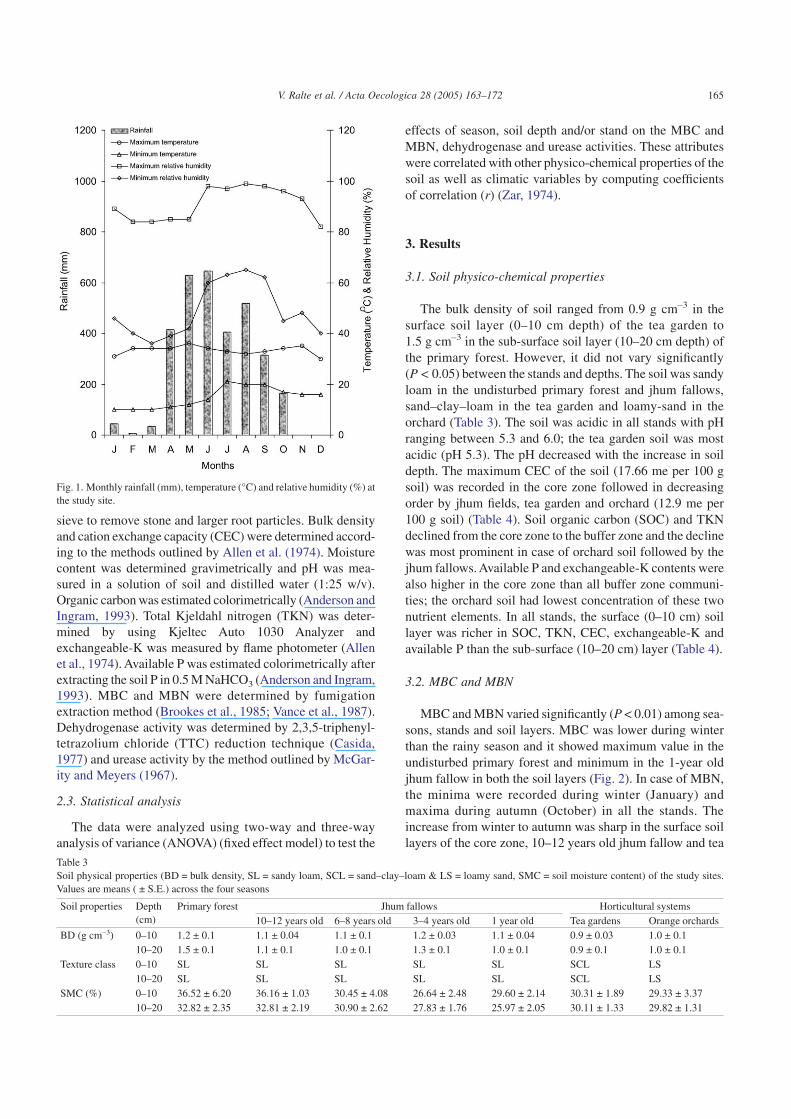
Eleaocarpus tectorius, Helicia nilagirica, Litsea lancifolia,Neolitsea umbrosia and Syzygium cumini. The fast growingweeds like Eupatorium adenophorum, Mikania micranthaand a variety of grasses were dominant in the 1-year old stand,while 3–12 years old stands were dominated by secondarysuccessional tree species such as Callicarpa vestita, Euryaacuminata, Macaranga indica, and Saurauia roxburghii. Thedensity and basal cover of trees, shrubs and herbs are givenin Table 2. The tree cover was highest in the undisturbed pri-mary forest followed by 10–12, 6–8 and 3–4 years old jhumfallows. The climate of the area is monsoonic with a longrainy season (April–September) and moderately cool short-dry winter (November–February). The transitional periodsbetween rainy and winter (i.e. October) and winter and sum-mer (i.e. March) seasons represent autumn and spring, respec-tively. The annual rainfall during the study period (2000) was3173 mm, with highest rainfall (644.6 mm) in June. May wasthe hottest month of the year with a mean maximum tempera-ture of 36 °C, while the mean minimum temperature (10 °C)was recorded during January. The relative humidity rangedbetween 36% (March) and 99% (June) (Fig. 1).
2.2. Soil sampling and analysis
Soil samples were collected from two to three representa-tive sites of each of the seven selected stands during January,March,August and October 2000, representing winter, spring,rainy and autumn seasons, respectively.At each sampling date,five soil samples were randomly collected from 0 to 10 cmand 10–20 cm depths at each site using a soil corer and mixedthoroughly to obtain a homogeneous sample for each depthin a given stand. The samples were passed through 2 mm
Table 1Characteristics of the experimental sites
Experimental sites Area(ha)
Slopeangle (°)
Aspect Altitude(m)
Primary forest 10 30–35 East 1250Jhum fallows10–12 years old 10 20–25 East 12286–8 years old 10 30–35 East 11333–4 years old 10 30–35 East 12261 year old 10 30–35 East 1291Tea gardens 6 35 Southeast 950Orange orchards 8 35 Southeast 938
Table 2Density (mean number of individuals ha–1) and basal area (m2 ha–1) of treespecies and density (mean number of individuals ha–1) of shrub and herbspecies at different sampling sites in Nokrek BR
Sampling sites Trees Shrubs HerbsDensity Basal area Density Density
Primary forest 2595 45.95 37,250 115,250Jhum fallows10–12 years old 5665 17.03 51,833 209,5006–8 years old 7560 14.61 44,833 279,5003–4 years old 10,280 5.71 45,000 449,0001 year old – – – 840,500Orange orchards – – – 1041,750
164 V. Ralte et al. / Acta Oecologica 28 (2005) 163–172
sieve to remove stone and larger root particles. Bulk densityand cation exchange capacity (CEC) were determined accord-ing to the methods outlined by Allen et al. (1974). Moisturecontent was determined gravimetrically and pH was mea-sured in a solution of soil and distilled water (1:25 w/v).Organic carbon was estimated colorimetrically (Anderson andIngram, 1993). Total Kjeldahl nitrogen (TKN) was deter-mined by using Kjeltec Auto 1030 Analyzer andexchangeable-K was measured by flame photometer (Allenet al., 1974). Available P was estimated colorimetrically afterextracting the soil P in 0.5 M NaHCO3 (Anderson and Ingram,1993). MBC and MBN were determined by fumigationextraction method (Brookes et al., 1985; Vance et al., 1987).Dehydrogenase activity was determined by 2,3,5-triphenyl-tetrazolium chloride (TTC) reduction technique (Casida,1977) and urease activity by the method outlined by McGar-ity and Meyers (1967).
2.3. Statistical analysis
The data were analyzed using two-way and three-wayanalysis of variance (ANOVA) (fixed effect model) to test the
effects of season, soil depth and/or stand on the MBC andMBN, dehydrogenase and urease activities. These attributeswere correlated with other physico-chemical properties of thesoil as well as climatic variables by computing coefficientsof correlation (r) (Zar, 1974).
3. Results
3.1. Soil physico-chemical properties
The bulk density of soil ranged from 0.9 g cm–3 in thesurface soil layer (0–10 cm depth) of the tea garden to1.5 g cm–3 in the sub-surface soil layer (10–20 cm depth) ofthe primary forest. However, it did not vary significantly(P < 0.05) between the stands and depths. The soil was sandyloam in the undisturbed primary forest and jhum fallows,sand–clay–loam in the tea garden and loamy-sand in theorchard (Table 3). The soil was acidic in all stands with pHranging between 5.3 and 6.0; the tea garden soil was mostacidic (pH 5.3). The pH decreased with the increase in soildepth. The maximum CEC of the soil (17.66 me per 100 gsoil) was recorded in the core zone followed in decreasingorder by jhum fields, tea garden and orchard (12.9 me per100 g soil) (Table 4). Soil organic carbon (SOC) and TKNdeclined from the core zone to the buffer zone and the declinewas most prominent in case of orchard soil followed by thejhum fallows. Available P and exchangeable-K contents werealso higher in the core zone than all buffer zone communi-ties; the orchard soil had lowest concentration of these twonutrient elements. In all stands, the surface (0–10 cm) soillayer was richer in SOC, TKN, CEC, exchangeable-K andavailable P than the sub-surface (10–20 cm) layer (Table 4).
3.2. MBC and MBN
MBC and MBN varied significantly (P < 0.01) among sea-sons, stands and soil layers. MBC was lower during winterthan the rainy season and it showed maximum value in theundisturbed primary forest and minimum in the 1-year oldjhum fallow in both the soil layers (Fig. 2). In case of MBN,the minima were recorded during winter (January) andmaxima during autumn (October) in all the stands. Theincrease from winter to autumn was sharp in the surface soillayers of the core zone, 10–12 years old jhum fallow and tea
Fig. 1. Monthly rainfall (mm), temperature (°C) and relative humidity (%) atthe study site.
Table 3Soil physical properties (BD = bulk density, SL = sandy loam, SCL = sand–clay–loam & LS = loamy sand, SMC = soil moisture content) of the study sites.Values are means ( ± S.E.) across the four seasons
Soil properties Depth(cm)
Primary forest Jhum fallows Horticultural systems10–12 years old 6–8 years old 3–4 years old 1 year old Tea gardens Orange orchards
BD (g cm–3) 0–10 1.2 ± 0.1 1.1 ± 0.04 1.1 ± 0.1 1.2 ± 0.03 1.1 ± 0.04 0.9 ± 0.03 1.0 ± 0.110–20 1.5 ± 0.1 1.1 ± 0.1 1.0 ± 0.1 1.3 ± 0.1 1.0 ± 0.1 0.9 ± 0.1 1.0 ± 0.1
Texture class 0–10 SL SL SL SL SL SCL LS10–20 SL SL SL SL SL SCL LS
SMC (%) 0–10 36.52 ± 6.20 36.16 ± 1.03 30.45 ± 4.08 26.64 ± 2.48 29.60 ± 2.14 30.31 ± 1.89 29.33 ± 3.3710–20 32.82 ± 2.35 32.81 ± 2.19 30.90 ± 2.62 27.83 ± 1.76 25.97 ± 2.05 30.11 ± 1.33 29.82 ± 1.31
165V. Ralte et al. / Acta Oecologica 28 (2005) 163–172

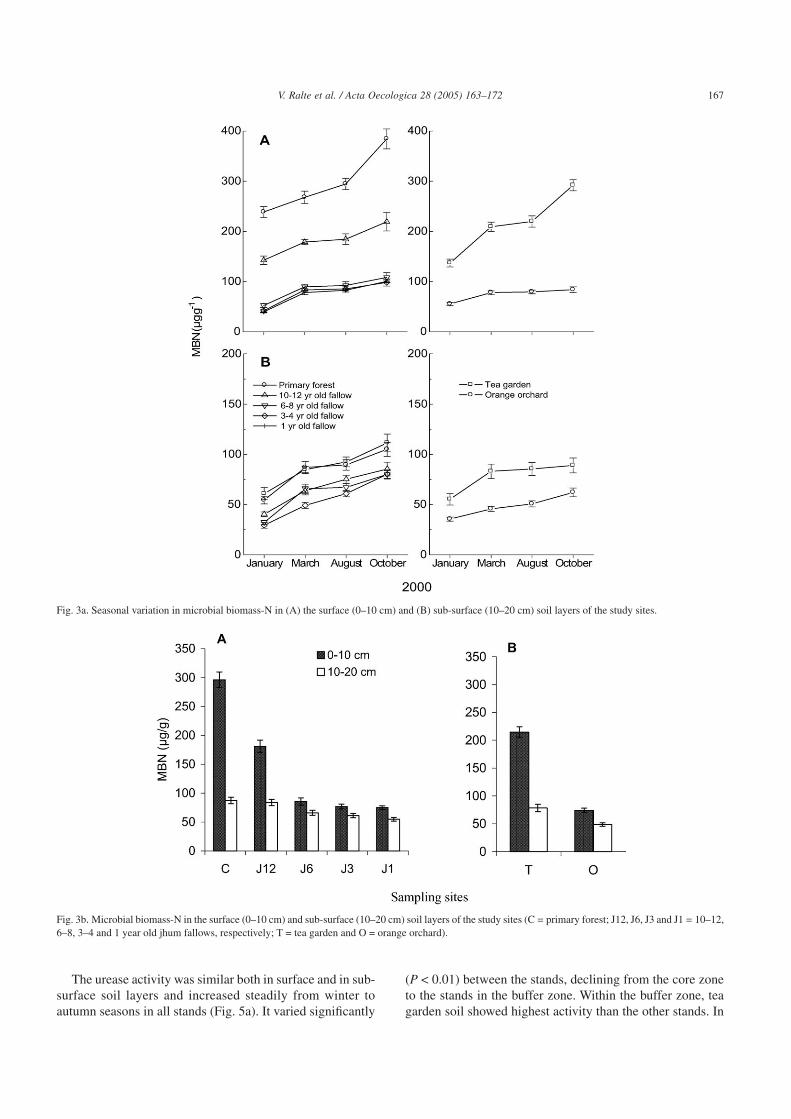
garden in the buffer zone whereas in the sub-surface soil layer,the increase from winter to autumn was gradual at all sites.The surface soil of the undisturbed primary forest had maxi-mum concentration of MBN followed in descending orderby the tea garden, jhum fallows and orchard in the buffer zone.The MBC and MBN in the jhum fallows gradually increasedwith the age of the fallow. Their values declined significantly(P < 0.01) from the surface to the sub-surface soil layer in allcases (Figs. 2 and 3).
3.3. Dehydrogenase and urease activity
Dehydrogenase activity varied significantly (P < 0.01)between the seasons; it was minimum during winter and maxi-mum during autumn, a trend similar to that of MBN (Fig. 4a).
It also showed significant (P < 0.01) variation between thestands. The undisturbed core zone recorded the maximumactivity. In the case of jhum fallows, it increased from the1 to 10–12 years old fallow during winter (January), spring(March) and rainy (August) seasons. However during autumn(October), 3–4 years old fallow had higher dehydrogenaseactivity than 6–8 years old fallow. The soil in tea garden hadgreater activity than the jhum fallows and orchard (Fig. 4b).The activity declined significantly (P < 0.01) from the sur-face to the sub-surface soil layer regardless of the stands. Italso showed a gradual increase from winter to autumn at sur-face layer. At sub-surface layer, the same trend was obtainedfor all the stands except 1 and 6–8 years old jhum fallowswhere the activity increased until spring (March) after whichit gradually declined. In orchard too, the activity started declin-ing after winter (January).
Table 4Soil chemical properties (CEC = cation exchange capacity, SOC = soil organic carbon, TKN = total Kjeldahl nitrogen, Av. P = available phosphorus and Ex.K = exchangeable potassium) of the study sites. Values are means ( ± S.E.) across the four seasons
Soil properties Depth(cm)
Primary forest Jhum fallows Horticultural systems10–12 yearsold
6–8 years old 3–4 years old 1 year old Tea gardens Orangeorchards
Ph 0–10 6.0 ± 0.52 5.9 ± 0.15 5.9 ± 0.24 5.9 ± 0.30 5.8 ± 0.17 5.4 ± 0.29 5.8 ± 0.1310–20 5.8 ± 0.30 5.7 ± 0.16 5.6 ± 0.20 5.5 ± 0.21 5.4 ± 0.16 5.3 ± 0.19 5.5 ± 0.16
CEC (me 100 g per soil) 0–10 17.7 ± 0.04 16.9 ± 0.06 16.2 ± 0.15 16.1 ± 0.49 14.0 ± 0.09 13.8 ± 0.27 12.9 ± 1.3110–20 16.3 ± 0.09 15.4 ± 0.06 14.6 ± 0.11 11.5 ± 0.74 10.1 ± 0.07 11.8 ± 0.10 10.4 ± 0.20
SOC (%) 0–10 6.59 ± 0.95 5.76 ± 0.77 4.91 ± 0.64 4.50 ± 0.40 4.46 ± 0.89 6.19 ± 0.71 4.49 ± 0.9110–20 3.07 ± 0.79 2.60 ± 0.29 2.58 ± 0.30 2.40 ± 0.26 2.08 ± 0.24 3.15 ± 0.37 2.23 ± 0.26
TKN (%) 0–10 0.44 ± 0.05 0.35 ± 0.04 0.33 ± 0.05 0.32 ± 0.02 0.25 ± 0.04 0.37 ± 0.06 0.26 ± 0.0410–20 0.34 ± 0.06 0.26 ± 0.05 0.25 ± 0.09 0.24 ± 0.04 0.23 ± 0.06 0.31 ± 0.05 0.23 ± 0.05
Av. P (µg g–1) 0–10 3.15 ± 0.30 2.53 ± 0.18 2.30 ± 0.30 2.10 ± 0.42 2.05 ± 0.70 3.15 ± 0.48 2.65 ± 0.4710–20 2.85 ± 0.70 2.13 ± 0.55 1.93 ± 0.20 1.84 ± 0.30 1.60 ± 0.45 2.08 ± 0.35 1.85 ± 0.30
Ex. K (µg g–1) 0–10 248 ± 10 200 ± 37 195 ± 12 170 ± 35 223 ± 20 212 ± 36 150 ± 910–20 153 ± 8 123 ± 16 110 ± 11 103 ± 16 115 ± 26 148 ± 24 88 ± 17
N-mineralization rate(µg g–1 per day)
0–10 1.07 ± 0.00 0.61 ± 0.03 0.42 ± 0.01 – 0.35 ± 0.04 – –
Fig. 2. Seasonal variation in MBC in (A) the surface (0–10 cm) and sub-surface (10–20 cm) soil layers of the BR (C = primary forest; J12, J6 and J1 = 10–12,6–8, and 1 year old jhum fallows, respectively).
166 V. Ralte et al. / Acta Oecologica 28 (2005) 163–172
The urease activity was similar both in surface and in sub-surface soil layers and increased steadily from winter toautumn seasons in all stands (Fig. 5a). It varied significantly
(P < 0.01) between the stands, declining from the core zoneto the stands in the buffer zone. Within the buffer zone, teagarden soil showed highest activity than the other stands. In
Fig. 3a. Seasonal variation in microbial biomass-N in (A) the surface (0–10 cm) and (B) sub-surface (10–20 cm) soil layers of the study sites.
Fig. 3b. Microbial biomass-N in the surface (0–10 cm) and sub-surface (10–20 cm) soil layers of the study sites (C = primary forest; J12, J6, J3 and J1 = 10–12,6–8, 3–4 and 1 year old jhum fallows, respectively; T = tea garden and O = orange orchard).
167V. Ralte et al. / Acta Oecologica 28 (2005) 163–172

the jhum fallows the urease activity increased during theregrowth of vegetation from 1 to 10–12 years old stand(Fig. 5b).
4. Discussion
4.1. Seasonal and spatial changes
Ecosystems with high organic matter input and easily avail-able organic matter compounds tend to have higher micro-bial biomass contents and activities because organic sub-stances are the preferred energy source for the microorganisms(Hassink, 1994). The high concentration of detrital materialin the surface soil layer (0–10 cm) in the subtropical forestincreases the availability of soil organic matter in the surfacelayer due to fast turnover rates of litter and fine roots(Arunachalam et al., 1998; John et al., 2002) (Table 4). Thechief contributory factor for the higher MBC and MBN in thecore zone soil than the jhum fallows, tea garden and orangeorchard seems to be the greater availability of organic nutri-ents in the core zone due to higher plant cover (Table 2). This
is clearly evident from the strong positive correlations betweenMBC and SOC (r = 0.71, P < 0.05), MBC and TKN (r = 0.92,P < 0.05), MBN and SOC (r = 0.95, P < 0.01) and MBN andTKN (r = 0.81, P < 0.05). Witter and Kanal (1998) alsoobserved a close relationship between concentrations oforganic C in microbial biomass in different soils of a 40 yearsold field. A positive relation between MBC and MBN(r = 0.96, P < 0.01) shows an intimate link between changesof N in soil and heterotrophic microbes, which use organic Cas their energy source (Arunachalam and Pandey, 2003).
Since microorganisms are mostly confined to the surfacesoil layer owing to better aeration and greater nutrient avail-ability, microbial biomass, and dehydrogenase and ureaseactivities were significantly greater in the surface soil layer(0–10 cm) compared to the sub-surface (10–20 cm) soil layerwhere the organic matter content and nutrient availability waslow and aeration was poor. Seasonal changes in soil mois-ture, soil temperature and available residue could have a strongeffect on soil microbial biomass and its activity (Diaz-Ravina et al., 1995). The seasonal trend of MBN observed inthe present study is in agreement with the findings of VonLützow et al. (1992) who recorded the highest MBN during
Fig. 4a. Seasonal variation in dehydrogenase activity in (A) the surface (0–10 cm) and (B) sub-surface (10–20 cm) soil layers of the study sites.
168 V. Ralte et al. / Acta Oecologica 28 (2005) 163–172

Fig. 4b. Dehydrogenase activity in the surface (0–10 cm) and sub-surface (10–20 cm) soil layers of the study sites (C = primary forest; J12, J6, J3 and J1 = 10–12, 6–8, 3–4 and 1 year old jhum fallows, respectively; T = tea garden and O = orange orchard).
Fig. 5a. Seasonal variation in urease activity in (A) the surface (0–10 cm) and (B) sub-surface (10–20 cm) soil layers of the study sites.
169V. Ralte et al. / Acta Oecologica 28 (2005) 163–172
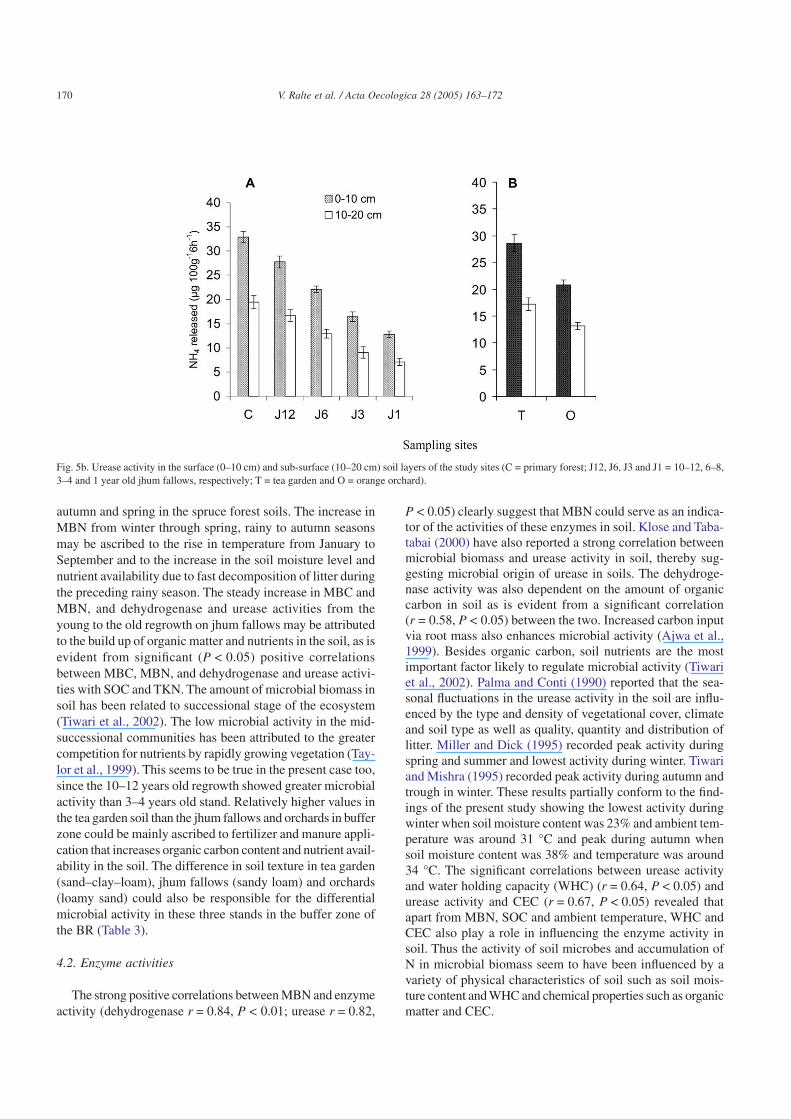
autumn and spring in the spruce forest soils. The increase inMBN from winter through spring, rainy to autumn seasonsmay be ascribed to the rise in temperature from January toSeptember and to the increase in the soil moisture level andnutrient availability due to fast decomposition of litter duringthe preceding rainy season. The steady increase in MBC andMBN, and dehydrogenase and urease activities from theyoung to the old regrowth on jhum fallows may be attributedto the build up of organic matter and nutrients in the soil, as isevident from significant (P < 0.05) positive correlationsbetween MBC, MBN, and dehydrogenase and urease activi-ties with SOC and TKN. The amount of microbial biomass insoil has been related to successional stage of the ecosystem(Tiwari et al., 2002). The low microbial activity in the mid-successional communities has been attributed to the greatercompetition for nutrients by rapidly growing vegetation (Tay-lor et al., 1999). This seems to be true in the present case too,since the 10–12 years old regrowth showed greater microbialactivity than 3–4 years old stand. Relatively higher values inthe tea garden soil than the jhum fallows and orchards in bufferzone could be mainly ascribed to fertilizer and manure appli-cation that increases organic carbon content and nutrient avail-ability in the soil. The difference in soil texture in tea garden(sand–clay–loam), jhum fallows (sandy loam) and orchards(loamy sand) could also be responsible for the differentialmicrobial activity in these three stands in the buffer zone ofthe BR (Table 3).
4.2. Enzyme activities
The strong positive correlations between MBN and enzymeactivity (dehydrogenase r = 0.84, P < 0.01; urease r = 0.82,
P < 0.05) clearly suggest that MBN could serve as an indica-tor of the activities of these enzymes in soil. Klose and Taba-tabai (2000) have also reported a strong correlation betweenmicrobial biomass and urease activity in soil, thereby sug-gesting microbial origin of urease in soils. The dehydroge-nase activity was also dependent on the amount of organiccarbon in soil as is evident from a significant correlation(r = 0.58, P < 0.05) between the two. Increased carbon inputvia root mass also enhances microbial activity (Ajwa et al.,1999). Besides organic carbon, soil nutrients are the mostimportant factor likely to regulate microbial activity (Tiwariet al., 2002). Palma and Conti (1990) reported that the sea-sonal fluctuations in the urease activity in the soil are influ-enced by the type and density of vegetational cover, climateand soil type as well as quality, quantity and distribution oflitter. Miller and Dick (1995) recorded peak activity duringspring and summer and lowest activity during winter. Tiwariand Mishra (1995) recorded peak activity during autumn andtrough in winter. These results partially conform to the find-ings of the present study showing the lowest activity duringwinter when soil moisture content was 23% and ambient tem-perature was around 31 °C and peak during autumn whensoil moisture content was 38% and temperature was around34 °C. The significant correlations between urease activityand water holding capacity (WHC) (r = 0.64, P < 0.05) andurease activity and CEC (r = 0.67, P < 0.05) revealed thatapart from MBN, SOC and ambient temperature, WHC andCEC also play a role in influencing the enzyme activity insoil. Thus the activity of soil microbes and accumulation ofN in microbial biomass seem to have been influenced by avariety of physical characteristics of soil such as soil mois-ture content and WHC and chemical properties such as organicmatter and CEC.
Fig. 5b. Urease activity in the surface (0–10 cm) and sub-surface (10–20 cm) soil layers of the study sites (C = primary forest; J12, J6, J3 and J1 = 10–12, 6–8,3–4 and 1 year old jhum fallows, respectively; T = tea garden and O = orange orchard).
170 V. Ralte et al. / Acta Oecologica 28 (2005) 163–172

4.3. Effect of human activities on vegetation regrowth
Repeated cycles of slash and burn agriculture on hill slopescoupled with high rainfall causing loss of soluble organic mat-ter and nutrients of surface soil layer through overland flowof rainwater appear to be responsible for low enzyme activityin the jhum fallows. On the other hand, the low input of detri-tal material and nutrients through litterfall in the orchardsresulted in the decreased MBN and enzyme activity in theorchards. On the contrary, the highest MBN and enzyme activ-ity in the undisturbed subtropical evergreen forest in the corezone followed by the older regrowth (10–12 years old) on thejhum fallows was due to greater organic matter and nutrientcontents, improved soil structure, and humid microclimateon the forest floor. The increase in MBC and MBN with theincreasing age of the plant regrowth on the jhum fallows maybe attributed to greater input of plant detritus that is ulti-mately incorporated into the soil and improves the nutrientpool (Maithani et al., 1998).
5. Conclusion
Human activities such as shifting agriculture and horticul-tural practices in the hilly buffer zone of the Nokrek BRcoupled with high rainfall cause depletion of the MBC andMBN, and reduction in their activities in soil except in the teagarden where fertilizer and manure were applied. Severalphysico-chemical properties of soil (WHC, pH, CEC, SOC,TKN) and ambient temperature also affected MBC, MBNand enzyme activities. As the vegetation regrows after aban-donment of cultivation on jhum land, MBC, MBN and enzymeactivity gradually increased with age of the secondary suc-cessional communities on the jhum fallows.
Acknowledgements
The authors are grateful to the Ministry of Environmentand Forests, Government of India for the financial supportunder the BR Programme (Sanction No.F.10/41/97-CS/BRdated 15.2.99). Thanks are due to the Forest Department, Gov-ernment of Meghalaya for granting permission to undertakefield study in the BR. We thank the Forest Officers attachedto the BR and villagers inhabiting the buffer zone of the BRfor their help and cooperation. Thanks are also due to Dr.Babu John for his help in the field studies, and soil samplingand analysis.
References
Allen, S.C., Grimshaw, H.M., Parkinson, J.A., Quarmby, C., 1974. Chemi-cal Analysis of Ecological Materials. Blackwell Scientific Publications,Oxford.
Anderson, J.M., Ingram, J.S.I., 1993. Tropical Soil Biology and Fertility—AHandbook of Methods, second ed. CAB International, Wallingford, UK.
Ajwa, H.A., Dell, C.J., Rice, C.W., 1999. Changes in enzyme activities andmicrobial biomass of tallgrass prairie soil as related to burning andnitrogen fertilization. Soil Biol. Biochem. 31, 769–777.
Arunachalam, A., Maithani, K., Pandey, H.N., Tripathi, R.S., 1998. Finelitter and nutrient changes during forest regrowth in humid sub-tropics ofnorth-eastern India. For. Ecol. Manage. 110, 209–219.
Arunachalam, A., Pandey, H.N., 2003. Ecosystem restoration of jhum fal-lows in Northeast India: microbial C and N along altitudinal and succes-sional gradients. Restor. Ecol. 11, 1–6.
Bonde, T.S., Schnurer, J., Rosswall, T., 1998. Microbial biomass as afraction of potentially mineralizable nitrogen in soil from long term fieldexperiments. Soil Biol. Biochem. 21, 447–452.
Brookes, D.C., Landman, A., Pruden, G., Jenkinson, D.S., 1985. Chloro-form fumigation and the release of soil nitrogen: a rapid direct extractionmethod for measuring microbial biomass nitrogen in soil. Soil Biol.Biochem. 17, 837–842.
Caravaca, F., Alguacil, M.M., Figueroa, D., Barea, J.M., Roldán, A., 2003.Re-establishment of Retama sphaerocarpa as a target species for recla-mation of soil physical and biological properties in a semi-arid Mediter-ranean land. For. Ecol. Manage. 182, 49–58.
Caravaca, F., Barea, J.M., Figueroa, D., Roldán, A., 2002. Assessing theeffectiveness of mycorrhizal inoculation and soil compost addition forreforestation with Olea europaea subsp. sylvestris through soil biologi-cal and physical parameters. Appl. Soil Ecol. 20, 107–118.
Casida, L.E., 1977. Microbial metabolic activity in soil as measured bydehydrogenase determination. Appl. Environ. Microbiol. 34, 630–636.
Diaz-Ravina, M., Acea, M.J., Carballas, T., 1995. Seasonal changes inmicrobial biomass and nutrient flush in forest soils. Biol. Fertil. Soils 19,220–226.
Dick, R.P., Rasmussen, P.E., Kerle, E.A., 1998. Influence of long-termresidues management on soil enzyme activity in relation to soil chemicalproperties of a wheat–fallow system. Biol. Fertil. Soils 6, 159–164.
Garcìa-Gil, J.C., Plaza, C., Soler-Rovira, P., Polo, A., 2000. Long-termeffects of municipal solid waste compost application on soil enzymeactivities and microbial biomass. Soil Biol. Biochem. 32, 1907–1913.
Grayston, S.J., Grifith, G.S., Mawdsley, J.L., Campbell, C.D.,Bardgett, R.D., 2001. Accounting for variability in soil microbiologicalcommunities of temperate upland grassland ecosystems. Soil Biol. Bio-chem. 33, 533–551.
Hassink, J., 1994. Effects of soil texture on the size of the microbial biomassand on the amount of C mineralized per unit of microbial biomass inDutch grassland soils. Soil Biol. Biochem. 26, 1573–1581.
Jenkinson, D.S., Ladd, J.N., 1981. Microbial biomass in soil: measurementand turnover. In: Paul, E.A., Ladd, J.N. (Eds.), Soil Biochemistry, vol. 5.Dekker, New York, pp. 415–471.
John, B., Pandey, H.N., Tripathi, R.S., 2002. Decomposition of fine roots ofPinus kesiya and turnover of organic matter, N and P of course and finepine roots and herbaceous roots and rhizomes in subtropical pine foreststands of different ages. Biol. Fertil. Soils 35, 238–246.
Kennedy, A.C., Papendick, R.I., 1995. Microbial characteristics of soilquality. J. Soil Water Conserv. 50, 243–248.
Klose, S., Tabatabai, M.A., 2000. Urease activity of microbial biomass insoils as affected by cropping systems. Biol. Fertil. Soils 31, 191–199.
Maithani, K., Arunachalam, A., Tripathi, R.S., 1998. Influence of leaf litterquality on N mineralization in soils of subtropical humid forestregrowths. Biol. Fertil. Soils 27, 44–50.
McGarity, J.W., Meyers, M.G., 1967. A survey of urease activity in soils ofnorthern New South Wales. Plant Soil 27, 217–238.
Miller, M., Dick, R.P., 1995. Changes of soil C and microbial biomass inwhole soil and aggregates in two cropping systems. Appl. Soil Ecol. 2,253–261.
Palma, R.M., Conti, M.E., 1990. Urease activity in Argentine soils: fieldstudies and influence of sample treatment. Soil Biol. Biochem. 22,105–108.
171V. Ralte et al. / Acta Oecologica 28 (2005) 163–172

Palma, R.M., Arrigo, N.M., Saubidet, M.E., Conti, M.E., 2000. Chemicaland biochemical properties as potential indicators of disturbances. Biol.Fertil. Soils 32, 381–384.
Taylor, L.A., Arthur, M.A., Yanai, R.D., 1999. Forest floor microbial biom-ass across a northern hardwood successional sequence. Soil Biol. Bio-chem. 31, 431–439.
Tiwari, S.C., Mishra, R.R., 1995. Seasonal variation in dehydrogenase andurease activities in hilly soils under grassland and forest. J. Indian Soc.Soil Sci. 43, 689–690.
Tiwari, S.C., Sorokhaibam, S.S., Dkhar, M.S., Mishra, R.R., 2002. Soildegradation affects dehydrogenase activity and microbial biomass C inhumid tropical forest soils of Arunachal Pradesh, North-Eastern India.Asian J. Microbiol. Biotechnol. Environ. Sci. 4, 143–148.
Vance, E.D., Brookes, P.C., Jenkinson, D.S., 1987. An extraction method formeasuring soil microbial biomass C. Soil Biol. Biochem. 19, 703–707.
von Lützow, M., Zelles, L., Scheunert, I., Ottow, J.C.G., 1992. Seasonaleffect of liming, irrigation and acid precipitation on microbial biomass Nin a spruce (Picea abies L.) forest soil. Biol. Fertil. Soils 13,130–134.
Witter, E., Kanal, A., 1998. Characteristics of soil microbial biomass in soilfrom a long term field experiment with different levels of C input. Appl.Soil Ecol. 10, 37–49.
Zar, J.H., 1974. Biostatistical Analysis, second ed. Prentice Hall, EnglewoodCliffs, NJ.
172 V. Ralte et al. / Acta Oecologica 28 (2005) 163–172




















