
Emotion and Attention Interaction Studied through Event-Related Potentials
20
Emotion and Attention Interaction Studied through Event-Related Potentials Luis Carretie ´ 1 , Manuel Martí ´n-Loeches 2 , Jose ´ Antonio Hinojosa 2 , and Francisco Mercado 1 Abstract & Several studies on hemodynamic brain activity indicate that emotional visual stimuli elicit greater activation than neutral stimuli in attention-related areas such as the anterior cingulate cortex (ACC) and the visual association cortex (VAC). In order to explore the temporo-spatial characteristics of the interaction between attention and emotion, two processes characterized by involving short and rapid phases, event-related potentials (ERPs) were measured in 29 subjects using a 60-electrode array and the LORETA source localization software. A cue/target paradigm was employed in order to investigate both expectancy-related and input processing- related attention. Four categories of stimuli were presented to subjects: positive arousing, negative arousing, relaxing, and neutral. Three attention-related components were finally analyzed: N280pre (from pretarget ERPs), P200post and P340post (both from posttarget ERPs). N280pre had a prefrontal focus (ACC and/or medial prefrontal cortex) and presented significantly lower amplitudes in response to cues announcing negative targets. This result suggests a greater capacity of nonaversive stimuli to generate expectancy-related attention. P200post and P340post were both elicited in the VAC, and showed their highest amplitudes in response to negative- and to positive-arousing stimuli, respectively. The origin of P200post appears to be located dorsally with respect to the clear ventral-stream origin of P340post. The conjunc- tion of temporal and spatial characteristics of P200post and P340post leads to the deduction that input processing-related attention associated with emotional visual stimulation in- volves an initial, rapid, and brief ‘early’ attentional response oriented to rapid motor action, being more prominent towards negative stimulation. This is followed by a slower but longer ‘late’ attentional response oriented to deeper processing, elicited to a greater extent by appetitive stimulation. & INTRODUCTION Current Data and Open Issues Though the neural activity associated with emotion and related to attention have mainly been studied separately, they constitute closely interrelated and mutually de- pendent processes. In natural environments, attention is preferentially sustained by stimuli that have affective significance, in contrast to routine, emotionally neutral events (e.g., Lang, Bradley, & Cuthbert, 1997). Some recent experiments using hemodynamic neuroimaging methods provide interesting data indicating that certain attention-related brain areas increase their activity in response to emotional visual stimuli. Firstly, there is a group of studies showing that several areas of the visual association cortex (VAC) are more highly activated in response to both positive (or pleasant) and negative (or unpleasant) images than in response to neutral pictures (Lane, Chua, & Dolan, 1999; Lang et al., 1998; Reiman et al., 1997). Another group of data, also obtained through functional hemodynamic methods, has indi- cated activation in the anterior cingulate cortex (ACC) in response to emotional visual stimulation, positive and negative (Northoff et al., 2000; Teasdale et al., 1999; Canli, Desmond, Zhao, Glover, & Gabrieli, 1998). Both groups of results have been interpreted as indicating increased attention towards affective stimuli. Results obtained in relation to hemodynamic changes associated with attention towards visual emotional stim- ulation should be complemented by data recorded using temporally agile physiological signals. Both attention and emotion are characterized by involving short, rapid subprocesses, some of the most important occurring within the first 500 msec after stimulus onset (e.g., Mangun & Hillyard, 1995; Halgren & Marinkovic, 1994). High temporal-resolution techniques are neces- sary to study several unexplored important issues. Firstly, it is necessary to study which aspects or types of attention are associated with the areas found to be activated in response to emotional visual stimuli. Usual tasks employed in neuroimaging studies are character- ized by the presentation of sets of pictures (with rela- tively long interstimulus intervals) during several seconds or even minutes, and probably involve different types of attention. In this regard, though the involve- 1 Universidad Auto ´noma de Madrid, 2 Universidad Complutense de Madrid © 2001 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 13:8, pp. 1109- 1128
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Emotion and Attention Interaction Studied through Event-Related Potentials

Emotion and Attention Interaction Studied throughEvent-Related Potentials
Luis Carretie1 Manuel Martiacuten-Loeches2Jose Antonio Hinojosa2 and Francisco Mercado1
Abstract
amp Several studies on hemodynamic brain activity indicatethat emotional visual stimuli elicit greater activation thanneutral stimuli in attention-related areas such as the anteriorcingulate cortex (ACC) and the visual association cortex(VAC) In order to explore the temporo-spatial characteristicsof the interaction between attention and emotion twoprocesses characterized by involving short and rapid phasesevent-related potentials (ERPs) were measured in 29 subjectsusing a 60-electrode array and the LORETA source localizationsoftware A cuetarget paradigm was employed in order toinvestigate both expectancy-related and input processing-related attention Four categories of stimuli were presentedto subjects positive arousing negative arousing relaxing andneutral Three attention-related components were finallyanalyzed N280pre (from pretarget ERPs) P200post andP340post (both from posttarget ERPs) N280pre had aprefrontal focus (ACC andor medial prefrontal cortex) and
presented significantly lower amplitudes in response to cuesannouncing negative targets This result suggests a greatercapacity of nonaversive stimuli to generate expectancy-relatedattention P200post and P340post were both elicited in theVAC and showed their highest amplitudes in response tonegative- and to positive-arousing stimuli respectively Theorigin of P200post appears to be located dorsally with respectto the clear ventral-stream origin of P340post The conjunc-tion of temporal and spatial characteristics of P200post andP340post leads to the deduction that input processing-relatedattention associated with emotional visual stimulation in-volves an initial rapid and brief lsquoearlyrsquo attentional responseoriented to rapid motor action being more prominenttowards negative stimulation This is followed by a slowerbut longer lsquolatersquo attentional response oriented to deeperprocessing elicited to a greater extent by appetitivestimulation amp
INTRODUCTION
Current Data and Open Issues
Though the neural activity associated with emotion andrelated to attention have mainly been studied separatelythey constitute closely interrelated and mutually de-pendent processes In natural environments attentionis preferentially sustained by stimuli that have affectivesignificance in contrast to routine emotionally neutralevents (eg Lang Bradley amp Cuthbert 1997) Somerecent experiments using hemodynamic neuroimagingmethods provide interesting data indicating that certainattention-related brain areas increase their activity inresponse to emotional visual stimuli Firstly there is agroup of studies showing that several areas of the visualassociation cortex (VAC) are more highly activated inresponse to both positive (or pleasant) and negative (orunpleasant) images than in response to neutral pictures(Lane Chua amp Dolan 1999 Lang et al 1998 Reimanet al 1997) Another group of data also obtainedthrough functional hemodynamic methods has indi-
cated activation in the anterior cingulate cortex (ACC)in response to emotional visual stimulation positive andnegative (Northoff et al 2000 Teasdale et al 1999Canli Desmond Zhao Glover amp Gabrieli 1998) Bothgroups of results have been interpreted as indicatingincreased attention towards affective stimuli
Results obtained in relation to hemodynamic changesassociated with attention towards visual emotional stim-ulation should be complemented by data recorded usingtemporally agile physiological signals Both attentionand emotion are characterized by involving short rapidsubprocesses some of the most important occurringwithin the first 500 msec after stimulus onset (egMangun amp Hillyard 1995 Halgren amp Marinkovic1994) High temporal-resolution techniques are neces-sary to study several unexplored important issuesFirstly it is necessary to study which aspects or typesof attention are associated with the areas found to beactivated in response to emotional visual stimuli Usualtasks employed in neuroimaging studies are character-ized by the presentation of sets of pictures (with rela-tively long interstimulus intervals) during severalseconds or even minutes and probably involve differenttypes of attention In this regard though the involve-
1Universidad Autonoma de Madrid 2Universidad Complutensede Madrid
copy 2001 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 138 pp 1109- 1128
ment of both the VAC and ACC in attention to visualstimulation has received solid theoretical and experi-mental support (Sturm et al 1999 Luck Chelazzi Hill-yard amp Desimone 1997 Shulman et al 1997 Mangunamp Hillyard 1995 Posner amp Raichle 1995) they havebeen proposed to reflect different aspects of attentionand to be preferentially activated with different taskrequirements or periods According to Posner and Pe-tersen (1990) we can distinguish between lsquolsquoa generalalert state and one in which attention is clearly orientedand engaged in processing informationrsquorsquo (p 33) Tasksor phases within a task in which a significant level ofexpectancy or alertness is necessary such as thoserelated to target detection have been proposed to elicitthe former type of attention (which may appear even inthe absence of stimulation) and involve the activation ofthe anterior cingulum (Posner amp Petersen 1990) Theinvolvement of the ACC in alertness has been supportedexperimentally (eg Sturm et al 1999) On the otherhand tasks requiring recognition of already presentedvisual stimuli are associated with the second type ofattention involving an increase in the activation of theVAC (Posner amp Petersen 1990 Posner amp Raichle 1995)Several experiments clearly support this involvement ofthe VAC in input processing-related attention towardsvisual stimuli (see review by Mangun amp Hillyard 1995) Itis important to distinguish both types of attention inresponse to emotional images because they have sepa-rate functions In order to achieve this distinction it isnecessary to analyze separately the activity in expect-ancy-related and in input processing-related periods ofthe experimental task through high temporal resolutionsignals
Secondly the temporal characteristics of subprocesseswithin each type of attention to emotional stimulationshould be described For example it is necessary todetermine whether or not the attentional reactions topositive and to negative stimuli are produced at the samemoment The fact that hemodynamic neuroimagingmethodologies have not found temporal differences inresponses to the two types of emotional stimulation maybe due to the fact that they record an accumulation ofactivity over relatively long periods In this regard severalbehavioral studies suggest that negative events elicitmore rapid or more prominent emotional responses(involving cognitive and physiological changes) thanneutral or positive events (see reviews by Cacioppo ampBernston 1994 Cacioppo amp Gardner 1999 Taylor1991) Recent brain activation data obtained with hightemporal resolution methodologies (magnetoencepha-lography in this particular case) support this lsquonegativitybiasrsquo (Northoff et al 2000) Attention-related brain ac-tivity in relation to this bias has not yet been studieddirectly though the privileged access of negative stim-ulation to attentional resources has been indirectly sug-gested by studies on autonomic physiological responses(Ohman Esteves Flykt amp Soares 1993) and behavioral
studies using reaction time (Pratto amp John 1991) orvisual fixation (Fiske 1980) as dependent variables Theadaptive and evolutionary advantages of this bias areobvious The consequences of a dangerous or injuriousevent are often much more dramatic than the conse-quences of ignoring or reacting slowly to neutral or evenappetitive stimuli (eg Ekman 1992 Ohman 1992)
The combination of high spatial and high temporalresolution may provide information on a third importantissue which areas are activated in each subprocesswithin each type of attention to emotional stimulationData exist indicating that in the VAC alone emotionalstimuli elicit activity in more than 15 sources or lsquoclustersrsquoalong with those activated by all (emotional and neutral)visual stimuli (see eg Lang et al 1998) It is importantto investigate which subprocess of attention they reflect(if any) by studying their temporal characteristics Thisissue may be closely related first to time-related neg-ativity bias since circuitry that computes urgent utility(eg avoidance of a dangerous event) often divergesfrom the circuitry subserving deep identification anddiscrimination (Shizgal 1999) Secondly it may also berelated to the current debate on whether the evaluativesystems (and the neural substrates) involved in theprocessing of positive and negative stimuli are inde-pendent or overlapping Emotional content of stimula-tion has been traditionally assumed to be defined by twodimensions valence (whose extremes are positive- neg-ative) and arousal (calming- arousing) Initially proposedby Russell (1979) the lsquovalence arousalrsquo or lsquocircumplexrsquomodel considers that both positive and negative eventsare processed (and trigger emotional reactions) in thesame and single valence-related evaluative circuit Theidea that positive and negative situations activate differ-ent evaluative systems (associated at least in part withseparate neural mechanisms) has recently been de-fended (Cacioppo amp Gardner 1999 Davidson amp Irwin1999 Tellegen Watson amp Clark 1999) This perspectivehas received experimental support (Northoff et al 2000Lane Reiman Ahern Schwartz amp Davidson 1997 Le-Doux 1995) The use of a methodology with both hightemporal and relatively high spatial resolution mayfacilitate the exploration of spatial differences even inthe rapid and transient attentional subprocesses whoseinvolvement in this positive- negative lsquospatial independ-encersquo has not yet been defined
The Present Study
Electroencephalographic (EEG) recording of event-re-lated brain potentials (ERPs) constitutes a powerful andnecessary tool for exploring these important issuesconcerning the interaction of attention and emotionApart from high temporal resolution ERPs are able toprovide acceptable spatial information if a dense andwide distribution of electrodes is used and subsequentrecordings are analyzed through source localization
1110 Journal of Cognitive Neuroscience Volume 13 Number 8
algorithms To the best of our knowledge and surpris-ingly ERP data on the interaction between emotion andvisual attention have not been provided to date There-fore the main goal of the present study was to remedythis situation A wide conception of this interactionshould take into account several forms of attention Inthis regard the first specific objective was to explore thetwo types of attention previously mentioned expect-ancy-related and input processing-related attention Thesecond objective was to study the temporal character-istics of both types of attention and particularly thoseassociated with the positive or negative valence of theemotional stimulation Finally the third objective was toexplore the possibility that subprocesses within eachtype of attention present to some extent different brainsources
Expectance is usually elicited in ERP research throughthe experimental paradigm employed to evoke thecontingent negative variation (CNV) In its traditionalform this paradigm (that due to its timing character-istics cannot be employed in hemodynamic studies)requires the presentation in each trial of a warningsignal (S1) and subsequently of an lsquoimperativersquo stimulus(S2) (Walter Cooper Aldridge McCallum amp Winter1964) The onset of S2 requires a rapid response fromsubjects (eg to press a button as rapidly as possible inorder to terminate S2) During the interval between S1and S2 in which the subject is alert the CNV appearsterminating just after S2 onset CNV amplitude relatesdirectly to expectancy-related attention to S2 (McCal-lum 1988) If S2 consists of a visual image and if itsvisual exploration is required by the task ERP elicited bythis second stimulus is also able to reflect input process-ing-related attention
Four emotional categories were studied in the presentexperiment in which an S1 (hereafter labeled lsquocuersquo)- S2(lsquotargetrsquo) type task was employed arousing-positive(A+) arousing-negative (A ) neutral (N) and relaxing(R) These categories are intended to cover the maincombinations of positivity negativity and arousal inorder to study their influence on attention (as explainedelsewhere [Lang et al 1997] certain combinations suchas R are virtually impossible) Cue stimuli were simplewhite-on-black schematic line drawings or symbols Asdescribed in detail in the Methods section each symbolwas identified in the instructions with a particular objector motif (eg insect building etc) Cue presentationconsisted of two drawings or symbols belonging to thesame emotional category one above the other (seeFigure 12) The ideographic-type structure of cues en-sured that the stimuli were physically homogeneouswith no significant differences in their complexity colorbrightness contrast or size Also this type of visual cuewas preferred to other types such as words becausethey provided the pictorial nature of the target stimuli(consisting of photographs as explained below) thusfacilitating the involvement of similar processing systems
in response to both cue and target stimuli Moreoversimple ideographic stimuli would be less prone tohaving deep intrinsic emotional significance at least bycomparison with photographs or realistic drawings Acolor photograph (target) was presented 1750 mseclater (see Figure 11) Photographs always representedone of the motifs symbolized by the two drawings of thecue presentation Participants were told to identifyverbally which of the two drawings of the cue corre-sponded to the target (verbal response was given 2 secafter target onset) The instructions did not explicitlyestablish that the experiment dealt with emotionalreactions and as explained the cue indicated only inan implicit way the emotional category of the followingtarget This strategy had two advantages First it helpedto avoid a situation whereby participants considered thatsome of the stimuli were more important for the taskthan others (eg emotional stimuli more importantthan neutral ones) and thus to avoid a relevance-for-task effect often described in previous studies (egDuncan-Johnson amp Donchin 1977 the stimuli on whichthe task focuses tend to elicit the highest amplitudes incertain endogenous ERP components) Second homog-enization of cognitive demands makes it easier to reachmore solid conclusions about the emotion-related cau-sation of possible attentional differences
In general tasks in which target information is pre-dicted (100 correctly announced) by a cue have acharacteristic that is very interesting and relevant tothe present study They allow for the random presenta-tion of different stimuli minimizing the effect of theorienting response at least in its relation to novelty ornonexpectance of stimulation Since only active selec-tive attention was of interest in the present study theorienting response could represent an important inter-ference for posttarget ERPs Moreover in natural envi-ronments situations in which subjects perceive somecues or signals that trigger affective-related expectanceand prepare them for possible forthcoming emotionalevents are relatively frequent These situations have asyet scarcely been investigated in relation to brain activityThe study of input processing-related attention requiredthat only attended stimuli (cues and targets) wereanalyzed To ensure this recordings obtained in trialsin which subjects responded erroneously were elimina-ted It was also necessary to avoid emotional categoriesbeing heterogeneous with respect to the level of atten-tion needed to carry out the task correctly differencesshould be due to the affective content of each categorybut not to its difficulty with respect to the task Controlanalyses were planned in order to discard if necessarythose categories differing in this aspect from the rest
Our first hypothesis is that expectancy-related andinput processing-related phases of the task which asexplained associate with different types of attention willbe characterized by the activation of different brainareas The former should associate with the activation
Carretie et al 1111
of prefrontal areas involving the ACC and the latterwith VAC areas Secondly we consider that expectancy-related attention elicited by cues and input processing-related attention elicited by targets will be differentiallyaffected by the negativity bias Previous data on expect-ance towards emotional stimuli indicate that cues an-nouncing the presentation of stimuli to subjects whohave previously rated them as highly negative elicitlower CNV (particularly lower early CNV) amplitudesthan nonnegative pictures (Yee amp Miller 1987 Klormanamp Ryan 1980) On the other hand ERP signs of greaterand more prominent responses after the presentation ofnegative stimulation have been recently reported (Car-retie Mercado Tapia amp Hinojosa 2001 Ito LarsenSmith amp Cacioppo 1998) These data suggest thatbiases favoring an urgent motor response to negative(eg aversive) stimulation mainly occur when the stim-ulus has already appeared On the other hand the cue isa signal not to respond but rather to process forth-coming stimuli Therefore this second hypothesis maybe formulated indicating that negativity bias will affectposttarget ERPs to a greater extent than pretargetrecordings The third hypothesis deals with the spatial
location of each subphase of the attention-related activ-ity We expect spatial characteristics of attention to beparallel to temporal characteristics and to negativity biasie spatial differences will be related to urgency ofwithdrawal motivation (when avoidance stimulation ispresented) or to the need for deeper processing (whennonaversive stimulation is presented) and will alsoaffect preferentially posttarget ERPs
RESULTS
Figures 1 and 2 in which 14 widely distributed recordingchannels have been selected from the 58 channelsactually used show the grand averages of pre- andposttarget (respectively) recordings once the baselinehad been subtracted from each ERP
Selection of Components
Temporal Principal Component Analyses
Components explaining most pre- and posttarget ERPvariance were extracted through temporal principalcomponent analyses (tPCAs) Two covariance-matrix-
Figure 1 Grand averages inresponse to the four groups ofcue (pretarget ERPs) obtainedfrom the 29 subjects at 14selected scalp locations fromthe 58 locations actually em-ployed A+=arousing-positiveA =arousing-negativeN=neutral R=relaxing
1112 Journal of Cognitive Neuroscience Volume 13 Number 8
based tPCAs were carried out one for pre- and the otherfor posttarget ERPs Since the original number of cases(58 channels 29 subjects 4 stimulus conditions) andthe number of variables (487 digitized points for pre-and 250 points for posttarget ERPs) were excessive forcomputations two operations for data reduction main-taining the relevant ERP information were carried outFirstly a representative widely distributed sample of 28channels was selected (see Methods) Secondly toreduce the amount of data (only for tPCAs) we averagedeach five adjacent digitized points for each ERP (eachfive adjacent points represent 20 msec) The decision onthe number of components to select was based on thescree test (see eg Cliff 1987 for a description)Extracted components were then submitted to varimaxrotation
Following this selection criterion two components orfactors were extracted from pretarget ERPs Factor 1explained 9597 of total variance and Factor 2 ex-plained 124 (6581 and 3140 respectively afterrotation) Figure 3 shows the rotated loadings plottedover time for these two components Additionally Fig-ure 4 presents the mean factor scores for each scalp
location and for both components Basically factorscores which are calculated for each individual ERPreflect the product of point loading by point amplitudeTherefore amplitude and factor score are directly-re-lated parameters Analysis of peak latencies observed inFigure 3 and topography deduced from Figure 4 asso-ciate Factor 1 with the homogeneously distributed longlatency negative variation marked with an arrow labeledslow negativity (SN) in grand averages (Figure 1) andFactor 2 with a relatively early negativity (peaking at 280msec) with frontal midline dominance (see the mostnegative locations in Figure 4) signaled as N280 in grandaverages
With respect to posttarget ERPs selection yielded fourfactors Factor 1 explained 5062 of total varianceFactor 2 explained 2599 Factor 3 explained 532and Factor 4 342 (3595 3444 1073 and 424respectively after rotation) Figure 3 shows the rotatedloadings plotted over time for these four componentsMean factor scores for each recording channel areshown in Figure 4 Factor peak latency and topographycharacteristics associate Factor 1 maximal at centrallocations with the negative slow wave signaled with an
Figure 2 Grand averages inresponse to the four groups oftargets obtained from the 29subjects at 14 selected scalplocations from the 58 locationsactually employed A+=arous-ing-positive A =arousing-ne-gative N=neutral R=relaxing
Carretie et al 1113
arrow labeled SW in grand averages (Figure 2) Factors 2(340 msec) and 3 (200 msec) both maximal at parietaland parieto-occipital sites can be associated with de-flections marked as P340 and P200 respectively inFigure 2 Finally Factor 4 (peak at 500 msec) maximalat centro-parietal sites with a slow positive deflectionlabeled as P500 in grand averages These labels will beemployed hereafter to make results more understand-able and do not necessarily imply the identification ofthese factors with lsquoclassicalrsquo components such as P300 ortraditional SW (in the present case SW could simplyreflect a CNV release)
Selection of VAC- andor ACC-Related Components
Though the experimental design ensured that subjectsattended to the stimuli the extent of the association ofthe six components extracted by the tPCAs with theattentional processes we are studying could vary con-siderably Before submitting factors to analyses on theexperimental effects it was necessary to discard thosemainly reflecting the activity of brain areas different from
those under study As explained in the Introduction thefocus of our interest were those components reflectingactivation of visual cortices (input processing-relatedattention) andor prefrontal structures associated withexpectancy-related attention such as the ACC In orderto analyze the extent to which each factor was associatedwith these relevant brain areas topographic and source-localization analyses were carried out taking into ac-count voltages recorded in the 58 original recordingchannels Figure 5 shows the maps of the averageactivity in the 200- 520- and 740- 1750-msec periods ofpretarget ERPs (corresponding to N280 and SN respec-tively) and in the 176- 224- 312- 368- 472- 528- and620- 800-msec periods of posttarget ERPs (P200 P340P500 and SW respectively) The size of these windowswas proportional to the component period both withrespect to its absolute value and in relation to the totalERP epoch and shape of components was also takeninto account For example N280pre presents two high-loaded peaks not separable according to tPCAs thatshould be included in the window in order to correctlytake into account the main activity of this component
PRE-TARGET
02
03
04
05
06
07
08
09
1
0 500 1000 1500
msec
Lo
ad
s
F1PRE
F2PRE
POST-TARGET
-04
-02
0
02
04
06
08
1
0 200 400 600 800
msec
F1POSTF2POSTF3POSTF4POST
Lo
ad
s
Figure 3 tPCA analysis factor loadings after Varimax rotation Toppretarget ERPs Bottom posttarget ERPs
POST-TARGET
-03
-02
-01
0
01
02
03
AF3
AF4 F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
F1POSTF2POSTF3POSTF4POST
PRE-TARGET
-08
-06
-04
-02
0
02
04
06
08
AF
3
AF
4
F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
Fa
ct
or
Sc
or
es
F1PREF2PRE
Fa
ct
or
Sc
or
es
Figure 4 tPCA analysis mean factor scores corresponding to the 28recording channels taken into account in this analysis Top pretargetERPs Bottom posttarget ERPs
1114 Journal of Cognitive Neuroscience Volume 13 Number 8
Additionally low-resolution brain electromagnetic to-mography (LORETA) was applied a 3-D discrete linearsolution for the EEG inverse problem (Pascual-MarquiMichel amp Lehman 1994 Pascual-Marqui 1999) In itscurrent version LORETA refers to a three-shell sphericalmodel registered to the Talairach human brain atlas(Talairach amp Tournoux 1988) Solution is given there-fore in three coordinates x is the distance in milli-meters to the right (+) or left ( ) of midline y is thedistance anterior (+) or posterior ( ) to the anteriorcommissure and z is the distance superior (+) orinferior ( ) to a horizontal plane through the anteriorand posterior commissures Registration between spher-ical and the realistic head model in which solutions aregiven by LORETA used EEG electrode coordinates re-ported by Towle et al (1993) Figure 6 illustrates thesources of activity for the six components as calculatedby LORETA Source analysis were carried out on theaverage ERPs in response to the four groups of stimuliScalp distribution and source analyses suggest thatP500 is not originated in visual cortices or in the ACCor related prefrontal areas (see Table 1) Therefore
components selected for later analyses on the exper-imental effects were N280 and SN from pretargetrecordings and P200 P340 and SW from posttargetrecordings
Effects of Emotional Content on SelectedComponents
Control Analyses
In order to better interpret the results on the effects ofemotional stimulation on selected VAC- andor ACC-related components two control analyses were previ-ously carried out Firstly ANOVAs on the assessmentsgiven by subjects to each group of stimuli were com-puted in order to confirm that their affective contentwas the one supposed a priori As explained in theMethods section each subject filled out a bidimensionalscaling test for each group of pictures after the recordingsessions This test assessed the valence and the arousalcontent of the pictures presented as targets Table 2shows means and standard error of means for each typeof stimulus One-way repeated-measure ANOVAs were
Figure 5 Topographic maps representing the mean values (in microvolts) for the periods (in milliseconds) indicated at the top of each columnThese windows correspond respectively to N280 and SN from pretarget recordings and to P200 P340 P500 and SW from posttarget recordingsA+=arousing-positive A =arousing-negative N=neutral R=relaxing
Carretie et al 1115
computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

ment of both the VAC and ACC in attention to visualstimulation has received solid theoretical and experi-mental support (Sturm et al 1999 Luck Chelazzi Hill-yard amp Desimone 1997 Shulman et al 1997 Mangunamp Hillyard 1995 Posner amp Raichle 1995) they havebeen proposed to reflect different aspects of attentionand to be preferentially activated with different taskrequirements or periods According to Posner and Pe-tersen (1990) we can distinguish between lsquolsquoa generalalert state and one in which attention is clearly orientedand engaged in processing informationrsquorsquo (p 33) Tasksor phases within a task in which a significant level ofexpectancy or alertness is necessary such as thoserelated to target detection have been proposed to elicitthe former type of attention (which may appear even inthe absence of stimulation) and involve the activation ofthe anterior cingulum (Posner amp Petersen 1990) Theinvolvement of the ACC in alertness has been supportedexperimentally (eg Sturm et al 1999) On the otherhand tasks requiring recognition of already presentedvisual stimuli are associated with the second type ofattention involving an increase in the activation of theVAC (Posner amp Petersen 1990 Posner amp Raichle 1995)Several experiments clearly support this involvement ofthe VAC in input processing-related attention towardsvisual stimuli (see review by Mangun amp Hillyard 1995) Itis important to distinguish both types of attention inresponse to emotional images because they have sepa-rate functions In order to achieve this distinction it isnecessary to analyze separately the activity in expect-ancy-related and in input processing-related periods ofthe experimental task through high temporal resolutionsignals
Secondly the temporal characteristics of subprocesseswithin each type of attention to emotional stimulationshould be described For example it is necessary todetermine whether or not the attentional reactions topositive and to negative stimuli are produced at the samemoment The fact that hemodynamic neuroimagingmethodologies have not found temporal differences inresponses to the two types of emotional stimulation maybe due to the fact that they record an accumulation ofactivity over relatively long periods In this regard severalbehavioral studies suggest that negative events elicitmore rapid or more prominent emotional responses(involving cognitive and physiological changes) thanneutral or positive events (see reviews by Cacioppo ampBernston 1994 Cacioppo amp Gardner 1999 Taylor1991) Recent brain activation data obtained with hightemporal resolution methodologies (magnetoencepha-lography in this particular case) support this lsquonegativitybiasrsquo (Northoff et al 2000) Attention-related brain ac-tivity in relation to this bias has not yet been studieddirectly though the privileged access of negative stim-ulation to attentional resources has been indirectly sug-gested by studies on autonomic physiological responses(Ohman Esteves Flykt amp Soares 1993) and behavioral
studies using reaction time (Pratto amp John 1991) orvisual fixation (Fiske 1980) as dependent variables Theadaptive and evolutionary advantages of this bias areobvious The consequences of a dangerous or injuriousevent are often much more dramatic than the conse-quences of ignoring or reacting slowly to neutral or evenappetitive stimuli (eg Ekman 1992 Ohman 1992)
The combination of high spatial and high temporalresolution may provide information on a third importantissue which areas are activated in each subprocesswithin each type of attention to emotional stimulationData exist indicating that in the VAC alone emotionalstimuli elicit activity in more than 15 sources or lsquoclustersrsquoalong with those activated by all (emotional and neutral)visual stimuli (see eg Lang et al 1998) It is importantto investigate which subprocess of attention they reflect(if any) by studying their temporal characteristics Thisissue may be closely related first to time-related neg-ativity bias since circuitry that computes urgent utility(eg avoidance of a dangerous event) often divergesfrom the circuitry subserving deep identification anddiscrimination (Shizgal 1999) Secondly it may also berelated to the current debate on whether the evaluativesystems (and the neural substrates) involved in theprocessing of positive and negative stimuli are inde-pendent or overlapping Emotional content of stimula-tion has been traditionally assumed to be defined by twodimensions valence (whose extremes are positive- neg-ative) and arousal (calming- arousing) Initially proposedby Russell (1979) the lsquovalence arousalrsquo or lsquocircumplexrsquomodel considers that both positive and negative eventsare processed (and trigger emotional reactions) in thesame and single valence-related evaluative circuit Theidea that positive and negative situations activate differ-ent evaluative systems (associated at least in part withseparate neural mechanisms) has recently been de-fended (Cacioppo amp Gardner 1999 Davidson amp Irwin1999 Tellegen Watson amp Clark 1999) This perspectivehas received experimental support (Northoff et al 2000Lane Reiman Ahern Schwartz amp Davidson 1997 Le-Doux 1995) The use of a methodology with both hightemporal and relatively high spatial resolution mayfacilitate the exploration of spatial differences even inthe rapid and transient attentional subprocesses whoseinvolvement in this positive- negative lsquospatial independ-encersquo has not yet been defined
The Present Study
Electroencephalographic (EEG) recording of event-re-lated brain potentials (ERPs) constitutes a powerful andnecessary tool for exploring these important issuesconcerning the interaction of attention and emotionApart from high temporal resolution ERPs are able toprovide acceptable spatial information if a dense andwide distribution of electrodes is used and subsequentrecordings are analyzed through source localization
1110 Journal of Cognitive Neuroscience Volume 13 Number 8
algorithms To the best of our knowledge and surpris-ingly ERP data on the interaction between emotion andvisual attention have not been provided to date There-fore the main goal of the present study was to remedythis situation A wide conception of this interactionshould take into account several forms of attention Inthis regard the first specific objective was to explore thetwo types of attention previously mentioned expect-ancy-related and input processing-related attention Thesecond objective was to study the temporal character-istics of both types of attention and particularly thoseassociated with the positive or negative valence of theemotional stimulation Finally the third objective was toexplore the possibility that subprocesses within eachtype of attention present to some extent different brainsources
Expectance is usually elicited in ERP research throughthe experimental paradigm employed to evoke thecontingent negative variation (CNV) In its traditionalform this paradigm (that due to its timing character-istics cannot be employed in hemodynamic studies)requires the presentation in each trial of a warningsignal (S1) and subsequently of an lsquoimperativersquo stimulus(S2) (Walter Cooper Aldridge McCallum amp Winter1964) The onset of S2 requires a rapid response fromsubjects (eg to press a button as rapidly as possible inorder to terminate S2) During the interval between S1and S2 in which the subject is alert the CNV appearsterminating just after S2 onset CNV amplitude relatesdirectly to expectancy-related attention to S2 (McCal-lum 1988) If S2 consists of a visual image and if itsvisual exploration is required by the task ERP elicited bythis second stimulus is also able to reflect input process-ing-related attention
Four emotional categories were studied in the presentexperiment in which an S1 (hereafter labeled lsquocuersquo)- S2(lsquotargetrsquo) type task was employed arousing-positive(A+) arousing-negative (A ) neutral (N) and relaxing(R) These categories are intended to cover the maincombinations of positivity negativity and arousal inorder to study their influence on attention (as explainedelsewhere [Lang et al 1997] certain combinations suchas R are virtually impossible) Cue stimuli were simplewhite-on-black schematic line drawings or symbols Asdescribed in detail in the Methods section each symbolwas identified in the instructions with a particular objector motif (eg insect building etc) Cue presentationconsisted of two drawings or symbols belonging to thesame emotional category one above the other (seeFigure 12) The ideographic-type structure of cues en-sured that the stimuli were physically homogeneouswith no significant differences in their complexity colorbrightness contrast or size Also this type of visual cuewas preferred to other types such as words becausethey provided the pictorial nature of the target stimuli(consisting of photographs as explained below) thusfacilitating the involvement of similar processing systems
in response to both cue and target stimuli Moreoversimple ideographic stimuli would be less prone tohaving deep intrinsic emotional significance at least bycomparison with photographs or realistic drawings Acolor photograph (target) was presented 1750 mseclater (see Figure 11) Photographs always representedone of the motifs symbolized by the two drawings of thecue presentation Participants were told to identifyverbally which of the two drawings of the cue corre-sponded to the target (verbal response was given 2 secafter target onset) The instructions did not explicitlyestablish that the experiment dealt with emotionalreactions and as explained the cue indicated only inan implicit way the emotional category of the followingtarget This strategy had two advantages First it helpedto avoid a situation whereby participants considered thatsome of the stimuli were more important for the taskthan others (eg emotional stimuli more importantthan neutral ones) and thus to avoid a relevance-for-task effect often described in previous studies (egDuncan-Johnson amp Donchin 1977 the stimuli on whichthe task focuses tend to elicit the highest amplitudes incertain endogenous ERP components) Second homog-enization of cognitive demands makes it easier to reachmore solid conclusions about the emotion-related cau-sation of possible attentional differences
In general tasks in which target information is pre-dicted (100 correctly announced) by a cue have acharacteristic that is very interesting and relevant tothe present study They allow for the random presenta-tion of different stimuli minimizing the effect of theorienting response at least in its relation to novelty ornonexpectance of stimulation Since only active selec-tive attention was of interest in the present study theorienting response could represent an important inter-ference for posttarget ERPs Moreover in natural envi-ronments situations in which subjects perceive somecues or signals that trigger affective-related expectanceand prepare them for possible forthcoming emotionalevents are relatively frequent These situations have asyet scarcely been investigated in relation to brain activityThe study of input processing-related attention requiredthat only attended stimuli (cues and targets) wereanalyzed To ensure this recordings obtained in trialsin which subjects responded erroneously were elimina-ted It was also necessary to avoid emotional categoriesbeing heterogeneous with respect to the level of atten-tion needed to carry out the task correctly differencesshould be due to the affective content of each categorybut not to its difficulty with respect to the task Controlanalyses were planned in order to discard if necessarythose categories differing in this aspect from the rest
Our first hypothesis is that expectancy-related andinput processing-related phases of the task which asexplained associate with different types of attention willbe characterized by the activation of different brainareas The former should associate with the activation
Carretie et al 1111
of prefrontal areas involving the ACC and the latterwith VAC areas Secondly we consider that expectancy-related attention elicited by cues and input processing-related attention elicited by targets will be differentiallyaffected by the negativity bias Previous data on expect-ance towards emotional stimuli indicate that cues an-nouncing the presentation of stimuli to subjects whohave previously rated them as highly negative elicitlower CNV (particularly lower early CNV) amplitudesthan nonnegative pictures (Yee amp Miller 1987 Klormanamp Ryan 1980) On the other hand ERP signs of greaterand more prominent responses after the presentation ofnegative stimulation have been recently reported (Car-retie Mercado Tapia amp Hinojosa 2001 Ito LarsenSmith amp Cacioppo 1998) These data suggest thatbiases favoring an urgent motor response to negative(eg aversive) stimulation mainly occur when the stim-ulus has already appeared On the other hand the cue isa signal not to respond but rather to process forth-coming stimuli Therefore this second hypothesis maybe formulated indicating that negativity bias will affectposttarget ERPs to a greater extent than pretargetrecordings The third hypothesis deals with the spatial
location of each subphase of the attention-related activ-ity We expect spatial characteristics of attention to beparallel to temporal characteristics and to negativity biasie spatial differences will be related to urgency ofwithdrawal motivation (when avoidance stimulation ispresented) or to the need for deeper processing (whennonaversive stimulation is presented) and will alsoaffect preferentially posttarget ERPs
RESULTS
Figures 1 and 2 in which 14 widely distributed recordingchannels have been selected from the 58 channelsactually used show the grand averages of pre- andposttarget (respectively) recordings once the baselinehad been subtracted from each ERP
Selection of Components
Temporal Principal Component Analyses
Components explaining most pre- and posttarget ERPvariance were extracted through temporal principalcomponent analyses (tPCAs) Two covariance-matrix-
Figure 1 Grand averages inresponse to the four groups ofcue (pretarget ERPs) obtainedfrom the 29 subjects at 14selected scalp locations fromthe 58 locations actually em-ployed A+=arousing-positiveA =arousing-negativeN=neutral R=relaxing
1112 Journal of Cognitive Neuroscience Volume 13 Number 8
based tPCAs were carried out one for pre- and the otherfor posttarget ERPs Since the original number of cases(58 channels 29 subjects 4 stimulus conditions) andthe number of variables (487 digitized points for pre-and 250 points for posttarget ERPs) were excessive forcomputations two operations for data reduction main-taining the relevant ERP information were carried outFirstly a representative widely distributed sample of 28channels was selected (see Methods) Secondly toreduce the amount of data (only for tPCAs) we averagedeach five adjacent digitized points for each ERP (eachfive adjacent points represent 20 msec) The decision onthe number of components to select was based on thescree test (see eg Cliff 1987 for a description)Extracted components were then submitted to varimaxrotation
Following this selection criterion two components orfactors were extracted from pretarget ERPs Factor 1explained 9597 of total variance and Factor 2 ex-plained 124 (6581 and 3140 respectively afterrotation) Figure 3 shows the rotated loadings plottedover time for these two components Additionally Fig-ure 4 presents the mean factor scores for each scalp
location and for both components Basically factorscores which are calculated for each individual ERPreflect the product of point loading by point amplitudeTherefore amplitude and factor score are directly-re-lated parameters Analysis of peak latencies observed inFigure 3 and topography deduced from Figure 4 asso-ciate Factor 1 with the homogeneously distributed longlatency negative variation marked with an arrow labeledslow negativity (SN) in grand averages (Figure 1) andFactor 2 with a relatively early negativity (peaking at 280msec) with frontal midline dominance (see the mostnegative locations in Figure 4) signaled as N280 in grandaverages
With respect to posttarget ERPs selection yielded fourfactors Factor 1 explained 5062 of total varianceFactor 2 explained 2599 Factor 3 explained 532and Factor 4 342 (3595 3444 1073 and 424respectively after rotation) Figure 3 shows the rotatedloadings plotted over time for these four componentsMean factor scores for each recording channel areshown in Figure 4 Factor peak latency and topographycharacteristics associate Factor 1 maximal at centrallocations with the negative slow wave signaled with an
Figure 2 Grand averages inresponse to the four groups oftargets obtained from the 29subjects at 14 selected scalplocations from the 58 locationsactually employed A+=arous-ing-positive A =arousing-ne-gative N=neutral R=relaxing
Carretie et al 1113
arrow labeled SW in grand averages (Figure 2) Factors 2(340 msec) and 3 (200 msec) both maximal at parietaland parieto-occipital sites can be associated with de-flections marked as P340 and P200 respectively inFigure 2 Finally Factor 4 (peak at 500 msec) maximalat centro-parietal sites with a slow positive deflectionlabeled as P500 in grand averages These labels will beemployed hereafter to make results more understand-able and do not necessarily imply the identification ofthese factors with lsquoclassicalrsquo components such as P300 ortraditional SW (in the present case SW could simplyreflect a CNV release)
Selection of VAC- andor ACC-Related Components
Though the experimental design ensured that subjectsattended to the stimuli the extent of the association ofthe six components extracted by the tPCAs with theattentional processes we are studying could vary con-siderably Before submitting factors to analyses on theexperimental effects it was necessary to discard thosemainly reflecting the activity of brain areas different from
those under study As explained in the Introduction thefocus of our interest were those components reflectingactivation of visual cortices (input processing-relatedattention) andor prefrontal structures associated withexpectancy-related attention such as the ACC In orderto analyze the extent to which each factor was associatedwith these relevant brain areas topographic and source-localization analyses were carried out taking into ac-count voltages recorded in the 58 original recordingchannels Figure 5 shows the maps of the averageactivity in the 200- 520- and 740- 1750-msec periods ofpretarget ERPs (corresponding to N280 and SN respec-tively) and in the 176- 224- 312- 368- 472- 528- and620- 800-msec periods of posttarget ERPs (P200 P340P500 and SW respectively) The size of these windowswas proportional to the component period both withrespect to its absolute value and in relation to the totalERP epoch and shape of components was also takeninto account For example N280pre presents two high-loaded peaks not separable according to tPCAs thatshould be included in the window in order to correctlytake into account the main activity of this component
PRE-TARGET
02
03
04
05
06
07
08
09
1
0 500 1000 1500
msec
Lo
ad
s
F1PRE
F2PRE
POST-TARGET
-04
-02
0
02
04
06
08
1
0 200 400 600 800
msec
F1POSTF2POSTF3POSTF4POST
Lo
ad
s
Figure 3 tPCA analysis factor loadings after Varimax rotation Toppretarget ERPs Bottom posttarget ERPs
POST-TARGET
-03
-02
-01
0
01
02
03
AF3
AF4 F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
F1POSTF2POSTF3POSTF4POST
PRE-TARGET
-08
-06
-04
-02
0
02
04
06
08
AF
3
AF
4
F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
Fa
ct
or
Sc
or
es
F1PREF2PRE
Fa
ct
or
Sc
or
es
Figure 4 tPCA analysis mean factor scores corresponding to the 28recording channels taken into account in this analysis Top pretargetERPs Bottom posttarget ERPs
1114 Journal of Cognitive Neuroscience Volume 13 Number 8
Additionally low-resolution brain electromagnetic to-mography (LORETA) was applied a 3-D discrete linearsolution for the EEG inverse problem (Pascual-MarquiMichel amp Lehman 1994 Pascual-Marqui 1999) In itscurrent version LORETA refers to a three-shell sphericalmodel registered to the Talairach human brain atlas(Talairach amp Tournoux 1988) Solution is given there-fore in three coordinates x is the distance in milli-meters to the right (+) or left ( ) of midline y is thedistance anterior (+) or posterior ( ) to the anteriorcommissure and z is the distance superior (+) orinferior ( ) to a horizontal plane through the anteriorand posterior commissures Registration between spher-ical and the realistic head model in which solutions aregiven by LORETA used EEG electrode coordinates re-ported by Towle et al (1993) Figure 6 illustrates thesources of activity for the six components as calculatedby LORETA Source analysis were carried out on theaverage ERPs in response to the four groups of stimuliScalp distribution and source analyses suggest thatP500 is not originated in visual cortices or in the ACCor related prefrontal areas (see Table 1) Therefore
components selected for later analyses on the exper-imental effects were N280 and SN from pretargetrecordings and P200 P340 and SW from posttargetrecordings
Effects of Emotional Content on SelectedComponents
Control Analyses
In order to better interpret the results on the effects ofemotional stimulation on selected VAC- andor ACC-related components two control analyses were previ-ously carried out Firstly ANOVAs on the assessmentsgiven by subjects to each group of stimuli were com-puted in order to confirm that their affective contentwas the one supposed a priori As explained in theMethods section each subject filled out a bidimensionalscaling test for each group of pictures after the recordingsessions This test assessed the valence and the arousalcontent of the pictures presented as targets Table 2shows means and standard error of means for each typeof stimulus One-way repeated-measure ANOVAs were
Figure 5 Topographic maps representing the mean values (in microvolts) for the periods (in milliseconds) indicated at the top of each columnThese windows correspond respectively to N280 and SN from pretarget recordings and to P200 P340 P500 and SW from posttarget recordingsA+=arousing-positive A =arousing-negative N=neutral R=relaxing
Carretie et al 1115
computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

algorithms To the best of our knowledge and surpris-ingly ERP data on the interaction between emotion andvisual attention have not been provided to date There-fore the main goal of the present study was to remedythis situation A wide conception of this interactionshould take into account several forms of attention Inthis regard the first specific objective was to explore thetwo types of attention previously mentioned expect-ancy-related and input processing-related attention Thesecond objective was to study the temporal character-istics of both types of attention and particularly thoseassociated with the positive or negative valence of theemotional stimulation Finally the third objective was toexplore the possibility that subprocesses within eachtype of attention present to some extent different brainsources
Expectance is usually elicited in ERP research throughthe experimental paradigm employed to evoke thecontingent negative variation (CNV) In its traditionalform this paradigm (that due to its timing character-istics cannot be employed in hemodynamic studies)requires the presentation in each trial of a warningsignal (S1) and subsequently of an lsquoimperativersquo stimulus(S2) (Walter Cooper Aldridge McCallum amp Winter1964) The onset of S2 requires a rapid response fromsubjects (eg to press a button as rapidly as possible inorder to terminate S2) During the interval between S1and S2 in which the subject is alert the CNV appearsterminating just after S2 onset CNV amplitude relatesdirectly to expectancy-related attention to S2 (McCal-lum 1988) If S2 consists of a visual image and if itsvisual exploration is required by the task ERP elicited bythis second stimulus is also able to reflect input process-ing-related attention
Four emotional categories were studied in the presentexperiment in which an S1 (hereafter labeled lsquocuersquo)- S2(lsquotargetrsquo) type task was employed arousing-positive(A+) arousing-negative (A ) neutral (N) and relaxing(R) These categories are intended to cover the maincombinations of positivity negativity and arousal inorder to study their influence on attention (as explainedelsewhere [Lang et al 1997] certain combinations suchas R are virtually impossible) Cue stimuli were simplewhite-on-black schematic line drawings or symbols Asdescribed in detail in the Methods section each symbolwas identified in the instructions with a particular objector motif (eg insect building etc) Cue presentationconsisted of two drawings or symbols belonging to thesame emotional category one above the other (seeFigure 12) The ideographic-type structure of cues en-sured that the stimuli were physically homogeneouswith no significant differences in their complexity colorbrightness contrast or size Also this type of visual cuewas preferred to other types such as words becausethey provided the pictorial nature of the target stimuli(consisting of photographs as explained below) thusfacilitating the involvement of similar processing systems
in response to both cue and target stimuli Moreoversimple ideographic stimuli would be less prone tohaving deep intrinsic emotional significance at least bycomparison with photographs or realistic drawings Acolor photograph (target) was presented 1750 mseclater (see Figure 11) Photographs always representedone of the motifs symbolized by the two drawings of thecue presentation Participants were told to identifyverbally which of the two drawings of the cue corre-sponded to the target (verbal response was given 2 secafter target onset) The instructions did not explicitlyestablish that the experiment dealt with emotionalreactions and as explained the cue indicated only inan implicit way the emotional category of the followingtarget This strategy had two advantages First it helpedto avoid a situation whereby participants considered thatsome of the stimuli were more important for the taskthan others (eg emotional stimuli more importantthan neutral ones) and thus to avoid a relevance-for-task effect often described in previous studies (egDuncan-Johnson amp Donchin 1977 the stimuli on whichthe task focuses tend to elicit the highest amplitudes incertain endogenous ERP components) Second homog-enization of cognitive demands makes it easier to reachmore solid conclusions about the emotion-related cau-sation of possible attentional differences
In general tasks in which target information is pre-dicted (100 correctly announced) by a cue have acharacteristic that is very interesting and relevant tothe present study They allow for the random presenta-tion of different stimuli minimizing the effect of theorienting response at least in its relation to novelty ornonexpectance of stimulation Since only active selec-tive attention was of interest in the present study theorienting response could represent an important inter-ference for posttarget ERPs Moreover in natural envi-ronments situations in which subjects perceive somecues or signals that trigger affective-related expectanceand prepare them for possible forthcoming emotionalevents are relatively frequent These situations have asyet scarcely been investigated in relation to brain activityThe study of input processing-related attention requiredthat only attended stimuli (cues and targets) wereanalyzed To ensure this recordings obtained in trialsin which subjects responded erroneously were elimina-ted It was also necessary to avoid emotional categoriesbeing heterogeneous with respect to the level of atten-tion needed to carry out the task correctly differencesshould be due to the affective content of each categorybut not to its difficulty with respect to the task Controlanalyses were planned in order to discard if necessarythose categories differing in this aspect from the rest
Our first hypothesis is that expectancy-related andinput processing-related phases of the task which asexplained associate with different types of attention willbe characterized by the activation of different brainareas The former should associate with the activation
Carretie et al 1111
of prefrontal areas involving the ACC and the latterwith VAC areas Secondly we consider that expectancy-related attention elicited by cues and input processing-related attention elicited by targets will be differentiallyaffected by the negativity bias Previous data on expect-ance towards emotional stimuli indicate that cues an-nouncing the presentation of stimuli to subjects whohave previously rated them as highly negative elicitlower CNV (particularly lower early CNV) amplitudesthan nonnegative pictures (Yee amp Miller 1987 Klormanamp Ryan 1980) On the other hand ERP signs of greaterand more prominent responses after the presentation ofnegative stimulation have been recently reported (Car-retie Mercado Tapia amp Hinojosa 2001 Ito LarsenSmith amp Cacioppo 1998) These data suggest thatbiases favoring an urgent motor response to negative(eg aversive) stimulation mainly occur when the stim-ulus has already appeared On the other hand the cue isa signal not to respond but rather to process forth-coming stimuli Therefore this second hypothesis maybe formulated indicating that negativity bias will affectposttarget ERPs to a greater extent than pretargetrecordings The third hypothesis deals with the spatial
location of each subphase of the attention-related activ-ity We expect spatial characteristics of attention to beparallel to temporal characteristics and to negativity biasie spatial differences will be related to urgency ofwithdrawal motivation (when avoidance stimulation ispresented) or to the need for deeper processing (whennonaversive stimulation is presented) and will alsoaffect preferentially posttarget ERPs
RESULTS
Figures 1 and 2 in which 14 widely distributed recordingchannels have been selected from the 58 channelsactually used show the grand averages of pre- andposttarget (respectively) recordings once the baselinehad been subtracted from each ERP
Selection of Components
Temporal Principal Component Analyses
Components explaining most pre- and posttarget ERPvariance were extracted through temporal principalcomponent analyses (tPCAs) Two covariance-matrix-
Figure 1 Grand averages inresponse to the four groups ofcue (pretarget ERPs) obtainedfrom the 29 subjects at 14selected scalp locations fromthe 58 locations actually em-ployed A+=arousing-positiveA =arousing-negativeN=neutral R=relaxing
1112 Journal of Cognitive Neuroscience Volume 13 Number 8
based tPCAs were carried out one for pre- and the otherfor posttarget ERPs Since the original number of cases(58 channels 29 subjects 4 stimulus conditions) andthe number of variables (487 digitized points for pre-and 250 points for posttarget ERPs) were excessive forcomputations two operations for data reduction main-taining the relevant ERP information were carried outFirstly a representative widely distributed sample of 28channels was selected (see Methods) Secondly toreduce the amount of data (only for tPCAs) we averagedeach five adjacent digitized points for each ERP (eachfive adjacent points represent 20 msec) The decision onthe number of components to select was based on thescree test (see eg Cliff 1987 for a description)Extracted components were then submitted to varimaxrotation
Following this selection criterion two components orfactors were extracted from pretarget ERPs Factor 1explained 9597 of total variance and Factor 2 ex-plained 124 (6581 and 3140 respectively afterrotation) Figure 3 shows the rotated loadings plottedover time for these two components Additionally Fig-ure 4 presents the mean factor scores for each scalp
location and for both components Basically factorscores which are calculated for each individual ERPreflect the product of point loading by point amplitudeTherefore amplitude and factor score are directly-re-lated parameters Analysis of peak latencies observed inFigure 3 and topography deduced from Figure 4 asso-ciate Factor 1 with the homogeneously distributed longlatency negative variation marked with an arrow labeledslow negativity (SN) in grand averages (Figure 1) andFactor 2 with a relatively early negativity (peaking at 280msec) with frontal midline dominance (see the mostnegative locations in Figure 4) signaled as N280 in grandaverages
With respect to posttarget ERPs selection yielded fourfactors Factor 1 explained 5062 of total varianceFactor 2 explained 2599 Factor 3 explained 532and Factor 4 342 (3595 3444 1073 and 424respectively after rotation) Figure 3 shows the rotatedloadings plotted over time for these four componentsMean factor scores for each recording channel areshown in Figure 4 Factor peak latency and topographycharacteristics associate Factor 1 maximal at centrallocations with the negative slow wave signaled with an
Figure 2 Grand averages inresponse to the four groups oftargets obtained from the 29subjects at 14 selected scalplocations from the 58 locationsactually employed A+=arous-ing-positive A =arousing-ne-gative N=neutral R=relaxing
Carretie et al 1113
arrow labeled SW in grand averages (Figure 2) Factors 2(340 msec) and 3 (200 msec) both maximal at parietaland parieto-occipital sites can be associated with de-flections marked as P340 and P200 respectively inFigure 2 Finally Factor 4 (peak at 500 msec) maximalat centro-parietal sites with a slow positive deflectionlabeled as P500 in grand averages These labels will beemployed hereafter to make results more understand-able and do not necessarily imply the identification ofthese factors with lsquoclassicalrsquo components such as P300 ortraditional SW (in the present case SW could simplyreflect a CNV release)
Selection of VAC- andor ACC-Related Components
Though the experimental design ensured that subjectsattended to the stimuli the extent of the association ofthe six components extracted by the tPCAs with theattentional processes we are studying could vary con-siderably Before submitting factors to analyses on theexperimental effects it was necessary to discard thosemainly reflecting the activity of brain areas different from
those under study As explained in the Introduction thefocus of our interest were those components reflectingactivation of visual cortices (input processing-relatedattention) andor prefrontal structures associated withexpectancy-related attention such as the ACC In orderto analyze the extent to which each factor was associatedwith these relevant brain areas topographic and source-localization analyses were carried out taking into ac-count voltages recorded in the 58 original recordingchannels Figure 5 shows the maps of the averageactivity in the 200- 520- and 740- 1750-msec periods ofpretarget ERPs (corresponding to N280 and SN respec-tively) and in the 176- 224- 312- 368- 472- 528- and620- 800-msec periods of posttarget ERPs (P200 P340P500 and SW respectively) The size of these windowswas proportional to the component period both withrespect to its absolute value and in relation to the totalERP epoch and shape of components was also takeninto account For example N280pre presents two high-loaded peaks not separable according to tPCAs thatshould be included in the window in order to correctlytake into account the main activity of this component
PRE-TARGET
02
03
04
05
06
07
08
09
1
0 500 1000 1500
msec
Lo
ad
s
F1PRE
F2PRE
POST-TARGET
-04
-02
0
02
04
06
08
1
0 200 400 600 800
msec
F1POSTF2POSTF3POSTF4POST
Lo
ad
s
Figure 3 tPCA analysis factor loadings after Varimax rotation Toppretarget ERPs Bottom posttarget ERPs
POST-TARGET
-03
-02
-01
0
01
02
03
AF3
AF4 F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
F1POSTF2POSTF3POSTF4POST
PRE-TARGET
-08
-06
-04
-02
0
02
04
06
08
AF
3
AF
4
F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
Fa
ct
or
Sc
or
es
F1PREF2PRE
Fa
ct
or
Sc
or
es
Figure 4 tPCA analysis mean factor scores corresponding to the 28recording channels taken into account in this analysis Top pretargetERPs Bottom posttarget ERPs
1114 Journal of Cognitive Neuroscience Volume 13 Number 8
Additionally low-resolution brain electromagnetic to-mography (LORETA) was applied a 3-D discrete linearsolution for the EEG inverse problem (Pascual-MarquiMichel amp Lehman 1994 Pascual-Marqui 1999) In itscurrent version LORETA refers to a three-shell sphericalmodel registered to the Talairach human brain atlas(Talairach amp Tournoux 1988) Solution is given there-fore in three coordinates x is the distance in milli-meters to the right (+) or left ( ) of midline y is thedistance anterior (+) or posterior ( ) to the anteriorcommissure and z is the distance superior (+) orinferior ( ) to a horizontal plane through the anteriorand posterior commissures Registration between spher-ical and the realistic head model in which solutions aregiven by LORETA used EEG electrode coordinates re-ported by Towle et al (1993) Figure 6 illustrates thesources of activity for the six components as calculatedby LORETA Source analysis were carried out on theaverage ERPs in response to the four groups of stimuliScalp distribution and source analyses suggest thatP500 is not originated in visual cortices or in the ACCor related prefrontal areas (see Table 1) Therefore
components selected for later analyses on the exper-imental effects were N280 and SN from pretargetrecordings and P200 P340 and SW from posttargetrecordings
Effects of Emotional Content on SelectedComponents
Control Analyses
In order to better interpret the results on the effects ofemotional stimulation on selected VAC- andor ACC-related components two control analyses were previ-ously carried out Firstly ANOVAs on the assessmentsgiven by subjects to each group of stimuli were com-puted in order to confirm that their affective contentwas the one supposed a priori As explained in theMethods section each subject filled out a bidimensionalscaling test for each group of pictures after the recordingsessions This test assessed the valence and the arousalcontent of the pictures presented as targets Table 2shows means and standard error of means for each typeof stimulus One-way repeated-measure ANOVAs were
Figure 5 Topographic maps representing the mean values (in microvolts) for the periods (in milliseconds) indicated at the top of each columnThese windows correspond respectively to N280 and SN from pretarget recordings and to P200 P340 P500 and SW from posttarget recordingsA+=arousing-positive A =arousing-negative N=neutral R=relaxing
Carretie et al 1115
computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

of prefrontal areas involving the ACC and the latterwith VAC areas Secondly we consider that expectancy-related attention elicited by cues and input processing-related attention elicited by targets will be differentiallyaffected by the negativity bias Previous data on expect-ance towards emotional stimuli indicate that cues an-nouncing the presentation of stimuli to subjects whohave previously rated them as highly negative elicitlower CNV (particularly lower early CNV) amplitudesthan nonnegative pictures (Yee amp Miller 1987 Klormanamp Ryan 1980) On the other hand ERP signs of greaterand more prominent responses after the presentation ofnegative stimulation have been recently reported (Car-retie Mercado Tapia amp Hinojosa 2001 Ito LarsenSmith amp Cacioppo 1998) These data suggest thatbiases favoring an urgent motor response to negative(eg aversive) stimulation mainly occur when the stim-ulus has already appeared On the other hand the cue isa signal not to respond but rather to process forth-coming stimuli Therefore this second hypothesis maybe formulated indicating that negativity bias will affectposttarget ERPs to a greater extent than pretargetrecordings The third hypothesis deals with the spatial
location of each subphase of the attention-related activ-ity We expect spatial characteristics of attention to beparallel to temporal characteristics and to negativity biasie spatial differences will be related to urgency ofwithdrawal motivation (when avoidance stimulation ispresented) or to the need for deeper processing (whennonaversive stimulation is presented) and will alsoaffect preferentially posttarget ERPs
RESULTS
Figures 1 and 2 in which 14 widely distributed recordingchannels have been selected from the 58 channelsactually used show the grand averages of pre- andposttarget (respectively) recordings once the baselinehad been subtracted from each ERP
Selection of Components
Temporal Principal Component Analyses
Components explaining most pre- and posttarget ERPvariance were extracted through temporal principalcomponent analyses (tPCAs) Two covariance-matrix-
Figure 1 Grand averages inresponse to the four groups ofcue (pretarget ERPs) obtainedfrom the 29 subjects at 14selected scalp locations fromthe 58 locations actually em-ployed A+=arousing-positiveA =arousing-negativeN=neutral R=relaxing
1112 Journal of Cognitive Neuroscience Volume 13 Number 8
based tPCAs were carried out one for pre- and the otherfor posttarget ERPs Since the original number of cases(58 channels 29 subjects 4 stimulus conditions) andthe number of variables (487 digitized points for pre-and 250 points for posttarget ERPs) were excessive forcomputations two operations for data reduction main-taining the relevant ERP information were carried outFirstly a representative widely distributed sample of 28channels was selected (see Methods) Secondly toreduce the amount of data (only for tPCAs) we averagedeach five adjacent digitized points for each ERP (eachfive adjacent points represent 20 msec) The decision onthe number of components to select was based on thescree test (see eg Cliff 1987 for a description)Extracted components were then submitted to varimaxrotation
Following this selection criterion two components orfactors were extracted from pretarget ERPs Factor 1explained 9597 of total variance and Factor 2 ex-plained 124 (6581 and 3140 respectively afterrotation) Figure 3 shows the rotated loadings plottedover time for these two components Additionally Fig-ure 4 presents the mean factor scores for each scalp
location and for both components Basically factorscores which are calculated for each individual ERPreflect the product of point loading by point amplitudeTherefore amplitude and factor score are directly-re-lated parameters Analysis of peak latencies observed inFigure 3 and topography deduced from Figure 4 asso-ciate Factor 1 with the homogeneously distributed longlatency negative variation marked with an arrow labeledslow negativity (SN) in grand averages (Figure 1) andFactor 2 with a relatively early negativity (peaking at 280msec) with frontal midline dominance (see the mostnegative locations in Figure 4) signaled as N280 in grandaverages
With respect to posttarget ERPs selection yielded fourfactors Factor 1 explained 5062 of total varianceFactor 2 explained 2599 Factor 3 explained 532and Factor 4 342 (3595 3444 1073 and 424respectively after rotation) Figure 3 shows the rotatedloadings plotted over time for these four componentsMean factor scores for each recording channel areshown in Figure 4 Factor peak latency and topographycharacteristics associate Factor 1 maximal at centrallocations with the negative slow wave signaled with an
Figure 2 Grand averages inresponse to the four groups oftargets obtained from the 29subjects at 14 selected scalplocations from the 58 locationsactually employed A+=arous-ing-positive A =arousing-ne-gative N=neutral R=relaxing
Carretie et al 1113
arrow labeled SW in grand averages (Figure 2) Factors 2(340 msec) and 3 (200 msec) both maximal at parietaland parieto-occipital sites can be associated with de-flections marked as P340 and P200 respectively inFigure 2 Finally Factor 4 (peak at 500 msec) maximalat centro-parietal sites with a slow positive deflectionlabeled as P500 in grand averages These labels will beemployed hereafter to make results more understand-able and do not necessarily imply the identification ofthese factors with lsquoclassicalrsquo components such as P300 ortraditional SW (in the present case SW could simplyreflect a CNV release)
Selection of VAC- andor ACC-Related Components
Though the experimental design ensured that subjectsattended to the stimuli the extent of the association ofthe six components extracted by the tPCAs with theattentional processes we are studying could vary con-siderably Before submitting factors to analyses on theexperimental effects it was necessary to discard thosemainly reflecting the activity of brain areas different from
those under study As explained in the Introduction thefocus of our interest were those components reflectingactivation of visual cortices (input processing-relatedattention) andor prefrontal structures associated withexpectancy-related attention such as the ACC In orderto analyze the extent to which each factor was associatedwith these relevant brain areas topographic and source-localization analyses were carried out taking into ac-count voltages recorded in the 58 original recordingchannels Figure 5 shows the maps of the averageactivity in the 200- 520- and 740- 1750-msec periods ofpretarget ERPs (corresponding to N280 and SN respec-tively) and in the 176- 224- 312- 368- 472- 528- and620- 800-msec periods of posttarget ERPs (P200 P340P500 and SW respectively) The size of these windowswas proportional to the component period both withrespect to its absolute value and in relation to the totalERP epoch and shape of components was also takeninto account For example N280pre presents two high-loaded peaks not separable according to tPCAs thatshould be included in the window in order to correctlytake into account the main activity of this component
PRE-TARGET
02
03
04
05
06
07
08
09
1
0 500 1000 1500
msec
Lo
ad
s
F1PRE
F2PRE
POST-TARGET
-04
-02
0
02
04
06
08
1
0 200 400 600 800
msec
F1POSTF2POSTF3POSTF4POST
Lo
ad
s
Figure 3 tPCA analysis factor loadings after Varimax rotation Toppretarget ERPs Bottom posttarget ERPs
POST-TARGET
-03
-02
-01
0
01
02
03
AF3
AF4 F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
F1POSTF2POSTF3POSTF4POST
PRE-TARGET
-08
-06
-04
-02
0
02
04
06
08
AF
3
AF
4
F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
Fa
ct
or
Sc
or
es
F1PREF2PRE
Fa
ct
or
Sc
or
es
Figure 4 tPCA analysis mean factor scores corresponding to the 28recording channels taken into account in this analysis Top pretargetERPs Bottom posttarget ERPs
1114 Journal of Cognitive Neuroscience Volume 13 Number 8
Additionally low-resolution brain electromagnetic to-mography (LORETA) was applied a 3-D discrete linearsolution for the EEG inverse problem (Pascual-MarquiMichel amp Lehman 1994 Pascual-Marqui 1999) In itscurrent version LORETA refers to a three-shell sphericalmodel registered to the Talairach human brain atlas(Talairach amp Tournoux 1988) Solution is given there-fore in three coordinates x is the distance in milli-meters to the right (+) or left ( ) of midline y is thedistance anterior (+) or posterior ( ) to the anteriorcommissure and z is the distance superior (+) orinferior ( ) to a horizontal plane through the anteriorand posterior commissures Registration between spher-ical and the realistic head model in which solutions aregiven by LORETA used EEG electrode coordinates re-ported by Towle et al (1993) Figure 6 illustrates thesources of activity for the six components as calculatedby LORETA Source analysis were carried out on theaverage ERPs in response to the four groups of stimuliScalp distribution and source analyses suggest thatP500 is not originated in visual cortices or in the ACCor related prefrontal areas (see Table 1) Therefore
components selected for later analyses on the exper-imental effects were N280 and SN from pretargetrecordings and P200 P340 and SW from posttargetrecordings
Effects of Emotional Content on SelectedComponents
Control Analyses
In order to better interpret the results on the effects ofemotional stimulation on selected VAC- andor ACC-related components two control analyses were previ-ously carried out Firstly ANOVAs on the assessmentsgiven by subjects to each group of stimuli were com-puted in order to confirm that their affective contentwas the one supposed a priori As explained in theMethods section each subject filled out a bidimensionalscaling test for each group of pictures after the recordingsessions This test assessed the valence and the arousalcontent of the pictures presented as targets Table 2shows means and standard error of means for each typeof stimulus One-way repeated-measure ANOVAs were
Figure 5 Topographic maps representing the mean values (in microvolts) for the periods (in milliseconds) indicated at the top of each columnThese windows correspond respectively to N280 and SN from pretarget recordings and to P200 P340 P500 and SW from posttarget recordingsA+=arousing-positive A =arousing-negative N=neutral R=relaxing
Carretie et al 1115
computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

based tPCAs were carried out one for pre- and the otherfor posttarget ERPs Since the original number of cases(58 channels 29 subjects 4 stimulus conditions) andthe number of variables (487 digitized points for pre-and 250 points for posttarget ERPs) were excessive forcomputations two operations for data reduction main-taining the relevant ERP information were carried outFirstly a representative widely distributed sample of 28channels was selected (see Methods) Secondly toreduce the amount of data (only for tPCAs) we averagedeach five adjacent digitized points for each ERP (eachfive adjacent points represent 20 msec) The decision onthe number of components to select was based on thescree test (see eg Cliff 1987 for a description)Extracted components were then submitted to varimaxrotation
Following this selection criterion two components orfactors were extracted from pretarget ERPs Factor 1explained 9597 of total variance and Factor 2 ex-plained 124 (6581 and 3140 respectively afterrotation) Figure 3 shows the rotated loadings plottedover time for these two components Additionally Fig-ure 4 presents the mean factor scores for each scalp
location and for both components Basically factorscores which are calculated for each individual ERPreflect the product of point loading by point amplitudeTherefore amplitude and factor score are directly-re-lated parameters Analysis of peak latencies observed inFigure 3 and topography deduced from Figure 4 asso-ciate Factor 1 with the homogeneously distributed longlatency negative variation marked with an arrow labeledslow negativity (SN) in grand averages (Figure 1) andFactor 2 with a relatively early negativity (peaking at 280msec) with frontal midline dominance (see the mostnegative locations in Figure 4) signaled as N280 in grandaverages
With respect to posttarget ERPs selection yielded fourfactors Factor 1 explained 5062 of total varianceFactor 2 explained 2599 Factor 3 explained 532and Factor 4 342 (3595 3444 1073 and 424respectively after rotation) Figure 3 shows the rotatedloadings plotted over time for these four componentsMean factor scores for each recording channel areshown in Figure 4 Factor peak latency and topographycharacteristics associate Factor 1 maximal at centrallocations with the negative slow wave signaled with an
Figure 2 Grand averages inresponse to the four groups oftargets obtained from the 29subjects at 14 selected scalplocations from the 58 locationsactually employed A+=arous-ing-positive A =arousing-ne-gative N=neutral R=relaxing
Carretie et al 1113
arrow labeled SW in grand averages (Figure 2) Factors 2(340 msec) and 3 (200 msec) both maximal at parietaland parieto-occipital sites can be associated with de-flections marked as P340 and P200 respectively inFigure 2 Finally Factor 4 (peak at 500 msec) maximalat centro-parietal sites with a slow positive deflectionlabeled as P500 in grand averages These labels will beemployed hereafter to make results more understand-able and do not necessarily imply the identification ofthese factors with lsquoclassicalrsquo components such as P300 ortraditional SW (in the present case SW could simplyreflect a CNV release)
Selection of VAC- andor ACC-Related Components
Though the experimental design ensured that subjectsattended to the stimuli the extent of the association ofthe six components extracted by the tPCAs with theattentional processes we are studying could vary con-siderably Before submitting factors to analyses on theexperimental effects it was necessary to discard thosemainly reflecting the activity of brain areas different from
those under study As explained in the Introduction thefocus of our interest were those components reflectingactivation of visual cortices (input processing-relatedattention) andor prefrontal structures associated withexpectancy-related attention such as the ACC In orderto analyze the extent to which each factor was associatedwith these relevant brain areas topographic and source-localization analyses were carried out taking into ac-count voltages recorded in the 58 original recordingchannels Figure 5 shows the maps of the averageactivity in the 200- 520- and 740- 1750-msec periods ofpretarget ERPs (corresponding to N280 and SN respec-tively) and in the 176- 224- 312- 368- 472- 528- and620- 800-msec periods of posttarget ERPs (P200 P340P500 and SW respectively) The size of these windowswas proportional to the component period both withrespect to its absolute value and in relation to the totalERP epoch and shape of components was also takeninto account For example N280pre presents two high-loaded peaks not separable according to tPCAs thatshould be included in the window in order to correctlytake into account the main activity of this component
PRE-TARGET
02
03
04
05
06
07
08
09
1
0 500 1000 1500
msec
Lo
ad
s
F1PRE
F2PRE
POST-TARGET
-04
-02
0
02
04
06
08
1
0 200 400 600 800
msec
F1POSTF2POSTF3POSTF4POST
Lo
ad
s
Figure 3 tPCA analysis factor loadings after Varimax rotation Toppretarget ERPs Bottom posttarget ERPs
POST-TARGET
-03
-02
-01
0
01
02
03
AF3
AF4 F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
F1POSTF2POSTF3POSTF4POST
PRE-TARGET
-08
-06
-04
-02
0
02
04
06
08
AF
3
AF
4
F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
Fa
ct
or
Sc
or
es
F1PREF2PRE
Fa
ct
or
Sc
or
es
Figure 4 tPCA analysis mean factor scores corresponding to the 28recording channels taken into account in this analysis Top pretargetERPs Bottom posttarget ERPs
1114 Journal of Cognitive Neuroscience Volume 13 Number 8
Additionally low-resolution brain electromagnetic to-mography (LORETA) was applied a 3-D discrete linearsolution for the EEG inverse problem (Pascual-MarquiMichel amp Lehman 1994 Pascual-Marqui 1999) In itscurrent version LORETA refers to a three-shell sphericalmodel registered to the Talairach human brain atlas(Talairach amp Tournoux 1988) Solution is given there-fore in three coordinates x is the distance in milli-meters to the right (+) or left ( ) of midline y is thedistance anterior (+) or posterior ( ) to the anteriorcommissure and z is the distance superior (+) orinferior ( ) to a horizontal plane through the anteriorand posterior commissures Registration between spher-ical and the realistic head model in which solutions aregiven by LORETA used EEG electrode coordinates re-ported by Towle et al (1993) Figure 6 illustrates thesources of activity for the six components as calculatedby LORETA Source analysis were carried out on theaverage ERPs in response to the four groups of stimuliScalp distribution and source analyses suggest thatP500 is not originated in visual cortices or in the ACCor related prefrontal areas (see Table 1) Therefore
components selected for later analyses on the exper-imental effects were N280 and SN from pretargetrecordings and P200 P340 and SW from posttargetrecordings
Effects of Emotional Content on SelectedComponents
Control Analyses
In order to better interpret the results on the effects ofemotional stimulation on selected VAC- andor ACC-related components two control analyses were previ-ously carried out Firstly ANOVAs on the assessmentsgiven by subjects to each group of stimuli were com-puted in order to confirm that their affective contentwas the one supposed a priori As explained in theMethods section each subject filled out a bidimensionalscaling test for each group of pictures after the recordingsessions This test assessed the valence and the arousalcontent of the pictures presented as targets Table 2shows means and standard error of means for each typeof stimulus One-way repeated-measure ANOVAs were
Figure 5 Topographic maps representing the mean values (in microvolts) for the periods (in milliseconds) indicated at the top of each columnThese windows correspond respectively to N280 and SN from pretarget recordings and to P200 P340 P500 and SW from posttarget recordingsA+=arousing-positive A =arousing-negative N=neutral R=relaxing
Carretie et al 1115
computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

arrow labeled SW in grand averages (Figure 2) Factors 2(340 msec) and 3 (200 msec) both maximal at parietaland parieto-occipital sites can be associated with de-flections marked as P340 and P200 respectively inFigure 2 Finally Factor 4 (peak at 500 msec) maximalat centro-parietal sites with a slow positive deflectionlabeled as P500 in grand averages These labels will beemployed hereafter to make results more understand-able and do not necessarily imply the identification ofthese factors with lsquoclassicalrsquo components such as P300 ortraditional SW (in the present case SW could simplyreflect a CNV release)
Selection of VAC- andor ACC-Related Components
Though the experimental design ensured that subjectsattended to the stimuli the extent of the association ofthe six components extracted by the tPCAs with theattentional processes we are studying could vary con-siderably Before submitting factors to analyses on theexperimental effects it was necessary to discard thosemainly reflecting the activity of brain areas different from
those under study As explained in the Introduction thefocus of our interest were those components reflectingactivation of visual cortices (input processing-relatedattention) andor prefrontal structures associated withexpectancy-related attention such as the ACC In orderto analyze the extent to which each factor was associatedwith these relevant brain areas topographic and source-localization analyses were carried out taking into ac-count voltages recorded in the 58 original recordingchannels Figure 5 shows the maps of the averageactivity in the 200- 520- and 740- 1750-msec periods ofpretarget ERPs (corresponding to N280 and SN respec-tively) and in the 176- 224- 312- 368- 472- 528- and620- 800-msec periods of posttarget ERPs (P200 P340P500 and SW respectively) The size of these windowswas proportional to the component period both withrespect to its absolute value and in relation to the totalERP epoch and shape of components was also takeninto account For example N280pre presents two high-loaded peaks not separable according to tPCAs thatshould be included in the window in order to correctlytake into account the main activity of this component
PRE-TARGET
02
03
04
05
06
07
08
09
1
0 500 1000 1500
msec
Lo
ad
s
F1PRE
F2PRE
POST-TARGET
-04
-02
0
02
04
06
08
1
0 200 400 600 800
msec
F1POSTF2POSTF3POSTF4POST
Lo
ad
s
Figure 3 tPCA analysis factor loadings after Varimax rotation Toppretarget ERPs Bottom posttarget ERPs
POST-TARGET
-03
-02
-01
0
01
02
03
AF3
AF4 F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
F1POSTF2POSTF3POSTF4POST
PRE-TARGET
-08
-06
-04
-02
0
02
04
06
08
AF
3
AF
4
F5
F1
F2
F6
FC
1
FC
5
FC
2
FC
6
C1
C5
C2
C6
CP
1
CP
5
CP
2
CP
6
P1
P5
P2
P6
PO
7
PO
1
PO
2
PO
8
O1
O2
Electrode Locations
Fa
ct
or
Sc
or
es
F1PREF2PRE
Fa
ct
or
Sc
or
es
Figure 4 tPCA analysis mean factor scores corresponding to the 28recording channels taken into account in this analysis Top pretargetERPs Bottom posttarget ERPs
1114 Journal of Cognitive Neuroscience Volume 13 Number 8
Additionally low-resolution brain electromagnetic to-mography (LORETA) was applied a 3-D discrete linearsolution for the EEG inverse problem (Pascual-MarquiMichel amp Lehman 1994 Pascual-Marqui 1999) In itscurrent version LORETA refers to a three-shell sphericalmodel registered to the Talairach human brain atlas(Talairach amp Tournoux 1988) Solution is given there-fore in three coordinates x is the distance in milli-meters to the right (+) or left ( ) of midline y is thedistance anterior (+) or posterior ( ) to the anteriorcommissure and z is the distance superior (+) orinferior ( ) to a horizontal plane through the anteriorand posterior commissures Registration between spher-ical and the realistic head model in which solutions aregiven by LORETA used EEG electrode coordinates re-ported by Towle et al (1993) Figure 6 illustrates thesources of activity for the six components as calculatedby LORETA Source analysis were carried out on theaverage ERPs in response to the four groups of stimuliScalp distribution and source analyses suggest thatP500 is not originated in visual cortices or in the ACCor related prefrontal areas (see Table 1) Therefore
components selected for later analyses on the exper-imental effects were N280 and SN from pretargetrecordings and P200 P340 and SW from posttargetrecordings
Effects of Emotional Content on SelectedComponents
Control Analyses
In order to better interpret the results on the effects ofemotional stimulation on selected VAC- andor ACC-related components two control analyses were previ-ously carried out Firstly ANOVAs on the assessmentsgiven by subjects to each group of stimuli were com-puted in order to confirm that their affective contentwas the one supposed a priori As explained in theMethods section each subject filled out a bidimensionalscaling test for each group of pictures after the recordingsessions This test assessed the valence and the arousalcontent of the pictures presented as targets Table 2shows means and standard error of means for each typeof stimulus One-way repeated-measure ANOVAs were
Figure 5 Topographic maps representing the mean values (in microvolts) for the periods (in milliseconds) indicated at the top of each columnThese windows correspond respectively to N280 and SN from pretarget recordings and to P200 P340 P500 and SW from posttarget recordingsA+=arousing-positive A =arousing-negative N=neutral R=relaxing
Carretie et al 1115
computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

Additionally low-resolution brain electromagnetic to-mography (LORETA) was applied a 3-D discrete linearsolution for the EEG inverse problem (Pascual-MarquiMichel amp Lehman 1994 Pascual-Marqui 1999) In itscurrent version LORETA refers to a three-shell sphericalmodel registered to the Talairach human brain atlas(Talairach amp Tournoux 1988) Solution is given there-fore in three coordinates x is the distance in milli-meters to the right (+) or left ( ) of midline y is thedistance anterior (+) or posterior ( ) to the anteriorcommissure and z is the distance superior (+) orinferior ( ) to a horizontal plane through the anteriorand posterior commissures Registration between spher-ical and the realistic head model in which solutions aregiven by LORETA used EEG electrode coordinates re-ported by Towle et al (1993) Figure 6 illustrates thesources of activity for the six components as calculatedby LORETA Source analysis were carried out on theaverage ERPs in response to the four groups of stimuliScalp distribution and source analyses suggest thatP500 is not originated in visual cortices or in the ACCor related prefrontal areas (see Table 1) Therefore
components selected for later analyses on the exper-imental effects were N280 and SN from pretargetrecordings and P200 P340 and SW from posttargetrecordings
Effects of Emotional Content on SelectedComponents
Control Analyses
In order to better interpret the results on the effects ofemotional stimulation on selected VAC- andor ACC-related components two control analyses were previ-ously carried out Firstly ANOVAs on the assessmentsgiven by subjects to each group of stimuli were com-puted in order to confirm that their affective contentwas the one supposed a priori As explained in theMethods section each subject filled out a bidimensionalscaling test for each group of pictures after the recordingsessions This test assessed the valence and the arousalcontent of the pictures presented as targets Table 2shows means and standard error of means for each typeof stimulus One-way repeated-measure ANOVAs were
Figure 5 Topographic maps representing the mean values (in microvolts) for the periods (in milliseconds) indicated at the top of each columnThese windows correspond respectively to N280 and SN from pretarget recordings and to P200 P340 P500 and SW from posttarget recordingsA+=arousing-positive A =arousing-negative N=neutral R=relaxing
Carretie et al 1115
computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

computed for valence and for arousal dimensions usingStimuli (four levels A+ A N and R) as factor TheGreenhouse- Geisser (GG) epsilon correction was ap-plied to adjust the degrees of freedom of the F ratiosPost hoc comparisons were made to determine thesignificance of pairwise contrasts using Tukeyrsquos one-factor HSD procedure ( a = 05) ANOVAs indicated thatStimuli differed in valence as in arousal [F(384) = 7950
p lt 001 GG e = 073 and F(384) = 7865 p lt 001 GGe = 086 respectively] Post hoc contrasts indicated thatA+ and A significantly showed distinct valence but notdistinct arousal A+ and A differed from N and R inarousal On the other hand A+ and R did not differ withrespect to valence but both differed from N and A inthis dimension N and R were distinct in both dimen-sions
Figure 6 Images of neuronalelectric activity averaging theresponses to the four types ofstimuli computed with LORE-TA for the periods and compo-nents (both pre- andposttarget) indicated at the leftside of the figure For eachcomponent three orthogonalbrain views in Talairach spaceare shown sliced through theregion of the maximum activityLeft slice axial seen fromabove nose up center slicesaggital seen from the leftright slice coronal seen fromthe rear Talairach coordinatesx from left (L) to right (R) yfrom posterior (P) to anterior(A) z from inferior to superiorThe location of maximum ac-tivity is given as (xyz) coordi-nates in Talairach space and isgraphically indicated by blacktriangles on the coordinateaxes Anatomical distribution ofthe most active neuronal gen-erators is shown in Table 1
1116 Journal of Cognitive Neuroscience Volume 13 Number 8
Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

Secondly differences with respect to the task difficultyassociated with each group of pictures were analyzed inorder to ensure that the possible effects elicited by thestimuli were not due to this factor The presence ofdifferences may indicate that level of difficulty andtherefore of attention demands is different for eachstimulus category Therefore the number of errors inthe task with respect to each group of stimuli wasanalyzed via one-way repeated-measures ANOVA onfactor Stimuli (means standard error of means andstandard deviations of incorrect answers in the taskappear in Table 2) Differences were not significant[F(384) = 131 p gt 25]
ANOVAs on Experimental Effects
Two-way repeated measures ANOVAs were computedon factor scores (as explained directly related to ampli-
tudes) for selected components The factors were Stim-uli (four levels A+ A N and R) and Channel (28 levelscorresponding to the same sample of 28 channelspreviously analyzed through tPCA) Also in this casethe GG epsilon correction was applied to adjust thedegrees of freedom of the F ratios Post hoc compar-isons were made to determine the significance of pair-wise contrasts using the Bonferroni correction ( a =05) Table 3 shows main results of these ANOVAs Withrespect to pretarget ERPs factor Stimuli produced sig-nificant effects only in N280 Post hoc comparisonsindicated that this negative component presents itshigher amplitudes in response to A+ which differedsignificantly from amplitudes to A but not from thoseto N and R The interaction Stimuli Channel was alsosignificant Figure 7 shows locations where post hoccomparisons found significant differences between stim-uli in the Stimuli Channel interaction mainly pro-
Table 1 Solutions Provided by LORETA with Respect to Components Extracted by the tPCA Both for Pre- and for PosttargetComponents
Component x y z Close Regions (d lt 5 mm)
PRE N280 (200- 520) 4 52 1 BA10 (medial frontal gyrus)
BA32 (ACC)
SN (740- 1750) 3 45 13 BA11 (medial frontal gyrus)
POST P200 (176- 224) 53 60 8 BA39 (posterior middle temporal gyrus)
BA37 (posterior middle temporal gyrus)
P340 (312- 368) 53 53 13 BA20 (inferior temporal gyrus)
BA37 (fusiform gyrus)
P500 (472- 528) 3 46 43 BA7 (precuneus)
BA31 (posterior cingulate gyrus)
SW (620- 800) 53 60 15 BA22 (superior temporal gyrus)
BA19 (middle temporal gyrus)
BA39 (middle temporal gyrus)
BA44 (superior temporal gyrus)
Talairach coordinates x from left to right y from posterior to anterior z from inferior to superior In brackets and just after the component namesthe temporal interval (in msec) of the analyzed windows
Table 2 Means and in Brackets Standard Error of Means (1) of Assessments Given by Subjects to Stimuli (Arousal and Valence)in the Scaling Test and (2) of Errors Committed in the Task (Percentages with Respect to the Total Number of Trials Are AlsoProvided)
Arousal(Relaxing 2 Arousing 2)
Valence(Negative 2 Positive 2) Number of Errors
Arousing-positive 0586 (0131) 1172 (0139) 0379 (0135) [0296]
Arousing-negative 0983 (0123) 0948 (0145) 0172 (0071) [0134]
Neutral 0000 (0093) 0000 (0124) 0207 (0077) [0162]
Relaxing 1293 (0125) 1638 (0082) 0379 (0104) [0296]
Carretie et al 1117
duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

duced at frontal and central sites bilaterally Post hocanalyses also indicated that differences between hemi-spheres (between symmetrical locations) were not sig-nificant for Factor Channel or for the Stimuli Channelinteraction
With respect to posttarget ERPs factor Stimuli pro-duced significant effects in P200 and P340 This factorhad opposite effects on both components P340 re-vealed that the highest amplitudes were produced inresponse to A+ which significantly differed from thoseelicited by A N and R On the other hand A elicitedthe highest amplitudes in P200 These amplitudes weresignificantly different from those elicited by A+ Theinteraction Stimuli Channel was also significant for thetwo factors As Figure 7 illustrates P340 shows its maindivergences with respect to stimulation at central pari-etal and occipital areas P200 also presents its greatestdifferences between conditions at those scalp areas butin this case they appear mainly at midline and righthemisphere regions Post hoc analyses indicated thatalso in this case differences between symmetrical loca-tions were not produced for Factor Channel or for theinteraction Stimuli Channel in any of the two compo-nents
Additional Spatial Analyses on N280pre P200post andP340post
Additional analyses were carried out in order to moreprecisely define the spatial characteristics of N280preP200post and P340post the three components thatshowed significant amplitude differences as a functionof the experimental effects Firstly LORETA was againapplied to these three components This time only
responses to the maximum-effect stimulus (accordingto ANOVAs) were analyzed The scope of these newlocalization analyses on the responses to the stimulieliciting the highest amplitudes in each of these compo-nents was to ensure the best signal-to-noise ratio for thelocalization of sources Therefore inverse solutionswere calculated from the ERPs elicited by A+ inN280pre and P340post windows and by A in P200postwindow (Figure 8) With respect to the 200- 520-msecwindow of pretarget ERPs (corresponding to N280pre)solution provided by LORETA is a prefrontal focuslocated near midline (x = 4 y = 45 z = 8) lessanteriorly located than in the analyses on the averageof all the stimuli (Table 1 Figure 6) This point is withinthe ACC Brodmannrsquos area (BA) 24 (d lt 1 mm) and BA32 (d lt 4 mm) are located within a radius of 5 mm fromit Moreover BA 10 corresponding to medial frontalgyrus is also close to the solution given by LORETA (d lt4 mm) Main sources provided for the two windows ofposttarget ERPs are the same that were calculated byanalyses on the average of all the stimuli (Table 1 Figure6) Two focuses are provided as solutions for thewindow corresponding to P200post The main focus ismore dorsally located (x = 53 y = 60 z = 8) than thesecond (x = 53 y = 60 z = 6) The posterior middletemporal gyrus (BA 39 d lt 5 mm BA 37 d lt 5 mm) isthe closest area to first focus The second focus is in themiddle occipital gyrus (BA19 d lt 3 mm BA37 d lt 3mm) Finally LORETArsquos solution for the window corre-sponding to P340post also posterior (x = 53 y = 53z = 13) is clearly ventral Inferior temporal gyrus (BA20 d lt 3 mm) and fusiform gyrus (BA 37 d lt 4 mm)both in the temporal lobe are the closest matches forthat source
Table 3 Results of ANOVAs on Component Scores both with Respect to Factor Stimuli and with Respect to the Interaction Stimuliby Channel
Component Factor Stimuli (df = 384)Interaction Stimuli
Channel (df = 812268)
PRE N280 F = 321 plt 05 F = 234 plt 05
GG e = 082 GG eee = 008
SN F = 175 ns F = 178 ns
GG e = 078 GG e = 008
POST P200 F = 412 plt 02 F = 465 plt 001
GG eee = 078 GG eee = 008
P340 F = 702 plt 005 F = 275 plt 05
GG eee = 068 GG eee = 006
SW F = 182 ns F = 204 ns
GG e = 070 GG e = 007
Significant contrasts are in bold
df = degrees of freedom GG = Greenhouse- Geisser ns = nonsignificant
1118 Journal of Cognitive Neuroscience Volume 13 Number 8
Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

Windows selected for source analyses may reflectthe activity of different components Though thesewindows involve the peak of components to analyzethe rest of components could also present significantactivity in them Moreover sources provided by theseanalyses may correspond to components that thoughpresent in the window are not those that are sensitiveto the experimental manipulations In order to testthese possibilities covariance-matrix-based spatial prin-cipal component analyses (sPCAs) were computed onthese three windows and one-way repeated-measuresANOVAs on resulting spatial component scores usingStimuli (four levels A+ A N and R) as factor werecarried out Variables in sPCAs were average record-ings in each of the 58 channels Extracted componentswere submitted to varimax rotation Also in this casethe decision on the number of components to selectwas based on the scree test According to this crite-rion two components were extracted for each win-dow Figure 9 shows in the form of scalp maps theloadings for each channel in each of the spatialfactors The existence of a prefrontal focus of activityin N280pre window and an occipito-parietal sourceboth for P200post and P340post windows was con-firmed by sPCAs Additionally ANOVAs confirmed thatthese spatial factors were sensitive to the experimentalmanipulations as can be seen in Table 4
Regression Analyses
An important question is to estimate the stimulusaspects explaining the differences observed in ANOVAsThough it is reasonable to deduce from these analysesthat valence more than arousal explains results concern-ing the three components since differences betweenA+ and A are clear in them these trends must bestatistically confirmed Therefore the association be-tween ratings given by subjects to stimuli in the ques-tionnaire (valence and arousal) and temporal factorscores was analyzed via multiple regression Three anal-yses were carried out one for each component(N280pre P340post and P200post) For each eightlocations with maximum temporal factor scores wereselected among those showing significant differencesbetween conditions (see Figure 7) in order to analyzethe lsquomost responsiversquo areas Mean temporal factor scorecorresponding to these locations was the dependent
Figure 7 Schematic representation of scalp locations where differ-ences among stimuli were significant in the interaction ChannelStimuli (Bonferroni correction a = 005) lsquoHeadsrsquo correspond toN280pre (top) P200post (middle) and P340post (bottom) Onlylocations where differences among stimuli were produced are clearlyvisible Differences in color for each location represent differencesamong stimuli A+ arousing-positive A arousing-negative Nneutral R relaxing Example upper-left scalp location of first head hasshown a significant difference between A and A+
Carretie et al 1119
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8
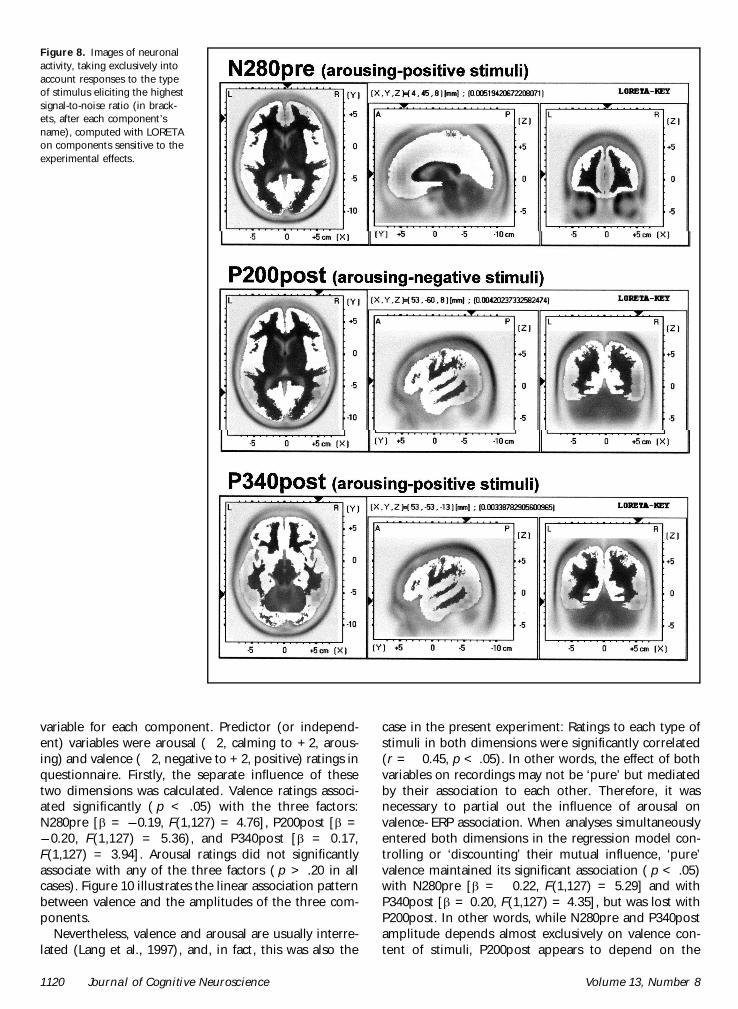
variable for each component Predictor (or independ-ent) variables were arousal ( 2 calming to +2 arous-ing) and valence ( 2 negative to +2 positive) ratings inquestionnaire Firstly the separate influence of thesetwo dimensions was calculated Valence ratings associ-ated significantly ( p lt 05) with the three factorsN280pre [ b = 019 F(1127) = 476] P200post [ b =
020 F(1127) = 536) and P340post [ b = 017F(1127) = 394] Arousal ratings did not significantlyassociate with any of the three factors ( p gt 20 in allcases) Figure 10 illustrates the linear association patternbetween valence and the amplitudes of the three com-ponents
Nevertheless valence and arousal are usually interre-lated (Lang et al 1997) and in fact this was also the
case in the present experiment Ratings to each type ofstimuli in both dimensions were significantly correlated(r = 045 p lt 05) In other words the effect of bothvariables on recordings may not be lsquopurersquo but mediatedby their association to each other Therefore it wasnecessary to partial out the influence of arousal onvalence- ERP association When analyses simultaneouslyentered both dimensions in the regression model con-trolling or lsquodiscountingrsquo their mutual influence lsquopurersquovalence maintained its significant association ( p lt 05)with N280pre [ b = 022 F(1127) = 529] and withP340post [ b = 020 F(1127) = 435] but was lost withP200post In other words while N280pre and P340postamplitude depends almost exclusively on valence con-tent of stimuli P200post appears to depend on the
Figure 8 Images of neuronalactivity taking exclusively intoaccount responses to the typeof stimulus eliciting the highestsignal-to-noise ratio (in brack-ets after each componentrsquosname) computed with LORETAon components sensitive to theexperimental effects
1120 Journal of Cognitive Neuroscience Volume 13 Number 8
combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

combination of valence and arousal contents In order toconfirm this fact the regression was calculated forP200post with respect to the Valence Arousal productIn this case arousal was rescaled from [ 2 to +2] to[0 to +4] in order to avoid artificial balances among
stimulus categories (with the original scale R stimulitend to be negative in arousal and positive in valencewhile A stimuli tend to be negative in valence andpositive in arousal the product of both dimensionswould yield similar values for R and A ) The associ-
Figure 9 Topographic representation of factor loadings for the spatial factors (SF) or components extracted through sPCAs Spatial factors thataccording to ANOVAs (see Table 4) were sensitive to the experimenta l effects
Table 4 Results of ANOVAs on Spatial Component Scores with Respect to Factor Stimuli
Spatial Factor 1 Spatial Factor 2
N280pre F = 216 ns F = 317 p lt 05
GG e = 089 GG eee = 090
[9736 9498] [492 254]
P200post F = 508 plt 01 F = 1020 plt 001
GG eee = 071 GG eee = 087
[9731 6740] [247 3238]
P340post F = 307 plt 05 F = 266 ns
GG eee = 079 GG e = 067
[9816 7688] [166 2285]
In brackets the percentage of variance explained before and after rotation respectively Significant contrasts are in bold
df = 384 in all cases GG = Greenhouse- Geisser ns = nonsignificant
Carretie et al 1121
ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

ation of P200post with [Valence Arousal] wassignificant [ b = 018 F(1127) = 444 p lt 05]
DISCUSSION
Attention-related ERP activity shows differences thatdepend on the emotional content of the stimulationThese differences have been produced even when stim-uli are physically similar (cues) and in a task in whichthe four stimulus categories may be considered identicalwith respect to their cognitive requirements (attentiondemanded by task perceptive and discriminative com-plexity cue memory encoding etc) Valence of theemotional stimulation appears to be the aspect thatpreferentially explains the differences observed in theattention-related ERP components selected in thepresent study However as regression analyses havedemonstrated the influence of arousal through itsinteraction with valence was also significant forP200post
Pretarget Activity
The lowest amplitude of N280pre was elicited by A The task requirements lead to the consideration that thetype of attention reflected in this component should beexpectancy-related The possibility that the observedN280pre effects associate with the input processing-related attention to the cues themselves seems improb-able Cues are simple and homogeneous drawings (Fig-ure 10) and it is difficult to find a characteristic in A+ Nand R cues themselves that could capture attention to agreater extent than A cues Secondly input processing-related attention should associate preferentially withthe VAC but experimental effects have been observed inthe frontal spatial component of N280pre Source anal-ysis suggests that this spatial component originates inthe ACC and medial prefrontal cortex
The ACC is associated as explained in the Introduc-tion to expectancy-related attention Additionally theACC has been reported to be activated in anticipationof affective events (Ploghaus et al 1999 MurthaChertkov Beauregard Dixon amp Evans 1996) Coordi-nates given by LORETA for the frontal focus in the twoanalyses (on average recordings for all the stimuli andon recordings elicited by the stimulus evoking thehighest N280pre amplitudes) mark a point in thevicinity of the lsquoaffective divisionrsquo of the ACC (ACad)one of the two functional and cytoarchitectural regions(the other is the lsquocognitive divisionrsquo) in which recentstudies suggest that the ACC could be subdivided(Bush Luu amp Posner 2000) In fact the ACad signifi-cantly increases its activity in tasks in which emotionalinformation is involved (eg emotional Stroop tasksWhalen et al 1998) The medial prefrontal cortex isactivated in a number of neuroimaging studies present-ing emotional stimulation (Teasdale et al 1999 LaneReiman et al 1997 Reiman et al 1997) It is impor-tant to underline here that recent data suggest it playsan important role in visual attention processes in
Figure 10 Scatterplots of valence assessments and N280pre (top)P200post (middle) and P340post (bottom) factor scores Regressionlines are also shown
1122 Journal of Cognitive Neuroscience Volume 13 Number 8
rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

rodents (Bussey Muir Everit amp Robbins 1997) and itsclose relation with the ACC (particularly BA 32) andattentional processes in humans has been proposed(Lane Fink Chau amp Dolan 1997 Lane et al 1999)Additionally it has been reported that the ventromedialsector of the prefrontal cortex is also involved in theanticipation of future affective consequences (see areview in Bechara Damasio amp Damasio 2000 inrelation to this the possibility that anticipation of theemotional state evoked by the thought of targetscontributes to the activity reflected in N280pre shouldbe taken into account) All these functional profilesdescribing the ACC and medial prefrontal cortex makethem candidate areas to develop an important role inrelation to emotion and attention interactions in ex-pectancy and anticipatory processes such as thosefacilitated by present task In this particular caseresults indicate that at least in anticipatory processesthe activity in these areas decreases as threat value ofexpected stimulation increases
It has been indicated that error-related negativity(ERN) a negative ERP component that appears whensubjects make an erroneous response (Gehring GossColes Meyer amp Donchin 1993) originates in the vicinityof the ACC (Dahaene Posner amp Tucker 1994) ERN hasbeen related to negative affect (Luu Collins amp Tucker2000) a result that could be interpreted as opposed topresent data However both the experimental method-ology by which the ERN is evoked (conspicuously differ-ent from the one employed here) and the functional andanatomical richness of the ACC (which is capable ofeliciting processes of very diverse nature in its differentareas) make it improbable that N280pre and ERN arerelated components N280pre should be related to CNVAs explained in the Introduction this negative compo-nent appears in S1- S2 experimental paradigms such asthe one used here and has often been described as anlsquoexpectancyrsquo wave The relationship of CNV amplitude toexpectancy-related attention to S2 is direct (for a reviewon attention and CNV see McCallum 1988) ParticularlyN280pre could be identified as an early CNV While lateCNV mainly reflecting preparation to respond to S2 ismaximal at central- parietal locations early CNV is mainlyfound over the frontal lobes (Leynes Allen amp Marsh1998 Damen amp Brunia 1994 Rohrbaugh amp Gaillard1983) Dipole localization studies have placed the originof early CNV in prefrontal areas (Basile Rogers Bour-bon amp Papanicolau 1994) Previous ERP data alsosuggest that early CNV decreases its amplitude whenS2 consists of negative images Klorman and Ryan(1980) and later Yee and Miller (1987) presented tonesas S1 and slides of negative and nonnegative images asS2 Early CNV amplitude was significantly lower whenforthcoming S2 was an unpleasant image in subjectswho ranked those images as highly negative These dataand those of the present study support the idea thatACCmedial frontal cortex activity decreases when neg-
ative events are expected or anticipated though resultson ERN suggest that a different pattern of activity maybe produced in these frontal regions in nonanticipatoryaffective processes This lsquodouble behaviorrsquo could explainwhy some hemodynamic studies fail to find significantdifferences in the response of these areas to positiveand negative stimuli (emotional vs neutral differencesbeing the only found in them Lane 1999 Reiman et al1997)
Posttarget Activity
The task requirements lead to the consideration that thetype of attention reflected in P200post and P340post ismainly associated with input processing-related atten-tion At this point of the task subjects had to process arelatively complex image (as compared to cue stimuli) inorder to provide a correct response Moreover compo-nents selected from these posttarget ERPs originated inthe VAC which is related to this sort of attention It isdifficult to compare the present data with those ob-tained from previous ERP studies and identification ofpresent components with previously reported compo-nents cannot be direct As explained in the Introductionand to our knowledge experiments specifically orientedto the study of visual attention and emotion interactionhave not been carried out up to the present Thusattentional components are usually outside the analysesof ERPs elicited by emotional images and emotionalvisual stimuli are usually not employed in ERP studies onattention Additionally the experimental tasks differgreatly in the majority of cases from the one employedhere Task is a critical factor and its design (physicalcomplexity of stimulation interstimulus interval sensorycategory of the stimulation stimulus parameter to beattended type of responsemdashif any manual verbal etc)significantly influences the characteristics (amplitude orlatency) of recorded components some componentssuch as CNV or P3b may not even appear at all Forexample current P200post presents a similar latency tothat of P2 (sometimes labeled lsquoselection positivityrsquo)often obtained in ERP research on selective attentionto nonemotional parameters of visual stimulation How-ever it is typically maximal at frontal scalp sites (seeeg the review by Mangun amp Hillyard 1995) so thatcurrent P200post and lsquotraditionalrsquo P2 cannot be identi-fied as the same component On the other handP340post presents similar latency to that of P3a (SquiresSquires amp Hillyard 1975) but the latter presents itsmaximum at fronto-central sites Moreover as indicatedP2 and P3a usually appear in tasks that are markedlydifferent from that used here Consequently currentP200post and P340post which could be defined ascomponents related to visual attention to previouslyannounced emotional stimuli with a maximum in pari-eto-occipital areas and originating in the VAC will bediscussed without reference to previous ERP data
Carretie et al 1123
The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

The first of the two posttarget VAC-related compo-nents has mainly reacted around 200 msec to negativestimulation This result suggests that at this stagenegative stimuli elicit to a greater extent than the otherstimuli the mobilization of visual processing resourcesThis lsquoearlyrsquo attentional response is followed by a lsquolatersquoattentional response some 140 msec later characterizedby the opposite trend (from the point of view ofstimulus valence) since it reflects the highest mobiliza-tion of perceptive resources towards appetitive (iepositive and arousing) stimuli Another important time-related difference exists between P200post andP340post The period is shorter in the former compo-nent (Figure 3) It can be clearly appreciated in parietaland occipital recordings that ERPs to A+ maintain arelatively constant level of amplitude for more than 200msec peaking in P340 (Figure 2) On the other handA elicits a high response in P200 but it decreasesrapidly in about 50 msec Since period of componentmay be understood to reflect the duration of the sub-jacent neural processes the present results suggest thatthe early attention response reflects a briefer andprobably less acute mobilization of visual processingresources towards the stimulation Taken together la-tency and period data support the idea mentioned inthe Introduction that the neural response to negativestimulation is more rapid and shorter than the responseto other types of stimulation and that this lsquonegativitybiasrsquo is already reflected in attentional phases
Along with this temporal distinction between maximalresponse to A+ and maximal response to A aninteresting result of the present experiment is that aspatial distinction has also been observed Thus themain focus for P200post was located in posterior areas ofthe middle temporal gyrus (BA 39 and 37) Meanwhilethe source for P340post was located in the inferiortemporal gyrus or in the fusiform gyrus (BA 20 and37) Even assuming the highest margin of error in EEG-based source localization it can be established thatP200post and P340post originated in different sourcesand that the early attentional component appears topresent one focus dorsally located with respect to thefocus of the late attentional component The proposedfocus for the latter component is located in the lsquoventralrsquovisual stream which projects from V1 to the temporalpole and is preferentially involved in the discriminationof objects analyzing attributes such as color or shape(Zeki amp Shipp 1988 Livingstone amp Hubel 1987) Theproposed focus for P200post the early attentional com-ponent is a relatively wide area known as the posteriormiddle temporal gyrus which belongs to the lsquodorsalstreamrsquo (see eg Tootell Dale Sereno amp Malach1996) a projection of fibers from the striate visual cortexto the parietal lobe Thus a spatial distinction appears tooccur between lsquoearly attentionrsquo a rapid subphase andlsquolate attentionrsquo a slower and longer subphase presum-ably oriented to detailed processing As a consequence
and indirectly this differentiation is also present be-tween maximum response to negative and maximumresponse to positive stimulation
The main role of the dorsal stream has been identifiedas the processing of motion and spatial location How-ever these are far from being its sole functions Forexample the processing of visual attributes lsquotypicallyrsquobelonging to the ventral pathway such as shape hasbeen found to be carried out in the dorsal stream ofnonhuman primates (Sereno amp Maunsell 1998) Inhumans data also exist that show high activation ofdorsal stream areas (including posterior middle tempo-ral gyrus) in response to static visual stimuli presentedwith invariant spatial locations (Mummery PattersonHodges amp Price 1998 Martin Haxby Lalonde Wiggsamp Ungerleider 1995 Martin Wiggs Ungerleider ampHaxby 1996) In relation to emotional stimulationactivation of dorsal stream areas such as the middletemporal gyrus (Kosslyn et al 1996) and inferior parietallobe (Lang et al 1998) has been observed to be greater(as the present data also suggest) in response to neg-ative than to positive and neutral stimulation A possibleexplanation is that the dorsal stream appears to beactivated in response to visual stimuli with motor ac-tion-associated meaning regardless of whether they arephysically in motion or not For example static wordsassociated with action elicit the activation of this stream(Mummery et al 1998 Martin et al 1995 1996)
Conclusion
A description of the complete lsquopicturersquo of the processesobserved in present research could be as follows Ex-pectancy-related attention associated with N280preappears to activate the ACC andor medial prefrontalareas responsible for organizing the processing of forth-coming events and the reaction to them (Posner ampRaichle 1995) Cues announcing nonharmful eventsgenerate greater activation of these cortices particularlyas compared with negative events That is to say expect-ancy-related attention towards nonharmful stimuli isgreater than towards negative stimuli In a differentinput processing-related process (once the stimulusappears) attention is responsible for organizing sensoryresources and mobilizing them at the time and placethat they are necessary in order to adequately processthe emotional visual events and it also decides theextent to which this mobilization must take place tobe effective The VAC is in this type of attention the keybrain area The onset of a negative event announced by acue may enhance the early attentional response re-flected in P200post originated in the dorsal streamThe objective of this early attentional response wouldbe not a deep processing but a rapid motoric andautonomic response Appetitive events enhance the lateattentional response reflected in P340post longer andmore thorough carried out in the ventral pathway and
1124 Journal of Cognitive Neuroscience Volume 13 Number 8
oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

oriented to deeper processing of stimuli that typicallytrigger approaching behavior
Main conclusions derived from present results arefirstly that expectancy-related and input processing-related phases of the task appear to activate differentbrain areas the former would associate with the activa-tion of the ACC andor medial prefrontal areas whereasthe later would activate the VAC regions Secondly clearsigns of a negativity bias have been observed in brainresponses reflecting input processing-related but notvigilance-related attention Finally results suggest thatcertain aspects within the neural substrates involved inthe processing of positive and negative stimulation arespatially segregated The actual range of these conclu-sions must be established taking into account twoconstraints inherent to the methodology employed inthis study On one hand it should not be discarded thatprocesses other than affective and attentional couldcontribute to configure the effects observed here Thusthe use of emotional pictures could bear the risk thatsome stimulus properties other than those controlled inthis study may influence the ERP shape On the otherhand though the conclusions dealing with the spatialorigin of the observed activity have been reached bymeans of some of the most sophisticated tools availableto obtain spatial information from the ERPs (LORETAand sPCAs) the fact that they constitute indirect ap-proaches to source localization must be rememberedhere Future research using more direct methodologiesshould contrast these findings
METHOD
Subjects
Thirty right-handed students from the UniversidadAutonoma de Madrid took part in this experimentThe data from only 29 of them were eventually analyzedas explained later These 29 subjects 23 women and 6men were aged between 18 and 30 years (mean = 224)They took part voluntarily in the experiment
Stimuli and Procedure
Figure 11 shows schematically the structure of theexperimental paradigm employed in the present experi-ment Due to the symbolic scarcely realistic character-istics of the drawings used as cues subjects had theopportunity to see them to identify their meaning and toassociate them with targets before the recording sessionguided by the experimenter Two drawings or symbolswere designed for each category The two drawingscorresponding to A+ represented a lsquolsquonude personrsquorsquo anda lsquolsquocakersquorsquo those corresponding to A represented anlsquolsquoinsectrsquorsquo and a wolf lsquolsquojawrsquorsquo those belonging to the Ncategory represented a lsquolsquobuildingrsquorsquo and a lsquolsquoglassrsquorsquo andfinally lsquolsquoforestrsquorsquo and lsquolsquosubmarinersquorsquo landscapes were rep-resented by the two drawings of the R group Cuepresentations consisted of the two drawings belongingto each category one above the other (see Figure 12)This presentation lasted 250 msec The dimensions ofeach cue presentation were 169 339 cm
A color photograph (target) was presented 1750 mseclater (285 423 cm) Color photographs were anopposite-sex nude or an ice cream cake if an A+ cuewas presented a tropical ortopteroid insect or an openmouth of a wolf in an aggressive expression if the cuebelonged to the A category a glass or a flat buildingfor N-type cues and wood or underwater images for Rcues In other words (as subjects were told in theinstructions) photographs represented always one ofthe objects or motifs represented by the two drawings
Figure 11 Schematic representation of the experimental paradigmemployed in the present experiment ITI = intertrial interval
Figure 12 Stimuli used as cueFrom left to right first cueannounced an arousing-positivetarget second announced anarousing-negative target thirdannounced a neutral target andfourth announced a relaxingtarget
Carretie et al 1125
of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

of the cue presentation Therefore the cue implicitlyinformed about the emotional content of the targetTwo seconds after the end of the target presentation abeep was presented to the subjects If the photographcorresponded to the drawing that appeared at the topof the cue presentation they were instructed to say lsquolsquoArsquorsquoand if it corresponded to the bottom drawing of thecue they had to say lsquolsquoBrsquorsquo Participants were instructed tolook continuously at a small mark located in the centerof the screen and to respond and to blink only after thebeep Intertrial interval lasted 2500 msec Thirty-twotrials were presented for each of the four groups ofstimuli A+ A N and R (ie a total of 128 trials waspresented)
Subjects completed a bidimensional scaling test foreach group of pictures after the recording sessions Thistest assessed the valence and the arousal content of thepictures two affective dimensions considered to explainthe principal variance of the emotional meaning (LangGreenwald Bradley amp Hamm 1993 Smith amp Ellsworth1985 Russell 1979 Osgood Succi amp Tannenbaum1957) Since analyses of ERPs were carried out groupingthe recordings elicited by the stimuli of each categorythe assessments made for each image separately werealso averaged for each category before being entered instatistical analyses
Recording and Data Analysis
Electroencephalographic data were recorded using anelectrode cap (ElectroCap International) with tin elec-trodes A total of 58 scalp locations were used Fp1 FpzFp2 AF3 AF4 F7 F5 F3 F1 Fz F2 F4 F6 F8 FC5 FC3FC1 FCz FC2 FC4 FC6 T7 C5 C3 C1 Cz C2 C4 C6T8 TP7 CP5 CP3 CP1 CPz CP2 CP4 CP6 TP8 P7 P5P3 P1 Pz P2 P4 P6 P8 PO7 PO3 PO1 POz PO2 PO4PO8 O1 Oz and O2 All scalp electrodes as well as oneelectrode at the left mastoid (M1) were originallyreferenced to one electrode at the right mastoid (M2)Electrooculographic (EOG) data were recorded supra-and infra-orbitally (vertical EOG) and from the leftversus right orbital rim (horizontal EOG) Electrodeimpedances were always kept below 3 k A bandpassof 01 to 50 Hz (3 dB points for 6 dBoctave roll-off)was used for the recording amplifiers The channelswere continuously digitizing data at a sampling rate of250 Hz for the duration of complete recording session
The continuous recording was divided into twoepochs for each trial The first epoch which involvedpretarget ERPs lasted 1950 msec beginning 200 msecbefore the cue onset The second epoch comprisedposttarget ERPs and lasted 1000 msec beginning 200msec before the target onset Trials where subjectsanswered incorrectly were eliminated A visual inspec-tion was also carried out eliminating epochs with eyemovements or blinks Results from one of the initial 30subjects had to be rejected due to the presence of an
excessive rate of ocular artifacts in the recordings ERPaverages were categorized according to each type ofstimulus For the entire sample of cephalic electrodesoriginally M2-referenced data were algebraically re-refer-enced off-line using a global average reference (Leh-mann 1987) ANOVAs and tPCAs required the selectionof 28 from the original 58 channels in order to avoid theloss of statistical power These 28 channels were AF3AF4 F5 F1 F2 F6 FC5 FC1 FC2 FC6 C5 C1 C2 C6CP5 CP1 CP2 CP6 P5 P1 P2 P6 PO7 PO1 PO2 PO8O1 and O2 They were selected on the basis that theyare topographically alternating across the scalp in a waythat every unselected scalp location is surrounded by atleast one or two immediately adjacent selected locations(interelectrode distances were short enough to admitthat the differences between adjacent locations wouldbe minimal)
Acknowledgments
The authors thank Francisco Munoz and Pilar Casado for theirhelp in data collection and R D Pascual-Marqui for his help inthe use of LORETA and for the adaptation to the 58 channelsused in the present study JAH is supported by a grant fromDireccion General de Investigacion Comunidad Autonoma deMadrid
Reprint requests should be sent to Luis Carretie Departamen-to de Psicolog a Biologica y de la Salud Facultad de Psicolog aUniversidad Autonoma de Madrid 28049 Madrid Spain or viae-mail carretieuames
REFERENCES
Basile L F H Rogers R L Bourbon W T amp PapanicolaouA (1994) Slow magnetic flux from human frontal cortexElectroencephalography and Clinical Neurophysiology 90157- 165
Bechara A Damasio H amp Damasio A R (2000) Emotiondecision making and the orbitofrontal cortex CerebralCortex 10 295- 307
Bush G Luu P amp Posner M I (2000) Cognitive andemotional influences in anterior cingulate cortex Trends inCognitive Sciences 4 215- 222
Bussey T J Muir J L Everitt B J amp Robbins T W (1997)Triple dissociation of anterior cingulate posterior cingulateand medial frontal cortices on visual discrimination tasksusing a touchscreen testing procedure for the ratBehavioral Neuroscience 111 920- 936
Cacioppo J T amp Bernston G G (1994) Relationshipbetween attitudes and evaluative space A critical reviewwith emphasis on the separability of positive and negativesubstrates Psychological Bulletin 115 401- 423
Cacioppo J T amp Gardner W L (1999) Emotion AnnualReview of Psychology 50 191- 214
Canli T Desmond J E Zhao Z Glover G amp Gabrieli J DE (1998) Hemispheric asymmetry for emotional stimulidetected with fMRI NeuroReport 9 3233- 3239
Carretie L Mercado F Tapia M amp Hinojosa J A (2001)Emotion attention and the lsquonegativity biasrsquo studiedthrough event-related potentials International Journal ofPsychophysiology 41 75- 85
Cliff N (1987) Analyzing multivariate data New YorkHarcourt Brace Jovanovich
1126 Journal of Cognitive Neuroscience Volume 13 Number 8
Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

Dahaene S Posner M I amp Tucker D M (1994) Localizationof a neural system for error detection and compensationPsychological Science 5 303- 305
Damen E J P amp Brunia C M H (1994) Is a stimulus con-veying task-relevant information a sufficient condition toelicit a stimulus-preceding negativity Psychophysiology 31129- 139
Davidson R J amp Irwin W (1999) The functional neuro-anatomy of emotion and affective style Trends in CognitiveSciences 3 11- 21
Duncan-Johnson C C amp Donchin E (1977) On quantifyingsurprise The variation of event-related potentials withsubjective probability Psychophysiology 14 456- 457
Ekman P (1992) An argument for basic emotions Cognitionand Emotion 6 169- 200
Fiske S T (1980) Attentional and weight in personperception The impact of negative and extreme behaviorJournal of Personality and Social Psychology 38 889- 906
Gehring W J Goss B Coles M G H Meyer D E ampDonchin E (1993) A neural system for error detection andcompensation Psychological Science 4 385- 390
Halgren E amp Marinkovic K (1994) Neurophysiologicalnetworks integrating human emotions In M S Gazzaniga(Ed) The cognitive neurosciences (pp 1137- 1151)London MIT Press
Ito T A Larsen J T Smith N K amp Cacioppo J T (1998)Negative information weighs more heavily on the brain Thenegativity bias in evaluative categorizations Journal ofPersonality and Social Psychology 75 887- 900
Klorman R amp Ryan R M (1980) Heart rate contingentnegative variation and evoked potentials during anticipationof affective stimulation Psychophysiology 17 513- 523
Kosslyn S M Shin L M Thompson W L McNally R JRauch S L Pitman R K amp Alpert N M (1996) Neuraleffects of visualizing and perceiving aversive stimuli A PETinvestigation NeuroReport 7 1569- 1576
Lane R D (1999) Functional neuroanatomy of pleasant andunpleasant emotion Dissertation Abstracts InternationalB The Sciences and Engineering 60 1907
Lane R D Chua P M L amp Dolan R J (1999) Commoneffects of emotional valence arousal and attention on neuralactivation during visual processing of pictures Neuropsy-chologia 37 989- 997
Lane R D Fink G R Chau P M L amp Dolan R (1997)Neural activation during selective attention to subjectiveemotional responses NeuroReport 8 3969- 3872
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J (1997) Neuroanatomical correlates of hap-piness sadness and disgust American Journal of Psychia-try 7 926- 933
Lang P J Bradley M M amp Cuthbert B N (1997) Motivatedattention Affect activation and action In P Lang R FSimons amp M T Balaban (Eds) Attention and orientingSensory and motivational processes (pp 97- 135) MahwahNJ Erlbaum
Lang P J Bradley M M Fitzsimmons J R Cuthbert B NScott J D et al (1998) Emotional arousal and activation ofthe visual cortex An fMRI analysis Psychophysiology 35199- 210
Lang P J Greenwald M K Bradley M M amp Hamm A O(1993) Looking at pictures Affective facial visceral andbehavioral reactions Psychophysiology 30 261- 273
LeDoux J E (1995) Emotion Clues from the brain AnnualReview of Psychology 46 209- 235
Lehmann D (1987) Principles of spatial analysis In A S Gevinsamp A Remond (Eds) Handbook of electroencephalographyand clinical neurophysiology (vol 1 pp 309- 354)Amsterdam Elsevier
Leynes P A Allen J D amp Marsh R L (1998) Topographicdifferences in CNV amplitude reflect different preparatoryprocesses International Journal of Psychophysiology 3133- 44
Livingstone M S amp Hubel D H (1987) Psychophysicalevidence for separate channels for the perception of formcolor movement and depth Journal of Neuroscience 73416- 3468
Luck S J Chelazzi L Hillyard S A amp Desimone R (1997)Neural mechanisms of spatial selective attention in areas V1V2 and V4 of macaque visual cortex Journal of Neuro-physiology 77 24- 42
Luu P Collins P amp Tucker D M (2000) Mood personalityand self-monitoring Negative affect and emotionality inrelation to frontal lobe mechanisms of error monitoringJournal of Experimental Psychology General 129 43- 60
Mangun G R amp Hillyard S A (1995) Mechanisms andmodels of selective attention In M D Rugg amp M G H Coles(Eds) Electrophysiology of mind (pp 40- 85) OxfordOxford University Press
Martin A Haxby J V Lalonde F M Wiggs C L ampUngerleider L G (1995) Discrete cortical regionsassociated with knowledge of color and knowledge ofaction Science 270 102- 105
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specific knowledgeNature 379 649- 652
McCallum W C (1988) Potentials related to expectancypreparation and motor activity In T W Picton (Ed)Handbook of electroencephalography and clinicalneurophysiology (vol 3 pp 427- 534) Amsterdam Elsevier
Mummery C J Patterson K Hodges J R amp Price C J(1998) Functional neuroanatomy of the semantic systemDivisible by what Journal of Cognitive Neuroscience 10766- 777
Murtha S Chertkow H Beauregard M Dixon R amp EvansA (1996) Anticipation causes increased blood flow to theanterior cingulate cortex Human Brain Mapping 4 103-112
Northoff G Richter A Gessner M Schlagenhauf F Fell Jet al (2000) Functional dissociation between medial andlateral prefrontal cortical spatiotemporal activation innegative and positive emotions A combined fMRIMEGstudy Cerebral Cortex 10 93- 107
Ohman A (1992) Orienting and attention Preferredpreattentive processing of potentially phobic stimuli InB A Campbell H Haynes amp R Richardson (Eds)Attention and information processing in infants andadults Perspectives from human and animal researchHillsdale NJ Erlbaum
Ohman A Esteves F Flykt A amp Soares J J F (1993)Gateways to consciousness Emotion attention andelectrodermal activity In J C Roy W Boucsein D C Fowlesamp J H Gruzelier (Eds) Progress in electrodermal research(pp 137- 157) New York Plenum
Osgood C Suci G amp Tannenbaum P (1957) Themeasurement of meaning Urbana University of Illinois
Pascual-Marqui R D (1999) Review of methods for solvingthe EEG inverse problem International Journal ofBioelectromagnetism 1 75- 86
Pascual-Marqui R D Michel C M amp Lehman D (1994) Lowresolution electromagnetic tomography A new method forlocalizing electrical activity in the brain InternationalJournal of Psychophysiology 18 49- 65
Ploghaus A Tracey I Gati J S Clare S Menon R S et al(1999) Dissociating pain from its anticipation in the humanbrain Science 284 1979- 1981
Posner M I amp Petersen S E (1990) The attention system of
Carretie et al 1127
the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8

the human brain Annual Review of Neuroscience 1325- 42
Posner M I amp Raichle M E (1995) Images of mindBehavioral and Brain Sciences 18 327- 383
Pratto F amp John O P (1991) Automatic vigilance Theattention-grabbing power of negative social informationJournal of Personality and Social Psychology 61380- 391
Reiman E M Lane R D Ahern G L Schwartz E GDavidson R J Friston K J Yun L S amp Chen K (1997)Neuroanatomical correlates of externally and internallygenerated human emotion American Journal of Psychia-try 154 918- 925
Rohrbaugh J amp Gaillard A (1983) Sensory and motoraspects of the contingent negative variation In A Gaillardamp W Ritter (Eds) Advances in psychology (vol 10 pp269- 310) Amsterdam Elsevier
Russell J A (1979) Affective space is bipolar Journal ofPersonality and Social Psychology 37 345- 356
Sereno A B amp Maunsell J H R (1998) Shape selectivity inprimate lateral intraparietal cortex Nature 395 500- 503
Shizgal P (1999) On the neural computation of utilityImplications from studies of brain stimulation reward InD Kahneman E Diener amp N Schwarz (Eds) Well-beingThe foundations of hedonic psychology (pp 500- 524)New York Russell Sage Foundation
Shulman G L Corbetta M Buckner R L Raichle M E Fiez JA Miezin F M amp Petersen S E (1997) Top-down modula-tion of early sensory cortex Cerebral Cortex 7 193- 206
Smith C A amp Ellsworth P C (1985) Patterns of cognitiveappraisal in emotion Journal of Personality and SocialPsychology 48 813- 838
Squires N K Squires K C amp Hillyard S A (1975) Twovarieties of long-latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387- 4101
Sturm W de Simone A Krause B J Specht KHesselmann V Radesmacher I Herzog H Tellmann L
Mueller-Gaertner H W amp Willmes K (1999) Functionalanatomy of intrinsic alertness Evidence for a fronto-parietal-thalamic- brainstem network in the right hemisphere Neu-ropsychologia 37 797- 805
Talairach J amp Tournoux P (1988) Co-planar stereotaxicatlas of the human brain Stuttgart Germany Thieme
Taylor S E (1991) Asymmetrical effects of positive andnegative events The mobilization- minimization hypothesisPsychological Bulletin 110 67- 85
Teasdale J D Howard R J Cox S G Ha Y Brammer M JWilliams S C R amp Checkley S A (1999) Functional MRIstudy on the cognitive generation of affect American Jour-nal of Psychiatry 156 209- 215
Tellegen A Watson D amp Clark L A (1999) On thedimensional and hierarchical structure of affect Psycho-logical Science 10 297- 309
Tootell R B H Dale A M Sereno M I amp Malach R (1996)New images from human visual cortex Trends inNeuroscience 19 481- 489
Towle V L Bolanos J Suarez D Tan K Grzeszczuk Ret al (1993) The spatial location of EEG electrodes Lo-cating the best-fitting sphere relative to cortical anatomyElectroencephalography and Clinical Neurophysiology86 1- 6
Walter W G Cooper R Aldridge V J McCallum W C ampWinter A L (1964) Contingent negative variation Anelectric sign of sensorimotor association and expectancy inthe human brain Nature 203 380- 384
Whalen P J Bush G McNally R J Wilhelm S McInerneyS C Jenike M A amp Rauch S L (1998) The emotionalcounting Stroop paradigm A functional magnetic resonanceimaging probe of the anterior cingulate affective divisionBiological Psychiatry 44 1219- 1228
Yee C M amp Miller G A (1987) Affective valence andinformation processing Electroencephalography andClinical Neurophysiology (Suppl 40) 300- 307
Zecki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311- 317
1128 Journal of Cognitive Neuroscience Volume 13 Number 8




















