1 Professional Knowledge ** 60 120 (Each question carries ...
Upload
independentCategory
view
3download
0
ORIGINAL ARTICLE
Ceramide and raft signaling are linked with each other in UVA
radiation-induced gene expression
S Grether-Beck1, M Salahshour-Fard1, A Timmer1, H Brenden1, I Felsner1, R Walli1, J Fullekrug2
and J Krutmann1
1Zellbiologie, Institut fuer Umweltmedizinische Forschung (IUF) at the Heinrich-Heine-University Duesseldorf gGmbH, Duesseldorf,Germany and 2Internal Medicine IV, University of Heidelberg, Heidelberg, Germany
Solar ultraviolet A (UVA) (320–400 nm) radiation-in-duced gene expression in keratinocytes is initiated at thelevel of the cell membrane via generation of singlet oxygenand subsequent formation of ceramide from sphingomye-lin. We now report that the UVA response also involvesraft signaling and that ceramide and raft signaling arelinked with each other. Upon UVA irradiation, the lipidcomposition of rafts decreased 40% in sphingomyelinand 60% in cholesterol (Chol). Also, decrease of Cholincreased the susceptibility towards UVA-induced geneexpression, whereas increase of Chol completely abolishedtheir capacity to generate signaling ceramides and tomount the subsequent UVA response. This inhibition wasnot associated with UVA-induced Chol oxidation and wasalso seen after treatment of cells with plant sterols. TheUVA responsiveness depended on the ratio of Chol versusceramide in rafts. A ratio smaller than 1 permittedinitiation and transduction of the signaling response,whereas a ratio greater than 1, for example, upon sterolpretreatment, abolished this response, indicating thatUVA radiation-induced ceramide signaling is controlledby the lipid composition of rafts.Oncogene (2008) 27, 4768–4778; doi:10.1038/onc.2008.116;published online 28 April 2008
Keywords: signaling; UVA; rafts; ceramide
Introduction
Solar ultraviolet A (UVA) (320–400 nm) radiation is awell-studied inducer of gene expression in human skincells (Tyrrell, 2004; Grether-Beck et al., 2005). Thephotobiological and molecular mechanisms are ofgeneral interest as UVA radiation-induced gene expres-sion is relevant for photoaging, photocarcinogenesisand pathogenesis of frequent photodermatoses such as
polymorphic light eruption (Schwarz, 2005; Tyrrell andReeve, 2006). Human epidermal keratinocytes representthe primary cellular target for UVA radiation. Inprevious studies, we have shown that the UVAradiation-induced stress response is initiated in this celltype by a photo-oxidative process, which causes thegeneration of singlet oxygen and that UVA radiation/singlet oxygen-induced gene expression critically dependon activation of transcription factor activator protein-2(AP-2) (Grether-Beck et al., 1996). The steps that occurdownstream of UVA radiation-induced singlet oxygengeneration and upstream of AP-2 activation involvegeneration of the second messenger ceramide from cellmembrane sphingomyelin indicating that UVA radia-tion-induced alterations at the level of cell membranelipids are critically involved in this signaling cascade(Grether-Beck et al., 2000).
Lipid microenvironments on the cell surface, knownas lipid rafts, participate in signal transduction processes(Simons and Toomre, 2000). Rafts or microdomainsconsist of sphingolipids such as sphingomyelin andcholesterol (Chol) and are located in the exoplasmicleaflet of the bilayer of biological membranes (Simonsand van Meer, 1988; Simons and Ikonen, 1997; Brownand London, 1998). Lipid rafts can change theircomposition in response to intra- or extracellular stimuli(Simons and Toomre, 2000; Simons and Ehehalt, 2002),for example, UV-induced induction of apoptosismediated by activation of acid sphingomyelinase (Zhanget al., 2001; Kashkar et al., 2005; Rotolo et al., 2005).This favors specific protein–protein interactions andsubsequent activation of signaling cascades. Raft func-tion has been shown to critically depend on the contentof Chol in the plasma membrane (Anderson, 1993;Schnitzer et al., 1994; Gustavsson et al., 1999).
As we had previously shown that UVA radiationresults in generation of signaling ceramides from plasmamembrane-localized sphingomyelin and as sphingo-myelin is associated with Chol in raft microdomains,we have assessed whether lipid composition of rafts isaltered by UVA irradiation and if yes, whether this maybe of functional relevance for UVA radiation-inducedsignaling.
Here we demonstrate that UVA radiation-inducedsignaling critically depends on the ratio of Chol versusceramide in cell membrane rafts. Our studies thus
Received 6 September 2007; revised 11 February 2008; accepted 14March 2008; published online 28 April 2008
Correspondence: Dr S Grether-Beck, Zellbiologie, Institut fuerUmweltmedizinische Forschung (IUF) at the Heinrich-Heine-University Duesseldorf gGmbH, Auf’m Hennekamp 50, DuesseldorfD-40225, Germany.E-mail: [email protected]
Oncogene (2008) 27, 4768–4778& 2008 Macmillan Publishers Limited All rights reserved 0950-9232/08 $30.00
www.nature.com/onc
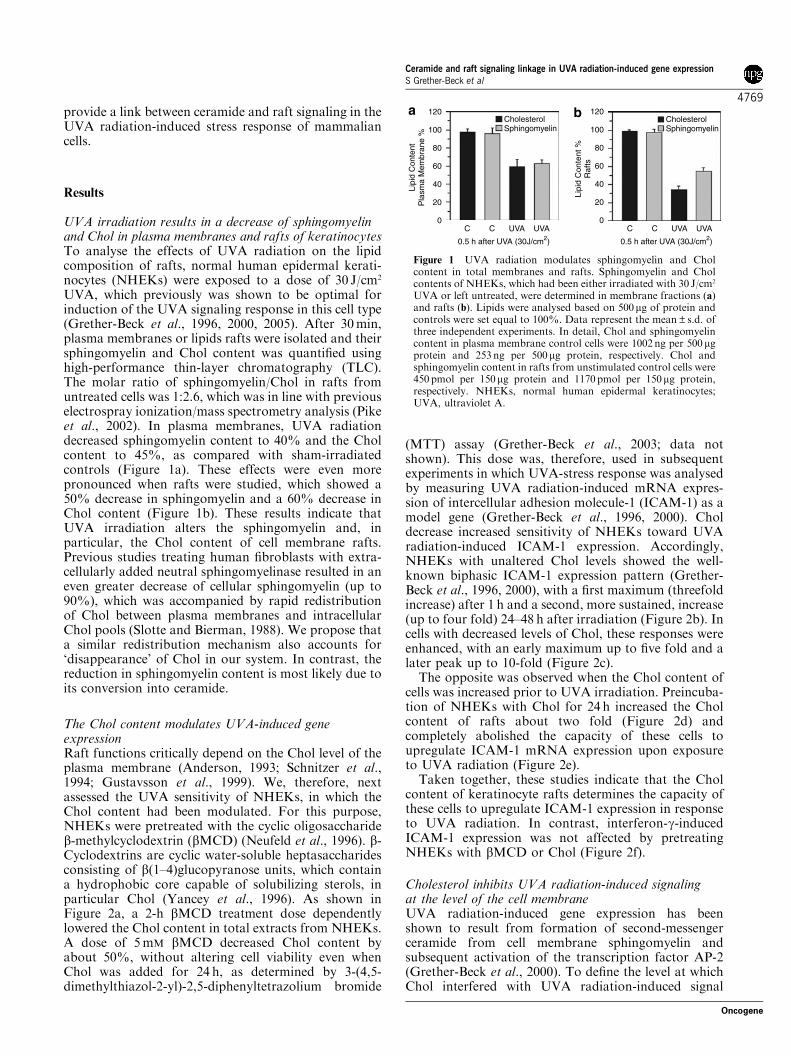
provide a link between ceramide and raft signaling in theUVA radiation-induced stress response of mammaliancells.
Results
UVA irradiation results in a decrease of sphingomyelinand Chol in plasma membranes and rafts of keratinocytesTo analyse the effects of UVA radiation on the lipidcomposition of rafts, normal human epidermal kerati-nocytes (NHEKs) were exposed to a dose of 30 J/cm2
UVA, which previously was shown to be optimal forinduction of the UVA signaling response in this cell type(Grether-Beck et al., 1996, 2000, 2005). After 30min,plasma membranes or lipids rafts were isolated and theirsphingomyelin and Chol content was quantified usinghigh-performance thin-layer chromatography (TLC).The molar ratio of sphingomyelin/Chol in rafts fromuntreated cells was 1:2.6, which was in line with previouselectrospray ionization/mass spectrometry analysis (Pikeet al., 2002). In plasma membranes, UVA radiationdecreased sphingomyelin content to 40% and the Cholcontent to 45%, as compared with sham-irradiatedcontrols (Figure 1a). These effects were even morepronounced when rafts were studied, which showed a50% decrease in sphingomyelin and a 60% decrease inChol content (Figure 1b). These results indicate thatUVA irradiation alters the sphingomyelin and, inparticular, the Chol content of cell membrane rafts.Previous studies treating human fibroblasts with extra-cellularly added neutral sphingomyelinase resulted in aneven greater decrease of cellular sphingomyelin (up to90%), which was accompanied by rapid redistributionof Chol between plasma membranes and intracellularChol pools (Slotte and Bierman, 1988). We propose thata similar redistribution mechanism also accounts for‘disappearance’ of Chol in our system. In contrast, thereduction in sphingomyelin content is most likely due toits conversion into ceramide.
The Chol content modulates UVA-induced geneexpressionRaft functions critically depend on the Chol level of theplasma membrane (Anderson, 1993; Schnitzer et al.,1994; Gustavsson et al., 1999). We, therefore, nextassessed the UVA sensitivity of NHEKs, in which theChol content had been modulated. For this purpose,NHEKs were pretreated with the cyclic oligosaccharideb-methylcyclodextrin (bMCD) (Neufeld et al., 1996). b-Cyclodextrins are cyclic water-soluble heptasaccharidesconsisting of b(1–4)glucopyranose units, which containa hydrophobic core capable of solubilizing sterols, inparticular Chol (Yancey et al., 1996). As shown inFigure 2a, a 2-h bMCD treatment dose dependentlylowered the Chol content in total extracts from NHEKs.A dose of 5mM bMCD decreased Chol content byabout 50%, without altering cell viability even whenChol was added for 24 h, as determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
(MTT) assay (Grether-Beck et al., 2003; data notshown). This dose was, therefore, used in subsequentexperiments in which UVA-stress response was analysedby measuring UVA radiation-induced mRNA expres-sion of intercellular adhesion molecule-1 (ICAM-1) as amodel gene (Grether-Beck et al., 1996, 2000). Choldecrease increased sensitivity of NHEKs toward UVAradiation-induced ICAM-1 expression. Accordingly,NHEKs with unaltered Chol levels showed the well-known biphasic ICAM-1 expression pattern (Grether-Beck et al., 1996, 2000), with a first maximum (threefoldincrease) after 1 h and a second, more sustained, increase(up to four fold) 24–48 h after irradiation (Figure 2b). Incells with decreased levels of Chol, these responses wereenhanced, with an early maximum up to five fold and alater peak up to 10-fold (Figure 2c).
The opposite was observed when the Chol content ofcells was increased prior to UVA irradiation. Preincuba-tion of NHEKs with Chol for 24 h increased the Cholcontent of rafts about two fold (Figure 2d) andcompletely abolished the capacity of these cells toupregulate ICAM-1 mRNA expression upon exposureto UVA radiation (Figure 2e).
Taken together, these studies indicate that the Cholcontent of keratinocyte rafts determines the capacity ofthese cells to upregulate ICAM-1 expression in responseto UVA radiation. In contrast, interferon-g-inducedICAM-1 expression was not affected by pretreatingNHEKs with bMCD or Chol (Figure 2f).
Cholesterol inhibits UVA radiation-induced signalingat the level of the cell membraneUVA radiation-induced gene expression has beenshown to result from formation of second-messengerceramide from cell membrane sphingomyelin andsubsequent activation of the transcription factor AP-2(Grether-Beck et al., 2000). To define the level at whichChol interfered with UVA radiation-induced signal
120
100
80
60
40
20
0
120
100
80
60
40
20
0C C UVA UVA
0.5 h after UVA (30J/cm2)
C C UVA UVA
0.5 h after UVA (30J/cm2)
Lipi
d C
onte
nt
Pla
sma
Mem
bran
e %
Lipi
d C
onte
nt %
Raf
ts
CholesterolSphingomyelin
CholesterolSphingomyelin
Figure 1 UVA radiation modulates sphingomyelin and Cholcontent in total membranes and rafts. Sphingomyelin and Cholcontents of NHEKs, which had been either irradiated with 30 J/cm2
UVA or left untreated, were determined in membrane fractions (a)and rafts (b). Lipids were analysed based on 500mg of protein andcontrols were set equal to 100%. Data represent the mean±s.d. ofthree independent experiments. In detail, Chol and sphingomyelincontent in plasma membrane control cells were 1002 ng per 500mgprotein and 253 ng per 500mg protein, respectively. Chol andsphingomyelin content in rafts from unstimulated control cells were450 pmol per 150mg protein and 1170pmol per 150mg protein,respectively. NHEKs, normal human epidermal keratinocytes;UVA, ultraviolet A.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4769
Oncogene
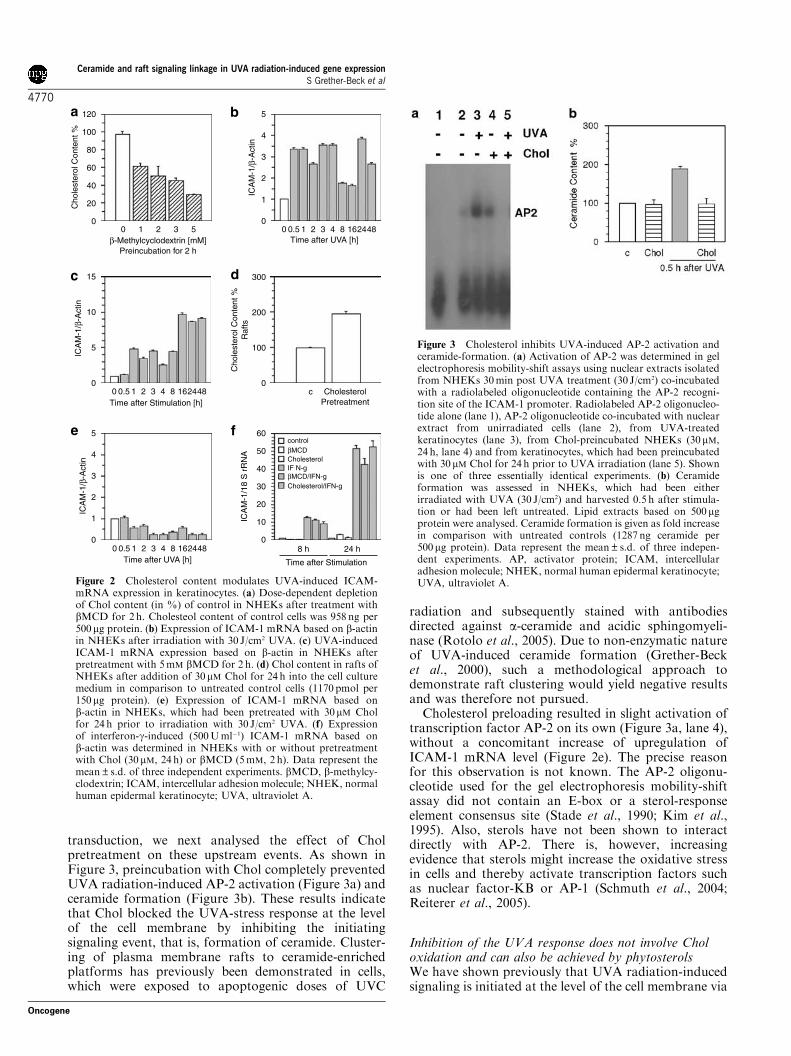
transduction, we next analysed the effect of Cholpretreatment on these upstream events. As shown inFigure 3, preincubation with Chol completely preventedUVA radiation-induced AP-2 activation (Figure 3a) andceramide formation (Figure 3b). These results indicatethat Chol blocked the UVA-stress response at the levelof the cell membrane by inhibiting the initiatingsignaling event, that is, formation of ceramide. Cluster-ing of plasma membrane rafts to ceramide-enrichedplatforms has previously been demonstrated in cells,which were exposed to apoptogenic doses of UVC
radiation and subsequently stained with antibodiesdirected against a-ceramide and acidic sphingomyeli-nase (Rotolo et al., 2005). Due to non-enzymatic natureof UVA-induced ceramide formation (Grether-Becket al., 2000), such a methodological approach todemonstrate raft clustering would yield negative resultsand was therefore not pursued.
Cholesterol preloading resulted in slight activation oftranscription factor AP-2 on its own (Figure 3a, lane 4),without a concomitant increase of upregulation ofICAM-1 mRNA level (Figure 2e). The precise reasonfor this observation is not known. The AP-2 oligonu-cleotide used for the gel electrophoresis mobility-shiftassay did not contain an E-box or a sterol-responseelement consensus site (Stade et al., 1990; Kim et al.,1995). Also, sterols have not been shown to interactdirectly with AP-2. There is, however, increasingevidence that sterols might increase the oxidative stressin cells and thereby activate transcription factors suchas nuclear factor-KB or AP-1 (Schmuth et al., 2004;Reiterer et al., 2005).
Inhibition of the UVA response does not involve Choloxidation and can also be achieved by phytosterolsWe have shown previously that UVA radiation-inducedsignaling is initiated at the level of the cell membrane via
120
100
80
60
40
20
00 1 2 3 5
Cho
lest
erol
Con
tent
%
β-Methylcyclodextrin [mM] Preincubation for 2 h
5
4
3
2
1
00 0.5 1 2 3 4 8 162448
Time after UVA [h]IC
AM
-1/β
-Act
in
5
4
3
2
1
00 0.5 1 2 3 4 8 162448
Time after UVA [h]
0 0.5 1 2 3 4 8 162448Time after Stimulation [h]
ICA
M-1
/β-A
ctin
300
200
100
0c Cholesterol
Pretreatment
Cho
lest
erol
Con
tent
%R
afts
control
Cholesterol
Cholesterol/IFN-g
βMCD
βMCD/IFN-gIF N-g
60
50
40
30
20
10
0
ICA
M-1
/18
S r
RN
A
8 h 24 h
Time after Stimulation
15
10
5
0
ICA
M-1
/β-A
ctin
Figure 2 Cholesterol content modulates UVA-induced ICAM-mRNA expression in keratinocytes. (a) Dose-dependent depletionof Chol content (in %) of control in NHEKs after treatment withbMCD for 2 h. Cholesteol content of control cells was 958 ng per500mg protein. (b) Expression of ICAM-1 mRNA based on b-actinin NHEKs after irradiation with 30 J/cm2 UVA. (c) UVA-inducedICAM-1 mRNA expression based on b-actin in NHEKs afterpretreatment with 5mM bMCD for 2 h. (d) Chol content in rafts ofNHEKs after addition of 30mM Chol for 24 h into the cell culturemedium in comparison to untreated control cells (1170 pmol per150mg protein). (e) Expression of ICAM-1 mRNA based onb-actin in NHEKs, which had been pretreated with 30mM Cholfor 24 h prior to irradiation with 30 J/cm2 UVA. (f) Expressionof interferon-g-induced (500Uml�1) ICAM-1 mRNA based onb-actin was determined in NHEKs with or without pretreatmentwith Chol (30 mM, 24 h) or bMCD (5mM, 2 h). Data represent themean±s.d. of three independent experiments. bMCD, b-methylcy-clodextrin; ICAM, intercellular adhesion molecule; NHEK, normalhuman epidermal keratinocyte; UVA, ultraviolet A.
Figure 3 Cholesterol inhibits UVA-induced AP-2 activation andceramide-formation. (a) Activation of AP-2 was determined in gelelectrophoresis mobility-shift assays using nuclear extracts isolatedfrom NHEKs 30min post UVA treatment (30 J/cm2) co-incubatedwith a radiolabeled oligonucleotide containing the AP-2 recogni-tion site of the ICAM-1 promoter. Radiolabeled AP-2 oligonucleo-tide alone (lane 1), AP-2 oligonucleotide co-incubated with nuclearextract from unirradiated cells (lane 2), from UVA-treatedkeratinocytes (lane 3), from Chol-preincubated NHEKs (30mM,24 h, lane 4) and from keratinocytes, which had been preincubatedwith 30 mM Chol for 24 h prior to UVA irradiation (lane 5). Shownis one of three essentially identical experiments. (b) Ceramideformation was assessed in NHEKs, which had been eitherirradiated with UVA (30 J/cm2) and harvested 0.5 h after stimula-tion or had been left untreated. Lipid extracts based on 500mgprotein were analysed. Ceramide formation is given as fold increasein comparison with untreated controls (1287 ng ceramide per500mg protein). Data represent the mean±s.d. of three indepen-dent experiments. AP, activator protein; ICAM, intercellularadhesion molecule; NHEK, normal human epidermal keratinocyte;UVA, ultraviolet A.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4770
Oncogene

generation of singlet oxygen (Grether-Beck et al., 1996).Chol can quench singlet oxygen (Korytowski andGirotti, 1999) or free radicals (Girotti, 1998), indicatingthe possibility that the UVA-inhibitory effect of Cholmight be due to its antioxidant properties. We thereforesearched for formation of the respective signatureoxysterols in lipid fractions isolated from UVA-irra-diated NHEKs. Peroxidation of Chol can occur via twodistinct pathways, (i) a type-I oxidation, which is freeradical-mediated and which leads to generation ofoxysterols oxidized at the carbon atom 7(C7), and (ii)a type-II oxidation, which is singlet oxygen-mediatedand which promotes formation of oxysterols oxidizedat carbon atom 5 (C5) (Girotti, 1992; Girotti andKorytowski, 2000). Small amounts of the singletoxygen-mediated oxysterol 3b-hydroxy-5a-cholest-6-ene-5-hydroperoxide (5a-OOH) could be detected insamples of pure Chol, which had been irradiated for1 h with a solar simulator emitting both UVB andUVA radiation (Figure 4a), thus confirming resultsfrom previous studies (Girotti and Korytowski,2000). In contrast, even loading the 15-fold amount oflipid extracts revealed no trace amounts of singletoxygen-generated 3b-hydroxy-5a-cholest-6-ene-5-hy-droperoxide (5a-OOH), 3b-hydroxycholest-4-ene-6a-hy-droperoxide (6a-OOH), 3b-hydroxycholest-4-ene-6b-hydroperoxide (6b-OOH) nor free radical-generated3b-hydroxycholest-5-ene-7a-hydroperoxide (7a-OOH)or 3b-hydroxycholest-5-ene-7b-hydroperoxide (7b-OOH)oxysterols in samples from UVA-irradiated keratinocytes(Figure 4b, lanes 5 and 11). Thus, inhibition of UVAradiation-induced signaling by Chol was not associatedwith Chol oxidation.
As chemical structures of Chol and plant sterols showsimilarities, we wondered whether inhibition of UVAradiation-induced signaling could also be achieved withnon-Chol sterols such as phytosterols. For this purpose,NHEKs were treated for 24 h with campesterol (Camp),b sitosterol (Sito) or stigmasterol prior to UVAirradiation. As is shown in Figure 5, phytosterolpretreatment suppressed UVA radiation-induced geneexpression (Figure 5a), AP-2 activation (Figure 5b) andceramide formation in rafts (Figure 5c). These resultsindicate that plant sterols function as Chol analogues inthis system by inhibiting the UVA radiation-inducedstress response in NHEKs.
The ratio of Chol versus ceramide within rafts determinesthe UVA responseTo understand the mechanism (s) by which Cholprotects cells from UVA stress, we assessed the effectsof Chol pretreatment on the lipid composition of rafts.As is shown in Figure 6, Chol pretreatment increasedChol and sphingomyelin content of rafts isolatedfrom unirradiated NHEKs up to 200%, and completelyprevented the UVA radiation-induced decrease inboth lipids.
We wondered whether inhibition of theUVA response by Chol pretreatment was caused byconcomitant increase of Chol and sphingomyelin or due
to the relative change in either Chol or ceramide orsphingomyelin. For this purpose, we assessed UVAresponsiveness of keratinocytes, which had alteredmolar ratios between Chol, sphingomyelin and ceramidewithin rafts as a consequence of their treatment with thechemical compounds listed in Table 1, all of which arewell known to interfere with the biosynthesis ofsphingomyelin or Chol.
As is shown in Table 1 and in Figures 5 and 6, Chol aswell as phytosterol pretreatment prevented the UVAresponse by increasing both Chol and sphingomyelinand without changing the molar ratio of sphingomyelinversus Chol. AY9944 (trans-1,4-bis(2-chlorobenzylami-nomethyl)cyclohexane dihydrochloride), an inhibitor forD7-dehydrocholesterol reductase, also abrogated theUVA response (Figure 7a), but this inhibition wasassociated with a decrease of Chol, a reciprocal increaseof sphingomyelin (Figure 7b) and thus a change of themolar ratio of sphingomyelin versus Chol from 1:2.6 to1:1.3. Essentially identical results (data not shown) wereobtained if BM15.766 was used, which also inhibitsD7-dehydrocholesterol reductase (Wolf et al., 1996).
frontChol
frontChol
1 2 3 4
CholOOH
CholOOH
7-Ketotchol
7-Ketochol
7�-OH7�-OH
7�-OH
start
start
Figure 4 Cholesterol oxidation products in UVA-irradiatedkeratinocytes. High-performance TLC separation of Chol photo-oxidation products using heptane/ethyl acetate (1:1, v/v): (a) lane 1,standard mix containing Chol, 7-Ketochol and 7b-OH; lane 2,Chol photo-oxidized with Rose Bengal; lane 3, 7-Ketocholpretreated with NaBH4 to reduce partially resolvable ChOOHs towell-resolved ChOHs (diols); lane 4, 7b-OH. (b) Lane 1, standardmix containing Chol (25mg), 7-Ketochol and 7b-OH (1.25mg each);lane 2, Chol (150 mg); lane 3, Chol (150mg) photo-oxidized withRose Bengal; lane 4, lipid extract from unirradiated NHEK(equivalent 375mg protein); lane 5, lipid extract from NHEKirradiated with 30 J/cm2 UVA (equivalent 375mg protein); lane 6,left free; lane 7, 7-Ketochol (7.5mg); lane 8, 7-ketochol aftertreatment with NaBH4; lane 9, extract of Rose Bengal-photo-oxidized Chol (150 mg) after treatment with NaBH4; lane 10, lipidextract from unirradiated NHEK (equivalent 375mg protein) aftertreatment with NaBH4; lane 11, lipid extract from NHEKirradiated with 30 J/cm2 UVA (equivalent 375mg protein) aftertreatment with NaBH4; lane 12, 7b-OH (7.5mg); lane 13, standardmix as lane 1. The chromatograms represent one of three essentiallyidentical experiments. 7-Ketochol, 7-ketocholesterol; NHEK,normal human epidermal keratinocyte; 7b-OH, 7b-hydroxycholes-terol; TLC, thin-layer chromatography; UVA, ultraviolet A.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4771
Oncogene
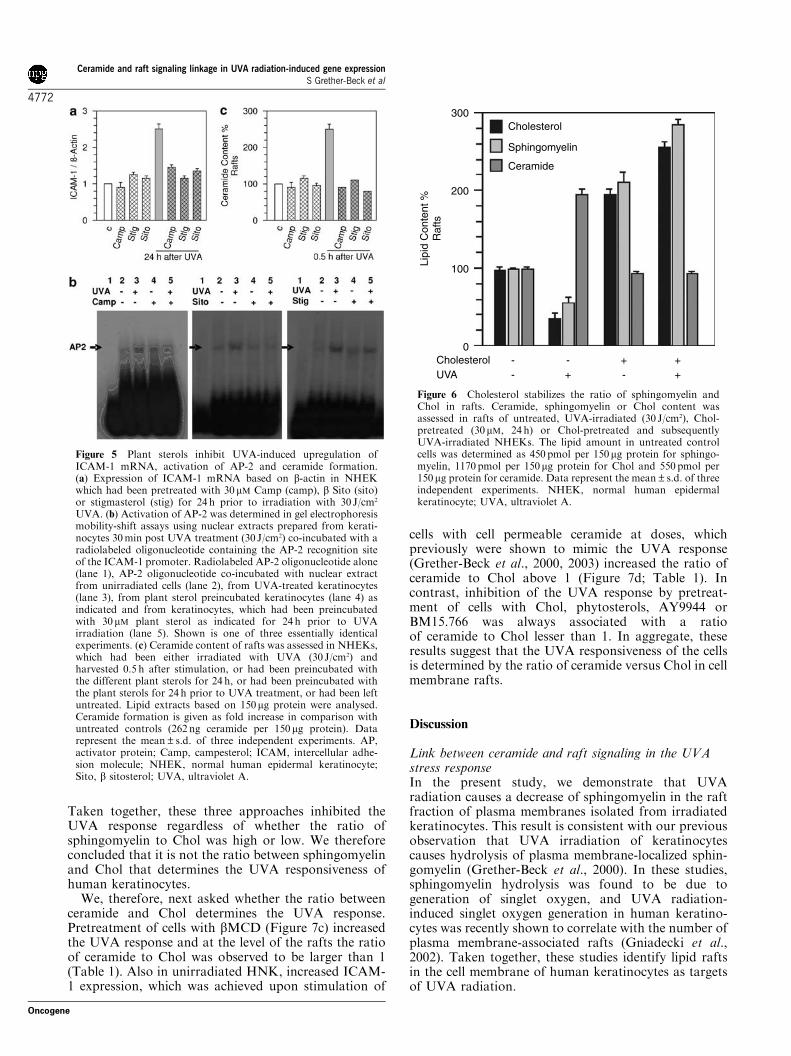
Taken together, these three approaches inhibited theUVA response regardless of whether the ratio ofsphingomyelin to Chol was high or low. We thereforeconcluded that it is not the ratio between sphingomyelinand Chol that determines the UVA responsiveness ofhuman keratinocytes.
We, therefore, next asked whether the ratio betweenceramide and Chol determines the UVA response.Pretreatment of cells with bMCD (Figure 7c) increasedthe UVA response and at the level of the rafts the ratioof ceramide to Chol was observed to be larger than 1(Table 1). Also in unirradiated HNK, increased ICAM-1 expression, which was achieved upon stimulation of
cells with cell permeable ceramide at doses, whichpreviously were shown to mimic the UVA response(Grether-Beck et al., 2000, 2003) increased the ratio ofceramide to Chol above 1 (Figure 7d; Table 1). Incontrast, inhibition of the UVA response by pretreat-ment of cells with Chol, phytosterols, AY9944 orBM15.766 was always associated with a ratioof ceramide to Chol lesser than 1. In aggregate, theseresults suggest that the UVA responsiveness of the cellsis determined by the ratio of ceramide versus Chol in cellmembrane rafts.
Discussion
Link between ceramide and raft signaling in the UVAstress responseIn the present study, we demonstrate that UVAradiation causes a decrease of sphingomyelin in the raftfraction of plasma membranes isolated from irradiatedkeratinocytes. This result is consistent with our previousobservation that UVA irradiation of keratinocytescauses hydrolysis of plasma membrane-localized sphin-gomyelin (Grether-Beck et al., 2000). In these studies,sphingomyelin hydrolysis was found to be due togeneration of singlet oxygen, and UVA radiation-induced singlet oxygen generation in human keratino-cytes was recently shown to correlate with the number ofplasma membrane-associated rafts (Gniadecki et al.,2002). Taken together, these studies identify lipid raftsin the cell membrane of human keratinocytes as targetsof UVA radiation.
Figure 5 Plant sterols inhibit UVA-induced upregulation ofICAM-1 mRNA, activation of AP-2 and ceramide formation.(a) Expression of ICAM-1 mRNA based on b-actin in NHEKwhich had been pretreated with 30 mM Camp (camp), b Sito (sito)or stigmasterol (stig) for 24 h prior to irradiation with 30 J/cm2
UVA. (b) Activation of AP-2 was determined in gel electrophoresismobility-shift assays using nuclear extracts prepared from kerati-nocytes 30min post UVA treatment (30 J/cm2) co-incubated with aradiolabeled oligonucleotide containing the AP-2 recognition siteof the ICAM-1 promoter. Radiolabeled AP-2 oligonucleotide alone(lane 1), AP-2 oligonucleotide co-incubated with nuclear extractfrom unirradiated cells (lane 2), from UVA-treated keratinocytes(lane 3), from plant sterol preincubated keratinocytes (lane 4) asindicated and from keratinocytes, which had been preincubatedwith 30mM plant sterol as indicated for 24 h prior to UVAirradiation (lane 5). Shown is one of three essentially identicalexperiments. (c) Ceramide content of rafts was assessed in NHEKs,which had been either irradiated with UVA (30 J/cm2) andharvested 0.5 h after stimulation, or had been preincubated withthe different plant sterols for 24 h, or had been preincubated withthe plant sterols for 24 h prior to UVA treatment, or had been leftuntreated. Lipid extracts based on 150mg protein were analysed.Ceramide formation is given as fold increase in comparison withuntreated controls (262 ng ceramide per 150mg protein). Datarepresent the mean±s.d. of three independent experiments. AP,activator protein; Camp, campesterol; ICAM, intercellular adhe-sion molecule; NHEK, normal human epidermal keratinocyte;Sito, b sitosterol; UVA, ultraviolet A.
Lipi
d C
onte
nt %
Raf
ts
Cholesterol
Sphingomyelin
Ceramide
CholesterolUVA
--
--++ +
+
300
200
100
0
Figure 6 Cholesterol stabilizes the ratio of sphingomyelin andChol in rafts. Ceramide, sphingomyelin or Chol content wasassessed in rafts of untreated, UVA-irradiated (30 J/cm2), Chol-pretreated (30mM, 24 h) or Chol-pretreated and subsequentlyUVA-irradiated NHEKs. The lipid amount in untreated controlcells was determined as 450 pmol per 150mg protein for sphingo-myelin, 1170 pmol per 150mg protein for Chol and 550 pmol per150mg protein for ceramide. Data represent the mean±s.d. of threeindependent experiments. NHEK, normal human epidermalkeratinocyte; UVA, ultraviolet A.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4772
Oncogene
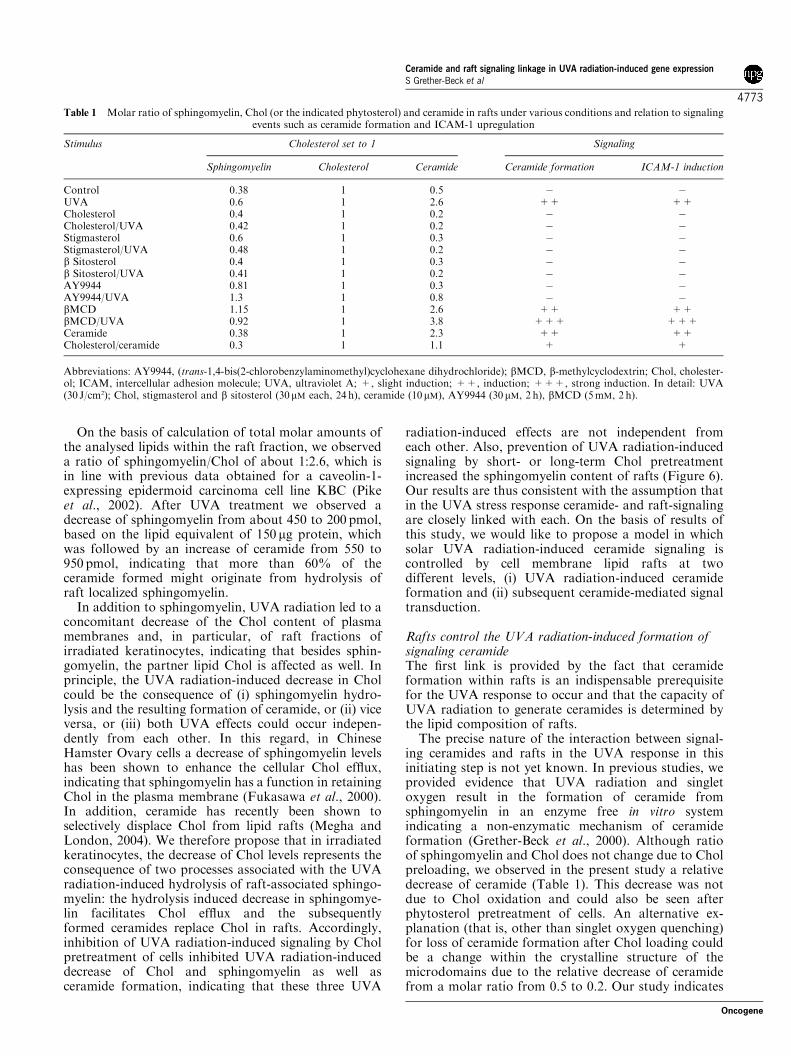
On the basis of calculation of total molar amounts ofthe analysed lipids within the raft fraction, we observeda ratio of sphingomyelin/Chol of about 1:2.6, which isin line with previous data obtained for a caveolin-1-expressing epidermoid carcinoma cell line KBC (Pikeet al., 2002). After UVA treatment we observed adecrease of sphingomyelin from about 450 to 200 pmol,based on the lipid equivalent of 150 mg protein, whichwas followed by an increase of ceramide from 550 to950 pmol, indicating that more than 60% of theceramide formed might originate from hydrolysis ofraft localized sphingomyelin.
In addition to sphingomyelin, UVA radiation led to aconcomitant decrease of the Chol content of plasmamembranes and, in particular, of raft fractions ofirradiated keratinocytes, indicating that besides sphin-gomyelin, the partner lipid Chol is affected as well. Inprinciple, the UVA radiation-induced decrease in Cholcould be the consequence of (i) sphingomyelin hydro-lysis and the resulting formation of ceramide, or (ii) viceversa, or (iii) both UVA effects could occur indepen-dently from each other. In this regard, in ChineseHamster Ovary cells a decrease of sphingomyelin levelshas been shown to enhance the cellular Chol efflux,indicating that sphingomyelin has a function in retainingChol in the plasma membrane (Fukasawa et al., 2000).In addition, ceramide has recently been shown toselectively displace Chol from lipid rafts (Megha andLondon, 2004). We therefore propose that in irradiatedkeratinocytes, the decrease of Chol levels represents theconsequence of two processes associated with the UVAradiation-induced hydrolysis of raft-associated sphingo-myelin: the hydrolysis induced decrease in sphingomye-lin facilitates Chol efflux and the subsequentlyformed ceramides replace Chol in rafts. Accordingly,inhibition of UVA radiation-induced signaling by Cholpretreatment of cells inhibited UVA radiation-induceddecrease of Chol and sphingomyelin as well asceramide formation, indicating that these three UVA
radiation-induced effects are not independent fromeach other. Also, prevention of UVA radiation-inducedsignaling by short- or long-term Chol pretreatmentincreased the sphingomyelin content of rafts (Figure 6).Our results are thus consistent with the assumption thatin the UVA stress response ceramide- and raft-signalingare closely linked with each. On the basis of results ofthis study, we would like to propose a model in whichsolar UVA radiation-induced ceramide signaling iscontrolled by cell membrane lipid rafts at twodifferent levels, (i) UVA radiation-induced ceramideformation and (ii) subsequent ceramide-mediated signaltransduction.
Rafts control the UVA radiation-induced formation ofsignaling ceramideThe first link is provided by the fact that ceramideformation within rafts is an indispensable prerequisitefor the UVA response to occur and that the capacity ofUVA radiation to generate ceramides is determined bythe lipid composition of rafts.
The precise nature of the interaction between signal-ing ceramides and rafts in the UVA response in thisinitiating step is not yet known. In previous studies, weprovided evidence that UVA radiation and singletoxygen result in the formation of ceramide fromsphingomyelin in an enzyme free in vitro systemindicating a non-enzymatic mechanism of ceramideformation (Grether-Beck et al., 2000). Although ratioof sphingomyelin and Chol does not change due to Cholpreloading, we observed in the present study a relativedecrease of ceramide (Table 1). This decrease was notdue to Chol oxidation and could also be seen afterphytosterol pretreatment of cells. An alternative ex-planation (that is, other than singlet oxygen quenching)for loss of ceramide formation after Chol loading couldbe a change within the crystalline structure of themicrodomains due to the relative decrease of ceramidefrom a molar ratio from 0.5 to 0.2. Our study indicates
Table 1 Molar ratio of sphingomyelin, Chol (or the indicated phytosterol) and ceramide in rafts under various conditions and relation to signalingevents such as ceramide formation and ICAM-1 upregulation
Stimulus Cholesterol set to 1 Signaling
Sphingomyelin Cholesterol Ceramide Ceramide formation ICAM-1 induction
Control 0.38 1 0.5 � �UVA 0.6 1 2.6 ++ ++Cholesterol 0.4 1 0.2 � �Cholesterol/UVA 0.42 1 0.2 � �Stigmasterol 0.6 1 0.3 � �Stigmasterol/UVA 0.48 1 0.2 � �b Sitosterol 0.4 1 0.3 � �b Sitosterol/UVA 0.41 1 0.2 � �AY9944 0.81 1 0.3 � �AY9944/UVA 1.3 1 0.8 � �bMCD 1.15 1 2.6 ++ ++bMCD/UVA 0.92 1 3.8 +++ +++Ceramide 0.38 1 2.3 ++ ++Cholesterol/ceramide 0.3 1 1.1 + +
Abbreviations: AY9944, (trans-1,4-bis(2-chlorobenzylaminomethyl)cyclohexane dihydrochloride); bMCD, b-methylcyclodextrin; Chol, cholester-ol; ICAM, intercellular adhesion molecule; UVA, ultraviolet A; +, slight induction; ++, induction; +++, strong induction. In detail: UVA(30 J/cm2); Chol, stigmasterol and b sitosterol (30mM each, 24 h), ceramide (10mM), AY9944 (30 mM, 2 h), bMCD (5mM, 2 h).
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4773
Oncogene
that in human epidermal keratinocytes a UVA responsecan be elicited if the ratio of ceramide versus Chol isgreater than 1 and that no response occurs if the ratio islesser than 1. Accordingly, tight packing of Chol tosphingomyelin in raft structures, which is likely to beenhanced after Chol pretreatment and the concomitant
increase in raft Chol content (Figure 6), is thought toaffect the crystalline structure of sphingomyelin (Brownand London, 2000) and might thereby determine thesensitivity of sphingomyelin molecules towards UVAradiation-induced, singlet oxygen-mediated formationof ceramides from sphingomyelin. As a consequence,Chol rich rafts would be relatively resistant againstUVA radiation-induced formation of ceramides, thesubsequent decrease of sphingomyelin levels and theresulting decrease in the Chol content, as observed in thepresent study (Figure 6). Our results are thus consistentwith the idea that the ceramide content within or aroundthe microdomains affects the sensitivity of sphingo-myelin towards singlet-oxygen-induced hydrolysis. Wepropose that prevention of the UVA stress response byChol is due to its ‘raft-stabilizing’ properties. Accord-ingly, a raft stabilizing function of Chol, which serves tofill the voids between the large and bulky glycero-sphingolipids has been described by Gulbins and Li(2006). According to this concept, pharmacologicalextraction of Chol destroys membrane rafts. Moreover,generation of ceramide within rafts, no matter by whichmechanism they are formed, dramatically alters thebiophysical properties of these membrane domains,since ceramide molecules have the tendency to sponta-neously self-associate to small ceramide-enriched mem-brane microdomains (Kolesnick et al., 2000).
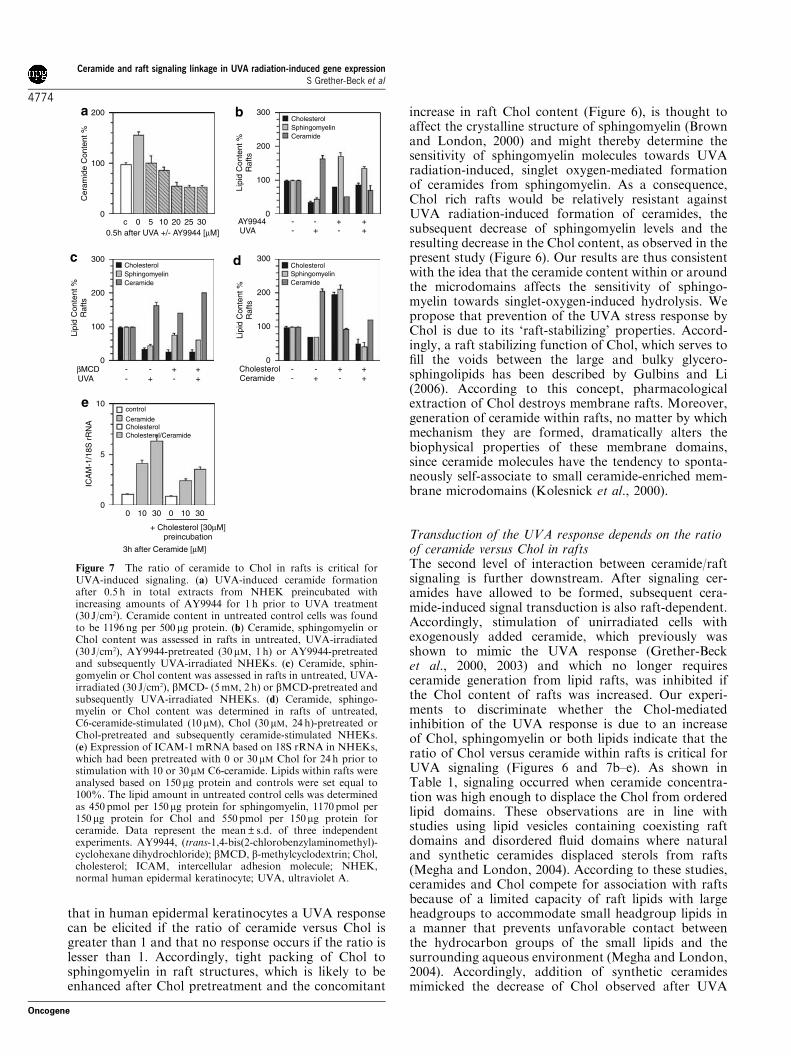
Transduction of the UVA response depends on the ratioof ceramide versus Chol in raftsThe second level of interaction between ceramide/raftsignaling is further downstream. After signaling cer-amides have allowed to be formed, subsequent cera-mide-induced signal transduction is also raft-dependent.Accordingly, stimulation of unirradiated cells withexogenously added ceramide, which previously wasshown to mimic the UVA response (Grether-Becket al., 2000, 2003) and which no longer requiresceramide generation from lipid rafts, was inhibited ifthe Chol content of rafts was increased. Our experi-ments to discriminate whether the Chol-mediatedinhibition of the UVA response is due to an increaseof Chol, sphingomyelin or both lipids indicate that theratio of Chol versus ceramide within rafts is critical forUVA signaling (Figures 6 and 7b–e). As shown inTable 1, signaling occurred when ceramide concentra-tion was high enough to displace the Chol from orderedlipid domains. These observations are in line withstudies using lipid vesicles containing coexisting raftdomains and disordered fluid domains where naturaland synthetic ceramides displaced sterols from rafts(Megha and London, 2004). According to these studies,ceramides and Chol compete for association with raftsbecause of a limited capacity of raft lipids with largeheadgroups to accommodate small headgroup lipids ina manner that prevents unfavorable contact betweenthe hydrocarbon groups of the small lipids and thesurrounding aqueous environment (Megha and London,2004). Accordingly, addition of synthetic ceramidesmimicked the decrease of Chol observed after UVA
200
100
00c 5 10 20 25 30
Cer
amid
e C
onte
nt %
0.5h after UVA +/- AY9944 [µM]
CholesterolSphingomyelinCeramide
CholesterolSphingomyelinCeramide
CholesterolSphingomyelinCeramide
300
200
100
0
Lipi
d C
onte
nt %
Raf
ts
AY9944UVA
--
--++ +
+
300
200
100
0
Lipi
d C
onte
nt %
Raf
ts
βMCDUVA
--
--++ +
+
300
200
100
0
Lipi
d C
onte
nt %
Raf
ts
CholesterolCeramide
--
--++ +
+
10
5
010 1030 3000
control
CeramideCholesterol
+ Cholesterol [30µM]preincubation
Cholesterol/Ceramide
3h after Ceramide [µM]
ICA
M-1
/18S
rR
NA
Figure 7 The ratio of ceramide to Chol in rafts is critical forUVA-induced signaling. (a) UVA-induced ceramide formationafter 0.5 h in total extracts from NHEK preincubated withincreasing amounts of AY9944 for 1 h prior to UVA treatment(30 J/cm2). Ceramide content in untreated control cells was foundto be 1196 ng per 500mg protein. (b) Ceramide, sphingomyelin orChol content was assessed in rafts in untreated, UVA-irradiated(30 J/cm2), AY9944-pretreated (30mM, 1 h) or AY9944-pretreatedand subsequently UVA-irradiated NHEKs. (c) Ceramide, sphin-gomyelin or Chol content was assessed in rafts in untreated, UVA-irradiated (30 J/cm2), bMCD- (5mM, 2 h) or bMCD-pretreated andsubsequently UVA-irradiated NHEKs. (d) Ceramide, sphingo-myelin or Chol content was determined in rafts of untreated,C6-ceramide-stimulated (10 mM), Chol (30mM, 24 h)-pretreated orChol-pretreated and subsequently ceramide-stimulated NHEKs.(e) Expression of ICAM-1 mRNA based on 18S rRNA in NHEKs,which had been pretreated with 0 or 30mM Chol for 24 h prior tostimulation with 10 or 30mM C6-ceramide. Lipids within rafts wereanalysed based on 150mg protein and controls were set equal to100%. The lipid amount in untreated control cells was determinedas 450 pmol per 150mg protein for sphingomyelin, 1170 pmol per150mg protein for Chol and 550pmol per 150mg protein forceramide. Data represent the mean±s.d. of three independentexperiments. AY9944, (trans-1,4-bis(2-chlorobenzylaminomethyl)-cyclohexane dihydrochloride); bMCD, b-methylcyclodextrin; Chol,cholesterol; ICAM, intercellular adhesion molecule; NHEK,normal human epidermal keratinocyte; UVA, ultraviolet A.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4774
Oncogene

irradiation (Figures 1b and 7d) and preloading ofkeratinocytes with Chol could partially inhibit cera-mide-induced gene expression (Figure 7e). These resultsare supported by studies using atomic-force microscopyto study the effects of ceramide generation by in situenzymatic hydrolysis of sphingomyelin in phase sepa-rated lipid bilayers that have sphingomyelin/Chol richdomains surrounded by a fluid phase (Johnston andJohnston, 2006). We showed that in situ generation ofceramide resulted in the restructuring of the bilayer togive areas of fluid phase, areas that have a distributionof domains similar to the original bilayer and areascontaining clusters or domains.
In conclusion, our study shows that ceramide and raftsignaling are tightly linked in solar UVA radiation-induced signal transduction in a way that the lipidcomposition of rafts controls the generation and activityof signaling ceramides. As raft microdomains alsocontain a variety of signaling proteins (Simons andToomre, 2000), further studies will have to address thequestion of how the ceramide signal is transposed fromthe outer leaflet of the membrane to the cytosol, forexample, by modulation of raft-associated proteins.
Clinical implications for photoprotection andphotodermatosisAbrogation of the UVA-stress response was not specificfor the Chol molecule, but could be mimicked by plantsterols, for example, Camp, b Sito and sigmasterol.These major plant sterols stabilized the cell membraneto a similar extend as Chol in our experiments (Figure 7).In contrast to invertebrates typically converting phytos-terols to Chol by de-alkylating C24 in the side chain,thereby furnishing Chol needed by membranes, verte-brates are not able to de-alkylate sterols with bulkierside chains such as Camp (24a-methylcholesterol), b-Sito (24a-ethylcholesterol) or stigmasterol (D22,24a-ethylcholesterol) (Xu et al., 2005). Our results are inline with recent observations that plant sterols such asCamp and b-Sito can substitute for Chol as the primarysterol in the plasma membrane without being convertedto Chol (Xu et al., 2005). UVA radiation-inducedsignaling in human keratinocytes is thought to con-tribute to a variety of solar UV radiation-induceddetrimental effects including photocarcinogenesis,photoaging and the pathogenesis of UVA-sensitive skindiseases such as polymorphic light eruption. The recentobservation that UVA radiation-induced gene expres-sion can be completely abrogated by pretreating humanepidermal keratinocytes in vitro with Chol or plantsterols indicates that topical application of thesemolecules in appropriate galenic formulations to humanskin may be used for photoprotection, for example, insunscreens in combination with UV filters.
A modulation in the lipid composition of rafts haspreviously been observed in human cells in response to avariety of exogenous stimuli including proinflammatorycytokines, hormones or growth factors (Liu andAnderson, 1995; Vainio et al., 2002; Yang et al., 2004).In these systems, alterations of the Chol content of lipid
rafts were of functional relevance for signal transduc-tion. Similarly, in the present study, modulation ofthe Chol content significantly affected the capacity ofhuman keratinocytes to mount a UVA response.Accordingly, a strong decrease in the Chol as observedby bMCD extraction was associated with an increasedsusceptibility towards UVA radiation, whereas Cholenrichment had the opposite effect. It should be notedthat patients with Smith–Lemli–Opitz syndrome fre-quently exhibit an increased susceptibility towards solarUV radiation (Charman et al., 1998; Anstey and Taylor,1999). This autosomal, recessive genetic disorder is dueto deletions as well as missense or nonsense mutations inthe gene encoding for 7-dehydrocholesterol reductase,which cause a lowered expression of the correspondingprotein D7-sterol-reductase (Fitzky et al., 1998). As aconsequence, Smith–Lemli–Opitz patients have de-creased Chol serum levels (Smith et al., 1964; Tintet al., 1994; Batta et al., 1995). Action spectrum studieshave revealed, that Smith–Lemli–Opitz patients areexquisitely sensitive to UVA radiation, whereas theirUVB response is normal (Anstey et al., 1999). Also, anenhanced induction of apoptosis upon UVA irradiationcould recently be observed in keratinocytes treated withAY9944, bMCD and 7-dehydrocholesterol (Valenciaand Kochevar, 2006). In addition, a recent case reportindicates that dietary supplementation with Cholsignificantly decreased UVA photosensitivity in anaffected child (Azurdia et al., 2001). These clinicalobservations are in line with our current observationthat a decrease in keratinocyte Chol content increases,whereas pretreatment with Chol decreases, the capacityof human keratinocytes to mount a UVA-stressresponse (Figures 2b, c and e). They also indicate thepossibility that UVA hypersensitivity in Smith–Lemli–Opitz patients is, at least in part, due to disturbed raftsignaling.
Materials and methods
MaterialsRecombinant human interferon-g was obtained from R&DSystems (Wiesbaden, Germany). Chol; the plant sterols Camp,b Sito and stigmasterol; AY9944; BM15.766 (4-[2-[4-[3-(4-chlorophenyl)-2-propenyl]-1-piperazinyl]ethyl]benzoic acidsulfate) and bMCD were purchased from Sigma-Aldrich(Munich, Germany). Lipid standards for ceramide, sphingo-myelin and ganglioside GM1 were from Merck Biosciences(Bad Soden, Germany) and for 7-ketosterol, as well as 7b-hydroxycholesterol (7b-OH), were from Sigma-Aldrich.b-Methylcyclodextrin was dissolved in distilled water as a250-mM stock solution. Sterols were dissolved in ethanol asa 10-mM stock solution and added to the cell culture medium24 h prior to irradiation in a final concentration of 30mM.
Cell cultureLong-term cultured, normal human keratinocytes preparedfrom neonatal foreskin were cultured under serum freeconditions for up to five passages (Grether-Beck et al., 1996,2000, 2003, 2005). Depletion of Chol from cytoplasmicmembranes was achieved through a 2-h preincubation ofcells with 5mM bMCD as previously described (Ridgwayet al., 1998).
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4775
Oncogene

UV irradiationFor UVA irradiation, medium was replaced by phosphate-buffered saline, lids were removed and cells were exposedto a dose of 30 J/cm2 UVA using a UVASUN 24 000 system(Sellas GmbH, Gevelsberg, Germany) as previously described(Grether-Beck et al., 1996, 2000, 2005).
Isolation of plasma membranesCell membranes were isolated from pelleted cells afterhomogenization by a Dounce homogenizer according tostandard differential centrifugation procedures. Identity ofthe plasma membrane was assessed by western blotting forvery-late antigen (VLA-2a¼ integrin-a2) (Emsley et al., 1997).In addition, isolated fractions were routinely analysed forcontamination with mitochondria and endoplasmic reticulumby western blotting for the BCL-2-family member myeloid cellleukemia 1 (Yang et al., 1995) and the 78-kDa glucose-regulated protein (GRP78/BiP; Linnik and Herscovitz, 1998),with negative results (data not shown).
Isolation of raftsKeratinocytes grown on 15-cm dishes were scraped off theplate and pelleted in phosphate-buffered saline. Rafts wereisolated as detergent-insoluble glycosphingolipid complexes(Benting et al., 1999). Detergent extractions were performed onice with pre-chilled solutions. Keratinocytes were resuspendedin 100ml 10mM Tris–HCl, pH 7.4, 150mM NaCl, 1mM EDTAwith CLAP (chymostatin, leupeptin, antipain and pepstatin A,25 mgml�1 each final), and then one volume of 2% TritonX-100 in the same buffer was added. After 30min of incubation,the lysate was adjusted to 40% Optiprep (Progen BiotechnikGmbH, Heidelberg, Germany) overlaid with 30 and 5%Optiprep and spun down for 4 h in an L8-M centrifuge(Beckman, Fullerton, CA, USA) with a 50.2-Ti rotor at40 000 r.p.m. (B168 448 g) at 4 1C. The resulting seven frac-tions were collected from top and the raft fraction wasidentified as the ganglioside GM1-positive fraction (data notshown) by high-performance TLC.
Lipid extractionCells were harvested at the indicated time points by scrapingthe cells off the plate on ice. Pellets were washed and sonicated.Quantification of lipids was performed using either 500 mg ofprotein (Bradford, 1976) of total extracts or 150 mg of proteinof rafts fraction for Folch extraction (Folch et al., 1957).Analysis of ceramides included a mild alkaline hydrolysis. Thelower phase of Folch extraction was evaporated undernitrogen. The lipids were dissolved in chloroform/methanol(2:1, v/v).
High-performance TLCSamples and standards were separated on silica-gel high-performance TLC plates (20� 10 cm) (Merck 60F 254s)prewashed for 60min in 2-propanol and dried 30min at120 1C. Samples and standards were applied to the TLC platesusing a CAMAG Linomat IV (CAMAG, Berlin, Germany) inbands of 0.8 cm via a spray-on technique (10 ml per sample) inwhich nitrogen carrier gas atomizes the sample from a syringeonto the plate, which is moving back and forth under theatomizer. Samples were separated using an Automated Multi-ple Development procedure on an Automated MultipleDevelopment 2 device (CAMAG). This procedure consistedof seven repeated developments of the chromatogram using astepwise elution gradient with methanol, dichloromethaneand n-hexane (methanol/dichloromethane/n-hexane: 100/0/0;10/90/0; 9/91/0; 8/92/0; 3/97/0; 2/98/0; 0/0/100) on a CAMAG
Automated Multiple Development 2 device. Visualization wasperformed by postchromatic derivatization, as describedpreviously (Grether-Beck et al., 2005). Quantification wasperformed with a CAMAG TLC Scanner II and CATSsoftware at 550 nm, with a plot of peak area versus weightspotted for a series of standards using a second-orderpolynomial calibration.
Determination of Chol photo-oxidation productSinglet oxygen-induced primary oxidation products of Cholare 3b-hydroxy-5a-cholest-6-ene-5-hydroperoxide (5a-OOH),3b-hydroxycholest-4-ene-6a-hydroperoxide (6a-OOH) and 3b-hydroxy-cholest-4-ene-6b-hydroperoxide (6b-OOH). Theseproducts were detected by TLC using heptane ethyl acetate(1:1, v/v) (Girotti and Korytowski, 2000). Free radical-mediated hydroperoxides 7a-hydroxycholesterol and 7b-OHwere generated from commercially available 7-ketocholesterol(7-Ketochol) by addition of NaBH4 (Bachowski et al., 1988).
Differential reverse transcriptase–PCRTotal RNA was isolated using RNeasy Total RNA kits(Qiagen, Hilden, Germany). Two-step real-time reversetranscriptase–PCR was performed to measure gene expression.SuperScript III First-Strand Synthesis System for reversetranscriptase–PCR (Invitrogen, Karlsruhe, Germany) wasused for the reverse transcription step with random hexamers.For each gene, a specific PCR primer pair was designed usingPrimer Express 2.0 software (Applied Biosystems, Darmstadt,Germany) based on the cDNA sequence published asreferenced: b-actin 50-CCTGGCACCCAGCACAAT-30/50-GCCGATCCACACGGAGTACT-30 (Ponte et al., 1984;Vandekerckhove and Weber, 1978); 18S rRNA 50-GCCGCTAGAGGTGAAATTCTTG-30/50-CATTCTTGGCAAATGCTTTCG0-30 (McCallum and Maden, 1985) and ICAM-1 50-CCTGGCACCCAGCACAAT-30/50-GCCGATCCACACGGAGTACT-30 (Staunton et al., 1988). The PCR reactions werecarried with on an Opticon 1 (MJ Research, Waltham, MA,USA) using SYBR Green PCR Master Mix (AppliedBiosystems). Each reverse transcriptase–PCR experiment wasperformed twice.
Nuclear extracts and gel electrophoresis mobility-shift assayNuclear extracts were prepared according to the proceduredescribed by Dignam et al. (1983). Gel electrophoresis mobility-shift assays to detect AP-2 activity were performed as previouslydescribed (Grether-Beck et al., 1996, 2000, 2003, 2005).
Abbreviations
�MCD, �-methylcyclodextrin; 7a-OH, 7a-hydroxycholesterol;7�-OH, 7�-hydroxycholesterol; 7-Ketochol, 7-ketocholesterol;AP-2, activator protein 2; Camp, campesterol; Chol, choles-terol; CholOOH, cholesterol hydroperoxide; ICAM-1, inter-cellular adhesion molecule-1; IFN-g, Interferon-g; NHEK,normal human epidermal keratinocytes; Sito, b sitosterol;Stigma, stigmasterol; UVA, ultraviolet A.
Acknowledgements
We thank H Sies and K-D Kroencke (Duesseldorf), as well asE Proksch (Kiel), for helpful discussion and critical review ofthe manuscript. This work was supported by the DeutscheForschungsgemeinschaft Grant SFB 503, project B2, andSFB 728.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4776
Oncogene

References
Anderson RGW. (1993). Caveolae: where incoming and outgoingmessengers meet. Proc Natl Acad Sci USA 90: 10909–10913.
Anstey AV, Ryan A, Rhodes LE, Charman CR, Arlett CF, TyrrellRM et al. (1999). Characterization of photosensitivity in the Smith–Lemli–Opitz syndrome: a new congenital photosensitivity syndrome.Br J Dermatol 141: 406–414.
Anstey AV, Taylor CR. (1999). Photosensitivity in the Smith–Lemli–Opitz syndrome: the US experience of a new congenital photo-sensitivity syndrome. J Am Acad Dermatol 41: 121–123.
Azurdia RM, Anstey AV, Rhodes LE. (2001). Cholesterol supple-mentation objectively reduces photosensitivity in the Smith–Lemli–Opitz syndrome. J Dermatol 144: 143–145.
Bachowski GJ, Thomas JP, Girotti AW. (1988). Ascorbate-enhancedlipid peroxidation in photooxidized cell membranes: cholesterolproduct analysis as a probe of reaction mechanism. Lipids 23:580–586.
Batta AK, Tint GS, Shefer S, Abuelo D, Salen G. (1995). Identificationof 8-dehydrocholesterol (cholesta-5,8-dien-3 beta-ol) in patients withSmith–Lemli–Opitz syndrome. J Lipid Res 36: 705–713.
Benting JH, Rietveld AG, Simons K. (1999). N-glycans mediate theapical sorting of a GPI-anchored, raft-associated protein in Madin–Darby canine kidney cells. J Cell Biol 146: 313–320.
Bradford MM. (1976). A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein–dye binding. Anal Biochem 72: 248–254.
Brown DA, London E. (1998). Functions of lipid rafts in biologicalmembranes. Annu Rev Cell Dev Biol 14: 111–136.
Brown DA, London E. (2000). Structure and function of sphingolipid-and cholesterol-rich membrane rafts. J Biol Chem 275: 17221–17224.
Charman CR, Ryan A, Tyrrell RM, Pearse AD, Arlett CF, Kurwa HAet al. (1998). Photosensitivity associated with the Smith–Lemli–Opitz syndrome. Br J Dermatol 138: 885–888.
Dignam JD, Martin PL, Skastry BS, Roeder RGG. (1983). Eukaryoticgene transcription with purified components. Methods Enzymol 101:582–598.
Emsley J, King SL, Bergelson JM, Liddington RC. (1997). Crystalstructure of the I domain from integrin alpha2beta1. J Biol Chem
272: 28512–28517.Fitzky BU, Witsch-Baumgartner M, Erdel M, Lee JN, Paik Y-K,Glossmann H et al. (1998). Mutations in the delta7-sterol reductasegene in patients with the Smith–Lemli–Opitz syndrome. Proc Natl
Acad Sci USA 95: 8181–8186.Folch J, Lees M, Sloane-Stanley GH. (1957). A simple method for theisolation and purification of total lipides from animal tissues. J Biol
Chem 226: 497–509.Fukasawa M, Nishijima M, Itabe H, Takano T, Hanada K. (2000).Reduction of sphingomyelin level without accumulation of ceramidein Chinese hamster ovary cells affects detergent-resistant membranedomains and enhances cellular cholesterol efflux to methyl-beta-cyclodextrin. J Biol Chem 275: 34028–34034.
Girotti AW. (1992). Photosensitized oxidation of cholesterol inbiological systems: reaction pathways, cytotoxic effects and defensemechanisms. J Photochem Photobiol 13: 105–118.
Girotti AW. (1998). Lipid hydroperoxide generation, turnover, andeffector action in biological systems. J Lipid Res 39: 1529–1542.
Girotti AW, Korytowski W. (2000). Cholesterol as a singlet oxygendetector in biological systems. Methods Enzymol 319: 85–100.
Gniadecki R, Christoffersen N, Wulf HC. (2002). Cholesterol-richplasma membrane domains (lipid rafts) in keratinocytes: importancein the baseline and UVA-induced generation of reactive oxygenspecies. J Invest Dermatol 118: 582–588.
Grether-Beck S, Bonizzi G, Schmitt-Brenden H, Felsner I, Timmer A,Sies H et al. (2000). Non-enzymatic triggering of the ceramidesignalling cascade by solar UVA radiation. EMBO J 19: 5793–5800.
Grether-Beck S, Felsner I, Brenden H, Krutmann J. (2003).Mitochondrial cytochrome c release mediates ceramide-inducedactivator protein 2 activation and gene expression in keratinocytes.J Biol Chem 278: 47498–47507.
Grether-Beck S, Olaizola-Horn S, Schmitt H, Grewe M, Jahnke A,Johnson JP et al. (1996). Activation of transcription factor AP-2mediates UVA radiation- and singlet oxygen-induced expression ofthe human intercellular adhesion molecule 1 gene. Proc Natl Acad
Sci USA 93: 14586–14591.Grether-Beck S, Timmer A, Felsner I, Brenden H, Brammertz D,
Krutmann J. (2005). Ultraviolet A-induced signaling involves aceramide-mediated autocrine loop leading to ceramide de novosynthesis. J Invest. Dermatol 125: 545–553.
Gulbins E, Li PL. (2006). Physiological and pathophysiological aspectsof ceramide. Am J Physiol Regul Integr Comp Physiol 290: R11–R26.
Gustavsson J, Parpal S, Karlsson M, Ramsing C, Thorn H, Borg Met al. (1999). Localization of the insulin receptor in caveolae ofadipocyte plasma membrane. FASEB J 13: 1961–1971.
Johnston I, Johnston LJ. (2006). Localization of the insulin receptor incaveolae of adipocyte plasma membrane. Langmuir 22:11284–11289.
Kashkar H, Wiegmann K, Yazdanpanah B, Haubert D, Kronke M.(2005). Acid sphingomyelinase is indispensable for UV light-inducedBax conformational change at the mitochondrial membrane. J Biol
Chem 280: 20804–20813.Kim JB, Spotts GD, Halvorsen Y-D, Shih H-M, Ellenberger T et al.
(1995). Dual DNA binding specificity of ADD1/SREBP1 controlledby a single amino acid in the basic helix–loop–helix domain. Mol
Cell Biol 15: 2582–2588.Kolesnick RN, Goni FM, Alonso A. (2000). Compartmentalization of
ceramide signaling: physical foundations and biological effects.J Cell Physiol 184: 285–300.
Korytowski W, Girotti AW. (1999). Singlet oxygen adducts ofcholesterol: photogeneration and reductive turnover in membranesystems. Photochem Photobiol 70: 484–489.
Linnik KM, Herscovitz H. (1998). Multiple molecular chaperonesinteract with apolipoprotein B during its maturation. The networkof endoplasmic reticulum-resident chaperones (ERp72, GRP94,calreticulin, and BiP) interacts with apolipoprotein b regardless ofits lipidation state. J Biol Chem 273: 21368–21373.
Liu P, Anderson RGW. (1995). Compartmentalized production ofceramide at the cell surface. J Biol Chem 270: 27179–27185.
McCallum FS, Maden BE. (1985). Human 18S ribosomal RNAsequence inferred from DNA sequence. Variations in 18S sequencesand secondary modification patterns between vertebrates. Biochem
J 232: 725–733.Megha, London E. (2004). Ceramide selectively displaces cholesterol
from ordered lipid domains (rafts): implications for lipid raftstructure and function. J Biol Chem 279: 9997–10004.
Neufeld EB, Cooney AM, Pitha J, Dawidowicz EA, Dwyer NK,Pentchev PG et al. (1996). Intracellular trafficking of cholesterolmonitored with a cyclodextrin. J Biol Chem 271: 21604–21613.
Pike LJ, Han X, Chung K-N, Gross RW. (2002). Lipid rafts areenriched in arachidonic acid and plasmenylethanolamine and theircomposition is independent of caveolin-1 expression: a quantitativeelectrospray ionization/mass spectrometric analysis. Biochemistry
41: 2075–2088.Ponte P, Ng SY, Engel J, Gunning P, Kedes L. (1984). Evolutionary
conservation in the untranslated regions of actin mRNAs:DNA sequence of a human beta-actin cDNA. Nucleic Acids Res
12: 1687–1696.Reiterer G, MacDonald R, Browning JD, Morrow J, Matveev SV,
Daugherty A et al. (2005). Zinc deficiency increases plasma lipidsand atherosclerotic markers in LDL-receptor-deficient mice. J Nutr
135: 2114–2118.Ridgway ND, Lagace TA, Cook HW, Byers DM. (1998). Differential
effects of sphingomyelin hydrolysis and cholesterol transport onoxysterol-binding protein phosphorylation and Golgi localization.J Biol Chem 273: 31621–31628.
Rotolo JA, Zhang J, Donepudi M, Lee H, Fuks Z, Kolesnick R.(2005). Caspase-dependent and -independent activation of acidsphingomyelinase signaling. J Biol Chem 280: 26425–26434.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4777
Oncogene

Schmuth M, Elias PM, Hanley K, Lau P, Moser A, Willson TM et al.(2004). The effect of LXR activators on AP-1 proteins inkeratinocytes. J Invest Dermatol 123: 41–48.
Schnitzer JE, Oh P, Pinney E, Allard J. (1994). Filipin-sensitivecaveolae-mediated transport in endothelium: reduced transcytosis,scavenger endocytosis, and capillary permeability of select macro-molecules. J Cell Biol 127: 1217–1232.
Schwarz T. (2005). Biological effects of UV radiation on keratinocytesand Langerhans cells. Exp Dermatol 14: 788–789.
Simons K, Ehehalt R. (2002). Cholesterol, lipid rafts, and disease.J Clin Invest 10: 597–603.
Simons K, Ikonen E. (1997). Functional rafts in cell membranes.Nature 387: 569–572.
Simons K, Toomre D. (2000). Lipid rafts and signal transduction.Nat Rev Mol Cell Biol 1: 31–39.
Simons K, van Meer G. (1988). Lipid sorting in epithelial cells.Biochemistry 27: 6197–6202.
Slotte JP, Bierman EL. (1988). Depletion of plasma-membranesphingomyelin rapidly alters the distribution of cholesterol betweenplasma membranes and intracellular cholesterol pools in culturedfibroblasts. Biochem J 250: 653–658.
Smith DW, Lemli L, Opitz JM. (1964). A newly recognized syndromeof multiple congenital anomalies. J Pediatr 64: 210–217.
Stade BG, Messer G, Riethmuller G, Johnson JP. (1990). Structuralcharacteristics of the 50 region of the human ICAM-1 gene.Immunobiology 182: 79–87.
Staunton DE, Marlin SD, Stratowa C, Dustin ML, Springer TA.(1988). Primary structure of ICAM-1 demonstrates interactionbetween members of the immunoglobulin and integrin supergenefamilies. Cell 52: 925–933.
Tint GS, Irons M, Elias ER, Batta AK, Frieden R, Chen ST et al.(1994). Defective cholesterol biosynthesis associated with theSmith–Lemli–Opitz syndrome. N Engl J Med 330: 107–113.
Tyrrell RM. (2004). Solar ultraviolet A radiation: an oxidizing skincarcinogen that activates heme oxygenase-1. Antioxid Redox Signal
6: 835–840.
Tyrrell RM, Reeve VE. (2006). Potential protection of skin by acuteUVA irradiation—from cellular to animal models. Prog Biophys
Mol Biol 92: 86–91.Vainio S, Heino S, Mansson J-E, Fredman P, Kuismanen E,
Vaarala O et al. (2002). Dynamic association of human insulinreceptor with lipid rafts in cells lacking caveolae. EMBO Rep 3:95–100.
Valencia A, Kochevar IE. (2006). Ultraviolet A induces apoptosisvia reactive oxygen species in a model for Smith–Lemli–Opitzsyndrome. Free Radic Biol Med 15: 641–650.
Vandekerckhove J, Weber K. (1978). Mammalian cytoplasmic actinsare the products of at least two genes and differ in primary structurein at least 25 identified positions from skeletal muscle actins. Proc
Natl Acad Sci USA 75: 1106–1110.Wolf C, Chevy F, Pham J, Kolf-Clauw M, Citadelle D, Mulliez Net al. (1996). Changes in serum sterols of rats treated with 7-dehydrocholesterol-delta 7-reductase inhibitors: comparison tolevels in humans with Smith–Lemli–Opitz syndrome. J Lipid Res
37: 1325–1333.XU F, Rychnovsky SD, Belani JD, Hobbs HH, Cohen JC, Rawson
RB. (2005). Dual roles for cholesterol in mammalian cells. Proc Natl
Acad Sci USA 102: 14551–14556.Yancey PG, Rodrigueza WV, Kilsdonk EPC, Stoudt GW, Johnson
WJ, Phillips C et al. (1996). Cellular cholesterol efflux mediated bycyclodextrins. Demonstration of kinetic pools and mechanism ofefflux. J Biol Chem 271: 16026–16034.
Yang N, Huang Y, Jiang J, Frank SJ. (2004). Caveolar and lipid raftlocalization of the growth hormone receptor and its signalingelements: impact on growth hormone signaling. J Biol Chem 279:20898–20905.
Yang T, Kozopas KM, Craig RW. (1995). The intracellular distri-bution and pattern of expression of Mcl-1 overlap with, but are notidentical to, those of Bcl-2. J Cell Biol 128: 1173–1184.
Zhang Y, Mattjus P, Schmid PC, Dong Z, Zhong S, Ma W-Y et al.(2001). Involvement of the acid sphingomyelinase pathway inUVA-induced apoptosis. J Biol Chem 276: 11775–11782.
Ceramide and raft signaling linkage in UVA radiation-induced gene expressionS Grether-Beck et al
4778
Oncogene
Copyright © 2022 FDOKUMEN