
Aspects of the functional morphology in the cranial and cervical skeleton of the sabre-toothed cat...
15
Zoological Journal of the Linnean Society, 2005, 144, 363–377. With 11 figures © 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377 363 Blackwell Science, LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082The Lin- nean Society of London, 2005? 2005 1443 363377 Original Article FUNCTIONAL MORPHOLOGY OF P. OGYGIAM. J. SALESA ET AL. *Corresponding author. E-mail: [email protected] Aspects of the functional morphology in the cranial and cervical skeleton of the sabre-toothed cat Paramachairodus ogygia (Kaup, 1832) (Felidae, Machairodontinae) from the Late Miocene of Spain: implications for the origins of the machairodont killing bite MANUEL J. SALESA 1 *, MAURICIO ANTÓN 2 , ALAN TURNER 1 and JORGE MORALES 2 1 School of Biological & Earth Sciences, Byrom Street, Liverpool John Moores University, Liverpool, L3 3AF, UK 2 Departamento de Palaeobiología, Museo Nacional de Ciencias Naturales-CSIC, José Gutiérrez Abascal, 2. 28006 Madrid, Spain Received January 2004; accepted for publication March 2005 The skull and cervical anatomy of the sabre-toothed felid Paramachairodus ogygia (Kaup, 1832) is described in this paper, with special attention paid to its functional morphology. Because of the scarcity of fossil remains, the anatomy of this felid has been very poorly known. However, the recently discovered Miocene carnivore trap of Batallones-1, near Madrid, Spain, has yielded almost complete skeletons of this animal, which is now one of the best known machairodontines. Consequently, the machairodont adaptations of this primitive sabre-toothed felid can be assessed for the first time. Some characters, such as the morphology of the mastoid area, reveal an intermediate state between that of felines and machairodontines, while others, such as the flattened upper canines and verticalized mandibular symphysis, show clear machairodont affinities. © 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363-377. ADDITIONAL KEYWORDS: Batallones – Carnivora – Felinae – functional anatomy – Mammalia – Miocene – pantherine – Turolian – Vallesian. INTRODUCTION Sabre-toothed predators have evolved several times among different orders of mammals and in somewhat different forms even among nonmammalian synapsids (Turner & Antón, 1997). The similarities in detail between often completely unrelated taxa are so remarkable that the sabre-tooth adaptation has become a text-book example of convergent evolution. However, exactly how it evolved in each case, what the main evolutionary pressures were and where the balance lay between adaptation and phylogenetic con- straint in each group, still remain essentially a mystery. One of the reasons for this lack of resolution is the fact that within each convergent group of sabre-tooth predators it is usually the most derived, crown taxa that are the best known anatomically while the basal taxa have much poorer fossil records. Thus, among the Machairodontinae (the sabre-toothed subfamily within the extant family Felidae), the crown taxa such as Homotherium and Smilodon from the Pliocene and Pleistocene have been known for many decades thanks to complete skulls and skeletons, while the fos- sil record of basal genera such as Paramachairodus from the Late Miocene have traditionally consisted of scarce and fragmentary material. The situation is not
Transcript of Aspects of the functional morphology in the cranial and cervical skeleton of the sabre-toothed cat...

Zoological Journal of the Linnean Society
, 2005,
144
, 363–377. With 11 figures
© 2005 The Linnean Society of London,
Zoological Journal of the Linnean Society,
2005,
144
, 363–377
363
Blackwell Science, Ltd
Oxford, UK
ZOJZoological Journal of the Linnean Society
0024-4082The Lin-
nean Society of London, 2005? 2005
144
3
363377Original Article
FUNCTIONAL MORPHOLOGY OF P. OGYGIAM. J. SALESA ET AL.
*Corresponding author. E-mail: [email protected]
Aspects of the functional morphology in the cranial and cervical skeleton of the sabre-toothed cat
Paramachairodus ogygia
(Kaup, 1832) (Felidae, Machairodontinae) from the Late Miocene of Spain: implications for the origins of the machairodont killing bite
MANUEL J. SALESA
1
*, MAURICIO ANTÓN
2
, ALAN TURNER
1
and JORGE MORALES
2
1
School of Biological & Earth Sciences, Byrom Street, Liverpool John Moores University, Liverpool, L3 3AF, UK
2
Departamento de Palaeobiología, Museo Nacional de Ciencias Naturales-CSIC, José Gutiérrez Abascal, 2. 28006 Madrid, Spain
Received January 2004; accepted for publication March 2005
The skull and cervical anatomy of the sabre-toothed felid
Paramachairodus ogygia
(Kaup, 1832) is described in thispaper, with special attention paid to its functional morphology. Because of the scarcity of fossil remains, the anatomyof this felid has been very poorly known. However, the recently discovered Miocene carnivore trap of Batallones-1,near Madrid, Spain, has yielded almost complete skeletons of this animal, which is now one of the best knownmachairodontines. Consequently, the machairodont adaptations of this primitive sabre-toothed felid can be assessedfor the first time. Some characters, such as the morphology of the mastoid area, reveal an intermediate state betweenthat of felines and machairodontines, while others, such as the flattened upper canines and verticalized mandibularsymphysis, show clear machairodont affinities. © 2005 The Linnean Society of London,
Zoological Journal of theLinnean Society
, 2005,
144
, 363
-
377.
ADDITIONAL KEYWORDS: Batallones – Carnivora – Felinae – functional anatomy – Mammalia – Miocene –
pantherine – Turolian – Vallesian.
INTRODUCTION
Sabre-toothed predators have evolved several timesamong different orders of mammals and in somewhatdifferent forms even among nonmammalian synapsids(Turner & Antón, 1997). The similarities in detailbetween often completely unrelated taxa are soremarkable that the sabre-tooth adaptation hasbecome a text-book example of convergent evolution.However, exactly how it evolved in each case, what themain evolutionary pressures were and where thebalance lay between adaptation and phylogenetic con-
straint in each group, still remain essentially amystery.
One of the reasons for this lack of resolution is thefact that within each convergent group of sabre-toothpredators it is usually the most derived, crown taxathat are the best known anatomically while the basaltaxa have much poorer fossil records. Thus, amongthe Machairodontinae (the sabre-toothed subfamilywithin the extant family Felidae), the crown taxa suchas
Homotherium
and
Smilodon
from the Pliocene andPleistocene have been known for many decadesthanks to complete skulls and skeletons, while the fos-sil record of basal genera such as
Paramachairodus
from the Late Miocene have traditionally consisted ofscarce and fragmentary material. The situation is not

364
M. J. SALESA
ET AL.
© 2005 The Linnean Society of London,
Zoological Journal of the Linnean Society,
2005,
144
, 363–377
very different across the various families of mam-malian sabre-tooths, including the Nimravidae andBarbourofelidae among the Carnivora (Peigné, 2002;Morlo, Peigné & Nagel, 2004) and the marsupialThylacosmilidae (Argot, 2003, 2004).
The result of this unbalanced fossil record is thatemphasis on a series of highly convergent crown taxahas led to an exaggerated perception of homogeneityin adaptation and to suggestions that the so-called‘sabre-tooth complex’ would be under ‘strong pleiotro-pic control’ (Dawson
et al
., 1986). A better fossil recordand a detailed study of basal taxa would greatly helpto clarify the origins of this predatory adaptation, inparticular of the ‘canine shear-bite’, the derived killingbite modality that has been proposed as a functionalexplanation of the morphology of crown sabre-toothedtaxa such as
Smilodon
and
Homotherium
(Akersten,1985; Antón & Galobart, 1999)
The genus
Paramachairodus
Pilgrim, 1913 includesprimitive, leopard-sized sabre-toothed cats knownfrom the Late Miocene faunas of Eurasia (Beaumont,1975; Morales & Soria, 1977; Montoya, 1994; Morlo,1997; Salesa
et al
., 2003). Two species have been tra-ditionally referred to this genus:
P. ogygia
(Kaup,1832), of Vallesian
-
Early Turolian age (MN 9–11 ofMein, 1975) and
P. orientalis
(Kittl, 1887) of Turolianage (MN 11–13), distinguished by several dentaltraits. The more primitive species,
P. ogygia
, had non-crenulated upper canines, a P
4
without ectostyle andwith a strong protocone, and a P
3
with a posterointer-nal expansion.
P. orientalis
had crenulated uppercanines, a P
4
with a well marked ectostyle as well as areduced, backwardly displaced protocone and a P
3
reduced in size and without posterointernal expansion(Salesa
et al
., 2003).In 1924, Zdansky created the species
P. maximiliani
for a damaged skull from the Late Miocene of China.The differences between this third species and
P. orientalis
are largely confined to the more curvedupper canines of
P. maximiliani
, where there arecrenulations on both keels (Zdansky, 1924). Thesecharacters are now considered to be of little systematicvalue, because crenulations are not equally evident inboth keels. Thus for most authors,
P. maximiliani
is asynonym of
P. orientalis
(Pilgrim, 1931; Beaumont,1978; Salesa, 2002; Salesa
et al
., 2003).The anatomy of
Paramachairodus
has been largelyunknown, because it is present in only a few fossillocalities such as Pikermi (Greece, MN 12), Maragha(Iran, MN 11) or Eppelsheim (Germany, MN 9) andrepresented by relatively scarce material. Most findshave been of dentitions. While it has long been clearthat
Paramachairodus
was a sabre-toothed cat, theabsence of more extensive anatomical information hasmeant that its machairodont adaptations and wideraspects of its palaeobiology have remained unknown.
Consequently, no studies of the functional morphologyof
Paramachairodus
have yet been published.The discovery of Batallones-1, a new Late Miocene
fossil locality near Madrid, Spain, has yielded a largenumber of specimens of
P. ogygia
, with at least 24individuals represented and including several skulland mandibles (Salesa, 2002). This locality has beeninterpreted as a carnivore trap based on its specialcharacteristics, such as the presence of an extremelyhigh percentage of carnivores (98%) and the morphol-ogy of the site, essentially a hole with semiverticalwalls (Antón & Morales, 2000; Morales
et al
., 2000).The carnivore guild of Batallones-1 also includes thelion-sized sabre-toothed felid
Machairodus aphanis-tus
, the bear-dog
Amphicyon
sp. aff.
A. castellanus
, theprimitive hyaenid
Protictitherium crassum
and othercarnivores such as mustelids and
Simocyon batalleri
,a medium-sized carnivore related to the extant redpanda (Antón & Morales, 2000; Morales
et al
., 2000;Salesa & Fraile, 2000; Salesa, 2002; Antón
et al
.,2004a; Peigné
et al
., 2005).In this paper we present a functional analysis of the
cranial, mandibular and cervical anatomy of
P. ogygia
,concentrating on aspects directly related to the canineshear-bite adaptation. This analysis has revealed anintermediate morphology between the extant felinesand the more derived sabre-toothed cats such as
Smilodon
or
Homotherium
.
MATERIAL AND METHODS
Functional study of the cranial and cervical anatomyof
Paramachairodus ogygia
has been possible becauseof the great number of newly available fossils of thisspecies: a total of 16 skulls, 12 mandibles and severalvertebrae belonging to the Batallones-1 assemblage.All the material is housed at the Museo Nacional deCiencias Naturales-CSIC in Madrid, Spain. Compari-sons with other Felidae have been made using theextant felines
Panthera leo
,
Panthera tigris
,
Pantherapardus
,
Panthera onca, Uncia uncia, Neofelis nebu-losa, Caracal caracal, Lynx pardina
and
Puma con-color
, belonging to the collections of the MuseoAnatómico de la Universidad de Valladolid and MuseoNacional de Ciencias Naturales (Madrid).
We also used published information on modern felidanatomy (Barone, 2000; Reinhard & Jennings, 1935;Sisson & Grossman, 1962) and on the morphology ofother, more derived sabre-toothed cats such as
Smilo-don fatalis
and
Homotherium latidens
. The latter taxawere chosen as the ideal reference for comparison with
P. ogygia
because they exemplify the most derivedstate for the sabre-toothed adaptations within theMachairodontinae, whilst
P. ogygia
occupies a nearbasal position in the same subfamily. The degree ofmachairodont adaptations appears to be essentially

FUNCTIONAL MORPHOLOGY OF
P. OGYGIA
365
© 2005 The Linnean Society of London,
Zoological Journal of the Linnean Society,
2005,
144
, 363–377
independent of such body size differences as thoseobserved between
P. ogygia
and the referred taxa.Differences in the height of the crown of the upper
canines between
P. ogygia
and reference taxa wereassessed using an ANOVA on the index between crownheight and basal skull length. All the measured uppercanines were unworn specimens. Differences in rela-tive elongation of the neck were assessed by calculat-ing an index between the corpus length of the atlasand the third cervical vertebra. The lack of associatedC3 and atlas of
P. ogygia
in Batallones-1 was overcomeby taking an average measurement between severalisolated C3 and atlas vertebrae and comparing theresultant index with similar indices of reference taxausing a Student’s
t
-test. These results provided themost reliable indication of elongation of the neck inthe absence of complete, associated cervical series of
P. ogygia
.
FUNCTIONAL MORPHOLOGY OF THE SKULL AND MANDIBLE
G
ENERAL
CRANIAL
MORPHOLOGY
The skull of
P. ogygia
shows an overall morphologysimilar to that of a pantherine cat (Figs 1, 6, 11),although there are some differences. The shape of thenasals is neither completely rectangular, as in
Smilo-don
or
Homotherium
, nor triangular, as in most of thepantherines, but shows an intermediate morphology.All nuchal, lambdoid and sagittal crests are welldeveloped, and the join between sagittal and frontalcrests is situated at the level of the postorbital pro-cesses of the frontal bone, as in pantherines andfelines in general. The zygomatic arch of
P. ogygia
isbroadly similar to that of a pantherine, but with apostorbital process that is almost vestigial, unlike thepantherines in which this process is large and pointed.The tympanic bullae of
P. ogygia
are similar to those ofthe pantherines. They are rounded, but less inflatedthan in the latter, and extend from the anterior mar-gin of the mastoid process to the posterior margin ofthe postglenoid process.
M
ASTOID
AREA
The morphology of the mastoid region in
P. ogygia
shows an intermediate condition between that of themore derived Pleistocene sabre-toothed cats, such as
Smilodon
or
Homotherium
, and that of the fossil andextant Felinae. The paraoccipital process of
P. ogygia
is relatively smaller than in the latter, whereas themastoid process has a similar size to that in thefelines, although it is displaced in an anteroventraldirection (Fig. 1). This morphology is plesiomorphic forthe sabre-toothed cats.
In the more derived members of this group, theparaoccipital process becomes very reduced, almostvestigial, and the mastoid process is hyper-developed(Emerson & Radinsky, 1980), overlapping the auditorybulla in its growth in some genera such as
Smilodon
(Fig. 1). Thus, the mastoid region of
P. ogygia
can beconsidered primitive for the Machairodontinae, but itis clearly within the machairodont lineage because itsmorphology is derived in relation to the earliest felidsof the genera
Proailurus
and
Pseudaelurus
. There arefew known basicranial remains of these genera, but inthe
Proailurus
-like skull from Ginn Quarry (Hunt,1998: fig. 19A), and in the holotype of
Pseudaelurusvalidus
(Rothwell, 2001: fig. 3), the paroccipital pro-cess can be seen to be well developed ventrally, sur-passing the level of the mastoid process; this conditionis also found in fossil and extant species of the Felinae.
The mastoid process is the attachment area forsome important muscles directly involved in head-depression movements, such as the
brachiocephalicus
(which has its origin on the humerus), part of thefibres of the
sternocephalicus
(extending from the
manubrium
of the sternum) and the
obliquus capitisanterior
(with its origin on the ventrolateral surface ofthe atlas wings) (Reinhard & Jennings, 1935; Barone,2000). The inferred position of these attachments onthe cranium of
P. ogygia
is shown in Figure 2.The anteroventral displacement of these attach-
ment areas in the Machairodontinae in relation to theFelinae caused by the modification of the mastoid pro-cess has an important consequence. The distancebetween the muscular attachment areas and theatlanto-cranial articulation enlarges, resulting in anincrease in the length of the lever arm of the flexormuscles of the head (Antón & Galobart, 1999) andthus producing a more powerful contraction.
Moreover, there is another interesting implication ofthis displacement of muscle attachment areas. In theprimitive model, exemplified by the felines, the mainfunction of the
obliquus capitis anterior
is the exten-sion of the head on the atlas (Sisson & Grossman,1962; Barone, 2000). This is because a great number ofthe fibres of this muscle are disposed above the pointof rotation of the head over the atlas (Fig. 3A) and thustheir contraction produces the extension of the head.In the sabre-toothed cats, most of the fibres of the
obliquus capitis anterior
are below the point of rota-tion of the head (Fig. 3B) because of the anteroventraldisplacement of the mastoid process. The function ofthis muscle is thus ventral flexion of the head, amotion directly involved in the canine shear-bite(Antón
et al
., 2004b), as discussed further in the finalsection.
The reduction of the paroccipital process in thesabre-toothed cats, including
P. ogygia
, can be relatedto the need to increase the gape of the mandible

366
M. J. SALESA
ET AL.
© 2005 The Linnean Society of London,
Zoological Journal of the Linnean Society,
2005,
144
, 363–377
because of the enlargement of the upper canines. Thefunction of the digastricus muscle, whose insertionareas are the paroccipital process and the ventralflange of the dentary, is to open the mandible (Sisson& Grossman, 1962; Barone, 2000). If the ventral pro-jection and size of the paramastoid process is reduced,which is the condition shown by sabre-toothed felids,the attachment of this muscle is displaced dorsally,increasing fibre lengths and thus allowing efficientcontraction at larger gapes (Emerson & Radinsky,1980; Antón & Galobart, 1999).
The morphology of the mastoid region in P. ogygiashows an early stage of the skull modifications thatwill reach their maximum development in the laterMachairodontinae, such as Smilodon or Homothe-rium. The remodelling of this area in P. ogygia pro-duces an increase in the force generated by the flexormuscles of the head and an increase in the maximumgape of the mandible. These changes suggest that thedevelopment of the canine shear-bite, the machairo-dont type of killing bite, had already started in thisLate Miocene species (Salesa, 2002). This kind of
Figure 1. Left mastoid morphology of some species of Felidae showing the different development of the mastoid process(m.p.) and paraoccipital process (p.p.). A, Panthera leo. B, Paramachairodus ogygia from Batallones-1, B-1377. C, Smilodonfatalis from Rancho La Brea. D, B-1377, skull of P. ogygia from Batallones-1 in left lateral view with mastoid area circled.
m.p. m.p.m.p.
p.p.
p.p.
p.p.
A B C
D5 cm

FUNCTIONAL MORPHOLOGY OF P. OGYGIA 367
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
attack consisted of a well-aimed bite to the throat ofthe prey, which damaged its blood vessels and tracheaand resulted in almost instantaneous death. This is incontrast with the feline killing method where the preyhas to be suffocated with a bite in the throat lastingseveral minutes (Turner & Antón, 1997).
These two different hunting methods explain thesignificant differences in the cranial and cervical mor-phology of the Felinae and Machairodontinae, andthey are already present in one of the most primitivemachairodonts, P. ogygia (Martin, 1980; Biknevicius& van Valkenburgh, 1996; Turner & Antón, 1997;Antón & Galobart, 1999). This kind of attack wasprobably developed in parallel by other ‘sabre-toothed’carnivores, such as the Barbourofelines and Thylacos-milinae, and their mastoid anatomy shows someanalogies with that of the true sabre-toothed felids(Turner & Antón, 1997; Morales et al., 2001).
MANDIBULAR SYMPHYSIS
The mandibular symphysis of P. ogygia is stronglyverticalized, forming an almost square angle with theventral flange of the mandibular corpus (Fig. 4B).Owing to this verticalization, its anterior surface isflat and describes a distinctive plane. This morphol-ogy, typical of the sabre-toothed cats (Fig. 4C), isclearly different from the feline model, in which theanteroventral surface of the mandible is gently curvedupwards (Fig. 4A). The morphology of the mandibularsymphysis of P. ogygia can be related to the canineshear-bite, in which the mandible acts as an anchorwhile the head is flexed downwards, sinking the uppercanines into the flesh of prey.
This morphological change in the mandibular sym-physis of P. ogygia relative to the feline model can be
correlated with the existence of a high vertical stressin that zone during the canine shear-bite, due in partto the use of the mandible as an anchor. The curvedmandibular symphysis of felines does not suffer suchstress during the attack, because this is achieved withthe maxilla and mandible biting at the same time(Biknevicius, van Valkenburgh & Walker, 1996).Another possible interpretation, compatible with theone outlined above, is that an increase in the verticalheight of the symphysis is an efficient way to countersymphyseal bending due to axial twisting of themandibular corpora, an argument that has been putforward to explain the vertically high symphyses ofsabre-toothed therapsids (Jenkins, Thomasson & Nor-man, 2002).
It is also possible that the verticalized mandibularsymphysis of P. ogygia and other sabre-toothed catscan be explained as a consequence of reorganization inthe alveolar zone of the lower canines. Because of thesmaller palate width of the sabre-toothed cats relativeto the felines, the lower canines had to become verti-calized to allow occlusion. Nevertheless, it is probablethat both mechanisms acted together, creating thespecialized symphysis model of the Machairodontinae.
Some sabre-toothed cat taxa, such as Homotheriumand Megantereon, developed a mandibular flange, akind of ventral projection in the symphysis that could
Figure 2. Inferred areas of muscular attachment on theskull of Paramachairodus ogygia.
brachiocephalicus
rectus capitislateralis
digastricus
obliquus capitisanterior
sterno-cephalicus
Figure 3. Anatomical disposition of the muscles brachio-cephalicus (Br.), obliquus capitis anterior (Ob. Cap. Ant.)and obliquus capitis posterior (Ob. Cap. Post.) in Felinaeand Machairodontinae. A, Panthera leo. B, Homotheriumlatidens (artwork by M. Antón).
A
B
Br.
Br.
Ob. Cap. Post.
Ob. Cap. Post.
Ob. Cap. Ant.
Ob. Cap. Ant.

368 M. J. SALESA ET AL.
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
have been a reinforcement of this area. However, thesignificance of this structure is not clear, because itsabsence or presence does not have clear phylogeneticor biomechanical implications. For example, withinthe tribe Smilodontini, Smilodon, the most stronglybuilt among the sabre-toothed cats, does not show thistrait and neither does P. ogygia, whereas it is presentin Megantereon (Fig. 5).
As we pointed out earlier, during the machairodontbite the mandibular symphysis supported a strongvertical tension, which seems to be the main evolu-tionary pressure that produced the verticalized sym-physis. If the morphology of the mandibular flange isobserved in detail, we can see that it is not the thickstructure that might be expected in such a biomechani-cally stressed area. Rather, it is a more or less fragilebony sheet, projected downwards only in the most lat-eral parts of the symphysis. This structure alsoappears in other groups of sabre-toothed carnivorans,such as the marsupial Thylacosmilus atrox and mostof the nimravids, such as Hoplophoneus or Eusmilus.
In summary, the significance of this structure isunclear, and its absence in P. ogygia clearly did notaffect the development of a verticalized, derived man-dibular symphysis.
CORONOID PROCESS
The coronoid process of the more derived sabre-toothed felids is highly reduced in relation to the felinepattern (Fig. 4A, C) as a consequence of reorganiza-tion in the fibres of the temporalis muscle, whichbecame longer and more vertical, a change related tothe need to increase the maximum gape (Emerson &
Figure 4. Left hemimandibles of Felidae showing differ-ences in the development of the mandibular coronoid pro-cess. A, Panthera leo. B, Paramachairodus ogygia fromBatallones-1. C, Smilodon fatalis from Rancho La Brea.
A
B
C5 cm
Figure 5. Comparative views of the skull and mandible oftwo Smilodontini species. A, Paramachairodus ogygia. B,Megantereon cultridens (artwork by M. Antón).
A
B

FUNCTIONAL MORPHOLOGY OF P. OGYGIA 369
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
Radinsky, 1980; Martin, 1980; Antón et al., 2004a).P. ogygia has a coronoid process that is only slightlyreduced in comparison with a pantherine cat (Fig. 4B).This morphology is also seen in the contemporaneoussabre-toothed felid Machairodus aphanistus (Antónet al., 2004a) and reflects the primitive pattern of thetemporalis muscle in these first machairodontines.
CONDYLAR PROCESS
The condylar process of P. ogygia shows a similarorientation to that of the pantherines (Fig. 4A, B) andclearly differs from that of Smilodon fatalis (Fig. 4C).In the more derived sabre-toothed cats, the condylarprocess is orientated posteroventrally, in order toallow great gapes, whereas in the pantherines thisorientation is posterodorsal. Thus, P. ogygia shows theplesiomorphic state of this character, which indicatesthat the gapes produced by its mandible would not bevery different from those produced by pantherine cats.This morphology of P. ogygia also suggests that thechanges in the coronoid process of the early machair-
odonts may have been more related to changes in theorientation of the fibres of the temporalis muscle thanto changes in the mandibular gape.
UPPER CANINES
An index of the basal length of the skull vs. the uppercanine crown height was calculated for P. ogygia andnine species of Felinae (Table 1). These data wereanalysed by ANOVA; the results are given in Table 2.It can be seen that there are significant differencesbetween both groups, in that P. ogygia has relativelylonger upper canines than the Felinae. The uppercanines of P. ogygia are also laterally flattened(Fig. 6), whereas in Felinae their section is rounded.They also lack crenulations, which is the primitivecondition, although these are not present in all sabre-toothed species. While crenulations in the uppercanines would help to penetrate the flesh, their pres-ence is obviously a minor requirement in the develop-ment of the canine shear-bite, as showed by theabsence of this trait in the large upper canines of other
Table 1. Skull and upper canine measurements for Paramachairodus ogygia and nine species of Felinae. Abbreviations:CH, canine height; ML, canine mesiodistal length; BB, canine buccolingual breadth; BL, skull basal length; BL/CH index,index between BL and CH
Number Species CH ML BB BL BL/CH index
- P. tigris 35.98 18.93 14.57 247.35 6.875- P. tigris 37.12 18.69 14.48 247.35 6.6642415 P. onca 32.69 16.25 13.30 197.25 6.034125 P. onca 36.04 17.68 14.43 204.80 5.683409 P. concolor 24.40 12.02 9.87 176.60 7.2381599 P. pardus 29.29 14.06 10.80 167.75 5.727MNCN-4259 P. pardus 28.61 12.36 9.24 157.74 5.513- U. uncia 30.11 11.93 10.08 156.85 5.209- N. nebulosa 36.67 12.25 10.30 157.10 4.284- N. nebulosa 36.30 12.85 10.08 157.10 4.328MNCN-4060 P. leo 40.89 20.63 14.21 262.30 6.415MNCN-4060 P. leo 40.90 20.31 14.18 262.30 6.413275 P. concolor 22.99 11.28 9.11 153.84 6.6922 P. concolor 27.04 13.42 10.83 171.15 6.330MNCN-60 L. wiedii 11.99 5.57 3.81 82.41 6.873MNCN-60 L. wiedii 11.97 5.72 3.87 82.41 6.885MNCN-16784 L. pardina 16.09 6.49 5.20 91.95 5.715MNCN-4260 P. leo 46.38 25.04 17.30 310.90 6.700MNCN-4260 P. leo 45.86 24.84 17.09 310.90 6.780MNCN-16825 P. leo 48.35 25.43 17.13 320.85 6.640B-1377 P. ogygia 39.94 - - 165.55 4.145B-847 P. ogygia 36.06 - - 168.15 4.663B-847 P. ogygia 36.03 15.15 - 168.15 4.667B-5197 P. ogygia 37.91 15.39 9.41 159.80 4.215B-5197 P. ogygia 39.84 15.72 - 159.80 4.011B-4869 P. ogygia 33.25 13.81 - 157.15 4.726

370 M. J. SALESA ET AL.
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
sabre-toothed cat such as Megantereon, a morederived species than P. ogygia.
The possession of flattened and elongated uppercanines is a clear machairodont trait, and its presencein P. ogygia is enough for us to infer that this specieshad already developed the specialized hunting methodof this group. If P. ogygia tried to kill prey as felinesdo, with either a nape bite or a suffocating bite, theneither violent contact with bone or the lateral motionsof the struggling prey could have caused breakage ofthe upper canines, which were inherently more fragilethan the rounded-section upper canines of a feline(Emerson & Radinsky, 1980; Turner & Antón, 1997;Antón & Galobart, 1999).
FUNCTIONAL MORPHOLOGY OF CERVICAL VERTEBRAE
The atlas wings of P. ogygia project backward a littlemore than in the pantherines, but not as far as inother, more derived sabre-toothed cats such asHomotherium or Smilodon, in which this posteriorprojection is extreme (Fig. 7). This morphologyincreases the breadth and fibre length of the obliquuscapitis anterior (Fig. 8), which runs from the ventralsurface of the atlas wings to the mastoid process, andthe obliquus capitis posterior, which extends from thedorsal surface of the atlas wings to the lateral surfaceof the spinous process of the axis (Fig. 8) (Barone,2000; Salesa, 2002; Antón et al., 2004b). In more
Table 2. Results of ANOVA for BL/CH index in Table 1
Sum of squares d.f. Mean square F P
Between groups 14.056 1 14.056 24,755 0.000Within groups 13.628 24 0.568Total 27.684 25
Figure 6. Skulls and mandibles of four individuals of Paramachairodus ogygia from Batallones-1. A, B-847. B, B-4322. C,B-4778. D, B-7022.
5 cm
A B
CD

FUNCTIONAL MORPHOLOGY OF P. OGYGIA 371
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
derived machairodonts, such as Smilodon andHomotherium, this process of the axis is lengthenedposteriorly, which also increases the breadth of theobliquus capitis posterior. Nevertheless, the spinousprocess in P. ogygia has a pantherine-like morphology,with little backward projection, thus showing aprimitive state for this character.
The enlargement in the breadth of the musclesinvolved in the flexion and lateral rotation of the head
produces an increase in their contraction strength. Inaddition, the increased distance between the origin andinsertion of these muscles produces a lengthening of thelever arm, which also serves to increase the strength ofaction (Akersten, 1985; Antón & Galobart, 1999).
In P. ogygia, the third to seventh cervical vertebraeseem to be long in relation to those of the pantherinecats (Figs 9, 10). To assess this, an index between thecorpus length of C3 and atlas was calculated forP. ogygia, Genetta genetta and six species of Felinae(Table 3), and these data analysed with a Student’s t-test. P. ogygia shows a low sexual dimorphism index(Salesa, 2002) and the selected vertebrae used in theanalysis were all very similar in size. The result isshown in Table 4, demonstrating that there are signifi-cant differences between P. ogygia and Felinae, such
Figure 7. Development of the atlas wings in dorsal view.A, Panthera pardus. B, Paramachairodus ogygia, aspecimen with an associated axis, before preparation.C, Smilodon fatalis from Rancho La Brea (modified fromMerriam & Stock, 1932).
A B
3 cm
C
Figure 8. Composite reconstruction of the skull, mandibleand cervical vertebrae of Paramachairodus ogygia, basedon material of several individuals from Batallones-1, show-ing the inferred position of the main cranio-cervical mus-cles relevant to the canine shear-bite (artwork by M.Antón).
obliquus capitisanterior
obliquus capitisposterior
brachiocephalicus
Table 3. Atlas and third vertebra (C3) measurements forParamachairodus ogygia, six species of Felinae and theviverrid Genetta genetta. Abbreviations: VCL. vertebral cor-pus length; C3/AT index, index between vertebral corpuslength of C3 and Atlas. The specimens of P. ogygia do notcorrespond to single individuals, so the index was calcu-lated with the average of each VCL, 38.60 for Atlas, and26.62 for C3
Number Element Species VCLC3/ATindex
B-3874 (1) Atlas P. ogygia 39.51B-4914 Atlas P. ogygia 41.69B-1274 Atlas P. ogygia 36.28B-4318 Atlas P. ogygia 38.78B-ss Atlas P. ogygia 36.76B-4318c C3 P. ogygia 24.71B-744 (5) C3 P. ogygia 28.53 0.6902440 Atlas P. pardus 44.952440 C3 P. pardus 24.19 0.5381599 Atlas P. pardus 42.761599 C3 P. pardus 21.37 0.5002415 Atlas P. onca 45.392415 C3 P. onca 22.77 0.502125 Atlas P. onca 49.80125 C3 P. onca 25.80 0.518409 Atlas P. concolor 41.29409 C3 P. concolor 21.01 0.509275 Atlas P. concolor 39.25275 C3 P. concolor 22.91 0.584- Atlas P. tigris 60.35- C3 P. tigris 29.40 0.487MNCN-16810 Atlas L. pardina 21.70MNCN-16810 C3 L. pardina 9.66 0.4451518 Atlas C. caracal 23.251518 C3 C. caracal 13.82 0.594MNCN-14233 Atlas G. genetta 15.62MNCN-14233 C3 G. genetta 12.72 0.814

372 M. J. SALESA ET AL.
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
that P. ogygia has a longer C3 in relation to the atlasthan the Felinae. G. genetta also shows significant dif-ferences in this index.
As several authors have discussed, a long neck is atrait typical of the more derived machairodonts, and
can be related to the necessity for greater accuracy inthe neck movements during the canine shear-bite(Schaub, 1925; Ballesio, 1963; Turner & Antón, 1997;Antón & Galobart, 1999). However, since primitiveviverrids such as G. genetta also have long necks, thistrait could be a primitive character retained by sabre-toothed cats. Greater knowledge of cervical mor-phology in early felids, such as Proailurus andPseudaelurus, is necessary in order to settle this issue.In Homotherium and Smilodon, a clear reinforcementin the transverse processes of the cervical vertebraecan also be observed in addition to this cervical length-ening. These processes are the insertion areas of thescalenus and longus capitis (Antón & Galobart, 1999),
Figure 9. Photographs of first to seventh cervical vertebrae (C1-C7) (anterior to left) in dorsal view. A, Paramachairodusogygia from Batallones-1, respectively, B-4561, B-5407, B-744 (5), B-5458, B-5459, B-707 (12) and B-707 (12) (the latterhave the same number); B, Panthera pardus, 1599.
A
B
5 cm
Table 4. Results of Student’s t-test for C3/AT index ofTable 3
t-test d.f.Sig.(2-tailed)
Meandifference
C3/AT index -10.939 8 0.000 -0.170

FUNCTIONAL MORPHOLOGY OF P. OGYGIA 373
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
the main flexors of the neck (Barone, 2000). P. ogygiadoes not show this change, its transverse processesbeing very similar in size and morphology to those ofthe pantherine cats.
In summary, the neck morphology of P. ogygia showsa combination of some machairodont traits, such asthe lengthening of the whole vertebrae, and primitiveones, such as the small backwards projection of theatlas wings and the overall morphology of the cervicalvertebrae.
PALAEOETHOLOGICAL IMPLICATIONS
The morphology of the skull and cervical region inP. ogygia shows the beginning of the typical adapta-tions of the Machairodontinae, which are closelyrelated to the highly specialized killing method of thisgroup by means of the so-called canine shear-bite(Akersten, 1985). These cranio-cervical modifications,
which reach their highest development in the morederived sabre-toothed felids of the Plio-Pleistocenesuch as Homotherium and Smilodon, can be summa-rized as follows:
1. Presence of elongated and flattened upper canines.2. Verticalization of the mandibular symphysis.3. Reduction of the coronoid process of the mandible.4. Enlargement and anteroventral displacement of
the mastoid process.5. Reduction of the paroccipital process.6. Strong backwards projection of the atlas wings and
lengthening of the spinous process of the axis.7. Lengthening of the corpus of the cervical vertebrae.8. Enlargement of the transverse processes of the cer-
vical vertebrae.
As described above, P. ogygia shows a clear develop-ment of characters 1 and 2 only; characters 3, 4, 5 and7 are only moderately derived relative to the feline
Figure 10. Photographs of the first to seventh cervical vertebrae (C1-C7) (anterior to left) in Fig. 9, in lateral view.A, Paramachairodus ogygia from Batallones-1. B, Panthera pardus.
A
B
5 cm

374 M. J. SALESA ET AL.
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
model, while character 6 is very moderately expressedand character 8 is not present.
Although P. ogygia did not develop a completemachairodont cranio-cervical plan, the presence ofelongated and flattened upper canines undoubtedlypoints towards the canine shear-bite as the prey-killing method. The extant felines kill with either aprolonged bite to the throat or muzzle of prey, result-ing in its strangulation or suffocation (Schaller, 1972;Turner & Antón, 1997; Antón et al., 2004b), or with astrong bite to the nape, which causes death by break-ing the skull or spinal cord (Leyhausen, 1979; Turner& Antón, 1997).
The first method is used with large prey, whereasthe second is only applied to small animals, whichhave cranial bones fragile enough to avoid damagingthe canines of the predator (Seidensticker & McDou-gal, 1993). If sabre-toothed cats tried to kill their preywith these feline techniques, the long and flattenedupper canines would probably have been broken,either by hitting bone or because of the lateral ten-sions caused by the struggling of prey during the bite(Emerson & Radinsky, 1980; Turner & Antón, 1997;Antón & Galobart, 1999).
Consequently, the upper canines of the sabre-toothed cats were likely to have been used in a differ-ent way, and were probably developed to bite thethroat of the prey with a head flexion movement thatcut blood vessels and trachea, thus producing rapiddeath through combined suffocation and rapid bloodloss. During this motion, the mandible acted as ananchor. That would be the reason why the morphologyof the mandibular symphysis developed into a verti-calized and stabilizing border. The use of the machair-odonts’ powerful forepaws for the immobilization ofprey during the bite further contributed to protectingthe fragile upper canines from violent lateral tensions(Rawn-Schatzinger, 1992; Lewis, 1997; Turner &Antón, 1997).
In general, all cranial modifications of sabre-toothedcats can be seen as related to their specialized huntingand killing method. Thus the hyper-development ofthe mastoid process and the backwards projection ofthe atlas wings increased the leverage arm andstrength of the head flexor muscles, while the dorsaldisplacement of the paroccipital process and thereduction of the coronoid process increased themaximum gape. Other modifications contributed toreinforce the neck, mandible and skull in order towithstand the development of powerful forces duringthe kill.
The cranio-mandibular morphology of P. ogygiashows that this species had developed some importantmachairodont traits, such as the elongated and flat-tened upper canines and verticalized mandibular sym-physis. Other regions, such as the mastoid area and
atlas wings, remained in a relatively primitive state.Nevertheless, this morphological mosaic reveals thatthe most important aspects of the canine shear-bitewere the flattened upper canines and the verticalizedsymphyseal morphology, which are directly related tothe cutting-action and to stabilization of the bite.Other aspects, such as the strength of the head flexormuscles, seem to have been less critical and were fur-ther emphasized only later in evolution.
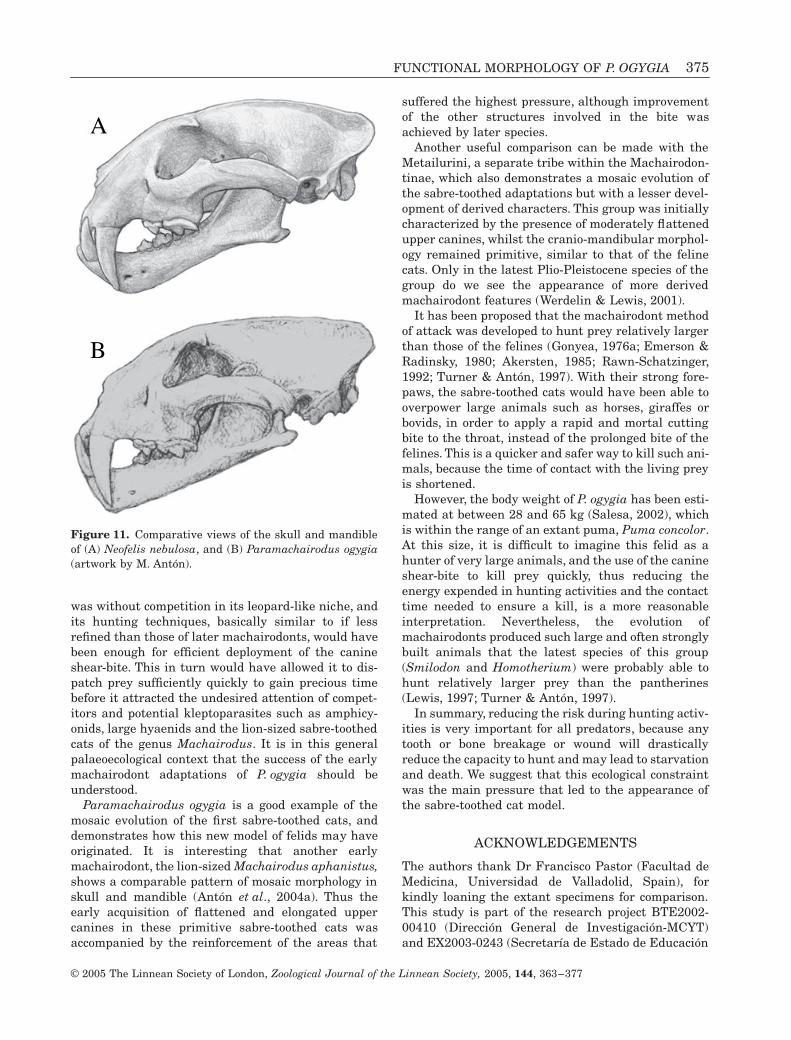
What does this morphology mean in the context ofmachairodont palaeoethology? P. ogygia had devel-oped the main morphological traits for the canineshear-bite, but it was also far from the highly derivedpattern shown by the latest sabre-toothed cats. Thereare several behavioural implications of the differencesbetween P. ogygia and both the pantherines and thecrown machairodontines. For example, if we comparethe morphology of the skull and neck in P. ogygia withthe superficially similar extant clouded leopard,Neofelis nebulosa (Fig. 11), we find that the pan-therine cat has an overall skull morphology compara-ble to that of the machairodont, with upper caninesalso of comparable crown height. However, in theextant species the upper canines are not flattened, themandibular symphysis is not as verticalized, themastoid process remains primitively small and with-out ventral projection, while the length of the neck istypical for a pantherine (Gonyea, 1976b). This set ofdifferences is enough to indicate the presence of cor-responding differences in killing behaviour, with theclouded leopard using a typical pantherine bite andthe early machairodont already relying more heavilyon the immobilization and quick killing of preythrough blood loss.
However, if we compare P. ogygia with Megantereon(a derived Plio-Pleistocene machairodontine, betweena leopard and a jaguar in size and possibly descendedfrom Paramachairodus), the main behavioural differ-ences would be that the younger taxon would kill preyeven faster and more efficiently than P. ogygia, andwould also have had access to relatively larger prey. Ifthey had been sympatric, then both sabre-toothedtaxa would have been in direct competition forresources due to their overlapping body sizes. Bothspecies would also have shared similar solitary habitsand a preference for wooded environments, asinferred from aspects of their body proportions thathave been shown to be related to ambush hunting(Kurtén, 1968; Van Valkenburgh, 1987; Marean, 1989;Turner & Antón, 1997; Agustí & Antón, 2002; Salesa,2002) and to closed-habitat preferences in felids (Gon-yea, 1976a, b).
In the event of hypothetical direct competition,Megantereon would have been at a distinct advantagebecause of its more efficient machairodont adapta-tions. However, in the Vallesian of Eurasia, P. ogygia
FUNCTIONAL MORPHOLOGY OF P. OGYGIA 375
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
was without competition in its leopard-like niche, andits hunting techniques, basically similar to if lessrefined than those of later machairodonts, would havebeen enough for efficient deployment of the canineshear-bite. This in turn would have allowed it to dis-patch prey sufficiently quickly to gain precious timebefore it attracted the undesired attention of compet-itors and potential kleptoparasites such as amphicy-onids, large hyaenids and the lion-sized sabre-toothedcats of the genus Machairodus. It is in this generalpalaeoecological context that the success of the earlymachairodont adaptations of P. ogygia should beunderstood.
Paramachairodus ogygia is a good example of themosaic evolution of the first sabre-toothed cats, anddemonstrates how this new model of felids may haveoriginated. It is interesting that another earlymachairodont, the lion-sized Machairodus aphanistus,shows a comparable pattern of mosaic morphology inskull and mandible (Antón et al., 2004a). Thus theearly acquisition of flattened and elongated uppercanines in these primitive sabre-toothed cats wasaccompanied by the reinforcement of the areas that
suffered the highest pressure, although improvementof the other structures involved in the bite wasachieved by later species.
Another useful comparison can be made with theMetailurini, a separate tribe within the Machairodon-tinae, which also demonstrates a mosaic evolution ofthe sabre-toothed adaptations but with a lesser devel-opment of derived characters. This group was initiallycharacterized by the presence of moderately flattenedupper canines, whilst the cranio-mandibular morphol-ogy remained primitive, similar to that of the felinecats. Only in the latest Plio-Pleistocene species of thegroup do we see the appearance of more derivedmachairodont features (Werdelin & Lewis, 2001).
It has been proposed that the machairodont methodof attack was developed to hunt prey relatively largerthan those of the felines (Gonyea, 1976a; Emerson &Radinsky, 1980; Akersten, 1985; Rawn-Schatzinger,1992; Turner & Antón, 1997). With their strong fore-paws, the sabre-toothed cats would have been able tooverpower large animals such as horses, giraffes orbovids, in order to apply a rapid and mortal cuttingbite to the throat, instead of the prolonged bite of thefelines. This is a quicker and safer way to kill such ani-mals, because the time of contact with the living preyis shortened.
However, the body weight of P. ogygia has been esti-mated at between 28 and 65 kg (Salesa, 2002), whichis within the range of an extant puma, Puma concolor.At this size, it is difficult to imagine this felid as ahunter of very large animals, and the use of the canineshear-bite to kill prey quickly, thus reducing theenergy expended in hunting activities and the contacttime needed to ensure a kill, is a more reasonableinterpretation. Nevertheless, the evolution ofmachairodonts produced such large and often stronglybuilt animals that the latest species of this group(Smilodon and Homotherium) were probably able tohunt relatively larger prey than the pantherines(Lewis, 1997; Turner & Antón, 1997).
In summary, reducing the risk during hunting activ-ities is very important for all predators, because anytooth or bone breakage or wound will drasticallyreduce the capacity to hunt and may lead to starvationand death. We suggest that this ecological constraintwas the main pressure that led to the appearance ofthe sabre-toothed cat model.
ACKNOWLEDGEMENTS
The authors thank Dr Francisco Pastor (Facultad deMedicina, Universidad de Valladolid, Spain), forkindly loaning the extant specimens for comparison.This study is part of the research project BTE2002-00410 (Dirección General de Investigación-MCYT)and EX2003-0243 (Secretaría de Estado de Educación
Figure 11. Comparative views of the skull and mandibleof (A) Neofelis nebulosa, and (B) Paramachairodus ogygia(artwork by M. Antón).
A
B

376 M. J. SALESA ET AL.
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
y Universidades del MECD). We thank the Comu-nidad Autónoma de Madrid (Dirección General de Pat-rimonio Histórico) for continuous funding support andresearch permissions. A.T. thanks the British Councilfor travel funding. Additional support was provided bythe National Geographic Society (Grant 6964–01). Wealso thank an anonymous referee, whose commentsimproved the quality of the manuscript.
REFERENCES
Agustí J, Antón M. 2002. Mammoths, sabertooths, andhominids. New York: Columbia University Press.
Akersten WA. 1985. Canine function in Smilodon (Mamma-lia; Felidae; Machairodontinae). Contributions in Science356: 1–22.
Antón M, Galobart A. 1999. Neck function and predatorybehaviour in the scimitar-toothed cat Homotherium latidens(Owen). Journal of Vertebrate Paleontology 19 (4): 771–784.
Antón M, Morales J. 2000. Inferencias paleoecológicas de laasociación de carnívoros del yacimiento de Cerro Batallones.In: Morales J, Nieto M, Amezua L, Fraile S, Gómez E, Her-ráez E, Peláez-Campomanes P, Salesa MJ, Sánchez IM,Soria D, eds. Patrimonio Paleontológico de la Comunidad deMadrid. Serie ‘Arqueología, Paleontología y Etnografía’,Vol. 6. Madrid: Dirección General de Patrimonio Histórico-Artístico, 190–201.
Antón M, Salesa MJ, Morales J, Turner A. 2004a. Firstknown complete skulls of the scimitar-toothed cat Machairo-dus aphanistus (Felidae, Carnivora) from the Spanish lateMiocene site of Batallones-1. Journal of Vertebrate Paleon-tology 24 (4): 957–969.
Antón M, Salesa MJ, Pastor JF, Sánchez IM, Fraile S,Morales J. 2004b. Implications of the mastoid anatomy oflarger extant felids for the evolution and predatory behav-iour of sabretoothed cats (Mammalia, Carnivora, Felidae).Zoological Journal of the Linnean Society 140: 207–221.
Argot C. 2003. Evolution of the South American mammalianpredators (Borhyaenoidea): anatomical and palaeobiologicalimplications. Zoological Journal of the Linnean Society 140:487–521.
Argot C. 2004. Functional-adaptive features and palaeobio-logic implications of the postcranial skeleton of the lateMiocene sabretooths borhyaenoid Thylacosmilus atrox(Metatheria). Alcheringa 28: 229–266.
Ballesio R. 1963. Monographie d’un Machairodus du gise-ment villafranchien de Senèze: Homotherium crenatidensFabrini. Traveaux du Laboratoire de Géologie de la Facultéde Sciences de Lyon 9: 1–129.
Barone R. 2000. Anatomie comparée des Mammifères domes-tiques, Tome 2, Arthrologie et Myologie. Paris: Éditions Vigot.
Beaumont G. 1975. Recherches sur les Félidés (Mammifères,Carnivores) du Pliocène inférieur des sables à Dinotheriumdes environs d’Eppelsheim (Rheinhessen). Archives des Sci-ences Physiques et Naturelles, Genève 28 (3): 369–405.
Beaumont G. 1978. Notes complémentaires sur quelques
Félidés (Carnivores). Archives des Sciences Physiques etNaturelles, Genève 31 (3): 219–227.
Biknevicius AR, van Valkenburgh B. 1996. Design for kill-ing: craniodental adaptations of mammalian predators. In:Gittleman JL, ed. Carnivore behavior, ecology and evolution,Vol. 2. London: Cornell University Press, 393–428.
Biknevicius AR, van Valkenburgh B, Walker J. 1996. Inci-sor size and shape: implications for feeding behaviors insaber-toothed cats. Journal of Vertebrate Paleontology 16 (3):510–521.
Dawson MR, Stucky RK, Krishtalka L, Black CC. 1986.Machaeroides simpsoni, new species, oldest known sabertoothcreodont (Mammalia), of Lost Cabin Eocene. Contributions toGeology, University of Wyoming, Special Paper 3: 177–182.
Emerson SB, Radinsky L. 1980. Functional analysis ofsabertooth cranial morphology. Paleobiology 6 (3): 295–312.
Gonyea WJ. 1976a. Behavioral implications of saber-toothedfelid morphology. Paleobiology 2: 232–342.
Gonyea WJ. 1976b. Adaptive differences in the body propor-tions of large felids. Acta Anatomica 96: 81–86.
Hunt RH Jr. 1998. Evolution of the aeluroid Carnivora: diver-sity of the earliest aeluroids from Eurasia (Quercy, Hsanda-Gol) and the origin of felids. American Musem Novitates3252: 1–65.
Jenkins I, Thomasson JJ, Norman DB. 2002. Primates andengineering principles: applications to craniodental mecha-nisms in ancient terrestrial predators. SenckenbergianaLethaea 82 (1): 223–240.
Kurtén B. 1968. Pleistocene mammals of Europe. Chicago:Aldine Publishing Co.
Lewis ME. 1997. Carnivoran paleoguilds of Africa: implica-tions of hominid food procurement strategies. Journal ofHuman Evolution 32: 257–288.
Leyhausen P. 1979. Cat behavior: the predatory and socialbehavior of domestic and wild cats. New York: STPM Press.
Marean CW. 1989. Sabretooth cats and their relevance forearly hominid diet and evolution. Journal of Human Evolu-tion 18: 559–582.
Martin LD. 1980. Functional morphology and the evolution ofcats. Transactions of the Nebraska Academy of Science 8:141–154.
Mein P. 1975. Biozonation du neogène Mediterranée a partirdes mammifères. Proceedings of the VI Congress of theRegional Committee on Mediterranean Neogene Stratigra-phy, Bratislava: 77–81.
Merriam JC, Stock C. 1932. The Felidae of Rancho la Brea.Washington: Carnegie Institution of Washington.
Montoya P. 1994. Los Macromamíferos del Mioceno superiordel área de Crevillente (Alicante). Unpublished DPhilThesis, Universitat de València, Facultat de CienciesBiologiques, Departament de Geologia.
Morales J, Alcalá L, Amezua L, Antón M, Fraile S, GómezE, Montoya P, Nieto M, Pérez B, Salesa MJ, SánchezIM. 2000. El yacimiento de el Cerro de los Batallones. In:Morales J, Nieto M, Amezua L, Fraile S, Gómez E, HerráezE, Peláez-Campomanes P, Salesa MJ, Sánchez IM, Soria D,eds. Patrimonio Paleontológico de la Comunidad deMadrid. Serie ‘Arqueología, Paleontología y Etnografía’,

FUNCTIONAL MORPHOLOGY OF P. OGYGIA 377
© 2005 The Linnean Society of London, Zoological Journal of the Linnean Society, 2005, 144, 363–377
Vol. 6. Madrid: Dirección General de Patrimonio Histórico-Artístico, 179–190.
Morales J, Salesa MJ, Pickford M, Soria D. 2001. A newtribe, new genus and two new species of Barbourofelinae(Felidae, Carnivora, Mammalia) from the Early Miocene ofEast Africa and Spain. Transactions of the Royal Society ofEdinburgh: Earth Sciences 91 (1): 97–102.
Morales J, Soria D. 1977. Presencia de la asociaciónMachairodus-Paramachairodus en Concud (Teruel). Teruel57–58: 1–9.
Morlo M. 1997. Die Raubtiere (Mammalia, Carnivora) ausdem Turolium von Dorn-Dürkheim 1 (Rheinhessen). Teil 1:Mustelidae, Hyaenidae, Percrocutidae, Felidae. CourierForschungs-Institut Senckenberg 197: 11–47.
Morlo M, Peigné S, Nagel D. 2004. A new species of Prosan-sanosmilus: implications for the systematic relationships ofthe family Barbourofelidae new rank (Carnivora, Mamma-lia). Zoological Journal of the Linnean Society 140: 43–61.
Peigné S. 2002. Systematic review of European Nimravinae(Mammalia, Carnivora, Nimravidae) and the phylogeneticrelationships ofd Paleogene Nimravidae. Zoologica Scripta32: 199–229.
Peigné S, Salesa MJ, Antón M, Morales J. 2005. Ailuridcarnivoran mammal Simocyon from the Late Miocene ofSpain and the systematics of the genus. Acta PaleontologicaPolonica 50 (2): 219–238.
Pilgrim GE. 1913. The correlation of the Siwaliks with mam-mal horizons of Europe. Records of the Geological Survey ofIndia 43 (4): 264–326.
Pilgrim GE. 1931. Catalogue of the Pontian Carnivora ofEurope. London: British Museum of Natural History.
Rawn-Schatzinger V. 1992. The scimitar cat Homotheriumserum Cope. Illinois State Museum Reports of Investigation47: 1–80.
Reinhard J, Jennings HS. 1935. Anatomy of the cat. NewYork: Henry Holt & Company.
Rothwell T. 2001. A partial skeleton of Pseudaelurus(Carnivora: Felidae) from the Nambé Member of the Tesuque
Formation, Española Basin, New Mexico. American MuseumNovitates 3342: 1–31.
Salesa MJ. 2002. Estudio anatómico, biomecánico, paleo-ecológico y filogenético de Paramachairodus ogygia (Kaup,1832) Pilgrim, 1913 (Felidae, Machairodontinae) delyacimiento vallesiense (Mioceno superior) de Batallones-1(Torrejón de Velasco, Madrid). Unpublished DPhil Thesis,Departamento de Biología Animal I, Facultad de CienciasBiológicas, Universidad Complutense de Madrid, Madrid.
Salesa MJ, Fraile S. 2000. Los carnívoros fósiles del Neógenomadrileño. In: Morales J, Nieto M, Amezua L, Fraile S,Gómez E, Herráez E, Peláez-Campomanes P, Salesa MJ,Sánchez IM, Soria D, eds. Patrimonio Paleontológico de laComunidad de Madrid. Serie ‘Arqueología, Paleontología yEtnografía’, Vol. 6. Madrid: Dirección General de PatrimonioHistórico-Artístico, 248–255.
Salesa MJ, Montoya P, Alcalá L, Morales J. 2003. Elgénero Paramachairodus Pilgrim, 1913 (Felidae, Machairo-dontinae) en el Mioceno superior español. Coloquios de Pale-ontología, Volumen Extraordinario 1: 603–615.
Schaller GB. 1972. The Serengeti lion. Chicago: University ofChicago Press.
Schaub S. 1925. Uber die osteologie von Machaerodus cul-tridens Cuvier. Eclogae Geologicae Helvetiae 19 (1): 255–266.
Seidensticker J, McDougal C. 1993. Tiger predatory behav-iour, ecology and conservation. Symposia of the ZoologicalSociety of London 65: 105–125.
Sisson S, Grossman JD. 1962. The anatomy of the domesticanimals, 4th edn. Philadelphia, PA: W.B. Saunders.
Turner A, Antón M. 1997. The big cats and their fossil rela-tives. New York: Columbia University Press.
Van Valkenburgh B. 1987. Skeletal indicators of locomotorbehavior in living and extinct carnivores. Journal of Verte-brate Paleontology 7 (2): 162–182.
Werdelin L, Lewis ME. 2001. A revision of the genus Dinofe-lis (Mammalia, Felidae). Zoological Journal of the LinneanSociety 132: 147–258.




















