
Castoroides in Ohio
10
UNLOCKING THE UNKNOWN PAPERS HONORING DR. RICHARD]. ZAKRZEWSKI edited by Greg H. Farley and Jerry R. Choate layout and design by Jennifer Higerd FORT HAYS STATE UNIVERSITY, HAYS, KANSAS
Transcript of Castoroides in Ohio

UNLOCKING THE UNKNOWNPAPERS HONORING DR. RICHARD]. ZAKRZEWSKI
edited by Greg H. Farley and Jerry R. Choatelayout and design by Jennifer HigerdFORT HAYS STATE UNIVERSITY, HAYS, KANSAS

-~
STERNBERG MUSEUM I MUSEUM OF THE HIGH PLAINS
ABSTRACTNew radiocarbon dates of the extinct giant beaver, Castoro ides ohioensis, from Clear Creek near Amanda, FairfieldCounty, and the Sheriden Pit at Indian Trail Caverns, Wyandott County, Ohio add to our knowledge of the timing andpattern of the extinction of this Pleistocene species. While it is still not possible to determine whether the primary causeof Castoro ides extinction was the result of environmental change or by the appearance of humans, an examination oflong-term changes in the distribution of the Castoroides lineage indicates a consistent decrease in its range from theHemphillian to the late Pleistocene. Any explanation for the disappearance of Castoro ides needs to include this long-term perspective rather then simply consider the record immediately prior to its disappearance.
INTRODUCTIONThe Pleistocene giant beaver, Castoroides
ohioensis, was among the more spectacular membersof the late Pleistocene fauna of North America. Sincethe first specimen was recovered from a peat bog nearNashport, Muskingum County, Ohio (Foster, 1838)our knowledge has greatly improved with its discov-ery from localities ranging from the Yukon to Florida.While 30 faunas with Castoroides were recorded bythe FAUNMAP Working Group (1994), the addition ofisolated records brings the number of records to over131 for the Late Rancholabrean (less then 150,000years B.P.) (McDonald in prep.). While the majority ofthe localities are found east of the Mississippi River, thegenus is also known from Irvingtonian and early Ran-cholabrean sites in the Great Plains and from the OldCrow Basin in the Yukon. Despite the relative abun-dance of specimens in the late Pleistocene, very fewhave good associated radiocarbon dates or have been
directly dated. The lack of this basic information haslimited our ability to determine the timing of the spe-cies extinction and to examine any patterns in changesin its distribution prior to its extinction that may allowus to infer whether its disappearance was environmen-tally driven or the result of the appearance of humans.We report here the recovery of new specimens of Cas-toroides ohioensis from Ohio that have been directlyradiocarbon dated and examine these new dates withinthe context of the few previously published radiocarbondates for the genus and the long term history of thelineage in order to provide a preliminary assessment ofthe extinction of Castoroides.
ABBREVIATIONSAll measurements of specimens are in mil-
limeters (mm). Institutions housing the specimensreferred to in the paper are identified by the followingacronyms: CMNH - Cleveland Museum of Natural
51

History; CinMNH - Cincinnati Museum of NaturalHistory; OHS - Ohio Historical Society; OSM - OhioState University Zoological Museum. P = premolar,M = molar with upper dentition indicated by an uppercase letter and lower dentition indicated by a lower caseletter. Ma - million years. B.P. - Before Present.
DISCOVERY AND LOCATIONS OF NEW SPECI-MENS OF CASTOROIDESClear Creek, Fairfield County
On 25 May 2003, Jeff Snyder Jr. of Amanda,Ohio discovered a new fossil of Castoro ides ohioen-sis on a gravel bar, mid-stream on Clear Creek. ClearCreek is a glacially reversed stream, with the reversalgorge located near the Fairfield/Hocking County line(Forsyth, 1962), about 17 kilometers downstream. Thesite where the specimen was found is roughly 1400 me-ters from the town-center of Amanda, Ohio. The gravelbar was approximately 300 to 400 meters upstream' ofAmanda-Southern Road, within Section 2, Clear CreekTownship, Fairfield County. Previous and subsequentheavy rains prevented fruitful search of the site andno other bones were found. The mandible probablywas eroded by these rains from the bank somewhereupstream. A nearly 90 degree bend lies 900 metersupstream from the discovery site. Determination of thelocation from maps places the site at about 39.64° Nand 82.76° W. Mr. Snyder later donated the mandibleto the natural history collections of the Ohio HistoricalSociety, where it has been designated as N 9109.The stream valley is listed as "Recent alluvian" byForsyth (1962). The area was mapped by Forsyth(1962) as alluvian and appears to match exactly withthe Algiers Silt Loam. The soils surrounding ClearCreek at the location of the find are entirely Algiers SiltLoam and these soils surround both sides ofthe streamfor at least 2.6 kilometers upstream, at a width varyingfrom 150 to 350 meters (measured from map in Meekeret al., 1960). These soils are described as being poorlydrained alluvial bottom lands with slopes typically at 0- 2% with a maximum of 6%. The surface soil is typi-cally 8 em thick, medium fine texture, and derived fromcalcareous Wisconsin glacial till. The lower layers aredarker and finer textured and neutral in pH. Both layersare medium to high in organic matter. Except wheredrain tiles have been installed, these soils typically arewater-covered part of the year, with runoff being slow.The surface and subsoil combined range up to 16.5 erndeep (Meeker et aI., 1960).Sheriden Pit, Wyandot County
The Sheriden Pit is part of a sinkhole-cavecomplex associated with Indian Trail Caverns, a com-mercial cave located 6.4 km west of Carey, WyandotCounty, Ohio on the south side of Ohio highway 568
52
(40.98° N, 83.44 0 W). The caverns are located inthe crest of a bedrock ridge formed from a reef in theMiddle Silurian Greenfield Formation. The ridge has anorthwest-southeast orientation and rises about 15 me-ters above the surrounding till plain. The cave is about3.2 km south of the Defiance Moraine, dated at 14,800years B.P. (Mickelson et al., 1983) which provides amaximum age of the fauna since the opening of thesinkhole and accumulation of vertebrate remains wouldhave postdated deglaciation of the region.
The sinkhole and associated cave passage ofthe Sheriden Pit were completely filled with sedimentsprior to excavation. Two distinct sediment types arepresent in the horizontal cave passage. The upper sedi-ments consist of a 45 em thick layer of finely laminatedsilts, deposited in standing water. Below this upperlayer are coarsely stratified layers of silt and clay mixedwith cobbles and boulders of dolomite derived from thesurrounding bedrock and smaller cobbles and gravelderived from reworked glacial deposits. It is this lowerlayer that contains the majority of the faunal remainsand from which all extinct megafauna including the
-----5cm
A
• • •FIGURE 1. A. Right lateral view of Clear Creek mandible (OHS N9109). B. Left lateral view of Sheri den Pit mandible (CinMNH VP1689) and left lower incisor. Scale is 5 cm.

Castoro ides have been recovered. Most of the uppersediments were removed during the initial excavationof the pit and cave passage by the commercial caveoperation prior to the recognition of the presence ofbones. The complete thickness of the remaining bonebearing sediments has yet to be determined but exceeds3 meters. The initial discoveries of Castoro ides wererecovered from disturbed sediments but additional ma-terial was recovered in situ. Following discovery of thePleistocene fauna initial excavations in the cave wereconducted by field crews of the Cincinnati Museum ofNatural History. These specimens are deposited in theVertebrate Paleontology Department of the CincinnatiMuseum of Natural History. Subsequent excavations atthe cave by crews from Kent State University lead byKen Tankersley recovered an additional specimen of thegiant beaver which is housed at the Cleveland Museumof Natural History.
SPECIMEN DESCRIPTIONSThe Clear Creek specimen is a partial right
mandible (OHS N 9109) (Fig. 1a). The incisor isalmost complete, lacking only a small chip at the distaltip, a section of enamel covering also near the tip, andanother small piece of enamel almost midway on thebuccal side of the incisor. The m 1 is present, and thealveoli for the p4, m2, and m3 are intact though theseteeth are missing. The bone along the ventral-buccaledge of the mandible has been broken and/or erodedaway along with part of the mandibular condyle and theentire angular process (Driesch, 1976) (gonial angle,Gilbert, 1990). This leaves approximately 88 milli-meters of the basal portion of the incisor root exposed.Measurements are provided in table I.
Prior to acquisition of the mandible by theOhio Historical Society, excessive drying caused sev-eral fragments to loosen. One fragment weighing 1.2grams was not able to be exactly re-associated duringconservation work. This fragment was carbon-dated byStafford Research Laboratories and a date of 12,040 ±35 years B.P. (UCIAMS-11219) was obtained.
The buccal side ofthe mandible between themandibular condyle and the angular process (Fig. 1) hasat least 6 partial tooth marks that cover an area about6.1 millimeters long. A second area includes 6 or 7 par-tial tooth marks covering about 8.0 mm of space, and athird area includes at least 5 partial tooth marks (partlyeroded away) that cover an area of about 8.3 mm. Therodent tooth marks each vary from 0.8 to 1.4 mm wide.
While multiple bones of Castoroides wererecovered from the Sheriden Pit, there is no duplicationof elements and it appears that only a single individualis represented. This is supported by their sizes andstages of epiphyseal fusion. The Castoroides material
housed at the Cincinnati Museum of Natural History(CinMNH) includes a left mandible (VP 1689) (Fig.Ib), lower left fourth premolar (VP 3954), right upperincisor (VP 1688), right scapula (VP 2003), right tibia(VP2004), two thoracic vertebrae (VP 3769, 1690) anda mid-caudal vertebrae (VPI691). Subsequent excava-tions by Ken Tankersley, formerly of Kent State Uni-versity, recovered a lumbar, (CMNH 1770AOI9-03-01).
Measurements of the mandible (CinMNH VP1689) and incisor (CinMNH VP 1688) are provided inTable I. Some of the bones from the Sheriden Pit suchas the scapula and the tibia show extensive modificationdue to gnawing. On the tibia this is most pronouncedaround the proximal end, making it difficult to obtainusable measurements for comparison with other speci-mens. Measurements of the tibia are provided in Table 2.
A sample of the Sheri den Pit mandible(CinMNH VP1689) produced an AMS date of 10,850 ±60 years B.P. (CAMS-26783) (Tankersley & Landefeld,1998). It is the youngest directly dated specimen ofCastoro ides utilizing this technique.
DISCUSSIONCurrently there are two recognized species
of Castoro ides, C. ohioensis and C. leiseyorum. C.ohioensis is only known from the Rancholabrean whileC. leiseyorum was first described from the Irvingto-nian Leisey fauna in Florida (Morgan & White, 1995).Parmalee & Graham (2002) described a skull of C.leiseyorum from the Cooper River, South Carolina andconsidered a number of alternatives in interpreting thespecies. One of these is that it may be a temporal spe-cies and the observed anatomical differences betweenit and C. ohioensis reflects its earl ier evolutionarystage in the lineage. However they also considered thepossibility that the Cooper River specimen was pos-sibly late Pleistocene in age and thus C. leiseyorummay have survived into the Rancholabrean and wasa contemporary of C. ohioensis. Unfortunately theCooper River specimen lacks both stratigraphic contextand age control. If however, C. leiseyorum did surviveinto the Rancholabrean it would then be considered thesoutheastern species of Castoro ides while C. ohioensismay have been restricted to the north around the GreatLakes and to the west. Alternatively C. leiseyorum maymerely be a geographic variant of C. ohioensis. For thepurposes of this paper we consider only the extinctionof C. ohioensis and the taxonomic, spatial and tempo-ral relationships of the two species still require criticalevaluation.
While the taxonomic questions concerning thenumber of species of Castoroides in the late Pleis-tocene remain unresolved based on currently knownspecimens, the broader question of the extinction of
53
the genus can still be examined. Given the paucity ofspecimens of Castoro ides with good radiocarbon dates,it is only possible to evaluate any patterns or trends ofchanges in its distribution prior to its extinction at theend of the Pleistocene in the broadest terms. As a partof this analysis we also consider long term trends withregard to distribution of not only Castoroides but itsevolutionary predecessors so as to provide a broadercontext in which to evaluate the extinction of Castoroi-
des, the terminal member ofthe subfamily Castoroidi-nae, at the end of the Pleistocene.
While Castoroides in the late Rancholabreanis considered to be widely distributed, with a range thatextends from Alaska and the Yukon in the northwest(Harington 1978) eastward to New Brunswick (Milleret a!., 2000) and south to Florida (Martin 1969, 1975),its distribution is more restricted than earlier membersof the subfamily. While the subfamily Castoroidinae
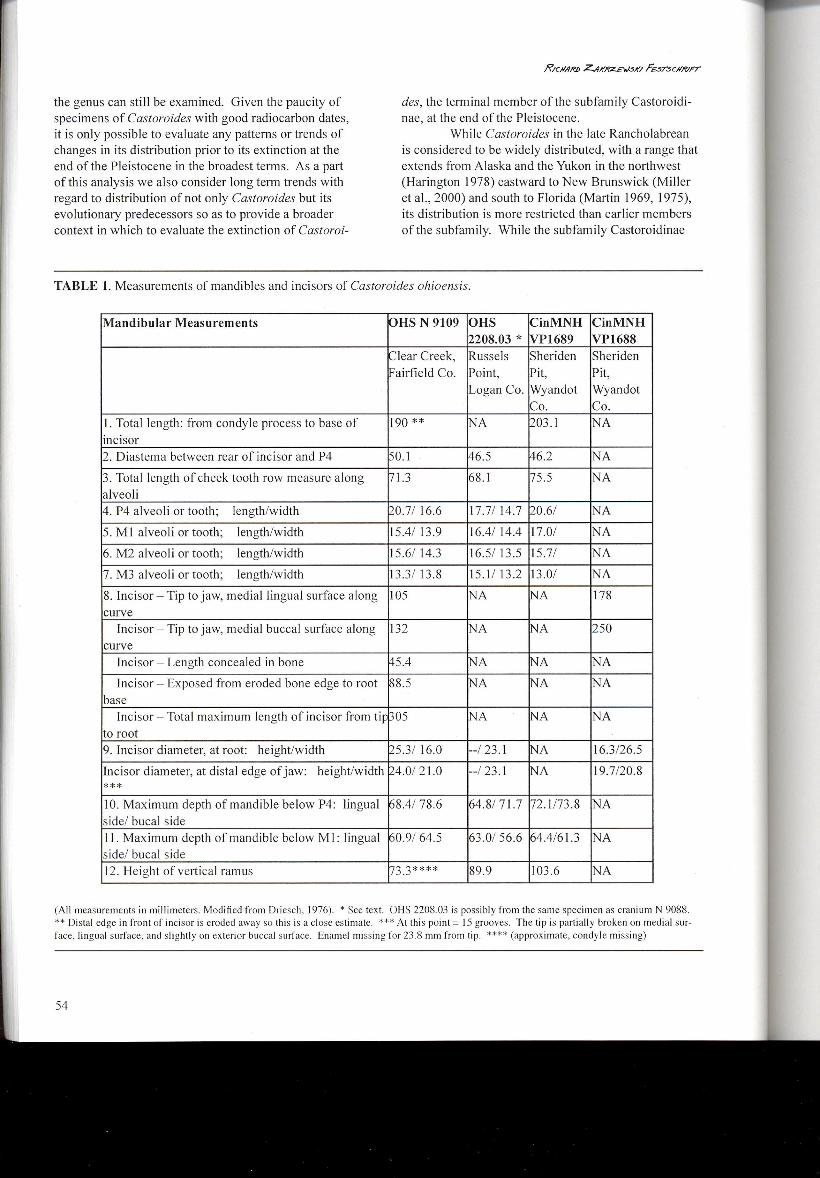
TABLE 1.Measurements of mandibles and incisors of Castoroides ohioensis.
Mandibular Measurements OHS N 9109 OHS CinMNH CinMNH2208.03 * VP1689 VP1688
Clear Creek, Russels Sheriden SheridenFairfield Co. Point, Pit, Pit,
Logan Co. Wyandot WyandotCo. Co.
I.Total length: from condyle process to base of 190 ** NA 203.1 NAincisor2. Diastema between rear of incisor and P4 50.1 46.5 ~6.2 NA
3. Total length of cheek tooth row measure along 71.3 68.1 75.5 NAalveoli4. P4 alveoli or tooth; length/width )0.7/16.6 17.7/14.7 ~0.6/ NA
5. Ml alveoli or tooth; length/width 15.4/13.9 16.4/14.4 17.0/ NA
6. M2 alveoli or tooth; length/width 15.6/14.3 16.5/13.5 15.7/ NA
7. M3 alveoli or tooth; length/width 13.3/13.8 15.1/13.2 13.0/ NA
8. Incisor - Tip to jaw, medial lingual surface along ]05 NA ~A 178curve
Incisor - Tip to jaw, medial buccal surface along 132 NA ~A 250curve
Incisor - Length concealed in bone ~5.4 NA ~A NA
Incisor - Exposed from eroded bone edge to root ~8.5 NA tNA NAbase
Incisor - Total maximum length of incisor from tit ~05 NA ~A NAto root9. Incisor diameter, at root: height/width ~5.3/16.0 --/23.1 tNA 16.3/26.5
Incisor diameter, at distal edge of jaw: height/width 174.0/21.0 --/23.1 tNA 19.7/20.8
***10. Maximum depth of mandible below P4: lingual ~8.4/78.6 64.8171.7 172.1173.8 NAside/ bucal sideII.Maximum depth of mandible below MI: lingual ~0.9/64.5 63.0/56.6 ~4.4/61.3 NAside/ bucal side12. Height of vertical ramus ~."),,**** 89.9 103.6 NA-' ..,
(All measurements in millimeters. Modified from Driesch, 1976). * See text. OHS 2208.03 is possibly from the same specimen as cranium N 9088.** Distal edge in front of incisor is eroded away so this is a close estimate. *** At this point = 15 grooves. The tip is partially broken on medial sur-face, lingual surface, and slightly on exterior buccal surface. Enamel missing for 23.8 rnrn from tip. **** (approximate, condyle missing)
54

Sangamonian) and Robert (late Wisconsinan) faunas(Schultz, 1967).
The inferred habitat of Castoroides was riverbottom, lakes, embayments and associated bogs andswamplands (Parmalee & Graham, 2002) and presum-ably the reduction of its range through time reflects adecrease in availability of this type of habitat in thewestern United States and Great Plains due to increas-ing aridity. The previous synopsis illustrates that thereduction in the range of the Castoroides lineage was along term trend that began well before the late Ran-cholabrean. By the late Rancholabrean (younger than150,000 years), the distribution of the lineage givingrise to Castoro ides had been effectively reduced andrestricted to the eastern United States and northwardinto Canada, specifically the Yukon, where presumablyits preferred habitat still persisted. There are currentlyno records in north-central Canada (Alberta, Saskatch-ewan, or Manitoba) to indicate if the two populations(Yukon and eastern North American) were contiguousalthough it is likely that suitable habitat existed betweenthe two regions.
With a range that extended into Alaska and theYukon in the Rancholabrean there is no question thatthe northern populations of Castoroides were directlyimpacted by the development of continental glaciers.While the Canadian records of Castoroides are scant,with only three localities: Old Crow Basin, Yukon;Don Valley Brickyard, Toronto, Ontario and IndianIsland in the Bay of Fundy, New Brunswick, they arestill instructive. The Old Crow Loc. 44 fauna, whichincludes both in situ Castoroides ohioensis and Cas-tor canadensis, is considered Sangamonian interglacial(Harington, 1990), as are the Ontario (Karrow, 1990)and New Brunswick (Miller et al., 2000) records. Withthe development of the last continental ice sheets in the
TABLE 2. Measurements of tibia of Castoro ides ohioensis. All measurements in millimeters.
Measurement CinMNH FM P15272 SMM P 69.26.1VP2004 after Galbreath, rightight 1938
Maximum length 753.5 237 230
Transverse diameter of 64 45.8'Proximal endA-P diameter at center 31of shaftTransverse diameter of 37.9 ~9 ~0.1distal endTransverse diameter of ~8 ~2.1distal articular surface
55
first appears in the early Miocene (Hemingfordian)(Korth, 2002) the earliest ancestor to Castoro idesconsidered here is Dipoides from the Hemphillian (ca.9 to 4.9 Ma). At this time its distribution extends as farwest as Arizona and Oregon (Shotwell, 1955; Wagner,1983). In the early Blancan the genus is still found inIdaho (Zakrzewski, 1969) but is not yet known fromArizona (Morgan & White, 2005). Dipoides gave riseto Procastoroides in the middle Blancan and westernrecords include Washington (Morgan & Morgan, 1995)and Idaho (Hearst, 1998). The first appearance ofthe genus Castoroides is in Irvingtonian I (1.9 Ma to900,000 years B.P.) in the Arkalon fauna of Kansas andLeisey fauna of Florida (Morgan & White, 1995). Dur-ing Irvingtonian II (900,000 to 400,000 years B.P.) thewesternmost record of the genus is from the CourtlandCanal fauna, also in Kansas (Eshelman & Hager, 1984).During Rancholabrean I(400,000 to 150,000 yearsB.P.) Castoroides is known from a number of faunason the Great Plains, four in Kansas (Adams, Kanopo-lis, Rezabek, and Sandahl), three in Nebraska (Angus,Hay Springs, and Mullen II)and one in Oklahoma(Berends). There are additional western records of Cas-toroides in central Nebraska and northeastern Colorado,but it should be noted that the ages of these faunas,such as the Colorado record, are currently unknown. Itappears that Castoroides disappeared from the GreatPlains by Rancholabrean II (younger than 150,000years) and its range had become restricted, with thewesternmost record in eastern Kansas on the KansasRiver, except for the population in the Yukon. One sitethat documents its disappearance in the Great Plains isthe four superposed faunas at Butler Springs, Kansas,where Castoroides is present in the Rancholabrean IAdams fauna, but absent in the younger RancholabreanII Butler Springs (late Illinoian), Cragin Quarry (early

Wisconsinan it appears that the Yukon and other Cana-dian populations were extirpated as there is currently noevidence that the giant beaver was present in Canada inthe Wisconsinan. The Wisconsinan glaciation com-pressed the range of Castoroides southward and east-ward so that by the height of the late glacial maximumits distribution had been effectively reduced by at leasthalf. In Ohio Castoro ides was able to rein vade someof its former range in the northern part of the state thatwas glaciated (Sheriden Pit and Clear Creek) but didnot disperse as far north as Canada despite the presenceof numerous proglacial lakes and associated wetlandhabitat which should have presumably provided anideal environment for the animal. This is in markedcontrast to other taxa with which it was commonly as-
sociated, such as the mastodon, (Mammut americanum)and extant beaver, Castor canadensis, both of whichdispersed northward into Canada in the late Wiscon-sinan following deglaciation (Harington 1978,2003).As a general trend it appears that there was a consistentdecrease in the range of Castoroides throughout its his-tory with only moderate gains in the late Pleistocene inreinvading territory impacted by continental glaciation.
RADIOCARBON DATES AND LATEST PLEIS-TOCENE DISTRIBUTION OF CASTOROIDES
There are few published radiocarbon dates forCastoroides, either based on associated organic mate-rial or direct dates from bone (Table 3). The first suchpublished date for Castoroides is 9,550 ± 375 years
TABLE 3. Radiocarbon dates associated with Castoro ides ohioensis.
Locality Latitude Longitude Age Age Source [Reference
Christensen Bog 39.87 ° N 85.83° W 14,080 ± 150 to Stratigraphic fGraham et aI.,Hancock Co. 12,060 ± 100 ISGS-502 1983Indiana ISGS-501Monmouth Brooks, 40.36°N 74.17 ° W 12,470 ± 250 Stratigraph ic Parris et al.,Monmouth Co. and poor 1995New Jersey 12,510 ± 180 association
GX-18769GX-25818
Kansas River, 39.07° N 95.78° W 12,150±80 AMS [This paperJohnson Co. CAMS-20004KansasClear Creek, 39.64° N 82.76° W 12,040 ± 35 AMS [This paperFairfield Co. Ohio UCLAMS-1121
9Dutchess Quarry 41.36°N 74.36° N 11,670 ± 70 AMS Funk &Cave 8, Orange Co 11,550 ± 60 CAMS-12587 Steadman, 1994New York r'AMS-33975Northern Lights, 40.03° N 82.97° W 11,480 ± 160 Y-526 Prufer & Baby,Franklin Co. Ohio 9,600 ± 500 M-66 1963
Associatedwood
Mississippi River, :44.9 ° N 93.2° W 10,320 ± 250 Whole bone !Erickson, 1967Ramsey Co.MinnesotaSheriden Pit, ~0.98° N 83.44° W 10,850 ± 60 AMS [Tankersley &Wyandot Co. Ohio CAMS-26783 Landefeld, 1998Ben Franklin, ~3.49° N ~5.77° W 9,550 ± 375 Charcoal and Slaughter &Dallas Co. Texas 11,135 ± 450 mollusk shell, Hoover, J 963
poorstratigraphicassociationSM-532SM-533
56

B.P. for a specimen from the Ben Franklin fauna, Texas(Slaughter & Hoover, 1963). The Ben Franklin faunawas recovered from four localities and the Castoro ideswas recovered from locality I. The radiocarbon dateof9,550 ± 375 was based on charcoal from a possiblehuman hearth near quarry 5 and another date of 11,135 ±450 was based on fresh water mussel shell from quarry 4.The lack of good stratigraphic association of either ofthese dates with the Castoro ides brings into questiontheir validity as being indicative ofthe true age of thespecimen.
A partial skeleton from along the MississippiRiver in Ramsey County, Minnesota was directly datedusing whole bone and an age of 10,320 ± 250 years B.P.was obtained (Erickson, 1967).
A radiocarbon date of 11,480 ± 160 years B.P.was reported by Prufer & Baby (1963) for wood associ-ated with the skull (OHS N 9090, formerly OSM 2211)from the Northern Lights locality off Cleveland Avenuein northeastern Columbus, 1.1 km west and 2.4 kmsouth of the northeast corner of Clinton Township, sec-tion I,Franklin County, Ohio (approximately N 40.03°and W 82.97° W).
Graham et a!. (1983) reported a Castoroidestooth from the Christensen Bog Mastodon site recov-ered from deposits dated between 14,080 ± 150 and12,060 ± 100 years B.P.
AMS dates were made directly from a Casto-roides tooth (NYSM-1188) recovered from DutchessQuarry Cave 8, Orange County, New York (41.36°N,74.36° W). The two dates were 11,670 ± 70 years B.P.(CAMS-12587) and 11,550 ± 60 (CAMS-33975) (Funk& Steadman, 1994).
FIGURE 2. Map showing approximate minimum range oi Castoroi-des at different time intervals in the late Pleistocene. Pairs of local i-ties correspond to those in table 3.
A mandible (KUVP-l 04035) from the KansasRiver near Lawrence (ca. 39.07° N, 95.78° W) has beenAMS dated at 12,150 ± 80 years 8.P. (Tom Stafford,pers. comm., 2005).
While the limited number of dates precludesin-depth analysis of any changes in the geographic dis-tribution of the giant beaver prior to its extinction, theydo at least provide the basis for a simple examinationwith regard to any changes in its distribution from about12,500 to 10,500 B.P. Based on the dates listed abovewe constructed four pairs of localities that provide fourtime slices at approximately 500 year intervals. Whilethese approximately coeval pairs of localities do notfully define the distribution of the species at each ofthese points in time they do provide at least some indi-cation ofthe animal's minimum distribution based onthe distance between each member of the pair.
While neither of the oldest pair of localities,Christensen Bog, Indiana (Graham et al., 1983) andBig Brook, New Jersey (Parris et al., 1995) is basedon direct dates their respective stratigraphic associa-tions can be considered roughly equivalent in time atca. 12,500 years B.P. These two localities are 993 kmapart. Younger specimens with direct dates include theClear Creek specimen (12,040 ± 35) and the KansasRiver specimen (12,150 ± 80) reported here and thesetwo localities are 1,120 km apart. The Dutchess QuarryCave 8 (11,670 ± 70) and Northern Lights localities(11,480 ± 160) are also widely separated at 740 km. At10,850 ± 60 the Sheriden Pit Cave specimen and theMinnesota specimen (10,320 ± 250) are currently theyoungest dates for Castoroides ohioensis based on di-rect bone dates. These two localities are 907 km apart.As discussed above, the date for the Ben Franklin speci-men is probably not an accurate indication of its age.
While these pairs of sites do not definitivelyprovide any indication of Castoroides' maximum dis-tribution for each point in time, they at least provide anindication of the minimum size of its range. The mini-mum distribution for the species has been representedas a circle for each set of localities (Fig. 2) with the twolocalities placed on the edge of the circle. This crudeapproximation will be modified as more radiocarbondates become available, but based on available informa-tion it provides a way to examine whether there wasany observable shift in the distribution of Castoroidesprior to its extinction. As illustrated in Figure 2, basedon this Iim ited data set it does not appear there was anysignificant change in the range of Castoro ides prior toits extinction or a consistent shift in any direction as allof these records broadly overlap.
A significant weakness of the above analysis isthe lack of radiocarbon dates for southern records of thegenus. As these become available they will certainly
57

provide a refinement of our knowledge as to its distri-bution in time and better show any trends in changesin its range prior to its extinction. It will be instructiveto see if dates from the southeast cluster with those inthe Great Lakes region provided here or if the datesobtained from specimens in the southeast are younger.If the Blitzkrieg model for Pleistocene extinctionsproposed by P. S. Martin (1967, 1984) is correct and itsextinction was human induced then specimens of Cas-toroides younger than the Sheriden Pit specimen shouldbe present in the southeast, and we should be able torecover material that clearly demonstrates evidence ofhunting and butchering.
Certainly more dates are required in order tobetter correlate the relationship between the regionaldisappearance of the giant beaver with local climatic orenvironmental change or the first appearance of humansin the area. It should be noted that two carved andbeveled bone rods and a Gainey style fluted point wererecovered from the Sheri den Pit (Redmond & Tankers-ley, 2005). Based on the tapered ends and overall mor-phology the bone rods were interpreted as indicative ofhaving been used as projectile points and not foreshafts.The bone points were recovered from sediments datedbetween 11,060 and 10,400 radiocarbon years and over-lap in time with that of the giant beaver. The Sheri denPit is thus the only North American site that clearlydocuments both temporal and geographic overlapbetween paleo-indians and the giant beaver. However,none of the giant beaver bones recovered indicate anyevidence of butchering or other human modification,so do not provide any evidence of direct interactionbetween the two species.
ACKNOWLEDG EMENTSThe authors wish to thank Jeff Snyder Jr. of
Amanda, Ohio; the Hendricks Family of Vanlue Ohio,particularly Richard Hendricks who allowed Greg Me-Donald to conduct excavations at the Sheri den Pit. KenTankersley, Northern Kentucky University, generouslyprovided information based on his work at the Sheri denPit, and Tom Stafford kindly allowed us to includethe information on the Kansas River specimen. Thepaper has benefited from the comments provided by thereviewers and we wish to express our appreciation.
REFERENCESDriesch, A. von den. 1976. A guide to the measure-
ment of animal bones from archaeologicalsites. Peabody Museum Bulletin I, \- 136.
Erickson, B.R. 1967: Paleontological evidence con-cerning some post glacial features of the Mis-sissippi River Valley. Scientific Publicationsof the Science Museum n.s. I, 1-4.
58
Eshelman, R. & Hager, M. 1984: Two Irvingtonian(Medial Pleistocene) vertebrate faunas fromnorth-central Kansas. In H.H. Genowaysand M.R, Dawson (eds.): Contributions inQuaternary vertebrate paleontology: A volumein memorial to John E. Guilday, 384-404.Carnegie Museum Natural History SpecialPublication No.8.
FAUN MAP Working Group. \994: FAUNMAP:A database documenting late Quaternarydistributions ofrnammal species in the UnitedStates. Illinois State Museum ScientificPapers 25 Volumes 1, 1-288, Volume 2, 289-690.
Forsyth, J.L. 1962: Glacial Map of Fairfield County,Ohio. In Wolfe, E.W., Forsyth, J.L. & Dove,G.D. Geology of Fairfield County. Plate 2.Department of Natural Resources, Division ofGeological Survey, Bulletin 60, 1-230, plus 3fold-out maps.
Foster, 1. W 1837: Organic Remains (Type descrip-tion of Castoro ides ohioensis within report).In Mather, W W 1838. Second Annual Reportof the Geological Survey ofthe State of Ohio.pages 79-83. Geological Survey. Columbus,Ohio.
Funk, R.E., & Steadman, D.W. 1994: Archaeologicaland paleoenvironmental investigations in theDutchess Quarry Caves, Orange County, NewYork. 125 pp. Persimmon Press Monographsin Archaeology. Buffalo, New York.
Galbreath, E.C. 1938: Post-glacial fossil vertebratesfrom east-central Illinois. Geological Seriesof the Field Museum of Natural History 6,303-313.
Gilbert, B. M. 1990: Mammalian Osteology. 428 pp.Missouri Archaeological Society, Columbia,Missouri.
Graham, R.W, HoLman, lA., & Parmalee, P.W 1983:Taphonomy and paleoecology of the Chris-tensen Bog mastodon bone bed, HancockCounty, Indiana. Illinois State Museum,Reports of Investigations 38,1-29.
Harington, C.R. 1978: Quaternary vertebrate faunasof Canada and Alaska and their suggestedchronological sequence. Syllogeus 15, 1-J 05.
Harington, C. R. J 990: Vertebrates of the last in-terglaciation in Canada: a review, with newdata. Geographic physique et Quaternaire 44,375-387.
Harington, C.R. (ed.). 2003: Annotated bibliographyof Quaternary vertebrates of northern N0I1hAmerica with radiocarbon dates. University ofToronto Press, Toronto.

Hearst, J. 1998: Depositional environments of theBirch Creek local fauna (Pliocene: Blancan),Owyhee County, Idaho. In W.A. Akersten,H.G. McDonald, J. Meldrum and M.T. Flint(eds.): And Whereas ... Papers on the Verte-brate Paleontology ofIdaho Honoring JohnA. White, Vol. 1,56-93. Idaho Museum ofNatural History Occasional Paper 36.
Karrow, P.F. 1990: Interglacial beds at Toronto On-tario. Geographie physique et Quaternaire 44,289-297.
Korth, W.W. 2002: Comments on the systematicsand classification of the beavers (Rodentia,Castoridae). Journal of Mammalian Evolution8,279-296.
Martin, P. S. 1967: Prehistoric overkill. In P. S. Martinand H. E. Wright (eds.): Pleistocene Extinc-tions: The Search for a Cause, 75-120: YaleUniversity Press, New Haven and London
Martin, P.S. 1984: Prehistoric overkill: the global model.In P.S. Martin and R.G. Klein (eds.). Quater-nary Extinctions: a prehistoric revolution, 354-403. University of Arizona Press, Tucson.
Martin, R. A. 1969: Taxonomy of the giant Pleistocenebeaver Castoroides from Florida. Journal ofPaleontology 43, 1033-) 04l.
Martin, R. A. 1975: Giant Pleistocene beavers and theWaccasassa River, Levy County, Florida. Bul-letin of the New Jersey Academy of Science20,26-30.
Meeker, R.L., lH. Petro, J.H., and Bone, S.W 1960: Soilsurvey of Fairfield County, Ohio. United StatesDepartment of Agriculture, Ohio AgriculturalExperiment Station. Series 1951, No.7, 1-77.
Mickelson, D.M., Clayton, L., Fullerton, D.S., & BornsJr., H.W 1983: The Late Wisconsin GlacialRecord of the Laurentide Ice sheet in the UnitedStates. In H.E. Wright Jr. (ed.): Late-Quater-nary Environments of the United States, Vol. IThe Late Pleistocene 3-37 (S.c. Porter ed.)
Miller, R. F., Harington, C.R., & Welch, R. 2000: Agiant beaver (Castoro ides ohioensis Foster)fossil from New Brunswick, Canada. AtlanticGeology 26, 1-5.
Morgan, G.S. & White, J.A. 1995: Small mammals(Insectivora, Lagomorpha, and Rodentia) fromthe early Pleistocene (Irvingtonian) LeiseyShell Pit local fauna, Hillsborough County,Florida. Bulletin of the Florida Museum ofNatural History 38 Pt. 11,397-461.
Morgan, G.S. & White Jr., R.S. 2005: Miocene andPliocene vertebrates from Arizona. NewMexico Museum of Natural History and Sci-ence Bulletin 29, 115-136.
Morgan, J.K. & Morgan, N.H. 1995: A new species ofCapromeryx (Mammalia: Artiodactyla) fromthe Taunton local fauna of Wash ington; andthe correlation with other Blancan faunas ofWashington and Idaho. Journal of VertebratePaleontology 15, 160-170.
Parmalee, P.W & Graham, R.W. 2002: Additionalrecords of the giant beaver, Castoroides, fromthe Mid-south: Alabama, Tennessee, and SouthCarolina. Smithsonian Contributions to Paleo-biology 93, 65-71.
Parris, D.C., Camburn, J. & Camburn, S. 1995: Amegafaunal radiocarbon date for a MonmouthCounty Brook locality, New Jersey. Journalof Vertebrate Paleontology 15 supplement,47A-48A.
Prufer, O. H. & Baby, R.S .. 1963: Palaeo-Indians ofOhio. Ohio Historical Society, Columbus,Ohio. 68 pp.
Redmond, B.G., & Tankersley, K.B .. 2005: Evidenceof early paleoindian bone modification anduse at the Sheriden Cave Site (33WY252),Wyandot County, Ohio. American Antiquity70,503-526.
Schultz, G.E. 1967: Four superimposed late-Pleisto-cene vertebrate faunas from southwest Kansas.In P.S. Martin and H.E. Wright Jr. (eds.):Pleistocene Extinctions, the Search for theCause, 321-336. Yale University Press, NewHaven.
Shotwell, l.A. 1955: Review of the Pliocene beaverDipoides. Journal of Paleontology 29, 129-144.
Slaughter, B. H., & Hoover, R. 1963: Sulphur RiverFormation and the Pleistocene mammals ofthe Ben Franklin local fauna. Journal of theGraduate Research Center, Southern MethodistUniversity 31, 132-148.
Tankersley, K.B. & Landefeld, C.S. 1998: Geochro-nology of the Sheriden Cave, Ohio: The 1997Field Season. Current Research in the Pleisto-cene 15, 136-138.
Wagner, H. M. 1983: The cranial morphology ofthe fossil beaver Dipoides smithi (Rodentia:Mammalia). Natural History Museum of LosAngeles County, Contributions in Science 346,1-6.
Zakrzewski, R. J. 1969: The rodents from the Hager-man local fauna, upper Pliocene of Idaho.Contributions from the Museum of Paleontol-ogy, University of Michigan 23, 1-36.
59




















