
CD30/CD30 Ligand (CD153) Interaction Regulates CD4+ T Cell-Mediated Graft-versus-Host Disease
10
of February 21, 2016. This information is current as Graft-versus-Host Disease T Cell-Mediated + Regulates CD4 CD30/CD30 Ligand (CD153) Interaction Podack and Patricia A. Taylor Roskos, Jonathan S. Serody, Hideo Yagita, Eckhard R. Panoskaltsis-Mortari, Hiromi Muta, Monica Jones, Melinda Bruce R. Blazar, Robert B. Levy, Tak W. Mak, Angela http://www.jimmunol.org/content/173/5/2933 doi: 10.4049/jimmunol.173.5.2933 2004; 173:2933-2941; ; J Immunol References http://www.jimmunol.org/content/173/5/2933.full#ref-list-1 , 21 of which you can access for free at: cites 33 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2004 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on February 21, 2016 http://www.jimmunol.org/ Downloaded from by guest on February 21, 2016 http://www.jimmunol.org/ Downloaded from
Transcript of CD30/CD30 Ligand (CD153) Interaction Regulates CD4+ T Cell-Mediated Graft-versus-Host Disease

of February 21, 2016.This information is current as
Graft-versus-Host Disease T Cell-Mediated+Regulates CD4
CD30/CD30 Ligand (CD153) Interaction
Podack and Patricia A. TaylorRoskos, Jonathan S. Serody, Hideo Yagita, Eckhard R.Panoskaltsis-Mortari, Hiromi Muta, Monica Jones, Melinda Bruce R. Blazar, Robert B. Levy, Tak W. Mak, Angela
http://www.jimmunol.org/content/173/5/2933doi: 10.4049/jimmunol.173.5.2933
2004; 173:2933-2941; ;J Immunol
Referenceshttp://www.jimmunol.org/content/173/5/2933.full#ref-list-1
, 21 of which you can access for free at: cites 33 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 21, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

CD30/CD30 Ligand (CD153) Interaction Regulates CD4� TCell-Mediated Graft-versus-Host Disease1
Bruce R. Blazar,2,3* Robert B. Levy,2† Tak W. Mak, ‡ Angela Panoskaltsis-Mortari,*Hiromi Muta, 4† Monica Jones,† Melinda Roskos,† Jonathan S. Serody,§ Hideo Yagita,¶
Eckhard R. Podack,† and Patricia A. Taylor*
CD30, a TNFR family member, is expressed on activated CD4� and CD8� T cells and B cells and is a marker of Hodgkin’slymphoma; its ligand, CD30L (CD153) is expressed by activated CD4� and CD8� T cells, B cells, and macrophages. Signaling viaCD30 can lead to proliferation or cell death. CD30-deficient (�/�) mice have impaired thymic negative selection and increasedautoreactivity. Although human alloreactive T cells preferentially reside within the CD30� T cell subset, implicating CD30 as aregulator of T cell immune responses, the role of CD30/CD153 in regulating graft-vs-host disease (GVHD) has not been reported.We used a neutralizing anti-CD153 mAb, CD30�/� donor mice, and generated CD153�/� recipient mice to analyze the effect ofCD30/CD153 interaction on GVHD induction. Our data indicate that the CD30/CD153 pathway is a potent regulator of CD4�,but not CD8�, T cell-mediated GVHD. Although blocking CD30/CD153 interactions in vivo did not affect alloreactive CD4� T cellproliferation or apoptosis, a substantial reduction in donor CD4� T cell migration into the gastrointestinal tract was readilyobserved with lesser effects in other GVHD target organs. Blockade of the CD30/CD153 pathway represents a new approach forpreventing CD4� T cell-mediated GVHD. The Journal of Immunology, 2004, 173: 2933–2941.
U pon T cell activation, CD28 and other costimulatory re-ceptors, including CD27, CD30, CD134, CD137, andCD154, are up-regulated. CD30, a 120-kDa type I trans-
membrane glycoprotein member of the TNFR family, is expressedon Reed-Sternberg cells in Hodgkin’s disease (1), B cells, andmitogen-stimulated T cells (2, 3). The peak time for CD30 expres-sion after TCR activation is 4–5 days in vitro (2), and either CD28or IL-4R signaling are required for CD30 up-regulation (4). CD30signals augment T cell proliferation at low levels of in vitro TCRstimulation (4, 5).
Similar to other members of the TNFR family, CD30 engage-ment may regulate T cell survival. For example, CD30 signalingregulates peripheral T cell responses, controlling T cell survivaland down-regulating cytolytic capacity (2, 6–12). CD30 also reg-ulates thymocyte survival. Thymic selection appears to be influ-enced by the level of CD30 expression. In one study, CD30-defi-cient (CD30�/�) mice were reported to express a negativeselection defect (9), although selection was not affected in anotherstudy using a different CD30�/� mouse strain (13). Conversely,
CD30-overexpressing mice had increased thymocyte apoptosis af-ter TCR engagement (10).
CD30 ligand (CD30L, CD153) is a 40-kDa type II membrane-associated glycoprotein belonging to the TNF family (3, 5, 14, 15).CD153 is expressed on activated T cells, primarily CD4� T cellsof both Th1 and Th2 phenotype, as well as on a subset of accessorycells (2, 3, 14, 15) and B cells (16–19). In addition, CD153 canprovide signals for B cell growth and differentiation (3, 15).
In vivo, CD30� T cells are increased in Omenn’s syndrome,which has features similar to graft-vs-host disease (GVHD)5 thatoccurs after bone marrow transplantation (BMT) (20). These find-ings, as well as the known function of CD30 on controlling T cellresponses, survival, and cytolytic effector cell function, promptedus to examine the role of the CD30/CD153 pathway in GVHD.Results obtained using a blocking anti-CD153 mAb, CD30�/�,and newly generated CD153�/� mice in well-characterized exper-imental models of allogeneic BMT indicate that the CD30/CD153pathway is a critical regulator of CD4� but not CD8� Tcell-mediated GVHD.
Materials and MethodsMice
BALB/c (H-2d), B6.C-H2bm12/KhEg (termed bm12), and B6.C-H2bm1/KhEg(termed bm1) recipients werepurchased from The Jackson Laboratory (BarHarbor, ME). C57BL/6 (termed B6) mice were purchased from the Na-tional Institutes of Health (Bethesda, MD). CD30�/� mice were generatedas described (9) and further backcrossed seven generations with B6. B6CD153�/� mice were generated as described below. Enhanced GFP(eGFP) transgenic mice, on the B6 background, were obtained from thelaboratory of J.S.S., and bred at the University of Minnesota (Minneapolis,MN) (21). Donors and recipients were 8–12 wk of age.
*Cancer Center and Department of Pediatrics, Division of Bone Marrow Transplan-tation, University of Minnesota, Minneapolis, MN 55455;†University of Miami, Mi-ami, FL 33101;‡Ontario Cancer Institute, Toronto, Ontario, Canada;§LinebergerComprehensive Cancer Center, School of Medicine, University of North Carolina,Chapel Hill, NC 27599; and¶Department of Immunology, School of Medicine, Jun-tendo University, Tokyo, Japan
Received for publication December 2, 2003. Accepted for publication June 10, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants R01 AI 34495,2R37 HL56067, R01 HL63452, AI46689, RR11576, P01 AI35225, CA39201, andAI/DK49829.2 B.R.B. and R.B.L. contributed equally to this work.3 Address correspondence and reprint requests to Dr. Bruce R. Blazar, University ofMinnesota, 460F CCRB, 425 East River Road, Minneapolis, MN 55455. E-mail ad-dress: [email protected] Current Address: Medical Institute of Bioregulation, Kyushu University, Japan.
5 Abbreviations used in this paper: GVHD, graft-vs-host disease; 7-AAD, 7-amino-actinomycin D; BM, bone marrow; BMT, BM transplantation; CD30L, CD30 ligand;CD95L, Fas ligand; eGFP, enhanced GFP; ES, embryonic stem cell; LN, lymph node;Neo, Neomycin; TBI, total body irradiation; TCD, T cell depleted.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
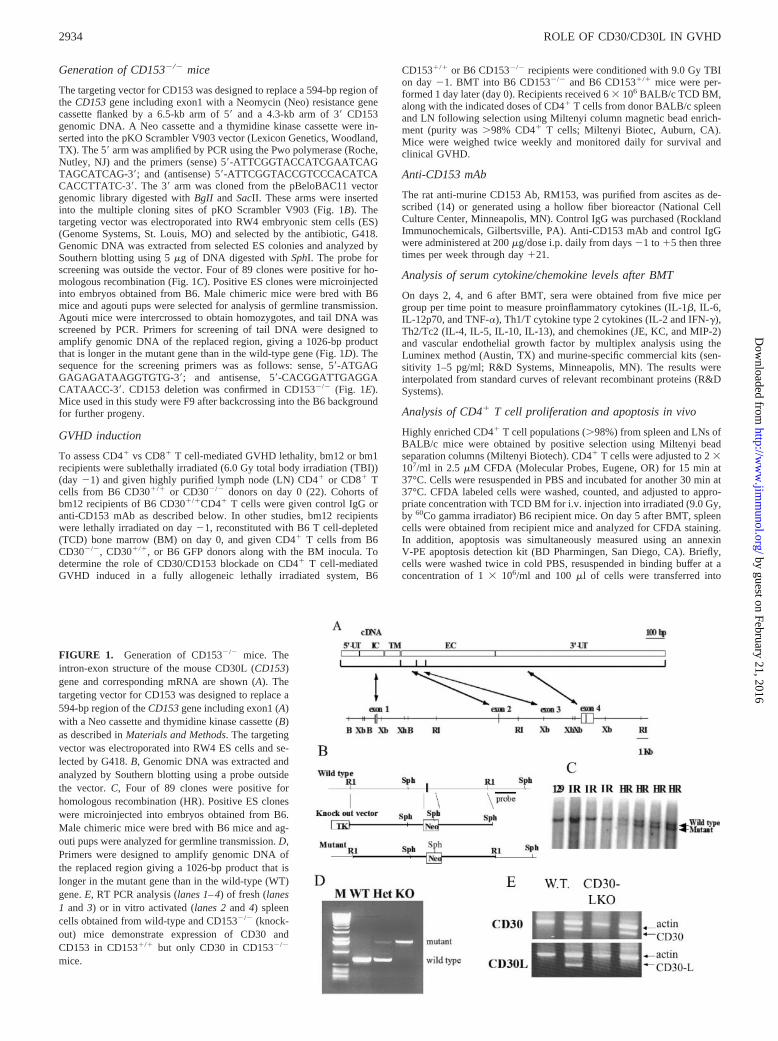
Generation of CD153�/� mice
The targeting vector for CD153 was designed to replace a 594-bp region ofthe CD153 gene including exon1 with a Neomycin (Neo) resistance genecassette flanked by a 6.5-kb arm of 5� and a 4.3-kb arm of 3� CD153genomic DNA. A Neo cassette and a thymidine kinase cassette were in-serted into the pKO Scrambler V903 vector (Lexicon Genetics, Woodland,TX). The 5� arm was amplified by PCR using the Pwo polymerase (Roche,Nutley, NJ) and the primers (sense) 5�-ATTCGGTACCATCGAATCAGTAGCATCAG-3�; and (antisense) 5�-ATTCGGTACCGTCCCACATCACACCTTATC-3�. The 3� arm was cloned from the pBeloBAC11 vectorgenomic library digested with BgII and SacII. These arms were insertedinto the multiple cloning sites of pKO Scrambler V903 (Fig. 1B). Thetargeting vector was electroporated into RW4 embryonic stem cells (ES)(Genome Systems, St. Louis, MO) and selected by the antibiotic, G418.Genomic DNA was extracted from selected ES colonies and analyzed bySouthern blotting using 5 �g of DNA digested with SphI. The probe forscreening was outside the vector. Four of 89 clones were positive for ho-mologous recombination (Fig. 1C). Positive ES clones were microinjectedinto embryos obtained from B6. Male chimeric mice were bred with B6mice and agouti pups were selected for analysis of germline transmission.Agouti mice were intercrossed to obtain homozygotes, and tail DNA wasscreened by PCR. Primers for screening of tail DNA were designed toamplify genomic DNA of the replaced region, giving a 1026-bp productthat is longer in the mutant gene than in the wild-type gene (Fig. 1D). Thesequence for the screening primers was as follows: sense, 5�-ATGAGGAGAGATAAGGTGTG-3�; and antisense, 5�-CACGGATTGAGGACATAACC-3�. CD153 deletion was confirmed in CD153�/� (Fig. 1E).Mice used in this study were F9 after backcrossing into the B6 backgroundfor further progeny.
GVHD induction
To assess CD4� vs CD8� T cell-mediated GVHD lethality, bm12 or bm1recipients were sublethally irradiated (6.0 Gy total body irradiation (TBI))(day �1) and given highly purified lymph node (LN) CD4� or CD8� Tcells from B6 CD30�/� or CD30�/� donors on day 0 (22). Cohorts ofbm12 recipients of B6 CD30�/�CD4� T cells were given control IgG oranti-CD153 mAb as described below. In other studies, bm12 recipientswere lethally irradiated on day �1, reconstituted with B6 T cell-depleted(TCD) bone marrow (BM) on day 0, and given CD4� T cells from B6CD30�/�, CD30�/�, or B6 GFP donors along with the BM inocula. Todetermine the role of CD30/CD153 blockade on CD4� T cell-mediatedGVHD induced in a fully allogeneic lethally irradiated system, B6
CD153�/� or B6 CD153�/� recipients were conditioned with 9.0 Gy TBIon day �1. BMT into B6 CD153�/� and B6 CD153�/� mice were per-formed 1 day later (day 0). Recipients received 6 � 106 BALB/c TCD BM,along with the indicated doses of CD4� T cells from donor BALB/c spleenand LN following selection using Miltenyi column magnetic bead enrich-ment (purity was �98% CD4� T cells; Miltenyi Biotec, Auburn, CA).Mice were weighed twice weekly and monitored daily for survival andclinical GVHD.
Anti-CD153 mAb
The rat anti-murine CD153 Ab, RM153, was purified from ascites as de-scribed (14) or generated using a hollow fiber bioreactor (National CellCulture Center, Minneapolis, MN). Control IgG was purchased (RocklandImmunochemicals, Gilbertsville, PA). Anti-CD153 mAb and control IgGwere administered at 200 �g/dose i.p. daily from days �1 to �5 then threetimes per week through day �21.
Analysis of serum cytokine/chemokine levels after BMT
On days 2, 4, and 6 after BMT, sera were obtained from five mice pergroup per time point to measure proinflammatory cytokines (IL-1�, IL-6,IL-12p70, and TNF-�), Th1/T cytokine type 2 cytokines (IL-2 and IFN-�),Th2/Tc2 (IL-4, IL-5, IL-10, IL-13), and chemokines (JE, KC, and MIP-2)and vascular endothelial growth factor by multiplex analysis using theLuminex method (Austin, TX) and murine-specific commercial kits (sen-sitivity 1–5 pg/ml; R&D Systems, Minneapolis, MN). The results wereinterpolated from standard curves of relevant recombinant proteins (R&DSystems).
Analysis of CD4� T cell proliferation and apoptosis in vivo
Highly enriched CD4� T cell populations (�98%) from spleen and LNs ofBALB/c mice were obtained by positive selection using Miltenyi beadseparation columns (Miltenyi Biotech). CD4� T cells were adjusted to 2 �107/ml in 2.5 �M CFDA (Molecular Probes, Eugene, OR) for 15 min at37°C. Cells were resuspended in PBS and incubated for another 30 min at37°C. CFDA labeled cells were washed, counted, and adjusted to appro-priate concentration with TCD BM for i.v. injection into irradiated (9.0 Gy,by 60Co gamma irradiator) B6 recipient mice. On day 5 after BMT, spleencells were obtained from recipient mice and analyzed for CFDA staining.In addition, apoptosis was simultaneously measured using an annexinV-PE apoptosis detection kit (BD Pharmingen, San Diego, CA). Briefly,cells were washed twice in cold PBS, resuspended in binding buffer at aconcentration of 1 � 106/ml and 100 �l of cells were transferred into
FIGURE 1. Generation of CD153�/� mice. Theintron-exon structure of the mouse CD30L (CD153)gene and corresponding mRNA are shown (A). Thetargeting vector for CD153 was designed to replace a594-bp region of the CD153 gene including exon1 (A)with a Neo cassette and thymidine kinase cassette (B)as described in Materials and Methods. The targetingvector was electroporated into RW4 ES cells and se-lected by G418. B, Genomic DNA was extracted andanalyzed by Southern blotting using a probe outsidethe vector. C, Four of 89 clones were positive forhomologous recombination (HR). Positive ES cloneswere microinjected into embryos obtained from B6.Male chimeric mice were bred with B6 mice and ag-outi pups were analyzed for germline transmission. D,Primers were designed to amplify genomic DNA ofthe replaced region giving a 1026-bp product that islonger in the mutant gene than in the wild-type (WT)gene. E, RT PCR analysis (lanes 1–4) of fresh (lanes1 and 3) or in vitro activated (lanes 2 and 4) spleencells obtained from wild-type and CD153�/� (knock-out) mice demonstrate expression of CD30 andCD153 in CD153�/� but only CD30 in CD153�/�
mice.
2934 ROLE OF CD30/CD30L IN GVHD
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
staining tubes and stained with 5 �l of annexin V-PE and 5 �l of 7-ami-noactinomycin D (7-AAD; BD Pharmingen). Cells were incubated at roomtemperature for 15 min in the dark, 400 �l of binding buffer was added toeach tube, and cells were analyzed by flow cytometry within 1 h. Unstainedand single-stained (i.e., annexin V-PE only and 7-AAD) cells were used toset compensation and quadrants for analysis.
In vivo imaging
As reported previously, images were taken with a Magnafire color camera(Optronics, Goleta, CA) mounted onto a Leica MZFLIII stereomicroscopeusing a GFP2-bandpass filter and a �0.63 transfer lens (Leica Microsys-tems, Bannockburn, IL) (23). Zoom factors from �3.5–10 were used. Ex-posure times were optimized for GVHD control mice for each organ andidentical times were used for all other groups. Mice receiving allogeneicBM only (non-GFP) served as negative controls for background autofluo-rescence in the green channel, and as demonstrated in a previous publica-tion, resulted in black images and are not shown in this publication to savespace (23). Instead, images of syngeneic controls consisting of irradiatedB6 mice transplanted with B6 BM and B6 GFP� T cells are shown todistinguish homeostatic expansion from the alloantigen-induced prolifera-tion in the GVHD control mice. Three mice per group were examined at 1wk and 2 wk after BMT. A representative image is illustrated.
Statistical analysis
The Kaplan-Meier product-limit method was used to calculate survivalrates. Differences between groups were determined using the log-rank testand were considered significant for p values � 0.05.
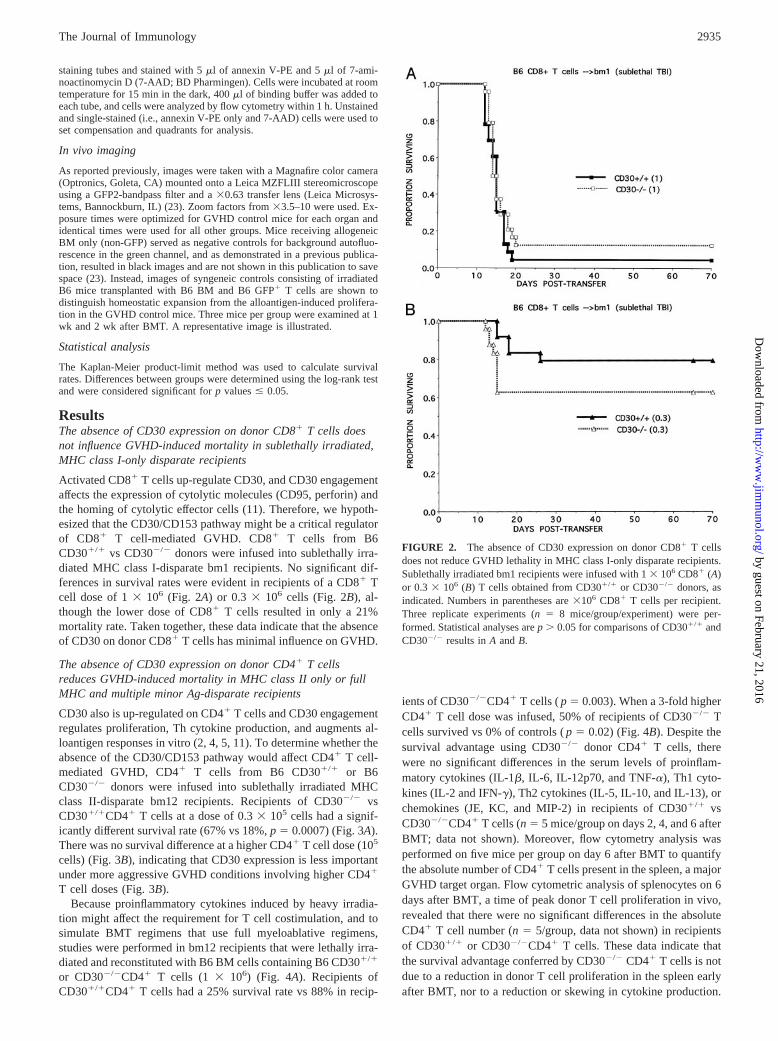
ResultsThe absence of CD30 expression on donor CD8� T cells doesnot influence GVHD-induced mortality in sublethally irradiated,MHC class I-only disparate recipients
Activated CD8� T cells up-regulate CD30, and CD30 engagementaffects the expression of cytolytic molecules (CD95, perforin) andthe homing of cytolytic effector cells (11). Therefore, we hypoth-esized that the CD30/CD153 pathway might be a critical regulatorof CD8� T cell-mediated GVHD. CD8� T cells from B6CD30�/� vs CD30�/� donors were infused into sublethally irra-diated MHC class I-disparate bm1 recipients. No significant dif-ferences in survival rates were evident in recipients of a CD8� Tcell dose of 1 � 106 (Fig. 2A) or 0.3 � 106 cells (Fig. 2B), al-though the lower dose of CD8� T cells resulted in only a 21%mortality rate. Taken together, these data indicate that the absenceof CD30 on donor CD8� T cells has minimal influence on GVHD.
The absence of CD30 expression on donor CD4� T cellsreduces GVHD-induced mortality in MHC class II only or fullMHC and multiple minor Ag-disparate recipients
CD30 also is up-regulated on CD4� T cells and CD30 engagementregulates proliferation, Th cytokine production, and augments al-loantigen responses in vitro (2, 4, 5, 11). To determine whether theabsence of the CD30/CD153 pathway would affect CD4� T cell-mediated GVHD, CD4� T cells from B6 CD30�/� or B6CD30�/� donors were infused into sublethally irradiated MHCclass II-disparate bm12 recipients. Recipients of CD30�/� vsCD30�/�CD4� T cells at a dose of 0.3 � 105 cells had a signif-icantly different survival rate (67% vs 18%, p � 0.0007) (Fig. 3A).There was no survival difference at a higher CD4� T cell dose (105
cells) (Fig. 3B), indicating that CD30 expression is less importantunder more aggressive GVHD conditions involving higher CD4�
T cell doses (Fig. 3B).Because proinflammatory cytokines induced by heavy irradia-
tion might affect the requirement for T cell costimulation, and tosimulate BMT regimens that use full myeloablative regimens,studies were performed in bm12 recipients that were lethally irra-diated and reconstituted with B6 BM cells containing B6 CD30�/�
or CD30�/�CD4� T cells (1 � 106) (Fig. 4A). Recipients ofCD30�/�CD4� T cells had a 25% survival rate vs 88% in recip-
ients of CD30�/�CD4� T cells ( p � 0.003). When a 3-fold higherCD4� T cell dose was infused, 50% of recipients of CD30�/� Tcells survived vs 0% of controls ( p � 0.02) (Fig. 4B). Despite thesurvival advantage using CD30�/� donor CD4� T cells, therewere no significant differences in the serum levels of proinflam-matory cytokines (IL-1�, IL-6, IL-12p70, and TNF-�), Th1 cyto-kines (IL-2 and IFN-�), Th2 cytokines (IL-5, IL-10, and IL-13), orchemokines (JE, KC, and MIP-2) in recipients of CD30�/� vsCD30�/�CD4� T cells (n � 5 mice/group on days 2, 4, and 6 afterBMT; data not shown). Moreover, flow cytometry analysis wasperformed on five mice per group on day 6 after BMT to quantifythe absolute number of CD4� T cells present in the spleen, a majorGVHD target organ. Flow cytometric analysis of splenocytes on 6days after BMT, a time of peak donor T cell proliferation in vivo,revealed that there were no significant differences in the absoluteCD4� T cell number (n � 5/group, data not shown) in recipientsof CD30�/� or CD30�/�CD4� T cells. These data indicate thatthe survival advantage conferred by CD30�/� CD4� T cells is notdue to a reduction in donor T cell proliferation in the spleen earlyafter BMT, nor to a reduction or skewing in cytokine production.
FIGURE 2. The absence of CD30 expression on donor CD8� T cellsdoes not reduce GVHD lethality in MHC class I-only disparate recipients.Sublethally irradiated bm1 recipients were infused with 1 � 106 CD8� (A)or 0.3 � 106 (B) T cells obtained from CD30�/� or CD30�/� donors, asindicated. Numbers in parentheses are �106 CD8� T cells per recipient.Three replicate experiments (n � 8 mice/group/experiment) were per-formed. Statistical analyses are p � 0.05 for comparisons of CD30�/� andCD30�/� results in A and B.
2935The Journal of Immunology
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
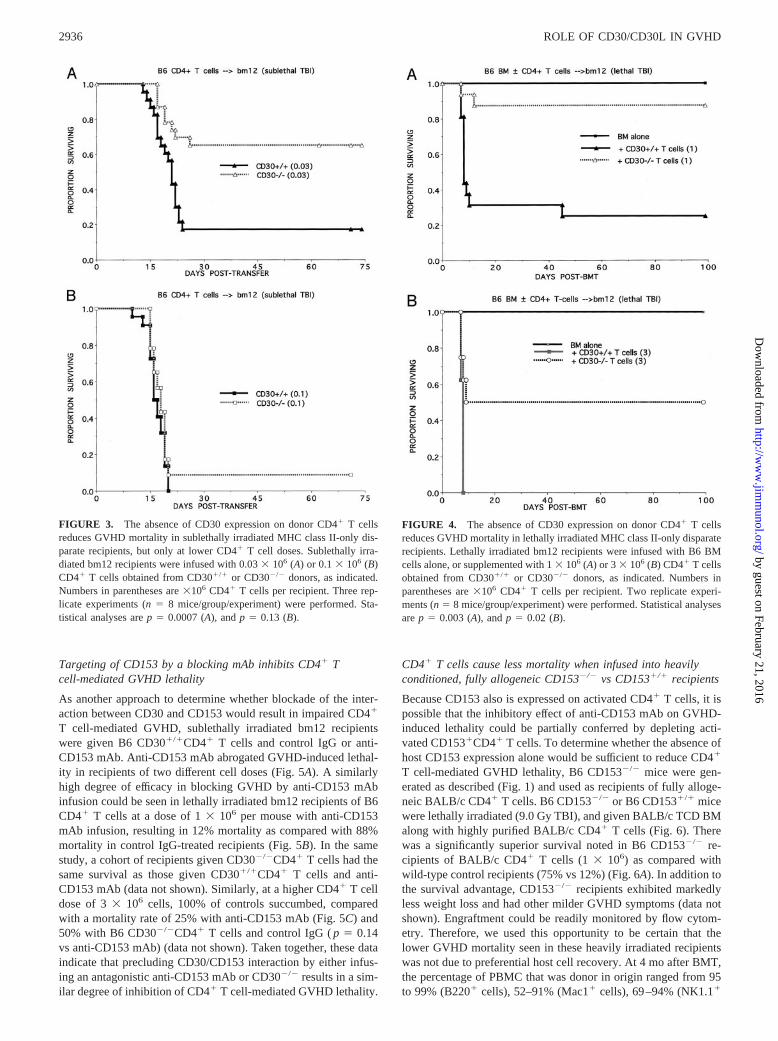
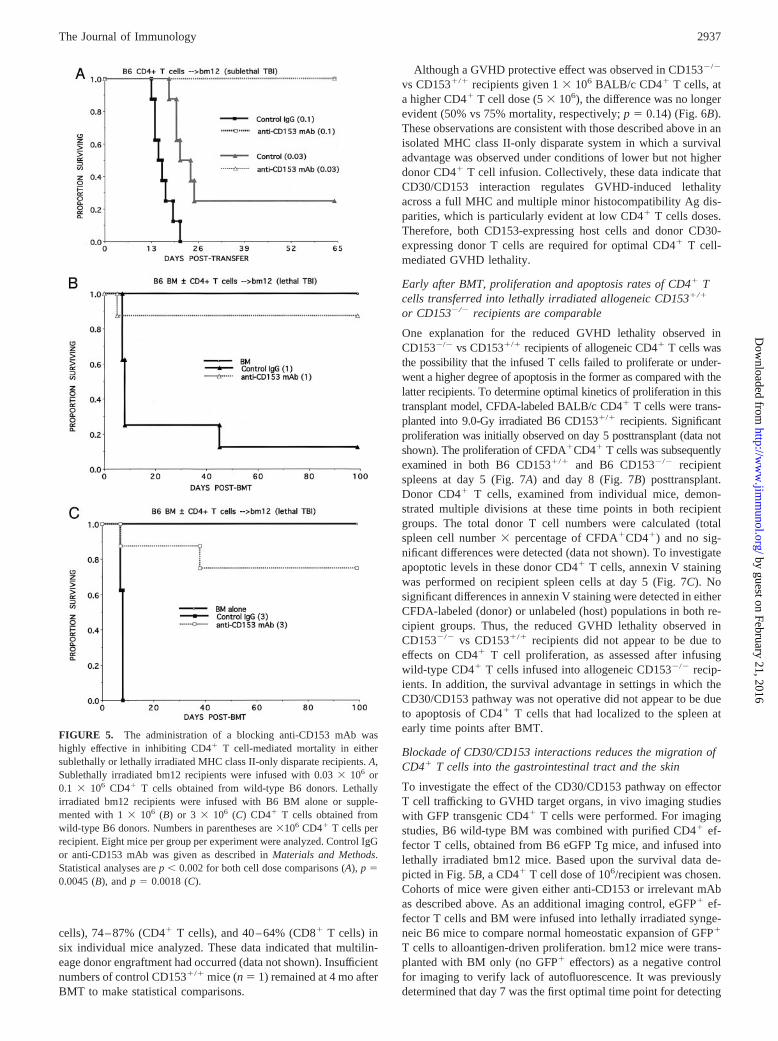
Targeting of CD153 by a blocking mAb inhibits CD4� Tcell-mediated GVHD lethality
As another approach to determine whether blockade of the inter-action between CD30 and CD153 would result in impaired CD4�
T cell-mediated GVHD, sublethally irradiated bm12 recipientswere given B6 CD30�/�CD4� T cells and control IgG or anti-CD153 mAb. Anti-CD153 mAb abrogated GVHD-induced lethal-ity in recipients of two different cell doses (Fig. 5A). A similarlyhigh degree of efficacy in blocking GVHD by anti-CD153 mAbinfusion could be seen in lethally irradiated bm12 recipients of B6CD4� T cells at a dose of 1 � 106 per mouse with anti-CD153mAb infusion, resulting in 12% mortality as compared with 88%mortality in control IgG-treated recipients (Fig. 5B). In the samestudy, a cohort of recipients given CD30�/�CD4� T cells had thesame survival as those given CD30�/�CD4� T cells and anti-CD153 mAb (data not shown). Similarly, at a higher CD4� T celldose of 3 � 106 cells, 100% of controls succumbed, comparedwith a mortality rate of 25% with anti-CD153 mAb (Fig. 5C) and50% with B6 CD30�/�CD4� T cells and control IgG ( p � 0.14vs anti-CD153 mAb) (data not shown). Taken together, these dataindicate that precluding CD30/CD153 interaction by either infus-ing an antagonistic anti-CD153 mAb or CD30�/� results in a sim-ilar degree of inhibition of CD4� T cell-mediated GVHD lethality.
CD4� T cells cause less mortality when infused into heavilyconditioned, fully allogeneic CD153�/� vs CD153�/� recipients
Because CD153 also is expressed on activated CD4� T cells, it ispossible that the inhibitory effect of anti-CD153 mAb on GVHD-induced lethality could be partially conferred by depleting acti-vated CD153�CD4� T cells. To determine whether the absence ofhost CD153 expression alone would be sufficient to reduce CD4�
T cell-mediated GVHD lethality, B6 CD153�/� mice were gen-erated as described (Fig. 1) and used as recipients of fully alloge-neic BALB/c CD4� T cells. B6 CD153�/� or B6 CD153�/� micewere lethally irradiated (9.0 Gy TBI), and given BALB/c TCD BMalong with highly purified BALB/c CD4� T cells (Fig. 6). Therewas a significantly superior survival noted in B6 CD153�/� re-cipients of BALB/c CD4� T cells (1 � 106) as compared withwild-type control recipients (75% vs 12%) (Fig. 6A). In addition tothe survival advantage, CD153�/� recipients exhibited markedlyless weight loss and had other milder GVHD symptoms (data notshown). Engraftment could be readily monitored by flow cytom-etry. Therefore, we used this opportunity to be certain that thelower GVHD mortality seen in these heavily irradiated recipientswas not due to preferential host cell recovery. At 4 mo after BMT,the percentage of PBMC that was donor in origin ranged from 95to 99% (B220� cells), 52–91% (Mac1� cells), 69–94% (NK1.1�
FIGURE 3. The absence of CD30 expression on donor CD4� T cellsreduces GVHD mortality in sublethally irradiated MHC class II-only dis-parate recipients, but only at lower CD4� T cell doses. Sublethally irra-diated bm12 recipients were infused with 0.03 � 106 (A) or 0.1 � 106 (B)CD4� T cells obtained from CD30�/� or CD30�/� donors, as indicated.Numbers in parentheses are �106 CD4� T cells per recipient. Three rep-licate experiments (n � 8 mice/group/experiment) were performed. Sta-tistical analyses are p � 0.0007 (A), and p � 0.13 (B).
FIGURE 4. The absence of CD30 expression on donor CD4� T cellsreduces GVHD mortality in lethally irradiated MHC class II-only disparaterecipients. Lethally irradiated bm12 recipients were infused with B6 BMcells alone, or supplemented with 1 � 106 (A) or 3 � 106 (B) CD4� T cellsobtained from CD30�/� or CD30�/� donors, as indicated. Numbers inparentheses are �106 CD4� T cells per recipient. Two replicate experi-ments (n � 8 mice/group/experiment) were performed. Statistical analysesare p � 0.003 (A), and p � 0.02 (B).
2936 ROLE OF CD30/CD30L IN GVHD
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
cells), 74–87% (CD4� T cells), and 40–64% (CD8� T cells) insix individual mice analyzed. These data indicated that multilin-eage donor engraftment had occurred (data not shown). Insufficientnumbers of control CD153�/� mice (n � 1) remained at 4 mo afterBMT to make statistical comparisons.
Although a GVHD protective effect was observed in CD153�/�
vs CD153�/� recipients given 1 � 106 BALB/c CD4� T cells, ata higher CD4� T cell dose (5 � 106), the difference was no longerevident (50% vs 75% mortality, respectively; p � 0.14) (Fig. 6B).These observations are consistent with those described above in anisolated MHC class II-only disparate system in which a survivaladvantage was observed under conditions of lower but not higherdonor CD4� T cell infusion. Collectively, these data indicate thatCD30/CD153 interaction regulates GVHD-induced lethalityacross a full MHC and multiple minor histocompatibility Ag dis-parities, which is particularly evident at low CD4� T cells doses.Therefore, both CD153-expressing host cells and donor CD30-expressing donor T cells are required for optimal CD4� T cell-mediated GVHD lethality.
Early after BMT, proliferation and apoptosis rates of CD4� Tcells transferred into lethally irradiated allogeneic CD153�/�
or CD153�/� recipients are comparable
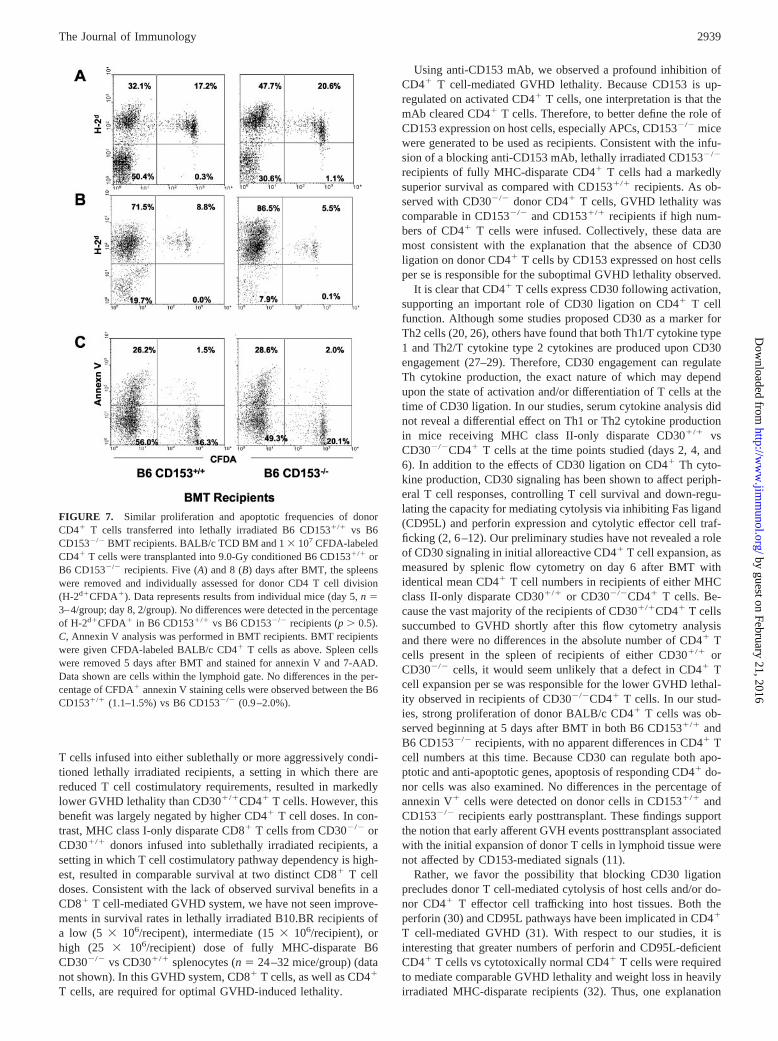
One explanation for the reduced GVHD lethality observed inCD153�/� vs CD153�/� recipients of allogeneic CD4� T cells wasthe possibility that the infused T cells failed to proliferate or under-went a higher degree of apoptosis in the former as compared with thelatter recipients. To determine optimal kinetics of proliferation in thistransplant model, CFDA-labeled BALB/c CD4� T cells were trans-planted into 9.0-Gy irradiated B6 CD153�/� recipients. Significantproliferation was initially observed on day 5 posttransplant (data notshown). The proliferation of CFDA�CD4� T cells was subsequentlyexamined in both B6 CD153�/� and B6 CD153�/� recipientspleens at day 5 (Fig. 7A) and day 8 (Fig. 7B) posttransplant.Donor CD4� T cells, examined from individual mice, demon-strated multiple divisions at these time points in both recipientgroups. The total donor T cell numbers were calculated (totalspleen cell number � percentage of CFDA�CD4�) and no sig-nificant differences were detected (data not shown). To investigateapoptotic levels in these donor CD4� T cells, annexin V stainingwas performed on recipient spleen cells at day 5 (Fig. 7C). Nosignificant differences in annexin V staining were detected in eitherCFDA-labeled (donor) or unlabeled (host) populations in both re-cipient groups. Thus, the reduced GVHD lethality observed inCD153�/� vs CD153�/� recipients did not appear to be due toeffects on CD4� T cell proliferation, as assessed after infusingwild-type CD4� T cells infused into allogeneic CD153�/� recip-ients. In addition, the survival advantage in settings in which theCD30/CD153 pathway was not operative did not appear to be dueto apoptosis of CD4� T cells that had localized to the spleen atearly time points after BMT.
Blockade of CD30/CD153 interactions reduces the migration ofCD4� T cells into the gastrointestinal tract and the skin
To investigate the effect of the CD30/CD153 pathway on effectorT cell trafficking to GVHD target organs, in vivo imaging studieswith GFP transgenic CD4� T cells were performed. For imagingstudies, B6 wild-type BM was combined with purified CD4� ef-fector T cells, obtained from B6 eGFP Tg mice, and infused intolethally irradiated bm12 mice. Based upon the survival data de-picted in Fig. 5B, a CD4� T cell dose of 106/recipient was chosen.Cohorts of mice were given either anti-CD153 or irrelevant mAbas described above. As an additional imaging control, eGFP� ef-fector T cells and BM were infused into lethally irradiated synge-neic B6 mice to compare normal homeostatic expansion of GFP�
T cells to alloantigen-driven proliferation. bm12 mice were trans-planted with BM only (no GFP� effectors) as a negative controlfor imaging to verify lack of autofluorescence. It was previouslydetermined that day 7 was the first optimal time point for detecting
FIGURE 5. The administration of a blocking anti-CD153 mAb washighly effective in inhibiting CD4� T cell-mediated mortality in eithersublethally or lethally irradiated MHC class II-only disparate recipients. A,Sublethally irradiated bm12 recipients were infused with 0.03 � 106 or0.1 � 106 CD4� T cells obtained from wild-type B6 donors. Lethallyirradiated bm12 recipients were infused with B6 BM alone or supple-mented with 1 � 106 (B) or 3 � 106 (C) CD4� T cells obtained fromwild-type B6 donors. Numbers in parentheses are �106 CD4� T cells perrecipient. Eight mice per group per experiment were analyzed. Control IgGor anti-CD153 mAb was given as described in Materials and Methods.Statistical analyses are p � 0.002 for both cell dose comparisons (A), p �0.0045 (B), and p � 0.0018 (C).
2937The Journal of Immunology
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

GFP� cells in this strain combination (P.A.T., unpublished data).By 1 wk after BMT, lymphoid organs (LNs, Peyer’s patches, andspleen) had a similar influx of GFP� cells in irrelevant and anti-CD153 mAb-treated GVHD mice, indicating that CD30/CD153did not appear to adversely affect homing into these organs (datanot shown). A representative illustration of the mesenteric LN isshown as imaged at 1 and 2 wk after BMT (Fig. 8A). A transientdecrement in GFP�CD4� T cells was observed in the skin (Fig.8B) and lung (data not shown) in anti-CD153 vs irrelevant mAb-treated recipients such that fewer GFP� cells were present in theformer at 1 wk but not 2 wk after BMT. Modest reductions inGFP� cells were seen in the liver at both time points (Fig. 8C). Themajor effects of CD30/CD153 pathway blockade were in the extentof GFP� T cell infiltration into the gastrointestinal tract, as illus-trated by images of the stomach (Fig. 8D), ileum (Fig. 8E), andcolon (Fig. 8F). There was a marked and persistent decrement inGFP�CD4� T cells that migrated to these sites in anti-CD153mAb-treated recipients. Notably, the LN adjacent to the colon im-aged at 1 wk after BMT reveals no substantial effect of anti-CD153vs irrelevant mAb. These findings suggest that the improved sur-vival in anti-CD153 mAb-treated recipients may be due to im-
paired donor CD4� T cell homing to or expansion within GVHDtarget organs, especially those of the gastrointestinal tract.
DiscussionIn this study, we demonstrate that the signaling via the CD30/CD153 pathway is required for optimal GVHD-induced lethalityby donor CD4� but not CD8� T cells. CD4� T cell-mediatedGVHD lethality was inhibited in situations in which there was anisolated MHC class II-only disparity or a full MHC plus multipleminor Ag disparities. However, the influence of CD30 ligation onGVHD-induced lethality was not evident at higher CD4� T celldoses, indicating that the requirement for CD30 engagement couldbe obviated if the vigor of GVHD induction was increased byinfusing large numbers of T cells. Collectively, these data providenew evidence that CD30 regulates alloresponses in vivo that resultin a clinically relevant disease process, GVHD.
The observation that alloreactive CD4� T cell-mediated GVHDlethality was more dependent than CD8� T cells upon CD30 li-gation was somewhat unexpected based upon the finding that al-loantigen- activation induces CD30 on CD8� and to a lesser extenton CD4� T cells (24, 25). MHC class II-disparate CD30�/�CD4�
FIGURE 6. Lethally irradiated CD153�/� recipientsof fully allogeneic donor BM and CD4� T cells have amarked reduction in GVHD-induced mortality as com-pared with CD153�/� recipients at lower but not higherCD4� T cell doses. B6 CD153�/� or B6 CD153�/� re-cipients (n � 8 mice/group) were lethally irradiated andinfused with BALB/c BM supplemented with BALB/cCD4� T cells at a dose of 1 � 106 (A) or 5 � 106 (B).Statistical analysis was p � 0.0018 (A), and p � 0.14 (B).
2938 ROLE OF CD30/CD30L IN GVHD
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
T cells infused into either sublethally or more aggressively condi-tioned lethally irradiated recipients, a setting in which there arereduced T cell costimulatory requirements, resulted in markedlylower GVHD lethality than CD30�/�CD4� T cells. However, thisbenefit was largely negated by higher CD4� T cell doses. In con-trast, MHC class I-only disparate CD8� T cells from CD30�/� orCD30�/� donors infused into sublethally irradiated recipients, asetting in which T cell costimulatory pathway dependency is high-est, resulted in comparable survival at two distinct CD8� T celldoses. Consistent with the lack of observed survival benefits in aCD8� T cell-mediated GVHD system, we have not seen improve-ments in survival rates in lethally irradiated B10.BR recipients ofa low (5 � 106/recipent), intermediate (15 � 106/recipient), orhigh (25 � 106/recipient) dose of fully MHC-disparate B6CD30�/� vs CD30�/� splenocytes (n � 24–32 mice/group) (datanot shown). In this GVHD system, CD8� T cells, as well as CD4�
T cells, are required for optimal GVHD-induced lethality.
Using anti-CD153 mAb, we observed a profound inhibition ofCD4� T cell-mediated GVHD lethality. Because CD153 is up-regulated on activated CD4� T cells, one interpretation is that themAb cleared CD4� T cells. Therefore, to better define the role ofCD153 expression on host cells, especially APCs, CD153�/� micewere generated to be used as recipients. Consistent with the infu-sion of a blocking anti-CD153 mAb, lethally irradiated CD153�/�
recipients of fully MHC-disparate CD4� T cells had a markedlysuperior survival as compared with CD153�/� recipients. As ob-served with CD30�/� donor CD4� T cells, GVHD lethality wascomparable in CD153�/� and CD153�/� recipients if high num-bers of CD4� T cells were infused. Collectively, these data aremost consistent with the explanation that the absence of CD30ligation on donor CD4� T cells by CD153 expressed on host cellsper se is responsible for the suboptimal GVHD lethality observed.
It is clear that CD4� T cells express CD30 following activation,supporting an important role of CD30 ligation on CD4� T cellfunction. Although some studies proposed CD30 as a marker forTh2 cells (20, 26), others have found that both Th1/T cytokine type1 and Th2/T cytokine type 2 cytokines are produced upon CD30engagement (27–29). Therefore, CD30 engagement can regulateTh cytokine production, the exact nature of which may dependupon the state of activation and/or differentiation of T cells at thetime of CD30 ligation. In our studies, serum cytokine analysis didnot reveal a differential effect on Th1 or Th2 cytokine productionin mice receiving MHC class II-only disparate CD30�/� vsCD30�/�CD4� T cells at the time points studied (days 2, 4, and6). In addition to the effects of CD30 ligation on CD4� Th cyto-kine production, CD30 signaling has been shown to affect periph-eral T cell responses, controlling T cell survival and down-regu-lating the capacity for mediating cytolysis via inhibiting Fas ligand(CD95L) and perforin expression and cytolytic effector cell traf-ficking (2, 6 –12). Our preliminary studies have not revealed a roleof CD30 signaling in initial alloreactive CD4� T cell expansion, asmeasured by splenic flow cytometry on day 6 after BMT withidentical mean CD4� T cell numbers in recipients of either MHCclass II-only disparate CD30�/� or CD30�/�CD4� T cells. Be-cause the vast majority of the recipients of CD30�/�CD4� T cellssuccumbed to GVHD shortly after this flow cytometry analysisand there were no differences in the absolute number of CD4� Tcells present in the spleen of recipients of either CD30�/� orCD30�/� cells, it would seem unlikely that a defect in CD4� Tcell expansion per se was responsible for the lower GVHD lethal-ity observed in recipients of CD30�/�CD4� T cells. In our stud-ies, strong proliferation of donor BALB/c CD4� T cells was ob-served beginning at 5 days after BMT in both B6 CD153�/� andB6 CD153�/� recipients, with no apparent differences in CD4� Tcell numbers at this time. Because CD30 can regulate both apo-ptotic and anti-apoptotic genes, apoptosis of responding CD4� do-nor cells was also examined. No differences in the percentage ofannexin V� cells were detected on donor cells in CD153�/� andCD153�/� recipients early posttransplant. These findings supportthe notion that early afferent GVH events posttransplant associatedwith the initial expansion of donor T cells in lymphoid tissue werenot affected by CD153-mediated signals (11).
Rather, we favor the possibility that blocking CD30 ligationprecludes donor T cell-mediated cytolysis of host cells and/or do-nor CD4� T effector cell trafficking into host tissues. Both theperforin (30) and CD95L pathways have been implicated in CD4�
T cell-mediated GVHD (31). With respect to our studies, it isinteresting that greater numbers of perforin and CD95L-deficientCD4� T cells vs cytotoxically normal CD4� T cells were requiredto mediate comparable GVHD lethality and weight loss in heavilyirradiated MHC-disparate recipients (32). Thus, one explanation
FIGURE 7. Similar proliferation and apoptotic frequencies of donorCD4� T cells transferred into lethally irradiated B6 CD153�/� vs B6CD153�/� BMT recipients. BALB/c TCD BM and 1 � 107 CFDA-labeledCD4� T cells were transplanted into 9.0-Gy conditioned B6 CD153�/� orB6 CD153�/� recipients. Five (A) and 8 (B) days after BMT, the spleenswere removed and individually assessed for donor CD4 T cell division(H-2d�CFDA�). Data represents results from individual mice (day 5, n �3–4/group; day 8, 2/group). No differences were detected in the percentageof H-2d�CFDA� in B6 CD153�/� vs B6 CD153�/� recipients (p � 0.5).C, Annexin V analysis was performed in BMT recipients. BMT recipientswere given CFDA-labeled BALB/c CD4� T cells as above. Spleen cellswere removed 5 days after BMT and stained for annexin V and 7-AAD.Data shown are cells within the lymphoid gate. No differences in the per-centage of CFDA� annexin V staining cells were observed between the B6CD153�/� (1.1–1.5%) vs B6 CD153�/� (0.9–2.0%).
2939The Journal of Immunology
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

for the lowered CD4� T cell-mediated GVHD observed whenCD30 engagement is precluded could be down-regulation of ex-pression of these molecules (11). In addition to regulation of cy-tolytic activity, CD30 signaling up-regulates CCR7 (11). Thus, analternative mechanism for a suboptimal GVHD lethality responseby CD30�/�CD4� T cells may involve effector cell traffickingresulting in inadequate homing of CD4� T cells to secondary lym-
phoid organs such as intestinal Peyer’s patches (33) or GVHDtarget organs. Although we did not observe differences in second-ary lymphoid organ homing, we did observe substantial reductionsin CD4� T cell migration in GVHD target organs, particularly thegastrointestinal tract, with lesser findings in the skin, liver, andlung. The gastrointestinal tract is a vital target tissue of GVHD andtissue injury can result in diarrhea and high levels of endotoxin
FIGURE 8. Anti-CD153 mAb infusionreduces allogeneic CD4� T cell infiltrationinto GVHD target organs, especially the gas-trointestinal tract. B6 GFP�CD4� T cells(106) were infused into lethally irradiatedMHC class II-disparate bm12 or syngeneicrecipients. Whole body imaging was per-formed on three mice per group at 1 and 2 wkafter BMT, as indicated. A representativeimage from each group is shown for mes-enteric LN (A), skin (B), liver (C), stomach(D), ileum (E), and colon (F). Anti-CD153mAb did not affect GFP�CD4� T cellinfiltration into the mesenteric LNs (A)or other LNs such as the inguinal LN orPeyer’s patches LN (data not shown). Inanti-CD153 vs irrelevant mAb-treated re-cipients, GFP�CD4� T cells were mark-edly reduced at both time points in thestomach (D), ileum (E), and colon (F), sug-gesting that the gastrointestinal tract wasthe major site of benefit for CD30/CD153blockade.
2940 ROLE OF CD30/CD30L IN GVHD
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

into the circulation. Thus, the profound reduction of GFP�CD4�
T cell infiltration into the stomach, ileum, and colon in anti-CD153mAb-treated recipients could explain the superior survival andlower weight loss seen in settings in which CD30/CD153 interac-tions are blocked.
In summary, we have shown that blockade of the CD30/CD153pathway reduces CD4� T cell-mediated GVHD. We further dem-onstrate that these findings can be observed using one of a varietyof approaches to preclude CD30/CD153 interactions including theadministration of CD30�/� donor CD4� T cells, CD30�/�CD4�
T cells together with the infusion of a blocking anti-CD153 mAb,or use of CD153�/� recipients. A marked effect of precludingCD30 ligation on CD4� T cell-mediated GVHD mortality wasobserved in either sublethally or lethally irradiated recipients, al-though the infusion of high CD4� T cell doses abrogated thissurvival advantage. The major effect of precluding CD30/CD153interactions is upon inhibiting the homing of CD4� T cells to thegastrointestinal tract, which was associated with increased survivaland lower weight loss.
Because CD30/CD153 specific reagents are in clinical trials,future studies may be considered to prevent or treat gastrointestinalGVHD by targeting the CD30/CD153 pathway.
AcknowledgmentsWe recognize the excellent technical assistance of Angela Jones for as-sessing chimerism in CD153�/� recipients and Melinda Berthold for cy-tokine determinations.
References1. Durkop, H., U. Latza, M. Hummel, F. Eitelbach, B. Seed, and H. Stein. 1992.
Molecular cloning and expression of a new member of the nerve growth factorreceptor family that is characteristic for Hodgkin’s disease. Cell 68:421.
2. Bowen, M. A., R. K. Lee, G. Miragliotta, S. Y. Nam, and E. R. Podack. 1996.Structure and expression of murine CD30 and its role in cytokine production.J. Immunol. 156:442.
3. Shanebeck, K. D., C. R. Maliszewski, M. K. Kennedy, K. S. Picha, C. A. Smith,R. G. Goodwin, and K. H. Grabstein. 1995. Regulation of murine B cell growthand differentiation by CD30 ligand. Eur. J. Immunol. 25:2147.
4. Gilfillan, M. C., P. J. Noel, E. R. Podack, S. L. Reiner, and C. B. Thompson.1998. Expression of the costimulatory receptor CD30 is regulated by both CD28and cytokines. J. Immunol. 160:2180.
5. Smith, C. A., H. J. Gruss, T. Davis, D. Anderson, T. Farrah, E. Baker,G. R. Sutherland, C. I. Brannan, N. G. Copeland, N. A. Jenkins, et al. 1993. CD30antigen, a marker for Hodgkin’s lymphoma, is a receptor whose ligand defines anemerging family of cytokines with homology to TNF. Cell 73:1349.
6. Duckett, C. S., and C. B. Thompson. 1997. CD30-dependent degradation ofTRAF2: implications for negative regulation of TRAF signaling and the controlof cell survival. Genes Dev. 11:2810.
7. Kurts, C., F. R. Carbone, M. F. Krummel, K. M. Koch, J. F. Miller, andW. R. Heath. 1999. Signalling through CD30 protects against autoimmune dia-betes mediated by CD8 T cells. Nature 398:341.
8. Telford, W. G., S. Y. Nam, E. R. Podack, and R. A. Miller. 1997. CD30-regulatedapoptosis in murine CD8 T cells after cessation of TCR signals. Cell. Immunol.182:125.
9. Amakawa, R., A. Hakem, T. M. Kundig, T. Matsuyama, J. J. Simard, E. Timms,A. Wakeham, H. W. Mittruecker, H. Griesser, H. Takimoto, et al. 1996. Impairednegative selection of T cells in Hodgkin’s disease antigen CD30-deficient mice.Cell 84:551.
10. Chiarle, R., A. Podda, G. Prolla, E. R. Podack, G. J. Thorbecke, and G. Inghirami.1999. CD30 overexpression enhances negative selection in the thymus and me-diates programmed cell death via a Bcl-2-sensitive pathway. J. Immunol.163:194.
11. Muta, H., L. H. Boise, L. Fang, and E. R. Podack. 2000. CD30 signals integrateexpression of cytotoxic effector molecules, lymphocyte trafficking signals, andsignals for proliferation and apoptosis. J. Immunol. 165:5105.
12. Grell, M., G. Zimmermann, E. Gottfried, C. M. Chen, U. Grunwald, D. C. Huang,Y. H. Wu Lee, H. Durkop, H. Engelmann, P. Scheurich, et al. 1999. Induction of
cell death by tumour necrosis factor (TNF) receptor 2, CD40 and CD30: a role forTNF-R1 activation by endogenous membrane-anchored TNF. EMBO J. 18:3034.
13. DeYoung, A. L., O. Duramad, and A. Winoto. 2000. The TNF receptor familymember CD30 is not essential for negative selection. J. Immunol. 165:6170.
14. Shimozato, O., K. Takeda, H. Yagita, and K. Okumura. 1999. Expression ofCD30 ligand (CD153) on murine activated T cells. Biochem. Biophys. Res. Com-mun. 256:519.
15. Wiley, S. R., R. G. Goodwin, and C. A. Smith. 1996. Reverse signaling via CD30ligand. J. Immunol. 157:3635.
16. Cerutti, A., A. Schaffer, R. G. Goodwin, S. Shah, H. Zan, S. Ely, and P. Casali.2000. Engagement of CD153 (CD30 ligand) by CD30� T cells inhibits classswitch DNA recombination and antibody production in human IgD� IgM� Bcells. J. Immunol. 165:786.
17. Younes, A., U. Consoli, V. Snell, K. Clodi, K. O. Kliche, J. L. Palmer,H. J. Gruss, R. Armitage, E. K. Thomas, F. Cabanillas, and M. Andreeff. 1997.CD30 ligand in lymphoma patients with CD30� tumors. J. Clin. Oncol. 15:3355.
18. Gattei, V., M. Degan, A. Gloghini, A. De Iuliis, S. Improta, F. M. Rossi,D. Aldinucci, V. Perin, D. Serraino, R. Babare, et al. 1997. CD30 ligand isfrequently expressed in human hematopoietic malignancies of myeloid and lym-phoid origin. Blood 89:2048.
19. Cerutti, A., L. Trentin, R. Zambello, R. Sancetta, A. Milani, C. Tassinari,F. Adami, C. Agostini, and G. Semenzato. 1996. The CD5/CD72 receptor systemis coexpressed with several functionally relevant counterstructures on human Bcells and delivers a critical signaling activity. J. Immunol. 157:1854.
20. Del Prete, G., M. De Carli, F. Almerigogna, C. K. Daniel, M. M. D’Elios,G. Zancuoghi, F. Vinante, G. Pizzolo, and S. Romagnani. 1995. Preferentialexpression of CD30 by human CD4� T cells producing Th2-type cytokines.FASEB J. 9:81.
21. Serody, J. S., S. E. Burkett, A. Panoskaltsis-Mortari, J. Ng-Cashin, E. McMahon,G. K. Matsushima, S. A. Lira, D. N. Cook, and B. R. Blazar. 2000. T-lymphocyteproduction of macrophage inflammatory protein-1� is critical to the recruitmentof CD8� T cells to the liver, lung, and spleen during graft-versus-host disease.Blood 96:2973.
22. Blazar, B. R., P. A. Taylor, A. Panoskaltsis-Mortari, A. H. Sharpe, andD. A. Vallera. 1999. Opposing roles of CD28:B7 and CTLA-4:B7 pathways inregulating in vivo alloresponses in murine recipients of MHC disparate T cells.J. Immunol. 162:6368.
23. Panoskaltsis-Mortari, A., A. Price, J. R. Hermanson, E. Taras, C. Lees,J. S. Serody, and B. R. Blazar. 2004. In vivo imaging of graft-versus-host-diseasein mice. Blood 103:3590.
24. Martinez, O. M., J. Villanueva, S. Abtahi, P. R. Beatty, C. O. Esquivel, andS. M. Krams. 1998. CD30 expression identifies a functional alloreactive humanT-lymphocyte subset. Transplantation 65:1240.
25. Chan, K. W., C. D. Hopke, S. M. Krams, and O. M. Martinez. 2002. CD30expression identifies the predominant proliferating T lymphocyte population inhuman alloimmune responses. J. Immunol. 169:1784.
26. Kim, M. Y., F. M. Gaspal, H. E. Wiggett, F. M. McConnell,A. Gulbranson-Judge, C. Raykundalia, L. S. Walker, M. D. Goodall, andP. J. Lane. 2003. CD4�CD3� accessory cells costimulate primed CD4 T cellsthrough OX40 and CD30 at sites where T cells collaborate with B cells. Immunity18:643.
27. Alzona, M., H. M. Jack, R. I. Fisher, and T. M. Ellis. 1994. CD30 defines a subsetof activated human T cells that produce IFN-� and IL-5 and exhibit enhanced Bcell helper activity. J. Immunol. 153:2861.
28. Nakamura, T., R. K. Lee, S. Y. Nam, B. K. Al-Ramadi, P. A. Koni, K. Bottomly,E. R. Podack, and R. A. Flavell. 1997. Reciprocal regulation of CD30 expressionon CD4� T cells by IL-4 and IFN-�. J. Immunol. 158:2090.
29. Harlin, H., E. Podack, M. Boothby, and M. L. Alegre. 2002. TCR-independentCD30 signaling selectively induces IL-13 production via a TNF receptor-asso-ciated factor/p38 mitogen-activated protein kinase-dependent mechanism. J. Im-munol. 169:2451.
30. Blazar, B. R., P. A. Taylor, and D. A. Vallera. 1997. CD4� and CD8� T cellseach can utilize a perforin-dependent pathway to mediate lethal graft-versus-hostdisease in major histocompatibility complex-disparate recipients. Transplanta-tion 64:571.
31. Graubert, T. A., J. F. DiPersio, J. H. Russell, and T. J. Ley. 1997. Perforin/granzyme-dependent and independent mechanisms are both important for thedevelopment of graft-versus-host disease after murine bone marrow transplanta-tion. J. Clin. Invest. 100:904.
32. Jiang, Z., E. Podack, and R. B. Levy. 2001. Major histocompatibility complex-mismatched allogeneic bone marrow transplantation using perforin and/or Fasligand double-defective CD4� donor T cells: involvement of cytotoxic functionby donor lymphocytes prior to graft-versus-host disease pathogenesis. Blood98:390.
33. Murai, M., H. Yoneyama, T. Ezaki, M. Suematsu, Y. Terashima, A. Harada,H. Hamada, H. Asakura, H. Ishikawa, and K. Matsushima. 2003. Peyer’s patchis the essential site in initiating murine acute and lethal graft-versus-host reaction.Nat. Immunol. 4:154.
2941The Journal of Immunology
by guest on February 21, 2016http://w
ww
.jimm
unol.org/D
ownloaded from




















