
Changes in ammonoid fauna and palaeoceanographic environment ...
163
Changes in ammonoid fauna and palaeoceanographic environment in the late Early Jurassic Northern Hemisphere Kentaro NAKADA Doctoral Program in Fundamental Science Graduate School of Science and Technology Niigata University
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Changes in ammonoid fauna and palaeoceanographic environment ...

Changes in ammonoid fauna and
palaeoceanographic environment in the late Early Jurassic
Northern Hemisphere
Kentaro NAKADA
Doctoral Program in Fundamental Science
Graduate School of Science and Technology
Niigata University

Abstract
The late Early Jurassic time-interval (about 187-175 Ma) is attracted by the Early
Toarcian Oceanic Anoxic Event (OAE), regarded as a global and dramatic
paleoenvironmental change. However the paleoceanographic environments during late Early
Jurassic have been analyzed vigorously in Europe, contributions from the Asian regions are
still limited. The lack ofthese data has disturbed the discussions of global paleoceanographic
changes in this time-interval. The aims of this study are to analyze the paleoceanographic
changes in the West Panthalassa (present East Asia) during late Early Jurassic based on the
geochemical analyses of the Early Toarcian OAE in Japan, the short-term
paleoenvironmental and faunal events, and the detailed transitional patterns of the ammonoid
paleobiogeography for tracing the long-term paleoceanographic change, and to discuss the
global paleoceanographic environment affected to the change of marine biota (ammonoids)
in the Northern Hemisphere.
In this study, the revision or the establishment of ammonoid zonal schemes in the Toyora
and the Kuruma Groups and the mutual/international correlation of these zonations are
discussed before analyzing the geochemistry and paleobiogeography because of the necessity
to propose the chronological standard. Six ammonoid zones in the Toyora Group, the Lower
to Middle Jurassic epicontinental deposits distributed in southwest Japan, and three zones in
the Kuruma Group, the Lower Jurassic continental shelf sediments exposed in central Japan,
are proposed in this study and are corresponded with the zonal schemes established outside
Japan, especially with the standard ammonoid zonation discussed in the Northwest European
province.
The transitional patterns of the Japanese ammonoid faunas are analyzed by utilizing the
assemblages from the Toyora and the Kuruma Groups. The late to middle Late Pliensbachian
ammonoid assemblage from Japan is associated with the Boreal elements and shows a high
similarity with the coeval Northeast Russian ammonoid fauna. However, in latest
Pliensbachian, a turnover from the Boreal fauna to the Tethyan fauna via the Tethyan-Boreal
mixed fauna is distinguished mainly in the assemblage from the Kuruma Group. This
turnover seems to be mainly affected by the coeval regression during latest Pliensbachian to
earliest Toarcian recognized in Europe, in addition to the starting of the warming event. In
contrast, a faunal mixing of the Tethyan and the Boreal faunas recognized in the middle Early
Toarcian assemblage is perhaps reflected by the abrupt transgression distinguished in this
time-interval. Consequently, the late Early Jurassic ammonoid transitions in the Northern
Hemisphere are closely related with the changes of eustatic sea level and seawater
temperature.

The concentrations of trace elements and the anomalies of REE are analyzed for
discussing the Early Toarcian GAE in the Lower Nishinakayama Formation of the Toyora
Group. The positive excursions of the V and Co concentrations are recognized in the middle
part of the P. palms Zone, corresponded to earliest Toarcian, with a negative shift of the Ni
concentration and a positive Ce anomaly. These geochemical anomalies suggest the
oxygen-depleted marine condition biostratigraphically corresponded to the coeval minor
anoxic environment in Europe. In contrast, the major Early Toarcian GAB recognized in
Europe and North American Cordillera is not identified in the Toyora Group. Therefore, the
effect of the major Early Toarcian GAE seemed to be limited only in the West Tethys and
East Panthalassa.
Moreover, a diversity fall of ammonoid assemblage is also identified during the
oxygen-depleted redox condition in earliest Toarcian. This diversity crisis is characterized by
the extinction of East Asian endemic Harpoceratinae, and the survival of only a species of
pandemic Arieticeratinae. Consequently, the ammonoid diversity crisis in the middle part of
the P. palms Zone is perhaps reflected by the anoxic event in East Asia. Thus, the
relationships between the marine redox conditions and the faunal associations of
Hildoceratidae probably suggest the difference of the anoxic tolerances between
Harpoceratinae and Arieticeratinae.

Contents
1. Introduction
1-1. Paleoenvironmental background of late Early Jurassic
1-1-1. General climatic changes in late Early Jurassic
1-1-2. Early Toarcian Oceanic Anoxic Event
1-2. Aims of this analysis
2. Geological framework
2-1. Toyora Group
2-2. Kuruma Group
p.l
p.7
3. Ammonoid biostratigraphy and its international correlation p.17
3-1. Aims of this chapter
3-2. Re-examination of ammonite biostratigraphy in the Toyora Group
3-2-1. Characteristics of materials
3-2-2. Methods
3-2-3. Stratigraphical repetition along the Sakuraguchidani Valley
3-2-4. Results
3-2-4-1. Amaltheus stokesi Assemblage Zone
3-2-4-2. Canavaria japonica Zone
3-2-4-3. Paltarpites paltus Zone
3-2-4-4. Dactylioceras helianthoides Zone
3-2-4-5. Harpoceras inouyei Zone
3-2-4-6. Pseudogrammoceras-Phlyseogrammoceras Assemblage Zone
3-2-5. Reexamination of the PIT boundary
3-2-6. Correlation with the previous zonation in the Toyora Group
3-3. Proposal of a new ammonoid zonation in the Kuruma Group
3-3-1. Characteristics of materials and methods
3-3-2. Differences between fossil-bearing rocks
3-3-3. Results
3-3-3-1. Stokesi-Repressus Assemblage Zone
3-3-3-2. Margaritatus-aff. Talrosei Assemblage Zone
3-3-3-3. Satoi Assemblage Zone
3-4. International correlation for the new ammonite zonations
3-4-1. Northwest European province
3-4-2. Mediterranean province

3-4-3. North American Cordillera province
3-4-4. South American province
3-4-5. Northeast Russia
4. Relationship between the ammonoid paleobiogeographic and the paleoceanographic
changes
p.53
4-1. Aims of this chapter
4-2. Paleobiogeographic changes of Japanese ammonoid assemblages
4-2-1. Faunal characters of the assemblages from the Kuruma Group
4-2-2. Faunal characters of the assemblages from the Toyora Group
4-2-3. Faunal changes of Japanese ammonoid assemblages
4-3. International correlation of Japanese ammonoid assemblages
4-3-1. Stokesi Standard Subzone
4-3-2. Subnodosus-Gibbosus Standard Subzones
4-3-3. Apyrenum Standard Subzone
4-3-4. Hawskerense Standard Subzone
4-3-5. Tenuicostatum Standard Zone
4-3-6. Serpentinum and Lower to Middle Bifrons Standard Zones
4-3-7. Fallaciosum Standard Subzone and Dispansum Standard Zone
4-4. Discussion of the paleoceanographic change in the Northern Hemisphere
4-4-1. Early Jurassic paleoenvironmental changes in the Northern Hemisphere
4-4-1-1. Sea level changes
4-4-1-2. Sea surface temperature changes
4-4-2. Paleoceanographic changes on the basis of the ammonoid faunal changes
4-4-2-1. Latest Pliensbachian faunal turnover
4-4-2-2. Early Toarcian faunal mixing
5. Geochemical analyses of the Early Toarcian OAE in East Asia p.83
5-1. Aims of this chapter
5-2. Materials and analytical methods
5-3. Results
5-3-1. Trace element concentrations
5-3-2. Rare Earth Element concentrations
5-4. Discussion
5-4-1. Redox conditions during the Lower Toarcian in the Toyora Group
5-4-2. Relationships between the anoxia and the ammonoid diversity changes
5-4-3. Early Toarcian OAE in the Northern Hemisphere

6. Systematic paleontology
6-1. Genus Amaltheus
7. Conclusions
7-1. Ammonoid biostratigraphy
7-2. Ammonoid paleobiogeography
7-3. Geochemical analyses of the Early Toarcian OAE
Acknowledgements
Reference
Plates
Appendices
p.99
p.109

1
1. Introduction
1-1. Paleoenvironmental background of late Early Jurassic
The Jurassic period (199.6±0.6-145.5±4.0 Ma, Gradstein et al., 2004; Fig. 1-1)
was an important time-interval in the studies of stratigraphy because some basic
frameworks, including "law of strata identified by fossils" and the biostratigraphic
units (biozone), were proposed in the European Jurassic successions (e. g. Smith,
1816-1819). Especially, late Early Jurassic time (equivalent to Late
Pliensbachian-Toarcian time-interval in this study; 187.0-175.6±2.0 Ma, Gradstein
et al., 2004; Fig. 1-1) was also characteristic in the beginning of the break-up of
Pangaea supercontinent and recently attracted by the dramatic paleoceanographic
events, represented by the Early Toarcian Oceanic Anoxic Event (OAE).
The analyses of the paleoceanographic environments during late Early Jurassic
have been discussed mainly in Europe. In contrast, the climate changes of East
Asia in this time-interval are not discussed sufficiently. In Japan, the existence of
the Early Toarcian OAE in the epicontinental depositional environment have been
suggested only in the Toyora Group on the basis of the characteristic macro faunas,
sedimentary structures and carbon isotope profile (Tanabe, 1991; Izumi & Tanabe,
2010), although the detailed distinctions of this phenomenon have not been
analyzed in this section. Moreover, the detailed faunal transitions of the late Early
Jurassic ammonoid assemblages have not been also analyzed in East Asia. The
paleobiogeographic faunal changes of ammonoid enable to trace the long term
paleoenvironmental changes with high biostratigraphic resolution. The absence of
these data in Eats Asia has been disturbed the global paleobiogeographic
discussion of ammonoid in late Early Jurassic time.
The geochemical data of the Early Toarcian OAE, as a proxy of short-term
marine environmental change, and the paleobiogeographic faunal transitions, as a
tracer of long-term climate changes, reveal the paleoceanographic changes in East
Asia during late Early Jurassic time-interval.

.7
.-- - - - - - - - - - 1 .0
,\,
,,,,,,,,,,
\,,\,,,,,,,,,,,,,,,,,,,,
\,,,,,\,,
\
C
__._IC
Fig . 1-1 . S Jura . iod . h a ut in t I. ( 2 ).
1-1-1. G ner I _ _~ of clim t d iot i I te rly Jur ic
Through the Jurassic period, the temperatures were generally warmer than
present and rose from Triassic toward the Cretaceous. On the basi s of the oxygen
isotope analyses, a cooling during Late Pliensbachian and an abrupt warming in
earliest Toarcian were recognized in late Early Jurassic time-interval (e.g. Bailey et

3
al., 2003). On the other hand, the eustatic sea level showed the higher stand than
present and was raised from Late Permian to Late Cretaceous (e.g. Haq et aI., 1988).
In late Early Jurassic, a regression during Late Pliensbachian and a transgression in
Early Toarcian were distinguishable on the basis of the Sr isotope ratio (e.g.
McArthur et al., 2000).
The Jurassic period was also characterized by the dramatic shifts of marine and
terrestrial biota, including the explosive development of ammonoids, the prosperity
of dinosaur and appearances of bony fishes and birds. The diversification of
ammonoids in this period enabled the high-resolution ammonoid biostratigraphy
and paleobiogeography.
1-1-2. Early Toarcian Oceanic Anoxic Event
The late Early Jurassic time has been attracted to the Early Toarcian OAE, the
global paleoenvironmental change proposed by Jenkyns (1988) on the basis of the
worldwide distribution of the organic black shale (Fig. 1-2).
The Oceanic Anoxic Event is the dramatic change of marine environment
characterized by the development of the oxygen-depleted water mass and has been
mainly recognized at the Permian/Triassic boundary (around 251Ma), at Early
Aptian (Early Cretaceous, 121-120 Ma) and at the Cenomanian/Turonian boundary
(Late Cretaceous, around 93.5Ma). The Early Toarcian OAE was one of the most
large-scale paleoceanographic changes through the Jurassic period, mainly
distinguished by the carbon and the oxygen isotopic anomalies (e.g. Hesselbo et al.,
2000). This phenomenon has been analyzed in detail mainly in Europe from various
viewpoints including lithostratigraphy, biostratigraphy, sedimentology and
chemostratigraphy (e.g. Jenkyns & Clayton, 1986, 1997; Jenkyns, 1988; Palfy &
Smith, 2000; Cecca & Macchioni, 2004). Palfy & Smith (2000) considered that this
OAE was caused by the global warming in earliest Toarcian, triggered by the
coeval Karoo-Ferrar flood basalt volcanism. On the other hand, paleontological
analyses have been carried out mainly on the relationship between the OAE and the
mass-extinction events, and the diversity crises of ammonoids, coeval with the

Fig. 1-2. Distribution of arly oarci n(1988) . Paleogeographic map is modiand Dera e al. (2009a).
nkyn(2007)
arly oarcian OA, er r c g d in ur p cca & Macchi i, 2 4.
In addition, the blac -colored ediment hav be n al
pelagic succession (e.g. ori et aI., 2000).
di tin ui hed in th
1-2. im Iy i
However the paleoceanographic environments during late arly Jurassic have
been analyzed vigorously in Europe, contributions from the Asian regions are still
limited . In Japan, the existence of the Early Toarcian OA in the epicontinental

5
successions were only discussed in the Nishinakayama Formation, the middle part
of the Toyora Group exposed in southwest Japan, by Tanabe (1991) and Izumi &
Tanabe (2010). This formation is known as the most major source of the late Early
Jurassic ammonoids in Japan. Thus, the ammonoid biostratigraphy has been
discussed in this formation (e.g. Hirano, 1973b, Nakada & Matsuoka, 2009).
According to the poor exposures of the Lower Jurassic epicontinental deposits in
East Asia, this area is regarded as a suitable section for analyzing the Early
Toarcian GAE in this region because of the distinguished biostratigraphic control
in theNishinakayama Formation. Tanabe (1991) suggested that the black
mudstones in the middle part of this formation (Nb Member; Tanabe, 1991) were
deposited under the oxygen-depleted bottom conditions and enabled to correspond
to the facies of the global Early Toarcian GAE on the basis of the sedimentological
and paleontological evidence. Izumi & Tanabe (2010) distinguished an abrupt
negative shift in the carbon isotope, fluctuation and suggested the effect of the
Early Toarcian GAE. However, the detailed stratigraphic position, scale and
mechanism of this phenomenon have not been analyzed in this section. The data of
the Early Toarcian GAE in East Asia enable to contribute the global-scale
discussion of this paleoenvironmental change in the Northern Hemisphere.
In contrast, the paleobiogeographic changes of the Jurassic ammonoids have
been discussed in a global scale (e.g. Page, 2008). The transitional pattern of
ammonoid fauna is an excellent proxy for tracing the long term paleoenvironmental
changes. In Japan, the detailed transitional pattern of late Early Jurassic ammonoid
assemblages have not been analyzed, and the lack of these data have disturbed the
discussions of global ammonoid paleobiogeography in this time-interval.
Ammonoid faunas from Japan, including the assemblages from the Toyora and the
Kuruma Groups, have been compared paleobiogeographically with the faunas from
outside Japan (Sato, 1956; Hirano, 1973b). However, the transitions of a
comprehensive Japanese ammonoid fauna during late Early Jurassic and their
paleobiogeographic significances have not been discussed in previous studies.
Consequently, the aims of this study are to analyze the paleoceanographic

6
changes in the West Panthalassa (present East Asia) during late Early Jurassic
time-interval based on the geochemical analyses of the Early Toarcian GAE in
Japan, the short-term paleoenvironmental and faunal extinction events, and the
detailed transitional patterns of the ammonoid paleobiogeography for tracing the
long-term paleoceanographic change, and to discuss the global paleoceanographic
environment affected to the change of marine biota (ammonoids) in the Northern
Hemisphere.
These paleoecological analyses should be discussed on the basis of the detailed
ammonite-based chronological schemes which are compared with the European
zonation. The zonal framework of the Toyora Group has been proposed by Hirano
(1973b), and three ammonite zones were established in the Nm Member (lower part
of the Nishinakayama Formation; Hirano, 1971): the Fontanelliceras fontanellense
Zone, the Protogrammoceras nipponicum Zone and the Dactylioceras helianthoides
Zone in ascending order. However, the necessity to refine the Hirano's zonation
was pointed out by Nakada & Matsuoka (2009) based on the difficulty of the
international correlation. In contrast, the ammonoid biostratigraphy of the Kuruma
Group was analyzed by Sato (1955). Although the Teradani Formation, the upper
part of the Kuruma Group, was corresponded to the Upper Pliensbachian, the
detailed depositional age of this formation has not been discussed because of the
poor ammonoid faunal association and the unidentified ammonoid-bearing horizon
in the previous study. Therefore, the revision or the establishment of ammonoid
zonal schemes in the Toyora and the Kuruma Groups and the mutual/international
correlation of these zonations should be discussed before analyzing the
geochemistry and paleobiogeography.

7
2. Geological framework
2-1. Toyora Group
The Toyora Group is the Lower to Middle Jurassic epicontinental sediments
exposed around the Tabe Basin in the western part of Yamaguchi Prefecture,
southwest Japan (Fig. 2-1). The distribution of the group is divided into the
northern district (Ishimachi district) and the southern district (Tabe district) by the
Kikugawa (Tabe) Fault. The Toyora Group exposed in the northern district yields
numerous ammonoid fossils (e.g. Matsumoto & Ono, 1947; Hirano, 1971, 1973a, b),
with bivalves, gastropods and echinoids (e.g. Hayami, 1958, 1959, 1960a, b, 1961,
1962). In contrast, the latter, where the lithofacies are similar to the former and
abundant plant fossils are obtained, are rare in marine biota (Hirano, 1971;
Kawamura, 2010). The study area includes the majority part of the north district,
3.5 km northwest-southeast (NW-SE) and 4.5 km northeast-southwest
(NE-SW)(Fig.2-1).
The lithostratigraphic classification of the Toyora Group follows Hirano's
study (1971). A geological map and a generalized geological column of the study
area are presented in Figs. 2-2 and 2-3, respectively. The group is divided into
three formations (in ascending order): the Higashinagano Formation, the
Nishinakayama Formation and the Utano Formation (Figs. 2-2, 2-3). Each
formation is subdivided into two to four members. The lithostratigraphic
denominations of these members are revised based on the International
Stratigraphic Guide (Salvador, 1994).
The Higashinagano Formation consists of mainly sandstone with conglomerate
and siltstone. It is subdivided into four members (in ascending order; Fig. 2-3); the
Chuzankei Conglomerate Member (basal conglomerate), the Higashinakayama
Sandstone Member (massive arkose medium to coarse sandstone), the Kido
Sandstone Member (bedded medium to fine sandstone with sandy siltstone and thin
coal beds) and the Koyagawa Sandy Mudstone Member (sandy siltstone). Some
ammonite specimens were obtained from the base of the Kido Sandstone Member

··· 1······
.i.'
:r"..'
r__------"--...., '.;.
".
Shira 0
)5 (
-,
'....~
Sea of Japan
o
.......
r
raz
2 m
~.:' \
.........'
o
o
Kuru a Gro
is io soGro is
oyora Group
. S di areaFi . 2-5.

9
(Hirano, 1971), and many bivalves and gastropods from this formation were
described by Hayami (1958, 1959, 1960a, b, 1961, 1962). The Higashinagano
Formation generally shows a fining upward sequence. The Chuzankei
Conglomerate Member covers the basement, the Permian Toyohigashi Group which
is composed of metamorphic sedimentary rock (merange), at an unconformity,
although the Higashinakayama Sandstone Member overlies the basement directly in
the Higashinagano area because the basal conglomerate pinches out to the north
(Fig. 2-2).
The Nishinakayama Formation, mainly analyzed in this study, is composed of
(in ascending order) the Sakuraguchidani Mudstone Member and the Ishimachi
Sandstone and Mudstone Member (Fig. 2-3). The Sakuraguchidani Mudstone
Member is mainly composed of silty clay and clay, often with parallel lamination
or bioturbation, and is characterized by abundant occurrence of ammonoid fossils.
Some thin sandstone (fine to medium .grain) and acidic tuff beds are alternated with
these black mudstones (Fig. 2-3). This member is well exposed along the
Sakuraguchidani Valley (Fig. 2-2). The Ishimachi Sandstone and Mudstone
Member is mainly consisted of black bioturbated siltstones with numerous
intercalates of fine to medium grained thin sandstones (Fig. 2-3). Ammonoid
fossils were obtained from three mudstone beds and a tuffaceous sandstone horizon
of this member exposed along the Sakuraguchidani Valley.
The Utano Formation consists mainly of alternating beds of sandstone and
sandy siltstone. This formation is subdivided into the Eragawa Sandy Mudstone
Member (well-laminated sandy siltstone), the Andadani Sandstone and Mudstone
Member (mudstone with many intercalates of sandstone), the Kodani Sandy
Mudstone Member (massive sandy siltstone), and the Kamiokaeda Sandstone and
Mudstone Member (sandstone with many intercalates of mudstone), in ascending
order (Fig. 2-3). The Utano Formation is generally composed of a coarsening
upward sequence and is overlain by the Cretaceous Kanmon Group, the
epicontinental sediment which is composed of coarse-grained clastic rocks, at an
angular unconformity.

,
131.03434.084N
,,
,,,
! 0 500 1,000 (m)
LEGE D -/
Higa hinag no F. nmon Group conformityKoy a a Sa m nt unconformity
c=J Kido M. Quartz porphyry Faultc=J Higa hina ayama M. Rhyolite Con al d fault" Chuzan ei Porphylit Inferred fault
It ~ Anticfin
/" Strike and dip Syncfin
Fig. 2-2 . Geological map of the study area in the Toyora Group .

11
t1y II
with
mo t part i compo
60125m
60 m+
60120m
60150m
50120m
oz ~ ~ I-__I- -t4:e4:zIen4:
"I
?
alenian
Sinemurian
Callo ian
Pre-Jurassic
(!)
0 Toarcian «CJ) a:CJ) aco >- rL- a::J., I- ~
IenZ
Pli ns-bac ian
D udstoneSandstoneAmmonoid fossils
o Calcareous nodule
D Sandy silt toneConglomerate
( Bivalve fossils-=- Coal beds
Fig. 2-3. Generalized geolog'cal column of the Toyora Group in the study area.

12
In the study area, the Toyora Group generally strikes NE-SW and dips
northwestward at 30-40° (Fig. 2-2). A NE-SW striking fault occurs near the Koya
River, and three NW-SE striking faults occur to offset the NE-SW striking fault. In
addition, two fold structures are observed: an anticline in the Ishimachi area and a
syncline across the Utano River (Fig. 2-2).
2-2. Kuruma Group
The Kuruma Group, the Early Jurassic continental shelf sediments, is exposed
around the northeastern part of the circum-Hida terrane in the eastern part of
Toyama Prefecture, the western part of Niigata Prefecture and the northern part of
Nagano Prefecture, central Japan (Fig. 2-1). The distribution of this group is
divided into the main area and the eastern area (Kumazaki & Kojima, 1998)(Fig.
2-1), and lithostratigraphy has been analyzed separately by Kobayashi et aI. (1957),
Chihara et aI. (1979), Takizawa (1984), Shiraishi (1992) and Kumazaki & Kojima
(1998)(Fig. 2-4). The ammonoid biostratigraphy in this study is analyzed along the
Daira River and the Teradani Valley, located at the western part of the main area in
Toyama Prefecture (Figs. 2-1, 2-5), because this section yields abundant Early
Jurassic ammonoid fossils (Sato, 1955; Kobayashi et aI., 1957).
The Kuruma Group exposed in the main area was divided into seven formations
by Kobayashi et al. (1957) in ascending order: the Jogodani, the Kitamatadani, the
Negoya, the Teradani, the Shinatani, the Otakidani and the Mizukamidani
Formations (Fig. 2-4). Thereafter, the Mizukamidani Formation was included into
the Tetori Group, the Middle Jurassic to Early Cretaceous continental shelf
sediments distributed in central Japan, by Takizawa (1984). This group is mainly
composed of sandstones, mudstones and conglomerates with numerous fossils, for
example, ammonoids, bivalves, gastropods, belemnites and plants, and is
characterized by alternation of the marine and non-marine sediments (Kobayashi et
aI., 1957). The Kuruma Group covers serpentinite melange and olistostrome of the
circum-Hida terrane and is mainly overlain by the Tetori Group, including the
Mizukamidani Formation, the Kurobishiyama Formation and the Siritakayama

Kobayashi at at (1957) Takizawa (1984) Shiraishi (1992) Kumazaki & Kojima (1996)main area main area eastern area main area eastern area
Mizukami- Km3Km2
danl F.Km1 ...... ..... -- ..... ---- -- ...... ",. ~ .... ......
Otakidani F. Otakidani F. Otakidani F.
Shinatani F.Ks2
Shinatani F. Shinatani F.Ks1
Teradani F. Teradani F. Teradani F.
Kn2
Negoya F. Negoya F. Negoya F.
Kn1
?Kk3 Yoshina ...
Kitamata- Yoshina- Kitamata-zawa F.
Kk2 Kitamata-dani F. dani F. zawa F. dant F.
Odokoro-Kk1 Odokoro- gawa F.
Kj.2gawa F.
Jogodani F. Jogodani F. Gamahara- Jogodani F. Gamahara-Kj1 zawa F. zawa F.
Fig. 2...4. Different views on the stratigraphic sequence of the Kuruma Group.

37.7073 ° 2
FO
. 2-5. I a &:! "' ;~In" .oJIlUr"-AI";.u ro es a d fa . locatrues

15
Formation, at an unconformity (Kobayashi et aI., 1957; Takizawa, 1984; Kumazaki
& Kojima, 1996).
In this study, the lithostratigraphic classification of the Kuruma Group follows
Kobayashi et al. (1957) and Takizawa (1984). Individual columnar sections of the
Kuruma Group in study area are presented in Fig. 2-6. The Kuruma Group
distributed in the study area are divided into four formations (Fig. 2-6), the Negoya
Formation (alternating medium to fine grained sandstone and mudstone with some
conglomerate or fine grained tuff intercalations), the Teradani Formation (sandy
siltstone with some fine grained sandstone beds), the Shinatani Formation (fine
grained sandstones with numerous sandy siltstones and some conglomerates) and
the Otakidani Formation (black mudstone with thin sandstone and conglomerate
intercalations) in ascending order. These formations exhibit lateral changes in
lithofacies, and the Teradani Formation pinches out to the west (Fig. 2-6). In the
study area, the Kuruma Group generally strikes W-E and dips northward at 25-50°.
A W-E striking fault occurs along the Daira River, and many NW-SE striking faults
occur to offset the W-E striking fault (Kobayashi et aI., 1957; Kumazaki & Kojima,
1996).
Ammonoid fossils have been obtained from the Teradani Formation (Sato,
1955) and the Otakidani Formation (Kobayashi et al., 1957). Most of the ammonoid
specimens analyzed in this study were yielded in the former formation. The
Teradani Formation is successively exposed along the Teradani Valley, the type
area of this formation, which mainly composed of black sandy mudstone without
sedimentary structure and bioturbation (Fig. 2-6). On the other hand, this formation
distributed along the Daira River is composed of alternating sandy mudstone and
fine to medium grained sandstone and is divided into some tectonic blocks by many
faults (Kobayashi et al., 1957; Kumazaki & Kojima, 1996). Abundant marine biota,
including ammonoids, bivalves, gastropods and echinoids, and plants are obtained
from this formation in this study.

Daira River 1
1
Te adani Vall y
congkHllerale
I'lIl 'Fft1i ft2
- - - - _.. '
-,: 100m,,IIIII1III
--I,: Daira Ri er 2II,I Daira RO r 3: Daira i rL ----------------------------
u.:cco~~
co(5
u.:cco"0coL-
~ca
:;::coEL-
au,co~aC)Q)
Z
u.:cco.....,coc
..c(f)
~(':tinr,~ co po de ee .... lIrT1 .....~rr 'esa e saFig. 2-6. Individual columnar sectio s of e ruto the fossil locality numbers in Fi s. 3 , 3-5.

17
3. Ammonoid biostratigraphy and its international correlation
3-1. Aims of this chapter
The ammonoid biostratigraphy in the Toyora Group has been discussed by
Matsumoto & Ono (1947), Hirano (1973b), Tanabe (1991) and Nakada & Matsuoka
(2009). The zonal framework of the Toyora Group was mainly established by
Hirano (1973b) and three ammonite zones were mentioned in the Nm Member
(lower part of the Nishinakayama Formation; Hirano, 1971). On the other hand,
Nakada & Matsuoka (2009) drew the Pliensbachian/Toarcian (P/T) boundary in the
Nishinakayama Formation and pointed out the necessity of refining the Hirano's
zonation.
In the Kuruma Group, although the Teradani Formation was compared roughly
to the Upper Pliensbachian based on the·· occurrences of Amaltheus sp. and
Canavaria sp. ex gr. C. geyeriana (HAAS) by Sato (1955), the detailed ammonoid
biostratigraphy has not been analyzed in the Kuruma Group because of their poor
faunal association. The occurrence of abundant ammonoid materials in this study
enables to propose the ammonoid zonal framework in the Teradani Formation.
This study aims to revise the ammonite zonal scheme of the Toyora Group, to
establish an ammonoid zonal scheme in the Kuruma Group and to correlate these
new frameworks with the zonations established outside Japan.
3-2. Re-examination of ammonite biostratigraphy in the Toyora Group
3-2-1. Characteristics of materials
Ammonoid biostratigraphic work of the Toyora Group was mainly carried out
along the Sakuraguchidani Valley, one of the tributary streams of the Koya River
(Fig. 2-2), because of the outcrop condition and abundance of ammonoid fossils. In
addition, in previous studies, ammonoid biostratigraphical analyses have been
mainly carried out along this route. A total of 304 ammonoid specimens (276
specimens from 48 horizons of the Sakuraguchidani Mudstone Member and 28
specimens from four horizons of the Ishimachi Sandstone and Mudstone Member)

18
obtained along the Sakuraguchidani Valley are utilized in this study, in addition to
a specimen from the Kido Sandstone Member described by Hirano (1971)(Fig. 3 in
Plate 4). Twenty two identified genera and 32 identified species are shown in
Appendices 3-1, 3-2, 3-3, 3-4 and Plates 1-4 and 8. Almost all of the specimens
from the Sakuraguchidani Mudstone Member were embedded and compressed
horizontally to the bedding plane but the ornaments were well preserved.
The faunal characteristics of the ammonoid assemblages change with
stratigraphic levels in the Sakuraguchidani Mudstone Member. The lower part of
the member is not as abundant in ammonoid fossils. The fauna from the middle part
is characterized by the abundance of Harpoceratinae, especially Protogrammoceras
nipponicum (MATSUMOTO)(Figs. 1-3 in Plate 2). In the upper part, the diversity
reaches its maximum level, and this assemblage is identified by the abundance of
"Cleviceras" chrysanthemum (YOKOYAMA)(Figs. 1-3 in Plate 3) and Dactylioceras
helianthoides (YOKOYAMA)(Figs. 1-3 in Plate 1). Except for the Eoderocerataceae
genera (Dactylioceras, Peronoceras and Amaltheus), the fauna from the upper part
of this member consists mostly of high-diversity Hildoceratoidea genera. These
specimens indicate successive ages from Late Pliensbachian to Early Toarcian
(Hirano, 1973b). On the other hand, most of the materials from the Ishimachi
Sandstone and Mudstone Member were obtained from bed 53-4 which consists of
tuffaceous fine grained sandstone. This assemblage is mainly associated with
Grammoceratinae (genera Grammoceras and Pseudogrammoceras) and the genus
Osperlioceras with some Phylloceratina (Plate 8; Appendices 3-4).
3-2-2. Methods
This study shows the necessity of recognizing ammonoid-bearing levels in
detail for establishing a new zonation, which can be compared to higher-resolution
biostratigraphic frameworks proposed outside Japan. The mudstones of the
Sakuraguchidani Mudstone Member exposed along the Sakuraguchidani Valley
were subdivided into a few meters thick layers on the basis of the following
indices: particle size, laminated or bioturbated, fissility, fragments of fossil woods

19
and calcareous nodules. A detailed route map and corresponding lithological
columnar sections of the valley are shown in Fig. 3-1.
Biostratigraphical analysis was based on the range charts of the
Sakuraguchidani Valley. The valley branches into the North Valley and the South
Valley. The analyses were carried out separately (Figs. 3-2, 3-3).
3-2-3. Stratigraphical repetition along the Sakuraguchidani Valley
As mentioned above, the previous ammonoid biostratigraphical analysis has
been mainly carried out along the Sakuraguchidani Valley (Fig. 2-2). This route has
been considered to have no repetition of strata but this analysis proposes a new
interpretation (Figs. 3-1, 3-2, 3-3). Around the fork of the North and the South
Valleys, a specimen of the genus Dactylioceras, the index taxa of the Lower
Toarcian, was obtained from bed 14-1 in the North Valley (D. helianthoides; Figs.
1-3 in Plate 1), corresponding with the lowest occurrence of this species. Therefore,
this horizon was regarded as the lower Toarcian by Nakada & Matsuoka (2009). In
contrast, a specimen of Amaltheus margaritatus MONTFORT was found in bed 36-1
along the South Valley (Figs. 3-1, 3-3) in our additional examination after the
working by Nakada & Matsuoka (2009). This species has been obtained from the
Margaritatus Standard Zone to the Apyrenum Standard Subzone, the Upper
Pliensbachian except for the uppermost part of the Northwest European zonation
(e.g. Meister, 1988). Based on the lithological correlation between these two
sections, the latter is obtained from a horizon stratigraphically slightly higher than
the former. This biostratigraphical disagreement suggests the repetition of strata by
an inferred fault that exists between these beds (Figs. 3-1, 3-2, 3-3). The
occurrence of Petranoceras sp. aff. P. rinaldinii VENTURI (Fig. 9 in Plate 2) from
bed 10-6 supports this tectonic interpretation because P. rinaldinii was described in
the Mirabile Zone (Lowermost Toarcian) of central Italy (Faraoni et aI., 1994). The
ammonoid biostratigraphy should be discussed separately above bed 17-5 and
below bed 14-1 along the North Valley, as well as above bed 36-1 and below bed
10-6 along the South Valley.

(a)
II
10 m
-: HlGllSH1NAC3ANO...... I I
n t c l lTCW"'t Fa
ISH I KAY. Fo on WSAKURAGUCHIDA I ~
ud 0 e em r
.......
............'CD. heliclzlIlthcfides
// " .:···············C__··~
ISouth Vall yl
..'
............
! I
.../ ..

21
3-2-4. Results
Four ammonoid zones in the Sakuraguchidani Mudstone Member in the Toyora
Group are proposed in this analysis as follows, in ascending order (Figs. 3-1, 3-2,
3-3): the Canavaria japonica Zone, the Paltarpites paltus Zone, the Dactylioceras
helianthoides Zone and the Harpoceras inouyei Zone. In addition, two ammonoid
assemblage zones are also established in this study as follows: the Amaltheus
stokesi Assemblage Zone In the Higashinagano Formation and the
Pseudogrammoceras-Phlyseogrammoceas Assemblage Zone in the Ishimachi
Sandstone and Mudstone Member (Figs. 3-1, 3-2).
3-2-4-1. Amaltheus stokesi Assemblage Zone
This zone is represented by Amaltheus stokesi (SOWERBY)(Fig. 4 in Plate 4),
widely distributed in the Northwest European, the North American Cordillera and
the Russian provinces and regarded as one of the typical Boreal elements in this
period (e.g. Meister, 1989).
This interval is characterized by two specimens (GK. G. 11293 and GK. G.
11292) described by Hirano (1971) as Amaltheus sp. cf. A. stokesi (SOWERBY) and
Arieticeras sp. aff. A. apertum MONESTIER. According to Hirano (1971), they
obtained from almost same horizon of the Kido Sandstone Member, the
Higashinagano Formation, exposed along the Chuzankei Valley (Fig. 2-2). The
occurrences of these two species are limited in this zone.
3-2-4-2. Canavaria japonica Zone
This zone is represented by Canavaria japonica (MATSUMOTO)(Fig. 8 in Plate
1). The base of this zone has not been observed along this route by the inferred
fault. In this study, the lowermost part of this zone is characterized by the
Occurrence of A. margaritatus (Figs. 9-10 in Plate 1), one of the most important
index taxa of Late Pliensbachian, from bed. 36-1 along the South Valley, and this
zone is not recognized along the North Valley because of the absence of outcrops.
In addition to the genus Amaltheus, this interval is characterized by the abundance

o
?
o
a.:
Fg. 3-2. Columnar section and biostratigraphic succession of ammonoid along the North Valleyof t e Sakuraguchidani Valley. The numbers beside the columnar section correspond to thenumber of the outcrop in Fig. 3-1. Thin broken lines: re-examined zonal boundaries . Thickbro en line: prensbachianIToarcian (PIT) boundary. The previous ammonite zonation followsHirano (1973b) and Tanabe (1991). The legend of the columnar section is as in Fig. 3-1.

23
of the index species from bed 36-2 immediately above the Amaltheus bed along the
South Valley. The genus Canavaria is common in the Mediterranean province, and
A. margaritatus is dominant in the Northwest European province in the Upper
Pliensbachian. Moreover, both of these two genera are limited to this zone.
3-2-4-3. Paltarpites paltus Zone
This zone is represented by Paltarpites paltus (BUCKMAN)(Fig. 8 in Plate 2),
also described in the Northwest European province (Howarth, 1957) and North
American Cordillera (Frebold, 1970). The base of the P. paltus Zone is defined by
the first occurrence of the index species and is drawn at bed 41-1 along the South
Valley (Fig. 3-3). In contrast, this zonal boundary seems to be drawn below bed
17-5 along the North Valley based on the stratigraphic correlation between the
North and the South Valleys of the Sakuraguchidani Valley (Figs. 3-1,3-2,3-3).
This interval is stratigraphically divided into two parts depending on the faunal
characters. The lower part is represented by low-diversity Hildoceratidae with rare
Lytoceratina. More precisely, the occurrence of ammonoid fossils is generally rare
in the lower part of the P. paltus Zone except for some diverse fauna including P.
paltus (Fig. 8 in Plate 2), Paltarpites toyoranus (MATSUMOTO)(Fig. 7 in Plate 2),
Fontanelliceras fontanellense (GEMELLARO)(Figs. 6-7 in Plate 1), Lytoceras sp.,
Lioceratoides aradasi (FUCINI)(Fig. 10 in Plate 2), and Lioceratoides yokoyamai
(MATSUMOTO) from bed 41-1 along the South Valley, and the occurrence of F.
fontanellense (Figs. 6-7 in Plate 1), which is a pandemic taxon distributed mainly
in Europe and ranges from the middle to the top of the lower part of this zone, from
bed 18-1a along the North Valley. Almost all taxa are common in the Mediterranean
province, except for the genus Tiltoniceras, and are limited to the lower part of the
P. paltus Zone except for L. yokoyamai, which extends into the H. inouyei Zone.
The upper part of theP. paltus Zone is characterized by high-diversity endemic
species of Harpoceratinae with some Lytoceratina and Phylloceratina. This part
almost corresponds to the range of P. nipponicum (Figs. 1-3 in Plate 1), the most
dominant species in the upper part of the P. paltus Zone. The lower half of this

oQ.c:3:J
enCD~
»33o:Joa:-oen~,
en
P.
,.,.tptpNt.I
I
~
La
roro
nt8\dense
-10.
o
N
HIGASHINAGANOKOYAGAW
c: r~;:;:
I I I ... =ren 0
en@~
(1)
CJ)
fI)
Q)
5
'<
c
0 '(ii ' fI)
Q)
o
0"-t\ - ,..... (0=r '(1)(,.)
I(,.)
to

25
upper part is composed of alternating sandstone and mudstone and is poor m
ammonoid fossils. Only two specimens of P. nipponicum are obtained from bed
18-1c, corresponding to the first occurrence of this species. In contrast, the abrupt
diversification of Harpoceratinae is recognized in bed 19-1b, corresponding to the
upper half of the upper part, along the North Valley. The fauna from this bed is
characterized by the abundance of P. nipponicum and the appearance of many
Harpoceratinae species, including Protogrammoceras yabei HIRANO (Fig. 5 in
Plate 2):> Fuciniceras nakayamense (MATSUMOTO)(Fig. 6 in Plate 2):> Fuciniceras
primordium (MATSUMOTO):> Polyplectus okadai (YOKOYAMA)(Fig. 6 in Plate 3):> and
"Cleviceras" chrysanthemum (Figs. 1-3 in Plate 3):> with Lytoceras sp. In addition
to the North Valley, L. yokoyamai, P. nipponicum (Figs. 1-3 in Plate 2) and
Calliphylloceras sp. from bed 48-2:> and "Cleviceras" chrysanthemum from bed
48-3 are recognized in the upper part of the P. paltus Zone along the South Valley.
Almost all species obtained from this interval extend into the superj acent D.
helianthoides Zone except for P. nipponicum and F primordium.
3-2-4-4. Dactylioceras helianthoides Zone
This zone is represented by D. helianthoides (Figs. 1-3 in Plate 1):> also
described in Germany and Spain (Schmidt-Effing, 1972). The base of the D.
helianthoides Zone is marked by the first occurrence of the index species and is
drawn at bed 19-5b along the North Valley and bed 48-4 along the South Valley.
This interval is characterized by the abundance of D. helianthoides (Figs. 1-3
in Plate 1) and high-diversity Harpoceratinae, and contains a large number of
"Cleviceras" chrysanthemum (Figs. 1-3 in Plate 3):> F nakayamense (Fig. 6 in Plate
2) and the index species, with some specimens of L. yokoyamai, P. okadai (Fig. 6 in
Plate 3), Protogrammoceras onoi HIRANO (Fig. 4 in Plate 2), P.yabei (Fig. 5 in
Plate 2), Lioceratoides matsumotoi HIRANO, Hildaites sp. (Fig. 7 in Plate 3),
Calliphylloceras sp. and Lytoceras sp. All these species extend into the superjacent
H. inouyei Zone, with the exception of the genus Protogrammoceras, which
disappears at bed 21-1 within this zone. The uppermost part of the D. helianthoides

26
Zone along the South Valley has not been observed because the distribution of the
outcrops along this route is limited at bed 48-9.
3-2-4-5. Harpoceras inouyei Zone
This zone is represented by Harpoceras inouyei (YOKOYAMA)(Fig. 4 in Plate 3),
the endemic species of the Toyora Group, and almost corresponds to the range of
the index species (Fig. 3-2). The base of the H. inouyei Zone is defined by the first
occurrence of the index species and is drawn at bed 23-1 along the North Valley,
synchronous with that of Cleviceras sp. cf. C. exaratum (YOUNG & BIRD)(Fig. 5 in
Plate 3). The upper boundary of this zone is difficult to define because of the
absence of proper index taxa, for example the genus Hildoceras.
This interval is characterized by diverse genera of Harpoceratinae and
Dactylioceratidae, which includes the genera Cleviceras, Harpoceras,
Lioceratoides, Polyplectus, Fuciniceras, Dactylioceras and Peronoceras, with
some Calliphylloceras and Lytoceras. The fauna of this interval is dominated by
"Cleviceras" chrysanthemum (Figs. 1-3 in Plate 3) and Peronoceras subfibulatum
(YOKOYAMA)(Fig. 5 in Plate 1) which appears at bed 25-3 in the H. inouyei Zone.
Lioceratoides yokoyamai, Calliphylloceras sp., Lytoceras sp., L. matsumotoi and P.
subfibulatum (Fig. 5 in Plate 1) disappear gradually from the middle (bed 26-4) to
upper part (bed 28-4) of this zone. The fauna obtained from the uppermost part of
the H. inouyei Zone at bed 29-3 along the North Valley consists of D. helianthoides
(Figs. 1-3 in Plate 1), P. okadai (Fig. 6 in Plate 3),F. nakayamense (Fig. 6 in Plate
2), "Cleviceras" chrysanthemum (Figs. 1-3 in Plate 3), C. sp. cf. C. exaratum (Fig.
5 in Plate 3), and H. inouyei (Fig. 2 in Plate 3).
3-2-4-6. Pseudogrammoceras-Phlyseogrammoceras Assemblage Zone
This zone IS represented by the genera Pseudogrammoceras and
Phlyseogrammoceras (Figs. 10-19 in Plate 8) which are especially dominant in
central Europe (Guex, 1975).
This interval is characterized by the occurrence of Harpoceratinae (genus

27
Osperlioceras), Grammoceratinae (genera Grammoceras, Pseudogrammoceras and
Phlyseogrammoceras) and Phylloceratina (genus Calliphylloceras). A total of eight
species are recognized as follows; Osperlioceras sp. A (Figs. 1-4 in Plate 8),
Osperlioceras sp. B (Figs. 5-7 in Plate 8), Pseudogrammoceras sp. A (Figs. 10-12
in Plate 8), Pseudogrammoceras sp. B (Figs. 13-14 in Plate 8), Grammoceras sp.
(Figs. 8-9 in Plate 8), Phlyseogrammoceras sp. (Fig. 19 in Plate 8), Dactylioceras
sp. (Figs. 20-21 in Plate 8) and Calliphylloceras sp. (Figs. 22-23 in Plate 8).
Almost all species are obtained from bed 53-4 only, except for Osperlioceras sp. A
which are recognized from beds 53-2, 53-3, 53-4 and 56-2 (Fig. 3-2).
3-2-5. Reexamination of the PIT boundary
The ammonoid zonation in the Toyora Group has been documented mainly in
the Sakuraguchidani Mudstone Member by Hirano (1973b), who established three
ammonite zones, in ascending order: the Fontanelliceras fontanellense Zone, the
Protogrammoceras nipponicum Zone and the Dactylioceras helianthoides Zone.
Nakada & Matsuoka (2009) recently reviewed the PIT boundary by adopting the
European index, the first occurrence of the genus Dactylioceras. It was drawn by
the first occurrence of D. helianthoides (Figs. 1-3 in Plate 1) within the F.
fontanellense Zone of Hirano (1973b). However, this index species was identified
as the subgenus Dactylioceras (Orthodactylites), and the first occurrence of this
species seemed to be correlated to a horizon within the Clevelandicum Standard
Subzone, the second subzone of the Toarcian stage, by Schmidt-Effing (1972). In
contrast, Paltarpites paltus (BUCKMAN)(Fig. 8 in Plate 2) obtained from this group
characterizes the first subzone of the Toarcian and has been utilized as an index
species of the PIT boundary in France (Gabilly, 1976; Elmi et aI., 1997).
Consequently, the first occurrence of P. paltus is the most suitable candidate for
indicating this stage boundary in the Toyora Group. Nakada & Matsuoka (2009)
pointed out problems of the previous studies as follows: the disagreement between
the zonal boundaries of Hirano (1973b) and the PIT boundary of Nakada &
Matsuoka (2009) and the zonal nomenclature of the D. helianthoides Zone. This

28
biostratigraphic study should be discussed for taking these Issues into
consideration.
The PIT boundary must be revised to consider this stratigraphic repetition. It is
marked at bed 41-1 in the South Valley above the fault by the first occurrence of P.
paltus and is revised to a higher stratigraphic level than that of Nakada & Matsuoka
(2009)(Figs. 3-1, 3-2, 3-3). In contrast, the interval below the fault is compared to
the Lower Toarcian based on the occurrence of some indicators such as P. paltus,
Dactylioceras and Petranoceras, and consequently has no stage boundary.
3-2-6. Correlation with the previous zonation in the Toyora Group
As mentioned above, three ammonite zones- the F. fontanellense Zone, the P.
nipponicum Zone and the D. helianthoides Zone- were established by Hirano
(1973b) in the Toyora Group. According to Hirano (1973b), the lower boundary of
the F. fontanellense Zone was marked by the appearance of Amaltheus, Arieticeras,
Canavaria and/or Dactylioceras, and the base of the P. nipponicum Zone was
probably defined by the first occurrence of the index species, drawn immediately
below the first occurrence of P. nipponicum in the range chart (Hirano, 1973b;
Table 1). The upper limit of the P. nipponicum Zone, equal to the basement of the D.
helianthoides Zone, was described immediately above the last occurrence of the
genera Protogrammoceras and Fuciniceras and was drawn immediately above the
last occurrence of P. yabei (Hirano, 1973b; Table 1). The indicator of this boundary
was revised by Tanabe (1991) to the last occurrence of the genus
Protogrammoceras, because the range of F. nakayamense (Fig. 6 in Plate 2)
extended to the higher stratigraphic level. The upper boundary of the D.
helianthoides Zone was drawn just below the Ishimachi Sandstone and Mudstone
Member (Na Member in Hirano, 1973b), characterized by the last occurrence of P.
subfibulatum in Table 1 (Hirano, 1973b).
Results of the correlation of the ammonite zonal scheme established in this
study with the previous ammonite biostratigraphic analyses in the Toyora Group
are presented in Figs. 4-2 and 4-3. The C. j aponica Zone and the lower part of the P.

29
paltus Zone nearly correspond with the F. fontanellense Zone by Hirano (1973 b),
and the upper part of the P. paltus Zone is compared with the lower half of the P.
nipponicum Zone by Hirano (1973b). The D. helianthoides Zone established in this
study corresponds to the upper half of the P. nipponicum Zone and lowermost part
of the D. helianthoides Zone proposed by Hirano (1973b). The H. inouyei Zone in
this study is nearly corresponds to the D. helianthoides Zone proposed by Hirano
(1973b).
The new zonal framework is established to take international correlation into
account. Therefore, the zonation is more suitable internationally compared to
previous studies (Hirano, 1973b; Tanabe, 1991), especially in the Toarcian.
Moreover, the definitions of the D. helianthoides Zone in this study differ from the
previous study, while these zonal names are the same. The D. helianthoides Zone in
this study is proposed to solve the problem about biostratigraphical nomenclature
of the previous D. helianthoides Zone as pointed out by Nakada & Matsuoka
(2009).
3-3. Proposal of a new ammonoid zonation in the Kuruma Group
3-3-1. Characteristics of materials and methods
Ammonoid biostratigraphic work of the Kuruma Group was carried out along
the Daira River and the Teradani Valley, one of the tributary streams of the Daira
River (Fig. 2-5), because the Teradani Formation is well distributed along these
routes (see Fig. 2 in Kumazaki & Kojima, 1996). A total of 90 ammonoid
specimens obtained along these routes are utilized in this study. Identified eight
genera and 16 species are shown in Appendix 3-5 and Plates 4-7. Although 36
materials are recognized from four beds exposed along the Teradani Valley and
eight beds along the Daira River (Figs. 2-6, 3-4, 3-5, Appendix 3-5), the others are
found from the floated rocks which are provided from Daira 6 and 7 sections (Fig.
3-4; Appendix 3-5). Almost all of the specimens were compressed horizontally to
the median section, but some well preserved specimens were not affected the
compaction (Figs. 3-4 in Plate 5).
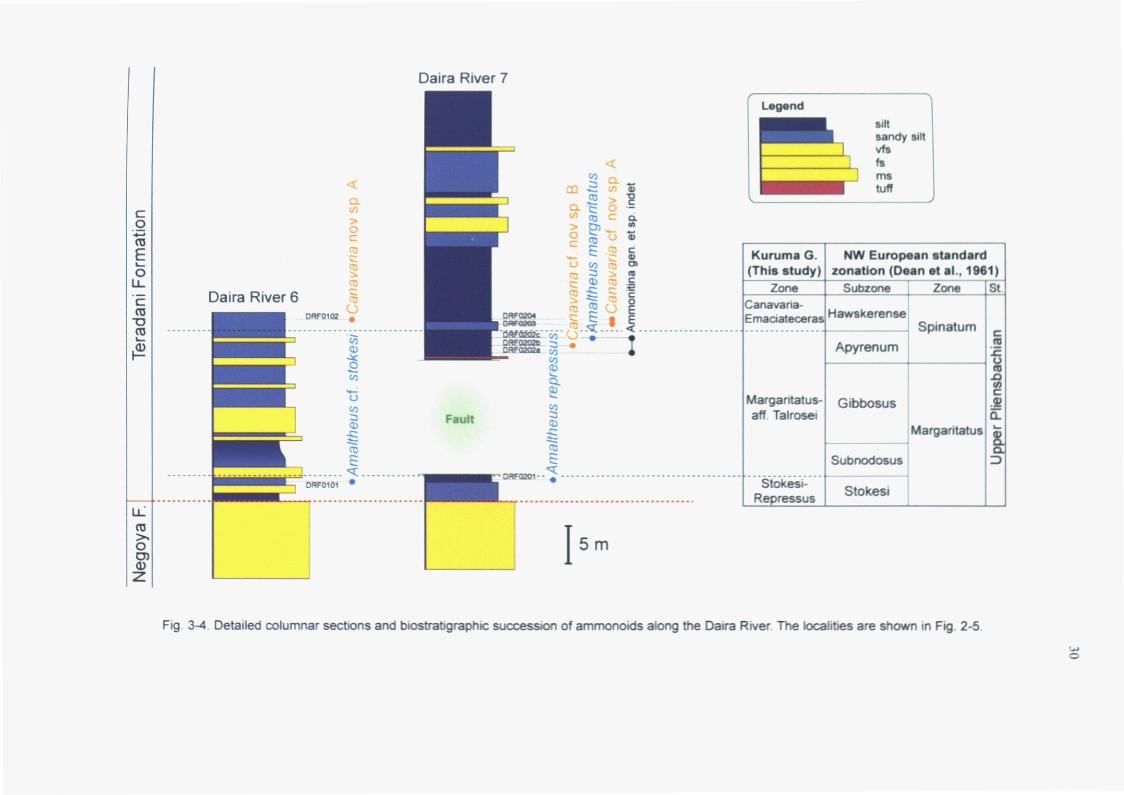
Daira River 7
mu
CI)
.2~
"2ci.en
E c
c~
~ ·coE ~ Emacia r
- ~ • • - • . <{ ._ •••••• -.- •• _------ ----------------+--------1
5
-~F
Da ira River 6
.................111111~---
co
+JroE~
oLL
cro'Uro~
~
u,ro~
o0)Q)Z
Fig. 3-4. Detailed colum a ec s a rnrnonoio s a e locaJitJes a e Fi . 2-5.

31
In this study, ammonoid fossils are obtained from the lower and the upper parts
of the Teradani Formation (Figs. 2-6, 3-5). The fauna from the lower part is
characterized by the occurrence of Harpoceratinae, including the genera Canavaria
and Emaciaticeras, and the genus Amaltheus, which is compared to the fauna
described by Sato (1955). Moreover, these genera indicate the Late Pliensbachian
(Sato, 1955). In contrast, the assemblage from the upper part of this formation is
associated with Fuciniceras nakayamense (MATSUMOTO), Protogrammoceras sp.
and Dactylioceras sp. This faunal association has a high similarity with that from
the Toyora Group. On the basis of the faunal characters, this assemblage enables to
compare chronologically to the Early Toarcian, although more detailed correlations
are difficult because of the impossibility of specific identification. In this study,
the ammonoid biostratigraphy of the Teradani Formation is discussed by using the
fauna from the lower part of this formation.
3-3-2. Relationship between fossil-bearing rocks and included ammonite taxa
Biostratigraphical analysis was based on the range charts of the Daira River
and the Teradani Valley (Figs. 3-4, 3-5). As mentioned above, most of the fossil
specimens are obtained from the floated rocks provided from Daira 6 and 7 sections
(Fig. 3-4; Appendix 3-5). The relationships between the ammonite-bearing clastic
rocks and the included taxa are distinguishable in the floated collections from
Daira 6 section.
The specimens identified as the genus Amaltheus are mainly obtained from
well-sorted mudstone with abundant pyrite. In Daira 6 section, the lowermost part
of the Teradani Formation, characterized by the occurrence of A. cf. stokesi (Fig. 6
in Plate 4) from bed DRFO101, is composed of black mudstone with sandy siltstone
and fine-grained sandstone (Fig. 3-4). In contrast, most of the specimens of the
genera Canavaria or Emaciaticeras (both are included in Arieticeratinae) are found
from the poorly-sorted sandy mudstone or tuffceous fine-grained sandstone blocks
with the fragments of plant fossils. This kind of sandy mudstone is mainly
recognizable in the upper part of Daira 6 section, including bed DRFO102 which

·~s:en
Margaritatu
Stokesi
Gibbo us
Subnodos u
..-Stokes,:..Re ressus
Margaritatuaft. Talrosei
J:Qf............ ... 3(I)
a. a.en enc CI) OJ
E mOJ a. C
CU 0g g en ECU -:.:::: E OJ ECU o E C «cCI) CU m OJ
~a 8>
C>OJQ) - ro
.~ 0 c:
.S a.. ~roo ~
u.. OJ
8-~
ro:::c
~cu"'Sc~Q.(I)Q)
1::W__.I.Of .Q:L . ~ . . . ..., . ..,.. .. Haw k r n.. ·tD~02··'~. . . 'Co' .,... . 3 Em ci ti r............... ... .. .c: .. « ci 'Ci) ci __ . - c
Q. . en c en ro~ a. CU Q> OJ Apyrenum :c;:) en ' t::: E ~ I--- - - -+- - - ----l U
..., >CUQ>c ro.TQEOj... 0 ~ ' - 0 .0
c: CU (I) E enCU c:: ~' t::: CU o E~O (1)«CU ~c:: ~
t3 ~-J
Fig, 3-5 . Columnar sections and biostratigraphic succession of ammonoids along the TeradaniValley. The localities are shown in Fig. 2-5 .

33
yielded to a specimen of Canavaria nov sp. A (Fig. 3-4). Thus, the floated
collections from Daira 6 section enable to divide into two groups, the genus
Amaltheus group and the Arieticeratinae group, on the basis of the lithofacies of
the fossil-bearing rocks. The biostratigraphic disagreement between the genera
Canavaria or Emaciaticeras and almost all species of the genus Amaltheus in
Europe (e.g. Page, 2003) supports this faunal division. Moreover, a specimen
identified as A. margaritatus is almost co-occurred with the genus Canavaria in
Daira 7 section (Fig. 3-4). According to this biostratigraphic data and the
phylogeny of the genus Amaltheus proposed in Europe and Russia (Meister, 1988;
Dagis, 1976), the genus Amaltheus group seems to be subdivided
biostratigraphically into two groups (in ascending order) as follows; A. repressus-A.
stokesi group and A. margaritatus-A. aff. talrosei group.
The ammonoid biostratigraphy in the Kuruma Group is discussed on the basis
of these lithostratigraphic and biostratigraphic evidence (Fig. 3-4).
3-3-3. Results
Three ammonoid assemblage zones in the Teradani Formation of the Kuruma
Group are proposed in this analysis as follows, in ascending order (Figs. 3-4, 3-5):
the Stokesi-Repressus Assemblage Zone, the Margaritatus-aff. Talrosei Assemblage
Zone and the Canavaria-Emaciaticeras Assemblage Zone.
3-3-3-1. Stokesi-Repressus Assemblage Zone
This zone is represented by Amaltheus stokesi (SOWERBY)(Figs. 1-2, 4-5 in
Plate 4), also recognized in the Northwest European, the North American
Cordillera, the Russian and the East Asian (Toyora Group) provinces, and
Amaltheus repressus DAGIS (Figs. 6-7 in Plate 4), described only in Northeast
Russia by previous studies (Dagis, 1976). The co-occurrence of A. stokesi and A.
repressus has been also recognized in Northeast Russia (Dagis, 1976).
This interval is identified by the occurrences of these two species. Amaltheus
cf. stokesi (Fig. 6 in Plate 4) was obtained from bed DRFO101 of the Daira River 6

34
section (Fig. 3-4), and A. repressus (Fig. 6 in Plate 4) was obtained from bed
DRF0201 of the Daira River 7 (Fig. 3-4). These two species were also found into
the floated rocks provided from these two sections (Appendix 3-5).
3-3-3-2. Margaritatus-aff. Talrosei Assemblage Zone
This zone is represented by Amaltheus margaritatus MONTFORT (Figs. 7-15 in
Plate 4) widely distributed in high latitudinal areas of the Northern Hemisphere and
Amaltheus sp. aff. A. talrosei REPIN (Fig. 1 in Plate 5), the similar species was
previously recognized only in Northeast Russian (Repin, 1968). These two species
were co-occurred within the Russian provinces reported by Repin (1988).
In the Daira River 7 section, a specimen of Canavaria sp. cf. Canavaria nov sp.
B was obtained from bed DRF0202b just below the occurrence of A. margaritatus
(Fig. 3-4). Canavaria nov sp. B (Fig. 7 in Plate 6) is characterized by the quite
rigid ribbing which has a high similarity with that of the genus Arieticeras,
co-occurred with A. margaritatus in central Europe (e.g. Meister, 1989). Therefore,
this interval is identified by the occurrences of A. margaritatus, A. aff. talrosei and
Canavaria nov sp. B (Fig. 3-4). Many specimens of these species were also found
in the floats from the Daira River 6 and 7 sections.

35
only floated rocks provided from the Daira River 6 and 7 sections (Fig. 3-4). As
mentioned above, these taxa were perhaps obtained from the stratigraphically
higher horizons than those of the genus Amaltheus based on the lithofacies of the
fossil-bearing rocks. In addition, Canavaria naxensis (GEMMELLARO), Canavaria
haugi (GEMMELLARO), Canavaria prognatum FUCINI and the genus Emaciaticeras
have been recognized only in the Elisa Subzone (Upper Emaciatum Zone) of the
Mediterranean province (Braga, 1983), equivalent to the Hawskerense Standard
Subzone (Upper Spinatum Standard Zone) of the Northwest European province
(Page, 2003), although the range of A. margaritatus was limited in the Apyrenum
Subzone, the Upper Spinatum Standard Zone, and the Margaritatus Standard Zone
(Meister, 1988). Therefore, this zone is located above the Margaritatus-aff.
Talrosei Assemblage Zone (Fig. 3-4).
3-4. International correlation for the new ammonite zonations
Ammonoid biostratigraphical analyses from Late Pliensbachian to Toarcian
have been discussed in Europe since the mid-19th century (e.g. d'Orbigny, 1842;
Oppel, 1856-1858). The zonal schemes of this time-interval have been developed
mainly in the Northwest European province and are subdivided into a sequence of
zones, subzones and horizons (Dean et aI., 1961; Gabilly et aI., 1971, 1974;
Howarth, 1973, 1978, 1980, 1992; Gabilly, 1976; Dommergues, 1979; Cope et aI.,
1980; Dommergues & Meister, 1987; Elmi et aI., 1991, 1994, 1997; Page, 2003). In
addition, the zonal frameworks of this period were also documented in the
Mediterranean province (Donovan, 1958; Braga et aI., 1982; Braga, 1983; Guex,
1973; Elmi et aI., 1974, 1991, 1994; Macchioni, 2002), the North American
Cordillera province (Smith et aI., 1988; Jakobs et aI., 1994), the South American
province (e. g. Hillebrandt, 1981, 1987) and the Northeast Russian province (e. g.
Dagis, 1968; Polubotko & Repin, 1974; Repin, 1988; Kalacheva, 1988), and these
zonations were correlated with the Northwest European zones (e.g. Dommergues et
aI., 1997a; Page, 2003-; Smith et aI., 1988; Fig. 3-6). These interprovincial
correlations, especially at subzone and horizon level, are usually difficult because

Romn'-""lZl
F
F
....
t-- - - t-- - - - - - - --t-- - - -+-- - - --+-- - - ---+ - - - - - - - - -
....Q)
~o
.....J
Q)
C. t-- - - t-- - - - - - - --t-- - - -+-- - - --t ... - - - - - - - - - - - - - - - - - - -c.:;)
c:
e
0 1--+--__-+-_~::;::..--__l--__4---__r--~I- 1---- --1
Fig. 3-6. Correia 'provi ceot e ort

37
of the faunal provincialism. In this section, the ammonite zonal schemes of the
Toyora and the Kuruma Groups proposed in this study are compared to these
zonations established outside Japan on the basis of a review of the ammonite zonal
schemes established in each province.
3-4-1. Northwest European province
Representative countries included In this province are Britain, France,
Germany and northern Spain (Page, 2003). The detailed ammonite zonal scheme
established in England has been regarded as the standard zonation of this period.
The general framework of zone and subzone in this province was documented by
Dean et al. (1961) and the details have been discussed by Gabilly et al. (1971,
1974), Howarth (1973, 1978, 1980, 1992), Gabilly (1976), Dommergues (1979),
Cope et al. (1980), Dommergues & Meister (1987), Elmi et al. (1991, 1994, 1997)
and Page (2002, 2003).
The PIT boundary has been traditionally defined by the first occurrence of the
genus Dactylioceras because of their pandemic and abundant occurrence (e. g. Dean
et al., 1961). Moreover, several definitions have been submitted, for example, the
first occurrence of Protogrammoceras paltum (BUCKMAN) in England (Howarth,
1973) and the occurrence of Paltarpites paltus (BUCKMAN) in addition to the genus
Dactylioceras in France (Gabilly, 1976; Elmi et al., 1997). As mentioned above,
the base of the P. paltus Zone corresponds to this boundary in the Toyora Group.
The zonal scheme of Late Pliensbachian has been documented on the basis of
the evolutionary lineage of Amaltheidae. The Late Pliensbachian was divided into
two zones (in ascending order): the Margaritatus Zone and the Spinatum Zone. The
Margaritatus Zone, introduced by Oppel (1856), was represented by A.
margaritatus and was subdividedinto three subzones on the basis of the lineage of
the genus Amaltheus (in ascending order): the Stokesi Subzone, the Subnodosus
Subzone and the Gibbosus Subzone. Some horizons were documented in each
sUbzone on the basis of lineage of the genus Protogrammoceras. The base of the
lowest subzone, the Stokesi Subzone, was clearly defined by the first occurrence of

38
the index species, Amaltheus stokesi (SOWERBy)(Dean et al., 1961). The base of the
middle and the upper zones were also marked by the first occurrence of each index
species, Amaltheus subnodosus (YOUNG & BIRD) and Amaltheus gibbosus
(SCHLOTHEIM). The Spinatum Zone was represented by Pleuroceras spinatum
(BRUGUIERE) and was subdivided into two subzoneson the basis of the lineage of
the genus Pleuroceras (in ascending order): the Apyrenum Subzone and the
Hawskerense Subzone. The base of the lower subzone was defined by the first
occurrence of Pleuroceras solare (PHILLIPS) and that of the upper zone was drawn
immediately above the last occurrence of P. solare.
In the Toyora Group, the A. stokesi Assemblage Zone is correlated to the
Stokesi Standard Subzone, the lowermost Margaritatus Standard Zone, based on the
occurrence of the index species, A. stokesi (Fig. 3-6). In the Nishinakayama
Formation, the base of the C. japonica Zone observed along the Sakuraguchidani
Valley in this study seems to be comparable with Late Pliensbachian. In this study,
the lowermost ammonite horizon of this zone is characterized by the occurrence of
A. margaritatus (Fig. 4-6) obtained from bed 36-1 along the South Valley. These
specimens (SA36-1-1, Fig. 9 in Plate 1; SA36-1-2, Fig. 10 in Plate 1) have a very
strong keel and the ribs projected on the keel. Although this species has been
recognized from the Subnodosus Standard Subzone to the Apyrenum Standard
Subzone (Meister, 1988), these characters are limited to the late type obtained from
the Apyrenum Standard Subzone (e.g. Fig. 3 in Plate 4; Meister, 1988). Thus, this
horizon corresponds to the Apyrenum Standard Subzone (Fig. 3-6). This
correlation suggests that the C. japonica Zone is compared to the Spinatum
Standard Zone of the Northwest European province. However, the biostratigraphic
correlation of the base of this zone is impossible along the Sakuraguchidani Valley
because the lowermost part of this zone is lacked by the inferred fault (Figs. 3-1,
3-2, 3-3). In addition, the absence of any other Amaltheidae specimens from the
Toyora Group in this study makes further detailed comparisons of these zones in
this section difficult.
In the Teradani Formation of the Kuruma Group, the Stokesi-Repressus

39
Assemblage Zone enables to compare with the Stokesi Subzone of the standard
zonation and the A. stokesi Assemblage Zone in the middle part of the
Higashinagano Formation of the Toyora Group proposed in this study (Fig. 3-6),
based on the occurrence of A. stokesi. As mentioned above, the Margaritatus-aff.
Talrosei Assemblage Zone is mainly represented by A. margaritatus. This index
species corresponded from the Subnodosus Standard Subzone to the Apyrenum
Standard Subzone is characterized by the chronological changes of surface
ornaments (Meister, 1988) as follows; the early type from the Subnodosus and
Gibbosus Standard Subzones has a low keel with fine crenulations and the weak
ribbing, and the late type from the Apyrenum Standard Subzone is characterized by
the individual keel with well prorsiradiated serrations and the connection of the
ribs and the keel. Based on these morphological characters of A. margaritatus from
the Margaritatus-aff. Talrosei Assemblage Zone, this interval is correlated from the
Subnodosus to the Apyrenum Subzones of the standard zonation (Fig. 3-6). The
Canavaria-Emaciaticeras Assemblage Zone is associated with the genera
Canavaria and Emaciaticeras, typical in the Mediterranean assemblage (Braga,
1983). Based on the correlation of the Northwest European and the Mediterranean
zonations, this zone is corresponded to the Hawskerense Standard Subzone (Fig.
3-6).
The Early Toarcian zonation of northwest Europe has mainly been established
on the basis of the evolutionary lineages of Dactylioceratidae and Hildoceratidae
and has been divided into three zones (in ascending order): the Tenuicostatum Zone,
the Serpentinum Zone, and the Bifrons Zone. The Tenuicostatum Zone was
represented by Dactylioceras tenuicostatum (YOUNG & BIRD)(Dean et al., 1961)
and was subdivided into four subzones (in ascending order): the Paltum Subzone,
the Clevelandicum Subzone, the Tenuicostatum Subzone and the Semicelatum
Subzone, which were discussed by the lineage of Dactylioceratidae except for the
lowest subzone (Howarth, 1973). The Serpentinum Zone, represented by
Harpoceras serpentinum (SCHLOTHEIM), was subdivided into two subzones based
on the lineage of Harpoceratinae (in ascending order): the Exaratum Subzone and

40
the Falciferum Subzone. The base of the Exaratum Subzone, represented by
Cleviceras exaratum (YOUNG & BIRD), was defined by the first occurrence of the
genus Eleganticeras (Howarth, 1992; Elmi et al., 1997). Howarth (1992) drew this
base by the first occurrence of Eleganticeras elegantulum (YOUNG & BIRD) in
Yorkshire coast (England), and described a sequence of E. elegantulum followed by
the first occurrence of C. exaratum and H. serpentinum, an early type of the genus
Harpoceras, which was approximately 3 m thick. The upper subzone, the
Falciferum Subzone, was represented by Harpoceras falciferum (SOWERBY) and
the base was defined by the first occurrence of the index species (Dean et al., 1961).
The Bifrons Zone, represented by Hildoceras bifrons (BRUGUIERE), was
documented on the basis of the evolutionary lineage of Dactylioceratidae and was
subdivided into three subzones (in ascending order): the Commune Subzone, the
Fibulatum Subzone and the Crassum Subzone. The bases of these subzones were
defined by the first occurrences of each index species, Dactylioceras commune
(SOWERBY), Peronoceras fibulatum (J. DE C. SOWERBY) and Catacoeloceras
crassum (YOUNG & BIRD).
In this study, a zonal boundary, the base of the P. paltus Zone, is proposed
above the PIT boundary in the Toyora Group. The base of the D. helianthoides
Zone, drawn at the first occurrence of D. helianthoides, seems to be correlated to a
horizon within the Clevelandicum Standard Subzone (Fig. 3-6), because the first
occurrence of this species was drawn immediately below the first occurrence of
Dactylioceras tenuicostatum (YOUNG & BIRD), the index species of the base of the
Tenuicostatum Standard Subzone (Fig. 31; Schmidt-Effing, 1972). The base of the
H. inouyei Zone, defined by the first occurrence of H. inouyei (Fig. 4 in Plate 3), is
equivalent to the first occurrence of Cleviceras sp. cf. C. exaratum. The first
Occurrence of the index 'species also corresponds to that of the genus Harpoceras in
the Toyora Group (Fig. 3-6). This relationship between C. exaratum and the genus
Harpoceras is very similar to the sequence of Harpoceratinae horizons into the
Exaratum Standard Subzone in the Northwest European province (Howarth, 1992).
The minor difference between the first occurrence of E. elegantulum and those of C.

41
exaratum and early Harpoceras may be disregarded. Therefore, the base of the H.
inouyei Zone corresponds to the base of the Exaratum Standard Subzone (Fig. 3-6).
As mentioned above, the upper boundary of the H. inouyei Zone is difficult to
define because of the absence of suitable index taxa. The international correlation
of the uppermost part of the H. inouyei Zone, represented by the occurrence of D.
helianthoides (Figs. 1-3 in Plate 1), P. okadai (Fig. 6 in Plate 3), F. nakayamense
(Fig. 6 in Plate 2), C. sp. cf. C. exaratum and H. inouyei (Fig. 2 in Plate 3), is also
complicated by the endemism of the Toyora assemblage that lacks the
biostratigraphic index species of the European province. Hirano (1973b) pointed
out the morphological similarity between P. subfibulatum (Fig. 5 in Plate 1) and P.
fibulatum, the index species of the Fibulatum Standard Subzone, and therefore,
proposed that the upper boundary of his D. helianthoides Zone which was equal to
the upper limit of the H. inouyei Zone of this study seems to be correlated to a
horizon within the subzone (Fig. 3-6). In addition, the biostratigraphic key genera,
which are indicated to be above the Fibulatum Standard Subzone, were absent in
this section. These characters suggest that the uppermost part of the H. inouyei
Zone is approximately correlated within the Fibulatum Standard Subzone by
following Hirano (1973b)(Fig. 3-6).
The Late Toarcian ammonoid zonal scheme of this province has been discussed
on the basis of the evolutionary lineages of Grammoceratinae, Phymatoceratinae
and Hammatoceratinae and has been divided into five zones (in ascending order):
the Variabilis Zone, the Thouarsense Zone, the Dispansum Zone, the Pseudoradiosa
Zone and the Aalensis Zone. The Variabilis Zone was represented by Haugia
variabilts (D'ORBIGNY)(Dean et al., 1961) and was subdivided into three subzones
(in ascending order): the Variabilis Subzone, the Illustris Subzone, and the Vitiosa
Subzone, which were discussed by the lineage of the genus Haugia (Gabilly et al.,
1971; Gabilly, 1976). The Thouarsense Zone, represented by Grammoceras
thouarsense (D'ORBIGNY), was subdivided into four subzones based on the lineage
of Grammoceratinae (in ascending order): the Bingmanni Subzone, the Striatulum
Subzone, the Fascigerum Subzone and the Fallciosum Subzone. The Dispansum

42
Zone was represented by Phlyseogrammoceras dispansum (LYCETT) and was
subdivided into two subzones (in ascending order): the Insigne Subzone and the
Gruneria Subzone. The Pseudoradiosa Zone, represented by Dumortieria
pseudoradiosa (BRANCO), was documented on the basis of the evolutionary lineage
of the genus Dumortieria, Grammoceratinae, and was subdivided into two subzones
(in ascending order): the Levesquei Subzone and the Pseudoradiosa Subzone. The
Aalensis Zone, represented by Pleydellia aalensis (ZIETEN), was subdivided into
two subzones based on the lineage of the genus Pleydellia, Grammoceratinae (in
ascending order): the Mactra Subzone and the Fluitans Subzone.
In the Ishimachi Sandstone and Mudstone Member of the Toyora Group, the
assemblage of the Pseudogrammoceras-Phlyseogrammoceras Assemblage Zone
contains the genera Grammoceras and Phlyseogrammoceras (Fig. 3-2, Appendix
3-4). The former has been mainly obtained from the Fallciosum Standard Subzone
(Gabilly, 1976), and the latter is characterized by the limited range in the
Dispansum Standard Zone (Rulleau, 2007). These biostratigraphical characters
suggest that the Pseudogrammoceras-Phlyseogralnmoceras Assemblage Zone
enables to compare from the Fallciosum Standard Subzone of the Thouarsense
Standard Zone to the Dispansum Standard Zone of the Northwest European zonal
scheme (Fig. 3-6).
3-4-2. Mediterranean province
Representative countries included in this province are Austria, Italy, southern
Spain and North Africa (Page, 2003). Ammonite zonal schemes employed in this
province are commonly different from those of Northwest Europe owing to the
faunal provincialism. The provincial zonations were mentioned by Donovan (1958),
Guex (1973), Elmi et al. (1974, 1991, 1994, 1997), Braga et al. (1982), Braga
(1983), Meister (1987), Dommergues et al. (1997a, b) and Meister et al. (1994).
However, the biostratigraphic correlations between the Northwest European and
the Mediterranean provinces that have been tabulated by Dommergues et al.
(1997b), Elmi et al. (1997), Macchioni (2002) and Page (2003), and the intra-zonal

43
comparison, especially at the horizonal level, still has many problems (Page,
2003).
Although a rework due to the regression around the PIT boundary has been
identified in the Northwest European province (Page, 2003), these phenomena are
not recognized in the Mediterranean province. Therefore, almost all candidates of
Global Stratotype Section and Point (GSSP) are selected from this province by the
International Commission on Stratigraphy (ICS). In this province, this boundary
has been defined by the first occurrence of the subgenus Dactylioceras
(Eodactylites) , including D. (E.) polymorphum, D. (E.) mirabile, D. (E.) simplex
and D. (E.) pseudocommune (Guex, 1973; Macchioni, 2002). Because this
subgenus has not been reported in the Toyora Group, the PIT boundary drawn in
this area is difficult to compare in detail with that of the Mediterranean province.
Late Pliensbachian fauna in the Mediterranean province are characterized by
the abundance of Hildoceratidae genera (Fuciniceras, Protogrammoceras,
Arieticeras, Emaciaticeras and so on), with some rare Amaltheidae specimens (e.g.
Elmi et al., 1974). Therefore, the zonations of this province have been discussed on
the basis of evolutionary lineages of Hildoceratidae. The zonal scheme of Late
Pliensbachian in this province has been divided into three zones (in ascending
order): the Lavinianum Zone, the Algovianum Zone and the Emaciatum Zone (Fig.
3-6). The presence of faunal differentiations makes interprovincial correlation
difficult at the intra-zonal level (Page, 2003). The lower zone, the Lavinianum(
Zone represented by Fuciniceras lavinianum (MENEGHINI), was compared with the
lower part of the Margaritatus Zone of the Northwest European zonation. Detailed
subdivision of this zone was different in the interprovincial area. The Algovianum
Zone, represented by Arieticeras algovianum (OPPEL), was subdivided into five
subzones (in ascending order): the Rogazzonii Subzone, the Bertrandi Subzone, the
Accuratum Subzone, the Meneghinii Subzone and the Levidorsatum Subzone. This
zone corresponds with the lower part of the Margaritatus Standard Zone to the
lower part of the Spinatum Standard Zone. The upper zone, the Emaciatum Zone
represented by Emaciaticeras emaciatum (CATULLO), was subdivided into the

44
Solare Subzone and the Elisa Subzone, in ascending order, and correlated with the
upper part of the Spinatum Standard Zone.
The zonal scheme of Early Toarcian was divided into three zones (in ascending
order): the Polymorphum Zone, the Levisoni Zone and the Bifrons Zone (Fig. 3-6).
The closest correlation between the Mediterranean and the Northwest European
provinces is enabled only within the Bifrons Zone (Page, 2003). The Polymorphum
Zone, represented by Dactylioceras polymorphum FUCINI, was mentioned by the
evolutionary lineage of Dactylioceratidae, especially the subgenus Eodactylites,
and was subdivided into the Mirabile Subzone and the Semicelatum Subzone, in
ascending order. This zone corresponds to the Tenuicostatum Zone of the
Northwest European zonation. The Levisoni Zone, the middle zone represented by
Harpoceras levisoni (SIMPSON), was mainly discussed on the basis of the
evolutionary lineage of Hildoceratidae. The base was defined by the first
occurrence of the index species. This zone was subdivided into two subzones (in
ascending order): the Levisoni Subzone and the Falciferum Subzone, and compared
to the Serpentinum Zone of the standard zonation. The Bifrons Zone, the upper
zone represented by H. bifrons, was subdivided into the Sublevisoni Subzone and
the Bifrons Subzone, in ascending order, and correlated with the Bifrons Zone of
the standard zonation.
The zonation of Late Toarcian was divided into five zones (in ascending
order): the Gradata Zone, the Bonarellii Zone, the Speciosum Zone, the Meneghini
Zone and the Aalensis Zone (Fig. 3-6). The zonal scheme of this period has been
discussed on the basis of the evolutionary lineages of Harpoceratinae,
Mercaticeratinae, Grammoceratinae, Phymatoceratinae, Hammatoceratinae and
Dactylioceratidae. The closest correlation between the Mediterranean province and
the Northwest European province (especially central Europe) enables only within
the Aalensis Zone (Page, 2003). The Gradata Zone was represented by Brodieia
gradata (MERLA) and was subdivided into the Gemma Subzone and the
Alticarinatus Subzone, in ascending order. This zone corresponds to the Variabilis
Zone of the Northwest European zonation. The Bonarellii Zone, represented by

45
Hammatoceras bonarellii PARISCH & VIALE, was mainly discussed on the basis of
the evolutionary lineage of the genus Pseudogrammoceras. This zone was
subdivided into two subzones (in ascending order): the Mediterraneum Subzone
and the Fallaciosum Subzone, and compared to the Thouarsense Zone of the
standard zonation. The Speciosum Zone, represented by Hammatoceras speciosum
JAMENSCH, was subdivided into the Speciosum Subzone and the Reynesi Subzone,
in ascending order, and correlated to the Dispansum Standard Zone. The Meneghini
Zone was represented by Dumortieria menegnihi HAUG, subzones were not
proposed. The Aalensis Zone of this province is the same as that of the Northwest
European province including the subdivision of the subzones (see above).
In general, the Toyora faunas, especially in Early Toarcian, shared many
similarities with the Mediterranean fauna (Hirano, 1973b). The faunal composition
characterized by the dominance of the high-diversity Harpoceratinae genera is
typical in the Mediterranean province, although the genus Cleviceras is more
common in the Northwest European province (Howarth, 1992; Page, 2008). In
contrast, correlation between these two zonations is difficult because of the
absence of the biostratigraphic key species of the Mediterranean zones from the
Toyora Group. On the basis of the correlation between the Northwest European and
the Mediterranean zonal schemes, the zonation of the SakuraguchidaniMudstone
Member, the Toyora Group, is compared to the Mediterranean zonal scheme, as
shown in Fig. 3-6. The A. stokesi Assemblage Zone is roughly corresponded to the
lower part of the Lavinianum Zone. The base of the C. japonica Zone seems to be
compared to the base of the Meneghinii Subzone, and the base of the P. paltus Zone
is corresponded to the base of the Polymorphum Zone (PIT boundary). The D.
helianthoides Zone is perhaps correlated to the Semicelatum Subzone of the
Mediterranean zonation. In the H. inouyei Zone, the base is correlated to the
Levisoni Zone, and the upper boundary is roughly compared to the middle part of
the Bifrons Subzone. The Pseudogralnlnoceras-Phlyseogrammoceras Assemblage
Zone enables to correspond to the upper part of the Fallaciosum Subzone and the
Speciosum Zone of the Mediterranean province.

46
The interval below the inferred fault along the North Valley contains several
ammonoid specimens as follows: P. sp. aff. P. rinaldinii (Fig. 9 in Plate 2), P.
paltus (Fig. 8 in Plate 2) and Fontanelliceras sp. from bed 10-6, and Lytoceras sp.
and D. helianthoides (Fig. 1 in Plate 1) from bed 14-1. In addition to the
occurrence of P. paltus, the occurrence of the genus Petranoceras, described in the
Mirabile Zone (lowermost Toarcian) of central Italy (Faraoni et al., 1994), and the
genus Fontanelliceras, characteristic taxa from the uppermost Pliensbachian to the
lowermost Toarcian, suggest that bed 10-6 is comparable to the P. paltus Zone. In
contrast, bed 14-1 is corresponded to the D. helianthoides Zone based on the
occurrence of the index species.
As mentioned above, the assemblages of the Teradani Formation, the Kuruma
Group, are mainly associated with the genus Amaltheus, except for that of the
Canavaria-Emaciaticeras Assemblage Zone. Therefore, correlation of the
Stokesi-Repressus and the Margaritatus-aff. Talrosei Assemblage Zones with the
Mediterranean zonal scheme is difficult because of the faunal disagreement. On the
basis of the correlation between the Northwest European and the Mediterranean
zonal schemes, these two assemblage zones are compared to the Mediterranean
zonal scheme, as shown in Fig. 3-6. The Stokesi-Repressus Assemblage Zone is
compared to the lower part of the Lavinianum Zone, and the Margaritatus-aff.
Talrosei Assemblage Zone is corresponded to the upper part of the Lavinianum
Zone, Algovianum Zone and the Solare Subzone. In contrast, the fauna of the
Canavaria-Emaciaticeras Assemblage Zone includes the affinitive species of C.
naxensis, C. haugi and C. prognatum, in addition to the occurrence of the genus
Emaciaticeras from this zone. On the basis of the ranges of these species and the
genus in the Mediterranean province, the Canavaria-Emaciaticeras Assemblage
Zone enables to compare to the Elisa Subzone (Upper Emaciatum Zone) of the
Mediterranean zonation (Fig. 3-6).
3-4-3. North American Cordillera province
In North America, the ammonoid biostratigraphy has been studied in Cordillera,

47
western North America. The assemblages from this area contain some endemic taxa
known from the circum-Pacific area, for example Dactylioceras kanense McLEARN
and Peronoceras pacificum HILLEBRANDT, and lack some European genera
including Nodicoeloceras, Hildoceras and Haugia. These characters necessitate the
establishment of regional zonal frameworks in this province. The ammonoid zonal
schemes were mentioned by Smith et al. (1988) and Jakobs et al. (1994), mainly
discussed in the Queen Charlotte Islands.
, The Upper Pliensbachian was divided into the Kunae Zone and the
Charlottense Zone in ascending order. The former, represented by Fanninoceras
kunae McLEARN, was corresponded to the Margaritatus Standard Zone, and the
latter, represented by Fanninoceras charlottense McLEARN, was compared to the
Spinatum Standard Zone.
The Toarcian was divided into five zones, the Kanense Zone, the Planulata
Zone, the Crassicosta Zone, the Hillebrandti Zone and the Yakounensis Zone in
ascending order. The Kanense Zone was represented by Dactylioceras kanense
McLEARN, and the base was defined by the first occurrence of the index species.
This zone was correlated to the Tenuicostatum and the Serpentinum Zones of the
Northwest European zonation (Fig. 3-6). The Planulata Zone, represented by
Rarenodia planulata VENTURI, was almost compared with the Bifrons Standard
Zone (Fig. 3-6), and the base was defined by the first occurrence of index species.
The Crassicosta Zone was represented by Phymatoceras crassicosta MERLA, and
the base was defined by the index species. This base was corresponded to the
lowermost Variabilis Zone of the standard zonation, and the upper boundary
equivalent to the base of the Thouarsense Standard Zone (Fig. 3-6). The
Hillebrandti Zone, represented by Phymatoceras hillebrandt JAKOBS, was
compared to the Thouarsense Zone of the standard zonation (Fig. 3-6). The base of
this zone was defined by the first occurrences of the genus Grammoceras,
Podagrosites latescens (SIMPSON) and the index species. The Yakounensis Zone
Was represented by Yakounia yakounensis JAKOBS & SMITH, and the base was
marked by the first occurrences of Yakounia silvae JAKOBS & SMITH,Pleydellia

48
maudensis JAKOBS & SMITH, Hammatoceras speciosum JANENSCH and Pleydellia
spp. This zone was compared to the Dispansum, the Pseudoradiosa and the Aalensis
Zones of the standard zonation (Fig. 3-6).
The North American Cordillera fauna is characterized by the high endemism,
for example the occurrence of the endemic genus Fanninoceras. Thus, the
zonations proposed in the Toyora and the Kuruma Groups are difficult to compare
to the North American zonal scheme directly because of the absence of the
biostratigraphic key species of the North American zones from these groups
without some exceptions, for example the occurrence of "Cleviceras" sp. cf. C.
chrysanthemum (YOKOYAMA) and Cleviceras sp. cf. C. exaratum (YOUNG & BIRD)
from the North American Cordillera province. On the basis of the correlation
between the Northwest European and the North American Cordillera zonal schemes
(Smith et aI., 1988; Jakobs et aI., 1994), the zonations of the Toyora and the
Kuruma Groups are compared to the North American Cordillera zonal scheme, as
shown in Fig. 3-6. The A. stokesi Assemblage Zone of the Toyora Group and the
Stokesi-Repressus Assemblage Zone of the Kuruma Group are correlated to the
lower part of the Kunae Zone, and the C. japonica Zone (Toyora Group) is almost
corresponded to the Charlottense Zone. In the Kuruma Group, the lower part of the
Margaritatus-aff. Talrosei Assemblage Zone is equivalent to the upper part of the
Kunae Zone, and the upper part of the Margaritatus-aff. Talrosei Assemblage Zone
and the Canavaria-Emaciaticeras Assemblage Zone are corresponded to the
Charlottense Zone of the North American Cordillera zonation. The P. paltus, the D.
helianthoides and the lower part of the H. inouyei Zones of the Toyora Group are
compared to the Kanense Zone, and the upper part of the H. inouyei Zone is coeval
with the lower part of the Planulata Zone in North American Cordillera. The
Pseudogrammoceras-Phlyseogrammoceras Assemblage Zone is roughly correlated
to the upper part of the Hillebrandti Zone and the lower part of the Yakounensis
Zone.

49
3-4-4. South American province
Ammonoid assemblages from the South American province also contain some
endemic species of circum-Pacific and lack some European species like the North
American Cordillera province. Hillebrandt (1987) proposed the regional ammonoid
zonal schemes in the Northern Atacama and the Northeast Antofagasta regions of
Northern Chile. The Upper Pliensbachian was subdivided into the Fannini and the
Disciforme Zones in ascending order, and the Toarcian was subdivided into 10
zones, the Tenuicostatum Standard Zone, the Hoelderi Zone, the Largaense Zone,
the Pacificum Zone, the Chlensis Zone, the Toroense Zone, the Copiapense Zone,
the Tenuicostatum Zone, the "Lotharingica" Zone and the "Fluitans" Zone in
ascending order. Although this zonation was well compared with the North
American Cordillera zonal scheme on the basis of the abundant common species
(Jakobs et aI., 1994; Hillebrandt et aI., 1992), the correlations of this zonal scheme
to the Toyora and the Kuruma zonations are difficult because the faunal
disagreement.
According to the comparisons among the North American Cordillera, the South
American and the standard zonations (Smith et aI., 1988; Jakobs et aI., 1994), the
zonal schemes of the Toyora and the Kuruma Groups are indirectly corresponded to
the South American zonation, as shown in Fig. 3-6. During Late Pliensbachian, the
A. stokesi, the Stokesi-Repressus and the lower part of the Margaritatus-aff.
Talrosei Assemblage Zones are correlated to the Fannini Zone, and the C. japonica
Zone, the upper part of the Margaritatus-aff. Talrosei Assemblage Zone and the
Canavaria-Emaciaticeras Assemblage Zone are compared to the Disciforme Zone
of the South American zonation. In the Toarcian of the Toyora Group, the P. paltus
and the D. helianthoides Zones are almost equal to the Tenuicostatum Standard
Zone of the South American zonal scheme, and the H. inouyei Zone is roughly
corresponded to the Hoelderi, the Largaense and the lower part of the Pacificum
Zones in South American province. The Pseudogrammoceras-Phlyseogrammoceras
Assemblage Zone seems to be compared to the upper part of the Copiapense Zone
and the lower part of the Tenuicostatum Zone represented by Phlyseogrammoceras

50
tenuicostatum (JAWORSKI).
3-4-5. Northeast Russia
In the Russian province, the ammonoid biostratigraphy from the Upper
Pliensbachian to the Toarcian have been mainly analyzed in five areas; the Russian
Platform, the Mediterranian Geosynclinal Belt, the Siberian Platform, the
Northeast Russia, and the Far East Russia. The regional ammonoid zonal schemes
were proposed by Dagis & Dagis (1965), Polbotko & Repin (1966, 1974) and Dagis
(1968, 1976) in the Omolon and the Korkadon Basin, the Northeast Russia, the
most maj or ammonoid source of the Russian province during this period.
The zonation of the Upper Pliensbachian was divided into three zones, the
Stokesi Zone, the Tarlosei Zone, and the Viligaensis Zone in ascending order. The
Late Pliensbachian assemblage from this province is characterized by the
co-occurrence of the Northwest European and the Russian endemic species of the
genus Amaltheus, the typical Boreal taxa in this period. Dagis (1976) discussed the
systematic evolution of the Russian endemic species and the biostratigraphic
correlation of the ranges of these species. The regional zonal scheme of the Upper
Pliensbachian was established on the basis of the evolutionary lineage of the genus
Amaltheus and was compared to the standard zonation. The Stokesi Zone,
represented by A. stokesi, was applied the Stokesi Subzone of the Northwest
European zonation. This zone was corresponded to the range of the index species,
and the fauna was associated with A. stokesi, Amaltheus bifurcus (HOWARTH) and
Amaltheus repressus DAGIS (Dagis, 1976; Repin, 1988). The Tarlosei Zone was
represented by Amaltheus tarlosei REPIN, the endemic species of the Russian
provinces. The fauna of this zone was associated with A. margaritatus, Amaltheus
subbifurcus REPIN, Amaltheus brodnaensis ventrocalvus REPIN, A. talrosei,
Amaltheus striatus asiaticus REPIN, Amaltheus bulunensis REPIN, Amaltheus
evolutus BUCKMAN, Amaltheus sensibilis DAGIS and Amaltheus conspectus DAGIS
(Dagis, 1976; Repin, 1988). This zone was corresponded to the Subnodosus Zone
and the lower part of the Gibbosus Subzones of the standard zonation. The

51
Viligaensis Zone was also represented by the Russian endemic species, Amaltheus
viligaensis (TUCHKOV), and was correlated to the upper part of the Gibbosus
Standard Subzone and the Spinatum Standard Zone. The assemblage of this zone
includes A. talrosei, Amaltheus brodnaensis brodnaensis REPIN, Amaltheus
borealis DAGIS, Amaltheus complanatus DAGIS, Amaltheus extremus REPIN and
Arieticeras (?) sp. aff. A. algovianum (OPPEL) in addition to the index species
(Dagis, 1976; Repin, 1988).
In the Kuruma Group, the Stokesi-Repressus Assemblage Zone is corresponded
to the Stokesi Zone of the Northeast Russian zonation on the basis of the
occurrence of the A. stokesi and A. repressus (Fig. 3-6). The Margaritatus-aff.
Talrosei Assemblage Zone is compared to the Talrosei Zone and the lower part of
the Viligaensis Zone because of the occurrence of A. margaritatus and A. aff.
talrosei from this assemblage zone (Fig. 3-6). The Canavaria-Emaciaticeras
Assemblage Zones is difficult to correlate directly to the Northeast Russian zones
because of the faunal disagreement. On the basis of the comparisons among the
Northeast Russian, the Mediterranean and the standard zonations in this study, this
assemblage zone is indirectly compared to the upper part of the Viligaensis Zone of
the Northeast Russian Zonation, as shown in Fig. 3-6.
In the Toyora Group, the A. stokesi Assemblage Zone is correlated to the
Russian Stokesi Zone because of the occurrence of A. stokesi from the Toyora
Group (Fig. 3-6). The base of the C. japonica Zone of the Toyora Group enables to
compare to the lower part of the Viligaensis Zone of the Northeast Russian
zonation because of the occurrence of A. margaritatus with the Apyrenum-type
surface ornaments (see above in detail)(Fig. 3-6).
The zonation of the Toarcian in Northeast Russia was subdivided into six zones,
the Propinquum Zone, the Falciferum Standard Zone, the Athleticum Zone, the
Monestieri Zone, the Polare Zone and the Rozenikrantzi Zone in ascending order.
The Propinquum Zone, represented by Tiltoniceras propinquum (WHITEAVES), was
subdivided into two beds. The lower bed was characterized by the occurrence of
Kedonoceras (=Orthodactylites) asperum DAGIS with Kedonoceras comptum DAGIS,

52
T. propinquum and Tiltoniceras costatum BUCKMAN, and the upper bed was
distinguished by Arctomercaticeras costatum REPIN associated with
Arctomercaticeras tenue REPIN and T. propinquum. The PIT boundary was drawn at
the base of the lower bed defined by the first occurrence of the subgenus
Orthodactylites, the genus Dactylioceras. The PIT boundary seems to be correlated
to the base of the Clevelandicum Subzone of the Northwest European zonation (Fig.
3-6), thus the Propinquum Zone is corresponded to the Clevelandicum, the
Tenuicostatum and the Semicelatum Subzones of the Tenuicostatum Zone proposed
in the Northwest European province (Fig. 3-6). Moreover, this boundary is
compared to the base of the D. helianthoides Zone proposed in the Toyora Group
(Fig. 3-6), because the base of this zone is also marked by the first occurrence of
the subgenus Dactylioceras (Orthodactylites), D. (0.) helianthoides. The
Falciferum Standard Zone was applied to the Falciferum Zone (=the Serpentinum
Zone) of the Northwest European zonal scheme. Thus, the base of the falciferum
Standard Zone in Northeast Russia enables to correlate to the base of the H. inouyei
Zone in the Toyora Group (Fig. 3-6). The Athleticum Zone, represented by
Dactylioceras athleticum (SIMPSON), was compared to the lower part of the Bifrons
Standard Zone (Fig. 3-6). The Monestieri Zone, represented by Zugodactylites
monestieri DAGIS, was corresponded to the middle to upper part of the Bifrons
Standard Zone, the Polare Zone, represented by Porpoceras polare (FREBOLD), was
correlated to the Uppermost Bifrons Zone, and the Rozenikrantzi Zone,
distinguished by Pseudolioceras rozenikrantzi (SIM:PSON), was compared to all over
the Upper Toarcian (Fig. 3-6). The upper boundary of the H. inouyei zone proposed
in the Toyora Group is roughly compared to the Monestieri Zone in Northeast
Russia, and the Pseudogrammoceras-Phlyseogrammoceras Assemblage Zone seems
to be corresponded to the upper part of the Rozenikrantzi Zone (Fig. 3-6).

4. Relationship between the ammonoid
paleoceanographic changes
53
paleobiogeographic and the
4-1. Aims of this chapter
The paleobiogeographic changes of marine biota enable to trace the long term
paleoceanographic changes, for example, global ocean circulation, sea water
temperature and sea level change. In Early Jurassic period, the paleobiogeography
of marine biota has been discussed in many taxa, including ammonoids (e.g.
Hallam, 1975; Macchioni & Cecca, 2002; Page, 2008), bivalves (e. g. Hallam, 1977,
1983; Liu et aI., 1998; Damborenea, 2002) and ostracods (Arias, 2006, 2007, 2008).
Above all, ammonoid is one of the most distinguished index taxa because of their
high biostratigraphic resolution.
The paleobiogeographic changes of the Jurassic ammonoid have been
discussed in a global scale. Arkell (1956) analyzed the worldwide Jurassic
ammonoid paleobiogeography by using the terms realm and province, and
recognized three realms as follows; the Boreal Realm (high latitudinal realm in the
Northern Hemisphere), the Tethyan Realm (low latitudinal realm) and the Pacific
Realm (endemic realm in the Eastern Pacific). Hallam (1975) decided the
boundaries between the Boreal and the Tethyan Realms globally on Pliensbachian,
Bathonian, Callovian and Tithonian. Page (2008) discussed the ammonoid
paleobiogeography through Jurassic period based on the recognition of marine
paleobiogeographic units (realm, subrealm, province and subprovince) proposed by
Westermann (2000a, b). In the Northern Hemisphere, four provinces including the
Arctic, the Subboreal, the Athabascan and the West Pacific provinces were
recognized in the Boreal Reahn, and four provinces, the Submediterranean, the
Mediterranean, the Ethiopian and the Andean provinces, were distinguished in the
Tethyan Realm during Late Pliensbachian and Toarcian. The East Pacific Realm
was also decided on the Athabascan and the Andean provinces by the occurrence of
endemic taxa in the East Pacific. Although these Early Jurassic ammonoid
paleobiogeographic studies have been analyzed mainly in Europe, North American

54
Cordillera and East Asia (Japan), it has not been discussed in West and Central
Asia because of the absence of ammonoid paleobiogeographic data. The lack of
these data has disturbed the discussions of ammonoid paleobiogeography in all
over the Northern Hemisphere. In contrast, the late Early Toarcian ammonoid
assemblage from Northern Tibet was recently described by Yin et al. (2006).
In Japan, the Early Jurassic ammonoid assemblages have been compared
individually with the European, the North American Cordillera, the West Asian and
the Madagascar faunas. Sato (1956) indicated that the assemblage from the
Teradani Formation, the Kuruma Group, was characterized by the mixing of a
Boreal taxon, the genus Amaltheus, and a Tethyan taxon, the genus Canavaria,
during Late Pliensbachian, although the fauna from the Nishinakayama Formation,
the Toyora Group, was' associated only with the Tethyan genera. Hirano (1973 b)
compared the faunas from the Toyora Group with that from outside Japan and
pointed out the faunal similarity between the Toyora and the Mediterranean or the
central European assemblages, associated with the Tethyan elements, during Late
Pliensbachian. According to Hirano (1973b), the Toyora assemblage showed high
similarity with the Alta Brianza, the Pyrenee and the Eastern Paris Basin faunas
during Early Toarcian, in addition to the high similarity between the Toyora and the
Eastern Caucasus faunas during Late Toarcian. However, a comprehensive and
detailed transitional pattern of late Early Jurassic ammonoid assemblages from
Japan has not been analyzed in previous studies. The global and the detailed
ammonoid paleobiogeography should be discussed by utilizing the more detailed
faunal association data of Japanese Early Jurassic ammonoid assemblages revealed
in this study. Therefore, the aims of this study are to analyze a comprehensive and
detailed chronological changes of the late Early Jurassic ammonoid faunal
compositions from Japan by using the assemblages from the Toyora and the
Kuruma Groups, to indicate the paleobiogeographical positions of each assemblage
by comparing with faunas from outside Japan, for example, the Northwest
European, the Mediterranean, the North Tibetan, the Iranian, the Southeast Asian,
the Northeast Russian and the Far East Russian provinces and to discuss the

55
paleoceanographic changes which affected the paleobiogeographic changes of late
Early Jurassic ammonoid assemblages in the Northern Hemisphere.
4-2. Paleobiogeographic changes of Japanese ammonoid assemblages
The paleobiogeographic changes of Japanese ammonoid assemblages are
discussed on the basis of Fig. 4-1, the ranges of each ammonite taxon obtained
from the Toyora and the Kuruma Groups are illustrated in the family or the
subfamily level.
4-2-1. Faunal characters of the assemblages from the Kuruma Group
As mentioned above, the ammonoid assemblage from the Stokesi-Repressus
and the Lower to Middle Margaritatus-aff. Talrosei Assemblage Zones of the
Kuruma Group, corresponded to the Margaritatus Standard Zone, is associated with
the genus Amaltheus (Fig. 4-1). The family Amaltheidae including the genera
Amaltheus, Amauroceras and Pleuroceras is dominant in the assemblages from
high latitudinal areas of the Northern Hemisphere, such as the Northwest European
and the Northeast Russian provinces. Dommergues et al. (1987) designated the
family Amaltheidae as a distinctive index taxon of the Boreal fauna. Thus, the
dominance of the genus Amaltheus in this assemblage indicates a high similarity
with the Boreal fauna of this period. In addition, the faunal specific composition of
this assemblage is characteristic in the mixing of Pan-Boreal species, A. stokesi and
A. margaritatus, and Russian endemic species, A. repressus and A. aff. talrosei.
The ammonoid assemblage from the upper part of the Margaritatus-aff.
Talrosei Assemblage Zone, corresponded to the Apyrenum Subzone of the standard
zonation, is mainly composed of the genus Canavaria, Canavaria nov sp. B, with a
few specimens of the genus Amaltheus, A. margaritatus (Fig. 4-1). The subfamily
Arieticeratinae (family Hildoceratidae), including the genus Canavaria and
Emaciaticeras, is typical in low latitudinal areas, for example, the Mediterranean
province (Braga, 1983), and has been regarded as one of the major Tethyan faunal
composition (e.g. Hallam, 1975). Therefore, this faunal association suggests that

l ,evesat.lei
So 5
Fig. 4-1 . Ranges of ach ammonit family or ub mily obt in from th Toyor nd h Kurum rouduring late Early Jura ic time (La e PH n b chi n-To rcian). Ammonoidzon I ch m in th orthwEuropean province folio Page (2003). u rieal g r from Gr d t in t I. (2004).
thi a embla e i charact nz d by th mrxm
fauna.
f th n th I thy' n
The assemblage from the anavarra- maciatic ra A mblag n ,
correlated to the Hawskerense ubzone of the standard nati n, i character ized
by a high di ersity of the subfamily Arieticeratinae associated with f u P C I of
the genus Canavaria Canavaria nov sp. A, C. aff. naxensis, aff. pro natum and
C. aff. haugi) and a species of the genus Emaciaticeras maciaticeras nov
sp. (Fig. 4-1. his assemblage is characterized by the dominance of the ethyan
index taxa.

57
The other ammonoid assemblage is obtained from the uppermost part of the
Teradani Formation exposed along the Teradani Valley (Figs. 3-4, 4-1), associated
with the subfamily Harpoceratinae, including Fuciniceras nakayamense and
Protogrammoceras sp., and the family Dactylioceratidae (Dactylioceras sp.). This
faunal association suggests the high similarity between this assemblage and that
from the Tethyan Realm. Although this assemblage enables to be roughly
corresponded to Early Toarcian on the basis of the existence of the genus
Dactylioceras, more detailed biostratigraphic correlations are disturbed by the poor
occurrence of the ammonoid specimens.
4-2-2. Faunal characters of the assemblages from the Toyora Group
The ammonoid specimens corresponded to the Stokesi Assemblage Zone were
obtained from the Higashinagano Formation, the Toyora Group. As mentioned
above, two materials were described as A. cf. stokesi and Arieticeras aff. apertum
by Hirano (1973b). These specimens suggest that the fauna from this assemblage
zone is characterized by the mixing of the Boreal and the Tethyan taxa (Fig. 4-1).
From the lower part of the C. japonica Zone correlated to the Apyrenum
Standard Subzone, two specimens of A. margaritatus are obtained along the South
Valley of the Sakuraguchidani Valley (Fig. 3-3). Therefore, this assemblage is
composed of the Boreal fauna (Fig. 4-1). In contrast, the assemblage from the
upper part of the C. japonica Zone, correlated to the Hawskerense Subzone of the
standard zonation, is associated with Canavaria japonica, the subfamily
Arieticeratinae (Figs. 3-3, 4-1). This faunal composition shows a high similarity
with that in the Tethyan Realm.
The ammonoid assemblages from the P. paltus and the D. helianthoides Zones,
biostratigraphically corresponded to the Tenuicostatum Standard Zone, is
dominated by a high-diversity of the subfamily Harpoceratinae (family
Hildoceratidae), such as the genera Protogrammoceras, Fuciniceras, Lioceratoides,
Paltarpites and Petranoceras, with some Arieticeratinae (F. fontanellense) in the
lower part of the P. paltus Zone and abundant Dactylioceratidae (D. helianthoides)

58
during the D. helianthoides Zone (Fig. 4-1). In addition to the subfamily
Arieticeratinae, some genera of the subfamily Harpoceratinae including all of these
taxa mentioned above are also dominant in the low latitudinal assemblages, for
example in the Submediterranean faunal province, thus this subfamily has been
regarded as the index taxa of the Tethyan Realm (e.g. Macchioni & Cecca, 2002;
Page, 2008). On the other hand, the family Dactylioceratidae has been regarded as
the pandemic paleobiogeographic character (e.g. Macchioni & Cecca, 2002).
Therefore, the faunal association indicates that this assemblage is the
Tethyan-derived fauna.
The ammonite assemblage from the H. inouyei Zone is dominated by some
Harpoceratinae genera, such as Harpoceras, Cleviceras, Polyplectus, Hildaites and
Lioceratoides, with some Dactylioceratidae (D. helianthoides, P. subfibulatum)(Fig.
4-1). During the early to middle Early Toarcian (Tenuicostatum and Serpentinum
Standard Zones), the ammonoid assemblages from the Boreal Realm, for example,
the Northwest European and the Northeast Russian provinces, are characterized by
the occurrence of the phylogeny of Cleviceras, constituted by the genera
Tiltoniceras, Eleganticeras and Cleviceras (Howarth, 1992). Some specimens of C.
cf. exaratum, also described previously in the Northwest European and the North
American provinces, are obtained from the H. inouyei Zone. In contrast, the other
genera of Harpoceratinae, such as Harpoceras, Polyplectus, Hildaites and
Lioceratoides, are mainly distributed in the Tethyan Realm. Thus, the assemblage
from the H. inouyei Zone is characteristic in the mixing of the Boreal and the
Tethyan faunas.
The Late Toarcian ammonoid assemblage fro In the Ishimachi Sandstone and
Mudstone Member, corresponded to the Fallaciosum Subzone (Thouarsense Zone)
and the Dispansum Zone of the standard zonation, is mainly composed of
Harpoceratinae (genus Osperlioceras) and Grammoceratinae (genera Grammoceras,
Pseudogrammoceras and Phlyseogrammoceras) with a few Dactylioceratidae and
Phylloceratina (Fig. 4-1). These taxa were mainly distributed in low and middle
latitudinal areas of the Northern Hemisphere during Late Toarcian. In contrast, the

59
fauna from the Boreal Realm including the Northwest European province and North
Alaska have been characterized by the occurrence of the genus Pseudolioceras in
the subfamily Harpoceratinae (e.g. Page, 2008). Consequently, the Late Toarcian
ammonoid fauna from the Toyora Group is associated only with the Tethyan index
taxa.
4-2-3. Faunal changes of Japanese ammonoid assemblages
On the basis of the paleobiogeographic faunal characters, seven ammonoid
assemblages are identified in East Asia (Japan) during late Early Jurassic time
(Late Pliensbachian-Toarcian).
In the Margaritatus Standard Zone, the first ammonoid assemblage associated
with the Boreal fauna is recognized from the Kuruma Group (Fig. 4-1). However,
this Boreal faunal composition is completely replaced by the Tethyan elements in
the third assemblage corresponded to the Hawskerense Standard Subzone, through
the second assemblage characterized by the mixing of the Boreal and the Tethyan
index taxa during the Apyrenum Standard Zone. The same faunal transition is also
detectable in almost coeval assemblage from the Toyora Group. Consequently, a
turnover from the Boreal fauna to the Tethyan fauna is recognizable in the Japanese
ammonite assemblages during the Apyrenum Standard Zone (Fig. 4-1). This
bio-event occurs in the Japanese assemblages is biostratigraphically earlier than
the timing of corresponded faunal change previously identified in the Northwest
European assemblages. The faunal replacement from the Boreal to the Tethyan
faunas, chronologically corresponded to Pliensbachian/Toarcian boundary, has
been mainly recognized in the Northwest European province (e.g. Dean et aI.,
1961; Macchioni & Cecca, 2002) and is characterized by the extinction of the
family Amaltheidae in whorl the Boreal Realm.
The Tethyan-derived faunal composition continues during the Tenuicostatum
Standard Zone (fourth assemblage obtained from the Toyora Group). However, the
faunal composition of the fifth assemblage corresponded to the Serpentinum and
the Lower-Middle Bifrons Standard Zones (Commune-Fibulatum Subzones) is

60
distinguished by the dominance of Tethyan Harpoceratinae with a few Boreal
elements, the genus Cleviceras. Therefore, a mixing of the Tethyan and the Boreal
faunal elements is recognized in East Asia during middle to late Early Toarcian
(Fig. 4-1). In contrast, the sixth ammonoid assemblage obtained from the middle
Upper Toarcian of the Toyora Group is composed of the Tethyan index taxa (Fig.
4-1), thus the faunal mixing seem to be limited in middle to late Early Toarcian.
During the Stokesi Standard Zone, the ammonoid assemblage from the Toyora
Group is characterized by the mixing of Boreal and the Tethyan faunal elements.
This faunal association differs from the Boreal-dominated faunal composition of
the coeval assemblage from the Kuruma Group (Fig. 4-1). This faunal disagreement
between the Kuruma and the Toyora assemblages during this time-interval seems to
reflect the latitudinal interval between these two localities.
4-3. International correlation of Japanese ammonoid assemblages
Faunal compositions of 13 faunal elements in seven time-intervals during Late
Pliensbachian to Toarcian and their geographic distribution in the Northern
Hemisphere are shown in Figs. 4-2, 4-3, 4-4, 4-5, 4-6, 4-7 and 4-8. The pie charts
are illustrated on the basis of the number of the specimens, fossil-bearing horizons
or associated species in each faunal element, distinguishable from the color of the
numbers shown at the center of these circles (to see Fig. 4-2 for the legend of the
colors). The paleogeographic map in late Early Jurassic time utilized in this
analysis is modified from the reconstructions of Damborenea (2002), Golonka
(2007) and Dera et al. (2010).
4-3-1. Stokesi Standard Subzone
As mentioned above, the Boreal fauna has been characterized by the
dominance of Amaltheidae during late Pliensbachian. In the Stokesi Standard
Subzone, the typical Boreal assemblages, such as Northeast Russia, Far East Russia,
Northern Caucasus, Brooks-Mackenzie Basin (North Alaska) and Western Scotland,
are composed of only Amaltheidae (Fig. 4-2). In addition, the fauna from Northeast

Fig. 4-2. Fau al com
Tul, B.C . (Sp)
1. ,1988:. 1992)
.)

62
Russia is characteristic in the dominant Russian-endemic species of the genus
Amaltheus, represent 93 % of the faunal composition.
The Tethyan assemblages distributed in latitude 300N or less is mainly
associated by Harpoceratinae and Arieticeratinae. In the Submediterranean faunal
province including Apennines (Italy), Subbetic (Southern Spain), Lusitania Basin
(Portugal), Bakony Basin (Hungary) and High-Atlas (Morocco), the assemblages
are dominated by Mediterranean Harpoceratinae, representing 42-92 % of the
faunal composition, with abundant Phylloceratina and Lytoceratina accounted for
94 % in maximum (Fig. 4-2). In the North American Cordillera province except for
Tulsequah (Northern British Columbia), it is difficult for the ammonoid
assemblages from this province to discuss the faunal correlation in subzonal level
because of the low biostratigraphic resolution of the North American regional
zonal scheme.Although the fauna from southern part of North American Cordillera,
such as Oregon (USA), Spatsizi (Northern British Columbia) and the Queen
Charlotte Islands, look to be characterized by the abundant occurrence of
Arieticeratinae, these faunal compositions seems to be mainly reflected by that of
the Subnodosus-Gibbosus Standard Subzones, because of the dominance of the
Tethyan elements, Harpoceratinae, in the Tulsequah fauna during the Ha wskerense
Standard Subzone. Moreover, the mixing of the Boreal and the Tethyan faunas are
recognizable in the assemblages from the Causses Basin (France) and Tulsequah
(Northern British Columbia), located around latitude 30oN.
The fauna from the Kuruma Group in the Stokesi Standard Subzone is
dominated by the Northwest European species of the genus Amaltheus,
representing 75 % of the faunal composition, with some Russian endemic
Amaltheus species (Fig. 4-2). This association indicates high faunal similarity
between the Kuruma and the Northeast Russian assemblages. In contrast, the
mixing of Amaltheidae and Arieticeratinae recognized in the fauna from the Toyora
Group is also distinguishable in the Tulsequah fauna. Thus, the assemblage from
the Toyora Group is somewhat similar with that from Northern British Columbia
during this time-interval.

63
4-3-2. Subnodosus-Gibbosus Standard Subzones
As well as the Stokesi Standard Subzone, the Boreal fauna during the
Subnodosus-Gibbosus Standard Subzones IS identified by the dominant
Amaltheidae, recognized in Scotland, England, Northeast Russia, Siberian Platform
(Russia) and Northern Caucasus assemblages (Fig. 4-3). The Russian endemic
species of the genus Amaltheus are mainly distinguishable in the Northeast Russian
and the Siberian Platform assemblages, representing about 55 % of the faunal
composition. The assemblage from the Kuruma Group in this time-interval is
dominated by the Northwest European species of the genus Amaltheus with some
Russian endemic Amaltheus (eight percents of the faunal composition), thus, is
similar with the Northeast Russia and the Siberian Platform assemblages (Fig. 4-3).
In contrast, the Tethyan fauna of the Subnodosus-Gibbosus Standard Subzone
IS generally characterized by the dominance of Arieticeratinae (representing
46-94 % of the composition) which is recognizable in the assemblages from
Apennines, Subbetic, the Bakony Basin, Jebel Zaghouan (Tunisia), Oregon,
Spatsizi and Eastern Caucasus. In addition to Arieticeratinae, the abundant
occurrences of Harpoceratinae are characteristic in High-Atlas, Lycia Allochton
(Turkey), the Queen Charlotte Islands and Vietnam assemblages (Fig. 4-3). The
dominance of Phylloceratina and Lytoceratina in the Bakony Basin, the Lusitania
Basin and Lycia Allochton, representing 27-58 % of the faunal composition,
enables to except for this paleobiogeographic discussion because of the pandemic
distribution of these suborders. The assemblages from North American Cordillera
are characteristic in also the genus Fanninoceras (6-37 % of the faunal
composition), the endemic genus of Harpoceratinae in Cordillera. According to the
distribution of the mixing fauna of the Boreal and the Tethyan elements
distinguished in the Causses Basin, the Lusitania Basin, Tulsequah and Far East
Russia, the boundary between the Boreal and the Tethyan Realms enables to be
drawn around latitude 300N as well as the Stokesi Standard Zone, except for the
remarkable distribution of the Far East Russian fauna (Fig. 4-3).

J
B kony B(Geczy &
1 8)
c;
Fig. -3. Faunal coSubzo e . Legend ofSpatsizi in 8ri . Colu

65
4-3-3. Apyrenum Standard Subzone
In the Apyrenum Standard Subzone, the assemblages from the Boreal Realm
are characterized by the dominance of Amaltheidae including the genus
Pleuroceras, the Russian endemic species of the genus Amaltheus and A.
margaritatus, in addition to the genus Tiltoniceras which is mainly distributed in
high latitudinal areas of the Northern Hemisphere. The typical Boreal fauna
recognized in Northeast Russia, Western Scotland, England and the Causses Basin
are composed of almost only Amaltheidae (Fig. 4-4), especially the Northeast
Russian fauna is dominated by the Russian endemic species of the genus Amaltheus
(97 % of the faunal composition).
The Tethyan faunas in this time-interval are mainly composed of
Harpoceratinae (distinguished in High-Atlas, Subbetic and the Queen Charlotte
Islands) or Arieticeratinae (recognizable in Oregon)(Fig. 4-4). The assemblage of
Far East Russia, biostratigraphically compared to this time-interval by Repin
(1988), is probably corresponded to the Hawskerense Standard Subzone on the
basis of their faunal association including the genus Paltarpites and C. japonica.
The assemblages from the Bakony Basin and Apennines, paleogeographically
located at the eastern edge of the Western Tethys, are dominated by Phylloceratina
and Lytoceratina (representing about 75 % of the faunal composition) with some
Hildoceratidae (Harpoceratinae and Arieticeratinae) and a few Amaltheidae. The
mixing of the Boreal and the Tethyan faunas is distinguishable in the assemblages
from Subbetic, the Queen Charlotte Islands, Spatsizi and Tulsequah. In North
American Cordillera except for Oregon, the Boreal elements are mainly associated
with the genus Tiltoniceras and the Northwest European Amaltheidae, however the
former taxon is absent in the coeval European faunas. During the Apyrenum
Standard Subzone, the boundary between the Boreal and the Tethyan Realms IS
removed southward and is drawn around latitude 300N (Fig. 4-4).
The fauna from the Kuruma Group in the Apyrenum Standard Subzone is
characterized by the dominant Arieticeratinae, representing 83 % of the faunal
composition, with some Northwest European species of the genus Amaltheus (Fig.

Or on (Or)(Iml y. 1968:
Smith l I. 1 8),
in (l u) /
~I
1. .2007) /I
I/
0
<> /
Fig. 4-4. Faunal coSubzone. Legend of e u er( orth American Cordillera). SEast (Russia) are com 0 0

67
4-4). Although this faunal association indicates the mixing of the Boreal and the
Tethyan faunas, no similar composition, characterized by the dominance of
Arieticeratinae with some Amaltheidae, are recognizable in the Northern
Hemisphere. In contrast, the assemblage from the Toyora Group in this
time-interval is composed of only the Northwest European species of the genus
Amaltheus, and show a high similarity with those of the Boreal fauna. However,
this assemblage does not seem to be reflected by the original faunal composition
sufficiently, because of the small amount of the specimens.
4-3-4. Hawskerense Standard Subzone
The genus Pleuroceras, the Russian endemic species of the genus Amaltheus
and the genus Tiltoniceras are regarded as the Boreal faunal elements during the
Hawskerense Standard Subzones. In the Northwest European province, the Boreal
faunas are characterized by the dominance of the genus Pleuroceras (representing
more than 86 % of the faunal composition), recognizable in Western Scotland,
England and the Causses Basin (Fig. 4-5). In contrast, the fauna from Northeast
Russia is dominated by the Russian endemic species of the genus Amaltheus. A
specimen of Amaltheus viligaensis TUCHKOV, one of a typical endemic species of
the Russian province in this time-interval, was also reported in the Queen Charlotte
Islands (Smith et al., 1988).
In the Tethyan Realm, the ammonoid assemblages are mainly composed of
Harpoceratinae, Arieticeratinae and Phylloceratina/Lytoceratina in this subzone.
The assemblages from the Submediterranean faunal province are generally
characterized by the dominant Arieticeratinae (36-84 % of the faunal composition),
especially in Subbetic, Apennines and High-Atlas (Fig. 4-5). This trend is also
recognized in Japanese assemblages, representing 94 % in the Kuruma Group and
100 % inthe Toyora Group. This means that the faunal composition of the Japanese
assemblages show a high similarity with that of the Submediterranean faunal
province, especially the High-Atlas fauna. In contrast, the faunas from Jebel
Zaghouan (Tunisia), Lycian Allochton (Turkey) and Far East Russia are dominated

68
by Harpoceratinae, representing 45-97 % of the faunal compositions.
The faunas from the Lusitania and the Bakony Basins are associated with
dominant Harpoceratinae, Arieticeratinae and Phylloceratina/Lytoceratina with rare
Boreal elements, such as the genera Pleuroceras and Tiltoniceras (Fig. 4-5). In the
North American Cordillera province, including the Queen Charlotte Islands,
Tulsequah and Spatsizi, the fauna is characterized by the mixing of the Tethyan
elements and the genus Tiltoniceras. These mixed faunas are distributed around
latitude 30 oN .

Engl nd (En)(Howarth . 1992
Simm et 1. .2010),Braa - c nz
B in (Br)(Paul on. 1991)
... )
Fig. 4-6. Faunal cornoosmonsZone. Lege d of e
S ard

70
The assemblage from the Toyora Group in the Tenuicostatum Standard Zone is
composed of the dominant Harpoceratinae (67 % of the faunal composition) with
Arieticeratinae (genus Fontanellicerasv, Dactylioceratidae (genus Dactylioceras)
and Lytoceratina (Fig. 4-6). This faunal association is similar with the
Submediterranean faunas. Especially, the diversified generic composition of
Harpoceratinae from the Toyora Group including the genera Paltarpites,
Protogrammoceras, Fuciniceras, Lioceratoides and Petranoceras has a high
similarity with that from Apennines in this time-interval.
4-3-6. Serpentinum and Lower to Middle Bifrons Standard Zones
As well as the Tenuicostatum Standard Zone, the pandemic distribution of the
genus Dactylioceras is also distinguishable in the Serpentinum and the Lower to
Middle Bifrons Zone (including the Commune and the Fibulatum Standard
Subzones).
The Boreal Realm fauna during the Serpentinum Zone is characterized by the
dominance of the genera Eleganticeras and Cleviceras. The phylogeny of
Cleviceras constituted by the genera Tiltoniceras, Eleganticeras and Cleviceras
was proposed by Howarth (1992) and has been regarded as a Boreal element from
latest Pliensbachian to middle Early Toarcian (e.g. Page, 2008). In addition, the
Boreal fauna from the Bifrons Standard Zone and the Upper Toarcian are identified
by the occurrence of the genus Pseudolioceras (Harpoceratinae)(e.g. Page, 2008).
The Boreal type assemblages of this time-interval are mainly recognized in the
Russian province, such as Northeast and Far East Russia (Fig. 4-7).
The Tethyan Realm during this time-interval is characterized by the dominant
occurrence of some Harpoceratinae genera, for example, Harpoceras, Hildaites,
Hildoceras and Ovaticeras (e.g. Macchioni & Cecca, 2002; Page, 2008). These
genera account for 56 % of the faunal composition and are co-occurred with some
characteristic taxa containing the subfamily Mercaticeratinae (Hildoceratidae) in
the Mediterranean province, including Apennines and Algeria (Fig. 4-7). The
Tethyan-derived fauna are also recognized in West and Southeast Asia (Iran,

a d
( 0 )ort
.C. )
in (C )S)
w. Scotl nd (Sc)(Howarth . 1992;
Simm at at., 2010)•
Fig. 4-7. Faunal compositio of e" a etementsMiddle Bifrons S andard Zo es. Lege doe n l u." h.c~r

72
Vietnam and Kalimantan).
The assemblage from the Toyora Group is associated with the dominant
Mediterranean Harpoceratinae, representing 57 % of the faunal composition, with
some Arctic Harpoceratinae (C. cf. exaratum), Dactylioceratidae (genera
Dactylioceras and Peronoceras) and Phylloceratina/Lytoceratina. This faunal
composition, characterized by the mixing of the Tethyan and the Boreal faunas,
shows a high similarity with that of the Northwest European province, especially
the Causses Basin fauna (Fig. 4-7). These mixed faunas are also distributed in
North American Cordillera and Northern Caucasus. In contrast, the assemblage
from Northern Tibet, paleogeographically near the Toyora Group, is dominated by
Arctic Harpoceratinae, such as the genus Cleviceras (Yin et al., 2006).
4-3-7. Fallaciosum Standard Subzone and Dispansum Standard Zone
As mentioned above, the Boreal ammonite fauna during Late Toarcian IS
distinguished by the dominance of the genus Pseudolioceras (e.g. Page, 2008). The
typical Boreal fauna recognized in the Brook-Mackenzie Basin, Northern Alaska, is
characterized by the occurrence of Pseudolioceras lectum (SIMPSON) and was
roughly corresponded to the Thouasense Standard Zone (Poulton, 1991).
In contrast, the Tethyan faunas in the Fallaciosum Standard Subzone
(Thouasense Standard Zone) and the Dispansum Zone are characteristic in the
dominance of Hildoceratidae including Harpoceratinae, Grammoceratinae and
Paroniceratinae, except for the genus Pseudolioceras (e.g. Page, 2008). The
Northwest European, the West Asian, the Northern Caucasian and the Southeast
Asian (Roti; Indonesia) faunas are dominated by the subfamily Grammoceratinae.
The assemblages from the southern part of the Northeast European province, such
as the Causses and Lusitania Basins, are composed of the various taxa including
Harpoceratinae, Grammoceratinae, Paroniceratinae and Hammatoceratidae (Fig.
4-8). The fauna from North American Cordillera, recognizable in the Queen
Charlotte Islands, is distinguished by the dominant Phymatoceratidae, representing
72 % of the faunal composition, with some Tethyan elements.

welatemnann.
Toyor G. ( 0)(Th' y)
••En I nd (En)
(Simm at al.. 2010)
e a S ar

74
The assemblage from the Toyora Group is characterized by the abundance of
the genus Osperlioceras (Harpoceratinae) and Grammoceratinae (including the
genera Grammoceras, Pseudogrammoceras and Phlyseogrammoceras)(see chapter
3 and Fig. 4-8). The generic composition of this assemblage shows a high
similarity with that of the assemblage from the Causses Basin, located in the
Submediterranean faunal province of the Tethyan Reahn.
4-4. Discussion of the paleoceanographic change in the Northern Hemisphere
4-4-1. Early Jurassic paleoenvironmental changes in the Northern Hemisphere
Among the various paleoceanographic elements, the fluctuations of sea level
and seawater temperature are attracted for considering the origin of the late Early
Jurassic ammonoid paleobiogeographic changes in the Northern Hemisphere. The
faunal changes of Japanese ammonoid assemblages analyzed in this study are
illustrated in Fig. 4-9, with the paleobiogeographic faunal changes of the European
ammonoid assemblages (Macchioni, 2002), the strontium isotope fluctuation
(87 Sr/86 Sr) analyzed in Yorkshire, England (Jones et al., 1994; McArthur et al.,
2000), the oxygen isotope excursion ( D 180 ) in Yorkshire (McArthur et aI., 2000;
Bailey et al., 2003), Dotterhausen and Aselfingen (both in Germany; Bailey et al.,
2003) and the seawater temperature fluctuations calculated by the composite D180
data (Dera et al., 2009a).
4-4-1-1. Sea level changes
The sea level changes during the Jurassic period have been analyzed in
utilizing various techniques by Hallam (1978, 1988, 2001), Vail & Todd (1981),
Haq et al. (1988), Sulyk (1991), Li & Grant-Mackie (1993), Legaretta & Uliana
(1996), de Graciansky et al. (1998) and Hardenbol et al. (1998). Hallam (1978,
1988) proposed the shallowing and deepening cycles in Europe by using the
combination of facies analyses. The sea level curves through the Jurassic time were
illustrated in the North Sea by Exxon team on the basis of the seismic stratigraphic
analysis (Vail & Todd, 1981) and sequence stratigraphic analysis (Haq et aI., 1988).
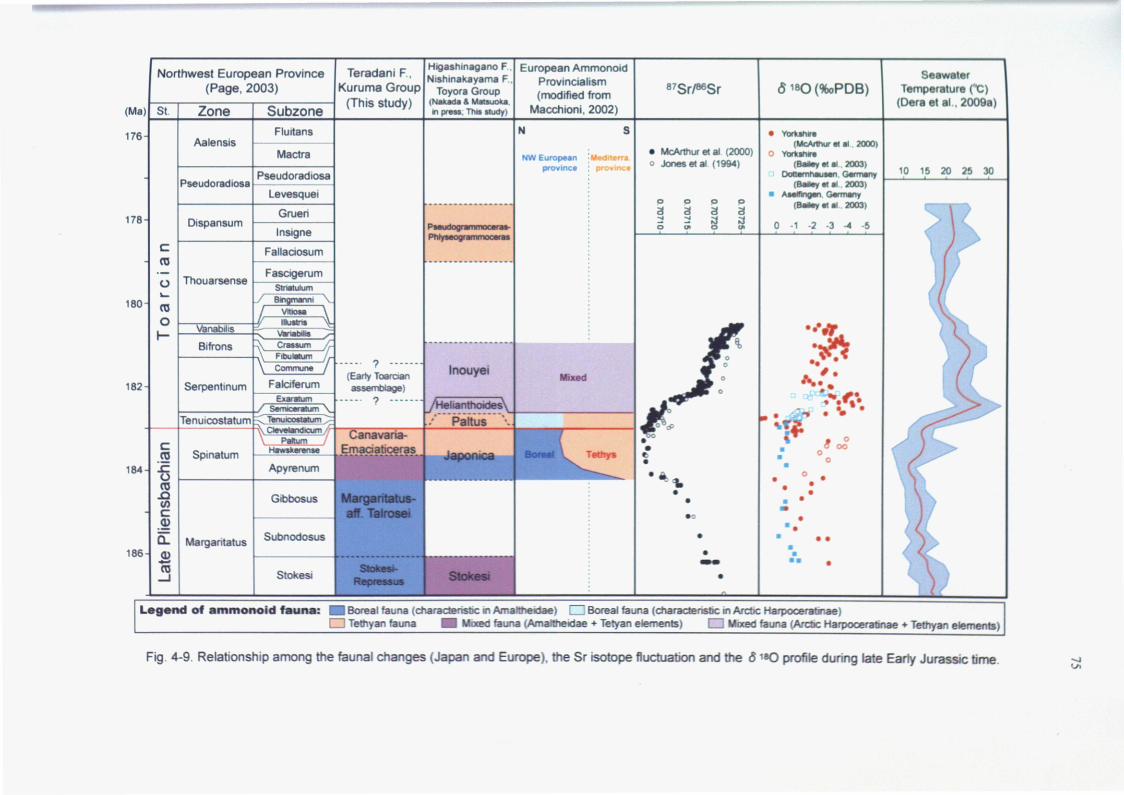
9 Ea Ju a ic
87Sr Sr 8 180 ( POS)(0
•0
0 0 0 0
~ ~ ~ ~~ ::! jj
0 ., ·2 -J0 (II 0 tit
Ter dani F.,Kuruma Group
(This study )
Mactra
FluitansAalensis
P udo dio
c:ro
Northwe t European Provine(Page , 2003)
roo~
182?
c:.~ Spin tums:oCO.c G' u(/)c:
.~a.. rg S
186 Q)-ro S...J
ILFig. 4-9. Relatio ip a eta E
180
(M) St. Zone Subzone
178
176

76
According to Haq et al. (1988), the sea level during Early Jurassic was generally
rising from Hettangian to Toarcian. Similar eustatic carves have been also
proposed in East Greenland (Sulyk, 1991), Southern Tibet (Li & Grant-Mackie,
1993) and west-central Argentina (Legaretta & Uliana, 1996). In contrast, the
second order transgressive-regressive cycles during late Early Jurassic time (Late
Pliensbachian and Toarcian) were revealed both in the Northwest European and the
Mediterranean provinces by de Graciansky et al. (1998). According to de
Graciansky et al. (1998), two regressions (Lower Pliensbachian-lower part of the
Margaritatus Standard Zone and Upper Toarcian-Aalenian) and a transgression
(middle to upper part of the Margaritatus Standard Zone-Bifrons Standard Zone)
were recognized in the Northwest European province, and two regressions (middle
to upper part of the Margaritatus Standard Zone-lower part of the Tenuicostatum
Standard Zone and Upper Toarcian-Aalenian) and two transgressions (Lower
Pliensbachian-lower part of the Margaritatus Standard Zone and upper part of the
Tenuicostatum Standard Zone-Bifrons Standard Zone) were distinguished in the
Mediterranean province.
On the other hand, the Sr isotope fluctuation has been considered to reflect the
eustatic sea level changes during late Early Jurassic (Hallam, 1997; Cope, 1998),
because the Sr isotope ratio is controlled by the amount of the detrital fluxes which
are relevant to the erosion of continents. Jones et al. (1994) analyzed the Sr isotope
ratio in the Lower Jurassic succession exposed in Yorkshire, England, for the first
time (Fig. 4-9), and recognized a negative excursion from Hettangian to
Pliensbachian and a positive shift in Toarcian. McArthur et al. (2000) showed the
high-resolution Sr isotope fluctuation of belemnites obtained from the same section
of Jones et al. (1994)(Fig. 4-9). Cohen et al. (2004) suggested that the abrupt
positive shift of the Sr isotope ratio in the Exaratum Standard Subzone, analyzed
by McArthur et al. (2000), was reflected in the global warming which has been
considered as the trigger of the Early Toarcian OAE and enhanced the chemical
weathering of continents.

77
4-4-1-2. Sea surface temperature changes
Oxygen isotope excursions (D 180 ) have been considered as an important
paleoenvironmental proxy for reflecting the seawater temperature. The numerous
data of D 180 have been analyzed in the European Lower Jurassic successions by
using the various taxa including belemnite, bivalve, planktonic foraminifera and so
on. Especially, the D 180 data from the Lower Toarcian have been attracted for
discussing the causes of the Early Toarcian OAE (Jenkyns & Clayton, 1986;
McArthur et aI., 2000; Bailey et aI., 2003; Rosales et aI., 2004; Gomez et aI., 2008;
Dera et aI., 2009b).
The D 180 profiles of belemnite have been regarded as the index of sea surface
temperature and have been examined in several regions in Europe, such as
Yorkshire (McArthur et aI., 2000; Bailey et aI., 2003), the Paris Basin (Dera et aI.,
2009b), the Southern German Basin (Bailey et aI., 2003) and Northern Spain
(Rosales et aI., 2004; Gomez et aI., 2008). According to the D 180 records of
belemnite in Yorkshire and the Southern German Basin analyzed by McArthur et al.
(2000) and Bailey et aI. (2003)(Fig. 4-9), a prominent negative shift was
distinguishable from the Apyrenum Standard Subzone to the Exaratum Standard
Subzones, in addition to two gentle positive excursions recognized in the
Subnodosus Standard Subzone and the Serpentinum Standard Zone. On the basis of
these geochemical fluctuations, the presence of a cooling during Late
Pliensbachian (Margaritatus Standard Zone) and an abrupt warming during Early
Toarcian (Tenuicostatum Standard Zone and Exaratum Standard Subzone) have
been distinguished in Europe (e.g. Bailey et aI., 2003; Dera et al., 2009b). This
strong positive excursion suggesting the abrupt warming event is stratigraphically
corresponded to the strong positive shift of the Sr isotope ratio which mentioned
above (Fig. 4-9). The profile of the sea surface temperature calculated by the
composite D 180 data of belemnite is illustrated in Fig. 4-9.

78
4-4-2. Paleoceanographic changes on the basis of the ammonoid faunal changes
4-4-2-1. Latest Pliensbachian faunal turnover
As mentioned above, a turnover from the Boreal to the Tethyan faunas in the
Apyrenum Standard Subzone is recognizable in the late Early Jurassic ammonoid
assemblages from Japan (Figs. 4-1, 4-9). This turnover is biostratigraphically
coeval with the beginning of the warming mainly distinguished from the
Tenuicostatum Standard Zone to the Exaratum Standard Subzone in Europe (Fig.
4-9). The fluctuation of seawater temperature enables to regard as one of the
important factors which affect the paleobiogeographic faunal changes of
ammonoids. These Tethyan-dominated faunal compositions of the Japanese
assemblages are quite different with the association of the Northwest European
fauna, characterized by the abundant occurrence of Amaltheidae, in the
Hawskerense Standard Subzone. The Northwest European province was perhaps
paleolatitudinally correspondent to Japan in late Early Jurassic time on the basis of
the paleogeographic continental distribution (e.g. Golonka, 2007). Therefore, the
dramatic faunal change of Japanese assemblages in the Apyrenum Standard
Subzone is chronologically discordant for the comparable faunal turnover,
characterized by the extinction of Amaltheidae and corresponded to the
Pliensbachian/Toarcian boundary, in the almost equal latitudinal regions in the
western Tethys, such as the Causses Basin and England (Figs. 4-4, 4-5, 4-9). The
additional paleoceanographic factors are required for accounting this disagreement
of faunal compositions between the Japanese and the Northwest European
assemblages during latest Pliensbachian.
The recognition of the Tethyan-derived fauna in Japanese assemblages is
synchronous with the eustatic low stand of sea level during latest Pliensbachian
(Spinatum Standard Zone) to earliest Toarcian (Tenuicostatum Standard Zone) on
the basis of the Sr isotope ratio in Yorkshire (Fig. 4-9). On the basis of the
high-resolution ammonoid biostratigraphy, the presence of reworking and the
condensation of ammonoids have been distinguished in England and Digne area
(South France) by de Graciansky et al. (1998) and Guex et al. (2001). Macchioni

(A) arga -t tu tan..ard 0 (
~-
Fig. 4-10. Schema ic repre enta 'on 0 e s eaStandard Subzone.
rnrn,nnJ"l.at"t· tau as ,.n~ r;'+~h S S a (B)

80
(2002) suggested that this phenomenon was caused by the regression recognized in
latest Pliensbachian-earliest Toarcian. The uppermost Pliensbachian-lowermost
Toarcian succession in the Northwest European province are mainly composed of
the shallow marine sediments, such as mudstone, sandstone, limestone and
ironstone (Guex et al., 2001; Simms et al., 2010).
The uppermost Pliensbachian-lowermost Toarcian Far East Russian and North
Alaskan successions are also composed of the shallow marine clastic deposits
(mudstone and sandstone)(Poulton et al., 1992; Sey et al., 1992). The coeval South
Alaskan successions are lacked by the unconformity (Poulton et al., 1992).
According to Golonka (2007), these sediments were deposited in the shallow
marine, distributed between the North American and the Eurasian continents and
divided the Anui Ocean (Zonenshain et al., 1990; Senger & Natalin, 1996; Golonka
et al., 2003) from the Panthalassa. In the Margaritatus Standard Zone, the Boreal
elements, such as Amaltheidae, were diffuse from the high to the middle latitudinal
areas through this seaway, decided Paleo-Bering Strait in this study (Fig. 4-10).
However, the shallowing of this seaway affected by the regression during latest
Pliensbachian and earliest Toarcian perhaps disturbed the spreading of the Boreal
faunal elements from the Anui Ocean to the Panthalassa (Fig. 4-10). Consequently,
the disappearance of Amaltheidae and the domination of the Tethyan elements
recognized in the latest Pliensbachian ammonoid assemblages from Japan reflected
the shallowing of Paleo-Bering Strait caused of the global regression.
4-4-2-2. Early Toarcian faunal mixing
The mixing of the Boreal and the Tethyan faunas of the Toyora assemblage is
distinguishable in the Exaratum Standard Subzone and the Lower to Middle Bifrons
Standard Zone. During this time-interval, the eustatic sea level had been rose in
high stand (McArthur et al., 2000), almost equal to that of the Margaritatus
Standard Zone (see Fig. 4-9). This transgression allowed the spreading of the Early
Toarcian Boreal elements (phylogeny of Cleviceras) to the Panthalassa through
Paleo-Bering Strait (Fig. 4-11). In addition, the slight and coeval cooling was

(C) nu·co t m ~~I"~~r Zo ( )
Fig. 4-11. Sc ernatic represeSerpentinum and the Ea y
. (C) nuicostatum S e a (0)

82
proposed on the basis of a small positive excursion of the 5 180 fluctuation in
Europe (Fig. 4-9). Thus, the faunal mixing of Japanese assemblage in this
time-interval is affected by the global transgression and the cooling. In the
Exaratum Standard Subzone, the faunal mixing in the European ammonoid
assemblages reduced the provincialisms In the European ammonoid
paleobiogeography (Dera et al., 2010). This faunal equation also seemed to be
affected by the coeval and abrupt transgression.

83
5. Geochemical analyses of the Early Toarcian OAE in East Asia
5-1. Aims of this chapter
Deposition of organic black shales in the Lower Toarcian has been recognized
in the Northwest European province. Jenkyns & Clayton (1986) analyzed the stable
carbon isotope record of bulk carbonate ( 8 13Ccarb) through the Lower Toarcian
succession and showed a positive shift of 013Ccarb at the black shale horizon. On the
basis of this fluctuation into the black shale, corresponded biostratigraphically in
the Falciferum Zone (= the Serpentinum Zone in this study), the sedimentation of
the black shale was considered as the Early Toarcian Oceanic Anoxic Event (OAE).
Jenkyns (1988) suggested that this phenomenon was the global environmental
change because of the 'worldwide distribution of the black shale, including the
Northwest European province, the Mediterranean province, the North American
Cordillera province, the South American province, the East Asian (Japan) province,
Madagascar and Australia (Fig. 1-2). This event has been studied in detail in
Europe from various viewpoints as follows; (i13C (e.g. Jenkyns & Clayton, 1986,
1997~ Jenkyns, 1988~ Hesselbo et al., 2000, 2007~ Schmid-Rohl et al., 2002~ Cohen
et al., 2004~ Gomez et al., 2008~ Hermoso et al., 2009), (i180 (e.g. McArthur et al.,
2000~ Bailey et al., 2003~ Rosales et al., 2004; Gomez et al., 2008; Dera et al.,
2009b), Os isotope (Cohen et al., 2004), major/trace elements (Bellanca et al.,
1999; McArthur et al., 2000; Bailey et al., 2003), trace fossil contents (Parisi et al.,
1996) and distribution of clay minerals (e.g. Dera et al., 2009a). Moreover, this
phenomenon is recorded not only in continental shelf sediments but also pelagic
successions. Hori et al. (2000) showed the positive excursions of V/Ah03 ratios in
the Toarcian black chart succession exposed in the Katsuyama Section, central
Japan, suggesting the effect of the Early Toarcian OAE. The pelagic black
mudstone were recognized in Slovenia by Gorican et al. (2003), also considered to
deposit under the Toarcian OAE.
In contrast, the relationship between the Early Toarcian OAE and the mass
extinction event in epicontinental marine environments have been also discussed in

84
various taxa including ammonoids, bivalves, rhynchonellid brachiopods,
ostracodes, belemnites and dinoflagellates (Raup & Sepkoski, 1984, 1986; Hallam,
1996; Hallam & Wignall, 1997; Aberhan & Fursich, 1997, 2000; Harries & Little,
1999; Fursich et al., 2001; Macchioni & Cecca, 2002; Cecca & Macchioni, 2004;
Dera et al., 2010). This extinction has been regarded as the global bio-event on the
basis of data from northern Siberia (Zakharov, 1994) and the Andean Basin
(Aberhan & Fursich, 1997, 2000). Moreover, this event also has been recognized in
pelagic environment with the extinctions of radiolarians and calcareous
nannofossils (Hori, 1997; Gorican et al., 2003). Especially in ammonoids, two
mass extinction events were recognized in both the Northwest European and the
Mediterranean provinces around the Upper Pliensbachian-Lower Toarcian (seven
ammonoid zones, about 4.7My) by Macchioni & Cecca (2002) and Cecca &
Macchioni (2004). The first bioevent took place at the Pliensbachian/Toarcian
boundary which is connected with the extinction of Boreal type ammonoids and sea
level fall, while the second mass extinction took place at the uppermost part of the
Semicelatum Subzone of the standard zonation which corresponds to the beginning
of the Early Toarcian GAE. This extinction is characterized by the diversity fall
and the increasing value of extinction. In addition, the diversity of endemic species
significantly fell at this horizon.
In Japan, the existence of the Toarcian GAE was discussed in the
Nishinakayama Formation of the Toyora Group by Tanabe (1991) and Izumi &
Tanabe (2010). The middle part of the Nishinakayama Formation, composed of
laminated, bituminous black mudstones with sedimentary pyrite, is characterized
by the macrofauna which consists of nekton and pseudoplanktonic bivalves. On the
basis of these faunal compositions, Tanabe (1991) suggested the existence of the
Early Toarcian GAE in the Toyora Group. In addition, Izumi & Tanabe (2010)
showed the effect of the Toarcian OAE in the middle to the upper part of the Lower
Nishinakayama Formation on the basis ofthe negative carbon isotope excursion of
organisms ( 0 13Corg) and the low trace fossil content. However, contributions about
the Toarcian GAE from the Asian regions are still limited, although the Toarcian

85
GAE and the related mass extinction have been regarded as the global event. The
aims of this study are to determine the paleochemical environment of the East
Asian province during Early Toarcian by the trace element and the rare earth
element (REE) analyses, and to discuss the effect of the OAE, including the
extinction of marine macro biota, in the western Panthalassa.
5-2. Materials and analytical methods
Twenty-eight samples of mudstones from the Sakuraguchidani Valley were
analyzed as whorl rock in terms of trace element and REE geochemistry. Almost all
of these specimens were collected from the North Valley of the Sakuraguchidani
Valley except for a material from bed 41-1 exposed along the South Valley (Fig.
5-1).
Major and trace element concentrations were measured by X-ray fluorescence
(XRF) spectrometry, using a Rigaku RIX 3000 system at the Faculty of Science,
Niigata University. Glass disks were prepared by fusing rock powder sample (0.5
g) with lithium tetraborate (LbB407 , 5.0 g) for major element analysis, and rock
powder sample (1. 8 g) with lithium tetraborate (2.88 g) and lithium metaborare
(LiB02, 0.72 g) for trace element analysis. Detailed analytical procedures have
been described by Takahashi & Shuto (1997).
REE and Li, Sc, Co, Zn, Ga, Cs, Hf, Ta, Th and U abundances were determined
by using an inductively coupled plasma-mass spectrometer (ICP-MS; Agilent
7500a) at the Faculty of Science, Niigata University. Whorl rock powder (0.1 g)
was dissolved in an HF -HN03 mixture on a hotplate, followed by fusion with
Na2C03 (0.5 g) at 1,050°C. The solution was neutralized with HN03+HCl and
diluted by a factor of 20,000 with a mixture of HF -HN03-HCl. A single solution of
USGS reference material (BHYO-2) was utilized as an external calibration
standard, using reference values of Eggins et al. (1997). Sensitivity variation
during the analytical runs was corrected by using four internal standards (In, Re,
Bi). External standardization was performed for individual elements in unknown
samples by interpolation of results for replicate analyses of BHVO-2 after 5-6

86
unknown samples. Analytical quality was examined by comparing the results of
this analysis for USGS reference materials, JG-1a and W-2a with literature values.
The data of this study are consistent with literature values within 5 % deviation for
all elements, although results for other than REE are not shown.
5-3. Results
Geochemical data for the black mudstone samples from the Sakuraguchidani
Valley, including V/Ah03, U/Ah03, Ba/Ah03, Ni/Ah03, Zr/Ah03, Cr/Ah03,
Co/Ah03, Th/U, Ce/Ce* and Eu/Eu*, are shown in Figs. 5-1, 5-2, 5-3 and
Appendix 5-1. The fluctuations of 8 13Corg and Lamina Preservation Index (LPI)
shown in Figs. 5-1 and 5-2 were also analyzed in the North Valley of the
Sakuraguchidani Valley by Izumi & Tanabe (2010). The ammonite zonal scheme of
the Toyora Group in Figs. 5-1 and 5-2 are proposed in this study (see chapter 3).
5-3-1. Trace element concentrations
The results of various trace element fluctuations illustrated in Figs. 5-1 and 5-2
are normalized by Ah03 because these elemental concentrations are affected by the
amount of the detrital flux. Al concentration is regarded as a good proxy for
analyzing the fluctuation of the flux (e.g. Bellanca et aI., 1999).
Overall, U, Zr and Cr concentrations are relatively uniform through the
Sakuraguchidani Mudstone Member (Figs. 5-1, 5-2). Th/U fluctuation mostly
follows the same pattern except for three small positive shifts at beds 19-5b, 21-1
and 24-4 and three small negative shifts at beds 19-1b, 20-3b and 28-1 (Fig. 5-2).
Concentrations of V, Ba, Ni and Co strongly increase during the lower part of
the Sakuraguchidani Mudstone Member, almost all of these shifts are corresponded
to the P. paltus Zone (Figs. 5-1, 5-2). Synchronous shifts to higher values for V and
Co are recognized at bed 18-1 c, which is associated with a small shift in Ni (Figs.
5-1, 5-2). The V concentration increases slightly during the upper part of the H.
inouyei Zone with two small positive shifts at beds 26-1 and 28-1 (Fig. 5-1). The
Co fluctuation also has some small positive peaks, for example, at beds 25-5 and

VIA120 3
Fig. 5-1 . Co u arin parallel colu s. ecolumnar ion is as i
U/A1203 UA1203 Zr/A1203
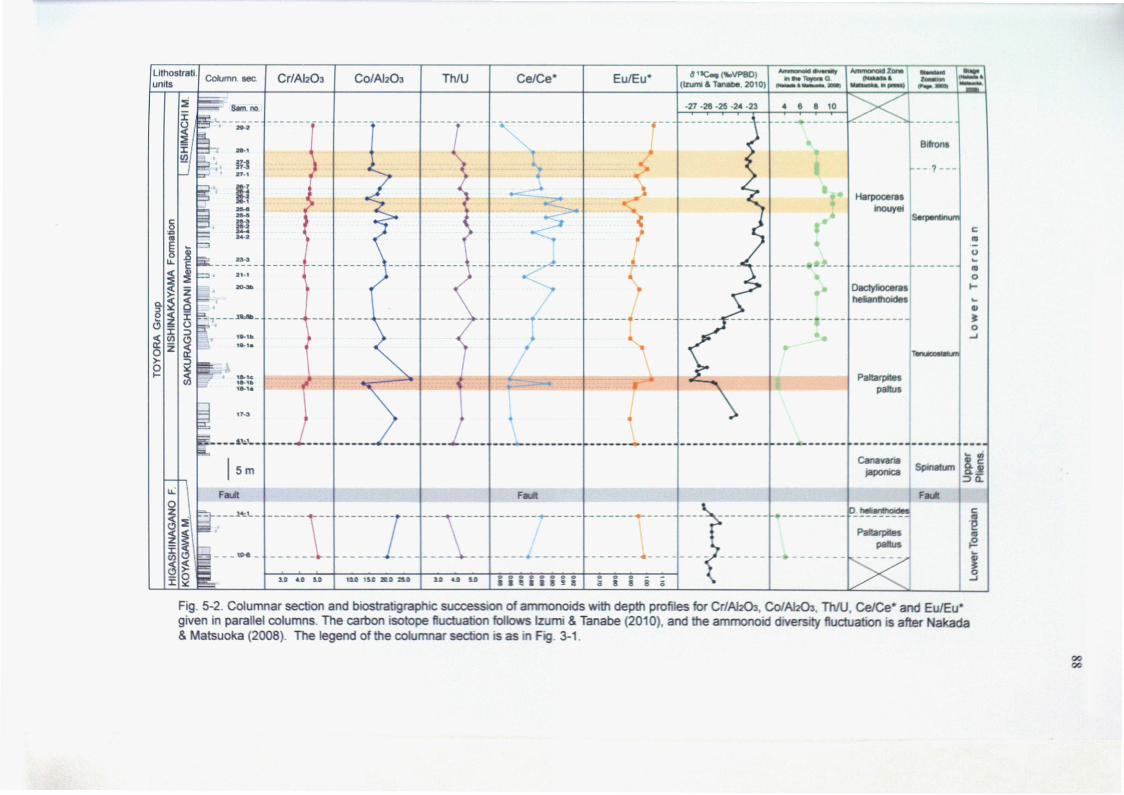
Cr/A120J Co /AI20J Th/U CeJCe Eu/Eu·
p
o...J
o...J

89
27-1 (Fig. 5-2). In contrast, the Ni concentration shows an abrupt increase at bed
17-3, immediately before the positive excursions of V and Co, with four small
positive shifts at beds 18-1c, 26-2,27-1 and 28-1, and with a negative shift at bed
18-1b associated with an increase of Ba concentration (Fig. 5-1). In addition to the
positive excursion at bed 18-1 b, the fluctuation of Ba also has two abrupt positive
shifts at beds 19-1b and 20-3b (Fig. 5-1).
5-3-2. Rare Earth Element concentrations
The North American Shale Composite (NASC)-normalized REE patterns using
values from McLennan (1989) are illustrated in Fig. 5-3. These fluctuations show
some minor variations in the REE patterns, for example a change in Eu data. I
attracted attention in Ce and Eu, considered as geochemical tracers of the GAE (e. g.
Hori et al., 2000), and analyzed the Ce and the Eu anomalies. These are calculated
as Ce/Ce*= Ceu/((Lan+Prn/2) and Eu/Eu*= Eun/((Smn+Gdn/2) after Hori et al.
(2000), and are shown in Fig. 5-2.
However the Ce/Ce* has little variation (Ce/Ce*=O. 853-0.923), some positive
shifts are recognized through the Sakuraguchidani Mudstone Member. During the P.
paltus and the D. helianthoides Zones, the Ce/Ce* value is relatively low except for
two positive Ce anomalies at beds 18-1band 20-3b (Fig. 5-2). In contrast, the
Ce/Ce* become high in the lower part of the H. inouyei Zone with some positive
anomalies at bed 25-3, 25-8 and 26-2, and decrease in the upper part of the H.
inouyei Zone.
Overall, the Eu/Eu* is uniform and increases from lower to the upper part of
the Sakuraguchidani Mudstone Member, except for a small positive anomaly at bed
18-1c and a negative anomaly at bed 26-1 (Fig. 5-2).
5-4. Discussion
5-4-1. Redox conditions during the Lower Toarcian in the Toyora Group
The geochemical data from the Sakuraguchidani Valley, the Toyora Group,
provide insights about stratigraphical changes of redox conditions during Early

01
1 1-1
1 -10
17
- 1 1 20 1 -1b 2en
18 1c 2
- 1 1 251 1 2 1-1
0
E00
s:1en
coG)
E
0
G)oecac~
.c
ww0::
0.1ro
.....J(1)
oL-
a.. "CZ E
(J)
:::JW
.0t- o o
J:L-
W Et-
.0>-
:::J......J
Fig. 5-3. orth American Shale Composit (NASC)-normarized REE patterns of black mud tone fromthe Sakuraguchidani udstone Member, exposed along the Sakuraguchidani Valley, in the ToyoraGroup. The ASC data are after cLennan (1989). The numbers of p cim n corre pond to thenumber of the outcrop in Fig. 3-1 and the number beside the columnar section in Fig. 5-2. The legendof the columnar section is as in Fig. 3-1.

91
Toarcian in the East Asian province.
As mentioned above, the concentrations of V and Co are strongly increase in
beds 18-1 c which is corresponded to the middle part of the P. paltus Zone,
associated with an abrupt negative shift of the Ni concentration and a positive
anomaly of Ce at bed 18-1b (Figs. 5-1,5-2). These elements have been recognized
as redox tracers of marine environments.
Under reducing sea water condition, V is reduced to V (III) by free H2S which
is generated by bacterial sulfate reduction and is precipitated as the solid oxide
V203 (Breit & Wanty, 1991; Wanty & Goldhaber, 1992; Hetzel et aI., 2009). In
addition, the V (III) is taken up by geoporphyrins and concentrated into the
sediments under the anoxic environment, although Ni ions are taken into the
porphyrins under the oxic condition (Moldowan et aI., 1986; Hatch & Leventhal,
1992). These excursions are stratigraphically corresponded to a negative shift of
the 013Corg and the low value of LPI analyzed by Izumi & Tanabe (201 O)(Fig. 5-1).
As a result, the positive peak of the V concentration at bed 18-1 c and the negative
excursion of the Ni fluctuation at bed 18-1b suggest that these beds were deposited
under the sulfide-bearing strongly anoxic paleoenvironment, just after the oxic
condition characterized by a positive shift of the Ni concentration at bed 17-3 (Fig.
5-1). The positive shift of the Co concentration at bed 18-1 c supports this
hypothesis (Fig. 5-2), because this element is precipitated as CoS under the
reduced condition with free hydrogen sulfide (Heggie & Lewis, 1984; Gendron et
aI., 1986). Moreover, the positive Ce anomaly at bed 18-1 b also suggests an anoxic
environment because of the enrichment of Ce (III) into the reducing water mass
(Liu et aI., 1988)(Fig. 5-2).
These geochemical peaks are also associated with a positive shift of the Ba
concentration (Fig. 5-1). Barium enrichment has been regarded as the indicator of
the high biogenic flux, which suggests the high productivity on the surface water
(Schmitz, 1987; Dymond et aI., 1992). Therefore, the anoxic marine condition
identified at beds 18-1band 18-1c seems to be affected by the high productivity on
the photic zone.

92
A strong negative excursion of the o13Corg fluctuation was also recognized
around bed 19-1 a by Izumi & Tanabe (2010), associated with the low LPI values
(Fig. 5-1). However, no distinct excursions of the redox elements at this bed
suggest that this negative shift of the o13Corg does not reflect the redox condition of
this period.
During the D. helianthoides and the H. inouyei Zones, the various excursions
of the redox elements are recognized in some beds. A small positive shift of the
concentration of V is identified at bed 20-3b corresponded to the upper part of the
D. helianthoides Zone, with the anomalies of Ba, Ni, Ce and 013 Corg (Figs. 5-1, 5-2).
Around the middle part of the H. inouyei Zone including beds 25-8, 26-1 and 26-2,
small excursions of V, Ni, Co and 013 Corg are recognized with the positive Ce
anomalies and a negative Eu anomaly (Figs. 5-1, 5-2). Moreover, the
concentrations of V and U slightly increases at bed 28-1 corresponded to the upper
part of the H. inouyei Zone (Fig. 5-1). These small shifts of the redox elements
seem to be affected by the dysoxic environments, although the o13 Corgfluctuation
increases gradually during overall these two zones (Figs. 5-1, 5-2). As a result, the
anoxic condition occurred in the P. paltus Zone was generally recovered through
the D. helianthoides and the H. inouyei Zones with some short term
oxygen-depleted marine environments.
5-4-2. Relationships between the anoxia and the ammonoid diversity changes
As mentioned above, the various epicontinental taxa, including ammonoid,
bivalve, ostracode, and so on, were affected by the Early Toarcian GAE in Europe.
Especially, the relationship between the diversity crises of ammonoid and the Early
Toarcian GAE has been discussed in both the Northwest European and the
Mediterranean provinces (Macchioni & Cecca, 2002~ Cecca & Macchioni, 2004).
The mass extinction recognized at the uppermost part of the Semicelatum Subzone
of the standard zonation was corresponded to the beginning of the Early Toarcian
GAE and was considered to be affected by this Early Toarcian paleoenvironmental
change.

93
The biodiversity fluctuation in ammonoid also has been analyzed in the Toyora
Group, southwest Japan by Nakada & Matsuoka (2008). The evolutionary trends of
diversity, origination and extinction are discussed at specific and generic levels in
the Nishinakayama Formation exposed along the Sakuraguchidani Valley. Nakada
& Matsuoka (2008) recognized a strong diversity fall at beds 18-1a, 18-1band
18-1c in the lower part of the P. paltus Zone, followed by an immediate
diversification at bed 19-1 b, a small diversity fall at bed 23-1 corresponded to the
base of the H. inouyei Zone, and a gradual diversity fall from bed 26-3 to bed 29-3,
compared to the upper part of the H. inouyei Zone (Figs. 5-2, 5-4). These events
mainly reflect the diversity fall of Harpoceratinae and that of endemic species.
The distinct diversity crisis recognized at beds 18-1 a, 18-1band 18-1 c in the P.
paltus Zone is stratigraphically corresponded to the most maj or anoxic event
characterized by the geochemical shifts of the V, Ni and Co concentrations, Ce/Ce*
and o13 Corg fluctuation (Fig. 5-4). This relationship suggests that the anoxic marine
condition in the P. paltus Zone caused the East Asian ammonoid assemblage to the
diversity crisis. In addition, this diversity crisis is characterized by the extinction
of the subfamily Harpoceratinae, and the survival of only a species of pandemic
Arieticeratinae. The ammonite assemblage from bed 41-1, corresponded just below
the horizons with geochemical anomalies and faunal extinction, is mainly
associated with moderately diversified Harpoceratinae, P. paltus (Fig. 8 in Plate 2),
P toyoranus (Fig. 7 in Plate 2), L. yokoyamai and L. aradasi (Fig. 10 in Plate 2),
with a few Arieticeratinae (F. fontanellense) and Lytoceratina (Lytoceras sp.)(see
Fig. 3-3). The specific composition of Harpoceratinae shows a high endemism of
the East Asian province. All of these endemic Harpoceratinae species are
disappeared in bed 18-1 by affecting the dramatic shift of the marine redox
condition, and only a pandemic species of the subfamily Arieticeratinae, F.
fontanellense (Figs. 6-7 in Plate 1) is recognizable during this oxygen-depleted
time-interval. The end of the anoxic marine condition is accompanied by the
dramatic recovery of the ammonoid diversity, especially the endemic
Harpoceratinae species. Seven endemic species of the subfamily Harpoceratinae
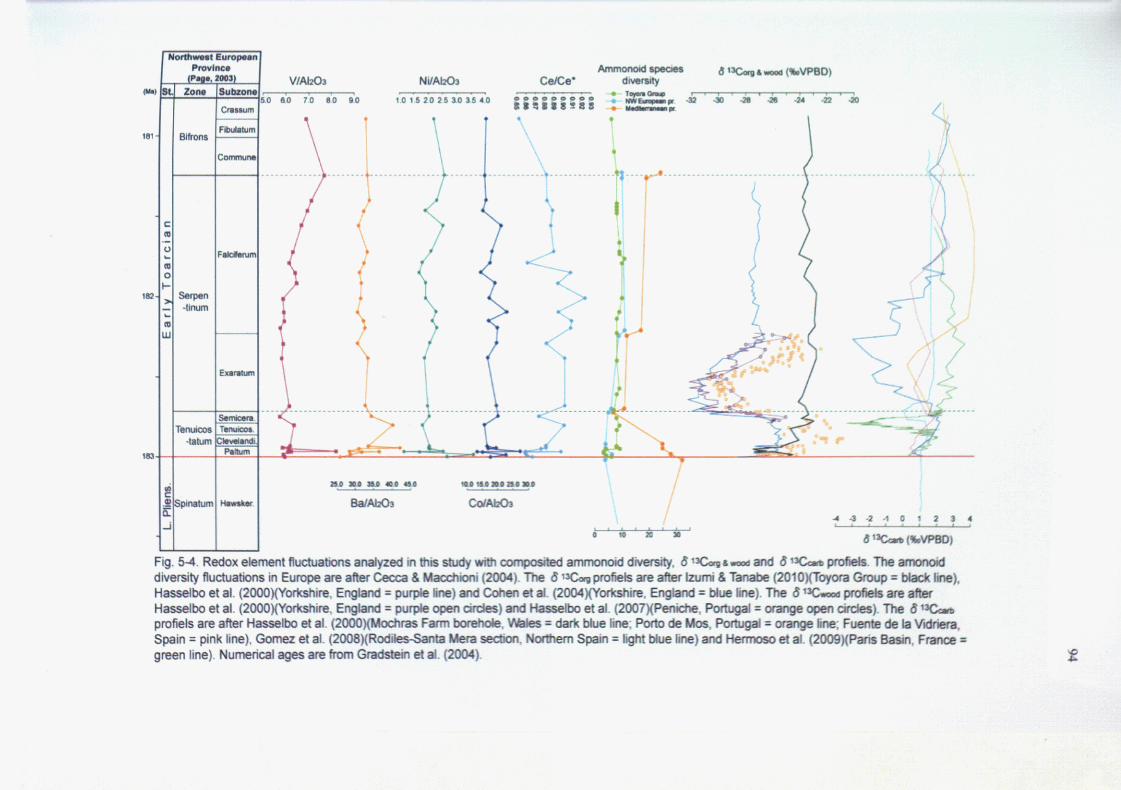
c:cu
0F"- rum
CU0I-
-"-cuW
VlA120J
80 7.'0 alo 9~O
NilA120 3
1.0 15 20 25 0 5" 0
4,

95
are obtained from bed 19-1, correlated to the uppermost part of the P. paltus Zone,
including P. nipponicum (Figs. 1-3 in Plate 2), P. yabei (Fig. 5 in Plate 2), F.
nakayamense (Fig. 6 in Plate 2), P. okadai (Fig. 6 in Plate 3) and "Cleviceras"
chrysanthemum (Figs. 1-3 in Plate 3). These relationships between the marine
redox conditions and the faunal associations of Hildoceratidae (including
Harpoceratinae and Arieticeratinae) probably suggest differences of anoxic
tolerances between these subfamilies.
The diversity falls of ammonoid assemblage in the upper part of the H. inouyei
Zone also seems to be related to the oxygen-depleted marine environment,
distinguished geochemically by a gradual increase of the V concentrations with
SOlne excursions of the Ni and Co concentrations (Fig. 5-4). Therefore, SOITIe minor
diversity fall affected by the dysoxic environments are accompanied during the
recovery of ammonoid diversity from the extinction in the lower part of the P.
paltus Zone.
5-4-3. Early Toarcian OAE in the Northern Hemisphere
Since the geochemical analysis of Jenkyns & Clayton (1986), the Early
Toarcian OAE has been mainly discussed on the basis of the 013C fluctuation in
Europe. Jenkyns & Clayton (1986, 1997) displayed the Ol3 C fluctuation in England
and recognized a positive shift in the Serpentinum Standard Zone following the
negative stage during the Spinatum and the Tenuicostatum Standard Zones. In the
Northwest European province, Hesselbo et al. (2000) analyzed the high-resolution
o13Ccarb, ol3Corg and 013Cwood through the Lower Whitby Mudstone exposed in
Yorkshire Coast, England, and identified an abrupt negative excursion in each of
these fluctuations, biostratigraphically corresponded to the uppermost part of the
Tenuicostatum Standard Zone (Semicelatum Subzone) and the Lower Serpentinum
Standard Zone (lower to middle part of the Exaratum Subzone)(Fig. 5-4). Cohen et
al. (2004) identified the anoxic condition during the uppermost Tenuicostatum
Zone and the Exaratum Subzone in England on the basis of the Ol3 Corg and the Os
isotope profiles, in addition to a negative excursion of the o13 Corg fluctuation in the

96
lower part of the Tenuicostatum Standard Zone (Fig. 5-4). Hermoso et al. (2009)
analyzed the 813Ccarb fluctuation in the southern Paris Basin and recognized that the
negative shift of carbon isotope occurs in four steps during the upper part of the
Tenuicostatum Standard Zone, in addition to a gentle negative excursion of that in
the lower part of the Tenuicostatum Standard Zone. In northern Spain, Gomez et al.
(2009) also recognized a negative excursion of the 813Ccarb fluctuation analyzed in
northern Spain during the upper part of the Semicelatum Subzone and the lower
part of the Elegantulum Zone (Fig. 5-4). As a result, the anoxic marine condition in
the Northwest European province was identified from the uppermost Tenuicostatum
Standard Zone to the Exaratum Standard Subzone of the Serpentinum Standard
Zone.
The negative shifts of 013 C have been also recognized in the Mediterranean
province. According to Hesselbo et al. (2007), negative shifts of the () 13Ccarb and
813Cwood fluctuations analyzed in Peniche, Portugal, were identified in the
lowermost Polymorphum Zone, almost equivalent to the Paltum Subzone of the
standard zonation, and the Lower Levisoni Zone, corresponded to the Exaratum
Standard Subzone (Fig. 5-4). In northern Italy, Bellanca et al. (1999) identified the
oxygen-depleted zone on the basis of the fluctuations of 813Ccarb and the some
redox element concentrations. Based on these analyses, the anoxic event in the
Mediterranean province has been confirmed from the uppermost Polymorphum
Zone to the lower part of the Levisoni Zone. The ammonoid diversity crisis mainly
identified in the Mediterranean province (e. g. Cecca & Macchioni, 2004) was
biostratigraphically corresponded to the Early Toarcian GAE (Fig. 5-4) and
therefore considered to be affected by this marine environmental change.
In the North American Cordillera province, the 813Corg profile was analyzed
along Yakoun River in the Queen Charlotte Islands by Caruthers et al. (2010). The
813Corg fluctuation of Caruthers et al. (2010) shows a dramatic negative excursion
that averages -30.94%0 with increasing of Total Organic Carbon value. These
geochemical anomalies are probably corresponded to the upper part of the Kanense
Zone of the North American Cordillera zonation. Although the detailed

97
biostratigraphic correlation of these curves with the European geochemical results
is disturbed by the poor occurrence of ammonoid fossils in the Yakoun River
succession, the negative shift of 0"c., is almost coeval with the Early Toarcian
GAE distinguished in Europe. The effect of the Early Toarcian OAE seemed to
have reached the coastal areas of North American Cordillera via the Viking
Corridor, a seaway from northwest Europe to the Anui Ocean, or the Hispanic
Corridor, which pass through the Central America. On the other hand, the al3 Core;
curve of North American Cordillera is stable in the lowermost Toarcian, and no
shifts are recognizable in this interval.
The Early Toarcian OAE has been also discussed in the Northern Tibet and the
pelagic area. Chen et al. (2005) displayed the a13Core; and the atomic C/N ratio
profiles, and recognized a negative shift of the a13Core; fluctuation. In the pelagic
area, this event has been identified in East Asia (the Katsuyama Section, central
Japan; Hori et al., 2000) and Slovenia (Gorican et al., 2003) on the basis of the
positive excursions of V/Ah03 ratios and the occurrence of the pelagic black
sediments. However, the detailed biostratigraphic correlation between these events
and the Early Toarcian OAE recognized in European epicontinental successions
and the Toyora Group are difficult because of the low-resolution of the
biostratigraphic age controls in the Northern Tibet and the pelagic successions.
The East Asian Toarcian OAE in this study is recognized in the middle part of
the P. paltus Zone (Fig. 5-4). This event is compared to the Paltum Subzone, the
lowermost subzone of the Toarcian Stage, in the standard zonation. This result
suggests that the OAE in East Asia is corresponded to the oxygen-depleted
condition identified by the al3Ccarb and the al3Core; negative shifts in both of the
Northwest European and the Mediterranean provinces (Cohen et al., 2004;
Hesselbo et al., 2007)(Fig. 5-4). Therefore, the anoxic event recognized in the
lowermost part of the Tenuicostatum Zone enables to regard as a global
paleoenvironmental change in the Tethys and the West Panthalassa. The oxygen
depleted conditions identified in Northern Tibet and the pelagic successions (Chen
et al., 2005; Hori et al., 2000; Gorican et al., 2003) seeln to be corresponded to this

98
earliest Toarcian dysoxic event. In contrast, the maj or Early Toarcian OAE from
the lowermost part of the Tenuicostatum Zone to the Exaratum Subzone,
recognized in Europe and North American Cordillera, is not identified clearly in
the Toyora Group. This means that the anoxic condition in this period was
developed only in the limited area, the western Tethys and the East Panthalassa.

99
6. Systematic paleontology
3-1. Genus Amaltheus
Remark: Almost all of the materials utilized in this analysis are crushed
horizontally to the bedding plane and the preservation is not so well.
Class Cephalopoda CUVIER, 1798
Sub-class Ammonoidea ZITTEL, 1884
Order Psiloceratida HOUSA, 1965
Amended by Dommergues (2002)
Superfamily Eoderoceratoidea NEUMAYR, 1875
Family Liparoceratidae HYATT, 1867
Amended by Dommergues & Meister (1999)
Subfamily Amaltheinae HYATT, 1867
Genus Amaltheus MONTFORT, 1808
Type species: Amaltheus margaritatus MONTFORT, 1808
Remark: The whorl section is compressed and subelliptical. Surface ornaments
are varied; some species have smooth whorl flank, and others are characterized by
the spiny or strong ribbing. Most of species have a crenulated keel.
Amaltheus stokesi (SOWERBY, 1818)
Plate 4, figs. 1-5
1818. Ammonites Stokesi SOWERBY, pI. 190.
1955. Amaltheus cf. nudus (QUENSTEDT). - IMLAY, pI. 10, fig. 5.
1958. Amaltheus stokesi (SOWERBY). - HOWARTH, pI. 1, figs. 5, 7, 12-14; pI. 2, figs.
1, 3, 10; text-figs. 4, 5.
1964. Amaltheus stokesi (SOWERBY). - FREBOLD, pI. 6, figs. 6, 13.
1967. Amaltheus stokesi (SOWERBY). - FREBOLD, et aI., pI. 1, figs. 1,7.
1971. Amaltheus cf. stokesi (SOWERBY). - HIRANO, pI. 17, fig. 11.

100
1974. Amaltheus (Amaltheus) stokesi (SOWERBY). - REPIN, pI. 1, figs. 5, 9.
1988. Amaltheus (Amaltheus) stokesi (SOWERBY). - KRYMHOLTS et aI., pI. 2, fig. 1.
1991. Amaltheus stokesi (SOWERBY). - POULTON, pI. 8, figs. 2, 7, 8.
1992. Amaltheus (Amaltheus) stokesi (SOWERBY). - In: WESTERMANN, pI. 17, figs.
1,3,6.
1992. Amaltheus stokesi (SOWERBY). - In: WESTERMANN, pI. 3, fig. 11; pI. 5, fig. 9;
pI. 6, fig. 4 (cf.).
1994. Amaltheus cf. stokesi (SOWERBY). - PALFY & HART, pI. 1, fig. 12.
1994. Amaltheus stokesi (SOWERBY). - JAKOBS & PALFY, pI. 1, fig. 9.
1996. Amaltheus stokesi (SOWERBY). - SMITH& TIPPER, pI. 19, fig. 1; pI. 20, fig. 3.
1997. Amaltheus stokesi (SOWERBY). - JOHANNSON et aI., pI. 2, fig. 13.
1998. Amaltheus stokesi (SOWERBY). - GECZY& MEISTER, pI. 6, fig. 10.
? 2002. Amaltheus stokesi (SOWERBY). - RAKUS & GUEX, pI. 27, fig. 8.
2003. Amaltheus stokesi (SOWERBY). - MEISTER & FRIEBE, pI. 16, figs. 9, 14.
2006. Amaltheus stokesi (SOWERBY). - FAURE, p. 43, fig. 8.
2006. Amaltheus stokesi (SOWERBY). - TOPCHISHVILI et aI., pI. 11, fig. 4.
2007. Amaltheus stokesi (SOWERBY). - RULLEAU et aI., pI. 34, fig. 5; pI. 35, fig. 1.
2007. Amaltheus stokesi (SOWERBY). - MOUTERDE et aI., pI. 1, figs. 9, 14.
Material: Four specimens, DRF0101, DRF0148, DRF0141 and DRF0209, from
the Kuruma Group, and a specimen, GK. G. 11293, from the Toyora Group.
Diagnosis: A species of the genus Amaltheus with one-to-one connection
between the each ribs and serrations of the crenulated keel.
Dimensions (mm):
No. D Wh U U/D Wh/D
DRFOI0l 47.7 23.0 11.2 0.235 0.482
DRF0148 29.2 14.2 6.0 0.206 0.486
DRF0141 35.2 15.6 9.1 0.259 0.443
GK. G. 11293 41.4+ 20.5 8.2 0.198 0.495

101
Description: The whorls are moderately to quite involute and enlarged rapidly,
therefore the umbilici are narrow (Fig. 6-1). Detailed characters of the whorl
section are unknown because of the compression. The venter bears a broad and
elevated keel with coarse crenulation. The radial and sometime irregular ribs are
somewhat strong in inner two-third of the whorl flank and become slightly weak
toward the venter. They are strongly projected forward at the ventrolateral part and
pass onto the keel as crenulation.
Discussion: This species is mainly distinguished from Amaltheus margaritatus
MONTFOR'!, one of the most typical species of this genus, by the connecting pattern
between the ribs and the crenulations and characters of the keel. The former
species has a round keel, and the ribs are connected with each serration of the
crenulation of keel. On the other hand, a keel of the later species is more individual,
and the crenulation becomes finer than former. In addition, the ribs of the latter
species are not connected with each serration of the crenulation one by one.
Amaltheus sensibilis DAGIS and Amaltheus subbifurcus REPIN, both are
obtained from the Northeast Russian province, are distinguished from the present
species by the larger umbilicus and stronger projection of the ribs at the
ventrolateral part.
The present species lS quite similar with Amaltheus bifurcus HOWARTH.
However, A. bifurcus lS characterized by the more coarse ribbing with a not
developed keel.
A specimen, DRFO143 (Fig. 6 in Plate 4) obtained from the Kuruma Group, has
some affinities with the present species, for example, the connection of some ribs
with the serration of the crenulated keel. However, this specimen is small with not
so well preservation. In this description, the specimen is treated as Amaltheus sp.
cf. A. stokesi (SOWERBY).
Age and distribution: Amaltheus stokesi is an index species of the base of the
Upper Pliensbachian. Therefore, this species characterizes the Stokesi Subzone of
the Margaritatus Zone, Upper Pliensbachian.
The present species has been recognized in the Northwest European province,

25~----------------------------------------.,
..-.EE"'-"....., 20.cC).-.c
-CG.~-:cE
::::>
5
Legend
• 0 Ama/theus stokes!• 0 Ama/theus magaritatusA ~ Ama/theus repressus• o Ama/theus aft. ta/rosei
16 specimens
•
Umbilical width
o 10 20 30
Diameter (mm)
40 50 60
Fig. 6-1. Variation in whorl height and umbilical width of the genus Amaltheus from Japan.

103
the Mediterranean province (Morocco in North Africa), East Asia (Japan), North
American Cordillera (British Columbia and Alaska), Caucasus {Georgia) and
Russia (the Northeast and the Far East provinces).
Amaltheus margaritatus MONTFORT, 1808, form margaritatus MONTFORT, 1808
Plate 1, figs. 9-10; Plate 4, figs. 7-16
1808. Amaltheus margaritatus MONTFORT, p. 90 (fig.), 91.
1812 . Ammonites acutus J. SOWERBY, pI. 17, fig. 1.
1845. Ammonites sedgwickii J. BUCKMAN, p. 40 (nomen nudum).
1852. Ammonites foliaceus GIEBEL, p. 540.
1885 Ammonites amaltheus nudus QUENSTEDT, pI. 41, figs. 1-2.
1885 Ammonites amaltheus compressus QUENSTEDT, pI. 41, fig. 17; pI. 42, fig. 8.
1955. Amaltheus sp. indet. SATO, pI. 18, figs. 1, 2.
1958 Amaltheus margaritatus de MONTFORT, 1808. - HOWARTH, text-fig. 8
(neotype), 9 (lectotype).
1964. Amaltheus stokesi (SOWERBY). - FREBOLD, pI. 6, fig. 7.
1974. Amaltheus margaritatus de MONTFORT. - ELMI et aI., pI. 1, fig. 1.
1976. Amaltheus margaritatus de MONTFORT. - POURMOTAMED & MOTAMED, p. 106,
fig.3.
1988. Amaltheus (Amaltheus) margaritatus de MONTFORT. - KRYMHOLTS et aI., pI. 2,
fig. 5.
1991. Amaltheus stokesi (SOWERBY). - POULTON, pI. 8, fig. 1.
1998. Amaltheus margaritatus de MONTFORT. - GECZY & MEISTER, p. 103 with
synonymy.
1997. Amaltheus stokesi (SOWERBY). - JOHANNSON et aI., pI. 2, fig. 12.
2002. Amaltheus margaritatus de MONTFORT. - FAURE, pI. 7~ fig. 16.
2006. Amaltheus margaritatus de MONTFORT. - TOPCHISHVILI et aI., pI. 11, figs. 1,
2.
2006-2007. Amaltheus margaritatus de MONTFORT. - FAURE, p. 43, figs. 1, 3, 4.

104
2007. Amaltheus margaritatus de MONTFORT. - RULLEAU et aI., pI. 35, figs. 3-5.
2007. Amaltheus margaritatus de MONTFORT. - MOUTERDE et aI., pI. 2, figs. 1, 8.)
2008. Amaltheus margaritatus de MONTFORT forme margaritatus de MONTFORT. -
DOMMERGUES et aI., pI. 10, fig. 4.
Material: Nine specimens, DRF0147, DRF0142, DRF0207, DRF0208,
FMM2007, FMM2008, FMM2009, MM2791 (T5303-1a in Sato, 1955) and
MM2792 (T5303-1 in Sato, 1955), from the Kuruma Group, and three specimens,
SA36-1-1,SA32-1-2 and THM0001, from the Toyora Group.
Dimensions (mm):
No. D Wh D DID Wh/D
DRF0147 33.0 16.6 6.8 0.206 0.503
DRF0142 24.7 11.0 5.5 0.223 0.445
DRF0207 40.2 19.2 9.2 0.229 0.478
FMM2007 41.3 18.1 11.2 0.271 0.438
FMM2008 36.4 13.6 11.4 0.313 0.374
MM2792 35.9 16.6 10.2 0.284 0.462
MM2791 25.2 12.4 5.5 0.218 0.492
SA36-1~1 .33.5 17.4 10.7 0.319 0.519
THMOOOI 41.7 18.6 11.0 0.264 0.446
Description: The whorls are moderately involute and enlarged rapidly, and the
umbilicus are slightly narrow (Fig. 6-1). Detailed characters of the whorl section
are unknown because of the compression. The venter bears an individual and
elevated keel with fine crenulation. The present species from Japan is characterized
by the lateral ribs. On the inner whorl, the radial and slightly sigmoidal ribs started
from umbilical edge are well developed on middle of the flank and are tending to
disappear near the venter, but not completely fade out. They are strongly projected
forward at the upper part of the flank until the keel, but each serration of the
crenulation of the keel are not connected with the ribs. A specimen, DRFO147 (Fig.
7 in Plate 4), has some longitudinal striae on the whorl flank. On the other hand,

105
the ribs become weak and fade out on the outer whorl.
Discussion: On the basis of the recognizable characters of surface ornament
and keel. all of our materials identified as the present species are similar to the
classic Amaltheus from Europe. The present species is distinguished from
Amaltheus stokesi (SOWERBY) by characters of the ribbing and the keel, as
mentioned above.
The specimens identified as the present species from Japan are similar to
Amaltheus margaritatus MONTFORT from the Northeast Russian province described
by Dagis (1976). However, the Northeast Russian specimens have the little more
evolute whorls and the more sick keels.
The present species is similar to Amaltheus conspectus DAGIS but is
distinguished by the more involute whorl and the connection between the serration
of the crenulated keel and the ribs recognized in the latter species.
The present species is also similar with Amaltheus talrosei REPIN, probably
one of the varieties of A. margaritatus. However, A. talrosei is distinguished from
the present species by the more evolute whorl and stronger projection of the ribs at
the ventrolateral part.
The specimens obtained from the Nishinakayama Formation of the Toyora
Group (SA36-1-1, Fig. 9 in Plate 1; SA36-1-2, Fig. 10 in Plate 1) and some
materials from the Teradani Formation (e.g. DRF0207, Fig. 12 in Plate 4) are
characterized by an elevated and individual keel with well prorsiradiated serrations
and the connection of the ribs and the keel, occasionally some ribs and the
serrations of the crenulation. These characters are limited to' the specimens
obtained from the Apyrenum Subzone of the Northwest European province (e.g.
Fig. 3 in Plate 4; Meister, 1988). The Japonica Zone established by Nakada &
Matsuoka (in press) in the Toyora Group was compared to the Apyrenum Standard
Subzone based on these morphological characters.
A specimen, DRF0210 (Fig. 2 in Plate 5) obtained from the Kuruma Group, is
similar to the present species in the characters of the ribbing but is only a large
fragment without the ventral part. Therefore, this specimen is described as

106
Amaltheus sp. in this study.
Age and distribution: This species ranges from the Subnodosus Subzone of the
Margaritatus Zone to the Apyrenum Subzone of the Spinatum Zone in the
Northwest European zonation, compared to the Upper Pliensbachian.
Amaltheus margaritatus was mainly distributed in high latitudinal area of the
Northern Hemisphere, equal to the Boreal Realm in Late Pliensbachian, as follows;
the Northwest European province, East Asia (Japan), North American Cordillera
(British Columbia and Alaska) and Russia (the Northeast and the Far East
provinces). In addition, this species is also recognized in the Mediterranean
province (Algeria and Morocco in North Africa) and Caucasus (Georgia).
Amaltheus repressus DAGIS, 1976
Plate 4, figs. 17-18
1976. Amaltheus (Proamaltheus) repressus DAGIS, pl. 11, figs. 1-5.
/Material: Two specimens, DRF0125 and DRF0202, from the Kuruma Group.
Diagnosis: A species of the genus Amaltheus with constrictions.
Dimensions (mm):
No.
DRF0202
D
27.1
Wh U
12.4 6.8
UID Wh/D
0.251 0.458
Description: The whorls are moderately involute and enlarged rapidly,
therefore the umbilicus are slightly narrow (Fig. 6-1). Details of the whorl section
are unknown. The venter has a slightly weak and crenulated keel, and the
crenulations are very fine and well prorsiradiated. However surface ornaments on
the whorl flank are generally weak, the present species is characterized by the
presence of constrictions. On the small whorl, a specimen of DRF0202 (Fig. 17 in
Plate 4), the radial and irregular ribs started from umbilical edge are well
developed in the inner half of whorl and become weak rapidly for the venter. They
are strongly projected forward at the ventrolateral part and connected with the keel.

107
A slightly sigmoidal constriction IS developed on the whorl flank and is also
strongly projected at the ventrolateral part. On the outer whorl examined In a
specimen DRFO125 (Fig. 18 in Plate 4)~ the ribs become very weak and two
constrictions per 45° of a whorl are developed near the aperture.
Discussion: In previous study, the Russian endemic species of the genus
Amaltheus was only recognized in Alaska of North America by Imlay (195'5)~
Amaltheus viligaensis (TUCHKOV)~ without the Russian province. Therefore, the
occurrences of the present species and Amaltheus sp. aff. A. talrosei REPIN as
mentioned below are the first records of the Russian endemic Amaltheus from Asia.
The present species is similar to Amaltheus margaritatus MONTFORT and
Amaltheus conspectus DAGIS in surface ornaments. However, the former IS
distinguished from the latter two species by the presence of constrictions.
Age and distribution: Amaltheus repressus co-occurred with Amaltheus stokesi
(SOWERBY) in the Northeast Russian province and correlated with the Stokesi
Standard Subzone of the Margaritatus Standard Zone, earliest Late Pliensbachian
(Dagis, 1976).
The present species was distributed in the Northeast Russian province and East
Asia (Japan).
Amaltheus sp. aff. A. talrosei REPIN, 1968
Plate 5, figs. 1a-b
DID Wh/D
0.407 0.328
D
19.6
Wh
15.8
Material: Only a specimen, DRFOI49~ from the Kuruma Group.
Dimensions (mm):
No. D
DRF0149 48.2
Description: The whorl is moderately evolute, and therefore the umbilicus is
slightly large (Fig. 3-1). Details of the whorl section are unknown because of the
compression. However the ventral part is not well preserved, a broad and
crenulated keel is partly recognized in the outer whorl (Fig. 1b in Plate 5)~ and the

108
serrations of the crenulation are coarse and well prorsiradiated. The rursiradiate
and irregular ribs started from umbilical edge are strong near the inner flank and
are tending to disappear from middle to outer flank. They are strongly projected
forward at the ventrolateral part, and some of them pass through onto the
crenulated keel.
Discussion: The present specimen is quite similar to Amaltheus talrosei REPIN,
described only in the Northeast Russian province by Repin (1968) and Dagis
(1976), in the evolute coiling, well prorsiradiated serration of the crenulated keel
and the connection of the ribs and the serration of the keel. However, the former
has more rigid and irregular ribbing than the latter. Consequently, the present
specimen is identified as Amaltheus sp. aff. A. talrosei REPIN in this description.
The present specimen is also similar to Amaltheus margaritatus MONTFORT and
Amaltheus viligaensis (TUCHKOV) in characters of the ribbing but is distinguished
by the more evolute whorl and the stronger projection of the ribs at the
ventrolateral part.
Age and distribution: In Russian, A. talrosei was obtained from the Viligaensis
Zone, equivalent to the Spinatum Zone of the Northwest European zonation (Dagis,
1976; Krymholts et al., 1988). However, the base of the Viligaensis Zone is
compared into the ~ Margaritatus Standard Zone because of the occurrence of
Arieticeras sp. aff. A. algovianum (OPPEL), ranges into the Subnodosus-Apyrenum
Standard Zone, from the Viligaensis Zone (Fig. 3-6). In addition to the zonal
correlation, the present specimen is co-occurred with Amaltheus margaritatus
MONTFORT from the Kuruma Group. Consequently, the range of the present species
is corresponded to the upper part of the Margaritatus Zone of the Northwest
European zonal shame (Fig. 3-6).
The present specimen was distributed in East Asia (Japan).

109
7. Conclusions
7-1. Ammonoid biostratigraphy
The ammonoid zonal scheme of the Toyora Group is revised in this study, and
six zones are proposed (in ascending order) as follows; the A. stokesi Assemblage
Zone, the Canavaria japonica Zone, the Paltarpites paltus Zone, the Dactylioceras
helianthoides Zone, the Harpoceras inouyei Zone and the Pseudogrammoceras
Phlyseogrammoceras Assemblage Zone.
In contrast, three ammonoid zones are established in the Teradani Formation,
the Kuruma Group, as follows; the Stokesi-Repressus Assemblage Zone, the
Margaritatus-aff. Talrosei Assemblage Zone and the Canavaria-Emaciaticeras
Assemblage Zone in ascending-order.
These zonations are corresponded to the zonal schemes of outside Japan,
especially the Northwest European standard zonation (Fig. 3-6). The A. stokesi
Assemblage Zone and the Stokesi-Repressus Assemblage Zone are compared to the
Stokesi Standard Subzone of the Margaritatus Standard Zone (Late Pliensbachian).
The Margaritatus-aff. Talrosei Assemblage Zone IS correlated to the
Subnodosus-Gibbosus Standard Subzones (Margaritatus Standard Zone) and the
Apyrenum Standard Subzone (Spinatum Standard Zone, Late Pliensbachian). The
Canavaria-Emaciaticeras Assemblage Zone is corresponded to the Hawskerense
Standard Subzone of the Spinatum Standard Zone. The base of the C. japonica
Zone is roughly correlated to the Apyrenum Standard Subzone. The base of the P.
paltus Zone is equivalent to the Pliensbachian/Toarcian boundary. The base of the
D. helianthoides Zone is compared to the base of the Paltum Standard Subzone
(Tenuicostatum Standard Zone, Early Toarcian). The base of the H. inouyei Zone is
corresponded to the Exaratum Standard Subzone of the Serpentinum Standard Zone,
and the upper limit seems to be corresponded to the Fibulatum Standard Subzone
(Bifrons Standard Zone, Early Toarcian).

110
7-2. Ammonoid paleobiogeography
The transitional patterns during late Early Jurassic time-interval (Late
Pliensbachian-Toarcian) and their paleobiogeographic correlation of a
comprehensive Japanese ammonoid fauna are analyzed by utilizing the ammonoid
assemblages from the Toyora and the Kuruma Groups.
During the Stokesi Standard Zone, the assemblage from the Kuruma Group,
characterized by the Russian endemic species of the genus Amaltheus, shows the
high similarity with the Northeast Russian fauna, although the coeval assemblage
from the Toyora Group is associated with the Boreal and the Tethyan elements.
This faunal disagreement reflects the paleolatitudinal interval between these
groups. The assemblage of the Subnodosus-Gibbosus Standard Subzone from the
Kuruma Group is continuously characteristic in the Boreal-derived fauna. The
faunal association of the Apyrenum Standard Subzone from the Kuruma Group, the
mixing of the Tethyan and the Boreal elements, is not recognized in the Northern
Hemisphere. The faunas from the Hawskerense Standard Subzone and the
Tenuicostatum Standard Zone, dominated by Arieticeratinae and Harpoceratinae,
are similar with those of the Mediterranean province. The fauna in the
Serpentinum-Lower to Middle Bifrons Standard Zones from the Toyora Group is
characterized by the mixing of the Tethyan and the Boreal faunas and shows the
high similarity with the Northwest European fauna. The generic faunal association
of the Fallaciosum Standard Subzone and the Dispansum Standard Zone is similar
with that from the Causses Basin in the Northwest European province.
As a result, a turnover from the Boreal fauna composed of the genus Amaltheus
(Amaltheidae) to the Tethyan fauna dominated by Arieticeratinae is distinguished
mainly in the Kuruma Group during the Apyrenum Standard Subzone (Fig. 4-1).
This faunal change is chronologically discordant for the comparable faunal
turnover in the almost equal paleolatudinal areas of the Northwest European
province, which is corresponded to the PIT boundary. The turnover recognized in
the latest Pliensbachian ammonoid fauna from Japan seems to be mainly affected
by the regression during latest Pliensbachian (Spinatum Standard Zone) to earliest

111
Toarcian (Tenuicostatum Standard Zone), in addition to the starting of the warming
event (Fig. 4-9).
In contrast, a faunal mixing of the Boreal and the Tethyan faunas is recognized
in the assemblage from the Toyora Group during the Exaratum Standard Subzone
(Fig. 4-1). This bio-event enables to correspond to the coeval faunal conformity in
the Northwest European and the Mediterranean provinces. The faunal mixing in the
middle Early Toarcian ammonoid assemblage is perhaps reflected by the abrupt
transgression during Early Toarcian, with the slight cooling event (Fig. 4-9).
7-3. Geochemical analyses of the Early Toarcian OAE
The concentrations of trace elements and the anomalies of REE are analyzed in
the Sakuraguchidani Mudstone Member exposed along the Sakuraguchidani Valley,
the Toyora Group. The positive excursions of V/Ah03, Ba/Ab03 and Co/Ah03 are
recognized in bed 18-1c, the middle part of the P. paltus Zone (earliest Toarcian),
with a negative shift of NilAh03 and a positive Ce anomaly (Figs. 5-1, 5-2). These
geochemical anomalies suggest the oxygen-depleted paleoceanographic conditions,
biostratigraphically corresponded to the coeval minor anoxic environment in the
Northwest European province (Fig. 5-4). The major Early Toarcian OAE from the
uppermost Tenuicostatum Standard Zone to the Exaratum Standard Subzone
recognized in Europe and North American Cordillera is not identified in the Toyora
Group. Thus, the anoxic water mass affected by this event was developed only in
the West Tethys and East Panthalassa.
Moreover, a diversity fall of ammonoid assemblage is also identified in bed
18-1. This diversity crisis is characterized by the extinction of Harpoceratinae, and
the survival of only a species of pandemic Arieticeratinae, Fontanelliceras
fontanellense. Consequently, the ammonoid diversity crisis in the middle part of
the P. paltus Zone is perhaps reflected by the anoxic event in East Asia. Moreover,
the relationships between the marine redox conditions and the faunal associations
of Hildoceratidae probably suggest the difference of anoxic tolerances between
Harpoceratinae and Arieticeratinae.

112
In addition to the faunal transitions affected by the changes of eustatic sea
level and seawater temperature, an ammonoid bioevent reflected by the marine
redox condition can be identified during earliest Toarcian (Tenuicostatum Standard
Zone).

113
Acknowledgements
I gratefully thank Prof. Atsushi Matsuoka (Niigata University) for his general
guidance for this study. I would like to thank Dr. Christian Meister (Natural
History Museum of the City of Geneva) for their kind guidance on the
classification and identification of ammonoids with many valuable comments about
ammonoid taxonomy, biostratigraphy and paleobiogeography. I also thank Mr.
Ken-ichi Miyakita (Niigata City), Mr. Kaku Nagata (Niigata City), Mr. Kazunobu
Terabe (Arabian Oil Company), Mr. Hideo Ishida (Mine City), Mr. Yoshihiko Kubo
(Nagano City) and Mr. Takahiro Aoki (Nagano City) for offering many ammonoid
specrmens.
I am grateful to Prof. Jean Guex (University of Lausanne), Prof. Jiarun Yin
(China University of Geosciences, Beijing), Dr. Naoki Watanabe (Niigata
University), Prof. Kazushige Tanabe (the University of Tokyo), Prof. Hiromichi
Hirano (Waseda University), Dr. Yasuhiro Iba (National Museum of Nature and
Science), Dr. Alain Morard (University of Fribourg) and Dr. Tadashi Sato (Fukada
Geological Institute) for their valuable comments. I also thank Prof. Jun-ichi
Tazawa, Dr. Isao Niikawa, Dr. Toshiyuki Kurihara, Dr. Naoto Ishida, Mr. Takashi
Nikaido and the other members of the Historical Earth Science Seminar in Niigata
University for their helpful suggestions. I wish to thank Dr. Takenori Sasaki (the
University Museum, the University of Tokyo), Prof. Hiroyoshi Sano (Kyushu
University), Dr. Akihiro Misaki (Kitakyushu Museum of Natural History and
Human History) and Mr. Yosuke Ibaraki (Fossa Magna Museum) for their kind
assistance in observation of specimens. Mr. Koshiro Takanasi (Niigata University)
is thanked for their guidance for the geochemical analyses. I thank Mr. Takayoshi
Fukutomi (Shimonoseki City), Mr. Keisuke Kawano (the Firefly Museum of Toyota
Town), Ms. Junko Masahara (Shimonoseki City) and Tenkyoji Youth Hostel for
their kind helps during my field work. The Asahi Townhouse is thanked for their
cooperation in the field. I express my gratitude to these persons.

114
References
Aberhan, M., Fursich, T.F., 1997. Diversity analysis of Lower Jurassic bivalves of
the Andean Basin and the Pliensbachian-Toarcian mass extinction. Lethaia, 29,
181-195.
Aberhan, M., Fursich, T.F., 2000. Mass origination versus mass extinction: the
biological contribution of the Pliensbachian-Toarcian extinction event. Journal
of the Geological Society, 157,55-60.
Arias, C., 2006. Northern and Southern Hemispheres ostracod palaeobiogeography
during the Early Jurassic: possible migration routes. Palaeogeography,
Palaeoclimatology, Palaeoecology, 233, 63-95.
Arias, C., 2006. Pliensbachian-Toarcian ostracod biogeography in NW Europe:
Evidence for water mass structure evolution. Palaeogeography,
Palaeoclimatology, Palaeoecology, 251, 398-421
Arias, C., 2008. Palaeoceanography and biogeography In the Early Jurassic
Panthalassa and Tethys Oceans. Gondwana Research, 14, 306-315.
Arkell, W.J., 1956. Jurassic geology of the World. Oliver and Boyd Ltd., 806p.
Bailey, T.R., Rosenthal, Y., McArthur, J.M., van de Schootbrugge, B., Thirlwall, M.
F., 2003. Paleoceanographic changes of the Late Pliensbachian- Early Toarcian
interval: a possible link to the genesis of an Oceanic Anoxic Event. Earth and
Planetary Science Letters, 212, 307-320.
Bellanca, A., Masetti, D., Neri, R., Venzia, F., 1999. Geochemical and
sedimentological evidence of productivity cycles recorded in Toarcian black
shales from the Belluno Basin, Southern Alps,Northern Italy. Journal
ofSedimentary Research, 69, no. 2, 466-476.
Braga, J.C., 1983. Ammonites del Domerense de la Zona Subbetica (Cordilleras
Beticas, sud de Espafia). 410p, Unpublished doctoral thesis, Universidad de
Granada, Espana.
Braga, lC., Comas Rengifo, M.J., Goy, A., Rivas, P., 1982. Comparaciones
faunisticas y correlaciones en el Pliensbachiense de la Zona subbetica,

115
Cordillera iberica. Boletln de la Real Sociedad Espanola de Historia Natural
(Geologia), 80,221-244.
Breit, G.N., Wanty, R.B., 1991. Vanadium accumulation in carbonaceous rocks. A
review of geochemical controls during deposition and diagenesis. Chemical
Geology, 91, 83-97.
Buckman, J., 1845. In: Murchison, R.I. (Ed.), Outline of the geology of the
neighbourhood of Cheltenham. 1-109.
Caruthers, A.H., Grocke, D.R., Smith, P.L., 2010. The Early Toarcian Oceanic
Anoxic Event in Western Canada. Short papers for the 8th International
congress on the Jurassic System, 338-339.
Cecca, F., Macchioni, P., 2004. The two Toarcian (Early Jurassic) extinction events
in ammonoids. Lethaia, 37, 35-56.
Chen, L., Yi, H., Hu, R., Zhong, H., Zou, Y., 2005. Organic Geochemistry of the
Early Jurassic Oil Shale from the Shuanghu Area in Northern Tibet and the
Early Toarcian Oceanic Anoxic Event. Acta Geologica Sinica, 79, no. 3,
392-397.
Chihara, K., Komatsu, M., Uemura, T., Hasegawa, Y., Shiraishi, S., Yoshimura, T.,
Nakamizu, M., 1979. Geology and tectonics of the Omi-Renge and the Joetsu
tectonic belts (5): Geology and tectonics of the Omi-Renge tectonic belt.
Science Reports ofNiigata University, Series E, no. 5, 1-61.
Cohen, A.S., Coe, A.L., Harding, S.M., Schwark, L., 2004. Osmium isotope
evidence for the regulation of atmospheric CO2 by continental weathering.
Geology, 32, 157-160.
Cope, J.C.W., Getty, T.A., Howarth, M.K., Morton, N., Torrens, H.S., 1980. A
correlation of Jurassic Rocks in the British Isles Part One: Introduction and
Lower Jurassic. Geological Society ofLondon, Special reports, no. 14, 73p.
Cope, J.C.W., 1998. Discussion on estimates of the amount and rate of sea-level
change across the Rhaetian-Hettangian and Pliensbachian-Toarcian boundaries
(latest Triassic to early Jurassic), Journal of the Geological Society, London,
155, 421-422.

116
Cuvier, G., 1795. Tableau elementaire de l'histoire naturelle des animaux. Paris.
Dagis, A.A., 1968. Toarcian Ammonites (Dactylioceratidae) of North Siberia.
Moscow: Nedra. (in Russian)
Dagis, A.A., 1976. Late Pliensbachian ammonites (Amaltheidae) of the North
Siberia. Academy of Science of the USSR, Siberian Branch, 309, 77p. (in
Russian)
Dagis, A.A., Dagis, A.S., 1965. On zonal subdivision of Toarcian deposits in the
North-East of USSR. In: Stratigraphy and Paleontology of Mesozoic Deposits
of Siberia, Moscow: Nauka, 15-26. (in Russian)
Damborenea, S.E., 2002. Jurassic evolution of Southern Hemisphere manne
palaeobiogeographic units based on benthonic bivalves. Geobios, Memoire
special, no. 24,51-71.
Dean, W.T., Donovan, D.T., Howarth, M.K., 1961. The Liassic ammonite zones and
subzones of the North-West Europe Province. Bulletin of the British Museum
(Natural History), Geology, 4, 435-505.
De Graciansky, P.,-C., Jacquin, T., Hesselbo, S.P., 1998. The Ligurian cycle: an
overview of Lower Jurassic 2nd-order transgressive/regressive facies cycles in
western Europe. In: de Graciansky, P.-C., Hardenbol, J., Jacquin, T., Vail, P.R.
(Eds.), Mesozoic and Cenozoic Sequence Stratigraphy of European Basins.
Society for Sedimentary Geology, Special Publication, 60, 467-479.
Dera, G., Pellenard, P., Neige, P., Deconinck, J.-F., Puceat, E., Dommergues, J.-L.,
2009a. Distribution of clay minerals in Early Jurassic Peritethyan seas:
Palaeoclimatic significance inferred from multiproxy comparisons.
Palaeogeography, Palaeoclimatology, Palaeoecology, 271, 39-51.
Dera, G., Puceat, F., Pellenard, P., Neige, P., Delsate, D., 2009b. Water mass
exchange and variations in seawater temperature in the NW Tethys during the
Early Jurassic: Evidence from neodymium and oxygen isotopes of fish teeth
and belemnites. Earth and Planetary Science Letters, 286, 198-207.
Dera, G., Neige, P., Dommergues, J.-L., Fara, E., Laffont, R., Pellenard, P., 2010.
High-resolution dynamics of Early Jurassic marine extinctions: the case of

117
Pliensbachian-Toarcian ammonites (Cephalopoda). Journal of the Geological
Society, London, 167, 21-33.
Dommergues, J.L., 1979. Le Carixien Bourguignon. 195p., Unpublished thesis
(These 3eme Cycle), L'Universite de Dijion, France.
Dommergues, J.-L., 2002, Les premiers Lytoceratatoidea du Nord-Ouest de
l'Europe (Ammonoidea, Sinernurien inferieur, France). Exemple de
convergence evolutive vers les morphologies "capricornes". Revue de
Paleobiologie, 21, no. 1,257-277.
Dommergues, J.L., Meister, C., 1987. La biostratigraphie des ammonites du
Carixien (Jurassique inferieur) d'Europe occidentale: un test de la methode des
associations unitaires. Eclogae Geologicae Halvetiae, 80, 919-938.
Dommergues, J.-L., 1987. L'evolution chez les Ammonitina du Lias moyen
(Carixien, Domerien basal) en Europe occidentale. Documents des
Laboratoires de Geologie de Lyon, 98, 1-297.
Dommergues, J.L., Meister, C., Mouterde, R., 1997a. Pliensbachian. 3rd
International Symposium on Jurassic stratigraphy (Poitiers 1991), Resumes,
126.
Dommergues, J.L., Meister, C., Schirolli, P., 1997b. Les successions des
ammonites du Sinemurien superieur au Toarcien basal dans les Prealpes de
Brescia (Italie). Memorie di Scienze Geolologiche, 49, 1-26.
Dommergues, J.-L., Meister, C., 1999. Cladistic formalisation of relationships
within a superfamily of Lower Jurassic Ammonitina: Eoderocerataceae. Revue
Paleobiologie 18, 1, 273-286.
Dommergues, J.-L., Meister, C., Bonneau, M., Poisson, A., Vrielinck, B., 2005. Les
ammonites pliensbachiennes des nappes Lyciennes (Turquie meridionale).
Description de faunes nouvelles, implications biostratigraphiques et
paleobiogeographiques. Geobios, 38, 407-435.
Dommergues, J.-L., Dugue, 0., Gauthier, H., Meister, C., Neige, P., Raynaud, D.,
Savary, X., Trevisan, M., 2008. Les ammonites du Pliensbachien et du
toarciuen basal dans la carriere de la Roche Blain (Fresnay-Ie-Puceux,

118
Calvados, Basse-Normandie, France). Taxonomie, implications
stratigraphiques et paleogeographiques. Revue deiPaleobtologie, 27, 1,
265-329.
Donovan, D.T., 1958. The ammonite zones of the Toarcian (Ammonitico Rosso
facies) of southern Switzerland. Eclogae Geologicae Helvetiae, 51, 33-60.
D' Orbigny, A., 1842-1849. Paleontologic francaise; terrains jurassiques 1:
Cephalopodes, 642p. Paris: Masson.
Dymond, J., Suess, E., Lyle, M., 1992. Barium in deep-sea sediments: a
geochemical proxy for paleoproductivity. Paleoceanography, 7, 163-181.
Eggins, S.M., Woodhead, J.D., Kinsley, L.P.J., Mortimer, G.E., Sylvester, P.,
McCulloch, M.T., Hergt, J.M., Handler, M.R., 1997. A simple method for the
precise determination of > 40 trace elements in geological samples by ICPMS
using enriched isotope internal standardization. Chemical Geology, 134,
311-326.
EI Hariri, K., Dommergues, J.-L., Meister, C., Souhel, A., Chafiki, D., 1996. Les
Ammonites du Lias inferieur et moyen du Haut-Atlas de Beni Mellal (Maroc);
taxonomie et biostratigraphie ahaute resolution. Geobios, 29, 537-576.
Elmi, S., Atrops, F., Mangold, C., 1974. Les zones d'ammonites du
Domerien-Callovien de I' Algeria occidentals 1: Domerien-Toarcien.
Documents des Laboratoires de Geologie de la Faculte des Science, 61, 1-84.
Lyon: L'Universite Claude Bernard.
Elmi, S., Gabilly, J., Mouterde, R., Rulleau, L., 1991. Toarcien. 3rd International
Symposium on Jurassic stratigraphy (Poitiers 1991), Resumes, 127.
Elmi, S., Gabilly, J., Mouterde, R., Rulleau, L., Rocha, R.B., 1994. L'etage
Toarcien de I'Europe et de la Tethys; divisions et correlations. Geobios
Memoire Special, 17, 149-159.
Elmi, S., Rulleau, L., Gabilly, J., Mouterde, R., 1997. Toarcien. In: Cariou, E.,
Hantzpergue, P. (Eds.), Groupe Francais d'Etude du Jurassique.
Biostratigraphie du Jurassique ouest-europeen et mediterraneen: zonations
paralleles et distribution des invertebres et microfossiles. Bulletin du Centre de

119
Recherches. ElfExploration Production Memoires, 17, 120-125.
Elmi, S., Mouterde, R., Rocha, R.B., Rulleau, L., 2007. Dne succession de
reference pour le Toarcien Moyen et Superieur: les "Margas calcarias de Sao
Giao" dans les environs de Cantanhede (sous-bassin nord lusitanien, Portugal).
Ciencias da Terra (UNLj, no. 16, 113-133.
Faraoni, P., Marini, A., Pallini, A., Venturi, F., 1994. Nuove faune ad ammonite
delle Zone a E. mirabilis ed H. serpentinus nella Valle del F. Bosso (PS) e loro
riflessi sulla biostratigrafia del limite Domeriano-Toarciano in Appennino.
Studi Geologici Camerti, vol. spec. Biostratigrafia dell' Italia Centrale,
247-197.
Faure, P., 2002. Le Lias des Pyrenees. Strata, Serie, 2,39, 1-761.
Faure P., 2006. Le Domerien (Pliensbachien superieur) des Corbieres (Aude,
France) Biostratigraphie, evolution sedimentaire, paleogeographie. Bulletin de
la Societe d'Etudes scientifiques de l'Aude Clll, 29-44.
Faure P., Almeras, Y., Sekatni, N., Zargouni, F., 2007. Le Pliensbachien de Jebel
Zaghouan (Tunisie). Nouvelles donnees fauniques. Implications
biostratigraphiques et paleobiogeographiques, Geodiversitas, 29, no. 4,
473-506.
Frebold, H., 1964. Lower Jurassic and Baj ocian Ammonoid faunas of Northwestern
British Columbia and Southern Yukon. Geological Survey of Canada, 116, 31
p.
Frebold, H., Mountjoy, E.W., Tempelman-Kluit, D.J., 1967. New occurrence of
Jurassic Rocks and Fossils in Central and Northern Yukon Territory.
Geological Survey ofCanada, paper 67-12.
Frebold, H., Mountjoy, E.W., Tempelman-Kluit, D.J., 1967. New occurrences of
Jurassic rocks and fossils in Central and Northern Yukon Territory. Geological
Survey of Canada, 67, no. 12, 28 p.
Frebold, H., 1970. Pliensbachian Ammonoids from British Columbia and southern
Yukon. Canadian Journal ofEarth Science, 7, no. 2, 435-456.
Fursich, F.T., Berndt, R., Scheuer, T., Gahr, M., 2001. Comparative ecological

120
analysis of Toarcian (Lower Jurassic) bentic faunas from southern France and
east-central Spain. Lethaia, 34, 169-199.
Gabilly, J., 1976. Le Toarcien a Thouars et dans le Centre-Ouest de la France,
evolution de la fauna a Harpoceratinae, Hildoceratinae. Editions du Centre
National de la Recherches Scientifiques. Les Stratotypes francais, 3, 1-217.
Gabilly, J., Elmi, S., Mattei, J., Mouterde, R., 1971. Les zones du Jurassique en
France: Toarcien. Bulletin de la Societe geologique de France, 2, 82-84.
Gabilly, J., Elmi, S., Mattei, J., Mouterde, R., Rioult, M., 1974. L'etage Toarcien
zones et sous-zones d'Ammonites. In: Colloque du Jurassique a Luxembourg
1967, Memoires du Bureau de Recherches Geologiques et Minieres, 75,
605-634.
Geczy, B., Meister, C., 1998. Les ammonites du Domerien de la montagne du
Bakony (Hongrie). Revue de Paleobiologie Geneve, 17,1,69-161.
Gendron, A., Silverberg, N., Sundby, B., Lebel, J., 1986. Early diagenesis of
cadmiumandcobalt in sediments of the Laurentian Trough. Geochimica et
Cosmochimica Acta, 50,741-747.
Giebel, C.G., 1852. Fauna der Vorwelt, Band 3, Die Cephalopoden. 856p, Leipzig.
G6mez, J.J., Goy, A., Canales, M.L., 2008. Seawater temperature and carbon
isotope variations in belemnites linked to mass extinction during the Toarcian
(Early Jurassic) in Central and Northern Spain. Comparison with other
European sections. Palaeogeography, Palaeoclimatology, Palaeoecology, 258,
28-58.
Golonka, J., 2007. Late Triassic and Early Jurassic palaeogeography of the world.
Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 297-307.
Golonka, J., Bocharova, N.Y., Ford, D., Edrich, M.E., Bednarczyk, J., Wildharber,
J., 2003. Paleogeographic reconstructions and basins development of the Arctic.
In: Golonka, J. (Ed.), Thematic Set on Paleogeographic Reconstruction and
Hydrocarbon Basins: Atlantic, Caribbean, South America, Middle East,
Russian Far East, Arctic. Marine and Petroleum Geology, 20, 211-248.
Gorican, S., Smuc, A., Baumgartneret, B.O., 2003. Toarcian Radiolaria from Mt.

121
Mangart (Slovenian-Italian border) and their paleoecological implications.
Marine Micropaleontology, 49, 275-30l.
Gradstein, P.M., Ogg, J.G., and Smith, A.G., Agterberg, F.P., Bleeker, W., Cooper,
R.A., Davydov, V., Gibbard, P., Hinnov, L.A., House, M.R., Lourens, L.,
Luterbacher, H.P., McArthur, J., Melchin, M.J., Robb, L.J., Shergold, J.,
Villeneuve, M., Wardlaw, B.R., Ali, J., Brinkhuis, H., Hilgen, F.J., Hooker, J.,
Howarth, R.J., Knoll, A.H., Laskar, J., Monechi, S., Plumb, K.A., Powell, J.,
Raffi, 1., Rohl, D., Sadler, P., Sanfilippo, A., Schmitz, B., Shackleton, N.J.,
Shields, G.A., Strauss, H., Van Dam, J., van Kolfschoten, T., Veizer, J., Wilson,
D., 2004. A Geologic Time Scale 2004. Cambridge University Press, 589p.
Guex, J., 1973. Apercu biostratigraphique sur Ie Toarcien inferieur du Moyen-Atlas
morocain et discussion sur la zonation de ce sous-etage dans les series
mediterraneennes. Eclogae Geologicae Helvetiae, 66, 493-523.
Guex, J., 1975. Description biostratigraphique du Toarcien superieur de la bordure
sud des Causses (France). Eclogae Geologicae Helvetiae, 68, 97-129.
Guex, J., Morard, A., Bartolini, A., Morettini, E., 2001. Decouverte d'une
importante lacune stratigraphique it la limite Domerieu-Toarcien: implications
paleo-oceanographiques. Bulletin de la Societe Vaudoise des Sciences
Naturelles, 345, 277-284.
Hallam, A., 1975. Jurassic Environments. Cambridge Univ, Press, Cambridge, 269
p.
Hallam, A., 1977. Jurassic bivalve biogeography. Paleobiology, 3, 58-73.
Hallam, A., 1978. Eustatic cycles In the Jurassic. Palaeogeography,
Palaeoclimatology, Palaeoecology, 23, 1-32.
Hallam, A., 1983. Early and mid-Jurassic molluscan biogeography and the
establishment of the Central Atlantic Seaway. Palaeogeography,
Palaeoclimatology, Palaeoecology, 43, 181-193.
Hallam, A., 1988. A re-evolution of Jurassic eustasy in the light of new data and
the revised Exxon curve. In: Wilgus, C.K., Hastings, B.S., Kendall, C.G.St.C.,
Posamatir, H.W., Ron, C.A., van Wagner, J.C. (Eds.), Sea-Level Changes. An

122
Intergated Approaches. Society for Sedimentary Geology, Special Publication,
42, 261-273.
Hallam, A., 1996. Recovery of the marine fauna in Europe after the end-Triassic
and early Toarcian mass extinction. In: Hart, M.B. (Ed.), Biotic recovery from
mass extinction events. Geological Society, Special Publication, 102, 231-236.
Hallam, A., 1997. Estimates of the amount and rate of sea-level changes across the
Rhaetian-Hettangian and Pliensbachian-Toarcian boundaries (latest Triassic to
early Jurassic). Journal of the Geological Society, London, 154,773-779.
Hallam, A., 2001. A review of the broad pattern of Jurassic sea-level changes and
their possible causes in the light of current knowledge. Palaeogeography,
Palaeoclimatology, Palaeoecology, 167, 23-37.
Hallam, A., Wignall, P., 1997. Mass Extinction and Their Aftermass. Oxford
University Press, Oxford, 320p.
Haq, B.U., Hardenbold, J.L., Vail, P.R., 1988. Mesozoic and Cainozoic
chronostratigraphy and cycles of sea-level changes. Sea-Level changes. Society
ofEconomic Palaeontologist and Mineralogist, 42, Special Publication, Tulsa,
Oklahoma, 71-108.
Hardenbold, J., Thierry, J., Farley, M.B., Jacquin, T., de Graciansky, P.-C., Vail, P.,
1998. Jurassique Sequence Chronostratigraphy, Chart 6. In: Graciansky P.-C.,
Hardenbold, J., Jacquin, T., Vail, P., Farley, M.B. (Eds.), Mesozoic and
Cenozoic Sequence Stratigraphy of European Basins. Mesozoic and Cenozoic
Sequence Stratigraphy of European Basins. Society for Sedimentary Geology,
Special Publication, 60, 3-13.
Harries, P.J., Little, C.T.S., 1999. The Early Toarcian (Early Jurassic) and the
Cenomanian-Turonian (Late Cretaceous) mass extinctions: similarities and
contrasts. Palaeogeography, Palaeoclimatology, Palaeoecology, 154, 39-66.
Hatch, J.R., Leventhal, J.S., 1992. Relationship between inferred redox potential of
the depositional environment and geochemistry of the Upper Pennsylvanian
(Missourian) Stark Shale Member of the Dennis Limestone, Wabaunsee County,
Kansas, U. S.A. Chemical Geology, 99, 65-82.

123
Hayami, I., 1958. Taxonomic notes on Cardinia with description of a new species
from the Lias of western Japan. Journal of the Faculty of Science, the
University of Tokyo, sec. 2,11, pt. 2,115-130.
Hayami, I., 1959. Lower Liassic Lamellibranch fauna of the Higashinagano
Formation in West Japan. Journal of the Faculty of Science, the University of
Tokyo, sec. 2, 12, pt. 1, 31-84.
Hayami, I., 1960a. Lower Liassic gastropods from the Higashinagano Formation in
West Japan. Japanese Journal of Geology and Geography, 31, nos. 2-4,
99-106.
Hayami, I., 1960b. Jurassic inoceramids in Japan. Journal of the Faculty ofScience,
the University of Tokyo, sec. 2, 12, pt. 2, 277-328.
Hayami, I., 1961. On the Jurassic pelectypod faunas In Japan. Journal of the
Faculty ofScience, the University of Tokyo, sec. 2, 13, pt. 2, 243-343.
Hayami, I., 1962. Jurassic Pelecypod faunas in Japan with special reference to their
stratigraphical distribution and biogeographical provinces. Journal of the
Geological Society ofJapan, 68, 96-108. (in Japanese, with English abstract)
Heggie, D., Lewis, T., 1984. Cobalt in pore waters of marine sediments. Nature,
311, 453-455.
Hermoso, M., Le Callonnec, L., Minoletti, F., Renard, M., Hesselbo, S.P., 2009.
Expression of the Early Toarcian negative carbon-isotope excursion in
separated carbonate microfractions (Jurassic, Paris Basin). Earth and
Planetary Science Letters, 277, 194-203.
Hesselbo, S.P., Grocke, D.R., Jenkyns, H.C., Bjerrum, C.J., Farrimond, P., Bell,
H. S.M., Green, O.R., 2000. Massive dissociation of gas hydrate during a
Jurassic oceanic anoxic event. Nature, 406, 392-395.
Hesselbo, S.P., Jenkyns, H.C., Duarte, L.V., Oliveira, L.C.V., 2007. Carbon-isotope
record of the Early Jurassic (Toarcian) Oceanic Anoxic Event from fossil wood
and marine carbonate (Lusitanian Basin, Portugal). Earth and Planetary
Science Letters, 253, 455-470.
Hetzel, A., Bottcher, M.E., Wortmann, D.G., Brumsack, H.-J., 2009. Paleo-redox


125
Howarth, M.K., 1958. A Monograph of the Ammonites of the Liassic Family
Amaltheidae in Britain. Palaeontographical Society: Part 1 (1957), 111, 1-26;
Part 2, 112, 27-53.
Howarth, M.K., 1973. The stratigraphy and ammonite fauna of the Upper Liassic
Grey Shales of the Yorkshire Coast. Bulletin of the British Museum (Natural
History), Geology, 24, 235-277.
Howarth, M.K., 1978. The stratigraphy and ammonite fauna of the Upper Lias of
Northamptonshire. Bulletin of the British Museum (Natural History), Geology
Series, 29, 235-288.
Howarth, M.K., 1980. Toarcian. In: Cope, J.C.W., Getty, T.A., Howarth, M.K.,
Morton, N., Torrens, H. S., 1980. A correlation of Jurassic Rocks in the British
Isles Part One: Introduction and Lower Jurassic. Geological Society ofLondon,
Special reports, no. 14,53-59.
Howarth, M.K., 1991-1992. The Ammonite family Hildoceratidae in the Lower
Jurassic of Britain. Monograph of the Palaeontographical Society: part 1
(1991), 1-106, part 2 (1992), 107-200.
Hyatt, A., 1867. The fossil cephalopods of the Museum of Comparative Zoology.
Bulletin of the Museum ofComparative Zoology, 3, 71-102.
Imlay, R.W., 1955. Characteristic Jurassic mollusks from northern Alaska.
Geological Survey Professional Paper, 274-D, 69-96.
Imlay, R. W., 1968. Lower Jurassic (Pliensbachian and Toarcian) Ammonites from
Eastern Oregon and California. Geological Survey Professional Paper, 593-C,
1-51.
Izumi, K., Tanabe, K., 2010. The Early Toarcian OAE in Northeast Tethys from the
Viewpoint of the Bioturbation (Toyora area, West Japan). Short papers for the
8th International congress on the Jurassic System, 357-358.
Jakobs, G.K.) 1992. Toarcian (Early Jurassic) ammonoids from western North
America. Bulletin of Geological Survey of Canada, 428,1-137.
Jakobs, G.K.,palfy, J., 1994. Upper Triassic to Middle Jurassic biostratigraphic
and facies studies in the Iskut River map area, northwestern British Columbia.

126
Cerrent Research 1994-E, Geological survey of Canada, 17-28.
Jakobs, G.K., Smith, P.L., Tipper, H.W., 1994. An ammonite zonation for the
Toarcian (Lower Jurassic) of the North American Cordillera. Canadian Journal
ofEarth Science, 31, no. 6,919-942.
Jenkyns, H.C., 1988. The Early Toarcian (Jurassic) Anoxic Event: stratigraphic,
sedimentary, and geochemical evidence. American Journal of Science, 288,
101-15I.
Jenkyns, H.C., Clayton, C.J., 1986. Black shales and carbon isotopes in pelagic
sediments from the Tethyan Lower Jurassic. Sedimentology, 33, 87-106.
Jenkyns, H.C., Clayton, C.J., 1997. Lower Jurassic epicontinental carbonates and
mudstones from England and Wales: chemostratigraphic signals and the early
Toarcian anoxic event. Sedimentology, 44, 687-706.
Johannson, G.G., Smith, P.L., Gordey, S.P., 1997. Early Jurassic evolution of the
northern Stikinian arc: evidence from Laberge Group, northwestern British
Columbia. Canadian Journal ofEarth Sciences, 34, 1030-1057.
Jones, C.B., Jenkyns, H.C., Hesselbo, S.P., 1994. Strontium isotopes In Early
Jurassic seawater. Geochimica Cosmochimica Acta, 58, 1285-130 I.
Lachkar, N., Dommergues, J.-L., Meister, C.; Lang, J., Chellar, E.H., 2007. Apport
des ammonites au cadre biostratigraphique et chronostratigraphique du
Haut-Atlas de Rich au cours du Lias (Maroc). African Geoscience Review, 14,
no. 1,95-105.
Kalacheva, E.D., 1988. Toarcian. In: Krymholts, G.Ya., Mesezhnikov, M.S.,
Westermann, G.B.G. (Eds.), The Jurassic Ammonite Zones of the Soviet Union.
Geological Society ofAmerica Special Pape Boulder (Colorado), 223, 14-18.
Kawamura, H., 2010. Stratigraphic revision of the Jurassic Toyora Group of the
southern part of the Tabe basin, Yamaguchi Prefecture, southwest Japan.
Journal of the Geological Society of Japan, 116, no. 1, 27-44. (in Japanese,
with English abstract)
Kobayashi, T., Konishi, K., Sato, T., Hayami, 1., Tokuyama, A., 1957. On the
Lower Jurassic Kuruma Group. Journal of the Geological Society ofJapan, 63,

127
182-194. (in Japanese, with English abstract)
Krymholts, G.Ya., Mesezhnikov, M.S., Westermann, G.E.G., 1988. The Jurassic
ammonite zones of the Soviet Union. Geological Society of America Special
Pape Boulder (Colorado), 223, 116 p.
Kumazaki, N., Kojima, S., 1996. Depositional history and structural development
of the Kuruma Group (lower Jurassic) on the basis of clastic rock composition.
Journal of the Geological Society of Japan, 102, no. 4, 285-302. (in Japanese,
with English abstract)
Legaretta, L., Uliana, M.A., 1996. The Jurassic succession In west-central
Argentina: stratal patterns, sequences and paleogeographic evolution.
Palaeogeography, Palaeoclimatology, Palaeoecology, 120, 303-330.
Li, X., Grant-Mackie, J.A., 1993. Jurassic sedimentary cycles and eustatic
sea-level changes in southern Tibet. Palaeogeography, Palaeoclimatology,
Palaeoecology, 101, 27-48.
Liu, C., Heinze, M., Fursich, F.T., 1998. Bivalve provinces in the Proto-Atlantic
and along the southern margin of the Tethys in the Jurassic. Palaeogeography,
Palaeoclimatology, Palaeoecology, 137, 127-151.
Liu, Y.-G., Miah, M.R.U., Schmitt, R.A., 1988. Cerium: A chemical tracer for
paleo-oceanic redox conditions. Geochimica et Cosmochimica Acta, 52,
1361-1371.
Macchioni, F., 2002. Myths and legends in the correlation between the Boreal and
Tethys. Implications on the dating of the Early Toarcian mass extinctions and
the Oceanic Anoxic Event. Geobios memoire special, 24, 150-164.
Macchioni, F., Cecca, F., 2002. Biodiversity and biogeography of middle-late
liassic ammonoids: implications for the early Toarcian mass extinction.
Geobios memoire special, 24, 165-175.
Macchioni, F., Meister, C., 2003. Ammonite biostratigraphy of some Mediterranean
section. 2: The succession of the Gola del F. Burano (Umbria-Machigiano
Basin, Apennine), a reference section for Tethyan Domerian. Revue de
Paleobiologie Geneve, 22, 363-420.

128
Matsumoto, T., Ono, A., 1947. A biostratigraphic study of the Jurassic Toyora
Group, with special reference to ammonites. Science Reports of the Faculty of
Science, Kyushu University, Geology, 2, no. 1, 20-31. (in Japanese)
McArthur, J.M., Donovan, D.T., Thirlwall, M.F., Fouke, B.W., Mattey, D., 2000.
Strantiurn isotope profile of the early Toarcian (Jurassic) oceanic anoxic event,
the duration of ammonite biozones, and belemnite palaeotemperatures. Earth
and Planetary Science Letters, 179, 269-285.
McLennan, S.M., 1989. Rare earth elements in sedimentary rocks: influence of
provenance and sedimentary processes. In: Lipin, B.R., McKray, G.A. (Eds.),
Geochemistry and mineralogy of rare earth elements. Mineralogical Society of
America, Washington DC, 169-200.
Meister, C., 1987. Comparison des faunas d'ammonites au Domerien (Jurassique
inferieur) entre le bassin des Causses et les Cordilleres Betiques; composition
faunique et elements de correlation. Comptes Rendus de I 'Academie des
Sciences (Paris), Serle 2a, 305, 425-428.
Meister, C., 1988. Ontogenese et evolution des Amaltheidae (Ammonoidea).
Eclogae Geologicae Halvetiae, 81,763-841.
Meister, C., 1989. Les ammonites du Domerien des Causses (France), Analyses
paleontologiques et stratigraphiques. Editions du Centre National de la
Recherche Scientifique, 15, 1-80.
Meister, C., Blau, J., Bohm, F., 1994. Ammonite biostratigraphy of the
Pliensbachian Stage in the Upper Austroalpine Jurassic. Eclogae Geologicae
Helvetiae, 87, 139-155.
Meister, C., Friebe, J.G., 2003. Austroalpine Liassic ammonites from Vorarlberg
(Austria, Nothern Calcareous Alps). Beitrage zur Palaontologie, 28, 9-99.
Moldowan, J.M., Sundararaman, P., Schoell, M., 1986. Sensitivity of biomarker
properties to depositional environment and/or source input in the Lower
Toarcian of SW-Germany. Organic Geochemistry, 10,915-926.
Montfort, D.de, 1808, Conchyliologie systematique et classification methodique
des coquilles. Paris, 410 p.

129
Morard, A., 2004. Les evenements du passage Domerien-Toarcien entre Tethys
occidentale et Europe du Nord-Ouest. 338p, Unpublished doctoral thesis,
Universite de Lausanne.
Mouterde, R., Dommergues, J.-L., Meister, C., Rocha, R.B., 2007. Atlas des
fossiles caracteristiques du Lias portugais. IlIa) Domerien (Ammonites).
Ciencias da Terra, 16, 67 -Ll l .
Nakada, K., Matsuoka, A., 2008. The Toarcian Oceanic Anoxic Event and
biodiversity fluctuation in Early Jurassic ammonoid assemblage in East Asia.
Abstract of the 33rd International Geological Congress, HPF13313L.
Nakada, K., Matsuoka, A., 2009. On the Pliensbachian/Toarcian boundary in the
Lower Jurassic Toyora Group in southwest Japan. Volumina Jurassica, 7,
47-54.
Neumayr, M., 1875. Die Ammoniten der Kreide und die Systematik der
Ammonitiden. Zeitschrift der . Deutschen geologischen Gesellschajt, 27,
854-892.
Oppel, A., 1856-1858. Die Juraformations Englands, Frankreichs und des
sudwestlichen Deutschlands. 1-438 (1856); 439-586 (1857); 587-857 (1858).
Page, K.N., 2002. A sequence of biohorizons for the Subboreal Province Lower
Toarcian in northern Britain and their correlation with a Submediterranean
Standard. In: Matire, L. (Ed.), 6th International Symposium on the Jurassic
System, Mondello, Italy, 12-22 September, 2002, Abstract and Program,
142-143.
Page, K.N., 2003. The Lower Jurassic of Europe: its subdivision and correlation.
Geological Survey ofDenmark and Greenland Bulletin, 1, 23-59.
Page, K.N., 2008. The evolution and geography of Jurassic ammonoids.
Proceedings of the Geologist' Association, 119, 35-57.
Palfy, J., Hart, C.J.R., 1994. Biostratigraphy of the Lower to Middle Jurassic
Laberge Group Whitehorse Map Area (105D), Southern Yukon. In: Yukon
Exploration and Geology, 1994, Exploration and Geological Services Division,
Yukon, Indian and Northern Affairs Canada. Part C, 73-86.

130
Palfy, J., Smith, P.L., 2000. Synchrony between Early Jurassic extinction, oceanic
anoxic event, and the Karoo-Ferrar flood basalt volcanism. Geology, 28, no. 8,
747-750.
Parisi, G., Ortega-Huertas, M., Nocchi, M., Palomo, 1., Monaco, P., Martinez, F.,
1996. Stratigraphy and geochemical anomalies of the early Toarcian
oxygen-poor interval in the Umbria-Marche Apennines (Italy). Geobios, 29, no.
4, 469-484.
Polubotko, LV., Repin, Y.S., 1966. Stratigraphy and ammonites of Toarcian stage
of the central part of Omolon Massif. In: Materials on Geology and Mineral
resources of the North-East USSR, issue 19, 30-55. (in Russian)
Polubotko, LV., Repin, Y.S., 1974. biostratigraphy of the Lower Jurassic. deposits
of the North-East USSR. In: The Main Problems of Biostratigraphy and
Paleogeography of the North-East USSR, 68-89. (in Russian)
Poulton, T.P., 1991. Hettangian through Aalenian (Jurassic) guide fossils and
biostratigraphy, Northern Yukon and adjacent northwest territories. Geological
Survey of Canada, 410, 1-95.
Poulton, T.P., Detterman, R.L., Hall, R.L., Jones, D.L., Peterson, J.A., Smith, P.,
Taylor, D.G., Tipper, H.W., Westermann, G.E.G., 1992. Western Canada and
United States. In: Westermann, G.E.G. (Ed.), The Jurassic of the
Circum-Pacific. Cambridge University Press, Cambridge, 29-92.
Pourmotamed, F., Motamed, A., 1976. Sur l'existence du Domerien marin dans
l'Elbourz central (Iran). Comptes rendus sommaires de la Societe Geologique
de France, 3, 105-108.
Quenstedt, F.A. 1882-85: Die Ammoniten des schwabischen Jura. 1. Der Schwarze
Jura (Lias) 1882: 1-48 (pI. 1-6); 1883: 49-96 ( pI.7-12); 1884: 97-240 (pI.
13-30); 1885: 241-440 (pI. 31-54). Schweizerbart, 440 p.
Rakus, M., Guex, J., 2002. Les ammonites du Jurassique inferieur et moyen de la
dorsale tunisienne. Memoires de Geologie, 39, 217 p.
Raup, DM., Sepkoski, J.J.Jr., 1984. Periodicity of extinction in the geologic past.
Proceedings of the National Academy ofSciences, 81,801-805.

131
Raup, DM., Sepkoski, J.J.Jr., 1986. Periodic extinction of families and genera.
Science, 231, 833-836.
Repin, J.S., 1968. Lower Jurassic ammonites and Middle. Field Atlas of the
Jurassic Flora and Fauna of North-East USSR, 5-18.
Repin, J.S., 1974. Amaltheidae representatives from the Upper Pliensbachian
deposits in the north-eastern parts of the USSR and their stratigraphical
significance. In: Sachs, V.N., Biostratigraphy of Boreal Mesozoic. Academy of
Sciences of the USSR Siberian Branch, Transactions of the Institute of
Geology and Geophysics, 136, 50-174.
Repin, Ju.S., 1988. Pliensbachian. In: Krymholts, G. Ya., Mesezhnikov, M.S.,
Westermann, G.E.G. (Eds.), The Jurassic Ammonite Zones of the Soviet Union.
Geological Society ofAmerica Special Pape Boulder (Colorado), 223, 10-14.
Rosales, 1., Quesada, S., Robles, S., 2004. Paleotemperature variations of Early
Jurassic seawater recorded in geochemical trends of belemnites from the
Basque-Cantabrian basin, northern Spain. Palaeogeography,
Palaeoclimatology, Palaeoecology, 203, 253-275.
Rulleau, 2007. Biostratigraphie et paleontologic du Lias superieur et du Dogger de
la region lyonnaise, Tome 1. Rulleau, L. et section Gee-Paleo du Comite
d'etablissement des carrieres Lafarge, Lozanne (ed.), 382 p.
Rulleau, L., Guiffray, A., Dommergues, J.-L., 2007. Biostratigraphie et
paleontologic de la region lyonnaise, Tome II: du socle au Lias moyen. Rulleau,
L. et section Gee-Paleo du Comite d'etablissement des carrieres Lafarge,
Lozanne (ed.), 229 p.
Salvador, A. (Ed.), 1994. International Stratigraphic Guide-A guide to stratigraphic
classification, terminology, and procedure, Second Edition. International
Union of Geological Sciences and Geological Society of America, 214p.
Sato, T., 1955. Les ammonites recueillies dans Ie Groupe de Kuruma, Nord du
Japon Central. Transactions and Proceedings of the Palaeontological Society
ofJapan, New Series, no. 20, 111-118.
Sato, T., 1956. Correlation du Jurassique inferieur japonais en basant sur les

132
ammonites fossiles. Journal of the Geological Society of Japan, 62, no. 732,
490-503. (in Japanese, with French abstract)
Sato, T., Westermann, G.E.G., 1991. Japan and South-East Asia. In: Westermann,
G.E.G., Riccardi, R.C. (Eds.), Jurassic Taxa Ranges and Correlation Charts for
the Circum-Pacific. Newsletters on Stratigraphy, 24,81-108.
Schmid-Rohl, A., Rohl, H.-J., Oschmann, W., Frimmel, A., Schwark, L., 2002.
Palaeoenvironmental reconstruction of Lower Toarcian epicontinental black
shales (Posidonia Shale, SW Germany): global versus regional control.
Geobios, 35, 13-20.
Schmidt-Effing, R., 1972. Die Dactylioceratidae, eme Ammoniten-Familie des
unteren Jura. Munstersche Forschungen zur Geologie und Palaontologie, 216p.
Schmitz, B.,1987. Barium, equatorial high productivity, and the northward
wandering of the Indian continent. Paleoceanography, 2, 63-77.
Senger, A.M.C., Natalin, B.A., 1996. Paleotectonics of Asia: fragment of a
synthesis. In: An, Yin, Harrison, T.M. (Eds.), The Tectonic Evolution of Asia.
Cambridge University Press, Cambridge, 486-640.
Sey, 1.1., Repin, Y.S., Kalacheva, E.D., Okuneva, T.M., Paraketsov, K.V.,
Polubotko, V., 1992. Eastern Russia. In: Westermann, G.E.G. (Ed.), The
Jurassic of the Circum-Pacific. Cambridge University Press, Cambridge,
225-245.
Seyed-Emami, K., Fursich, F.T.~ Wilmsen, M., Schairer, G., Majidifard, M.R., 2004.
First record of Jurassic (Toarcian-Bajocian) ammonites from the northern Lut
Block, east-central Iran. Acta Geologica Polonica, 54, no. 1, 77-94.
Seyed-Emami, K., Fursich, F.T., Wilmsen, M., Cecca, F., Majidifard, M.R.,
Schairer, G., Shekarifard, A., 2006. Stratigraphy and ammonite fauna of the
upper Shemshak Formation (Toarcian-Aalenian) at Tazareh, eastern Alborz,
Iran. Journal ofAsian Earth Sciences, 28, 259-275.
Shiraishi, S., 1992. The Hida Marginal Tectonic Belt in the middle reaches of the
River Hime-kawa with special reference to the lower Jurassic Kuruma Group.
Earth Science (Chikyu kagaku) , 46, no. 1, 1-20. (in Japanese, with English

133
abstract)
Simms, M.J., Chidlaw, N., Morton, N., Page, K.N., 2010. British Lower Jurassic
Stratigraphy. Joint Nature Conservation Committee, 458p.
Smith, P.L., Tipper, H.W., Taylor, D.G., Guex, J., 1988. An ammonite zonation for
the Lower Jurassic of Canada and the United States: the Pliensbachian.
Cnadian Journal ofEarth Science, 25, 1503-1523.
Smith, P.L., Tipper, H.W., 1996. Pliensbachian (Lower Jurassic) Ammonites of the
Queen Charlotte Islands, British Columbia. Bulletins of American
Paleontology, 108, 348, 122 p.
Smith, W., 1816-1819. Strata Identified by Organized Fossils. London.
Sowerby, J., 1812-22. The Mineral Conchology of Great Britain; or coloured
figures and descriptions of those remains of testaceous animals or shells, which
have been preserved at various times and depths in the earth. Meredith (ed.),
London, .1-4, 1-383.
Sulyk, F., 1991. Squence stratigraphy of the Jurassic-lowermost Cretaceous in East
Greenland. Bulletin of the American Association of Petroleum Geologists, 75,
1468-1488.
Takahashi, T., Shuto, K., 1997. Major and trace element analyses of silicate rocks
using X-ray fluorescence spectrometry RIX3000. Rigaku-Denki Journal, 28,
25-37. (in Japanese)
Takizawa, F., 1984. Upper boundary and veneer rock of the Kuruma Group,
Hida-Gaien Belt. Abstract of the 91st Annual Meeting of the Geological Society
ofJapan, 202. (in Japanese)
Tanabe, K., 1991, Early Jurassic macrofauna of the oxygen-depleted epicontinental
marine basin in the Toyora Area, West Japan. Proceedings of Shallow Tethys,
no. 3, 147-157.
Thomson, R. C., Smith, P."L., 1992. Pliensbachian (Lower Jurassic) biostratigraphy
and ammonite fauna of the Spatsizi area, North-Central British Columbia.
Geological Survey ofCanada Bulletin, 437, 1-87.
Topchishvili, M., Lominadze, T., Tsereteli, 1., Todria, V., Nadareishvili, G., 2006.

134
Stratigraphy of the Jurassic of Georgia. Proceedings of the Georgian Academy
ofSciences, new series, 122, 455 p.
Vail, P.R., Todd, R.G., 1981. Northern North Sea Jurassic unconformities,
chronostratigraphy and sea-level changes from seismic stratigraphy. In: Illing,
L.V., Hobson, G. (Eds.), Petroleum Geology of the Continental Shelf of
North-West Europe. Keydon, London, 216-235.
Wanty, R.B., Goldhaber, M.B., 1992. Thermodynamics and kinetics of reactions
involving vanadium in natural systems - accumulation of vanadium In
sedimentary rocks. Geochimica et Cosmochimica Acta, 56, 1471-1483.
Westermann, G.E.G., 1992. The Jurassic of the circum-Pacific. New York-Oakleigh,
Cambridge University Press, 676 p.
Westermann, G.E.G., 2000a. Biochore classification and nomenclature in
palaeobiogeography: an attempt at order. Palaeogeography, Palaeoclimatology,
Palaeoecology, 158, 1-13.
Westermann, G.E.G., 2000b. Marine faunal realsm of the Mesozoic: review and
revision under the new guidelines for biogeographic classification and
nomenclature. Palaeogeography, Palaeoclimatology, Palaeoecology, 163,
49-68.
Yin, J., Gao, J., Wang, Y., Zhang, S., Zheng, C., Xu, D., Bai, Z., Sun, L., Su, X.,
2006. Jurassic ammonites in anoxic black shales from Sewa and Amdo,
Northern Tibet. Acta Palaeontologica Sinica, 45, 311-331. (in Chinese)
Zakharov, V.A., 1994. Climatic fluctuations and other events in the Mesozoic of the
Siberian Arctic. In: Thurston, D.K., Fujita, K. (Eds.), Proceedings of the
International Conference on Arctic Margin, Ancorage, Alaska, US Department
of the Interior, 23-28.
Zittel, K.A. von, 1884. Cephalopoda. In: Zittel, K.A. von, Handbuch der
Palaontologie 1, Abteilung 2, 1-893.
Zonenshain, L.P., Kuzmin, M.L., Natapov, L.N., 1990. Geology of the USSR: a
plate-tectonic synthesis. American Geophysical Union, Geodynamics Series 21,
1-242.

135
Plate 1
Dactylioceras helianthoides (YOKOYAMA)
la-b. SA14-l-l, North Valley, locality 14 *la: Rubber cast, 1b: external mould
2. SA27-3-l, North Valley, locality 27
3. SA27-3-2, North Valley, locality 27
Dactylioceras sp. aff. D. helianthoides (YOKOYAMA)
4. SA2B-1-4, North Valley, locality 28 *Rubber cast
Peronoceras subfibulatum (YOKOYAMA)
5. SA26-5-3, North Valley, locality 26
Fontanelliceras fontanellense (GEMMELLARO)
6. SA18-lh-l, North Valley, locality 18
7. SA18-la-3,North Valley, locality 18
Canavaria japonica (MATSUMOTO)
8. SA36-2-3, South Valley, locality 36
Amaltheus margaritatus MONTFORT
9. SA36-l-l, South Valley, locality 36
10. SA36-l-2, South Valley, locality 36
The locality numbers correspond to that of Fig. 3-1. Scale bars are equal to 1 em.
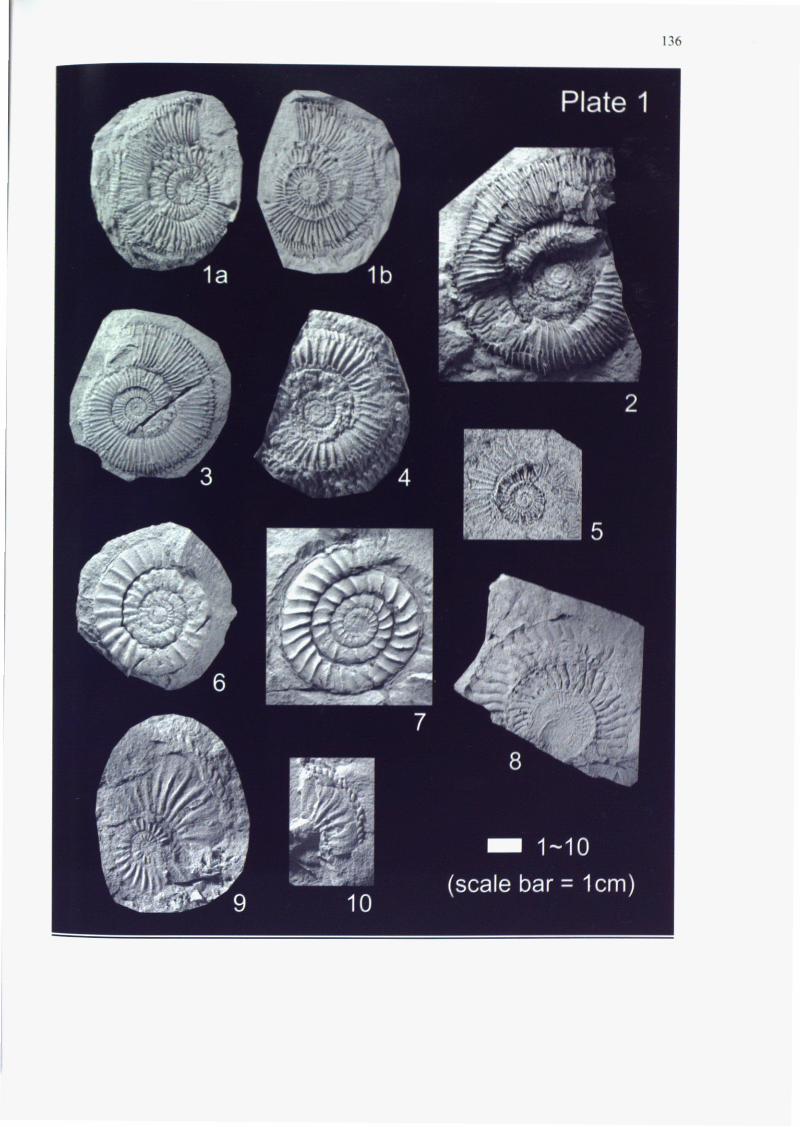

137
Plate 2
Protogrammoceras nipponicum (MATSUMOTO)
1. KTI9-1b-l, North Valley, locality 19
2. SAI9-1c-2, North Valley, locality 19
3. SA18-1c-1, North Valley, locality 18
Protogrammoceras onoi HIRANO
4. SA19-1b-2, North Valley, locality 1.9
Protogrammoceras yabei HIRANO
5. HI48-8-1, South Valley, locality 48
Fuciniceras nakayamense (MATSUMOTO)
6. SA19-1b-20, North Valley, locality 19
Paltarpites toyoranus (MATSUlVIOTO)
7. SA41-1-3, South Valley, locality 41
Paltarpites paltus (BUCKMAN)
8. SA41-1-2, South Valley, locality 41
Petranoceras sp. aff. P. rinaldinii VENTURI
9. SAI0-6-2, South Valley, locality 10
Lioceratoides aradasi (FUCINI)
10. KT41-1-2, South Valley, locality 41
The locality numbers correspond to that of Fig. 3-1. Scale bars are equal to 1 em.


139
Plate 3
"Cleviceras" chrysanthemum (YOKOYAMA)
1. SA19-1b-30, North Valley, locality 19
2. SA20-3b-3, North Valley, locality 20
3. SA26-4-4, North Valley, locality 26
Harpoceras inouyei (YOKOYAMA)
4, SA23 -1- 7, North Valley, locality 23
Cleviceras sp. cf. C. exaratum (YOUNG & BIRD)
5. SA23-1-3, North Valley, locality 23
Polyplectus okadai (YOKOYAMA)
6. SA20-3b-6, North Valley, locality 20
Hi/daites sp.
7. SA23-5-1, North Valley, locality 23
The locality numbers correspond to that of Fig. 3-1. Scale bars are equal to 1 em.


141
Plate 4
Amaltheus stokesi (SOWERBY)
1. DFFO101, Daira River 6 section, floated rock
2. DRFO148, Daira River 6 section, floated rock
3. GK. G. 11293, loco 50 in Hirano (1971), Chuzankei Valley *Rubber cast
4a-b. DRF0209, Daira River 6 section, floated rock
5. DRE0141, Daira River 6 section, floated rock
Amaltheus sp. cf. A. stokesi (SOWERBY)
6. DRF0143, Daira River 6 section, horizon: DRF0101
Amaltheus margaritatus MONTFORT
7. DRFO147, Daira River 6 section, floated rock
8. FMM2007, Daira River 6 section, floated rock
9. FMM2008, Daira River 6 section, floated rock
10. FMM2009, Daira River 6 section, floated rock
11. DRFO142, Daira River 6 section, floated rock
12. DRF0207, Daira River 7 section, floated rock "Apyrenum-type
13. MM2791 (T5303-1a in Sato, 1955), locality: unknown *Rubber cast
14. MM2792 (T5303-1 in Sato, 1955), locality: unknown *Rubber cast
15. DRF020 8, Daira River 7 section, floated rock *Rubber cast
16. THM0001, Toyora Group, locality: unknown
Amaltheus repressus DAGIS
17. DRF0202, Daira River 7 section, horizon: DRF0201
18. DRF0125, Daira River 6 section, floated rock
The numbers of section correspond to that of Fig. 2-5. The horizon numbers
correlate to Figs. 2-6 and 4-4. Scale bars are equal to 1 em.


143
Plate 5
Amaltheus sp. aff. A. talrosei (REPIN)
la-b. DRFOl49, Daira River 6 section, floated rock
Amaltheus sp.
2. DRF0210, Daira River 7 section, floated rock
Emaciaticeras nov sp .
.3a-Q~ DRFOl02, Daira River 6 section, floated rock
4a-b. DRFOl03, Daira River 6 section, floated rock
Juraphyll ites planulata FUCINI
5. TDFOIOl, Teradani Valley, horizon: TDFOI
The numbers of section correspond to that of Fig. 2-5. The horizon numbers
correlate to Figs. 2-6, 4-4 and 4-5. Scale bars are equal to 1 em.

I 4

145
Plate 6
Canavaria nov sp. A
1. TDF0210, Teradani Valley, horizon: TDF02
2. DRFO124, Daira River 6 section, floated rock
3. DRF0150, Daira River 6 section, floated rock
4. DRFOI23, Daira River 6 section, floated rock
5. I2RFO 121, Daira River 6 section, floated rock
Canavaria sp. aff. C. haugi (GEMMELLARO)
6. DRFO111, Daira River 6 section, floated rock
Canavaria nov sp. B
7. DRFO134, Daira River 6 section, floated rock
Canavaria sp. aff. C. naxensis (GEMMELLARO)
8. DRFOI04, Daira River 6 section, floated rock
Canavaria sp. aff. C. prognatum FUCINI
9. DRF020 1, Daira River 7 section, floated rock
Canavaria sp.
lOa-b. DRFO128, Daira River 6 section, floated rock
11a-b. DRFO129, Daira River 6 section, floated rock
12a-b. DRFO110, Daira River 6 section, floated rock
13a-c. TDF0206, Teradani Valley, horizon: TDF02
The numbers of section correspond to that of Fig. 2-5. The horizon numbers
correlate to Figs. 2-6, 4-4 and 4-5. Scale bars are equal to 1 em.
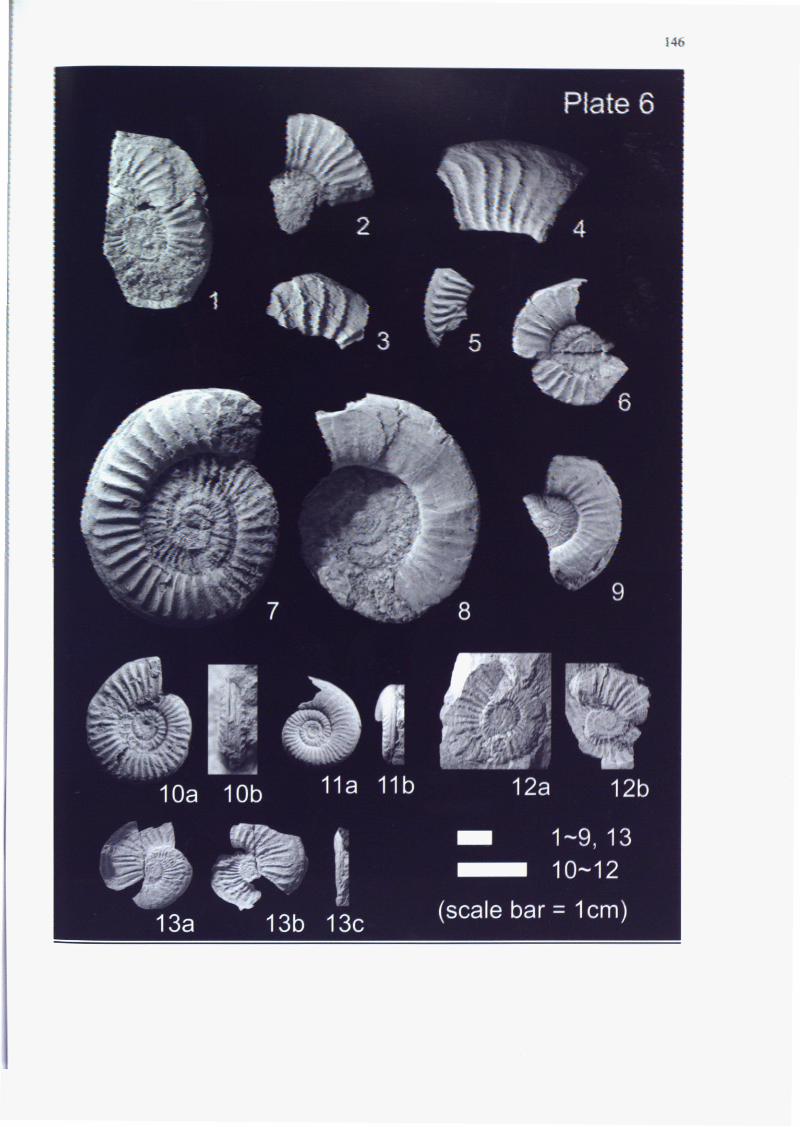
14

147
Plate 7
Lytoceras sp. cf. L. siemensi (DEKMAN)
1. TDF0302, Teradani Valley, horizon: TDF03
The horizon number corresponds to that of Figs. 2-5, Figs. 2-6 and 4-5. Scale bars
are equal to 10 em.

Pia 7
(scale bar =10em)

Plate 8
149

150
Dactylioceras sp. juv
20. SA53-4-15, North Valley, locality 53
21. SA53-4-17, North Valley, locality 53
Calliphylloceras sp.
22. SA53-4-18, North Valley, locality 53
23. SA53-4-1, North Valley, locality 53
The locality numbers correspond to that of Fig. 3-1. Scale bars are equal to 1 em.

Plate 8
6
75
3
4
1
89
10 11 12
15
14
18 1923
1-16,18-19,22-2317,20-21
20 21 22(scale bar =1cm)

2120
Sakuraguchidani Mudstone Member, Nishinakayama Formation
17 18 191410~. Outcrop No.Species I-----.-----.-----,.--=:....----,-----~---=-------=~--------,-----:----I
~ Fontanellicerasfontanellense~ (GEMELLARO)Q)o.~
~
SA10-S-1 SA18-1a-1SA18-1a-2SA18-1a-3SA18-1a-4SA18-1a-5SA18-1b-1
Petranoceras art: rinaldiniiVENTURI
SA1Q..6..2
Pallarpitespallus (BUCKMAN) SA10-6-6
Protogrammocerus nipponicllm(MATSUMOTO)
SA18-1c-1 SA19-1a-1 SA19-1b-11 SA19-1b-23SA18-1c-2 SA19-1a-3 SA19-1b-15 SA19-1b-25
SA19-1a-4 SA19-1b-16 SA19-1b-35SA19-1b-3 SA19-1b-17 SA19·1b-36SA19-1b-8 SA19-1b-18 SA19-1b-37SA19-1b-9 SA19-1b-19 SA19-1c-1
SA19-1c-2SA19-1c-3SA19-1c-4SA19-5a-1SA19-5a-2KT19-1b-1
KT19-1b-3KT19-1b-4KT19-1b-6KT19-1b-8
SA20-3b-7
SA20-3b-1 SA21-1-4
SA19-1b-26 KT19-1b-7
SA19-1b-1 SA19-1h-32 KT19-1b-2SA19-1b-29 SA19-1b-34SA19-1b-27
SA19·1b-2
SA19-1b-5
SA19·1b-10 SA19-1b-33SA19-1b-20 KT19-1c-1
Cll Q) Protogranrmo. cr. nippotlicllnl
~ ~ Protogramn,oceras )!abei
~ ~ ~ HIRANO8 g ~ Protogrammoceras cr. yabei
:si! ~ Protogrttmmoceras anoi HIRANO!:'! :r: 0:E e- hF;:;:u='cl;':':·nT:ie":':er='as::"'p:"":rT:im:"":o:":"rd:n"ill:::":m-::----I---+----l----+----+-----------------------4---+---1
~ (MATSUMOTO)Fuclniceras nakayamense
(MATSUMOTO)Lioceratoldesmafsllmotoi
HIRANOSA19-5b-1
TtltOl,iceras sp. SA17-5-1
"Cleviceras" chrysanthemum(MATSUMOTO)
SA19-1c-S SA19-1b-30SA19-1b-21
SA20-3b-3 SA21-1-1SA20-3a-3 SA21-1-3
"Co" afr. Chrl!SQllthemllm SA19-1b-28PolyplectllS okadai
(yOKOYAMA)SA19-1a-2SA19-1b-31
SA20-3b-2SA20-3b-6SA20-3b-8
Eoderoce- Dactyliocerasheliallthoidesrataceae NOKOYAMA)
SA14-1-1 SA19-5b-3 SA19-5b-5SA19-5b-4
SA20-3a-1SA20-3a-2
LJ'toceras sp. SA14-1-4 SA19-1b-4 SA19-1b-24SA19-1b-7 SA19-1b-38
Ammonitesp. SA10-S-3 SA14-1-2 SA17-4-1 SA18-8-1SA10-6-4 SA14-1-3
SA14-1-5
SA19-1b-6 SA19-5b-2SA19-1b-22 KT19-1b-5SA19-1c-6
SA20-3b-1 SA21-1-2
Appendix 3-1. Occurrence list of ammonoid fossils from the Sakuraguchidani Mudstone Member along the North Valley of the SakuraguchidaniValley, part 1. The inventory numbers of specimens are presented by each outcrop. The outcrop numbers correspond to those of Fig. 3-1.

~. Sakuraguchidani Mudstone Member. Nishinakayama FormationSpecies 23 24 25 26 27 28 29
FuciniCl!rosnaka)'amense SA23-1-1 SA24-1-2 SA26-4-9 SA27-4-1 SA29-3-8(MATSUMOTO) SA23-3-4
Lioceratoidesyokoyamai SA25-6-1 SA26-4-10(MATSUMOTO)
Liocerotoides matsumotoi SA26-1-4 SA27-5-2HIRANO SA27-5-10
"Ciesicems It chfJ'sllIfthtmum SA23-2-1 SA24-1-1 SA25-2-2 SA26-1-1 SA26-S-7 SA27-2-2 SA28-1-3 SA29-3-1(I) (MATSUMOTO) SA23-2-3 SA25-3-2 SA26-1-2 SA26-S-2 SA27-S-4 SA28-4-1 SA29-3-2co (I) co
SA26-4-4 SA26-S-4 SA27-S-8 SA28-4-7 SA29-3-3CI> IV .5:2 ;g ~ SA26-4-6 SA27-5-90tti ~ s SA26-4-7 SA27-5-1S~ § 0 "e. "cr. cJlrysonthemume- SA27-5-17 SA29-3-9 SA29-3-110 :2 IV32 I I Clnicerascr. exaralum SA23-1-3 SA28-4-2 SA29-3-4I (yOUNG & BIRD) SA28-4-4 SA29-3-7
Harpoceras ino'J'e; SA23-1-7 SA26-5-8 SA29-3-6(YOKOYAMA) SA23-3-3
Harpoceras sp. SA23-2-2 SA27-5-16
Polypleclus okadai SA24-S-1 SA29-3-5(YOKOYAMA)
* Hi/daltes sp. SA23-5-1
Dactylioceras helianthoides SA23-1-S SA26-1-3 SA27-3-1 SA29-3-10(YOKOYAMA) SA23-1-6 SA26-4-5 SA27-3-2
co SA23-2-S SA27-5-S
-8 SA23-3-1 SA27-5-11:§ SA23-3-2 SA27-S-12~ Dactylioceras afT. heliandloidts SA28-1-4se DactJ'/ioceras sp, SA26-1-6 SA28-4-8-8 Peronocems subflbulatum SA25-3-1 SA26-1-5 SA27-2-1 SA28-4-50w (YOKOYAMA) SA26-3-1 SA27-4-2
SA26-5-3 SA27-5"()SA26-7-1 SA27-5-7
Calliph).lloceras sp. SA26-4-1
Lywcefassp. SA26-7-2
Ammonite sp, SA23-1-2 SA25-2-1 SA26-5-S SA26-7-3 SA26-4-3 SA27-5-1 SA27-5-14 SA28-1-1 SA28-4-6 SA29-4-1SA23-1-4 SA26-5-6 SA26-7-4 SA26-4-8 SA27-5-3 SA28-1-2 SA28-4-9SA23-2-4 SA26-S-9 SA26-4-2 SA27-5-13 SA28-4-3
Appendix 3-2. Occurrence list of ammonoid fossils from the Sakuraguchidani Mudstone Member along the North Valley of the Sakuraguchidani Valley.part 2. The inventory numbers of specimens are presented by each outcrop. The outcrop numberscorrespond to those of Fig. 3-1. *: Hildoceratinae.

o. Sakuraguchidani Mudstone Memberl Nishinakayama FormationSpecies 36 40 41 43 48
Q,) Fonlanefliceras!ontalleUense SA41-1-11as (GEMELLARO).;;~ Call1lvarla japollica SA36-2-1~ (MATSUMOTO) SA36-2-2
:;:: SA36-2-3(l)'C SA36-2-4-c
SA36-2-5Paltarpitespallus (BUCKMAN) SA41·1·2 SA41-1-19Paltarpites tOJ'oranus SA41-1-3 KT41-1-1
(MATSUMOTO)PrologramnlOceras nipponicum SA48-2-5
(MATSUMOTO)
ProtograRlmoceras )labe; HIRANO HI48-8b-1Profogrammoceras sp. SA48-2·3
m Fudniceras nakaJYlmellse SA48-8b-4 SA48-8c-1Q,) m (MATSUMOTO) SA48·9-1 KT48-8a-3:2 "0
(l) Lioceratoides matsumoto;0 1= KT48-8c-2'@ e as
s .!: HIRANO
~1ii LioceratoidesJ'Okoyamai0~
SA41-1-8 SA48-2-1 KT48-8c-1:g :2 (MATSUMOTO) SA48-8a-4I I 0
~ Lioceratoides aradasi(FUCINI) KT41-1-2J:
Liocerato;des sp, SA41-1-10 SA41-1-12"Cfeviceras" chfJ'santhemuII' SA48-3-1
(MATSUMOTO) SA48-8a-8Clev;ceras sp. SA48-8a-10Polyplectus okadai SA48-6-1
(YOKOYAMA)
Amoftheus margorltatus HI36-1-1MONTFORT HI36-1-2
Eoderoce- Dactylioceras he/ianthoides SA48-4-1 SA48-8a-9 KT48-8a-1ratoidea (yOKOYAMA) SA48-4-2 SA48-8b-3 KT48-8a-2
SA48-8a-2 SA48-8c-5 KT48-8a-4
DacfJ'lioceras sp. KT4B-8a-5
CalfiphJ'lfoceras sp. SA48-2-4 SA48·8a-1 SA48·8b-1SA48-2-6 SA48-8a-7 SA48-8b-2
~'toceras sp, SA41-1-18 SA48-8a-6Ammonite sp. SA40-4-1 SA41-1-1 SA41-1-6 SA41-1-13 SA41-1-16 SA41-1-21 SA43-2-1 SA43-4-1 SA43-4-4 SA48-2-2 SA48-8c-2
SA41-1-4 SA41-1-7 SA41·1-14 SA41-1-17 KT41-1-3 SA43-2-2 SA43+2 SA48-8a-3 SA48-8C-3SA41-1-5 SA41-1-9 SA41-1-15 SA41-1-20 KT41-1-4 SA43-3-1 SA43+3 SA48-8a-5 SA48-8c-4
Appendix 3-3. Occurrence list of ammonoid fossilsfromthe Sakuraguchidani Mudstone Memberalongthe South Valley of the Sakuraguchidani Valley.The inventory numbers of specimens are presented by eachoutcrop. The outcrop numbers correspond to thoseof Fig.3-1.

o. Ishimachi Sandstone and Mudstone Member. Nishinakayama F.Species 53-2 53-3 53-4
Osperlioceras sp, A SA53-2-1 SA53-3-1 SA53-4-4al SA53-4.7c: SA53-4.8e SA53-4·9I Osperlioceras sp, B SA53-4.2iv SA53-4·3J: SA53-4-5
Grommoceras sp. SA53-4·11SA53-4·14
as ID-2l as PseudograntnJoceras sp.A SA53-4·10.:2:§ m SA53·4·12e B SA53-4-19S 00 30! al30! :fJ: c:
Pseudogrontmoceras sp. B SA53-4·20~ SA53-4·21S0EEe(!)
SA53-4.13 SA53-4-22Pseudogrammoceros sp,juvSA53-4.16 SA53-4·23
Phlyseogramntoceras sp, SA53-4-6
Dact}'/ioceras sp,juv SA53-4-15SA53-4.17
Eoderoce·ratoldea
Calliph}'lloceras sp, SA53-4·1SA53-4.18
Appendix 3-4.Occurrence list of ammonoid fossils fromthe Ishimachi Sandstone and Mudstone Member along the North Valley of the SakuraguchidaniValley. The inventory numbers of specimens are presented by each horizon. The outcrop numbers correspond to those of Fig.3-1.

~'Teradanl Formation Kuruma GrOUD
SpeciesDaira River 6 section Daira River 7 section Teradani Vallev Locality
DRFOf01 DRF0102 floated collecriol'l DRF0201 DRF0202a DRF0202b DRF0202c DRF0203 DRF0204 floated couecnon TDF01 TDF02 TDF03 TDF04 unknown
Canavaria novsp.A DRF0150 DRF0108 DRF0120 020301 020401 DRF0203 TDF0210DRF0110 DRF0121 020302 020402 TDF0211DRF0131 DRF0122
DRF0123DRF0124
Canavarta novsp.B DRF0134 DRF0144 020101 DRF0211DRF0135 DRF0146 DRF0213DRF0136DRF0137DRF0138DRF0104 DRF0132 DRF0204
DRF0133C. aff. haugl DRF0111C. aff. oroanatum DRF0201
020202 DRF0205 TDF0202Canavarta sp. DRF0206 TDF0203
TDF0204TDF0205TDF0206TDF0207TnF:O?QR
Emaciattceras sp. DRF0102 DRF0212DRF0103
Arieticeratinae gen.et sp. DRF0106 DRF0109 020201
indet DRF0107 DRF0112DRF0126 DRF0129DRF0127 DRF0130DRF0128
Amaitheus stokest DRF0101 DRF0148 DRF0209DRF0141
A. cf. stokes! DRF0143 DRF0207DRF0147 FMM2Q07 0202a02 DRF0208 T5303·1
A.margarttatus DRF0142 FMM2008 T5303·1aFMM2009
A.reoressus DRF0125 DRF0202A. aff. talroset DRF0149Amaitneu» sp. DRF0210Juraphyllites planulata DRF0214 TDF0101Lytoceras cf. siemenst. TDF0302
DRF0113 DRF0139 0202a01DRF0114 DRF0140
Ammonite sp, DRF0115 DRF0118DRF0116 DRF0119DRF0117
Appendix 3-5.Occurrence list of ammonoid fossils fromthe Teradani Formation, the Kuruma Group. The inventory numbers of specimens are presentedby each section and horizon. The horizon numbers correspond to those of Figs. 3-4 and 3-5.

sample No. 10-6 14-1 41·1 17-3 18·1a 18-1b 18·1c 19-1a 19-1b 19-5b 20-3b 21·1 23·3 24-2 24-4 25-2 25-3 25·5 25-8 26-1 26-2 26-4 26-7 27-1 27·3 27-5 28-1 29·2{malor elementsl
5102 60.58 61.58 61.34 59.75 61.41 62.79 53.04 59.35 60.59 57.97 60.80 54.75 58.27 56.08 61.78 56.38 57.58 60.34 55.95 57.63 61.50 56.65 60.59 57.60 59.32 60.62 55.34 54.94TI02 0.74 0.70 0.70 0.72 0.71 0.76 0.89 0.73 0.73 0.68 0.74 0.64 0.69 0.68 0.76 0.68 0.69 0.74 0.67 0.70 0.74 0.67 0.72 0.67 0.73 0.75 0.65 0.70
AI203 17.25 16.70 17.18 15.96 17.03 17.36 17.33 17.42 15.39 16.88 16.19 15.10 15.86 15.58 17.54 15.81 16.22 17.22 15.96 16.13 17.49 15.69 16.78 15.61 16.09 17.24 15.21 15.94FeO· 5.54 5.98 5.38 5.35 5.32 4.17 6.62 5.72 5.87 5.01 4.47 4.93 4.77 4.94 4.96 4.99 4.98 5.56 4.94 4.76 4.98 4.67 4.95 5.01 4.87 5.01 4.64 4.58MnO 0.02 0.03 0.02 0.05 0.02 0.02 0.04 0.03 0.03 0.04 0.01 0.05 0.04 0.05 0.02 0.04 0.04 0.03 0.05 0.04 0.02 0.05 0.03 0.04 0.03 0.03 0.05 0.04MgO 2.43 2.47 1.86 2.38 1.97 1.66 2.98 2.,26 2.50 2.31 2.06 2.03 2.20 2.20 2.01 2.25 2.28 2.40 2.27 2.25 2.15 2.21 2.34 2.39 2.54 2.45 2.18 2.15CaO 0.36 0.32 0.18 1.58 0.24 0.23 0.36 0.26 0.26 3.25 0.25 5.53 2.87 4.23 0.25 3.93 3.96 0.79 4.96 3.92 0.41 4.36 1.66 4.27 2.12 1.89 5.67 5.14Na20 2.49 2.50 2.50 2.29 2.04 2.05 1.67 1.84 1.97 2.21 1.76 1.92 2.24 2.19 2.42 2.12 2.14 2.35 2.15 2.24 2.32 2.11 2.21 2.04 2.06 2.18 1.97 1.92K20 2.80 2.53 2.72 2.43 2.93 3.13 2.71 3.22 2.30 2.84 2.76 2.49 2.48 2.48 2.84 2.55 2.68 2.83 2.63 2.66 2.82 2.55 2.76 2.53 2.51 2.81 2.45 2.70P205 0.13 0.12 0.15 0.14 0.16 0.15 0.16 0.16 0.15 0.14 0.14 0.14 0.15 0.15 0.15 0.15 0.16 0.16 0.15 0.16 0.16 0.15 0.16 0.15 0.16 0.16 0.17 0.16
H20tiLOI+Fe203-FeO' 6.56 7.10 7.47 6.94 7.78 7.11 10.67 8.13 7.63 7.54 7.96 9.76 7.76 8.84 6.87 8.45 8.08 6.95 8.88 7.71 7.14 8.85 6.46 8.10 7.23 6.40 9.68 9.90Total 98.89 100.04 99.51 97.60 99.59 99.42 96.46 99.12 97.42 98.88 97.15 97.34 97.33 97.43 99.60 97.34 98.81 99.38 98.62 98.21 99.72 97.95 98.66 98.42 97.65 99.56 98.01 98.18
(trace elementsl
Nb 9.821 9.629 9.794 9.897 11.32 11.408 9.859 12.231 11.059 10.062 11.762 9.911 10.5 9.933 11.359 9.507 11.05 10.926 10.465 10.16 10.66 10.375 10.055 9.613 10.651 10.301 9.376 11.026NI 56.2 62.2 51 63.5 33.6 24.5 52.1 41.1 36.3 39.7 34.7 38.7 36.7 34.7 40.7 42.5 39.9 43.9 35.8 35.8 34 33.6 40.1 44.8 35.8 44.1 45.5 41.2Pb 16.903 21.356 20.894 23.885 25.136 22.309 40.599 28.443 30.179 21.129 27.283 28.24 26.173 27.227 19.07 27.638 17.721 20.257 16.339 16.776 17.812 27.069 18.891 19.564 28.897 17.392 22.387 24.141Rb 118.34 111.22 123.1 112.76 131.82 140.64 126.19 147.79 110.04 128.5 134.92 112.08 113.82 113.13 130.4 116.64 121.88 127.1 118.8 118.08 128.23 118.17 126.81 118.48 117.88 127.52 111.8 122.61Sr 156.56 157.2 148.84 228.49 121.49 123.48 133.19 107.05 136.99 317 116.67 376.06 279.88 361.56 144.88 313.04 306.3 171.31 381.07 326.9 164.88 367.49 221.26 348.83 247.74 236.43 443.94 437.17Th 11.22 10.61 13.35 12.06 11.48 13.78 9.36 11.06 10.76 10.89 12.02 9.22 11.08 10.20 12.90 10.69 10.16 12,18 10.80 10.12 11.87 10.06 12.39 10.03 11.06 10.64 9.78 10.24Y 22.613 26.419 38.278 38.083 32.843 29.4 28.602 27.244 29.798 25.418 28.9 26.439 28.819 28.372 23.994 27.467 28.948 28.851 29.09 31.122 24.582 26.122 27.946 29.418 26.142 29.798 31.088 28.574Zr 181.68 176.94 160.56 168.97 163.98 171.81 156.36 169.17 168.56 164.86 159.48 147.28 181.64 152.53 173.52 152.29 180.58 167.72 180.21 168.41 165.78 154.92 162.55 158.93 153.47 185.61 149.1 156.49Sa 581 528.7 484.4 503.8 590.7 884.4 671.8 593 723.1 819.8 729.5 593.3 678.8 569.1 678.8 580.5 5n.4 569.2 559.8 565 589.9 570.6 807.2 534.7 576.5 627.6 573.7 694.4Cr 95.2 84.1 75.2 n,8 78.8 83.3 93.1 84.9 78.9 80.8 81.5 74.3 76.8 79.5 82 n,7 79.6 81.8 77.9 85.1 85.8 81.6 84.2 81.3 88.2 91.7 81.2 86.5V 127.43 114.06 111.34 104.29 113.44 117.6 166.28 117.67 100.69 114.51 115.96 99.18 109.28 102.75 111.17 102.79 106.66 110.48 106.09 116.2 121.61 108,86 116.56 116.03 124.29 132.4 132.82 125.1
Co 20.31 23.20 17,6947 22.70 15.04 13.36 27.22 17.01 19.41 16.62 15.68 20.17 19,55 16.69 19;66 19.96 16.99 23.04 17.27 19.13 14.45 17.52 18,11 21.21 15.21 16,25 15.85 16.22Th 12.20 11.60 12.3167 12.31 14.24 14.10 13.94 14.65 12.29 14.06 12.86 14.02 14.66 14.14 13.96 13.82 13.92 13.64 14.09 13.40 13.74 14.14 13.14 13.00 13.77 13.04 13.14 14.33U 2.80279 3.24378 3.17074 2.79196 3.30991 3.37115 3.20764 3.1561 2.92916 2.76661 3.20001 2.87518 3.10039 3.12033 2.83296 2.94707 3.03522 2.90145 3.00158 2.9509 2.90163 3.03036 3.07402 2.7905 3.10768 2.86791 3.33876 3.42735
I'lIre earth elementsLa 0.99104 0.88541 0.98396 1.19736 1.12712 1.13143 1.24023 1.13947 1.03652 1.06678 1.11484 1.01206 1.14605 1.00785 0.97263 0.99721 1.08096 1.03702 1,08021 1.02803 0.93814 1.03748 1.08787 1.03406 1.06139 1.0546 1.08217 1.05271Ct 0.85081 0.78449 0.86901 1.02661 0.96197 0.94522 1.0939 0.96072 0.909 0.90924 0.96239 0.90207 0.99803 0.91242 0.82529 0.87101 0.948 0.93907 0.942 0.94078 0.8248 0.93381 0.92103 0.90249 0.93112 0.93117 0.95851 0.91044
Pr 0.94902 0.87641 1.03118 1.16968 1.10937 1.07016 1.19837 1.07334 1.03778 0.99637 1.04631 0.98999 1.13957 1.01643 0.85846 0.97808 1.027211 1.02893 1.02645 1.0097 0,90694 1.01964 1.04943 0.9941 1.03847 1.04099 1.08592 1.0119
Nd 0.81296 0.80049 1.00856 1.10208 0.99428 0.96156 1.10463 0.96398 0.98179 0.88539 0.93671 0.89721 1.04987 0.93328 0.n654 0.90082 0.94334 0.93802 0.92522 0.94096 0.81316 0.92142 0.93474 0.91104 0.94494 0.92407 1.00395 0.896438m 0.86744 0.91325 1.34181 1.37854 1.15158 1,01356 1.23099 1.01113 1.179n 0.94478 1.09984 1.04523 1.20694 1.1169 0.80221 1.06638 1.07915 1.08597 1,03378 1.10062 0.86247 1.10889 1.07206 1.06988 1.11961 1.08063 1.21956 1.04771
Eu 0.80452 0.85286 1.47953 1.29271 0.98636 0.90745 1.07461 1.00749 1.12341 0.8346 0.96294 0.98312 1.06032 0.98066 0.74017 1.02439 1.03261 0.99998 0.99628 0.99398 0.73906 0.99107 1.04678 1.06393 1.01677 1.06121 1.15668 1.08431
Gd 0.n138 0.88079 1.48831 1.41686 1.05816 0.94255 1.08816 0.93246 1.1291 0.90859 1.04844 1.01611 1.14638 1.02753 0.76452 1.04486 1.06106 1.01942 1.0231 1.06227 0.85914 1.00622 1.03788 1.07161 1.04637 1.00937 1.17886 1.02564Tb 0.71541 0.87092 1.4066 1.34273 0.98148 0.87134 1.0676 0.86138 1.08982 0.8301 1.02137 0.94989 1.08228 0.97402 0.70044 0.98143 0.96563 0.94817 0.99256 1.0089 0.76947 0.93206 0.94881 0.95568 1.003 0.98272 1.11412 0.98806
Dv 0.63869 0,7687 1.15643 1.09824 0.89989 0.79302 0.91753 0.75707 0.9357 0.74088 0,91352 0.81362 0.91803 0.83025 0.62976 0.81097 0.79852 0.82291 0.8568 0.89927 0.66907 0.79599 0.80544 0.82901 0.84275 0.832 0.93295 0.87446
Ho 0.7698 0.90789 1.31743 1.22267 1.03239 0.92766 1.09888 0.88204 1.06316 0.87108 1.07556 0.92381 1.04613 0.96911 0.78862 0.95233 0.94314 0.93696 0.98867 1.00466 0.83262 0.91844 0.9485 0.96916 0.95885 0.91964 1.06119 1.01623
Er 0.70054 0.82743 1.06626 1.03853 0.94003 0.89198 1.00318 0.78811 0.93631 0.n676 0.93682 0.82948 0.92661 0.8349 0.71985 0.83198 0.83527 0.83694 0.86084 0.89355 0.73688 0.80623 0.83439 0.84807 0.85538 0.87096 0.92669 0.91887
Tm 0.79694 0.92139 1.02227 1.0n69 0.94936 0.81781 1.12063 0.81831 0.95484 0.83001 1.01666 0.83169 0.98485 0.87198 0.1766 0.87356 0.90117 0.88971 0,92132 0.93778 0.8238 0.87881 0.86133 0.83856 0.90999 0.90678 0.94071 0.9365
Yb 0.60234 0.96867 1.05887 1.02902 0.93616 0.92486 1.15686 0.82951 0.96541 0.84834 1.03921 0.8874 0.99848 0.93123 0.8181 0.89382 0.87387 0.91631 0..92648 0.90969 0.82818 0.87833 0.88903 0.86809 0.92284 0.927n 0.96879 0.99024Lu 0.82443 0.97516 1.06569 1.03663 0.94639 0.9189 1.1763 0.82887 0.95619 0.82427 1.02939 0.8796 0.99496 0.91447 0.80837 0.89069 0.86761 0.88004 0.91811 0.88948 0.79607 0.86803 0.90019 0.85835 0.89467 0.92538 0.95526 0.96751
Appendix 5-1.Analytical results of majorelements, traceelements and rareearth elements in mudstone samples fromthe Toyora Group.Sample numbers correspond to the horizon numbers in Figs. 5-1, 5-2 and 5-3.




















