
Emotion and Attention Interaction Studied through Event-Related Potentials
Upload
independentCategory
view
0download
0
Offprint from "Marine Biology" International Journal on Life in Oceans and Coastal Waters, Vol. 14, No.2, May 1972, Pages 130-142
© by Springer- Verlag 1972 . Printed in Germany
Carbohydrases of some marine invertebrates with notes on their food and on the natural occurrence of the carbohydrates studied
J. HYLLEBERG KRISTENSEN
Zoological Institute, Laboratorium B, Ecology, University of Aarhus; Aarhus, Denmark

130 J. HYLLEBERG KRISTENSEN: Carbohydrases of ma.rine invertebrates 111ar. Bioi.
Abstract
Extracts of 22 marine invertebrates were incubated with 29 different carbobydrates, and enzyme speotra estimated from chromatograms. Enzymatic activities were quantified as reducing sugar in 14 of these species. Significant hydrolysis of laminaran, glycogen, and amylose was found in nearly all species, while hydrolysis of oligosaccharides and structural polysaccharides in most cases was weak or absent. The strongest overall degradation of carbohydrates was found among crustaceans. Enzymatic degradation of laminaran shows some relation to food, but the spectra of carbohydrases are not directly predictable from knowledge of natural food sources. Some carbohydrases (sucmse, cellobiase, lactase) could not be explained by ecological considerations. The results indicate that structural polysaccharides are utilized only to a small extent.
Introduction
FENCREL (1970) observed that bacteria and other microflora were removed from detrital particles during their passage through amphipods, while the detritus itself remained undigested. This observation, and studies by CROSSLEY et al. (1963), GEORGE (1964), NEWELL (1965), HARGRAVE (1970), and KrSSELEvA and VITYUR (1970), indicate that detritus, defined as dead organic matter, plays a minor role in the nutrition of detritus feeders.
This paper presents the in vitro activity of carbohydrases extracted from benthlc animals mainly ingesting detritus composed of inert substances (Fox, 1950) such as structural carbohydrates. The enzymatic splitting of cellulose, hemicellulose and algal poly-uronic acids was, therefore, especially studied. An attempt is made to estimate the significance of these substances as energy sources of detritus feeders by comparing the activity of carbohydrases. In addition, some commonly used oligo- and polysaccharides were included in the study to permit comparison with more readily hydrolysed substrates.
An increasing interest in carbohydrases during the last decades has resulted in a number of papers presenting different approaches. Comparative studies on degradation of carbohydrates have been carried out on groups of ecologically comparable species. NIELSEN (1962) made a qualitative comparison of
carbohydrases in 34 soil and litter invertebrates, and B. O. NIELSEN (1966) in 6 wrack invertebrates. The present work is, to my knowledge, the first of this kind which compares carbohydrases in marine invertebrates, many of which have common feeding habits. Other papers deal with the spectrum of carbohydrases of single taxonomical groups or of species; these studies deal chiefly with the quantitative aspect of enzymatic hydrolysis. Degradation has also been investigated in selected animals with respect to particular carbohydrates. Many papers deal with cellulases (references by YOKOE and YASUMASU, 1964) chitinases (extensively treated by JENIAUX, 1963), laminaranases (SOVA et a.!., 1970), and alginases (FRANSSEN and JENIAUX, 1965; FAVARO V and VASKOVSKY, 1971).
Materials and methods
For the qualitative studies, 19 detritus feeders and omnivorous species were collected during spring and early summer from the estuarine Niva Bay (Muus, 1967), i.e., the bivalves Macoma balthica (L.), Mya arenaria L., Mytilus edulis L., and Cardium edule L., the gastropods Hydrobia ulvae (PENNANT), H. ventrosa (MONTAGU), and Litto1'ina littorea L., the amphlpods Gammarus za.ddachi SEXTON, Bathyporeia pilosa LINDSTROM, B. sarsi WATKlN and Corophium vollltatar (PALLAS), the decapods Carcinus maenas L., a.nd Crangon crangon (FABR.), the isopod ldothea balthica (PALLAS), the barnacle Balanus crenatus BRUG., the polychaetes Nereis divel'sico/{)l' O. F. MULLER, Pygospio elegans CLAPAREDE, Arenicola marina L., and the oligochaete Paranais littoralis MULLER. Three species, the gastropod Nassarius reticulatus (L.) the sea star Asterias rllbens L. and the brittle star Ophiacomina nigra ABILDG., were collected in the 0resund and used for comparative purposes. They were fed mussel meat and kept for varying periods of time in aquaria. The other species were dissected shortly after collection, and digestive organs (Fig. 1) were ground at 6°C with HCl-washed quartz. Some small species (Fig. 1) were washed in buffer and homogenized in to/.().

Vol. 14, No.2, 1972 J. HYLLEBERG KRISTENSEN: Carbohydrases of marine invertebrates 131
Samples were diluted to ca. 1: 25 with citric acidphosphate buffer, pH 5.6. Some extracts were also made at pH 7.6. (Hydrobia ulvae H. ventrosa, Nassarius reticulatus and Asterias rubens) and pH 4.3. (N. reticulatus, Ophiocomina nigra and A. ru,bens).
After centrifugation, the supernatant was used as enzyme preparation and 20 fJ.I added to each of the following 29 commercial carbohydrates (about 0.5 mg of each): the <x-glucosides - maltose, sucrose and trehalose (Merck), melezitose (Sigma); the ,a-glucoside cellobiose (Merck); the <x-galactosides raffinose (Merck) melibiose (Sigma); the ,a-galactoside lactose (Merck); the polysaccharides amylose (B.D.H.), laminaran (Nutritional Biochemical Corporation, Cleveland, USA) dextran and xylan (Fluka), glycogen and cellulose (Merck), kappa carrageenan and 2 types of citrus pectin (Copenhagen Pectin Factory), carrageenan, agar-agar, furcellaran, hypnean, iridophycan, Naalginate and eucheuman (Litex, Copenhagen), chitin and hyaluronic acid (Sigma), alginic acid, gum arabic and methylcellulose (origin unknown).
Incubations were made under a cover of toluene at 32°C for 12 to 16 h. The degree of hydrolysis was estimated with paper chromatography by visual comparison with control preparations, 1 fJ.l samples were spotted on Whatman No.1 paper and run at room temperature (ca. 22 DC) with n-butanol, glacial acetic acid, and distilled water (4:1:1) as a solvent, and developed according to TREVELYAN et al. (1950).
In the quantitative study, 14 of the mentioned species (Table 1) were collected during spring and early summer along the east coast of Jutland in Kysing Fjord (Muus, 1967) and in Kal0 Vig.
Extracts were made shortly after collection, in distilled water, and filtered by serial filtration through Millipore prefilter AP 25, SS 3 fJ. and HA 0.45 fJ. freezedried (Hetosicc CD3) and stored at 5°C. No measurable decrease in activity was found after storage up to 3 months. Immediately before use, the dry powder was dissolved in a phosphate-citrate buffer, pH 5.6 or 7.6. The reaction mixtures contained 0.7 mg extract in 100 fJ.l buffer and 5 mg oligosaccharide or 0.5 mg polysaccharide. The former was added as solution, and the latter weighed on a micro balance (Mettler M5) and added as dry material.
All incubations were made at 32°C under toluene. The first measurement of reducing sugar (RS) was made within 15 min. At least 4 additional samples were analysed at intervals during the following 6 h. The samples were heat-inactivated, deproteinized and RS-measured according to NELSON (1944). After centrifugation, half of the supernatant (1 mI) was diluted to 20 mI and extinction measured at 520 nm (Beckman B). RS was calculated as glucose or Nacetyl-glucosamin equivalents (chitinolysis) after subtraction of a 9arbohydrate and an enzyme control.
Parallel incubations of identical reaction mixtures gave a standard deviation of ±8.3% (n= 10) in
17'
calculated RS. Incubations were read at constant intervals.
Methods based on alkaline copper tartrate have been commonly used in similar studies, and HARNDEN (1968) reports an accuracy higher than ±1.5% in the range of RS measured by the author. Possibly, the lower accuracy in this study is due to a higher concentration of extract used in order to ensure sufficient activity of all enzymes present in crude extracts. Thus, many enzymes were present in concentrations far exceeding the amount of protein, giving direct proportionality between enzyme concentration and hydrolysis. Specific activity was not calculated, therefore, and the absolute RS quantity was used as a measure of carbohydrase activity.
Other methods to estimate RS were tried, but neither colorimetry nor titration according to SoniOGYI (1945) gave better reproducibility.
Many enzyme preparations decreased the viscosity of pectins, mucoid, and algal colloids, but these hydrolytic reactions were not studied.
Results
<x-glucosides
Maltose
This sugar interferes with liberated glucose and, therefore, hydrolysis was studied only by chromatography (Fig. 1). Hydrolysis was strong in all incubations at pH 5.6, pH 4.3, and 7.6. Maltase was used as a control of enzyme preparations.
The main digestive function of maltase in vivo is evidently to complete break-down of maltose liberated by digestion of starch, but free maltose may be a minor constituent of the food. Maltose occurred in extracts of a red alga (NAGASHIMA et aI., 1969a) and is excreted by certain symbiotic algae (CERNICHIARI et aI., 1969). In contrast, free maltose is common in higher plants (KARRER, 1958). In animals, maltose is accumulated in a crayfish (SPECK, 1969).
Sucrose
In the qualitative study, medium or strong hydrolysis was found in most of the species (Fig. 1). The quantitative study showed low activity in Balanus crenatus and a still weaker activity in Arenicola marina. As a rule, crustaceans showed the strongest activity, e.g. ldothea balthica and Oorophium Volutator (Table 1). This relatively strong sucrose-splitting is remarkable, as there is no indication of a higher level of sucrose in their food.
The pH of 5.6 is close to the optimum value found by HORIUCIII (1963) in a marine bivalve. KOOIMAN (1964) reports that the pH optima for both maltase and sucrase ranged between pH 5.0 and 6.0. Sucrose
132 J. HYLLEBERG KRISTENSEN : Carbohydrases of marine invertebrate. Mar. Bioi.
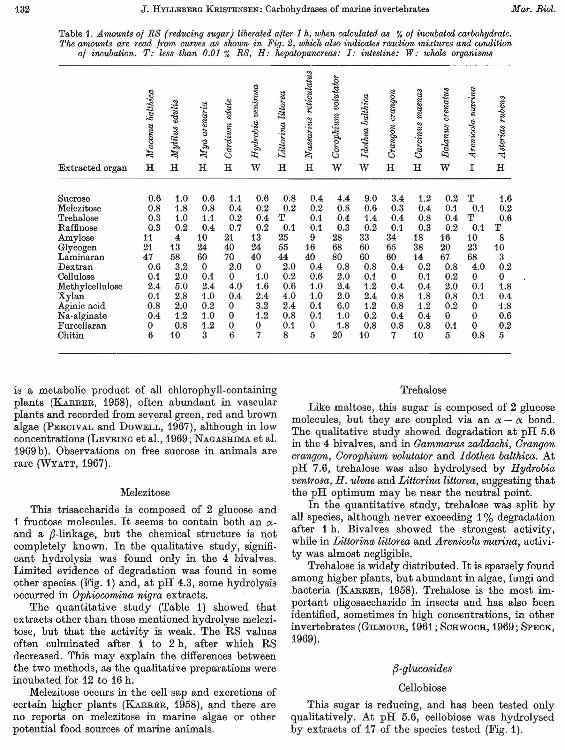
Table 1. A mounts 0/ RS (reducing sugar) liberated alter 1 h, when caku1ated as % 0/ incubated carbohydrate. The amounts are read from curves as shown in Fig. 2, which atso indicates reaction mixtures and condition
0/ incubation. T: less than 0.01 % RS, H: hepatopancreas: I: intestine: W: whole organisms
t!
.§ ~ j ~ .~ ~
~ ... .§ ~ g
.,
J Ol .~
! .~ "" " t!
~ 0{;
~ ~ Extracted organ H H H H W
Sucrose 0.6 1.0 0.6 1.1 0.6 Melezitose 0.8 1.8 0.8 0.4 0.2 Trehalose 0.3 1.0 1.1 0.2 0.4 Raffinose 0.3 0.2 0.4 0.7 0.2 Amylose 11 4 10 21 13 Glycogen 21 13 24 40 24 Laminaran 47 58 60 70 40 Dextran 0.6 3.2 0 2.0 0 Cellulose 0.1 2.0 0.1 0 1.0 Methylcellulose 2.4 5.0 2.4 4.0 1.6 Xylan 0.1 2.8 1.0 0.4 2.4 Aginic acid 0.8 2.0 0.2 0 3.2 Na·alginate 0.4 1.2 1.0 0 1.2 FurceIJaran 0 0.8 1.2 0 0 Chitin 6 10 3 6 7
is a metabolic product of all chlorophyll. containing plants (KAB.:aER, 1958), often abundant in vascular plants and recorded from several green, red and brown algae (PERCIVAL and DOWELL, 1967), although in low concentrations (LEVRING et aI., 1969; NAGASHIMA et al. 1969b). Observations on free sucrose in animals are rare (WYATT, 1967).
Melezitose
This trisaccharide is composed of 2 glucose and 1 fructose molecules. It seems to contain both an IX
and a p-linkage, but the chemical structure is not completely known. In the qualitative study, significant hydrolysis was found only in the 4 bivalves. Limited evidence of degradation was found in some other species (Fig. 1) and, at pH 4.3, some hydrolysis occurred in Ophiocomina nigra extracts.
The quantitative study (Table 1) showed that extracts other than those mentioned hydrolyse melezitose, but that the activity is weak. The RS values often culminated after 1 to 2 h, after which RS decreased. This may explain the differences between the two methods, as the qualitative preparations were incubated for 12 to 16 h.
Melezitose occurs in the cell sap and excretions of certain higher plants (KARRER, 1958), and there are no reports on melezitose in marine algae or other potential food sources of marine animals.
rl j ! I J ~
~ .§ 5 ~ lS i
.!':
::! .. .~
~ ~ f . ~
c c
.~ "" 5 '"
J .!l ...,
~ .~ .§ .~ t! 1S' §
~ ~ ~
~ ::; 8 ~ ~ t.:'3 "'i "'i H H W W H H W I H
0.8 0.4 4.4 9.0 3.4 1.2 0.2 T 1.6 0.2 0.2 0.8 0.6 0.3 0.4 0.1 0.1 0.2
T 0.1 0.4 1.4 0.4 0.8 0.4 T 0.6 0.1 0.1 0.3 0.2 0.1 0.3 0.2 0.1 T
25 9 28 33 34 18 16 10 8 55 16 68 60 65 38 20 23 10 44 40 80 60 60 14 67 68 3 2.0 0.4 0.8 0.8 0.4 0.2 0.8 4.0 0.2 0.2 0.6 2.0 0.1 0 0.1 0.2 0 0 0.6 1.0 2.4 1.2 0.4 0.4 2.0 0.1 1.8 4.0 1.0 2.0 2.4 0.8 1.8 0.8 0.1 0.4 2.4 0.1 6.0 1.2 0.8 1.2 0.2 0 1.8 0.8 0.1 1.0 0.2 0.4 0.4 0 0 0.6 0.1 0 1.8 0.8 0.8 0.8 0.1 0 0.2 8 5 20 10 7 10 5 0.8 5
Trehalose
Like maltose, this sugar is composed of 2 glucose molecules, but they are coupled via an IX - IX bond. The qualitative study showed degradation at pH 5.6 in the 4 bivalves, and in Gammarus zadr1achi, Grangon crangon, Gorophium vol;utator and ldothea balthica. At pH 7.6, trehalose was also hydrolysed by Hydrobia ventrosa, H. ulvae and Littorina littorea, suggesting that the pH optimum may be near the neutral point.
In the quantitative study, trehalose was split by all species, although never exceeding 1 % degradation after 1 h. Bivalves showed the strongest activity, while in Littorina littorea and Arenicola marina, activity was almost negligible.
Trehalose is widely distributed. It is sparsely found among higher plants, but abundant in algae, fungi and bacteria (KARRER, 1958). Trehalose is the most important oligosaccharide in insects and has also been identified, sometimes in high concentrations, in other invertebrates (GILMOUR, 1961; SCHWOCH, 1969; SPECK,
1969).
p-glucosides
Cellobiose
This sugar is reducing, and has been tested only qualitatively. At pH 5.6, cellobiose was hydrolysed by extracts of 17 of the species tested (Fig. 1).
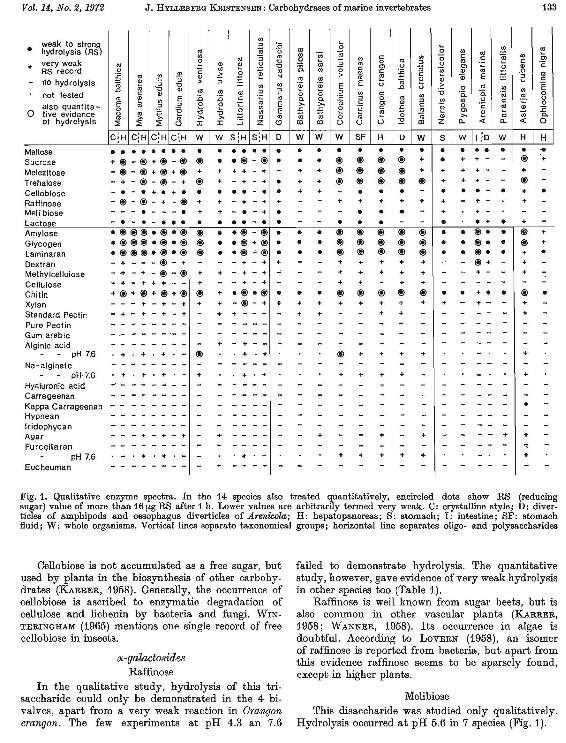
Vol. 14, No.2, 1972 J. HYLLEllERG KRISTENSEN: Carbohydrases of marine invertebrates 133
weak to strong • hydrolysis (RS)
very weak RS record no hydrolysis
not tested also quantita ~ o tive evidence of hydrolysis
Maltose Sucrose Melezitose Trehalose Cellobiose Raffinose Meli biose Lactose Amylose Glycogen Laminaran Dextran Methylcellulose Cellulose Chitin Xylan Standard Pectin Pure Pectin Gum arabic Alginic acid
pH 7.6
Na-alginate pH-7.6
Hyaluronic acid Carrageenan Kappa Carrageenan Hypnean IridOPhycan Agar Furcellaran
pH 7.6 Eucheuman
• • • • • • •• • +@~@+@~@ @
-@-@+@+@ + -+-@~@-+ @
- ." - • + • +. • ~@-@-+-@ +
---e---e + -e-e- •••• e@@@e@e@ @
e@@@e@e@ @
e@@@e@e@ @
-+ -@-+ -.+-+-@-@ + - ~ - + + + - - +
+@+@+@+@ @
+-+-+ + + - + - +
+ . + . + . - @
+ • + . + +
- + - +
- . + • +
"' :J
:~
"' "' '" z
:c o
'" "0 "0
'" N
"' '" '" E E '" (!)
• ••••• • • @ - @ • + + +
- + e · .. -. . + - +
• - + · .. -. . e e@ - @ e e e @ + @ • e • @ ~ @ e
- - + + + - + - + - - + - + + e@e@.
+ - @ - +
+ +-
+ - + -. + +
+ . +
. +
'" (/)
-'1 0.
'" .~
o C. ". .c OJ to
w e e +
+
e e e
e + +
w
e + + +
• e
•
e
+
+
o ro :J
o >
E :J :c c. o (; ()
w e
SF
@ @ @ @ @ @
e +
• e e
@ @ @ @ @ @ + + +
@ @ + +
+
+
+
c: o 0> c: ~ o c: o 0> c: ~
U
H D w e e
@ @ +
@ @ + @ @ @ e e
+ +
• e e @ @ @
@ @ @ @ @ @
+ + + + + +
@ @ @
+ + +
+
+ +
+ +
+ + +
~
o o o ~ Q)
> :0
"' '§ Q)
z
s e
• + + e
+
• e
•
e
'" c:
'" 0> Q)
Q)
o C. en o 0> ". a.
.. c:
'" E
'" '0 o 'c Q)
<
'" '0; c: ~
'" a.
W liD W
• •• • + +-+ +-+ +-· . - .
e + e
• @ ••
• @ •• • '@ •• - @ +
+ -
• +. • + -
- - +
"' c: <D .0
~
"' .~. Q)
iii <:
H
~ C>
'c '" c: 'E o o o :c c. o
H e .. @ '+
+ @
+ e +
+ @ +
@ +
• +
Ii> e
+
+
+
+
+
+
Fig. 1. Qualitative enzyme spectra. In the 14 species also treated quantitatively, encircled dots show RS (reducing sugar) value of more than 16 [Lg RS after 1 h. Lower values are a.rbitrarily termed very weak. C: orystalline style; D: divertic\es of amphipods and oesophagus divertic\es of Arenicola; H: hepatopanoreas; S: stomach; I: intestine; SF: stomach fluid; W: whole organisms. Vertical lines separate taxonomical groups; horizontal line separates oligo- and polysaccharides
Cellobiose is not accumulated as a free sugar, but used by plants in the biosynthesis of other carbohydrates (KARRER, 1958). Generally, the occurrence of cellobiose is ascribed to enzymatic degradation of cellulose and lichenin by bacteria and fungi. WIN
TERINGHAM (1965) mentions one single record of free cellobiose in insects.
O<.-galactosides Raffinose
In the qualitative study, hydrolysis of this trisaccharide could only be demonstrated in the 4 bivalves, apart from a very weak reaction in Grangon crangon. The few experiments at pH 4.3 an 7.6
failed to demonstrate hydrolysis. The quantitative study, however, gave evidence of very weak hydrolysis in other species too (Table 1).
Raffinose is well known from sugar beets, but is also co=on in other vascular plants (KARRER,
1958; WANNER, 1958). Its occurrence in algae is doubtful. According to LOVERN (1958), an isomer of raffinose is reported from bacteria, but apart from this evidence raffinose seems to be sparsely found, except in higher plants.
Melibiose This disaccharide was studied only qualitatively.
Hydrolysis occurred at pH 5.6 in 7 species (Fig. 1).

134 J. HYLLEBERG KRISTENSEN: Carbohydrases of marine invertebrates Mar. Biol.
In nature, it is restricted in occurrence to secretions, leaves and fruits of higher plants.
{J -galactoside.s
Lactose
Like melibiose, lactose gives galactose and glucose on hydrolysis, but the 2 molecules are coupled via a {J-linkage. The sugar is reducing, and was only studied by chromatography. At pH 5.6, it was hydrolysed by 15 species (Fig. 1). At pH 4.3, hydrolysis was evident in Ophiocornina nigra. At the same pH, extracts of N Msarius reticulatus and AsteriM rubens also showed lactolytic activity. At pH 7.6, Littorina littona extracts hydrolysed lactose.
Milk of mammals is the only lactose source known in animals, but it has occasionally been isolated in higher plants (KARRER, 1958).
Polysaccharides, re.serve carbohydra,te.s
Amylose
This 1,4-linked (X-glucan was well hydrolysed at pH 5.6 by all extracts except from Ophiocornina nigra, where only weak activity could be demonstrated.
Animal amylases belong to the (X type, giving oligosaccharides (finally maltose) at hydrolysis. A step-wise hydrolysis could be observed on the chromatograms of all species, except the echinoderms AsteriM rubens and Ophiocom.ina nigra, where only glucose spots were identified.
The quantitative study showed a variation in RS-liberation after 1 h, ran"oing from 33 % in ldothea balthica to 4% in Mytilus edulis (Table 1). AGRAWAL (1963) found an amylase optimum near pH 5.8 in Corophium volutator, and BLANDAMEER and BEECHEY (1964) an optimum at 7.0 in CMcinus m.aena8. I found an optimum near 6.2 in M. edulis, but a pH of 5.6 was retained throughout, as the pH effect itself appeared to be small. This finding can be compared with observations by BLANDAMEER and BEECHEY (1964), who found that the specific activity in hepatopancreas extracts of C. m.aenM rose from 1.3 fJ. mol maltose/min/ mg protein at pH 5.6 to 1.8 at the pH 7.0 optimum (figures taken from their Fig. 1).
Amylose is a reserve carbohydrate of all chlorophyll-containing plants. Probably all green algae contain starch similar to that of land plants, comprising both amylopectin and amylose (PERCIVAL and DowELL, 1967). The starch of Rhodophycea (floridean starch) and of Cyanophycea also contain an (X-l,4-linked glucan (MEEUSE, 1962; PERCIVAL, 1968), while the starch of Phaeophycea consists of the i'i-l,3-linked glucan laminaran.
Many forms of bacteria produce starch-like glucans identical in structure to those produced by yeast and higher plants (FOSTER and STACEY, 1958).
Glycogen
In the qualitative study, this 1.4 and 1.6-linked (X-glucan was split by all species, but hydrolysis in Ophiocom.ina nigra was weak compared with the other species.
The quantitative study showed hydrolysis of glycogen, which generally followed the pattern of amylase activity. In 11 species, the ratio of glycogen to amylose (G/A) hydrolysis was about 2: 1. In Mytilus edulis, the ratio was nearly 3: 1, and in Balanus CTenatus and AsteriM Tubens, about 1: 1. This result can be compared with the finding of FmQUET and DANDRIFOSSE (1967), who studied G/A ratios in vertebrates and found them to be about 1: 2 in herbivores and 1: 1 in carnivores.
The G/A ratios computed from the present data appear to be independent of the nutritional type of the species investigated.
Amylase is known to act on glycogen but, according to WYATT (1967), a further enzyme is required for glucose production. MARZLUFF (1969) found that (X-glucosidase, in casu sucrase might fulfill this expected role. The present study gives little evidence of such correlation.
Glycogen is similar in structure to amylopectin from plants, but the molecule is more branched and contains more glucose units (KARLSON, 1962). Glycogens from bacteria are similar to those of animal origin (FOSTER and STACEY, 1958; ANTOINE and TEpPER, 1969).
Dextran
This glucan contains a high proportion of (X-1,6-glucosidic linkages. At pH 5.6, the chromatograms had distinct spots in A1·enicola marina, Balanus crenatus and Corophium volutator, while only weak hydrolysis could be observed in 6 other species.
The quantitative study revealed the highest RS value in Arenicola 1narina, although only 4 % RS was liberated after 1 h (Table 1).
A specificity of the dextran-splitting enzyme cannot be estimated from my investigation, but BRUNI et al. (1969). found that a highly purified acid (X-glucosidase hydrolysed glycogen, maltose and dextran at pH 4.5
Dextran is a constituent of certain chain-forming bacteria (FOSTER and STACEY, 1958). It is likely to be rare in marine environments.
Laminaran
In the qualitative study, the {J-1,3-linked glucan laminaran was significantly split by all species except AsteriM 1·ubens. A weak hydrolysis was found in extracts tested at pH 4.3 and 7.6. The laminaran was split into glucose and laminamn dextrins, mainly laminarabiose.
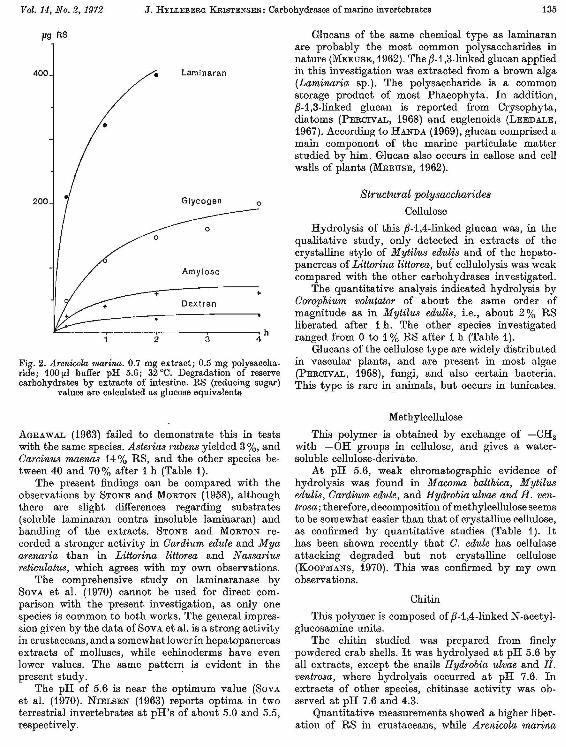
Laminaran was found to be efficiently split by Compkium volutato-r (80% RS after 1 h), while
Vol. 14, No.2, 1972 J. HYLLEBERG KRISTENSEN: Carbohydrases of marine invertebrates 135
JIg AS
400 • Laminaran
•
200 Glycogen o
o
Amylose
+ +
+ Dextran
2 3
Fig. 2. Arenicola marina. 0.7 mg extract; 0.5 mg polysaccharide; 100 V-l buffer pH 5.6; 32°C. Degradation of reserve carbohydrates by extracts of intestine. RS (reducing sugar)
values are calculated as glucose equivalents
AGRAWAL (1963) failed to demonstrate this in tests with the same species. Asterias l'ubens yielded 3 %, and Cal'cinus maenas 14% RS, and the other species between 40 and 70% after 1 h (Table 1).
The present findings can be compared with the observations by STONE and MORTON (1958), although there are slight differences regarding su bstra tes (soluble laminaran contra insoluble laminaran) and handling of the extracts. STONE and MORTON recorded a stronger activity in Cardium edule and Mya arenaria than in Littorina littorea and Nassarius reticulatus, which agrees with my own observations.
The comprehensive study on laminaranase by SOYA et al. (1970) cannot be used for direct comparison with the present investigation, as only one species is common to both works. The general impression given by the data of SOYA et al. is a strong activity in crustaceans, and a somewhat lower in hepatopancreas extracts of molluscs, while echinoderms have even lower values. The same pattern is evident in the present study. .
The pH of 5.6 is near the optimum value (SOYA et al. (1970). NIELSEN (1963) reports optima in two terrestrial invertebrates at pH's of about 5.0 and 5.5, respectively.
GJucans of the same chemical type as laminaran are probably the most common polysaccharides in nature (MEEUSE, 1962). The J'i-1,3-linked glucan applied in this investigation was extracted from a brown alga (Laminaria sp.). The polysaccharide is a common storage product of most Phaeophyta. In addition, J'i-1,3-Jinked glucan is reported from Crysophyta, diatoms (PERCIVAL, 1968) and euglenoids (LEEDALE, 1967). According to HANDA (1969), glucan comprised a main component of the marine particulate matter studied by him. Glucan also occurs in callose and cell walls of plants (MEEUSE, 1962).
Structural polysaccharides
Cellulose
Hydrolysis of this J'i-1,4-Jinked glucan was, in the qualitative study, only detected in extracts of the crystalline style of Mytilus edulis and of the hepatopancreas of Littorina littorea, but cellulolysis was weak compared with the other carbohydrases investig!Lted.
The quantitative analysis indicated hydrolysis by Corophium volutator of about the same order of magnitude as in Mytilus edulis, i.e., about 2% RS liberated after 1 h. The other species investigated ranged from 0 to 1 % RS after 1 h (Table 1).
GJucans of the cellulose type are widely distributed in vascular plants, and are present in most algae (PERCIVAL, 1968), fungi, and also certain bacteria. This type is rare in .. animals, but occurs in tunicates.
Methylcellulose
This polymer is obtained by exchange of -CHa with -OH groups in cellulose, and gives a watersoluble cellulose-derivate.
At pH 5.6, weak chromatographic evidence of hydrolysis was found in .Macoma balthica, Mytilus edulis, Cardium edule, and Hydrobia ulvae and H. ventrosa; therefore, decomposition of methyl cellulose seems to be somewhat easier than that of crystalline cellulose, as confirmed by quantitative studies (Table 1). It has been shown recently that C. edule has cellulase attacking degraded but not crystalline cellulose (KOOPMANS, 1970). This was confirmed by my own observations.
Chitin
This polymer is composed of J'i-1,4-linked N-acetylglucosamine units.
The chitin studied was prepared from finely powdered crab shells. It was hydrolysed at pH 5.6 by all extracts, except the snails Hydrobia ulvae and H. ventrosa, where hydrolysis occurred at pH 7.6. In extracts of other species, chitinase activity was observed at pH 7.6 and 4.3.
Quantitative measurements showed a higher liberation of RS in crustaceans, while Arenicola marina

136 J. HYLLE1lERG KRISTENSEN: Carbohydrases of marine invertebrates Mar. BiOI.
had the lowest activity (Table 1). The estimated chitinnolysis comprises the activities both of chitinase and chitobiase calculated as N-acetylglucosamin equivalents. The reducing capability of N-acetylglucosamine is only about i of that of glucose.
Chitin is a structural component in several invertebrate phyla, especially arthropods, molluscs and annelids (JENIAUX, 1963). The polymer is also known to occur in fungi and certain algae, e.g. diatoms (McLACHLAN et aI., 1965).
Xylan
This pentosan is principally a ,B-1,4.linked polymer of xylose, but also 1,3-linkages occur. Only one case of xylanase activity was observed in the qualitative incubatious. A spot with an R j value corresponding to xylose indicated degradation of xylan by ldothea balthica. The chromatographic records were not in accordance with the RS measurements, where all extracts indicated hydrolysis of xylose ranging from 4% RS in Littorina littorea to 0.1 % in Arenicola marina after 1 h (Table 1).
Xylan is widely distributed in cell walls of vascular plants. Smaller amounts of other compounds are usually found associated with xylans, their nature depending on the origin of the polymer. There a,re several reports on xylans in marine algae (KREGER, 1962), especially in Rhodophyta (PERCIVAL and DowELL,1967).
Pectin
The constituants, i.e., sugars, uronic acids, the degree of estrification and percentage of insoluble protopectin, varies according to the origin of the pectin, but the polymer is principally a chain of iX-1,4-linked galacturonic-acid units.
Hydrolysis of pectin was only tested by ohromatography. The high-ester pectin (estrificated in 67.6% of the carboxyl groups) was not hydrolysed in any of the incubations. An unspecified standard pectin tested had spots on the chromatograms with an R j value corresponding to glucose in about half of the incubations. In this case, however, it is obvious that the spots are due to hydrolysis of oligosaccharides present in the standard pectin.
Pectins are widely distributed, but found mainly in vascular plants.
Gum arabic
This acidic polysaccharide occurs in Acacia species. It contains several sugars and uronic acids (HIRST and JONES, 1958), and varies in composition depending on origin.
Incubation with gum arabic did not reveal reducing carbohydrates in the qualitative tests, and quantitative studies were not carried out.
Alginic acid and alginate
These polysaccharides are polymers of mannuronic and guluronic acids connected by 1,4 and possible also 1,3 linkages. Sodium alginate is a water-soluble salt of alginic acid, but varies in uronic-acid proportions as well as accompanying sugars.
In the qualitative analysis, degradation of alginic acid was not observed in the species tested at pH 5.6, but it was split at pH 7.6 by Littorina littorea and, to a lesser degree, by Hydrobia ventrosa. The sodium salt was split by the same species both at pH 5,6 and 7.6. At pH 7.6, weak hydrolysis of Na-alginate was observed in NassariU8 nticulatU8.
Measurements at pH 7.6 gave a higher liberation of RS from alginic acid than from Na-alginate (Table 1). The pH 7.6 is close to the optimal values recorded by FRANSSEN and JENIAUX (1965).
A polysaccharide similar to alginic acid has been isolated from bacteria grown under artificial conditions but, apart from this record, the occurrence of this polysaccharide seems to be restricted to brown algae (PERCIVAL and DOWELL, 1967).
Hyaluronic acid
This polymer consists of glucuronic acid and Nacetylglucosamine connected via 1,3-linkages to chainformed macromolecules.
Degradation of hyaluronic acid was tested only by chromatography. The viscosity of the mucoid was obviously decreased by many of the extracts, but complete degradation never occurred, and oligoglycosides were never observed. KITAMIKADO and YA· MAMOTO (1969, 1970) investigated hyaluronidase activity in fish, some invertebrates and bacteria, and found chromatographic evidence of di- and tetrasaccharides as products of hydrolysis. The pH optima of enzymes of fish and bacteria were 3.5 and 6.2, respectively.
Hyaluronic acid comprises a significant part of connective tissue and many slimy secretions in animals.
Mucilage-like polysaccharides
None of the polysaccharides, carrageenan, kappa carrageenan, hypnean, eucheuman and iridophycan, liberated RS except to a small extent in Asterias rubens (Fig. 1). An unidentified reducing-sugar (Rj
close to glucose) was found in agar incubations in some of the species at pH 5.6 (Fig. 1). Furcellaran was negative on all chromatograms, but incubations investigated photometrically showed an increases in RS from 0.1 to 1,8% after 1 h at pH 7.6 (Table 1).
The polysaccharides are characterized by linkage of sulfate esters with a galactan, although several sugars other than galactose have been identified in hydrolysates.

Vol. 14, No.2, 1972 J. HYLLEBERG KRISTENSEN: Carbohydrases of marine invertebrates 137
Carrageenan is a crude extract of the red algae Chondrus stellata. Kappa carrageenan is the residue in the supernatant after HCl extraction of carrageenan.
Extracts from the red algae Hypnea and Iridea are termed hypnean and iridophycan, respectively, and are similar in composition to carrageenan.
Agar-agar is extracted from red algae of the genera Gelidium and Gracillaria. Furcellaran (Danish agar) is a 1,3-linked galactoglycane sulphate isolated from the red alga Furcellaria jastigiata. Eucheuman is extracted from Eucheuman cottonii.
Discussion
Evaluation of the metabolic significance of enzymes from crude extracts is not easy since their activity can be interpreted in different ways. Firstly, enzymes play an important role in digestion when acting on readily hydrolysable material. Secondly, certain enzymes may act as a "tool" to open cell walls and make hydrolysable matter more readily accessible. Thirdly, enzymes may be produced by microorganisms in the gut of the animals studied.
The methods of the present work should cover all three possibilities. However, the following discussion is based chiefly on the first-mentioned possibility, as a relatively strong hydrolysis is likely to occur in carbohydrates with a potential food value. Enzymes with "tool" function may be difficult to evaluate, but it is possible that some of the enzymatic activities termed "very weak RS records" in Fig. 1 belong to this category. Enzymes such as cellulase may be important in the digestive process, even when present in small quantities.
As regards the enzymes of microorganisms in the gut, it is evident that these enzymes cannot be distinguished from the host enzymes by the technique used here. However, bacterial enzymes are believed to occur only in small quantities in the extracts, as there are no morphological adaptations of the gut of the animals studied which would enable quantities of microorganisms to be harboured.
It has often been discussed whether enzyme spectra may provide information on the food sources of animals or not. According to WEEL (1961), the general opinion is that enzyme equipment and secretion "fit" the diet. An evaluation of the enzymefood relation from the present study seems to depend on whether the carbohydrases are treated "one by one" or on whether the total number of significantly hydrolysed carbohydrates is considered. A one-by-one analysis only gives limited information in this respect. This conclusion coincides with the result of NIELSEN (1962), who found a poor - if any - correlation between the type of diet and the carbohydrases present in the alimentary tract of soil and litter invertebrates.
This poor correlation can be explained by two facts: (1.) A number of carbohydrates are of universal
18 Marine Biology. Vol. 14
occurrence in plants, and may be found in animals also. In other words, demonstration of, e.g. trehalase activity in crude extracts, cannot show whether the tested animals posses the trehalase to digest plants or other animals, or whether the enzyme is used elsewhere in the metabolism of these species; (2) according to earlier theories (VEIBEL, 1958), only 5 different enzymes are necessary to hydrolyse all glycosides. Nei· ther this. theory nor the view that hydrolysis of each glycoside demands a specific enzyme are accepted today. There are many records of highly purified enzymes attacking a number of substrates with identical linkage between the molecules (cf. BRUNI et aI., 1969; HASEGAWA and NORDIN, 1969).
Enzymes hydrolysing carbohydrates which are unlikely to occur in natural food may be considered from this view point. If a specific cellobiase exists (VEIBEL, 1958; OKADA et al., 1966), the strong cellobiase activity found in most of the investigated species is amazing, as cellobiose is only known to occur free in the last step of the decomposition of cellulose ..
NIELSEN (1962), PENTREATH (1969), and TELFORD (1970) also observed that cellobiose was strongly hydrolysed, even when degradation of cellulose could not be demonstrated by the sll-me extracts. The natural substrate is likely to have universal occurrence, since cellobiase has been demonstrated in terrestrial, limnic, and marine invertebrates. Possible compounds would be glucons connected via {i-linkage to an aglucon. Amygdalin is a {i-glucoside of this type, and is hydrolysed by the {i-glucosidase emulsin, which also acts on cellobiose.
The glucon of amygdalin is gentiobiose, which has the glucose molecules· connected via {i-1,6-linkages, indicating that the con£guration at - 0 - is more important for the action of the enzyme than the linkage number. Emulsin is a typical plant enzyme and, according to GOTTSCHALK (1958), it has been established that plant {i-glucosidases from different sources vary in specificity. Some act on cellobiose, gentiobiose and other {i-D-glycosides, while others have more restricted activity.
JENIAUX and DEVIGNE (1960) observed that commercial {i-glucosidase hydrolyses chitobiose, and this ought to be considered in future studies on the natural function of cellobiase. Chitobiose consists of two Nacetylglucosamine molecules linked by a {i-1,4-linkage, and it is split by chitobiase which, by some authors, is considered identical with N-acetylglucosaminidase (JENIAUX, 1963). The latter enzyme is common in digestive tissues, for example in various granules in the hepatopancreas of molluscs (SUMNER, 1969). It is known that digestive cell extracts of Mytilus calijornianus posses lytic activity dissolving bacteria rapidly (ZOBELL and LANDON, 1937). Cell walls of bacteria contain peptidoglycan (TIPPER et a!., 1967); lysozyme, (muramidase) shows N-acetylglucosaminidase activity towards chitin, various saccharides, and

138 J. HYLLEBERG KRISTENSEN: Carbohydrases of marine invertebrates Mar. Bioi.
synthetic substrates, and it acts on bacteria ce11-walls, cleaving the glycosidic bond of the N-acetylmuramic acid (W ADSTROM and HISATSUNE, 1970). My suggestion is, therefore, that hydrolysis of cellobiose may be caused by lysozyme.
The strong p-galactosidase activity (lactase) of most of the species tested is also interesting. Here, as in cellobiose, hydrolysis is conspicuous, but the occurrence of lactose in the natural diet is not known. It occurs in milk and in pollen of certain flowering plants. It is likely that lactase, if it occurs in marine invertebrates, acts on p-galactosides as it does in yeast, bacteria etc. (GOTTSCHALK, 1958).
Other substrates could be galactolipids of algae, which according to BENSON and SHIBUYA (1962) can be enzymatically degraded by IX- and p-galactosidases.
The slight hydrolysis of alginic acid and alginate by the carnivores Asterias rubens and Nassarius reticulatus is another puzzling example (Table 1), as alginic acid is restricted in occurrence to brown algae which are not a food source of these two species. FRANSSEN and JENIAUX (1965) found a hepatopancreas alginase activity of 1.2 A.U. (alginolytic units) in N. Teticulatus compared with 72 A.U. per g tissue of Littorina littorea (herbivore). In L. littorea, alginic acid may have a nutritional importance or the enzyme is here a tool to open up ingested cell walls. However, in carnivores, the importance of alginase is not known. The enzyme is either present as a reminiscence of ancestral functions, or it attacks a chemical bond which occurs in natural food as well as in alginic acid. Algal polysaccharides similar in structure to animal polysaccharides exist (PERCIVAL and DOWELL, 1967).
The highest number of carbohydrases was found in detritus feeders and microherbivores (bivalves) and the lowest in the presumably carnivorous Ophiocomina nigm. In my opinion, the total number of carbohydrases should permit a rough estimation of the natural food source of a species, but there is no evidence of a sharp classification between herbivores, carnivores, omnivores and detritus feeders on the basis of different carbohydrase spectra.
Fig. 1. shows that bivalves are able to degrade most glucosides, galactosides and a number of polysaccharides. Macoma balthica (deposit feeder) ingests the top layer of the sediment (NEWELL, 1965). Mya arenaria, Cardium edule and M ytilus edulis feed on suspended material. In all 4 species, diatoms and other minute plant material are frequently found in the intestines (BLEGVAD, 1914). No doubt exists that plant material is also digested as demonstrated in several feeding experiments, e.g. by WERNSTEDT (1942). WERNSTEDT fed Macoma balthica and Cardium edule with detritus and diatom cultures, and found natural detritus rich in bacteria to be more favourable for growth in M. balthica than in C. edule. With
diatoms, he observed a reverse result. The quantitative data on carbohydrases (Table 1) indicate a more effective degradation of carbohydrates by C. edule than by M. balthica. The stronger activity in the former species make my carbohydrase results comparable to the experiments of WERNSTEDT. A more effective degradation of laminaran in C. edule might be expected, since lamiDaran is an important reserve in many diatoms (MEEUSE, 1962).
Some of the crustaceans hydrolysed several carbohydrates. The omnivorous ldothea balthica degraded 10 different carbohydrates. This species readily ingests filamentous algae (RAVANKO, 1969), but appears to utilize animal food better (SOLDATOVA et aI., 1969). The qualitative spectra and the liberation of RS were found to be much the same as in Cmngon cmngon (omnivore, BLEGVAD, 1914) and Corophium volutator, a selective detritus feeder which sorts-out diatoms (THAMDRUP, 1935) and various microorganisms from mucous films on sand grains (MEADOWS, 1964). In contrast, Carcinus maenas, an omnivore with preference for animal food (ROPES, 1968), has a similar spectrum (11 carbohydrases) but a much lower RS liberation (Table 1). A comparison of RS liberation in the four crustaceans, therefore, indicates a different nutritive value of carbohydrates. Again, lamiDaran degradation seems to give good evidence for a food and enzyme relationship. LamiDaran may be significant in the nutrition of C. volutator, less so in that of I. balthica and C. crangon and of no significance in C. maenas.
The narrow spectrum of carbohydrates degraded by Bathyporeia pilosa and B. sarsi is remarkable, as they are known exclusively to ingest matter adhering to sand grains (NICOLAISEN and KANNEWORFF, 1969) covered by diatoms, blue-green algae and bacteria (ANDERSON and MEADOWS, 1968, 1969). This low number of carbohydrases is perhaps due to a specialized diet, since these amphipods live in a special environment of well-sorted sand.
The lowest number of carbohydrases in crustaceans was found in Balanus crenatus. HARNDEN (1968) investigated the carbohydrases of B. nubilis (27 carbohydrates tested), and found a low number of hydrolytic enzymes, viz. maltase, lamiDaranase, chitinase, and glycogenase active at pH 5.5 These carbohydrases were also found to be significantly active in the present study on B. crenatus. The small RS liberation (apart from lamiDaranase) may indicate that zooplankton is a major food source of sessile barnacles. BARNES (1969) found diatoms and sand grains only occasionally in the stomach of this species. Experiments by BOOKHOUT and COSTLOW (1959) have shown that animal food alone or in combination with Chlamydomonas support growth of older barnacles to a greater extent than unialgal cultures alone, indicating that, even if the algae are ingested, only a minor part of the carbohydrates are digested.

Vol. 14, No.2, 1972 J. HYLLEBERG KRISTENSEN: CarbohydraJJeS of marine invertebrates 139
Four gastropods were investigated, but the relationship between carbohydrases and diet is not evident. Littorina littorea, an omnivore (BLEGVAD, 1914) which feeds mostly on fucoids (OWEN, 1966), possesses the widest spectrum (10 carbohydrases). However, Nassarius reticulatus is capable of hydrolysing nearly the same carbohydrates, although quantitative differences were obvious. The intestine of N. reticulatus is often filled with bottom material containing many diatoms (BLEGVAD, 1914). This material may orginate from ingested food (bivalves, crustaceans), but the carbohydrases indicate that plant material is also perhaps digested by N. reticulatus. The Hydrobia species ingest bottom material and mucous film scraped off with the radula (NEWELL, 1965). The quantitative spectrum (Table 1) is not only very similar to that of the detritus feeding M acoma balthica, but also to the spectrum of N. reticulatus.
Arenicola marina hydrolysed 8 carbohydrates. The small detritus feeding Pygospio elegans hydrolysed 6, but the spectrum is not complete. Nereis diversicolor hydrolysed 8 and the oligochaete Paranais littoralis at least 7 carbohydrates. A comparison of food and enzyme spectra, respectively, is not possible in this group due to lack of data, except in A. marina and N. diversicolor. The former species ingests sand together with filtered particles (KRUGER, 1971), while the latter is omnivorous (GOERKE, 1971) with a preference for animal food (KONIG, 1964, p. 220).
The poor RS liberation obtained with Arenicola marina . extracts indicates that plant material other than diatoms and bacteria is of minor importance in the nutrition. This conclusion coincides with experiments by HOBSON (1967), who found that little of the available organic carbon in the sediment was utilized in two North American Arenicola species.
For comparative reasons, two echinoderms collected at deeper water were included in the study, as they are both known to take animal food. The sea-star Asterias 1"ubens hydrolysed 6 carbohydrates, and the brittle star Ophiocomina nigra only 4.
Asterias rubens preys on small bivalves, crustaceans etc., and detritus in the stomach (BLEGVAD, 1914) may, as in Nassarius reticula/us, originate from ingested prey. Here again, part of the carbohydrate may be utilized by the predator, although RS measurements indicate low efficiency only.
Ophiocomina nigra had the lowest number of carbohydrases of the species investigated. However, the spectrum would doubtless be extended if RS measurements were carried out. The poor hydrolytic activity (Fig. 1) is surprising, as the brittle star is considered to be omnivorous (REESE, 1966). VEVERS (1956) found that O. nigra occasionally browses on algae. According to FONTAINE (1965), the ophiuroid is capable of extracting plankton by filter feeding, while ROUSHDY and HANSEN (1960) did not find evidence of filter feeding in this species. At any rate, the spectrum of
carbohydrases may call attention to the possibility that O. nigra mainly digests animal food. The significance of animal food is easily demonstrated in aquaria, where mussel meat stimulates the feeding activity of O. nigra violently.
It appears from the present investigation that carbohydrases bear some relation to the food and trophical classification of marine invertebrates. However, oligosaccharides and reserve carbohydrates of plants may be of nutritive significance in most species, while utilization of structural carbohydrate is obviously poor, with the few exceptions of alginic acid and alginate. This conclusion is in agreement with other investigations. HUANG and GIESE (1958) investigated some marine herbivores, and found that only readily assimilable material such as reserve carbohydrates were digested. In contrast, BOOLOOTIAN and LASKER (1964) reported 80% of the dry matter of the brown alga Macrocystis to be removed by digestion in vivo in the sea urchin Strongylocentrotus pU1·puratus. However, they found other algae digested with lower efficiences by the same species (44 to 62%).
The present study also seems to fit available experimental data on the utilization of dead organic material by detritus feeders. NEWELL (1965) suggests that bacteria are an important food source for Macoma balthica. The detritus per se (dead material) only served as a substrate of attached flora. Experiments of FENCHEL (1970) on a marine detritus-feeding amphipod, and by HARGRAVE (1970) on a freshwater amphipod, gave similar evidence. FENCHEL (1970) found that detritus particles were completely free from attached Inicroflora (bacteria) during the passage through the alimentary tract of the amphipod. CROSSLEY et a1. (1963) state that terrestrial Inicroarthropods only fragment detritus, but that bacteria and fungi are responsible for the actual decay.
Enzymes hydrolysing cellulose, polyuronic acids and mucilage-like substances are co=only reported in marine invertebrates. However, it appears to be a general characteristic of enzymes acting on cellulose that they need a long exposure time of the substrate for complete digestion, and such long exposure is not likely in the detritus-feeding animals studied. KRUGER (1971) mentions 14 min as an average time necessary for Arenicola marina to renew its intestinal contents. I have measured the renewal time in the deposit-feeding bivalve Abra alba S. WOOD by fecalpellet countings, and have found that the gut content was renewed in 0.3 g animals in 1 h at 18°C. In other words, it is believed that structural carbohydrates are not degraded during passage through the gut of these animals.
A number of works on cellulases have been published and, according to Y OKOE and YASUMASU (1964), cellulase is a widely distributed enzyme among invertebrates. Cellulases which may be significant in metabolism have been shown in wood-ingesting spe-

140 J. HYLLEBERG KRISTENSEN: Carbohydrases of marine invertebrates MM. Bioi.
cies such as Teredo (GREENFmLD, 1959), Bankia indica (NAIR, 1955) and Limnoria (several references in ARVY, 1969). However, much work on degradation of carbohydrates is based on viscosimetry or turbidity readings, and great variety in applied substrates may, in addition, make comparisons impossible. The difficulties are demonstrated by EpPLEY and LAsKER
(1959), who found viscosimetric evidence of alginase in St·rongylocentrotus purpura/us, butfailed to show RS as a result of the alginolysis.
It is emphasized that the significance of carbohydrases in the metabolism and turnover of structural elements is, on the whole, unknown in marine invertebrates, although there are several reports on enzymatic degradation of these compounds (cf. MEEUSE and FLUEGEL, 1958; GALLI and GIESE, 1959 ; HORIUcm, 1963). However, it can be assumed that bulk degradation of structural polysaccharides will not normally occur through digestive enzymes produced by the herbivores themselves, but possibly in some cases through associated microorganisms. In general, a herbivore will ingest large amounts of plants but only digest a fraction; an omnivore will prefer animal to plant food; and detritus feeders will pass quantities of detritus through the alimentary tract in order to secure the nessessary intake of easily assimilable matter.
Summary
1. Hydrolysis of oligosaccharides was mostly found to be weak in extracts of the species studied.
2. The reserve carbohydrates amylose, glycogen and laminaran were significantly split in nearly all species. Dextran was poorly, if at all, split.
3. Hydrolysis of structural carbohydrates was very poor, with few exceptions formed by methycellulose and alginic acid. An exception was chitinolysis, which occurred in all species.
4. The spectrum of the carnivorous Nassu'rius reticulatus was very similar to that of the detritusfeeding Jiydrobia ventrosu, indicating that the spectra of carbohydrases cannot be predicted from information on ingested food .
5. This latter result, as well as the whole problem of a food and enzyme relation, is discussed together with the significant hydrolysis of carbohydrate which does not occur in the natural food .
Adc7WWledgemenia. Finanoial support from Statens NaturvidenskabJige ForskingsrM is gratefully acknowledged. Thanks are also due to the Copenhagen Pectin Factory and the Litex Faotory, Glostrup, for gifts of algal polysaccharides. My sincere gratitude is due to my colleague Dr. T. FENCHEL for his encouragement and interest in this work, and to Miss E. GLOB, who carefully carried out the laborious quantitative work.
Literature cited
AGRAWAL, V. P .: Studiea on the physiology of digestion in Corophium valutat<>r. J . mar. bio!. Ass. U.K. 41, 125--128 (1963) .
ANDERSON, J. G. and P. S. MEADOWS: 1rIicro-organisms a.nd organic matter attaohed to the surfaces of marine sand grains. J . mar. bio!. Ass. U.K. 48, 161-175 (1968).
- - Bacteria on intertidal sand grains. Hydrobiologia 38, 33---46 (1969).
ANTOINE, A. D. and B. S. TEPPER: Characterization of glycogens from mycobacteria. Arcbs Biochem. Biophys. 134, 207-213 (1969).
ARvy, L.: Lea enzymes chez lea crustacea. Annee BioI. 8, 505-580 (1969).
BARNES, H.: Stomach contents ",nd microfeeding of some common cirripedes. Can. J. Zoo!. 87, 231-236 (1959).
BENSON, A. A. and I. SBIBUYA: Surfactant lipids. In: Physiol. ogy and biocht'mistry of algae, pp 371-383. Ed. by R. A. LEWIN. New York and London: Academic Press 1962.
BLANDAMEER, A. and R. B. BEECIlEY: The indentification of an a-amylase in aqueous extracts of the hepatopanoreas of Carcinus maenas, the common shore crab. Comp. Biochem. Physiol. 13, 97-105 (1964).
BLEGV AD, H.: Food and condition of nourishment among the communities of invertebrate animals on the sea bottom in Danish waters. Bep. Dan. bioI. Stu 22, 41-78 (Appendix) (1914).
BOOKHOUT, C. G. and J . D. COSTLow: Feeding, molting, and growth in barnacles. In: Marine boring and fouling organisms, pp 212-225. Ed. by D. L. BAY. Sep,ttle: University of Washington Press 1959.
BOOLOOTIAN, R. A. and B. LASKER: Digestion of brown algae and distribution of nutrients in the purple sea urchin Strongylocentrotus purpura/us. Compo Biochem. Physiol. 11, 273-289 (1964).
BRUNI, C. B., F. AURlccmo and L COVELLI: Acid a·D.glucosidase glucohydrolase from cattle liver. J. bio!. Chem. 244, 4735-4742 (1969).
CEBNIC1IIABI, E., L . MUS<JATINE and D. C. SMITH: Maltose excretion by the symbiotic algae of Hydra viridis. Proc. R. Soc. (Ser. B) 178, 657-576 (1969).
CROSSLEY, D. A. Jr., M. WrrlUMP and G. J . DODSON : GroSs effects of arthropods and microflora on rates of leaf litter breakdown. In: Progress in terrestrial and fresh water ecology, pp 98-99. Health Physics Div. A. Prog. Rep. OBNL - 3492, U.s. Atomio Energy Commission 1963.
DUCKWOBTH, M. and J. B. TuRVEY: An extra~ellular a.garase from a Cytopha{ja species. Biochem. J. 113, 139-142 (1969).
EPPLEY, R. W. and R. LASKER: Alginase in the sea urchin Strongylocentrotus purpuratus. Science, N.Y. 129, 214-215 (1959).
FAVOROV, V. V. and V. E . VASKOVSKY: Alginases of marine invertebrates. Comp. Biochem. Physiol. 38, 689-696 (1971).
FENCHEL, T.: Studies on the decomposition of organic detritus derived from the turtle grass TJuilassia tesl1ulim,m. Limnol. Oceanogr. 16, 14-20 (1970).
FmQUET, M.-P. et G. DANDRIFOSSE : Influence dll regime alimentaire sur lea proprietes catalytiques de I'amylase pancreatique. Archs into Physiol. Biochim. 78, 347-355 (1967).
FONTAINE, A. B .: The feeding mechanism of the ophiuroid Ophiocomina ni{J'ra. J. mar. bio!. Ass. U.K. 45, 373-385 (1965).
FOSTER, A. B. and M. STACEY: The polysaccharides from lower plants such as oacteria,.algae, fungi and liohens etc. In: Enoyolopedia of plant physiology,'Vol. 6. pp 331-334. Ed. by W. BUllLAND. Springer Verlag: Berlin 1958.
Fox, D. L.: Comparative metabolism of organic betritus by inshore animals. Ecology 31, 100-108 (1950).
FRANSSEN, J. et CR. JEN1AUX: Digestion de I'adde alginique ohez les invertebrea. Cah. BioI. mar. 6, 1-21 (1965) .
GALLI, D. R. and A. C. GIESE : Carbohydrate digestion in a herbivorous snrul Tegula junehralis. J . expo Zool. 140, 415--440 (1959).

Vol. 14. No.2, 1972 J. HYLLEBERG KRISTENSEN: Carbohydrases of marine invertebrates 141
GEORGE. D.: Organic matter available to the polychaete Cirri/ormia /entaculata {MONTAGU} living in an intertidal mud flat. Limnol. Oceanogr. 9, 453-455 {1964}.
GILMOUR, D.: Biochemistry of insects, 343 pp. New York: Academio Press 1961.
~ERKE, H.: Die Erniihrnngsweise der Nereis·Arten {Polychaeta. Nereidae} der deutschen Kiisten. VercH£. Inst. Meeresforsch. Bremerh. 13, 1-50 {1971}.
GOTTSORALK, A.: The enzymes controlling hydrolytic, phosphorolytic and transfer reactions of the oligosaooharides. In: Enoyclopedia of plant physiology, Vol. 6. pp 87-124. Ed. by W. RUHLAND. Berlin: Springer Verlag 1958.
GREENFIELD. L. J.: Cellulolytic activity in Teredo. In: Marine boring and fouling organisms, pp 363-371. Ed. by D. L . RAY. Seattle: University of Waahington Press 1959.
ILumA, N. : Carbohydrate metabolism in the marine diatom Skek/,on,etna OO8latum. Mar. BioI. 4. 208-214 (1969).
HARGRAVE. B. T.: The effect of a deposit-feeding amphipod on the metabolism of benthic microflora. Limnol. Oceanogr. 16.21-30 (1970) .
HARNDEN. D. G.: Digestive carbohydra.ses of BaJa,71/U8 nubilis (DABWIN. 1854). Comp. Biochem. Physiol. 26. 303-309 {1968}.
HASEGAWA. S. and J. H. NORDIN: Enzymes that hydrolyze fungal cell wall polysaooharides. I. Purifioation ~d properties of an endo-",.D{1,3}-glucanase from Tnchoderma wide. J. bioI. Chem. 244, 5460-5470 (1969).
HmST. E. and J . K. JONES: The gums and mucilages of plants. In: Enoyclopedia of plant physiology. Vol. 6. pp 500-517. Ed. by W. RUHLAND. Berlin: Springer Verlag 1958.
HOBSON, K. D.: The feeding and ecology of two North Paoifio Arenicola speoies. BioI. Bull. mar. bioI. Lab., Woods Hole 188, 343-354 (1967).
HORrocm, S.: On the nature of digestive diverticula in the marine lamellibranch Venerupis philippinarum ADAMS et REEVE. Sci. Rep. Tokyo Kyoiku Daig. (Sect. B) 11. 133-152 (1963).
HUANG, H. and A. C. GIESE: Tests for digestion of algal polysaccharides by some marine herbivores. Science, N. Y. 127, 475 (1958).
JENUUX, CR.: Chitine et chitinolyse. 181 pp. Paris: Masson et Cle 1963.
- et J. DEVlGNE: Sur la liberation d'acetylglucosamine il. partir de chitine pulverisee. sous l'action des chitobiases. Archs into Physio!. Biochim. 68. 685-686 (1960).
KARLSON. P. : Kurzes Lehrbuch der Biochemie. 360 pp. Stuttgart: G. Thieme Verlag 1962.
KARRER. W.: Konstitution und Vorkommen der organischen PfIanzenstoffe. 1207 pp. Basel und Stuttgart: Birkhiinser Verlag 1958.
KIssELEVA. M. I. and D. M. VlTYUK: The feeding of Arenicola grubii (Arenicolidae) in the Black Sea. Zool. Zh. 49,219-222 (1970).
KITAMIKADO, M. and H. YAMAMOTO: Distribution of hyaluronidase in fish tissues. Bull. Jap. Soc. scient. Fish. 85. 466-470 (1969).
- - Action mechanism of fish gastric hyaluronidn.se. Bull. Jap. Soc. scient. Fish. 86, 385-390 (1970).
KOODlAN, P.: The occurrence of carbohydrases in digestive juice and in hepatopancreas of AslaGu8 fluviatilis F ABR. and of HomaMJ.$ vulgaris M-E. J. cell. compo Physio!. 68, 197-201 (1964) . .
KOOPMANS. J. J. C. : I. The nature of the cellulases in Helix pomata and Cardium eduk. Neth. J. Zoo!. 20. 445-463 (1970).
KONIG, D.: Einige neue Beobachtungen tiber die Rolle der Diatomeen am Wattenmeer. In: C.r. Congr. into Algaes mar. No.4. pp 215-221. Ed. by A. D. DE VmVlLLE. Oxford and Paris: Pergamon Press 1964.
KREGER. D. R: Cell walls. In: Physiology and biochemistry of algae, pp 315-335. Ed. by R A. LEWIN. New York and London: Academic Press 1962.
KRUGER, F.: Bau und Leben des Wattwurmes Arenicola marina. Helgoliinder wiss. Meeresunters. 22, 149-200 (1971).
LEEDALE, G. F.: Euglenoid flagellates, 242 pp. New York: Prentice Hall Inc. 1967.
LEVRING. T., H. A. HOPPE and O. J. SCHMID: Marine algae. 421 pp. Hamburg: Gram de Gruyter & Co. 1969. .
LOVERN, J . A.: Glyoolipids. In : Encyolopedia of plant phYSIOlogy, Vol. 6. pp 815-816. Ed. by W. RUHLAND. Berlin: Springer Verlag 1958.
MAIIZLUFJr. G. A . : Studies of trehalose and sucrose of Drosophila melanogaster. Arohs Biochem. Biophys. 184. 8-18 {1969}.
MAYER. D. und H. KihILWEIN: Ein Beitrag zur Kenntnis der cellulolytischen Aktiviiit von Archangium violauum. Zentb!. Bakt. ParasitKde (Abt. II) 123. 361-368 {1969}.
McLACHLAN. J . A.. A. G. McINNES and M. FALK: Studies on the chitan (ohitin: poly-N-a.cetylglucosamine) fibers of the diatom T1uzUu8iosira fluviatilis HUSTEDT. I. Production and isolation of ohitan fibers. Can. J. Bot. 48, 707-713 (1965) .
MEADows, P. S.: Experiments on substrate selection by Corophium species: films and bacteria on sand particles. J. expo BioI. 41. 499-511 (1964).
MEEUSE. B. J . D. : Storage products. In: Physiology and biochemistry of algae. pp 289-313. Ed. by R A. LEWIN. New York and London: Academic Press 1962.
- and W. FLUEGEL : Carbohydrate-digesting enzymes in the sugar gland juice of Cryptochiton s/dlm MIDDENDORF. Archs neerl. Zool. 13. 301-313 (1958).
Muus, B. : The fauna of Danish estuaries and lagoons. Meddr Kommn Danm. Fisk.·og Havunders. 0, 1-316 (1967).
NAGASBIMA, H., S. NAKAMURA and K. NISIZAwA: Isolation and identification of low molecular weight carbohydrates from a red alga, Serraticardia maxima. Bot. Mag., Tokyo 82, 379-386 (1969a).
-, H. OZAKI, S. NAXAMUBA and K. NISIZAwA: Physiological studies on f10ridean starch, f1oridoside and trehalose in a red alga, Serraticardia maxima. Bot. Mag.. Tokyo 82. 462-473 {1969b}.
NAIR. N. B.: Cellulaae activity of the crystalline style of the wood-boring pelecypod Bankia indica NAIR. Curr. Sci. 24. p. 201 (1955) .
NELSON, N.: A photometric adaptation of the Somogyi method for the determination of glucose. J. bioI. Chem. 108. 375-380 {1944}.
NEWELL, R.: The role of detritus in the nutrition of two marine deposit feeders the prosobranch Hydrobia ulvae and the binlve Mawma balthica. Proc. ZOOI. Soc. Lond. 144, 25-45 (1965) .
NICOLAISEN, W. and E. KANNEWORFF: On the burrowing and feeding habits of the amphipods Bathyporeia pilosa LINDSTBOM and Bathyporeia sarsi WATKIN. Ophelia 6, 231-250 {1969}.
NIELSEN, B. 0.: Carbohydrases of some wrack invertebrates. Nature jutl. 12, 191-194 (1966).
NIELSEN. C. 0.: Carbohydrases in soil and litter invertebrates. Oikos 13, 200-215 (1962).
- Laminariases in soil and litter invertebrates. Nature, Lond. 199, p. 1001 (1963).
OKADA, G. T .• T. NISIZAwA and K. NISIZAwA: Cellulases of a marine molluso Dolahella sp. Biochem. J. 99, 214-221 (1966).
OWEN, G.: Digestion. In : Physiology of Mollusca. Vol. II. pp 53--96. Ed. by K. M. WILBOUR and C. M. Y ONGE. New York and London: Academic Press 1966.
PENTBEATH, R J.: The morphology of the gut and a qualitative review of digestive enzymes in Bome New Zealand ophiuroids. J. Zoo!. London 109, 413--423 (1969).
PEROIVAL, E.: Marine algal carbohydrates. Ocea.nogr. mo.r. BioI. A. Rev. 6, 137-161 (1968).

142 J. HYLLEBERG KRISTENSEN: Carbohydrases of ma.rine invertebrates Mar. Bioi.
- and R. H. DOWELL: Chemistry and enzymology of marine algal polysaccharides, 219 pp. New York & London: Academio Press 1967.
RAVANKo, 0.: Benthic algae as food for some evertebrates in the inner part of the Baltic. Limnologica 7, 203-205 (1969).
REESE, E. S.: The complex behaviour of echinoderms. In: Physiology of Echinodermata, pp 157-218. Ed. by R. A. BOOLOOTIAN. New Yor)<, London and Sydney: Interscience 1966.
ROPES, J. W.: The feeding of the green crab Carcinus maenas (L.) Fishery Bull. Fish. WildI. Servo U.S. 67, 183-203 (1968).
RoUSHDy,H. M. and V. HANSEN: Ophiuroids feeding on phytoplankton. Nature, Lond. 188, 517-518 (1960).
SCHWOCH, G.: EinfluB von Augenstielextract auf Trehalose, Fructose und Glucose in der Hamolymphe von Orwnectes limosus. Experientia 26, 1324-1325 (1969).
SOLDATOVA, I. N., E. A. TSIKHON-LUKANINA, G. G. NIKOLAEVA and T. A. LUlU.8HEVA: On the transformation of the energy of food in marine crustaceans. Okeanologia 9, 1087-1094 (1969).
SOMOGYI, M.: A new reagent for the determination of sugars. J. bioI. Chem. 160, 61-68 (1945).
SOVA, V. V., L. A. ELYAKOVA and V. E. VASKOVSKY: The distribution of laminarinases in marine invertebrates. Comp. Biochem. PhysioI. 32, 459---464 (1970).
SPECK, U.: Das Kohlenhydratspektrum in den Organen des FluBkrebses Orwnectes limosus und seine Veranderungen im Jahresablauf. Z. vergI. Physiol. 66, 51-69 (1969).
STONE, B. A. and J. E. MORTON: The distribution of cellulases and related enzymes in the mollusca. Proc. malac. Soc. Lond. 33, 127-141 (1958).
SmINER, A. T.: The distribution of some hydrolytic enzymes in the cells of the digestive gland of certain lamellibranchs and gastropods. J. Zool. London 168, 277-291 (1969).
TELFORD, M.: Comparative carbohydrase activities of some crustacean tissue and whole animal homogenates. Comp. Biochem. Physiol. 34, 81-90 (1970).
THAMDRUP, H. M.: Beitrage zur Okologie der Wattenfauna auf experimenteller Grundlage_ Meddr Kommn Danm. Fiskog Havunders. Ser. Fisk.) 10, 1-125 (1935).
TIPPER, D. J., J. STROMINGER and J. C. ENSIGN: Structure of the cell wall of Staphylococcus aureus, Strain Copenhagen.
VII. Mode of action of the bacteriolytic peptidase from Myxobacter and the isolation of intact cell wall polysaccharides. Biochemistry, N.Y. 6, 906-920 (1967).
TREVELYAN, W. E., D. P. PROOTER and J. S. HARRISON: Detection of sugars on paper chromatograms. Nature, Lond. 166, 444---445 (1950).
VEIBEL, S.: Die Fermente des Auf- und Abbans (besonders die speziellen Heterosidasen). In: Encyclopedia of plant phy. siology, Vol. 6. pp 780--798. Ed. by W. RUHLAND. Berlin: Spriuger Verlag 1958.
VEVERS, H. G.: Observations on the feeding mechanism in some echinoderll1ll. Proc. zool. Soo. Lond. 126, 484---485 (1956).
W ADSTROM, T. and K. lhsATSUNE: Bacteriolytic enzymes from Staphylococcus aureus. Biochem. J. 120, 735-744 (1970).
WANNER, H.: Die Speicherung von Koblenhydrat im Blatt. In: Enoyclopedia of plant physiology, Vol. 6. pp 841-854. Ed. by W. RUHLAND. Berlin: Springer Verlag 1958.
WEEL, P. B. VAN: The comparative physiology of digestion in molluscs. Am. Zool. 1, 245-252 (1961).
WERNSTEDT, C.: Studies on the food of Macoma baltMca and Cardium edule. Vidensk. MOOdr dansk naturh. Foren. 106, 241-252 (1942).
WINTERINGllAM, F. P. W.: Some distinctive features of inseot metabolism. In: Aspects of insect biochemistry, pp 29-37. Ed. by T. W. GOODWIN. New York and London: Academio Press 1965.
WYATT, G. R.: The biochemistry of sugars and polysaccharides in insects. In: Advances in inseot physiology, Vol. 4. pp 287-360. Ed. by J. W. L. BEAMENT. London & New York: Aoademio Press 1967.
YOKOE, Y. and I. YASUMASU: The distribution of cellulase in invertebrates. Comp. Bioohem. Physiol. HI, 323-338 (1964).
ZOBELL, C. E. and W. A. LANDON: Bacterial nutrition of the california mussel. Proc. Soo. expo BioI. IIiOO. 116, 607-609 (1937).
Author's address: J. HYLLEBERG KRISTENSEN Zoological Institute Laboratorium B, Ecology University of Aarhus 8000 Aarhus C Denmark
Date of final manuscript acceptance: February 4, 1972. Communicated by B. SWEDMARK, Fiskebiickskil
Copyright © 2022 FDOKUMEN




















