
Carnivores (Canidae, Felidae and Ursidae) of Romain-la-Roche (Doubs, France)
108
1661-5468 VOL. 29, N° 2, 2010
Transcript of Carnivores (Canidae, Felidae and Ursidae) of Romain-la-Roche (Doubs, France)

1661-5468
VOL. 29, N° 2, 2010

ISSN 0253-6730
1 LAMPEA, UMR 6636 du CNRS, Institut Dolomieu, 15 rue Maurice Gignoux, F-38031 Grenoble cedex, France. [email protected]
I. IntroductIon
Le site de Romain-la-Roche (Doubs) a livré à Patrick PauPe et à son équipe franc-comtoise un matériel paléontologique considérable. Les carnivores y figurent en bonne place. Plus d’une dizaine d’espèces ont fréquenté les abords de l’aven. Les animaux y sont tombés, livrant maintenant un matériel d’étude comptant des milliers de pièces. Ce matériel a été collecté méthodiquement, traité et enregistré avec soin. Il constitue maintenant un repère important pour les faunes pléistocènes. Les canidés, les ursidés, les félidés abondent. Si les hyénidés n’ont laissé aucun vestige, quelques coprolithes attestent cependant de leur passage sur le site. Les mustélidés sont également représentés. Leur étude en sera assurée par Philippe Fosse.La présence de ces carnivores, dominée par les loups, les ours et les lions des cavernes n’a rien de surprenant. Les ours hivernent dans la grotte quand elle fonctionne en
système fermé. La présence occasionnelle des loups et des lions reste une constante de ce type de site. Quand l’érosion a pour conséquence l’effondrement du toit de la galerie, cette dernière devient un piège de type aven s’ouvrant sur le plateau qui domine la vallée de l’Ognon. La température interne d’un karst reste relativement régulière, chaude en hiver et fraîche en été. Les grands herbivores recherchent généralement les abords de tels avens. L’herbe, à proximité de l’ouverture, pousse plus tôt dans la saison car l’air chaud fait fondre la neige plus vite au dégel et elle croît plus rapidement. Ces grands herbivores possèdent en général une vue faciale médiocre, rendant forte la probabilité de tomber dans un trou béant au ras de la surface du plateau. L’aven de Romain-la-Roche a fonctionné comme un piège efficace, sur la durée : les mammouths, les rhinocéros, les grands bovidés, les cervidés et les chevaux en font les frais. Leurs cadavres en décomposition, un de temps en temps, ont alerté par l’odeur les carnivores, grands et
Revue de Paléobiologie, Genève (décembre 2010) 29 (2) : 495-601
carnivores (canidae, Felidae et ursidae) de romain-la-roche (doubs, France)
alain ARgANt1
résuméLe gisement de Romain-la-Roche (Doubs) livre une faune abondante où les carnivores figurent en bonne place. Il constitue maintenant un repère important pour les canidés, les ursidés et les félidés représentés par un matériel considérable. Les canidés correspondent à Vulpes lagopus, Vulpes vulpes et principalement à Canis lupus. Deux Ursidés coexistent comme souvent, Ursus arctos et Ursus spelaeus de loin le plus abondant. trois espèces de félidés sont présentes, Felis silvestris, Panthera pardus et surtout Panthera spelaea, dont trois crânes et un squelette réparti sur quelques m2 sont connus. La biochronologie de Canis lupus, d’Ursus spelaeus et de Panthera spelaea apporte des éléments de datation convergents, centrés autour du stade 5 (Interglaciaire Eémien). La méthode de RabedeR appliquée aux quatrièmes prémolaires d’Ursus spelaeus indique un stade évolutif comparable à celui de l’ours des cavernes des couches profondes d’Herdengelhöhle (Autriche), daté entre 112 000 et 135 000 ans.
Mots-clésRomain-la-Roche, aven, paléontologie, carnivores, Pléistocène.
Abstractcarnivores (canidae, Felidae et ursidae) of romain-la-roche (doubs, France).- In the Romain-la-Roche (Doubs) fossil site the carnivores are an important part in the great amount of fauna remains excavated. they form now an important reference for the Canidae, Ursidae and Felidae, which occur there in a considerable quantity. the Canidae correspond to Vulpes lagopus, Vulpes vulpes and mainly to Canis lupus. As often, two Ursidae coexist, Ursus arctos and Ursus spelaeus, which is greatly the most abundant. three species of Felidae occur, Felis silvestris, Panthera pardus and essentially Panthera spelaea. there are three skulls and one skeleton of P. spelaea scattered throughout some square meters. the biochronology of Canis lupus, of Ursus spelaeus and of Panthera spelaea provides convergent dating elements, pointing around stage 5 (Eemian Interglacial). the RabedeR’s method applied on the fourth premolars of Ursus spelaeus indicates an evolutive stage comparable to that of the cave bear found in the deep layers of Herdengelhöhle (Austria) which is dated between 112 000 and 135 000 years.
KeywordsRomain-la-Roche, sink hole, palaeontology, carnivores, Pleistocene.

496 a. ARgANt
petits, qui peuvent la détecter à des kilomètres à la ronde. Ils sont venus pour charogner. Les risques de chute, les confrontations, engendrent à nouveau une mortalité nettement supérieure à la moyenne et leurs vestiges sont venus se mêler à ceux des herbivores. Il existe un peu d’industrie lithique à Romain-la-Roche, mais elle reste épisodique et n’explique en rien l’accumulation des ossements qui demeure totalement naturelle. tout au plus l’homme préhistorique se comporte-t-il en concurrent des charognards, exploitant peut-être la viande et sans doute les fourrures comme le prouvent quelques traces de dépeçage sur des ossements de loup (Pl. VIII-1).Le matériel de Romain-la-Roche apparaît assez bien conservé, rarement roulé, gris blanc à beige, souvent pris dans du sédiment induré ou des concrétions, avec parfois des traces noires de dioxyde de manganèse et/ou de fer traduisant une stagnation dans l’eau. Des éléments fragiles comme les omoplates, les côtes, les vertèbres ne sont pas rares. P. PauPe a précisé l’origine stratigraphique du matériel (PauPe, 1994-1995, 1996, 1997). Les restes de carnivores proviennent essentiellement des couches VI – VII b et c, VIII a et b les plus inférieures. La Figure 1 (PauPe, 1994-1995, p. 15) montre la répartition de l’essentiel du matériel pour chacune des grandes familles de ces carnivores (canidés, ursidés et félidés). On peut constater que les félidés (Felis silvestris, Panthera pardus et Panthera spelaea) correspondent principalement aux couches inférieures VIII a et b. Les ours, et donc Ursus spelaeus, proviennent essentiellement des couches situées juste au-dessus, VI-VII b et c. Les canidés par contre proviennent de l’ensemble de ces couches. Il n’est pas possible de répartir ce matériel en fonction de la stratigraphie d’après les documents dont je dispose. Les indications des listings respectifs, souvent très vagues, voire incertaines (?) ne le permettent pas. Il faut donc étudier globalement chaque espèce. Mais il s’agit de couches consécutives, sans doute proches chronologiquement, et quelques dizaines
de milliers d’années ne représentent qu’une fraction de temps insignifiante en terme d’évolution des espèces.Le matériel déposé au Musée CuvieR de Montbéliard (Responsable des Collections, thierry Malvesy) a fait l’objet d’un enregistrement morphométrique systématique de ma part. Ce système d’enregistrement, dont le principe a été amorcé en 1991, correspond à un plan-type que j’ai adapté à chaque famille de carnivore. Il a pour but d’assurer une standardisation des prises de mesures et des observations morphologiques (PUEMA pour les ursidés, PEMAC pour les canidés, PFEMA pour les félidés, P = plan, EM = enregistrement morphométrique, A = aRgant), aRgant, 1991, 2004a. Chaque type de pièce fossile porte un code de détermination, possède un schéma et un plan de mesurage et d’observation avec des numéros de transfert.La préparation et le marquage du matériel de la fouille de P. PauPe et de son équipe sont remarquables et jouent correctement leur rôle indispensable. Par contre les différents listings informatiques (CAN, URS, FEL) sont à manier avec précaution. Ils ont été réalisés par des non-spécialistes et il existe des erreurs de détermination (bien compréhensibles) ou d’attributions abusives sur la base de la taille. Le listing CAN, par exemple, est loin de contenir réellement 1927 pièces si on enlève les éléments de chat sauvage, de panthère, d’ours, de glouton, attribués trop rapidement et surtout si on ne tient pas compte des nombreux fragments approximativement « taille loup ou renard », indéterminables avec précision mais qui peuvent être tout aussi bien de la panthère, du glouton ou même de petits bovidés ou cervidés. Les remontages et collages, difficiles compte tenu de l’état du matériel à l’origine, présentent aussi un certain nombre d’anomalies : canines inversées, dents s’adaptant à la taille de l’alvéole et collées mais avec un stade d’usure incompatible avec le reste de la denture, qu’il convient de déceler et de prendre en compte à l’étude pour éviter des surprises.
Fig. 1 : Romain-la-Roche. Répartition stratigraphique principale des restes de canidés, d’ursidés et de félidés (d’après PauPe, 1994-1995, p. 15).
FÉLIDÉS CANIDÉSURSIDÉS

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 497
Avertissement :Les mesures données au cours de cette étude sont toujours en mm et l’unité n’apparaît pas dans les tableaux.Une échelle empirique d’estimation de l’usure dentaire, d’application simple et rapide, apporte des indications utiles : U = 0, usure nulle ; U = 1, usure perceptible mais faible ; U = 2, usure moyenne de la couronne et l’émail a parfois disparu ; U = 3, usure très prononcée mais le relief initial reste encore visible ; U = 4, couronne pratiquement arasée ; U = 5, la table d’usure appartient à la racine (aRgant, 1991). Une décimale, U = 0,3 par exemple, traduit simplement un état intermédiaire. Elle n’a surtout pas de valeur mathématique et ne peut en aucun cas donner lieu à des moyennes. Elle apporte simplement dans cet exemple deux informations utiles. La pointe de la canine n’a plus son état originel U = 0 mais n’atteint pas, loin s’en faut, l’usure U = 1. Sa durée de vie a été courte car l’animal est mort jeune.La hauteur de la couronne (HC) d’une dent jugale n’est donnée que dans le cas de dents à usure nulle ou très faible. Elle est prise sur la face externe, en ligne directe et non en projection (trop aléatoire) entre le sommet principal de cette couronne et l’amorce inférieure du collet.
Abréviations utilisées :A ou (A) : mesure sur alvéole(s) d’une dent, (A) mesure entre alvéoles pour une série* = environ (à la suite d’une mesure)cv : corps vertébralDenture : I = incisive ; C = canine ; P = prémolaire ; M = molaireP2/ = deuxième prémolaire supérieureP/2 = deuxième prémolaire inférieureext. prox. = extrémité proximale ; dist = extrémité distalefragt ou fgt = fragmentg ou d = gauche ou droite (parfois g ou D)inf. ; sup. ; ant. ; post. = inférieur(e) ; supérieur(e) ; antérieur(e) ; postérieur(e)juv. = juvénileLat ou lat : latéralisation d’une pièce (g ou d)mand. = mandibulemin. ; max. = minimum ; maximum∆l = min. –max.morpho. = morphologiepart. = partielDAP = diamètre antéro-postérieur = longueurDt = diamètre transversal = largeurDDP = diamètre dorso-plantaire = hauteurDCC = diamètre crânio-caudal = longueur dans le cas d’une vertèbre par exempleMD = diamètre mésio-distal = longueur (pour une dent)VL = diamètre vestibulo-lingual = largeur (pour une dent)HC = hauteur de la couronne MHNL = Muséum d’Histoire Naturelle de Lyon (L = Lyon ; B = Bâle ; Z = Zurich ; P = Paris…)NR = nombre de restes (entiers ou fragmentaires)
NMI = nombre minimum d’individus (nombre strict d’une certaine pièce)NMIE = nombre minimum d’individus exprimé prenant en compte tous les éléments de différentiation disponibles : latéralisation, similitudes d’aspect, morphologiques ou d’usure... (= NMI de combinaison)Mtc., Mtt. = métacarpien, métatarsien (suivi du numéro de rang du doigt concerné), ex. : Mtc.3 = troisième métacarpienNT = numéro de transfert (dans un tableau, une figure, une légende)x3 ou Serr.x3 : prise de mesure par serrage trois points, en appuyant le calibre à coulisse en 2 points évidents d’un même côté et l’autre branche en contact avec la limite maximum opposée. La mesure ne correspond plus à une prise en projection, trop aléatoire dans quelques cas.La numérotation des listes informatiques CAN, URS, FEL est reprise et sert à caractériser les pièces étudiées. Leur « identité », RO.87-R.18-31 par exemple, n’est pas indiquée sauf si c’est nécessaire pour gagner de la place. On peut toujours la retrouver en consultant le listing PauPe par Familles où se trouvent également les coordonnées dans l’espace et l’attribution stratigraphique éventuelle de chaque pièce.
II. étude pAléontologIque
II.1. ordre carnivoraSuper-famille canoidea
Famille canidaeaSous-famille caninaeII.1.1. genre Vulpes
espèce Vulpes lagopus (Linné, 1758)
Le matériel appartenant à des canidés de taille bien infé-rieure à celle de Canis lupus correspond manifestement aux renards. La part principale en est attribuable au renard commun Vulpes vulpes, mais il existe de rares pièces traduisant des individus de très petite taille, ne pouvant correspondre qu’à du renard polaire, Vulpes lagopus. La coexistence des deux espèces, signalée maintes fois dans les gisements d’Europe occidentale, reste tout à fait clas-sique.Bien que le terme Alopex soit encore utilisé, toutes les études systématiques existantes ou en cours montrent que le renard polaire est mieux placé à l’intérieur du genre Vulpes et doit être appelé Vulpes lagopus. Il n’y a aucune raison en matière de systématique ou d’évolution (lignée Vulpes alopecoides – V. praeglacialis – « Alopex » lagopus largement admise), ni même de morphologie ou d’écologie, pour retenir une distinction de genre et de ne pas utiliser Vulpes (Werdelin, in letteris). Je souscris entièrement à cette opinion.F. PoPlin (1976) fait le point sur la distinction entre ces deux espèces lors de l’étude de gönnersdorf (Allemagne) où Vulpes lagopus apparaît bien représenté (une trentaine

498 a. ARgANt
d’individus). Les caractères distinctifs font l’objet d’une étude critique validant les meilleurs caractères discriminants.
II.1.1.1. données morphométriques :La liste d’inventaire du matériel disponible apparaît en annexe à la fin. Il est ici des plus réduit (NR = 5, NMI = 1). tout le matériel dentaire disponible appartenant à des renards peut-être attribué au vu des dimensions à Vulpes vulpes, à l’exception de deux canines supérieures non usées de petite taille laissant planer le doute (CAN.1019, CAN.1022). Ce ne sont malheureusement pas les meilleures pièces discriminantes comme le souligne F. PoPlin. L’état de ces canines au collet ne permet pas non plus de prendre des mesures valables et rend impossible la comparaison des diamètres antéro-postérieurs comme il serait souhaitable.Ces deux canines non usées, peu allongées et de petite taille (CAN.1019 et CAN.1022) font penser à Vulpes lagopus. Elles sont d’habitude plus longues et plus fines chez Vulpes vulpes. F. PoPlin mesure la hauteur de la couronne sur le flanc distal de quelques canines (PoPlin, 1976, p. 31) :Vulpes vulpes : C. sup. 18, 19, 22 mmVulpes lagopus : C. sup. 14, 15, 17 mmSur les deux exemplaires de Romain-la-Roche je trouve respectivement 16,25 et 16,24 mm. Je les attribue donc à Vulpes lagopus.Une septième vertèbre dorsale endommagée CAN.1807, épiphysée, appartient à un renard. Sa morphologie élimine totalement Felis silvestris dont la vertèbre équivalente est à peine plus petite. Cette taille, particulièrement réduite par rapport à Vulpes vulpes actuel me conforte dans la détermination de Vulpes lagopus. Dimensions du corps vertébral : DDC =12,1 ; DT max. = 12,6.Parmi ce matériel, seule une moitié distale de fémur d. (CAN.787) appartenant à un renard de très petite dimension (DT dist. = 16,8 ; DT méd. = 6,9) apporte la certitude de la présence de Vulpes lagopus (Fig. 2). Il existe également un très petit métatarsien 3 d. (CAN.1788) : DT prox. = 6,04, DAP. Prox. = 9,96, DT.
Méd. = 4,25, DAP. Méd. = 3,55. Malheureusement il manque l’extrémité distale. On ne peut donc en connaître la longueur, élément important de la détermination. J’attribue sans certitude cette pièce également au renard polaire.
II.1.1.2. Biochronologie :La lignée évolutive Vulpes alopecoides à Vulpes praeglacialis àVulpes lagopus reste à démontrer, mais n’a rien d’impossible. Vulpes alopecoides du Villafranchien de Saint-Vallier (Drôme) (viRet, 1954 ; aRgant, 2004) est un vrai renard de petite taille qui pourrait être à l’origine des Vulpes actuels. Vulpes praeglacialis du Pléistocène moyen (Escale, Montoussé 4, Arago, Achenheim) (boniFay, 1971 ; CRégut, 1979) est souvent considéré comme l’ancêtre du renard polaire actuel. Les premières mentions de Vulpes lagopus se rencontrent dans les faunes « rissiennes » (tornewton Cave). Au Pléistocène supérieur sa présence est souvent signalée dans des couches livrant une industrie moustérienne. Sa fréquence maximum se situe cependant à la fin de la dernière période glaciaire, lors des périodes les plus froides.Le tableau 1 compare l’extrémité distale du fémur d. (CAN. 787) à celles de Vulpes praeglacialis et de Vulpes
Fig. 2 : Romain-la-Roche, petits Canidés, fémur. DT minimum de la diaphyse en fonction du Dt distal (d’après Clot, 1980, fig. 69, p. 139).
mm
mm
7
8
9
10
16 17 18 19 20 21 22 23
Vulpes vulpes
Vulpes lagopus
= Romain-la-Roche
DT
min
.
DT dist.
Fémur
Tableau 1 : Romain-la-Roche, Vulpes lagopus, comparaison du fémur CAN.787 à Vulpes praeglacialis (boniFay, 1971 ; delPeCh, 1989) et à Vulpes vulpes (Clot, 1980).
Fémur Vulpes lagopus romain-la-roche
V. p. escale
V. cf. p.Vaufrey
V.vgerde
V.v.enlène
V.v pair-non-pair
V.v actuelBigorre
dt distal 16,8 17,5 18,0
819,7-23,7
21,351,16
619,5 à 22,5
21,11,08
720,0 à 21,6
20,860,56
5019,0 à 24,0
20,951,30
dt méd. 6,9 7,6 7,8 - - - -
V. p. = Vulpes praeglacialis V. cf. p. = Vulpes cf. praeglacialis V.v. = Vulpes vulpes

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 499
lagopus. Le Dt dist. apparaît de très petite dimension, inférieure même à la forme ancienne du Pléistocène moyen de L’Escale et de la grotte Vaufrey. Ce fémur de Romain-la-Roche n’atteint pas non plus les limites inférieures des valeurs du renard polaire de gerde (Hautes-Pyrénées), gisement peut-être anté-éemien (Clot & duRanthon, 1987, p. 94), ni à plus fortes raisons celles des renards polaires du Pléistocène supérieur (stades 4, 3 et 2). Mais il s’agit d’un unique exemplaire insuffisant pour permettre des comparaisons sérieuses et avancer une hypothèse autre que la fourchette chronologique la plus large.
II.1.1.3. paléoécologie : Par son adaptation au froid rigoureux, le renard polaire est un bon marqueur climatique. Sa présence lors des périodes les plus froides de la dernière glaciation, en association avec le renne, constitue une preuve suffisante. Mais rien n’empêche qu’il déborde au sud sur les zones tempérées froides et se rencontre associé au renard commun. C’est le cas dans de nombreux gisements d’Europe occidentale. A Romain-la-Roche, il apparaît largement minoritaire, ce qui permet de supposer qu’il se situe en limite de son aire de répartition de l’époque.
II.1.2. genre Vulpesespèce Vulpes vulpes (Linné, 1758)
L’essentiel du matériel de Romain-la-Roche attribuable à de petits canidés correspond à Vulpes vulpes, le renard roux. On peut dénombrer 42 restes pour un nombre minimum de 4 individus (4 ext. prox. de cubitus g.). Le matériel crânien et dentaire est dominé par les demi-mandibules et les dents inférieures isolées. A. Clot
(1980) faisait déjà cette remarque à propos du matériel de gerde (Hautes-Pyrénées) en signalant que cela se produit souvent dans les gisements du Pléistocène européen. A Romain-la-Roche, le matériel post-crânien apparaît très fragmenté et seuls 2 métacarpiens sont complets. Il faut sans doute invoquer l’action des nécrophages et des broyeurs d’os (canidés plutôt que hyénidés) que la petite taille du renard et la moindre résistance de ses ossements devait fortement intéresser.
II.1.2.1. données morphométriques :La liste d’inventaire du matériel disponible apparaît en annexe à la fin (NR = 42, NMI = 4).demi-mandibule : Tabl. 2- CAN.17+44 : demi-mandibule d. avec branche
horizontale privée de l’extrémité antérieure jusqu’à la C inf, branche montante complète avec condyle d’articulation, alvéoles vides de C. inf., de P/2, de P/3, de M/3, présence de P/4, M/1 (CAN. 44), M/2. P/1 manque totalement (aucune trace d’alvéole entre la canine et P/2).
- CAN.1773 : demi-mandibule d., branche horizontale complète depuis l’alvéole de la C. inf. avec alvéoles vides de C., P/1, P/2, P/3, P/4 et M/3. La M/1 et la M/2 sont présentes. La branche montante manque.
- CAN. 20 : moitié de demi-mandibule g. avec branche horizontale endommagée au niveau de P/3, branche montante cassée à mi-hauteur. Condyle présent mais incomplet, P/4 cassée à la moitié antérieure, M/1 cassée, M/2 et M/3 présentes. Correspond très certainement à CAN.17 (taille, morphologie, aspect, couleur identiques).
- CAN. 169 : petit fragment de demi-mandibule d. avec M/2 et alvéole de M/3.
CAN. 17 est la seule pièce pratiquement complète à
Tableau 2 : Romain-la-Roche, Vulpes vulpes, demi mandibule. Données disponibles (A = entre alvéoles), Pemac 20 (Fig. 7).
Vulpes vulpesdemi mandibulepemac 20
4 5 7 8 9 10 11 12 15 20 21 22 23
CAN.17 d. 90,33* 84,53 58,98 (A) - 55,31
(A) 26,28 - 30,0 14,82 5,74 33,66 12,54 12,46
CAN.1773 d. - 88,63 62,53 (A)
58,59 (A)
54,10 (A) 24,37 33,68
(A)29,84 (A) 13,01 5,64 - 14,05 10, 63
CAN.20 g. - - - - - 25,54 - - 15,0 5,98 - 12,66* -
4 - L. C. post. – condyle (milieu)5 - L. C. post. – incisure angulaire6 - L. C. post. – processus angulaire7 - L. C. post. – M/3 post..8 - L. P/1-M/39 - L. P/2-M/310 - L. molaires
11 - L. prémolaires 12 - L. P/2-P/415 - L. alvéole M/120 - Dt mandibule derrière M/121 - Hauteur processus angulaire-coronion22 - Hauteur mandibule derrière M/1 (face externe)23 - Hauteur mandibule derrière P/2

500 a. ARgANt
l’exception de la partie antérieure manquante. La P/1 n’existe pas. Elle est par contre nettement présente sur CAN.1773. Dans tous les cas d’observations possibles, les dents se touchent parfaitement sauf sur CAN.1773 où existe un espace de 3,2 mm entre les alvéoles vides de P/2 et de P/3. La branche horizontale apparaît longue et relativement fine, assez aplatie latéralement par rapport au renard actuel. L’apophyse coronoïde de la branche montante de CAN.17 monte moins haut et se courbe un peu dans sa partie supérieure. La fosse deltoïdienne est profonde, plus creuse que sur Vulpes vulpes actuel. Le condyle d’articulation (CAN.17, CAN.20) apparaît plus étroit transversalement et plus massif en son centre que sur l’actuel.
etude dentaire :dents supérieures : Tabl. 3- P4 / : la carnassière supérieure du renard de Romain-
la-Roche est connue par deux dents isolées, non usées, à couronne creuse (jeune individu) (CAN. 45 g. ; CAN. 1135 d.). Elle montre un paracône développé, très redressé, avec une arête rectiligne descendant jusqu’au léger rebord limitant la couronne à l’avant. Le métacône, en contrebas, correspond à une lame un peu arquée atteignant l’arrière de la couronne. Un cingulum net le borde côté lingual. Le tubercule antéro-interne habituel apparaît bien présent mais sans être particulièrement volumineux comme sur Vulpes vulpes actuel. Il est déporté vers l’avant. La morphologie et la taille correspondent au renard actuel tout en donnant l’impression d’une plus grande étroitesse accentuant l’aspect coupant de la denture (paracône élevé et étroit).
- M1/ : les deux premières molaires supérieures
isolées CAN.46 g et CAN.36 d, peu usées (U = 0,3) appartiennent à Vulpes vulpes (Clot, 1980, fig. 61, p. 130). Le paracône domine le métacône, mais les deux possèdent un plus fort développement sur CAN.36 et paraissent nettement plus grêles sur CAN.46. Le bord externe de la couronne de CAN.36 possède un développement beaucoup plus important avec des angles beaucoup plus saillants. Un cingulum net borde le côté vestibulaire, également beaucoup développé sur CAN.36. Le reste de la couronne de cette M1/ reste conforme à Vulpes vulpes avec un protocône en arc de cercle et un hypocône bien développé formant le rebord le plus interne du talon. On ne peut suspecter une autre espèce comme Nyctereutes par exemple.
dents inférieures : Tabl. 3- C : la canine inférieure d. CAN. 1928 U = 0,2, longue
et fine comme chez Vulpes vulpes n’a pas été prise en compte car le recollage de la racine n’est pas correct et la limite couronne-racine manque de netteté. La hauteur de la couronne élimine en tout cas le renard polaire.
- P/3 : deux troisièmes prémolaires (CAN. 642 d. U = 1 et CAN. 1204 d. U = 0) nous montrent une dent à protoconide redressé, haut et pointu quand il n’est pas usé. A l’avant, un petit renflement amorce ce protoconide au bord antérieur de la couronne. Une cuspide nette émerge à mi-pente le long de l’arête postérieure qui aboutit à un renflement au niveau de la partie terminale de la dent. CAN.1204 non usée apparaît particulièrement étroite et coupante.
- P/4 : deux quatrièmes prémolaires (CAN. 17 d. U = 0 et CAN. 1020 d. U = 0) non usées traduisent une dent étroite, coupante, au protoconide redressé également,
Tableau 3 : Romain-la-Roche, Vulpes vulpes, denture.
Vulpes vulpesdenture lAt. réF.cAn. Md Vl Hc 1 2 3 4
P4/ g. CAN. 45 15,52 6,06 7,72 8,87d. CAN. 1135 14,04 6,22 8,49 8,61
M1/ g. CAN. 46 9,46 11,32 4,87d. CAN. 36 10,25 11,70 5,00
P/3 d. CAN. 642 9,28 3,98 (U = 1)d. CAN. 1204 9,31 4,07 6,11
P/4 d. CAN. 1020 9,91 4,48 6,57
M/1d. CAN. 17 16,16 6,08 8,97 6,45 10,53 5,36 5,91d. CAN. 1773 14,11 5,49 8,57 6,47 9,76 4,96 5,29d. CAN. 136 15,89 5,80 9,24 7,27 10,94 5,04 5,51
M/2
d. CAN. 17 7,17 5,08 4,54d. CAN. 1773 6,90 5,22 4,16g. CAN. 20 7,42 5,14 4,38d. CAN. 169 7,67 5,89 4,51
M/3 g. CAN. 20 3,31 2,57 2,54
1- MD paracône P4/ ou protoconide M/12 - MD trigonide M/1
3 - MD talonide (= L. de l’hypoconide), denticule externe4 -VL talonide seul

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 501
pointu et légèrement arqué vers l’intérieur dans sa partie haute. La morphologie est semblable à celle de la P/3, mais le denticule accessoire apparaît beaucoup plus développé.
- M/1 : trois premières molaires d’usure faible (CAN. 17, CAN. 1773, CAN. 136) assurent la documentation. Le protoconide flanqué d’un métaconide bien développé, côté lingual, domine largement le paraconide. Sur le talonide, s’observent un hypoconide dominant et, côté lingual, un endoconide bien présent, moins volumineux cependant que le précédent. Côté lingual toujours, un petit tubercule visible mais très réduit se rencontre entre le métaconide et l’endoconide, comme le soulignent de nombreux auteurs, sur CAN. 17 et CAN. 1773. Par contre, il faut vraiment la loupe pour l’observer sur CAN. 136. Ces trois M/1 apparaissent étroites avec un aspect très coupant du trigonide (Fig. 3-5).
- M/2 : quatre secondes molaires (CAN. 17 d, CAN. 1773 d, CAN. 20 g, CAN. 169 d) présentent toutes deux cuspides antérieures sensiblement égales en amorce de la couronne et deux cuspides réduites en partie postérieure terminale, avec au centre une dépression longitudinale. Le rebord antéro-externe apparaît enfin plus volumineux et descend un peu plus bas sur la racine, rendant la forme de la couronne asymétrique (Fig. 6).
- M/3 : un exemplaire de troisième molaire g. est resté en place à cause de la brèche qui en a assuré le blocage sur CAN. 20. C’est une dent très réduite à couronne conique un peu allongée longitudinalement. L’alvéole vide existe dans les trois autres cas.
Fig. 3 : Romain-la-Roche, petits canidés, dispersion selon la longueur (MD) et la largeur (VL) de la M/1 (d’après PoPlin, 1976).
Vulpes lagopus
Vulpes vulpes
VLmm M/1
7
6
5
MD mm
= Romain-la-Roche
= Romain-la-Roche
Vulpes lagopus
Vulpes vulpes
L
mm
15
10
M/1mm
1510
5L M/2+3
Fig. 4 : Romain-la-Roche, petits canidés, dispersion selon la longueur (MD) de la M/1 et la longueur des deux molaires suivantes M/2+3 (d’après PoPlin, 1976).
Vulpes lagopus
Vulpes vulpes
L du trigonide
mm15
10
M/1mm
105
L M/1+2+3 = Romain-la-Roche
Vulpes lagopusactuel
Vulpes vulpes actuel
VL
Vulpes lagopusde Gönnersdorf
4 7 85 6
5
6
4
3
M/2mm
MD mm
= Romain-la-Roche
Fig. 5 : Romain-la-Roche, petits canidés, dispersion selon la longueur du trigonide de la M/1 et la longueur de la portion broyeuse de la série jugale inférieure M/1+2+3 (d’après PoPlin, 1976).
Fig. 6 : Romain-la-Roche, petits canidés, dispersion selon la longueur (MD) et la largeur (VL) de la M/2 (d’après PoPlin, 1976).

502 a. ARgANt
Squelette post-crânien :colonne vertébrale : Deux vertèbres cervicales, atlas CAN.22 (Tabl. 4) et axis CAN.947 (Tabl. 5) bien conservées appartiennent à un même individu (même aspect, compatibilité des surfaces articulaires). Elles présentent la taille et les caractéristiques de celles de renards. Quelques petites différences par rapport au renard actuel laissent planer un doute quant à l’attribution à Vulpes vulpes ou à Vulpes lagopus :- longueur plus importante du corps vertébral de l’axis
du renard de Romain-la-Roche,- fort développement et fort relief de l’arête centrale et
des ailes de la face plantaire de cet axis.Je pense que l’on peut écarter le renard polaire à cause de la taille comparable à celle d’un fort renard mâle actuel. Ces différences seraient dans ce cas un élément de caractérisation du Vulpes vulpes de Romain-la-Roche.Membres : tabl. 6Les os longs entiers du renard de Romain-la-Roche permettent d’utiliser la fig. 20 de F. PoPlin (PoPlin, 1976, p. 33). Le radius (CAN. 26), les deux métacarpiens 5 (CAN. 1864, CAN.1785), le tibia (CAN.1637), tous complets, se placent incontestablement dans l’aire de répartition des valeurs de Vulpes vulpes, souvent au niveau des valeurs supérieures et toujours hors de la zone de chevauchement avec Vulpes lagopus.L’étude des carnivores de gerde (Hautes-Pyrénées) de A. Clot (Clot, 1980) livre également de nombreuses données et comparaisons. L’humérus de Romain-la-Roche (CAN. 1700) s’intègre bien à la population de Vulpes vulpes de la fig.64, p. 134 (DT min. diaphyse / DT distal). Il en est de même pour le radius (CAN.26), fig. 65, p. 135 (Dt méd. / DAP. dist.). Les tibias (CAN. 1782, CAN. 1028) se retrouvent dans les valeurs supérieures de Vulpes vulpes de la fig. 71, p. 141 (DAP. dist. / DT dist.). Les calcanéums complets (CAN.27, CAN.1018) se répartissent aussi dans l’ensemble Vulpes vulpes (Dt max. / Longueur) de la fig. 72, p. 142 (Clot, 1980).Le renard roux de Romain-la-Roche apparaît de plus petite taille que le renard actuel, plus gracile, mais le tibia (CAN.1637) est un peu plus long et moins massif que
celui d’un renard actuel de bonne taille. Il faut peut-être y voir des membres plus allongés. Quelques différences morphologiques méritent d’être signalées.cubitus : tout en étant de taille inférieure à celui d’un fort renard mâle de Bourgogne, les dimensions accessibles dépassent les valeurs rencontrées sur Vulpes lagopus. La morphologie de l’extrémité proximale, bien représentée car très résistante, reste semblable à celle de Vulpes vulpes actuel, à la différence de l’incisure radiale où s’insert l’extrémité proximale du radius, qui présente un Dt plus faible. La pointe interne qui la limite possède un plus faible relief et un moindre développement.
II.1.2.2. Biochronologie :Le Vulpes vulpes jansoni (boniFay, 1971) de l’Escale (Saint-Estève-Janson, Bouches-du-Rhône) du Pléistocène moyen médian marque une des premières apparitions de Vulpes vulpes. On rencontre le renard roux également à la Caune de l’Arago (CRégut, 1979), à Es taliens (Clot & duRanthon, 1990), à la grotte Vaufrey (Dordogne). Le renard de Lunel-Viel (Hérault) (boniFay, 1971), stades isotopiques 13, 14 ou 15 (Interglaciaire « Mindel-Riss ») correspond probablement au renard roux mais de taille plus forte qu’à l’Escale. Cette espèce est aussi bien représentée en Espagne à La Sima de los Huesos (n = 23 individus) (gaRCia, 2003), gisement daté d’au moins 530 000 ans (bisChoFF et al., 2007). Vulpes vulpes est
Tableau 4 : Romain-la-Roche, Vulpes vulpes, vertèbre cervicale atlas (Pemac 30, schéma de mesurage idem Pfema 30 de Panthera spelaea, Tableau 84).
Vulpes vulpesAtlas, pemac. 30 1 2 3 4 5 6 7 8
CAN.22 17,1 47,8 23,6 29,6 21,5 19,8 14,3 8,7
1- DDP max. 5- Dt articulaire caudal2- Dt max. 6- DAP max. interarticulaire3- DAP max. 7- DAP interarticulaire min. ventral4- DT articulaire crânial 8- DAP interarticulaire min. dorsal
Tableau 5 : Romain-la-Roche, Vulpes vulpes, vertèbre cervicale axis (Pemac 31, schéma de mesurage idem Pfema 31 de Panthera spelaea, tableau 85).
Vulpes vulpes Axis
pemac. 311 2 3 4 5
CAN.947 28,3 24,3* 39,6 20,4 12,5
1- DDP max. 4- DT articulaire crânial2- Dt max. avec apophyses 5- Dt articulaire caudal3- DAP max.

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 503
régulièrement signalé à la grotte Vaufrey depuis le stade 9 (delPeCh, 1989). On le trouve par la suite au Lazaret (Alpes-Maritimes), à Châtillon-Saint-Jean (Drôme), à gerde (Hautes-Pyrénées) par exemple. Au Pléistocène supérieur c’est un élément banal de la faune.Le renard roux de Romain-la-Roche comparé à Vulpes vulpes actuel apparaît plus gracile, avec une denture plus coupante, un condyle d’articulation de la mandibule plus massif et possède sans doute des membres plus allongés, plus fins. Ceci le distingue des formes du Pléistocène supérieur.
II.1.2.3. paléoécologie :L’habitat du renard roux reste très variable, dans ou
à proximité des forêts, des broussailles ou de toute végétation haute. Son aire de répartition est sans doute une des plus vaste de toutes celles des mammifères sauvages actuels. Il occupe toute l’Europe, la plupart des îles. Il est largement présent en Amérique du Nord, en Asie. Il se rencontre même dans des agglomérations. Il n’est pas possible de tirer des informations précises de sa présence dans les gisements, si ce n’est l’existence d’un couvert végétal assez dense et d’un climat tempéré, éventuellement tempéré-froid. Le renard roux peut avoir un impact direct sur d’autres espèces de carnivores qu’il tend à évincer pour éviter la concurrence directe, par exemple belettes, hermines et Vulpes lagopus (aRtois, 1988).
Tableau 6 : Romain-la-Roche, Vulpes vulpes, matériel post-crânien.
Vulpes vulpesSquelette post crânien
1 2 3 4 5 6 7 8 9 10 11 12 13 14
l dt prox.
dAp prox.
dt méd.
dAp méd.
dt dist.
dAp dist.
Humérus CAN. 1700 - - - 8,21* 8,53* 19,40 15,43*
cubitus
CAN. 1472 - 6,71 15,72 - - - - 6,78 13,74 8,77 11,46 6,05CAN. 1182 - - - - - - - - - 8,60 11,47 5,30CAN. 1157 - 7,67 17,17 7,18 5,34 - - 5,77 14,91 8,96 11,70 5,74CAN. 28 - 7,01 16,51 - - - - 5,76 14,89 9,06 11,22 5,94CAN. 1516 - 6,73 15,60 - - - - 6,00* 13,41 8,05 11,02 5,62CAN. 427 - 6,94 15,26 - - - - 5,86 13,41 8,89 11,63 5,75
radiusCAN. 26 120,35 11,99 7,65 8,94 5,20 15,64 8,73CAN. 949 - 11,79 7,44 9,55 5,24 - -
Mtc.5CAN. 1864 46,10 7,88 6,89 5,27 3,86 7,14 6,08CAN. 1785 44,72 7,60 6,56 5,00 4,08 7,33 6,35
Fémur
CAN. 1176 - 27,72 - - - - - 12, 43 11,97CAN. 1791 - - - - - - - 12,65 11,36CAN. 1791 - - - - - 21,40 23,22CAN. 1790 - - - - - - 22,39
tibia
CAN. 1637 160,0* 25,32 27,17 10,25 10,99 - 12,25CAN. 1027 - 22,00 25,08 9,41 8,62 - -CAN. 1774 - 23,85 26,03 - - - -CAN. 1782 - - - - - 15,83 11,32CAN. 1028 - - - - - 16,33 11,29
calcanéum
CAN. 1743 - 10, 67 11,62 - - - -CAN. 27 33,16 12,68 11,91 5,32 9,16 8,54 10,15 20,90CAN. 1018 32,47 12,76 11,80 5,06 8,76 7,57 9,95 21,27CAN. 1005 30,94 - - 5,08 - 7,54 8,24 19,91CAN. 1002 - 13,10 11,59 6,01 8,90 - - -
Astragale CAN. 1485 19,40 12,39 10,28 9,60 6,30 6,12 8,41
cubitus :2 - Dt maxi. olécrâne (serrage de l’ensemble)3 - DAP maxi. olécrâne-processus anconé10 - Dt sommet de l’olécrâne11 - DDP sommet de l’olécrâne12 - DT petite cavité sigmoïde (A’N : hue, 1907)13 - DAP cavité sigmoïde (AA’ : hue, 1907)14 - DT processus anconé
Fémur :8 - Dt boule articulaire, ext. proximale9 - DAP boule articulaire, ext. proximalecalcanéum :2 - Dt max.3 - DAP max.5 - DAP min.8 - L. du manubrium
Astragale :2 - Dt max.3 - DDP max.4 - DT articulaire poulie5 - Dt min. col6 - DAP tête7 - Dt tête

504 a. ARgANt
II.1.3. genre Canisespèce Canis lupus Linné, 1758,
Le site de Romain-la-Roche livre un important matériel de canidés largement dominé par Canis lupus, le loup. Ce sera à l’avenir un solide jalon dans l’histoire de cette espèce. Il existe également, avec certitude, un peu de Cuon alpinus. La distinction des deux espèces sur du matériel dentaire ou squelettique isolé reste délicate. Il n’est pas impossible que certains éléments de ce dernier se retrouvent attribués par défaut à Canis lupus.
II.1.3.1. données morphométriques :La liste d’inventaire du matériel disponible apparaît en
annexe à la fin (NR = 1616, NMI = 20 demi-mandibules gauches.crâne : Le matériel crânien du loup de Romain-la-Roche est bien représenté mais hélas très fragmenté. Il n’y a aucun crâne complet. Malgré tout, ce matériel apporte de nombreuses informations morphométriques (tabl. 7). Il s’agit d’un loup de petite taille, au museau plutôt allongé et à la denture puissante. La détermination du sexe des individus selon les critères habituels comme le développement de la crête sagittale n’a pu se faire à cause de l’état de fragmentation du matériel crânien.denture supérieure : Tabl. 8, Fig. 8-1, Fig. 8-3, Fig. 8-5, Pl. I, fig. 5-6 (CAN.842, CAN.631)Incisives : la pièce CAN.1914 (Pl. I, fig. 3), fragment
Tableau 7 : Romain-la-Roche, Canis lupus, mesures du crâne. Pemac 10 reprend et complète OStEO canidés (desse et al., 1986). Les numéros de transfert (Nt) correspondent aux différents plans de mesurage d’OStEO.
crânes pemac 10 nt
Canis lupus
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
CAN.372Code 3 123,3 140,1 105,6 104,4 111,9 110,2 39,3 37,6Code 4 59,5 46,3Code 6 82,5 103,2 25,4 62,9
CAN.1703Code 4 73,2* 52,6
Code 6g 83,0 102,4 27,3 65,0d 81,9 98,5 27,2 61,7
CAN.378 Code 6 89,6A
104,9 26,4 70,5A
CAN705 Code 6 76,5A
g92,3
A
g24,3*
CAN.620
Code 3 125,3 107,0*
Code 6d
79,2A
d95,3
A
d23,3
d63,1
CAN.842 Code 6 59,7A
CAN.1274 Code 6 78,9 92,3 21,3 62,8nt 18 19 20 21 22 23 24 25 26
CAN.791 Code 3 69,1 43,7 24,1 16,3
code 3 :1 - Longueur totale acrocranion-prosthion2 - Longueur condylobasale3 - Longueur basale basion – prosthion4 - Longueur basicrâniale basion-synsphénion5 - Longueur basifaciale synsphénion-prosthion6 - Longueur du neurocrâne basion-nasion7 - Longueur du neurocrâne acrocranion-point frontal médian8 - L. du viscérocrâne nasion-prosthion9 - L. faciale point frontal médian-prosthion
10 - L. maxi. des nasaux nasion-rhinion 12 - L. du museau13 - L. médiane du palais staphylion-prosthion14 - L. du palais15 - L. staphylion-palatinooral16 - L. horizontale du palais (cf. mesure 14)18 - Dt otion-otion20 - Dt max. aux condyles occipitaux22 - Dt max. du trou occipital23 - H. du trou occipital basion opisthion
code 4 :3 - Dt min. entre les orbites entorbital-entorbital6 - Dt du palais aux alvéoles des canines
code 6 :1 - L. des dents jugales2 - C-M3/ (bord oral de C, extrémité aborale de M3/)3 - L. des molaires4 - L. des prémolaires

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 505
d’os incisif d. encore pris partiellement dans de la brèche, a conservé en place l’ensemble des trois incisives et la canine. Elle montre les trois incisives I1/, I2/, I3/ légèrement usées. On peut en observer la forme typique avec la collerette un peu dentelée en relief tout autour de la face occlusale de la couronne. Les racines des deux premières I1/ et I2/ visibles montrent un aplatissement latéral et une terminaison tronquée. La I3/ possède la couronne caniniforme caractéristique avec un bourrelet nettement marqué, sauf en face externe et un pincement antéro-interne laisse apparaître une petite échancrure à la jonction avec l’arête antérieure limitant cette couronne. L’usure toujours forte sur des incisives de loup est maximum pour I1/.canine supérieure :Les canines supérieures constituent un lot conséquent (44 dents, 25 C sup g et 19 C sup d, NMI = 25 individus). La Figure 8-1 montre la répartition MD/VL des C sup du Canis lupus de Romain-la-Roche. Leur dispersion en deux ensembles harmonieusement répartis (faible chevauchement) indique une bonne répartition entre mâles et femelles. Les deux sexes apparaîssent bien représentés. La domination apparente des mâles dans ce cas sera inversée pour les canines inférieures. Il faut y voir le rôle du hasard des chutes dans l’aven et de la conservation des vestiges.prémolaires :p1/ - CAN.790 est une dent simple, d’assez forte taille, unicuspide, uniradiculée, courte et massive. La couronne dissymétrique montre une partie externe largement développée, enveloppante sur l’avant. Un plan presque vertical scinde cette couronne en deux et la partie linguale, réduite, possède un rebord incliné marquant la base. Un faible bourrelet souligne ce bord lingual en déterminant un petit pointement à l’avant, au contact de l’arête antéro-interne et un pincement à l’arrière. On ne peut confondre cette première prémolaire avec son homologue inférieure bien différente.p2/ et p3/ - Ces prémolaires supérieures se distinguent des inférieures équivalentes par une plus grande longueur d’où l’impression d’étroitesse qu’elles donnent. toutes les deux (P2/ et P3/) présentent un paracône élevé, abrupt à l’arrière, plus particulièrement sur P3/. Sa partie antérieure se termine par une arête fortement déjetée côté lingual, beaucoup plus que sur les prémolaires inférieures. La pente antérieure du protocône est raide sur P2/ (angle d’environ 45°) mais plus longue et plus progressive sur P3/ (angle d’environ 30°). Dans les deux cas l’arrière de la couronne porte deux cuspides qui pointent, une à mi-pente, l’autre plus réduite à l’extrémité postérieure de la couronne. Cette dernière amorce un cingulum net sur P3/ mais plus atténué sur P2/, qui court côté lingual jusqu’au contact avec l’arête antérieure du paracône. On note sur la P3/ une inflexion importante vers le bas au niveau du bord antérieur de la racine postérieure avec une striction et un sillon à l’arrière de ce décrochement. Ce caractère sans être totalement absolu apparaît comme
une constante de la P3/ du loup de Romain-la-Roche (23 observations sur 29 possibles).p4/ - Pl. I, fig. 4 (CAN.921)C’est une dent puissante, au talon antéro-interne (protocône) nettement marqué malgré une certaine variabilité allant du petit pointement séparé en relief, à un simple élargissement de la couronne en pente régulière. Le paracône bien développé, fortement redressé, domine le métacône. A l’avant une arête que gomme progressivement l’usure, court jusqu’au bord antérieur. Une courte ramification s’en détache pour rejoindre le tubercule antéro-interne. La couronne est parfois bordée par un faible renflement côté vestibulaire mais pas toujours. Côté lingual un cingulum net souligne le métacône en remontant classiquement jusqu’à sa terminaison postérieure.M1/ - Le paracône et le métacône constituent un bloc vestibulaire puissant, à l’aspect triangulaire aux deux extrémités. Le paracône apparaît nettement plus volumineux et domine largement le métacône en hauteur. Un bourrelet modérément développé souligne la base de la couronne côté vestibulaire et se termine sous forme d’un liseré côté antérieur du paracône et côté postérieur du métacône. Un resserrement médian diminue fortement la largeur de la dent. Le talon interne modérément arqué paraît beaucoup moins large que le bloc vestibulaire. Une cuvette profonde et peu étendue, marque le centre de cette molaire. Elle est limitée par un protocône bien présent, mais souvent usé dans sa partie la plus élevée. L’hypocône participe au relief qui constitue la bordure linguale, nette, ceinturant tout le talon. Cette morphologie apparaît conforme à ce que l’on attend d’une première molaire de loup mais l’aspect général reste moins massif, plus tranchant que chez les loups de la dernière glaciation (OIS. 4, 3, 2).M2/ - La structure de la M2/ rappelle évidemment celle de la M1/ précédente mais avec un développement moindre du bloc paracône – métacône. Ce massif paraît peu développé, toujours dominé par le paracône. On ne retrouve plus le resserrement médian de la couronne, ni l’aspect anguleux du bloc vestibulaire fortement arrondi. Le protocône se situe au milieu du talon interne. L’hypocône est réduit à un simple rebord lingual de la couronne.demi-Mandibule : Tabl. 9, Fig. 7, Pl. II, fig. 1-2 (CAN.1187)Elles sont longues et relativement fines. Les plus fortes, hautes et plus massives, correspondent évidemment aux mâles. Les prémolaires ont la place de se positionner sans chevauchement. Il existe toujours un diastème d’environ 8 à 10 mm entre la première prémolaire et la canine.J’ai recherché sur un lot important les différents points analysés habituellement :- toutes les demi-mandibules ou fragments susceptibles
de livrer l’information montrent la présence de la P/1, à une exception près CAN.1318 qui n’en possède pas. Un long diastème de 14 mm sépare seulement la canine de la P/2.

506 a. ARgANt
Tableau 8 : Romain-la-Roche, Canis lupus, denture supérieure.
Canis lupusdenture
supérieureMd Vl Hc Ht Vltalon
I
I1/
n n = 17 n = 16 n = 2 n = 1min.-max. 6,4-8,9 5,3-8,4 10,3-12,8moy. 7,35 6,62 11,54 22,70écart-type 0,71 0,81 1,71 -
I2/
n n = 13 n = 13 n = 6 n = 5min.-max. 6,8-9,2 7,2-8,5 12,7-15,2 24,7-26,8moy. 8,2 7,76 13,37 25,71écart-type 0,69 0,44 0,95 0,91
I3/
n n = 43 n = 41 n = 15 n = 11min.-max. 8,5-10,8 6,7-8,4 12,6-16,0 28,8-33,6moy. 9,68 7,57 14,74 31,14écart-type 0,67 0,42 1,02 1,38
c c. sup.
n n = 43 n = 41 n = 18 n = 15min.-max. 10,3-13,8 6,4-8,9 18,5-27,0 43,5-53,6moy. 12,44 7,72 23,36 49,89écart-type 1,02 0,59 2,33 2,92
pM
p1/
n n = 18 n = 18 n = 12 n = 9min.-max. 7,0-9,8 5,0-6,3 6,2-8,6 16,0-20,0moy. 7,86 5,38 7,32 18,39écart-type 0,68 0,36 0,73 1,21
p2/
n n = 34 n = 31 n = 18 n = 8min.-max. 11,8-16,5 5,4-7,0 6,9-10,2 18,4-21,6moy. 14,01 6,17 8,04 19,86écart-type 0,98 0,46 0,74 1,08
p3/
n n = 30 n = 31 n = 12 -min.-max. 14,0-17,5 6,1-8,8 7,5-11,9 -moy. 15,89 7,32 8,63 -écart-type 1,00 0,79 1,20 -
p4/
n n = 42 n = 36 n = 15 - n = 36min.-max. 21,7-27,3 11,1-14,9 12,3-15,6 - 8,4-11,1moy. 24,23 12,68 14,26 - 9,48écart-type 1,33 1,08 0,98 - 0,63
M
M1/
n n = 52 n = 45 - -min.-max. 14,0-18,1 16,6-21,0 - -moy. 15,63 18,98 - -écart-type 1,00 1,21 - -
M2/
n n = 39 n = 39 n = 29 n = 4min.-max. 6,7-10,0 9,9-13,8 3,9-6,0 10,6-11,8moy. 8,44 11,77 5,25 11,12écart-type 0,73 1,03 0,50 0,67

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 507
- toutes les demi-mandibules ou fragments susceptibles de livrer l’information possèdent une troisième molaire ou une alvéole vide car cette molaire unicuspide tombe facilement au cours de la fossilisation (n = 44 observations possibles sur les demi-mandibules de canidés, 44 présences de M/3, y compris pour le cuon). Parfois la racine est cassée au ras de la mandibule et remplie de sédiment induré. Sans user l’os au Dremel en insistant et sans observation à la loupe binoculaire pour voir apparaître un indice de racine, il aurait été facile de conclure trop rapidement à une absence. Cette observation de la M/3 écarte à priori la possibilité de présence de demi-mandibules de cuon parmi le lot, ce dernier ne possèdant pas habituellement de troisième molaire inférieure bien que ce caractère ne soit pas absolu. Une M/3 peut bien se développer chez les cuons actuels comme le souligne stehlin (dubois & stehlin, 1933) dans l’étude sur le cuon de Cotencher (MajoR, 1900) et sa présence est parfois signalée chez des formes fossiles au cours du Pléistocène.
- la position du trou mentonnier a parfois été avancée comme moyen de distinguer le loup du cuon. Sur le matériel de Romain-la-Roche le trou principal le plus antérieur (souvent conique et oblique) se situe à la verticale du contact P/1-P/2 dans 13 cas sur 18 observations possibles et sous la racine antérieure de P/3 dans les 5 cas restants. Je ne l’ai pas observé sous la P/1. Ce n’est pas à mon avis un caractère discriminant car on retrouve sur les différentes figurations de
demi mandibules de Cuon alpinus des situations comparables.
- l’apophyse angulaire à la base de la branche montante présente sur le cuon une morphologie particulière des fosses ptérygoïdes supérieure et inférieure, l’expansion de la fosse inférieure repoussant l’attache du muscle masséter vers l’avant (gaRCia, 2003, fig. III.3.16, p.150). Sur le loup de Romain-la-Roche,
Tableau 9 : Romain-la-Roche, Canis lupus, mandibule, Pemac 20 (Fig. 7).
Canis lupusMandibulepemac 20
n min max moy écart-type
1 5 153,50 179,10 166,96 11,66 L. totale milieu condyle-infradental2 4 151,40 180,60 163,55 13,42 L. processus angulaire -infradental3 4 147,80 170,40 161,68 10,30 L. incisure angulaire-infradental4 6 131,24 158,70 145,86 11,94 L. condyle-C.post.5 8 127,86 151,10 140,55 9,08 L. incisure angulaire-C.post.6 7 131,71 160,70 147,92 10,73 L. processus angulaire-C.post.7 19 87,00 103,40 96,63 4,46 L. C.post.-M/3 post.8 20 84,00 97,77 91,17 3,82 L. P/1-M/39 24 78,80 89,44 84,13 3,36 L. P/2-M/310 30 37,50 50,74 43,23 3,12 L. des molaires11 28 46,20 54,70 50,43 2,00 L. des prémolaires P/1-P/412 31 38,30 48,70 43,68 2,17 L. P/2-P/415 3 27,30 28,80 28,03 0,75 L. de l’alvéole de la carnassière20 26 10,20 14,10 12,25 0,87 Dt de la mandibule derrière M/1 21 2 44,00 71,00 57,50 19,09 H. processus angulaire-coronion22 35 24,10 43,80 29,38 3,39 H. derrière M/123 30 20,70 34,60 24,86 2,91 H. entre P/2 et P/326 31 11,30 15,70 13,27 1,12 DT maxi 1/2 mdb entre P/4 et M/1
1
23
4
5
6
78
9
1011
21
2226
1223
Fig. 7
Fig. 7 : Pemac 30, plan de mesurage de la mandibule des canidés.

508 a. ARgANt
sur 8 observations possibles, je retrouve le type C de gasPaRd (1964) avec des fosses très nettes, aux limites anguleuses, mais rien qui rappelle le Cuon alpinus de la trinchera galeria d’Atapuerca (gaRCia, 2003, fig. III.3.23, p167). Le développement du processus angulaire reste très variable. Il est par exemple deux fois plus volumineux sur CAN.1215 que sur CAN. 1318. Les deux condyles d’articulation respectifs confirment cette variation de taille. La deuxième pièce (demi mandibule d complète) appartient manifestement à un fort mâle et la première (fragment d isolé) correspond probablement à une femelle aux muscles ptérygoïdes et masséters nettement moins puissants.
- La morphologie de la fosse massétérienne me semble un meilleur critère de distinction entre Canis lupus et Cuon alpinus. Elle est profonde sur le loup de Romain-la-Roche, s’avance en général jusque sous la M/3, parfois à la verticale de la limite M/2 – M/3. Par contre elle ne descend pas très bas et elle est bordée en limite inférieure par un relief régulier, assez large (10 mm environ). Elle ne va pas, en tout cas partiellement, jusqu’à la base de l’os mandibulaire comme elle le fait sur une mandibule de cuon actuel. L’axe longitudinal de cette fosse paraît par ailleurs plus oblique sur le loup par rapport au grand axe longitudinal de la branche horizontale de la mandibule. Il semble plus redressé chez Cuon alpinus (gaRCia, 2003, fig. III.3.23, p.167, photo b). Aucune des mandibules du loup de Romain-la-Roche pouvant fournir cette information ne correspond au type cuon.
- C’est toujours la P/2 qui est la plus usée à l’arrière du protoconide. On pourrait parfois croire à la non-existence d’un petit tubercule secondaire situé à mi-talon à l’arrière de la cuspide principale. Dans 6 cas (CAN.269, 455, 419, 898, 1467, 1119) la P/2 n’en possède cependant pas et on ne peut invoquer une quelconque usure. Dans deux cas supplémentaires (CAN.1196, 1072), il s’agit d’un simple pointement de la grosseur d’une tête d’épingle métallique. Dans les 16 autres cas observables ce tubercule est présent.
denture inférieure : Tabl. 10, Fig. 8-2, Fig. 8-4, Fig. 8-6Incisives : La demi-mandibule d CAN.1318 montre le bloc incisif complet avec I/1, I/2 et I/3. Une usure nette affecte ces trois dents, maximum pour I/3 (U = 1). La I/1 la plus petite, très étroite, à racine mince et très aplatie, possède la couronne faiblement divisée habituelle avec l’épaulement latéral qui s’appuie un peu en contrebas de la pointe principale, côté externe. La I/2 reproduit la même forme de couronne mais avec une taille nettement supérieure et une racine plus large, restant aplatie. L’épaulement latéral prend une ampleur croissante qui va s’amplifier encore sur la I/3 en donnant cette forme caractéristique en mitaine. La base interne de la couronne est soulignée simplement par un petit renflement à la base. On ne constate pas en tout cas pas de bourrelet continu bordant la couronne comme sur les incisives supérieures.
canine inf. :La canine inférieure apparaît moins puissante que la supérieure. Sa couronne, moins élevée, se courbe davantage et le profil longitudinal rappelle un S contrairement aux canines supérieures. La Figure 8-2 montre la répartition MD/VL de ces dents. La large dispersion des points traduit une forte variabilité et l’existence de quelques cas isolés mériterait un examen plus approfondi. Deux ensembles se distinguent cependant nettement. Ils traduisent bien la répartition entre mâles et femelles, somme toute assez équilibrée. prémolaires :p/1 :Elle se distingue facilement de la première prémolaire supérieure par sa plus petite taille et la forme dissymétrique de la couronne déséquilibrée par une cuspide haute, très à l’avant et une longue pente raide qui descend assez bas sur la racine côté lingual. Un petit pointement imperceptible en relief marque l’arrière de la couronne. C’est une dent uniradiculée.p/2, p/3, p/4 :Moins allongée que sur les prémolaires supérieures équivalentes, leur couronne paraît également moins déjetée du côté lingual à l’avant du protoconide. L’arête antérieure de ce tubercule principal possède cependant une légère courbure interne. Les trois dernières prémolaires sont construites sur le même type, la cuspide principale (protoconide) et à l’arrière, un pointement médian suivi d’un relief plus faible, en bord postérieur de la couronne. La taille des prémolaires augmente régulièrement de P/2 à P/4 de même que l’importance prise par les tubercules à l’arrière du protoconide, avec relèvement du talon postérieur. Côté lingual, un bourrelet marque la base de la couronne sans que l’on puisse parler vraiment de cingulum.Molaires :M/1 :La carnassière inférieure apparaît puissante. Elle possède un trigonide élevé dominé par le protoconide que flanque côté postéro-interne, un métaconide net atteignant au maximum la moitié de sa hauteur. Le talonide broyeur est constitué par un hypoconide volumineux côté vestibulaire qui occupe environ les deux tiers de la surface et qui finalement se trouve en position centrale compte tenu de la pente externe. L’entoconide constitue le rebord lingual du talon et est parfois constitué de deux cuspides (une fois 3) décroissantes en direction du métaconide (n = 17 cas sur 35 observables).M/2 : La deuxième molaire est nettement plus réduite que la M/1 tout en prolongeant directement son talon broyeur. Le relief antérieur transversal, constitué d’une barre paraconide-métaconide est précédé d’un bourrelet où se termine l’arête qui part du paraconide, arête visible seulement sur les dents d’usure nulle ou faible. Ce dernier tubercule, de loin le plus volumineux des deux, se retrouve pratiquement en position centrale à cause du développement latéral de la pente qui descend très bas

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 509
Tableau 10 : Romain-la-Roche, Canis lupus, denture inférieure.
Canis lupusdenture inférieure Md Vl Hc Ht
I
I/1
n n = 17 n = 13 n = 5 n = 5min.-max. 4,7-6,4 3,2-4,3 7,7-8,2 20,6-21,5moy. 5,41 3,84 7,89 21,25écart-type 0,46 0,31 0,22 0,40
I/2
n n = 19 n = 19 n = 3 n = 1min.-max. 5,9-8,5 4,8-6,2 8,4-12,2 25,96moy. 7,37 5,53 10,64écart-type 0,79 0,38 1,96
I/3
n n = 37 n = 37 n = 5 n = 4min.-max. 6,9-9,5 5,9-8,0 11,4-12,1 28,4-29,8moy. 7,53 6,99 11,63 28,92écart-type 0,58 0,60 0,54 0,59
c c. inf.
n n = 55 n = 55 n = 22 n = 14min.-max. 11,3-15,0 7,1-10,0 19,3-25,3 45,3-54,2moy. 13,03 8,42 22,48 48,58écart-type 0,98 0,72 1,61 2,79
pM
p/1
n n = 24 n = 24 n = 14 n = 2min.-max. 5,4-7,3 4,0-5,4 4,8-6,3 15,4-15,9moy. 6,22 4,65 5,50 15,68écart-type 0,44 0,37 0,48 0,35
p/2
n n = 43 n = 43 n = 19 -min.-max. 10,8-13,9 5,2-7,0 6,1-9,0 -moy. 12,34 6,07 7,91 -écart-type 0,85 0,45 0,67 -
p/3
n n = 32 n = 33 n = 13 -min.-max. 12,3-16,3 6,0-8,2 6,7-9,5 -moy. 14,01 6,92 8,34 -écart-type 1,12 0,74 0,78 -
p/4
n n = 46 n = 47 n = 18 -min.-max. 13,4-18,0 6,6-8,9 9,0-10,9 -moy. 15,61 7,76 9,47 -écart-type 1,11 0,62 0,60 -
M
M/1
n n = 53 n = 51 n = 29 -min.-max. 24,7-30,9 9,7-12,5 14,2-17,8 -moy. 27,23 10,91 16,09 -écart-type 1,35 0,67 0,86 -
M/2
n n = 49 n = 48 n = 22 -min.-max. 9,2-13,1 7,0-9,3 4,7-9,1 -moy. 10,8 8,02 6,65 -écart-type 0,79 0,53 1,10 -
M/3
n n = 7 n = 7 n = 4 -min.-max. 4,5-5,4 4,4-5,2 3,0-4,3 -moy. 5,02 4,71 3,68 -écart-type 0,35 0,30 0,61 -

510 a. ARgANt
en position antéro-externe sur la racine antérieure. Le métaconide, bien que net, reste cependant plus faible. L’hypoconide bien marqué prolonge le protoconide vers l’arrière en position encore presque centrale malgré un rétrécissement au niveau du talon. Un léger bourrelet termine la couronne côté lingual jusqu’au métaconide sans autre relief particulier. M/3 :C’est une toute petite molaire résiduelle, présente dans tous les cas où l’observation reste possible, en place (n = 4 cas), sous forme de racine cassée au ras de l’os mandibulaire (n = 8 cas) ou sous forme d’alvéole vide ou bien remplie de sédiment induré (n = 36). Elle est toujours uniradiculée. Sa couronne, pratiquement ronde, avec un faible relief central résultant de la conjonction de trois rides se termine par une faible dépression à l’arrière de la couronne.
Squelette post-crânien :colonne vertébrale : La colonne vertébrale fait rarement l’objet d’études. C’est dommage car les vertèbres, une fois déterminées avec précision (condition indispensable), peuvent comme tout autre élément squelettique apporter des éléments de détermination de l’espèce et de comparaison, en particulier de la taille des individus. A Romain-la-Roche les vertèbres de canidés de grande taille abondent, souvent très bien conservées.Vertèbres cervicales :Une vertèbre atlas CAN.1117 parfaitement conservée fournit toutes les dimensions (tabl. 11).Curieusement, l’axis (Pl. 5-5, CAN.526) apparaît bien mieux représentée, traduisant un NMI de 17 individus (tabl. 12).Les autres vertèbres cervicales se reconnaissent aisément par leur corps vertébral long et aplati, fortement arqué
Fig. 8 : Romain-la-Roche, Canis lupus, rapport MD/VL pour : 1- Canine supérieure ; 2- Canine inférieure ; 3- P4/ ; 4- P/3 ; 5- M1/ ; 6- M/1.
4 -Romain-la-Roche, Canis lupus , P3 inférieure5
5,5
6
6,5
7
7,5
8
8,5
9
12 12,5 13 13,5 14 14,5 15 15,5 16 16,5 17
MD en mm
VL e
n m
m

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 511
et remontant vers l’arrière. De nombreuses pièces sont inscrites dans l’inventaire en annexe. Je n’ai retenu que les mieux conservées, celles dont la détermination exacte est possible (tabl. 13). Un bloc de brèche lie l’axis CAN.451 à la 3e vertèbre cervicale d’un même individu. L’axis s’est simplement déboîté et a pivoté sur le côté en se tournant vers l’arrière.Vertèbres dorsales :Les vertèbres dorsales sont également nombreuses (voir inventaire en annexe). Comme précédemment, seules les plus complètes dont le rang est assuré, seront prises en compte à titre d’exemple (Tabl. 14).Vertèbres lombaires :Il n’est pas possible de mesurer toutes les vertèbres lombaires. Le tableau 15 fournit les données pour la plus complète de chaque catégorie (rang).Sacrum :trois pièces seulement correspondent à des sacrums de Canis lupus. Aucun n’est intact et seules les dimensions
de la surface articulaire crâniale peuvent être acquises (tabl. 16).Vertèbres caudales :De nombreuses vertèbres caudales correspondent aux canidés. Les apophyses en étoile bien caractéristiques ne se disposent pas exactement comme chez les félidés. Mais compte tenu de l’amincissement progressif de la queue et de la variation morphologiques des vertèbres les plus terminales, il est bien difficile d’être catégorique pour leur rang et même dans leur détermination. Je me contente de les inscrire à l’inventaire en annexe.ceintures :- Omoplate : Aucune omoplate ne s’est conservée
entièrement, rien de surprenant compte-tenu de la fragilité de ce type d’ossement. On ne rencontre que des extrémités articulaires plus ou moins étendues. Le tableau 17 fournit les indications métriques. Sur le plan morphologique se retrouvent toutes les caractéristiques des omoplates de Canis lupus. Seul le
Tableau 11 : Romain-la-Roche, Canis lupus, atlas.
Atlaspemac. 30 1 2 3 4 5 6 7 8
CAN.1117 29,0 90,7 47,5 47,2 34,5 37,2 12,1 20,2
CAN.982 28,9 - - 45,4 36,4 34,6 9,4 16,3
CAN.1809 - - - - 35,6 36,8 - 16,5
1- DDP max. 5- Dt articulaire caudal2- Dt max. 6- DAP max. interarticulaire3- DAP max. 7- DAP interarticulaire min. ventral4- DT articulaire crânial 8- DAP interarticulaire min. dorsal
Tableau 12 : Romain-la-Roche, Canis lupus, axis.
Axispemac. 31 1 2 3 4 5
CAN.539 43,2 - 71,7 33,7 20,2
CAN.690 47,6 - - 37,0 21,6
CAN.530 45,5 - 73,8 34,2 20,2
CAN.526 42,2 38,3 69,6 31,2 18,3
CAN.451 46,4 - 72,0 35,2 20,5
CAN.1810 - - 72,0 35,2 22,2*
CAN.954 42,0 - 70,0* 34,2 -
CAN.622 - - 78,0* 32,9 20,2
1-DDP max. 4- DT articulaire crânial2- Dt max. avec apophyses 5- Dt articulaire caudal3- DAP max.

512 a. ARgANt
Tableau 13 : Romain-la-Roche, Canis lupus, autres vertèbres cervicales.
Vert. cervicalespemac. 32 1 2 3 4 5 6 7
CAN.451 3 35,6 - 43,2* - 16,7* 34,8CAN.1053 3 - - - 16,5 19,4 33,8CAN.1726 3 52,0 40,0 17,6 18,6 34,6CAN.486 4 - 40,8 44,4 17,2 19,3 32,8CAN.1120 4 44,-5* 52,0* 45,0* 17,7 20,5 33,8CAN.1107 4 - - - 18,7 20,4* 31,5CAN.1118 5 - - 45,5 18,2 19,9 32,9CAN.572 6 - - 28,6 15,4 18,0 26,2CAN.1102 6 - - 38,4 18,0 20,0 33,2CAN.944 6 - - 38,5 16,2 18,7 32,3CAN.1121 7 - 52,8 33,2 17,0 26,0* 26,3
1- Rang de la vertèbre si connu 5- Dt articulaire crânial2- DDP max. avec apophyses 6- Dt articulaire caudal3- Dt max. avec apophyses. 7- DAP max. interarticulaire du corps vertébral (CV)4- DAP max. avec apophyses
Tableau 14 : Romain-la-Roche, Canis lupus, vertèbres dorsales.
V. dorsalespemac. 33 1 2 3 4 5 6 7
CAN.383 1 - 47,7* - 27,9 24,0* 22,0CAN.769 3 73,0 40,4* 38,8 25,4 27,3 19,3CAN.1491 4 - - - 25,2 26,4 20,0CAN.1778 5 - - - 23,9* 25,5 19,0CAN.577 6 56,5* - 52,0* 22,6 25,5 21,5CAN.1216 7 - - - 20,0 20,3 18,5CAN.522 8 50,0* - 53,5* 21,6 22,2 19,3CAN.1101 10 40,0* 34,0* 30,5* 21,4 23,8 21,4CAN.1104 11 - 33,8* - 20,0 22,9 22,5CAN.382 12 - - - 24,3 23,8 24,3CAN. 13 - - - 28,6 25,3 28,6
1- Rang de la vertèbre si connu 5- Dt articulaire crânial, fovea costale comprise2- DDP max. avec apophyses 6- Dt articulaire caudal, fovea costale comprise3- Dt max. avec apophyses 7- DAP max. interarticulaire (CV)4- DAP max. avec apophyses (en projection)

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 513
tubercule infraglénoïdal d’attache du triceps brachial, situé longitudinalement par rapport au rebord inférieur de la cavité glénoïde permet une remarque. Il présente sur le loup de Romain-la-Roche une forme triangulaire allongée à base étroite saillant nettement, sur tous les cas observables (n = 13).
- Coxal : Aucun os coxal du bassin n’est complet. La zone de l’acétabulum, plus résistante, s’est mieux conservée. Seuls les éléments assez complets pour permettre un minimum de mesures sont pris en compte dans le Tableau 18. Le coxal g CAN.495 ressort du lot (n = 13 g et 17 d) par sa petite taille et une massivité moindre. Il possède quelques petites différences morphologiques par rapport aux coxaux de Canis lupus et je l’attribue à Cuon alpinus europaeus. tous les autres éléments restent conformes entre eux et caractérisent le loup de Romain-la-Roche.
Membre antérieur :Humérus : Tabl. 19La taille reste moyenne et relativement constante pour tout le lot des humérus. La morphologie ne présente pas de différences notables avec celle de Canis lupus. Seule la tubérosité deltoïdienne paraît relativement moins
large et descend un peu moins bas sur la diaphyse du loup de Romain-la-Roche. Le foramen supratrochléaire caractéristique des canidés apparaît toujours très nettement.Radius : Tabl. 20trois radius complets seulement nous renseignent sur la longueur. On retrouve la morphologie habituelle de Canis lupus. On peut simplement souligner sur tous les cas d’observation possible, la netteté et le relief de l’arête de l’attache musculaire du muscle carré pronateur qui court le long de la diaphyse en position latérale externe (baRone, 1976, p. 293 fasc. 2).Cubitus : Tabl. 21Un seul élément entier (CAN.1158) nous permet de connaître la longueur de cet os chez le loup de Romain-la-Roche. Deux autres cubitus (CAN.464, CAN.725) sont presque complets et la partie manquante peut être valablement estimée sur des bases sûres. Rien d’étonnant à la fragmentation de la plupart des cubitus dont la longueur et la finesse rendent la diaphyse particulièrement fragile. La partie proximale par contre présente une grande résistance. Elle reste particulièrement difficile à caractériser métriquement. Morphologiquement, ce matériel correspond classiquement à Canis lupus. A Romain-la-Roche, il n’atteint ni les dimensions, ni la puissance des grandes formes du Pléistocène supérieur. Carpe : Tabl. 22Métacarpiens : Tabl. 23Les métacarpiens du loup de Romain-la-Roche sont nombreux, souvent fragmentés mais les pièces complètes ne sont pas rares. La morphologie en est celle des canidés, diaphyses droites, allongées et à bords parallèles pour les deux rayons centraux, légèrement arquées vers l’extérieur pour les deux rayons latéraux. Le métacarpien 1 correspond au doigt 1 latéral, vestigial chez les canidés.
Tableau 15 : Romain-la-Roche, Canis lupus, vertèbres lombaires.
Vert. lombairespemac. 34
1 2 3 4 5 6 7
CAN.593 1 - - - 22,1 23,9 30,0CAN.1123 2 46,9 - - 26,1 28,4 35,2CAN.400 3 50,2 - - 23,3 24,5 29,5CAN.443 4 51,7 - - 22,3 24,15 29,1CAN.1291 5 53,4 64,5* 44,0 23,3 26,4 32,7CAN.1930 6 48,8 - - 25,5 27,8 33,4CAN.994 7 - - - 25,0 26,6 25,4
1- Rang de la vertèbre si connu 5- Dt articulaire crânial2- DDP max. avec apophyses 6- Dt articulaire caudal3- Dt max. avec apophyses 7- DAP max. interarticulaire (CV)4- DAP max. avec apophyses (en projection)
Tableau 16 : Romain-la-Roche, Canis lupus, sacrum.
Sacrumpemac. 35
5dt articulaire
crânial
6dAp articulaire
crânialCAN.1149 28,5 12,6CAN.739 28,8 12,4CAN.1225 24,1 11,9

514 a. ARgANt
Tableau 17 : Romain-la-Roche, Canis lupus, omoplate.
Canis lupusOmoplatepemac 40
5dAp glénoïdal avec processus coracoïde
6dAp articul.
7dt articul.
8dt min. col
n 9 12 14 9
min 31,1 26,5 19,0 28,3
max. 37,8 33,0 22,2 31,5
moy. 34,71 29,65 20,61 30,11
écart-type 2,13 2,04 0,92 1,23
Tableau 18 : Romain-la-Roche, Canis lupus, coxal.
Canis lupuspemac 41coxal
1 2 3 4 5 6 7 8 9 10 11 12 13
n 1 20 4 - - - 21 - 18 19 14 18 18
min-max - 21,5-26,0-
28,7-33,6 - - - 14,0-
18,3 - 22,4-26,9
21,4-24,2
17,4-20,8
9,5-12,3
8,4-13,4
moy. 161,7* 24,17 31,65 - - - 16,95 - 25,04 23,06 19,03 10,75 10,23écart-type - 1,22 2,41 - - - 1,22 - 1,64 0,81 1,06 0,77 1,05
1- L. max.2- DAP acétabulum3- DAP foramen obtus4- L. symphyse pubienne5- Dt min. bassin complet au niveau de la petite échancrure
sciatique6- Dt min. milieu bord sup. des 2 cavités cotyloïdes
7- Dt épine sciatique au milieu de la cavité cotyloïde8- Dt min. bassin complet au niveau de la grande
échancrure sciatique9- DDP acétabulum10-DDP min. pédoncule de l’ilion11-DDP min. pédoncule de l’ischion12-Dt pédoncule de l’ilion13-Dt pédoncule de l’ischion
Tableau 19 : Romain-la-Roche, Canis lupus, humérus.
Canis lupusHuméruspemac 50
1l
2dt prox.
3dAp prox.
4dt méd.
5dAp méd.
6dt dist.
7dAp dist.
n 5 10 10 38 25 27 31min-max 177,4-183,8 31,7-40,2 44,8-52,9 13,1-15,8 14,9-21,0 29,2-42,3 26,9-35,6moy . 180,84 34,69 47,45 23,08 18,19 37,54 30,24écart-type 2,41 2,33 2,32 1,46 1,46 2,86 2,06

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 515
Tableau 20 : Romain-la-Roche, Canis lupus, radius.
Canis lupus radius
pemac 51
1l
2dt prox.
3dAp prox.
4dt méd.
5dAp méd.
6dt dist.
7dAp dist.
8dt articul.
dist.n 3 29 29 14 14 15 18 12
min-max 176,7-198,2 18,5-23,6 11,8-15,6 13,3-17,8 9,0-10,9 24,2-31,3 13,3-16,4 18,6-22,6
moy . 186,13 21,02 14,04 15,19 10,11 28,41 15,17 21,07écart-type 10,98 1,43 1,06 1,12 0,56 1,75 0,93 1,23
Tableau 21 : Romain-la-Roche, Canis lupus, cubitus.
Canis lupus cubitus pemac 52
1l
2dt art. prox. min.
3dAp prox. max.
4dt
méd.
5dAp méd.
6dt dist.
7dAp dist.
8dt
articul. prox. max.
9dAp art.
prox.
10dt
olécrâne prox.
11
n 3 31 17 7 7 - - 25 23 14 40
min-max 209,1-220,3 7,4-11,1 26,4-33,8 7,2-10,5 8,5-11,9 6,9-8,2 9,6-10,5 13,6-22,9 17,4-24,4 10,0-16,7 22,3-30,1
moy. 215,8 9,11 29,88 9,01 10,11 - - 18,25 20,97 13,38 25,59écart-type 5,91 0,96 2,32 1,11 1,32 - - 2,31 1,72 1,61 1,97
11 - DAP max. au bord distal de l’incisure trochléaire (serrage direct)
Tableau 22 : Romain-la-Roche, Canis lupus, carpe.
Canis lupuscarpe
pemac 53
1ddp
(hauteur)
2dt
(largeur)
3dAp
(longueur)
4 5 6
Scapholunairepemac 531
n 5 4 5 5 5min-max 11,3-14,2 21,9-22,7 13,8-16,1 17,0-19,9 13,0-14,7
moy. 12,65 22,30 14,57 17,78 13,80écart-type 1,27 0,35 1,01 1,21 0,78
pisiformepemac 532
n 8 8 8 8 8 8min-max 17,6-20,5 9,3-10,9 11,1-13,0 8,4-10,14 10,0-12,8 7,8-9,9
moy. 18,98 9,83 12, 9,40 10,89 8,55écart-type 0,94 0,48 0,62 0,50 0,89 0,67
Pyramidalpemac 533
n 3 3 3min-max 8,0-10,6 11,1-15,5 11,4-16,0
moy. 9,34 12,92 13,48écart-type 1,26 2,31 2,36
unciformepemac 534
n 6 6 6min-max 13,2-15,0 11,7-13,6 10,3-12,1
moy. 13,98 12,49 11,37écart-type 0,70 0,75 0,74
Magnumpemac 535
n 1 1 1min-max 12,6 10,9 19,4
Scapholunaire : 4- DT articulaire distal 5- DAP articulaire distal
pisiforme :4- DT dist.5- DAP dist.6- Dt min. col

516 a. ARgANt
Il se reconnaît à sa faible longueur qui le distingue des autres métacarpiens, à sa faible largeur avec un aspect déprimé latéralement et à la forme très particulière de ses deux extrémités. La partie proximale possède une forme globalement triangulaire munie d’une profonde encoche articulaire répondant au trapèze. L’extrémité distale se termine en biseau dont le pan coupé regarde vers l’extérieur et s’adapte à la forme caractéristique de la partie proximale de la première des deux phalanges du pouce.Membre postérieur :Fémur : Tabl. 24Quatre fémurs complets fournissent toutes les dimensions. Les extrémités proximales ou distales, très robustes, apparaissent nombreuses. On retrouve la morphologie habituelle des canidés et de Canis lupus en particulier. Signalons simplement la largeur et la pente régulière, dans sa partie basse, de la surface allant de la fosse trochantérique jusqu’au niveau du petit trochanter avec la base de la crête intertrochantérique comme limite latérale externe, semblant correspondre à un caractère général (sur tous les cas observables).Rotule : Tabl. 25Je n’ai trouvé que trois rotules dans le matériel. C’est un os court, épais, plein et massif. Il n’y a pas de raisons qu’il n’y en ait pas plus au vu des séries des fémurs et des
tibias. C’est par contre un élément de petite taille qui peut échapper facilement à la fouille ou à la détermination.Tibia : Tabl. 26, Fig. 9C’est le tibia qui présente le maximum d’éléments entiers (n = 9). Nous retrouvons les caractères habituels des tibias de Canis lupus. A noter l’importance de la crête tibiale haute, avec une rupture brutale en raccord avec la diaphyse (type A) dans 10 cas sur 17 observations possibles. Dans les autres cas (type B) la crête est moins haute avec un raccord en pente progressive. La première observation semble s’appliquer aux tibias les plus robustes, mais pas toujours les plus longs (CAN.1927 par ex.). Faut-il y voir un caractère sexuel secondaire lié au développement musculaire du biceps fémoral plus puissant chez les mâles ? Pour le vérifier j’ai mis en relation la longueur du tibia au périmètre de la partie médiane de la diaphyse pour déterminer un indice de robustesse, mis ensuite en rapport avec le type A ou B de la crête tibiale. Pour déterminer le périmètre de cette diaphyse j’utilise un mince ruban de toile adhésive qui entoure le milieu strict du tibia (longueur divisée par 2) puis le déroule et mesure sa longueur en le collant sur la table. C’est la technique mise au point par F. PRat pour mesurer le périmètre au collet des canines et des I3/ de l’ours de La Romieu (PRat & thibault, 1976). Les tibias étant les os longs les mieux conservés, il devient possible de faire un
Tableau 23 : Romain-la-Roche, Canis lupus, métacarpiens.
Canis lupusMétacarpiens
pemac 54
1l
2dt prox .
3dAp prox .
4dt méd.
5dAp méd.
6dt dist.
7dAp dist.
8dt art. dist.
Mtc.1pemac 541
n 5 5 4 5 5 6 5 6min-max 24,5-27,8 5,0-7,2 6,0-6,9 3,5-4,5 4 ,4-5,9 5,8-7,3 5,7-6,3 6,0-7,4
moy. 25,75 5,71 6,49 3,95 5,11 6,74 6,05 6,90écart-type 1,26 0,87 0,40 0,41 0,61 0,58 0,35 0,51
Mtc.2pemac 542
n 4 10 10 6 6 5 5 5min-max 61,9-71,5 8,3-10,2 10,6-13,9 7,3-8,8 5,6-8,6 10,1-11,0 8,9-10,5 9,9-10,5
moy. 66,27 9,38 12,48 7,88 6,60 10,67 9,79 10,38écart-type 4,44 0,60 0,97 0,48 1,11 0,37 0,61 0,29
Mtc.3pemac 543
n 7 12 12 11 10 6 6 6min-max 71,3-82,4 8,8-11,0 10,5-13,9 6,8-8,8 5,2-8,0 9,4-11,0 9,8-11,5 8,5-9,8
moy. 77,35 9,88 12,73 7,72 6,11 10,39 10,97 9,26écart-type 4,38 0,67 0,91 0,60 0,79 0,56 0,59 0,43
Mtc.4pemac 544
n 5 10 12 10 10 6 5 6min-max 67,6-80,5 7,8-9,2 10,6-12,8 6,5-8,1 5,7-7,0 9,2-10,6 9,9-11,4 9,0-9,9
moy. 73,14 8,60 12,01 7,00 6,32 10,04 10,55 9,11écart-type 4,66 0,50 0,63 0,47 0,45 0,53 0,52 0,42
Mtc.5pemac 545
n 6 13 13 11 11 11 10 11min-max 60,5-70,2 11,1-13,3 10,4-12, 8 6,6-8,3 5,3-7,5 10,3-12,2 9,2-10,9 10,1-11,5
moy. 63,59 12,25 11,41 7,54 6,34 11,08 10,22 10,62écart-type 3,67 0,65 0,75 0,54 0,59 0,62 0,54 0,49

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 517
diagramme de répartition sur 9 exemplaires auxquels je rajoute les deux tibias du même individu de Cuon alpinus europaeus (Fig. 9). Les types A et B de la crête tibiale se répartissent manifestement sans classement, sans rapport véritable entre la crête et le sexe de l’animal. Il faut donc conclure à un caractère individuel variable. Par contre les deux tibias de cuon se distinguent et confirment le bien fondé de leur détermination. Notons également que des restes ou des traces plus ou moins développées de péroné soudé au tibia (en position centrale ou extrémité distale) s’observent dans 3 cas sur 35 observations possibles. Cela arrive plus fréquemment chez les canidés que chez les autres carnivores.Péroné : Tabl. 27C’est un os long particulièrement fragile. Aucun n’est complet. Seules trois extrémités distales, les plus caractéristiques, ont été cataloguées au listing des Canidés de Romain-la-Roche.Tarse : Le calcanéum (tabl. 28) et l’astragale (tabl. 29) représentent l’essentiel du tarse du loup de Romain-la-Roche. Ce sont des pièces robustes, facilement reconnaissables par rapport aux autres éléments, ce qui a dû faciliter leur incorporation dans le listing La pièce CAN.1294 réunit le calcanéum d. et l’astragale d’un même individu, soudés par de la brèche en position anatomique et en parfait état de conservation (Pl. II, fig. 3, 4, 5). Les autres éléments du tarse apparaissent plus rarement (tabl. 30).Métatarsiens : Tabl. 31Les métatarsiens sont bien représentés comme l’étaient les métacarpiens. Ils apparaissent, au niveau des moyennes, toujours plus longs et plus massifs que ces
derniers à de rares exceptions près pour les extrémités distales plus fortes. Rien d’étonnant à cela. Le pied d’un loup est plus long que sa main, par contre cette dernière se montre puissante pour lui permettre de creuser. La morphologie des métatarsiens du loup de Romain-la-Roche correspond bien à celle du loup. L’impression générale est ici l’absence de massivité. Les métatarsiens des rayons latéraux (2 et 5) présentent moins de décalages de longueur par rapport aux rayons centraux (3 et 4) que chez les métacarpiens. Leur diaphyse reste relativement droite, avec des bords parallèles.phalanges et sésamoïdes :Les phalanges, relativement résistantes, ont été bien conservées. Malheureusement, sans une détermination précise de la patte et du doigt, il n’est pas possible de les utiliser pour des comparaisons. Des phalanges de Cuon alpinus europaeus coexistent probablement dans les lots respectifs. Leur distinction s’avère presque impossible, sauf pour deux (phalanges 1 : CAN.1469 et CAN.141) qui ressortent par une très faible taille par rapport à Canis lupus mais sont nettement trop grandes pour être Vulpes vulpes. Phalanges 1 : Tabl. 32 et 33Les phalanges 1 postérieures de Canis lupus correspondent aux phalanges les plus longues et les plus fines. Les antérieures plus courtes et plus larges se repèrent assez facilement. La détermination du doigt ne pose pas de problème entre doigts centraux (rayons 3 et 4) et doigts latéraux (rayons 2 et 5) à cause de la forme arquée et comprimée latéralement de ces derniers. Elle reste plus difficile après et il faut disposer d’un bon matériel de comparaison. Je me suis contenté de déterminer et de mesurer les phalanges 1 complètes les plus évidentes à situer (n = 20). Les autres apparaissent seulement sur la liste du matériel en annexe à la fin (n = 65).Phalanges 2 : Tabl. 34Elles se classent facilement entre antérieures et postérieures. Les phalanges 2 antérieures apparaissent courtes et larges et les postérieures beaucoup plus longues et moins larges. Le classement gauche-droite ne pose pas de problème grâce à la pente de la partie distale de la diaphyse (dissymétrique à cet endroit) dirigée vers l’extérieur. Par contre la distinction des doigts latéraux
Tableau 25 : Romain-la-Roche, Canis lupus, rotule.
Canis lupus rotule
pemac 61
1longueur
2largeur
3épaisseur
CAN.1929 D 19,3 12,7 10,2CAN.1693 D 18,3 12,0 9,8CAN.1389 D 19,3 10,4 8,7
Tableau 24 : Romain-la-Roche, Canis lupus, fémur.
Canis lupusFémur
pemac 60
1l
2dt prox.
max.
3dt max.
tête fémur
4Distance
2 trochanters
5dt min.
col fémur.
6dt
méd.
7dAp méd.
8dt dist.
9dAp dist.
10dt maxart. dist.
n 5 12 19 9 15 15 15 20 18 13min 190,70 40,64 19,84 32,40 16,07 13,10 13,96 28,81 36,09 30,41max 206,20 48,68 24,40 41,63 21,24 17,34 17,50 39,77 44,67 39,70moy 196,78 45,40 22,09 36,84 18,82 15,44 15,48 36,41 41,45 34,36
écart-type 7,11 3,02 1,18 3,24 1,4 1,28 1,03 2,61 2,18 2,374- Distance directe entre le bord externe du grand trochanter et le sommet du petit trochanter.

518 a. ARgANt
Tableau 26 : Romain-la-Roche, Canis lupus, tibia.
Canis lupus tibia
pemac 62
1l
2dt prox.
3dApprox.
4dt
médian
5dAp
médian
6dt dist.
7dAp dist.
8dt
art. dist.
9dAp
art. dist.n 8 13 12 28 26 27 28 23 23
min 185,60 35,96 39,20 12,83 12,65 23,18 17,83 15,42 14,63max 220,80 46,00 44,89 17,26 15,78 29,44 21,48 20,04 19,33moy 203,68 39,89 42,51 14,70 14,58 26,02 19,65 18,03 17,77
écart-type 11,81 2,79 1,86 1,05 0,85 1,73 1,12 1,29 1,27
Fig. 9 : Romain-la-Roche, Canis lupus, tibia. Périmètre médian de la diaphyse/longueur. Mise en relation avec le type A ou B de la crête tibiale. Position des deux tibias de Cuon alpinus europaeus.

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 519
Tableau 27 : Romain-la-Roche, Canis lupus, péroné.
Canis lupus péronépemac 63
6 7dt dist. dAp distal
CAN.1355 D 8,3 13,6CAN.38 D 7,7 13,1
CAN.1087 g 8,3 13,1
Tableau 28 : Romain-la-Roche, Canis lupus, tarse, calcanéum.
Canis lupuscalcanéumpemac 641
1l
2dt max.
3dApmax.
4dt
min. méd.
5dAp min.
méd.
6dt dist.
7dAp dist.
8distance
hors artic.n 13 14 16 17 17 15 15 16
min-max 45,9-54,8 18,5-22,6 19,8-23,3 5,7-10,1 11,3-15,0 10,7-16,6 13,3-16,0 30,1-35,4moy 50,09 20,58 21,26 8,33 13,63 13,54 14,75 32,73
écart-type 2,9 1,4 1,2 0,9 0,9 1,5 0,9 1,5
Tableau 29 : Romain-la-Roche, Canis lupus, tarse, astragale.
Canis lupus Astragale
pemac 642
1l
2dt max.
3ddpmax.
4dt
articulaire
5dt col
6dAp tête
n 12 11 9 15 12 12min-max 26 ,5-32,0 17,1-23,2 15,6-18,4 13,7-17,6 11,2-13,2 8,0-11,0
moy 30,14 21,07 16,72 16,29 11,98 9,33écart-type 1,8 1,6 1,0 1,2 0,7 0,9
Tableau 30 : Romain-la-Roche, Canis lupus, tarse, autres.
Canis lupus Tarse
Pemac 64
1dAp
2dt
3dp
cuboïdepemac 643
n 7 6 7min-max 17,0-20,3 14,9-17,7 14,7-16,8
moy 18,99 16,09 15,30écart-type 1,10 1,25 0,79
naviculairepemac 644
n 4 4 5min-max 9,9-12,4 16,1-20,4 13,4-14,9
moy 11,26 18,31 14,16écart-type 1,10 1,80 0,70
3e cunéiformepemac 647
n 2 2 2min-max 11,5-11,6 9,7-10,0 14,9-15,1
moy 11,53 9,85 15,01écart-type 0,02 0,17 0,03

520 a. ARgANt
Tableau 31 : Romain-la-Roche, Canis lupus, métatarsiens.
Canis lupus Métatarsiens pemac 65
1l
2dt prox.
3dAp prox.
4dt méd.
5dAp méd.
6dt dist.
7dAp dist.
8dt art.
dist
Mtt.2pemac 652
n 6 12 9 10 13 8 9 8min-max 68,2-79,7 7,7-10,3 13,9-16,0 6,6-8,2 6,1-9,3 9,9-12,3 9,5-10,7 9,6-10,6
moy 75,26 8,79 14,77 7,38 7,03 10,76 9,91 10,04écart-type 4,31 0,76 0,79 0,45 0,83 0,76 0,42 0,34
Mtt.3pemac 653
n 7 17 11 11 11 7 7 7min-max 73,4-88,5 9,3-11,3 13,6-16,7 7,8-9,0 5,6-7,6 9,5-10,7 9,9-11,4 8,6-9,8
moy 80,61 10,25 15,34 8,23 6,41 10,20 10,55 9,21écart-type 5,60 0,58 0,91 0,35 0,71 0,44 0,55 0,39
Mtt.4pemac 654
n 3 10 9 7 7 4 4 4min-max 81,8-85,6 8,4-10 ,5 12,9-14,8 6,5-7,8 5,9-6,9 9,4-10,4 9, 8-11,0 8 ,7-9,2
moy 83,41 9,56 13,71 7,32 6,40 9,76 10,28 8,93écart-type 1,95 0,74 0,57 0,44 0,37 0,50 0,50 0,23
Mtt.5pemac 655
n 3 5 5 7 7 6 5 5min-max 76,5-82,3 10,8-12,7 11,7-13,3 5,1-6,5 6,6-8,0 8,9-10,9 8,5-10,2 8,6-9,9
moy 78,73 11,66 12,40 5,90 7,36 9,80 9,24 9,11écart-type 3,12 0,83 0,67 0,50 0,51 0,69 0,71 0,55
Tableau 32 : Romain-la-Roche, Canis lupus, phalanges 1 antérieures déterminées.
Canis lupusphalanges 1antérieurespemac 55
1l
2dt prox.
3dAp prox.
4dt méd.
5dAp méd.
6dt dist.
7dAp dist.
8dAp dist.
art.
doigt 2
n 7 7 7 7 7 7 7 7min 24,9 9,11 8,63 5,34 6,23 7,72 5,84 7,71max 27,94 10,83 9,98 6,91 7,13 9,51 7,3 8,65moy 26,72 9,98 9,10 6,21 6,71 8,31 6,68 8,06
écart type 1,00 0,61 0,53 0,53 0,35 0,62 0,49 0,40
doigt 3
n 4 4 4 4 4 4 4 4min 28,65 9,02 7,98 5,75 5,4 7,12 5,6 6,88max 31,22 10,01 9,38 6,5 6,21 8,44 6,82 8,48moy 29,76 9,61 8,76 6,19 5,68 7,66 6,29 7,59
écart type 1,14 0,44 0,62 0,32 0,38 0,57 0,51 0,67
doigt 4
n 2 2 2 2 2 2 2 2min 29,1 10,08 9,1 6,34 5,8 8,25 6,6 8,16max 31,09 10,41 9,46 6,5 5,84 8,61 6,77 8,55moy 30,10 10,25 9,28 6,42 5,82 8,43 6,69 8,36
écart type - - - - - - - -
doigt 5
n 7 7 7 7 7 7 7 7min 24,93 10,45 8,91 6,48 6,5 8,16 6,49 7,96max 27,97 11,45 9,88 6,92 6,96 9,65 7,32 9,11moy 26,63 10,88 9,53 6,71 6,74 8,76 6,90 8,59
écart type 1,22 0,33 0,34 0,17 0,16 0,46 0,32 0,35

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 521
et des doigts centraux reste beaucoup moins évidente. Je me limite à la distinction des phalanges 2 antérieures (n = 10) et postérieures (n = 13), en mesurant deux lots représentatifs de phalanges intactes. Les autres apparaissent sur la liste du matériel en annexe (n = 26).Phalanges 3 : Tabl. 35
Il est malheureusement très difficile de déterminer plus précisément les phalanges portant les griffes. Seule la distinction de la phalange de l’extrémité du doigt 1 résiduel, forcément antérieur, beaucoup plus petite et bien plus courte, reste évidente. On ne peut même pas distinguer avec sécurité les phalanges antérieures des phalanges postérieures. Comme précédemment j’ai réuni un lot représentatif de phalanges 3 complètes pour les mesurer (n = 14). Les quatre dernières phalanges du doigt 1 antérieur (CAN.462, 1663, 218, 1425) ressortent nettement du lot par leur faible taille et présentent un relatif aplatissement latéral par rapport aux autres phalanges 3. Celles non mesurées apparaissent sur la liste du matériel en annexe (n =20).Sésamoïdes : Tabl. 36Six sésamoïdes de canidés appartiennent à Canis lupus. Leur forme arquée et leur petite taille les rendent bien caractéristiques. Il ne peut s’agir de Vulpes vulpes dont la taille des sésamoïdes reste bien inférieure. Par contre on ne peut éliminer totalement le cuon pour les plus petits.
II.1.3.2. Biochronologie :La lignée évolutive du loup s’utilise fréquemment comme indicateur biochronologique et ce depuis
Tableau 34 : Romain-la-Roche, Canis lupus, phalanges 2 antérieures et postérieures.
Canis lupus 1l
2dt
prox.
4dt
méd.
6dtdist.
phalanges 2antérieuresPemac 56
n 10 10 10 10min 14,45 7,93 5,45 7,32max 17,19 9,7 7,64 9,34moy 15,80 8,85 6,56 8,33
écart type 0,94 0,61 0,67 0,71
phalanges 2postérieures
pemac 67
n 13 13 13 13min 18,36 7,39 4,95 7,73max 23,05 10,16 6,86 9,68moy 20,96 8,71 5,98 8,54
écart type 1,39 0,76 0,61 0,67
Tableau 33 : Romain-la-Roche, Canis lupus, phalanges 1 postérieures déterminées.
Canis lupus phalanges 1postérieures
pemac 66
1l
2dt prox.
3dAp prox.
4dt méd.
5dAp méd.
6dt dist.
7dAp dist.
8dAp dist. art.
doigt 2
n 5 5 5 5 5 5 5 5min 26,68 8,94 7,75 5,07 5,79 7,43 5,85 7,47max 27,68 9,92 9,06 5,91 6,6 8,39 6,38 8,24moy 27,12 9,52 8,39 5,40 6,08 7,92 6,14 7,91
écart type 0,43 0,39 0,47 0,33 0,34 0,34 0,22 0,29
doigt 3
n 5 5 5 5 5 5 5 5min 28,84 9,05 8,69 5,92 5,14 7,81 6,27 7,69max 32,19 9,99 9,82 6,45 6,04 8,6 7,5 8,55moy 30,34 9,48 9,27 6,19 5,77 8,15 6,76 8,16
écart type 1,34 0,40 0,45 0,21 0,37 0,39 0,55 0,44
doigt 4
n 10 10 10 10 10 9 10 10min 28,59 9,16 8,41 5,73 5,19 7,75 5,93 7,56max 34,00 10,57 9,68 6,47 6,17 8,75 7,39 8,73moy 31,02 9,80 9,17 6,09 5,65 8,18 6,49 8,00
écart type 1,97 0,47 0,41 0,27 0,36 0,38 0,41 0,42
doigt 5
n 5 5 5 5 5 5 5 5min 22,72 8,41 7,8 4,71 5,84 6,86 5,14 6,86max 24,93 9,46 8,45 5,72 6,28 7,78 6,14 7,77moy 24,09 8,94 8,05 5,13 5,98 7,25 5,80 7,28
écart type 0,86 0,49 0,28 0,38 0,18 0,40 0,39 0,38

522 a. ARgANt
longtemps (lehMan, 1954 ; boniFay, 1971 ; Patou, 1984 ; aRgant, 1991…) :
Canis etruscus àCanis mosbachensis à Canis lupus lunellensis àCanis lupus santenaisiensis à Canis lupus lupus.
Elle traduit une augmentation relativement régulière de la taille moyenne des individus (jugée sur une population suffisante) pour atteindre les dimensions maximum au Pléistocène supérieur avant de rediminuer finalement à l’Holocène. Depuis Canis etruscus, les dents se sont allongées et épaissies, la M/1 présente un paraconide élevé. Les vrais loups, au sens actuel du terme, apparaissent au Pléistocène moyen. Le Canis lupus lunellensis boniFay de Lunel-Viel (Hérault) traduit un animal d’assez bonne stature (OIS 13, 14 ou 15 ? « Mindel-Riss »). Il devient possible si on dispose d’une série suffisante d’éléments dentaires (les mieux étudiés) d’utiliser les moyennes comme marqueurs chronologiques. Il s’agit bien sûr d’un schéma à prendre avec la prudence nécessaire car il est impensable d’imaginer dans toute l’Europe une évolution
linéaire uniforme et régulière. Pour cette raison, j’ai évité le plus possible les comparaisons à grande distance, à l’échelle de l’Europe. J’ai volontairement donné la priorité aux sites proches (Franche-Comté, Bourgogne) pour rester dans le domaine médio-européen et conserver le maximum de paramètres communs ou très proches pour les gisements choisis. Cela ne va pas sans un certain nombre d’inconvénients, comme celui de se priver de sites bien datés de référence mais l’avantage de la cohérence des paramètres me semble préférable.Nuria gaRCia a précisé la transition entre le petit canidé cromérien Canis mosbachensis (= Canis lupus mosbachensis) et la forme typique Canis lupus (gaRCia, 2003, p. 137-138). Le type A correspond à Canis lupus, protoconide de la M/1arrondi, vertical et paraconide bas. Le protoconide de la P/4 est aussi arrondi, bas et la seconde cuspide postérieure confondue avec le bourrelet arrière terminal. Le type B appartient à la forme ancestrale Canis mosbachensis avec, sur la M/1, un protoconide pointu et orienté vers l’arrière, un paraconide haut. Pour la P/4, le type primitif possède un protoconide pointu et haut, une deuxième cuspide postérieure se distinguant du bourrelet terminal (fig. III-3-8 et 3-9, p. 137). La forme d’Atapuerca – trinchera galeria tD.8a (Espagne) correspond à cette forme ancienne (type B), tout comme celle de Château – Brèche 4, la plus ancienne (Saône-et-Loire, Bourgogne, France). A Romain-la-Roche le type A est nettement reconnaisssable, il s’agit bien de Canis lupus.J’utilise de préférence la première molaire inférieure (M/1) pour tenter les comparaisons en évitant par exemple la P4/ et la M1/ pour lesquelles on ne connaît pas toujours exactement la façon de mesurer des auteurs. La Figure 10 présente le diagramme de répartition des moyennes (MD/VL) de cette M/1 pour des sites de référence retenus pour leur nombre de données disponibles et leur position chronologique largement admise. La population du Canis lupus de Romain-la-Roche apparaît proche de celle de Santenay (Côte-d’Or), gisement malheureusement assez mal daté que l’on attibue le plus souvent à la fin du stade isotopique 6, voire au stade 5 (Chaline, 1969, p. 81 ; aRgant, 1991), « antéwürmien » en tout cas. Que vaut cette indication biochronologique ? Plusieurs datations U/th ont été tentées sur des ossements de Romain-la-Roche (Lab. de géochronologie, Paris). Seule la date de 150 000 ± 18 000 peut être retenue (éch.13, z = 460 cm, couche VI – VII c) indiquant la fin du stade isotopique 6. Les couches VI à VIII b présentent la plus grande abondance et le meilleur état de conservation des restes de Canis lupus (PauPe, 1996). Le loup de Romain-la-Roche ne se rattache pas en tout cas aux formes anciennes Canis mosbachensis du Pléistocène moyen médian (Escale, Hundsheim, Mosbach, Vergranne, Château, Atapuerca tD.8a). Ce n’est pas non plus une forme du Pléistocène supérieur Canis lupus lupus, dont il n’atteint ni la grande taille, ni la denture massive et plus broyeuse (tabl. 37).La biochronologie situe le Canis lupus de Romain-
Tableau 35 : Romain-la-Roche, Canis lupus, phalanges 3.
Canis lupus phalanges 3
1l
2dt prox.
4dt méd.
doigt 1antérieur pemac 57
n 4 12 12min 12,96 6,06 9,38max 16,48 9,59 12,19moy 15,01 7,93 10,91
écart type 1,58 0,89 0,90
doigt non déterminépemac 75
n 10 10 10min 19,27 7,62 9,38max 23,18 9,59 12,19moy 20,99 8,23 11,11
écart type 1,21 0,57 0,83
Tableau 36 : Romain-la-Roche, Canis lupus, sésa-moïdes.
Canis lupus Sésamoïdespemac 72
1dAp
2dt
3ddp
CAN. 460 11,9 4,4 6,6CAN. 1430 10,0 3,8 5,7CAN.1432 10,1 3,8 5,5CAN.1534 10,0 4,3 5,5CAN.1425 9,8 3,8 5,6CAN.1431 8,2 4,3 5,3

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 523
L’Escale La Fage Santenay
Romain-la-Roche
Villereversure
LazaretBalaruc VII
Jaurens
CHRONO
RELATIVELunel-Viel
VL
Canis lupus M1 inférieure
8
8,5
9
9,5
10
10,5
11
11,5
12
12,5
23 24 25 26 27 28 29 30 31
MD (mm)
n<5
n>5Linéaire (n>5)
L'Escale
Hundsheim
MosbachLa Fage
Balaruc VII
LazaretRomain-la-Roche Santenay
Villereversure
Jaurens
Tourville
Château (EN. Br.4)
(mm
)
Vergranne
Château (ES. c. 2)
Vergisson II
Lunel-Viel
Fig. 10
Vergisson II
Vergisson II
Fig. 10 : Comparaison des moyennes de la longueur et de la largeur de la M/1 des populations de Canis lupus de gisements du Pléistocène moyen et du Pléistocène supérieur.
Tableau 37 : Romain-la-Roche, Canis lupus, os longs du membre antérieur (humérus, radius, métacarpiens). Comparaison de la longueur avec d’autres gisements européens.
Sites 1Huméruslongueur
2radius
longueur
3Mtc.2
longueur
4Mtc.3
longueur
5Mtc.4
longueur
6Mtc.5
longueur
romain-la-roche
n 5 3 4 7 5 6min-max 177,4-183,8 176,7-198,2 61,9-71,5 71,3-82,4 67,6-80,5 60,5-70,2
moy. 180,84 186,13 66,27 77,35 73,14 63,59écart-type 8,41 10,98 4,44 4,48 4,66 3,67
Actuel groënland et danemark
n 10 11min-max 201,0-233,0 198,0-234,0
moy. 214,19 212,11
Jaurens pléist. sup.
n 1 2 6 4 2 3min-max 225,0 217,0-219,0 77,0-81,0 90,0-95,0 90,0-97,0 75,0-78,0
moy. 77,8 91,8 76,7
pléist. moyen récentn 1 2 1 2 2
min-max 199,0 61,5-62,0 71,0 70,8-74,0 61,0-64,5l’escale
pléist. moyen méd.Canis etruscus mosbaschensis
n 4 6 2 2 2 2min-max 145,5-160,0 148,9-168,9 56,3-56,4 63,8-64,8 63,6-65,8 54,1-55,0
moy. 153,1 156,3

524 a. ARgANt
la-Roche à la fin du Pléistocène moyen récent ou à l’interglaciaire Eémien, au Pléistocène supérieur ancien (Fig. 10).
II.1.3.3. paléoécologie :Canis lupus possède une formidable capacité d’adaptation puisqu’il se rencontre dans tous les climats extrêmes, des régions froides de l’arctique jusqu’aux déserts tropicaux et forêts à mousson de l’Inde avec toutes les transitions imaginables : bois, forêts, steppes, landes des régions tempérées, du niveau de la mer jusqu’à plus de 5000 m d’altitude dans l’Himalaya, en Amérique du Nord, en Europe et en Asie. Seules, l’Afrique et l’Amérique du Sud présentent un certain isolement génétique (beauFoRt, 1987). Il apparaît donc illusoire de tirer une signification paléoécologique de la présence du loup à Romain-la-Roche.
II.1.4. Sous-Famille cuoninaegenre Cuon
espèce Cuon alpinus PaLLas, 1811
Le site de Romain-la-Roche, grâce à l’abondance des vestiges de Canis lupus qu’il renferme, permet la connaissance d’une population statistiquement fiable. On peut donc approcher les données métriques et morphologiques sur des séries significatives. Dans ce contexte, tout élément peu représenté (le cuon est toujours rare), sortant du cadre statistique de la population de Canis lupus la plus nombreuse, peut être considéré comme appartenant à une espèce différente. Par exemple les deux tibias complets (CAN.595 g et CAN.507 d) présentent une longueur inhabituelle, inférieure à la valeur de la série des tibias complets de Canis lupus (n = 11). Il y a tout lieu de supposer une autre espèce de canidé aux membres plus courts et plus grêles, ce qui est le cas de Cuon alpinus. Une fois la présence du cuon
assurée, il devient plus facile d’accorder aux variations morphologiques une valeur déterminante en recherchant les variations qui dépassent l’ampleur des variations individuelles de Canis lupus. Mais la distinction de Canis lupus et de Cuon alpinus reste encore aléatoire dans de nombreux cas, en particulier sur du matériel dentaire isolé.
II.1.4.1. données morphométriques :La liste d’inventaire du matériel disponible apparaît en annexe à la fin.demi-mandibule : CAN.1069 d. (Pl. I, fig. 1-2) avec sa branche horizontale compète, sauf à l’avant de la canine : C. inf, racine de P/1 cassée à ras, P/2, P/3, P/4, M/1 endommagée, M/2 et l’alvéole vide de M /3. Il reste juste le bord antérieur de la base de la branche montante et le processus angulaire n’est pas présent (Tabl. 38, Pl. I, fig. 1-2).CAN.1069 se distingue de la série des demi-mandibules de Canis lupus de Romain-la-Roche. Sa taille correspond aux plus petites valeurs rencontrées. Seules deux autres mandibules de loup, probablement de petites femelles, sont comparables. La P/1, dont l’alvéole est présente remplie de sédiment induré, paraît de taille relativement importante. Elle correspond à une dent uniradiculée et n’est séparée de la canine que par un diastème de 3 mm. La partie incisive manque. Les trois prémolaires suivantes portent un denticule net sur le flanc postérieur du paraconide comme celles de Canis lupus et correspondent parfaitement à leur description tout en donnant une impression de plus forte étroitesse. Ce tubercule accessoire sur la P/1 est net. Cette prémolaire reste isolée ici de la P/2 par un espace vide de 3 mm. La quatrième prémolaire présente un denticule antérieur au paraconide bien individualisé. C’est le seul cas sur 37 observations de P/4 à Romain-la-Roche. Sans cette information, il aurait été difficile de déterminer Cuon alpinus, car la M/3 ou du moins son alvéole (dent
Tableau 38 : Romain-la-Roche, Cuon alpinus europaeus, mandibule, Pemac 20 (Fig. 7).
Cuon alpinus europaeusMandibulepemac 20
romain-la-rochecAn. 1069
7 93,4 L. C.post.-M/3 post.8 89,3 L. P/1-M/39 83,5 L. P/2-M/310 41,2 L. des molaires11 50,4 L. des prémolaires P/1-P/412 43,0 L. P/2-P/420 11,6 Dt de la mandibule derrière M/1 22 25,9 H. derrière M/123 21,7 H. entre P/2 et P/326 12,7 DT maxi 1/2 mdb entre P/4 et M/1

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 525
uniradiculée), est bien présente ici alors que son absence constitue souvent un argument déterminant de l’espèce. D’autres différences viennent confirmer cette distinction.Denture inférieure : Tabl. 39La M/1 malheureusement endommagée, montre cepen-dant un trigonide court, un protoconide moins abrupt à l’arrière que chez le loup, qui descend moins bas sur la face externe. Le quart postéro-interne étant cassé on ne peut malheureusement pas observer valablement le talo-nide, ni vérifier si un métaconide flanquait côté interne
l’arrière du protoconide. Un très léger renflement sou-ligne le bord externe de la couronne sans que l’on puisse parler de cingulum. Cette molaire donne, malgré la cas-sure, l’impression d’une plus grande étroitesse que chez le loup et a un aspect plus coupant. La M/2 de CAN.1069 (usure nulle) se distingue nettement de son homologue du loup par une plus grande étroitesse, un moindre développement de la partie antéro-externe qui descend beaucoup plus bas, en chevauchement sur la racine, chez le loup. L’arête antérieure forme une barre
Tableau 39 : Romain-la-Roche, Cuon alpinus europaeus, comparaison de la denture inférieure. Rosières (gRossouvRe & stehlin, 1912), Cuon stehlini.
Cuon stehlini Cuon priscus
Cuon alpinus fossilis
Cuon alpinus europaeus
ro
MA
In-
lA- r
oc
He
SIteS 1 2 3 4 5 6 7 8 9 10 11 12 cAn.1069
c.infMd - - - - - - 10,7 12,0 - - - - 11,88Vl - - - - - - 6,7 - - - - - 7,7
p/4
Md 18,0 -
a-13,7
14,8 14,8
a-13,6 a-14,0
15,0 - 14,9 13,6
n =42
14,7b-14,0 b-14,8 b-13,8 10,7-13,6
c-15,2 12,370,81
Vl - -
a-6,8
7,0 7,2
a-7,3 a-6,5
- - - 6,9
n=42
7,0b-6,8 b-6,7 b-6,5 5,1-7,3
c-7,5 6,010,46
M/1
Md 24,6 25,1
a-23,2 a-24,2 a-24,0 a-22,2
23,5* 24,3 22,0 26,8 21,4
n=42
26,2b-23,3 b-23,3 b-24,5 b-23,6 18,8-23,3c-24,0 c-23,9 c-24,6 c-24,2 21,42
d-24,5 1,06
Vl - 9,7
a-9,8 a-9,5 a-9,0 a-8,7
- 9,0 9,0 10,1 8,8
n=42
9,8b-9,8 b-9,2 b-9,0 b-8,9 7,1-9,3c-9,5 c-9,2 c-9,3 c-9,6 8,27
d-9,6 0,49
M/2
Md - 10,1
a-8,9 a-9,0
8,15
a-8,2
- - - - 8,0*
n=42
9,8b-9,0 b-9,3 b-8,2 6,0-9,0
c-8,4 7,520,64
Vl - 7,6
a-6,9
7,4 6,45
-
- - - - -
n=42
7,3b-7,2 b-6,6 3,1-7,1
- 5,940,68
1- Rosière (stehlin, 1912), Cuon stehlini 7- Vaufrey, grotte 15 (delPeCh in Rigaud et al., 1988), Cuon sp.2- Mosbach (tobien, 1957), Cuon priscus 8- Fontechevade (aliMen et al., 1958), Cuon alpinus europaeus3- Hundsheim (thénius, 1954), Cuon priscus 9- Cotencher (dubois & stehlin, 1933), Cuon alpinus europaeus4- Lunel-Viel (boniFay, 1971), Cuon priscus 10- Obarresta (gaRCia, 2003, tabl. III.3.12, p.155), Cuon5- Heppenloch (adaM, 1959), Cuon alpinus fossilis alpinus europaeus6- Atapuerca, trinchera galeria (tg.1, tg.2, tg.3) 11- Chokier (CoRdy, 1983), Cuon alpinus europaeus gaRCia, 2003 (tabl. III.3.12, p.155), Cuon alpinus 12- Cuon alpinus actuels in gaRCia, 2003 (tabl. III.3.12, p.155) europaeus

526 a. ARgANt
en fort relief. Elle est constituée d’un protoconide plus volumineux mais de même niveau que le métaconide fort développé. L’hypoconide assez élevé participe à une bordure en arc de cercle continu, de relief décroissant à partir de lui et qui ceinture une fosse occupant la partie postérieure de la dent.Squelette post-crânien :ceintures : Tabl. 40Le coxal g CAN.495 bien qu’endommagé, reste relativement assez complet pour montrer la tubérosité ischiatique, l’acétabulum et le col de l’ilion. Ses dimensions le rangent parmi les trois pièces de plus petite taille de la série des coxaux de Canis lupus (n = 30), en particulier par son diamètre de la cavité cotyloïde (DAP acétabulum = 22,4 mm). La tête du fémur g CAN.508 s’y emboîte parfaitement, contrairement aux têtes de fémurs appartenant à Canis lupus de taille plus importante. Ce fémur CAN.508 possède d’ailleurs des dimensions moindres que celles des autres fémurs, moindre longueur, proportionnellement moindre largeur des extrémités articulaires qui paraissent moins massives.Membres :« Les os des membres du cuon se distinguent de ceux des loups ou des vrais chiens parce qu’ils sont relativement courts, l’animal étant relativement plus bas sur patte » (boule, 1927, p. 52-53).cubitus : Tabl. 41Un seul cubitus CAN.12 droit sort du lot des cubitus du loup de Romain-la-Roche. Il est presque complet, un peu encroûté de calcite qu’il a fallu dégager pour pouvoir observer valablement l’extrémité proximale. L’extrémité distale manque, mais en tenant compte des indications fournies par la terminaison de l’arête latérale, il est possible de reconstituer la longueur totale avec l’aide du matériel de comparaison sur des bases sérieuses. Sans être exacte totalement, cette longueur ne peut s’écarter
de plus de 2 à 3 mm de la réalité. Sa taille reste faible, comprise entre celle du cubitus d’un fort renard et celle d’un cubitus du loup de Romain-la-Roche. La courbure générale apparaît nettement plus accentuée que chez Canis lupus (CAN.725, 1158, 397, 216...) et la diaphyse particulièrement fine distalement. La partie proximale, relativement bien conservée, présente une incisure trochléaire beaucoup moins développée dans sa partie antérieure, plus aplatie que chez Canis lupus, en donnant l’impression d’être comprimée latéralement.carpe : Tabl. 42CAN.1432 correspond à un magnum droit de canidé plus fort que Vulpes vulpes, mais de très petite taille par rapport à CAN.1716 appartenant à Canis lupus (Tabl. 42). Je l’attribue à Cuon alpinus europaeus. De petites différences morphologiques au niveau des facettes articulaires et un moindre développement relatif de la partie la plus large apportent une confirmation.Fémur : Tabl. 43Les deux fémurs identiques CAN.508 g. et CAN.757 d ressortent du lot des fémurs de Canis lupus de Romain-la-Roche. Le premier est complet, à peine endommagé à l’extrémité distale. L’extrémité distale du second manque. Ils correspondent aux deux plus petits spécimens. La tête du fémur g. s’emboîte bien dans l’acétabulum du coxal g. précédent CAN.495, déjà attribué à Cuon. Les extrémités distales de ces deux fémurs s’adaptent bien aux plateaux tibiaux correspondants de CAN.595 g. et CAN.507 d. Là encore, les extrémités paraissent moins massives, moins larges que chez le loup. La distance entre les deux trochanters est bien moindre. La fosse trochantérique s’ouvre moins largement au niveau du petit trochanter, paraît finalement plus creuse et la tête du fémur plus détachée sur le Cuon alpinus europaeus de Romain-la-Roche.tibia : Tabl. 44
Tableau 40 : Romain-la-Roche, Cuon alpinus europaeus, coxal.
Cuon alpinuscoxal pemac 41 1 2 3 4 5 6 7 8 9 10 11 12 13
CAN.495g
- 22,4 - - - - 16,2 - 22,4 22,0 18,2 10,2 8,7
1- L. max.2- DAP acétabulum3- DAP foramen obtus4- L. symphyse pubienne5- Dt min. bassin complet au niveau de la petite échancrure
sciatique6- Dt min. milieu bord sup. des 2 cavités cotyloïde7- Dt épine sciatique au milieu de la cavité cotyloïde
8- Dt min. bassin complet au niveau de la grande échancrure sciatique
9- DDP acétabulum10-DDP min. pédoncule de l’ilion11-DDP min. pédoncule de l’ischion12-Dt pédoncule de l’ilion13-Dt pédoncule de l’ischion

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 527
Les deux tibias complets CAN.595 g. et CAN.507 d. se distinguent des tibias de Canis lupus par une longueur moindre, des épiphyses articulaires moins volumineuses et moins larges. Ils ont tous les deux les mêmes dimensions, le même aspect, une provenance proche (Carrés S.19 et R.19). Ils correspondent à deux astragales CAN.596 g et CAN.1095 d qui s’emboîtent parfaitement à leurs extrémités articulaires distales. Le tibia g et l’astragale g sont d’ailleurs tous deux enregistrés dans le même carré et sous le même numéro (RO.86. S.19-59). Il s’agit bien d’un même individu de Cuon alpinus.
Les différences morphologiques par contre résistent mal à la comparaison avec la série complète des tibias de Canis lupus (NR = 33 dont 11 complets). La variabilité individuelle ressort plus sur des séries importantes et il ne reste finalement que de rares points de différenciations morphologiques si on exclut les proportions et les dimensions absolues moindres, déjà signalées sur Cuon. L’amorce proximale de la face postérieure du tibia paraît cependant plus plane, sur une plus grande longueur, pour les deux tibias du Cuon (même individu). La crête tibiale en vue latérale externe paraît plus haute, relativement
Tableau 41 : Romain-la-Roche, Cuon alpinus europaeus, cubitus droit (CAN. 12).
Cuon alpinus pemac
52cubitus
1l
2dt art.
min.
3dAp prox.
4dt
méd.
5dAp méd.
6dt dist.
7dAp dist.
8dt
articul. prox. max.
9dAp art.
prox.
10dt
olécrâne prox.
11ddp
CAN.12 196,3* 7,6 26,5 9,7 10,4 - - 13,4 18,1 10,9 23,2
Tableau 42 : Romain-la-Roche, Cuon alpinus europaeus, carpe. Magnum d. (CAN. 1422), comparaison (Vulpes vulpes actuel correspond à un mâle de forte taille).
Cuon alpinus Magnum
pemac 535
1ddp
2dt
3dAp
Canis lupusCAN. 1716 12,6 10,9 19,4
Cuon alpinus europaeusCAN. 1432 9,7 7,9 14,6
Vulpes vulpes 6,1 5,9 10,7
Tableau 43 : Romain-la-Roche, Cuon alpinus europaeus, fémur.
Cuon alpinus pemac 60
Fémur
1l
2dt
prox.
3dt
max.tête
fémur
4distance 2
trochanters
5dt min.
col fémur.
6dt
méd.
7dApméd.
8dtdist.
9dApdist.
10dtart. dist.
11dAp
art. dist
CAN. 508 g 181,0 43,4 20,4 33,2 16,4 13,9 14,4 - 39,0* - 39,3*CAN. 757 D - - - - - 14,4 14,2 34,3 38,4 32,5 39,2
4- Distance directe entre le bord externe du grand trochanter et le sommet du petit trochanter.
Tableau 44 : Romain-la-Roche, Cuon alpinus europaeus, tibia.
Cuon alpinus tibia
pemac 62
1l
2dt prox.
3dApprox.
4dt
médian
5dAp
médian
6dt dist.
7dAp dist.
8dt
art.dist.
9dAp
art. dist.CAN. 595 D 190,6 40,7 41,8 14,2 14,5 26,7 18,2 19,1 17,6CAN. 507 g 185,0 36,5 40,3 14,3 13,0 24,6 18,6 16,8 15,4

528 a. ARgANt
plus courte et la fosse tibiale formée plus creuse. Elle se termine par une rupture distale avec la diaphyse plus raide. A l’extrémité distale, proportionnellement moins large et moins haute, la partie articulaire répondant à l’astragale est plus étroite et plus profonde chez le cuon que chez le loup. Côté externe, la cochlée tibiale possède un développement moindre chez le cuon. Ceci correspond évidemment aux observations faites sur les deux astragales.Signalons enfin une trace résiduelle de soudure du péroné à la base du tibia d. CAN.507, mais pas sur le tibia gauche CAN.595.tarse : Astragale : Tabl. 45Il faut souligner la petite taille des deux astragales de cuon qui se comparent aux plus petits éléments de la série de Canis lupus. Ils se ressemblent morphologiquement mais le col de la tête de l’astragale apparaît nettement moins long et moins large chez le cuon que chez le loup. La trochlée paraît un peu plus profonde, relativement moins dissymétrique que chez le loup. La surface articulaire répondant au calcanéum, côté interne, est plus allongée, plus étroite et surtout relie la tête côté proximal au bord distal de la trochlée sur Cuon alpinus europaeus.phalange 1 : Tabl. 46Deux phalanges de canidés (CAN.1469 et CAN.1413) paraissent vraiment petites et grêles par rapport au lot des phalanges 1 de Canis lupus. Elles sont nettement plus grosses que celles d’un fort renard. Dans le contexte précédent, la seule possibilité de détermination reste Cuon alpinus europaeus.
II.1.4.2. Biochronologietrois espèces différentes de Cuon sont connues, Cuon stehlini thénius, 1954, Cuon priscus thénius, Cuon
alpinus europaeus bouRguignat, 1868, formant une même lignée (adaM, 1959) ou indépendantes et correspondant à trois vagues de migration différentes (boniFay, 1971). La première espèce de grande taille se rencontre au Pléistocène inférieur et moyen (Rozière, Vallonnet et peut-être à l’Escale). La seconde espèce, Cuon priscus toujours de forte taille caractérise le Pléistocène moyen (Mosbach, Caune de l’Arago, Hundsheim, Lunel-Viel, Verzé). Cuon alpinus correspond à la forme la plus récente. Ses premières apparitions datent du « Riss » et on le connaît ensuite en Europe occidentale jusqu’à la fin du Pléistocène supérieur. Ses dimensions sont moindres et il se rapproche des cuons actuels. Plusieurs sous-espèces ont été avancées, par exemple Cuon alpinus fossilis pour le cuon d’Heppenloch (nehRing, 1890) souvent considéré comme une forme archaïque de Cuon alpinus europaeus. Dans le cas précis de Romain-la-Roche, il convient de déterminer s’il s’agit de Cuon priscus, de Cuon alpinus fossilis ou de C. alpinus europaeus. La Figure 11 classe les différentes espèces et sous-espèces selon le rapport des longueurs de M/2 et de M/1. CAN.1069 possède une M/1 très longue et une M/2 relativement moins développée. Le point correspondant se place sans ambiguïté parmi Cuon priscus. A contrario la présence d’une véritable cuspide à la partie antérieure de P/4 constitue souvent un argument déterminant de la sous-espèce Cuon alpinus europaeus bouRguignat 1868. Que penser de cette contradiction ? L’histoire des cuons fossiles, malgré de nombreuses études, apparaît encore bien fragmentaire à cause de la rareté des sites et de la rareté du matériel dans les gisements quand il y en a. Il est encore impossible dans ces conditions de cerner réellement la part évolutive et la part des variations individuelles sur un matériel suffisant. De plus, il y a toutes les probabilités pour
Tableau 45 : Romain-la-Roche, Cuon alpinus europaeus, astragale.
Cuon alpinus Astragalepemac 642
1l
2dt prox.
3ddpmax.
4dt
articulaire
5dt col
6dAp tête
7dt tête
CAN. 596 g 26,5 20,0 15,3 15,4 11,0 9,3 13,8CAN. 1095 D 27,0 20,6 15,0 - - 9,2 13,8
Tableau 46 : Romain-la-Roche, Cuon alpinus europaeus, phalange 1
Cuon alpinus phalanges 1
pemac 73
1l
2dt prox.
3dAp prox.
4dt méd.
5dAp méd.
6dt dist.
7dApdist.
CAN. 1469 23,6 7,8 6,3 5,5 4,0 5,9 4,5CAN. 1413 24,5 8,4 7,3 5,0 5,6 6,7* 5,6*

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 529
que cette histoire des cuons fossiles corresponde à de multiples vagues d’expansion de populations asiatiques.Dans l’état actuel des connaissances, on peut seulement constater que le cuon de Romain-la-Roche a une denture dépassant nettement les valeurs des formes du Pléistocène supérieur et de la forme actuelle. La P/4 possède un denticule antérieur, ce qui permet d’éliminer la forme Cuon priscus du Pléistocène moyen. Il s’agit vraisemblablement de Cuon alpinus europaeus. Il n’y a qu’un seul individu à Romain-la-Roche et ce n’est pas l’idéal sur le plan statistique. On constate cependant que les exemplaires de Cuon alpinus d’Atapuerca, trinchera galeria tg.3 (gaRCia, 2003, fig.III.3.22 et 23, p. 166-167) possèdent d’assez fortes dimensions et que l’on rencontre sur les individus représentés un denticule antérieur nettement développé à la quatrième prémolaire inférieure (ATA.89.GSU12.F.26-1 ; ATA.84.gSU.11.F.17-1). Ce gisement est rapporté au Pléistocène moyen. Il existe très peu de mentions du squelette post-crânien dans la littérature. F. delPeCh (delPeCh, 1989, p. 233) signale une extrémité distale de tibia droit (couche V/VI) s’articulant bien avec un astragale de cuon de la couche VIII du point de vue dimensionnel. Elle donne :DAP articulaire distal = 16,0 mm (CAN.595 g = 17,4 ; CAN.507 d = 16,5)
DT articulaire distal = 21,4 mm (CAN.595 g = 20,9 ; CAN.507 d = 21,2)C’est compatible avec le matériel de Romain-la-Roche. Ces couches sont attribuées à une période du milieu du « Riss ».Ces deux dernières indications chronologiques suggèrent que le cuon de Romain-la-Roche se situe probablement vers la fin de l’avant-dernière glaciation, au stade OIS 6 ou au tout début du dernier interglaciaire Eémien. Il ne correspond pas aux formes plus récentes.
II.1.4.3. paléoécologieLes cuons actuels évitent les paysages découverts et se plaisent dans les zones montagneuses couvertes de forêts épaisses et giboyeuses (boule, 1927 ; boniFay, 1971). Mais lorsqu’on regarde de façon plus globale la répartition chronologique et spatiale des cuons fossiles tout au long du Pléistocène en Europe, on s’aperçoit rapidement qu’ils présentent une large adaptabilité à de nombreux biotopes, y compris dans les zones de basse altitude et quasiment en plaine (Chokier, 300 m d’altitude, CoRdy, 1983). Le dhole actuel (Cuon alpinus), comme de nombreux carnivores, possède une capacité exceptionnelle d’adaptation à son environnement. On le trouve aussi bien dans les forêts tropicales d’Asie ou dans la jungle indienne que dans les steppes de Sibérie ou dans
Fig. 11 Fig. 11 : Romain-la-Roche, représentation graphique de la longueur et de la largeur de la M1 inférieure (variables standardisées) de
Cuon priscus et Cuon alpinus ssp. de différents sites européens (tabl. 39).

530 a. ARgANt
les régions montagneuses himalayennes jusqu’à 3000 m d’altitude. A l’état fossile on ne peut donc guère, à mon avis, lui accorder de signification climatique précise.
II.2. ordre carnivoraSuper-famille ursoidea
Famille ursidaeSous-famille ursinaeII.2.1. genre Ursus
espèce Ursus arctos Linné, 1758
Parmi l’important matériel d’ours des cavernes de Romain-la-Roche on trouve de rares éléments appartenant à l’ours brun Ursus arctos. Cela n’a rien d’exceptionnel et se produit dans un grand nombre de gisements car les deux espèces coexistent depuis longtemps (1 Ma, voire plus). La difficulté réside dans la séparation des deux espèces, facile pour un certain nombre de pièces significatives mais beaucoup plus difficile dans de nombreux cas (des incisives usées par exemple, des pièces endommagées). Il est probable qu’un certain nombre d’éléments d’U. arctos se retrouvent déterminés par défaut comme Ursus spelaeus, mais l’essentiel est que sa présence soit assurée, ce qui est le cas ici, sans aucun doute possible.
II.2.1.1. données morphométriques :La liste d’inventaire du matériel disponible apparaît en annexe à la fin (NR = 19, NMI = 1, NMIE = 2).La demi-mandibule g URS.1058 (RO.86-R.17-69) bien conservée est longue et fine. Elle porte les alvéoles vides de P/4 et de M/3. Les molaires présentes M/1 et M/2 apparaissent noircies, fortement usées. Par contre la canine en parfait état, blanche, non usée, me paraît incompatible avec ces deux molaires précédentes qui traduisent un individu très âgé. Je la traite comme une dent isolée car je suppose un recollage abusif sur la base de la compatibilité de taille entre la racine et l’alvéole vide. On peut observer la présence d’une alvéole vide de P/1 très nette ne pouvant en aucun cas correspondre à l’alvéole vide d’une canine lactéale fraîchement tombée et qui ne s’est pas encore refermée comme on le voit parfois chez les très jeunes individus. Le même cas se reproduit sur la demi-mandibule d URS.271 (RO.85-R.19-50) qui porte la canine en place, usée, et la P/4. Une alvéole vide de P/1 est présente également. Il n’y a pas de traces d’autres prémolaires antérieures à P/4 (Pl.3-1 a, CAN.271). La P/1 est souvent la seule prémolaire inférieure sur la mandibule d’Ursus arctos comme le montre la Planche III, fig. b (Ursus arctos subactuel, Espagne). On ne rencontre jamais de P/1 sur les mandibules d’Ursus spelaeus du gisement. Il y a au moins deux individus d’ours brun à Romain-la-Roche car ces deux mandibules g et d ne correspondent pas par leurs dimensions (Tabl. 47, Tabl. 48, Fig. 12).denture supérieure : dents isolées
P3/ : URS.1740 g (RO.87-Q.10-22), MD = 6,6, VL = 5,2, HC = 4,4.URS.1743 d (RO.86-Q.19-41), MD = 7,0, VL = 6,8, HC = 4,9.denture inférieure : : Tabl. 48Ensembles : URS.271 (RO.85-R.19-50), URS.1769 (RO.86-R.17-69)Dents isolées :P/1 : URS.1741 g (RO.87-Q.17-5), MD = 6,6, VL = 4,4, HC = 3,4, HT = 14,6.P/3 : URS.1742 g (RO.86-R.19-131), MD = 5,5, VL = 4,8, HC = 4,6, HT = 13,1.P/4 : URS.1743 dSquelette post-crânien :tibia : Tabl. 49Un tibia g pratiquement complet URS.1187 se distingue du lot des tibias d’Ursus spelaeus par une plus grande longueur, plus de gracilité. Ces proportions sont caractéristiques d’Ursus arctos aux membres plus allongés, plus fins.Métapodes : Mtc.1 g URS.629 (RO.86-S.17-21), Mtc.2 g URS.1384 (RO.87-Q.17-14), Mtc.5 d URS.658 (RO.86-R.19-41), Mtt.1 d URS.1422 (RO.87-P.18-9), Mtt.3 g URS.1072 (RO.86-R.17-94), issus de carrés proches (tabl. 50).Quelques métapodes ressortent du lot de ceux d’Ursus spelaeus par une plus grande longueur, par leur gracilité et les bords plus parallèles de leur diaphyse. Ils appartiennent à Ursus arctos.phalange 1 : URS.1353 (RO.87-HS-23), Tabl.51phalanges 3 : Tabl. 52, Pl. 4-6 (URS.844)URS.518 (RO.86-S.18-39), URS.632 (RO.86-S.17-27), URS.844 (RO.86-R.18-76), URS.850 (RO.86-R.18-10) Nettement plus longues et plus fines chez Ursus arctos, les phalanges 3 distinguent bien les deux espèces d’ours. A Romain-la-Roche, plusieurs phalanges 3 très nettement allongées appartiennent avec certitude à l’ours brun, comme par exemple URS.844 (Pl. IV, fig. 6).
II.2.1.2. BiochronologieLe matériel d’ours brun de Romain-la-Roche n’est pas assez abondant d’une part et, d’autre part, la biochronologie d’Ursus arctos est loin d’être documentée par des séries suffisantes. Il apparaît illusoire de tenter d’utiliser ce matériel comme moyen de comparaison et à plus forte raison comme élément de datation.La coexistence de l’ours brun et de l’ours des cavernes se rencontre dans presque tous les gisements livrant suffisamment de vestiges d’ursidés. Le gisement Paléolithique moyen de Biache-Saint-Vaast (Pas-de-Calais), daté de 170 Ka. environ, livre par exemple les deux espèces (auguste, 1988). Un des plus anciens ours brun connu en Europe, celui de l’Aven de Vergranne (Doubs), du Pléistocène moyen médian coexiste avec Ursus deningeri, l’ancêtre d’Ursus spelaeus (Chagneau & PRat, 1983). Le site de Vergranne se situe seulement à quelques kilomètres de Romain-La-Roche.

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 531
Tableau 47 : Tableau 47 : Romain-la-Roche, Ursus spelaeus et Ursus arctos, mandibule, Puema 20 (Fig.n12).
MandibuleUrsus
puema 20
Ursus spelaeus Ursus arctos
n Min Max Moy Ecart-type URS.271 URS.1058
1 16 249,8 323,9 282,2 19,5 249,7 271,0*2 9 255,7 311,5 279,2 19,9 250,6 -3 16 214,7 278,5 246,0 16,9 217,2 232,34 17 237,8 307,2 272,6 19,8 239,3 265,95 9 214,2 272,4 245,8 18,8 217,1 233,06 25 127,6 165,0 145,5 9,1 134,1 -7 24 151,9 188,4 172,3 10,9 157,8 134,68 24 85,4 108,9 96,8 5,3 96,9 85,89 23 35,2 61,5 47,9 6,9 39,8 51,910 24 68,2 99,1 81,8 7,3 82,0 71,911 13 11,8 15,9 14,2 1,0 13,4 -12 18 81,0 198,1 106,9 26,1 81,7 100,513 10 107,0 141,5 120,3 11,2 - 116,014 24 48,3 71,9 60,8 6,2 48,7 55,615 24 47,6 75,2 62,2 7,0 52,6 55,516 24 46,2 59,8 54,1 4,1 47,6 46,717 22 48,0 69,0 56,5 5,0 52,0 52,418 20 44,4 66,9 56,7 5,2 50,3 52,319 24 48,1 70,7 59,5 6,5 51,9 50,820 21 51,0 79,9 65,3 7,8 54,6 56,521 18 18,4 30,2 25,6 2,8 22,0 18,222 14 44,0 69,3 57,6 9,0 44,0 48,623 20 69,8 101,3 86,4 8,2 72,8 -24 20 33,2 47,3 40,2 4,4 33,525 28 17,4 23,4 20,2 1,8 17,8 19,026 25 21,2 30,0 24,1 2,6 25,0 20,527 10 110,0 153,7 132,9 14,6 - 128,628 12 29,6 42,5 36,1 4,0 31,229 13 51,9 92,7 80,6 11,0 69,130 14 162,8 198,9 178,0 9,8 166,7
1- Longueur totale, Condyle-Infradental2- Distance processus angulaire-Infradental3- Distance Condyle – arrière de la canine4- Distance Condyle – avant de la canine5- Distance Processus angulaire – arrière de la canine6- Distance entre arrière de la canine et la dernière molaire7- Distance entre avant de la canine et l’arrière de la
dernière molaire8- Longueur de la rangée des dents jugales9- Longueur du diastème10-Longueur des molaires11-Longueur des prémolaires12-Distance condyle – arrière de la dernière molaire13-Hauteur du Processus angulaire – Coronion14-Hauteur de la branche horizontale au niveau de l’arrière
de la M/1, côté lingual15-Hauteur de la branche horizontale au niveau de l’arrière
de la M/2, côté lingual
16-Hauteur de la mandibule au niveau du diastème17-Hauteur de la mandibule sous la P/4, côté labial18-Hauteur de la mandibule sous la M/1, côté labial19-Hauteur de la mandibule sous la M/2, côté labial20-Hauteur de la mandibule sous la M/3, côté labial21-Hauteur du condyle22-Largeur du condyle23-Diamètre maximum de la symphyse24-Diamètre minimum de la symphyse25-Epaisseur de la branche horizontale entre P/4 et M/126-Epaisseur de la branche horizontale entre M/2 et
M/327-Hauteur maximum de la mandibule28-Longueur Infradental- arrière de la canine29-Longueur Infradental – avant de la P/430-Longueur Infradental – arrière de la dernière molaire

532 a. ARgANt
1
2
3
4
5
6
7
891011
12
13
14 1516
17181920
21
22
23 24
2526
27
28
293012
Mandibule - Puema 20
Fig. 12 : Plan de mesurage des demi-mandibules d’ursidés (Puema.20)
1- Longueur totale, Condyle-Infradental
2- Distance processus angulaire-Infradental
3- Distance Condyle – arrière de la canine
4- Distance Condyle – avant de la canine
5- Distance Processus angulaire – arrière de la canine
6- Distance entre arrière de la canine et la dernière molaire
7- Distance entre avant de la canine et l’arrière de la dernière molaire
8- Longueur de la rangée des dents jugales
9- Longueur du diastème10- Longueur des molaires11- Longueur des prémolaires12- Distance condyle – arrière de la
dernière molaire13- Hauteur du Processus angulaire
– Coronion14- Hauteur de la branche horizontale
au niveau de l’arrière de la M/1, côté lingual
15- Hauteur de la branche horizontale au niveau de l’arrière de la M/2, côté lingual
16- Hauteur de la mandibule au niveau du diastème
17- Hauteur de la mandibule sous la P/4, côté labial
18- Hauteur de la mandibule sous la M/1, côté labial
19- Hauteur de la mandibule sous la M/2, côté labial
20- Hauteur de la mandibule sous la M/3, côté labial
21- Hauteur du condyle22- Largeur du condyle23- Diamètre maximum de la
symphyse24- Diamètre minimum de la symphyse25- Epaisseur de la branche horizontale
entre P/4 et M/126- Epaisseur de la branche horizontale
entre M/2 et M/327- Hauteur maximum de la mandibule28- Longueur Infradental- arrière de la
canine29- Longueur Infradental – avant de la
P/430- Longueur Infradental – arrière de la
dernière molaire

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 533
Tableau 48 : Romain-la-Roche, Ursus arctos, denture inférieure.
Ursus arctosdenture inférieure Md Vl Hc u lat.
C inf URS.271 17,8 14,5 27,7 cassure dP/4 URS.271 14,0 8,8 8,5 0 d
P/4 isolée URS.1743 13,1 7,5 7,5 0 dM/1 URS.1058 24,2 - - 4 gM/2 URS.1058 26,9 16,6 - 4 g
Tableau 49 : Romain-la-Roche, Ursus arctos, tibia.
Ursus arctostibia g puema 62 l dt prox dAp prox dt méd dAp méd dt dist dAp dist
URS.1187 327,1 - 83,5 29,3 33,0 74,0 41,3
Tableau 50 : Romain-la-Roche, Ursus arctos, métapodes.
Ursus arctosMétapodes
puema 54 et 65
l dt prox dAp prox
dt méd dAp méd dt dist dAp dist dt dist articul
Mtc.1 URS.629 81,1 27,2 21,9 12,2 12,5 21,4 - -Mtc.2 URS.1384 80,2 17,0 26,5 13,5 10,9 21,5 19,0 19,8Mtc.5 URS.658 80,0* 24,4* - 12,8 16,0 23,6 17,4 21,5Mtt.1 URS.1422 76,0* 27,8 20,6 12,4 12,0 - - -Mtt.3 URS.1072 85,9 20,3 30,7 14,2 11,7 22,7 20,2 18,8
Tableau 51 : Romain-la-Roche, Ursus arctos, phalanges 1.
Ursus arctos phalange 1puema 73
l dt prox dAp prox dt méd dAp méd dt dist dAp dist
URS.1353 40,6 18,6 14,7 10,4 10,4 13,6 10,0
Tableau 52 : Romain-la-Roche, Ursus arctos, phalanges 3
Ursus arctos phalanges 3puema 75
l dt prox dAp prox
URS.518 56,4 27,6 17,5URS.632 53,4 25,4 -URS.844 56,0* 29,0 17,3URS.850 52,0* 29,9 17,4

534 a. ARgANt
II.2.1.3. paléoécologieUrsus arctos a une signification plus tempérée que l’ours des cavernes. Il a connu à la fin de la dernière période glaciaire une expansion liée à l’amélioration climatique holocène alors qu’Ursus spelaeus disparaît par exemple de Chartreuse au Dryas récent vers 13000 BP (aRgant et al., 2009). Si son habitat actuel montagnard et forestier ne reflète que l’espace concédé par l’homme, il a vécu auparavant sur tout le territoire en milieu forestier ou assimilé. Sa présence conjointe avec celle d’Ursus spelaeus à Romain-la-Roche indique donc une couverture végétale dominée par des arbres, probablement sous un climat de type tempéré, ce que ne contredit pas la palynologie.
II.2.2. genre Ursusespèce Ursus spelaeus RosenmüLLeR &
HeinRotH, 1794
L’ours des cavernes Ursus spelaeus apparaît bien représenté à Romain-la-Roche, pas tout à fait autant que Canis lupus, mais en nombre de restes déterminés avec certitude, il occupe largement le second rang des espèces de carnivores. Il constitue également une série de référence en fournissant un jalon non négligeable dans l’histoire de l’espèce.
II.2.2.1. données morphométriquesLa liste d’inventaire du matériel disponible apparaît en annexe à la fin (NR = 1313, NMI = 20).crânes : Tabl. 53, Tabl. 54, Fig. 13, Pl. V, fig. 1 (URS.01)Au moins 17 crânes sont attestés mais comme souvent à Romain-la-Roche dans un état de conservation médiocre. Quatre seulement peuvent fournir les principales données morphométriques attendues, les autres apportant essentiellement des informations dentaires.Crâne URS.001 : couche c.7 bC’est le seul crâne complet, d’ailleurs exposé au Musée dans la Galerie Cuvier (Pl. V, fig. 1). Il possède la largeur et l’arrondi de l’arrière-crâne, la bosse frontale bien marquée et le museau court d’Ursus spelaeus typique. Il montre un très fort angle frontal (148 gr) et un fort enfoncement glabellaire (173 gr) bien caractéristiques. On peut noter une alvéole vide de P3/ aux maxillaires gauche et droit. Cette prémolaire antérieure à P4/ n’existe pas ou alors très exceptionnellement sur Ursus spelaeus, mais se rencontre plus fréquemment sur la forme ancestrale Ursus deningeri.Crâne URS.973 : présente également une alvéole vide de P3/ à gauche et à droite.Crânes URS.157 et 158 : couche c.7 : ils présentent aussi un fort angle frontal et l’enfoncement glabellaire caractéristiques.URS. 681 : ce fragment montre une alvéole vide de P3/ d.URS.1438 : ce fragment montre une alvéole vide de P3/ d.
Les crânes indiquent nettement que le stade Ursus spelaeus typique est atteint à Romain-la-Roche, mais la présence d’une prémolaire antérieure à P4/ dans plusieurs cas apporte l’indication d’une relative ancienneté.demi-mandibules : Fig. 12, Tabl. 45, Pl. IV, fig. 7 (URS.1439), Pl. IV, fig. 1 (URS.1085) et Pl. IV, fig. 8 (URS.1439)trente-cinq demi-mandibules ou fragments (20 g et 15 d) existent, souvent bien conservées, car au moins 16 d’entre elles permettent la plupart des observations et les mesures importantes (Tabl. 45, Fig. 12). On retrouve les variations morphologiques habituelles, de la demi-mandibule longue, plutôt droite et gracile (type A) à la demi-mandibule plutôt courte, à branche horizontale haute et épaisse, relativement courbe (type B). La Figure 15 met en rapport ces deux types morphologiques et le sexe, déterminé grâce au diagramme de répartition des canines inférieures, pour vérifier si ces deux formes correspondent bien à une différenciation à caractère sexuel ou non. Romain-la-Roche apporte une confirmation. trois mandibules de type A, dont une en limite de répartition des deux lots (grosse femelle) se retrouvent dans l’ensembles des canines appartenant à des femelles. Les cinq demi-mandibules de type B appartiennent à l’ensemble des mâles. Les autres points de ce diagramme correspondent aux cas où la détermination du type ou du sexe n’est pas possible. L’étude d’un lot important de demi-mandibules de la population d’Ursus spelaeus deningeroides d’Azé 1-3 (Saône-et-Loire) concluait à un aspect gracile chez tous les jeunes (A) puis, à une différenciation d’ordre sexuel (A femelles, B mâles) à partir de la maturité sexuelle des individus. Les âges relatifs utilisés sont déterminés par les stades d’usure dentaire (aRgant, 1991, p. 114).denture :denture lactéale :Les dents lactéales ne sont connues dans le matériel de Romain-la-Roche que par deux canines déciduales, une supérieure (CAN.544) et une inférieure (CAN.1160). Elles ont une couronne non usée et leur racine complète est sans traces de résorption. Il ne s’agit pas de la chute des canines lactéales remplacées par des dents définitives. Elles traduisent ici la mort au moins d’un ourson tombant dans le piège de l’aven. Cette faible représentation conforte l’existence du piège et infirme la présence d’une galerie profonde ouverte par la suite et servant de lieu d’hivernation pour les ours.denture supérieure : tabl. 55Incisives : Les incisives ont une racine simple expliquant qu’elles se détachent facilement du crâne. Il en existe à Romain-la-Roche, le plus souvent sous forme de dents isolées. La I1/ et la I2/ se reconnaissent facilement à leur couronne arrondie et courbée, flanquée de deux reliefs plus ou moins dissymétriques sur la pente linguale. La I3/ est la plus fréquente. Son aspect caniniforme et sa massivité lui confèrent une bonne résistance. Elle distingue bien Ursus arctos et Ursus spelaeus chez qui elle possède une

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 535
Tableau 53 : Romain-la-Roche, Ursus spelaeus et Ursus arctos, crâne, Puema 10 (Fig. 13).
urS. 001 157 158 972 159 160 409 973 972 17171 456,9 356,2 347,7*2 438,73 406,9456 237,2 210,0 210,0*7 144,28 254,3 194,6 176,0*9 80,5 140,0*10 186,7 139,01112 168,8 142,013 88,0 85,2 83,6 85,414 209,5 216,015 26,3 37,0 33,6 45,316 14,3 24,8 36,617 120,6 106,4 123,3 104,1 118,4 108,718 278,0 240,0* 242,0*19 86,1 85,0* 84,0* 84,0 96,0* 71,320 151,1 132,0* 119,8 108,0* 11721 105,7 80,022 118,7 130,0* 110,0 119,023 143,9 132,724 148,0 130,025 173,0 166,028 1 1 2 2 ? 2
1- Longueur totale, Acrocranion-Prosthion2- Longueur condylobasale, Condyle occipital-Prosthion3- Longueur basale, Condyle occipital-Prosthion4- Longueur basicraniale, Basion-Synsphénion5- Longueur basifaciale, Synsphénion-Prosthion6- Longueur du neurocrâne, Acrocranion-milieu du
frontal7- Longueur du viscérocrâne, Nasion-Prosthion8- Longueur faciale, milieu du Frontal-Prosthion9- Longueur du nasal, Nasion-Rhinion10-Longueur du museau, Infraorbital-Prosthion11-Longueur du palatin, Staphylion-Prosthion12-Largeur Otion-Otion13-Largeur du condyle occipital14-Largeur du processus paraoccipital
15-Largeur du trou occipital16-Hauteur du trou occipital, Basion-Opisthion17-Largeur maximum du neurocrâne, Euryon-Euryon18-Largeur des arcades zygomatiques, Zygion-Zygion19-Largeur minimum du neurocrâne, à la constriction20-Largeur maximum du frontal, Ectorbital-Ectorbital21-Largeur minimale au niveau des orbites, Entorbital-
Entorbital22-Hauteur du triangle occipital, Acrocranion-Basion23-Hauteur maximum du crâne, Staphylion-Frontal24-Angle frontal en grades25-Angle glabellaire en grades26-Dt maximum ouverture nasale27-Dt facial entre foramens intraorbitaires28-Sexe (mâle = 1, femelle = 2)

536 a. ARgANt
1
23
4
5
6
7
8
910
11
12
1314
15
16
18
19
2021
22
23
26
27
1
2
3
4
56
78
9
Crâne - Puema 10
17
Maxillaire - Puema 100
N
F
E E
P
R
P
SOt Ot
St
A
F
St OtP
P
P
Op
Z
B
Z
A
Fig. 13 : Plan de mesurage du crâne et du maxillaire supérieur d’ursidés (Puema 10 et Puema 100).
1- Dt maximum du maxillaire2- Dt minimum du maxillaire3- Dt maximum entre les canines4- DT maximum entre les M 2/5- Longueur entre avant de la C et arrière de la M2/6- Longueur entre arrière de la C et arrière de la M2/7- Longueur des dents jugales
8- Longueur des molaires9- Longueur du diastème entre C et P4/10- PM antérieure à P/4 (oui = 1, non = 2)11- MD de la P/3 ; si la dent est absente MD de l’alvéole12- VL de la P/3 ; si la dent est absente VL de l’alvéole13- Sexe mâle (1) ou femelle (2) ; C sup. g et/ou d

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 537
couronne moins élevée, plus large. La fossa lunaris située entre l’arête mésiale et l’arête linguale est bien présente à la base de la couronne, côté lingual. L’arête distale porte parfois un très léger renflement (kalyx distalis) à peine perceptible dans le cas de CAN.1484 (type 1, RabedeR, 1999, p. 72, Abb. 42), mais absent sur CAN.1373. La fréquence va en augmentant chez les formes spéléennes les plus récentes. La taille de cette dent s’accroît au cours de l’évolution. Le type spéléen typique de grande taille atteint des dimensions bien supérieures à celles rencontrées à Romain-la-Roche.canine supérieure : Fig. 14Les canines sont classiquement utilisées pour déterminer le sexe des individus, les mâles possédant théoriquement des crocs plus forts (Koby, 1949). La répartition des points sur le diagramme de répartition VL/MD (Fig. 14) montre bien deux ensembles distincts sans problème de chevauchement. A Romain-la-Roche, les effectifs donnent ici un léger avantage aux mâles (n = 13 contre 10 femelles) pour les canines supérieures, mais la situation s’inverse et traduit pratiquement l’égalité pour les canines inférieures (n = 15 mâles pour 16 femelles). C’est la preuve de non hivernation de cette population, car il y aurait dans ce cas une nette majorité de femelles venant mettre bas dans la grotte à cette période, ce qui confirme la cause accidentelle de l’origine du matériel paléontologique.p4/ : Pl. IV, fig. 5 (URS.681)Chez Ursus etruscus il s’agit d’une dent simple à trois
cuspides bien séparées que l’on retrouve chez U. arctos. Chez U. deningeri, cette structure évolue vers la voie spéléenne en se compliquant. Chez U. spelaeus le deutérocône, en position latérale interne, prend un développement plus important, des tubercules accessoires se dessinent et se multiplient. Les crêtes du paracône et du métacône forment un angle de moins en moins marqué. A Romain-la-Roche, la P4/varie comme souvent entre deux extrêmes, allant d’une dent relativement simple telle URS. 1372 (des morphotypes A ou B ; RabedeR, 1992, fig.1, p.134) à une dent plus large, avec des tubercules secondaires plus ou moins développés (morphotype C). Le deutérocône ici paraît volumineux mais toujours simple, nettement séparé du protocône et du métacône. Les crêtes de ces deux tubercules forment un angle moyen de 171 gr (tabl. 55). Le protocône domine toujours largement le métacône. Un cingulum un peu crénelé borde pratiquement la base de la couronne.M1/ : Pl. IV, fig. 3 (URS.442)C’est une dent large, un peu globuleuse. Les reliefs principaux correspondent au paracône et au métacône. Le premier domine toujours nettement, mais les deux sont volumineux. Le parastyle et le métastyle, qui épaulent les deux tubercules principaux, ont une importance variable selon les dents examinées. Ils ne sont jamais très développés. La surface d’occlusion possède de multiples rides et granulations. Le bord lingual de la molaire présente une pente de 50° environ, couverte de rides. Un cingulum bien marqué, fortement crénelé, longe tout le flanc interne de la M1/ à mi-pente du bord.
Tableau 54 : Romain-la-Roche, Ursus spelaeus, maxillaire supérieur, Puema 100 (Fig.n13).
Ursus spelaeus Maxillaire supérieur
puema 1001 2 3 4 5 6 7 8 9 10 11 12 13
URS.404 80URS.1 116,3 68,7 96,5 56,2 170,3 144,3 97,9 69,6 46,7 1 5,6 a 5,3a 1URS.158 107,5 79,2 61,1 164,2 138,4 92,0 73,3 41,4 0URS.157 105,7URS.159 103 0URS.160 98,3 54,6 80,7 0URS.409 111 65,6 86,0 71,3 0URS.973 142 122 77,0 50,0 1 1URS.814 121,6 86,6 71,7 35,3URS.1194 98,4URS.1438 84,34 67,7 24,2
1. Dt maximum du maxillaire 8. Longueur des molaires2. DT minimum du maxillaire 9. Longueur du diastème entre C et P4/3. DT maximum entre les canines 10. PM antérieure à P/4 (oui = 1, non = 0)4. T maximum entre les M 2/ 11. MD de la P/3 ; si la dent est absente MD de l’alvéole5. Longueur entre avant de la C et arrière de la M2/ 12. VL de la P/3 ; si la dent est absente VL de l’alvéole6. Longueur entre arrière de la C et arrière de la M2/ 13. Sexe mâle (1) ou femelle (2) ; C sup. g et/ou d7. Longueur des dents jugales
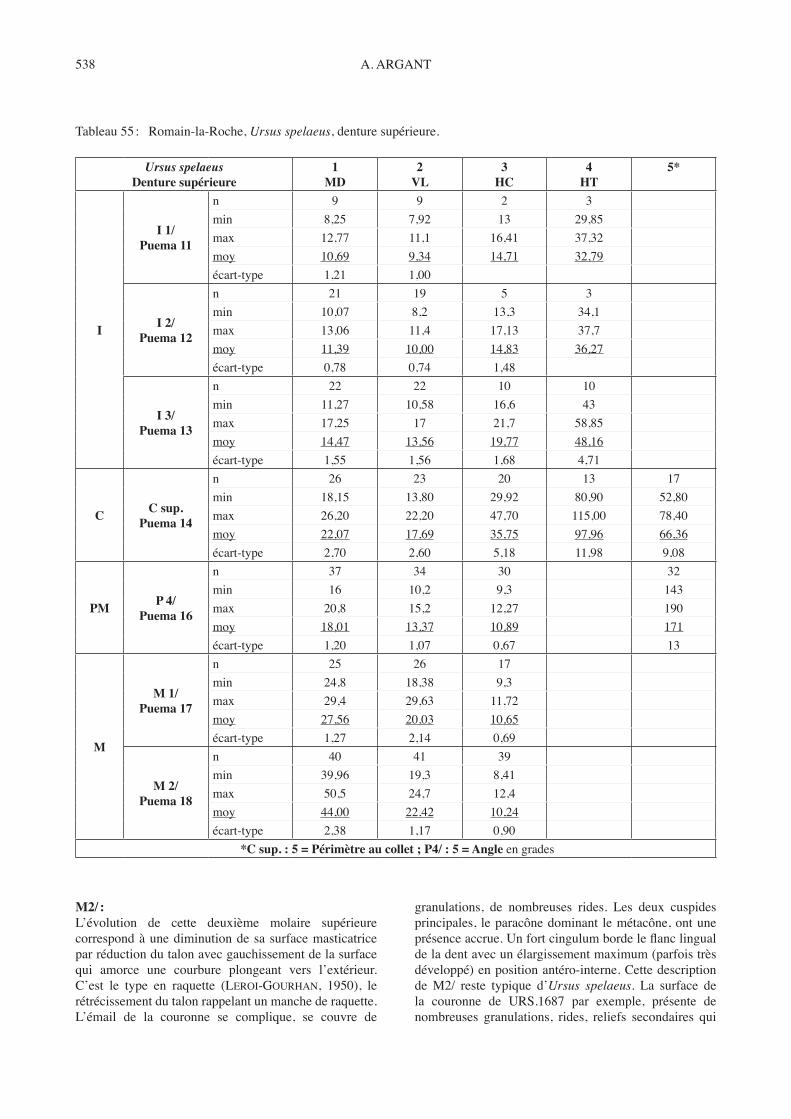
538 a. ARgANt
M2/ :L’évolution de cette deuxième molaire supérieure correspond à une diminution de sa surface masticatrice par réduction du talon avec gauchissement de la surface qui amorce une courbure plongeant vers l’extérieur. C’est le type en raquette (leRoi-gouRhan, 1950), le rétrécissement du talon rappelant un manche de raquette. L’émail de la couronne se complique, se couvre de
granulations, de nombreuses rides. Les deux cuspides principales, le paracône dominant le métacône, ont une présence accrue. Un fort cingulum borde le flanc lingual de la dent avec un élargissement maximum (parfois très développé) en position antéro-interne. Cette description de M2/ reste typique d’Ursus spelaeus. La surface de la couronne de URS.1687 par exemple, présente de nombreuses granulations, rides, reliefs secondaires qui
Tableau 55 : Romain-la-Roche, Ursus spelaeus, denture supérieure.
Ursus spelaeus denture supérieure
1Md
2Vl
3Hc
4Ht
5*
I
I 1/puema 11
n 9 9 2 3min 8,25 7,92 13 29,85max 12,77 11,1 16,41 37,32moy 10,69 9,34 14,71 32,79écart-type 1,21 1,00
I 2/puema 12
n 21 19 5 3min 10,07 8,2 13,3 34,1max 13,06 11,4 17,13 37,7moy 11,39 10,00 14,83 36,27écart-type 0,78 0,74 1,48
I 3/puema 13
n 22 22 10 10min 11,27 10,58 16,6 43max 17,25 17 21,7 58,85moy 14,47 13,56 19,77 48,16écart-type 1,55 1,56 1,68 4,71
c c sup.puema 14
n 26 23 20 13 17min 18,15 13,80 29,92 80,90 52,80max 26,20 22,20 47,70 115,00 78,40moy 22,07 17,69 35,75 97,96 66,36écart-type 2,70 2,60 5,18 11,98 9,08
pM p 4/puema 16
n 37 34 30 32min 16 10,2 9,3 143max 20,8 15,2 12,27 190moy 18,01 13,37 10,89 171écart-type 1,20 1,07 0,67 13
M
M 1/puema 17
n 25 26 17min 24,8 18,38 9,3max 29,4 29,63 11,72moy 27,56 20,03 10,65écart-type 1,27 2,14 0,69
M 2/puema 18
n 40 41 39min 39,96 19,3 8,41max 50,5 24,7 12,4moy 44,00 22,42 10,24écart-type 2,38 1,17 0,90
*c sup. : 5 = périmètre au collet ; p4/ : 5 = Angle en grades

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 539
affectent même le cingulum prenant un aspect crénelé. Sa morphologie est proche du type 3 de g. RabedeR (1999, p. 55, Abb. 30).denture inférieure : tabl. 56Incisives : Les incisives inférieures se distinguent facilement des incisives supérieures par leur aspect anguleux, rectiligne, contrastant avec les formes globalement arrondies des supérieures. La I/3 avec sa couronne en forme de mitaine est particulièrement reconnaissable. tout comme les autres dents, les incisives enregistrent l’évolution. La I/3 URS.1366 par exemple correspond au morphotype C de g. RabedeR (1999, p. 74, Abb. 44) avec un sulcus et une arête linguale qui s’étendent de la base de la couronne jusqu’à la moitié supérieure de la couronne, mais sans atteindre la pointe.canines : Fig. 15Les canines inférieures se distinguent des supérieures par un plus fort rayon de courbure de la couronne et une plus grande largeur relative. Le profil longitudinal en S, imposé par la sortie de la mandibule avec dévers externe de la couronne, apporte souvent une confirmation. A Romain-la-Roche, la racine quand elle est visible reste fine et allongée dans 7 cas sur 10 observations possibles et est grosse, large et arrondie (« patatiforme ») dans les 3 cas restants.prémolaires : p/4 : on observe la variabilité habituelle allant de la P/4 simple « arctoïde », avec le protoconide comme seul relief
important et quelques pointements à l’extrémité distale de la couronne (par exemple CAN.716, CAN.709), aux types nettement spéléens avec deux forts tubercules (parfois trois), un protoconide et un métaconide en position antéro-interne avec un fort élargissement et multiplication anarchique des tubercules secondaires (par exemple CAN.826). Cela va des types C.1, C.2 aux types D.2 ou D.3 de g. RabedeR (1999).M/1 : Pl. IV, fig. 4 a et b (URS. 282 et 284)La première molaire inférieure de l’ours des cavernes de Romain-la-Roche apparaît courte, ramassée, avec un talon large. URS.1365, par exemple, possède un protoconide large, peu élevé. Les métastyles forment une rangée de cuspides reliant le métaconide au bord lingual du paraconide à l’avant de la couronne. Ce dernier, peu élevé, au bord antérieur bien vertical, présente une ligne médiane en relief au centre de sa partie occlusale triangulaire. Le métaconide est bien séparé du protoconide. L’hypoconide reste faible, de peu de relief. L’endoconide, toujours important pour préciser le degré d’évolution, est formé de trois cuspides de taille décroissant régulièrement de l’arrière vers l’avant, rappelant Ursus deningeri.D’après l’analyse des morphotypes de g. RabedeR (RabedeR, 1999, p. 20 à 23) :- métastyles bordant le protoconide côté lingual : type 2
(Abb. 3, p. 21)- endoconide (ou entoconide) en position linguale du
talon : type A.3 (Abb. 4, p. 21)
Fig. 14 : Romain-la-Roche, Ursus spelaeus, répartition de MD x VL des canines supérieures.

540 a. ARgANt
Tableau 56 : Romain-la-Roche, Ursus spelaeus, denture inférieure.
Ursus spelaeusdenture inférieure
1Md
2Vl max.
3Hc
4Ht
5pc
I
I /1puema 21
n 10 10 5 3min 7,65 5,09 8,19 26,56max 10,27 8,35 11,67 32,1moy 8,71 6,27 10,51 29,84écart-type 0,96 1,28 1,37 2,91
I /2puema 22
n 10 9 2min 8,57 7,53 11,24 max 11,34 9,29 13,55 moy 9,96 8,42 12,40 écart-type 1,01 0,53
I /3puema 23
n 21 15 6 5min 10,38 10,8 14,8 36,08max 13,14 12,86 18,12 42,83moy 11,31 11,87 16,19 39,23écart-type 0,68 0,59 1,27 3,03
c c inf.puema 24
n 34 33 14 5 17min 17,84 13,40 27,74 79,34 52,57max 29,33 21,74 40,20 96,09 84,01moy 23,51 17,19 35,03 89,09 66,30écart-type 3,73 2,41 4,80 8,33 10,21
pM p /4puema 26
n 25 27 16 min 12,72 7,47 7,52 max 33,4 21 17,75 moy 14,42 9,03 8,58 écart-type 0,91 0,69 0,69
M
M /1puema 27
1 2 3 4*n 22 24 8 18min 24,49 11,84 10,14 8,18max 31,68 15,88 12,24 12,67moy 28,26 13,84 11,01 10,71écart-type 2,02 1,08 0,60 1,07
M /2puema 28
n 32 29 18 31min 25,14 15,1 9,5 15max 32,44 21 17,75 19,12moy 28,34 17,60 10,72 17,09écart-type 1,73 1,32 1,85 1,13
M /3puema 29
n 26 27 24 min 20,75 15,9 7,17 max 33,4 20,6 10,5
moy 25,10 18,41 8,52 écart-type 2,58 1,20 0,74
*M/1 et M/2 : 4 = VL antérieur

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 541
- enthypoconide en bordure vestibulaire du talon : type A (Abb. 5, p. 22)
- piliers de l’endoconide : type B (Abb. 6, p. 22).M/2 : La couronne de cette deuxième molaire inférieure possède une striction suivie d’un élargissement du lobe postérieur. globalement, mais avec quelques exceptions du domaine de la variation individuelle, la largeur du lobe postérieur renseigne sur l’ancienneté. Ursus etruscus possède un lobe antérieur dominant ou égal au postérieur. Plus tard Ursus deningeri montre un lobe postérieur plus large ou égal à l’antérieur. Chez Ursus spelaeus le lobe postérieur s’élargit encore plus nettement. A Romain-la-Roche le lobe postérieur correspond à la plus grande largeur dans 19 cas sur les 25 observations possibles. C’est le lobe antérieur qui domine, très légèrement, dans les 6 autres cas (URS.35, URS.406, URS.1435 g et d, URS.1483). Sur URS.1483 par exemple, le protoconide et le métaconide s’étendent vers l’intérieur de la couronne en formant une crête transversale barrant la dent au premier tiers antérieur. L’hypoconide est simple et forme un massif peu élevé. L’endoconide correspond à deux cuspides subégales.D’après l’analyse des morphotypes de g. RabedeR (1999, p. 20 à 23) :- trigonide, tout à l’avant de la couronne : type 3 (Abb.
10, p. 29)- métalophe, crête protoconide – métaconide : type A
(Abb. 9, p. 29)
- métastyle, en vue linguale : intermédiaire entre les types 1 et 2 (Abb. 11, p. 29)
- mésolophe au milieu de la surface occlusale : type B’ (Abb. 12, p. 30)
- enthypoconide dans la partie centrale du lobe postérieur : type B (Abb. 13, p. 30).
M/3 : Pl. IV, fig. 2 (URS.285)La forme générale de la troisième molaire inférieure de l’ours des cavernes de Romain-la-Roche correspond à la forme en « haricot » (leRoi-gouRhan, 1950, p. 116) avec une légère constriction côté vestibulaire dans 22 cas sur 27 observations possibles. Dans les 5 autres cas, il s’agit d’une forme dérivée proche. C’est souvent le méta-conide, côté lingual, qui paraît le relief dominant, le pro-toconide formant une sorte de massif complexe, bas, où s’amorcent deux ou trois sillons (type C.2 de RabedeR).D’après l’analyse des morphotypes de g. RabedeR (1999, p. 36 à 39) :- forme de la couronne : type B (Abb. 16, p. 36)- complexe du protoconide : type C.2 (Abb. 17, p. 37)- flanc intérieur du métaconide : type C à D (Abb. 18,
p. 38)- développement de l’hypoconide : type C.2 ou C.3
(Abb. 19, p. 38)- développement du centrolophide : type E (Abb. 20,
p. 38)- développement de l’endoconide : type C (Abb. 21,
p. 39)- développement du talon : type C (Abb. 22, p. 39).
Fig. 15 : Romain-la-Roche, Ursus spelaeus, répartition de MD x VL des canines inférieures. Mise en relation avec le type A ou B de la mandibule.

542 a. ARgANt
L’apport des morphotypes de g. RabedeR (1999) n’est plus à souligner. Il n’est pas possible pour des raisons de place de faire figurer ici tous ces morphotypes mais le renvoi à la publication permet cependant d’atteindre un niveau de précision remarquable, compte tenu de la complexité du système.Squelette post-crânien :colonne vertébrale :- Vertèbres cervicales : Quatorze individus d’Ursus spelaeus au moins ont laissé leur atlas, souvent assez bien conservé, au fond de l’aven de Romain-la-Roche. Le tableau 57 en permet une bonne représentation, au moins du corps central. Par contre la seconde cervicale, l’axis, présente un taux de conservation nettement moindre. Sept pièces fragmentaires seulement sont disponibles et n’apportent que peu de données morphométriques (tabl. 58). Il est possible également qu’il y ait eu un problème de détermination au moment de la constitution des différents inventaires sur lesquels sont fondées les collections. Les autres vertèbres cervicales bien reconnaissables à leur corps vertical à facettes articulaires en biseaux parallèles sont nombreuses. Il est malheureusement difficile de les situer avec précision et leur apport morphométrique devient dans ce cas illusoire.- Vertèbres dorsales et lombaires : l’inventaire en annexe montre l’abondance de ces vertèbres, relativement bien conservées mais elles restent également difficiles à déterminer avec précision. Elles ne peuvent apporter toutes les données souhaitables, ni permettre des comparaisons à cause du manque d’études sur ce type de matériel dans d’autres sites.
- Vertèbre caudale : URS.1773La queue d’un ours est remarquablement courte. Une première vertèbre caudale, bien caractéristique est représentée. Il s’agit d’un jeune individu car le corps vertébral n’est pas encore épiphysé : DT = 42,5, DAP corps = 24,4, DAP max = 31,8, DDP max = 18,6.ceintures scapulaire et pelvienne :omoplate (Fig. 16) : les omoplates complètes restent rares car ce sont des pièces fragiles. Il s’agit essentiellement de fragments comportant la partie articulaire (12 g, 8 d). Seule URS.1494 (RO.87-Q.20-4) apparaît assez complète pour figurer dans le Tableau 59, en complément des données statistiques.coxal (Fig. 17) : le bassin, os pourtant solide, n’est jamais complet. Sa partie centrale résiste mieux à l’action des Canidés broyeurs d’os et la zone portant l’acétabulum n’est pas rare (8 coxaux g, 10 d). Le tableau 60 fournit la statistique des données existantes. URS.258 (RO.85-S.18-14) correspond à un bassin avec ses deux os coxaux en place. La partie gauche mieux conservée livre un maximum de données dans ce cas particulier.Membres : Tabl. 61-64On retrouve les caractéristiques bien connues d’Ursus spelaeus dans tous les éléments squelettiques, mais l’ours des cavernes de Romain-la-Roche ne correspond jamais aux formes lourdes et de grande taille typiques du stade OIS.4 (« Würm ancien ») que l’on trouve dans le Jura à cette époque.
Tableau 57 : Romain-la-Roche, Ursus spelaeus, atlas.
Atlas U. spelaeuspuema 30
1ddp max
2dt max
3dAp max
4dt art. crânial
5dt art. caudal
6dAp art. max. x3
7dAp min
dors.
8dAp min ventral
n 11 3 3 10 11 9 9 13min 46,4 165 75 72,15 64,68 53,31 23 20,06max 72 234 89 94,2 90,9 90,98 35,29 39,05moy 55,77 206,67 80,00 81,81 75,95 66,55 28,23 29,53écart-type 7,74 - - 7,15 8,10 10,84 4,60 5,61
Tableau 58 : Romain-la-Roche, Ursus spelaeus, axis.
Axis U. spelaeus puema 31
1Hauteur
max
2largeur
3largeur
ant.-post.
4dt art. crânial
5dt tête art.
crâniale max.
6dt tête art.
crâniale min.
7dt
caudal
8dAp caudal
n 2 - 1 4 5 6 1 1min 88,5 -
70,3264,3 11,89 21,01
37,9 23,29max 106,45 - 77,5 15,68 27moy 97,48 - - 72,87 14,49 23,65 - - écart-type - - - 6,03 1,62 1,94 - -

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 543
Les données métriques apparaissent sous forme de tableaux de présentation globale et des remarques complètent la morphologie dans quelques cas particuliers. Afin de les rendre plus concrets, certains tableaux sont complétés par des valeurs individuelles d’os longs, de rotules, de carpe et de tarse, parmi les mieux conservés et les plus complets. Les humérus sont nombreux, 5 complets et 32 partiels déterminables. On ne peut affirmer si les parties proximales et distales proviennent ou non d’un même élément tant qu’elles ne sont pas jointives. C’est sans doute le cas de nombreux fragments et le même problème se répètera pour tous les os longs. La détermination du NMI ne pourra se faire que sur un unique élément g ou d, le mieux représenté, NMI = 12 (12 DT médian g) pour l’humérus.Les radius apparaissent moins nombreux, 4 complets et 20 partiels pour un NMI de 10 individus (10 DT médian g).
L’enregistrement métrique de l’extrémité proximale des cubitus d’Ursus spelaeus n’est pas des plus faciles et oblige à une définition rigoureuse de la façon de prendre les mesures sinon il n’a guère d’efficacité. A Romain-la-Roche 5 cubitus complets et 30 partiels sont conservés, pour un NMI de 15 individus (15 DT médian d).Les fémurs correspondent aux os longs les mieux conservés avec les tibias. Le péroné, os fragile, l’est beaucoup moins. La rotule pourtant très solide apparaît moins représentée. Fémur : n= 7 complets (2 g et 5 d) et 27 partiels (13 g et 14 d), NMI = 14 (14 DT méd d)Rotule : n = 9 complètes (5 g et 4 d) et 6 partielles (2 g et 4 d), NMI = 8 (8 DAP prox d)Tibia : n = 7 complets (5 g et 2 d) et 27 partiels (13 g et 14 d), NMI = 14 (14 DT méd g)Péroné : n = 2 complets (1 g et 1 d) et 18 partiels (9 g et 9 d), NMI = 6 (6 DT méd g)
Tableau 59 : Romain-la-Roche, Ursus spelaeus, omoplate.
Ursus spelaeus
Omoplate Puema 40
1DDP max.(L)
2DDP avec
acromion
3DAP max.distal
4DT max.
avecacromion
5DAP glénoïdal
avec tubercule et processus coracoïde
6DAP art.
7DT art.
8DAP
min. col
9DAP
acromion
n 6 2 0 6 6 10 12 4 8min 245 - - 87,81 75 60,27 37,42 89,06 20,68max 333,7 - - 106 92,1 106,1 55,9 112 52,4moy 276,12 286,00 - 95,49 83,40 71,24 44,50 95,78 36,21écart-type 31,75 - - 6,61 6,00 12,84 5,23 - 12,20
URS.1494 272,5 286,0 - 91,4 81,3 65,6 40,3 91,0 40,0
Tableau 60 : Romain-la-Roche, Ursus spelaeus, coxal.
Ursus spelaeus Coxal
Puema 41
2 3 4 5 6 7 8 9 10 11 12 13
n 11 2 1 1 2 12 2 10 6 5 5 4min 46,21 81,2 130 145 114,5 23,8 163 45,16 39 28,66 16,74 24,66max 70,28 81,2 130 145 189 47,4 180 76,2 59,93 54,9 38,68 47,05moy 60,07 81,20 130,00 145,00 151,75 36,55 171,50 63,78 51,37 42,87 27,74 33,53écart-type 9,14 - - - - 8,40 - 9,76 9,73 9,37 8,23 -URS.258 56,4 81,2 130,0 145,0 114,5 23,8 163,0 60,7 39,0 - - -
1- Longueur maxi.2- DAP int. acétabulum3- DAP foramen obturatum4- L. symphyse pubienne5- DT min. bassin complet6- DT bassin complet entre le milieu des 2 bords supérieurs de l’acétabulum7- DT bord sup. acétabulum
8- DT bassin complet au niveau du contact sacrum9- Hauteur acétabulum (perpendiculaire à 2)10- DT min. ilion11- DT min. ischion12- Epaisseur pédoncule ilion13- Epaisseur pédoncule ischion

544 A. ARgANT
On peut constater que la partie médiane des os longs apparaît toujours la mieux conservée et donc la plus mesurée (DT médian). Cela traduit une plus grande résistance de cette partie non spongieuse et une moins grande attractivité des diaphyses pour les Canidés.Je ne peux entrer dans les détails pour chaque os du carpe et du tarse, mais AltunA (1973), PrAt & thibAult, (1976) et ChAgneAu (1985) ont bien montré toute l’utilisation que l’on peut en faire pour déterminer l’espèce ou comme marqueur de l’évolution.Je ne prendrai que l’exemple du calcanéum (Pl. III, fig. 2, URS.798) et de l’astragale (Pl. III, fig. 3, URS.438).
Les données métriques figurent dans le Tableau 64. Le calcanéum apparaît assez peu représenté, l’astragale à peine un peu mieux. Ce sont pourtant des os épais, massifs, d’habitude résistants. Peut-être y a-t-il un problème d’ordre taphonomique qui m’échappe ? Je ne pense pas à un problème de détermination lors de la constitution du listing par les fouilleurs car ce sont des ossements évidents à reconnaître.Sur le calcanéum d’Ursus spelaeus, la tubérosité d’insertion du ligament calcanéo-cuboïdien est sensiblement oblique par rapport à l’axe principal. Elle l’est plus modérément chez Ursus deningeri, comme chez U. arctos. Cet angle augmente au cours de l’évolution, probablement en rapport avec l’alourdissement des formes typiques des ours des cavernes, conjointement à l’angle de torsion du tibia en lien direct avec l’accentuation du varus du pied (Couturier, 1954 ; AltunA, 1973 ; PrAt & thibAult, 1976).A Romain-la-Roche, l’angle de torsion du tibia, mesuré comme l’indiquent PrAt & thibAult (1976, p. 69) correspond à n = 6, ext. 25-40°, valeur moyenne 34°, écart-type 5,84 (Tabl. 62). F. PrAt constate pour l’Ursus deningeri romeviensis de La Nauterie (c.11 + c.8) un angle de 41° (n = 16) contre 45° pour Ursus spelaeus (n = 22). L’ours de Romain-la-Roche présente donc des valeurs plus faibles que celles de La Nauterie pour ce qui est de la torsion du tibia.A Romain-la-Roche, l’angle de la tubérosité du ligament du calcanéum (n = 3) correspond à 37°- 48°, moy.42°, écart-type 5,50 (Tabl. 64). L’angle mesuré sur le dessin de la figure 35, p. 72 de PrAt & thibAult, 1976 correspond à 35°. L’ours de Romain-la-Roche apparaît donc cette fois plus spéléen que celui de La Nauterie, mais le varus de son pied est sans doute moins marqué.
Fig. 16 : Plan de mesurage de l’omoplate (Puema 40).
3
7
65
9
42 1
Omoplate - Puema 40
8
Fig. 17 : Plan de mesurage du coxal (Puema 41).
10
12
1
2
34
5
67
8
11
Coxal ours - Puema 41
13
9
épaisseurpédoncule ischion
épaisseurpédoncule ilion
cavité cotyloïde

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 545
Tableau 61 : Romain-la-Roche, Ursus spelaeus, membre antérieur.
Membre antérieurUrsus spelaeus
1L
2DT
prox.
3DAP prox.
4DT
méd.
5DAP
médian
6DT dist
7DAP dist. max.
8DT art.
dist. max.
9DT art. prox.
10DAP art. prox
11 12
Humérus Puema 50
n 5 9 5 22 19 10 7 10 11 8 8min 370 60,8 89,5 24,1 25,5 104,8 62,4 63,3 58,6 77,4 102,5max 439,2 97,5 114 51,7 53,6 133,9 79 98,9 91,3 92,5 144,4moy 387,48 84,01 102,87 37,07 37,74 116,74 73,47 82,07 72,38 83,79 132,15écart-type 29,44 10,84 8,88 7,62 6,05 9,57 7,16 11,37 10,32 6,05 14,00URS.128 374,6 77,4 - 40,7 37,2 109,6 62,4 75,4 74,6 - 140,0URS.462 439,2 91,5 104,2 47,3 - 133,9 79,0 92,9 91,3 77,4 -
L DT prox.
DAP prox.
DT méd.
DAP médian
DT dist DAP dist.
DT art.dist. max.
DAP art. dist. max.
RadiusPuema 51
n 5 12 14 18 16 5 6 4 4min 277,6 39,1 27,2 22,9 15 64 33,3 43,6 28,1max 346,9 54,1 55,6 40 23,9 75,5 51,1 57,1 32,4moy 315,02 44,88 36,31 28,54 18,59 69,80 42,42 50,83 30,72écart-type 33,75 4,59 8,78 4,79 2,53 5,32 6,16 6,03 1,95URS.1181 346,9 41,5 31,1 40,0 20,5 71 41,3 48,4 28,1URS.190 279,2 46,7 35,3 31,2 17,3 64,0 39,4 43,6 30,4URS.479 335,6 54,1 40,4 34,0 22,8 75,5 51,1 57,1 32,4URS.493 335,8 39,6 55,6 34,2 23,9 74,0 47,0 54,2 32,0
L DT prox.
DAP prox.
DT méd.
DAP médian
DT dist DAP dist.
CubitusPuema 52
n 5 7 13 28 30 6 6 0 7 8 7 10min 312,7 65,9 30,7 19,4 18,3 22,2 40,5 - 24,6 34,8 80,1 28,7max 392,5 86,6 62,6 41,3 42,8 33,8 50,5 - 36,5 52,8 101,4 47moy 346,90 72,19 48,31 26,68 31,40 27,22 44,95 - 28,45 43,77 88,97 38,84écart-type 35,70 7,77 9,16 4,85 6,20 5,03 3,61 - 4,30 5,71 8,39 5,67URS.1390 392,5 79,2 62,6 23,8 35,8 33,0 47,2 - 24,7 43,3 94,1 47,0URS.1346 312,7 68,7 50,6 25,2 32,9 23,7 42,5 - 27,9 40,4 80,1 40,8URS.404 377,2 86,6 46,2 41,3 31,4 33,8 50,5 - 36,5 47,3 101,4 44,0
Humérus : 11- Distance pointe distale de l’empreinte deltoïdienne au milieu bord inf. ext. art. distale
Cubitus :9- DAP min. diaphyse côté distal10- DAP articulaire prox. min.(serrage int.)11- DAP articulaire prox. max (serrage ext.)12- DT olécrâne (serrage x 3)

546 A. ARgANT
Tableau 62 : Romain-la-Roche, Ursus spelaeus, membre postérieur.
Membre postérieurUrsus spelaeus
1L
2DT
prox.
3DAPprox.
4DT
méd.
5DAP méd.
6DT dist.
7DAP dist.
8DT art.
dist.
9 10
Fémur Puema 60
n 9 10 12 26 24 11 12 8 3 9min 375,0 96,9 46 25,2 19,1 80,7 54,0 75,9 80,3 59,2max 469,2 225,7 60,8 67,09 37,55 107,0 88,2 103,8 91,2 97,6moy 422,81 123,67 53,78 38,59 27,80 95,39 74,67 87,89 86,17 77,51écart type 32,79 36,90 5,00 7,53 4,10 7,81 9,53 8,73 5,50 13,54URS.1026 441,6 114,1 60,0 40,6 31,2 93,0 81,0 89,3 91,2 92,0URS.136 375,0 11,6 48,7 37,9 27,1 90,6 70,5 75,9 - 59,2URS.129 388,5 108,7 52,6 40,0 30,2 89,2 73,4 81,0 - 65,0
Rotule Puema 61
n 13 14 15 9 12min 49,9 36,0 21,8 38,2 36,0max 67,4 49,0 31,1 51,5 46,9moy 59,71 42,26 26,43 45,64 40,94écart type 5,21 4,57 2,67 3,98 3,92URS.1081 49,9 39,6 25,5 38,2 40,7URS.781 67,4 49,0 30,7 51,5 46,6URS.538 57,5 37,4 22,9 45,2 37,2
Tibia Puema 62
n 8 7 10 24 23 11 11 7 9 6min 253,0 75,8 57,7 22,2 21,0 56,4 34,7 47,9 24,2 25,0max 325,1 105,4 90,6 36,6 38,9 75,3 47,8 58,5 35,4 40,0moy 284,46 90,55 73,64 28,18 29,72 68,52 40,18 54,25 30,85 34,17écart type 25,83 10,77 9,77 3,27 4,79 6,15 4,33 4,17 3,71 5,85URS.472 301,6 105,4 78,5 32,8 38,9 74,4 47,5 58,5 33,8 25,0URS.513 253,3 86,8 60,0 29,2 - 62,5 39,3 58,4 29,2 40,0
Péroné Puema 63
n 2 5 5 11 11 13 12min 223,2 17, 9 23,3 10,3 11,4 16,1 24, 9max 263,3 28,9 31,2 14,7 14,6 25,6 33,0moy 243,25 24,98 25,97 11,77 13,02 20,05 29,32écart type 4,30 3,18 1,31 0,99 2,54 2,41URS.1369 263,3 27,6 26,4 14,7 14,6 25,6 33,0URS.1169 223,2 17,9 25,1 10,3 13,5 18,4 24,9
Fémur : 9- Distance entre les deux trochanters (sommet au milieu du petit trochanter)10- Longueur du col
Rotule :9- L. articulaire10- DT articulaire
Tibia :9- DAP articulaire distal (serrage en 3 points)10- Angle de torsion en grades

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 547
Tableau 63 : Romain-la-Roche, Ursus spelaeus, carpe (Fig. 18).
Ursus spelaeus CARPE
1DDP
2DT
3DAP
4DT art.prox.
5DAP art.
dist.
6DAP art.
dist.
7DT art.
dist.
Scapholunaire Puema 531
n 5 8 8 9 11 7 7min 28,9 47,9 49,38 46 26,8 25,62 42,7max 32,28 58 65,8 58 38,3 30,6 50,29moy 30,20 50,81 55,79 51,02 31,99 28,42 45,06écart-type 1,36 3,36 6,34 4,47 3,55 1,90 2,57
PyramidalPuema 532
DAP DT DDP Distance max. de la pièce ligne
directe
Distance de la pièce ppd à l’axe de 4
n 8 7 7 7 7min 19,6 28,67 28,74 40,27 32,53max 26,3 47,26 37,3 48 40,62moy 22,50 36,94 32,84 44,98 36,49écart-type 1,95 6,61 2,91 2,75 2,60
PisiformePuema 533
L max. L min. largeurtête
DT art. DAP art. DT min. col
n 4 4 4 7 8 6min 36,5 35,3 25,7 27,9 13,7 14,41max 51,2 47,55 33 34,75 20,98 22,02moy 45,35 42,81 29,20 31,28 18,14 17,83écart-type 6,62 5,79 3,74 2,52 2,42 2,65
Unciforme Puema 534
DAP DT DDPn 7 7 7min 27,55 31,73 25,14max 34,32 38,02 32,69moy 30,35 34,97 28,10écart-type 2,33 2,11 2,41
MagnumPuema 535
DDP DT DAPn 10 10 9min 24,56 17,98 28,54max 31,6 23,5 36,46moy 27,88 20,20 31,94écart-type 2,88 2,08 2,65
Trapézoïde Puema 536
DDP DT DAPn 7 7 6min 10,96 11,63 23,58max 17,41 19,84 30moy 14,42 16,75 25,81écart-type 2,16 3,57 2,70
TrapèzePuema 537
DAP DT DDPn 5 4 5min 19,54 13,37 20,23max 25,28 17,22 28,3moy 21,92 15,41 23,98écart-type 2,23 2,01 3,54

548 A. ARgANT
Fig. 18 : Plan de mesurage d’os du carpe et du tarse, et de la rotule (Puema 64, 53 et 61).
21
3
5
2
1 3 4
5
6
7
8
4
6
8
45
2
1 3 6
7
2
1
3
4
5
6
2
1 3
2
1
3
9
10
7
AstragalePuema 642
CalcanéumPuema 641
Tarse
Carpe
ScapholunairePuema 531
PisiformePuema 533
UnciformePuema 534
RotulePuema 61

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 549
Tableau 64 : Romain-la-Roche, Ursus spelaeus, tarse (Fig. 18).
Ursus spelaeus Tarse
L DT art. avec
astrag.
DT tuber calcanei
DAP tuber
calcanei
DT prox. max.
DDP prox. max.
DT min. diaphyse
DAP min. diaphyse
Angle en degrés
Calcanéum Puema 641
n 5 3 4 5 10 11 4 4 3min 89,13 43,65 32 31,86 54 37,7 16,24 31,81 37max 110,7 52,78 38,32 42,2 71,85 51,53 20,47 37,87 48moy 97,10 47,95 36,15 36,96 60,64 44,68 17,77 34,13 42,33écart-type 8,10 4,59 2,87 3,85 5,95 4,81 1,86 2,64 5,51
Astragale Puema 642
DAP DT DDP DDPmini
DT art. max.
trochlée
DT mini col
DT max. tête
L diagonale max(ligne directe)
n 12 12 11 12 11 12 11 11min 46,2 51,18 30 19,42 35,5 26,96 30,6 52,48max 62,1 66,2 36,8 29,63 67 35,56 39,86 76,4moy 51,52 56,01 33,51 26,83 42,88 29,73 35,76 63,05écart-type 4,28 4,61 2,12 3,03 8,34 2,15 2,80 6,00
Cuboïde Puema 643
DAP DT max. DDP n 9 9 8 min 24,93 30,4 27,02 max 29,8 40 42,2 moy 27,17 35,08 35,17 écart-type 1,83 2,72 6,18
Naviculaire Puema 644
DAP DT DDP n 7 8 7 min 1 2 3 max 19,3 40,3 43,4 moy 17,11 36,16 38,34 écart-type 2,52 2,94 4,02
1er cunéiforme Puema 645
n 8 7 7 min 14,25 13,09 26,35 max 19,5 16,6 30,3 moy 16,38 14,46 27,04 écart-type 1,49 1,21 1,44
2ème cunéiforme Puema 646
n 3 3 3 min 11,13 13 20,09 max 14,73 16,5 25,62 moy 13,05 14,82 22,66 écart-type 1,81 1,75 2,78
3ème cunéiforme Puema 647
n 6 6 6 min 12,3 17,4 25,9 max 16,7 22,1 30,8 moy 15,36 20,24 28,58 écart-type 1,59 1,62 1,71

550 A. ARgANT
Les facettes articulaires de la face plantaire de l’astragale présentent une courbure moins accentuée chez Ursus spelaeus que chez Ursus arctos et Ursus deningeri. La facette médiane approche l’angle plat (180°) dans certains cas chez l’ours des cavernes. A Romain-la-Roche la concavité reste accusée et se rapproche plus de celle d’Ursus deningeri. On n’observe jamais pour ces deux éléments du tarse, les fortes dimensions que l’on rencontre chez les formes du Pléistocène supérieur moyen (« Würm ancien »).Métapodes :Métacarpiens : Tabl. 65Un lot de quatre métacarpiens d consécutifs appartient à un même individu :- 1 Mtc.2 : URS.1421, RO.87-P.18-19- 1 Mtc.3 : URS.1374, RO.87-P.19-1- 1 Mtc.4 : URS.1420, RO.87-P.18-19- 1 Mtc.5 : URS.1375, RO.87-P.19-1Ils sont endommagés distalement de la même façon pour les trois derniers, les tailles correspondent et ils ont été
découverts dans deux carrés proches (P.18 et P.19), dans la même couche 8a.Métatarsiens : Tabl. 66Phalanges et sésamoïdes : de nombreuses phalanges et de nombreux sésamoïdes sont déterminables au genre Ursus si on admet qu’il n’est pas possible de trancher avec certitude entre U. arctos et U. spelaeus. Ils figurent dans l’inventaire en annexe (à U. spelaeus) sans être mesurés.
II.2.2.2. BiochronologieLes grandes tendances évolutives de la lignée de l’ours des cavernes Ursus deningeri-Ursus spelaeus ont été bien définies :- augmentation de la taille,- alourdissement des formes,- développement des bosses frontales modifiant le profil
supérieur du crâne au niveau du front et de la glabelle,- disparition progressive des prémolaires antérieures aux
quatrièmes prémolaires,
Tableau 65 : Romain-la-Roche, Ursus spelaeus, métacarpiens.
Ursus spelaeus Métacarpiens 1L
2DT prox
3DAP prox.
4DT méd.
5DAP méd.
6DT dist.
7DAP dist.
8DT art.
dist.
Mtc.1 Puema
541
n 10 12 12 11 11 11 6 7min 56,56 18,75 16,53 7,08 4,79 12,47 11,17 14,9max 71,04 28 24,65 14,67 13,98 21,53 16,4 20,95moy 62,71 24,21 20,00 12,05 10,64 18,31 15,15 17,46écart-type 4,84 2,60 2,44 2,03 2,55 2,51 2,00 1,93
Mtc.2 Puema
542
n 12 15 1 17 17 13 10 10min 61 13,9 - 11,4 9 17,7 17,28 16,7max 83,77 23,5 - 17,68 14,62 25,82 21,1 21,98moy 74,08 18,09 24,41 14,41 12,13 22,22 18,90 19,42écart-type 6,25 2,32 - 1,71 1,55 2,01 1,11 1,43
Mtc.3 Puema
543
n 7 10 7 10 10 8 5 6min 73,4 17,8 25,8 14,2 11,9 22,1 19,4 17,8max 86,53 21,3 30,75 17,15 14,55 26,9 21,25 24,22moy 78,32 19,74 28,06 15,69 13,06 23,91 19,91 21,18écart-type 5,45 1,36 2,02 1,21 0,79 1,87 0,77 2,43
Mtc.4 Puema
544
n 12 15 10 13 13 10 9 7min 72,8 14,96 24,72 14,76 11,54 23,89 18,41 2,9max 88,73 25,39 34,65 18,61 14,77 28,8 24,38 25,82moy 78,91 20,65 29,16 16,40 13,50 25,39 20,96 20,03écart-type 5,02 2,34 3,22 1,16 0,89 1,77 1,82 7,84
Mtc.5 Puema
545
n 7 11 13 14 14 12 7 7min 75,2 25,21 25,96 11,33 12,22 21,71 17,38 22,8max 89,86 31,2 35,61 19,5 17,5 31,9 22,23 28,6moy 81,51 27,93 30,63 16,79 14,44 26,92 19,76 25,83écart-type 5,84 2,17 2,70 2,48 1,73 2,79 1,78 2,17

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 551
- apparition de tubercules secondaires aux P4/ et P/4,- complication croissante du relief des couronnes des
différentes molaires et réduction des arrière-molaires,- raccourcissement relatif des segments distaux des
membres,- augmentation de la brièveté relative des métapodes 1
par rapport aux métapodes 5,- augmentation de la torsion du tibia et modification des
os du carpe et du tarse, notamment du calcanéum et de l’astragale.
II.2.2.2.1. Comparaison dentaireg. rAbeder a étudié minutieusement l’évolution de la denture de l’ours des cavernes comme marqueur de l’évolution. La méthode morphodynamique qu’il a mise au point sur les quatrièmes prémolaires (P4/, P/4) (rAbeder 1983, 1989a, 1991) a été ensuite étendue à toute la denture (rAbeder, 1999). La comparaison avec un corpus de sites bien datés présentant tous des séries dentaires numériquement importantes (20 individus constituant un strict minimum), fournit un cadre de comparaison rigoureux. L’application de cette méthode
sur le matériel de la Balme à Collomb (Savoie) (P4/, P/4) m’a déjà permis d’avancer un élément de datation satisfaisant de cette population d’Ursus spelaeus (ArgAnt, 1995). Le gisement de Romain-la-Roche fournit plus de 20 quatrièmes prémolaires supérieures (n = 27) et inférieures (n = 22) d’Ursus spelaeus. Il devient possible d’appliquer la méthode de rAbeder à cette population.Les Figures 19 et 20 présentent les morphotypes des P4/ et P/4 dont la présence statistique associée à un coefficient évolutif permet de calculer les indices morphodynamiques de ces deux prémolaires (Tabl. 67 et 68). Le croisement des valeurs de ces deux indices place le gisement de Romain-la-Roche parmi les gisements ayant atteint un degré évolutif comparable de leur population d’Ursus spelaeus (Fig. 21). L’ours de Romain-la-Roche se situe chronologiquement après les formes de la fin du Pléistocène moyen comme Schartzfeld, Hunas, Repolüst en Allemagne et se retrouve proche des formes des couches profondes d’Herdengelhöhle (Autriche), entre 112 et 135000 ans (döPPes et al., 2008). Les gisements de Château (Saône-et-Loire) et d’Azé (Saône-et-Loire)
Tableau 66 : Romain-la-Roche, Ursus spelaeus, métatarsiens.
Ursus spelaeusMétatarsiens 1
L2
DT prox.3
DAP prox.4
DT méd.5
DAP méd.6
DT dist.7
DAP dist.
8DT art.
dist.
Mtt.1 Puema 651
n 5 7 7 6 6 5 4 3min 48,65 19,99 19,29 9,36 10,27 15,71 13,29 14,88max 59,45 27,77 25,29 11,99 13,97 20,04 15,3 16,55moy 52,47 22,37 22,15 10,31 11,88 17,27 14,20 15,98écart-type 4,32 2,63 2,09 0,98 1,45 1,70 0,94 0,95
Mtt.2 Puema 652
n 7 9 9 9 9 7 7 5min 56,42 9,92 17,64 7,18 5,26 11,68 9,63 16,23max 68,15 16,48 23,76 12,96 10,79 19,49 16,24 17,45moy 63,47 13,84 21,24 11,01 9,02 16,89 14,04 16,80écart-type 3,87 2,25 2,34 1,88 1,94 3,07 2,17 0,51
Mtt.3 Puema 653
n 8 10 9 9 10 9 6 7min 63,78 12,47 20,43 8,37 6,62 14,71 14,6 15,7max 85,7 22,3 32,7 16,2 14,1 23,1 17,5 20,5moy 73,20 17,38 25,71 13,03 10,49 19,68 16,23 17,56écart-type 6,52 2,97 4,16 2,11 2,11 2,40 1,26 1,75
Mtt.4 Puema 654
n 8 13 9 13 13 9 6 6min 70,99 12,9 22,4 9,59 8,29 17,21 15,98 18,31max 87,21 23,2 33,4 18,4 16,3 26,5 18,76 23,1moy 79,64 19,64 27,95 14,72 12,63 22,00 17,74 20,11écart-type 4,68 2,85 2,94 2,25 1,93 2,49 1,03 1,66
Mtt.5 Puema 655
n 4 8 7 9 9 6 3 2min 75,24 21,14 24,33 8,49 9,07 20,49 17,42 21,57max 94,39 33,14 33,1 15,1 16,7 25,9 19,49 25,29moy 83,02 27,67 28,95 13,03 13,65 24,06 18,70 23,43écart-type 8,32 3,69 3,45 2,04 2,13 1,98 1,12 2,63

552 A. ARgANT
morp
hodynamisc
her index
0
1
2
3
4
Höcker 1 Höcker 2
Proto
loph.
Met
alo
ph
.
A/D
D/F
D E
F
A B C
Fig. 19 : Schéma morphodynamique de la couronne de la P4/ d’Ursus spelaeus : morphotypes A, B, C, D, E, F (rAbeder, 1992, fig. 1, p. 134).
0
1
2
3
3,5
4
4,5
5
1,5
2,5
0,5
E1 E2 E3
F2 F3
D1 D2 D3
C1 C2
B2
A
B1
C3
morp
hodynamisc
her index
Hypolophid
Mphd
Hyd.
Pad
Med
access. Höcker
Fig. 20 : Schéma morphodynamique de la couronne de la P/4 d’Ursus spelaeus : morphotypes A, B, C, D, E, F (rAbeder, 1992, fig. 2, p. 135).

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 553
sont nettement antérieurs (ArgAnt, 2002, 2009) et le gisement de la Balme-à-Collomb (Savoie) bien plus récent (ArgAnt, 2004).Dans l’étude dentaire, je me suis contenté d’indiquer pour chaque dent les morphotypes correspondant à Ursus spelaeus de Romain-la-Roche en renvoyant à la publication de g. rAbeder. Je n’ai pas calculé les indices morphodynamiques pour les molaires car cela devient très complexe Mais les morphotypes rencontrés (plusieurs types par dent) correspondent avec cette position chronologique indiquée par les quatrièmes prémolaires. Ils se rapportent généralement à des formes intermédiaires entre Ursus deningeri de Hunas (Allemagne) de la fin du Pléistocène moyen et Ursus
spelaeus de Herdengelhöhle (Autriche) du « Würm moyen » dont les dessins schématiques des couronnes respectives servent d’exemple (rAbeder, 1991).M/1 :La première molaire inférieure M/1 enregistre également bien l’évolution, à condition toujours de disposer d’une série numériquement suffisante pour représenter les gisements. Le Tableau 69 et la Figure 22 montrent la position théorique de l’ours de Romain-la-Roche. Ursus etruscus est représenté par Saint-Vallier (Drôme) et le Val d’Arno (Italie) (in PrAt & thibAult, 1976), Ursus deningeri par Mosbach 2 (in PrAt & thibAult, 1976), Sprimont (Belgique) (KleCzKo, 1999), Château (Saône-et-Loire) (ArgAnt, 1991), Nauterie c.11 (PrAt
Tableau 67 : Romain-la-Roche, Ursus spelaeus, P4 supérieure. Calcul de l’indice morphodynamique (RAbeder, 1990, p. 108).
P4/Morphotypes G.
Rabeder
(1)Nombre
(2)Coefficient évolutif
(3)Produit (1) x (2) = 3
(4)Fréquence du type
(%)A 16 0 0 59,2
A/D 3 1 3 11,1B 7 1 7 25,9C 1 2 2 3,7D 0 2 0 0E 0 3 0 0
D/F 0 3 0 0F 0 4 0 0
Total 27 - 12 100Indice morphodynamique : Im = (3) x 100 : (1) = 12 x 100 : 27 = 44,4
Tableau 68 : Romain-la-Roche, Ursus spelaeus, P4 inférieure. Calcul de l’indice morphodynamique (RAbeder, 1990, p. 108).
P/4Morphotypes G.
Rabeder
(1)Nombre
(2)Coefficient évolutif
(3)Produit (1) x (2) = 3
(4)Fréquence du type
(%)A 1 0 0 4,5
B.1B.2C.1 12 1 12 54,5C.2 4 2 8 18,2C.3D.1 4 1,5 6 18,2D.2E.1 1 2 2 4,5
Total 22 - 28 100Indice morphodynamique : Im = (3) x 100 : (1) = 28 x 100 : 22 = 127,2

554 A. ARgANT
& thibAult, 1976) du Pléistocène moyen médian et Nauterie c.8 (PrAt & thibAult, 1976), gigny c.XXVI (ChAgneAu, 1989), Azé 1-3 et 1-2 (ArgAnt, 1991) du Pléistocène moyen récent. Ursus spelaeus du Pléistocène supérieur correspond aux Furtins-Ossuaire 2 (Saône-et-Loire) (leroi-gourhAn, 1950), La Balme-à-Collomb (Savoie) (ArgAnt, 1995), gamsulzenhöhle Ens.1 et Ens.2 (Autriche) (rAbeder, 1995 a et b), Arcy-sur-Cure (Yonne) (bAryshniKov & dAvid, 2000 : Pm = Paléolithique moyen, Ps = Paléolithique supérieur), Châteaubourg (Ardèche) (PAutret-homerville, 2000) et große grotte (Allemagne) (WeinstoCK, 1999). Ces documents rendent bien compte de la croissance relativement progressive en longueur et en largeur de la dent, en classant les gisements dans l’ordre chronologique attendu. Il est vrai que dans le domaine des datations, on ne dispose pas toujours de données uniformes et de fiabilités comparables. Cette progression ne présente pas sans doute non plus toute la régularité théorique supposée.
L’attribution stratigraphique du matériel n’a pas toujours la finesse souhaitable ou n’est pas indiquée avec précision. En tout cas, la longueur moyenne des M/1 de la population de l’ours des cavernes de Romain-la-Roche se situe après Azé (1-3 et 1-2), approximativement entre les ours de La Nauterie c.8 du Pléistocène moyen plutôt récent et les ours de la grotte des Furtins, bien avant les grandes formes typiques du Pléistocène supérieur moyen ou récent (gamssulzenhöhle, Châteaubourg, Arcy, große grotte).
II.2.2.2.2. Comparaison des métapodesLes métapodes voient leur massivité augmenter au cours de l’évolution. Pour comparer le matériel de Romain-la-Roche, les gisements médioeuropéens et plus spécialement les plus proches géographiquement (Bourgogne, Franche-Comté, Jura, Alpes du Nord) ont été privilégiés, pour limiter au maximum la variabilité géographique. De nombreux gisements régionaux livrent
RO
Azé 1-3
Fig. 21 : Utilisation des indices morphodynamiques des quatrièmes prémolaires pour déterminer le degré évolutif de l’Ursus spelaeus de Romain-la-Roche. Datation par comparaison avec de nombreux gisements d’Europe moyenne, souvent datés par des méthodes absolues, 14C ou Uranium-Thorium (rAbeder, 1991, p. 119).

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 555
de l’ours des cavernes Ursus deningeri ou Ursus spelaeus : Vergranne (Doubs) du Pléistocène moyen médian, Château-Ensemble Nord (Brèches 2 et 3) du Pléistocène moyen médian (Saône-et-Loire), Villereversure (Ain), Santenay (Côte-d’Or), Les Furtins-Ossuaire 2 (Berzé-la-Ville, Saône-et-Loire), Saint-Romain (Côte-d’Or), Arcy-sur-Cure (Yonne)… Certains ne fournissent que des séries limitées et il n’est malheureusement pas possible de les utiliser tous pour la comparaison des métapodes.L’ours des cavernes de Romain-la-Roche est bien Ursus spelaeus, il n’y a aucun doute là-dessus. Le gisement
d’Azé 1-3 en Bourgogne constitue pour cette comparaison le jalon chronologique du Pléistocène moyen récent (OIS.6). Les termes les plus récents seront constitués par le matériel de la Balme-à-Collomb (Entremont-le-Vieux, Savoie) que j’étudie et celui de goyet (Condroz, Belgique) dans la vallée de la Meuse, altitude 350 m, « Chambre B », lot B.4 (NR = 2184, NMI = 89, datation : 14C-AMS : 36500 +1040/-920 et 35470+780/-710 BP) (germonPré & sAblin, 2001 ; Athen et al., 2005). La grotte des Furtins (Saône-et-Loire) fouilles de Leroi-gourhan (leroi-gourhAn, 1950), ossuaire 2, appartient
Tableau 69 : Romain-la-Roche, Ursus spelaeus, M/1 (MD x VL), degré évolutif de la population comparé à celui de populations de différents sites européens.
1- Saint-Vallier (Drôme) in PrAt & thibAult, 19762- Val d’Arno (Italie) ibid.3- Mosbach 2 (Allemagne) in bAllesio, 19834- Belle-Roche, Sprimont (Belgique), KleCzKo, 19995- Château Ensemble Nord, Brèche 2, ArgAnt, 19916- Nauterie, la Romieu (c.11), PrAt & thibAult, 19767- gigny c. XXVI inférieure, ChAgneAu, 19898- Azé 1-3 (Saône-et-Loire), ArgAnt, 19919- Azé 1-2 (Saône-et-Loire), ArgAnt, 199110- Nauterie, la Romieu (c.8), PrAt & thibAult, 197611- Furtins, ossuaire 2 (Berzé-la-Ville, Saône-et-Loire),
ArgAnt, 1991
12- Balme à Collomb (Entremont-le-Vieux, Savoie), ArgAnt, 1995
13- gamssulzenhöhle (Autriche), secteur 1, rAbeder, 1995 14- gamssulzenhöhle (Autriche), secteur 3, rAbeder, 1995 15- Arcy-sur-Cure (Yonne), Paléolithique moyen,
bArysChniKov & dAvid, 200016- Arcy-sur-Cure (Yonne), Paléolithique supérieur, ibid.17- Châteaubourg (Ardèche), PAutret-homerville, 200018- große grotte (Allemagne), WeinstoCK, 199919- Ursus spelaeus, Sud France, PrAt & thibAult, 197620- Ursus spelaeus, Europe centrale, rode in bAllesio,
198321- Romain-la-Roche (Doubs), comparaison
M/1Ursus etruscus Ursus deningeri -------------------------------------------------->
1 2 3 4 5 6 7
Sites SV Val d’Arno Mosbach2 Sprimont CHA.1 Naut. c.11 gigny.c.XXVI
MDn 8 8 26 127 17 17 13moy. 23,80 23,86 27,00 27,50 27,33 27,70 27,77
VLn 8 7 26 127 24 17 13moy. 11,75 10,68 13,09 13,10 13,17 13,17 13,24
M/1Ursus spelaeus --------------------------------------------------------------------------------->>
8 9 10 11 12 13 14Sites Azé 1-3 Azé 1-2 Naut. c.8 Furtins B à C. gS.1 gS.3
MDn 81 12 28 5 79 46 28moy. 27,37 27,49 28,24 28,76 28,87 29,80 30,01
VLn 79 12 25 5 82 44 28moy. 13,51 13,76 13,66 13,76 13,84 14,27 14,29
M/1Ursus spelaeus ----------------------------------------------------------------------------------->
15 16 17 18 19 20 21Sites Arcy P.m. Arcy P.s. Chatbg.07 große grotte Ursus spelaeus Romain-la-Roche
MDn 8 25 17 23 83 208 22moy. 30,30 30,70 31,03 31,10 30,79 30,3 28,26
VLn 8 25 18 22 83 208 24moy. 14,90 15,00 14,81 14,80 14,8 14,66 13,84

556 A. ARgANT
probablement à un épisode tempéré antéwürmien (« Préwürm ») correspondant à la phase Saint-germain II de la grande Pile comme le suggère la palynologie (ArgAnt, 1991).La Baume d’Echenoz-la-Méline (couche VIII) couche fossilifère inférieure, est datée « d’une phase plutôt interstadiaire du Pléistocène moyen » (CAmPy, 1982). L’ours est Ursus deningeri (ChAgneAu, 1985).Le Tableau 70 et la Figure 23 montrent la position des indices de massivité de tous les métapodes (Mtc et Mtt) d’Ursus spelaeus de Romain-la-Roche, comparés à ceux d’ autres gisements. L’indice de massivité Im traduit le rapport entre la largeur de la diaphyse en son milieu strict et la longueur maximum de la longueur de ce métapode (Im = DT med x 100 / L).Une première remarque concerne les effectifs. La population de Romain-la-Roche ne fournit pas une série très étoffée, variant de 4 à 12 éléments selon le métapode, ce qui reste un peu faible sur le plan statistique et peut apporter un certain déphasage. La série des courbes obtenue traduit bien la complexité de cette évolution, tout en dégageant de grandes tendances évolutives. Selon le rayon et selon que le métapode soit antérieur ou
postérieur, la position de Romain-la-Roche varie. C’est le Mtc.4 qui classe le mieux les gisements dans l’ordre chronologique admis pour les sites de référence. Dans ce cas supposé favorable, l’indice de massivité du quatrième métacarpien de l’ours des cavernes de Romain-la-Roche vient s’insérer entre Azé 1-3 et Les Furtins-Ossuaire 2 (Fig. 24).L’ours des cavernes de Romain-la-Roche n’atteint pas la grande taille, ni la massivité des formes du stade OIS.4 (« Würm ancien »). Tous les éléments de comparaison montrent que d’après son degré évolutif cette population se situe dans la fourchette chronologique allant de la fin du stade OIS.6 (« Riss final ») à la fin du stade OIS.5 (« Riss-Würm »).
II.2.2.2.3. PaléoécologieL’analyse palynologique prouve la présence de l’ours des cavernes en milieu forestier, sous climat très tempéré dans la grotte des Furtins, ossuaire 2 (ArgAnt, 1991). A contrario, en Bourgogne à gissey-sur-Ouche (Côte d’Or) ou à Châteaubourg (Ardèche) l’analyse pollinique par J. ArgAnt de coprolithes de hyènes associés aux vestiges d’Ursus spelaeus montrent des conditions climatiques
10
11
12
13
14
15
16
23 24 25 26 27 28 29 30 31 32MD
VL
U. etruscusU. deningeriU. spelaeusRomain-la-RocheLinéaire (Ensemble)
SV
Val d'Arno
Mosbach 2CHA.1
Azé 1-2Azé 1-3
Sprimont
Naut.c8
Naut.C11
FurtinsB à C
Gigny
GS.1GS.3
Arcy PmArcy Ps
Chatbg
Große Grotte
Romain la Roche
Degré évolutif de la M1 inférieure
Fig. 22 : Diagramme de répartition de MD x VL des M/1 d’Ursus etruscus, U. deningeri et U. spelaeus de différents sites européens. Place de Romain-la-Roche dans cet ensemble.

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 557
Tableau 70 : Indice de massivité (Im) des métapodes d’Ursus spelaeus de plusieurs gisements.
Ursus spelaeusIndice de massivité des
métapodes
Romainla-Roche Azé 1-3 Echenoz c.8 Furtins Balme à
Collomb Goyet B4
Mtc1 18,80 19,90 20,70 21,10 21,30 19,85Mtc2 20,00 21,40 21,80 23,00 23,40 22,69Mtc3 19,90 20,30 20,40 20,60 20,90 20,99Mtc4 21,14 20,60 20,30 21,40 22,07 22,17Mtc5 21,20 22,20 21,50 22,20 22,26 22,53Mtt1 19,90 20,60 19,90 23,20 20,92 22,60Mtt2 17,50 20,30 19,20 20,40 20,10 20,50Mtt3 18,10 18,70 18,40 20,00 19,80 20,99Mtt4 17,60 18,60 18,00 18,90 18,89 19,24Mtt5 14,90 16,30 16,60 16,50 15,28 16,25
Fig. 23 : Comparaison de l’indice de massivité (Im) des métapodes de Romain-la-Roche à celles de différentes populations d’Ursus
spelaeus.
Echenoz c.8
Azé 1-3
Romain-la-Roche
Furtins Balme à Collomb
Goyet B.4
Mtc.4 CHRONO
Fig. 24 : Position d’Ursus spelaeus de Romain-la-Roche par rapport à différentes populations d’Ursus spelaeus, d’après l’indice de massivité des métapodes.

558 A. ARgANT
particulièrement rigoureuses (pelouses à armoises). Ursus spelaeus présente donc un caractère ubiquiste s’adaptant à un large éventail de biotopes, allant du milieu forestier au milieu plus ou moins steppique. Mais il ne faut pas oublier qu’il est très largement végétarien. Il a besoin d’une réelle biodiversité végétale pour se nourrir. On peut supposer une dynamique de ses populations adaptée à la recherche des conditions locales les plus favorables lors des péjorations climatiques sévères. En tout cas l’analyse palynologique des coprolithes de hyènes de Romain-la-Roche indique un milieu tempéré frais dominé par le pin (Pinus silvestris), le frêne (Fraxinus) et le chêne (Quercus). Les espèces héliophiles qui les accompagnent excluent une formation arbustive serrée et évoquent plutôt un boisement lâche avec des espaces ouverts (J. ArgAnt, ce volume).
II.3. Ordre CarnivoraSuper-famille Feloidae
Famille FelidaeSous-famille Felinae
Trois espèces sont représentées à Romain-la-Roche : Felis silvestris le chat sauvage, Panthera pardus, la panthère et surtout Panthera spelaea, le lion des cavernes. Les deux premières espèces ne sont finalement attestées que par un matériel réduit.Le lion des cavernes se rencontre par contre dans de nombreux gisements paléontologiques, habituellement par de rares pièces. Il apparaît cette fois bien représenté à Romain-la-Roche qui livre un beau matériel appartenant à quatre individus au moins, dont trois crânes « complets ». Les vestiges post-crâniens de l’individu 3 de la couche VIII b par exemple, traduisent une dispersion réduite à quelques carrés de fouille contigus (PAuPe, 1996, p. 56-57, fig. 28).
II.3.1. Genre FelisEspèce : Felis silvestris Schreber, 1777
Un chat sauvage est documenté à Romain-la-Roche. Les vestiges proviennent de la couche VIII de la base. Mise à part la phalange 1 (FEL. 249) trouvée dans le carré T. 17 (z = 621 cm de profondeur) toutes les autres pièces proviennent de carrés proches (z compris entre 662 et 665 cm de profondeur) et correspondent manifestement à un seul et unique individu. La liste d’inventaire du matériel disponible apparaît en annexe à la fin (NR = 10, NMI = 1).
Canine sup. g : FEL.226 (RO.86-R18-101) (Tabl. 71, Pl. V, fig. 2)Cette canine non usée montre nettement la présence des doubles sillons parallèles à l’intérieur et à l’extérieur des bords vestibulaire et lingual de la couronne.
Le radius FEL.248, à extrémité distale manquante, de couleur brun-noir, très fossilisé. Il présente une forte taille et de puissantes attaches musculaires (Tabl. 72).Mtc.2 g : FEL.164 (RO.88-P.13-10) (Tabl. 73)Tarse : Calcanéum : FEL.228 g (RO.89-P.14-3) (Tabl. 74)Classiquement, le manubrium du calcanéum FEL.228 apparaît relativement long et déprimé latéralement et le tuber calcanei dissymétrique avec bord médial nettement relevé. Le bord antérieur médial de la surface articulaire répondant à l’astragale paraît par contre un peu plus étiré, ce qui repousse légèrement vers l’extérieur la surface articulaire antérieure répondant au cuboïde.Cuboïde : FEL.160 g (RO.88-P.13-10) (Tabl. 75)3° cunéiforme : FEL.238 g (RO.88-HS.152) (Tabl. 75)Phalanges :Phalanges 1 : FEL.249 (doigt 4 ant. d) (RO.87-P.13-1), FEL.260 (RO.87-T.17-41) (Tabl. 76)Phalanges 2 : FEL.158 g (RO.88-P.13-8), FEL.159g (RO.88-P.13-9) (Tabl. 76)Il est malheureusement impossible d’utiliser un matériel aussi réduit pour rechercher des informations biochronologiques. La seule indication de forte taille sur un unique individu à Romain-la-Roche (un fort mâle ?) limite la valeur des comparaisons. Le calcanéum par exemple apparaît de taille nettement supérieure à celui de la Caune de l’Arago (Pyrénées-Orientales), séquence supérieure, sol g, H.17 HQY3 2275 (Crégut, 1979) et à plus forte raison à celui d’un Felis silvestris actuel.Du point de vue paléoenvironnemental, le chat sauvage traduit un milieu forestier alternant avec des terrains découverts et des broussailles, mais également des savanes boisées (vAn den brinK, 1967) et apporte ici une indication de contexte climatique tempéré-froid à tempéré.
II.3.2. Genre PantheraEspèce : Panthera pardus (Linné, 1758)
Quelques pièces attestent de la présence de Panthera pardus à Romain-la-Roche. La liste du matériel disponible apparaît en annexe à la fin (NR = 7, NMI = 1).La diaphyse FEL. 251 seule conservée, indique un tibia droit long et relativement fin (RO. 87-S.17-25). Il manque les informations des épiphyses, mais ce tibia appartient manifestement à un félidé. Il a une taille trop forte pour être celle d’un lynx et elle est deux fois plus petite que celle d’un lion des cavernes. Je l’attribue sans certitude, mais avec une bonne probabilité, à Panthera pardus (DT méd = 19,7 ; DAP méd = 22,2).Pisiforme : FEL.252 g (RO.88-Q.19-52) (Tabl. 77)Astragale : FEL.250 d (RO.87-HS-66) (Tabl. 78)Mtt.4 : FEL.227 g (RO.83-Q.17-3) (Tabl. 79, Pl. V, fig. 4)Phalanges 1 : FEL.254 (RO.87-T.17-1) doigt 5 d ; FEL.255 (RO.86-R.18-96) doigt 3 d (Tabl. 80)Phalange 2 : FEL.41 (RO.86-Q.18-127) (Tabl. 80)

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 559
Tableau 71 : Romain-la-Roche, Felis silvestris, canine supérieure gauche.
Felis silvestrisCanine sup. g
Pfema 14 MD VL HC HT U
FEL.226 9,7 13,1 29,4 - 0
Tableau 72 : Romain-la-Roche, Felis silvestris, radius.
Felis silvestris Radius g Pfema 51
L DT prox DAP prox DT méd DAP méd DT dist DAP dist
FEL.248 - 7,9 10,8 5,0 8,7 - -
Tableau 73 : Romain-la-Roche, Felis silvestris, métacarpien 2.
Felis silvestris Mtc.2 g
Pfema542L DT prox DAP prox DT méd DAP méd DT dist DAP dist
FEL.164 39,9 5,2 8,7 4,0 3,6 5,5 6,3
Tableau 74 : Romain-la-Roche, Felis silvestris, calcanéum.
Felis silvestris Tarse calcanéum gPfema 641
L DT articulaire DT tuber calcanei
DDP tuber calcanei
DT max projection
DDP max
FEL.228 40,9 14,3 10,2 11,2 17,7 14,7Arago sol g 37,9 14,2
actuel 33,4 14,2
Tableau 75 : Romain-la-Roche, Felis silvestris, cuboïde et 3e cunéiforme.
Felis silvestris -Tarsecuboïde gPfema 643
DAP DT DDP
FEL.160 11,8 - -3° cunéiforme g
Pfema 647 DAP DT DDP
FEL.238 9,0 8,8 14,2

560 A. ARgANT
Tableau 76 : Romain-la-Roche, Felis silvestris, phalanges 1 et 2.
Felis silvestris Phalanges 1
Pfema 66L DT prox DAP prox DT méd DAP méd DT dist DAP dist
FEL.249doigt 4 ant. d 20,1 6,4 5,4 4,2 3,2 4,8 4,5
FEL.260 18,2 7,4 6,1 3,9 4,2 5,7 4,5Phalanges 2
Pfema 74 L DT prox DAP prox DT méd DAP méd DT dist DAP dist
FEL.158 16,3 5,7 5,4 3,0 3,9 5,0 4,2FEL.159 16,8 6,1 5,6 3,4 3,5 5,2 4,7
Tableau 77 : Romain-la-Roche, Panthera pardus, pisiforme.
Panthera pardus Pisiforme g Pfema 533
DDP (x3) DAP max DT articulaire DAP articulaire DT min col
FEL.252 23,3 13,8 15,0 7,6 7,4
Tableau 78 : Romain-la-Roche, Panthera pardus, astragale.
Panthera pardus Astragale d Pfema 642
L DT max DDPmax
DT artic. trochlée
DT min col DT max tête
DDP tête L maxdiagon.
FEL.250 37,4 35,2 21,6 22,5 16,1 21,2 17,3 42,8
Tableau 79 : Romain-la-Roche, Panthera pardus, métatarsien 4.
Panthera pardus Mtt.4 g
Pfema 654L DT prox DAP prox DT méd DAP méd DT dist DAP dist
FEL.227 - 15,3 18,3 10,4 9,3 - -actuel 14,1 16,9 8,2 8,2
Tableau 80 : Romain-la-Roche, Panthera pardus, phalanges 1 et 2.
Panthera pardus Phalange 1Pfema 73
L DT prox DAP prox DT méd DAP méd DT dist DAP dist
FEL.254 doigt 5 d 33,8 11,2 10,7 7,2 7,7 9,6 7,3FEL.255 doigt
3 d.40,0 14,2 11,7 9,5 7,8 11,1 9,2
Phalange 2Pfema 74 L DT prox DAP prox DT méd DAP méd DT dist DAP dist
FEL.41 24,0 10,4 9,3 6,7 6,9 10,8 7,8

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 561
La panthère se rencontre dans la couche VIIIa, c’est-à-dire à la base du remplissage, tout comme Felis silvestris et Panthera spelaea. La taille de l’individu de Romain-la-Roche se situe légèrement au-dessus de celle d’un individu actuel comme le montre la comparaison du Mtt.4.Du point de vue paléoenvironnemental, la panthère est un animal ubiquiste et particulièrement eurytherme. Elle n’apporte guère d’indication dans ce cas précis, si ce n’est par les indications du contexte général de la couche où on la rencontre.
II.3.3. Genre PantheraEspèce : Panthera spelaea (GoLdfuss, 1810)
Le lion des cavernes de Romain-la-Roche fournit un important matériel paléontologique, rare d’habitude. Trois individus jeunes adultes livrent tous les éléments du squelette. Un très jeune individu s’y ajoute, avec une moins bonne conservation il est vrai. Il a même été possible de réunir un squelette (composite) représentatif de l’espèce pour l’exposition permanente de la galerie Cuvier du Musée illustrant la paléontologie régionale, ce qui est loin d’être fréquent.
II.3.3.1. Données morphométriquesLa liste d’inventaire du matériel disponible apparaît en annexe à la fin (NR = 258, NMI = 4).Crâne : Tabl. 81, Fig. 25, Fig. 26, Pl. VI, fig. 1-2 (FEL.3)Les trois crânes découverts (FEL.1, FEL.2, FEL.3) appartiennent bien au lion des cavernes Panthera (Leo) spelaea. Ils en possèdent la forme générale globuleuse et la denture caractéristique. Il s’agit d’un vrai lion Panthera, sous-genre Leo. Si le lion des cavernes est bien un lion, il appartient incontestablement à une espèce différente
du lion actuel. L’observation du crâne FEL.3 (Pl. V, fig. 1-2), en particulier de la forme de l’extrémité postérieure des palatins au niveau des choanes (veresCAgin, 1969) confirme la distinction entre les deux espèces déjà faite à propos du crâne du lion des cavernes d’Azé (ArgAnt, 1988). Ce sont également les conclusions de l’étude de l’ADN mitochondrial ancien (burger et al., 2004) qui n’en tire malheureusement pas les conséquences sur le plan taxonomique. La dénomination Panthera (Leo) spelaea, et non pas Panthera leo spelaea faisant du lion des cavernes une simple sous-espèce du lion actuel Panthera leo, me semble la seule utilisable si on veut respecter la réalité paléontologique. C’est l’avis, depuis longtemps d’un certain nombre d’auteurs, auprès desquels je me range sans hésitations (boule, 1906 ; veresCAgin, 1969 ; bonifAy, 1971 ; bAllesio, 1975 ; ArgAnt, 1991 et bien d’autres).- individu 1 : RO.86.Q.19-77.FEL.1, femelle probable. Crâne pratiquement complet à l’exception des deux os
des nasaux, d’une partie des palatins et de l’os incisif droit. La denture est assez complète. Manquent les M1/ g et d, les P2/ g et d (mais les alvéoles restent visibles), ainsi que la I1/ d et les I2/ g et d. L’arrière-crâne apparaît court, abaissé, plutôt fuyant vers l’arrière. Il présente des crêtes nucale et sagittale faibles. La protubérance occipitale n’est que faiblement développée. Le museau paraît court et large. La largeur des arcades zygomatiques plutôt faible donne un aspect moins triangulaire au crâne. Les bulles tympaniques partiellement endommagées présentent malgré cela un net relief traduisant un développement important.
- individu 2 : RO.86.R.20-54.FEL.2, mâle probable Crâne le plus endommagé, en grande partie par
vandalisme sur le terrain (23 octobre 1986) et qui a défrayé la chronique à l’époque. Le maxillaire droit ainsi qu’une grande partie de l’arcade zygomatique
Fig. 25 : Schéma de la forme de l’extrémité postérieure des palatins au niveau des choanes (d’après VeresCAgin, 1969, vol.1, p.464).
ROMAIN-LA-ROCHEindividu n°3RO.88-P.20-8
Forme de steppe Forme de forêt
Pantheraspelaea
Pantheraleo
Pantheratigris
Pantheraonca

562 A. ARgANT
Tableau 81 : Romain-la-Roche, Panthera spelaea, crâne, Pfema 10 (Fig. 26).
Panthera spelaea Crâne – Pfema 10
Individu 1 Individu 2 Individu 3RO.86-Q.19-77 RO.86-R.20-54 RO.88-P.20-8
g d g d g d1 Longueur prosthion - basion 344,5 400,9 364,82 Longueur faciale prosthion - point F 201,8 233,0 221,33 Longueur sup. neurocrâne point F - acrocranion 173,0 207,8 186,54 Longueur viscérocrâne prosthion - cranion / 181,0 168,65 Longueur latérale du museau 140,0 - 124,5* / 151,3 _ 152,06 D.T. maximum ouverture nasale 49,2 / 44,0*7 D.T. maximum au niveau des canines 103,6* / 94,08 D.T. de la face entre foramens infraorbitaires 95,5 / 93,79 D.T. minimum entre les orbites entorbital-entorbital 75,0 89,4 79,410 D.T. frontal ectorbital-ectorbital 108,0* 127,4* 110,011 D.T. minimum de la boîte crânienne 73,3 76,3 71,012 D.T. maxi. neurocrâne à la suture pariéto-temporale 99,0 112,6 92,013 D.T. maxi. de la protubérance occipitale externe 75,0 96,6 77,5*14 D.T. bizygomatique zygion - zygion 223,9 / 225,615 Longueur neurocrâne nasion - nasion / / 237,616 Hauteur maxi. arcade zygomatique 56,4 - 56,0 / 55,6 - /17 Hauteur mini. arcade zygomatique 31,3* - / / 34,8 - /18 Longueur maximum de l’orbite 70,0 - / / 75,6 - 76,219 Hauteur maximum de l’orbite 46,2 - / / 55,0 - 54,020 Hauteur du triangle occipital acrocranion -basion 103,8 121,2 104,221 Longueur des dents jugales P2/ - M1/ ou alvéoles 78,0 -79,0 / 80,7 - 87,322 Longueur des prémolaies P3/ - P4/ ou alvéoles 78,4 - 77,8 / 76,3 - 81,223 Longueur diastème entre alvéoles C - P3/ 7,0 - 6,9 / 8,8 - 10,0*24 Longueur condylo-basale 307,0 / 332,725 Longueur totale prosthion - acrocranion 290,6 / 310,526 Longueur basifaciale prosthion-synsphénion 197,7* / 214,627 Longueur basifaciale synsphénion - basion 106,0* 105,5 198,028 Longueur médiane maxi. du palais prosthion-staphylion 165,6 / 173,229 Longueur médiane mini. du palais prosthion-staphylion / / 161,530 Longueur rangée d’incisives ou alvéoles 58,9 51,6 52,831 D.T. interne entre les racines des canines 63,3 / 58,232 D.T. maximum du palais 143,0 / 125,033 D.T. maximum au niveau des ptérygoïdes 43,7 44,6 39,034 D.T. maximum du trou occipital 32,2 28,7 29,535 Hauteur du trou occipital basion-opisthion 15,0* 13,5 18,436 D.T. maximum aux condyles occipitaux 65,0 64,3 66,837 D.T. otion - otion 135,5 155,0 137,938 Longueur de la bulle tympanique / - 48,5* 46,7 44,5*-39,039 Largeur de la bulle tympanique / - 33,5* 27,4 27,2* - /40 Largeur maxi. cavité glénoïde (serrage externe) 26,3 - 25,4 33,5* 27,6 -25,6

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 563
12 3
4
5
6 7 8 9
10 11 12 13 14
PN
N
FA
A
15
16
18
19
20
2122
23
2425
26 2728
29
303132
33 3435 36 37
3839
40
17
P SPPO
Fig. 26
Fig. 26 : Plan de mesurage du crâne des félidés (Pfema.10)

564 A. ARgANT
du même côté manquent. Les palatins sont absents. Malgré la cassure, on a l’impression d’une plus grande largeur des arcades zygomatiques et d’un aspect nettement triangulaire de ce crâne. La denture apparaît très incomplète, présence seulement des I1/ g et d, I2/ g et d, I3/ g et d et de la C sup. g. Sur le maxillaire gauche (intact à cet endroit), la première racine (sectionnée) de prémolaire se situe à 32 mm de la canine. C’est un diastème bien long entre la canine et la P2/, jamais rencontré ailleurs. Il est plus vraisemblable de supposer l’absence de P2/ et d’attribuer la racine à P3/. L’arrière-crâne apparaît allongé, plus relevé que sur le crâne précédent et porte une crête sagittale et une protubérance fortes. Le museau reste assez allongé. Les bulles tympaniques sont peu renflées, moins saillantes que sur FEL.1.
- individu 3 : RO.88.P.20-8. FEL.3, femelle probable, Planche VI, fig. 1-2.
Crâne recouvert d’un léger encroûtement calcaire, qui présente le meilleur état de conservation des trois, mis à part les nasaux et une partie du maxillaire droit endommagés. La largeur des arcades zygomatiques est plutôt faible et l’aspect du crâne moins triangulaire. La denture apparaît pratiquement complète à l’exception des P2/ g et d dont on voit les alvéoles vides et des M1/ g et d. L’arrière crâne apparaît bas, plongeant, à crête sagittale et protubérance occipitale faibles, comme chez l’individu n°1. Le museau se montre relativement allongé. Les bulles tympaniques sont nettes, en relief.Ces trois crânes de Panthera spelaea de Romain-la-Roche permettent de distinguer par leur morphologie et les données métriques le sexe des animaux :- plus grande largeur des arcades zygomatiques chez
les mâles conférant un aspect plus triangulaire en vue inférieure
- arrière-crâne plus allongé, plus redressé aussi chez les mâles, avec une crête sagittale et une protubérance occipitale plus fortes. Au contraire, aspect plus fuyant vers le bas, plus arrondi de l’arrière crâne des femelles.
- curieusement, les bulles tympaniques, tout en ayant des dimensions comparables, paraissent plus en relief dans le cas des deux crânes de femelles et plus diffuses et plus aplaties sur le crâne du mâle de Romain-la-Roche (comme sur celui d’Azé, ArgAnt, 1988). Faut-il y voir un caractère sexuel stable ou une simple coïncidence ?
- les dimensions des canines (L x l de la couronne), malgré le manque des C sup. sur FEL.2, confirment aussi l’attribution de ces crânes à un mâle et à deux femelles (Fig. 28).
Par contre, d’autres caractères comme la longueur et la largeur du museau, la largeur du renflement pariétal, le développement des apophyses entourant les bulles tympaniques relèvent de variations individuelles et non sexuelles.Mandibule : Tabl. 82, Fig. 27, Pl. VI, fig. 3 (FEL.04)- la mandibule de l’individu 1 n’est pas connue avec
certitude (FEL.47, FEL.49 ?)- individu 2 : RO.87.R.20-15.FEL.4, demi-mandibules
g et d, mâle probable, Pl. VI, fig. 3. Les deux branches de la mandibule sont complètes, en
bon état de conservation, longues et assez grêles, avec une faible hauteur et une faible largeur de la branche horizontale. Présence des deux canines inférieures g et d, de P/3 d, P/4 d et de M/1 d, le côté gauche ayant été endommagé par l’acte de vandalisme. L’extrémité antérieure de la fosse massétérienne (moins profonde
Tableau 82 : Romain-la-Roche, Panthera spelaea, demi-mandibule, Pfema 20 (Fig.27).
Panthera spelaea Mandibule – Pfema 20
Individu 2 Individu 3RO.86-R.20-54 RO.88-P.20-8
gauche droite 1 Longueur infradental - milieu du condyle 277,2 275,0 235,12 Longueur infradental - apophyse angulaire 281,0 283,0 232,73 Longueur infradental - apophyse coronoïde / 275,0 244,14 Hauteur processus angulaire - coronion / 136,3 120,65 Longueur P/3-M/1 ou alvéoles 80,2 81,0 75,46 Longueur diastème C - P/3 (entre alvéoles) 37,6 37,6 30,77 Hauteur branche mandibulaire derrière M/1 56,2 55,6 56,08 Hauteur branche mandibulaire devant P/3 49,3 49,3 46,19 Longueur alvéole P/3 / 19,2 18,810 Longueur alvéole P/2 27,4 27,0 /11 Longueur alvéole M/1 29,0 28,7 27,912 D.T. au niveau de la canine 31,6 31,6 20,013 D.T. avant P/3 23,2 22,4 20,714 D.T. arrière M/1 25,0 25,7 21,015 D.T. condyle d’articulation 69,7 67,3 53,6

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 565
que chez l’exemplaire d’Azé) ne s’engage pas sous la M/1 sur la demi-mandibule gauche, mais le fait nettement sur le côté droit ce qui laisse supposer un développement plus important du muscle masséter droit. Cela doit rendre prudent quant à l’utilisation de ce caractère souvent considéré comme indicateur d’ancienneté. La saillie symphysaire n’apparaît pas particulièrement proéminente. Le profil longitudinal du bord inférieur de la mandibule présente, à gauche comme à droite, une convexité assez nette située à la verticale du contact P4/-M/1.
- individu 3 : RO.88.P.19-16.FEL.207, demi-mandibule g.
Seule la demi-mandibule gauche est connue, concré-tionnée elle aussi comme le crâne. Elle est complète même si son état de conservation apparaît moins bon que dans le cas précédent. Elle reste assez grêle, mais ce caractère est atténué par une moins grande longueur. Elle porte la canine inférieure gauche ainsi que la M/1 g. La P/3 et la P/4 manquent depuis longtemps et les alvéoles ont été concrétionnées. L’extrémité antérieure de la fosse massétérienne ne s’engage pas sous la M/1. La saillie symphysaire apparaît nettement développée comme à Jaurens (bAllesio, 1980). Le profil inférieur plus rectiligne présente quand même une légère conve-xité dont le maximum se situe sous le milieu de M/1.
Ce matériel présente l’intérêt de permettre la comparaison des côtés gauche et droit de la mandibule d’un même individu (individu 2, FEL.2), ce qui doit nous rendre prudent avant de tirer des conclusions chronologiques de la position de l’extrémité antérieure de la fosse massétérienne par rapport à la M/1. La relative gracilité de ces mandibules d’individus mâle et femelles de Romain-la-Roche permet d’éliminer déjà les formes les plus anciennes de lion des cavernes, particulièrement massives, ce que les crânes suggèrent également. Le Panthera spelaea de Romain-la-Roche est forcément plus récent que l’exemplaire d’Azé attribuable au Pléistocène moyen récent.
Denture : Tabl. 83- Incisives :Incisives supérieures : elles sont toutes représentées, pratiquement sur chacun des crânes. Ces dents à racine unique se détachent pourtant facilement. C’est une preuve de l’absence de transport et d’enfouissement rapide des vestiges. Les faibles dimensions de ces dents, au rôle réduit chez les félidés, se retrouvent évidemment sur le matériel de Romain-la-Roche. Leur usure reste faible et maximum pour la I3/ plus caniniforme (U = 1 pour FEL.1 et FEL.3, 1,5 pour FEL.2).Incisives inférieures : aucune n’a été trouvée en place et seules deux incisives isolées (FEL.42, FEL.214) pourraient correspondre, mais leur état d’usure ne permet pas une détermination précise.- Canines :Canines supérieures : FEL.1 : canines assez grêles, avec couronne à arête postérieure partiellement effacée et à émail fendillé dans la partie visible restante, double sillon caractéristique sur les faces internes et externes de cette couronne, usure U = 0 à g et U = 0,6 à droite.FEL.2 : canine gauche seule, à la couronne endommagée non mesurable, plus massive que la précédente, sillons non visibles effacés par le frottement, usure U = 0,6.FEL.3 : canines assez grêles, traces d’usure sur la face antéro-interne des couronnes par frottement contre la canine inférieure, avec recoupement de l’usure de pointe. L’arête postérieure de la couronne présente un émail fendillé, non affectée par l’usure. Le double sillon se rencontre sur les faces externe et interne de la couronne. L’usure reste faible U = 0,3.Canines inférieures :FEL.2 : présence des deux canines inférieures, traces nettes d’usure de la couronne par frottement contre les canines supérieures face postéro-interne recoupant l’usure de pointe. L’arête postérieure de la couronne possède un émail fendillé. Le double sillon n’existe que sur la face externe de la couronne, usure U = 0,3.FEL.3 : canine gauche seule connue, à aspect grêle, faible usure postéro-interne de la couronne frottant contre la canine supérieure avec un léger recoupement de l’usure de pointe, arête postérieure non usée, à émail fendillé. Double sillon sur la face externe seulement, usure U = 0,3.- Prémolaires :Prémolaires supérieures :P2/ : ces dents à racine simple se détachent facilement. Elles existaient bien sur FEL.1 et FEL.3 car on retrouve les alvéoles vides. Par contre elles manquaient probablement chez FEL.2, du moins celle du maxillaire supérieur gauche seul connu.P3/ : la cuspide antérieure se trouve classiquement déjetée du côté interne. Côté lingual, l’arrière de la couronne descend largement sur la racine postérieure particulièrement élargie à cet endroit. Aucune différence ne se constate entre FEL.1 et FEL.3.
Fig. 27 : Plan de mesurage de la mandibule des félidés (Pfema.20)
Fig. 27
1
2
3
4
56
78
9 10 11

566 A. ARgANT
P4/ : pas de différences notables entre les P4/ de FEL.1 et FEL.2 seules disponibles. Elles n’ont ni la largeur, ni la massivité des dents équivalentes du Panthera spelaea d’Azé. Le talon antéro-interne apparaît bien développé. Le bord lingual de la couronne est rectiligne. Le rebord vestibulaire présente une légère inflexion au niveau du paracône, mais n’a pas l’aspect en accolade que l’on rencontre à Azé (ArgAnt, 1988). L’usure de ces dents, encore faible n’affecte de façon sensible ni le paracône, ni le métacône.Prémolaires inférieures :P/3 : seule la P/3 droite de FEL.2 est conservée. Le tubercule principal bien net, aux arêtes antérieure et postérieure peu marquées, est encadré par deux tubercules secondaires d’égale importance. La cuspide postérieure s’incurve classiquement du côté lingual.P/4 : seul FEL.2 livre les P/4. Un protoconide bien affirmé, au sommet légèrement usé (U = 0,2), est entouré de deux tubercules d’égale importance. On peut noter un élargissement sensible de la moitié postérieure de la dent.
- Molaires :Molaire supérieure : seules les alvéoles de la M1/ sont présentes, en position transversale classique à l’arrière de la P4/ sur les deux crânes FEL.1 et FEL.3. Observation impossible sur FEL.2. Aucune M1 / isolée n’a été trouvée dans le matériel, mais c’est une dent toute petite et qui relève presque de la recherche de microfaune. Molaire inférieure : la morphologie de cette dent est importante pour l’étude de Panthera spelaea. Aucune différence morphologique ne peut-être notée entre les différentes M/1 dont je dispose à Romain-la-Roche. Elles paraissent relativement allongées du fait d’une largeur assez faible n’atteignant pas, et de loin, le développement des formes anciennes du Pléistocène moyen (Château, Azé, ArgAnt, 1991). Le talon postérieur net mais peu développé, le rebord lingual rectiligne de la couronne, sans le renflement caractéristique des formes anciennes, indiquent plutôt les formes récentes.Toutes les dents de Panthera spelaea de Romain-la-Roche présentent une bonne stabilité morphologique et
Tableau 83 : Romain-la-Roche, Panthera spelaea, denture supérieure et inférieure.
Panthera spelaea – Denture
DENTURESUPERIEURE
Individu 1 Individu 2 Individu 3DENTURE
INFERIEURE
Individu 1 Individu 2 Individu 3RO.86-Q.19-77 RO.86-R.20-54 RO.88-P.20-8 RO.86-Q.19-77 RO.86-R.20-54 RO.88-P.20-8
g d g d g d g d g d g d
I1/M.D. 8,5 / 8,0 8,0 8,1 8,2
C. inf.
M.D. / / 24,9 24,1 21,7 /V.L. 5,7 / / 5,8 5,7 5,3 V.L. / / 18,6 17,7 15,4 /
I2/M.D. / / 8,7 9,2 9,6 10,0 H.C. / / / / 37 /V.L. / / 6,8 6,8 6,2 6,6 P.C. / / 70,0 68,6 64 /
I3/M.D. 12,9 13,0 11,6 11,6 12,8 13,0 H.T. / / / / / /V.L. 10,9 11,0 13,7 13,3 11,0 10,8
P/3
M.D. / / / 19 / /
C.sup
M.D. 24,4 24,8 / / 22,7 22,0 V.L.post. / / / 9,6 / /V.L. 17,9 18,2 / / 16,8 16,8 V.L. ant. / / / 7 / /H.C. 47,8 / / / / / H.C. / / / 10,6 / /P.C. 72,0 72,0 / / 66,6 64,5 P/4 M.D. / / 27,4 27,7 / /H.T. / / / / 104,7 98,9 H.C. / / / 16,9 / /
P2/M.D. 9,0 (A) 9,2 (A) / / 8,8 11,5*
M.D. tuber. princ.
/ / / 12,4 / /
V.L. 6,8 (A) 6,4 (A) / / 8,8 8,7* V.L. post. / / 14,1 13,8 / /
P3/M.D. 27,3 27,2 / / 26,3 26,3 V.L. ant. / / 10,8 11,3 / /
V.L.maxi. 14,0 13,7 / / 13,5 13,6 V.L. mini. / / 10,4* 11,3 / /V.L. ant. 10,8 10,8 / / 11,0 /
M/1
MD / / 28,4 29,0 27,9 /
P4/
MD ext. 38,2 39,6 / / 38,1 38,7 MD pad / / / 15,0 16,3 /MD méd. 36,4 36,8 / / 35,6 36,3 MD prd / / / 16,2 16,8 /MD int. 38,6 39,0 / / 37,3 37,7 VL / / 14,6 14,2 14,4 /MD pa 15,2 15,3 / / 13,3 13,9 HC prd / / / 17,0 15,8 /MD me 14,8 14,9 / / 14,3 14,4 HC sous
sillon/ / / / 7,3 /
VL ant. 20,0 20,4 / / 20,6 20,7VL post. 14,4 14,2 / / 13,7 13,4
A= mesure de l’alvéole, pa = paracône, me = métacône, tal. = talon,pad = paraconide, prd= protoconideDT min.
tal. 13,2 12,0 / / 13,6 13,3

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 567
dimensionnelle. L’âge que l’on peut approcher par l’usure dentaire, traduit des animaux à denture définitive depuis un certain temps mais avec une usure encore faible. Ce sont tous les trois de jeunes adultes. Parmi eux, le mâle FEL.2 semble légèrement plus âgé que les deux femelles FEL.1 et FEL.3.Squelette post-crânien :Colonne vertébrale :L’atlas, l’axis et le sacrum sont souvent les seuls éléments mentionnés. Cela s’explique par la rareté des vertèbres bien conservées avec leurs apophyses, permettant seules une détermination précise de leur rang. Les corps vertébraux, de loin les plus fréquents, n’apportent que peu d’informations, si ce n’est une vague indication de taille. Je n’ai retenu à Romain-la-Roche que les restes permettant d’assurer la détermination de Panthera spelaea. L’étude morphométrique est réservée aux seules vertèbres suffisamment bien conservées et bien localisées dans la colonne vertébrale. Le groupement des vestiges de l’individu 3 (RO. 88-P. 20-8) appartenant à un même secteur (carrés : P. 18, P. 19, Q. 19), permet de prendre en compte avec certitude des vertèbres d’un même individu (femelle adulte) :vertèbres cervicales : Tabl. 84-86.1. Atlas : FEL. 145 – RO. 89 – P. 19 – 202. Axis : FEL. 169 – RO. 89 – P. 19 – 343. V. c. 4 : FEL. 205 - RO. 89 – P. 19 – 254. V. c. 5 : FEL. 143 – RO. 89 – P. 19 – 175. V. c. 6 : FEL. 206 – RO. 89 – P. 19 – 316. V. c. 7 : FEL. 148 – RO. 89 – P. 19 – 31vertèbres dorsales : Tabl. 871. V. d. 1 : FEL. 156 – RO. 89 – Q. 19 – 172. V. d. 2 : FEL. 155 – RO. 89 – Q. 19 – 173. V. d. 4 : FEL. 146 – RO. 89 – P. 19 – 244. V. d. 9 : FEL. 204 – RO. 89 – P. 19 – 255. V. d. 10 : FEL. 203 – RO. 89 – P. 19 – 256. V. d. 12 : FEL. 150 – RO. 89 – P. 19 – 327. V. d. 13 : FEL. 202 – RO. 89 – P. 19 – 15vertèbres lombaires : Tabl. 881. V. l. 2 : FEL. 220 – RO. 89 – P. 18 – 402. V. l. 3 : FEL. 219 – RO. 89 – P. 18 – 403. V. l. 5 : FEL. 218 – RO. 89 – P. 18 – 404. V. l. 6 : FEL. 221 – RO. 89 – P. 18 – 40
sacrum : Tabl. 89Les nombreuses vertèbres caudales de la longue queue des félidés et donc de Panthera spelaea se repèrent facilement par leur longueur et la morphologie (extrémités articulaires bien particulières) : FEL. 35 : longueur 59,7 mm 8e ou 9e vertèbre caudale. Ceintures scapulaire et pelvienne :- omoplate : NR = 1 d, Tabl. 90, Pl. VII, fig. 1 (FEL.200)L’omoplate d FEL.200 bien conservée, apporte le maximum de données métriques possibles, ce qui est rare.- coxal : NR = 2Parmi de nombreux fragments pour lesquels la détermination entre Ursus et Panthera reste aléatoire, deux fragments FEL.147 et FEL.200 appartiennent à Panthera spelaea, mais ne fournissent pas de données métriques.Membre antérieur :- Humérus : NR = 1 g et 1 d, Tabl. 91.La mauvaise conservation des humérus est à souligner. Seule une diaphyse d FEL.142 fournit de rares données.- Radius : NR = 4 g et 1 d, Tabl. 92.Deux radius complets g FEL.116 et FEL.211 livrent des informations métriques.- Cubitus : NR = 2 g et 1 d, Tabl. 93.Trois cubitus sont représentés dont deux complets FEL.247 et FEL.111+109. Ce dernier (FEL.111+109) provient du recollage de deux parties trouvées à la fouille, la même année, dans le même carré P.18, dans la même couche et à 20 cm de distance environ (P.18-22 et P.19-23).- Carpe :Les os du carpe, comme du tarse, restent en général bien représentés dans les gisements. Ce sont des os courts, robustes, sans moelle, qui n’attirent pas les canidés. A Romain-la-Roche, ils sont tous représentés et fournissent un maximum de données. Deux individus distincts peuvent être suivis dans la série grâce à leur aspect et à la concordance des tailles. Un carré noir et un cercle noir les indiqueront dans les tableaux respectifs. Il est par contre plus difficile, en l’absence d’étude taphonomique poussée (répartition spatiale globale des vestiges de
1
2
3
4
5
6
7
8
vue dorsale
vue crâniale
ATLAS - PFEMA 30
1 2 3 4 5 6 7 8
FEL.45 51,3 - - 69,0 76,7 81,4 26,2 37,4
FEL.145 50,0 - 80,5 72,0 75,6 65,6 24,1 31,8
Atlas (vert. cerv. 1) – PF. 301- DD P ma x.2- DT max.3- DA P ma x.4- DT articulaire crânial5- DT articulaire caudal6- DA P ma x. interarticulaire7- DA P interarticulaire min. ventral8- DA P interarticulaire min. dorsal
AtlasPfema 30
FEL. même individu n°3
Tableau 84 : Romain-la-Roche, Panthera spelaea, atlas.

568 A. ARgANT
1 2 3 4 5
FEL.46 - - 100,7 68,5 44,0
FEL.169 74,4 - 93,0 71,4 43,0
Axis (vert. cerv. 2) – PF. 311-DDP m ax.2- DT max. avec apophyses3- DA P ma x. 4- DT articulaire crânial5- DT articulaire caudal
1
2
3
4
AXIS - PFEMA 31
vue crâniale
vue latérale
AxisPfema 31
FEL. même individu n°3
Tableau 85 : Romain-la-Roche, Panthera spelaea, axis.
Tableau 86 : Romain-la-Roche, Panthera spelaea, autres vertèbres cervicales.
vue crâniale
vue dorsale
2
3
4
5
7
Autres VERT. CERVICALESPFEMA 32
Autres v ert. cerv. PF. 321- Rang de la vertèbre si connu2- DD P ma x. avec apophyses3- DT max. avec apophyses.4- DA P ma x. avec apophyses5- DT articulaire crânial6- DT articulaire caudal7- DA P ma x. interarticulaire du corps
vertébral
1 2 3 4 5 6 7
FEL.205 4e - - - 43,0 - 37,7
FEL.143 5e 75,3 - 40,0 38,0 36,5
FEL.206 6e - - 57,2 - 36,6 34,5
FEL.148 7e 98,0 101,6 55,0 37,0 38,5 35,0
V. cerv.Pfema 32
FEL. même individu n°3
Tableau 87 : Romain-la-Roche, Panthera spelaea, vertèbres dorsales.
FEL. même individu n°3 (RO.88-P.20-8)
1 2 3 4 5 6 7
FEL.156 1e 133,3 98,0 40,5 36,5 33,4 36,4
FEL.155 2e 143,0 82,0 39,1 33,5 33,7 38,5
FEL.121 3e - - - 40,2 44,0 50,0
FEL.146 4e - 77,0 15,1 30,2 33,0 37,0
FEL.262 6e 125,0* 75,2 105,4 37,8 58,0* 40,0
FEL.204 9e - - - 34,0 45,0 41,9
FEL.203 10e 81,5 64,0 46,4 35,4 45,0 43,2
FEL.150 12e - 52,7 68,2 46,0 49,4 47,0
FEL.246 12e 86,3 - - 47,4 48,6 52,2
FEL.263 12e 86,3 51,5* 80,0 48,0 48,5 52,3
FEL.202 13e 83,3 54,6 78,0 47,7 49,0 50,2
,
,
Vertèbres dorsales – PF. 331- Rang de la vertèbre si connu2- DD P ma x. avec apophyses3- DT max. avec apophyses.4- DA P ma x. avec apophyses
(en projection)5- DT articulaire crânial fovea
costale com prise6- DT articulaire caudal fovea
costale com prise7- DA P ma x. interarticulaire (CV )
vue latérale vue crâniale
53
4
2
57
6
VERT. DORSALE - PFEMA 33
V. dors.Pfema 33

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 569
Panthera spelaea) de faire le lien avec les individus 1, 2 ou 3 des crânes.scapholunaire : NR = 3 g et 1 d, Tabl. 94.pisiforme : NR = 3 g et 1 d, Tabl. 95.trapèze : NR = 2 g, Tabl. 96.trapèzoïde : NR = 2 g et 1 d, Tabl. 97.magnum : NR = 1 g, Tabl. 98.unciforme : NR = 1 g et 1 d, Tabl. 99.pyramidal : NR = 2 g et 1 d, Tabl. 100.Métacarpiens
Si on se base sur trois individus adultes tombés dans l’aven, comme il y a 5 métacarpiens par main et 2 mains par individu, on devrait trouver 30 métacarpiens confondus à Romain-la-Roche. Seuls vingt métacarpiens, souvent bien conservés, peuvent être dénombrés et 18 fournissent des données dans le tableau (Tabl. 101). Ce n’est déjà pas si mal.Un bel ensemble de quatre métacarpiens g complets appartient à un même individu : Pl. VII, fig. 2- le Mtc.2 : FEL. 100 - RO.89-P.18-19
11
2
4
7
89
10 12
SACRUM - PFEMA 35
1- L totale max .2- L entre le p rocessus articulaire crânial et le processus épineux de la
dernière vertèbre sacrale (Barone, 1976, p.214)3- DCC a rticulaire max.4- DT max. ailes5- DT articulaire crânial6- DA P articulaire crânial7- L. max. semi-articulation/coxal (g. ou d.) sur l’aile du sacrum8- DT max. semi-articulation/coxal (g. ou d.) sur l’aile du sacrum, x39- Distance entre les 2 p remiers forame ns sacraux pelviens (entre axes)
10- DT a rticulaire distal11- DDP m ax. avec la 1 ère épine dorsale et le bord ventral du corps du
sacrum12- DDP entre le processus épineux sacral de la d ernière vertèbre
dorsalem ent e t le bord inférieur de l’apex
Sacrum - PF.35
1 3
1 2 3 4 5 6 7 8 9 10 11 12
FEL.261 - 115,6 103,4 91,6 58,8 29,6 56,2 28,6 39,4 31,2* - 39,0*
Pantheraleo actuel(Feme lle)Afrique
105,5 95,6 92,4 83,6 45,3 24,6 24,2 (G)26,2 (D) 32,6 23,7 59,2 31,9
6 5
SacrumPfema 35
Vue dorsale
Vue latérale G
2
7
6
4
53
7
4
VERT. LOMBAIRE PFEMA 34
FEL. même individu n°3 (RO.88-P.20-8)
1 2 3 4 5 6 7
FEL.166 1e 89,7 80,0 52,2 46,9 48,9 56,5
FEL.201 2e 100,4 - - - 48,0* 56,0
FEL.220 2e - - - 61,0* 55,0 66,4
FEL.219 3e - - - 54,3 55,4 68,3
FEL.222 4e 103,4* - - 48,7 53,4 70,6
FEL.218 5e 104,3 - - 53,2 52,6 60,4
FEL.221 6e - - - - - 63,3
V. lomb.Pfema 34
Vertèbres lombaires – PF. 341- Rang de la vertèbre si connu2- DD P ma x. avec apophyses3- DT max. avec apophyses.4- DA P ma x. avec apophyses
(en projection)5- DT articulaire crânial6- DT articulaire caudal7- DA P ma x. interarticulaire (CV )
Tableau 88 : Romain-la-Roche, Panthera spelaea, vertèbres lombaires.
Tableau 89 : Romain-la-Roche, Panthera spelaea, sacrum.

570 A. ARgANT
Tableau 90 : Romain-la-Roche, Panthera spelaea, omoplate.
Tableau 91 : Romain-la-Roche, Panthera spelaea, humérus.
Panthera spelaeaHumérusPfema. 50
1 2 3 4 5 6 7 8 9 10 11 12
FEL. 142 - - - - - 36,5* 41,8* 93,7* 70,3* - - D
1- Longueur max. (en projection)2- DT proximal max.3- DAP proximal max.4- DT articulaire prox.5- DAP articulaire prox.6- DT milieu diaphyse (le plus mince)
7- DAP milieu diaphyse (le plus fort)8- DT distal9- DAP distal10 - DT trochlée articulaire distale11- DAP trochlée articulaire distale12- Latéralisation
Tableau 92 : Romain-la-Roche, Panthera spelaea, radius.
Panthera spelaeaRadius
Pfema. 511 2 3 4 5 6 7 8
FEL. 116 330,8 46,1 32,6* 33,6 22,4* 67,7 39,2 g
FEL.211 349,4 42,7 31,2 37,0 21,3 69,3 40,8 g
1- Longueur 2- DT proximal 3- DAP proximal 4- DT médian
5- DAP médian6- DT distal7- DAP distal8- Latéralisation
Omoplate – PF. 401- Longueur max. (en projection)2- DAP min. col3- DAP max. en projection4- DT max avec acromion5 - DAP max. proximal6- DAP articulaire7- DT articulaire8- Latéralisation
1
2
3
4
OMOPLATE - PFEMA 40
67
5
1 2 3 4 5 6 7 8
FEL.200 273,5* 63,3 218,6* - 70,4 61,3 46,5 D
OmoplatePfema 40

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 571
Tableau 93 : Romain-la-Roche, Panthera spelaea, cubitus.
Panthera spelaeaCubitus
Pfema 521 2 3 4 5 6 7 8
FEL. 210 416,1 49,6 70,8 22,8 42,4* 23,3 37,7 g
FEL.111 + 109 408,0* 47,3 71,3 22,2 33,0 21,0 35,2 g
FEL. 210 416,1 49,6 70,8 22,8 42,4* 23,3 37,7 g
FEL. 247 - - - - - 30,29 46,97 D
1- Longueur 2- DT proximal 3- DAP proximal 4- DT médian
5- DAP médian6- DT distal7- DAP distal8- Latéralisation
Tableau 94 : Romain-la-Roche, Panthera spelaea, scapholunaire.
Panthera spelaeaScapholunaire
Pfema. 5311 2 3 4 5 6 7 8
FEL. 37<
30,0 52,8 39,7 47,6 31,6 45,6 28,6 g
FEL.211 33,4 56,0 44,6 51,3 34,0 51,2 30,5 g
FEL. 104n
32,0 54,9 39,4 46,0 29,8 46,0 33,3 g
FEL. 114n
- 54,9 39,0* 46,4* - - - D
Dans tableaux 94 à 100 : < et n individus distincts ; FEL. = même individu
1- DDP max 2- DT max. 3- DAP max. 4- DT articulaire prox.
5- DAP articulaire prox.6- DT max. articulaire distal7- DAP max. articulaire distal8- Latéralisation
Tableau 95 : Romain-la-Roche, Panthera spelaea, pisiforme.
Panthera spelaea PisiformePfema 533
1 2 3 4 5 6 7 8
FEL. 16<
46,1 25,7 14,6 13,4 - - 23,7 g
FEL.93 45,8 24,3 14,8 13,7 - - 21,4 g
FEL.106 47,5 25,0 15,0 - - 22,3 D
1- DDP (hauteur) max 2- DT prox. max. 3- DAP prox. max. 4- DT milieu col
5- DAP milieu col6- DT ext. distale (tête)7- DAP ext. distale (tête)8- Latéralisation

572 A. ARgANT
- le Mtc.3 : FEL. 99 - RO.89-P.18-19- le Mtc.4 : FEL. 101 - RO.89-P.18-19- le Mtc.5 : FEL. 101 - RO.89-P.18-19Ces deux derniers sont soudés par la brèche. Ils proviennent tous du même carré de fouille P.18-19. Le Tableau 101 permet d’en connaître les dimensions respectives.Membre postérieur - Fémur : NR = 3 g et 1 d, Tabl. 102.Un seul fémur d FEL.147 est complet et fournit le maximum de données.- Rotule : NR = 1 g et 2 d, Tabl. 103, Pl. V, fig. 3 (FEL.215)- Tibia : NR = 2 g et 1 d, Tabl. 104.- Péroné : NR = 2 g et 2 d, Tabl. 105.- Tarse :Tous les os du tarse, sauf le premier cunéiforme, sont également bien représentés :calcanéum : NR = 2 g et 1 d, Tabl. 106.astragale : NR = 2 g et 1 d, Tabl. 107, Pl. V, fig. 6 (FEL.223).naviculaire : NR. 1 g et 1 d, Tabl. 108.cuboïde : NR = 1 g et 1 d, Tabl. 109.cunéiforme 2 : NR = 2 d, Tabl. 110.cunéiforme 3 : NR = 3 g et 1 d, Tabl. 111.- Métatarsiens : Douze métatarsiens confondus, sur 24 possibles, ont été déterminés et mesurés (Tabl. 112). Ils sont plus longs que les métacarpiens et donc plus fragiles, d’où une moins bonne représentation.Phalanges et sésamoïdes : Tabl. 113-116Ces petits éléments du squelette apparaissent bien représentés à Romain-la-Roche, mêlés directement aux autres éléments de faune. Ce sont les premiers éléments mobilisés en cas de transport par l’eau. Leur abondance indique ici l’absence de transport particulier du matériel dans l’intervalle de temps entre la mise en place et le dégagement par la fouille. (ArgAnt, 2004 b, p. 815).La détermination précise des phalanges reste possible à condition de disposer d’un matériel de comparaison sûr et complet, cela reste très long et parfois hasardeux. Sans
Tableau 96 : Romain-la-Roche, Panthera spelaea, tra-pèze.
Panthera spelaeaTrapéze
Pfema 5371 2 3 4
FEL. 38<
19,7 16,0 31,0 g
FEL.88 17,1 20,0 31,3 g
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.
Tableau 97 : Romain-la-Roche, Panthera spelaea, tra-pézoïde.
Panthera spelaeaTrapézoïdePfema. 536
1 2 3 4
FEL. 39<
15,1 27,0 23,7 g
FEL.87 16,8 27,5* 25,1 g
FEL. 103 14,4 24,9 25,3 D
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.
Tableau 98 : Romain-la-Roche, Panthera spelaea, mag-num.
Panthera spelaeaMagnum
Pfema 5351 2 3 4
FEL. 23<
26,2 25,4 36,7 g
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.
Tableau 99 : Romain-la-Roche, Panthera spelaea, unci-forme.
Panthera spelaeaUnciforme Pfema 534
1 2 3 4
FEL. 24<
30,0 23,9 33,3 g
FEL.199 30,0* 24,0* 31,5* D
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.
Tableau 100 : Romain-la-Roche, Panthera spelaea, pyramidal.
Panthera spelaeaPyramidal Pfema 532
1 2 3 4
FEL. 22<
16,2 21,4 27,1 g
FEL.110 14,2 21,0 28,0 g
FEL. 86 - 20,8 28,0 D
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 573
Tableau 101 : Romain-la-Roche, Panthera spelaea, métacarpiens.
Panthera spelaea –Métacarpiens – Pfema 54Mtc. 1 1 2 3 4 5 6 7 8FEL. 10 41,2 24,3 18,7 19,0 15,0 14,8 18,3 gFEL. 108 44,0 22,3 17,3 17,5 - 18,6 17,0 gFEL. 174 39,1 24,0 16,3 17,5* 14,0* 21,3 18,5 DMtc. 2FEL. 33 126,2 27,8 35,9 19,1 19,2 27,8 - gFEL. 31 126,4 26,2 35,9 19,2 19,4 27,4 25,0 DFEL.11 111,4 23,2 32,4 15,6 18,9 25,0 22,5 gFEL. 115 117,2 24,4 34,0 - - 24,6 22,6 DFEL. 100 n 119,0 24,5 33,2 16,8 17,5 26,0 22,2 gMtc. 3FEL. 12 118,0 22,4 - 16,3 16,0 24,4 23,4 DFEL. 20 - 23,7 29,4 18,6 17,0 - - gFEL. 99 n 130,2 30,4 28,8 17,0 15,0 27,0 24,6 gFEL. 157 124,7* 29,2 - 17,3 15,6* 26,3 - gMtc. 4FEL. 6 123,2 23,5 27,2 17,5* 17,0* 24,3* 23,0* DFEL. 101 n (1) 128,3 22,8* 28,6 16,0 15,2* 24,7 23,3 gFEL. 96+98 135,5 23,7 22,5 18,0 14,5 - - DMtc. 5FEL. 13 103,7 27,2 29,0 15,0* 15,5* 23,7 23,2 gFEL. 101 n (1) 109,0 - 23,4 16,5* 15,5 22,5 22,1 gFEL. 224 - 27,98 - 16,21 17,27 26,0 - D
1- Longueur max.2- DT proximal3- DAP proximal4- DT milieu diaphyse5- DAP milieu diaphyse
6- DT distal7- DAP distal8- Latéralisation
(1) soudés par de la brèche (RO.89-P.18-19)n même individu
Tableau 102 : Romain-la-Roche, Panthera spelaea, fémur.
1 2 3 4 5 6 7 8 9 10 11
FEL. 147 423,0 100,4 50,0 - - 34,0* 34,7 81,4 87,9 - D
FEL.225 - - - - - - - 84,4 84,1 - G
FEL. 230 - 95,00* 78,23 - - - - - - G
FEL.231 - 97,4* - 83,3 46,56 - - - - - G
Fémur PF. 601- DA P (longueur) m ax.2- DT (largeur) ma x.3- DA P prox.4- Longueur grand trochanter-petit trochanter5- DT tête
6- DT médian7- DA P mé dian8- DT distal9- DA P distal
10- DT a rticulaire distal11- Latéralisation
5
4
grand trochanter
petit trochanter
FémurPfema 60
-
P. spelaea

574 A. ARgANT
Tableau 103 : Romain-la-Roche, Panthera spelaea, rotule.
Panthera spelaeaRotule
Pfema. 611 2 3 4 5 6
FEL. 212 67,6 46,9 29,1 46,4 46,7 g
FEL.135 64,6 45,0 28,5 42,0 43,1 D
FEL. 215 62,03 45,40 24,86 44,96 40,19 D
1- DDP (longueur) max.2- DT (largeur) max. 3- DAP médian (épaisseur) max..
4- DDP articulaire max.5- DT articulaire max.6- Latéralisation
Tableau 104 : Romain-la-Roche, Panthera spelaea, tibia.
Panthera spelaeaTibia
Pfema 621 2 3 4 5 6 7 8
FEL. 113 375,0* 90,0* 97,8 32,5 - 65,2 45,9 D
FEL.165 376,0* 88,3 99,4 - - 69,0* 46,2* g
1- DAP (longueur) max.2- DT prox.. 3- DAP prox.4- DT médian
5- DAP médian6- DT distal7- DAP distal8- Latéralisation
Tableau 105 : Romain-la-Roche, Panthera spelaea, péroné.
Panthera spelaeaPéroné
Pfema 631 2 3 4 5 6 7 8
FEL. 139 13,0 15,8 22,0 34,3 g
FEL.191 17,2 26,0 D
FEL. 189 18,7 32,2 D
FEL. 245 9,50 10,00 16,64 31,19 g
1- DAP (longueur) max.2- DT prox.. 3- DAP prox.4- DT médian
5- DAP médian6- DT distal7- DAP distal8- Latéralisation

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 575
détermination précise, phalanges et sésamoïdes perdent alors beaucoup de leur pouvoir de comparaison.D’une manière générale les sésamoïdes de Panthera spelaea se distinguent de ceux d’Ursus spelaeus par une plus grande longueur, une forme plus arquée en croissant et par une plus grande étroitesse. La pièce FEL. 123 montre une phalange 1 de Panthera spelaea soudée par de la brèche à un grand sésamoïde collé à la partie proximale et assure la détermination.Divers :- côtes : Parmi les centaines de fragments de côtes du matériel de Romain-la-Roche, il en existe très certainement appartenant à Panthera spelaea. Mais l’état fragmentaire rend pratiquement impossible leur détermination. La taille constitue souvent un élément déterminant, mais les dimensions comparables pour Panthera spelaea et Ursus
spelaeus ne permettent pas d’être catégorique. Les cas de côtes bien conservées, suffisamment longues, portant la tête et le tubercule costal articulaires, sont tellement rares que seule la pièce FEL. 199 peut être attribuée à Panthera spelaea : section partie haute : 30 mm x 20 mm.- sternèbres : Les sternèbres de Panthera spelaea ont un aspect prismatique caractéristique :FEL. 138 : L = 61,0 ; DT max. = 23,9 ; DDP max. = 28,4 Il s’agit probablement d’une sternèbre en position centrale, mais je ne peux déterminer son rang plus précisément.
II.3.3.2. BiochronologieLa lignée évolutive Panthera spelaea fossilis - Panthera spelaea spelaea est maintenant bien connue (bAllesio, 1975, 1980 ; sChütt & hemmer, 1978 ; ArgAnt 1991,
Tableau 106 : Romain-la-Roche, Panthera spelaea, calcanéum.
Panthera spelaeaCalcanéumPfema 641
1 2 3 4 5 6 7 8
FEL. 43 126,7 55,0 54,7 34,8 43,3 65,0* - g
FEL.50<
128,5 47,7 54,7 32,6 39,7 - - g
FEL. 51<
126,0* - 52,0 34,3 39,0 70,2 - D
1- Longueur max.2- DT prox.. 3- DAP max.4- DT tuber calcanei max.
5- DAP tuber calcanei max.6- L. manubrium7- L. sustentaculum tali8- Latéralisation
Tableaux 106 à 111 : FEL< = même individu
Tableau 107 : Romain-la-Roche, Panthera spelaea, astragale.
Panthera spelaeaAstragalePfema 642
1 2 3 4 5 6 7 8 9
FEL. 51<
64,8 52,3 36,0* 41,2* D
FEL.119 63,6 52,0 41,2* g
FEL. 223 65,9 56,3 38,5 34,34 39,31 31,25 38,66 76,38 g
1- Longueur max.2- DT (largeur) max. 3- DAP (épaisseur) max.4- DAP min.5- DT max. trochlée
6- DT col7- DT tête8- Longueur diagonale max.9- Latéralisation

576 A. ARgANT
p. 246, 2007, p. 127), même s’il existe encore des zones d’ombre au niveau des formes intermédiaires (Panthera spelaea intermedia ?). La forme ancienne Panthera spelaea fossilis arrive en Europe au Pléistocène moyen médian (Isernia, Forest-Bed, Mosbach, Vértesszölös, Château, Sprimont) vers 0,7 ou 0,6 Ma. Elle correspond à des animaux de très grande taille, à la musculature puissante. Les individus de Panthera spelaea fossilis de La Sima de los Huesos (500 Ka), de Trinchera galeria et Trinchera Dolina (TD.10 = 400 Ka.) en Espagne possèdent toujours une grande taille (gArCiA, 2003). La taille décroît par la suite assez régulièrement, tout en restant encore importante lors de l’interglaciaire rapporté à l’Holsteinien (Lunel-Viel, Aldène). Quelques exceptions cependant dans cette décroissance, des formes plus petites sont connues à la fin du Pléistocène moyen (La Fage, bAllesio, 1975) et plus tard des formes de grande taille à l’Eémien, au début du Pléistocène supérieur. Au Pléistocène supérieur moyen et récent (OIS.4, 3, 2) la taille du lion des cavernes décroît pour atteindre finalement celle d’un lion actuel de moyenne stature. En France, les derniers spécimens sont datés de 12 248 ± 66 BP dans le Bassin Parisien et de 11 150 ± 220 BP dans le Sud-Ouest de la France. Ce schéma théorique doit évidemment être repris avec prudence. La taille est à manier avec précaution en paléontologie et les Félidés ne livrent jamais un matériel abondant. La sécurité statistique de 30 individus reste impensable. Les variations morphologiques n’apportent guère d’éléments non plus. Outre la grande stabilité morphologique des félidés, elles sont pratiquement impossibles à vérifier sur un nombre suffisant d’individus. Ce sont les critères dentaires, mieux conservés et mieux étudiés, qui constituent finalement le meilleur outil de comparaison et malgré ses imperfections, la taille reste encore l’élément fondamental de comparaison. Un complément de comparaison sur le squelette post-crânien sera tenté.Romain-la Roche livre les restes de plusieurs individus (NMI = 4), bien conservés pour trois d’entre eux et relativement groupés dans l’espace. Ce n’est pas fréquent dans les gisements paléontologiques de pouvoir suivre un individu « complet » et de relier entre eux certains éléments squelettiques avec une bonne sécurité (individu N° 3) Les Figures 28 et 29 représentent le diagramme de répartition de toutes les canines supérieures et inférieures (MDxVL) de Panthera spelaea dont j’ai pu disposer. Elles placent de façon indiscutable Panthera spelaea de Romain-la-Roche parmi les formes de petite taille qui sont à priori des formes du Pléistocène supérieur. Mais un doute subsiste car il existe également des formes de petite taille au Pléistocène moyen récent comme à La Fage, à l’Abri Suard (bAllesio, 1975), à l’Igue des Rameaux que j’étudie. Les valeurs très importantes atteintes par les canines des grandes formes de Panthera spelaea fossilis du Pléistocène moyen médian prouvent leur antériorité. La correspondance avec l’Igue des
Tableau 108 : Romain-la-Roche, Panthera spelaea, navi-culaire.
Panthera spelaeaNaviculaire Pfema
6441 2 3 4
FEL. 53<
26,3 35,5 45,7 D
FEL.119 - 35,4 - g
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.
Tableau 109 : Romain-la-Roche, Panthera spelaea, cu-boïde.
Panthera spelaeaCuboïde
Pfema 6431 2 3 4
FEL. 55<
33,4 29,9 33,0 D
FEL.187 31,8 32,3 35,0 g
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.
Tableau 110 : Romain-la-Roche, Panthera spelaea, 2e cunéiforme.
Panthera spelaea2e cunéiforme
Pfema 6461 2 3 4
FEL. 102 14,3 14,8 24,3 D
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.
Tableau 111 : Romain-la-Roche, Panthera spelaea, 3e cunéiforme.
Panthera spelaea3e cunéiforme
Pfema 6471 2 3 4
FEL. 51<
24,9 25,9 47,2 D
FEL.119 25,0 28,3 45,8 g
1- DAP (longueur) max. 3- DDP (hauteur) max2- DT (largeur) max. 4- Latéralisation.

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 577
Tableau 112 : Romain-la-Roche, Panthera spelaea, métatarsien.
Panthera spelaea – Métatarsiens – Pfema 65
Mtt.2 1 2 3 4 5 6 7 8FEL. 5 135,0* 21,0* 32,0* 17,7* 18,5* 24,8* 24,3* DFEL. 30 - 19,5 30,0 16,0 17,0 - - gFEL. 57(1) 134,8 25,2 31,0 17,8 - 24,0 23,0 DFEL.184 - 19,4 33,6* 15,4 16,4 - - DMtt.3FEL. 8 145,5 28,0 40,0 23,8 17,1 28,2 - gFEL. 57 151,2 32,7 39,0 20,6 17,0 24,2 17,0 DFEL.182 150,0 31,4 41,0* 21,0 17,3 28,4 24,2 gMtt.4FEL. 7 149,2 28,8 31,0 - - - 21,4 gFEL. 54+56 - 26,0 36,4 - - 24,6 24,3 DFEL. 186 - 25,0* 37,7* - - - - gMtt.5FEL. 58 141,8 27,0 17,7 12,8 16,0 21,7 21,0 DFEL. 183 - - - - - 21,4 - g
1- Longueur max.2- DT proximal3- DAP proximal4- DT milieu diaphyse
5- DAP milieu diaphyse6- DT distal7- DAP distal8- Latéralisation(1) soudés par de la brèche (RO.89-P.18-10)
Tableau 113 : Romain-la-Roche, Panthera spelaea, phalange 1.
Phalanges 1 Pfema 73
1L
2DT prox
3DAP prox
4DT méd
5DAP méd
6DT dist
7DAP dist
n 16 14 15 13 13 14 16min. 35,4 19,2 17,5 12,4 13,1 16,0max. 65,4 25,5 21,5 20,7 23,8 21,8moy. 55,98 23,17 19,73 16,50 15,17 18,87 15,26
écart-type 7,60 1,95 1,03 2,26 2,84 1,58 1,77
Tableau 114 : Romain-la-Roche, Panthera spelaea, phalange 2.
Phalanges 2 Pfema 74
1L
2DT prox
3DAP prox
4DT méd
5DAP méd
6DT dist
7DAP dist
n 12 11 10 9 2 12 11min. 37,0 18,2 17,0 11,9 13,0 17,1 10,6max. 49,8 20,4 21,0 13,6 14,0 20,3 16,1moy. 42,55 19,34 19,17 12,92 13,50 18,53 13,91
écart-type 4,60 0,76 1,29 0,57 0,70 1,02 1,47

578 A. ARgANT
Rameaux, fouilles F. rouzAud (Saint-Antonin-Noble-Val, Tarn-et-Garonne) me paraît plus significative. Le site est complexe. Les éléments de datation avancés indiquent une fourchette chronologique allant du stade OIS.9 (300 000 ans environ) à l’Eémien (stade 5), vers 150 000 ans environ selon les couches. Le matériel de Panthera spelaea y est particulièrement abondant et bien conservé. Il permet de réunir les données concernant 17 C sup. et 14 C inf, toutes ces dents appartenant aux couches de la série 6 (60, 62, 63), sauf deux de la couche 53. La datation de ces deux ensembles est à rapporter au stade 5 selon l’étude de la microfaune par M. JeAnnet (JeAnnet, 2005), sous un climat globalement tempéré frais avec influence océanique.Le radius des félins se conserve bien. La comparaison du radius du lion de Romain-la-Roche avec les formes anciennes de la Brèche de Château, les formes intermédiaires d’Azé 1-3, de l’Igue des Rameaux et l’actuel (MHNL + 1 femelle Afrique) montre qu’il présente en fin de compte une taille appréciable contredisant l’impression dentaire. Ce radius est largement de taille supérieure au lion actuel, se rapproche de celui d’Azé du Pléistocène moyen récent. Le radius, bien représenté à l’Igue des Rameaux (n = 10, couche 62) correspond à une taille inférieure. Romain-la-Roche dépasse même la limite supérieure des variations dans pratiquement tous les cas, sauf dans le cas des formes anciennes de taille nettement supérieure. Les formes les plus récentes de Panthera spelaea spelaea des stades 3 et 2, apparaissent de taille inférieure, ce qui permet de les éliminer. Romain-la-Roche se place en position intermédiaire, entre la fin
du stade OIS. 6 et la fin du stade 5. Il ne doit pas être très éloigné chronologiquement du lion des cavernes de l’Igue des Rameaux.Le sacrum de Panthera spelaea, court et large, permet une grande souplesse de l’arrière-train. Il a le mérite d’être un bon indicateur de la taille des individus moins dépendant des facteurs individuels que ne le sont les dents et les os longs. A Romain-la-Roche, le sacrum FEL.261 légèrement endommagé, mais néanmoins complet, permet une comparaison avec celui du Panthera spelaea fossilis de la Brèche de Château pour les formes anciennes, de l’Igue des Rameaux de la fin du Pléistocène moyen récent, et avec l’actuel (Panthera leo, Afrique, femelle). On s’aperçoit que l’exemplaire de Romain-la-Roche dépasse nettement les dimensions d’un sacrum de lion actuel, tout en n’atteignant pas les valeurs du lion de Château. Les quelques données comparables (DCC articulaire max) présentent à Romain-la-Roche des valeurs proches de celles de l’Igue des Rameaux (Tabl. 89).Toutes ces informations concourent à situer chronologiquement le lion de Romain-la-Roche autour de l’Eémien, soit un peu avant vers la fin du stade 6, soit en fin d’Interglaciaire au tout début du stade 4, au moment de l’amorce de la décroissance de taille de la forme typique Panthera spelaea spelaea. La datation des gisements de cette période reste particulièrement délicate. Il existe également une période de fort refroidissement au milieu du stade OIS. 5. Il faut peut-être enfin nuancer le modèle théorique admis et reconnaître un rôle climatique venant perturber les tendances évolutives.
II.2.3.3. Paléoécologie Traditionnellement, le lion des cavernes est considéré comme un animal de milieu découvert des régions tempérées ou tempérées froides (ArgAnt, 1991). Les analyses palynologiques de J. ArgAnt dans la Brèche de Château (Saône-et-Loire) (ArgAnt et al., 2007) et de plusieurs autres gisements concluent souvent à la présence d’un milieu où se côtoient des arbres, conifères et feuillus, mais aussi des espaces ouverts. L’analyse pollinique des coprolithes de hyène de Romain-la-Roche arrive aussi à des conclusions similaires.Mais il ne faut pas oublier que de grands félins actuels comme le tigre de Sibérie peuvent résister à des conditions climatiques très rigoureuses en hiver et certaines autres populations de la même espèce être parfaitement adaptées à des conditions tropicales. L’expansion eurasiatique de Panthera (Leo) spelaea fossilis au cours du Pléistocène moyen, depuis 700 000 ans, s’est poursuivie jusqu’en Amérique du Nord forcément par le détroit de Behring. Elle est à l’origine des lions américains (Kurtén & Andersen, 1980). Ce passage par le détroit nécessite qu’il soit dépourvu de sa tranche d’eau de l’ordre de 60 m. Seul, un climat froid capable de causer une baisse importante du niveau marin peut l’expliquer, en liaison peut-être avec la tectonique mais cette dernière ne peut être seule en cause. A l’évidence, Panthera spelaea
Tableau 115 : Romain-la-Roche, Panthera spelaea, pha-lange 3.
Phalanges 3 Pfema 75
1DT prox. max.
2DDP prox. max.
n 17 8min. 11,6 36,4max. 19,0 55,0moy. 14,98 43,98
écart-type 1,83 5,80
Tableau 116 : Romain-la-Roche, Panthera spelaea, sésa-moïdes.
Sésamoïdes Pfema 76
1L max.
2l max.
3Hauteur (x3)
n 16 10 10min. 17,9 8,4 11,2max. 27,5 10,9 14,0moy. 23,68 9,74 12,89
écart-type 3,70 0,72 0,33

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 579
RO. Ind.3RO. Ind.1
Romain-la-RocheP. s. fossilisP. s. interm.P. s. spelaea
Romain-la-Roche, Panthea spelaea. Canines supérieures, comparaison
V.L.
32,0
30,0
28,0
26,0
24,0
22,0
20,0
18,0
19,0
16,0
14,021,0 23,0 27,0 29,0 31,0 33,0 35,0 37,025,0
M.D.
Fig. 28 : Rapport longueur/largeur des canines supérieures de Panthera spelaea depuis le Pléistocène moyen médian jusqu’au Pléistocène supérieur de nombreux sites d’Europe occidentale. Position des lions de Romain-la-Roche (étoiles).
RO. Ind.3
RO. Ind.1
Romain-la-RocheP. s. fossilisP. s. interm.P. s. spelaea
Romain-la-Roche, Panthea spelaea. Canines inférieures, comparaison
V.L.
32,0
30,0
28,0
26,0
24,0
22,0
20,0
18,0
19,0
16,0
14,021,0 23,0 25,0 27,0 29,0 31,0 33,0 35,035,0 37,025,0
M.D.
Fig. 29 : Rapport longueur/largeur des canines inférieures de Panthera spelaea depuis le Pléistocène moyen médian jusqu’au Pléistocène supérieur de nombreux sites d’Europe occidentale. Position des lions de Romain-la-Roche (étoiles).

580 A. ARgANT
fossilis et le jaguar Panthera gombaszoegensis qui a suivi le même trajet, ont, à une époque au moins, prouvé une adaptation aux climats les plus rigoureux des périodes glaciaires. Il faut donc nuancer la première impression et admettre que Panthera spelaea résiste bien dans certains cas à des climats froids. La prudence doit donc être de règle avant de lui accorder une signification climatique précise.
III. CONCLUSION
Cette étude des carnivores (mustélidés exceptés) de Romain-la-Roche, montre toute l’importance des séries de vestiges piégées dans l’aven. Les informations morphométriques qu’il est possible d’en tirer constituent une base de données conséquente précisant les faunes européennes. Les carnivores fournissent tout comme d’autres groupes des indications biochronologiques. A Romain-la-Roche, les degrés évolutifs des canidés (Cuon alpinus europaeus et Canis lupus), des ursidés (Ursus spelaeus) et des félidés (Panthera spelaea), permettent d’avancer des éléments de datation du gisement.Rappelons ces éléments :- Cuon alpinus europaeus :Le cuon de Romain-la-Roche possède une dentition dépassant en taille les formes du Pléistocène supérieur du dernier épisode glaciaire. Malgré cette forte taille, il ne s’agit pas de Cuon priscus du Pléistocène moyen. Il présente par contre une bonne ressemblance (grande taille, denticule antérieur à P/4) avec les individus d’Atapuerca Tg.3 (Trinchera galeria) en Espagne, rapportés à la fin du Pléistocène moyen.- Canis lupus :Le loup de Romain-la-Roche apparaît proche du point de vue dentaire de la population de Canis lupus santanaisiensis de Santenay, Pointe-du-Bois (Côte d’Or) en Bourgogne. Ce gisement célèbre, malheureusement mal daté, est le plus souvent attribué à la fin du Pléistocène moyen récent (OIS. 6), soit au dernier Interglaciaire Eémien (OIS. 5), au Pléistocène supérieur ancien. Il n’atteint pas en tout cas la grande taille, ni la denture massive et plus broyeuse des loups du Pléistocène supérieur moyen ou récent (stades OIS. 4 ou 3).
- Ursus spelaeus :L’ours des cavernes de Romain-la-Roche est très nettement spéléen. Il est antérieur aux formes les plus grandes, lourdes et massives typiques du dernier glaciaire (stades OIS. 4 et 3). Son degré évolutif le place après la population de l’Ursus spelaeus denigeroides d’Azé 1-3, datée du Pléistocène moyen récent (stade OIS. 6), mais il n’en est pas très éloigné et des caractères ancestraux d’Ursus deningeri ressortent parfois ici, comme à Santenay ou dans des gisements du Pléistocène supérieur ancien de Bourgogne (Argant, 1991). La population de la grotte des Furtins-Ossuaire 2 (Berzé-la-Ville, Saône-et-Loire) paraît plus récente. Elle correspond probablement à un des épisodes tempérés « Saint-Germain » de la fin de l’Eémien ou du début du Pléistocène supérieur moyen (« Würm ancien »).- Panthera spelaea :La taille du lion des cavernes de Romain-la-Roche place cette population au Pléistocène supérieur récent (stades OIS. 3 ou 2) si on admet une décroissance régulière de taille au cours de l’évolution. Il subsiste cependant dans cette évolution des zones d’ombre pour les formes de la fin du Pléistocène moyen. Des individus de petite taille sont connus, par exemple à La Fage (Corrèze) au cours du stade OIS. 6. Le Panthera spelaea de Romain-la-Roche peut tout aussi bien se rattacher chronologiquement à la fin de cette période. - Un argument indirect vient conforter ces éléments. En Bourgogne, la hyène des cavernes Crocuta spelaea ne devient vraiment présente dans les gisements qu’à partir du Pléistocène supérieur moyen (« Würm ancien »). Sans être totalement absents auparavant, les hyénidés sont tellement rares qu’on ne rencontre que peu d’indices, parfois seulement leurs coprolithes très caractéristiques, comme justement à Romain-la-Roche.Finalement, la fourchette chronologique que l’on peut retenir pour la datation du site grâce aux carnivores se place entre l’extrême fin du Pléistocène moyen récent et la fin du Pléistocène supérieur ancien (Eémien). La méthode de l’indice morphodynamique de g. rAbeder situe l’ours des cavernes de Romain-la-Roche au même degré d’évolution que celui des populations anciennes d’Herdengelhöhle en Autriche, entre 135 Ka et 112 Ka, c’est-à-dire au cours du stade OIS. 5.
Planche I
Fig. 1: Romain-la-Roche, Cuon alpinus europaeus, demi-mandibule droite CAN. 1069, vue latérale externe.Fig. 2: Romain-la-Roche, Cuon alpinus europaeus, demi-mandibule droite CAN. 1069, vue latérale interne.Fig. 3: Romain-la-Roche, Canis lupus, os incisif supérieur droit CAN. 1914, canine supérieure, I1/, I2/, I3/, vue occlusale.Fig. 4: Romain-la-Roche, Canis lupus, P4/ droite, CAN. 921, vue linguale.Fig. 5: Romain-la-Roche, Canis lupus, fragment de maxillaire supérieur droit CAN.842 avec C. supérieure, alvéole de
P1/, P2/, P3/, P4/ droites, vue vestibulaire.Fig. 6: Romain-la-Roche, Canis lupus, fragment de maxillaire supérieur gauche CAN.631 avec P4/, M1/, et M2/ gauches,
vue occlusale.

Planche I
1
2
4
3
5
6
1cm
1cm
1cm
1cm
1cm

582 A. ARgANT
REMERCIEMENTS
Il m’est agréable de remercier très sincèrement tous ceux qui ont participé à cette longue « chaîne opératoire » conduisant de l’extraction des fossiles aux cartons d’ossements des réserves du Musée Cuvier de Montbéliard qui conserve maintenant le matériel. Je remercie tout spécialement Thierry mAlvesy, Responsable des Collections, pour sa disponibilité et son accueil chaleureux toutes les fois où je suis allé sur place étudier le matériel paléontologique. Un grand merci également à Claude guérin pour m’avoir confié cette étude, il y a bien longtemps en 1987. Jacqueline ArgAnt a eu la lourde tâche de la mise au point informatique de tous les tableaux, figures, photos du manuscrit final. Elle a droit à toute ma reconnaissance. Je remercie également Nuria gArCiA et Lars Werdelin pour leurs commentaires et leurs judicieux conseils.Je garde enfin une pensée émue pour Patrick PAuPe, personnage hors du commun, sans qui rien de tout cela n’aurait pu arriver.
RéFéRENCES BIBLIOgRAPHIQUES AdAm, K.d. (1959) - Mittelpleistozäne caniden aus den
Heppenloch bei gutenberg (Würtemberg). Stuttgarter Beitrage zur naturkunde, 27 : 1-46.
Alimen, h., C. ArAmbourg & A. sChreuder (1958) - La grotte de Fontéchevade (3e partie). Archives de l’Institut de Paléontologie humaine, Mémoire 29 : 1-262.
AltunA, J. (1973) - Hallazgos de Oso pardo (Ursus arctos, Mammalia) en cuevas del Pais Vasco. Munibe, San sebastian, Ano XXV, 2-4 : 121-170.
ArgAnt, A. (1988) - Etude de l’exemplaire de Panthera spelaea (goldfuss, 1810) (Mammalia, Carnivora, Felidea) du gisement pléistocène moyen récent de la grotte d’Azé (Saône-et-Loire). Revue de Paléobiologie, Genève, 7 (2) : 449-466.
ArgAnt, A. (1991) - Carnivores quaternaires de Bourgogne. Documents du Laboratoire de Géologie de Lyon, 115 : 1-301.
ArgAnt, A. (1995) - Un essai de biochronologie à partir de l’évolution dentaire de l’ours des cavernes. Datation du site de la Balme à Collomb (Entremont-le-Vieux, Savoie, France). Quaternaire, 6 (3-4), Paris : 139-149.
ArgAnt, A. (2004a) - Les Carnivores du gisement Pliocène final de Saint-Vallier (Drôme, France). In : fAure, M.
& C. guérin (Eds). Le gisement pliocène final de Saint-Vallier (Drôme, France). Geobios, MS 26 : S133-S182.
ArgAnt, A. (2004 b) - Rapports Hommes-Carnivores au Paléolithique inférieur d’Azé I-1 (Saône-et-Loire, France) : comparaison taphonomique des assemblages de faune des différents secteurs d’Azé I. Actes du XIVe Congrès UISPP « Hommes et Carnivores au Paléolithique », Liège 2001, Revue de Paléobiologie, Genève, 23 (2) : 803-819.
ArgAnt, A. (2009) - Biochronologie et grands Mammifères au Pléistocène moyen et supérieur en Europe occidentale : l’apport des Canidés, des Ursidés et des Carnivores en général. Quaternaire, 20 (4) : 467-480.
ArgAnt, A. & J. ArgAnt (2002) - Die Bären von Château (Burgund, Frankreich). In : rosendAhl, W., m. morgAn & m. lóPez CorreA (Ed.). Cave-Bear-Researches/ Höhlen-Bären-Forschungen, Abhandlung zur Karst- und Höhlenkunde, München, Heft 34 : 57-63.
ArgAnt, A., m. JeAnnet, J. ArgAnt & m. erbAJevA (2007) - The big cats of the fossil site Château Breccia Northern Section (Saône-et-Loire, Burgundy, France) : stratigraphy, palaeoenvironment, ethology and biochronological dating. Courier Forschunginstitut Senckenberg, Frankfurt a. M., 259 : 121-140.
ArgAnt, A., M. PhiliPPe & J. ArgAnt (2009) - Chronologie de la faune fossile quaternaire de Chartreuse. Implications paléoenvironnementales. Rapport d’étude du projet Oursalp-Chartreuse, Parc naturel régional de Chartreuse, Saint-Pierre-de-Chartreuse, 3 : 1-29.
ArgAnt, J. (2010) - Palynologie des coprolithes d’hyène de Romain-la-Roche (Doubs, France) : apport paléoenvironnemental. In : guérin, C. & Th. mAlvesy, L’aven pléistocène moyen final de Romain-la-Roche (Doubs, France). Revue de Paléobiologie, 29 (2) : 473-476.
Artois, m. (1989) - Le renard roux (Vulpes vulpes linnAeus, 1758). Encyclopédie des Carnivores de France. Société française pour l’étude et la protection des Mammifères, Nort-sur-Erdre, 3 : 1-90.
Athen, K., C. frömKe & m. germonPré (2005) - Analysis of postcranial elements of cave bear material (Ursus spelaeus) from goyet (Condroz, Belgium). Bulletin de l’Institut royal des sciences naturelles de Belgique, Sciences de la Terre, 75 : 267-283.
Auguste P. (1988) - Etude des restes osseux des grands Mammifères des niveaux D, D1, II base. In : tuffreAu, A. & J. sommé (dir.). Le gisement paléolithique moyen de Biache-Saint-Vaast (Pas-de-Calais), vol. I, Stratigraphie, environnement, Etudes archéologiques (1ère partie). Mémoires de la Société Préhistorique Française, 21 : 133-169.
bAllesio, r. (1975) - Etude de Panthera (Leo) spelaea (goldfuss) nov. sub sp. (Mammalia, Carnivora, Felidae)
Planche II
Fig. 1: Romain-la-Roche, Canis lupus, demi-mandibule gauche CAN.1187, vue latérale externe.Fig. 2: Romain-la-Roche, Canis lupus, demi-mandibule gauche CAN.1187, vue latérale interne.Fig. 3: Romain-la-Roche, Canis lupus, ensemble droit calcanéum et astragale soudés CAN. 1294, vue latérale externe. Fig. 4: Romain-la-Roche, Canis lupus, ensemble droit calcanéum et astragale soudés CAN. 1294, vue latérale interne.Fig. 5: Romain-la-Roche, Canis lupus, ensemble droit calcanéum et astragale soudés CAN. 1294, vue dorsale.

Planche II
1 cm
1 cm
1 cm
1
2
3 4
5

584 A. ARgANT
du gisement pléistocène moyen des Abîmes de la Fage à Noailles (Corrèze). Nouvelles archives du Muséum d’histoire naturelle de Lyon, 13 : 47-55.
bAllesio, r. (1980) - Le gisement pléistocène supérieur de la grotte de Jaurens à Nespouls, Corrèze, France : les Carnivores (Mammalia, Carnivora), Felidae. Nouvelles archives du Muséum d’histoire naturelle de Lyon, 18 : 61-102.
bAllesio, r. (1983) - Le gisement pléistocène supérieur de la grotte de Jaurens à Nespouls, Corrèze, France : les Carnivores (Mammalia, Carnivora), III- Ursidae, Ursus arctos Linnaeus. Nouvelles archives du Muséum d’histoire naturelle de Lyon, 21 : 9-43.
bArone, r. (1976) - Anatomie comparée des Mammifères domestiques. Vigot Frères édit., Paris, 1 : 1-269, 2 : 1-428.
bAryshniKov, g.f. & f. dAvid (2000) - les ours des cavernes à Arcy-sur-Cure (Yonne, France) – Ursus (spelearctos) spelaeus Rosenmüller et Heinroth, 1974. Quaternaire, 11 (1) : 65-79.
beAufort (de) f. (1987) - Le loup en France : éléments d’écologie historique. Encyclopédie des Carnivores de France. Société française pour l’étude et la protection des Mammifères, Nort-sur-Erdre, 1 : 1-32.
berger, g.W., A. Pérez-gonzález, e. CArbonell, J-l. ArsuAgA, J.-m. bermúdez de CAstro & t.-l. Ku (2008) - Luminescence chronology of cave sediments at the Atapuerca paleoanthropological site, Spain. Journal of Human Evolution, 55 : 300-311.
bisChoff, J.l., r.W. WilliAms, r.J. rosenbAuer, A. ArAnburu, J.l. ArsuAgA, n. gArCiA & g. CuenCA-besCós (2007) - High resolution U-series dates from the Sima de los Huesos hominids yields 600+∞/-66 kyrs : implications for the evolution of the Neanderthal lineage. Journal of Archaeological Science, 34 : 763-770.
bonifAy, M.-F. (1971) - Carnivores quaternaires du sud-est de la France. Mémoires du Muséum National d’Histoire naturelle, Paris, n.s., série C, 21 (2) : 1-377.
boule, M. (1906) - Les grands chats des cavernes. Annales de Paléontologie, Paris, 1 : 1-27.
boule, M. & L. villeneuve (de) (1927) - La grotte de l’Observatoire à Monaco. Archives de l’Institut de Paléontologie humaine, Paris, 1 : 1-113.
burger, J., W. rosendAhl, o. loreille, h. hemmer, t. eriKsson, A. götherström, J. hiller, m.J. Collins, m.J.t. Wess & K.W. Alt (2004) - Molecular phylogeny of the extinct cave lion Panthera leo spelaea. Molecular Phylogeny and Evolution, 30 : 841-849.
CAmPy M. (1982) - Le Quaternaire franc-comtois. Essai chronologique et paléoclimatique. Thèse de Doctorat ès Sciences naturelles, Université de Franche-Comté, n°159 : 575 p.
CAmPy, M. (Dir.) (1983) - L’Aven de Vergranne (Doubs). Un site du Pléistocène moyen à Hominidé. Annales scientifiques de l’Université de Besançon, Géologie, 4e série, 5 : 1-92.
ChAgneAu, J. (1985) - Contribution à l’étude des os des extrémités des pattes d’Ursus deningeri romeviensis. Comparaison avec Ursus arctos et Ursus spelaeus. Bulletin de la Société d’Anthropologie du Sud-Ouest, 20 (2-3) : 61-107.
ChAgneAu, J. (1989) - Les Ursidés. In : CAmPy, m., J. ChAline & m. vuillemey. La Baume de Gigny (Jura). XXVIIe supplément à gallia Préhistoire, CNRS éditeur : 69-76.
ChAgneAu, J. & C. PrAt (1983) - Les Ursidés de l’aven de Vergranne (Doubs). In : L’aven de Vergranne (Doubs). Un site du pléistocène moyen à Hominidé. Annales scientifiques de l’Université de Besançon, Géologie, 4e série, 5 : 93-109.
ChAline, J. (1969) - Les faunes de Mammifères du Pléistocène de Bourgogne. In : Ciry r., Livret guide excursion A1, Alsace-Vosges-Bourgogne, 8ème Congrès INQUA, Paris : 1-100
ChAPut, F. (2000) - La faune moustérienne du gisement de Vergisson 2 (Saône-et-Loire, France). Mémoire de licence, Institut de préhistoire et des sciences de l’Antiquité, Université de Neufchâtel : 1-230.
Clot, A. (1980) - La grotte de la Carrière (gerde, Hautes-Pyrénées). Stratigraphie et paléontologie des Carnivores. Travaux du Laboratoire de Géologie de l’Université Paul Sabatier, Toulouse, 1 : 1-239, 2 : 1-263.
Clot, A. (1987) - La grotte de Gerde (Hautes-Pyrénées), site préhistorique et paléontologique. Société Ramond, Bagnères-de-Bigorre : 1-210.
Clot, A. & f. durAnthon (1990) - Les mammifères fossiles du Quaternaire des Pyrénées. Muséum d’histoire naturelle de Toulouse : 1-159.
Cordy, J-M. (1983) - Découverte de Cuon alpinus europaeus bourguignAt dans le Quaternaire de Belgique. Mémoire de la Société préhistorique française, Paris, 16 : 49-59.
Couturier, m. (1954) - L’ours brun. Couturier M. édit., grenoble : 1-903.
Crégut-bonnoure E., 1996 - Ordre des Carnivores : Canidae. In : guérin, C. & M. PAtou-mAthis (Dir.). Les grands mammifères plio-pléistocènes d’Europe, Masson édit, Paris : 155-166.
Crégut, E. (1979) - La faune de Mammifères du Pléistocène moyen de la Caune de l’Arago à Tautavel (Pyrénées-Orientales). Travaux du laboratoire de Paléontologie humaine et de Préhistoire, Marseille, 3 (1) : 1-381.
delPeCh, f. (1989) - Les grands mammifères, à l’exception des Ursidés. In : rigAud et al. La grotte Vaufrey (Dordogne).
Planche III
Fig. 1 : Romain-la-Roche, Ursus arctos, demi-mandibule droite URS.271 : a. Vue occlusale ; b. Comparaison avec une mandibule d’Ursus arctos actuel, vue occlusale.Fig. 2 : Romain-la-Roche, Ursus spelaeus, calcanéum droit URS. 798 : a. Vue dorsale b.Vue plantaire.Fig. 3 : Romain-la-Roche, Ursus spelaeus, astragale droit URS. 438, vue plantaire.

Planche III
1a
b
2
3
a b
1 cm
1 cm
1 cm

586 A. ARgANT
Paléoenvironnement, chronologie, activités humaines. Mémoires de la Société préhistorique française, 19 : 213-289.
Desse, J., l. ChAix & n. desse-berset (1986) - « Ostéo », Base-réseau de données ostéométriques pour l’archéozoologie. Procédures, codages, exploitation des données, organisation du réseau. Editions du C.N.R.S., Valbonne : 161 p.
döPPes, d., s. KemPe & W. rosendAhl (2008) - Dated Paleontological cave sites of Central Europe from Late Middle Pleistocene to early Upper Pleistocene (OIS 5 to OIS 8). Quaternary International, 187 : 97-104.
döPPes, d. & g. rAbeder (eds) (1997) - Pliozäne und pleistozäne Faunen Österreichs. Verlag der Österreichischen Akademie der Wissenschaften, 10 : 1-411.
dubois, A. & h.g. stehlin (1933) - La grotte de Cotencher, station moustérienne. Mémoires de la Société paléontologique Suisse, Bâle, 52-53 : 1-292.
gArCiA, N. (2003) - Osos y otros carnivoros de la Sierra de Atapuerca. Fondation oso de Asturias : 1-575.
gAsPArd, M. (1964) - La région de l’angle mandibulaire chez les Canidae. Mammalia, 28 : 249-329.
germonPré, m. & m.v. sAblin (2001) - The cave bear (Ursus spelaeus) from goyet, Belgium). Bulletin de l’Institut royal des sciences naturelles de Belgique, Sciences de la Terre, 71 : 209-233.
grossouvre A. & stehlin h. g. (1912) - Les sables de Rosières, près Saint-Florent (Cher). II. Paléontologie. Bulletin de la Société Géologique de France, Paris, 4e
série, 12 : 194-212.hue, E. (1907) - Musée ostéologique. Etude de la faune
quaternaire. Ostéologie des mammifères. Ed. Schleicher, Paris, 2 fascicules, album de 186 planches.
JeAnnet, M. (2005) - La microfaune de l’Igue des Rameaux à Saint-Antonin-Noble-Val (Tarn-et-garonne, France). Essai de Biostratigraphie. Préhistoire du Sud-Ouest, 2 : 109-125.
KleCzKo, S., 1999 - Position du gisement de la Belle-Roche dans le contexte de l’évolution des faunes mammaliennes continentales en Europe. Mémoire de licence de biologie animale, Université de Liège, 52 p.
Koby, F.E. (1949) - Le dimorphisme sexuel des canines d’Ursus arctos et d’Ursus spelaeus. Revue suisse de Zoologie, 56 (36) : 675-687.
Kurtén, b. & e. Anderson (1980) - Pleistocene mammals
of North America. Columbia University press, New-York : 1-442.
LehmAnn, U. (1954) - Die Fauna des « Vogelherds » bei Stettten ob Lontal (Würtemberg). Neues Jahrbuch für Geologie und Paläontologie Abh., Stuttgart, 99 : 33-146.
leroi-gourhAn, A. (1950) - La caverne des Furtins. Préhistoire. Presses Universitaires de France, Paris, 11 : 17-125.
mAJor, C.J.F. (1900) - Remarks on remains of Cyon sardous from a cave in Sardinia. Proceedings of the zoological Society of London.
nehring, A. (1890) - Über Cuon alpinus foss. Aus dem Heppenloch in Württemberg. Sitzungsberichte der Gesellschaft naturforschender Freunde zu Berlin.
PAtou, M. (1984) - Contribution à l’étude des Mammifères des couches supérieures de la grotte du Lazaret (Nice, Alpes-Maritimes). Thèse de Doctorat 3e cycle, Université P. et M. Curie, Paris 6 : 1-437.
PAuPe, P. (1994-1995) - L’aven de Romain-la-Roche. Rapport d’activités 1994-1995. Association pour la sauvegarde des sites préhistoriques et paléontologiques du canton de Rougemont et du pays de Montbéliard : 1-45.
PAuPe, P. (1996) - L’aven de Romain-la-Roche : un site archéologique européen. Extrait du Bulletin de la Société d’Émulation de Montbéliard, 119 : 1-96.
PAuPe, P. (1997) Il y a 150 000 ans, un site paléontologique et préhistorique : l’aven de Romain-la-Roche. Bibliothèque de travail, Publications de l’Ecole moderne française, Cannes, 1085 : 1-33.
PAutret-homerville, C. (2000) - Les Carnivores pléistocènes de la Grotte de Châteaubourg en Ardèche. Mémoire de Maîtrise d’Archéologie, Université Paris I : 1-77.
PoPlin, F. (1976) - Les grands vertébrés de Gönnersdorf. Fouilles 1968. Franz Steiner. Verlag gmbH, Wiesbaden : 1-212.
PrAt, f. & C. thibAult (1976) - Le gisement de Nauterie à la Romieu (gers). Fouilles de 1967 à 1973. Nauterie I. Mémoires du Muséum d’histoire naturelle de Paris, Paris, N.S., série C Sciences de la Terre, 35 : 1-82.
rAbeder, g. (1983) - Neues vom Hölhlenbären : Zur Morphogenetik der Backenzähne. Die Höhle, Wien, 34 (2) : 67-85.
rAbeder, g. (1989) - Modus und gesschwindigkeit der Höhlenbären-Evolution. Schriften des Vereins zur
Planche IV
Fig. 1: Romain-la-Roche, Ursus spelaeus, rangée des dents jugales de la demi-mandibule gauche URS.1085, vue linguale.
Fig. 2: Romain-la-Roche, Ursus spelaeus, M/3 gauche URS. 285, vue occlusale.Fig. 3: Romain-la-Roche, Ursus spelaeus, M1/ droite URS.442, vue occlusale.Fig. 4: Romain-la-Roche, Ursus spelaeus, M/1, vues occlusales : a. URS. 282 ; b. URS. 284.Fig. 5: Romain-la-Roche, Ursus spelaeus, fragment de maxillaire supérieur droit URS. 681 avec P4/ et alvéole vide de
P3/, vue occlusale.Fig. 6: Romain-la-Roche, Ursus arctos, phalange 3 URS. 844.Fig. 7: Romain-la-Roche, Ursus spelaeus, mandibule URS. 1439, vue occlusale.Fig. 8: Romain-la-Roche, Ursus spelaeus, même mandibule URS. 1439, détail de la partie incisive et des deux canines
cassées puis usées.

Planche IV
1
2
46
7
5 a
b
3
8
1 cm
1 cm
1 cm
1 cm
5 cm
1 cm

588 A. ARgANT
Verbreitunq naturwissenschaftlicher Kenntnisse, Wien, 127 : 105-126.
rAbeder, g. (1991) - Die Höhlenbären von Conturines. Entdeckung und Erforschung einer Dolomiten-Höhle in 2800 m Höhe. Athesia Verlag, Bozen : 1-125.
rAbeder, g. (1992) - Das Evolutionsniveau des Höhlenbären aus dem Nixloch bei Losenstein-Ternberg (O.O.). Mitteilungen der Kommission für Quartärforschung der Österreichischen Akademie der Wissenschaften, Wien, 8 : 133-141.
rAbeder, g. (1995a) - Evolutionsniveau und Chronologie der Höhlenbären aus der gamssulzen-Höhle im Toten gebirge (Oberösterreich). Mitteilungen der Kommission für Quartärforschung der Österreichischen Akademie der Wissenschaften, 9 : 69-81.
rAbeder, g. (1995b) - Chronologie der gamssulzen-Höhle im Toten gebirge (Oberösterreich). Mitteilungen der Kommission für Quartärforschung der Österreichischen Akademie der Wissenschaften, 9 : 129-133.
rAbeder, g. (1999) - Die Evolution des Höhlenbärengebisses. Mitteilungen der Kommission für Quartärforschung der Österreichischen Akademie der Wissenschaften, 11 : 1-102
sChütt, g. & H. hemmer (1978) - Zur Evolution des Löwen (Panthera leo L.) im europäischen Pleistozän. Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, (4) : 228-255.
thenius, E. (1954) - Die Caniden (Mammalia) aus dem Altquartär von Hundsheim (Niederösterreich) nebst Bemerkungen zur stammesgeschichte der Gattung Cuon. Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, 99, 2 : 230-286.
tobien, H. (1957) - Cuon Hodg. und Gulo Frisch (Carnivora, Mammalia) aus den altpleistozän Sanden von Mosbach bei Wiesbaden. Acta zoologica cracoviensiai, II, 18 : 433-451.
vAn den brinK, f.h. & P. bArruel (1967) - Guide des Mammifères sauvages d’Europe. Delachaux et Niestlé édit. : 1-263.
veresCAgin, N.K. (1969) - Le lion des cavernes : Panthera (Leo) spelaea goldfuss et son histoire dans l’Holarctique. Etudes sur le quaternaire dans le monde, VIIIe Congrès INQUA, Paris, 1 : 463-464.
viret, J. (1954) - Le loess à bancs durcis de Saint-Vallier (Drôme) et sa faune de mammifères villafranchiens, Nouvelles Archives du Muséum d’Histoire naturelle de Lyon : 1-20.
WeinstoCK, J. (1999) - The Upper Pleistocene mammalian fauna from the große grotte near Balubeuren (southwestern germany). Stuttgarter Beitrage Naturkunde, Serie B, Stuttgart, 277 : 1-49.
ANNEXE : inventaire du matériel par espèce
La liste d’inventaire, mise en annexe à cause de sa longueur, a néanmoins un rôle important. Elle constitue le corpus d’étude de chacune des espèces des trois Familles de carnivores de cet article. Elle permet, si nécessaire, de retrouver la détermination exacte et la latéralisation de chaque pièce référencée tenue en main. Prenons un exemple – CAN.726 – la recherche, par tâtonnement il est vrai, peut avancer assez rapidement si on part du constat qu’il s’agit d’une dent faisant partie d’un ensemble (ici fragment d’os incisif d’un crâne). On arrive à I 3/g de Canis lupus. Cette liste donne une idée précise du volume du matériel et peut permettre le décompte des pièces de chaque espèce pour une étude taphonomique par exemple. Dans certains cas enfin, c’est la seule mention de pièces encore déterminables, mais trop fragmentaires pour être utilisées.
CAN.1 : Vulpes lagopus (NR = 5, NMI = 1)
- CAN.1019 : C. sup. g. - dent isolée, U = 0- CAN.1022 : C. sup. d. - dent isolée, U = 0- CAN.1807 : 7° vertèbre lombaire endommagée- CAN. 1788 : Mtt. 3 d. - extrémité distale manquante- CAN. 787 : ext. dist. fémur d.
CAN.2 : Vulpes vulpes (NR = 42, NMI = 4)
Demi-mandibule : CAN.17+44 d. ; CAN.1773 d. ; CAN. 20 g. endommagée ; CAN. 169 d fragmentDents isolées :Dents supérieures :- CAN. 45 : P4/ g. – dent isolée, complète, U = 0.- CAN. 1135 : P4/ d. – dent isolée, complète, U = 0.Dents inférieures : - CAN. 1928 : C. inf. d. – dent isolée, couronne endommagée,
U = 0,2- CAN. 642 : P/3 d. – dent isolée, U = 1.- CAN. 1204 : P/3 d. – dent isolée, couronne seule, creuse
(jeune individu), U = 0.- CAN. 1020 : P/4 d. – dent isolée, couronne seule, creuse
(même individu que CAN. 1204 probable), U = 0.- CAN. 136 : M/1 d. – dent isolée, couronne seule, creuse, U =
0.
Planche V
Fig. 1: Romain-la-Roche, Ursus spelaeus, crâne complet URS. 01 (exposition), vue de dessus.Fig. 2: Romain-la-Roche, Felis silvestris, canine supérieure gauche FEL. 226, vue linguale.Fig. 3: Romain-la-Roche, Panthera spelaea, rotule droite FEL. 215, vue de face.Fig. 4: Romain-la-Roche, Panthera pardus, 4e métatarsien gauche FEL. 227, moitié proximale, vue dorsale.Fig. 5: Romain-la-Roche, Canis lupus, axis CAN. 526, vue dorsale.Fig. 6: Romain-la-Roche, Panthera spelaea, astragale gauche FEL. 223, vue dorsale.

Planche V
1
2
3
4
5 61 cm
5 cm
1 cm
1 cm
1 cm
1 cm

590 A. ARgANT
Squelette post-crânien :Colonne vertébrale : atlas (CAN.22) ; axis (CAN.947)Humérus : CAN.1700 : moitié dist. humérus g.Cubitus : CAN.1472 : tiers proximal cubitus g.- CAN.1182 : ext. prox. cubitus d. endommagée.- CAN.1157 : cubitus g. à extrémité dist. manquante- CAN.28 : tiers prox. cubitus g.- CAN.1516 : ext. prox. cubitus d.- CAN.427 : ext. prox. cubitus g.Radius : CAN.26 : radius g. complet- CAN.949 : moitié prox. radius d.Métacarpien 5 : CAN.1864 : Mtc.5 g. complet.- CAN.1785 : Mtc.5 d. complet. Fémur : CAN.1176 : ext. prox. fémur g. endommagée.- CAN.1791 : ext. prox. fémur d. très endommagée.- CAN.1791 : ext. dist. fémur g.- CAN.1790 : ext. dist. fémur d.Tibia : CAN.1637 : tibia g. complet- CAN.1027 : tibia d. endommagé- CAN.1774 : ext. prox. tibia g.- CAN.1769 : moitié dist. tibia g. endommagée.- CAN.1782 : ext. dist. tibia d.- CAN.1690 : ext. dist. tibia d. endommagée.- CAN.1028 : ext. dist. tibia d. Calcanéum : CAN.1743 : calcanéum g. très endommagé.- CAN.27 : calcanéum d. complet.- CAN.1018 : calcanéum g. complet.- CAN.1005 : calcanéum d. endommagé.- CAN.1002 : calcanéum d. endommagé.Astragale : CAN.1485 : astragale g.
CAN.3 – Canis lupus (NR = 1616, NMI = 25)
Matériel crânien :Aucun crâne n’est complet. Seuls les fragments significatifs apportant des données sont utilisés : CAN.392, CAN.1703, CAN.378, CAN.705, CAN.791, CAN.620, CAN.842, CAN.1274, NEA.05. Soit au moins 10 crânes et sans doute nettement plus.Denture supérieure : I1/ - sur fragment d’os incisif CAN.1914 d, CAN.273 g. Dents isolées : CAN.1509, CAN.1902, CAN.1900, CAN.836, CAN.1325, CAN.763, CAN.151, CAN.1320 g ; CAN.417, CAN.561, CAN.1261, CAN.447, CAN.1323, CAN.733, CAN.647, CAN.656 d. Soit n = 9 g et 9 d.I2/ - sur fragment d’os incisif CAN.1914, CAN.866 d. Dents isolées : CAN.237, CAN.145, CAN.1926, CAN.1506, CAN.1836, CAN.762, CAN.1050 g ; CAN.1505, CAN.703, CAN.237, CAN.646 d. Soit n = 7 g et 6 d.I3/ - sur fragment d’os incisif CAN.620, CAN.392, CAN.662, CAN.726 g ; CAN.620, CAN.392, CAN.1914, CAN.866 d. Dents isolées : CAN.1497, CAN.1457, CAN.1475, CAN.1926,
CAN.162, CAN.83, CAN.206, CAN.1322, CAN.779, CAN.415, CAN.330, CAN.1897, CAN.1210, CAN.539, CAN.648, CAN.1249, CAN.1572, CAN.1498 g ; CAN.1659, CAN.886, CAN.637, CAN.1687, CAN.743, CAN.1136, CAN.693, CAN.1127, CAN.635, CAN.88, CAN.1310, CAN.1478, CAN.138, CAN.761, CAN.525, CAN.1321, CAN.1260, CAN.1569, CAN.532 d. Soit n = 22 g et 23 d.C sup. - ensemble : CAN.1703, CAN.392, CAN.1274, CAN.1792, NEA.06 g ; CAN.1703, CAN.392, CAN.842, CAN.1274, CAN.1914 d. Dents isolées : CAN.951, CAN.620, CAN.1201, CAN.323, CAN.1309, CAN.1926, CAN.926, CAN.262, CAN.750, CAN.35, CAN.795, CAN.815, CAN.1259, CAN.1538, CAN.816, CAN.692, CAN.418, CAN.110, CAN.1921, CAN.599 g ; CAN. 1134, CAN.620, CAN.1479, CAN.933, CAN.1111, CAN.924, CAN.600, CAN.1349, CAN.502, CAN.1070, CAN.334, CAN.1865, CAN.611, CAN.771 d. Soit n = 25 g et 19 d.P1/ - ensemble : CAN.1703, CAN.1274, CAN.790 g ; CAN.392 d. Dents isolées : CAN.960, CAN.827, CAN.625, CAN.1744, CAN.410 g ; CAN.163, CAN.1074, CAN.1644, CAN.923, CAN.872, CAN.1552, CAN.942, CAN.746, CAN.1319 d. Soit n = 8 g et 10 d.P2/ - ensemble : CAN.705, CAN.1274, CAN.187, CAN.1270, CAN.277, CAN.790, CAN.1348, NEA.06 g ; CAN.392, CAN.1703, CAN.842, CAN.1274, CAN.1271, CAN.171, CAN.273 d. Dents isolées : CAN.749, CAN.1610, CAN.1179, CAN.1645, CAN.1926, CAN.135, CAN.321, CAN.77, CAN.331, CAN.207, CAN.160, CAN.1874, CAN.1666 g ; CAN.1863, CAN.885, CAN.748, CAN.73, CAN.911, CAN.609 d. Soit : n = 21 g et 13 d.P3/ - ensemble : CAN.1793, CAN.63, CAN.490, CAN.1270, CAN.165, CAN.1533, CAN.726, CAN.705, CAN.392, CAN.662, CAN.1703, CAN.192 g ; CAN.1271, CAN.273, CAN.1793, CAN.171, CAN.1605, CAN.842, CAN.726, CAN.864, CAN.740, CAN.705, CAN.1703, CAN.504, CAN.68, CAN.921, CAN.1445 d.. Dents isolées : NEA.07, NEA.08 g ; CAN.1106, CAN.332, CAN.800 d. Soit n = 14 g et 18 d.P4/ - ensemble : CAN.1703, CAN.705, CAN.620, CAN.726, CAN.790, CAN.1794, CAN.273, CAN.165, CAN.791, CAN.1274, CAN.1926, CAN.1882, CAN.632, CAN.1271, CAN.1793, CAN.1824, CAN.1270, NEA.05 g ; CAN.392, CAN.1703, CAN.378, CAN.705, CAN.842, CAN.740, CAN.662, CAN.1518, CAN.563, CAN.1757, CAN.922, CAN.1223, CAN.1882, CAN.1084, CAN.726, CAN.620, CAN.384, CAN.273, CAN.1844, CAN.1926, CAN.658, CAN.1793, CAN.1274, NEA.07 d. Dents isolées : CAN.869 g ; CAN.865 d. Soit n = 19 et 25 d.M1/ - ensemble : CAN.631, CAN.392, CAN.662, CAN.273, CAN.1703, CAN.705, CAN.620, CAN.926, CAN.1882, CAN.1926, CAN.1084, CAN.165, CAN.790, CAN.1793, CAN.1270, CAN.791, CAN.1792, CAN.1274, CAN.107, NEA.05 g ; CAN.1703, CAN.705, CAN.726, CAN.620,
Planche VI
Fig. 1: Romain-la-Roche, Panthera spelaea, crâne complet FEL. 03, individu n°3, vue latérale droite.Fig. 2: Romain-la-Roche, Panthera spelaea, crâne complet FEL. 03, individu n°3, vue inférieure.Fig. 3: Romain-la-Roche, Panthera spelaea, demi-mandibule droite FEL. 04, individu n°2, vue latérale externe.

Planche VI
1
2
3
1 cm

592 A. ARgANT
CAN.378, CAN.864, CAN.740, CAN.1084, CAN.1793, CAN.922, CAN.1223, CAN.273, CAN.1271, CAN.563, CAN.662, CAN.337, CAN.1274, CAN.1757, NEA.08 d. Dents isolées : CAN.1141, CAN.58, CAN.1888, CAN.1833 g ; CAN.237, CAN.660, CAN.75, CAN.518 d. Fragments : CAN.114+97, CAN.166+115, CAN.92, CAN.122, CAN.121, CAN.52 g ; CAN.1864, CAN.162, CAN.117, CAN.871 d. Soit n = 30 g et 27 d.M2/ - ensemble : CAN.1703, CAN.620, CAN.726, CAN.790, CAN.1274, CAN.273, CAN.165, CAN.1882, CAN.631, CAN.1793, CAN.1270, CAN.1084, NEA.05 g ; CAN.392, CAN.378, CAN.705, CAN.740, CAN.662, CAN.791, CAN.563, CAN.1757, CAN.1223, CAN.1882, CAN.1084, CAN.726, CAN.620, CAN.1605, CAN.106, NEA.07 d. Dents isolées : CAN.1793, CAN.864, CAN.392, CAN.98, CAN.1899, CAN.66, CAN.1889 g ; CAN.47, CAN.565, CAN.659, CAN.74, CAN.649, CAN.1334 d. Soit n = 20 g et 22 d.Demi-mandibules :g : CAN.1187, CAN.1072, CAN.492, CAN.1642, CAN.1624, CAN.269, CAN.645, CAN.270, CAN.419, CAN.1515, CAN.1784, CAN.1313, CAN.1317, CAN.1196, CAN.10, CAN.898, CAN.1608, CAN.1145, CAN.1919, CAN.1486, CAN.1629. Soit n = 21.d : CAN.652, CAN.455, CAN.493, CAN.1318, CAN.897, CAN.1362, CAN.1467, CAN.704, CAN.1119, CAN.1622, CAN.1193, CAN.688, CAN.704, CAN.251, CAN.822, CAN.168, CAN.1524, CAN.310, CAN.11, CAN.1309, CAN.1614, CAN.850, CAN.813, CAN.1069, CAN.1486. Soit n = 25.Denture inférieure :I/1 - ensemble : CAN.1317 g ; CAN.1318 d. Dents isolées : CAN.679, CAN.606, CAN.1507, CAN.1415, CAN.1650, CAN.659, CAN.425, CAN.1544, CAN.907 g ; CAN.935, CAN.1640, CAN.1413, CAN.1623, CAN.1700, CAN.995 d. Soit n = 10 g et 7 d.I /2 - ensemble : CAN.492, CAN.645, CAN.1317 g ; CAN.1318, CAN.684 d. Dents isolées : CAN.665, CAN.1541, CAN.489, CAN.653, CAN.1618 g ; CAN.487, CAN.605, CAN.1803, CAN.1236, CAN.1547, CAN.172, CAN.1508, CAN.1617, CAN.655, CAN.1847 d. Soit n = 8 g et 12 d.I/3 - ensemble : CAN.1624, CAN.645, CAN.1317 g ; CAN.1318, CAN.704, CAN.1622, CAN.684 d. Dents isolées : CAN.1489, CAN.1580, CAN.603, CAN.777, CAN.224, CAN.1533, CAN.903, CAN.1918, CAN.372, CAN.104, CAN.1502, CAN.423, CAN.543, CAN.772, CAN.701, CAN.654, CAN.832 g ; CAN.1612, CAN.1068, CAN.900, CAN.170, CAN.1702, CAN.1357, CAN.325, CAN.1208, CAN.604, CAN.317, CAN.1461, 30 d. Soit n = 20 g et 16 d.C inf. – ensemble : CAN.1072, CAN.1624, CAN.645, CAN.1515, CAN.1317, CAN.1486 g ; CAN.455, CAN.1318, CAN.897, CAN.1622, CAN.934, CAN.1069 d. Dents isolées : CAN.1166, CAN.601, CAN.1564, CAN.1501, CAN.699, CAN.1125, CAN.420, CAN.814, CAN.851, CAN.676,
CAN.269, CAN.200, CAN.899, CAN.78, CAN.327, CAN.1534, CAN.55, CAN.1636, CAN.1834, CAN.560, CAN.269, NEA.10 g ; CAN.766, CAN.1904, CAN.158, CAN.538, CAN.828, CAN.684, CAN.1783, CAN.317, CAN.1240, CAN.1686, CAN.81, CAN.25, CAN.1235, CAN.1526, CAN.1202, CAN.817, CAN.1635, CAN.111, CAN.1359, CAN.62, CAN.112, CAN.79, CAN.159, CAN.1187 d. Soit : n = 28 g et 30 d.P/1 – ensemble : CAN.1187, CAN.492, CAN.1624, CAN.645, CAN.270, CAN.419, CAN.1317, CAN.1609, CAN.1486 g ; CAN.652, CAN.455, CAN.493, CAN.1193, CAN.1528, CAN.269, CAN.1486 d. Dents isolées : CAN.1550, CAN.414, CAN.902, CAN.1060, NEA.09 g ; CAN.50, CAN.904, CAN.91 d. Soit n = 14 g et 10 d.P/2 – ensemble : CAN.1187, CAN.1072, CAN.492, CAN.1642, CAN.1624, CAN.645, CAN.270, CAN.419, CAN.21, CAN.1532, CAN.1317, CAN.1196, CAN.898, CAN.1609, CAN.1486, CAN.846 g ; CAN.652, CAN.455, CAN.493, CAN.1318, CAN.1467, CAN.704, CAN.1119, CAN.1622, CAN.1193, CAN.704, CAN.251, CAN.310, CAN.1309, CAN.850, CAN.1528, CAN.269, CAN.1069, CAN.1486 d. Dents isolées : CAN.936, CAN.681, CAN.1781 g ; CAN.1869, CAN.1494, CAN.809, CAN.1462, CAN.1535 d. Soit n = 16 g et 23 d.P/3 – ensemble : CAN.1187, CAN.1072, CAN.492, CAN.1624, CAN.269, CAN.645, CAN.270, CAN.419, CAN.94, CAN.1317, CAN.1196, CAN.898, CAN.1436 g ; CAN.652, CAN.455, CAN.493, CAN.1318, CAN.1467, CAN.704, CAN.1119, CAN.1622, CAN.1193, CAN.704, CAN.251, CAN.822, CAN.310, CAN.1309, CAN.1069, CAN.1486 d. Dents isolées : CAN.10, CAN.319, CAN.1447, CAN.56, CAN.1577, CAN.205 d. Soit n = 13 g et 22 d.P/4 – ensemble : CAN.1187, CAN.492, CAN.1642, CAN.1624, CAN.269, CAN.645, CAN.270, CAN.419, CAN.1515, CAN.1815, CAN.1525, CAN.1317, CAN.1196, CAN.10, CAN.898, CAN.1609, CAN.1486, CAN.1145, CAN.1441 g ; CAN.652, CAN.455, CAN.493, CAN.1318, CAN.897, CAN.1072, CAN.704, CAN.1119, CAN.1622, CAN.1193, CAN.688, CAN.704, CAN.1362, CAN.251, CAN.822, CAN.168, CAN.310, CAN.1309, CAN.850, CAN.14, CAN.1873, CAN.1069 d. Dents isolées : CAN.68, CAN.89, CAN.806 g ; CAN.812, CAN.1492, CAN.848, CAN.215 d. Soit n = 22 g et 26 d. M/1 – ensemble : CAN.419, CAN.221, CAN.1515, CAN.9, CAN.645, CAN.916, CAN.270, CAN.1525, CAN.492, CAN.269, CAN.1187, CAN.1072, CAN.1624, CAN.898, CAN.1642, CAN.1609, CAN.1317, CAN.1441, CAN.1145, CAN.10, CAN.1196 g ; CAN.1622, CAN.1119, CAN.850, CAN.1614, CAN.1309, CAN.1193, CAN.897, CAN.1069, CAN.1629, CAN.310, CAN.1467, CAN.652, CAN.1318, CAN.493, CAN.704, CAN.455, CAN.688, CAN.1524, CAN.813, CAN.1362, CAN.822, CAN.1530, CAN.168, CAN.11 d. Dents isolées : CAN.1776, CAN.1241, CAN.800,
Planche VII
Fig. 1: Romain-la-Roche, Panthera spelaea, omoplate droite FEL. 200, vue externe.Fig. 2: Romain-la-Roche, Panthera spelaea, ensemble Mtc.2 FEL. 100, Mtc.3 FEL. 101, Mtc.4 FEL.101, Mtc.5 FEL.101
gauches d’une même main, vue dorsale.

Planche VII
2
11 cm
1 cm

594 A. ARgANT
CAN.677 g ; CAN.80, CAN.315, CAN.134, CAN.834, CAN.40 d. Fragments : CAN.336, CAN.829 g ; CAN.126, CAN.251 d. Soit n = 27 g et 31 d.M/2 – ensemble : CAN.1187, CAN.492, CAN.1642, CAN.1624, CAN.1559, CAN.270, CAN.419, CAN.1515, CAN.1784, CAN.9, CAN.221, CAN.798, CAN.93, CAN.1317, CAN.10, CAN.1609, CAN.1919 g ; CAN.652, CAN.455, CAN.493, CAN.1318, CAN.897, CAN.1467, CAN.704, CAN.1119, CAN.1622, CAN.1193, CAN.688, CAN.704, CAN.1629, CAN.1362, CAN.269, CAN.1524, CAN.310, CAN.11, CAN.1309, CAN.1098, CAN.930, CAN.1069, CAN.1486 d. Dents isolées : CAN.1207, CAN.808, CAN.1077, CAN.908 g ; CAN.803, CAN.1553, CAN.127, CAN.1561, CAN.117 d. Soit n = 21 g et 28 d.M/3 – ensemble : CAN.1515, CAN.1317, CAN.10 g ; CAN.897 d. Dents isolées : CAN.905, CAN.1543 g ; CAN.1496, CAN.1504 d. Soit n = 5 g et 3 d.Squelette post-crânien :Colonne vertébrale :vertèbres cervicales :Atlas : CAN.1117, CAN.982, CAN.1809, CAN.938, CAN.490Axis : CAN.539, CAN.690, CAN.530, CAN.526, CAN.451, CAN.1810, CAN.954, CAN.622, CAN.1327, CAN.1621, CAN.1115, CAN.1026, CAN.1051, CAN.687, CAN.1620, CAN.1727Autres vertèbres cervicales : CAN.584, CAN.891, CAN.1730, CAN.856, CAN.1915, CAN.967, CAN.720, CAN.1582, CAN.1476, CAN.477, CAN.1464, CAN.1725, CAN.1175, CAN.1281 CAN.1513, CAN.483, CAN.1906, CAN.480, CAN.1499, CAN.418, CAN.1328, CAN.421 non situéesv. cerv. 3 : CAN.451, CAN.1053, CAN.1726v. cerv. 4 : CAN.486, CAN.1120, CAN.1107v. cerv. 5 : CAN.1118v. cerv. 6 : CAN.572, CAN.1102v. cerv. 7 : CAN.1121vertèbres dorsales : CAN., CAN., CAN., CAN., CAN., CAN., CAN., v. dors. 1 : CAN.1451, CAN.383, CAN.1113, CAN.1585v. dors. 3 : CAN.769, CAN.1777, CAN.1683, CAN.1342, CAN.1103v. dors. 4 : CAN.1491v. dors. 5 : CAN.1778, CAN.1257, CAN.1304v. dors. 6 : CAN.577, CAN.1756, CAN.1754, CAN.1343, CAN.1253v. dors. 7 : CAN.1216v. dors. 8 : CAN.522v. dors. 10 : CAN.1101, CAN.756
v. dors. 11 : CAN.1104, CAN.1256, CAN.1122v. dors. 12 : CAN.382, CAN.1295, CAN.1112, CAN.1116v. dors. 13 : CAN.1364vertèbres lombaires :v. lomb. 1 : CAN.593, CAN.401, CAN.559, CAN.1474v. lomb. 2 : CAN.1123, CAN.524, CAN.1299, CAN.1238v. lomb. 3 : CAN.765, CAN.1185, CAN.1258, CAN.591, CAN.1286, CAN.400, CAN.1290v. lomb. 4 : CAN.514, CAN.1188, CAN.970, CAN.1192, CAN.1691, CAN.1114, CAN.443, CAN.536, CAN.396, CAN.729, CAN.1287, CAN.483, CAN.1699, CAN.788v. lomb. 5 : CAN.889, CAN.496, CAN.1291, CAN.592v. lomb. 6 : CAN.890, CAN.1288, CAN.1066, CAN.510, CAN.1315, CAN.1750, CAN.1930v. lomb. 7 : CAN.1289, CAN.509, CAN.13, CAN.994, CAN.953, CAN.1955, CAN.1168, CAN.398, CAN.698sacrum : CAN.1149, CAN.739, CAN.1225vertèbres caudales : CAN.1284, CAN.1224, CAN.682, CAN.1226, CAN.1477, CAN.1219, CAN.506, CAN.776, CAN.353, CAN.1230, CAN.1222, CAN.575, CAN.784Sternèbre : CAN.552Ceintures scapulaire et pelvienne :Omoplate :g : CAN.1606, CAN.959, CAN.16, CAN.1048, CAN.1042, CAN.1033, CAN.1912, CAN.1082, CAN.1010 ; soit n = 9.d : CAN.717, CAN.1599, CAN.1465, CAN.1583, CAN.1751, CAN.1911, CAN.985, CAN.18, NEA.19 ; soit n = 8.Coxal :g : CAN.1285, CAN.858, CAN.377, CAN.363, CAN.1682, CAN.1146, CAN.1186, CAN.453, CAN.307, CAN.1269, CAN.1267, CAN.537 ; soit n = 12d : CAN.379, CAN.1242, CAN.371, CAN.858, CAN.537, CAN.377, CAN.1718+439, CAN.313, CAN.778, CAN.1276, CAN.1711, CAN.494, CAN.1771, CAN.1283, CAN.440, CAN.453, CAN.698 ; soit n= 17.Humérus :Complets – CAN.435, 515 g ; CAN.368, 485 d.Fragments – CAN.186, CAN.1514, CAN.887, CAN.616, CAN.252, CAN.1514, CAN.231, CAN.1658, CAN.958, CAN.184, CAN.429, CAN.1741, CAN.1765, CAN.1013, CAN.1308, CAN.472, CAN.185, CAN.1885, CAN.1814, CAN.211, CAN.724, CAN.450, CAN.364, CAN.339, CAN.361 g ; CAN.299, CAN.663, CAN.230, CAN.1519, CAN.1276, CAN.365, CAN.489, CAN.987, CAN.1648, CAN.465, CAN.613, CAN.234, CAN.260, CAN.339, CAN.1273, CAN.1300, CAN.663, CAN.1825, CAN.1818, CAN.253, CAN.1884, CAN.579, CAN.747, CAN.338, CAN.1879,
Planche VIII
Fig. 1: Romain-la-Roche, Canis lupus, fémur droit CAN. 1197, traces anthropiques de dépeçage sur l’extrémité distale, vue latérale interne.
Fig. 2: Romain-la-Roche, Ursus spelaeus, fragment proximal de cubitus gauche URS. 1457, vue latérale interne, « pit » circulaire (diamètre 11 mm).
Fig. 3: Romain-la-Roche, Canis lupus, extrémité proximale de fémur droit CAN. 1628, traces de rognage et « pit » circulaire (dimension 5 x 6 mm).
Fig. 4: Romain-la-Roche, Canis lupus, 5e vertèbre lombaire CAN. 889, vue latérale gauche, « puncture » allongée.Fig. 5: Romain-la-Roche, Canis lupus, 5e vertèbre lombaire CAN. 889, vue latérale droite, « puncture » circulaire
(diamètre 6,2 mm) avec esquilles osseuses en place.

Planche VIII
1
2
3
4
5
1 cm
1 cm
1 cm
1 cm

596 A. ARgANT
CAN.284, CAN.978, CAN.1039, CAN.1024, CAN.361 d. Soit n = 27 g et 32 d.Radius :Complets – CAN.540 g ; CAN.1185, CAN.641 d.Fragments – CAN.386, CAN.1189, CAN.174, CAN.785, CAN.845, CAN.1030, CAN.1685, CAN.1724, CAN.399, CAN.833, CAN.860, CAN.217, CAN.1676, CAN.302, CAN.386, CAN.1698, CAN.1221, CAN.531, NEA.11, NEA.15 g ; CAN.1001, CAN.1641, CAN.176, CAN.178, CAN.586, CAN.963, CAN.1453, CAN.997, CAN.1842, CAN.520, CAN.274, CAN.650, CAN.436, CAN.638, CAN.244, CAN.1292, CAN.1241, CAN.1452, CAN.298, CAN.965, CAN.175, CAN.173, CAN.669, CAN.1767, CAN.274, CAN.309, CAN.172, CAN.860, CAN.1742, NEA.12, NEA.13, NEA.16, NEA.17 d. Soit n = 21 g et 35 d.Cubitus Complets – CAN.1158 g ; CAN.464, CAN.725 d.Fragments – CAN.397, CAN.1190, CAN.1031, CAN.197, CAN.229, CAN.664, CAN.1577, CAN.1261, CAN.974, CAN.767, CAN.755, CAN.996, CAN.479, CAN.362, CAN.1591, CAN.22, CAN.442, CAN.1600, CAN.245, CAN.1710, CAN.1397, NEA.18 g ; CAN.1247, CAN.840, CAN.1237, CAN.216, CAN.12, CAN.250, CAN.212, CAN.290, CAN.30, CAN.358, CAN.220, CAN.668, CAN.1330, CAN.311, CAN.280, CAN.222, CAN.1593, CAN.977, CAN.1040, CAN.1758, CAN.1817, CAN.263, CAN.977, CAN.859, CAN.289 d. Soit n = 23 g et 27 d.Carpe :Scapholunaire – CAN.1719, CAN.1630, CAN.463 g ; CAN.1371, CAN.1772 d. Soit n = 3 g et 2 d.Pyramidal – CAN.1789, CAN.1806, CAN.1143 d. Soit n = 3 d.Unciforme – CAN.670, CAN.1867, CAN.457 g ; CAN.1721 d. Soit n = 3 g et 1 d.Magnum – CAN.1716 d. Soit n = 1 d.Métacarpiens :Mtc.1 – CAN.1403, CAN.1798, CAN.1712 g ; CAN.1717, CAN.1421, CAN.973 d. Soit n = 3 g et 3 d.Mtc.2 – CAN.656, CAN.404, CAN.1701, CAN.1681, CAN.1312, CAN.37 g ; CAN.1382, CAN.707, CAN.640, CAN.1597, CAN.308, CAN.893, CAN.1749, CAN.246 d. Soit n = 6 g et 8 d.Mtc.3 – CAN.444, CAN.1638, CAN.1293, CAN.1748, CAN.694, CAN.1803, CAN.422, CAN.1872 g ; CAN.1365, CAN.1651, CAN.388, CAN.431, CAN.41, CAN.1307 d. Soit n = 8 g et 6 d.Mtc.4 – CAN.1218, CAN.1652, CAN.1487, CAN.24, CAN.529, CAN.666, CAN.990, CAN.1450 g ; CAN.293, CAN.1851, CAN.1381, CAN.1375, CAN.569 d. Soit n = 8 g et 5d.Mtc.5 – CAN.639, CAN.1632, CAN.31, CAN.1376, CAN.948, CAN.1770, CAN.1702, CAN.1598, CAN.248 g ; CAN.355, CAN.718, CAN.2170, CAN.1746, CAN.564, CAN.1390, CAN.1827, CAN.140, CAN.255, CAN.295 d. Soit n = 9 g et 10 d.Fémur :Complets – CAN.508, CAN.1647 g ; CAN.770, CAN.1197, CAN.1628 d.Partiels – CAN.265, CAN.1167, CAN.1604, CAN.387, CAN.239, CAN.281, CAN.236, CAN.32, CAN.265, CAN.1816, CAN.233, CAN.1058, CAN.1183, CAN.1065, CAN.8, CAN.1440, CAN.295, CAN.272, CAN.301, CAN.1012, CAN.1156, CAN.278, NEA.20 g ; CAN.1689, CAN.981, CAN.1086, CAN.932, CAN.271, CAN.1169, CAN.1801, CAN.275, CAN.757, CAN.232, CAN.356,
CAN.223, CAN.238, CAN.1063, CAN.1265, CAN.441, CAN.1248, 5 d. Soit n = 25 g et 21 d.Tibia :Complets – CAN.395, CAN.430, CAN.595, CAN.1927 g ; CAN.553, CAN.507, CAN.535, CAN.576, CAN.982 d.Partiels – CAN.934, CAN.267, CAN.277, CAN.279, CAN.618, CAN.275, CAN.956, CAN.1891, CAN.988, CAN.570, CAN.1714, CAN.182, CAN.213, CAN.929, CAN.267, CAN.362, CAN.1081, CAN.180, CAN.181, CAN.1846, CAN.1581, CAN.1036, CAN.277, CAN.456, NEA.01, NEA.02 g ; CAN.369, CAN.1244, CAN.1657, CAN.268, CAN.589, CAN.303, CAN.1044, CAN.1050, CAN.283, CAN.1607, CAN.1280, CAN.983, CAN.779, CAN.1025, CAN.1607, CAN.1720, CAN.357, CAN.1264, CAN.437, CAN.1004, CAN.979, CAN.1050 d, soit n = 30 g et 27 d.Péroné :CAN. 1355, CAN.38 d, soit n = 2 d.Tarse : Pemac 75Calcanéum complets : CAN.980, CAN.422, CAN.254, CAN.1043 g ; CAN.1294, CAN.1307, CAN.594, CAN.1890, CAN.780, CAN.1064, CAN.714, CAN.1448 d.partiels : CAN.1797, CAN.647, CAN.19, CAN.1080, CAN.1738 g ; CAN.1860 d, soit n = 9 g et 9 d.Astragale complet : CAN.405, CAN.1344, CAN.984 g ; CAN.1294, CAN.1164, CAN.1063, CAN.1296, CAN.1133, CAN.962 d.partiels : CAN.1006, CAN.1017, CAN.1726, CAN.975 g ; CAN.754, CAN.1029, CAN.1032, CAN.33 d, soit n = 7 g et 10 d.CuboïdeCAN.550, CAN.1520, CAN.1183, CAN.1385 g ; CAN.1373, CAN.1661, CAN.694 d, soit n = 4 g et 3 d.NaviculaireCAN.459, CAN.1090 g, soit 2 g.3° cunéïformeCAN.1336 g, soit n = 1 g.Métatarsiens :Mtt.2 – CAN.589, CAN.1217, CAN.1660, CAN.366, CAN.1165, CAN.1398, CAN.1272, CAN.349, CAN.1446, CAN.360, CAN.1835, CAN.1272, CAN.1384 g ; CAN. 209, CAN.1510, CAN.567, CAN.1795, CAN.1670 d, soit 13 g et 5 d.Mtt.3 – CAN.1366, CAN.1089, CAN.1509, CAN.745, CAN.1616, CAN.247, CAN.861, CAN.992, CAN.391, CAN.259, CAN.1643 g ; CAN.1194, CAN.1130, CAN.1684, CAN.226, CAN.347, CAN.741, CAN.315, CAN.1239 d, soit n = 11 g et 8 d.Mtt.4 – CAN.585, CAN.1368, CAN.554, CAN.1138, CAN.1823 g ; CAN.288, CAN.192, CAN.610, CAN.350, CAN.1399, CAN.1512 d, soit n = 5 g et 6d.Mtt.5 – CAN.1181, CAN.1653, CAN.312, CAN.296, CAN.1386, CAN.1383 g ; CAN.1703, CAN.555, CAN.1401, CAN.191, CAN.1378 d, soit n = 6 g et 5 d.Phalanges :Phalange 1 :Phalanges 1 antérieures mesurées :Doigt 2 – CAN.562, CAN.512 g ; CAN.142, CAN.710, CAN.42, CAN.35, CAN.340 d.Doigt 3 – CAN.511, CAN.445, CAN.1466, CAN.588 g.Doigt 4 – CAN.521 ; CAN.1184 d.Doigt 5 – CAN.461, CAN.769, CAN.484 g ; CAN.145, CAN.194, CAN.993, CAN.671 d.

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 597
Phalanges 1 postérieures mesurées :Doigt 2 – CAN.375, CAN.1093, CAN.1654, CAN.541, CAN.1413, CAN.1922 g.Doigt 3 – CAN.527, CAN.1611, CAN.208, CAN.39 g ; CAN.719 d.Doigt 4 – CAN.630, CAN.925, CAN.551, CAN.346, CAN.96, CAN.1400 g ; CAN.566, CAN.82, CAN.1137, CAN.1385 d.Doigt 5 – CAN.1094, CAN.1370, CAN.1170, CAN.1357 g ; CAN.843 d.Phalanges 1 indéterminées non mesurées : CAN.545, CAN.1254, CAN.1231, CAN.227, CAN.151, CAN.141, CAN.1303, CAN.1406, CAN.858, CAN.389, CAN.1388, CAN.1662, CAN.546, CAN.578, CAN.543, CAN.1762, CAN.1391, CAN.256, CAN.986, CAN.214, CAN.1868, CAN.1387, CAN.1923, CAN.446, CAN.1203, CAN.249, CAN.1667, CAN.1420, CAN.1352, CAN.1396, CAN.1468, CAN.1374, CAN.1729, CAN.1418, CAN.945, CAN.1433.Phalange 2 :Phalanges 2 antérieures mesurées : CAN.1041, CAN.1410, CAN.481, CAN.1723 g ; CAN.731, CAN.1153, CAN.345, CAN.1405, CAN.1672, CAN.498 d.Phalanges 2 postérieures mesurées : CAN.1631, CAN.1377, CAN.458, CAN.1229, CAN.1129, CAN.1216 g ; CAN.190, CAN.1404, CAN.1678, CAN.1356, CAN.1392, CAN.448, CAN.1212 d.Phalanges 2 indéterminées non mesurées : Antérieures – CAN.1411, CAN.972, CAN.1417, CAN.884, CAN.1409, CAN.547, CAN.1414, CAN.820.Postérieures – CAN.557, CAN.1901, CAN.1097, CAN.1454, CAN.1861, CAN.977, CAN.1395, CAN.219, CAN.976, CAN.1854, CAN.464, CAN.1924, CAN.1415, CAN.1857, CAN.1056, CAN.1096, CAN.730.Phalange 3 :Doigt 1 : CAN.1425 g ; CAN.218, CAN.1663, CAN.462, d.Phalanges 3 indéterminées mesurées : CAN.1488, CAN.1669, CAN.804, CAN.1061, CAN.468, CAN.1665, CAN.694, CAN.768, CAN.1204, CAN.598.Phalanges 3 indéterminées non mesurées : CAN.201, CAN.1423, CAN.1855, CAN.500, CAN.556, CAN.501, CAN.608, CAN.1408, CAN.1412, CAN.928, CAN.1298, CAN.412, CAN.1424, CAN.153, CAN.1856, CAN.1233, CAN.1163, CAN.462, CAN.1407, CAN.1426.Sésamoïdes : CAN.460, CAN.1430, CAN.1432, CAN.1534, CAN.1425, CAN.1431.
VI.4. CAN.4 : Cuon alpinus (NR = 13, NMI = 1)
Demi-mandibule :- CAN.1069 – demi-mandibule d. Ceintures :- coxal – CAN.495 g.Squelette post-crânien :Colonne vertébrale : axis fragment CAN.470.Membres :- cubitus : CAN.12 d.- fémur : CAN.508 g ; CAN.757 d.- carpe : magnum , CAN.1432 d.- tibia : CAN.595 g ; CAN.507 d.- tarse : astragale, CAN.596 g ; CAN.1095 d.- phalange 1 : CAN.1469, CAN.1413.
VI.5. URS.1 – Ursus arctos (NR = 19, NMIE = 2)
Denture supérieure :P3/ : dents isolées URS.1740 g ; URS.1743 dDemi mandibule : URS.1058 g complèteDenture inférieure : dents isoléesP/1 : URS.1741 g ; P/3 : URS.1742 g ; P/4 : URS.1743 dPost-crânien :- métacarpiens : Mtc.1 : URS.629 g ; Mtc.2 : URS.1384 g ;
Mtc.5 : URS.658 d- tibia : URS.1187 g- métatarsiens : Mtt.1 – URS.1422 d, Mtt.3 – URS.1072 g- phalange 1 : URS.1353- phalanges 3 : URS.355, URS.518, URS.632, URS.844,
URS.850, URS.1351.
URS.2 – Ursus spelaeus (NR = 1313, NMI = 20)Crânes : URS.001, URS.157, URS.158, URS.159, URS.160, URS.409, URS.972, URS.973, URS.1717, URS.1194, URS. 247, URS.339, URS.404, URS.1224 , URS.119, URS.681, URS.1438.Denture supérieure :Incisives :I1/ :Ensembles : URS.398, URS.106, URS.528, URS.533, URS.758 g ; URS.656, URS.634 d.Dents isolées : URS.1011, URS.1451 d.I2/ : Dents isolées : URS.1709, URS.1718 g ; URS.1192, URS.1490, URS.1769 d.I3/ : Ensembles : URS.112, URS.103, URS.825, URS.867 g ; URS.150, URS.27, URS.566, URS.432, URS.731, URS.826, URS.824, URS.823, URS.866 d.Dents isolées : URS.1013, URS.1168, URS.1373, URS.945, URS.934 g ; URS.1056, URS.1160, URS.1484, URS.208, URS.40 d.Canines supérieures :Ensembles : URS.339, URS.404, URS.1, URS.118, URS.157, URS.158, URS.18, URS.409, URS.973 g ; URS.404, URS.1, URS.156, URS.157, URS.160, URS.178, URS.191, URS.28, URS.409, URS.582, URS.973 d.Dents isolées : URS.321, URS.401, URS.105, URS.125, URS.148, URS.52, URS.62, URS.888, URS.886 g ; URS.402, URS.540, URS.677 d.Canine déciduale sup isolée : URS.1767 d. Prémolaires :P3/ : Alvéoles vides, ensembles : URS.973 g ; URS.681, URS.973, URS.1438 d.Dents isolées : URS.1744, URS.1745 g ; URS.1746, URS.1747 d.P4/ :Ensembles : URS.160, URS.22, URS.409, URS.973, URS.1194, URS.339, URS.1, URS.118, URS.158 g ; URS.1438, URS.160, URS.25, URS.409, URS.67, URS.681, URS.814, URS.1194, URS.339, URS.1, URS.156, URS.158 d.Dents isolées : URS.879, URS.1372, URS.1770, URS.54, URS.96, URS.400, URS.114, URS.115 g ; URS.1452, URS.55, URS.631, URS.789, URS.210, URS.216, URS.218, URS. 399 d.

598 A. ARgANT
Molaires :M1/ :Ensembles : URS.1717, URS.409, URS.67, URS.973, URS.1194, URS.1687, URS.339, URS.404, URS.413, URS.118, URS.158, URS.159 g ; URS.409 , URS.67, URS.973, URS.814, URS.1194, URS.1116, URS.1689, URS.1438, URS.339, URS.156, URS.158, URS.159 d.Dents isolées : URS.61, URS.70, URS.113, URS.116, URS.24, URS.574, URS.1442 g ; URS.442, URS.679, URS.214.M2/ :Ensembles : URS.973, URS.1194, URS.1687, URS.1717, URS.158, URS.159, URS.160, URS.409, URS.45, URS.67, URS.972, URS.339, URS.404, URS.413, URS.1, URS.118, URS.1194, URS.157 g ; URS.973, URS.814, URS.832, URS.1194, URS.1118, URS.1698, URS.1686, URS.1438, URS.159, URS.160, URS.409, URS.46, URS.67, URS.339, URS.404, URS.1, URS.1194, URS.157, URS.158 d.Dents isolées : URS.1733, URS.26, URS.59, URS.70, URS.151 g ; URS.680, URS.171, URS.64, URS.215, URS.209, URS.156 d.Demi-mandibules : URS.1154, URS.1147, URS.1134, URS.1027, URS.119, URS.142, URS.1439, URS.17, URS.173, URS.406, URS.444, URS.507, URS.472, URS.1025, URS.1102, URS.1085, URS.328, URS.344, URS.217, URS.782 g ; URS.1483, URS.1683, URS.1770, URS.1153, URS.138, URS.1439, URS.69, URS.480, URS.845, URS.995, URS.904, URS.272, URS.271, URS.423, URS.424 d.Denture inférieure :Incisives :I/1 :Ensembles : URS.588, URS.829 g ; URS.752, URS.830 d.Dents isolées : URS.1753, URS.1754 g ; URS.1338, URS.1342, URS.1755, URS.1757 d.I/2 :Ensembles : URS.567, URS.1154 g ; URS.541, URS.738 d.Dents isolées : URS.1760 g ; URS.1702, URS.1448, URS.1756, URS.1758, URS.1759 d.I/3 :Ensemble : URS.536 g.Dents isolées : URS.1108, URS.1366, URS.1367, URS.1771, URS.999, URS.925, URS.1009, URS.1062, URS.317, URS.360, URS.371, URS.134, URS.19, URS.537 g ; URS.1068, URS.1736, URS.763, URS.673, URS.831, URS.883, URS.263, URS.614 d.Canines :Ensembles : URS.782, URS.1439, URS.1025, URS.1102, URS.1085, URS.1147, URS.1134, URS.1154, URS.217, URS.344, URS.1027, URS.173 g ; URS.1683, URS.1730, URS.845, URS.1439, URS.904, URS.995, URS.1153, URS.1483, URS.272, URS.271, URS.138, URS.1439 d.Dents isolées : URS.1443, URS.1058, URS.1144, URS.283, URS.416, URS.406, URS.71, URS.787 g ; URS.1721, URS.1140, URS.213, URS.17, URS.668 d.Canine déciduale inf, isolée : URS.1768 d.Prémolaires : P/4Ensembles : URS.344, URS.1027, URS.1439, URS.406, URS.444, URS.1102, URS.1085, URS.1147, URS.1154 g ; URS.271, URS.138, URS.1439, URS.23, URS.480, URS.904, URS.995, URS.1153 d.Dents isolées : URS.211, URS 882, URS.1745, URS.1746, URS.1747, URS.1748 g ; URS.212, URS.1743, URS.1744 d.Molaires :M/1 :Ensembles : URS.344, URS.1027, URS.1439, URS.406,
URS.1025, URS.1058, URS.1085, URS.1147, URS.1134 g ; URS.207, URS.1439, URS.480, URS.845, URS.904, URS.995 d.Dents isolées : URS.282, URS.284, URS.396, URS.43, URS.939, URS.1165 g ; URS.153, URS.31, URS.546, URS.1164, URS.1365 d.M/2 :Ensembles : URS.1439, URS.1025, URS.1058, URS.1102, URS.1085, URS.1147, URS.1134, URS.1154, URS.217, URS.344, URS.1027, URS.173, URS.406, URS.444 g ; URS.845, URS.1439, URS.904, URS.1153, URS.1483, URS.1690, URS.207, URS.138 d.Dents isolées : URS.786, URS.369, URS.385, URS.13, URS.35 g ; URS.1691, URS.799, URS.313, URS.20, URS.37, URS.36 d.M/3 :Ensembles : URS.1102, URS.1085, URS.1147, URS.1134, URS.1154, URS.217, URS.344, URS.1027, URS.1439, URS.406 g ; URS.1153, URS.1483, URS.207, URS.138, URS.1439, URS.904, URS.995 d.Dents isolées : URS.1071, URS.1020, URS.285, URS.324, URS.174, URS.21 g ; URS.1447, URS.1731, URS.1368, URS.30 d.Squelette post-crânien :Colonne vertébrale :- Vertèbres cervicales :Atlas (V.C.1) : URS.336, URS.465, URS.49, URS.531, URS.720, URS.734, URS.874, URS.997, URS.920, URS.1171, URS.1196, URS.1711, URS.1668, URS.1772Axis (V.C.2) : URS.465, URS.858, URS.996, URS.1075, URS.1177, URS.1225, URS.1459Autres vertèbres cervicales : rang indéterminéURS.121, URS.465, URS.542, URS.593, URS.60, URS.624, URS.635, URS.448, URS.452, URS.641, URS.811, URS.838, URS.848, URS.1051, URS.1049, URS.1063, URS.1040, URS.1040, URS.1151, URS.456, URS.1332, URS.1603, URS.1656, URS.1548- Vertèbres dorsales : rang indéterminéURS.494, URS.467, URS.464, URS.642, URS.843, URS.853, URS.894, URS.876, URS.1070, URS.1099, URS.1407, URS.1662, URS.242, URS.239, URS.276, URS.287, URS.412, URS.122, URS.133, URS.186, URS.189, URS.270, URS.509, URS.544, URS.520, URS.625, URS.428, URS.451, URS.450, URS.1505, URS.1184, URS.1641, URS.1649, URS.1642, URS.1640, URS.1625, URS.1611, URS.1281, URS.1710, URS.1704, URS.1590, URS.1584, URS.1586, URS.1581, URS.1585, URS.1664, URS.1469, URS.1455, URS.1542, URS.1548, URS.1541, URS.1545, URS.1399, URS.1679, URS.1700, URS.1701- Vertèbres lombaires : rang indéterminéURS.139, URS.177, URS.516, URS.551, URS.456, URS.506, URS.465, URS.896, URS.1068, URS.1115, URS.1040, URS.1042, URS.1190, URS.1130, URS.1123, URS.1157, URS.344, URS.1189, URS.1626, URS.1201- Sacrum : URS.258, URS.863, URS.1076, URS.1129- Vertèbres caudales : rang 1, URS.1773Ceintures :Omoplate :complètes : URS.463 g ; URS.1494, URS.278 d.partielles : URS.1670, URS.332, URS.341, URS.162, URS.184, URS.68, URS.785, URS.857, URS.860, URS.1047, URS.1041 g ; URS.1519, URS.246, URS.274, URS.176, URS.126, URS.1074 d.Coxaux :

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 599
partiels : URS.330, URS.127, URS.161, URS.258, URS.550, URS.872, URS.1117, URS.1198 g ; URS.1219, URS.335, URS.127, URS.258, URS.869, URS.864, URS.871, URS.998, URS.1106, URS.1198 d.Membres :humérus :complets : URS.1406, URS.462 g ; URS.1408, URS.128, URS.512 d.partiels : URS.517, URS.447, URS.499, URS.841, URS.820, URS.889, URS.1345, URS.1595, URS.1227, URS.1209, URS.1583, URS.1266, URS.1466, URS.224, URS.248, URS.303, URS.345, URS.120, URS.197, URS.407 g ; URS.558, URS.1138, URS.1178, URS.1633, URS.1413, URS.1391, URS.227, URS.294, URS.318, URS.107, URS.188, URS.487 d.radius :complets : URS.190, URS.479, URS.493 g ; URS.1181 d.partiels : URS.1082, URS.1084, URS.1509, URS.204, URS.273, URS.420, URS.202, URS.562, URS.79, URS.852, URS.906 g ; URS.1097, URS.1410, URS.1440, URS.1720, URS.329, URS.102, URS.591, URS.598, URS.426 d.cubitus :complets : URS.1390, URS.201, URS.405 g ; URS.445, URS.1346 d.partiels : URS.219, URS.327, URS.137, URS.192, URS.198, URS.523, URS.581, URS.599, URS.65, URS.425, URS.784, URS.819, URS.1457, URS.1456, URS.331 g ; URS.286, URS.337, URS.338, URS.408, URS.130, URS.172, URS.48, URS.74, URS.9, URS.504, URS.915, URS.900, URS.1492, URS.487, URS.427 d.Carpe :Scapholunaire :complets : URS.772, URS.1069, URS.1399 g ; URS.220, URS.267, URS.95, URS.485, URS.885 d.partiels : URS.8, URS.1216 g ; URS.1203 d.Pyramidal : complets : URS.630, URS.640, URS.647, URS.775, URS.921 g ; URS.1279, URS.1376 d.partiels : URS.569, URS.1245 g ; URS.152, URS.980, URS.1680 d.Pisiforme :complets : URS.94 g ; URS.311, URS.979 d.partiels : URS.988, URS.1146, URS.545 g ; URS.880, URS.1681 d.Unciforme : complets : URS.616, URS.639, URS.833 g ; URS.974, URS.1591, URS.1377 d.partiel : URS.1513 g.Magnum :complets : URS.773, URS.1515, URS.1635, URS.1577, URS.1252 g ; URS.539, URS.651, URS.983, URS.1655, URS.1378 d.Trapèzoïde :complets : URS.774, URS.1614, URS.1634 g ; URS.729, URS.1608, URS.1599, URS.1423 d.Trapèze :Complets : URS.383, URS.765, URS.926, URS.1328 g ; URS.737 d.Métacarpiens :Mtc.1 :complets : URS.75, URS.76, URS.646, URS.743, URS.916, URS.1433 g ; URS.319, URS.377, URS.579, URS.745 d.partiels : URS.29, URS.1005 g ; URS.1355, URS.1697 d.
Mtc.2 : complets : URS.296, URS.623, URS.82, URS.645, URS.721, URS.849 g ; URS.530, URS.589, URS.437, URS.473, URS.891, URS.1421 d.partiels : URS.226, URS.104, URS.1694 g ; URS.627, URS.1037 d.Mtc.3 :complets : URS.291, URS.83, URS.726, URS.933 g ; URS.561, URS.436, URS.976 d.partiels : URS.238, URS.235, URS.1152 g ; URS.1374 d.Mtc.4 :complets :URS.14, URS.262, URS.572, URS.724, URS.1086 g ; URS.297, URS.607 , URS.443, URS.803, URS.975 d.partiels : URS.204, URS.80, URS.433, URS.1073, URS.1461 g ; URS.1420, URS.362, URS.1705 d.Mtc.5 :complets : URS.81, URS.722, URS.917, URS.1080 g ; URS.617, URS.978 d.partiels : URS.1386, URS.204, URS.237, URS.51, URS.790, URS.1287 g ; URS.1375, URS.325, URS.44, URS.475, URS.1012, URS.1107 d.fémur :complets : URS.1026, URS.124 g ; URS.1113, URS.129, URS.136, URS.166, URS.483 d.partiels : URS.1193, URS.1427, URS.1434, URS.1190, URS.333, URS.415, URS.1180, URS.132, URS.135, URS.169, URS.196, URS.600, URS.471 g ; URS.1148, URS.1453, URS.322, URS.334, URS.364, URS.131, URS.175, URS.266, URS.564, URS.595, URS.619, URS.66, URS.482, URS.818 d.rotule :complètes : URS.498, URS.875, URS.1081, URS.358, URS.92 g ; URS.486, URS.781, URS.293, URS.73 d.partielles : URS.1176, URS.389 g ; URS.1732, URS.675, URS.230, URS.676 d.tibia :complets : URS.340, URS.10, URS.1187, URS.478, URS.513 g ; URS.856, URS.472 d.partiels : URS.496, URS.837, URS.661, URS.890, URS.1031, URS.1471, URS.221, URS.244, URS.295, URS.193, URS.194, URS.58, URS.78 g ; URS.455, URS.902, URS.897, URS.908, URS.1149, URS.1137, URS.1183, URS.1197, URS.1552, URS.320, URS.108, URS.181, URS.182, URS.596 d.péroné :complets : URS.1369 g ; URS.1169 d.partiels : URS.1550, URS.374, URS.538, URS.990, URS.1007, URS.1511, URS.1161, URS.1212, URS.1204 g ; URS.1716, URS.801, URS.1006, URS.1521, URS.1363, URS.1337, URS.1275, URS.1205, URS.1212 d.Tarse :calcanéum :complets : URS.6 g ; URS.431, URS.501, URS.779, URS.798 d.partiels : URS.231, URS.683, URS.1573 g ; URS.620, URS.436, URS.966, URS.1014, URS.1600 d.astragale :complets : URS.240, URS.366, URS.807, URS.1185 g ; URS.222, URS.597, URS.438, URS.439, URS.884, URS.1002 d.partiels : URS.12, URS.145 g ; URS.110, URS.548, URS.1713 d.Cuboïde :complets : URS.7, URS.708, URS.1091, URS.1643, URS.1371 g ; URS.618, URS.691, URS.1638 d.partiels : URS.1606 g ; URS.149 d.

600 A. ARgANT
Naviculaire : complets : URS.693, URS.1383 g ; URS.578, URS.665, URS.712, URS.1004, URS.1032 d.partiels : URS. 1604+1607 g ; URS.1596, URS.474 d.Cunéiforme 1 :complets : URS.384, URS.1357, URS.1339 g ; URS.111, URS.898, URS.1516, URS.1343 d.partiel : URS.970 d.Cunéiforme 2 :complets : URS.1340, URS.1614 g ; URS.1631 d.Cunéiforme 3 :complets : URS.717, URS.1109, URS.1669 g ; URS.610, URS.1632, URS.1546 d.Métatarsiens :Mtt.1 :complets : URS.381, URS.602, URS.684 g ; URS.1003, URS.1429 d.partiels : URS.502, URS.938 g ; URS.969 d.Mtt.2 :complets : URS.387, URS.601, URS.1685, URS.1485 g ; URS.382, URS.964, URS.1018, URS.1170 d.partiels : URS.373, URS.1722 d.Mtt.3 :complets : URS.386, URS.615 g ; URS.15, URS.563, URS.585, URS.590, URS.952, URS.1015 d.partiels : URS.395, URS.265, URS.1320 g ; URS.1464 d.Mtt.4 :complets : URS.326, URS.375, URS.388, URS.571 g ; URS.622, URS.440, URS.662, URS.1008 d.partiels : URS.1382, URS.628, URS.718, URS.859, URS.1708 g ; URS.1463, URS.144, URS.16, URS.503, URS.778, URS.1265 d. Mtt.5 : complets : URS.368, URS.604, URS.1098 g ; URS.692 d.partiels : URS.307, URS.365, URS.397, URS.260, URS.793, URS.911 g ; URS.290, URS.347, URS.497, URS.977, URS.1035 d.Phalanges :phalange 1 : doigt indéterminéURS.223, URS.228,URS.227, URS.289, URS.292, URS.312, URS.379, URS.367, URS.411, URS.100, URS.117, URS.264, URS.268, URS.42,5 URS.19, URS.529, URS.53, URS.535, URS.547, URS.573, URS.575, URS.577, URS.586, URS.621, URS.613, URS.606, URS.435, URS.434, URS.429, URS.495, URS.650, URS.654, URS.655, URS.727, URS.744, URS.747, URS.723, URS.749, URS.689, URS.739, URS.695, URS.753, URS.704, URS.757, URS.752, URS.706, URS.690, URS.705, URS.697, URS.777, URS.804, URS.815, URS.428, URS.913, URS.894, URS.901, URS.887, URS.892, URS.865, URS.981, URS.984, URS.968, URS.936, URS.944, URS.1022, URS.1053, URS.1052, URS.1055, URS.1054, URS.1065, URS.1111, URS.1043, URS.1039, URS.1186, URS.1162, URS.1181, URS.1163, URS.1159, URS.1358, URS.1348, URS.1356, URS.1354, URS.1347, URS.1327, URS.1353, URS.1359, URS.1469, URS.1431, URS.1432, URS.1437, URS.1725, URS.1728.phalange 2 : doigt indéterminéURS.229, URS.316, URS.309, URS.352, URS.357, URS.461, URS.56, URS.576, URS.587, URS.592, URS.77, URS.85, URS.88, URS.91, URS.99, URS.470, URS.466, URS.746, URS.728, URS.742, URS.741, URS.703, URS.716, URS.696, URS.797, URS.847, URS.846, URS.910, URS.982, URS.1000, URS.932, URS.960, URS.961, URS.1048, URS.1087,
URS.1044, URS.1158, URS.1350, URS.1349, URS.1352, URS.1487, URS.1488, URS.1489, URS.1200, URS.1672, URS.1430, URS.1724, URS.1729, URS.1370.phalange 3 : doigt indéterminéURS.225, URS.288, URS.101, URS.527, URS.549, URS.568, URS.583, URS.584, URS.605, URS.84, URS.86, URS.89, URS.90, URS.94, URS.97, URS.948, URS.927, URS.1016, URS.1023, URS.1024, URS.1067, URS.1110, URS.1105, URS.1092, URS.1179, URS.1172, URS.1167, URS.1351, URS.1341, URS.1198, URS.1359, URS.1486, URS.1411, URS.1434, URS.1738.Sésamoïdes : doigt indéterminéURS.306, URS.376, URS.261, URS.748, URS.750, URS.769, URS.764, URS.759, URS.771, URS.733, URS.755, URS.760, URS.756, URS.761, URS.762, URS.768, URS.767, URS.732, URS.796, URS.795, URS.907, URS.1912, URS.985, URS.994, URS.987, URS.992, URS.965, URS.928, URS.923, URS.963, URS.1166, URS.1331, URS.1199, URS.1622, URS.1618, URS.1619, URS.1620, URS.1616, URS.1623, URS.1615, URS.1621, URS.1617, URS.1609, URS.1307, URS.1306, URS.1571, URS.1412, URS.1394.Côtes : rang indéterminéURS.250 d, URS.449 g, URS.458 d, URS.460 g, URS.543 d, URS.644 g, URS.648 g, URS.652 g, URS.862, URS.937, URS.1001, URS.1045 g, URS.1131, URS.1418, URS.1419, URS.1585… et une multitude de fragments indéterminables spécifiquement.Os hyoïde : URS.1547 (épihyal).
FEL.1 – Felis silvestris (NR = 10, NMI = 1)
Denture : - canine sup : FEL.226 g.Post-crânien :- radius : FEL.248 g endommagé.- métacarpien 2 : FEL.164 g endommagé.- tarse : calcanéum FEL.228 g complet, cuboïde FEL.160 g fragment, 3° cunéiforme FEL.238 g complet.- phalanges 1 : FEL.260, FEL.249 (phal. ant., doigt 4, d).- phalanges 2 : FEL.158, FEL.159 g.
FEL.2 – Panthera pardus (NR = 7, NMI = 1)
Post-crânien :- carpe : pisiforme FEL.252 g complet.- tibia : FEL251 diaphyse d.- tarse : astragale FEL250 d complet.- métatarsien 4 : FEL.227 g moitié prox.- phalanges 1 : FEL.254 (post., doigt 5, d), FEL.255 (post.,
doigt 3, d) entières.- phalange 2 : FEL.41 d entière.
FEL.3 – Panthera spelaea :- 3 crânes : FEL.1, FEL.2, FEL.3- 1 mandibule complète g et d (FEL.4), 3 demi-mandibules g
(FEL.47, FEL.49, FEL.207)- denture supérieure : incisives : - I1/ : g FEL.1 ; g et d FEL.2 ; g et d FEL.3- I2/ : g et d FEL.2 ; g et d FEL.3

Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France) 601
- I3/ : g FEL.1 ; g et d FEL.2 ; g et d FEL.3canines sup. : g et d FEL.1 ; g FEL.2 ; g et d FEL.3 ; FEl.209 juv.prémolaires : - P2/ : g FEL.1 ; g et d FEL.3 ; g FEL.233- P3/ : g et d FEL.1 ; g et d FEL.3- P4/ : g et d FEL.1 ; g et d FEL.3molaires : M1/ : FEL.235- denture inférieure :canines inf. : g et d FEL.4 ; g FEL.47 ; d FEL.49 ; g FEL.207prémolaires : - P/3 : g FEL.47, d FEL.4, isolées g FEL.153, d FEL.154- P/4 : g et d FEL.4 ; g FEL.47 ; d FEL.49 ; g FEL.207 ; g
FEL.234molaire : - M/1 : g et d FEL.4 ; g FEL.47 ; g FEL.207dents indéterminées : 4 I isolées, très usées, indéterminables (FEL.42, FEL.214, FEL.236, FEL.237)Post-crânien :- colonne vertébrale :vertèbres cervicales : 2 atlas (FEL.45, FEL.145) ; 2 axis (FEL.46, FEL .169) ; 4 autres (FEL.143, FEL.148, FEL.205, FEL.206)vertèbres dorsales : n°1 – FEL.156 ; n°2 – FEL.155 ; n°3 – FEL.121 ; n°4 – FEL.146 ; n°6 – FEL.262 ; n°9 – FEL.201, FEL.204 ; n°10 – FEL.203 ; n°12 – FEL.150, FEL.246, FEL.263 ; n°13 – FEL.202. vertèbres lombaires : n°1 - FEL.166 ; n°2 - FEL.201, FEL.219, FEL.220 ; n°3 - FEL.218 ; n°4 - FEL.222 ; n°5 - FEL.217 ; n°6 - FEL.221 ; rang indét. FEL.213.sacrum : FEL.261.1 vertèbre caudale : n°8 - FEL.35.- ceintures :omoplate : FEL.200 d.coxal : 2 fragments FEL.200, FEL147.- membre antérieur :humérus : FEL.232 g, FEL.142 d.radius : FEL.116, FEL.211, FEL.212, FEL.217 g, FEL.135 d.cubitus : FEL.109+111, FEL.210 g, distal FEL.247 d.carpe : scapholunaire : FEL.37, FEL.104, FEL.211 g, FEL.114 d.- pisiforme : FEL.16, FEL.93, FEL.186 g, FEL.106 d.- trapèze : FEL.38, FEL.88 g.- trapèzoïde : FEL.39, FEL.87 g, FEL.103 d.- magnum : FEL.23 g.- unciforme : FEL.24 g, FEL.199 d.
- pyramidal : FEL.22, FEL.110 g, FEL.86 d.- métacarpiens : - mtc.1 : FEL.10, FEL.108 g, FEL.174 d.- Mtc.2 : FEL.11, FEL.12, FEL.33, FEL.100 g, FEL.31, FEL.115 d.- Mtc.3 : FEL.11, FEL.20, FEL.99, FEL.157 g, FEL.12d.- Mtc.4 : FEL.101 g, FEL.6, FEL.96+98 d.- Mtc.5 : FEL.13, FEL.101 g, FEL.224d.- membre postérieur : fémur : FEL.225, FEL.230, FEL.231 g, FEL.147 d.rotule : FEL.212 g, FEL.135, FEL.215 d.tibia : FEL.165, FEL.229 g ; FEL.113 d.péroné : proximal FEL.191 d ; distal FEL.139, FEL.245 g ; FEL.189 d.tarse : - astragale : FEL.119, FEL.223 g ; FEL.51 d.- calcanéum : FEL.43, FEL.50 g ; FEL.51 d.- naviculaire : FEL.119 g ; FEL.53 d.- cuboïde ; FEL.187 g ; FEL.55 d.- cunéiforme 2 : FEL.102 d.- cunéiforme 3 : FEL.119 g ; FEL.51 d.métatarsiens : - mtt.2 : FEL.30 g ; FEL.5, FEL.57, FEL.184 d.- Mtt.3 : FEL.8, FEL.182 g ; FEL.57 d.- Mtt.4 : FEL.7, FEL.186 g ; FEL.54+56 d.- Mtt.5 : FEL.183 g ; FEL.58 d.côtes : partielle FEL.199.sternèbres : FEL.138.phalanges 1 : doigt 5 antérieur FEL.26 ; doigt 5 post FEL.14 ; doigt indéterminé FEL.17, FEL.59, FEL.61, FEL.62, FEL.65, FEL.66, FEL.81, FEL.94, FEL.123, FEL.124, FEL.127, FEL.133, FEL.171, FEL.175, FEL.176, FEL.208, FEL.216.phalanges 2 :doigt 3 antérieur g FEL.172 ; doigt indéterminé FEL.6, FEL.21, FEL.27, FEL.28, FEL.29, FEL.36, FEL.48, FEL.63, FEL.67, FEL.71, FEL.78, FEL.125, FEL.177, FEL.192.phalanges 3 :FEL.15, FEL.19, FEL.25, FEL.28, FEL.34, FEL.64, FEL.70, FEL.74, FEL.75, FEL.76, FEL.77, FEL.80, FEL.82, FEL.122, FEL.136, FEL.173, FEL.179, FEL.194.sésamoïdes :FEL.85, FEL.89, FEL.90, FEL.92, FEL.128, FEL.129, FEL.130, FEL.131, FEL.132, FEL.134, FEL.239, FEL.240, FEL.241, FEL.242, FEL.243, FEL.244.




















