
Carbon isotope fractionation and changes in the flux and composition of particulate matter resulting...
14
Limnol. Oceanogr., 32(l), 1987, 83-96 0 1987, by the American Society of Limnology and Oceanography, Inc. Carbon isotope fractionation and changes in the flux and composition of particulate matter resulting from biological activity during a sediment trap experiment in Lake Greifen, Switzerland’ Cindy Lee2 Woods Hole Oceanographic Institution, Woods Hole, Massachusetts 02543 Judith A. McKenzie Geological Institute, Swiss Federal Institute of Technology, 8092 Zurich Michael Sturm Federal Institute for Water Resources and Water Pollution Control (EAWAG), Swiss Federal Institute of Technology, 8600 Diibendorf Abstract Sediment traps were deployed in Lake Greifen, a hard-water eutrophic, Swiss lake, for 2 weeks in May and June 1983. Oxygen, pH, and suspended particle concentrations indicated a sharp increase in productivity during the experiment. Carbon isotope composition of the dissolved inorganic carbon recorded the balance between photosynthesis and respiration and showed possible evidence of difhtsion of atmospheric CO, across the water-atmosphere interface, most likely to replace CO, used during photosynthesis. Allochthonous mineral debris from resuspended sediments or river input was a major component of the sedimenting material. Due to removal of COz, large, newly formed calcite crystals precipitated when photosynthesis increased. The carbon isotope ratio of trap calcite and the composition of trap amino acids showed the influence of the detrital input on the more dominant isotopic signal of authigenic calcite. Sediment calcite is enriched in 13C relative to authigenic calcite in the sediment traps. This enrichment is likely due to fractionation during precipitation in the surface waters or during dissolution in the water and sediments due to CaCO, undersaturation. We found extensive loss of material from unpoisoned sediment traps in the euphotic zone, most likely due to zooplankton feeding. Stable carbon isotopes have been used to study paleoproductivity and the sedimen- tary history of lake (Stuiver 1975; Fritz et al. 1975; McKenzie 1982, 1985) and ocean (e.g. see Sundquist and Broecker 1985) ba- sins. In most of these studies, the stable car- bon isotope ratios found in precipitated car- bonates were correlated with environmental changes which affected the carbon isotope ratio of the dissolved inorganic carbon (DIC) at the time of precipitation. Such a corre- lation requires assumptions about the iso- topic fractionation factors governing pre- p.- 1 Contribution 5998 from the Woods Hole Ocean- ographic Institution, Contribution 254 from the Lab- oratory for Experimental Geology, Zurich, and also a contribution from EAWAG. This research was sup- ported by the Swiss National Science Foundation (Limnogeology Project), the U.S. Office of Naval Re- search, and the U.S. National Science Foundation (OCE 83-00018). * Present address: Marine Science Research Center, SUNY, Stony Brook, New York 11794. cipitation. These enrichment factors appear to be better known for inorganic carbonate precipitation (Emrich et al. 1970; Turner 1982) than for biogenic carbonate forma- tion (Grossman 1984). McKenzie (1982, 198 5) conducted paleoproductivity studies in Lake Greifen, a hard-water, eutrophic Swiss lake where inorganic calcite precipi- tates seasonally during times of high bio- logical productivity. We present results here from a short-term sediment trap experi- ment in Lake Greifen which show the re- lationships between biological activity and the isotopic composition of DIC and be- tween the carbon isotope ratios of calcite in the water column and in the sediments. The carbon isotope composition of DIC in lakes is influenced internally by the de- gree of biological activity in the water and externally by the isotopic signal of DIC in the rivers and groundwaters feeding the lake (Oana and Deevey 1960; Takahashi et al. 1968; Rau 1978; McKenzie 1982, 1985). 83
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Carbon isotope fractionation and changes in the flux and composition of particulate matter resulting...

Limnol. Oceanogr., 32(l), 1987, 83-96 0 1987, by the American Society of Limnology and Oceanography, Inc.
Carbon isotope fractionation and changes in the flux and composition of particulate matter resulting from biological activity during a sediment trap experiment in Lake Greifen, Switzerland’
Cindy Lee2 Woods Hole Oceanographic Institution, Woods Hole, Massachusetts 02543
Judith A. McKenzie Geological Institute, Swiss Federal Institute of Technology, 8092 Zurich
Michael Sturm Federal Institute for Water Resources and Water Pollution Control (EAWAG), Swiss Federal Institute of Technology, 8600 Diibendorf
Abstract
Sediment traps were deployed in Lake Greifen, a hard-water eutrophic, Swiss lake, for 2 weeks in May and June 1983. Oxygen, pH, and suspended particle concentrations indicated a sharp increase in productivity during the experiment. Carbon isotope composition of the dissolved inorganic carbon recorded the balance between photosynthesis and respiration and showed possible evidence of difhtsion of atmospheric CO, across the water-atmosphere interface, most likely to replace CO, used during photosynthesis. Allochthonous mineral debris from resuspended sediments or river input was a major component of the sedimenting material. Due to removal of COz, large, newly formed calcite crystals precipitated when photosynthesis increased. The carbon isotope ratio of trap calcite and the composition of trap amino acids showed the influence of the detrital input on the more dominant isotopic signal of authigenic calcite. Sediment calcite is enriched in 13C relative to authigenic calcite in the sediment traps. This enrichment is likely due to fractionation during precipitation in the surface waters or during dissolution in the water and sediments due to CaCO, undersaturation. We found extensive loss of material from unpoisoned sediment traps in the euphotic zone, most likely due to zooplankton feeding.
Stable carbon isotopes have been used to study paleoproductivity and the sedimen- tary history of lake (Stuiver 1975; Fritz et al. 1975; McKenzie 1982, 1985) and ocean (e.g. see Sundquist and Broecker 1985) ba- sins. In most of these studies, the stable car- bon isotope ratios found in precipitated car- bonates were correlated with environmental changes which affected the carbon isotope ratio of the dissolved inorganic carbon (DIC) at the time of precipitation. Such a corre- lation requires assumptions about the iso- topic fractionation factors governing pre-
p.- 1 Contribution 5998 from the Woods Hole Ocean-
ographic Institution, Contribution 254 from the Lab- oratory for Experimental Geology, Zurich, and also a contribution from EAWAG. This research was sup- ported by the Swiss National Science Foundation (Limnogeology Project), the U.S. Office of Naval Re- search, and the U.S. National Science Foundation (OCE 83-00018).
* Present address: Marine Science Research Center, SUNY, Stony Brook, New York 11794.
cipitation. These enrichment factors appear to be better known for inorganic carbonate precipitation (Emrich et al. 1970; Turner 1982) than for biogenic carbonate forma- tion (Grossman 1984). McKenzie (1982, 198 5) conducted paleoproductivity studies in Lake Greifen, a hard-water, eutrophic Swiss lake where inorganic calcite precipi- tates seasonally during times of high bio- logical productivity. We present results here from a short-term sediment trap experi- ment in Lake Greifen which show the re- lationships between biological activity and the isotopic composition of DIC and be- tween the carbon isotope ratios of calcite in the water column and in the sediments.
The carbon isotope composition of DIC in lakes is influenced internally by the de- gree of biological activity in the water and externally by the isotopic signal of DIC in the rivers and groundwaters feeding the lake (Oana and Deevey 1960; Takahashi et al. 1968; Rau 1978; McKenzie 1982, 1985).
83
84 Lee et al.
-13 -12 -II -10 -9 -8 -7 DIC SlJCp& Pko/
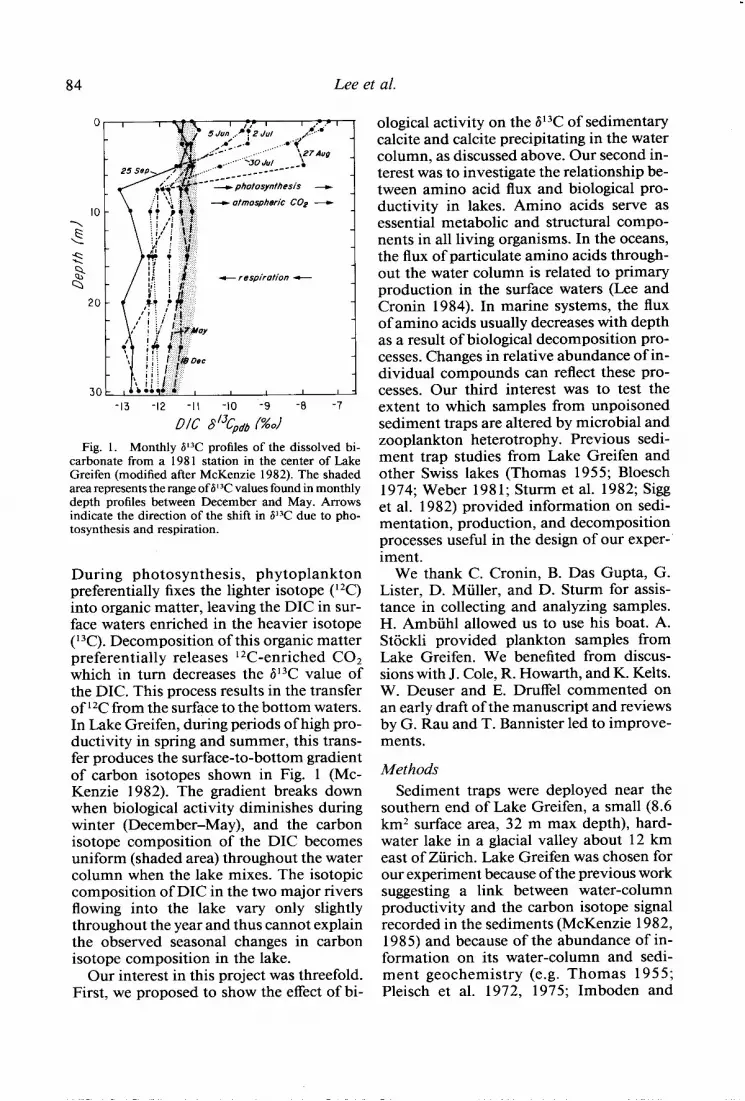
Fig. 1. Monthly P3C profiles of the dissolved bi- carbonate from a 198 1 station in the center of Lake Greifen (modified after McKenzie 1982). The shaded area represents the range of P3C values found in monthly depth profiles between December and May. Arrows indicate the direction of the shift in P3C due to pho- tosynthesis and respiration.
During photosynthesis, phytoplankton preferentially fixes the lighter isotope ( 12C) into organic matter, leaving the DIC in sur- face waters enriched in the heavier isotope (l 3C). Decomposition of this organic matter preferentially releases l 2C-enriched CO, which in turn decreases the 613C value of the DIC. This process results in the transfer of 12C from the surface to the bottom waters. In Lake Greifen, during periods of high pro- ductivity in spring and summer, this trans- fer produces the surface-to-bottom gradient of carbon isotopes shown in Fig. 1 (Mc- Kenzie 1982). The gradient breaks down when biological activity diminishes during winter (December-May), and the carbon isotope composition of the DIC becomes uniform (shaded area) throughout the water column when the lake mixes. The isotopic composition of DIC in the two major rivers flowing into the lake vary only slightly throughout the year and thus cannot explain the observed seasonal changes in carbon isotope composition in the lake.
Our interest in this project was threefold. First, we proposed to show the effect of bi-
ological activity on the 613C of sedimentary calcite and calcite precipitating in the water column, as discussed above. Our second in- terest was to investigate the relationship be- tween amino acid flux and biological pro- ductivity in lakes. Amino acids serve as essential metabolic and structural compo- nents in all living organisms. In the oceans, the flux of particulate amino acids through- out the water column is related to primary production in the surface waters (Lee and Cronin 1984). In marine systems, the flux of amino acids usually decreases with depth as a result of biological decomposition pro- cesses. Changes in relative abundance of in- dividual compounds can reflect these pro- cesses. Our third interest was to test the extent to which samples from unpoisoned sediment traps are altered by microbial and zooplankton heterotrophy. Previous sedi- ment trap studies from Lake Greifen and other Swiss lakes (Thomas 1955; Bloesch 1974; Weber 1981; Sturm et al. 1982; Sigg et al. 1982) provided information on sedi- mentation, production, and decomposition processes useful in the design of our exper- iment .
We thank C. Cronin, B. Das Gupta, G. Lister, D. Miiller, and D. Sturm for assis- tance in collecting and analyzing samples. H. Ambiihl allowed us to use his boat. A. Stiickli provided plankton samples from Lake Greifen. We benefited from discus- sions with J. Cole, R. Howarth, and K. Kelts. W. Deuser and E. Druffel commented on an early draft of the manuscript and reviews by G. Rau and T. Bannister led to improve- ments.
Methods Sediment traps were deployed near the
southern end of Lake Greifen, a small (8.6 km2 surface area, 32 m max depth), hard- water lake in a glacial valley about 12 km east of Zurich. Lake Greifen was chosen for our experiment because of the previous work suggesting a link between water-column productivity and the carbon isotope signal recorded in the sediments (McKenzie 1982, 1985) and because of the abundance of in- formation on its water-column and sedi- ment geochemistry (e.g. Thomas 1955; Pleisch et al. 1972, 1975; Imboden and
Swiss sediment trap experiment 85
0 2 4 6 8 10 12 14 16 18
p/f 6 65 7 75 8 85 9 I I I 1 I
Tf’C) 5 6 7 8 9 IO il i2 i4 , , , , , , , ,
i3 ,’ ,
2;. 15
02fmg 1-7 0 2 4 6 8 10 12 14 i6 18 o> ” ’ ” ” ‘.I
0, T//’ 1 ‘PM I’
2-
- trap A-
4- :
I 0 0
: ,,,‘/
8-
10 i
A 12
I
10 I
I s
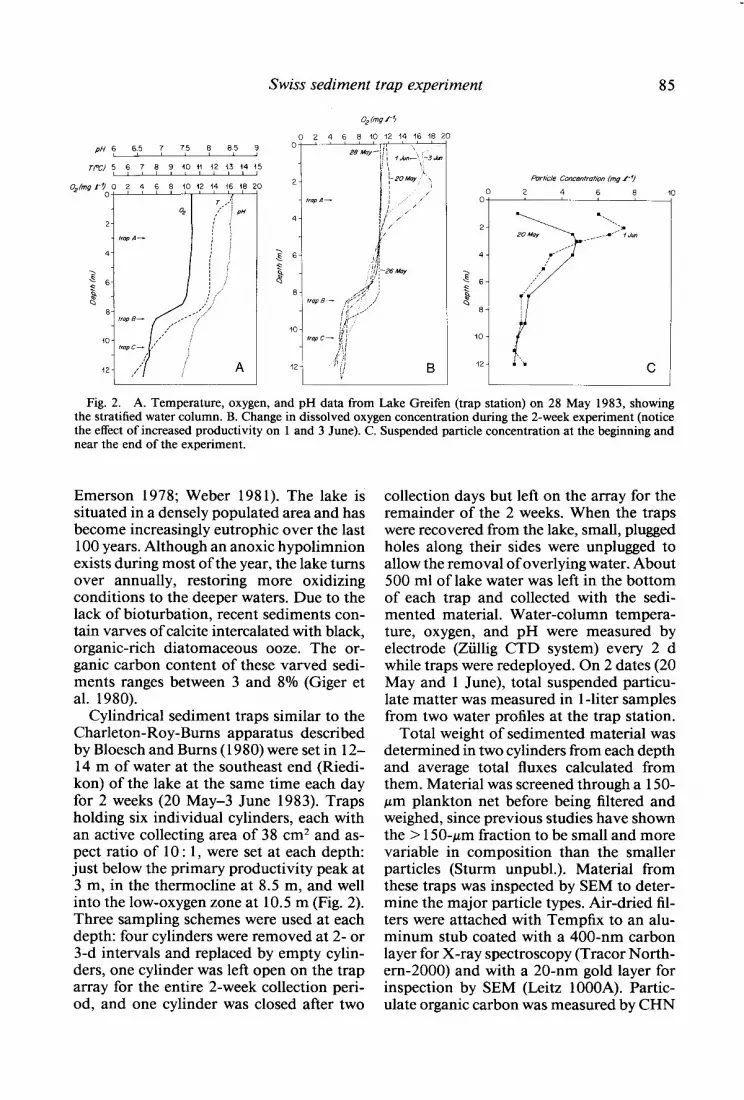
Fig. 2. A. Temperature, oxygen, and pH data from Lake Greifen (trap station) on 28 May 1983, showing the stratified water column. B. Change in dissolved oxygen concentration during the 2-week experiment (notice the effect of increased productivity on 1 and 3 June). C. Suspended particle concentration at the beginning and near the end of the experiment.
Emerson 1978; Weber 198 1). The lake is situated in a densely populated area and has become increasingly eutrophic over the last 100 years. Although an anoxic hypolimnion exists during most of the year, the lake turns over annually, restoring more oxidizing conditions to the deeper waters. Due to the lack of bioturbation, recent sediments con- tain varves of calcite intercalated with black, organic-rich diatomaceous ooze. The or- ganic carbon content of these varved sedi- ments ranges between 3 and 8% (Giger et al. 1980).
Cylindrical sediment traps similar to the Charleton-Roy-Bums apparatus described by Bloesch and Bums (1980) were set in 12- 14 m of water at the southeast end (Riedi- kon) of the lake at the same time each day for 2 weeks (20 May-3 June 1983). Traps holding six individual cylinders, each with an active collecting area of 38 cm2 and as- pect ratio of 10 : 1, were set at each depth: just below the primary productivity peak at 3 m, in the thermocline at 8.5 m, and well into the low-oxygen zone at 10.5 m (Fig. 2). Three sampling schemes were used at each depth: four cylinders were removed at 2- or 3-d intervals and replaced by empty cylin- ders, one cylinder was left open on the trap array for the entire 2-week collection peri- od, and one cylinder was closed after two
collection days but left on the array for the remainder of the 2 weeks. When the traps were recovered from the lake, small, plugged holes along their sides were unplugged to allow the removal of overlying water. About 500 ml of lake water was left in the bottom of each trap and collected with the sedi- mented material. Water-column tempera- ture, oxygen, and pH were measured by electrode (Ziillig CTD system) every 2 d while traps were redeployed. On 2 dates (20 May and 1 June), total suspended particu- late matter was measured in l-liter samples from two water profiles at the trap station.
Total weight of sedimented material was determined in two cylinders from each depth and average total fluxes calculated from them. Material was screened through a 150- pm plankton net before being filtered and weighed, since previous studies have shown the > 150-pm fraction to be small and more variable in composition than the smaller particles (Sturm unpubl.). Material from these traps was inspected by SEM to deter- mine the major particle types. Air-dried fil- ters were attached with Tempfix to an alu- minum stub coated with a 400-nm carbon layer for X-ray spectroscopy (Tracer North- em-2000) and with a 20-nm gold layer for inspection by SEM (Leitz 1000A). Partic- ulate organic carbon was measured by CHN
86 Lee et al.
Collection Period
16
12
Time ldqs-1
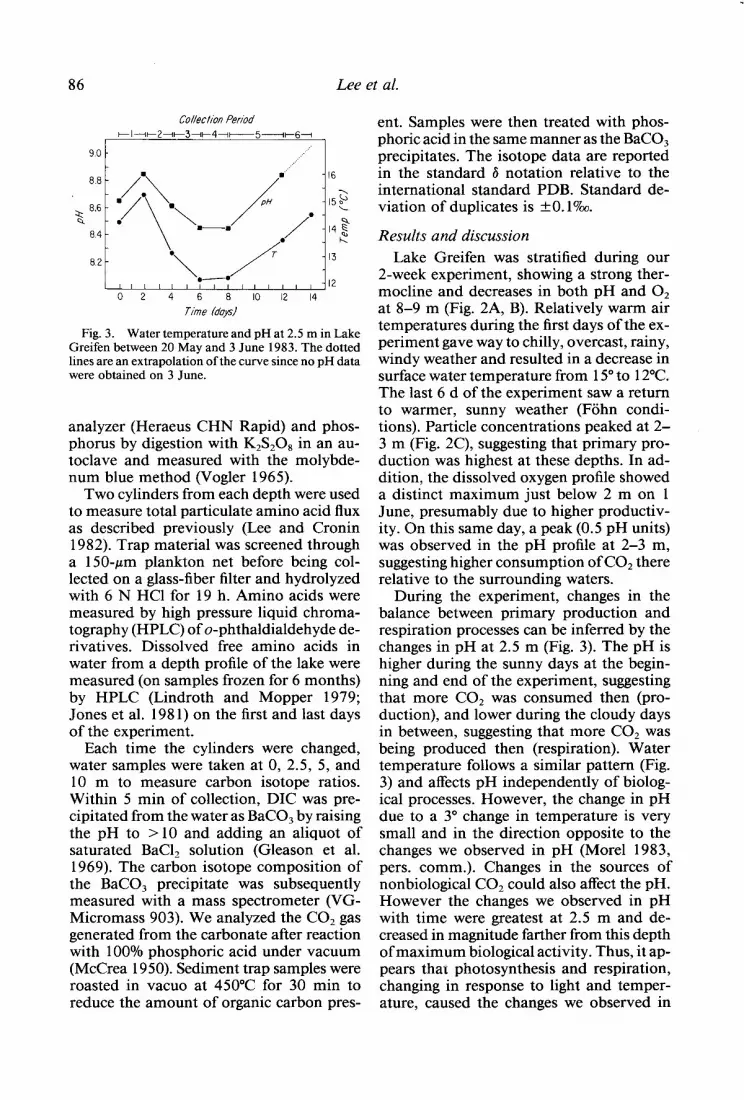
Fig. 3. Water temperature and pH at 2.5 m in Lake Greifen between 20 May and 3 June 1983. The dotted lines are an extrapolation of the curve since no pH data were obtained on 3 June.
analyzer (Heraeus CHN Rapid) and phos- phorus by digestion with K&O8 in an au- toclave and measured with the molybde- num blue method (Vogler 1965).
Two cylinders from each depth were used to measure total particulate amino acid flux as described previously (Lee and Cronin 1982). Trap material was screened through a 150-pm plankton net before being col- lected on a glass-fiber filter and hydrolyzed with 6 N HCl for 19 h. Amino acids were measured by high pressure liquid chroma- tography (HPLC) of o-phthaldialdehyde de- rivatives. Dissolved free amino acids in water from a depth profile of the lake were measured (on samples frozen for 6 months) by HPLC (Lindroth and Mopper 1979; Jones et al. 198 1) on the first and last days of the experiment.
Each time the cylinders were changed, water samples were taken at 0, 2.5, 5, and 10 m to measure carbon isotope ratios. Within 5 min of collection, DIC was pre- cipitated from the water as BaCO, by raising the pH to > 10 and adding an aliquot of saturated BaCl, solution (Gleason et al. 1969). The carbon isotope composition of the BaCO, precipitate was subsequently measured with a mass spectrometer (VG- Micromass 903). We analyzed the CO2 gas generated from the carbonate after reaction with 100% phosphoric acid under vacuum (McCrea 1950). Sediment trap samples were roasted in vacua at 450°C for 30 min to reduce the amount of organic carbon pres-
ent. Samples were then treated with phos- phoric acid in the same manner as the BaCO, precipitates. The isotope data are reported in the standard 6 notation relative to the international standard PDB. Standard de- viation of duplicates is k-0. lY&
Results and discussion Lake Greifen was stratified during our
2-week experiment, showing a strong ther- mocline and decreases in both pH and O2 at 8-9 m (Fig. 2A, B). Relatively warm air temperatures during the first days of the ex- periment gave way to chilly, overcast, rainy, windy weather and resulted in a decrease in surface water temperature from 15” to 12°C. The last 6 d of the experiment saw a return to warmer, sunny weather (Fiihn condi- tions). Particle concentrations peaked at 2- 3 m (Fig. 2C), suggesting that primary pro- duction was highest at these depths. In ad- dition, the dissolved oxygen profile showed a distinct maximum just below 2 m on 1 June, presumably due to higher productiv- ity. On this same day, a peak (0.5 pH units) was observed in the pH profile at 2-3 m, suggesting higher consumption of CO2 there relative to the surrounding waters.
During the experiment, changes in the balance between primary production and respiration processes can be inferred by the changes in pH at 2.5 m (Fig. 3). The pH is higher during the sunny days at the begin- ning and end of the experiment, suggesting that more CO2 was consumed then (pro- duction), and lower during the cloudy days in between, suggesting that more CO2 was being produced then (respiration). Water temperature follows a similar pattern (Fig. 3) and affects pH independently of biolog- ical processes. However, the change in pH due to a 3” change in temperature is very small and in the direction opposite to the changes we observed in pH (Morel 1983, pers. comm.). Changes in the sources of nonbiological CO2 could also affect the pH. However the changes we observed in pH with time were greatest at 2.5 m and de- creased in magnitude farther from this depth of maximum biological activity. Thus, it ap- pears that photosynthesis and respiration, changing in response to light and temper- ature, caused the changes we observed in
Swiss sediment trap experiment 87
24 3m
4 123456
E
@
S.5m
8 0.5
A 0 B 0 123456 123456 123456
Co//ection Pefiod Colection Period
15 1 1 3m 3m
05-
s k 123456 123456 s
9 k 1.5 1 3 or 50 -- $2 E I 40 X5m
16 Q 1
Collection Period
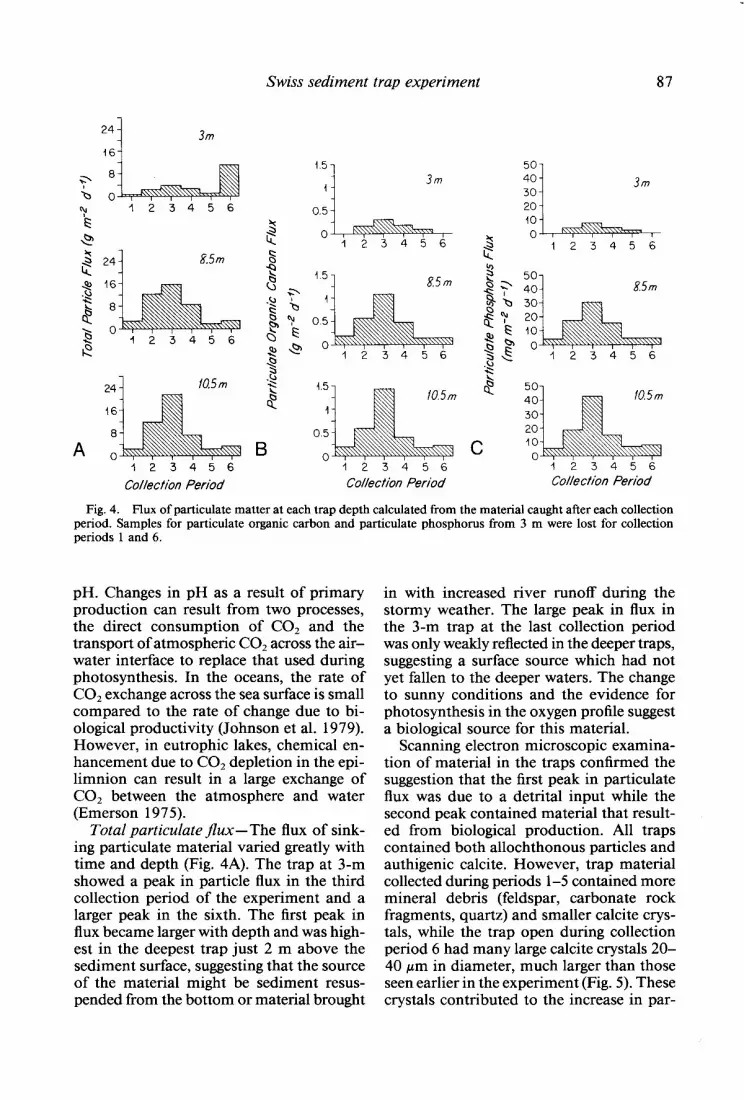
Fig. 4. Flux of particulate matter at each trap depth calculated from the material caught after each collection period. Samples for particulate organic carbon and particulate phosphorus from 3 m were lost for collection periods 1 and 6.
pH. Changes in pH as a result of primary production can result from two processes, the direct consumption of CO2 and the transport of atmospheric CO, across the air- water interface to replace that used during photosynthesis. In the oceans, the rate of CO, exchange across the sea surface is small compared to the rate of change due to bi- ological productivity (Johnson et al. 1979). However, in eutrophic lakes, chemical en- hancement due to CO, depletion in the epi- limnion can result in a large exchange of CO, between the atmosphere and water (Emerson 1975).
Total particulate flux-The flux of sink- ing particulate material varied greatly with time and depth (Fig. 4A). The trap at 3-m showed a peak in particle flux in the third collection period of the experiment and a larger peak in the sixth. The first peak in flux became larger with depth and was high- est in the deepest trap just 2 m above the sediment surface, suggesting that the source of the material might be sediment resus- pended from the bottom or material brought
in with increased river runoff during the stormy weather. The large peak in flux in the 3-m trap at the last collection period was only weakly reflected in the deeper traps, suggesting a surface source which had not yet fallen to the deeper waters. The change to sunny conditions and the evidence for photosynthesis in the oxygen profile suggest a biological source for this material.
Scanning electron microscopic examina- tion of material in the traps confirmed the suggestion that the first peak in particulate flux was due to a detrital input while the second peak contained material that result- ed from biological production. All traps contained both allochthonous particles and authigenic calcite. However, trap material collected during periods l-5 contained more mineral debris (feldspar, carbonate rock fragments, quartz) and smaller calcite crys- tals, while the trap open during collection period 6 had many large calcite crystals 20- 40 pm in diameter, much larger than those seen earlier in the experiment (Fig. 5). These crystals contributed to the increase in par-

Swiss sediment trap experiment 89
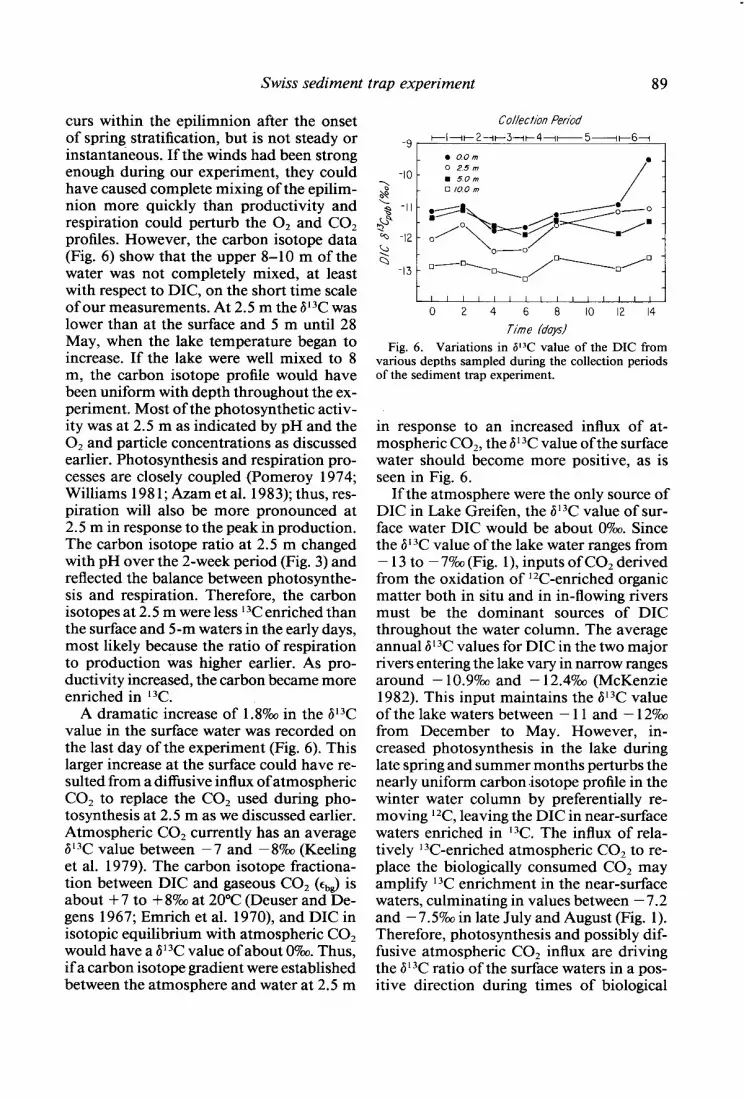
curs within the epilimnion after the onset of spring stratification, but is not steady or instantaneous. If the winds had been strong enough during our experiment, they could have caused complete mixing of the epilim- nion more quickly than productivity and respiration could perturb the 0, and CO2 profiles. However, the carbon isotope data (Fig. 6) show that the upper 8-10 m of the water was not completely mixed, at least with respect to DIC, on the short time scale of our measurements. At 2.5 m the 613C was lower than at the surface and 5 m until 28 May, when the lake temperature began to increase. If the lake were well mixed to 8 m, the carbon isotope profile would have been uniform with depth throughout the ex- periment. Most of the photosynthetic activ- ity was at 2.5 m as indicated by pH and the O2 and particle concentrations as discussed earlier. Photosynthesis and respiration pro- cesses are closely coupled (Pomeroy 1974; Williams 198 1; Azam et al. 1983); thus, res- piration will also be more pronounced at 2.5 m in response to the peak in production. The carbon isotope ratio at 2.5 m changed with pH over the 2-week period (Fig. 3) and reflected the balance between photosynthe- sis and respiration. Therefore, the carbon isotopes at 2.5 m were less 13C enriched than the surface and 5-m waters in the early days, most likely because the ratio of respiration to production was higher earlier. As pro- ductivity increased, the carbon became more enriched in 13C
A dramatic increase of 1.8%~~ in the 613C value in the surface water was recorded on the last day of the experiment (Fig. 6). This larger increase at the surface could have re- sulted from a diffusive influx of atmospheric CO, to replace the CO, used during pho- tosynthesis at 2.5 m as we discussed earlier. Atmospheric CO, currently has an average 613C value between - 7 and - 8%0 (Keeling et al. 1979). The carbon isotope fractiona- tion between DIC and gaseous CO, (~,d is about + 7 to + 8%0 at 20°C (Deuser and De- gens 1967; Emrich et al. 1970), and DIC in isotopic equilibrium with atmospheric CO2 would have a ii13C value of about 07~. Thus, if a carbon isotope gradient were established between the atmosphere and water at 2.5 m
Collection Period
-9 [ I-III-2-1~3---1~41~5---11-6-1
-10 - a s L -II- 8 @ - -& -12 - 4 Q
-13 -
tl I I I I I I I I I I I I I Ii
0 2 4 6 8 IO I2 I4
Time fdayd
Fig. 6. Variations in P3C value of the DIC from various depths sampled during the collection periods of the sediment trap experiment.
in response to an increased influx of at- mospheric C02, the 6 l 3C value of the surface water should become more positive, as is seen in Fig. 6.
If the atmosphere were the only source of DIC in Lake Greifen, the 613C value of sur- face water DIC would be about 0%~ Since the 6 l 3C value of the lake water ranges from - 13 to - 7% (Fig. l), inputs of CO, derived from the oxidation of 12C-enriched organic matter both in situ and in in-flowing rivers must be the dominant sources of DIC throughout the water column. The average annual 613C values for DIC in the two major rivers entering the lake vary in narrow ranges around - 10.9%~ and - 12.4700 (McKenzie 1982). This input maintains the 613C value of the lake waters between - 11 and - 127~ from December to May. However, in- creased photosynthesis in the lake during late spring and summer months perturbs the nearly uniform carbon isotope profile in the winter water column by preferentially re- moving l 2C, leaving the DIC in near-surface waters enriched in 13C. The influx of rela- tively 13C-enriched atmospheric CO2 to re- place the biologically consumed CO2 may amplify 13C enrichment in the near-surface waters, culminating in values between - 7.2 and - 7.5?& in late July and August (Fig. 1). Therefore, photosynthesis and possibly dif- fusive atmospheric CO2 influx are driving the 6 l 3C ratio of the surface waters in a pos- itive direction during times of biological

90 Lee et al.
productivity. In oligotrophic lakes where the effects of biological production are not as strong, river and groundwater input of DIC can be the more important influences on carbon isotope signal (Rau 1978).
Isotopic signal in calcite-The 613C val- ues of calcite collected in sediment traps could only be measured during the last col- lection period, when authigenic calcite pre- cipitation was high. Insufficient authigenic material was collected during the other trap deployments. Carbon isotope ratios during this last period were enriched in 13C relative to DIC values and became more enriched with depth: -8.34Ymat 3 m, -7.49Ymat 8.5 m, and -7.23YZ~ at 10.5 m. Part of the en- richment of the calcite relative to DIC is expected, due to equilibrium fractionation during precipitation. Grossman (1984) re- cently recalculated the Emrich et al. (1970) temperature-dependent enrichment rela- tionship to take into consideration fraction- ation differences between aragonite and cal- cite and Turner’s (1982) newer values for ccg (between calcium carbonate and gaseous C02). During the last trap collection period in Lake Greifen, the temperature averaged 14°C at 2.5 m, the depth of maximum pro- ductivity where most of the calcite would have precipitated. This temperature would cause a small enrichment of O-13?& in calcite relative to DIC according to Grossman’s equation.
If we assume that calcite was enriched in 13C by 0.13?& during precipitation at the time of our measurements, the trap calcite was still enriched by 2.6-3.7?&~ (the DIC ratio averaged - ll.O8o/oo), with the enrich- ment larger in the deeper traps. There are two possible explanations. The enrichment may show the extent to which the authigenic calcite is diluted by detrital carbonates from the catchment basin. A detrital influence would raise the 613C value measured in the trap since detrital carbonates in the incom- ing rivers are relatively enriched in 13C (-2o/oo: McKenzie unpubl. data). A simple mass balance allows us to calculate that 29% of the calcite in the 3-m trap is detrital rath- er than authigenic. This calculation assumes a trap 613C value of -8.34Y& a detrital car- bonate value of -~V&J, and an authigenic carbonate value of - 10.957~ (DIC correct-
ed for fractionation during precipitation). Similar calculations show detrital inputs of 35% at 8.5 m and 38% at 10.5 m. A second possible source of material enriched in 13C is the sediment. If surface sediments were being resuspended, the traps could reflect a mixture of authigenic calcite (- 10.95Ym) and surface sediment (- 6?&: McKenzie 1982). A mass balance suggests that for resuspen- sion to be the sole source of 13C enrichment in the trap material, 53% of the calcite in the 3-m trap would have to come from the sediments, 70% at 8.5 m, and 75% at 10.5 m. Either resuspension or river input of de- trital material could cause the increase in enrichment with depth. However, SEM in- spection of trap material from collection pe- riod 6 (Fig. 5 and unpubl.) showed a small contribution of nonauthigenic calcite. And in similar trap experiments in Lake Greifen during May and June 1978, Weber (1981) found detrital carbonate contributions of 1 O- 20% at 3 m. Thus, we believe that the en- richment of l 3C in trap carbonate during our experiment is most likely due to an input from detrital rather than resuspended ma- terial.
Turner (1982) found that fractionation during calcite precipitation was dependent on reaction kinetics and suggested that pre- vious measurements may have underesti- mated the extent of fractionation for slowly precipitating systems. Enrichment of 13C in calcite was much greater when the precip- itation rate was very slow, but was small and constant at faster rates. Turner found that calcite was enriched by 3-47~ relative to DIC in lakes slowly precipitating calcite in Australia and Canada. Given the pres- ence of detrital carbonates in the sediment trap material we collected, isotopic frac- tionation during calcite precipitation at the time of our experiment was most likely small, close to the 0.137~ calculated from Grossman’s equation. This result suggests that precipitation of calcite was rapid during period 6 when we collected our sample. However, it is possible that slower precip- itation occurs at other times of the year and may affect the isotopic signal recorded in the sediments.
Isotopic signal in DIC vs. sediments- McKenzie (1982) found sediment calcite in

Swiss sediment trap experiment 91
Lake Greifen to have an average 6’ 3C of - 7.1o/oo over the past 200 yr, with values of - 5 to - 6. ~Y&J more common in the past 50 yr when the lake was more eutrophic. These sediment values were corrected for the river input of detrital carbonate. As mentioned earlier, detrital carbonates (smaller crystals in Fig. 5 j are enriched in 13C (613C = - 2o/oo) relative to authigenic calcite (613C = - 10.95%0); thus, dilution of the sediment with this detrital material can raise the 613C value. Given that the sediment values above reflect only the calcite precipitating in the lake, they are quite enriched in 13C relative to modem values for DIC. As Fig. 1 shows, recently measured carbon isotope ratios for DIC are around - 1 10/00 for much of the year and are never more positive than - 7Vm even during the highest summer production. One possible explanation for this enrichment in 13C in the sediments over DIC is that much of the calcite in the lake sediments was slow- ly precipitated and enriched in 13C by - 3 or -4o/oo, as Turner found in the lakes he studied. This explanation might be likely if much of the calcite in the sediments was precipitated chemically throughout the year. However, Weber (198 1) found that most of the authigenic calcite precipitation in Lake Greifen occurred during periods of peak productivity during spring and summer. An alternative explanation for enrichment of sediment calcite is that perhaps more ex- treme excursions in carbon isotope ratio oc- cur during photosynthesis than were mea- sured by McKenzie (1982). If a substantial amount of the calcite in the sediment were formed during short-term extreme periods of calcite supersaturation (and thus possibly carbonate precipitation), enrichment in 13C could result. The trend toward 13C-enriched sediment calcite over the past 50 yr could result from either of these processes. With increasing eutrophication, higher phos- phate concentrations which can inhibit cal- cite precipitation could have slowed the pre- cipitation rate, thus allowing more fractionation to occur. Or, as eutrophication increased, higher incursions of atmospheric CO2 as a result of depletion of DIC during photosynthesis could have resulted in en- richment of 13C in the surface waters and thus of precipitating calcite.
It is also possible that diagenetic pro- cesses could explain both the 13C enrich- ment in the sediment relative to DIC and the increasing 13C enrichment of sediment trap calcite with depth. Preferential loss of the lighter isotope, 12C, may occur during calcite dissolution. As calcite crystals settle through the water column, they pass through a hypolimnion which is less CaCO, satu- rated than the surface waters; theoretically, they could partially dissolve and become enriched in 13C. However, Kelts and Hsii (1978) estimated that only 10% of a calcite crystal is dissolved as it falls through the 130-m water column of Lake Zurich. Erez ( 1978) found only a small isotopic fraction- ation of carbon due to dissolution of fora- minifera settling through an oceanic water column. Thus, the 1.1%0 difference we ob- served in trap calcite 613C between 3 and 10.5 m is more likely caused by detrital in- put than by dissolution. In the sediments, however, calcite dissolution can be more extensive. Emerson (1976) found that when authigenic calcite precipitating in Lake Greifen waters reached the sediments, it rapidly dissolved in interstitial waters rich in CO, from organic matter oxidation. Dis- solution occurred in the top 4 cm of sedi- ment which were undersaturated with re- spect to calcite, while pore waters from sediments deeper than 4 cm had reached equilibrium with calcite; Emerson also found that the top 50 cm of Lake Greifen sedi- ments were > 50% CaCO,. We do not know whether sufficient dissolution of authigenic calcite (formed from 613C of DIC between - 7 and - 11 o/00) occurs in the sediments or whether this dissolution causes sufficient 13C enrichment to produce a sediment calcite 613C of -6%0. This is difficult to determine; measuring changes in the a1 3C of pore-water CO, will not differentiate between CO, pro- duced during dissolution of CaCO, and CO2 produced during the oxidation of 12C-enriched organic matter, since both would serve to enrich the pore waters in 12C. This question must be further investigated in the lake before we can determine the in- fluence of dissolution on the sedimentary history of the lake. For example, as men- tioned earlier, sedimentary calcite became increasingly enriched in 13C over the last 50
92 Lee et al.
300
200 3m
100
3 0
k 123456
123456
123456
Collection Period
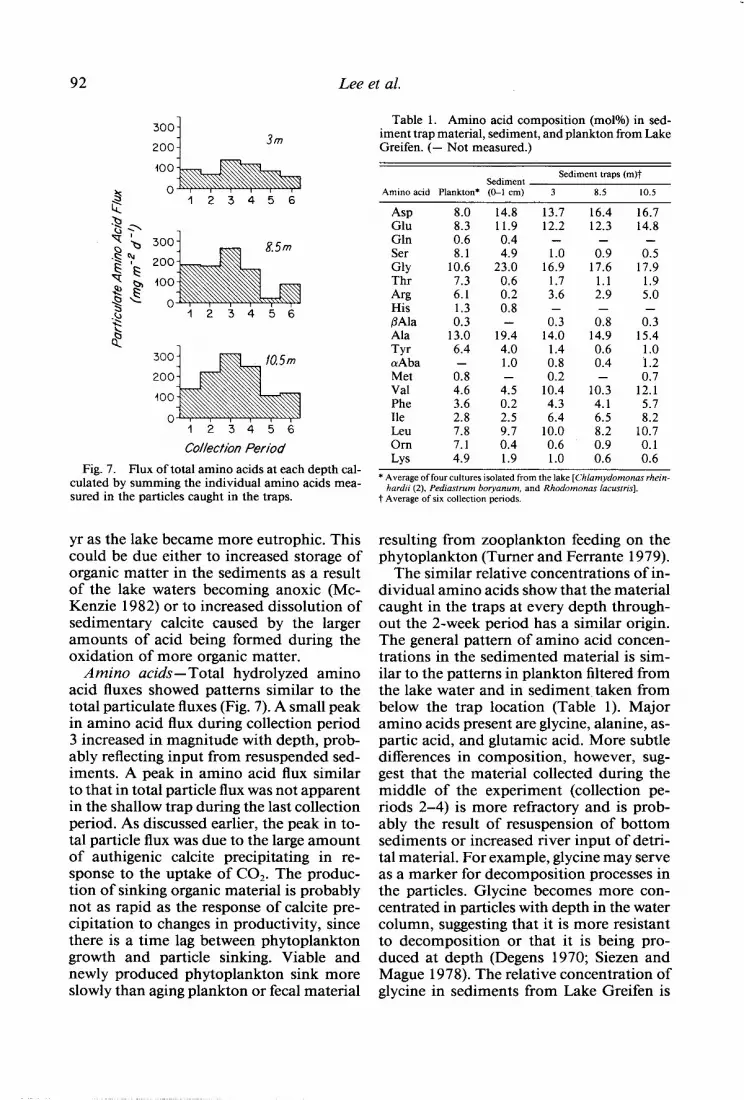
Fig. 7. Flux of total amino acids at each depth cal- culated by summing the individual amino acids mea- sured in the particles caught in the traps.
yr as the lake became more eutrophic. This could be due either to increased storage of organic matter in the sediments as a result of the lake waters becoming anoxic (Mc- Kenzie 1982) or to increased dissolution of sedimentary calcite caused by the larger amounts of acid being formed during the oxidation of more organic matter.
Amino acids- Total hydrolyzed amino acid fluxes showed patterns similar to the total particulate fluxes (Fig. 7). A small peak in amino acid flux during collection period 3 increased in magnitude with depth, prob- ably reflecting input from resuspended sed- iments. A peak in amino acid flux similar to that in total particle flux was not apparent in the shallow trap during the last collection period. As discussed earlier, the peak in to- tal particle flux was due to the large amount of authigenic calcite precipitating in re- sponse to the uptake of C02. The produc- tion of sinking organic material is probably not as rapid as the response of calcite pre- cipitation to changes in productivity, since there is a time lag between phytoplankton growth and particle sinking. Viable and newly produced phytoplankton sink more slowly than aging plankton or fecal material
Table 1. Amino acid composition (mol%) in sed- iment trap material, sediment, and plankton from Lake Greifen. (- Not measured.)
Sediment Sediment traps (m)t
Amino acid Plankton* (O-l cm) 3 8.5 10.5
Asp 8.0 14.8 13.7 16.4 16.7 Glu 8.3 11.9 12.2 12.3 14.8 Gln 0.6 0.4 - - - Ser 8.1 4.9 1.0 0.9 0.5 GUY 10.6 23.0 16.9 17.6 17.9 Thr 7.3 0.6 1.7 1.1 1.9 Arg 6.1 0.2 3.6 2.9 5.0 His 0.8 @Ala 0:: - 0; O< 0.3 Ala 13.0 19.4 14.0 14.9 15.4 Tyr 6.4 4.0 1.4 0.6 1.0 crAba - 1.0 0.8 0.4 1.2 Met 0.8 - 0.2 - 0.7 Val 4.6 4.5 10.4 10.3 12.1 Phe 3.6 0.2 4.3 4.1 5.7 Ile 2.8 2.5 6.4 6.5 8.2 LeU 7.8 9.7 10.0 8.2 10.7 Ol-ll 7.1 0.4 0.6 0.9 0.1 LYS 4.9 1.9 1.0 0.6 0.6
* Average of four cultures isolated from the lake [Chlamydomonas rhein- hardu (2), Pediastrum boryanum, and Rhodomonas lacustns].
t Average of six collection periods.
resulting from zooplankton feeding on the phytoplankton (Turner and Ferrante 1979).
The similar relative concentrations of in- dividual amino acids show that the material caught in the traps at every depth through- out the 2-week period has a similar origin. The general pattern of amino acid concen- trations in the sedimented material is sim- ilar to the patterns in plankton filtered from the lake water and in sediment, taken from below the trap location (Table 1). Major amino acids present are glycine, alanine, as- partic acid, and glutamic acid. More subtle differences in composition, however, sug- gest that the material collected during the middle of the experiment (collection pe- riods 2-4) is more refractory and is prob- ably the result of resuspension of bottom sediments or increased river input of detri- tal material. For example, glycine may serve as a marker for decomposition processes in the particles. Glycine becomes more con- centrated in particles with depth in the water column, suggesting that it is more resistant to decomposition or that it is being pro- duced at depth (Degens 1970; Siezen and Mague 1978). The relative concentration of glycine in sediments from Lake Greifen is
Swiss sediment trap experiment 93
Table 2. Relative concentration of glycine (mol gly : total mol) in sediment trap material, sediments, and algae from Lake Greifen.
Plank- Traps (m)
Sedi- ton 3 8.5 10.5 ment
Avg mol% gly 10.6 16.9* 17.6* 17.9* 23.0 Mel% gly,
collection period 3
Increase (%) 18.4 20.8 23.2
over avg, collection period 3 9 18 30
* Average of mol% in the six individual traps.
higher than in plankton or in trap material (Table 2). The average mol% of glycine in- creases with water depth, showing the in- fluence of increased reworking of material or of sediment resuspension. The mol% gly- cine was highest for each depth during col- lection period 3, with the 10.5-m sample having the same composition as the sedi- ment, 23 mol%. During collection period 3, mol% gly increased and increased more strongly at greater depths, again indicating that this collection period was most influ- enced by sediment resuspension or detrital input. The lowest mol% gly values were dur- ing collection periods 5 and 6.
Relative glycine concentrations can be used to estimate the maximum contribution of resuspended sediments to the trap ma- terial. If we assume that all the glycine in the sediment traps results from simply mix- ing the surface plankton (10.6 mol%) with sediment (23 mol%), a simple mass balance with the average mol% gly from all six traps suggests that the average contribution of re- suspended sediments can be no more than 51% at 3 m, 56% at 8.5 m, and 59% at 10.5 m. These percentages are lower (3, 35, 37%) during collection period 6 when productiv- ity was high and so much authigenic calcite was formed, and much higher (6 3,82,100%) during period 3 when particle flux was so high and resuspension was likely. During period 6, for resuspension to account for the 613C in the calcite as we discussed earlier, 53-75% of the calcite would have to come from resuspended sediments, while an input of only 29-38% of detrital material could
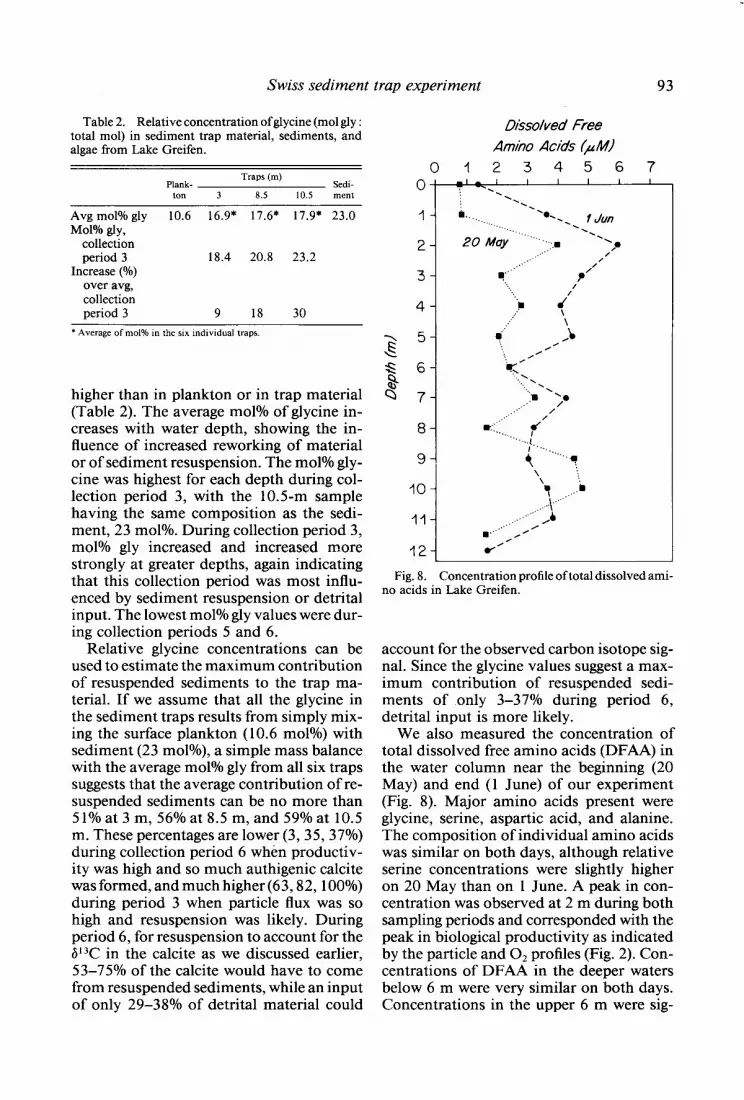
Dissolved Free Am/ho Acids (i&I)
0 1 234567 0 7’ ==.t ’
I I I I
1 - 8.. i . . . .._.
-.
‘..._ .‘..,_ =-•-.._ f&f)
2 - 20 May “’ ‘y.:.. . .
.A,. ,... ,_: /
. . . . ,
3- !:“’ t /’ ‘*..
4- “I .:. (I’ .:’ \
s 5 4“ ,b
//’ S 6- ‘K
/’ 8
‘._, \ ‘_ \ Q 7-
‘. .\ “;m ,:’ ‘f ,:’ / ,. ,’
a- =::... :” ““‘..;.. .., . . . . . .
f : ” \ ‘7 ,,,.;
,.: ,,,,_.... .\“”
. . . . A 8..““, //
/
w’
Fig. 8. Concentration profile of total dissolved ami- no acids in Lake Greifen.
account for the observed carbon isotope sig- nal. Since the glycine values suggest a max- imum contribution of resuspended sedi- ments of only 3-37% during period 6, detrital input is more likely.
We also measured the concentration of total dissolved free amino acids (DFAA) in the water column near the beginning (20 May) and end (1 June) of our experiment (Fig. 8). Major amino acids present were glycine, serine, aspartic acid, and alanine. The composition of individual amino acids was similar on both days, although relative serine concentrations were slightly higher on 20 May than on 1 June. A peak in con- centration was observed at 2 m during both sampling periods and corresponded with the peak in biological productivity as indicated by the particle and O2 profiles (Fig. 2). Con- centrations of DFAA in the deeper waters below 6 m were very similar on both days. Concentrations in the upper 6 m were sig-

Lee et al.
Table 3. Total particle flux (g m-2 d-l) from sedi- ment traps exposed over 2-week period.
Water depth of trap (m)
3 8.5 10.5
Trap open for 2 weeks* 1.8 6.5 7.5 Traps recovered every 2 dt 3.1 6.7 7.6 Recovery (%) in open trap 57 97 99
* Calculated from half the material in one open cylinder. t Average flux per day based on the total mass (average of two cylinders)
of particles caught during all six collection periods.
nificantly higher on 1 June between trap collection periods 5 and 6-the time of presumed increase in productivity. The concentration of DFAA may not so clearly reflect primary production since phyto- plankton release during photosynthesis is not the only source of DFAA, and microbial uptake of DFAA is more rapid than the removal of particulate amino acids.
Problems with the use of unpoisoned sed- iment traps -By deploying open and closed trap cylinders for the entire 2-week period, we hoped to address the question of loss from the traps by zooplankton or bacterial heterotrophy. The cylinder at each depth that was left open throughout the experi- ment should have collected as much ma- terial as the sum of the individual cylinders deployed at that depth for only 2 d. Table 3 shows that much (43%) of the material in the open surface trap was gone after 2 weeks while deeper traps did not lose as much ma- terial (2-3%). These losses can best be ex- plained by heavier zooplankton grazing on the trap particles in the surface waters. Zoo- plankton was very abundant in the surface trap cylinders during all collection periods.
Although we believe that zooplankton feeding was responsible for the major loss of material from the traps, the relative ami- no acid composition of material in the open cylinder suggests that bacteria were growing in the traps as well. One particular amino acid, P-glutamic acid, is thought to be a mi- crobial transformation product of glutamic acid in sediments (Henrichs and Farrington 1979). We found this unusual amino acid only in the 8.5- and 10.5-m cylinders that had been left open for the entire 2 weeks and which collected the most material. It is possible that these cylinders were simulating
Table 4. Particle fluxes (g m-2 d-l) from traps re- covered after 2 d and traps closed after 2 d and left in situ for the rest of the 2-week period.
Water depth of trap (m)
3 8.5 10.5
Trap closed after 2 d* 0.60 2.3 -j- Trap recovered after 2 d$ 0.49 2.8 2.8 Recovery (%) in closed trap 120 81 -
* Calculated from half the material in one cylinder. ‘/’ Sample for total particle weight was lost. $ Average of two cylinders.
sediment conditions because of the large amount of material sitting in the traps, and microbial processes were taking place which normally occur only after the particles have reached bottom.
Fluxes calculated from the cylinders closed after only 2 d of collecting are less precise than fluxes calculated from the open cylinders, since the mass of particles col- lected in the closed traps was so low that the error in our calculations becomes poten- tially large. At 3 m, the particle flux calcu- lated from the cylinder that was closed and left sitting in the lake for 12 more days ap- pears to be higher than from the cylinders that were recovered immediately (Table 4). Instead of seeing bacterial decomposition of the material resulting in a decreased flux, we see an increased flux most likely due to settling phytoplankton and zooplankton fe- cal pellets and carcasses. In the deeper trap (8.5 m), the flux calculated from the closed cylinder was lower than the flux calculated from cylinders recovered after 2 d. This may be due to enhanced bacterial decomposition in the closed cylinder. Our results indicate that over a very short period organisms can both contribute and remove material from unpoisoned sediment traps in ways that can greatly change the calculated fluxes.
Conclusions During a 2-week sediment trap experi-
ment in Lake Greifen, we observed a short- term change from lower to higher biological activity as reflected in profiles of dissolved oxygen and suspended particle concentra- tions. The composition of the inorganic matter caught in the sediment trap corre- spondingly changed from a dominant de-

Swiss sediment trap experiment
trital component to one enriched in authi- genic calcite, a by-product of photosynthetic activity. The carbon isotope composition of the dissolved inorganic carbon in the sur- face waters within the photic zone recorded the balance between photosynthesis and respiration. During periods with dimin- ished photosynthetic activity, respiration dominated and 12C-enriched CO2 was re- leased to the surface waters. When photo- synthetic activity increased, the P3C value increased due to preferential fixation of 12C into organic matter. In addition, the high 613C value of the DIC at the water-atmo- sphere interface suggested that atmospheric CO, might be diffusing across the boundary to replenish the CO2 utilized during the pe- riod of heightened photosynthetic activity. Although atmospheric CO, may be a sig- nificant component of the carbon cycle in the lake, the carbon isotope ratio of DIC throughout the water column, as measured in this study and previous work, suggests strongly that river input and carbon recy- cled in situ are the major sources. The car- bon isotope signal in sediment trap carbon- ate collected during the productive period at the end of the experiment was consistent with an input of rapidly precipitating au- thigenic calcite, with a smaller (N 30%) con- tribution from detrital sources. Amino acid data also suggested a detrital rather than resuspended-sediment source. Sediment calcite was enriched in 13C relative to DIC and to authigenic calcite in the traps. This enrichment may be due to fractionation during precipitation of calcite in the surface waters or during dissolution in the water column and sediments.
References AZAM, F., AND OTHERS. 1983. The ecological role of
water-column microbes in the sea. Mar. Ecol. Progr. Ser. 10: 257-263.
BLOESCH, J. 1974. Sedimentation und Phosphor- haushalt im Vierwaldstattersee (Horwer Bucht) und im Rotsee. Schweiz. Z. Hydrol. 36: 71-186.
-, AND N. M. BURNS. 1980. A critical review of sedimentation trap technique. Schweiz. Z. Hy- drol. 42: 15-55.
DEGENS, E. T. 1970. Molecular nature of nitrogenous compounds in seawater and recent marine sedi- ments, p. 77-106. In Organic matter in natural waters. Inst. Mar. Sci. (Alaska) Occas. Publ. 1.
DEUSER, W. G., AND E. T. DEGENS. 1967. Carbon isotope fractionation in the system CO,(gas)- CO, (aq.)-HCO,-(aq.). Nature 215: 1033-1035.
EMERSON, S. 1975. Chemically enhanced CO, gas ex- change in a eutrophic lake: A general model. Lim- nol. Oceanogr. 20: 743-753.
-. 1976. Early diagenesis in anaerobic lake sed- iments: Chemical equilibria in interstitial waters. Geochim. Cosmochim. Acta 40: 925-934.
-, AND G. WIDMER. 1978. Early diagenesis in anaerobic lake sediments- 2. Thermodynamic and kinetic factors controlling the formation of iron phosphate. Geochim. Cosmochim. Acta 42: 1307- 1316.
EMRICH, K., D. H. EHHALT, AND J. C. VOGEL. 1970. Carbon isotope fractionation during the precipi- tation of calcium carbonate. Earth Planet. Sci. Lett. 8: 363-37 1.
EREZ, J. 1978. The influence of differential produc- tion and dissolution on the stable isotope com- position of planktonic foraminifera. Ph.D. thesis, Woods Hole Oceanogr. Inst.-Mass. Inst. Technol. Program. 119 p.
FRITZ, P., T. W. ANDERSON, AND C. F. LEWIS. 197 5. Late-Quaternary climatic trends and history of Lake Erie from stable isotope studies. Science 190: 267-269.
G~HTER, R., AND A. MARES. 1985. Does settling seston release soluble reactive phosphorus in the hypolimnion of lakes? Limnol. Oceanogr. 30: 364- 371.
The flux data suggest that unpoisoned sediment traps in the euphotic zone may lose significant amounts of collected mate- rial due to zooplankton feeding, apparently at a faster rate than microbial loss. On the other hand, the lack of change in the amino acid composition in sinking particles sug- gests minimal alteration before reaching the lake bottom. Variations of the amino acid composition of material caught in the traps seem to be related to the source, i.e. resus- pended bottom material during windy pe- riods vs. settling phytoplankton during pe- riods of higher productivity.
G~GELER, H., H. W. VON GUNTEN, AND K. NYFFIZLER. 1976. Determination of 210Pb in sediments and in air samples by direct gamma-ray measurement. Earth Planet. Sci. L&t. 33: 119-121.
GIGER, W., C. SCHAFFNE R, AND S. G. WAKEHAM. 1980. Aliphatic and olefinic hydrocarbons in recent sed- iments of Greifensee, Switzerland. Geochim. Cos- mochim. Acta 44: 119-129.
GLEASON, J. O., I. FRIEDMAN, AND B. B. HANSHAW. 1969. Extraction of dissolved carbonate species from natural water for carbon-isotope analysis. U.S. Geol. Surv. Prof. Pap. 650-D, p. 248-250.
GROSSMAN, E. L. 1984. Carbon isotopic fractionation in live benthic foraminifera-comparison with in- organic precipitate studies. Geochim. Cosmo- chim. Acta 48: 1505-l 5 12.
HENRICHS, S. M., AND J. W. FARRINGTON. 1979. Ami- no acids in interstitial waters of marine sediments. Nature 279: 3 19-322.
95

96 Lee et al.
IMBODEN, D. M., AND S. EMERSON. 1978. Natural radon and phosphorus as limnologic tracers: Hor- izontal and vertical eddy diIIusion in Greifensee. Limnol. Oceanogr. 23: 77-90.
JOHNSON, K. S., R. M. PYTKOWICZ, AND C. S. WONG. 1979. Biological production and the exchange of oxygen and carbon dioxide across the sea surface in Stuart Channel, British Columbia. Limnol. Oceanogr. 24: 474-482.
JONES, B. N., S. PKABo, AND S. STEIN. 198 1. Amino acid analysis and enzymatic sequence determi- nation of peptides by an improved o-phthaldi- aldehyde precolumn labeling procedure. J. Liq. Chromatogr. 4: 565-586.
KEELING, C. D., W. G. MOOK, AND P. P. TANS. 1979. Recent trends in the 13CY2C ratio of atmospheric carbon dioxide. Nature 277: 12 l-l 23.
KELTS, K., AND K. Hsii. 1978. Freshwater carbonate sedimentation, p. 295-323. In A. Let-man [ed.], Lakes: Chemistry, geology, physics. Springer.
KUNZ, B., AND W. STUMM. 1984. Kinetik der Bildung und des Wachstums von Calciumcarbonat. Vom Wasser 62: 279-293.
LEE, C., AND C. CRONIN. 1982. The vertical flux of particulate organic nitrogen in the sea: Decom- position of amino acids in the Peru upwelling area and the equatorial Atlantic. J. Mar. Res. 40: 227- 251.
-, AND -. 1984. Particulate amino acids in the sea: Effects of primary productivity and bi- ological decomposition. J. Mar. Res. 42: 1075- 1097.
LINDROTH, P., AND K. MOPPER. 1979. High perfor- mance liquid Chromatographic determination of subpicomole amounts of amino acids by precol- umn fluorescence derivatization with o-phthal- dialdehyde. Anal. Chem. 51: 1667-l 674.
MCCREA, J. M. 1950. The isotopic chemistry of car- bonates and a paleotemperature scale. J. Chem. Phys. 18: 849-857.
MCKENZIE, J. A. 1982. Carbon- 13 cycle in Lake Grei- fen: A model for restricted ocean basins, p. 197- 207. In S. 0. Schlanger and M. B. Cita [eds.], Nature and origin of cretaceous carbon-rich facies. Academic.
-. 1985. Carbon isotopes and productivity in the lacustrine and marine environment, p. 99- 118. In W. Stumm [ed.], Chemical processes in lakes. Wiley-Interscience.
MINDER, L. 1922. Ueber biogene Entkalkung im Zii- richsee. Int. Ver. Theor. Angew. Limnol. 1: 20- 23.
MOREL, F. M. 1983. Principles of aquatic chemistry. Wiley-Interscience.
OANA, S., AND E. S. DEEVEY. 1960. Carbon- 13 in lake waters and its possible bearing on paleolimnology. Am. J. Sci. 258: 253-272.
PLEISCH, P., H. A. MATTENBERGER, AND A. PEYER. 1972, 1975. Dokumentation “Greifensee Was-
ser” Verband zum Schultze des Greifenzee. Uster, Switzerland.
POMEROY, L. R. 1974. The ocean food web, a chang- ing paradigm. Bioscience 24: 499-504.
RAU, G. 1978. Carbon-l 3 depletion in a subalpine lake: Carbon flow implications. Science 201: 90 l- 902.
SIEZEN, R. J., AND T. H. MAGUE. 1978. Amino acids in suspended particulate matter from oceanic and coastal waters of the Pacific. Mar. Chem. 6: 2 15- 231.
SIGG, L., M. STURM, W. STUMM, L. MART, AND H. W. N~NBERG. 1982. Schwermetalle im Bodensee, Mechanismen der Konzentrations-regulierung. Naturwissenschaften 69: 546-548.
STUIVER, M. 1975. Climate versus changes in 13C content of the organic component of lake sedi- ments during the Late Quaternary. Quat. Res. 5: 25 l-262.
STURM, M. 1984. Suspended particles in lakes. EAWAG News 16/17: 3-6.
-, U. ZEH, J. MILLER, L. SIGG, AND H. H. STABEL. 1982. Schwebstoffintersuchungen im Bodensee mit Interval1 - Sedimentationsfallen. Eclogae Geol. Helv. 75: 579-588.
SUNDQUIST, E., AND W. BROECKER. 1985. The carbon cycle and atmospheric CO,: Natural variations Ar- chean to Present. Geophys. Monogr. 32. Am. Geo- phys. Union.
TAKAHASHI, T., W. BROECKER, Y. H. LI, AND D. THUR- BER. 1968. Chemical and isotopic balances for a meromictic lake. Limnol. Oceanogr. 13: 272-292.
THOMAS, E. A. 1955. StolIhaushaltundSedimentation in oligotrophen Aegerisee und im eutrophen Pfaf- fiker- und Greifensee. Mem. 1st. Ital. Idrobiol. S(supp1.): 357-465.
TURNER, J. T., AND J. G. FERRANTE. 1979. Zooplank- ton fecal pellets in aquatic ecosystems. Bioscience 29: 670-677.
TURNER, J. V. 1982. Kinetic fractionation of carbon- 13 during calcium carbonate precipitation. Geo- chim. Cosmochim. Acta 46: 1183-l 19 1.
VOGLER, P. 1965. Beitrage zur Phosphatanalytik in der Limnologie. 2. Die Bestimmung des gel&ten Orthophosphates. Fortschr. Wasserchem. Ihrer Grenzgeb. 2: 109-l 19.
WEBER, H. P. 198 1. Sedimentologische und geo- chemische Untersuchungen im Greifensee (Kan- ton ZiirichSchweiz). Ph.D. thesis, Swiss Fed. Inst. Technol., Zurich. 204 p.
WETZEL, R. G. 1975. Limnology. Saunders. WILLIAMS, P. J. 198 1. Incorporation of microhetero-
trophic processes into the classical paradigm of the planktonic food web. Kiel. Meeresforsch. Son- derh. 5: l-28.
Submitted: 8 August 1985 Accepted: 5 August 1986




















