
Effect of ozone on bronchial mucosal inflammation in asthmatic and healthy subjects
Upload
independentCategory
view
0download
0
Int. J . Cancer: 30, 265-272 (1982)
CELLULAR INGESTION, TOXIC EFFECTS, AND LESIONS OBSERVED IN HUMAN BRONCHIAL EPITHELIAL TISSUE AND CELLS CULTURED WITH ASBESTOS AND GLASS FIBERS Aage HAUGEN194, Paul W. SCHAFER~, John F. LECHNERl, Gary D. STONER''5, Benjamin F. TRUMP3 and Curtis C. HARRIS Laboratory of Human Carcinogenesis, National Cancer Institute, Bethesda, Maryland, 20205; Department of
Surgery, Veterans Administration Medical Center, Washington, DC, 20422; Department of Pathology, Univer- sity of Maryland, Baltimore, Maryland, 21202, USA; Present address: Department of Toxicology, National Institute of Public Health, Oslo 1, Norway; Present address: Department of Pathology, Medical College of Ohio, C.S. 10008, Toledo, Ohio 43614, USA.
Although exposure to asbestos has been shown in previ- ous epidemiological studies to synergistically increase the risk of bronchogenic carcinoma for cigarette smokers, the pathobiological machanism(s) responsible for the cocar- cinogenic action of asbestos is not known. Therefore, the effects of asbestos fibers on normal human tracheobron- chial epithelium are of interest. Asbestos fibers, ie., amo- site, crocidolite and chrysotile (UICC samples) and glass fibers were initially assayed for their cytotoxicity. Bron- chial epithelial cells exposed to either asbestos or glass fibers displayed inhibition of cell growth as a function of fiber concentration. When compared to glass fibers, as- bestos fibers caused a statistically significant (w0.05) de- crease in cell population doubling rate. Chrysotile was approximately 10 times more cytotoxic than either amo- site or crocidolite and more than I00 times more cytotox- ic than the glass fibers. Epithelial calls were 10 to I 5 times more sensitive to the cytotoxic effects of asbestos fibers than bronchial fibroblasts from the same donor. Using electron microscopy, asbestos fibers were seen to be in- gested by epithelial cells. After exposure for 2 h short fibers (< I2 pn) were found both in the cytoplasm and within phagosomes. High-voltage electron microscopy combined with stereo-microscopy revealed asbestos fi- bers in the cytoplasmic matrix. A single exposure to amo- site asbestos (IWIOOO Wml) was found to induce focal hyperplasia and epidermoid metaplasia with cellular aty- pia in human tracheobronchial explants. Surface fine structure observations revealed asbestos fibers protrud- ing from these lesions. lntrasytoplasmic and intranuclear fibers of armsite were identified in these focal lesions us- ing X-ray microanalysis in combination with transmission electron microscopy. With increasing time, fibers were also seen deposited within the submucosa of the explants. The preferential cytotoxicity of asbestos for epithelial calk and the metaplastic response of bronchial epithelium may be important in the cacarcinogenic action of as- bestor.
Asbestos, a naturally occurring mineral fiber, has found widespread application. For example, more than 30 million tons of asbestos have been used in the United States since 1900. Lynch and Smith (1935) and Doll (1955) reported on correlations between occupa- tional exposure to asbestos and risk of bronchogenic carcinoma in humans. Asbestos fibers have been de- tected in the lungs, liver, kidney, spleen, lymph nodes, pancreas and gastrointestinal tract of exposed individuals (Evans et al., 1973; Langer et al., 1974). Because of their cytotoxic, fibrogenic and car- cinogenic properties, asbestos fibers are one of the most important environmental pollutants.
Exposure to large amounts of asbestos dust has been associated with the development of asbestosis, bronchogenic carcinomas, pleural and peritoneal mesotheliomas, and carcinomas of the gastrointestinal tract and larynx of humans (Selikoff et al., 1968;
Selikoff, 1977; Selikoff and Lee, 1978a; Harington, 1981). Asbestos is considered as a cocarcinogenic agent in the etiology of bronchogenic cancer. The combination of cigarette smoking and exposure to as- bestos increased the risk of bronchial cancer in such a way that 90% of the asbestos-related cancers are a result of its combined effect with cigarette smoke (Selikoff et al., 1968).
In experimental animals, a positive correlation be- tween exposure to various asbestos fibers and lung cancer has been demonstrated (Wagner et al., 1974). Chrysotile fibers were both cocarinogenic and weakly carcinogenic with a rat tracheal heterograft model (Topping and Nettesheim, 1980; Topping et al., 1980).
In vitro studies of fiber toxicity using cultures of tissues and cells have been reported by several laboratories. Whereas there have been only a few re- ports on the interaction of asbestos fibers with ex- planted tissue from experimental animals (Mossman and Craighead, 1981), studies with macrophages (Alli- son, 1971; Miller and Harington, 1972), fibroblastic and epithelioid cell lines (Harington et al., 1975; Richards and Jacoby, 1976; Neugut et al., 1978; Reiss et al., 1980) have revealed that chrysotile is more toxic than amosite and crocidolite.
Primary cultures of normal epithelial tissue and cells are not transformed and thus provide certain ad- vantages over studies using cell lines. Effects of asbes- tos fibers on human bronchial epithelial cells and tis- sue have not been previously examined due to the lack of suitable culture systems. Recently, methods to ob- tain replicative adult human bronchial epithelial cell cultures have been developed (Lechner et al., 1981). We have used these cells to examine the cytotoxic effects of asbestos fibers at the cellular level. In addi- tion, morphological lesions in cultured human re- spiratory musoca caused by asbestos fibers are de- scribed.
Fibers The UICC standard reference samples, provided by
Dr. V. Timbell, Medical Research Council, England, were Canadian chrysotile B, crocidolite and amosite.
MATERIAL AND METHODS
Abbreviations: 6EGF, epidermal growth factor; FBS, fetal bovine serum; HVEM, high-voltage electron microscopy; SEM, scanning electron microscopy; TEM, transmission electron microscopy; PBS, phosphate-buffered saline; STEM, scanning transmission electron microscopy.
Received: February 22,1982, and in revised form June 30, 1982.

266 HAUGEN ET AL
Homogenized GFID Whatman filters washed in 1 N HCl and water were used as glass fibers. Aliquots of fibers were weighed immediately prior to use and sterilized by autoclaving at 125°C for 20 min. Explant culture
Human bronchial explants were prepared from “im- mediate autopsy” and cultured in chemically defined medium by methods described previously (Harris et al., 1978). Beginning on day 7 of culture, 0.5-cm2 ex- plants were placed on the bottom of 60-mm bac- teriological dishes in modified M199 medium [0.6 mM Ca2+, 20 m~ HEPES, 20 pglml EGF6 (Collaborative Research, Inc., Waltham, MA), 5 X l O ” M hydrocor- tisone (Steraloids, Inc., Wilton, NH), 0.2 pglml insu- lin (Eli Lilly and Co.), trace elements, 50 ,@ml gen- tamicin, and 1.25% Sephadex G-10 filtered FBS] (Lechner et al., 1980). M199 medium was prepared by the NIH Media Unit. Tissue explants were exposed to amosite asbestos by pipetting 200 pl of a fiber suspen- sion (10, 100, or 1,000 pglml) onto the epithelial sur- face.
The culture medium was replaced with medium without fibers after 24 h incubation and subsequently at 4-day intervals for the duration of the experiment. After periods of 2 ,4 and 8 weeks, four explants from each group were processed for histology and electron microscopy. Cell culture
Monolayers of human bronchial epithelial cells were established using methods previously described (Lechner et al., 1981). To obtain cell outgrowths of fibroblast, the explants were cultured in CMRL 1066 medium supplemented with 10 % FBS. Two weeks af- ter explantation, asbestos fibers (100 ,@ml) were ad- ded to both epithelial cells and fibroblast cell out- growths and to secondary cultures of each cell type. The cells were fixed 1, 6 , 24, 48, 72 and 158 h after asbestos exposure and processed for SEM, TEM and HVEM examination. Cytotoxicity assay
Clonal growth rate was used to measure cytotoxici- ty. Sixty-mm tissue culture dishes were precoated with 50 pg fibronectin (Collaborative Research, Inc., Waltham, MA). Each dish was inoculated with either 10,000 human bronchial epithelial cells and 3 X lo5 growth-arrested Swiss mouse 3T3 feeder-cells or 1,000 human bronchial fibroblast cells. Twenty-four h after the cells were seeded, the medium was replaced by medium containing various concentrations of fibers. Each dose of fiber was assayed in triplicate. Following
TABLE I - ASBESTOS TOXICITY IN CULTURES OF HUMAN BRONCHIAL EPITHELIAL AND FIBROBLASTIC CELLS’
50% toxicity (&d)
Epithelial cells Fibroblastic cells Fiber
Chrysotile 550.2 95 f3 Amosite 1521 22555.8 Crocidolite 30f0.7 250+11 Glassfiber 10052.3 None * ‘The cytotodcity assay was done as described under “Material and Me-
thods”. - significant inhibition of clonal growth rate at 1W &d.
L AMOSITE - CROCIDOLITE
0 0.3 1 3 10 100 FIBERS, pg/ml
FIGURE 1 - Clonal growth rate of human bronchial epithe- lial cells as a function of fiber concentration. Growth rate based upon number of cells per 18 randomly selected colo- nies for each datum point.
3 days’ exposure, the cultures were washed twice and reincubated in fiber-free medium. Ten days post incu- bation, the cells were fixed in 10% formalin and the epithelial cell colonies were selectively stained with Rhodanile blue (Rheinwald, 1980). Clonal growth rates were expressed as the average number of popu- lation doublings (log, number of cellskolony) per 18 randomly selected colonies divided by days of incuba- tion (Lechner and Kaighn, 1979). Examination of tissue and cells by HVEM, SEM, TEM, STEM, and X-ray microanalysis
Cells incubated on glass coverslips and tissues were fixed for 2 h at room temperature in PBS (PH 7.2) containing 4 % paraformaldehyde and 1 % glutaral- dehyde (4FlG) (McDowell and Trump, 1976). After postfixation for 30 min in 1 % Os04, they were serially dehydrated in ethanol and embedded in Epon. Ultra- thin sections of both explants and cell pellets were cut on an LK3 8800 Ultratome and then double-stained with uranyl acetate and lead citrate. In order to meet requirements for X-ray microqnalysis, the section thickness ranged from 500-1000 A. Ultrastructural ex- amination and X-ray analysis were made using using a
FIGURE 2 - SEM of epithelial cell in a monolayer culture showing the initial phase of phagocytosis of an amosite asbes- tos fiber (100 pg/ml) 1 h after exposure. X22,OOO.
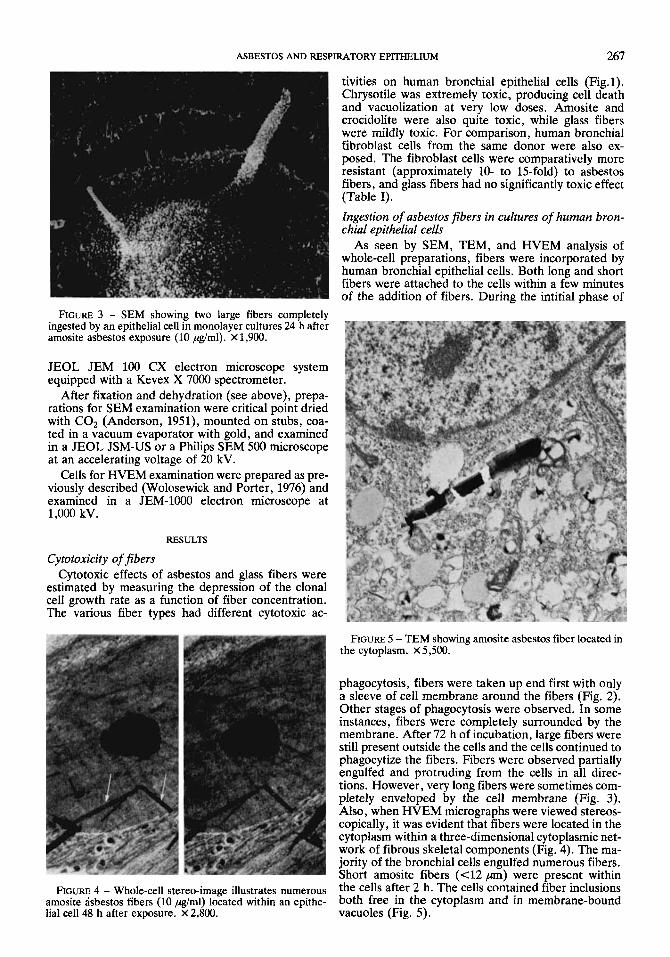
ASBESTOS AND RESPIRATORY EPITHELIUM 267
tivities on human bronchial epithelial cells (Fig. 1). Chrysotile was extremely toxic, producing cell death and vacuolization at very low doses. Amosite and crocidolite were also quite toxic, while glass fibers were mildly toxic. For comparison, human bronchial fibroblast cells from the same donor were also ex- posed. The fibroblast cells were comparatively more resistant (approximately 10- to 15-fold) to asbestos fibers, and glass fibers had no significantly toxic effect (Table I). Ingestion of asbestos fibers in cultures of human bron- chial epithelial cells
As seen by SEM, TEM, and HVEM analysis of whole-cell preparations, fibers were incorporated by human bronchial epithelial cells. Both long and short fibers were attached to the cells within a few minutes of the addition of fibers. During the intitial phase of
FIGURE 3 - SEM showing two large fibers completely ingested by an epithelial cell in monolayer cultures 24 h after amosite asbestos exposure (10 pg/ml). X 1,900.
JEOL JEM 100 CX electron microscope system equipped with a Kevex X 7000 spectrometer.
After fixation and dehydration (see above), prepa- rations for SEM examination were critical point dried with COz (Anderson, 1951), mounted on stubs, coa- ted in a vacuum evaporator with gold, and examined in a JEOL JSM-US or a Philips SEM 500 microscope at an accelerating voltage of 20 kV.
Cells for HVEM examination were prepared as pre- viously described (Wolosewick and Porter, 1976) and examined in a JEM-1000 electron microscope at 1,000 kV.
RESULTS
Cytotoxicity of fibers Cytotoxic effects of asbestos and glass fibers were
estimated by measuring the depression of the clonal cell growth rate as a function of fiber concentration. The various fiber types had different cytotoxic ac-
FIGURE 5 - TEM showing amosite asbestos fiber located in the cytoplasm. X 5,500.
phagocytosis, fibers were taken up end first with only a sleeve of cell membrane around the fibers (Fig. 2). Other stages of phagocytosis were observed. In some instances, fibers were completely surrounded by the membrane. After 72 h of incubation, large fibers were still present outside the cells and the cells continued to phagocytize the fibers. Fibers were observed partially engulfed and protruding from the cells in all direc- tions. However, very long fibers were sometimes com- pletely enveloped by the cell membrane (Fig. 3). Also, when HVEM micrographs were viewed stereos- copically, it was evident that fibers were located in the cytoplasm within a three-dimensional cytoplasmic net- work of fibrous skeletal components (Fig. 4). The ma- jority of the bronchial cells engulfed numerous fibers. Short amosite fibers ( e l2 pn) were present within
nGURE 4 - Whole-cell stereo-image illustrates numerous the cells after 2 h. The cells contained fiber inclusions amosite asbestos fibers (10 pg/ml) located within an epithe- both free in the cytoplasm and in membrane-bound lial cell 48 h after exposure. X 2,800. vacuoles (Fig. 5).
268 HAUGEN ET AL
Human bronchial explant cultures exposed to asbestos The response of tracheobronchial epithelium to the
presence of fibers was investigated using explants of cultured human bronchus. Amosite asbestos fibers were observed protruding from the explant shortly af- ter exposure to the fibers and frequently in areas of ciliated cells that contained few cilia. In areas contain- ing highly ciliated cells, most of the fibers were re- moved by ciliary action. However, fibers were occa- sionally observed on the ciliated epithelium 8 weeks after exposure to amosite.
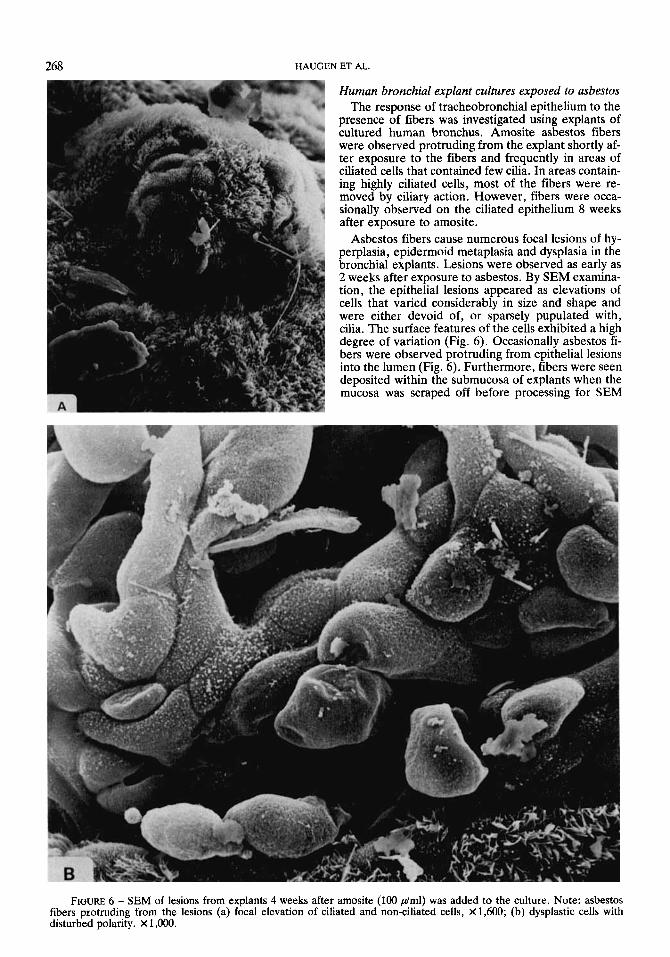
Asbestos fibers cause numerous focal lesions of hy- perplasia, epidermoid metaplasia and dysplasia in the bronchial explants. Lesions were observed as early as 2 weeks after exposure to asbestos. By SEM examina- tion, the epithelial lesions appeared as elevations of cells that varied considerably in sue and shape and were either devoid of, or sparsely pupulated with, cilia. The surface features of the cells exhibited a high degree of variation (Fig. 6). Occasionally asbestos fi- bers were observed protruding from epithelial lesions into the lumen (Fig. 6). Furthermore, fibers were seen deposited within the submucosa of explants when the mucosa was scraped off before processing for SEM
FIGURE 6 - SEM of lesions from explants 4 weeks after amosite (100 phnl) was added to the culture. Note: asbestos fibers protruding from the lesions (a) focal elevation of ciliated and non-ciliated cells, X1,600; (b) dysplastic cells with disturbed polarity. X 1,000.

ASBESTOS AND RESPIRATORY EPlTHELIUM 269
FIGURE 7 - Stratified cuboidal epithelium with loss of cilia- ted cells 4 weeks after exposure to amosite (100 d m l ) . x1,900.
(data not shown). Stratified cuboidal epithelium (Fig. 7) and metaplastic areas were seen (Fig. 8). The area surrounding the lesions was covered by Closely Packed FIGURE 9 - Absence of normal respiratory epithelium 2 cilia. CYtoPathic alterations of the bronchial weeks after amosite (10 d m l ) exposure. Variation in size of epithelium were manifested by irregular arrangements cells, cellular atypia, and loss of normal polarity were ob- of cells, polymorphism, variation in nuclear size, and served. X660. nuclear hyperchromasia (Fig. 9). In individual areas, normal cellular differentiation persisted. The mor- phology of the control tissues was essentially normal and only rare areas of metaplasia were seen (Fig.10).
RGURE 8 - Area of flattened metaplastic cells 4 weeks FIGURE 10 - Control culture of normal bronchial tissue 2 after exposure to amosite (10 pglml). X1,900. weeks after sham exposure. X 220.
270 HAUGEN ET AL.
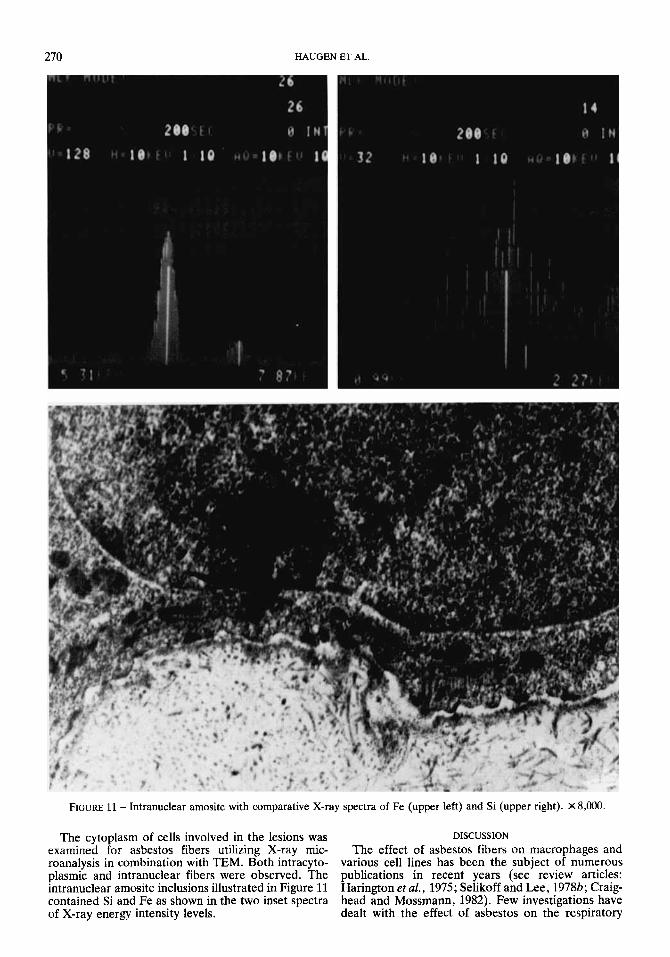
FIGURE 11 - Intranuclear amosite with comparative X-ray spectra of Fe (upper left) and Si (upper right). X8,ooO.
The cytoplasm of cells involved in the lesions was DISCUSSION examined for asbestos fibers utilizing X-ray mic- The effect of asbestos fibers on macrophages and roanalysis in combination with TEM. Both intracyto- various cell lines has been the subject of numerous plasmic and intranuclear fibers were observed. The publications in recent years (see review articles: intranuclear amosite inclusions illustrated in Figure 11 Harington et al., 1975; Selikoff and Lee, 19786; Craig- contained Si and Fe as shown in the two inset spectra head and Mossmann, 1982). Few investigations have of X-ray energy intensity levels. dealt with the effect of asbestos on the respiratory

epithelium of experimental animals (Mossman et al., 1977; Topping and Nettesheim, 1980; Topping et al., 1980). We have extended these studies to human re- spiratory epithelium maintained as explant and cell culture.
Four types of fibers (amosite,crocidolite, chrysotile and glass fibers) were analyzed for their cytotoxic ef- fects on cultured bronchial epithelial and fibroblast cells. Our findings, indicating chrysotile to be the most cytotoxic form of fiber dust tested, correspond to data previously obtained with various cell lines and mac- rophages (Harington, 1975; Wade et al., 1976; Neugut et al., 1978; Reiss et al., 1980; DiPaolo et al., 1981). Previous studies have shown a parallelism between cytotoxicity and hemolytic potency (Harington et al., 1975), probably due to damaging interactions with the plasma membrane. The marked hemolytic activity of chrysotile and the low activity of amosite and crocido- lite indicate that magnesium ion content might be an important factor in the hemolytic action of asbestos (Morgan et al., 1977; Reiss et al., 1980).
Although asbestos fibers produce profound effects in plasma membranes, the reaction with one cell type may not be identical to its reaction with another cell type. All three types of asbestos and glass fibers were more toxic to bronchial epithelial cells than bronchial fibroblastic cells from the same donor. A possible ex- planation for this difference in toxicity could be that the cell surface of bronchial epithelial cells permits a more efficient uptake of fibers than does the surface of fibroblasts.
There is no direct correlation between in vitro cy- totoxicity and carcinogenesis since all major types of asbestos are able to cause respiratory cancer (Rom and Palmer, 1974; Harington et al., 1975). Asbestos fibers might, however, induce changes in the plasma membrane similar to those evidenced in tumorigenic cells. High-molecular-weight membrane proteins were significantly reduced by asbestos treatment (Newman et al., 1980). Loss of high-molecular-weight glycopro- teins and glycolipid simplification have been corre- lated with cell transformation in vitro by chemical car- cinogens (Nicolson, 1976). The interaction between the plasma membrane and the fiber might also lead to increased permeability of carcinogens through the membrane.
Glass fibers were only midly toxic to bronchial cells. Recent findings have shown glass fibers to be car- cinogenic when implanted into pleura of rats (Stanton et al., 1977, 1981). Although there is no evidence as yet that glass fibers cause tumors in humans (Enterline and Henderson, 1975; Bayliss et al., 1976), a definite answer to this question will have to await further epidemiological studies since production of long fibers
with a diameter of 1 p or less started only a decade ago.
Electron microscopy and X-ray microanalysis re- vealed that human bronchial cells are able to phagocy- tize asbestos fibers. Treated cultures contained numerous fibers which were randomly orientated throughout the cell. There are differences between macrophages and epithelial cells in the recognition of asbestos fibers. Epithelial cells show no increased sur- face activity, and the fibers are generally phagocytized along the short axis. In contrast, the surface activity of macrophages was increased by exposure to asbestos fibers, and the fibers were held in place by cytoplasmic processes and phagocytized (Mckmore et al., 1980). Short fibers located in the cytoplasm may be less cy- totoxic to cells than partially ingested long fibers since these fibers interact with the plasma membrane over a prolonged period of time.
These experiments demonstrate that asbestos fibers are capable of stimulating abnormal growth of human bronchial epithelial cells as manifested by the appear- ance of discrete morphologic lesions in cultured ex- plants. Asbestos fibers produced histological lesions commonly observed in premalignant human bron- chus, such as hyperplasia and epidermoid metaplasia. Similar results were obtained by adding crocidolite as- bestos to cultured explants from hamster trachea (Mossman et al., 1977). Although the relevance of these morphological changes in the carcinogenic pro- cess has not been established, studies suggest that at least some of the epithelial cells emerging from the lesions after a fairly brief carcinogen exposure appear to have acquired a rather strikingly different growth potential (Topping et al., 1979a, b). Further, his- topathological studies on bronchial lesions in smokers indicate that the incidence of hyperplasia, metaplasia, atypia, carcinoma in situ and invasive carcinoma is related to cigarette consumption and age (Kundtson, 1960; Auerbach et al., 1961).
ACKNOWLEDGEMENTS
The authors wish to thank Drs. K.R. Porter and M. Fotina at the Department of Molecular, Cellular, and Developmental Biology, University of Colorado, for placing the HVEM facilities at our disposal, and Dr. H. Dalen for the use of his SEM facilities. This work was supported in part by (a) the Defense Civil Pre- paredness Agency, now a part of the Federal Emergency Management Agency, Intra-agency Ag- reement DCPA-01-77-C-0248, and (b) contract CP1- 5738. The technical assistance of Mrs. LA. McClen- don, Mrs. G.A. Myers, Mrs. M. Gladden, Mr. P. Meola and Mr. F. Jackson, and the secretarial assis- tance of Mrs. N. Paige were greatly appreciated.
ASBESTOS AND RESPIRATORY EPITHELIUM 27 1
REFERENCES
ALLISON, A.C., Effects of silica and asbestos on cells in culture. In: W.H. Walton (ed.), Inhaled particles, Vol. III, pp. 437-445, Pergamon, Oxford (1971).
ANDERSON, T.F., Techniques for preservation of 3-dimensional structure in preparing specimens for electron microscopy. Trans. N.Y. Acad. Sci., U, 130-133 (1951).
AUEREIACH, O., STOUT, A.P., and H ~ O N D , E.C., Changes in bronchial epithelium in relation to cigarette smoking and in rela- tion to lung cancer. N . Engl. J . Med., 265, 253-267 (1961).
BAYLIQS, D.L., DEMENT, J.M., WAGONER, J.K., and BLEJER, H.P., Mortality pattern among fibrous glass production workers. Ann. N . Y. Acad. Sci., 271, 324-335 (1976). CRAIGHEAD, J.E., and MOSSMAN, B.T., The pathogenesis of asbestos-associated diseases. N . Engl. J . Med., 306, 1446-1455 (1982). DIPAOLO, J.A., DEMARINIS, A.J., and DONIGER, J . , Asbestos and benzo(a)pyrene synergism in the transformation of Syrian hamster embryo cells. J. environ. Path. Toxicol. 5 , 535-543 (1 982).

272 HAUGEN ET AL.
DOLL, R., Mortality from lung cancer in asbestos workers. Brit. J . ind. Med., 12, 81-86 (1955). ENTERLINE, P.E., and HENDERSON, V., The health of retired fibrous glass workers. Arch. environ. Hlth, 30, 113-116 61975). EVANS, J.E., EVANS, R.J., HOLMES, A., HOUNAM, R.T., JONES, D.M., MORGAN, A., and WALSH, M., Studies on the deposition of inhaled fibrous material in the respiratory tract of the rat and its subsequent clearance using radioactive tracer techniques. Environ. Res., 6,180-201 (1973). HARINGTON, J.S., ALLISON, A.C., and BADAMI, D.V., Mineral fibers: chemical, physicochemical and biological properties. Advanc. Pharmacoi. Chemother., 12,291-402 (1975). HARINGTON, J.S., Fiber carcinogenesis: epidemiologic observa- tions and the Stanton hypothesis. J . nat. Cancer Znst., 67,977-990 (1981). HARRIS, C.C., AUTRUP, H., STONER, G.D., and TRUMP, B.F., Carcinogenesis studies in human respiratory epithelium; an ex- perimental model system. In: C.C. Harris (ed.), Pathogenesis and therapy of lung cancer, pp. 559-597, Marcel Dekker, New York (1978). KUNDTSON, K.P., The pathologic effects of smoking tobacco on the trachea and bronchial mucosa. Amer. J. clin. Path., 33, 310- 317 (1960). LANCER, A.M., MACKLER, A.D., and POOLEY, F.D., Electron microscopical investigation of asbestos fibers. Environ. Hlth Perspect., 9, 63-80 (1974). LECHNER, J.F., BABCOCK, M.S., MARNELL, M., NARAYAN, K.S., and KAIGHN, M.E., Normal human prostatic epithelial cell cul- tures. In: C.C. Harris, B.F. Trump and G.D. Stoner (ed.), Methods in cell biology, Vol. 21B, pp. 195-225, Academic Press, New York (1980). LECHNER, J.F., HAUGEN, A., AUTRUP, H., MCCLENDON, LA., TRUMP, B.F., and HARRIS, C.C., Clonal growth of epithelial cells from normal adult human bronchus. Cancer Res., 41, 2294-2304 (1981). LECHNER, J.F., and KAIGHN, M.E., Applications of the princi- ples of enzyme kinetics to clonal growth rate assays: an approach for delineating interactions among growth-promoting agents. J , Cell Physiol., 100, 519-530 (1979. LYNCH, K.M., and SMITH, W.A., Pulmonary asbestosis. 111. Car- cinoma of lung in asbesto-silicosis. Amer. J . Cancer, 24, 56-64 (1935). MCLEMORE, T.L., MACE, M.L., ROGGLI, V., MARSHALL, M.V., LAWRENCE, E.C., WILSON, R.K., MARTIN, R.R., BRINKLEY, B.B., and GREENBERG, S.D., Asbestos body phagocytosis by human free alveolar macrophages. Cancer Lett., 9,8593 (1980). MCDOWELL, E.M., and TRUMP, B.F., Histological fixative suit- able for diagnostic light and electron microscopy. Arch. Path. Lab. Med., 100, 405-414 (1976). MILLER, K., and HARINGTON, J.S., Some biochemical effects of asbestos on macrophages. Brit. J . exp. Path., 53,397-405 (1972). MORGAN, A,, DAVIES, P., WAGNER, J.C., BERRY, G., and HOLMES, A., The biological effects of magnesium-leached chry- sotile asbestos. Brit. J. exp. Path., 58, 465-473 (1977). MOSSMAN, B.T., and CRAIGHEAD, J.E., Mechanisms of asbestos carcinogensis. Environ. Res., 25, 269-280 (1981). MOSSMAN, B.T., KESSLER, J.B., LEY, B.W., and CRAIGHEAD, J.E., Interaction of crocidolite asbestos with hamster respiratory mucosa in organ culture. Lab. Invest., 36, 131-139 (1977). NEUGUT, A.I., EISENFJERG, D., SILVERSTEIN, M., PULKRABEK, P., and WEMSTEIN, I.B., Effects of asbestos on epithelioid lines. Environ. Res., 17, 256-265 (1978). NEWMAN, H.A., SAIT, Y.A., and HART, R.W., Putative inhibi- tory effects of chrysotile, crocidolite, and amosite mineral fibers
on the more complex surface membrane glycolipids and glyco- proteins. Environ. Hlth. Perspect., 34, 103-111 (1980). NICOLSON, G.L., Trans-membrane control of the receptors on normal and tumor cells: 11. Surface changes associated with transformation and malignancy., Biochem. Biophys. Acta., 458,
REISS, B., SOLOMON, S. WEISBURGER, J.H., and WILLIAMS, G.M., Comparative toxicities of different forms of asbestos in a cell culture assay. Environ. Res., 22, 109-129 (1980). RHEINWALD, J.G., Serial cultivation of normal human epidermal keratinocytes. In: C.C. Hams, B.F. Trump, and G.D. Stoner (ed.), Methods in cell biology, Vol. 21A, pp. 229-254, Academic Press, New York (1980). RICHARDS, R.J., and JACOBY, F., Light microscope studies on the effects of chrysotile asbestos and fiber glass on the morpholo- gy and reticulin formation of cultured lung fibroblasts. Environ. Res., 11, 112-121 (1976). ROM, W.N., and PALMER, P.E.S., The spectrum of asbestos- related disease. West. J. Med., l21, 10-21 (1974). SELIKOFF, I .J . , Cancer risk of asbestos exposure. In: H.H. Hiatt, J.D. Watson, and J.A. Winstein (ed.), Origins of human cancer, pp. 1765-1785, Cold Spring Harbor, New York (1977). SELIKOFF, I.J., HAMMOND, E.C., and CHURG, J., Asbestos expo- sure, smoking, and neoplasia. J. Amer. med. Ass., 2Q4, 106-112 (1968). SELIKOFF, I.J., and LEE, D.H.K., Characteristics, etiology, and prevalence of asbestos-associated carcinoma. In: 1. J. Selikoff and D.H.K. Lee (ed.), Asbestos and disease, pp. 307-336, Academic Press, New York (1978a). SELIKOFF, I.J., and LEE, D.H.K., General tissue reaction and cytotoxicity. In: I.J. Selikoff and D.H.K. Lee (ed.), Asbestos and disease, pp. 377-392, Academic Press, New York (1978b). STANTON, M.F., LAYARD, M., TEGERIS, A., MILLER, E., MAY, M., and KENT, E., Carcinogenicity of fibrous glass: pleural re- sponse in the rat in relation to fiber dimension. J. nat. Cancer
STANTON, M.F., LAYARD, M., TEGERIS, A,, MILLER, E., MAY, M., MORGAN, E., and SMITH, A, , Relation of particle dimension to carcinogenicity in amphibole asbestoses and other fibrous min- erals. 1. nut. Cancer Inst., 67, 965-976 (1981). TOPPING, D.C., GIESEMER, R.A., and NETIZSHEIM, P., Quantita- tive assessment of generalized epithelial changes in trachea mu- cosa following exposure to 7,12-dimethylbenz(a)anthracene. Cancer Res., 39, 4823-4828 (1979a). TOPPING, D.C., GIESEMER, R. A., and NETTESHEIM, P., Develop- ment and fate of local epithelial lesions in tracheal mucosa fol- lowing exposure to 7,12-dimethylbenz(a)antracene. Cancer Res.,
TOPPING. D.C.. and NEITESHEIM. P.. Two-staee carcinoeenesis
1-72 (1972).
Inst., 58, 587-602 (1977).
39,4829-4837 (19796).
studies with asbestos in Fisher 344 rats. J. nat.-Cancer Z&., 65, 627-630 (1980). TOPPING, D.C., NETTESHEIM, P., and MARTIN, D.H., Toxic and tumorigenic effects of asbestos in tracheal mucosa. Environ. Path. Toxicol., 3, 261-275 (1980). WADE, M.J., LIPKIN, L.E., and FRANK, A.L., Studies of in vitro asbestos-cell interaction. J . environ. Path. Toxicol., 2,1029-1039 (1976). WAGNER, J.C., BERRY, G., SKILDMORE, J.W., and TIMEIRELL, V., The effects of the inhalation of asbestos in rats. Brit. J. Cancer, 29,252-269 (1974). WOLOSEWICK, J.J., and PORTER, K.R., Stereo high-voltage elec- tron microscopy of whole cells of the human diploid line, WI-38. Amer. J . Anat., 147, 303-324 (1976).
Copyright © 2022 FDOKUMEN




















