
Taxonomic revision of Astragalus angustifolius group Fabaceae
Upload
independentCategory
view
3download
0
Captorhinus magnus, a new captorhinid (Amniota:Eureptilia) from the Lower Permian of Oklahoma,with new evidence on the homology of theastragalus
Richard A. Kissel, David W. Dilkes, and Robert R. Reisz
Abstract: The new species of single tooth-rowed captorhinid reptile Captorhinus magnus n. sp. from the Lower Permianfissure fillings at Richards Spur, Oklahoma differs from Captorhinus aguti in body size and dental and femoral morphology.Linear measurements of fully mature C. magnus elements range in size from 1.5 to 2.3 times as great as those of C. aguti,and the proximal articular surface of the femur, which is convex in C. aguti, is concave throughout ontogeny. C. magnuspossesses ogival cheek teeth aligned in a single row, indicating that ogival dentition can no longer be considered uniqueto C. aguti. A phylogenetic analysis of captorhinid interrelationships indicates that C. aguti and C. magnus form aclade that possesses a sister-group relationship with Captorhinus laticeps. Incompletely ossified astragali referred toC. magnus provide unequivocal evidence that the astragalus of Captorhinus formed through the fusion of three, originallyseparate ossifications, the tibiale, intermedium, and proximal centrale, rather than from a single ossification center. Atthe Richards Spur locality, C. magnus is the most abundant Captorhinus species produced from the deeper,stratigraphically lower sediments of the quarry. It is rare, however, in the uppermost, presumably younger deposits,where C. aguti represents the most numerous Richards Spur captorhinid.
Résumé : Une nouvelle espèce de reptile Captorhinidé à rangée de dents simple, Captorhinus magnus n. sp., provenantde remplissage de fissures au Permien inférieur à Richards Spur, en Oklahoma, diffère de Captorhinus aguti par ladimension de son corps et la morphologie du fémur. Les mesures linéaires d’un C. magnus à pleine maturité vont de1,5 à 2,3 fois celles du C. aguti et la surface articulaire proximale du fémur, laquelle est convexe chez C. aguti, estconcave à travers l’entière ontogenèse. C. magnus possède des dents jugales ogivales alignées en rangée simple, indiquantque la dentition ogivale ne peut plus être considérée unique à C. aguti. Une analyse phylogénétique des interrelationsde Captorhinidés indique que C. aguti et C. magnus forment un clade qui possède une relation de parenté de groupeavec Captorhinus laticeps. Des astragales non complètement ossifiés attribués à C. magnus fournissent des évidencesnon équivoques que l’astragale de Captorhinus s’est formé par la fusion de trois ossifications originalement séparées, letibial, l’intermédiaire et le naviculaire proximal, plutôt qu’à partir d’un centre unique d’ossification. À la localité de RichardSpurs, C. magnus est l’espèce de Captorhinus la plus abondante provenant de sédiments plus profonds, stratigraphiquementinférieurs, de la carrière. Cette espèce est toutefois rare dans les dépôts supérieurs, probablement plus jeunes, où C. agutireprésente le Captorhinidé le plus abondant de Richards Spur.
[Traduit par la Rédaction] Kissel et al. 1372
Introduction
The Dolese Brothers limestone quarry at Richards Spur,Oklahoma has yielded the remains of a diverse assemblageof fossil vertebrates. The site, which has been well documentedby Olson (1967, 1991), May and Cifelli (1998), Sullivan andReisz (1999a), and Sullivan et al. (2000), has produced thegreatest number of specimens of Permian vertebrate fossilsin Oklahoma. Preserved within Early Permian fissure filldeposits in the Ordovician Arbuckle Limestone, the fossil
material consists primarily of terrestrial amniotes andanamniotes, and increasingly larger faunal lists have beenproduced by Bolt (1980), Olson (1991), and Sullivan andReisz (1999a). Three captorhinid taxa have been describedfrom the locality thus far. Remains of Captorhinus aguti arevery common and constitute the majority of the fossil materialidentified from the site (May and Cifelli 1998; Modesto1999; Sullivan and Reisz 1999b). As a result of extensivetreatments by Fox and Bowman (1966) and Modesto (1998),the cranial and postcranial morphology of C. aguti is known
Can. J. Earth Sci. 39: 1363–1372 (2002) DOI: 10.1139/E02-040 © 2002 NRC Canada
1363
Received 12 March 2002. Accepted 12 June 2002. Published on the NRC Research Press Web site at http://cjes.nrc.ca on12 September 2002.
Paper handled by Associate Editor H.-D. Sues.
R.A. Kissel,1 D.W. Dilkes, and R.R. Reisz. Department of Biology, University of Toronto, 3359 Mississauga Road, Mississauga,ON L5L 1C6, Canada.
1Corresponding author (e-mail: [email protected]).
1

in great detail. In sharp contrast to C. aguti, the other RichardsSpur captorhinid taxa are based on far fewer specimens.May and Cifelli (1998) described Baeotherates fortsillensison the basis of a partial right mandible, and Modesto (1996)attributed a nearly complete right maxilla and a partial rightdentary to an unnamed basal, or plesiomorphic, captorhinid.Heaton (1979) assigned all single tooth-rowed captorhinidelements recovered from Richards Spur to Eocaptorhinus(= Captorhinus) laticeps. However, this material has sincebeen referred to C. aguti (Bolt 1980; Modesto 1998), therebyeliminating C. laticeps from the Richards Spur captorhinidassemblage.
Recent examination of fossils produced from the fissurefills has yielded the remains of a fourth Richards Spurcaptorhinid taxon, a new species of Captorhinus firstreported by Kissel et al. (2001). The purpose of this study isto describe this new species and compare it to the threecaptorhinid taxa previously identified from Richards Spur. Inaddition, numerous astragali are present among the specimensof this new captorhinid, providing the basis for a reevaluationof the homology of the astragalus.
Institutional abbreviationsFMNH, Field Museum of Natural History, Chicago, Illinois;
MCZ, Museum of Comparative Zoology, Harvard University,Cambridge, Massachusetts; OMNH, Sam Noble OklahomaMuseum of Natural History, Norman, Oklahoma; UCLA VP,University of California Vertebrate Paleontology, Los Angeles,California.
Systematic paleontology
Eureptilia Olson, 1947Captorhinomorpha Watson, 1917Captorhinidae Case, 1911Captorhinus Cope, 1896
Captorhinus magnus, new species
HolotypeOMNH 55387, complete right femur.
Referred specimensOMNH 55386, complete right maxilla; OMNH 55390,
complete left fibula; OMNH 55391, complete left mandible;OMNH 55392, anterior portion of right mandible; OMNH55396, left astragalus; OMNH 55397, right astragalus;OMNH 55399, left astragalus lacking proximal centrale;OMNH 55400, right intermedium; OMNH 56820, dorsalportion of cranium lacking jugals, maxillae, left postfrontal,postorbitals, quadrates, quadratojugals, squamosals,supratemporals, braincase, and palate; OMNH 56821, partialskull lacking nasals, premaxillae, and portions of the dentaries,lacrimals, maxillae, and prefrontals; OMNH 56881, completeleft humerus; OMNH 56884, left femur lacking extremeproximal end; OMNH 56885, complete left femur; OMNH56886, complete right femur; OMNH 56888, right femurlacking distal end of tibial condyle; OMNH 56891, left sideof pelvis; OMNH 56893, left side of pelvis; OMNH 56894,left side of pelvis; OMNH 56895, right side of pelvis;OMNH 56896, proximal end of left ulna; OMNH 56897,
proximal end of right ulna; OMNH 56900, proximal end ofleft ulna; OMNH 56909, glenoid portion of left scapulocoracoid;OMNH 56910, glenoid portion of left scapulocorocoid;OMNH 56911, glenoid portion of left scapulocoracoid;OMNH 56912, glenoid portion of right scapulocoracoid;OMNH 56913, glenoid portion of right scapulocoracoid;OMNH 56917, right mandible lacking proximal and distalends; OMNH 56921, left mandible lacking distal end;OMNH 56923, left dentary; OMNH 56929, proximal end ofleft femur; OMNH 56934, proximal end of left femur;OMNH 56943, proximal end of right femur; OMNH 56961,distal end of right humerus; OMNH 56963, distal end ofright humerus; OMNH 56965, distal end of left humerus;OMNH 56967, distal end of left humerus; OMNH 56970,distal end of left humerus; OMNH 56973, proximal end ofright humerus; OMNH 56975, proximal end of right humerus;OMNH 56983, proximal end of left humerus; OMNH56984, proximal end of left humerus.
EtymologyMagnus, Latin, large; referring to the large size of this
species relative to Captorhinus aguti, the type species of thegenus Captorhinus.
Locality and horizonOMNH V51 (Richards Spur Locality), the Dolese
Brothers limestone quarry (SW1/4 Section 31, Township 4N,Range 11W), located �10 km north of Fort Sill, ComancheCounty, Oklahoma. Fissure fill deposit stratigraphicallyequivalent to the lowermost Clear Fork Group (Leonardian:Lower Permian) of Texas; correlation based on vertebratefauna and is tentative.
DiagnosisA single tooth-rowed captorhinid distinguishable from
Captorhinus aguti on the basis of three features: (1) asignificantly larger adult size, (2) the presence of ogivalmarginal dentition aligned in a single row, and (3) concaveproximal articular surface of the femur. Linear measurementsof available well-ossified Captorhinus magnus elements(two nearly complete skulls; several mandibles; and isolatedpostcranial elements, including scapulocoracoids, pelves, humeri,femora, ulnae, a fibula, and astragali) from fully matureindividuals are 1.5 to 2.3 times larger than those in well-ossified,fully mature individuals of C. aguti.
Description
Since the skeletal elements of Captorhinus magnus possessa nearly identical morphology to those of C. aguti figured byFox and Bowman (1966), a complete description of C. magnusis not required. Measurements of available C. magnus elementsare listed in Tables 1 and 2, and a comparison of the averagemeasurements of selected C. magnus and C. aguti elements isprovided in Table 3. To reduce features associated withontogenetic variation, all descriptions are of well-ossified,presumably fully mature individuals. The orientation of limbelements follows Romer (1956), and the descriptions “singletooth-rowed” and “multiple tooth-rowed” are hereafter ab-breviated “single-rowed” and “multiple-rowed,” respectively.
Description of the skull of Captorhinus magnus is based upon
© 2002 NRC Canada
1364 Can. J. Earth Sci. Vol. 39, 2002
2
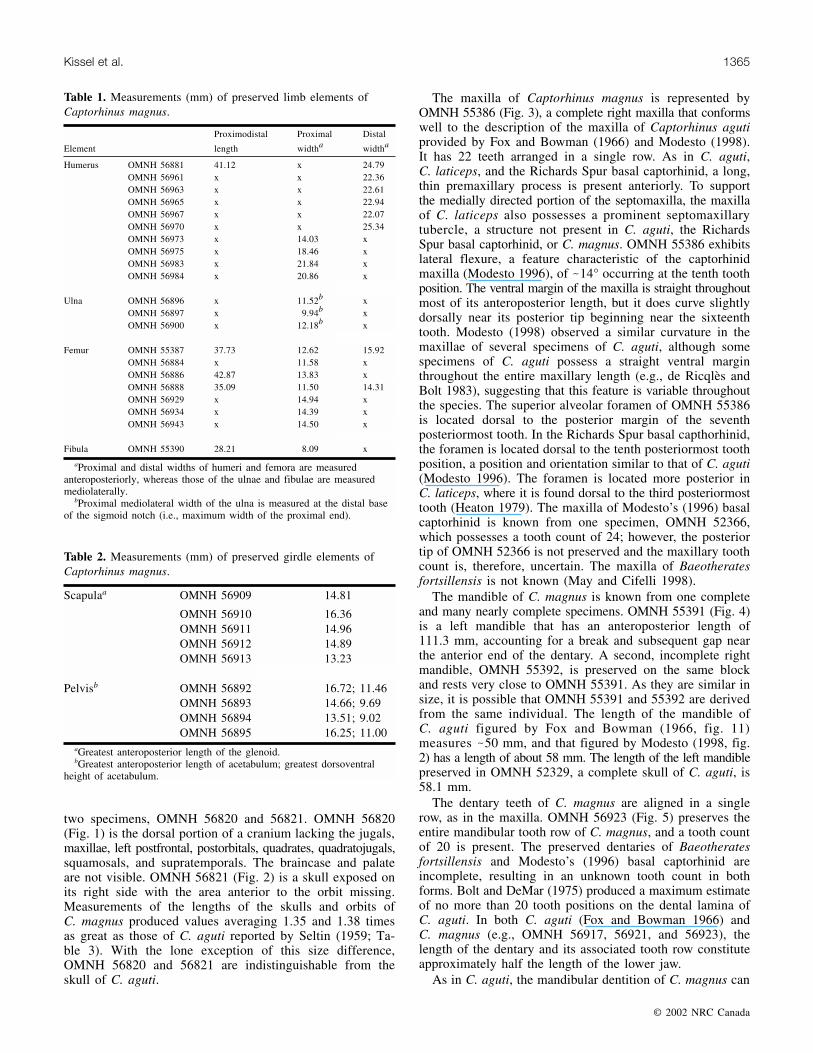
two specimens, OMNH 56820 and 56821. OMNH 56820(Fig. 1) is the dorsal portion of a cranium lacking the jugals,maxillae, left postfrontal, postorbitals, quadrates, quadratojugals,squamosals, and supratemporals. The braincase and palateare not visible. OMNH 56821 (Fig. 2) is a skull exposed onits right side with the area anterior to the orbit missing.Measurements of the lengths of the skulls and orbits ofC. magnus produced values averaging 1.35 and 1.38 timesas great as those of C. aguti reported by Seltin (1959; Ta-ble 3). With the lone exception of this size difference,OMNH 56820 and 56821 are indistinguishable from theskull of C. aguti.
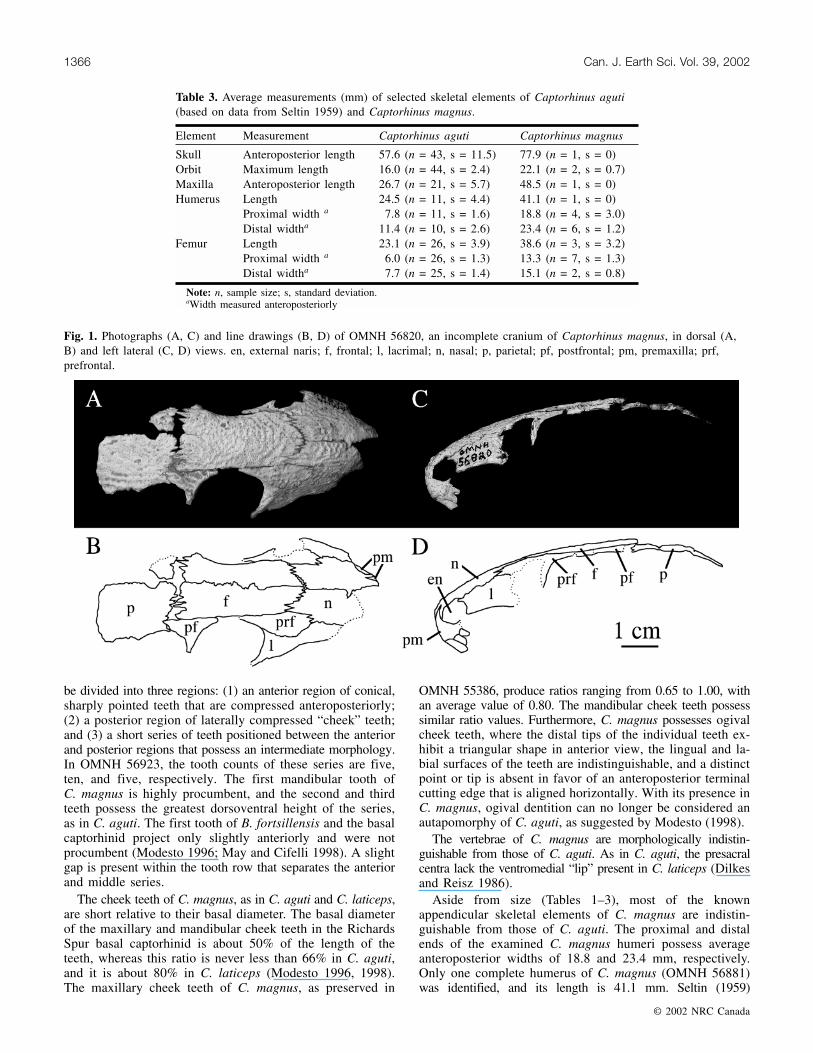
The maxilla of Captorhinus magnus is represented byOMNH 55386 (Fig. 3), a complete right maxilla that conformswell to the description of the maxilla of Captorhinus agutiprovided by Fox and Bowman (1966) and Modesto (1998).It has 22 teeth arranged in a single row. As in C. aguti,C. laticeps, and the Richards Spur basal captorhinid, a long,thin premaxillary process is present anteriorly. To supportthe medially directed portion of the septomaxilla, the maxillaof C. laticeps also possesses a prominent septomaxillarytubercle, a structure not present in C. aguti, the RichardsSpur basal captorhinid, or C. magnus. OMNH 55386 exhibitslateral flexure, a feature characteristic of the captorhinidmaxilla (Modesto 1996), of �14° occurring at the tenth toothposition. The ventral margin of the maxilla is straight throughoutmost of its anteroposterior length, but it does curve slightlydorsally near its posterior tip beginning near the sixteenthtooth. Modesto (1998) observed a similar curvature in themaxillae of several specimens of C. aguti, although somespecimens of C. aguti possess a straight ventral marginthroughout the entire maxillary length (e.g., de Ricqlès andBolt 1983), suggesting that this feature is variable throughoutthe species. The superior alveolar foramen of OMNH 55386is located dorsal to the posterior margin of the seventhposteriormost tooth. In the Richards Spur basal capthorhinid,the foramen is located dorsal to the tenth posteriormost toothposition, a position and orientation similar to that of C. aguti(Modesto 1996). The foramen is located more posterior inC. laticeps, where it is found dorsal to the third posteriormosttooth (Heaton 1979). The maxilla of Modesto’s (1996) basalcaptorhinid is known from one specimen, OMNH 52366,which possesses a tooth count of 24; however, the posteriortip of OMNH 52366 is not preserved and the maxillary toothcount is, therefore, uncertain. The maxilla of Baeotheratesfortsillensis is not known (May and Cifelli 1998).
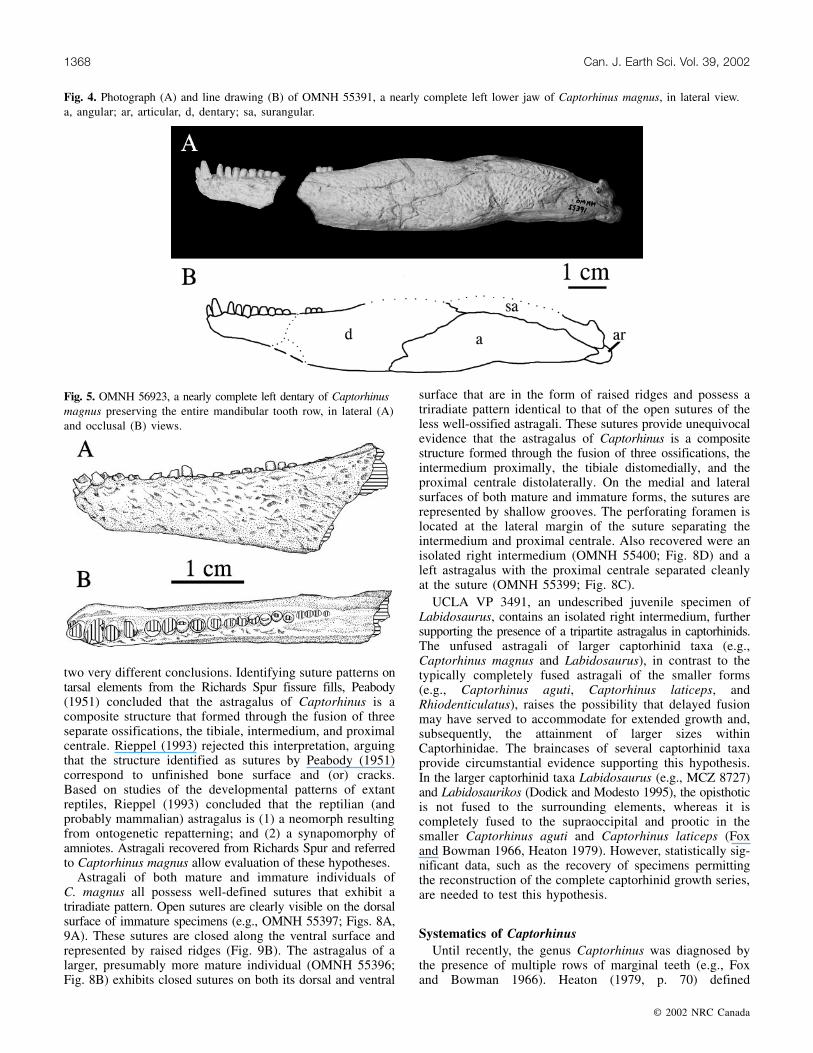
The mandible of C. magnus is known from one completeand many nearly complete specimens. OMNH 55391 (Fig. 4)is a left mandible that has an anteroposterior length of111.3 mm, accounting for a break and subsequent gap nearthe anterior end of the dentary. A second, incomplete rightmandible, OMNH 55392, is preserved on the same blockand rests very close to OMNH 55391. As they are similar insize, it is possible that OMNH 55391 and 55392 are derivedfrom the same individual. The length of the mandible ofC. aguti figured by Fox and Bowman (1966, fig. 11)measures �50 mm, and that figured by Modesto (1998, fig.2) has a length of about 58 mm. The length of the left mandiblepreserved in OMNH 52329, a complete skull of C. aguti, is58.1 mm.
The dentary teeth of C. magnus are aligned in a singlerow, as in the maxilla. OMNH 56923 (Fig. 5) preserves theentire mandibular tooth row of C. magnus, and a tooth countof 20 is present. The preserved dentaries of Baeotheratesfortsillensis and Modesto’s (1996) basal captorhinid areincomplete, resulting in an unknown tooth count in bothforms. Bolt and DeMar (1975) produced a maximum estimateof no more than 20 tooth positions on the dental lamina ofC. aguti. In both C. aguti (Fox and Bowman 1966) andC. magnus (e.g., OMNH 56917, 56921, and 56923), thelength of the dentary and its associated tooth row constituteapproximately half the length of the lower jaw.
As in C. aguti, the mandibular dentition of C. magnus can
© 2002 NRC Canada
Kissel et al. 1365
Element
Proximodistal
length
Proximal
widthaDistal
widtha
Humerus OMNH 56881 41.12 x 24.79OMNH 56961 x x 22.36OMNH 56963 x x 22.61OMNH 56965 x x 22.94OMNH 56967 x x 22.07OMNH 56970 x x 25.34OMNH 56973 x 14.03 xOMNH 56975 x 18.46 xOMNH 56983 x 21.84 xOMNH 56984 x 20.86 x
Ulna OMNH 56896 x 11.52b xOMNH 56897 x 9.94b xOMNH 56900 x 12.18b x
Femur OMNH 55387 37.73 12.62 15.92OMNH 56884 x 11.58 xOMNH 56886 42.87 13.83 xOMNH 56888 35.09 11.50 14.31OMNH 56929 x 14.94 xOMNH 56934 x 14.39 xOMNH 56943 x 14.50 x
Fibula OMNH 55390 28.21 8.09 x
aProximal and distal widths of humeri and femora are measuredanteroposteriorly, whereas those of the ulnae and fibulae are measuredmediolaterally.
bProximal mediolateral width of the ulna is measured at the distal baseof the sigmoid notch (i.e., maximum width of the proximal end).
Table 1. Measurements (mm) of preserved limb elements ofCaptorhinus magnus.
Scapulaa OMNH 56909 14.81
OMNH 56910 16.36OMNH 56911 14.96OMNH 56912 14.89OMNH 56913 13.23
Pelvisb OMNH 56892 16.72; 11.46OMNH 56893 14.66; 9.69OMNH 56894 13.51; 9.02OMNH 56895 16.25; 11.00
aGreatest anteroposterior length of the glenoid.bGreatest anteroposterior length of acetabulum; greatest dorsoventral
height of acetabulum.
Table 2. Measurements (mm) of preserved girdle elements ofCaptorhinus magnus.
3
be divided into three regions: (1) an anterior region of conical,sharply pointed teeth that are compressed anteroposteriorly;(2) a posterior region of laterally compressed “cheek” teeth;and (3) a short series of teeth positioned between the anteriorand posterior regions that possess an intermediate morphology.In OMNH 56923, the tooth counts of these series are five,ten, and five, respectively. The first mandibular tooth ofC. magnus is highly procumbent, and the second and thirdteeth possess the greatest dorsoventral height of the series,as in C. aguti. The first tooth of B. fortsillensis and the basalcaptorhinid project only slightly anteriorly and were notprocumbent (Modesto 1996; May and Cifelli 1998). A slightgap is present within the tooth row that separates the anteriorand middle series.
The cheek teeth of C. magnus, as in C. aguti and C. laticeps,are short relative to their basal diameter. The basal diameterof the maxillary and mandibular cheek teeth in the RichardsSpur basal captorhinid is about 50% of the length of theteeth, whereas this ratio is never less than 66% in C. aguti,and it is about 80% in C. laticeps (Modesto 1996, 1998).The maxillary cheek teeth of C. magnus, as preserved in
OMNH 55386, produce ratios ranging from 0.65 to 1.00, withan average value of 0.80. The mandibular cheek teeth possesssimilar ratio values. Furthermore, C. magnus possesses ogivalcheek teeth, where the distal tips of the individual teeth ex-hibit a triangular shape in anterior view, the lingual and la-bial surfaces of the teeth are indistinguishable, and a distinctpoint or tip is absent in favor of an anteroposterior terminalcutting edge that is aligned horizontally. With its presence inC. magnus, ogival dentition can no longer be considered anautapomorphy of C. aguti, as suggested by Modesto (1998).
The vertebrae of C. magnus are morphologically indistin-guishable from those of C. aguti. As in C. aguti, the presacralcentra lack the ventromedial “lip” present in C. laticeps (Dilkesand Reisz 1986).
Aside from size (Tables 1–3), most of the knownappendicular skeletal elements of C. magnus are indistin-guishable from those of C. aguti. The proximal and distalends of the examined C. magnus humeri possess averageanteroposterior widths of 18.8 and 23.4 mm, respectively.Only one complete humerus of C. magnus (OMNH 56881)was identified, and its length is 41.1 mm. Seltin (1959)
© 2002 NRC Canada
1366 Can. J. Earth Sci. Vol. 39, 2002
Element Measurement Captorhinus aguti Captorhinus magnus
Skull Anteroposterior length 57.6 (n = 43, s = 11.5) 77.9 (n = 1, s = 0)Orbit Maximum length 16.0 (n = 44, s = 2.4) 22.1 (n = 2, s = 0.7)Maxilla Anteroposterior length 26.7 (n = 21, s = 5.7) 48.5 (n = 1, s = 0)Humerus Length 24.5 (n = 11, s = 4.4) 41.1 (n = 1, s = 0)
Proximal width a 7.8 (n = 11, s = 1.6) 18.8 (n = 4, s = 3.0)Distal widtha 11.4 (n = 10, s = 2.6) 23.4 (n = 6, s = 1.2)
Femur Length 23.1 (n = 26, s = 3.9) 38.6 (n = 3, s = 3.2)Proximal width a 6.0 (n = 26, s = 1.3) 13.3 (n = 7, s = 1.3)Distal widtha 7.7 (n = 25, s = 1.4) 15.1 (n = 2, s = 0.8)
Note: n, sample size; s, standard deviation.aWidth measured anteroposteriorly
Table 3. Average measurements (mm) of selected skeletal elements of Captorhinus aguti(based on data from Seltin 1959) and Captorhinus magnus.
Fig. 1. Photographs (A, C) and line drawings (B, D) of OMNH 56820, an incomplete cranium of Captorhinus magnus, in dorsal (A,B) and left lateral (C, D) views. en, external naris; f, frontal; l, lacrimal; n, nasal; p, parietal; pf, postfrontal; pm, premaxilla; prf,prefrontal.
4
measured eleven humeri of C. aguti, which produced an averageproximal width, distal width, and length of 7.8, 11.4, and25.5 mm, respectively. No complete C. magnus ulnae havebeen identified thus far, but the proximal portions of three
are known. In these specimens, the mediolateral width of theproximal end, which is measured at the distal base of thesigmoid notch (i.e., the maximum width of the proximalend), yields a mean value of 11.2 mm, whereas the left ulnaof C. aguti figured by Fox and Bowman (1966, fig. 29) hasa proximal width of 5.4 mm. The only C. magnus fibulaidentified thus far (OMNH 55390) has a total length of28.2 mm. The right fibula of C. aguti figured by Fox andBowman (1966, fig. 35) is 14.1 mm in length. No completepectoral or pelvic girdles are known for C. magnus, but severalspecimens do possess a complete glenoid fossa or acetabulum.The glenoid fossae of five C. magnus specimens possess anaverage anteroposterior length of 14.9 mm. Four pelvesproduced an average acetabular anteroposterior length anddorsoventral height of 15.3 and 10.3 mm, respectively. Inthe C. aguti girdles figured by Fox and Bowman (1966, figs.24, 32), these values are 10.0, 8.4, and 5.5 mm, respectively.
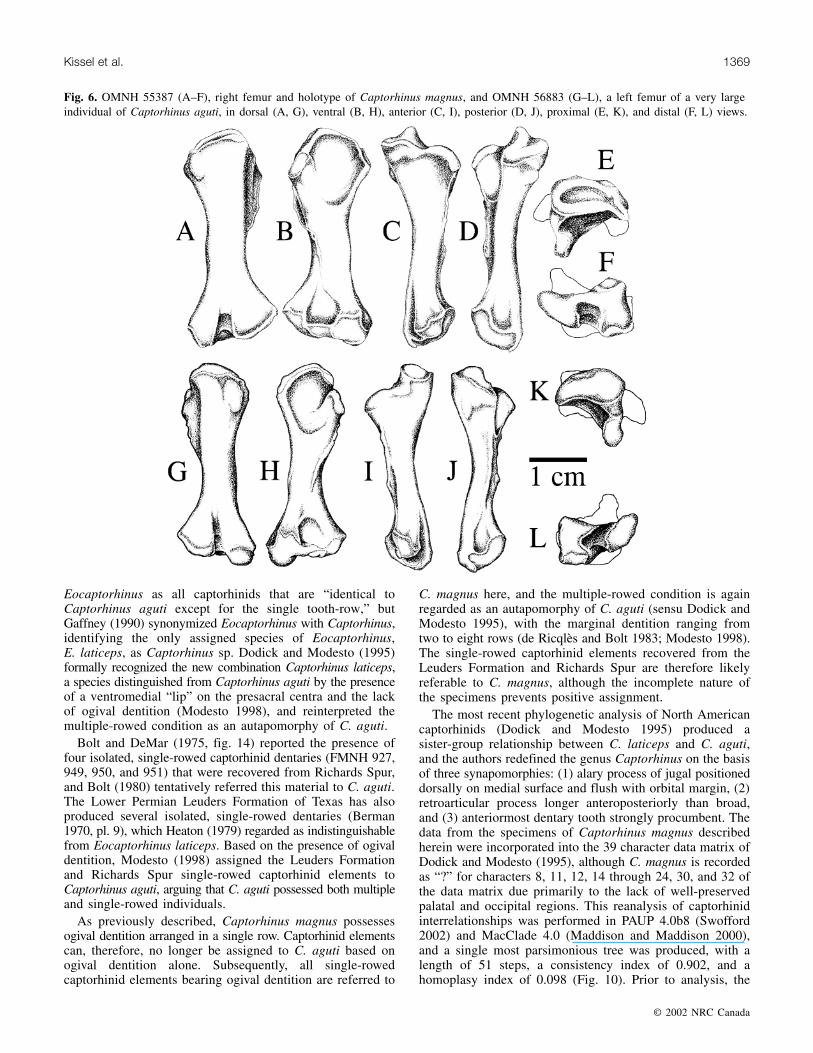
The average length and proximal and distal widths of themeasured C. magnus femora are 1.67, 2.22, and 1.96 timesas great as those of the 26 C. aguti femora examined bySeltin (1959; Table 3), respectively. Unlike most other skeletalelements, however, the femur of C. magnus can be furtherdistinguished from that of C. aguti based on features otherthan size. The femur of C. magnus (Fig. 6A–6F) possesses aconcave proximal articular surface in both immature andmature individuals. In contrast, the proximal femoral surfaceof C. aguti (Fig. 6G–6L) is strongly convex in all specimensexamined (n = 30), regardless of femoral size. Thus, theshape of the proximal articular surface of the femur allowsfor the distinction between similar-sized femora of C. agutiand C. magnus, regardless of the ontogenetic stage of theindividuals preserved (Fig. 6, 7). The femur of thecaptorhinid Labidosaurus, although somewhat similar in sizeto that of C. magnus, possesses a convex proximal articularsurface (Sumida 1989), and it has a much larger internaltrochanter than that of C. magnus.
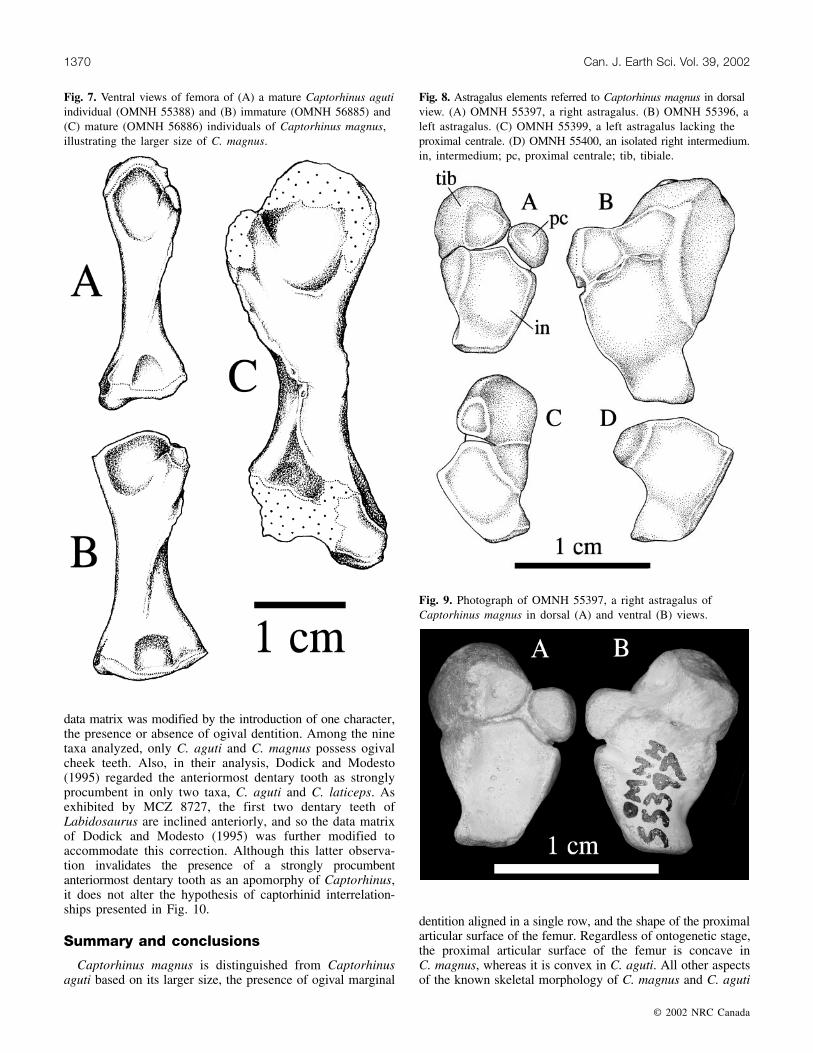
Except for its much greater size, the astragalus of Captorhinusmagnus (Figs. 8A–8B) is indistinguishable from that ofCaptorhinus aguti (Rieppel 1993, Figs. 5–7). It is stout andL-shaped in dorsal view, with the horizontal arm of the Lextending medially. A narrow proximal extension terminatesin a gently concave oval articular surface, which possesses adorsomedially orientated long axis, for the distal end of thefibula. The lateral surface is flat and forms the articular surfacefor the calcaneum along its length. The foramen for theperforating artery is located along the lateral margin near thedistal end of the bone. Distal to this foramen, the corner ofthe astragalus is a square-shaped process. The medial surfaceis characterized by the large, convex, and dorsomedially facingarticular surface for the tibia. The dorsal surface is smoothand concave, with an essentially flat surface rising at themargins, whereas the ventral surface is more irregular,possessing a rough, textured surface, a deep pit near thelaterodistal corner, and a deep groove that extendsproximomedially from the perforating foramen.
Discussion
On the ontogeny of the astragalus in captorhinidsThe homology of the astragalus of Captorhinus has been a
topic of considerable debate, with previous studies yielding
© 2002 NRC Canada
Kissel et al. 1367
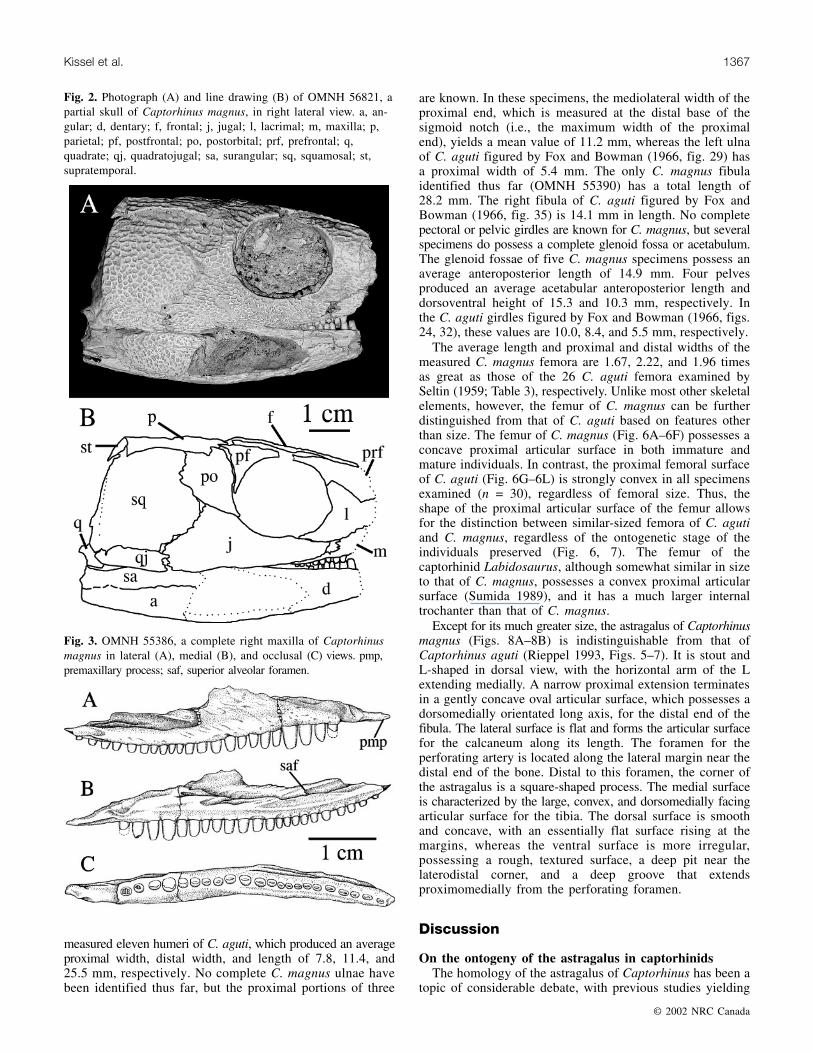
Fig. 2. Photograph (A) and line drawing (B) of OMNH 56821, apartial skull of Captorhinus magnus, in right lateral view. a, an-gular; d, dentary; f, frontal; j, jugal; l, lacrimal; m, maxilla; p,parietal; pf, postfrontal; po, postorbital; prf, prefrontal; q,quadrate; qj, quadratojugal; sa, surangular; sq, squamosal; st,supratemporal.
Fig. 3. OMNH 55386, a complete right maxilla of Captorhinusmagnus in lateral (A), medial (B), and occlusal (C) views. pmp,premaxillary process; saf, superior alveolar foramen.
5
two very different conclusions. Identifying suture patterns ontarsal elements from the Richards Spur fissure fills, Peabody(1951) concluded that the astragalus of Captorhinus is acomposite structure that formed through the fusion of threeseparate ossifications, the tibiale, intermedium, and proximalcentrale. Rieppel (1993) rejected this interpretation, arguingthat the structure identified as sutures by Peabody (1951)correspond to unfinished bone surface and (or) cracks.Based on studies of the developmental patterns of extantreptiles, Rieppel (1993) concluded that the reptilian (andprobably mammalian) astragalus is (1) a neomorph resultingfrom ontogenetic repatterning; and (2) a synapomorphy ofamniotes. Astragali recovered from Richards Spur and referredto Captorhinus magnus allow evaluation of these hypotheses.
Astragali of both mature and immature individuals ofC. magnus all possess well-defined sutures that exhibit atriradiate pattern. Open sutures are clearly visible on the dorsalsurface of immature specimens (e.g., OMNH 55397; Figs. 8A,9A). These sutures are closed along the ventral surface andrepresented by raised ridges (Fig. 9B). The astragalus of alarger, presumably more mature individual (OMNH 55396;Fig. 8B) exhibits closed sutures on both its dorsal and ventral
surface that are in the form of raised ridges and possess atriradiate pattern identical to that of the open sutures of theless well-ossified astragali. These sutures provide unequivocalevidence that the astragalus of Captorhinus is a compositestructure formed through the fusion of three ossifications, theintermedium proximally, the tibiale distomedially, and theproximal centrale distolaterally. On the medial and lateralsurfaces of both mature and immature forms, the sutures arerepresented by shallow grooves. The perforating foramen islocated at the lateral margin of the suture separating theintermedium and proximal centrale. Also recovered were anisolated right intermedium (OMNH 55400; Fig. 8D) and aleft astragalus with the proximal centrale separated cleanlyat the suture (OMNH 55399; Fig. 8C).
UCLA VP 3491, an undescribed juvenile specimen ofLabidosaurus, contains an isolated right intermedium, furthersupporting the presence of a tripartite astragalus in captorhinids.The unfused astragali of larger captorhinid taxa (e.g.,Captorhinus magnus and Labidosaurus), in contrast to thetypically completely fused astragali of the smaller forms(e.g., Captorhinus aguti, Captorhinus laticeps, andRhiodenticulatus), raises the possibility that delayed fusionmay have served to accommodate for extended growth and,subsequently, the attainment of larger sizes withinCaptorhinidae. The braincases of several captorhinid taxaprovide circumstantial evidence supporting this hypothesis.In the larger captorhinid taxa Labidosaurus (e.g., MCZ 8727)and Labidosaurikos (Dodick and Modesto 1995), the opisthoticis not fused to the surrounding elements, whereas it iscompletely fused to the supraoccipital and prootic in thesmaller Captorhinus aguti and Captorhinus laticeps (Foxand Bowman 1966, Heaton 1979). However, statistically sig-nificant data, such as the recovery of specimens permittingthe reconstruction of the complete captorhinid growth series,are needed to test this hypothesis.
Systematics of CaptorhinusUntil recently, the genus Captorhinus was diagnosed by
the presence of multiple rows of marginal teeth (e.g., Foxand Bowman 1966). Heaton (1979, p. 70) defined
© 2002 NRC Canada
1368 Can. J. Earth Sci. Vol. 39, 2002
Fig. 4. Photograph (A) and line drawing (B) of OMNH 55391, a nearly complete left lower jaw of Captorhinus magnus, in lateral view.a, angular; ar, articular, d, dentary; sa, surangular.
Fig. 5. OMNH 56923, a nearly complete left dentary of Captorhinusmagnus preserving the entire mandibular tooth row, in lateral (A)and occlusal (B) views.
6
Eocaptorhinus as all captorhinids that are “identical toCaptorhinus aguti except for the single tooth-row,” butGaffney (1990) synonymized Eocaptorhinus with Captorhinus,identifying the only assigned species of Eocaptorhinus,E. laticeps, as Captorhinus sp. Dodick and Modesto (1995)formally recognized the new combination Captorhinus laticeps,a species distinguished from Captorhinus aguti by the presenceof a ventromedial “lip” on the presacral centra and the lackof ogival dentition (Modesto 1998), and reinterpreted themultiple-rowed condition as an autapomorphy of C. aguti.
Bolt and DeMar (1975, fig. 14) reported the presence offour isolated, single-rowed captorhinid dentaries (FMNH 927,949, 950, and 951) that were recovered from Richards Spur,and Bolt (1980) tentatively referred this material to C. aguti.The Lower Permian Leuders Formation of Texas has alsoproduced several isolated, single-rowed dentaries (Berman1970, pl. 9), which Heaton (1979) regarded as indistinguishablefrom Eocaptorhinus laticeps. Based on the presence of ogivaldentition, Modesto (1998) assigned the Leuders Formationand Richards Spur single-rowed captorhinid elements toCaptorhinus aguti, arguing that C. aguti possessed both multipleand single-rowed individuals.
As previously described, Captorhinus magnus possessesogival dentition arranged in a single row. Captorhinid elementscan, therefore, no longer be assigned to C. aguti based onogival dentition alone. Subsequently, all single-rowedcaptorhinid elements bearing ogival dentition are referred to
C. magnus here, and the multiple-rowed condition is againregarded as an autapomorphy of C. aguti (sensu Dodick andModesto 1995), with the marginal dentition ranging fromtwo to eight rows (de Ricqlès and Bolt 1983; Modesto 1998).The single-rowed captorhinid elements recovered from theLeuders Formation and Richards Spur are therefore likelyreferable to C. magnus, although the incomplete nature ofthe specimens prevents positive assignment.
The most recent phylogenetic analysis of North Americancaptorhinids (Dodick and Modesto 1995) produced asister-group relationship between C. laticeps and C. aguti,and the authors redefined the genus Captorhinus on the basisof three synapomorphies: (1) alary process of jugal positioneddorsally on medial surface and flush with orbital margin, (2)retroarticular process longer anteroposteriorly than broad,and (3) anteriormost dentary tooth strongly procumbent. Thedata from the specimens of Captorhinus magnus describedherein were incorporated into the 39 character data matrix ofDodick and Modesto (1995), although C. magnus is recordedas “?” for characters 8, 11, 12, 14 through 24, 30, and 32 ofthe data matrix due primarily to the lack of well-preservedpalatal and occipital regions. This reanalysis of captorhinidinterrelationships was performed in PAUP 4.0b8 (Swofford2002) and MacClade 4.0 (Maddison and Maddison 2000),and a single most parsimonious tree was produced, with alength of 51 steps, a consistency index of 0.902, and ahomoplasy index of 0.098 (Fig. 10). Prior to analysis, the
© 2002 NRC Canada
Kissel et al. 1369
Fig. 6. OMNH 55387 (A–F), right femur and holotype of Captorhinus magnus, and OMNH 56883 (G–L), a left femur of a very largeindividual of Captorhinus aguti, in dorsal (A, G), ventral (B, H), anterior (C, I), posterior (D, J), proximal (E, K), and distal (F, L) views.
7
data matrix was modified by the introduction of one character,the presence or absence of ogival dentition. Among the ninetaxa analyzed, only C. aguti and C. magnus possess ogivalcheek teeth. Also, in their analysis, Dodick and Modesto(1995) regarded the anteriormost dentary tooth as stronglyprocumbent in only two taxa, C. aguti and C. laticeps. Asexhibited by MCZ 8727, the first two dentary teeth ofLabidosaurus are inclined anteriorly, and so the data matrixof Dodick and Modesto (1995) was further modified toaccommodate this correction. Although this latter observa-tion invalidates the presence of a strongly procumbentanteriormost dentary tooth as an apomorphy of Captorhinus,it does not alter the hypothesis of captorhinid interrelation-ships presented in Fig. 10.
Summary and conclusions
Captorhinus magnus is distinguished from Captorhinusaguti based on its larger size, the presence of ogival marginal
dentition aligned in a single row, and the shape of the proximalarticular surface of the femur. Regardless of ontogenetic stage,the proximal articular surface of the femur is concave inC. magnus, whereas it is convex in C. aguti. All other aspectsof the known skeletal morphology of C. magnus and C. aguti
© 2002 NRC Canada
1370 Can. J. Earth Sci. Vol. 39, 2002
Fig. 7. Ventral views of femora of (A) a mature Captorhinus agutiindividual (OMNH 55388) and (B) immature (OMNH 56885) and(C) mature (OMNH 56886) individuals of Captorhinus magnus,illustrating the larger size of C. magnus.
Fig. 8. Astragalus elements referred to Captorhinus magnus in dorsalview. (A) OMNH 55397, a right astragalus. (B) OMNH 55396, aleft astragalus. (C) OMNH 55399, a left astragalus lacking theproximal centrale. (D) OMNH 55400, an isolated right intermedium.in, intermedium; pc, proximal centrale; tib, tibiale.
Fig. 9. Photograph of OMNH 55397, a right astragalus ofCaptorhinus magnus in dorsal (A) and ventral (B) views.
8

are identical. Thus, without the presence of the femur or atooth-bearing element, distinguishing between individuals ofC. magnus and potentially very large individuals of C. agutiwould prove difficult. However, because skeletal elementsof C. magnus average nearly twice as large (1.8 times) asthose of C. aguti, the two species can likely be differentiatedwith size as the sole criterion. Furthermore, a larger samplesize consisting of more complete specimens of C. magnusmay permit the recognition of additional morphologicaldistinctions.
With its presence in C. magnus, ogival dentition can nolonger be considered an autapomorphy of C. aguti, and allsingle-rowed specimens of Captorhinus possessing ogivaldentition are referred to C. magnus. A phylogenetic analysis
of captorhinid interrelationships indicates that C. aguti andC. magnus form a clade that possesses a sister-grouprelationship with Captorhinus laticeps.
Incompletely ossified astragali referred to Captorhinusmagnus, as well as an isolated intermedium attributed toLabidosaurus, demonstrate that the astragalus of captorhinidswas formed through the fusion of three originally separateossifications, the tibiale, intermedium, and proximal centrale,rather than from a single ossification center. Berman andHenrici (in press) indicate an identical ontogenetic origin ofthe astragalus in Diadectidae, a clade within Diadectomorpha,the sister-group to Amniota (Reisz 1997). This apparent delayedfusion in larger members of Captorhinidae may have servedto permit the attainment of larger sizes within the taxon.
Although a statistical analysis is not possible because ofinadequate sample sizes, it was possible to compare theontogenetic stages of C. aguti and C. magnus. Femora ofsimilar ontogenetic stages of the two taxa remain vastly differentin size, and they exhibit different proportions (Figs. 6, 7).The femora of C. magnus are more robust than those ofC. aguti, possessing greater expansion of the proximal anddistal ends relative to the proximodistal length of the femur.In C. magnus, the average ratio of femoral proximalanteroposterior width to femoral length is 0.35, and the averageratio of femoral distal anteroposterior width to femoral lengthis 0.39. In C. aguti, these ratios are 0.26 and 0.33, respectively.This greater robustness, however, may simply reflect thelarger size of C. magnus, rather than a true morphologic feature,and exceptionally large individuals of C. aguti may also possessmore robust femora than those of smaller C. aguti individuals.
Captorhinus magnus has thus far been positively identifiedfrom only the Richards Spur locality, a site that also producesthe remains of C. aguti. Both species are represented throughoutthe entire stratigraphic sequence exposed at the locality, butthe relative abundance of C. magnus and C. aguti remainschanges throughout the interval. Fossils of C. magnus areknown predominately from the deeper regions of the fissurecomplex, where they are much more abundant than those ofC. aguti. In the upper, presumably younger sediments of thefillings, C. magnus remains are extremely rare and fossils ofC. aguti dominate the Richards Spur captorhinid assemblage.This relative abundance shift within the Richards SpurCaptorhinus population structure may record the ecologicalreplacement of the larger, single-rowed C. magnus by thesmaller, multiple-rowed C. aguti as the numerically dominantlocal Captorhinus species during the Early Permian. Alter-natively, if C. magnus and C. aguti shared an overlappinggeographic range during the Early Permian, then a change inthe local environment may have shifted the zone of overlappingranges away from the immediate area sampled at RichardsSpur. Such a shift could also result in the relative abundancechange recorded in the Richards Spur fossil assemblage.Unfortunately, collection at Richards Spur has not includedconsistent documentation of the stratigraphic position of thefossils; thus, the exact relationship, both stratigraphically andgeographically, between the C. magnus and C. aguti dominatedzones is not known precisely. Regardless, this distributionrefutes the possibility that C. magnus and C. aguti are sexualdimorphs of one species. If C. magnus and C. aguti are suchdimorphs, then a near equal distribution would be expectedthroughout the entire Richards Spur section.
© 2002 NRC Canada
Kissel et al. 1371
Fig. 10. Hypothesis of captorhinid interrelationships, modifiedfrom Dodick and Modesto (1995). Captorhinus aguti, Captorhinuslaticeps, Captorhinus magnus, Labidosaurikos meachami, andLabidosaurus hamatus form the ingroup. Protorothyris archeri,Romeria texana, Protocaptorhinus pricei, and Rhiodenticulatusheatoni represent the outgroups, with Protorothyris archeri servingas the non-captorhinid outgroup. Branches 1–3 are diagnosed bythe following autapomorphies: (1) maxillary dentition ends posteriorlyat level of orbital midpoint, postorbital region expanded laterally,skull table posterior margin with single median embayment, pterygoidtransverse flange narrow and tongue-like, parasphenoid with deepgroove present between cristae ventrolaterales, mandibular posteriorend acuminate, marginal “cheek” teeth chisel-shaped, anterior dentaryteeth procumbent; (2; diagnosing Captorhinus) jugal medial processpositioned dorsally on anterior process, retroarticular process elongate;(3) ogival marginal “cheek” teeth. The braincase and palate areunknown for C. magnus; however, since the known skull elementsof C. magnus are so similar to those of C. aguti, similarity in thesecharacters is assumed.
9

Acknowledgments
We thank Diane Scott of the University of Toronto atMississauga for advice regarding the preparation and illustrationof specimens. For the loan of specimens, we thank RichardCifelli of the Sam Noble Oklahoma Museum of Natural Historyat The University of Oklahoma, Norman, Oklahoma. Wealso thank Jeff Person of the Sam Noble Oklahoma Museumof Natural History for assistance with the cataloguing ofOMNH specimens. Reviews and comments by Drs. OlivierRieppel, Hans-Dieter Sues, and Stuart Sumida were greatlyappreciated and improved an earlier version of the manuscript.Finally, we would like to extend special thanks to WilliamMay for his continuing contribution to the understanding ofthe Lower Permain vertebrates of Oklahoma.
References
Berman, D.S. 1970. Vertebrate fossils from the Leuders Formation,Lower Permian of north-central Texas. University of CaliforniaPublications in Geological Sciences, 86: 1–61.
Berman, D.S., and Henrici, A.C. In press. Homology of the astragalusand structure and function of the tarsus of Diadectidae. Journalof Paleontology.
Bolt, J.R. 1980. New tetrapods with bicuspid teeth from the FortSill Locality (Lower Permian, Oklahoma). Neues Jahrbuch furGeologie und Palaeontologie, Monatshefte, 8: 449–459.
Bolt, J.R., and DeMar, R. 1975. An explanatory model of the evolutionof multiple rows of teeth in Captorhinus aguti. Journal ofPaleontology, 49: 814–832.
Case, E.C. 1911. A revision of the Cotylosauria of North America.Carnegie Institution of Washington Publication, 145: 1–122.
Cope, E.D. 1896. The reptilian order Cotylosauria. Proceedings ofthe American Philosophical Society, 34(149): 436–457.
Dilkes, D.W., and Reisz, R.R. 1986. The axial skeleton of the EarlyPermian reptile Eocaptorhinus laticeps (Williston). CanadianJournal of Earth Sciences, 23: 1288–1296.
Dodick, J.T., and Modesto, S.P. 1995. The cranial anatomy of thecaptorhinid reptile Labidosaurus meachami from the LowerPermian of Oklahoma. Palaeontology, 38: 687–711.
Fox, R.C., and Bowman, M.C. 1966. Osteology and relationshipsof Captorhinus aguti (Cope) (Reptilia: Captorhinomorpha). TheUniversity of Kansas Paleontological Contributions, Vertebrata,11: 1–79.
Gaffney, E.S. 1990. The comparative osteology of the Triassic turtleProganochelys. Bulletin of the American Museum of NaturalHistory, 194: 1–263.
Heaton, M.J. 1979. Cranial anatomy of primitive captorhinid reptilesfrom the Late Pennsylvanian and Early Permian of Oklahomaand Texas. Bulletin of the Oklahoma Geological Survey, 127: 1–84.
Kissel, R.A., Dilkes, D.W., and Reisz, R.R. 2001. A new captorhinidfrom the Lower Permian of Oklahoma, with comments on thehomology of the astragalus of Captorhinus. Journal of VertebratePaleontology, 21(3, Supplement): 68A.
Maddison, W.P., and Maddison, D.R. 2000. MacClade: analysis ofphylogeny and character evolution. Sinauer, Sunderland, Mass.
May, W.J., and Cifelli, R.L. 1998. Baeotherates fortsillensis, a newcaptorhinid reptile from the Fort Sill fissures, Lower Permian ofOklahoma. Oklahoma Geological Notes, 58: 128–137.
Modesto, S.P. 1996. A basal captorhinid reptile from the Fort Sillfissures, Lower Permian of Oklahoma. Oklahoma GeologicalNotes, 56: 4–14.
Modesto, S.P. 1998. New information of the skull of the EarlyPermian reptile Captorhinus aguti. PaleoBios, 18: 21–35.
Modesto, S.P. 1999. Colobomycter pholeter from the Lower Permianof Oklahoma: a parareptile, not a protorothyridid. Journal ofVertabrate Paleontology, 19: 466–472.
Olson, E.C. 1947. The family Diadectidae and its bearing on theclassification of reptiles. Fieldiana: Geology, 11: 1–53.
Olson, E.C. 1967. Early Permian vertebrates. Oklahoma GeologicalSurvey, Circular 74.
Olson, E.C. 1991. An eryopid (Amphibia: Labyrinthodontia) fromthe Fort Sill fissures, Lower Permian, Oklahoma. Journal ofVertebrate Paleontology, 11: 130–132.
Peabody, F.E. 1951. The origin of the astragalus of reptiles. Evolution,5: 339–344.
Ricqlès, A. de, and Bolt, J.R. 1983. Jaw growth and tooth replacementin Captorhinus aguti (Reptilia: Captorhinomorpha): a morphologicaland histological analysis. Journal of Vertebrate Paleontology,3: 7–24.
Reisz, R.R. 1997. The origin and early evolutionary history ofamniotes. Trends in Ecology & Evolution, 12: 218–222.
Rieppel, O. 1993. Studies on skeleton formation in reptiles. IV.The homology of the reptilian (amniote) astragalus revisited.Journal of Vertebrate Paleontology, 13: 31–47.
Romer, A.S. 1956. The Osteology of Reptiles. University of ChicagoPress, Chicago, Ill.
Seltin, R.J. 1959. A review of the family Captorhinidae. Fieldiana:Geology, 10: 461–509.
Sullivan, C., and Reisz, R.R. 1999a. First record of Seymouria(Vertebrata: Seymouriamorpha) from the Early Permian fissurefills at Richards Spur, Oklahoma. Canadian Journal of EarthSciences, 36: 1257–1266.
Sullivan, C., and Reisz, R.R. 1999b. The age, depositional environmentand tetrapod paleofauna of the Lower Permian fissure fills atRichards Spur, Oklahoma. Journal of Vertebrate Paleontology,19(Supplement to Number 3): 79A.
Sullivan, C., Reisz, R.R, and May, W.J. 2000. Large dissorophoidskeletal elements from the Lower Permian Richards Spur fissures,Oklahoma, and their paleoecological implications. Journal ofVertebrate Paleontology, 20: 456–461.
Sumida, S.S. 1989. The appendicular skeleton of the Early Permiangenus Labidosaurus (Reptilia, Captorhinomorpha, Captorhinidae)and the hind limb musculature of captorhinid reptiles. Journal ofVertebrate Paleontology, 9: 295–313.
Swofford, D.L. 2002. PAUP: phylogenetic analysis using parsimony.Version 4.0b8. Smithsonian Institution, Washington, D.C.
Watson, D.M.S. 1917. The evolution of the tetrapod shoulder girdleand forelimb. Journal of Anatomy, 52: 1–63.
© 2002 NRC Canada
1372 Can. J. Earth Sci. Vol. 39, 2002
10
All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.
Copyright © 2022 FDOKUMEN




















