
cDNA-amplified fragment length polymorphism to study the transcriptional responses of Lactobacillus...
10
ORIGINAL ARTICLE cDNA-amplified fragment length polymorphism to study the transcriptional responses of Lactobacillus rhamnosus growing in cheese-like medium C.G. Bove, C. Lazzi, V. Bernini, B. Bottari, E. Neviani and M. Gatti Department of Genetics, Biology of Microorganisms, Anthropology and Evolution, University of Parma, Parma, Italy Introduction Lactic acid bacteria (LAB) play different roles in cheese making. Starter LAB (SLAB) participate in the fermenta- tion process, whereas nonstarter LAB (NSLAB), probably present in all cheeses, are involved in the maturation, influencing flavour development (Beresford et al. 2001; Beresford and Williams 2004; Broadbent and Steele 2005). Although the role of NSLAB in cheese ripening has not been clarified yet, different authors have suggested their importance in this production step (Settanni and Mosch- etti 2010). In Parmigiano Reggiano cheese, it was demon- strated that SLAB are dominant until the second month of ripening, and after cheese brining, the species NSLAB, especially Lactobacillus rhamnosus, can grow and increase in number, while SLAB cells undergo autolysis (De Dea Lindner et al. 2008; Gatti et al. 2008). Lact. rhamnosus was shown to be the dominant species present when essential nutrients, such as sugars, are lacking. Therefore, this species seems to well adapt to the absence of lac- tose in cheese, confirming an optimal adaptability to unfavourable growth conditions. Presumably, this charac- teristic is because of the ability of Lact. rhamnosus to use nitrogen fraction as an alternative energy source. Banks and Williams (2004) reported that a potential energy sources in maturing cheese include amino acids generated during ripening. Peptides and amino acids can be catabolized by lactobacilli via transamination reaction (Kieronczyk et al. 2001; Yvon and Rijnen 2001; Liu et al. 2003). So far, only few studies have been carried out on Lact. rhamnosus in cheese ripening (De Dea Lindner et al. 2008; Gatti et al. 2008; Neviani et al. 2009). The latest Keywords cDNA-AFLP, cheese-like conditions, gene expression, Lactobacillus rhamnosus, transcription profiling. Correspondence Claudio G. Bove, Department of Genetics, Biology of Microorganisms, Anthropology and Evolution, University of Parma, Viale Usberti 11 ⁄ A, 43100 Parma, Italy. E-mail: [email protected] 2011 ⁄ 0271: received 16 February 2011, revised 30 May 2011, accepted 27 June 2011 doi:10.1111/j.1365-2672.2011.05101.x Abstract Aims: Lactobacillus rhamnosus is a dominant species during Parmigiano Reggiano cheese ripening and exhibits a great adaptability to unfavourable growth conditions. Gene expression of a Lact. rhamnosus, isolated from Parmigiano Reggiano cheese, grown in a rich medium (MRS) and in a cheese- like medium (CB) has been compared by a novel cDNA-amplified fragment length polymorphism (cDNA-AFLP) protocol. Methods and Results: Two techniques, capillary and gel electrophoresis cDNA- AFLP, were applied to generate unique transcript tags from reverse-transcribed messenger RNA using the immobilization of biotinylated 3¢-terminal cDNA fragments on streptavidin-coated Dynabeads. The use of three pairs of primers allowed detecting 64 genes expressed in MRS and 96 in CB. Different tran- scripts were observed when Lact. rhamnosus was cultured on CB and MRS. Conclusions: The cDNA-AFLP approach proved to be able to show that Lact. rhamnosus modifies the expression of a large part of genes when cultivated in CB compared with growth under optimal conditions (MRS). In particular, the profiles of the strain grown in CB were more complex probably because the cells activate different metabolic pathways to generate energy and to respond to the environmental changes. Significance and Impact of Study: This is the first research on Lact. rhamnosus isolated from cheese and represents one of the few concerning bacterial tran- scriptomic analysis towards cDNA-AFLP approaches. Journal of Applied Microbiology ISSN 1364-5072 ª 2011 The Authors Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology 855
Transcript of cDNA-amplified fragment length polymorphism to study the transcriptional responses of Lactobacillus...

ORIGINAL ARTICLE
cDNA-amplified fragment length polymorphism to studythe transcriptional responses of Lactobacillus rhamnosusgrowing in cheese-like mediumC.G. Bove, C. Lazzi, V. Bernini, B. Bottari, E. Neviani and M. Gatti
Department of Genetics, Biology of Microorganisms, Anthropology and Evolution, University of Parma, Parma, Italy
Introduction
Lactic acid bacteria (LAB) play different roles in cheese
making. Starter LAB (SLAB) participate in the fermenta-
tion process, whereas nonstarter LAB (NSLAB), probably
present in all cheeses, are involved in the maturation,
influencing flavour development (Beresford et al. 2001;
Beresford and Williams 2004; Broadbent and Steele 2005).
Although the role of NSLAB in cheese ripening has not
been clarified yet, different authors have suggested their
importance in this production step (Settanni and Mosch-
etti 2010). In Parmigiano Reggiano cheese, it was demon-
strated that SLAB are dominant until the second month
of ripening, and after cheese brining, the species NSLAB,
especially Lactobacillus rhamnosus, can grow and increase
in number, while SLAB cells undergo autolysis (De Dea
Lindner et al. 2008; Gatti et al. 2008). Lact. rhamnosus
was shown to be the dominant species present when
essential nutrients, such as sugars, are lacking. Therefore,
this species seems to well adapt to the absence of lac-
tose in cheese, confirming an optimal adaptability to
unfavourable growth conditions. Presumably, this charac-
teristic is because of the ability of Lact. rhamnosus to use
nitrogen fraction as an alternative energy source. Banks
and Williams (2004) reported that a potential energy
sources in maturing cheese include amino acids generated
during ripening. Peptides and amino acids can be
catabolized by lactobacilli via transamination reaction
(Kieronczyk et al. 2001; Yvon and Rijnen 2001; Liu et al.
2003). So far, only few studies have been carried out on
Lact. rhamnosus in cheese ripening (De Dea Lindner et al.
2008; Gatti et al. 2008; Neviani et al. 2009). The latest
Keywords
cDNA-AFLP, cheese-like conditions, gene
expression, Lactobacillus rhamnosus,
transcription profiling.
Correspondence
Claudio G. Bove, Department of Genetics,
Biology of Microorganisms, Anthropology and
Evolution, University of Parma, Viale Usberti
11 ⁄ A, 43100 Parma, Italy.
E-mail: [email protected]
2011 ⁄ 0271: received 16 February 2011,
revised 30 May 2011, accepted 27 June 2011
doi:10.1111/j.1365-2672.2011.05101.x
Abstract
Aims: Lactobacillus rhamnosus is a dominant species during Parmigiano
Reggiano cheese ripening and exhibits a great adaptability to unfavourable
growth conditions. Gene expression of a Lact. rhamnosus, isolated from
Parmigiano Reggiano cheese, grown in a rich medium (MRS) and in a cheese-
like medium (CB) has been compared by a novel cDNA-amplified fragment
length polymorphism (cDNA-AFLP) protocol.
Methods and Results: Two techniques, capillary and gel electrophoresis cDNA-
AFLP, were applied to generate unique transcript tags from reverse-transcribed
messenger RNA using the immobilization of biotinylated 3¢-terminal cDNA
fragments on streptavidin-coated Dynabeads. The use of three pairs of primers
allowed detecting 64 genes expressed in MRS and 96 in CB. Different tran-
scripts were observed when Lact. rhamnosus was cultured on CB and MRS.
Conclusions: The cDNA-AFLP approach proved to be able to show that
Lact. rhamnosus modifies the expression of a large part of genes when
cultivated in CB compared with growth under optimal conditions (MRS). In
particular, the profiles of the strain grown in CB were more complex probably
because the cells activate different metabolic pathways to generate energy and
to respond to the environmental changes.
Significance and Impact of Study: This is the first research on Lact. rhamnosus
isolated from cheese and represents one of the few concerning bacterial tran-
scriptomic analysis towards cDNA-AFLP approaches.
Journal of Applied Microbiology ISSN 1364-5072
ª 2011 The Authors
Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology 855

study demonstrated that the intraspecies heterogeneity,
found for 66 Lact. rhamnosus strains isolated during the
same Parmigiano Reggiano cheese making, seems to be
correlated with their abilities to adapt to the hostile envi-
ronment of the cheese throughout the ripening (Bove
et al. 2011). It was also found that the great part of
Lact. rhamnosus strains belonged to few biotypes that
were present in cheese from the first or second month of
ripening up to 10 months and beyond (Bove et al. 2011).
The detection of biotypes correlated with specific steps in
cheese ripening suggested that these strains may have spe-
cific metabolic activities, which allow an adaptation to
the microenvironment of cheese ripening.
The high degree of adaptation of Lact. rhamnosus to
different environments and conditions has been studied
towards several approaches (Succi et al. 2005; Koskenni-
emi et al. 2009) and probably involves many metabolic
and physiological changes. To identify and analyse the
genes involved in biological processes, genome-wide
expression analysis is considered one of the most efficient
tool (Breyne and Zabeau 2001). In this regard, cDNA-
amplified fragment length polymorphism (cDNA-AFLP)
is a method for genome-wide expression analysis often
proposed as an alternative to microarrays methods. In
fact, with respect to microarrays, at lower cost, cDNA-
AFLP has the advantage that any unknown genome or set
of genomes can be studied without prior sequence knowl-
edge and offers the possibility to detect also species-
specific genes that could be lacking in the reference strain.
Further, the cDNA-AFLP approach allows the detection
of lowly expressed genes and may allow the discrimina-
tion between homologous genes. Above all, the applica-
tion of cDNA-AFLP is suggested to study organisms with
complete genome sequence or cDNA collection unavail-
ability (Breyne and Zabeau 2001).
cDNA-AFLP is carried out according to the principle
of AFLP. The AFLP technique is based on the digestion
of DNA templates followed by the ligation of adapters to
restriction fragments and the selective PCR amplification
of subsets of these fragments using selective AFLP primers
containing a variable number of selective nucleotides.
Similarly, the original cDNA-AFLP method (Bachem et al.
1996) involves the reverse transcription of mRNA into
double-stranded cDNA, followed by the generation of a
complex mixture of transcript-derived fragments (TDFs)
by restriction enzyme digestion and ligation of specific
adapters, selective PCR amplification and, finally, the
visualization of the TDFs on high-resolution (sequence)
gels (Vuylsteke et al. 2007).
Recently, different researches were published to
improve the original cDNA-AFLP method (Decorosi et al.
2005; Kadota et al. 2007; Weiberg et al. 2008; Xiaohu
et al. 2009). Nevertheless, the most important variant is
the modification of the original protocol based on ‘one-
gene-multiple-tag’ into ‘one-gene-one-tag’ as reported by
Vuylsteke et al. (2007) and Breyne and Zabeau (2001).
The ‘one-gene-one-tag’ cDNA-AFLP protocol involves a
reduction in TDFs to a single fragment for each cDNA by
selecting the 3¢-terminal restriction fragment of each tran-
script before selective amplification. Recently, Levterova
et al. (2010) have developed a cDNA-AFLP strategy to
study the gene transcription in fungi based on the use of
capillary electrophoresis.
Although cDNA-AFLP technique has been extensively
applied to analyse transcription profiles of eukaryotic
organisms (Reijans et al. 2003; Bensch and Akesson 2005;
Ekkapongpisit et al. 2007; Neveu et al. 2007), there are
few data regarding cDNA-AFLP application to bacteria
(Dellagi et al. 2000; Noel et al. 2001). This is probably
due to the difficulties of working with bacterial mRNA. A
major technical challenge for transcriptome sequencing is
the low relative abundance of mRNAs in total cellular
RNA (1-5%) (Neidhardt and Umbarger 1996), made of
rRNAs and tRNAs that are, respectively, 86 and 14%
(Karpinets et al. 2006). Furthermore, unlike eukaryotic
mRNAs, bacterial mRNA generally lacks poly(A)-tail,
which makes their isolation and analysis complicated (He
et al. 2010).
In this study, we developed, for the first time, a cDNA-
AFLP protocol to study the gene expression profiles of
Lact. rhamnosus strain isolated from Parmigiano Reggiano
cheese. The novel transcriptomic approach was applied to
evaluate the changes in gene expression profile during the
growth of Lact. rhamnosus either in a rich medium
(MRS) or in a cheese-like medium, a medium based on
grated Parmigiano Reggiano ripened cheese, characterized
by the absence of milk sugars and rich in peptides, amino
acids and NaCl (Neviani et al. 2009).
Materials and methods
Bacterial strains and media
Lactobacillus rhamnosus 1473, previously isolated from
20-month ripened Parmigiano Reggiano (PR) cheese,
belonging to biotype no. 1 (Bove et al. 2011) was used in
this study. The strain, isolated on MRS agar (Oxoid Ltd,
Basingstoke, UK), was maintained as stock culture at
)80�C in MRS broth supplemented with 15% glycerol
(w ⁄ v). For RNA extraction, it was first propagated twice,
with a 2% inoculum in 10 ml of MRS broth at 30�C for
24 h; then, the culture was anaerobically (Gas Generating
kit; Oxoid Ltd) propagated twice at 30�C, until the top of
logarithmic phase was reached, in two different media:
MRS broth (De Man et al. 1960) and a cheese-like broth
called cheese broth (CB). CB was prepared partially
cDNA-AFLP for Lactobacillus rhamnosus C.G. Bove et al.
856 Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology
ª 2011 The Authors

modifying the protocol described by Neviani et al. (2009).
Fine-grain grated 20-month ripened PR cheese was dis-
solved (120 g l)1) in 0Æ07 mol l)1 sodium citrate pH 7Æ5(Sigma-Aldrich Co., St Louis, MO, USA). The suspension
was incubated at 42�C for 50 min under stirring condi-
tion (150 g); then, it was centrifuged (8000 g for 15 min
at 4�C) and filtered through sterile gauze to retain the
surfaced fat layer. Filtered liquid was acidified to pH 4Æ6by adding 1 mol l)1 HCl, sterilized for 15 min at 121�C
and centrifuged at 8000 g for 15 min to remove unhydro-
lysed proteins (casein and heat-denatured serum pro-
teins); 1 mol l)1 NaOH was added to supernatant until
pH 6Æ1 was reached. Freeze-dried cell lysate (protein con-
centration of 4 mg ml)1 determined using Bradford pro-
tein assay) of Lactobacillus helveticus PR775, previously
isolated from PR cheese (University of Parma collection),
was added at a final concentration corresponding to
8 log CFU ml)1. The volume of this freeze-dried cell
lysate added was 5 ml in 100 ml of CB (final protein
concentration of 0Æ2 mg ml)1). The resulting medium
was sterilized by filtration on 0Æ22-lm membrane filters
(Millipore Corporation, Bedford, MA, USA). For the
preparation of freeze-dried cell lysate, Lact. helveticus
PR775 was maintained as stock culture at )80�C in MRS
broth supplemented with 15% glycerol (w ⁄ v). It was first
propagated twice, with a 2% inoculum, in MRS broth at
30�C for 24 h; then, the culture was propagated in MRS
at 30�C until the stationary phase was reached. Harvested
cells were washed in 0Æ05 mol l)1 Tris ⁄ HCl pH 7Æ5,
centrifuged (15000 g for 15 min at 4�C) and frozen at
)80�C or directly resuspended in 0Æ05 mol l)1 Tris ⁄ HCl
pH 7Æ5. Cells were disrupted using a Branson model B15
Sonifier by 3 cycles of sonication (1 min each). After
pelleting of unbroken cells (15 000 g for 15 min at 4�C),
the supernatant was maintained as stock at )20�C.
RNA extraction and cDNA synthesis
The growth of Lact. rhamnosus 1473 at 30�C in both MRS
and CB was monitored by optical density (OD) measure-
ment (Jasco, Tokyo, Japan) until the top of logarithmic
phase was reached. About 109 cells at the top of logarith-
mic phase were harvested, and total RNA was stabilized
using RNeasy Protect Bacteria Mini kit (Qiagen, Valencia,
CA, USA) and isolated using RNeasy Mini Kit (Qiagen).
The protocol included the removal of DNA from the sam-
ples by DNAseI using the RNase-Free DNase Set (Qiagen).
Total RNA was resuspended in 50 ll of RNAse-free water
and checked on 0Æ8% agarose gel (Sigma-Aldrich). The
concentration and purity of RNA were assessed by OD
measurement at 260 and 280 nm (Jasco). Prior to cDNA
synthesis, total RNA was enriched in mRNA and then the
mRNA was polyadenylated (Fig. 1).
Using MicrobExpress bacterial mRNA enrichment kit
(Ambion, Applied Biosystems, Monza, Italy), 10 lg of
total RNA from three independent biological replicates
was enriched in mRNA by removing the 16S and 23S
ribosomal RNAs (rRNA). The enriched mRNA was puri-
fied from 5S RNA using the MEGAclear Purification kit
(Ambion) and quantified by spectrophotometer (Jasco).
To add a poly(A) tail to RNA transcripts, we used the
Poly(A) Tailing kit (Ambion). The poly-A RNA (2 lg)
3′5′
(A)n5′
biotinilated5′
PolyA 3′
5′ PolyA 3′
Total RNA
Capture oligo
Oligo MagBead
rRNA molecule
mRNA enrichmentEnriched mRNA
Poly(A) tail added
cDNA synthesis
First restriction digestion
3′ end capturing Streptavidin bead
Second restriction digestion
Adapter ligation
Selective PCR amplificationPrimer 1
Primer 2
(T)n oligo(dT)
Adaptator Adaptator
ge-cDNA-AFLP ce-cDNA-AFLP
Figure 1 Schematic overview of the cDNA-AFLP procedure with 3¢end capture applied to RNA isolated from Lactobacillus rhamnosus
1473. Total RNA isolated from Lact. rhamnosus 1473 is enriched for
mRNA by subtractive hybridization as indicated by the manufacturer
(MicrobExpress; Ambion, Huntington, UK). Poly(A) tail is added to
RNA transcripts using the Poly(A) Tailing kit (Ambion, Applied Biosys-
tems). cDNA is synthesized by reverse transcription using a biotinylat-
ed oligodT, followed by second-strand synthesis. Digestion with first
restriction enzyme captures with the aid of streptavidin-coated
magnetic beads followed by the digestion with the second restriction
enzyme, the removal of the 3¢ fragments and ligation of adaptors.
Selective amplification and separation of the fragments on a polyacryl-
amide gel and capillary electrophoresis and visualization.
C.G. Bove et al. cDNA-AFLP for Lactobacillus rhamnosus
ª 2011 The Authors
Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology 857

was used in the synthesis of double-stranded cDNA
according to the protocol reported by Vuylsteke et al.
(2007) except for the following modifications.
For first-strand complementary DNA synthesis, 1 ll of
100 lmol l)1 oligo dT25bio primer (Eurofins MWG
Operon, Ebersberg, Germany) was added to 20 ll of RNA
sample, and the mixture was heated to 65�C for 5 min
and quick-chilled on ice. The sample was collected by
brief centrifugation, and 4 ll of 5X buffer, 2 ll of DTT
(0Æ1 mol l)1) and 1 ll of dNTPs mix (10 mmol l)1) (In-
vitrogen Life Technologies, Carlsbad, CA, USA) were
added. The sample was kept at 42�C for 2 min to equili-
brate the temperature and then was added with 2 ll of
SuperScript� II RT (200 U ll)1) (Invitrogen Life Tech-
nologies) and incubated at 42�C for 2 h. For second-
strand cDNA synthesis, 32 ll of 5X second-strand buffer,
3 ll of dNTPs mix (10 mmol l)1), 6 ll of DTT
(0Æ1 mol l)1), 1Æ5 ll of E. coli ligase (10 U ll)1), 0Æ2 ll of
ribonuclease H (10 U ll)1), 5 ll of DNA polymerase I
(10 U ll)1) (Invitrogen Life Technologies) and 72Æ3 ll of
H2O were mixed, and all the mixture was added to 40 ll
of first-strand cDNA product. After an incubation of 2 h
at 16�C, the double-stranded cDNA was purified and
concentrated in 30 ll of elution buffer NE using the
NucleoSpin Extract II kits (Macherey-Nagel, Duren,
Germany). The cDNA was checked on 0Æ8% agarose gel
(Sigma-Aldrich) and quantified by spectrophotometer
(Jasco).
Preparation of template for cDNA-AFLP analysis
The template for cDNA-AFLP analysis was prepared
making some modifications to the procedure reported by
Vuylsteke et al. (2007) (Fig. 1). This protocol describes
the generation of cDNA-AFLP fragments detected using
infrared dye (IRD) detection technology and the Odyssey
Infrared Imaging System. A first reaction mixture con-
taining 1 ll of EcoRI (10 U ll)1), 4 ll of NEBuffer 4,
0Æ2 ll of BSA 100X (New England Biolabs, Ipswich, MA,
USA) and 14Æ8 ll of free-nuclease water was added to
20 ll of double-stranded cDNA (about 500 ng). This
reaction mixture was incubated for 2 h at 30�C. Subse-
quently, the mixture was added to 40 ll of solution con-
taining 10 ll of Dynabeads M-280 streptavidin
(Invitrogen Life Technologies) and 40 ll of 2X STEX buf-
fer (40 ml NaCl 5 mol l)1, 2 ml Tris–HCl 1 mol l)1 pH
8Æ0, 400 ll EDTA 0Æ5 mol l)1 pH 8Æ0 and water up to
100 ml). This reaction mixture was incubated for 30 min
at room temperature. This step allows the immobilization
of biotinylated 3¢-terminal cDNA fragments on strepta-
vidin-coated Dynabeads. The Dynabeads were collected,
washed and resuspended in 100 ll of 1X STEX buffer.
Subsequently, a second reaction mixture containing 1 ll
of MseI (10 U ll)1), 4 ll of NEBuffer 4, 0Æ1 ll of BSA
100X (New England Biolabs) and 4Æ9 ll of free-nuclease
water was added. This mixture was incubated for 2 h at
30�C under stirring condition. The supernatant (40 ll)
containing the digested cDNA fragment was added to
10 ll of ligation mixture. This consists in a total volume
of 10 ll containing 1 ll EcoRI adapter (Vuylsteke
et al. 2007) (5 pmol ll)1) (Eurofins MWG Operon), 1 ll
MseI adapter (50 pmol ll)1) (Vuylsteke et al. 2007)
(Eurofins MWG Operon), 1 ll NEBuffer 4 (New England
Biolabs), 1 ll T4 DNA ligase (1 U ll)1), 1 ll ATP
(10 mmol l)1), 0Æ1 ll of BSA 100X (Invitrogen Life Tech-
nologies) and 4Æ9 ll of free-nuclease water. This reaction
mixture was incubated for 3 h at 30�C. Subsequently, the
mixture was diluted to 100 ll with 0Æ1 mol l)1 TE buffer
and stored at )20�C.
Nonselective PCR
The ‘nonselective’ primers EcoRI-0 and MseI-0 (Eurofins
MWG Operon) were used for the pre-amplification of the
diluted template. Each pre-amplification mixture con-
tained 5 ll of the diluted template previously described,
1Æ5 ll of unlabelled MseI-0 primer (10 lmol l)1), 1Æ5 ll
of labelled IRD700EcoRI-0 primer (10 lmol l)1), 0Æ2 ll
of AmpliTaq (5 U ll)1) (Applied Biosystem-Pe Corpora-
tion, Foster City, CA, USA), 5 ll 10X PCR buffer, 5 ll
MgCl2 (25 mmol l)1), 2 ll dNTPs (5 mmol l)1) and
29Æ8 ll of free-nuclease water. The reaction was subjected
to the following PCR conditions: initial denaturation at
94�C for 2 min, 25 cycles consisted of 30-s denaturation
at 94�C, 1-min annealing at 56�C, 1-min extension at
72�C and 1 cycle of final extension for 10 min at 72�C.
All amplifications were performed in a Mastercycler Ep
Gradient S (Eppendorf, Hamburg, Germany). The pre-
amplification product was diluted 600 times with TE
0Æ1 mol l)1 buffer and stored at )20�C.
Selective PCR and capillary electrophoresis (ce-cDNA-
AFLP)
The ‘selective’ primers 5¢FAM-EcoRI-N (N stands for A
or T or AC), labelled with 5¢-carboxy fluorescein (FAM),
and unlabelled MseI-AC ⁄ T (Table 1) (Eurofins MWG
Operon) were used for selective amplification of the
diluted pre-amplification product. PCR mixture con-
tained 5 ll of diluted pre-amplification product, 0Æ8 ll
of labelled 5¢IRD700EcoRI-N primer (1 lmol l)1), 3 ll
of unlabelled MseI-AC ⁄ T primer (2 lmol l)1), 0Æ2 ll of
AmpliTaq (5 U ll)1) (Applied Biosystem-Pe Corpora-
tion), 2 ll of 10X PCR buffer, 2Æ4 ll of MgCl2(25 mmol l)1), 0Æ8 ll of dNTPs (5 mmol l)1) (Invitrogen
Life Technologies) and 5Æ8 ll of free-nuclease water.
cDNA-AFLP for Lactobacillus rhamnosus C.G. Bove et al.
858 Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology
ª 2011 The Authors

The thermocycling conditions were designed as fol-
lows: initial denaturation at 94�C for 2 min, 13 cycles
consisting of 10-s denaturation at 94�C, 30-s annealing
at 65�C (reduced each cycle by 0Æ7�C), 1-min extension
at 72�C, 23 cycles consisting of 10-s denaturation at
94�C, 30-s annealing at 56�C, 1-min extension at 72�C
(extended 1 s per cycle) and 1 cycle of final extension
for 10 min at 72�C. All amplifications were performed
in a Mastercycler Ep Gradient S (Eppendorf). The
cDNA-AFLP analysis of the amplified products was per-
formed making some modification to the procedure
reported by Lazzi et al. (2009). The amplified products
from selective amplification were added to 1 ll of Gene-
Scan-500 (ROX) size standard (Applied Biosystem-Pe
Corporation) and 24 ll of deionized formamide. The mix-
ture was heated for 5 min and cooled for 10 min on ice.
Samples were loaded and run on the ABI Prism 310
(Applied Biosystem-Pe Corporation) according to the
AFLP Microbial Fingerprinting protocol (Applied Bio-
system-Pe Corporation) and analysed using Genemapper
Analysis Software (Applied Biosystem-Pe Corporation)
according to the manufacturer’s instructions. The data for
each run were saved as an individual GeneScan file and
displayed as an electropherogram. A threshold of 100 RFU
(relative fluorescent unit) was considered in scoring to
highlight only sharp and easily distinguishable peaks; all
signals under this value were treated as background and
not scored.
Selective PCR and gel electrophoresis (ge-cDNA-AFLP)
The ‘selective’ primers 5¢IRD700EcoRI-N, labelled with a
infrared dye (IRDye� 700 phosphoramidite), and unla-
belled MseI-AC ⁄ T (Table 1) (Eurofins MWG Operon)
were used for selective amplification of the diluted pre-
amplification product. PCR mixture contained 5 ll of
diluted pre-amplification product, 0Æ8 ll of labelled
5¢IRD700EcoRI-N primer (1 lmol l)1), 3 ll of unlabelled
MseI-AC ⁄ T primer (2 lmol l)1), 0Æ2 ll of AmpliTaq
(5 U ll)1) (Applied Biosystem-Pe Corporation), 2 ll of
10X PCR buffer, 2Æ4 ll of MgCl2 (25 mmol l)1), 0Æ8 ll of
dNTPs (5 mmol l)1) (Invitrogen Life Technologies) and
5Æ8 ll of free-nuclease water.
The thermocycling conditions were the same as
described in ‘Selective PCR and capillary electrophoresis’
section. Electrophoresis was carried out through a 6%
Long Range gel as reported by Vuylsteke et al. (2007).
Amplification products (6 ll) obtained from selective
amplification were mixed with 3 ll of IR2 Stop Solution
Red (LI-COR Biosciences, Lincoln, NE, USA), denatured
at 95�C for 3 min and then cooled on ice for 10 min.
The 50- to 700-bp IRDye700 Sizing Standard (LI-COR
Biosciences) was denatured at 95�C for 2 min and cooled
on ice for 10 min. Samples were loaded onto the pre-
running gel (30 min at 40 mA), and 50- to 700-bp
IRDye700 Sizing Standard (LI-COR Biosciences) was
loaded every 6 lane for normalization purposes. The gel
was run for 4Æ5 h at 20 mA, and after electrophoresis,
Odissey (LI-COR Biosciences) at 700 nm was used for
gel scanning.
Results
In this study, the application of cDNA-AFLP methodol-
ogy for studying the adaptation of Lact. rhamnosus to
cheese environment is reported. Lact. rhamnosus was
grown in CB and MRS media. The top of logarithmic
phase, corresponding to the beginning of stationary
phase, was reached after 20 h by cells growing in MRS
and after 40 h by cells growing in CB (data not shown).
Two techniques (Fig. 1), capillary electrophoresis
cDNA-AFLP (ce-cDNA-AFLP) and gel electrophoresis
cDNA-AFLP (ge-cDNA-AFLP), have been applied to
Table 1 Sequences of the primers and adaptors used for cDNA-AFLP
Primers
and
adaptors Initials Sequences (5¢–3¢) Applications
EcoRI
adaptor
EcoRI Forw CTCGTAGACTGCGTACC Ligation
EcoRI
adaptor
EcoRI Rev AATTGGTACGCAGTCTAC Ligation
MseI
adaptor
MseI Forw GACGATGAGTCCTGAG’ Ligation
MseI
adaptor
MseI Rev TACTCAGGACTCAT Ligation
EcoRI-0 EcoRI-0 GACTGCGTACCAATTC Nonselective
PCR
EcoRI-A 5¢FAM-EcoRI-A GACTGCGTACCAATTCA Selective
PCR
EcoRI-A 5¢IRD-EcoRI-A GACTGCGTACCAATTCA Selective
PCR
EcoRI-T 5¢FAM-EcoRI-T GACTGCGTACCAATTCT’ Selective
PCR
EcoRI-T 5¢IRD-EcoRI-T GACTGCGTACCAATTCT Selective
PCR
EcoRI-AC 5¢FAM-EcoRI-AC GACTGCGTACCAATTCAC Selective
PCR
EcoRI-AC 5¢IRD-EcoRI-AC GACTGCGTACCAATTCAC Selective
PCR
MseI-0 MseI-0 GATGAGTCCTGAGTAA Nonselective
PCR
MseI-T MseI-T GATGAGTCCTGAGTAAT Selective
PCR
MseI-AC MseI-AC GATGAGTCCTGAGTAAAC Selective
PCR
cDNA-AFLP, cDNA-amplified fragment length polymorphism.
C.G. Bove et al. cDNA-AFLP for Lactobacillus rhamnosus
ª 2011 The Authors
Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology 859

obtain and compare the Lact. rhamnosus genome
expressed at maximum growing level in CB and in MRS.
In the first instance, ce-cDNA-AFLP was carried out to
monitor transcriptional changes related to the growth in
two different media. Subsequently, ge-cDNA-AFLP
allowed us to confirm the different expression profiles in
MRS and CB and to obtain fragments that could be
extracted and identified. Principal changes to the original
cDNA-AFLP procedure (Vuylsteke et al. 2007), consisting
in the isolation of mRNA from total RNA and in the
addition of a 3¢ poly(A) tail to mRNA, were made to
develop a novel protocol. This allowed to use an oligodT
biotinylated in the retrotranscription reaction. In this
way, it was possible to retrotranscribe all mRNA using a
unique primer and, subsequently, to capture only one
cDNA fragment deriving from each mRNA species during
the reactions of enzymatic digestion and immobilization
of biotinylated 3¢-terminal cDNA fragments on streptavi-
din-coated Dynabeads.
Double-stranded cDNA synthesized as previously
described from Lact. rhamnosus grown in CB and MRS
was used as template for AFLP analysis. Three combina-
tions of fluorescently labelled primers (Table 2) were
chosen to obtain 50- to 500-bp length fragments. The
selectivity of the primer pairs was determined by one or
two additional nucleotides to increase the stringency of
the PCR amplification step and the sensitivity of the anal-
ysis. In this way, simpler expression profiles were
obtained and analysed on the basis of the pres-
ence ⁄ absence of transcripts. Thanks to the addiction of
selective nucleotides to the primers, low abundance tran-
scripts were also detected as already shown by Breyne
et al. 2003. The different selective primer combinations
generated profiles characterized both by a different num-
ber of fragments and by fragments with different molecu-
lar weight.
In Table 2, the number of fragments as resulting from
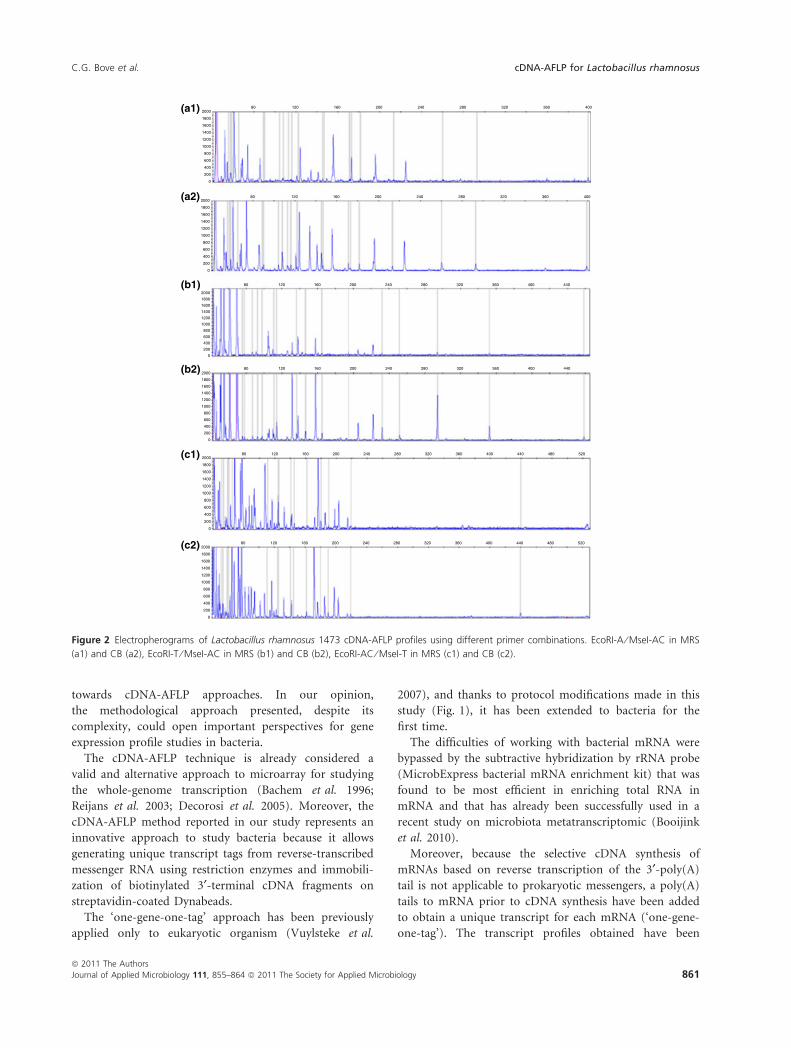
capillary and gel electrophoresis is reported. A significant
higher number of fragments are obtained when perform-
ing PCR amplification with the use of the FAM fluores-
cently labelled primer in ce-cDNA-AFLP protocol
(Fig. 2). The fragments generated by ce-cDNA-AFLP,
visualized as peaks in the electropherograms after selective
amplification, ranged from 15 (Fig. 2b1) to 35 (Fig. 2c2)
depending upon primer combinations and growth condi-
tions (Fig. 2).
The highest number of peaks in ce-cDNA-AFLP and
fragments in ge-cDNA-AFLP was produced both in MRS
and in CB using EcoRI-AC ⁄ MseI-T primer combination
(Table 2; Figs 2c1, 2c2 and 3, lanes 5 and 6). Neverthe-
less, the primer pairs EcoRI-A ⁄ MseI-AC and EcoRI-T ⁄MseI-AC originated the higher number of different frag-
ments between the strain grown in MRS and in CB, both
for gel electrophoresis and for capillary electrophoresis
(Table 2). The number of peaks detected in CB through
capillary electrophoresis was always higher than in MRS,
and on the contrary, the number of fragments detected in
CB through gel electrophoresis was lower than in MRS
using EcoRI-AC ⁄ MseI-T (Table 2).
Overall, the use of three pairs of primers allowed
detecting 64 genes expressed in MRS and 97 in CB by
ce-cDNA-AFLP technique. Comparing profiles by gene
mapper software, we found that nine transcripts were
expressed (peak present) by Lact. rhamnosus only when
grown in MRS, while 42 transcripts were expressed by
Lact. rhamnosus only when grown in CB. As a whole, the
profiles of the strain grown in CB were more complex
(Fig. 3).
Discussion
Adaptation of bacteria to their environment involves
many metabolic and physiological changes, and for this
reason, it is considered a very interesting research field.
Lact. rhamnosus species, among lactobacilli, demonstrated
to be able to adapt to very unfavourable environments
such as the ripening of PR cheese and to grow better than
other species becoming a dominant species.
In this study, a novel cDNA-AFLP protocol was
applied to study the profiles of gene expression of one
Lact. rhamnosus strain isolated from 20-month ripened
Parmigiano Reggiano cheese. This is the first research on
Lact. rhamnosus isolated from cheese and represents one
of the few concerning bacterial transcriptomic analysis
Table 2 Primer combinations used in cDNA-AFLP and number of fragments obtained
Primer combination
ce-cDNA-AFLP peaks ge-cDNA-AFLP fragments
MRS CB MRS CB
EcoRI-A ⁄ MseI-AC 16 (Fig. 2a1) 32 (Fig. 2a2) 16 (Fig. 3 lane 1) 24 (Fig. 3 lane 2)
EcoRI-T ⁄ MseI-AC 15 (Fig. 2b1) 29 (Fig. 2b2) 11 (Fig. 3 lane 3) 16 (Fig. 3 lane 4)
EcoRI-AC ⁄ MseI-T 33 (Fig. 2c1) 36 (Fig. 2c2) 33 (Fig. 3 lane 5) 27 (Fig. 3 lane 6)
Total peaks 64 97 60 67
CB, Cheese-like medium, cDNA-AFLP, cDNA-amplified fragment length polymorphism; MRS, rich medium.
cDNA-AFLP for Lactobacillus rhamnosus C.G. Bove et al.
860 Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology
ª 2011 The Authors
towards cDNA-AFLP approaches. In our opinion,
the methodological approach presented, despite its
complexity, could open important perspectives for gene
expression profile studies in bacteria.
The cDNA-AFLP technique is already considered a
valid and alternative approach to microarray for studying
the whole-genome transcription (Bachem et al. 1996;
Reijans et al. 2003; Decorosi et al. 2005). Moreover, the
cDNA-AFLP method reported in our study represents an
innovative approach to study bacteria because it allows
generating unique transcript tags from reverse-transcribed
messenger RNA using restriction enzymes and immobili-
zation of biotinylated 3¢-terminal cDNA fragments on
streptavidin-coated Dynabeads.
The ‘one-gene-one-tag’ approach has been previously
applied only to eukaryotic organism (Vuylsteke et al.
2007), and thanks to protocol modifications made in this
study (Fig. 1), it has been extended to bacteria for the
first time.
The difficulties of working with bacterial mRNA were
bypassed by the subtractive hybridization by rRNA probe
(MicrobExpress bacterial mRNA enrichment kit) that was
found to be most efficient in enriching total RNA in
mRNA and that has already been successfully used in a
recent study on microbiota metatranscriptomic (Booijink
et al. 2010).
Moreover, because the selective cDNA synthesis of
mRNAs based on reverse transcription of the 3¢-poly(A)
tail is not applicable to prokaryotic messengers, a poly(A)
tails to mRNA prior to cDNA synthesis have been added
to obtain a unique transcript for each mRNA (‘one-gene-
one-tag’). The transcript profiles obtained have been
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
80 120 160 200 240 280 320 360 400 440 480 520
80 120 160 200 240 280 320 360 400 440 480 520
80 120 160 200 240 280 320 360 400 440
80 120 160 200 240 280 320 360 400 440
80 120 160 200 240 280 320 360 400
80 120 160 200 240 280 320 360 400
200
400
600
800
1000
1200
1400
1600
1800
2000
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
200
400
600
800
1000
1200
1400
1600
1800
2000(a1)
(a2)
(b1)
(b2)
(c1)
(c2)
Figure 2 Electropherograms of Lactobacillus rhamnosus 1473 cDNA-AFLP profiles using different primer combinations. EcoRI-A ⁄ MseI-AC in MRS
(a1) and CB (a2), EcoRI-T ⁄ MseI-AC in MRS (b1) and CB (b2), EcoRI-AC ⁄ MseI-T in MRS (c1) and CB (c2).
C.G. Bove et al. cDNA-AFLP for Lactobacillus rhamnosus
ª 2011 The Authors
Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology 861

detected using capillary electrophoresis (ce-cDNA-AFLP)
and gel electrophoresis (ge-cDNA-AFLP).
The two applied detection techniques appear to have
different performances: the former (ce-cDNA-AFLP) is
more sensitive and can evidence a greater number of
genes and the latter (ge-cDNA-AFLP) is necessary to
identify them.
The sensitivity of these two techniques is different
because the capacity of gene separation using the gel is
lower than that obtained using capillary electrophoresis.
Furthermore, the lower number of amplification products
generated from cDNA templates in the ge-cDNA-AFLP
methods may have been due to the presence of tran-
scripts with the same or similar molecular weight that
lead to a unique band in the gel. For this reason, the
total number of gel fragments could be lower than the
total number of genes. Thus, fragment detection and
separation through the gel will be improved to optimize
the correspondence between transcripts obtained by two
techniques.
Considering the number of genes checked to date, the
cDNA-AFLP approach resulted able to show that
Lact. rhamnosus modifies the expression of several genes
when cultivated in cheese-like conditions (CB) compared
with growth under in vitro optimal conditions (MRS).
These media have different composition; in fact, MRS is a
standard laboratory complete medium containing a com-
plex nitrogen source consisting of peptone (protein
hydrolysate) and extracts (from meat and yeast), while
CB is a medium based on grated PR ripened cheese, rich
in peptides, amino acids and NaCl but devoid of milk
sugars (Neviani et al. 2009). Although Lact. rhamnosus
was observed to grow better in MRS, it resulted able to
grow as well in the poorer CB. The larger number of
amplification products generated in ce-cDNA-AFLP from
CB-derived template, with respect to those generated
from MRS-derived template, determined a more com-
plex profiles when Lact. rhamnosus grown in cheese like-
environment.
The transcriptomic profiles resulted to be affected by
the growth medium probably because it induces the
activation of different metabolic pathways in the cell to
generate energy and to respond to the environmental
changes. Different studies demonstrated that the adapta-
tion to dairy niches is associated either with a metabolic
simplification as loss of genes involved in carbohydrate,
amino acid and cofactor metabolisms or with an increase
in the expression of genes involved in peptide hydrolysis
and amino acid catabolism (Bolotin et al. 2004; Hols et al.
2005; van de Guchte et al. 2006; Callanan et al. 2008).
In this regard, further analysis will have to be focused
on gel extraction followed by the identification of
expressed genes. This could be interesting to deepen the
knowledge of the basal metabolism through the identifica-
tion of constitutive genes, as well as to analyse how cells
react to the dairy environment through the identification
of differently expressed genes in the two culture media.
50 bp
100 bp
145 bp
200 bp
204 bp
255 bp
300 bp
350 bp
364 bp
400 bp
700 bp
M 1 2 3 4 5 6 M
Figure 3 cDNA-AFLP fingerprint obtained with two different primer
combinations. M, 50- to 700-bp IRDye700 Sizing Standard; lanes 1
and 2, cDNA-AFLP fingerprinting of Lactobacillus rhamnosus cultured
in MRS and CB, respectively, using EcoRI-A ⁄ MseI-AC primer combina-
tion; lanes 3 and 4, cDNA-AFLP fingerprinting of Lact. rhamnosus
cultured in MRS and CB, respectively, using EcoRI-T ⁄ MseI-AC primer
combination; lanes 5 and 6, cDNA-AFLP fingerprinting of
Lact. rhamnosus cultured in MRS and CB using EcoRI-AC ⁄ MseI-T
primer combination.
cDNA-AFLP for Lactobacillus rhamnosus C.G. Bove et al.
862 Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology
ª 2011 The Authors

Moreover, the expected results could lead to under-
stand whether Lact. rhamnosus physiological mechanisms
of adaptation to cheese-like substrates, which are
fundamental for survival in cheese during long ripening,
could be involved both in ripening processes and in the
definition of organoleptic characteristics of Parmigiano
Reggiano cheese.
Acknowledgements
The authors are grateful to Dr Elisa Sgarbi and Antonietta
Cirasolo for technical assistance.
References
Bachem, C.W., van der Hoeven, R.S., de Bruijn, S.M., Vreug-
denhil, D., Zabeau, M. and Visser, R.G. (1996) Visualiza-
tion of differential gene expression using a novel method
of RNA fingerprinting based on AFLP: analysis of gene
expression during potato tuber development. Plant 9, 745–
753.
Banks, J.M. and Williams, A.G. (2004) The role of nonstarter
lactic acid bacteria in Cheddar cheese ripening. Int J Dairy
Technol 57, 145–152.
Bensch, S. and Akesson, M. (2005) Ten years of AFLP in
ecology and evolution: why so few animals? Mol Ecol 14,
2899–2914.
Beresford, T. and Williams, A. (2004) The microbiology of
cheese ripening. In Cheese: Chemistry, Physics and Micro-
biology ed Fox, P.F., McSweeney, P.L.H., Cogan, T.M. and
Guinee, T.P. pp. 287–318. the Netherlands: Elsevier,
Amsterdam.
Beresford, T.P., Fitzsimons, N.A., Brennan, N.L. and Cogan,
T.M. (2001) Recent advances in cheese microbiology. Int
Dairy J 11, 259–274.
Bolotin, A., Quinquis, B., Renault, P., Sorokin, A., Ehrlich,
S.D., Kulakauskas, S., Lapidus, A., Goltsman, E. et al.
(2004) Complete sequence and comparative genome analy-
sis of the dairy bacterium Streptococcus thermophilus. Nat
Biotechnol 22, 1554–1558.
Booijink, C.C.G.M., Boekhorst, J., Zoetendal, E.G., Smidt, H.,
Kleerebezem, M. and de Vos, W.M.V. (2010) Metatran-
scriptome analysis of the human fecal microbiota reveals
subject-specific expression profiles, with genes encoding
proteins involved in carbohydrate metabolism being
dominantly expressed. Appl Environ Microbiol 76,
5533–5540.
Bove, C.G., De Dea Lindner, J., Lazzi, C., Gatti, M. and
Neviani, E. (2011) Evaluation of genetic polymorphism
among Lactobacillus rhamnosus nonstarter Parmigiano
Reggiano cheese strains. Int J Food Microbiol 144, 569–572.
Breyne, P. and Zabeau, M. (2001) Genome-wide expression
analysis of plant cell cycle modulated genes. Curr Opin
Plant Biol 4, 136–142.
Breyne, P., Dreesen, R., Cannoot, B., Rombaut, D., Vandepo-
ele, K., Rombauts, S., Vanderhaeghen, R., Inze, D. et al.
(2003) Quantitative cDNA-AFLP analysis for genome-wide
expression studies. Mol Gen Genomics 269, 173–179.
Broadbent, J.R. and Steele, J.L. (2005) Cheese flavour and the
genomics of lactic acid bacteria. ASM News 71, 121–125.
Callanan, M., Kaleta, P., O’Callaghan, J., O’Sullivan, O., Jor-
dan, K., McAuliffe, O., Sangrador-Vegas, A., Slattery, L.
et al. (2008) Genome sequence of Lactobacillus helveticus,
an organism distinguished by selective gene loss and inser-
tion sequence element expansion. J Bacteriol 190, 727–735.
De Dea Lindner, J., Bernini, V., De Lorentiis, A., Pecorari, A.,
Neviani, E. and Gatti, M. (2008) Parmigiano Reggiano
cheese: evolution of cultivable and total lactic microflora
and peptidase activities during manufacture and ripening.
Dairy Sci Technol 88, 511–523.
De Man, J.C., Rogosa, M. and Sharpe, M.E. (1960) A medium
for the cultivation of lactobacilli. J Appl Bacteriol 23, 130–
135.
Decorosi, F., Vitia, C., Mengoni, A., Bazzicalupo, M. and
Giovannetti, L. (2005) Improvement of the cDNA-AFLP
method using fluorescent primers for transcription analysis
in bacteria. J Microbiol Methods 63, 211–215.
Dellagi, A., Birch, P.R., Heilbronn, J., Lyon, G.D. and Toth,
I.K. (2000) cDNA-AFLP analysis of differential gene
expression in the prokaryotic plant pathogen Erwinia
carotovora. Microbiology 146, 165–171.
Ekkapongpisit, M., Wannatung, T., Susantad, T.,
Triwitayakorn, K. and Smith, D.R. (2007) cDNA-AFLP
analysis of differential gene expression in human hepatoma
cells (HepG2) upon dengue virus infection. J Med Virol
79, 552–561.
Gatti, M., De Dea Lindner, J., De Lorentiis, A., Bottari, B.,
Santarelli, M., Bernini, V. and Neviani, E. (2008)
Dynamics of whole and lysed bacterial cells during
Parmigiano-Reggiano cheese production and ripening.
Appl Environ Microbiol 74, 6161–6167.
van de Guchte, M., Penaud, S., Grimaldi, C., Barbe, V.,
Bryson, K., Nicolas, P., Robert, C., Oztas, S. et al. (2006)
The complete genome sequence of Lactobacillus bulgaricus
reveals extensive and ongoing reductive evolution. Proc
Natl Acad Sci U S A 103, 9274–9279.
He, S., Wurtzel, O., Singh, K., Froula, J.I., Yilmaz, S., Tringe,
S.G., Wang, Z., Chen, F. et al. (2010) Validation of two
ribosomal RNA removal methods for microbial metatrans-
criptomics. Nat Methods 7, 807–813.
Hols, P., Hancy, F., Fontaine, L., Grossiord, B., Prozzi, D.,
Leblond-Bourget, N., Decaris, B., Bolotin, A. et al. (2005)
New insights in the molecular biology and physiology of
Streptococcus thermophilus revealed by comparative
genomics. FEMS Microbiol Rev 29, 435–463.
Kadota, K., Araki, R., Nakai, Y. and Abe, M. (2007) GOGOT:
a method for the identification of differentially expressed
fragments from cDNA-AFLP data. Algorithms Mol Biol 2,
5.
C.G. Bove et al. cDNA-AFLP for Lactobacillus rhamnosus
ª 2011 The Authors
Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology 863

Karpinets, T.V., Greenwood, D.J., Sams, C.E. and Ammons,
J.T. (2006) RNA: protein ratio of the unicellular organism
as a characteristic of phosphorous and nitrogen stoichio-
metry and of the cellular requirement of ribosomes for
protein synthesis. BMC Biol 4, 30.
Kieronczyk, A., Skeie, S., Olsen, K. and Langsrud, T. (2001)
Metabolism of amino acids by resting cells of non-starter
lactobacilli in relation to flavour development in cheese.
Int Dairy Journal 11, 217–224.
Koskenniemi, K., Koponen, J., Kankainen, M., Savijoki, K.,
Tynkkynen, S., de Vos, W.M., Kalkkinen, N. and
Varmanen, P. (2009) Proteome analysis of Lactobacillus
rhamnosus GG using 2-D DIGE and mass spectrometry
shows differential protein production in laboratory and
industrial-type growth media. J Proteome Res 8, 4993–
5007.
Lazzi, C., Bove, C.G., Sgarbi, E., Gatti, M., La Gioia, F.,
Torriani, S. and Neviani, E. (2009) Application of AFLP
fingerprint analysis for studying the biodiversity of
Streptococcus thermophilus. J Microbiol Methods 79, 48–
54.
Levterova, V., Panaiotov, S., Brankova, N. and Tankova, K.
(2010) Typing of genetic markers involved in stress
response by fluorescent cDNA-amplified fragment length
polymorphism technique. Mol Biotechnol 45, 34–38.
Liu, S.-Q., Holland, R. and Crow, V.L. (2003) The potential of
dairy lactic acid bacteria to metabolise amino acids via
non-transaminating reactions and endogenous transamina-
tion. Int J Food Microbiol 86, 257–269.
Neidhardt, F.C. and Umbarger, H.F. (1996) Chemical compo-
sition of Escherichia coli. In Escherichia coli and Salmonella
typhimurium, Cellular and Molecular Biology, 2nd edn. ed.
Neidhart, F.C., Ingraham, J.L., Low, K.B., Magasanik, B.,
Schaechter, M. and Umbarger, H.E. pp. 13–17. Washington,
DC: ASM Press.
Neveu, C., Charvet, C., Fauvin, A., Cortet, J., Castagnone-
Sereno, P. and Cabaret, J. (2007) Identification of
levamisole resistance markers in the parasitic nematode
Haemonchus contortus using a cDNA-AFLP approach.
Parasitology 134, 1105–1110.
Neviani, E., De Dea Lindner, J., Bernini, V. and Gatti, M.
(2009) Recovery and differentiation of long ripened cheese
microflora through a new cheese-based cultural medium.
Food Microbiol 26, 240–245.
Noel, L., Thieme, F., Nennstiel, D. and Bonas, U. (2001)
cDNA-AFLP analysis unravels a genome-wide hrpG-
regulon in the plant pathogen Xanthomonas campestris pv.
vesicatoria. Mol Microbiol 41, 1271–1281.
Reijans, M., Lascaris, R., Groeneger, A.O., Wittenberg, A.,
Wesselink, E., van Oeveren, J., de Wit, E., Boorsma, A.
et al. (2003) Quantitative comparison of cDNA-AFLP,
microarrays, and GeneChip expression data in
Saccharomyces cerevisiae. Genomics 82, 606–618.
Settanni, L. and Moschetti, G. (2010) Non-starter lactic acid
bacteria used to improve cheese quality and provide health
benefits. Food Microbiol 27, 691–697.
Succi, M., Tremonte, P., Reale, A., Sorrentino, E., Grazia, L.,
Pacifico, S. and Coppola, R. (2005) Bile salt and acid
tolerance of Lactobacillus rhamnosus strains isolated from
Parmigiano Reggiano cheese. FEMS Microbiol Lett 244,
129–137.
Vuylsteke, M., Peleman, J.D. and van Eijk, M.J.T. (2007)
AFLP-based transcript profiling (cDNAAFLP) for
genome-wide expression analysis. Nat Protoc, 2, 1399–
1413.
Weiberg, A., Pohler, D., Morgenstern, B. and Karlovsky, P.
(2008) Improved coverage of cDNA-AFLP by sequential
digestion of immobilized cDNA. BMC Genomics 9, 480.
Xiaohu, X., Heping, L. and Chaorong, T. (2009) A silver-
staining cDNA-AFLP protocol suitable for transcript
profiling in the latex of Hevea brasiliensis (para rubber
tree). Mol Biotechnol 42, 91–99.
Yvon, M. and Rijnen, L. (2001) Cheese flavour formation by
amino acid catabolism. Int Dairy J 11, 185–201.
cDNA-AFLP for Lactobacillus rhamnosus C.G. Bove et al.
864 Journal of Applied Microbiology 111, 855–864 ª 2011 The Society for Applied Microbiology
ª 2011 The Authors




















