
cDNA cloning and expression of the phytoene synthase gene in sunflower
6
Journal of Plant Physiology 162 (2005) 479—484 SHORT COMMUNICATION cDNA cloning and expression of the phytoene synthase gene in sunflower Mariangela Salvini a,b , Andrea Bernini b , Marco Fambrini b , Claudio Pugliesi b, a Scuola Normale Superiore, I-56124 Pisa, Italy b Dipartimento di Biologia delle Piante Agrarie, Sezione di Genetica, Universita` di Pisa, I-56124 Pisa, Italy Received 8 December 2003; accepted 25 April 2004 Summary A PCR approach was used to isolate a phytoene synthase (Psy) cDNA from sunflower (Helianthus annuus L.). A reconstructed full-length sequence (1598 bp) of the Psy cDNA was obtained; it contained a 1242 bp CDS, 172-nucleotides of 5 0 -untranslated region (UTR), and 170-nucleotides of 3 0 -UTR. The predicted protein (46.8 kDa) displayed a sequence of 414 amino acid residues with a putative transit sequence for plastid targeting in the N-terminal region. Phylogenetic analysis demonstrated that the sunflower Psy (HaPsy) clustered with the marigold (Tagetes erecta) Psy gene, with which it showed an overall amino acid identity of 93.7%. Moreover, the HaPsy sequence relates closely with other Psy sequences of higher plants. HaPsy was highly expressed in cotyledons, and young and mature leaves. In contrast, HaPsy transcript levels were comparatively lower in the stem and almost absent in the roots. The HaPsy transcript levels were influenced by leaf expansion, which suggested that the expression of the HaPsy gene is regulated during the process of leaf development. The role of HaPsy in controlling carotenoid biosynthesis is demonstrated by the concurrent increase of HaPsy transcript levels with the light-dependent enhanced carotenoid production in green tissues of sunflower. & 2004 Elsevier GmbH. All rights reserved. Introduction Carotenoid biosynthesis takes place in the plastid, but all known enzymes in the pathway are nuclear- encoded and post-translationally imported into the organelle (reviewed in Sandmann, 2001). In plas- tids, carotenoid biosynthesis depends on the 2-C- methyl-D-erythritol 4-phosphate (MEP) pathway (Lichtenthaler et al., 1997; Lichtenthaler, 1999; Rodrı ´quez-Concepcio´nandBoronat, 2002). The first ARTICLE IN PRESS www.elsevier.de/jplph KEYWORDS Carotenoid biosynthesis; Helianthus annuus; Phytoene synthase; Relative RT-PCR 0176-1617/$ - see front matter & 2004 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2004.04.011 Abbreviations: PPFD, photosynthetically active photon flux density; PSY, phytoene synthase; Psy , phytoene synthase gene; RACE, rapid amplification of cDNA ends; RT-PCR, reverse transcriptase polymerase chain reaction Corresponding author. E-mail address: [email protected] (C. Pugliesi).
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of cDNA cloning and expression of the phytoene synthase gene in sunflower

ARTICLE IN PRESS
Journal of Plant Physiology 162 (2005) 479—484
KEYWORDCarotenoidbiosynthesHelianthusPhytoeneRelative R
0176-1617/$ - sdoi:10.1016/j.
Abbreviationrapid amplifica�CorrespondE-mail addr
www.elsevier.de/jplph
SHORT COMMUNICATION
cDNA cloning and expression of the phytoenesynthase gene in sunflower
Mariangela Salvinia,b, Andrea Berninib, Marco Fambrinib,Claudio Pugliesib,�
aScuola Normale Superiore, I-56124 Pisa, ItalybDipartimento di Biologia delle Piante Agrarie, Sezione di Genetica, Universita di Pisa, I-56124 Pisa, Italy
Received 8 December 2003; accepted 25 April 2004
S
is;annuus;
synthase;T-PCR
ee front matter & 200jplph.2004.04.011
s: PPFD, photosynthetion of cDNA ends; RTing author.ess: [email protected]
SummaryA PCR approach was used to isolate a phytoene synthase (Psy) cDNA from sunflower(Helianthus annuus L.). A reconstructed full-length sequence (1598 bp) of the PsycDNA was obtained; it contained a 1242 bp CDS, 172-nucleotides of 50-untranslatedregion (UTR), and 170-nucleotides of 30-UTR. The predicted protein (46.8 kDa)displayed a sequence of 414 amino acid residues with a putative transit sequence forplastid targeting in the N-terminal region. Phylogenetic analysis demonstrated thatthe sunflower Psy (HaPsy) clustered with the marigold (Tagetes erecta) Psy gene, withwhich it showed an overall amino acid identity of 93.7%. Moreover, the HaPsysequence relates closely with other Psy sequences of higher plants. HaPsy was highlyexpressed in cotyledons, and young and mature leaves. In contrast, HaPsy transcriptlevels were comparatively lower in the stem and almost absent in the roots. The HaPsytranscript levels were influenced by leaf expansion, which suggested that theexpression of the HaPsy gene is regulated during the process of leaf development. Therole of HaPsy in controlling carotenoid biosynthesis is demonstrated by the concurrentincrease of HaPsy transcript levels with the light-dependent enhanced carotenoidproduction in green tissues of sunflower.& 2004 Elsevier GmbH. All rights reserved.
Introduction
Carotenoid biosynthesis takes place in the plastid,but all known enzymes in the pathway are nuclear-encoded and post-translationally imported into the
4 Elsevier GmbH. All rights rese
tically active photon flux densit-PCR, reverse transcriptase poly
i.it (C. Pugliesi).
organelle (reviewed in Sandmann, 2001). In plas-tids, carotenoid biosynthesis depends on the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway(Lichtenthaler et al., 1997; Lichtenthaler, 1999;Rodrıquez-Concepcion and Boronat, 2002). The first
rved.
y; PSY, phytoene synthase; Psy, phytoene synthase gene; RACE,merase chain reaction

ARTICLE IN PRESS
M. Salvini et al.480
committed step of carotenoid synthesis, the head-to-head condensation of geranylgeranyl dipho-sphate (GGPP) molecules to produce phytoene(colorless), is mediated by the enzyme phytoenesynthase (PSY). Membrane-localized enzymes carryout subsequent steps of the pathway leading to thecolored carotenoids. Phytoene undergoes fourdesaturation reactions with the production oflycopene (red). In this part of the pathwayphytofluene, z-carotene and neurosporene are theintermediates. Lycopene is converted to cycliccarotenoids b-carotene and a-carotene by cyclases.Subsequently, substitutions by hydroxyl, oxo, and/or epoxy groups produce xanthophylls with brightorange/yellow colors.
Genes encoding many of the carotenogenicenzymes have been isolated in bacteria, algae,fungi, and higher plants (reviewed in Sandmann,2001, 2002; Giuliano et al., 2002), but theregulatory mechanisms that control carotenoidbiosynthesis are poorly understood. In this study,we report the isolation of a cDNA corresponding tothe phytoene synthase gene of sunflower and theanalysis of its expression in relation to differentorgans and light-dependent pigment accumulationin chloroplasts.
Materials and methods
Sunflower (Helianthus annuus L.) seeds of theinbred line Nd-1 were germinated at 2371 1C inthe dark in Petri dishes and transferred, after 3 d,to 8 cm diameter pots containing a mixture of soiland sand. Seedlings were grown in a growthchamber at 2371 1C in the dark or under a 16 hphotoperiod. Several photosynthetically activephoton flux densities (PPFDs) were used: 5, 100,and 1500 mmolm�2 s�1. Irradiances were providedby a mercury vapour lamp (Osram HQI-TS 250 W/NDN, Wembley, England).
Cotyledon samples of 50–100mg (fresh weight)were collected and chloroplast pigments wereextracted as previously described (Fambrini etal., 1993). Each value was the mean of threeindependent experiments (five cotyledons for eachexperiment). The data were treated using analysisof variance procedures and means were separatedby Tukey’s test (P ¼ 0:05).
For cloning cDNA of the sunflower phytoenesynthase gene (Psy), amino acid sequences forpreviously cloned Psy genes were aligned. Regionswith the highest conservation were identified andtwo oligonucleotides primers were designed: P-A,50-TTCCKGGGASTTTGRGYTTGTTG-30 (forward) and
P-B, 50-GCCMAYACMGGCCATCTRCTAGC-30 (reverse).Total RNA was extracted from sunflower cotyledonsof 10-day-old seedlings grown under100 mmolm�2 s�1 (Chomczynski and Sacchi, 1987).First strand cDNA was made with 5 mg of total RNAusing the Superscript preamplification kit (Invitro-gen S.R.L, Life Technologies, Milan, Italy) followingconditions recommended by the manufacturer. ThecDNA was amplified with P-A and P-B primers. Afteran initial 94 1C for 4min denaturation step, 30cycles were run each with 30 s of denaturation at94 1C, followed by 30 s annealing at 55 1C, and 45 sextension at 72 1C. The PCR reaction yielded aproduct of about 750 bp, which was cloned into thepCRII vector (Invitrogen) and sequenced by thedideoxy method with an automated sequencer(clone Ha1Psy).
The rapid amplification of cDNA end (RACE)approach was used to isolate the 50 and 30 ends ofthe Psy cDNA. All reactions were performed withkits according to the manufacturer’s instructions(Invitrogen). First-strand synthesis was performedwith Superscript II retrotranscriptase. SpecificcDNAs from 30 and 50 RACE were subcloned intothe TOPO pCR 2.1 vector (Invitrogen) and se-quenced. The overlapping region with the cloneHa1Psy was confirmed. A putative full-length PsycDNA clone was obtained by PCR using a specificprimer at the extreme 50 end, 50-TCACTGAATGT-GAAGTGGG-30 (P-5, located 110 nucleotides up-stream the ATG) and 30 end, 50-CTCTTGTGCAACAACTTTC-30 (P-3, located 116 nu-cleotides downstream from the translational stopcodon). A 1492 bp product was cloned into the pCRIIvector (Invitrogen). The clone, named Ha2Psy andcontaining the entire sequence, was selected andsequenced.
The deduced amino acid sequence of the H.annuus PSY was compared to other PSY sequencesof higher plants, bacteria, cyanobacteria, algae,and fungi. The sequences were aligned by ClustalWversion 1.7 (Thompson et al., 1994) and manuallyadjusted. Phylogenetic analysis was performedusing programs from the PHYLIP group, PHYLogenyInference Package, Version 3.6 (Felsenstein, 1985).A distance matrix was computed according to theDayhoff PAM model by the program Protdist andthen it was used as input for the program Neighbor,where the UPGMA method of clustering wasselected. To calculate the amino acid identity, theregion corresponding to the transit peptide wasexcluded.
For expression analyses, total RNA extractionswere carried out from different organs: cotyledons(C) of 2-week-old seedlings, young leaves (2 cmlong, YL) of 3-week-old plants, mature leaves

ARTICLE IN PRESS
cDNA cloning and expression of the phytoene synthase gene in sunflower 481
(15 cm long, ML), stems (S), and roots (R) of 60-day-old plants grown in a growth chamber under100 mmolm�2 s�1. Moreover, total RNA extractionwas also carried out from cotyledons of 10-day-oldseedlings grown in the dark or under three differentPPFDs (5, 100 and 1500 mmolm�2 s�1). First strandcDNAs were made with a Superscript preamplifica-tion kit (Invitrogen) using 5 mg of total RNA for eachsample. PCRs were performed using gene-specificprimers from cDNA of Psy and b-actin: A21 (50-GCATCGCATATAACTCCCAAAGC-30) and A1105 (50-GCCGGATTGTCAGATGAAGACAT-30) for Psy, ACTINA5 (50-GATTCCGTTCA/TGAGGTC/T-30) and ACTINA 3(50-TC/TTCTGGA/TGGA/TGCAACCACC-30) for b-ac-tin. Primers were designed to yield a 437 bpfragment and a 243 bp fragment for Psy and b-actin, respectively. PCRs were carried out with 28cycles where the cDNAs were exponentially ampli-fied. PCRs were performed in 50 ml of 1� buffer(Applied Biosystem, Applera Italia, Monza, Italy)containing 0.2mM dNTPs, 0.2 mM of each primer (1primer pair for b-actin ad 1 primer pair for Psy),2mM MgCl2, 1 U AmpliTaq DNA polymerase (AppliedBiosystem) and 1.0 ml of single-strand cDNA. Ampli-fication was carried out according to the followingtemperature profile: 95 1C for 4min for denatura-tion, then 94 1C for 30 s, 55 1C for 30 s, 72 1C for 40 sand a final extension of 7min at 72 1C. The relativeamounts of each PCR product were readily quanti-fied by direct scanning with a densitometer ofethidium-stained 2% TAE-agarose gels with a UVPImage Store 5000 (Ultra Violet Products Ltd,Cambridge, England) equipped with the UVPGelBase-GelBlot TM Windows Software. To normal-ize for equal amounts of total RNA and efficiencyof cDNA synthesis from various tissue samples,the intensities of the bands were normalized withthe average intensity of the b-actin productacross the samples investigated. An arbitraryvalue of 100% was assigned to the b-actin productlevel of each sample. Each percentage value is themean of four independent experiments. The datawere treated using analysis of variance proceduresand means were separated by Tukey’s test(P ¼ 0:05).
Results and discussion
The reconstructed full-length cDNA sequence(GenBank accession number AJ304825), 1598 bplong, obtained from 30 and 50 RACE, was confirmedfrom the sequence of the clone Ha2Psy (1492 bp)and it contained a 1242 bp CDS, 172-nucleotides of50-untranslated region (UTR), and 170-nucleotides
of 30-UTR (data not shown). The predicted proteindisplayed a sequence of 414 amino acid residues(Fig. 1A), with a calculated molecular mass of46.8 kDa. A putative transit sequence for plastidtargeting was predicted to be the N-terminalresidues 1–67 by concordance with the predictedtransit peptides of daffodil (GenBank X78814), bellpepper (Romer et al., 1993), Arabidopsis (Scolnikand Bartley, 1993), and tomato PSY1 (Ray et al.,1992), which are 45, 47, 70, and 42 residues,respectively. A ChloroP 1.1 neural network-basedmethod (Emanuelsson et al., 1999) predicts atransit peptide cleavage site between residues 67and 68.
The phylogenetic analysis clearly demonstratedthat the sunflower Psy is clustered to Tagetes Psygene (GenBank AY099482), with which it showed anoverall amino acid identity of 93.7% (Fig. 1B).Moreover, the sunflower Psy sequence relatesclosely with other Psy sequences of higher plants,including PSY2 (Bartley and Scolnik, 1993) and PSY1(Ray et al., 1992) of tomato and PSY of Capsicum(Romer et al., 1993), which showed overall iden-tities of 86.1%, 83.0% and 83.6%, respectively. Alower identity was showed for Arabidopsis (Scolnikand Bartley, 1993) and maize (GenBank U32636)PSY, respectively, 81.0% and 75.2%. A significantidentity was also observed for the PSY from the algaDunaliella bardawil (GenBank U91900) and forphytoene synthase (CRTB) of cyanobacterium Spir-ulina platensis (AB001284), respectively, 58.2% and48.7%. By contrast, a low identity (11.5–30.2%) wasobserved with the phytoene desaturase (CRTE) ofbacteria and fungi.
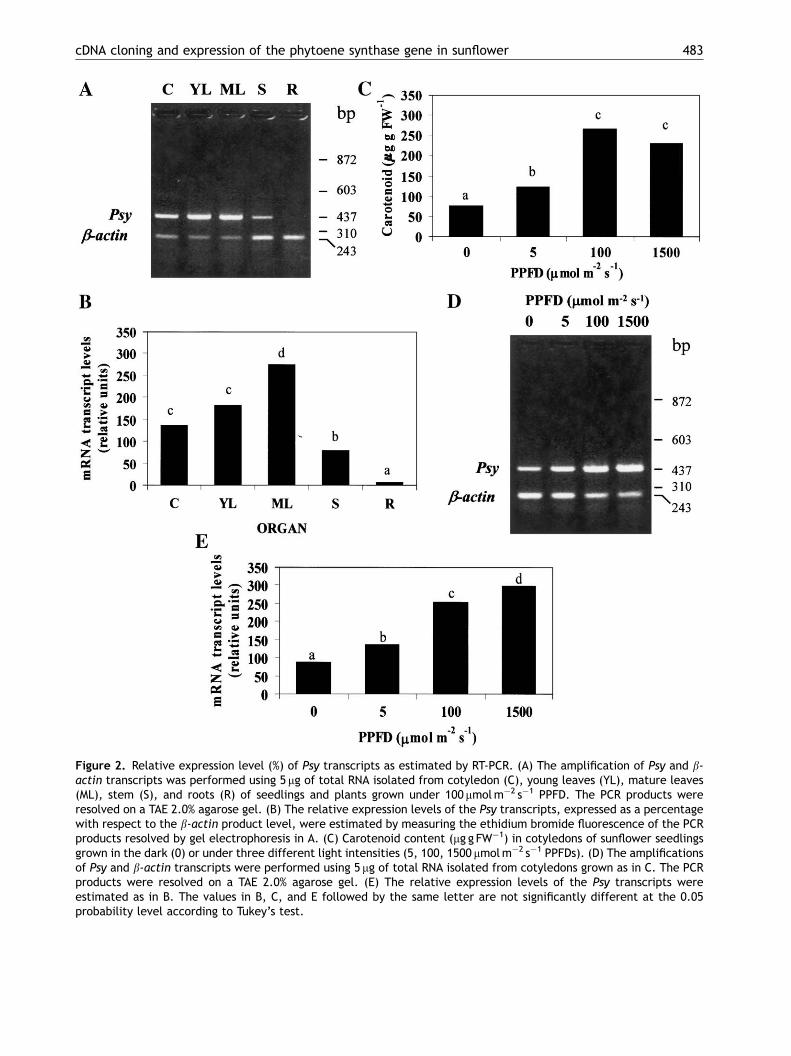
The organ-specific expression of sunflower Psywas analysed by relative RT-PCR assays (Fig. 2Aand B). The HaPsy expression was very high incotyledons, and young and mature leaves. Incontrast, mRNA transcript levels were compara-tively lower in the stem and nearly absent inroots. Giuliano et al. (1993) reported that tomatoPsy expression was relatively constant andrather down-regulated during leaf development.In contrast, we observed that the HaPsytranscript level was changed during leaf expansion,which suggested that the HaPsy expression isregulated during the process of leaf development.In fact, a higher Psy transcript accumulation wasobserved from mature expanded leaves, likely dueto the expression pattern displayed by Citrusunshiu Psy (Ikoma et al., 2001; Kim et al., 2001).The elevated accumulation of HaPsy transcripts inmature leaves might provide to the carotenoidcontent increase that characterizes the sunflowerleaf expansion (M. Fambrini et al., unpublishedresults).

ARTICLE IN PRESS
Figure 1. (A) Deduced amino acid sequence of novel phytoene synthase (Psy) cDNA from H. annuus. The putative signalof chloroplastic transit peptide is underlined, (B) UPGMA tree based on a Dayhoff PAM distance matrix generated usingProtdist and Neighbor programs (Phylip package version 3.572). The GenBank accession numbers are indicated inbrackets.
M. Salvini et al.482
The role of Psy in controlling carotenoid bio-synthesis in plants is evident during fruit ripening(Giuliano et al., 1993; Bramley, 2002) and flowerdevelopment (Moehs et al., 2001; Zhu et al., 2002)when an increase of the corresponding mRNAcontent correlates with enhanced carotenoid pro-duction. Light and light intensity are two of themost important factors involved in the regulation ofcarotenoid synthesis in the chloroplast (von Lintiget al., 1997; Welsch et al., 2000; Simkin et al.,2003b). Light-dependent increases of phytoenesynthase mRNA have been reported in severalspecies (Bartley and Scolnik, 1993; von Lintig etal., 1997; Welsch et al., 2003; Simkin et al.,2003b). It is also demonstrated that the increaseof Psy mRNA during the etioplast/chloroplastconversion is mediated via the phytochrome system(von Lintig et al., 1997; Welsch et al., 2003). Inaddition, the carotenoid content can influence theexpression of some carotenogenic genes (e.g.phytoene desaturase and z-carotene desaturase)through feedback inhibition by end-product accu-mulation (Romer et al., 2000; Fraser et al., 2000;Bramley, 2002; Simkin et al., 2003a). To evaluatethe relationship between the light-dependentcarotenoid accumulation and the HaPsy expression
we have analyzed the HaPsy transcript levels onseedlings grown in dark or under different lightconditions. We observed that with increasing lightintensity, the content of total carotenoid increasedwith a concurrent increase of the HaPsy transcriptaccumulation (Figs. 2C and E). In particular, fromdark to 1500 mmolm�2 s�1 of PPFD a continuousincrease of the HaPsy transcripts of three- to four-fold was observed. The rise of the HaPsy transcriptaccumulation from 100 to 1500 mmolm�2 s�1, whencarotenoids are already constant, is difficult toexplain. In green pepper leaves, it has been shownthat after transfer from light of 150–280mmolm�2 s�1,the rate of carotenoid photo-oxidation exceeds that ofsynthesis (Simkin et al., 2003a). It is likely that thehigher HaPsy activity might contribute to the biosyn-thetic replacement of those carotenoids destroyed byphoto-oxydation processes in sunflower cotyledonsgrown under 1500mmolm�2 s�1. Carotenogenesis ingreen tissues must be linked to the formation of acompetent membrane structure (Welsch et al., 2000),as well as the formation of chlorophylls, proteins,and lipids (Bramley, 2002). We cannot rule out thatone or more post-transcriptional control pointsmust be decisive in the regulation of the PSYenzymatic activity in sunflower cotyledons.
ARTICLE IN PRESS
Figure 2. Relative expression level (%) of Psy transcripts as estimated by RT-PCR. (A) The amplification of Psy and b-actin transcripts was performed using 5 mg of total RNA isolated from cotyledon (C), young leaves (YL), mature leaves(ML), stem (S), and roots (R) of seedlings and plants grown under 100 mmolm�2 s�1 PPFD. The PCR products wereresolved on a TAE 2.0% agarose gel. (B) The relative expression levels of the Psy transcripts, expressed as a percentagewith respect to the b-actin product level, were estimated by measuring the ethidium bromide fluorescence of the PCRproducts resolved by gel electrophoresis in A. (C) Carotenoid content (mg g FW�1) in cotyledons of sunflower seedlingsgrown in the dark (0) or under three different light intensities (5, 100, 1500 mmolm�2 s�1 PPFDs). (D) The amplificationsof Psy and b-actin transcripts were performed using 5 mg of total RNA isolated from cotyledons grown as in C. The PCRproducts were resolved on a TAE 2.0% agarose gel. (E) The relative expression levels of the Psy transcripts wereestimated as in B. The values in B, C, and E followed by the same letter are not significantly different at the 0.05probability level according to Tukey’s test.
cDNA cloning and expression of the phytoene synthase gene in sunflower 483

ARTICLE IN PRESS
M. Salvini et al.484
Acknowledgement
M. Salvini was supported by a Scuola NormaleSuperiore Grant.
References
Bartley GE, Scolnik PA. J Biol Chem 1993;268:25718–21.Bramley PM. J Exp Bot 2002;53:2107–13.Chomczynski P, Sacchi N. Anal Biochem 1987;162:156–9.Emanuelsson O, Nielsen H, von Hejne G. Protein Sci
1999;8:978–84.Fambrini M, Pugliesi C, Vernieri P, Giuliano G, Baroncelli
S. Theor Appl Genet 1993;87:65–9.Felsenstein J. Evolution 1985;39:783–91.Fraser PD, Schuch W, Bramley PM. Planta
2000;211:361–9.Giuliano G, Bartley GE, Scolnik PA. Plant Cell
1993;5:379–87.Giuliano G, Giliberto L, Rosati C. Trends Plant Sci
2002;7:427–9.Ikoma Y, Komatsu A, Kita M, Ogawa K, Omura M, Yano M,
Moriguchi T. Physiol Plant 2001;111:232–8.Kim I-J, Ko K-C, Kim C-S, Chung WI. J Plant Physiol
2001;158:795–800.Lichtenthaler HK. Annu Rev Plant Physiol Plant Mol Biol
1999;50:47–65.Lichtenthaler HK, Schwender J, Disch A, Rohmer M. FEBS
Lett 1997;400:271–4.
Moehs CP, Tian L, Osteryoung KW, DellaPenna D. Plant MolBiol 2001;45:281–93.
Ray J, Moureau P, Bird C, Bird A, Grierson D, Maunders M,Truesdale M, Bramley P, Schuch W. Plant Mol Biol1992;19:401–4.
Rodrıquez-Concepcion M, Boronat A. PIant Physiol2002;130:1079–89.
Romer S, Hugueney P, Bouvier F, Camara B, Kuntz M.Biochem Biophys Res Commun 1993;196:1414–21.
Romer S, Fraser PD, Kiano JW, Shipton CA, Misawa N,Schuch W, Bramley P. Nature Biotech 2000;18:666–9.
Sandmann G. Arch Biochem Biophys 2001;385:4–12.Sandmann G. Physiol Plant 2002;116:431–40.Scolnik PA, Bartley GE. Plant Physiol 1993;103:
1475.Simkin AJ, Laboure A-M, Kuntz M, Sandmann G. Z
Naturforsch 2003a;58c:371–80.Simkin AJ, Zhu C, Kuntz M, Sandmann G. J Plant Physiol
2003b;160:439–43.Thompson JD, Higgins GD, Gibson TJ. Nucl Acids Res
1994;22:4673–80.von Lintig J, Welsch R, Bonk M, Giuliano G, Batshauer A,
Kleinig H. Plant J 1997;12:625–34.Welsch R, Beyer P, Hugueney P, Kleinig H, von Lintig J.
Planta 2000;211:846–54.Welsch R, Medina J, Giuliano G, Beyer P, von Lintig J.
Planta 2003;216:523–34.Zhu C, Yamamura S, Koiwa H, Nishihara M, Sandmann G.
Plant Mol Biol 2002;48:277–85.




















