
O Dever de Mitigar o Prejuízo (Duty to Mitigate the Loss) e a Responsabilidade Civil do Estado
Upload
khangminh22Category
view
2download
0
Behaviour and crop-raiding patterns of Asian Elephants
(Elephas maximus): Can beehive fences help mitigate
human-elephant conflict in Sri Lanka?
Kylie Michelle Butler
B.Sc.(Ecology and Sustainability) (Hons.)
M. Env
A thesis submitted in total fulfillment of the requirements
for the degree of Master of Philosophy (Environmental Science)
Faculty of Science
School of Environmental and Life Sciences
University of Newcastle
New South Wales, Australia
August 2019
Acknowledgement of Government Support
This research was supported by an Australian Government Research Training (TRP) Scholarship.

Statement of Originality
I hereby certify that the work embodied in the thesis is my own work, conducted under normal
supervision. The thesis contains no material which has been accepted, or is being examined, for the
award of any other degree or diploma in any university of other tertiary institution and, to the best of
my knowledge and belief, contains no material previously published or written by another person,
except where due reference has been made. I give consent to the final version of my thesis being
made available worldwide when deposited in the University’s Digital Repository, subject to the
provisions of the Copyright Act 1968 and any approved embargo.
X. Kylie M. Butler
Ethical Note
All field work was conducted under the University of Newcastle’s Animal Ethics Committee Reference
No. A-2014-413, Human Ethics Committee Reference No. H-2014-0101, with Approval from the
Department of Wildlife and Conservation, Sri Lanka, Reference No. WL/3/2/66/14.
Funding Bodies
I gratefully acknowledge funding support from the following organisations: Elephant Action League;
Save the Elephants; Chester Zoo Conservation Grant; Phoenix Zoo Conservation and Science Grant;
Rufford Small Grants for Nature Conservation; Taronga Zoo Field Conservation Grant; Zoos Victoria
Conservation Grant; and The Transpetrol Foundation.

Table of Contents
List of Figures vi
List of Tables viii
Acknowledgements x
Abstract xiii
Chapter 1: Introduction
1.1 Human-Wildlife Conflict 2
1.2 Human-elephant conflict 3
1.2.1 Human-elephant conflict in Sri Lanka 4
1.3 Characteristics and behaviour of elephants in high human-elephant conflict areas 7
1.3.1 Risk taking behaviour 7
1.4 Human-elephant conflict mitigation 9
1.4.1 Beehive fencing 11
1.5 Rationale 13
1.6 Thesis Overview 14
1.7 References 16
Chapter 2: General behaviour patterns of wild elephants utilising areas of varying anthropogenic
risk to elephants, in Sri Lanka
2.1 Abstract 29
2.2 Introduction 29
2.3 Methods 36
2.3.1 Study site 36
2.3.1.1 Wasgamuwa National Park (low risk zone) 37
2.3.1.2 Weheragala Tank (medium risk zone) 38
2.3.2 Study subjects 40
2.3.3 Data collection: elephant observations 45
2.3.4 Behaviour coding from video footage 48
2.3.5 Statistical analysis 51
2.4 Results 52
2.4.1 General behavioural activities 52

2.4.2 Specific ‘reactive’ behaviours 60
2.4.3 Influence of prior negative interactions with humans (EHI levels) 61
2.5 Discussion 64
2.5.1 General behaviour patterns in Wasgamuwa National Park and Weheragala Tank 64
2.5.2 Effects of context and sex on elephant behaviour 66
2.6 Conclusion 70
2.7 References 72
Chapter 3: Crop-raiding in a high human-elephant conflict location in Sri Lanka: an assessment of
the characteristics and patterns of crop-raiding in Dewagiriya Village
3.1 Abstract 81
3.2 Introduction 81
3.3 Methods 86
3.3.1 Study site 86
3.3.2 Data collection 89
3.3.3 Statistical analysis 92
3.4 Results 93
3.4.1 Elephant events by raid days 93
3.4.2 Factors influencing elephant presence in Dewagiriya Village 96
3.4.3 Factors influencing within-site variations in elephant events 100
3.4.4 Damage inflicted by elephants 101
3.5 Discussion 105
3.5.1 Characteristics of crop-raiding elephants 106
3.5.2 Influences of crop-raiding behaviour 107
3.5.3 Within-site variations in elephant events 110
3.6 Conclusion 112
3.7 References 113
Chapter 4: Beehive fences as a human-elephant conflict mitigation tool for farmers in rural Sri
Lanka
4.1 Abstract 119
4.2 Introduction 119
4.2.1 Human-elephant conflict mitigation 120

4.2.2 Elephant interactions with bees and beehive fences – Africa 122
4.2.3 Elephant interactions with bees and beehive fences – Asia 124
4.3 Methods 126
4.3.1 Study site 126
4.3.2 Introducing the concept of beehive fencing and participant selection for the beehive fence
trial 128
4.3.3 Establishment of beehive fences 132
4.3.4 Data collection: elephant events 136
4.3.5 Fence maintenance 137
4.3.6 Beekeeping 139
4.3.7 Cost analysis 140
4.3.8 Farmer acceptance and willingness to participate in using beehive fencing 140
4.3.9 Statistical analysis 140
4.4 Results 141
4.4.1 Elephant events and the deterrent effect of beehive fences 141
4.4.2 Fence maintenance 145
4.4.3 Beekeeping and honey harvests 147
4.4.5 Cost of beehive fences 149
4.4.6 Farmer acceptance and willingness to participate 152
4.5 Discussion 153
4.6 Conclusion 157
4.7 References 158
Chapter 5: Discussion
5.1 Introduction 167
5.2 Summary of results 167
5.3 Methodological challenges 169
5.3.1 Behavioural assessments 169
5.3.2 Profiling of crop-raiding 171
5.3.3 Beehive fence trials 171
5.4 Closing statements 173
5.5 References 175

List of Figures
Figure 1.1: Sri Lanka: National Parks and elephant distribution 5
Figure 1.2: Example of a beehive fence 12
Figure 2.1: Overview of study site 37
Figure 2.2: South-eastern section of WNP 39
Figure 2.3 Weheragala Tank ‘medium risk zone’ 39
Figure 2.4: Example from Elephant ID File. Mohan 41
Figure 2.5: Elephant with a high elephant-human interaction rating 43
Figure 2.6: Elephant with a medium elephant-human interaction rating 44
Figure 2.7: Elephant with a low elephant-human interaction rating 44
Figure 2.8: Elephant observations in Wasgamuwa National Park 46
Figure 2.9: Elephant observations to the north of Weheragala Tank 47
Figure 2.10: Elephant observations from south-west of Weheragala Tank 47
Figure 2.11: % time spent standing still (without feeding) in WNP 55
Figure 2.12: % time spent in locomotion at WT 57
Figure 2.13: % time spent feeding at WT 58
Figure 2.14: % time spent standing still (without feeding) at WT 59
Figure 2.15: Vehicles and elephants in WNP (LRZ) 67
Figure 2.16: Elephant smells towards research vehicle in WNP (LRZ) 67
Figure 2.17: Elephants fleeing from the clearing towards the forest at WT 68
Figure 3.1: Location of study site: Dewagiriya Village position in Sri Lanka 87
Figure 3.2: Map of the Dewagiriya Village study area 90
Figure 3.3: Camera trap footage of elephants in Dewagiriya Village 95
Figure 3.4: Elephant events per month based on ‘best’ model predictors 97
Figure 3.5: Elephant events per year based on ‘best’ model predictors 98
Figure 3.6: Relationship between raid day rain and elephant events based on ‘best’ model
predictors 99
Figure 3.7: Elephant damage in Dewagiriya Village 102
Figure 4.1: Diagram of a beehive fence 123
Figure 4.2: A beehive fence 123
Figure 4.3: Study site location 127

Figure 4.4: Map of Dewagiriya Village showing experimental and control households 131
Figure 4.5: Langstroth beehive secured with coconut rope 132
Figure 4.6: Langstroth beehive secured with ‘z’ clips 132
Figure 4.7: Painting fence posts with old engine oil to protect from termites 133
Figure 4.8: A beehive fence surrounding the home garden of the Jayasena family 133
Figure 4.9: Beehive fence shade roofs 134
Figure 4.10: Layout of the ten beehive fences 135
Figure 4.11: Fence maintenance issues 138
Figure 4.12: Number of elephant events: Beehive fenced (experimental) vs. non-beehive fenced
(control) home gardens 142
Figure 4.13: Frequency of elephants entering gardens during an elephant event 143
Figure 4.14: Relationship between elephant breakthroughs and number of occupied beehives 145

List of Tables
Table 2.1: Elephant-human interaction (EHI) ratings and definitions 43
Table 2.2: Elephant ethogram 48
Table 2.3: % time spent in general behavioural activities 53
Table 2.4: LMM summary: effects of context * sex on general behaviour in WNP 54
Table 2.5: Relationship between treatment * sex of elephants in the MRZ 56
Table 2.6: Relationship between sex, context and the interaction between sex and context for
reactive behaviours in WNP and WT 60
Table 2.7: Relationship between treatment * EHI levels of elephants in WNP 61
Table 2.8: Relationship between treatment * EHI levels of elephants in WNP and reactive
behaviours 62
Table 2.9: Relationship between treatment * EHI levels of elephants at WT 63
Table 2.10: Relationship between treatment * EHI levels of elephants at WT and reactive
behaviours 64
Table 3.1: Moon phase definitions 91
Table 3.2: Results from models predicting likelihood of elephant events in Dewagiriya Village
96
Table 3.3: Results from models predicting within-site variations in frequency of elephant events
per household 101
Table 3.4: Crop and plant types grown and damaged in Dewagiriya Village 103
Table 3.5: Results from models predicting likelihood of damage during elephant events in
Dewagiriya Village 104
Table 3.6: Comparison of annual crop-raiding counts per km² across HEC locations 105
Table 4.1: List of households experiencing the most negative impacts of crop-raiding in
Dewagiriya Village 130
Table 4.2: Fence establishment and data collection periods for each experimental and control
household 136
Table 4.3: Beekeeping training and workshop record 139
Table 4.4: GLMM summary: the effect of hive numbers and occupied hives on fence break-
throughs 144
Table 4.5: Presence/absence counts of different pests found in beehives 146

Table 4.6: Asian honey bee colony introductions and abscondments 148
Table 4.7: Number of active beehives per fence 148
Table 4.8: Honey harvesting records per household 149
Table 4.9: Cost of beehive fence materials and equipment per fence 150
Table 4.10: Cost estimates for colony establishment and feeding per fence 151
Table 4.11: Cost estimate for the fencing materials, tools/equipment and colonies overall (per
fence) 151

Acknowledgements
This study has been an exciting, challenging, sometimes frustrating and exceedingly beautiful journey
into the world of Sri Lankan wildlife, conservation and communities. There are many people who have
played a significant role in this adventure, and many who deserve my warmest gratitude.
My academic supervisors Dr. Andrea Griffin and Assoc. Prof. Natalie Moltchaniswkyj, I thank you for
taking me on board, for your encouragement and for pushing me beyond my academic comfort zone.
I know I haven’t been the easiest student but there are many valuable lessons I have gained from your
expertise. My industry supervisor Dr. Lucy King, I could not have asked for a better mentor and am
forever grateful for your faith in allowing me to develop and lead the beehive fence trial in Sri Lanka.
There were many hurdles along the way: some I was prepared for but many for which I was not. This
project opened my eyes to many facets of life in conservation and your support was invaluable both
professionally and personally. I extend my gratitude to the entire Save the Elephants and Elephants
and Bees teams – it’s been an honour.
To my absolute superstar of a research assistant Supun Herath: you joined our team on day one and I
don’t know what I would have done without you. Thank you isn’t enough to express my appreciation
for your endless hard work, your English-Sinhalese translations, for putting up with the beestings and
constantly fixing fences, for keeping me safe from the elephants, and for letting Arrow in your tuk tuk.
A huge thank you also to Sarath and Siriya for always being available to help with anything. The three
of you were my A-Team and now lifelong friends: I miss our sunset hangs on the rock.
Many thanks to all from the Sri Lankan Wildlife Conservation Society who contributed, especially
Chathuranga Dharmarathne for your support during a particularly challenging field season, and to
Swarna and Leela for your endless smiles, curries and kindness towards me and my posse of village
dogs. Your friendship is always in my heart. I am also extremely grateful to local and international
beekeepers Dr. R.W.K. Punchihewa, Mr. Frank Ryde, Dr. David Gormley-O’Brien and Ms. Emma Collins.
Supun and I learned so much from your expertise, as did the Dewagiriya community. David and Emma,
that you came all the way from Australia to help a complete beekeeping novice, on a project you’d
just heard of, is beyond amazing.

Thank you also to my fantastic team of interns who helped with tasks big and small at various stages
of the project: Lauren Ross, Lorna McCallister, Manon van der Meer and Louise Jory. You guys slogged
it out through torrential rains, intense heat, charging elephants, sleeping in leaky rooms, power
outages, snakes and scorpions. And to those who took it one step further and came out to lead the
field team after I returned to Australia: Jessica van Fleteren, Rebecca Sargeant, Robin Cook and Zaineb
Akbarally. You often had to put up with far more than you’d bargained for and I am so grateful for all
your hard work. Special extra thanks to intern and project leader Brianna Stoyle who had probably the
hardest job of all wrapping up our final field season. All of you are absolute gems.
I am also thankful for the support, practical assistance, and company of new and old friends from
around the globe. Jo Heaton, Simon King, Jinalee de Silva, Hollis Burbank-Hammarlund, Yufei Gao,
Sarah Reid-Anderson and Natalie Gonzalez – thank you for helping in the field, for the great
conversations, and the sunset Lions. To friends who visited me on-site and often found themselves
digging holes and taking notes – I appreciate it immensely. That means you: Adam ‘Squire’ Mattinson,
James Ahern, Tim ‘BC’ Hunt, Lynsey Humes, Alistair ‘Stunt’ Hunt, and Joel Graham.
To the beautiful Dewagiriya community, I know you won’t read this but I want send my heartfelt
gratitude out to the universe. I was a stranger introducing a seemingly crazy idea about bees and
elephants, and I was welcomed warmly. Thank you to everyone who shared their stories and
experiences, who built and monitored fences, and for the endless cups of sugary black tea. I have no
words for how lucky I feel to have met you all. Special thanks to the Sobana, Jaysinghe, Sirisena,
Somathilaka, Ariyadasa, Guneris, Somanawathi, Dharmadasa, Ghanawathi, Karunadasa, and Jayasena
families for participating in the experimental trial. I wish you all happiness in your families, success in
your harvests, and safety living in proximity to the elephants.
A big shout-out to my brainy ‘cuz’ Nicky Butler who let me rant and rave, read her excellent thesis for
guidance, and found me an awesome statistics tutor. And to Alex Masey – without you, my data would
still be sitting in excel spreadsheets. Thank you for making ‘R’ less scary and for your patience and the
time you took away from your own work to help me out.
There are only a few people who truly ‘get’ why someone wants to gallivant around the globe chasing
elephants and bees and living a never-ending student life. Everyone mentioned here understood

something unique to this experience: either what it’s like to work in this industry or why I sometimes
have to do things the way I do: Tamsin Webb and Sundari Sitaram, there are some things that can only
be truly understood by people doing something along the same lines and I am so glad we connected.
Kirby Smith, Camille Royal and Susan Gorecki thank you for listening to my elephant tales and helping
with some tough decisions. Sarah Leatham and Dianne Butler, thanks for visiting me in Sri Lanka, for
the wine dates, and for ‘getting’ me.
To my husband Morgan Tipper, thank you for everything: for coming out to Sri Lanka and doing
whatever was needed in the field, for your technical mapping and computer assistance, for supporting
us financially, for your patience with having an often-absent partner and wife, for never getting tired
of me blabbing on about my work, and most importantly for always being on my side. Whether
intended or not, this adventure has been as much yours as mine. Let’s see what’s next!
Finally, I would like to dedicate this project to the late Mr. Jayasena and his wife Mrs. Jaysena. Both
respected elders in Dewagiriya Village, Mr. Jayasena passed away before his time in 2016 from Chronic
Kidney Disease (CKD). Sadly, many community members are afflicted by this due to contaminated
water sources. Their joint support of Mr. and Mrs. Jayasena and their enthusiasm for the beehive
fence project was instrumental in making it a reality. Their smiles and shared stories will remain in my
heart forever.
Mrs. Jayasena holding a photo of her
husband and I next to their beehive
fence. © Zaineb Akbarally

Abstract
Human-elephant conflict (HEC) in the form of crop-raiding, is a major conservation challenge to the
long-term survival of elephant populations, simultaneously threatening the livelihoods and personal
safety of people living in proximity to elephants. The widespread problem of HEC has led to a great
deal of research into the causes, consequences and predictors of elephant crop-raiding activity.
However, despite similarities across HEC situations, site-specific differences are also apparent.
Furthermore, most studies focus on one facet of HEC when it is a complex issue requiring
understanding of local elephant behaviour, identification of the characteristics and patterns of crop-
raiding at the local scale, and careful implementation and monitoring of mitigation strategies.
In this study, I selected a region of Sri Lanka experiencing high levels of HEC and sought to provide an
in-depth assessment of the site-specific situation generated over a three-year period. Specifically, we
aimed to: identify general patterns of behaviour occurring in local areas representing differing levels
of anthropogenic disturbance to elephants; profile patterns and predictors of crop-raiding activity in
a village heavily impacted by HEC; and test the effectiveness of beehive fencing as an Asian elephant
deterrent tool.
First, I provide initial evidence that elephants inhabiting areas of ‘medium’ level anthropogenic
disturbance outside of protected boundaries, interrupt feeding and increase ‘reactive’ behaviours
such as smelling and holding the ‘vigilance’ posture in response to immediate anthropogenic threats
in the environment. In the absence of any known disturbances, there was no difference in general
behaviours of male or female elephants between the two risk zones. Secondly, I show that elephant
crop-raiding in Dewagiriya Village occurs year-round and follows no clear seasonal patterns. Similar to
other HEC situations, male elephants are the predominant crop-raiders, and crop-raiding occurs
almost exclusively at nights. Within-site variations in crop-raiding intensity were also identified, with
properties closest to water tanks and forest habitat the most vulnerable. Finally, our three-year
beehive fence trial showed that households using beehive fences around their gardens had
significantly less elephant visits into their gardens then households without. Still, difficulties in
attracting natural colonies, poor honey production, set-up costs, and farmer motivations were barriers
to success.

This study contributes to the general body of knowledge on elephant behaviour in anthropogenically
influenced contexts, and specifically on patterns of crop-raiding and mitigation efforts. Further
research into the potential of beehive fences as an Asian elephant deterrent, preferably in a location
more amenable to beekeeping, would help to determine the value of expanding this technique further
in Sri Lanka, and elsewhere in Asia.

1
Chapter 1: Introduction

2
1.1 Human-wildlife conflict
Human-wildlife conflict (HWC), where the actions of wildlife negatively impact the well-being of
humans or vice versa, is an ongoing and increasing global environmental concern (Manfredo 2015;
Baruch-Mordo et al. 2008; Madden 2004). An array of wildlife from rodents to large mammals, are
implicated in HWC (Harich et al. 2013; Nijman and Nekaris 2010; Woodroffe et al. 2005b; Gillingham
and Lee 2003), encompassing varying situations from squirrels eating crops (Harich et al. 2013), to
wild dogs predating on livestock (Woodroffe et al. 2005a), to bears posing threats to park-goers (Lewis
et al. 2015; Baruch-Mordo et al. 2008). On the other side of the coin, humans often drive conflict by
hunting or poaching, such is the case with the illegal killing or capturing of wild animals for 'bush meat'
(Ripple et al. 2016) or private pet trades (Menard et al. 2013).
Wildlife entering farmlands to forage on crops, commonly known as 'crop-raiding', has been a
prominent HWC concern for decades and remains topical with no signs of abating (examples: Saraswat
et al. 2015; Hoffmeier-Karimi and Schulte 2014; Thapa 2010; Gillingham and Lee 2003; Bandara and
Tisdell 2002; Rao et al. 2002; Studsrod and Wegge 1995; Sukumar 1990). Communities of lower socio-
economic status, particularly those who live on the fringe of traditional wildlife habitat and rely on
subsistence farming, are often disproportionately affected by crop-raiding (Barua et al. 2013; Ogra
2008; Rao et al. 2002), and many animals have been described, fairly or otherwise, as 'agricultural
pests' including rodents (e.g. squirrels, Sciuridae spp: Harich et al. 2013), primates (e.g. rhesus
macaques, Macaca mulatta, Saraswat et al. 2015; and baboons, Papio anubis and Papio cynocephalus:
Hill 2000), ungulates (e.g. barking deer, Muntiacus muntjak: Rao et al. 2002, and peccaries, Tayassu
tajacu: Perez and Pacheco 2006) and large terrestrial mammals (e.g. elephants, Loxodonta africana,
Hoffmeier-Karimi and Schulte 2014; Elephas maximus: Santiapillai et al. 2006; and rhinoceros,
Rhinoceros unicornis: Thapa 2010).
Injury or death to both humans and wildlife is a serious consequence of HWC involving crop-raiding
(Barua et al. 2013; Choudhury 2004; Rao et al. 2002). Additionally, substantial economic losses (e.g.
when property or crops are damaged) and social costs (e.g. interruption of schooling or work due to
the presence of potentially dangerous wildlife) can have ongoing deleterious effects on people (Barua
et al. 2013; Harich et al. 2013; Hartter et al. 2011; Ogra 2008; Thirgood et al. 2005; Hill 2000). As
humans struggle to cope with consequences of HWC, negative perceptions of co-existence with

3
certain wildlife species grow (Dickman 2010; Madden 2004). Although not always responsible for the
most serious damage, species of a larger size which typically present a higher level of perceived
physical danger to humans, tend to receive the least tolerance and elicit harmful retaliatory actions
from people (Hoffmeier-Karimi and Schulte 2014; Ogra 2008; Naughton-Treves and Treves 2005; Hill
2004; Bandara and Tisdell 2002).
With the continuing growth of human populations, and expansion of communities into land inhabited
by wildlife, it seems inevitable that human-wildlife conflicts will only increase (Manfredo 2015; Harich
et al. 2013; Redpath et al. 2013; Fernando et al. 2005; Madden 2004). As a result, the conservation
efforts and long-term survival of several endangered and iconic wildlife species are being threatened
(Graham et al. 2010; Naughton-Treves and Treves 2005; Loe and Roskaft 2004; Thirgood et al. 2005;
Madden 2004). A prime example of this is the conflict involving humans and both African and Asian
elephants (Graham et al. 2010; Perera 2009; Sitati and Walpole 2006; Bandara and Tisdell 2002).
1.2 Human-elephant conflict
There are various forms of conflict involving humans and elephants, the two most common being the
illegal poaching of elephants to harvest tusks for the ivory trade, and crop-raiding by elephants.
Poaching primarily occurs in Africa where both male and female elephants typically have tusks (Ihwagi
et al. 2018; Burn et al. 2011; Kahindi et al. 2009), whereas crop-raiding is a major HEC issue in almost
all African and Asian elephant range countries (Okello 2016; Davies et al. 2011; Webber et al. 2011;
Graham et al. 2010; Perera 2009; Sitati and Walpole 2006; Sitati et al. 2003). Both are major
conservation challenges to the long-term survival of elephant populations however it is the issue of
crop-raiding that forms the focus of this study.
Although African and Asian elephants are distinct species, with many unique morphological and
behavioural characteristics, there are several fundamental characteristics of elephant crop-raiding
that transcend locations, countries and continents. In most locations, the primary drivers of HEC are
the loss or fragmentation of traditional elephant habitat and disruption of elephant movement
patterns, mainly due to deforestation, agricultural expansion and human encroachment into, or very

4
close to, elephant inhabited areas (Amwata and Mganga 2014; Chakraborty and Mondal 2013;
Campos-Arceiz et al. 2009; Fernando et al. 2005; Choudhury 2004; Sitati et al. 2003). Common
consequences of HEC are personally significant economic losses to farmers when crops are eaten or
trampled (Okello et al. 2016; Amwata and Mganga 2014; Gubbi 2012; Ekanayaka et al. 2011; Fernando
et al. 2005), damage to property as elephants attempt to access crops stored inside (Campos-Arceiz
et al. 2009), injury or death to humans and elephants (Chakraborty and Mondal 2013; Gubbi 2012;
Choudhury 2004; Sitati et al. 2003), disturbed sleep and work patterns as people are required to
continuously guard crops (Barua et al. 2013; Hill 2004), and negative impacts on human physical and
mental health (Barua et al. 2013; Ogra 2008; Sukumar 2006; Hoare 2000). Unsurprisingly, HEC fuels
negative perceptions of co-existing with elephants and hinders local conservation efforts (Amwata
and Mganga 2014; Chakraborty and Mondal 2013; Fernando et al. 2005).
Several common factors that influence the likelihood of elephants raiding crops have also been
identified across multiple sites. These include the proximity of villages and farms to elephant habitat
(Barnes et al. 2015; Gubbi 2012; Graham et al. 2010; Sitati et al. 2003), the types of crops grown and
the growth stage of crops, with ripening maize and rice particularly susceptible (Chakraborty and
Mondal 2013; Gubbi 2012; Webber et al. 2011; Graham et al. 2010; Campos-Arceiz et al. 2009; Chiyo
et al. 2005), and seasonal changes and rainfall (Gubbi 2012; Webber et al. 2011; Campos-Arceiz et al.
2009; Fernando et al. 2005). It is typical for male elephants to raid crops more frequently than female
elephants (Thaufeek et al. 2014; Das et al. 2014; Graham et al. 2010) and crop-raiding occurs almost
exclusively at nights (Das et al. 2014; von Gerhardt et al. 2014; Graham et al. 2010).
Despite the aforementioned commonalities across sites, it is wise to exercise caution when making
generalisations, even if the crop-raiding sites in question are in the same country or the same area of
a given country, and particularly when making management or mitigation decisions.
1.2.1 Human-elephant conflict in Sri Lanka
Sri Lanka is a country with a long and complicated history of human-elephant interactions. Although
elephants are traditionally revered as a cultural and religious icon (Fernando et al. 2011; Santiapillai

5
et al. 2006; Bandara and Tisdell 2003), wild elephants have historically been subjected to high levels
of negative interactions with humans. From the hunting and capture of elephants in the early 1900's
(Agar n.d; Hennessey 1929), to being caught in the crossfire of a lengthy and violent civil war
(Santiapillai et al. 2006; de Silva 1998; Alahakoon and Santiapillai 1997), to deforestation and habitat
loss (Haturusinghe and Weerakoon 2012; Wikramanayake et al. 2004), the relationship between
humans and elephants in Sri Lanka has been fraught with conflict. Crop-raiding has been discussed in
popular media since the mid 1900's (Hennessey n.d; Tutein-Nolthenius n.d) however it was around
the mid 1990's when it began to gain global attention as a serious elephant conservation issue (de
Silva 1998; Santiapillai 1996; Santiapillai and de Silva 1994). In 2005, HEC was recognised as the
principal threat to the survival of Sri Lanka's wild elephant population (Fernando et al. 2005) and it
continues to be a major issue in almost all elephant inhabited areas of Sri Lanka (Fernando et al. 2019;
Santiapillai et al. 2010) (Figure. 1.1).
Figure 1.1: Sri Lanka: National Parks and elephant distribution © Morgan Tipper

6
As in other locations where crop-raiding occurs, the disruption of traditional elephant habitat and
ongoing expansion of agriculture closer to forests perpetuate interactions between humans and
elephants (Haturusinghe and Weerakoon 2012; Fernando et al. 2011; Campos-Arceiz et al. 2009;
Santiapillai 1996). Specifically, large-scale development including major damming projects for
irrigation and the resettlement of people near forest boundaries, are bringing humans and elephants
into direct competition for resources (Fernando et al. 2011; Santiapillai et al. 2006; de Silva 1998).
It is estimated that more than 200 elephant (Fernando et al. 2011) and 50 human (Campos-Arceiz et
al. 2009; Perera 2009) deaths occur annually in Sri Lanka as a consequence of HEC. Non-fatal injuries
to elephants and humans (Thaufeek et al. 2014; Campos-Arceiz et al. 2009; Wikramanayake et al.
2004), crop loss (Thaufeek et al. 2014; Haturusinghe and Weerakoon 2012; Campos-Arceiz et al. 2009;
Wikramanayake et al. 2004), property damage (Thaufeek et al. 2014; Haturusinghe and Weerakoon
2012; Campos-Arceiz et al. 2009), and disruption of people's regular activities due to crop-guarding
efforts and being unable to safely move about the village when elephants are present, are further
serious ramifications (Santiapillai et al. 2010; de Silva 1998).
Elephants raid a wide variety of fruits, vegetables and grains in Sri Lanka during all seasons
(Haturusinghe and Weerakoon 2012; Ekanayaka et al. 2011; Campos-Arceiz et al. 2009;
Wikramanayake et al. 2004) however major crops such as rice are prime attractants and crop-raiding
often peaks as rice plants mature (Thaufeek et al. 2014; Haturusinghe and Weerakoon 2012; Fernando
et al. 2011). As with other areas experiencing high HEC throughout Asia, elephant crop-raiding
behaviour is almost entirely nocturnal (Campos-Arceiz et al. 2009; Wikramanayake et al. 2004;
Bandara and Tisdell 2003) and it is more common for male than female elephants to enter farmlands
to crop raid (Thaufeek et al. 2014; Haturusinghe and Weerakoon 2012; Ekanayaka et al. 2011; Bandara
and Tisdell 2002).
Most of the villages in close proximity to elephants are home to low-income farming families reliant
on annual or bi-annual harvest of rice crops for subsistence and income generation (Santiapillai et al.
2010; de Silva 1998). Ongoing crop-raiding activity increases desperation of communities to protect
their livelihoods, and people resort to more and more violent methods to deter elephants
(Haturusinghe and Weerakoon 2012; Santiapillai et al. 2010). Consequently, elephants respond more
aggressively to human presence, and a cycle of conflict ensues (Fernando et al. 2011). Despite
substantial efforts by conservationists and researchers to understand and mitigate crop-raiding in Sri
Lanka, it remains a prominent issue.

7
1.3 Characteristics and behaviour of elephants in high human-elephant conflict areas
The widespread problem of HEC has led to a great deal of research into the causes, consequences and
influencing predictors of elephant crop-raiding activity. However, for many locations experiencing
high levels of HEC, less is known about the general behaviour patterns (e.g. movement and foraging
habits, and responses to disturbance) of the overall local elephant population, especially in proximity
to farms and villages.
Knowledge of even relatively simple demographic characteristics can be hazy. For example, the best
available estimate of elephant abundance in Sri Lanka comes from a 2011 census conducted by the Sri
Lankan Department of Wildlife and Conservation (DWC). This census produced a population count of
5,879 wild elephants (Fernando et al. 2011; The Guardian 2011). However, it is likely population figures
have changed since then.
This lack of knowledge of precise elephant abundance near farms and villages clearly impedes wildlife
management planning. Only recently has a country-wide survey been completed revealing that
elephant range extends over almost 60% of Sri Lanka with people residing in almost 70% of this range
and conflict (especially involving male elephants) occurring almost everywhere humans and elephants
co-exist, (Fernando et al. 2019). This aligns with prior research identifying almost 65% of suitable
elephant habitat as existing outside of designated protected areas (PA’s) (Santiapillai et al. 2006) and
positing that the majority of elephants in Sri Lanka spent some or all of their time outside of PA’s
(Weerakoon et al. 2004).
In the absence of precise population figures and thus accurate estimates as to how many elephants
are involved in conflict, generating demographic and behavioural information of elephants utilising
habitat near farms and villages can provide valuable insight for predicting future crop-raiding activity
or even how elephants might react to deterrents or human presence.
1.3.1 Risk taking behaviour
Essentially, elephant crop-raiding is a high-risk, high-gain foraging strategy whereby elephants leave
the relative safety of PA's or forest habitat to enter farmlands and feed on crops. This foraging strategy

8
puts elephants in close proximity to humans. Given that people scare, injure or even kill elephants to
deter them from farms, interactions with humans present a substantial threat (Evans and Adams
2018). On the other hand, elephants may see proximity to human inhabited areas as a source of
opportunity, where nutritious fields of poorly guarded or unguarded crops are readily available (Evans
and Adams 2018).
Several behaviours of wild animals, including foraging strategies, are altered due to increasing human-
animal interactions, particularly conflict events (Fischhoff et al. 2008; Lacy and Martins 2003; Anthony
and Blumstein 2000; Olson et al. 1997). In three separate examples, lizards habituating to areas of
high human use showed more aggression than lizards in low human use areas (Lacy and Martins 2003),
Alaskan brown bears altered seasonal foraging patterns according to their degree of habituation to
humans (Olson et al. 1997), and zebra reduced use of grassland habitat when lions were observed in
the area on the same day (Fischhoff et al. 2008).
Crop-raiding, where animals make the decision to enter a potentially risky area to forage, is often
characterised by strong male-biased sex differences in frequency and intensity of raiding activity, with
males typically raiding more frequently, for longer durations, and causing more damage than females
(e.g. chimpanzees: Wilson et al. 2007; vervet monkeys: Saj et al. 1999). This is certainly true of
elephants. While crop-raiding is not exclusively a male activity, males display a much greater
propensity to exploit this foraging strategy then do females, even though all elephants within the
population could theoretically access crops (de Silva et al. 2013; Haturusinghe and Weerakoon 2012;
Fernando et al. 2011). Furthermore, in areas where both sexes do crop-raid, male elephants tend to
raid year-round while females limit crop-raiding activity to times when crops are nearing maturation
and to locations closest to forest boundaries (Williams et al. 2001).
In one of the few in-depth studies investigating the impacts of risk on Asian elephant behaviour,
Srinivassaiah et al. (2012) used areas of varying degrees of human disturbance to represent low to
high levels of risk, and monitored elephant behaviour patterns in each area. It was found that time
spent in-and outside of PA's, time spent feeding and specific behaviours all reflected the level of
perceived risk present. For example, elephants spent most of their time foraging or moving. However,
as the level of human disturbance (risk) became higher, time spent foraging decreased but time spent
moving increased as did vigilance behaviours. Adult male elephants spent more time in high risk areas
than any other elephants, demonstrating sex and age-based influences on risk-taking behaviour for
this population (Srinivassaiah et al. 2012).

9
Other studies relating to behaviour in the presence of risk have shown that elephants possess the
capacity to assess varying levels of risk and adjust their behaviour accordingly. African elephants used
olfactory and visual cues to distinguish between the perceived presence of men from the Masai tribe
(who represent a threat to elephants) and men from the Kamba tribe (who do not represent a threat
to elephants) and fled from the Masai stimuli (Bates et al. 2007). Similarly, Asian elephants exposed
to auditory cues of tiger growls (representing a real threat) with immediate silent retreat, and to
leopard growls (representing no real threat) with more investigative behaviours rather than avoidance
(Thuppil and Coss 2013).
With reference to elephant foraging behaviour in areas of high human presence, it could be that
females adjust their behaviour to the perceived level of risk by spending more time in PA's and
avoiding high-risk areas, whilst males fine-tune specific behaviours, moving faster across human-
dominated landscapes, entering farms under the cover of darkness, and selecting crop fields with the
least guarding effort present (Evans and Adams 2018; Graham et al. 2009).
Identifying behavioural differences of elephants in areas of varying degrees of human-elephant
interaction and potential influences of sex, age, and prior experience with humans can help to predict
the behaviour of individuals, or groups of individuals, across time and contexts (Freeman et al. 2013).
1.4 Human-elephant conflict mitigation
For as long as elephants have been raiding crops, farmers have been utilising a wide array of mitigation
methods with varying degrees of success. Since the 1990's, much research has focused on the
development, monitoring and assessment of elephant deterrent techniques (e.g. Africa: Hoare 2015;
Graham 2010; Graham and Ochieng 2008; Sitati and Walpole 2006; Dublin and Hoare 2004; Osborn
and Parker 2003; Asia: Gunaryadi et al. 2017; Thirunavukarasu 2014; Davies et al. 2011; Thapa 2010;
Perera 2009; Fernando et al. 2008; Wikramanayake et al. 2003; de Silva 1998; Santiapillai 1996).
Mitigation efforts can broadly be categorised as large-scale (e.g. methods implemented at
government level), traditional (e.g. methods that have long been used by individual farmers), or
community-based (e.g. methods implemented at a village level, often with the help of NGO's or other
research/conservation organisations). Most deterrents have been tried at numerous locations,
spanning both Africa and Asia (Hoare 2015; Fernando et al. 2008; Nelson et al. 2003). However, despite

10
a long history of tried and tested HEC mitigation methods, crop-raiding levels show no sign of abating.
What is becoming increasingly apparent is that there is no 'one size fits all' solution and that a
combination of deterrents based on site-specific knowledge of crop-raiding activity, the local elephant
population, and community perceptions may be the best approach (van de Water and Matteson 2018;
Pozo et al. 2017; Karidozo and Osborn 2015; Fernando et al. 2008).
Crop-raiding mitigation methods commonly implemented at a government level include confinement
of elephants into PA's through elephant drives, translocation of problem individuals, and large-scale
electric fencing (van Eden et al. 2016; Ponnusamy et al. 2016; Fernando et al. 2011; Santiapillai et al.
2006; Nelson et al. 2003). These methods are expensive and difficult to maintain and often do not
take into account the behaviour and movement patterns of elephants (Fernando et al. 2011; Perera
2009; Santiapillai 1996). Farmers are often encouraged to create 'elephant barriers' such as digging a
trench (Fernando et al. 2008) or planting unpalatable crops (Gross et al. 2017; 2016; Fernando et al.
2008; Nelson et al. 2003). Trenches have a high failure rate as they are generally either prohibitively
expensive or not strong enough to withstand elephant encroachment (Fernando et al. 2008). Planting
unpalatable crops has its merits and some success has been shown (Gross et al. 2017; 2016; Fernando
et al. 2008), however convincing farmers to change their cultivation habits is difficult and may not
always be economically viable (Fernando et al. 2008).
'Traditional' small-scale methods commonly employed by farmers include hanging tin cans from wire,
guarding crops from a tree hut or watch tower at night, making loud noises, shining bright flashlights,
and throwing firecrackers or fire sticks (sticks with an oil soaked rag set alight at one end) in the
direction of elephants (van de Water and Matteson 2018; Haturusinghe and Weerakoon 2012;
Fernando et al. 2011; Hedges and Gunayardi 2009; Perera 2009). Over time elephants habituate to, or
find means of overcoming, most of these methods (Fernando et al. 2011; Osborn and Parker 2003).
Many farmers struggle to protect their crops using only traditional methods, and over time may resort
to using increasingly harmful means to deter elephants. Some farmers set trap guns, plant live electric
wires, hide poison in crops, or use shot guns or explosives to injure or kill elephants (Haturusinghe and
Weerakoon 2012). As humans increase their aggressiveness towards elephants, elephants appear to
be adjusting their behaviour by reacting more aggressively towards humans. Thus, a spiral of HEC
escalation ensues (Fernando et al. 2011; Perera 2009).
Over the last two decades, a shift towards community-based mitigation encouraging farmers to use
non-violent methods has gained traction (van de Water and Matteson 2018; Changa et al. 2016;

11
Karidozo and Osborn 2015; Hedges and Gunaryadi 2009; Graham and Ochieng 2008; Sitati and
Walpole 2006). Ideally, community-based deterrents are relatively cheap, require little time to
establish and maintain, and encourage ownership and responsibility by the farmer (Changa et al. 2016;
Karidozo and Osborn 2015; King et al. 2011; 2009). Effectiveness of a community-based deterrent
depends not only the techniques ability to deter elephants but on how feasible it is for farmers to
implement and maintain, and their willingness to adopt the technique (King et al. 2011; 2009). Some
success has been demonstrated with initiatives using chilli based deterrents (Botswana: Pozo et al.
2017; India: Baishya et al. 2012; Sumatra: Hedges and Gunaryadi 2009; Tanzania: Changa et al. 2016),
bio-barriers of unpalatable crops (Nepal: Gross et al. 2017; Zambia: Gross et al. 2016), and beehive
fencing (Kenya: King et al. 2017; 2011; India: Nair and Jayson 2016) however further research and trials
are required to properly understand where and when these methods may be appropriate.
1.4.1 Beehive fencing
Beehive fencing is a relatively new community-based mitigation method which has shown promise in
Africa (King et al. 2017, 2011; Scheijen et al. 2018), and is gaining popularity not only for its properties
as an elephant deterrent but also due to recent worldwide concerns about the collapse of honey bee
colonies and the global push for promoting beekeeping and honey bee health (Meixner and Conte
2016). This novel solution came about in the early 2000's when researchers from conservation
organisation Save the Elephants (Kenya) discovered that African elephants avoided resting under trees
containing African honey bees (Apis mellifera scutellata) (Vollrath and Douglas-Hamilton 2002). This
was followed by a series of experiments where elephants were exposed to audio playback of bee
sounds and responded by rapidly retreating from the sounds, performing exploratory (e.g. smelling)
or risk avoidance (e.g. shaking heads to potentially keep bees away from sensitive areas) behaviours,
and producing rumble vocalisations (King et al. 2007). These rumble vocalisations were recorded and
played back to elephants who again responded with retreat and/or exploratory and avoidance
behaviours. This indicated that not only were elephants aware of the danger presented by the
perceived presence of bees, but that elephants could communicate this threat to conspecifics who
also responded accordingly (King et al. 2010).
Based on the concept that crop-raiding could be reduced by capitalising on the bee-avoidance
behaviour elephants exhibited during playback experiments, the 'beehive fence' was designed (King

12
et al. 2011; 2009). A beehive fence is a series of beehives hung from posts and connected to one
another by wire linking one beehive to the next, with beehives approximately 8 m apart (Figure. 1.2).
Elephants attempting to enter the area inside the beehive fence, will hit the connecting wire between
beehives, causing the attached beehives on either side to swing and, if occupied, disturb the bees
causing them to exit the hive and swarm at the elephants (King et al. 2011).
Figure 1.2: Example of a beehive fence: from Dr. Lucy King’s ‘Elephants and Bees’ research site in Sagalla, Kenya
© Kylie M. Butler
Today in Africa, beehive fencing is being utilised as an effective, relatively inexpensive, and low-
maintenance elephant deterrent, capable of deterring elephants from entering the fence perimeter
for up to 80% of approaches (King et al. 2017). In addition to providing protection of crops against
elephant depredation, farmers derive further benefits as honey and other bee products can provide
an additional income source or be kept for personal use, and honey bees provide pollination services
(King et al. 2017; 2011; 2009). Following on from initial success at multiple locations in Kenya (King et
al. 2011; 2009), results from a beehive fence trial in Tanzania using a linear fence to divide the park
boundary from nearby farmlands, also showed the potential of beehive fencing as a crop-raiding

13
mitigation tool (Scheijen et al. 2017). Additionally, beehives are being used in South Africa (Cook et al.
2017) and Gabon (Ngama et al. 2016) to protect valuable trees from elephant damage.
The success of beehive fencing in Africa, has led to questions about its potential applicability as an
elephant deterrent in Asia. Although there are many locations across the two continents with similar
crop-raiding characteristics, the species of elephant differs and more importantly, notable behavioural
differences exist between the African honey bee and Asian honeybee (Apis cerana cerana). Of most
concern is the difference in aggression, with African honey bees known to sting much more readily
and to swarm for longer distances when disturbed (Winston 1987), while Asian honey bees are
comparatively more placid (Punchihewa 1994). However, initial audio playback experiments testing
the response of wild elephants in Sri Lanka to the perceived presence of Asian honeybees, suggest
that avoidance responses are typical, although not as marked as in their African counterparts (King et
al. 2018).
To date, the only existing evidence of beehive fencing exhibiting success as an Asian elephant
deterrent is presented in two anecdotal newspaper reports from India (Dutta 2013; Manoj 2012), and
one three-month research trial, also in India where beehives were arranged in a short linear formation
blocking elephant pathways to fields from a nearby forest (Nair and Jayson 2016). Although this trial
showed promise, only 14 elephant approaches to the fence with two breaches were recorded and an
individual male elephant was responsible for ten of these approaches. Longer term studies over
multiple seasons are required to measure how the beehive fence performs over time and how
individual elephants respond to it, before it can be accurately assessed as a successful deterrent or
otherwise.
1.5 Rationale
Although extensive research exploring various facets of HEC from a multitude of sites exists, the vast
majority of studies focus on particulars: for example, crop-raiding patterns, or demographic
characteristics of the local population, or a specific mitigation technique. Even when multiple aspects
of the conflict situation are examined, knowledge of the behaviour patterns of the local elephant
population is often lacking. Information from these studies makes a valid contribution to the HEC body
of knowledge, however the importance of generating as complete a picture as possible of site-specific

14
conflict situations cannot be understated. This is particularly so when deciding on mitigation
techniques to implement or when recommending specific mitigation techniques based on success or
failure at a given location.
1.6 Thesis overview
In this study, I selected a region of Sri Lanka experiencing high levels of HEC in the form of crop-raiding
(see Chapters 2 and 3 for detailed site descriptions) and sought to provide an in-depth assessment of
the site-specific situation generated over a three-year period. The overarching objectives of this study
were:
a) To describe the general behaviour patterns of an elephant population in a high HEC
region of Sri Lanka, both inside and outside of the local protected area, Wasgamuwa
National Park (WNP)
b) To create a profile of the crop-raiding situation in a village near to WNP, including
identifying predictors of crop-raiding activity
c) To test the effectiveness of a new and potential HEC mitigation technique for Asia –
beehive fencing, and evaluate its overall potential as an Asian elephant deterrent
In Chapter 2, I use video footage of members of the local elephant population captured over two years
of observations to score general behaviour (e.g. feeding and small-scale movement patterns) and
specific actions (e.g. smelling and head-shaking) from a pre-determined ethogram. I compare these
behaviours and actions for elephants inside and outside of WNP, and also in the presence and absence
of known disturbances (e.g. approach of motorbike, tuk tuk or other vehicle). The potential influence
of sex and prior level of negative interaction with humans (using wounds/scars as a proxy) on
behaviour was also examined.
In Chapter 3, I move into the centre of the conflict zone: a small village on the outskirts of WNP called
Dewagiriya, which typically experiences high levels of crop-raiding year-round. Before implementing
mitigation techniques, it is vital to have a sense of the patterns and influencing factors of crop-raiding

15
activity, and this has been described for many crop-raiding sites in both Africa and Asia. Here, I use
farmer collected data and farmer interviews to determine seasonal, night/day, and lunar patterns of
crop-raiding, as well as typical group size and sex of crop-raiders (if known) and damage incurred. I
look at the potential influences of crop-type and farm proximity to forest habitat on susceptibility to
elephant encounters. These factors are commonly examined in crop-raiding studies and incorporating
them into this study facilitates meaningful comparison across sites. Additionally, I placed night-vision
camera traps in strategic locations throughout the village and in forest habitat on the periphery to
capture footage of elephants utilising these areas with the aim of identifying specific individuals, or at
least their gender and age class.
Finally, in Chapter 4, I present findings from a three-year beehive fence trial in Dewagiriya village,
assessing the effectiveness of beehive fences by comparing elephant events and entry into home
garden areas for experimental (beehive fence) and control (non-beehive fence) properties during the
same periods. Assessing deterrent effect is an important element of determining the success of any
mitigation technique however there are multiple factors essential to evaluating overall feasibility of a
deterrent tool, many of which are commonly overlooked in the literature. I incorporate not only the
deterrent effect of beehive fences but community perceptions of its usefulness and their willingness
to participate; cost and time analysis of set-up and maintenance; and benefits and challenges of
beekeeping, in an overall assessment of its potential as an Asian elephant deterrent tool in Sri Lanka.
Chapter 5 concludes this study by providing a discussion on our findings from the aforementioned
chapters, including the benefits of a multi-faceted approach that simultaneously investigates the
behaviour of the local elephant population, with in-depth analysis of crop-raiding patterns and
influencing factors, and a comprehensive assessment of a newly implemented mitigation technique.
We discuss the limitations and challenges of the study, provide recommendations as to the future
potential of beehive fencing as an Asian elephant deterrent, and suggest areas where further research
could be conducted.

16
1.7 References
Agar, RS n.d. 'Proclaimed elephants', IN: (eds: Ranasinghe, D, Ratnapala, S, Seneviratne, L,
Schwammer, H, Schwammer, G and Perera, BH) Saga of the elephant in Sri Lanka: Extracts from Loris
Volume 1 (1936) to Volume 10 (1964), Nedimala, YN Print Graphics, pp. 17 - 19.
Alahakoon, J & Santiapillai, C 1997, 'Elephants: unwitting victims in Sri Lanka's civil war', Gajah, vol.
18, pp. 63 – 65.
Amwata, DA & Mganga, KZ 2014, 'The African elephant and food security in Africa: experiences from
Baringo District, Kenya', Pachyderm, vol. 55, pp. 22 – 28.
Anthony, LL & Blumstein, DT 2000, 'Integrating behaviour into wildlife conservation: the multiple ways
that behaviour can reduce Ne', Biological Conservation, vol. 95, pp. 303 – 315.
Baishya, HK, Dey, S, Sarmah, A, Sharma, A, Gogoi, S, Aziz, T, Ghose, D & Williams, AC 2012, 'Use of chilli
fences to deter Asian elephants – a pilot study', Gajah, vol. 36, pp. 11 – 13.
Bandara, R & Tisdell, C 2003, 'Comparison of rural and urban attitudes to the conservation of Asian
elephants in Sri Lanka: empirical evidence', Biological Conservation, vol. 110, pp. 327 – 342.
Bandara, R & Tisdell, C 2002, 'Asian elephants as agricultural pests: economics of control and
compensation in Sri Lanka', Natural Resources Journal, vol. 42, pp. 491 – 519.
Barnes, RFW, Danquah, E, Hema, EM, Dubiure, UF, Manford, M, Nandjui, A & Boafo, Y 2015,
'Retrospective versus prospective designs for studies of crop-raiding by elephants in Kakum, Ghana',
Pachyderm, vol. 56, pp. 44 – 50.
Barua, M, Bhagwat, SA & Jadhav, S 2013, 'The hidden dimensions of human-wildlife conflict: health
impacts, opportunity and transaction costs', Biological Conservation, vol. 157, pp. 309 – 316.
Baruch-Mordo, S, Breck, SW, Wilson, KR & Theobald, DM 2008, 'Spatiotemporal distribution of black
bear-human conflicts in Colorado, USA', Journal of Wildlife Management, vol. 72, no. 8, pp. 1853 –
1862.

17
Bates, LA, Sayialel, KN, Njiraini, NW, Moss, CJ, Poole, JH and Byrne, RW 2007, 'Elephants classify human
ethnic groups by odor and garment colour', Current Biology, vol. 17, no. 22,
doi:10.1016/j.cub.2007.09.060.
Burn, RW, Underwood, FM & Blanc, J 2011, 'Global trends and factors associated with the illegal killing
of elephants: a hierarchical Bayesian analysis of carcass encounter data', PLoS ONE, vol. 6, no. 9,
e24165. doi:10.1371/journal.pone.0024165.
Campos-Arceiz, A, Takatsuki, S, Ekanayaka, SKK & Hasegawa, T 2009, 'The human-elephant conflict in
southeastern Sri Lanka: type of damage, seasonal patterns, and sexual differences in the raiding
behaviour of elephants', Gajah, vol. 31, pp. 5 – 14.
Chakraborty, K & Mondal, J 2013, 'Perceptions and patterns of human-elephant conflict at Barjora
block of Bankura district in West Bengal, India: insights for mitigation and management',
Environmental Development and Sustainability, vol. 15, pp. 547 – 565.
Changa, A, de Souza, N, Muya, J, Keyyu, J, Mwakatobe, A, Malugu, L, Ndossi, HP, Konuche, J, Omondi,
R, Mpinge, A, Hahn, N, Palminteri, S and Olson, D 2016, 'Scaling-up the use of chilli fences for reducing
human-elephant conflict across landscapes in Tanzania', Tropical Conservation Science, vol. 9, no. 2,
pp. 921 – 930.
Chiyo, PI, Cochrane, EP, Naughton, L & Basuta, GI 2005, 'Temporal patterns of crop raiding by
elephants: a response to changes in forage quality or crop availability?', African Journal of Ecology,
vol. 43, pp. 48 – 55.
Choudhury, A 2004, 'Human-elephant conflicts in northeast India', Human Dimensions of Wildlife, vol.
9, no. 4, pp. 261 – 270.
Cook, RM, Witkowski, ETF, Helm, CV, Henley, MD & Parrini, F 2017, 'Recent exposure to African
elephants after a century of exclusion: rapid accumulation of marula tree impact and mortality, and
poor regeneration', Forest Ecology and Management, vol. 401, pp. 107 – 116.

18
Das, BJ, Saikia, BN, Baruah, KK, Bora, A & Bora, M 2014, 'Nutritional evaluation of fodder, its preference
and crop-raiding by wild Asian elephant (Elephas maximus) in Sonitpur District of Assam, India',
Veterinary World, vol. 7(12), pp. 1082-1089.
Davies, TE, Wilson, S, Hazarika, N, Chakrabaty, J, Das, D, Hodgson, DJ & Zimmerman, A
2011,'Effectiveness of intervention methods against crop-raiding elephants', Conservation Letters,
doi: 10.1111/j.1755-263X.2011.00182.x
de Silva, M 1998, 'Status and conservation of the elephant (Elephas maximus) and the alleviation of
man-elephant conflict in Sri Lanka', Gajah, vol. 19, pp. 1 – 68.
de Silva, S, Webber, CE, Weerathugna, US, Pushpakumara, TV, Weerakoon, DK and Wittemyer, G 2013,
'Demographic variables for wild Asian elephants using longitudinal observations', PLoS ONE, vol. 8, no.
12, e82788. doi: 10.1371/journal.pone.0082788.
Dickman, AJ 2010, 'Complexities of conflict: the importance of considering social factors for effectively
resolving human-wildlife conflict', Animal Conservation, vol. 13, pp. 458 - 466.
Dublin, HT & Hoare, RE 2004, 'Searching for solutions: the evolution of an integrated approach to
understanding and mitigating human-elephant conflict in Africa', Human Dimensions of Wildlife, vol.
9, no. 4, pp. 271 – 278.
Dutta, A 2013, 'Bees, acrid smoke from chillis keep elephants away and reduce conflict', The Times of
India, June 24, 2013.
Ekanayaka, SKK, Campos-Arceiz, A, Rupasinghe, M, Pastorini, J & Fernando, P 2011, 'Patterns of crop
raiding by Asian elephants in a human-dominated landscape in southeastern Sri Lanka', Gajah, vol. 34,
pp. 20 -25.
Evans, LA and Adams, WM 2018, 'Elephants as actors in the political ecology of human-elephant
conflict', Transactions of the Institute of British Geographers, doi: 10.1111/tran.12242.

19
Fernando, P, de Silva, MKCR, Jayasinghe, LKA, Janaka, HK & Pastorini, J 2019, ‘First country-wide survey
of the endangered Asian elephant: toward better conservation and management in Sri Lanka’, Oryx,
pp. 1 – 10, https://doi.org/10.1017/s0030605318001254 .
Fernando, P, Jayewardene, J, Prasad, T, Hendavitharana, W & Pastorini, J 2011, ‘Current status of Asian
Elephants in Sri Lanka’, Gajah, vol. 25, pp. 93 – 103.
Fernando, P, Kumar, MA, Williams, AC, Wikramanayake, E, Aziz, T & Singh, SM 2008, Review of human-
elephant conflict mitigation measures practiced in South Asia, AREAS Technical Support Document,
World Wide Fund for Nature.
Fernando, P, Wikramanayake, E, Weerakoon, D, Jayasinghe, LKA, Gunawardene, M & Janaka, HK 2005,
'Perceptions and patterns of human-elephant conflict in old and new settlements in Sri Lanka: insights
for conservation and management', Biodiversity and Conservation, vol. 14, pp. 2465 – 2481.
Fischhoff, IR, Sundaresan, SR, Cordingley, J and Rubenstein, DI 2007, 'Habitat use and movements of
plains zebra (Equus burchelli) in response to predation danger from lions', Behavioural Ecology, doi:
10.1093/beheco/arm036.
Freeman, HD, Brosnan, SF, Hopper, LM, Lambeth, SP, Schapiro, SJ & Gosling, SD 2013, 'Developing a
comprehensive and comparative questionnaire for measuring personality in chimpanzees using a
simultaneous top-down/bottom-up design', American Journal of Primatology, vol. 75, pp. 1042 –
1053.
Gillingham, S & Lee, PC 2003, 'People and protected areas: a study of local perceptions of wildlife crop-
damage conflict in an area bordering the Selous Game Reserve, Tanzania', Oryx, vol. 37, pp. 316 – 325.
Graham, MD, Notter, B, Adams, WM, Lee, PC & Ochieng, TN 2010, 'Patterns of crop-raiding by
elephants, Loxodonta africana, in Laikipia, Kenya, and the management of human-elephant conflict',
Systematics and Biodiversity, vol. 8, no. 4, pp. 435 – 445.
Graham, MD, Douglas-Hamilton, I, Adams, WM, and Lee, PC 2009, 'The movement of African
elephants in a human-dominated land-use mosaic', Animal Conservation, vol. 12, pp. 445 – 455.

20
Graham, MD & Ochieng, T 2008, 'Uptake and performance of farm-based measures for reducing crop-
raiding by elephants Loxodonta Africana among smallholder farms in Laikipia District, Kenya', Oryx,
vol. 42, pp. 76 – 82.
Gross, EM, Drouet-Hoguet, N, Subedi, N & Gross, J 2017, 'The potential of medicinal and aromatic
plants (MAPs) to reduce crop damages by Asian elephants (Elephas maximus)', Crop Protection, vol.
100, pp. 29 – 37.
Gross, EM, Mc Robb, R and Gross, J 2016, 'Cultivating alternative crops reduces crop losses due to
African elephants' Journal of Pest Science, vol. 89, no. 2, pp. 495 – 506.
Gubbi, S 2012, 'Patterns and correlates of human-elephant conflict around a south Indian reserve',
Biological Conservation, vol. 148, pp. 88 – 95.
Gunaryadi, D, Sugiyo, Hedges, S 2017, 'Community-based human-elephant conflict mitigation: the
value of an evidence-based approach in promoting the uptake of effective methods', PLoS ONE, vol.
12, no. 5, e0173742.
Harich, FK, Treydte, AC, Sauerborn, J & Owusu, EH 2013, 'People and wildlife: conflicts arising around
the Bia Conservation Area in Ghana', Journal for Nature Conservation, vol. 21, pp. 342 – 349.
Hartter, J, Goldman, A & Southworth, J 2011, 'Responses by households to resource scarcity and
human-wildlife conflict: issues of fortress conservation and the surrounding agricultural landscape',
Journal for Nature Conservation, vol. 19, pp. 79 – 86.
Haturusinghe, HS & Weerakoon, DK 2012, ‘Crop raiding behaviour of elephants in the Northwestern
region of Sri Lanka’, Gajah, vol. 36, pp. 26 – 31.
Hedges, S & Gunaryadi, D 2009, 'Reducing human-elephant conflict: do chillies help deter elephants
from entering crop fields?', Oryx, vol. 44, no. 1, pp. 139 – 146.
Hennessey, DJG n.d, 'I watch in a chena', IN: (eds: Ranasinghe, D, Ratnapala, S, Seneviratne, L,
Schwammer, H, Schwammer, G and Perera, BH) Saga of the elephant in Sri Lanka: Extracts from Loris
Volume 1 (1936) to Volume 10 (1964), Nedimala, YN Print Graphics, pp. 51 – 55.

21
Hennessey, DJG 1929, 'Elephant shikar', IN: (eds: Ranasinghe, D, Ratnapala, S, Seneviratne, L,
Schwammer, H, Schwammer, G and Perera, BH) Saga of the elephant in Sri Lanka: Extracts from Loris
Volume 1 (1936) to Volume 10 (1964), Nedimala, YN Print Graphics, pp. 9 -13.
Hill, CM 2004, 'Farmers' perspectives of conflict at the wildlife-agricultural boundary: some lessons
learned from African subsistence farmers', Human Dimensions of Wildlife, vol. 9, pp. 279 – 286.
Hill, CM 2000, 'Conflict of interest between people and baboons: crop raiding in Uganda', International
Journal of Primatology, vol. 21, no. 2, pp. 299 – 315.
Hoare, R 2015, 'Lessons from 20 years of human-elephant conflict mitigation in Africa', Human
Dimensions of Wildlife, vol. 20, pp. 289 – 295.
Hoare, R 2000, 'African elephants and humans in conflict: the outlook for co-existence', Oryx, vol. 34,
pp. 34 – 38.
Hoffmeier-Karimi, RR & Schulte, BA 2014, 'Assessing perceived and documented crop damage in a
Tanzanian village impacted by human-elephant conflict (HEC)', Pachyderm, vol. 56, pp. 51 – 60.
Ihwagi, F, Thouless, C, Wang, T, Skidmore, AJ, Omondi, P & Douglas-Hamilton, I 2018, 'Night-day speed
ratio of elephants as indicator of poaching levels', Ecological Indicators, vol. 84, pp. 38 – 44.
Kahindi, O, Wittemyer, G, King, J, Ihwagi, F, Omondi, P & Douglas-Hamilton, I 2009, 'Employing
participatory surveys to monitor the illegal killing of elephants across diverse land uses in Laikipia-
Samburu, Kenya', African Journal of Ecology, doi: 10.1111/j.1365-2028.2009.01.200.x.
Karidozo, M & Osborn, FV 2015, 'Community based conflict mitigation trials: results of field tests of
chilli as an elephant deterrent', Biodiversity and Endangered Species, vol. 3 no. 1,
http://dx.doi.org/10.4172/2332-2543.1000144.
King, L, Pardo, M, Weerathunga, S, Kumara, TV, Jayasena, N, Soltis, J & de Silva, S 2018, ‘Wild Sri Lankan
elephants retreat from the sound of disturbed Asian honey bees’, Current Biology, vol. 28, pp 64 – 65.

22
King, LE, Lala, F, Nzumu, H, Mwambingu, E & Douglas-Hamilton, I 2017, 'Beehive fences as a
multidimensional conflict-mitigation tool for farmers coexisting with elephants', Conservation Practice
and Policy, vol. 31, no. 4, pp. 743 – 752.
King, LE, Douglas-Hamilton, I & Vollrath, F 2011, 'Beehive fences as effective deterrents for crop-
raiding elephants: field trials in northern Kenya', African Journal of Ecology, doi: 10.1111/j.1365-
2028.2011.01275.x.
King, LE, Soltis, J, Douglas-Hamilton, I, Savage, A & Vollrath, F 2010, 'Bee threat elicits alarm call in
African elephants', PLoS ONE, vol. 5, no. 4, e10346. doi:10.1371/journal.pone0010346.
King, LE, Lawrence, A, Douglas-Hamilton, I & Vollrath, F 2009, 'Beehive fence deters crop-raiding
elephants', African Journal of Ecology, vol. 47, pp. 131 – 137.
King, LE, Douglas-Hamilton, I & Vollrath, F 2007, 'African elephants run from the sounds of disturbed
bees', Current Biology, vol. 17, no. 19, R832.
Lacy, KE & Martins, EP 2003, 'The effect of anthropogenic habitat usage on the social behaviour of a
vulnerable species, Cyclura nubila', Animal Conservation, vol. 6, pp. 3 – 9.
Lewis, DL, Baruch-Mordo, S, Wilson, KR, Breck, SW, Mao, JS & Broderick, J 2015, 'Foraging ecology of
black bears in urban environments: guidance for human-bear conflict mitigation', Ecosphere, vol. 6,
no. 8, http://dx.doi.org/10.1890/ES15-00137.1.
Loe, J & Roskaft, E 2004, 'Large carnivores and human safety: a review', Ambio, vol. 6, pp. 283 – 288.
Lord, A, Waas, JR & Innes, J 1997, 'Effects of human activity on the behaviour of northern New Zealand
dotterel Charadrius obscurus aquilonius chicks', Biological Conservation, vol. 82, pp. 15 – 20.
Madden, F 2004, 'Creating coexistence between humans and wildlife: global perspectives on local
efforts to address human-wildlife conflict', Human Dimensions of Wildlife, vol. 9, no. 4, pp. 247 – 257.
Manfredo, MJ 2015, 'Essays on human-wildlife conflict 10 years after the Durban World Parks
Congress: an introduction', Human Dimensions of Wildlife, vol. 20, pp. 285 – 288.

23
Manoj, EM 2012, 'Jumbos get a sting of defeat', The Hindu Newspaper, February 25, 2012.
Meixner, MD and Conte, YL 2016, 'A current perspective on honey bee health', Apidologie, vol. 47, pp.
273 – 275.
Menard, N, Foulquier, A, Vallet, D, Qarro, M, Le Gouar, P & Pierre, JS 2013, 'How tourism and
pastoralism influence population demographic changes in a threatened large mammal species',
Animal Conservation, vol. 17, pp. 115-124.
Nair, RP and Jayson, EA 2016, 'Effectiveness of beehive fences to deter crop raiding elephants in
Kerala, India', International Research Journal of Natural and Applied Sciences, vol. 3, no. 1, pp. 14 – 19.
Naughton-Treves, L & Treves, A 2005, 'Socio-ecological factors shaping local support for wildlife: crop-
raiding by elephants and other wildlife in Africa', IN: (eds: Woodroffe R, Thirgood S and Rabinowitz A)
People and wildlife: conflict or coexistence? Cambridge, Cambridge University Press, pp. 252 – 277.
Nelson, A, Bidwell, P and Sillero-Zubiri 2003, 'A review of human-elephant conflict mitigation
strategies', People and Wildlife Initiative, Wildlife Conservation Research Unit, Oxford University.
Ngama, S, Korte, L, Bindelle, J, Vermeulen, C & Poulsen, JR 2016, 'How bees deter elephants: beehive
trials with forest elephants (Loxodonta africana cyclotis) in Gabon', PLoS ONE, vol. 11, no. 5, e0155690.
doi:10.1371/journal.pone.0155690.
Nijman, V & Nekaris, KA 2010, 'Testing a model for predicting primate crop-raiding using crop-and-
farm specific risk values', Applied Animal Behaviour Science, vol. 127, pp. 125 – 129.
Ogra, MV 2008, 'Human-wildlife conflict and gender in protected area borderlands: a case study of
costs, perceptions and vulnerabilities from Uttarakhand (Uttaranchal), India', Geoforum, vol. 39, pp.
1408 – 1422.
Okello, MM, Kiringe, JW, Njumbi, SJ and Isiche, J 2016, 'Prevalence of human-elephant conflicts in
Amboseli ecosystem, Kenya: current opinions of local communiy', International Journal of Biodiversity
and Conservation, vol. 8, no. 3, pp. 60 – 71.

24
Olson, TL, Gilbert, BK & Squibb, RC 1997, 'The effects of increasing human activity on brown bear use
of an Alaskan river', Biological Conservation, vol. 82, pp. 95 – 99.
Osborn, FV & Parker, GE 2003, 'Towards an integrated approach for reducing the conflict between
elephants and people: a review of current research', Oryx, vol. 37, no. 1, doi.
10.1017/90030605302000000.
Perera, BMAO 2009, 'The human-elephant conflict: a review of current status and mitigation
methods', Gajah, vol. 1, pp. 341 – 344.
Perez, E & Pacheco, LF 2006, 'Damage by large mammals to subsistence crops within a protected area
in a montane forest of Bolivia', Crop Protection, vol. 25, pp. 933 – 939.
Ponnusamy, V, Chackrapani, P, Lim, TW, Saaban, S and Campos-Arceiz, A 2016, 'Farmers' perceptions
and attitudes towards government constructed electric fences in Peninsular Malaysia', Gajah, vol. 45,
pp. 4 – 11.
Pozo, RA, Coulson, T, McCulloch, G, Strongza, A & Songhurst, A 2017, 'Chilli-briquettes modify the
temporal behaviours of elephants, but not their numbers', Oryx, doi: 10.1017/S0030605317001235.
Punchihewa, RWK 1994, Beekeeping for honey production, Sarvodaya Visha Lekha Publishing, Sri
Lanka.
Redpath, SM, Young, J, Evely, A, Adams, WM, Sutherland, WJ, Whitehouse, A, Amar, A, Lambert, RA,
Linnell, JDC, Watt, A & Gutierrez, RJ 2013, 'Understanding and managing conservation conflicts',
Trends in Ecology and Evolution, vol. 28, no. 2, pp. 100 – 109.
Rao, KS, Maikhuri, RK, Nautiyal, S & Saxena, KG 2002, 'Crop damage and livestock depredation by
wildlife: a case study from Nanda Devi Biosphere Reserve, India', Journal of Environmental
Management, vol. 66, pp. 317 – 327.
Ripple, WJ, Abernethy, K, Betts, MG, Chapron, G, Dirzo, R, Galetti, M, Levi, T, Lindsey, PA, Macdonald,
DW, Machovina, B, Newsome, TM, Peres, CA, Wallach, AD, Wolf, C & Young, H 2016, 'Bushmeat

25
hunting and extinction risk to the world's mammals', Royal Society of Open Science, vol. 3, pp. 160498.
http://dx.doi.org/10.1098/rsos.160498.
Saj, T, Sicotte, P & Paterson, JD 1999, 'Influence of food consumption on the time budget of vervets',
International Journal of Primatology, vol. 20, pp. 977 – 994.
Santiapillai, C, Wijeyamohan, S, Bandara, G, Athurupana, R, Dissanayake, N & Read, B 2010, 'An
assessment of the human-elephant conflict in Sri Lanka', Ceylon Journal of Biological Sciences, vol. 39,
no. 1, pp. 21 – 33.
Santiapillai, C, Fernando, P and Gunewardene, M 2006, 'A strategy for the conservation of the Asian
elephant in Sri Lanka', Gajah, vol. 25, pp. 91 – 102.
Santiapillai, C 1996, 'Mitigation of human-elephant conflicts in Sri Lanka', Gajah, vol. 15, pp. 1 – 7.
Santiapillai, C & de Silva, M 1994, 'An action plan for the conservation and management of elephant
(Elephas maximus) in Sri Lanka', Gajah, vol. 13, pp. 1 – 24.
Saraswat, R, SInha, A & Radhakrishna, S 2015, 'A god becomes a pest? Human-rhesus macaque
interactions in Himachal Pradesh, northern India', European Journal of Wildlife Research, vol. 61, pp.
435 – 443.
Scheijen, CPJ, Richards, SA, Smit, J, Jones, T & Nowak, K 2018, 'Efficacy of beehive fences as barriers
to African elephants: a case study in Tanzania', Oryx, doi: 10.1017/S0030605317001727.
Sitati, NW & Walpole, MJ 2006, 'Assessing farm-based measures for mitigation of human-elephant
conflict in Transmara District, Kenya', Oryx, vol. 40, no. 3, pp. 279 – 286.
Sitati, NW, Walpole, MJ, Smith, RJ & Leader-Williams, N 2003, 'Predicting spatial aspects of human-
elephant conflict', Journal of Applied Ecology, vol. 40, pp. 667 – 677.
Srinivasaiah, NM, Anand, VD, Vaidyanathan, S & Sinha, A 2012, 'Usual populations, unusual
individuals: insights into the behaviour and management of Asian elephants in fragmented
landscapes', PLoS ONE, vol. 7, no. 8, e42571. doi: 10.1371/journal.pone.0042571.

26
Studsrod, JE & Wegge, P 1995, 'Park-people relationships: the case of damage caused by park animals
around the Royal Bardia National Park, Nepal', Environmental Conservation, vol. 22, no. 2, pp. 132 –
142.
Sukumar, R 2006, 'A brief review of the status, distribution and biology of wild Asian elephants Elephas
maximus', International Zoo Yearbook, vol. 40, pp. 1 – 8.
Sukumar, R 1990, 'Ecology of the Asian elephant in southern India. II. Feeding habits and crop raiding
patterns', Journal of Tropical Ecology, vol. 6, pp. 33 – 53.
Thapa, S 2010, 'Effectiveness of crop protection methods against wildlife damage: a case study of two
villages at Bardia National Park, Nepal', Crop Protection, vol. 29, pp. 1297 – 1304.
Thaufeek, UL, Padmalal, UKGK and Fernando, P 2014, 'Land use and human-elephant conflict in the
Sigiriya Sanctuary, Sri Lanka', Gajah, vol. 40, pp. 26 – 30.
The Guardian, 2011, 'Sri Lanka elephant survey puts population at 5,879', The Guardian (online) 2
September, https://www.theguardian.com/world/2011/sep/02/sri-lanka-elephant-survey-
population (Accessed 24 September 2018).
Thirgood, S, Woodroffe, R and Rabinowitz, A 2005, 'The impact of human-wildlife conflict on human
lives and livelihoods', IN: (eds: Woodroffe R, Thirgood S and Rabinowitz A) People and wildlife: conflict
or coexistence? Cambridge, Cambridge University Press, pp. 13 – 26.
Thirunavukarasu, V 2014, Assessment of human elephant conflicts and mitigation measures to resolve
the conflict in Dharamapuri Forest Division, Tamil Nadu, India, MCT Phase IV Assignment, District
Forest Office, Tamil Nadu, India.
Thuppil, V and Coss RG 2013, 'Wild Asian elephants distinguish aggressive tiger and leopard growls
according to perceived danger', Biology Letters, Vol. 9, 20130518.
http://dx.doi.org/10.1098/rsbl.2013.0518.

27
Tutein-Nolethnius, AC n.d. 'Is our emblem doomed?' IN: (eds: Ranasinghe, D, Ratnapala, S,
Seneviratne, L, Schwammer, H, Schwammer, G and Perera, BH) Saga of the elephant in Sri Lanka:
Extracts from Loris Volume 1 (1936) to Volume 10 (1964), Nedimala, YN Print Graphics, pp. 20 – 21.
van de Water, A and Matteson, K 2018, 'Human-elephant conflict in western Thailand: socio-economic
drivers and potential mitigation strategies', PLoS ONE, vol. 13, no. 6, e0194736.
Van Eden, M, Ellis, E and Bruyere, BL 2016, 'The influence of human-elephant conflict on electric fence
management and perception among different rural communities in Laikipia County, Kenya', Human
Dimensions of Wildlife, vol. 21, no. 4, pp. 283 – 296.
Vollrath, F & Douglas-Hamilton, I 2002, 'African bees to control African elephants',
Naturwissenschaften, vol. 89, pp. 508 – 511.
von Gerhardt, K, Niekerk, AV, Kidd, M, Samways, M & Hanks, J 2014, 'The role of elephant Loxodonta
africana pathways as a spatial variable in crop-raiding location', Oryx, vol. 48(3), pp. 436 – 444.
Webber, CE, Sereivathana, T, Maltby, MP and Lee, PC 2011, 'Elephant crop-raiding and human-
elephant conflict in Cambodia: crop selection and seasonal timing of raids', Oryx, vol. 45, no. 2, pp.
243 – 251.
Weerakoon, DK, Gunawardene, MD, Janaka, HK, Jayasinghe, LKA, Perera, RAR, Fernando, P and
Wikramanayake, E 2004, 'Ranging behaviour and habitat use of elephants in Sri Lanka', in: (ed)
Jayewardene, J Endangered Elephants: Past, present and future, Proceedings of the symposium for
human-elephant relationships and conflicts, Sri Lanka, September 2003. Biodiversity and Conservation
Trust, Colombo, pp. 68 - 70.
Wikramanayake, ED, Hathurusinghe, HS, Janaka, HK, Jayasinghe, LKA, Fernando, P, Weerakoon, DK &
Gunawardene, MD 2004, 'The human-elephant conflict in Sri Lanka: lessons for mitigation,
management, and conservation from traditional land-use patterns', in: (ed) Jayewardene, J
Endangered Elephants: Past, present and future, Proceedings of the symposium for human-elephant
relationships and conflicts, Sri Lanka, September 2003. Biodiversity and Conservation Trust, Colombo,
pp. 164-169.

28
Williams, AC, Johnsingh, AJT & Krausman, PR 2001, 'Elephant-human conflicts in Rajaji National Park,
north-western India', Wildlife Society Bulletin, vol. 29, no. 4, pp. 1097 – 1104.
Wilson, ML, Hauser, MD & Wrangham, RW 2007, 'Chimpanzees (Pan troglodytes) modify grouping and
vocal behaviour in response to location-specific risk', Behaviour, vol. 144, pp. 1621 – 1653.
Winston, ML 1987, The biology of the honey bee, Harvard University Press 2nd Edition, USA, pp. 4 – 12.
Woodroffe, R, Lindsey, PA, Romanach, SS, Stein, A, Ranah, SMKO 2005a, 'Livestock predation by
endangered African wild dogs (Lycaon pictus) in northern Kenya', Biological Conservation, vol. 124, pp.
225 – 234.
Woodroffe, R, Thirgood, S & Rabinowitz, A (Eds) 2005b, People and wildlife: conflict or coexistence?
Cambridge University Press, Cambridge.

29
Chapter 2: General behaviour patterns of wild elephants utilising areas of varying anthropogenic risk to elephants, in Sri Lanka

30
2.1 Abstract
Many species alter their behaviour in response to perceived levels of anthropogenic disturbance, or
risk, in a given environment. Risk-avoidance responses such as fleeing are typical, and often have
fitness-consequences as the time available for important activities such as feeding, mating or resting
is reduced. In this study, the behaviour of Asian elephants observed in areas representing a low or
medium level of anthropogenic risk was investigated. The results provided no evidence that general
behavioural patterns including time spent in locomotion, feeding or standing still differed between
elephants observed inside National Park boundaries (low-risk zone) or in nearby external forest
(medium-risk zone). The frequency with which elephants performed typical ‘reactive’ behaviours,
such as smelling or dusting, was also similar. However, when presented with an immediate
anthropogenic disturbance, such as an approaching vehicle, elephants would reduce the time spent
feeding and increase the frequency of ‘reactive’ behaviours regardless of whether they were observed
in the low- or medium risk zone. Overall, this study provides initial evidence that certain human-
influenced stimuli are perceived as a disturbance or threat by elephants, while also highlighting the
challenging nature of investigating elephant response to anthropogenic disturbance in the wild.
2.2 Introduction
A by-product of worldwide human population growth and the correlating infrastructure and
agricultural development is a blurring of the delineation between human and wildlife habitats
(Fernando et al. 2011; Graham et al. 2009). Inevitably, this results in an increase of interactions
between the species, which is influencing a habitat of conflict between human and wildlife
communities (Lacy and Martins 2003). Anthropogenically induced alterations reducing the size and
quality of wildlife habitat, drive competition for resources both within and between species and force
many non-human animals to adjust to unprecedented levels of human activity (Lacy and Martins
2003). Termed ‘anthropogenic disturbance’ these interactions challenge the health and long-term
viability of numerous wildlife species (Setsaas et al. 2007; Frid and Dill 2002; Lima 1998).
Anthropogenic disturbance often leads to situations or environments of perceived or actual riskiness
to animals (Lacy and Martins 2003; Frid and Dill 2002; Anthony and Blumstein 2000; Mattson et al.
1992), and many species alter their behaviour to attend to varying risks in their environment (Lacy and

31
Martins 2003; Frid and Dill 2002). For species which are hunted, captured or otherwise injured by
humans, the risks associated with anthropogenic disturbance are particularly acute (Caro 2005; Frid
and Dill 2002). In such situations, animals often respond in a manner akin to responses provoked by
the threat of predation, of which risk-avoidance or minimisation strategies are common (Frid and Dill
2002; Steidl and Anthony 2000). However, the obvious benefits of increased safety assumed by risk-
avoidance, often incur costs to fitness including less time available for feeding, resting or mating
(Blumstein et al. 2005; Lacy and Martins 2003; Frid and Dill 2002; Anthony and Blumstein 2000; Lima
1998), excess energy expended while guarding or fleeing (Kiffner et al. 2014; Frid and Dill 2002), or
reduced access to optimal habitat (Blumstein et al. 2005; Frid and Dill 2002; de la Torre et al. 2000;
Lima 1998; Lima and Dill 1990). Such compromises to vital activities can have both short- and long-
term negative consequences on body condition, reproductive success and even survival for individual
animals (White et al. 2002; Lima 1998; Lima and Dill 1990).
Identifying the ways in which wild animals respond to anthropogenic disturbance is fraught with
difficulty. Behavioural change can often happen slowly, and by the time it is suspected it is often too
late to acquire a baseline of formerly typical behaviour by which to compare. Researchers have
addressed this challenge by comparing the behaviour of wildlife thought to be, or at risk of being,
affected by anthropogenic disturbance at locations representing varying levels of human activity. This
can include comparisons of areas of low to high hunting pressure (e.g. de Boer et al. 2004), areas
representing varying levels of non-lethal human recreational activity (e.g. Lusseau et al. 2009; White
et al. 1999), or inside and outside of designated wildlife protected areas (PA’s) (Crosmary et al. 2012).
Such studies provide numerous examples of behavioural differences according to perceived level of
anthropogenic risk, across a wide array of species (e.g. Crosmary et al. 2012; Lusseau et al. 2009;
Griffin et al. 2007; Setsaas et al. 2007; Donadio and Buskirk 2006; Williams et al. 2006; Caro 2005;
Lusseau 2003; White et al. 1999).
For example, recent studies show that heavily exploited mammals in Tanzania (including buffalo,
giraffe, roan antelope, topi, waterbuck, warthog and zebra) are more likely to flee or increase vigilance
in the presence of human observers outside of PA’s (Caro 2005). Similar avoidance behaviours were
exhibited in marine mammals who modified diving patterns to spend more time deeper underwater,
in response to the presence of tourist vessels (e.g. dolphins: Lusseau 2003; and killer whales: Lusseau
et al. 2009; Williams et al. 2006). For dolphins specifically, this change in locomotive patterns

32
interrupts important foraging opportunities (Lusseau 2003). Similarly, in the United States of America
(USA), grizzly bears increase time spent moving, thus reducing available foraging time, in areas where
human recreational activities such as mountain climbing are popular (White et al. 1999). This inverse
relationship between movement and feeding is also exhibited in camelid species inhabiting reserves
in Argentina where poaching frequently occurs (Donadio and Buskirk 2006). Other species, while not
necessarily altering locomotive patterns, exhibit increased vigilance behaviours, which can also
interrupt foraging. Heightened vigilance levels have been observed in roe deer occupying areas of
frequent hunting in the Netherlands (de Boer et al. 2004), in several species of ungulates utilising
habitat outside of PA’s in Zimbabwe (Crosmary et al. 2012) and Tanzania (Setsaas et al. 2007), and by
Olympic marmots in areas of high human recreation activity in the USA (Griffin et al. 2007).
As highlighted above, many species demonstrate a capacity to recognise variations in risk between
areas representing different levels of anthropogenic disturbance. Additionally, many species can also
differentiate between fluctuations in anthropogenic risk within a given location (Ferrari et al. 2009;
Bates et al. 2007), particularly if the risk is heightened by the physical presence of humans rather than
just anthropogenically induced changes to the ecosystem. Fluctuations in risk can occur daily (e.g. the
physical presence of a predator, or increasing visitor proximity to self), or seasonally (e.g. consistent
increased human presence during peak tourist holidays or legal hunting seasons) (Lima and Dill 1990).
An ability to discriminate between risk levels enables an animal to respond appropriately to the
specific situation, rather than expending excess energy in a constant state of high alert (Ferrari et al.
2009; Bates et al. 2007; Lima and Bednekoff 1999). For example, European roe deer increase vigilance
behaviours during assigned hunting seasons, then relax these behaviours when hunting ceases
(Behhaiem et al. 2008). Similarly, Alaskan brown bears who were not yet habituated to human
presence, would delay salmon feeding if the tourist season was extended, despite high availability of
food (Olson et al. 1997).
While changes in locomotion, foraging and vigilance are common responses to anthropogenic
disturbance, they are not necessarily uniform within individuals of the species present, and sex-biased
differences in response are quite common (Bunnerfeld et al. 2006; Lusseau 2003; Saj et al. 1999).
Generally speaking, males are likely to engage in riskier behaviour than females, trading off costs of
potential capture or injury for rewards including greater quantities of forage, or more nutritionally
rich foods (Ekananyaka et al. 2011; Bunnerfeld et al. 2006; Saj et al. 1999, Sukumar and Gadgil 1988).

33
This was evident in the Eurasian lynx, where male lynx spent more time in proximity to human
habitations than did female lynx, presumably driven by the increased density of prey species in these
locations and more willing to accept the associated anthropogenic risks (Bunnerfeld et al. 2006). A
similar story presents with male vervet monkeys who actively seek out anthropogenically disturbed
areas to forage on crops far more frequently than females (Saj et al. 1999). However, there are
exceptions. For example, while dolphins of both sexes interrupted foraging patterns and altered
locomotion by deep-diving in response to approaching tourist vessels, males were the most risk-
averse and initiated diving earlier than females (Lusseau 2003).
Elephants, both African and Asian, exemplify species whose long-term survival and wellbeing is
threatened by increasing levels of anthropogenic disturbance (Graham et al. 2010; Perera 2009; Sitati
and Walpole 2006; Bandara and Tisdell 2002). Elephant habitat in all elephant range countries has
been severely depleted due to human driven development and encroachment forcing humans and
elephants into ever-increasing proximity (Amwata and Mganga 2014; Chakraborty and Mondal 2013;
Campos-Arceiz et al. 2009; Fernando et al. 2005; Choudhury 2004; Sitati et al. 2003). Aside from a rare
few records of lions or tigers preying on young or weak elephants (lions: Davidson et al. 2013; Power
and Compion 2009; Loveridge et al. 2006, tigers: Kumaraguru et al. 2011), humans are the only
‘predator’ of elephants. Historically, humans hunted elephants for ‘sport’ and population control, and
captured elephants for entertainment, warfare and labour (Rangarajan 1996; Sukumar 1989).
Presently, poaching for body parts (Ihwagi et al. 2018; Burn et al. 2011; Kahindi et al. 2009) and
retaliatory attacks by farmers, who are frustrated and irate following elephant raids on their crops,
remain major threats to elephants (Okello 2016; Chakraborty and Mondal 2013; Gubbi 2012; Davies
et al. 2011; Webber et al. 2011; Graham et al. 2010; Perera 2009; Sitati et al. 2003). Thus, it makes
sense that elephants perceive humans as a threat (Thuppil and Goss 2013). Understanding how these
long-lived and cognitively sophisticated animals adapt behaviours to negotiate a world where their
habitat is increasingly altered by anthropogenic disturbance, could be key to developing effective
management strategies and promoting a more harmonious co-existence between humans and
elephants.
As it stands, evidence exists from multiple locations supporting the notion that elephants are aware
of the risks of anthropogenic disturbance and activity, and that behavioural adjustments are made to
minimise the risks presented by interactions with humans (Wilkie and Douglas-Hamilton 2019; Gunn

34
et al. 2013; Graham et al. 2009; Barnes et al. 2006). In rural communities across both Africa and Asia
where crop-raiding by elephants is common, elephants forage on crops almost exclusively at night
(Gunn et al. 2013; Graham et al. 2009; Barnes et al. 2006), despite wild foraging occurring often during
the day (Sukumar 1989). African elephants have also been known to increase their average speed of
locomotion when traversing areas of high human density (Wilkie and Douglas-Hamilton 2019).
Furthermore, in Kenya, elephants responded to the perceived presence of humans from specific tribes
known to present a threat, with increased vigilance and locomotion when compared to other less
harmful stimuli (Soltis et al. 2014; Bates et al. 2007). However, these examples typically focus on the
upper and lower scales of risk, comparing behaviour in the riskiest situations (e.g. foraging on crops in
a human occupied village) and the least risky (e.g. within the confines of a PA). More and more,
elephants are spending increased time outside of PA’s, especially in Asia where electric fences are less
common and often poorly maintained (Ponnusamy et al. 2016; Fernando et al. 2011; Santiapillai et al.
2006). The majority of elephants in Sri Lanka spend at least some of their time outside of PA’s, which
is not surprising given that more than half of suitable elephant habitat falls outside of protected
boundaries (Fernando et al. 2019; Santiapillai et al. 2006; Weerakoon et al. 2004). This increase in
proximity to humans certainly poses additional risks and exposure to anthropogenic disturbance for
elephants however is not as extreme as when elephants come into direct conflict with humans, such
as during crop raids. Identifying whether elephants adapt behaviours in this interim environment of
risk, and if so how, would provide clues to a largely unexplored element of human-elephant
coexistence.
When investigating and comparing behavioural patterns of any wild animal across multiple sites, it is
impossible to account for all confounding variables. This is, of course, the situation when studying wild
elephant behaviour in areas representing different levels of anthropogenic disturbance. A range of
factors, from even subtle environmental variations (e.g. vegetative structure, availability of resources,
microclimates, presence or absence of other species) (Griffin 2007) to individual animal personalities
(Hollander et al. 2008; Freeman et al. 2007) can potentially influence behaviour. However, logistic and
ethical constraints render it nigh on impossible to experimentally manipulate anthropogenic
disturbance in the wild. Captive studies can be useful but come with their own practical and ethical
considerations, and comparability may be debatable particularly in relation to long-lived and
intelligent species. That studies spanning numerous continents and species have demonstrated

35
changes in behaviour consistent with the scale of anthropogenic disturbance presented at different
locations, shows merit in such comparative approaches despite some limitations.
In this study, we provide a comparison of general behaviour patterns, specifically locomotion, foraging
and standing still, for Asian elephants utilising habitat representing different levels of anthropogenic
disturbance or risk to elephants. We selected an area of ‘low’ anthropogenic disturbance (hereafter
‘low risk zone’) located within designated wildlife PA boundaries, and an area of ‘medium’
anthropogenic disturbance (hereafter ‘medium risk zone’), located outside of the PA. Specifically, we
aimed to:
- Compare the time elephants spent foraging, in locomotion, and ‘standing still without feeding’
in the low and medium risk zones
- Compare frequency of ‘reactive behaviours’ (backing away, dusting, smelling, ‘tail up’ posture,
throwing objects, trunk touch to own mouth, vigilance/freeze posture) observed in the low
and medium risk zones
- Compare the above behaviours and actions for elephants in the presence and absence of
identified disturbances (approach of vehicle, pedestrian or dog), for each risk zone
Furthermore, researchers investigated whether behaviour differed across males and females for each
risk zone, and in the presence of known disturbances. We also investigated the potential influence of
prior negative interactions with humans, using wounds and scars as a proxy.
Information generated from this study will assist in identifying how, or indeed if, Asian elephants are
altering certain behavioural patterns, when inhabiting areas closer to human communities, and how
they are responding to the associated perceived anthropogenic risks. Contributing to the knowledge
base of animal behaviour inside and outside of protected areas generally, and specifically in response
to anthropogenic disturbance and risk, facilitates comparisons across both sites and species, and
enables retro-active evaluation of the impact of human disturbance on elephants. The future well-
being of elephant populations depends at least partially on facilitating a more peaceful coexistence
between humans and elephants living in proximity to one another. The more information garnered

36
about general behaviour patterns in human dominated landscapes, and specific responses to
anthropogenic risk and disturbance, the better equipped conservationists, managers and other policy
makers are to develop management strategies with the welfare of both elephants and humans at the
forefront.
2.3 Methods
2.3.1 Study site
This study was conducted within the dry zone of Central Sri Lanka, at two neighbouring locations which
nonetheless varied in levels of anthropogenic disturbance. The Central Sri Lankan dry zone has an
average annual temperature of 28°C, with little fluctuation throughout the year, and is characterised
by two wet seasons: the ‘long rains’ from October to January, and the ‘short rains’ from March to May.
The driest time of year is between July and September (de Silva and de Silva 2007; Fernando et al.
2005; Wikramanayake et al. 2004).
The two specific locations chosen for this research were: a) inside the south-eastern boundary of
Wasgamuwa National Park (park entrance: 07°39.051' N, 080°55.633' E), and b) an area outside of
park boundaries and in between villages called ‘Weheragala Tank’ (Figure 2.1). Both of these locations
are characterised by tropical dry mixed evergreen forest habitat bordering grassy clearings, with
access to permanent, although fluctuating, water sources. Elephants are commonly observed in both
areas, and it was considered highly likely that many individual elephants within the overall population
would utilise both areas. This provided a unique opportunity to study the behaviour of elephants from
the one population in areas of similar habitat but with varying degrees of anthropogenic activity and
low and medium disturbance. Additionally, both sites were easily accessible by vehicle with clearings
providing relatively unobstructed and safe observation opportunities. Further information on each
location is detailed below.
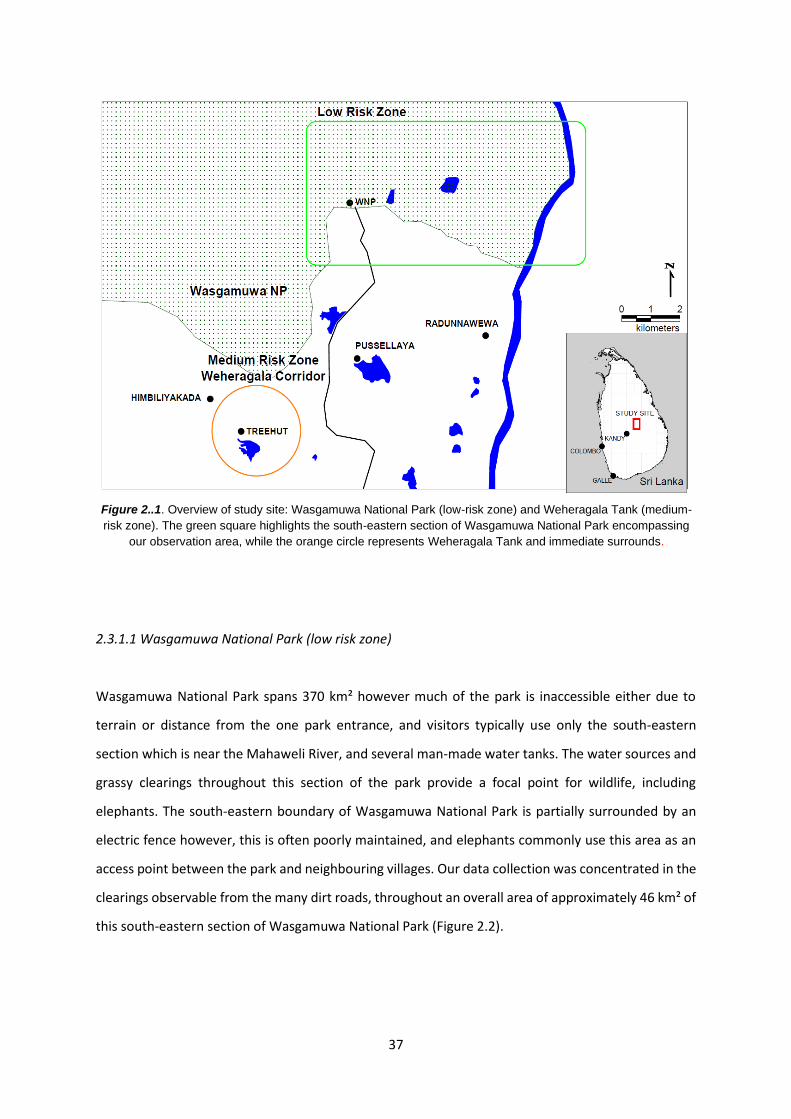
37
2.3.1.1 Wasgamuwa National Park (low risk zone)
Wasgamuwa National Park spans 370 km² however much of the park is inaccessible either due to
terrain or distance from the one park entrance, and visitors typically use only the south-eastern
section which is near the Mahaweli River, and several man-made water tanks. The water sources and
grassy clearings throughout this section of the park provide a focal point for wildlife, including
elephants. The south-eastern boundary of Wasgamuwa National Park is partially surrounded by an
electric fence however, this is often poorly maintained, and elephants commonly use this area as an
access point between the park and neighbouring villages. Our data collection was concentrated in the
clearings observable from the many dirt roads, throughout an overall area of approximately 46 km² of
this south-eastern section of Wasgamuwa National Park (Figure 2.2).
Figure 2..1. Overview of study site: Wasgamuwa National Park (low-risk zone) and Weheragala Tank (medium-
risk zone). The green square highlights the south-eastern section of Wasgamuwa National Park encompassing
our observation area, while the orange circle represents Weheragala Tank and immediate surrounds.

38
Wasgamuwa National Park was defined as a ‘low-risk zone’ for elephants, as by law, human-elephant
interactions within park boundaries should be limited to rangers and visitors travelling in vehicles
along designated tracks and minimising behaviour that could disturb the elephants. Furthermore,
visitors are only permitted in the park between 7 am and 6 pm, and at all times elephants can seek
refuge in forest away from humans.
2.3.1.2 Weheragala Tank (medium risk zone)
Located approximately two kilometres from the nearest park boundary and six kilometres from the
park entrance, Weheragala Tank is situated between three villages, all which experience some degree
of human-elephant conflict in the form of crop-raiding (pers. comm). Weheragala Tank is
characterised by a large man-made water tank, surrounded by a grassy clearing and forest habitat
(Figure 2.3). Water levels in the tank fluctuate widely throughout the year, generally coinciding with
the rain seasons, although local government departments can release or take water at any time. At its
most full, the tank covers an area of approximately 0.254 km² with water reaching up to the forest
edge. At its driest, water spans an area of only approximately 0.045 km² and a grassy clearing of up to
250 m length separates the forest and tank edges. When the tank is full, the western and southern
sides are not accessible by vehicle.
Weheragala Tank was defined as a ‘medium risk zone’ for elephants, as humans and elephants both
use resources such as water, forest edges and paths or roads. Negative interactions do occur,
particularly in the early mornings or late afternoons. At this time, people are often traveling between
villages for school or work, while elephants are often emerging from or returning to forests, before or
after foraging in the clearings and drinking from the water tanks. However, elephants are not exposed
to the same high level of negative human-elephant interactions as when they directly enter villages to
forage on cultivated crops. Thus, the anthropogenic disturbance and risk presented at Weheragala
Tank falls somewhere in the middle of the scale between PA’s and high conflict zones.

39
Figure 2.3 Weheragala Tank ‘medium risk zone’, showing the water tank and surrounding forest.
Figure 2.2 South-eastern section of Wasgamuwa National Park,showing tracks driven during
observation sessions, water reservoirs and the Mahaweli River.

40
2.3.2 Study subjects
Subjects were 72 wild and free-ranging adult Asian elephants (Elephas maximus) (25 females, 47
males), from a population inhabiting areas inside and outside of the south-eastern boundaries of
Wasgamuwa National Park. No male elephants exhibiting outward signs of musth (their reproductive
state) were included in the study. Elephants were observed in group sizes ranging from a single
elephant to groups of up to 52 individuals. Following the protocol outlined in Chiyo et al. (2011), a
group was defined as a spatially cohesive and behaviourally coordinated aggregation of two or more
elephants. ‘Spatially cohesive’ meant individuals were aggregated within a radius of 100 m and
oriented in the same general direction while moving, and ‘behaviourally coordinated’ meant the
majority of the group performed similar actions and/or interacted with other group members.
Individual adult elephants were identified using a photographic identification file which I developed
using photographs captured between May 2014 and November 2016 (e.g. Figure 2.4). Prior to this
study, no photographic identification file or other record of individuals from the local elephant
population existed. Consequently, precise ages of individuals were unknown. Elephants were
recognised as adults using a combination of cues based on body size, degree of ear folding, and
depigmentation (de Silva et al. 2011; Avirazhagen and Sukumar 2008; Gunawardene et al. 2004).
Each adult elephant was also given an ‘Elephant-Human Interaction (EHI)’ rating, using the presence
and absence of prominent lumps and wounds on their body (including head, limbs and tail) as a proxy
for negative interactions with humans. As confirmed by elephant behaviour and veterinarian experts
in Sri Lanka, these lumps are typical of those inflicted by humans using guns (including homemade or
shrapnel guns) and other catapults, often to chase elephants away from their homes or property.
Elephants were given a low, medium or high EHI score depending on the number of lumps present. If
both sides of the body could not be observed (and the elephant had less than 25 lumps visible on one
side – in which case an automatic high score is awarded), no rating was given (Table 2.1; Figures 2.5,
2.6 and 2.7). Due to small sample sizes, elephants with low and medium EHI scores were grouped
together, forming a low/medium category which we termed ‘low’ for the purposes of analysis.

41
Figure 2.4 Example from Elephant ID File. Mohan: page 1. This page shows left and right profiles and a front-on
view of Mohan’s face. Distinguishable depigmentation patterns, notches and holes in both ears are very obvious.

42
Figure 2.4 Example from Elephant ID File. Mohan: page 2. This page shows his full body left and right profiles,
illustrating numerous scars and lumps scattered about his body, and also an easily distinguishable shape for the
tail hair. All these features, in addition to ear shape and depigmentation provide important clues when attempting
to recognise individual elephants in the field.

43
EHI
score
Description
Low Elephant has ≤ 5 lumps anywhere on its body, head, limbs or tail
Medium Elephant has 6 to 24 lumps present, generally scattered sporadically across body,
head, limbs and/or tail but can also be clustered together in one or two locations
High Elephant has 25 ≥ lumps, generally across all areas of body, head, limbs and tail
Table 2.1 Elephant-human interaction (EHI) ratings and definitions
Figure 2..5 Elephant with a high Elephant-Human Interaction rating. This is an adult male elephant named
Abeeshan. This picture shows the numerous large lumps, bumps and wounds present on just the left side of his
body. Overall, I counted 46 lumps on his body and rated him as having a high level of previous negative
interactions with humans (high EHI). © Kylie M. Butler.

44
Figure 2.6 Elephant with a medium Elephant-Human Interaction rating. This is an adult male elephant
names Jagulan. This picture shows several lumps scattered about the right side of his body, including
near his rump. Overall, I counted 18 lumps on Jagulan’s body (the right side being worse affected than
the left). Jagulan was rated as having a medium level of previous negative interaction with humans
(medium EHI). © Kylie M. Butler.
Figure 2.7 Elephant with a low elephant-human interaction rating. This is an adult male
elephant named Amila. The picture shows that the right side of his body and face is free of
lumps or wounds. No lumps were counted on either side of his body and I rated him as having
a low level of previous negative interaction with humans (low EHI) © Kylie M. Butler.

45
2.3.3 Data collection: elephant observations
This study was conducted over an 18-month period between May 2015 and June 2016. Elephant
observations and recording of video footage in Wasgamuwa National Park were performed between
May 2015 and June 2016 in Wasgamuwa National Park, and between May 2015 and October 2016 at
Weheragala Tank. Elephant observations were typically performed in Wasgamuwa National Park once
per week, and at Weheragala Tank three times per week, during three ten-week field seasons. The
difference in sampling effort was due to reduced accessibility of the park following or during heavy
rain, the availability of a reliable research vehicle, and the availability of one of our two regular park
trackers.
The precise methods of approaching elephants and determining observations spots differed between
the two locations. This was due to differences in size, with Wasgamuwa National Park being much
larger and requiring us to drive around in order to find elephants, and Weheragala Tank being much
smaller requiring only stationary observation spots. Furthermore, in Wasgamuwa National Park we
were required to remain inside a closed-roof vehicle at all times, and to drive only along designated
dirt roads (refer back to Figure 2.2). Whereas, at Weheragala Tank we were limited in where we could
drive, as few tracks existed in the forest. Furthermore, it was not safe to drive through the forest in
the late afternoon when elephants were active. Therefore, stationary observation spots in the clearing
were selected and we waited to see if elephants would appear, rather than actively searching for them
(refer back to Figure 2.3).
Observations in Wasgamuwa National Park took place between 3 pm and 6:30 pm (when the park
closed). The exact route and distance covered during each session depended upon weather, road
condition and how many elephants were observed. Often park staff had information about where
elephants were located, and as we aimed to collect as much footage of elephant behaviour as possible,
this also influenced the route. For personal safety, and out of respect for the elephants and to
minimise disturbance caused by our presence, we always tried to place ourselves more than 30 m to
the nearest elephant, and to park parallel to the group, switching the engine off as soon as possible.
However, on some occasions the elephants were not sighted until we were within 30 m due to thick
vegetation or bends in the road. Video recording began as soon as we were stationary and continued
for 20 minutes, or until the elephant moved out of sight (if less than 20 minutes). If it was unsafe or
would cause disturbance to the elephants to leave after 20 minutes (e.g. elephants were on the road,

46
elephants were close to the vehicle, or a group with juveniles were separated on both sides of the
road) video recording continued until it was deemed appropriate to leave (Figure 2.8).
Observations at Weheragala Tank took place between 3:30 pm and 6:30 pm, when it would start to
become too dark for filming. Whether we observed from set locations to the north of the tank or to
the south-west of the tank depended on the likelihood of elephants being present, as determined by
recent observations and information from residents of neighbouring villages. In the area to the north
of the tank, observations were performed from a 4.5 m high platform erected in a tree hut with a 360°
view (Figure 2.9). At the south-western location, observations were conducted from inside, or
standing next to, the research vehicle between the tank edge and the forest (at least 50 m from the
forest edge) (Figure 2.10). When the water tank was full, the areas to the south and west of the tank
were inaccessible, and all observations were done from the north. At Weheragala Tank, video
recording began when elephants were first sighted until elephants were out of sight, or it was too dark
to continue observing.
Figure 2.8 Elephant observations in Wasgamuwa National Park
Left: A group of elephants foraging on grass and interacting, as seen from research vehicle © Kylie M. Butler
Right: Researcher Kylie M. Butler photographing elephants in Wasgamuwa National Park © Supun Herath

47
Figure 2.9 Elephant observations to the north of
Weheragala Tank
Left: The Tree Hut platform used for elephant
observations. To the left of the tree hut is forest habitat
and to the right the water tank. © Lacy Wittman.
Right: Field assistant Supun Herath and researcher
Kylie Butler filming and photographing a group of
elephants from the tree hut. © Tim Hunt.
Figure 2.10 Elephant observations from south-west of Weheragala Tank
Researcher Kylie Butler film a group of elephants at a distance of approximately 100 m from atop
the research vehicle © Lacy Wittman.

48
All video footage was filmed by a trained research assistant or myself, using a Sony Handycam HDR-
CX240 HD, Canon Legria HFR706, and/or Panasonic Lumix DMC-T 280 Ultra Zoom digital camera. Every
attempt was made to video record all elephants in the group, with the aim of collecting data on as
many individual elephants as possible. However, this was dependent upon group cohesion, and
individual movement patterns during the observation session. For example, large groups often
scattered over a wide area to feed, some individuals would walk back into the forest and its vegetative
cover, or a group would surround the vehicle rendering it impossible to film all elephants.
Furthermore, elephants who ventured furthest from forest habitat and thus closest to our observation
spots, were the most easily visible to film. Due to restrictions on where we were allowed to drive in
Wasgamuwa National Park, and where it was safe to drive at Weheragala Tank, our ability to
manoeuvre into position to give all elephants equal opportunity to be filmed was limited. However,
at no stage was filming concentrated on any individual elephant, or group of elephants, because they
were seen to be doing something more active or ‘interesting’ than others.
While video footage was being recorded, another observer would record the data and time of the
observation, the number of elephant’s present, whether they were in an all-male, cow-calf or mixed
groups, and the predominant activity of the group. Field notes detailing general group behaviours and
interactions or reactions to external stimuli were also documented, and proved useful when cross-
checking with video footage for disturbance events.
2.3.4 Behaviour coding from video footage
Using field notes to help determine on which dates and when disturbances occurred during elephant
observations, video footage was viewed and categorised into five to ten-minute segments, and divided
into two contexts: a) baseline and b) disturbance. Baseline meant that no known disturbances
occurred during the selected video coding period, or five minutes before the beginning of the video
coding period, and that observed behaviour was therefore not influenced by any external negative
stimuli identifiable by the observer.
Disturbance was defined as when an event presumed to represent a perceived threat or disturbance
to elephants occurred. Such events included approach of another vehicle (safari jeeps in Wasgamuwa
National Park, or general vehicles traveling in between villages at Weheragala Tank including jeeps,
tuk tuks and motorbikes), people moving on foot within sight of elephants and approaching to less

49
than 50 m (Weheragala Tank only), and dogs approaching elephants to within 50 m (Weheragala Tank
only). All disturbance events were recorded opportunistically, and at no time did we encourage or
manipulate the onset of a disturbance.
Overall, video footage from 85 elephant sightings was recorded (30 from Wasgamuwa National Park,
55 from Weheragala Tank). From these 85 recordings, 34 videos had segments suitable for extraction
of behavioural data during the baseline context (14 Wasgamuwa National Park, 20 Weheragala Tank),
and 26 videos had segments suitable for extraction of behavioural data during the disturbance context
(8 Wasgamuwa National Park, 18 Weheragala Tank). Some videos were suitable for only one of
baseline or disturbance data, and some were suitable for both. When videos were suitable for both
contexts, baseline data was extracted from video segments occurring before the disturbance event,
or if this was not possible, a minimum seven-minute interval between the end of the disturbance
event, and the beginning of the video segment used in coding for baseline behaviour was applied.
For the baseline context, videos were deemed to be of suitable quality for analysis if a minimum of
one focal individual was visible for a minimum period of five minutes. For the disturbance context,
videos were deemed to be of suitable quality for analysis if a minimum of one focal individual was
visible when the disturbance occurred and then either remained visible in the clearing, or was
confirmed to be behind vegetation (e.g. was seen entering the forest, and video footage and/or field
notes confirmed he/she did not re-emerge) for a minimum five minute period following the onset of
the disturbance. For each context, videos were assigned a numerical code and order of video analysis
was selected randomly. Once a video was selected for analysis the group type, size and composition
were confirmed, and all focal individuals identified. Following this, behavioural data was extracted one
focal individual at a time, with order of focal individuals also selected randomly.
For each context, the time an elephant spent in locomotion, counted in seconds from the first step
taken to the last, was recorded and expressed as a percentage of overall time, to account for variations
in observation durations between focal individuals. Scan samples at 30 second intervals were also
recorded to estimate durations of feeding bouts and time elephants spent ‘standing still without
feeding’, the assumption being that if elephants were observed performing the same behaviour for
consecutive scan samples they were considered to have been doing so for the overall period. These
were also converted to percentages. Counts of other behaviours, specifically backing away, dusting,

50
Behaviour Operational definition
Back away Elephant backward locomotion of three paces or more, while facing and within
ten metres of a vehicle, person or other (non-conspecific) animal when
backwards locomotion begins
Dusting Elephant picks up dirt/dust with its trunk and throws dirt/dust anywhere over its
body, face, legs or behind ears
Feeding Elephant picks up vegetation with its trunk, then places vegetation into its mouth
and chews within 30-s of picking vegetation up. Between picking up vegetation
and placing it in its mouth, the elephant often spends time ‘preparing’ food. This
can involve swinging/flicking it from side-to-side and/or breaking it with foot, or
hitting it on the ground. Elephant often chews for a long time in between placing
more food in mouth.
Freeze Elephant stands with head, ears, trunk and body motionless for five seconds or
more. Posture is usually stiff.
Locomotion:
running
Fast-paced forward locomotion in a specific direction, five paces or more.
Elephants knees are noticeably raised from the ground as it moves. Tail is often
(but not always) help horizontal to the body or higher.
Locomotion:
walking
Elephant forward locomotion (without running) in a specific direction (left, right
or straight) five paces or more
Smell Elephant holds trunk in the air or extends trunk in front of face and above knee
height, with trunk tip pointed in a direction of interest for two seconds or more
Standing still Elephant stands still in one location without feeding (and without holding freeze
posture). Can include shuffling footsteps from side-to-side or back-and-forth.
Tail up Elephant holds its tail horizontal to body or higher in a stiff posture, for a
minimum of three seconds.
Trunk to
mouth (self)
Elephant touches trunk tip to its own mouth (when it is not feeding or drinking).
Vigilance Elephant stands with head raised and either held motionless or slowly scanning
from side-to-side, plus one or more of the following: ears held perpendicular to
body, tail base rigid, and/or base of trunk thrust forward. Posture is held for a
minimum of three seconds.

51
Table 2.2. Elephant ethogram. Developed, adapted and expanded from: Freeman et al. 2010; Horback et al.
2013, and researchers personal field observations of wild African and Asian elephants: Kylie Butler 2011 – 2016.
smelling, ‘tail up’, throwing objects, trunk touch to own mouth, and vigilance/freeze posture, were
also recorded (Table 2.2). These behavioural responses were selected as they are typical of those
performed when an elephant is exploring a perceived external stimulus or feeling stressed, and we
termed these ‘reactive behaviours’(Table 2.2). These were converted to a ‘rate per minute’ (RPM)
score, however counts of individual ‘reactive behaviours’ were very low. To account for low counts,
and increase comparability across risk zone, sex and context, all ‘reactive behaviours’ were grouped
together into a single count for each elephant, from which an RPM score was calculated.
2.3.5 Statistical analysis
The potential effects of context (baseline or disturbance) and the sex of elephants (female or male),
on: 1) % time spent feeding; 2) % time spent in locomotion; 3) % time spent standing still (without
feeding); and 4) reactive behaviours (RPM) were measured using linear mixed effects models (LMM).
For RPM scores, the duration of the video segment measured in seconds was weighted to account for
variations in observation time which could have a bearing on the number of reactive behaviours
performed. For each of the four behaviours of interest, the interaction between context and sex were
included as predictor variables, with the identity of individual elephants fitted as a random intercept
term to allow for the repeated measure (paired) comparison of individual elephants who were
observed in both contexts. Percentage time spent in locomotion, and percentage time spent standing
still (without feeding) were both log(10) transformed before analysis to improve normality of data
(Warton and Hui 2011). Significance of terms in the model (p-value = 0.05), and post-hoc tests of
differences in least-squares means between categories were assessed using the Satterthwaite
approximation of degrees of freedom obtained from the lmer package. The proportion deviance
explained by the fixed effects and combined random fixed effects respectively, were calculated using
the marginal and conditional r-squared (Nakagawa and Schielzeth 2013).
Although we were interested in whether elephants differed in their response to disturbance relative
to baseline according to sex as well as risk level, final sample sizes were too small to allow inclusion of
the three-way interaction (risk * context * sex) in statistical models. For this reason, we opted to

52
examine responses for each risk zone separately, including only the two -way (context * sex)
interaction.
Finally, the potential effects of prior negative interactions with humans (EHI levels) were tested for
male elephants only. This is because EHI levels were confidently determined for all males but not all
females. Furthermore, the effect of EHI on the behaviour of male elephants inside and outside of PA’s
is of particular interest, as male-biases in HEC events are common. As only two male elephants
received a low EHI score, these were grouped with ‘medium’ EHI elephants to form a combined
low/medium EHI category. To test for effects of EHI levels, we fitted linear mixed models for each
response variable with EHI level and context as explanatory variables. The potential effects of prior
negative interactions with humans (EHI levels) were also tested by fitting linear mixed models for each
response variable with EHI level and context as explanatory variables.
Statistical analyses were conducted with the R Studio Statistical Package 3.5.1 (R core team 2018).
LMM’s were constructed using the package ‘lme4’ (Bates et al. 2014), with model outputs generated
with the ‘effects’ package (Fox 2003) and visualised with ‘ggplot2’ (Wickham 2016).
2.4 Results
2.4.1 General behavioural activities
The time elephants spent engaged in the three main behaviours recorded during afternoon
observations barely deviated between Wasgamuwa National Park (low risk zone) and Weheragala
Tank (medium risk zone). At both locations, the predominant activity was feeding, and elephants (n =
72) spent approximately half of the observed time doing this. Time spent in locomotion and standing
still (without feeding) was approximately 20% for each, and for both locations. One-way between
group analyses of variance (ANOVA) on each activity separately showed no significant differences
between the mean time elephants spent in any of these activities between Wasgamuwa National Park
and Weheragala Tank (p = 0.05) (Table 2.3).
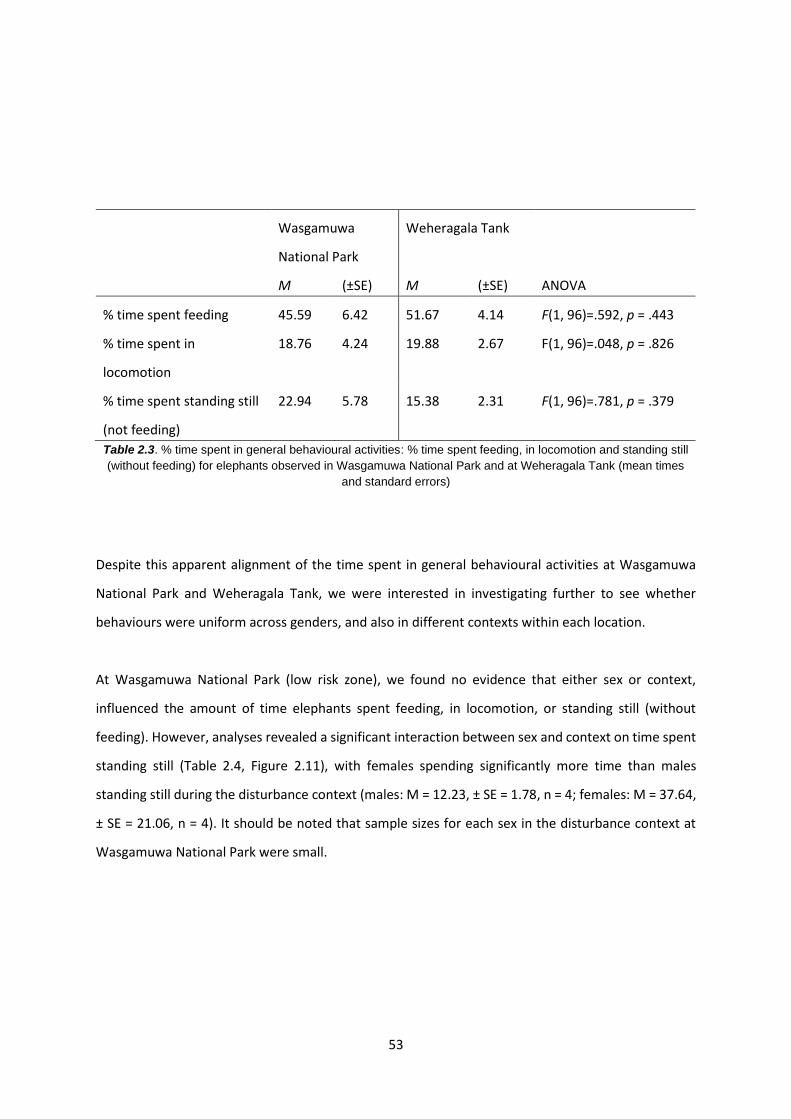
53
Wasgamuwa
National Park
Weheragala Tank
M (±SE) M (±SE) ANOVA
% time spent feeding 45.59 6.42 51.67 4.14 F(1, 96)=.592, p = .443
% time spent in
locomotion
18.76 4.24 19.88 2.67 F(1, 96)=.048, p = .826
% time spent standing still
(not feeding)
22.94 5.78 15.38 2.31 F(1, 96)=.781, p = .379
Table 2.3. % time spent in general behavioural activities: % time spent feeding, in locomotion and standing still
(without feeding) for elephants observed in Wasgamuwa National Park and at Weheragala Tank (mean times
and standard errors)
Despite this apparent alignment of the time spent in general behavioural activities at Wasgamuwa
National Park and Weheragala Tank, we were interested in investigating further to see whether
behaviours were uniform across genders, and also in different contexts within each location.
At Wasgamuwa National Park (low risk zone), we found no evidence that either sex or context,
influenced the amount of time elephants spent feeding, in locomotion, or standing still (without
feeding). However, analyses revealed a significant interaction between sex and context on time spent
standing still (Table 2.4, Figure 2.11), with females spending significantly more time than males
standing still during the disturbance context (males: M = 12.23, ± SE = 1.78, n = 4; females: M = 37.64,
± SE = 21.06, n = 4). It should be noted that sample sizes for each sex in the disturbance context at
Wasgamuwa National Park were small.

54
Response Predictor variable Coefficient SE t R²
(marginal)
R²
(conditional)
Locomotion
(log₁₀)
(Intercept) 0.93 0.14 5.56 0.06 0.75
Context – Disturbance 0.11 0.19 0.60
Sex – Female 0.29 0.24 1.21
Context – Disturbance: Sex
– Female
-0.21 0.29 -0.72
Feeding (Intercept) 54.13 9.65 5.61 0.05 0.53
Context – Disturbance -17.49 16.08 -1.09
Sex – Female -6.28 16.54 -0.38
Context – Disturbance: Sex
- Female
7.63 24.32 0.31
Standing still
(not feeding)
(log₁₀)
(Intercept) 0.82 0.22 3.8 0.15 0.98
Context – Disturbance -0.08 0.08 -0.91
Sex – Female -0.16 0.35 -0.45
Context - Disturbance: Sex
- Female
1.07 0.13 8.01
Table 2.4. LMM summary: effects of context * sex on general behaviour in Wasgamuwa National Park
Parameter coefficients, standard errors (SE) and t-values for the fixed effects in linear mixed models examining
the relationship between context * sex of elephants in the low risk zone, and R² values of the fixed terms
(marginal) and fixed and random terms (conditional). Coefficients for which the 95% confidence interval does not
include zero are shown in bold. Reference level of factors: context (baseline), sex (male).
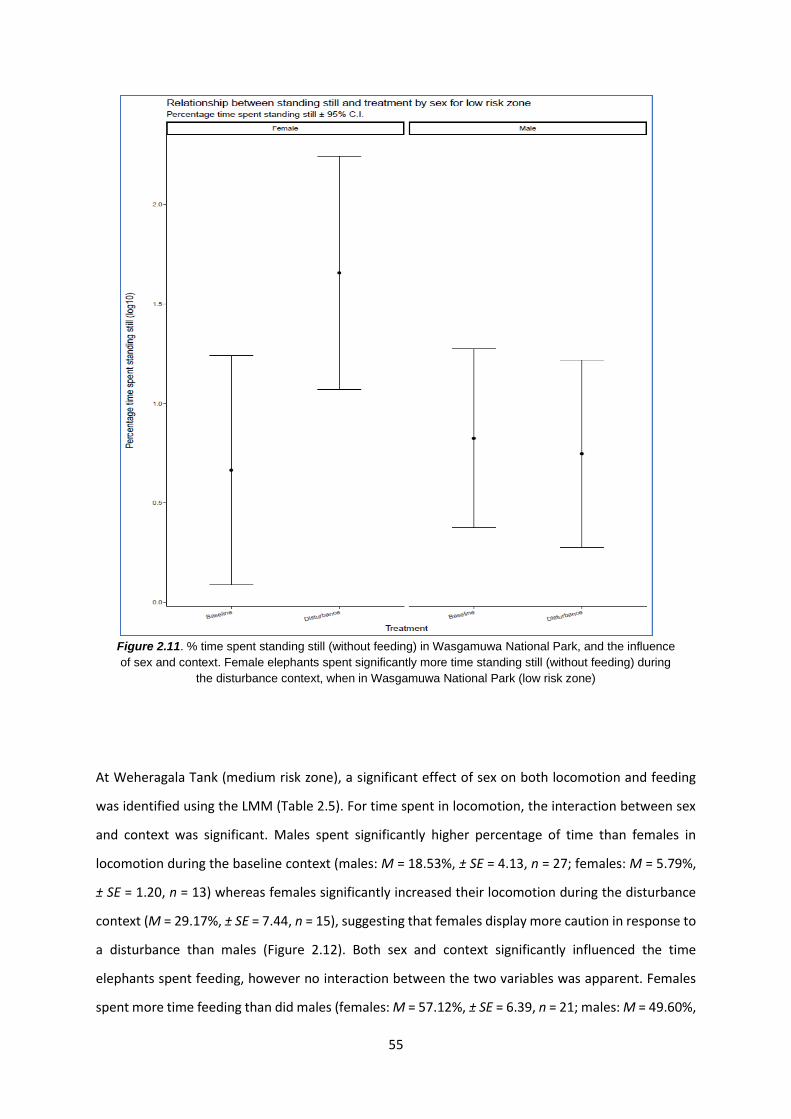
55
At Weheragala Tank (medium risk zone), a significant effect of sex on both locomotion and feeding
was identified using the LMM (Table 2.5). For time spent in locomotion, the interaction between sex
and context was significant. Males spent significantly higher percentage of time than females in
locomotion during the baseline context (males: M = 18.53%, ± SE = 4.13, n = 27; females: M = 5.79%,
± SE = 1.20, n = 13) whereas females significantly increased their locomotion during the disturbance
context (M = 29.17%, ± SE = 7.44, n = 15), suggesting that females display more caution in response to
a disturbance than males (Figure 2.12). Both sex and context significantly influenced the time
elephants spent feeding, however no interaction between the two variables was apparent. Females
spent more time feeding than did males (females: M = 57.12%, ± SE = 6.39, n = 21; males: M = 49.60%,
Figure 2.11. % time spent standing still (without feeding) in Wasgamuwa National Park, and the influence
of sex and context. Female elephants spent significantly more time standing still (without feeding) during
the disturbance context, when in Wasgamuwa National Park (low risk zone)
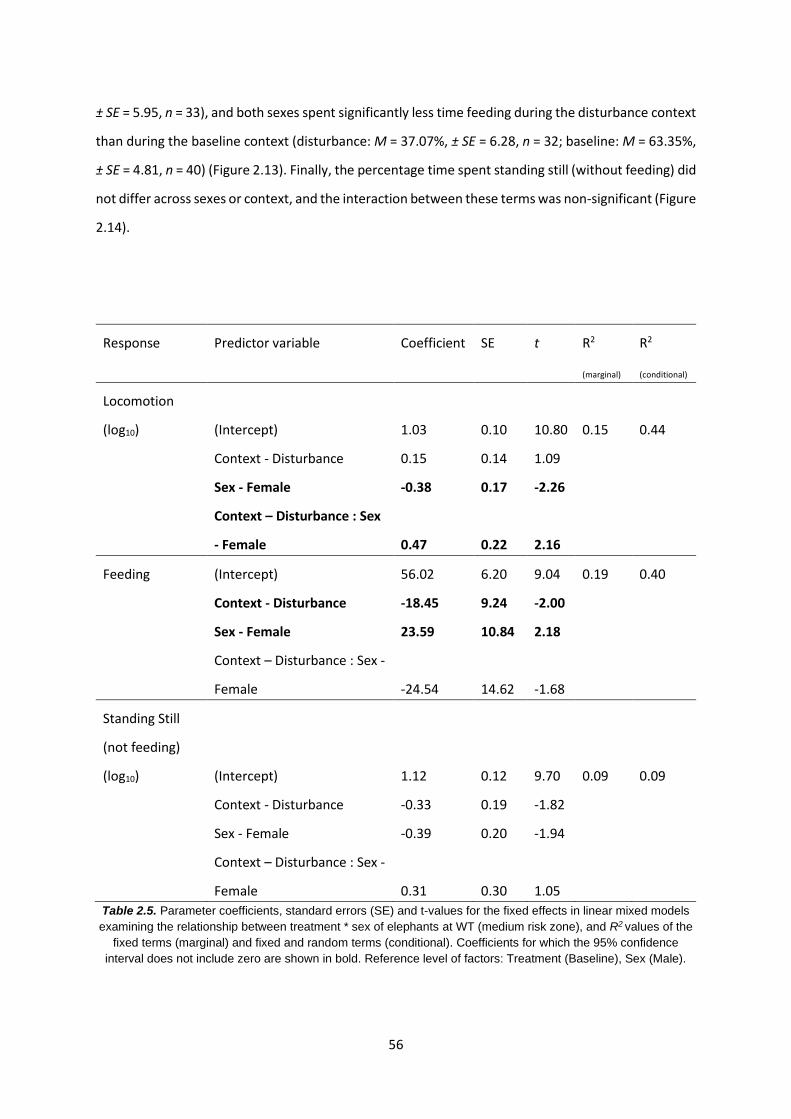
56
± SE = 5.95, n = 33), and both sexes spent significantly less time feeding during the disturbance context
than during the baseline context (disturbance: M = 37.07%, ± SE = 6.28, n = 32; baseline: M = 63.35%,
± SE = 4.81, n = 40) (Figure 2.13). Finally, the percentage time spent standing still (without feeding) did
not differ across sexes or context, and the interaction between these terms was non-significant (Figure
2.14).
Response Predictor variable Coefficient SE t R2
(marginal)
R2
(conditional)
Locomotion
(log10) (Intercept) 1.03 0.10 10.80
0.15
0.44
Context - Disturbance 0.15 0.14 1.09
Sex - Female -0.38 0.17 -2.26
Context – Disturbance : Sex
- Female 0.47 0.22 2.16
Feeding (Intercept) 56.02 6.20 9.04 0.19 0.40
Context - Disturbance -18.45 9.24 -2.00
Sex - Female 23.59 10.84 2.18
Context – Disturbance : Sex -
Female -24.54 14.62 -1.68
Standing Still
(not feeding)
(log10)
(Intercept) 1.12 0.12 9.70
0.09
0.09
Context - Disturbance -0.33 0.19 -1.82
Sex - Female -0.39 0.20 -1.94
Context – Disturbance : Sex -
Female 0.31 0.30 1.05
Table 2.5. Parameter coefficients, standard errors (SE) and t-values for the fixed effects in linear mixed models
examining the relationship between treatment * sex of elephants at WT (medium risk zone), and R2 values of the
fixed terms (marginal) and fixed and random terms (conditional). Coefficients for which the 95% confidence
interval does not include zero are shown in bold. Reference level of factors: Treatment (Baseline), Sex (Male).
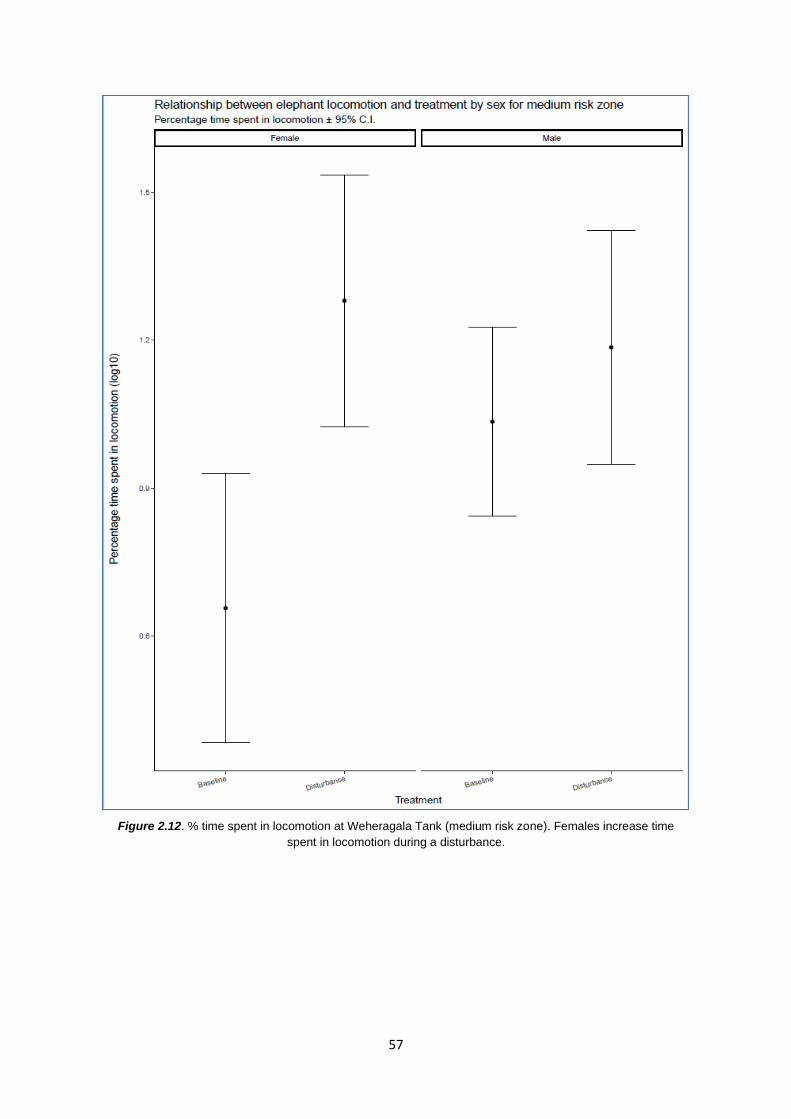
57
Figure 2.12. % time spent in locomotion at Weheragala Tank (medium risk zone). Females increase time
spent in locomotion during a disturbance.

58
Figure 2.13. % time spent feeding at Weheragala Tank (medium risk zone). Females spent the most time
feeding but this was reduced during the disturbance context.
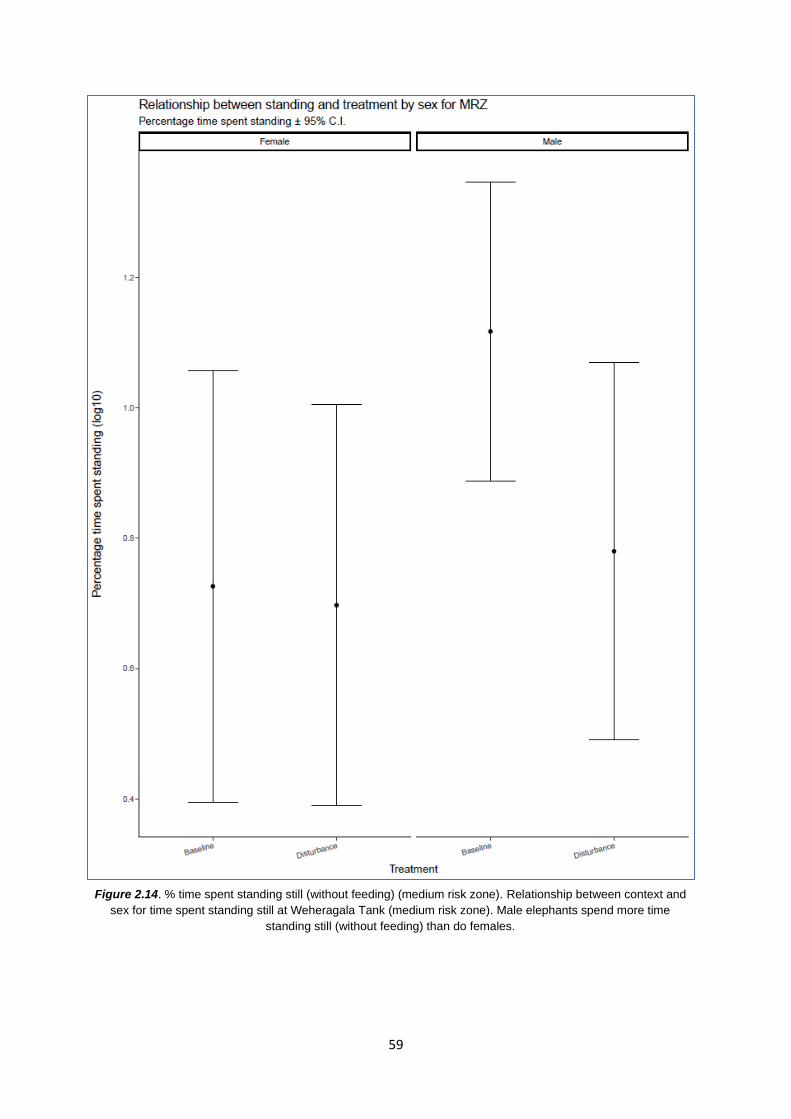
59
Figure 2.14. % time spent standing still (without feeding) (medium risk zone). Relationship between context and
sex for time spent standing still at Weheragala Tank (medium risk zone). Male elephants spend more time
standing still (without feeding) than do females.
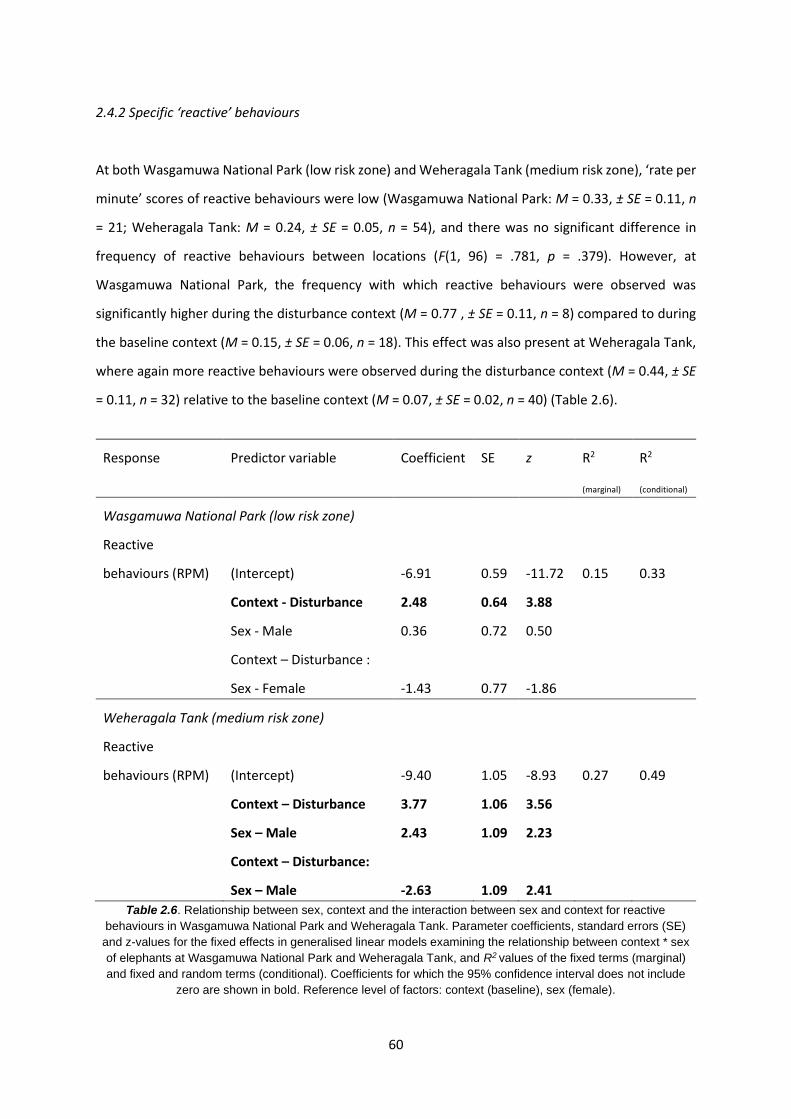
60
2.4.2 Specific ‘reactive’ behaviours
At both Wasgamuwa National Park (low risk zone) and Weheragala Tank (medium risk zone), ‘rate per
minute’ scores of reactive behaviours were low (Wasgamuwa National Park: M = 0.33, ± SE = 0.11, n
= 21; Weheragala Tank: M = 0.24, ± SE = 0.05, n = 54), and there was no significant difference in
frequency of reactive behaviours between locations (F(1, 96) = .781, p = .379). However, at
Wasgamuwa National Park, the frequency with which reactive behaviours were observed was
significantly higher during the disturbance context (M = 0.77 , ± SE = 0.11, n = 8) compared to during
the baseline context (M = 0.15, ± SE = 0.06, n = 18). This effect was also present at Weheragala Tank,
where again more reactive behaviours were observed during the disturbance context (M = 0.44, ± SE
= 0.11, n = 32) relative to the baseline context (M = 0.07, ± SE = 0.02, n = 40) (Table 2.6).
Response Predictor variable Coefficient SE z R2
(marginal)
R2
(conditional)
Wasgamuwa National Park (low risk zone)
Reactive
behaviours (RPM)
(Intercept) -6.91 0.59 -11.72
0.15
0.33
Context - Disturbance 2.48 0.64 3.88
Sex - Male 0.36 0.72 0.50
Context – Disturbance :
Sex - Female -1.43 0.77 -1.86
Weheragala Tank (medium risk zone)
Reactive
behaviours (RPM) (Intercept) -9.40 1.05 -8.93
0.27
0.49
Context – Disturbance 3.77 1.06 3.56
Sex – Male 2.43 1.09 2.23
Context – Disturbance:
Sex – Male -2.63 1.09 2.41
Table 2.6. Relationship between sex, context and the interaction between sex and context for reactive
behaviours in Wasgamuwa National Park and Weheragala Tank. Parameter coefficients, standard errors (SE)
and z-values for the fixed effects in generalised linear models examining the relationship between context * sex
of elephants at Wasgamuwa National Park and Weheragala Tank, and R2 values of the fixed terms (marginal)
and fixed and random terms (conditional). Coefficients for which the 95% confidence interval does not include
zero are shown in bold. Reference level of factors: context (baseline), sex (female).
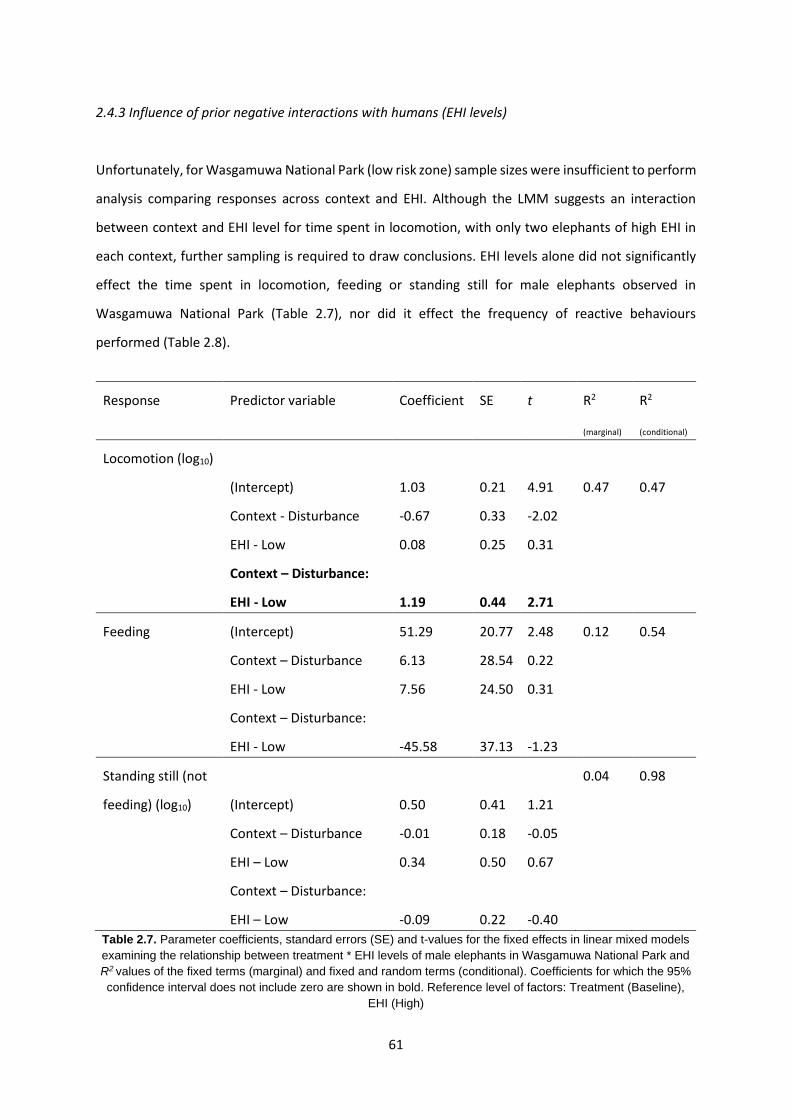
61
2.4.3 Influence of prior negative interactions with humans (EHI levels)
Unfortunately, for Wasgamuwa National Park (low risk zone) sample sizes were insufficient to perform
analysis comparing responses across context and EHI. Although the LMM suggests an interaction
between context and EHI level for time spent in locomotion, with only two elephants of high EHI in
each context, further sampling is required to draw conclusions. EHI levels alone did not significantly
effect the time spent in locomotion, feeding or standing still for male elephants observed in
Wasgamuwa National Park (Table 2.7), nor did it effect the frequency of reactive behaviours
performed (Table 2.8).
Response Predictor variable Coefficient SE t R2
(marginal)
R2
(conditional)
Locomotion (log10)
(Intercept) 1.03 0.21 4.91
0.47
0.47
Context - Disturbance -0.67 0.33 -2.02
EHI - Low 0.08 0.25 0.31
Context – Disturbance:
EHI - Low 1.19 0.44 2.71
Feeding (Intercept) 51.29 20.77 2.48 0.12 0.54
Context – Disturbance 6.13 28.54 0.22
EHI - Low 7.56 24.50 0.31
Context – Disturbance:
EHI - Low -45.58 37.13 -1.23
Standing still (not
feeding) (log10) (Intercept) 0.50 0.41 1.21
0.04 0.98
Context – Disturbance -0.01 0.18 -0.05
EHI – Low 0.34 0.50 0.67
Context – Disturbance:
EHI – Low -0.09 0.22 -0.40
Table 2.7. Parameter coefficients, standard errors (SE) and t-values for the fixed effects in linear mixed models
examining the relationship between treatment * EHI levels of male elephants in Wasgamuwa National Park and
R2 values of the fixed terms (marginal) and fixed and random terms (conditional). Coefficients for which the 95%
confidence interval does not include zero are shown in bold. Reference level of factors: Treatment (Baseline),
EHI (High)

62
Response Predictor variable Coefficient SE z R2
(marginal)
R2
(conditional)
Reactive
Behaviour (RPM) (Intercept) -8.57 1.50 -5.73
0.10
0.52
Treatment - Disturbance 2.04 0.82 2.49
EHI - Low 1.93 1.51 1.28
Treatment –Disturbance:
EHI - Low -1.87 0.97 -1.92
Table 2.8. Parameter coefficients, standard errors (SE) and z-values for the fixed effects in generalized linear
mixed models examining the relationship between treatment * EHI levels of elephants in the low risk zone, and
R2 values of the fixed terms (marginal) and fixed and random terms (conditional). Coefficients for which the 95%
confidence interval does not include zero are shown in bold. Reference level of factors: Treatment (Baseline),
EHI (High).
Sample sizes were higher at Weheragala Tank (medium risk zone), and of the 33 individual male
elephants for whom behaviour was scored, 13 had a high EHI level and 19 fell within the combined
low/medium category. One elephant was excluded from this analysis as his EHI level could not be
determined. The percentage time spent in locomotion, feeding and standing still (without feeding)
differed significantly between elephants of the two EHI categories (Table 2.9). Male elephants with a
high EHI level spent more time in locomotion than those with a low EHI level (high EHI: M = 33.92, ±
SE = 6.78, n = 19; low EHI: M = 13.09, ± SE = 3.23, n = 19), but less time feeding (high EHI: M = 28.77, ±
SE = 7.65, n = 19; low EHI: M = 66.58, ± SE = 7.15, n = 19). Male elephants with a high EHI level also
spent more time standing still (without feeding) than those with a low EHI level (high EHI: M = 25.81,
± SE = 6.26, n = 13; low EHI: M = 11.20, ± SE = 2.13, n = 19). Furthermore, elephants with a high EHI
level were observed performing reactive behaviours more frequently (high EHI: M = 0.45, ± SE = 0.17,
n = 13; low EHI: M = 0.08, ± SE = 0.03, n = 19) (Table 2.10).

63
Response Predictor variable Coefficient SE t R2
(marginal)
R2
(conditional)
Locomotion (log10)
(Intercept) 1.25 0.16 7.66
0.21
0.41
Context - Disturbance 0.24 0.21 1.11
EHI - Low -0.34 0.20 -1.70
Context – Disturbance :
EHI - Low -0.33 0.29 -1.11
Feeding (Intercept) 38.31 10.62 3.61 0.26 0.58
Context – Disturbance -20.06 13.08 -1.53
EHI - Low 27.59 13.08 2.11
Context – Disturbance:
EHI - Low 14.27 18.03 0.79
Standing still (not
feeding) (log10) (Intercept) 1.37 0.18 7.71
0.17
0.37
Context – Disturbance -0.38 0.23 -1.61
EHI – Low -0.37 0.22 -1.72
Context – Disturbance:
EHI – Low -0.04 0.32 -0.13
Table 2.9. Parameter coefficients, standard errors (SE) and t-values for the fixed effects in linear mixed models
examining the relationship between treatment * EHI levels of male elephants at Weheragala Tank and R2 values
of the fixed terms (marginal) and fixed and random terms (conditional). Coefficients for which the 95% confidence
interval does not include zero are shown in bold. Reference level of factors: Treatment (Baseline), EHI (High).

64
Response Predictor variable Coefficient SE
R2
(marginal)
R2
(conditional)
Reactive
Behaviour (RPM) (Intercept) -5.89 0.36
-
16.26
0.24
0.39
Treatment - Disturbance 0.96 0.29 3.31
EHI - Low -1.86 0.54 -3.47
Treatment – Disturbance:
EHI - Low 0.33 0.62 0.53
Table 2.10. Parameter coefficients, standard errors (SE) and z-values for the fixed effects in generalized linear
mixed models examining the relationship between treatment * EHI levels of elephants at Weheragala Tank, and
R2 values of the fixed terms (marginal) and fixed and random terms (conditional). Coefficients for which the 95%
confidence interval does not include zero are shown in bold. Reference level of factors: Treatment (Baseline),
EHI (High).
2.5 Discussion
2.5.1 General behaviour patterns in Wasgamuwa National Park and Weheragala Tank
This study revealed no evidence that elephants observed at Wasgamuwa National Park (low risk zone)
and Weheragala Tank (medium risk zone) differed in general behavioural patterns including time
spent in locomotion, feeding, or standing still (without feeding), or in the frequency of reactive
behaviours performed. Elephants spent most of their time feeding, which is not surprising considering
that to meet their nutritional requirements, Asian elephants consume up to 150 kg of vegetation every
day and often forage for up to 18 hours per day (Sukumar 1989; Vancuylenberg 1977, McKay 1973).
Although elephants are general feeders and consume a wide variety of plants, grass forms an
important component of their diet (Alahakoon et al. 2017; Sukumar 1989; Vancuylenberg 1977) and
both Weheragala Tank and the south-eastern section of Wasgamuwa National Park provide ample
opportunity for grazing. Elephants spent approximately 20% of their time in locomotion, generally
walking between clearings and the forest, or towards water tanks to drink. About the same proportion
of time was spent standing still (without feeding). Elephants very rarely performed ‘reactive’
behaviours.

65
The lack of difference in behaviour across the two risk zones, contrasts with numerous other studies
that have demonstrated changes in behaviour relative to the level of anthropogenic disturbance
present, for a wide array of species (Caro 2005; Lacy and Martins 2003; Frid and Dill 2002). Many of
these studies show a reduction in foraging activity at higher levels of anthropogenic disturbance
(Donaldio and Buskirk 2006; White et al. 1999) however, most studies do focus on more extreme
differences in risk across sites. However, in one study looking at the behaviour of large mammals inside
and outside of PA’s in Tanzania, elephants did not adjust their behaviour in the presence of a potential
anthropogenic disturbance, although most of the other species in the area did (Kiffner et al. 2014).
When an animal alters its usual behaviour as a response to negative external stimuli, costs to fitness
or health are often incurred (Blumstein et al. 2005; Lacy and Martins 2003; Frid and Dill 2002; Anthony
and Blumstein 2000; Lima 1998). This means that whether consciously or unconsciously, behavioural
choices are often governed by some level of cost-benefit analysis or trade-off (Bunnerfeld et al. 2006).
Given the abundance of available forage at Weheragla Tank, coupled with the shelter (plus additional
food sources) provided by the forest on the outskirts of the grassy clearings, elephants could in fact
be drawn to leave the protected boundaries of Wasgamuwa National Park because of the resources
available here. It could be that the ‘medium’ level of risk presented at Weheragala Tank is below the
threshold at which significant behavioural changes would be likely to occur, and that the benefits of
easily accessible food, water and shelter outweigh the slightly increased risk of interactions with
humans.
Thus, the lack of obvious behavioural differences between Wasgamuwa National Park and Weheragala
Tank could indicate that the level of risk represented by each site was not as divergent as presumed.
Whilst Weheragala Tank certainly appears to accurately represent an area of ‘medium’ anthropogenic
risk, there were indications that Wasgamuwa National Park did not fully represent the ‘low risk’ zone
it theoretically should be. Visitor numbers at Wasgamuwa National Park are considerably lower than
at other elephant inhabited parks in Sri Lanka (Senervirathna & Perera 2013), therefore elephants
within this population may be less habituated to tourists and consider their presence a risk or
disturbance, just as they would outside a PA. Furthermore, it was not uncommon to see vehicles
revving their engines or approaching elephants quickly, driving between mothers and offspring, and
talking loudly. These are all human behaviours which elephants could perceive as a disturbance or
threat (researcher pers. obs).

66
Identifying ‘medium’ risk zones near other National Parks within Sri Lanka and comparing behaviour
inside and outside of these PA’s would provide a useful comparison to our study in the Wasgamuwa
National Park region and provide further information as to how, or indeed if, Sri Lankan elephants
adjust their behaviour at an interim level of anthropogenic disturbance.
2.5.2 Effects of context and sex on elephant behaviour
Whether elephants were located in the low or medium risk zone did not appear to influence
behaviour, however the presence of an immediate anthropogenic risk or disturbance did. At
Weheragala Tank, we opportunistically witnessed disturbances including vehicles, pedestrians or dogs
approaching the elephants at a distance closer than 50 metres (Figure 2.15). Often people would be
revving engines of motorbikes or tuk tuks, playing loud music, or shouting loudly as they approached.
In response to this direct presence of a disturbance, elephants reduced the time they spent feeding
and increased the frequency of reactive behaviours. The most common reactive behaviours
performed were smelling (Figure 2.16) (26 times by 14 individual elephants) and assuming the
vigilance posture (16 times by 7 individual elephants), both of which are used to gather information
about the surrounding environment (Bates et al. 2007; King et al. 2007). Nine elephants were also
seen holding their tails in a stiff upright position), which is a posture commonly seen before elephants
flee. A sex-bias in behaviour was also observed with only female elephants increasing the time spent
in locomotion following a disturbance and being more likely to walk or run back towards the forest.
Inside Wasgamuwa National Park, the effect of disturbance was less marked, with no change in
feeding or locomotion for either sex, and only females altering time spent standing still, which
decreased during the disturbance context. As at Weheragala Tank, specific reactive behaviours
increased, and again smelling was the most common response. However, results from Wasgamuwa
National Park should be treated cautiously, as only eight elephants were able to be scored for
behaviour during a disturbance, and the only opportunistic disturbance available was the approach of
tourist vehicles (Figure 2.17).

67
Figure 2.16. Elephant smells towards the research vehicle in Wasgamuwa National Park (low risk zone) ©
Kylie M. Butler
Figure 2.15. Vehicles and elephants in Wasgamuwa National Park (low risk zone). A tourist vehicle approaches
an elephant to within 20 m without stopping © Kylie M. Butler

68
Given that research observations were conducted from a vehicle, the elephants from our Wasgamuwa
National Park (low-risk zone) sample were likely biased to individuals who were more comfortable in
the presence of vehicles than others. Of the 54 elephant groups encountered in Wasgamuwa National
Park during the study period, 17 groups (31.5%) either fled into the forest as we approached, charged
at the car, or both. However, most fleeing or charging occurred before we were able to set up
recording equipment thus we were unable to collect behavioural data on individuals within these
groups. Observations from hides at stationary locations throughout the park would enable less biased
sampling however we did not have permission to leave the vehicle or establish such look-outs. Future
research perhaps in collaboration with park staff is encouraged.
Akin to what was witnessed at Weheragala Tank, interruptions to foraging as a response to
disturbance are common among species exposed to the physical presence of humans through wildlife
tourism or hunting (e.g. bears (White et al. 1999; Olson et al. 2007), deer (de Boer et al. 2004), dolphins
(Lusseau 2003), whales (Lusseau et al. 2009; Williams et al. 2006)), and sex-biases in response also
commonly occur (Bunnerfeld et al. 2006; Saj et al. 1999). For most of these species, the potential long-
term effects of altered foraging patterns include loss of body condition and reduced health and fitness
Figure 2.17. Elephants fleeing from the clearing towards the forest at Weheragala Tank © Kylie M. Butler

69
(Blumstein et al. 2005; Lacy and Martins 2003; Frid and Dill 2002; White et al. 2002; Anthony and
Blumstein 2000; Lima 1998). For elephants specifically, female investment in offspring is high with an
18 to 20-month gestation period, followed by up to four years before a calf is completely weaned, and
the nutritional intake required to support this is high (Sukumar 1989). Male elephants also have high
energetic requirements relating to their reproductive states and success (Sukumar 1989).
Consequently, disruptions to foraging patterns could have costly effects on the health and fitness of
both female and male elephants. It is difficult, if not impossible, however, to conclusively determine
at what intensity changes to foraging would have to be for such consequences would occur. Identifying
long-term changes in body condition or reproductive fitness would take years, if not decades to
properly quantify, and accounting for confounding variables (e.g. food availability, seasonal changes
in the environment, and fluctuations in anthropogenic pressure) would be extremely difficult in the
wild. Still the facts remain that Asian elephants are listed as endangered on the IUCN Red List of
Threatened Species (Choudhury et al. 2008), their natural habitat is under ongoing threat (Fernando
et al. 2014; Haturusinghe and Weerakoon 2012; Campos-Arceiz et al. 2009; de Silva and de Silva 2007;
Fernando et al. 2005), and human-elephant conflict is threatening their long-term survival in the wild
(Fernando et al. 2011; Campos-Arceiz et al. 2009; Fernando et al. 2005). Given that significant changes
to feeding behaviour were evident, as a precautionary measure it would be best-practice to reduce
anthropogenic disturbance where possible. Educating people about the dangers of deliberately
disturbing elephants (e.g. by driving towards them, revving engines, or throwing firecrackers while
elephants are feeding peacefully) and perhaps imposing penalties on those found doing so, would be
a good first step.
Of additional concern, is whether elephants who have previous exposure to negative interactions with
humans, will increase displays of aggressive behaviour towards humans, as has been suggested by
some researchers (Fernando et al. 2011; Perera 2009). Most human-elephant interactions at
Weheragala Tank are incidental, occurring as people travel to or from villages (for work, school or
markets), and elephants are foraging in the clearings or walking from the forest to the water tank.
These interactions usually involve either (or both) parties receiving a fright, and generally humans
have to wait patiently for elephants to move on or drive slowly past when it is safe to do so. However,
intentional anthropogenic disturbances, where people rev engines, drive off-road towards elephants,
or throw missiles, are not uncommon, and elephants often respond with threat displays or by charging
(researcher pers. obs.). If elephants were to associate all people with these types of disturbances,

70
consequences for innocent villagers could be dire. Fortunately, we found no evidence that elephants
with high levels of prior negative interaction with humans, were responding to the observed
disturbances more reactively than others. Still, our proxy of using wounds and scars to determine
elephant-human interaction levels, may be more relevant in the specific context where these wounds
were inflicted, which is most likely inside villages while crop-raiding. In the interest of facilitating a
more peaceful co-existence between humans and elephants, in a world where increased interactions
are inevitable, we advocate showing basic respect to wild elephants by not intentionally disturbing
them, regardless of whether disturbing them increases aggression by elephants to humans, or not.
2.6 Conclusion
This study provides initial evidence that elephants inhabiting areas of ‘medium’ level anthropogenic
disturbance outside of Wasgamuwa National Park boundaries are reducing time spent feeding, as a
response to immediate anthropogenic threats in the environment. That elephants increased specific
‘reactive’ behaviours, including smelling and assuming the vigilance posture when vehicles
approached, both inside and outside of Wasgamuwa National Park further indicates that certain
human actions are viewed as a threat or disturbance by the elephants.
Additionally, this study highlights some of the difficulties involved in assessing behaviours of wild
animals in a wild context, according to a perceived level of anthropogenic risk. Although theoretically
the distinction in anthropogenic risk present inside and outside of PA boundaries should be clear, we
found that irresponsible behaviour of people in tourist vehicles inside Wasgamuwa National Park
often mirrored the behaviour of people outside the park, thus potentially blurring the distinction
between ‘low’ and ‘medium’ risk zones. Furthermore, when relying on opportunistic disturbances, it
can be difficult to attain sufficient sample sizes however it is the only ethical way to attempt this.
Undoubtedly, the question of influences of anthropogenic disturbance on wild elephant behaviour is
an interesting one. However, the time required to venture beyond initial evidence supporting
hypotheses that elephants do indeed alter their behaviour according to human-influenced activity,
and the difficulties in managing confounding variables, are significant challenges to research in the
wild. Nonetheless, further studies examining wild elephant behaviour across gradients of human
disturbance are fundamental in understanding the effect of anthropogenic disturbance and activity

71
on elephants living in close cohabitation with humans. That said, management plans for locations
such as the Wasgamuwa National Park/Weheragala Tank area, cannot afford to wait until longer-term
studies are complete. Management plans, to reduce anthropogenic disturbance where possible and
facilitate a safer environment for both humans and elephants where the two co-exist, should be
developed with the input of local authorities, researchers and conservationists, and local
communities. Management plans can then be informed and adapted, as research into this most
interesting yet complicated area progresses.

72
2.7 References
Alahakoon, AMDB, Pushpakumara, EMAB, Ellepola, G & Ranawana, KB 2017, ‘Food and feeding
patterns of Asian elephants in Uda Walawe National Park, Sri Lanka’, Gajah, vol. 46, pp. 4 – 13.
Amwata, DA & Mganga, KZ 2014, 'The African elephant and food security in Africa: experiences from
Baringo District, Kenya', Pachyderm, vol. 55, pp. 22 – 28.
Anthony, LL & Blumstein, DT 2000, 'Integrating behaviour into wildlife conservation: the multiple ways
that behaviour can reduce Ne', Biological Conservation, vol. 95, pp. 303 – 315.
Bandara, R & Tisdell, C 2002, 'Asian elephants as agricultural pests: economics of control and
compensation in Sri Lanka', Natural Resources Journal, vol. 42, pp. 491 – 519.
Barnes, RFW, Dubiure, UF, Danquah, E, Boafo, Y, Nandjui, A, Hema, EM & Manford, M 2006, 'Crop-
raiding elephants and the moon', African Journal of Ecology, vol. 45, pp. 112-115.
Bates, LA, Sayialel, KN, Njiraini, NW, Moss, CJ, Poole, JH and Byrne, RW 2007, 'Elephants classify human
ethnic groups by odor and garment colour', Current Biology, vol. 17, no. 22,
doi:10.1016/j.cub.2007.09.060.
Benhaiem, S, Delon, M, Lourtet, B, Cargnelutti, B, Aulagnier, S, Hewison, AJM, Morellet, N &
Berheyden, H 2008, 'Hunting increases vigilance levels in roe deer and modified feeding site selection',
Animal Behaviour, vol. 76, pp. 611-618.
Blumstein, DT, Fernandez-Juricic, E, Zollner, PA & Garity, SC 2005, ‘Inter-specific variation in avian
responses to human disturbance’, Journal of Applied Ecology, vol. 42, no. 5, pp. 943 – 953.
Bunnefeld, N, Linnell, JDC, Odden, J, van Duijin, MAJ & Andersen, R 2006, 'Risk taking by Eurasian lynx
(Lynx lynx) in a human-dominated landscape: effects of sex and reproductive status', Journal of
Zoology, doi: 10.1111/j.1469-7998.2006.00107.x.

73
Burn, RW, Underwood, FM & Blanc, J 2011, 'Global trends and factors associated with the illegal killing
of elephants: a hierarchical Bayesian analysis of carcass encounter data', PLoS ONE, vol. 6, no. 9,
e24165. doi:10.1371/journal.pone.0024165.
Campos-Arceiz, A, Takatsuki, S, Ekanayaka SKK & Hasegawa, T 2009, ‘The human-elephant conflict in
Southeastern Sri Lanka: Type of damage, seasonal patterns, and sexual differences in the raiding
behaviour of elephants’, Gajah, vol. 31, pp. 5 – 14.
Caro, TM 2005, 'Behavioural indicators of exploitation', Ethology, Ecology and Evolution, vol. 17, no.
2, pp. 189-194.
Caro, TM 1999, 'Demography and behaviour of African mammals subject to exploitation', Biological
Conservation, vol. 91, no. 1, pp. 91 – 97.
Chakraborty, K & Mondal, J 2013, 'Perceptions and patterns of human-elephant conflict at Barjora
block of Bankura district in West Bengal, India: insights for mitigation and management',
Environmental Development and Sustainability, vol. 15, pp. 547 – 565.
Choudhury, A 2004, 'Human-elephant conflicts in northeast India', Human Dimensions of Wildlife, vol.
9, no. 4, pp. 261 – 270.
Crosmary, WG, Makumbe, P, Cote, SD & Fritz, H 2012, 'Vulnerability to predation and water
constraints limit behavioural adjustments of ungulates in response to hunting risk', Animal Behaviour,
vol. 83, pp. 1367-1376.
Davidson, Z, Valiex, M, Van Kesteren, F, Loveridge, AJ, Hunt, JE, Murindagomo, F & Macdonald, DW
2013, ‘Seasonal diet and prey preference of the African lion in a waterhole-driven semi-arid savannah’,
PLoS ONE, vol. 8, no. 2, e55182. https://doi.org/10.1371/journal.pone.0055182.
Davies, TE, Wilson, S, Hazarika, N, Chakrabaty, J, Das, D, Hodgson, DJ & Zimmerman, A
2011,'Effectiveness of intervention methods against crop-raiding elephants', Conservation Letters,
doi: 10.1111/j.1755-263X.2011.00182.x

74
de Boer, HY, van Breikelen, L, Hootsmans, MJM & van Wieren, SE 2004, 'Flight distance in roe deer
Caperoulus capreolus and fallow deer Dama dama as related to hunting and other factors', Wildlife
Biology, vol. 10, no. 1, pp. 35 – 41.
de la Torre, S, Snowdon, CT & Bejarano, M 2000, ‘Effects of human activities on wild pygmy marmosets
in Ecuadorian Amazon’, Biological Conservation, vol. 94, pp. 153 – 163.
de Silva, M & de Silva, PK 2007, The Sri Lankan elephant: Its evolution, ecology and conservation, WHT
Publications, Colombo.
Donadio, E and Buskirk, SW 2006, 'Flight behaviour in guanacos and vicunas in areas with and without
poaching in western Argentina', Biological Conservation, vol. 127, pp. 139-145.
Ekanayaka, SKK, Campos-Arceiz, A, Rupasinghe, M, Pastorini, J & Fernando, P 2011, 'Patterns of crop
raiding by Asian elephants in a human-dominated landscape in southeastern Sri Lanka', Gajah, vol. 34,
pp. 20 -25.
Fernando, P, de Silva, MKCR, Jayasinghe, LKA, Janaka, HK & Pastorini, J 2019, ‘First country-wide survey
of the endangered Asian elephant: toward better conservation and management in Sri Lanka’, Oryx,
pp. 1 – 10, https://doi.org/10.1017/s0030605318001254 .
Fernando, P, Jayewardene, J, Prasad, T, Hendavitharana, W & Pastorini, J 2011, ‘Current status of Asian
Elephants in Sri Lanka’, Gajah, vol. 25, pp. 93 – 103.
Fernando, P, Wikramanayake, E, Weerakoon, D, Jayasinghe, LKA, Gunawardene, M & Janaka, HK 2005,
'Perceptions and patterns of human-elephant conflict in old and new settlements in Sri Lanka: insights
for conservation and management', Biodiversity and Conservation, vol. 14, pp. 2465 – 2481.
Freeman, HD & Gosling, SD 2010, ‘Personality in non-human primates: a review and evaluation of past
research’, American Journal of Primatology, vol. 72, pp. 653 – 671.
Frid, A & Dill, L 2002, 'Human-caused disturbance: stimuli as a form of predation risk', Conservation
Ecology; vol. 6 (1), http://www.consecol.org/vol6/iss1.art11.

75
Graham, MD, Notter, B, Adams, WM, Lee, PC & Ochieng, TN 2010, 'Patterns of crop-raiding by
elephants, Loxodonta africana, in Laikipia, Kenya, and the management of human-elephant conflict',
Systematics and Biodiversity, vol. 8, no. 4, pp. 435 – 445.
Graham, MD, Douglas-Hamilton, I, Adams, WM & Lee, PC 2009, 'The movement of African elephants
in a human-dominated land-use mosaic', Animal Conservation, vol. 12, pp. 445-455.
Griffin, SC, Valois, T, Taper, ML & Mills, LS 2007, 'Effects of tourists on behaviour and demography of
Olympic marmots', Conservation Biology, vol. 21, no. 4, pp. 1070-1081.
Gubbi, S 2012, 'Patterns and correlates of human-elephant conflict around a south Indian reserve',
Biological Conservation, vol. 148, pp. 88 – 95.
Gunn, J, Hawkins, D, Barnes, RFW, Mofulu, F, Grant, RA & Norton, GW 2013, ‘The influence on lunar
cycles on crop-raiding elephants: evidence for risk avoidance’, African Journal of Ecology, vol. 52, pp.
129 – 137.
Haturusinghe, HS & Weerakoon, DK 2012, ‘Crop raiding behaviour of elephants in the Northwestern
region of Sri Lanka’, Gajah, vol. 36, pp. 26 – 31.
Hollander, FA, van Overveld, T, Tokka, I & Matthysen, E 2008, ‘Personality and nest defence in the
great tit (Parsus major)’, Ethology, vol. 114, pp. 405 – 412.
Ihwagi, F, Thouless, C, Wang, T, Skidmore, AJ, Omondi, P & Douglas-Hamilton, I 2018, 'Night-day speed
ratio of elephants as indicator of poaching levels', Ecological Indicators, vol. 84, pp. 38 – 44.
Kahindi, O, Wittemyer, G, King, J, Ihwagi, F, Omondi, P & Douglas-Hamilton, I 2009, 'Employing
participatory surveys to monitor the illegal killing of elephants across diverse land uses in Laikipia-
Samburu, Kenya', African Journal of Ecology, doi: 10.1111/j.1365-2028.2009.01.200.x.
Kiffner, C, Kioko, J, Kissui, B, Painter, C, Serota, M, White, C & Yager, P 2014, 'Interspecific variation in
large mammal responses to human observers along a conservation gradient with variable hunting
pressure', Animal Conservation, doi: 10.1111/acv.12131.

76
King, LE, Douglas-Hamilton, I & Vollrath, F 2007, ‘African elephants run from the sound of disturbed
bees’, Current Biology, vol. 17, pp. R832 – R833.
Kumaraguru, A, Brinda, K & Asokan, S 2011, ‘Prey preference of large carnivores in Anamalai Tiger
Reserve India’, vol. 57, European Journal of Wildlife, pp. 627 – 637.
Lacy, KE & Martins, EP 2003, ‘The effect of anthropogenic habitat usage on the social behaviour of
vulnerable species, Cychura nubila’, Animal Conservation, vol. 6, pp. 3 – 9.
Lima, SL 1998, 'Nonlethal effects in the ecology of predator-prey interactions', Bioscience, vol. 48, no.
1, pp. 25 – 34.
Lima, SL and Dill, LM 1990, 'Behavioral decisions made under the risk of predation: a review and
prospectus', Canadian Journal of Zoology, vol. 68, pp. 619-640.
Loveridge, AJ, Hunt, HE, Murindagomo, F & Macdonald, DW 2006, ‘Influence of drought on predation
of elephant (Loxodonta africana) calves by lions (Panthera leo) in an African wooded savannah’,
Journal of Zoology, vol. 2770, no. 3, pp. 523 – 530.
Lusseau, D, Bain, DE, Williams, R & Smith, JC 2009, 'Vessel traffic disrupts the foraging behaviour of
southern resident killer whales Orcinus orca', Endangered Species Research, vol. 6, pp. 211-221.
Lusseau, D 2003, 'Male and female bottlenose dolphins Tursiops spp. have different strategies to avoid
interactions with tour boats in Doubtful Sound, New Zealand', Marine Ecology Progress Series, vol.
257, pp. 267 – 274.
Mattson, DJ, Blanchard, BM & Knight, RR 1992, ‘Yellowstone grizzly bear mortality, human
habituation, and whitebark pine seed crops’, The Journal of Wildlife Management, vol. 56, no. 3, pp.
432 – 442.
McKay, GM 1973, ‘Behaviour and ecology of the Asiatic Elephants in Southeastern Ceylon’,
Smithsonian Contributions to Zoology, vol. 125, Smithsonian Institution Scholarly Press, Washington,
D.C.

77
Okello, MM, Kiringe, JW, Njumbi, SJ and Isiche, J 2016, 'Prevalence of human-elephant conflicts in
Amboseli ecosystem, Kenya: current opinions of local communiy', International Journal of Biodiversity
and Conservation, vol. 8, no. 3, pp. 60 – 71.
Olson, TL, Gilbert, BK & Squibb, RC 1997, ‘The effects of increasing human activity on brown bear use
of an Alaskan river’, Biological Conservation, vol. 82, pp. 95 – 99.
Perera, BMAO 2009, 'The human-elephant conflict: a review of current status and mitigation
methods', Gajah, vol. 1, pp. 341 – 344.
Ponnusamy, V, Chackrapani, P, Lim, TW, Saaban, S and Campos-Arceiz, A 2016, 'Farmers' perceptions
and attitudes towards government constructed electric fences in Peninsular Malaysia', Gajah, vol. 45,
pp. 4 – 11.
Power, RJ & Compion, RXS 2009, ‘Lion predation on elephants in the Savuti, Chobe National Park,
Botswana’, African Zoology, vol. 44, no. 1, pp. 36-44.
R Core Team 2018, R: A language and environment for statistical computing, R Foundation for
Statistical Computing, Austria.
Rangarajan, M 1996, ‘The politics of ecology: the debate on wildlife and people in India 1970 – 1995’,
Economics of Politics Weekly, vol. 31, pp. 2391 – 2409.
Saj, T, Sicotte, P & Paterson, JD 1999, ‘Influence of human food consumption on the time budget of
vervets’, International Journal of Primatology, vol. 20, pp. 977 – 994.
Santiapillai, C, Fernando, P and Gunewardene, M 2006, 'A strategy for the conservation of the Asian
elephant in Sri Lanka', Gajah, vol. 25, pp. 91 – 102.
Senevirathna, HMMC & Perera, PKP 2013, ‘Wildlife viewing preferences of visitors to Sri Lanka’s
National Parks: implications for visitor management and sustainable tourism planning’, Journal of
Tropical Forestry and Environment, vol. 3, no. 2, pp. 1 – 10.

78
Setsaas, TH, Holmern, T, Mwakalebe, G, Stokke, S & Roskaft, E 2007, 'How does human exploitation
affect impala populations in protected and partially protected areas? – A case study from the Serengeti
Ecosystem, Tanzania', Biological Conservation, vol. 136, pp. 563-570.
Sitati, NW & Walpole, MJ 2006, 'Assessing farm-based measures for mitigation of human-elephant
conflict in Transmara District, Kenya', Oryx, vol. 40, no. 3, pp. 279 – 286.
Sitati, NW, Walpole, MJ, Smith, RJ & Leader-Williams, N 2003, ‘Predicting spatial aspects of human-
elephant conflict’, Journal of Applied Ecology, vol. 40, pp. 667 – 677.
Soltis, J, King, LE, Douglas-Hamilton, I, Vollrath, F & Savage, A 2014, ‘African elephant alarm calls
distinguish between threats from humans and bees’, PLoS ONE, vol. 9, no. 2, e89403.
doi:10.1371/journal.pone.0089403.
Steidl, RJ and Anthony, RG 2000, ‘Experimental effects of human activity on breeding bald eagles’,
Ecological Applications, vol. 10, no. 1, pp. 258 – 268.
Sukumar, R 1989, The Asian elephant: ecology and management, Cambridge University Press, USA.
Sukumar, R & Gadgil, M 1988, ‘Male-female differences in foraging on crops by Asian elephants’,
Animal Behaviour, vol. 36, no. 4, pp. 1233 – 1235.
Thuppil, V and Goss, RG 2013, 'Wild Asian elephants distinguish aggressive tiger and leopard growls
according to perceived danger' Biology Letters, vol. 9, 20130518,
http://dx.doi.org/10.1098/rsbl.2013.0518.
Vancuylenberg, BWB 1977 ‘Feeding behaviour of the Asiatic elephant in South-East Sri Lanka in
relation to conservation’, Biological Conservation, vol. 12, no. 1, pp. 33 – 54.
Webber, CE, Sereivathana, T, Maltby, MP and Lee, PC 2011, 'Elephant crop-raiding and human-
elephant conflict in Cambodia: crop selection and seasonal timing of raids', Oryx, vol. 45, no. 2, pp.
243 – 251.

79
Weerakoon, DK, Gunawardene, MD, Janaka, HK, Jayasinghe, LKA, Perera, RAR, Fernando, P and
Wikramanayake, E 2004, 'Ranging behaviour and habitat use of elephants in Sri Lanka', in: (ed)
Jayewardene, J Endangered Elephants: Past, present and future, Proceedings of the symposium for
human-elephant relationships and conflicts, Sri Lanka, September 2003. Biodiversity and Conservation
Trust, Colombo, pp. 68 - 70.
Wilkie, RD & Douglas-Hamilton, I 2018, ‘High-resolution tracking technology reveals distinct patterns
in nocturnal crop-raiding behaviour of an African elephant (Loxodonta africana) in Amboseli, Kenya’,
Pachyderm, vol. 59, pp. 41 – 48.
Williams, RM 1994, Behavioural responses of killer whales to whale-watching: opportunistic
observations and experimental approaches, Masters Thesis, Department of Zoology, University of
British Colombia.
White, D, Kendall, KC and Picton, HD 1999, 'Potential energetic effects of mountain climbers on
foraging grizzly bears', Wildlife Society Bulletin, vol. 27, no. 1, pp. 146-151.

80
Chapter 3: Crop-raiding in a high human-elephant conflict location in Sri Lanka: an assessment of the characteristics and patterns of crop-raiding in Dewagiriya Village

81
3.1 Abstract
Crop-raiding is a major form of human-elephant conflict, threatening the long-term viability of
elephant populations and the livelihoods of communities living in proximity to both Asian and African
elephants. In Sri Lanka, crop raiding has been identified as a predominant threat to elephant
conservation. We conducted an intensive three-year study into the crop-raiding patterns and
predictors of elephants entering a small rural village called Dewagiriya on the outskirts of Wasgamuwa
National Park. We found crop-raiding to be prevalent, occurring year-round with no clear seasonal
patterns. Male elephants either alone or in small groups were the most likely to crop-raid, and almost
exclusively at nights. Homes and crop fields closest to the water tanks and forest edges were most
vulnerable to elephant damage. We suggest that to best protect the community from elephant
damage, crop-guarding efforts be concentrated in these ‘higher’ risk areas.
3.2 Introduction
The facilitation of peaceful co-existence between humans and wildlife is an ongoing challenge for
conservation scientists, wildlife managers and communities worldwide (Goswami and Vasudev 2017).
In many locations, agriculture and development has destroyed or fragmented traditional wildlife
habitat. A reduction of suitable habitat in conjunction with human population growth and more
people being encouraged to settle near wildlife areas, has resulted in intensifying levels of human-
wildlife conflict (HWC) over resources including space, water and food (Puyravaud et al. 2019;
Manfredo 2015; Harich et al. 2013; Redpath 2013).
Crop-raiding, where wildlife forages on, or otherwise damages, agricultural crops, is a major source of
HWC (Saraswat et al. 2015; Chakraborty and Modal 2013; Ekanayake et al. 2011; Thapa 2010), and
sadly it is often the most vulnerable human and wildlife communities that are most negatively
impacted by this. In many areas where crop-raiding is a predominant issue, local communities are of
low socio-economic status relative to the rest of the country, and people struggle with the daily
realities of living alongside certain wildlife species (Barua et al. 2013; Ogra 2008; Rao et al. 2002). This
is particularly so for families who are reliant upon income generated from farming for basic living
expenses (e.g. health and education) and where the crops they grow are their main source of food
(Barua et al. 2013; Ogra 2008; O'Connell-Rodwell et al. 2000).

82
On the other side, although animals of varied species, shapes and sizes are known to cause havoc by
crop-raiding, those which are large-bodied and capable of inflicting physical injuries on humans, are
typically at the forefront of HWC concerns. Examples include both African and Asian elephants,
Loxodonta africana and Elephas maximus, (Hoffmeier-Karimi and Schulte 2014; Santiapillai et al.
2006); Indian rhinoceros, Rhinoceros unicornis, (Thapa 2010) and several primate species (e.g.
chimpanzees, Pan troglodytes: McLennan and Hill 2012; Mountain gorillas, Gorilla beringei: Hockings
and Hunle 2009; Sumatran orang-utans, Pongo abelii: Campbell-Smith et al. 2010). All these
aforementioned species are globally recognised as requiring protection to ensure their long-term
survival in the wild (IUCN Red List of Threatened Species), and unfortunately HWC (specifically crop-
raiding) presents yet another challenge to their future existence.
Crop-raiding by both African and Asian elephants has been a hot topic of conservation and
humanitarian concern for several decades, yet shows little sign of abating (Davies et al. 2011; Perera
2009; Sitati and Walpole 2006). Globally hundreds of human and elephant injuries and deaths are
reported each year (Thirgood et al. 2005), and farmers continue to experience severely reduced crop
yields and damage to property as a result of elephant visits (Okello et al. 2016; Amwata and Mganga
2014; Gubbi 2012; Ekanayaka et al. 2011; Campos-Arceiz et al. 2009). As farmers become more
desperate to defend their livelihoods, they often resort to more violent and harmful means of
deterring elephants, which can perpetuate the problem they are trying to alleviate. In Asia, where an
estimated 95% of traditional elephant habitat has been lost primarily to agriculture (Sukumar 2003;
2006; Hedges et al. 2005), human-elephant conflict (HEC) influenced by crop-raiding is recognised as
the biggest threat to the survival of wild elephant populations (Perera 2009; Sukumar 2006; Fernando
et al. 2005; Hedges et al. 2005; Leimgruber et al. 2003).
This complicated scenario is perhaps no better exemplified than by the crop-raiding issues identified
in Sri Lanka. With an estimated elephant population of over 5,800 wild elephants, Sri Lanka is home
to more than 10% of the global wild Asian elephant population and has the highest density of Asian
elephants (Fernando et al. 2011). Although Sri Lanka has more than 8,500 km² (13%) of land
designated as Wildlife Protected Area's (PA's) (DWC 2017), attempts to restrict wild elephants within
PA's have largely failed (Fernando et al. 2011; 2005) and it is not uncommon for elephants to be found
in higher densities outside of PA's with the majority of elephants spending a proportion of their time
in close proximity to rural communities (Fernando et al. 2011; Campos-Arceiz et al. 2009). Elephant

83
inhabited areas, including those on the periphery of villages, are being decimated by ongoing
deforestation for development, including dams and agricultural expansion (Fernando et al. 2014;
Haturusinghe and Weerakoon 2012; Campos-Arceiz et al. 2009; de Silva and de Silva 2007; Fernando
et al. 2005). This means that humans and elephants are being forced to meet their often overlapping
resource needs in increasingly close proximity. Consequently, crop-raiding appears to be worsening
and is having an overwhelming negative influence on the lives of many farmers, with the resulting
conflict the principle threat to the survival of Sri Lanka's elephants (Fernando et al. 2011; Campos-
Arceiz et al. 2009; Fernando et al. 2005).
Understanding how, when and why some elephants crop-raid is essential to predicting the
vulnerability of farms to crop depredation, and to identifying and implementing appropriate
mitigation strategies (Webber et al. 2011; Sitati et al. 2005). Predicting factors vary, however across
the vast global spatial scale in which elephant crop-raiding occurs, some general patterns have
emerged, and these can provide a useful comparative basis when investigating specific crop-raiding
conflict situations.
In many locations throughout Asia, elephants raid crops year-round with fruits (e.g. banana, coconut,
watermelon), vegetables (e.g. eggplant, pumpkin, sweet potato), legumes and grains (e.g. cowpeas,
maize, mung beans) and sugar cane commonly consumed (Chen et al. 2016; Ekanayaka et al. 2011;
Fernando et al. 2011; Webber et al. 2011; Barnes et al. 2005). Annually, seasonal variations in
frequency and intensity of crop-raiding are common, usually peaking towards the end of rain seasons
when major crops such as rice or maize are maturing (Chen et al. 2016; Das et al. 2014; Ekanayaka et
al. 2011; Webber et al. 2011). Within the 24-hour daily cycle, crop-raiding occurs almost exclusively at
night (Das et al. 2014; von Gerhardt et al. 2014; Graham et al. 2010; Sitati et al. 2003) yet typical
elephant feeding patterns show that the majority of foraging occurs during hours of daylight (Sukumar
2006). This tendency to crop-raid nocturnally indicates that elephants possess an awareness of the
risks associated with crop-raiding and adjust their behaviour accordingly (Gunn et al. 2013; Graham
et al. 2009, Barnes et al. 2006). Further supporting this risk-avoidance theory, at multiple locales in
Africa, the light afforded by lunar phase was found to influence crop-raiding activity. During new lunar
phases when nights are at their darkest, the occurrence of crop-raiding increased, and during full
moon phases when night-time visibility is greatest, crop-raiding decreased (Gunn et al. 2013; Barnes
et al. 2006). This has yet to be investigated in Asia.

84
Although both male and female elephants are often found outside of PA's and in proximity to villages,
sex-biases in crop-raiding are evident, with male elephants raiding far more frequently than females,
and typically doing so alone or in small bachelor groups (Thaufeek et al. 2014; Das et al. 2012; Sukumar
and Gadgil 1988). This suggests that in most locations, male elephants are more willing than female
elephants to exploit crops as part of a high-risk, high-gain foraging strategy (Sukumar and Gadgil 1988).
In Sri Lanka, where the importance of mitigating HEC is well-recognised, only seven peer-reviewed
papers specifically investigating predictors and patterns of crop-raiding behaviour at various conflict
locations throughout the country, have been published since 2002 (see Thaufeek et al. 2014;
Haturusinghe and Weerakoon 2012; Ekanayaka et al. 2011; Campos-Arceiz et al. 2009; Fernando et al.
2005; Wikramanayake et al. 2004; Bandara and Tisdell 2002). To the best of my knowledge, these
studies provide the only peer-reviewed data from which to compare crop-raiding characteristics in Sri
Lanka with other HEC zones globally, and for highlighting commonalities and differences in crop-
raiding within Sri Lanka. However, given the likelihood of a continuing increase in HEC throughout the
country (Fernando et al. 2011; Campos-Arceiz et al. 2009; Fernando et al. 2005), further research from
a wider array of conflict sites would contribute to the establishment of effective mitigation methods
by enhancing our understanding of the influencing factors and patterns of crop-raiding behaviour.
Almost all food crops grown in Sri Lanka are vulnerable to elephant damage (Thaufeek et al. 2014;
Haturusinghe and Weerakoon 2012), and at all sites assessed except one, various crops were subject
to some degree of elephant depredation throughout the entire year (Thaufeek et al. 2014;
Haturusinghe and Weerakoon 2012; Ekanayaka et al. 2011; Campos-Arceiz et al. 2009;
Wikramanayake et al. 2004; Bandara and Tisdell 2002). Rice, which is the country's primary agricultural
crop, was the most heavily targeted, and seasonal peaks in overall raiding coincided with the
approaching maturation of rice crops, typically beginning towards the end of the main rain season in
January (Thaufeek et al. 2014; Haturusinghe and Weerakoon 2012; Ekanayaka et al. 2011; Campos-
Arceiz et al. 2009; Fernando et al. 2005; Wikramanayake et al. 2004; Bandara and Tisdell 2002). At two
locations, a second crop-raiding peak was observed in August. These locations were in south-eastern
Sri Lanka and during this peak, permanent crops such as bananas, were the most frequently consumed
(Ekanayaka et al. 2011; Campos-Arceiz et al. 2009). High levels of attacks on houses, presumably so
elephants could access stored food inside, were evident in north-western, south-eastern and central

85
Sri Lankan conflict zones (Thaufeek et al. 2014; Haturusinghe and Weerakoon 2012; Campos-Arceiz et
al. 2009).
As with crop-raiding activity in other Asian and African locations, elephants in Sri Lanka used this
foraging strategy nocturnally and were rarely, if ever, seen in or approaching fields during daylight
hours (Campos-Arceiz et al. 2009; Wikramanayake et al. 2004; Bandara and Tisdell 2002). Male
elephants, either alone or in small all-male groups, were responsible for the majority of crop-raids,
and groups containing adult females raided far less frequently (Thaufeek et al. 2014; Haturusinghe
and Weerakoon 2012; Ekanayaka et al. 2011; Bandara and Tisdell 2002). Still, sex-biases in raiding
behaviour were not uniform across locations. At one site in north-central Sri Lanka, both male
elephants and female-led groups raided frequently, whereas at another site in southern Sri Lanka no
females at all were identified as raiders (Wikramanayake et al. 2004).
The Sri Lankan studies described above demonstrate broad patterns in targeted crop types, seasonal
and diurnal crop-raiding peaks, general sex-biases and group types of raiding elephants, yet also show
that even within a relatively small country, site-specific differences in crop-raiding do occur.
Furthermore, crop-raiding patterns or characteristics can differ within even an individual conflict
location. Very few studies in Africa or Asia provide an in-depth account of within-site variations (but
see: Chen et al. 2016; Barnes et al. 2005; Sitati et al. 2005), yet factors such as the proximity of a farm
to forest habitat or water sources, number of neighbouring farms or homes, plus socio-demographic
and economic factors, can all influence a farms vulnerability to crop-raiding (Chen et al. 2016; Sitati et
al. 2005; Hill 2004).
To properly identify the driving factors and likely consequences of crop-raiding, one cannot simply
look at patterns or predictors from other conflict zones, even within the same country or district. Such
information can provide a useful platform to build upon, however if the goal is to help communities
successfully alleviate HEC, the importance of identifying site-specific and within site characteristics
cannot be understated (Barnes 2009; Hill 2004).
In this study, we provide an in-depth investigation into the patterns and predictors of crop-raiding in
a single conflict location within a high HEC region of Sri Lanka. Specifically, we aimed to:

86
- quantify the frequency of elephant sightings in fields and home gardens and to describe the
patterns of damage incurred
- identify temporal patterns in crop-raiding at seasonal, lunar and diurnal scales
- determine sex-biases in raiding behaviour, as well as group type and size
Furthermore, we investigated whether significant within-site variations were present and the
potential spatial (e.g. location of field/home in relation to main road and forest periphery/water
tanks), socio-demographic and economic (e.g. household size, income) factors influencing these.
Information generated from this study will help to predict vulnerability of crops and homes to
elephants throughout the overall site and at individual farms. This can be used to develop mitigation
strategies tailored to specific temporal and spatial variations, hopefully encouraging the concentration
of oft-limited resources where they are most required, and will be most effective, for individual
farmers and the overall community. Additionally, we consider our findings in light of patterns and
predictors identified in existing crop-raiding studies in Sri Lanka and other affected areas. The more
information procured for a wider array of individual conflict locations, the more meaningful cross-site
comparisons will become in identifying and developing site-appropriate and targeted mitigation
strategies, based on commonalities and differences.
3.3 Methods
3.3.1 Study site
This study was conducted in the rural community of Dewagiriya Village, located in the dry zone of
Central Sri Lanka (Figure 3.1). Overall, the dry zone encompasses approximately 60%, or almost 40,000
km², of Sri Lanka (Fernando et al. 2011), spanning the north to central provinces then stretching south
down the east coast. The dry zone typically experiences annual rainfall between 1650 – 2100 mm, and
is characterised by two wet seasons: the 'long rains' from October to January, and the 'short rains'
from March to May (Fernando et al. 2005; Wikramanayake et al. 2004). The driest time of the year is
between July and September. The average annual temperature is 28°C, with little variation throughout
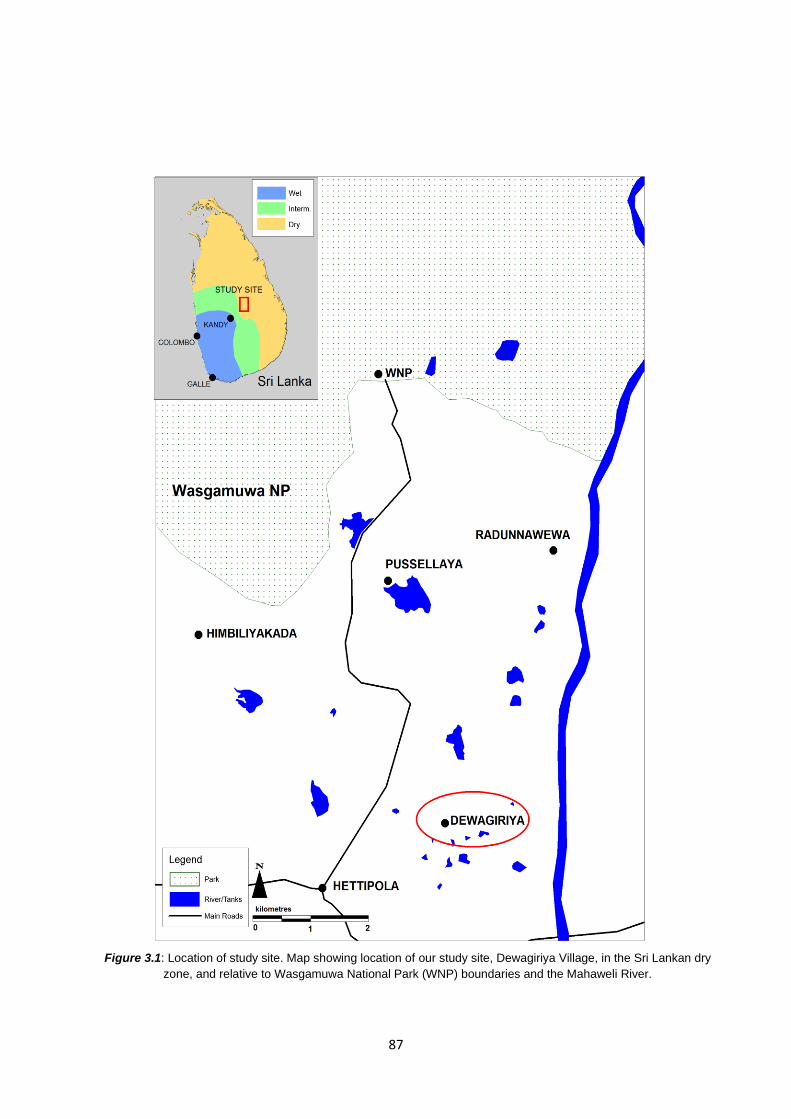
87
Figure 3.1: Location of study site. Map showing location of our study site, Dewagiriya Village, in the Sri Lankan dry
zone, and relative to Wasgamuwa National Park (WNP) boundaries and the Mahaweli River.

88
the year (de Silva and de Silva 2007). Wild elephants inhabit almost this entire zone, and are commonly
found both in and outside of designated PA's (Fernando et al. 2011).
Since the mid 1900's, the environment of the north-central and central provinces has been
dramatically altered by large scale development (Wikramanayake et al. 2004). Major hydro-projects,
particularly along the Mahaweli River, have seen the construction of many dams and reservoirs, and
facilitated the widespread expansion of irrigated agriculture. Large tracts of forest, which were
historically prime elephant habitat, have been cleared and converted into agricultural fields (de Silva
and de Silva 2007; Fernando et al. 2005). As the forests diminish, elephants are forced into smaller
areas of suitable habitat. At the same time, new human communities are being established and farms
are expanding closer to both the remaining forest and PA boundaries (de Silva and de Silva 2007).
Many of these projects continue today and increasing overlap of human and elephant communities is
inevitable.
The specific study location of Dewagiriya Village (07°33.928' N, 080°56.866' E) was established in 1958
(A. Dingira, pers. comm.), and is situated approximately ten kilometres below the southern boundary
of Wasgamuwa National Park, and two kilometres to the west of the Mahaweli River (Figure 3.1).
Although exact population numbers are unknown, at least 400 elephants inhabit Wasgamuwa
National Park and/or utilise the numerous forest patches, river banks and small reservoirs, in between
the southern park boundaries and Dewagiriya Village (researcher pers. obs. 2014 - 2016).
Covering an area of approximately 2 km², Dewagiriya village is home to around 40 permanent
households, with houses and fields scattered on either side of a dirt road running east to west through
the centre of the village. Four semi-permanent man-made reservoirs (known as ‘tanks’) are located in
the south area and are bordered by forest (Figure 3.2). These forests provide an excellent, albeit small,
habitat for elephants who often seek refuge there during the day, moving into the village to access
water sources and crops at night (BKG. Jayasena, pers. comm.).
The community is of a low socio-economic status and individual small-scale agriculture is the main
livelihood. Most families have a small garden next to their house of approximately ¼ acre where fruits
trees (e.g. banana, coconut, papaya), and small vegetable plots (e.g. long beans, eggplant, okra) are
grown. Each family also has an adjoining field, typically between one to three acres although

89
neighbouring fields often border each other, making the overall continuous field area much larger.
Rice is the predominant crop type cultivated, and is used for both subsistence and income generation.
Almost every family plants rain-fed rice fields during the 'long rains' and those with access to irrigated
water supplies will often plant a second rice crop during the 'short rains'.
At an initial community meeting I attended in June 2014, the 20 participants unanimously agreed that
HEC is a pressing issue for the entire community. Given the villages proximity to elephant habitat and
water sources, and the communities dependence on rice cultivation – a frequently raided crop in many
high HEC zones worldwide – Dewagiriya Village was deemed an ideal location for a long-term,
intensive investigation into the characteristics and patterns of crop-raiding.
3.3.2 Data Collection
This study was conducted over a three-year period from January 01st 2015 to December 31st 2017,
using resident reports of 'elephant events' collected during frequent home visits. I defined an
'elephant event' as any sighting of an elephant within the village, as well as any successful crop raids
that occurred.
There were a total of 40 households residing in the village at the conclusion of the study, however
four of these families had moved in part way through, one family lived in the village only during crop
growing seasons, and a further eight families were not home or contactable often enough to provide
consistent data. A household was deemed as not providing consistent data if I was not able to
interview them for a consecutive period of three months or more at any stage of the data collection
period. These thirteen households were excluded from analysis, leaving 27 participating households
scattered throughout the village (Figure 3.2).
Initially I distributed data sheets (translated into Sinhalese) to each household and trained an adult
family member how to answer each question. Each household was then visited every two to four
weeks to collect data sheets, discuss any elephant visits reported, and distribute new data sheets.
However, it was soon found that the majority of households failed to fill out their data sheets and
instead preferred to relay information directly during visits. To accommodate for this preference by
households, myself or my research assistants would scribe the participant's information directly onto
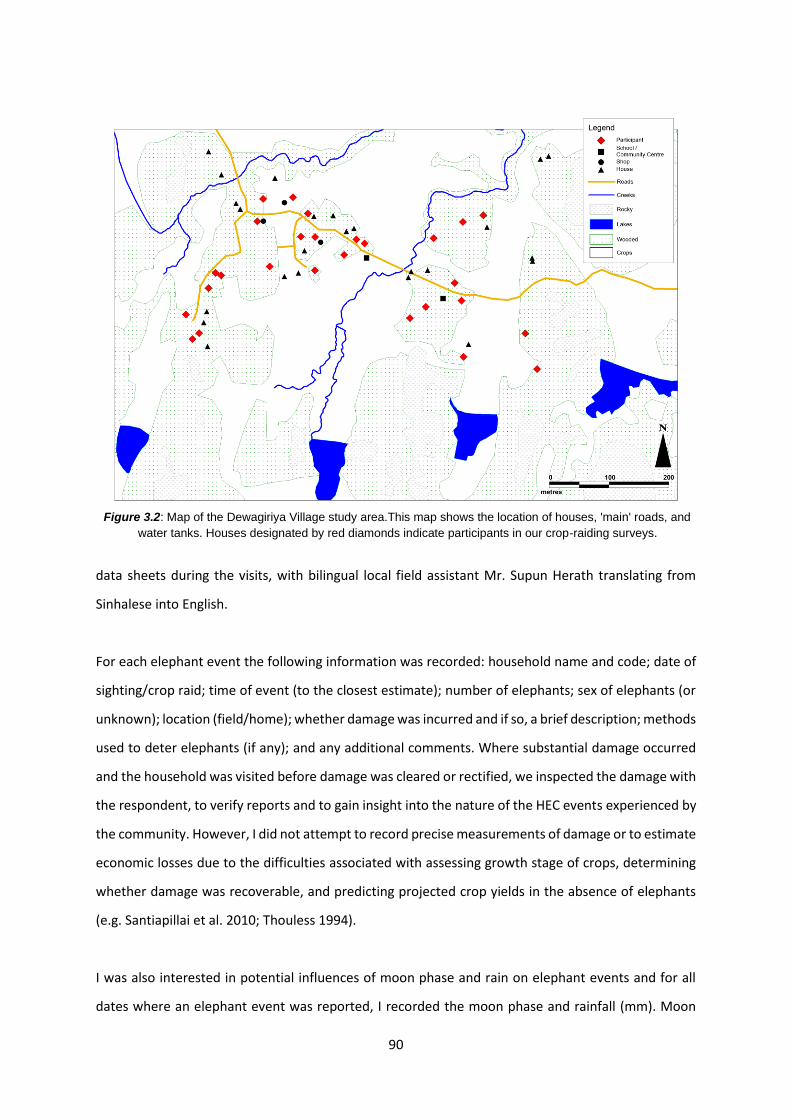
90
data sheets during the visits, with bilingual local field assistant Mr. Supun Herath translating from
Sinhalese into English.
For each elephant event the following information was recorded: household name and code; date of
sighting/crop raid; time of event (to the closest estimate); number of elephants; sex of elephants (or
unknown); location (field/home); whether damage was incurred and if so, a brief description; methods
used to deter elephants (if any); and any additional comments. Where substantial damage occurred
and the household was visited before damage was cleared or rectified, we inspected the damage with
the respondent, to verify reports and to gain insight into the nature of the HEC events experienced by
the community. However, I did not attempt to record precise measurements of damage or to estimate
economic losses due to the difficulties associated with assessing growth stage of crops, determining
whether damage was recoverable, and predicting projected crop yields in the absence of elephants
(e.g. Santiapillai et al. 2010; Thouless 1994).
I was also interested in potential influences of moon phase and rain on elephant events and for all
dates where an elephant event was reported, I recorded the moon phase and rainfall (mm). Moon
Figure 3.2: Map of the Dewagiriya Village study area.This map shows the location of houses, 'main' roads, and
water tanks. Houses designated by red diamonds indicate participants in our crop-raiding surveys.

91
phase data was collected from the freely available ‘Time and Date’ website (www.timeanddate.com)
and was categorised as full, waxing, new or waning following the protocol described in Gunn et al.
(2013) (Table 3.1). Presence or absence of rain (hereafter ‘raid day rain’) plus the cumulative total of
the previous 28 days rain (hereafter ‘previous month rain’) were recorded using rainfall
measurements (mm) purchased from the Department of Meteorology, Sri Lanka, however were
unavailable for December 2014 and July 2016. This unfortunately meant that I could not calculate
cumulative rainfall between the 1st and 28th of January 2015, and between the 1st July and 28th August
2016, and these dates were excluded from analysis.
Moon phase category Definition
Full Moon Night of the full moon, plus three nights before and three nights after
New moon Night of the new moon, plus three nights before and three nights after
Waning moon Nights between the full moon and the new moon (as defined above)
Waxing moon Nights between the new moon and the full moon (as defined above)
Table 3.1. Moon phase definitions
To facilitate a comparison of dates when elephant events did and did not occur, I generated an equal
number of randomly-chosen dates when elephants were not present in the village, with the
RANDBETWEEN command in Microsoft Excel using Julian dates for the duration of the study period,
and recorded moon phase and rainfall data for these also.
On any given date, it was possible for elephants to travel to multiple fields or household areas. As I
could not individually identify most elephants entering the village, I could not determine with certainty
if elephant events reported by multiple households on the same date were caused by the same
elephant, or group of elephants, or if separate groups of elephants were responsible. Thus, in order
to identify the frequency of elephant events throughout Dewagiriya village, the protocol described in
Gunn et al. (2013) was followed, where a 'raid day' period (24-hour period starting at 6 am) was used
to combine all records of elephant sightings and crop-raids within the village into a single count. The
majority of the time, reports from a given 'raid day' period consisted of the same or similar counts of
elephants, often concentrated in a localised pocket of the village. Based on this, the assumption that
multiple elephant events reported on any given 'raid day' period constitute only one event is
considered reasonable, while also avoiding the potential problem of over-estimating frequency of
elephant presence.

92
I did however count the frequency with which elephants visited each household's property, to identify
potential HEC 'hotspots' throughout the village. To help determine potential influencing factors of
crop-raiding vulnerability within Dewagiriya Village, the location of each household in relation to the
main road (north or south of, and adjacent to main road or not) was recorded. This provided a measure
of proximity to forest/water tank periphery, as the south of the village and specifically those houses
not located adjacent to the main road, were closer to the forest and water tanks. I used this measure
rather than distance in metres due to the changeable nature of water tank boundaries influenced by
rainfall and irrigation. Additionally, the number of adults per household and estimated annual
household income was documented during semi-structured one-on-one interviews which were used
to assess community perceptions of co-existence with elephants for a separate element of this overall
study.
Additionally, between January 2015 and December 2017, 15 camera traps were mounted at strategic
locations on posts or trees within the village, and at the forest periphery, to capture footage of
elephants with the aim of determining sex, group size and individual identity. Due to accessibility,
water/elephant damage and availability of charged batteries, the number of camera traps operational
at any given time varied between seven and 15.
3.3.3 Statistical analysis
To address the questions of whether certain factors influenced a) the likelihood of elephants entering
Dewagiriya Village, b) within-village variations in frequency of elephant events, and c) the likelihood
of elephant-inflicted damage occurring, I selected predictor variables for each based on
environmental, temporal, spatial or social conditions deemed likely to influence elephant behaviour
(table 3.1). The relationship of the associated predictor variables to: a) the likelihood of elephants
entering Dewagiriya Village, and c) the likelihood of elephant-inflicted damage occurring was tested
using generalised linear models, assuming a binomial distribution. The relationship of predictor
variables to b) within-village variations in frequency of elephant events, was tested using a generalised
linear mixed model, assuming a poisson distribution.
Analysis for the datasets relating to each of the above questions followed the same basic procedure.
First, I checked for collinearity of predictor variables using Spearman’s Rank Correlation, as correlated

93
predictors can produce unreliable parameter estimates (Symonds and Moussalli 2011; Quinn and
Keough 2002). All correlation coefficients were <0.40 indicating that no strong correlations existed,
thus all predictor variables were included in the modelling process. For all combinations of models, an
information theoretic approach was employed (Burnham and Andersen 2002), specifically Akaike’s
Information Criterion, adjusted for small sample sizes (AICc). Akaike weights (wi) were calculated to
gauge the level of support for each model, given the sample data (Burnham and Andersen 2002). AICc
differences (∆ i), which show the variation between the highest ranked models and other models in
the candidate set, were calculated to measure support for each candidate model. Models with ∆i <2
were considered to have substantial support (Burnham and Andersen 2002) and predictor variables
were considered influential when the 95% confidence interval (CI) of their coefficient did not overlap
zero.
Statistical analyses were conducted with the R Studio Statistical Package 3.5.1 (R core team 2018).
Generalised linear models (GLM’s) and generalised linear mixed models (GLMM’s) were constructed
using the package ‘lme4’ (Bates et al. 2014), with model outputs generated with the ‘effects’ package
(Fox 2003) and visualised with ‘ggplot2’ (Wickham 2016). Models were ranked and assessed for ‘best
fit’ using the ‘AICcmodavg’ package (Mazerolle 2016).
3.4 Results
3.4.1 Elephant events by raid days
During the three-year study period, one or more elephants were observed in Dewagiriya Village on
293 (25.7%) of 1096 possible raid days. Elephant events occurred almost exclusively at night,
specifically between six pm and three am, with the heaviest traffic occurring between nine pm and
three am. Elephant sightings during daylight hours were rare, comprising only 5.8% of all elephant
events. Predominantly, elephants entered the village in small groups, with a mean (±SE) group size of
1.89 (±0.06) elephants. The maximum number of elephants observed together was eight however
groups of more than four individuals were rarely observed (<4% of elephant events). Lone elephants
were the most common accounting for 47.1% of elephant events.

94
Given the primarily nocturnal nature of elephant events, group type and sex could be difficult to
ascertain. At times people were unaware of elephant presence until evidence was seen the next
morning, and some people were simply unable to differentiate between the sexes. Understandably,
people were more focused on deterring elephants from their property than determining sex and group
type. Still, for 52.2% of events reported, the participant confidently recorded group type. Results
supported a strong male sex bias in elephant activity within Dewagiriya Village with just over 50% of
elephant events attributed to lone males (36.9%) or small all-bull groups (13.3%) and only 2%
attributed to mixed-sex groups. No female-led family groups, or lone females, were identified.
Although camera trap footage represented only a snapshot of total elephant events, further support
was lent to the existence of male sex bias in elephant events. Of the 50 times elephants were captured
on camera, 82% showed bull groups of between one to five individuals, and the remaining footage
was of insufficient quality to determine sex (Figure 3.3)

95
.
Figure 3.3 Camera trap footage of elephants in Dewagiriya Village
Top: An adult male elephant forages in the forest on the periphery of Dewagiriya Village.
Bottom: An adult male elephant walks through Mr. Sobana’s field.

96
3.4.2 Factors influencing elephant presence in Dewagiriya Village
I selected month, year, moon phase, raid day rain, and previous month rain as likely predictors
influencing elephant events in Dewagiriya Village. Specifically, I hypothesised that: yearly and monthly
fluctuations in frequency of elephant events would be observed; that elephant events would peak
during the new moon phase when nights are at their darkest; and that elephant events would increase
when the previous month rain was high but that on nights of actual rainfall, elephants would enter
the village less frequently.
Using data collected on dates of elephant events plus the randomly generated no-elephant event
dates, I developed and compared (using AICc) a total of 22 models encompassing all possible
combinations of the predictor variables. The resulting model selection was four models with Δ i <2,
comprising the 95% confidence interval (CI) set (Table 3.2). Model fit was low for all models (D²≤0.10).
Adopting a backwards (step-down) selection process (Zuur et al. 2009), I fitted each of these models
separately and selected all influential variables (those whose 95% CI did not overlap 0) to include in
the final ‘best’ model from which to draw influence. Results indicated that three variables were
important predictors of elephant events: month, year and raid day rain.
Candidate model df LogLik AICc Δi wi cum. wi D2
Intercept + Raid day rain + Month 14 -361.51 751.82 0.00 0.36 0.36 0.10
Intercept + Month + Year 15 -360.91 752.74 0.92 0.23 0.59 0.10
Intercept + Previous month rain +
Raid day rain + Month
15 -361.08 753.09 1.27 0.19 0.78 0.10
Moon phase + Raid day rain +
Month
17 -359.22 753.62 1.80 0.15 0.93 0.09
Table. 3.1: Results from models predicting likelihood of elephant events in Dewagiriya Village.
All models that gained substantial support (Δi AICc <2 and/or cumulative wi = ≤ 0.95) are included. Variables
are described in methods. Also listed are degrees of freedom (df), LogLikelihood (LogLik), AIC values (AICc),
Delta AICc (Δi), Akaike weight (wi), cumulative Akaike weight (cum. wi) and deviance explained (D2) for each
model.
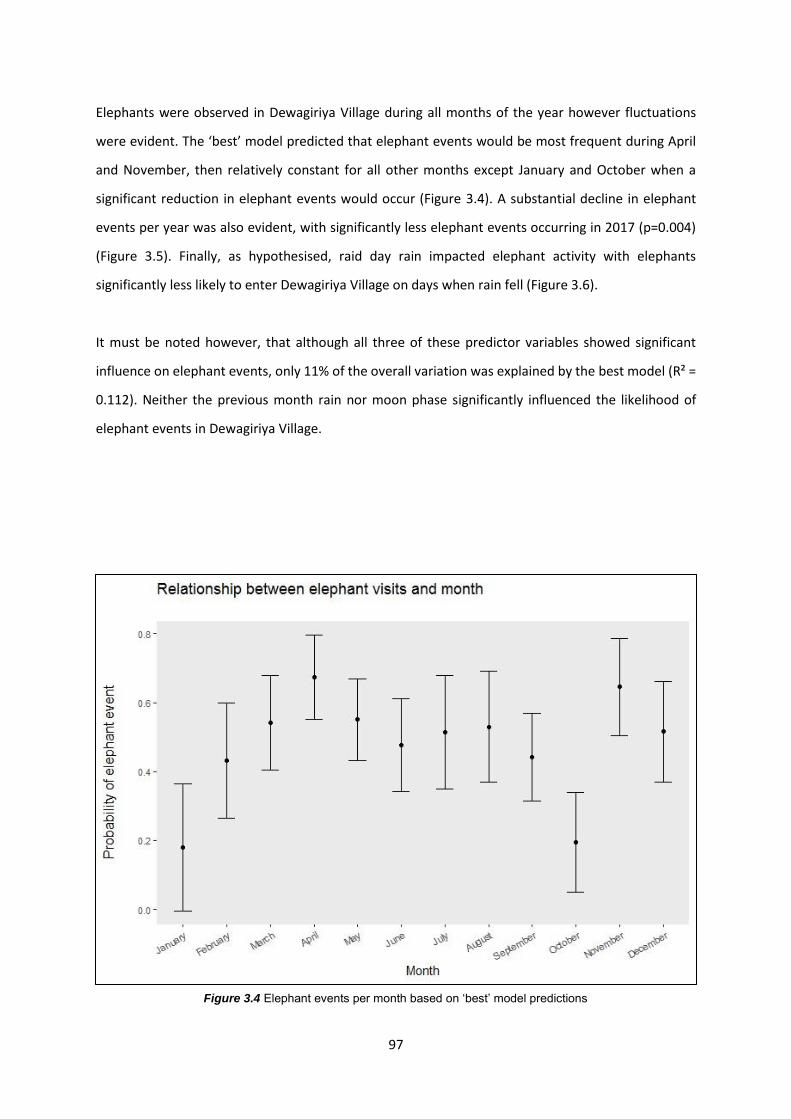
97
Elephants were observed in Dewagiriya Village during all months of the year however fluctuations
were evident. The ‘best’ model predicted that elephant events would be most frequent during April
and November, then relatively constant for all other months except January and October when a
significant reduction in elephant events would occur (Figure 3.4). A substantial decline in elephant
events per year was also evident, with significantly less elephant events occurring in 2017 (p=0.004)
(Figure 3.5). Finally, as hypothesised, raid day rain impacted elephant activity with elephants
significantly less likely to enter Dewagiriya Village on days when rain fell (Figure 3.6).
It must be noted however, that although all three of these predictor variables showed significant
influence on elephant events, only 11% of the overall variation was explained by the best model (R² =
0.112). Neither the previous month rain nor moon phase significantly influenced the likelihood of
elephant events in Dewagiriya Village.
Figure 3.4 Elephant events per month based on ‘best’ model predictions
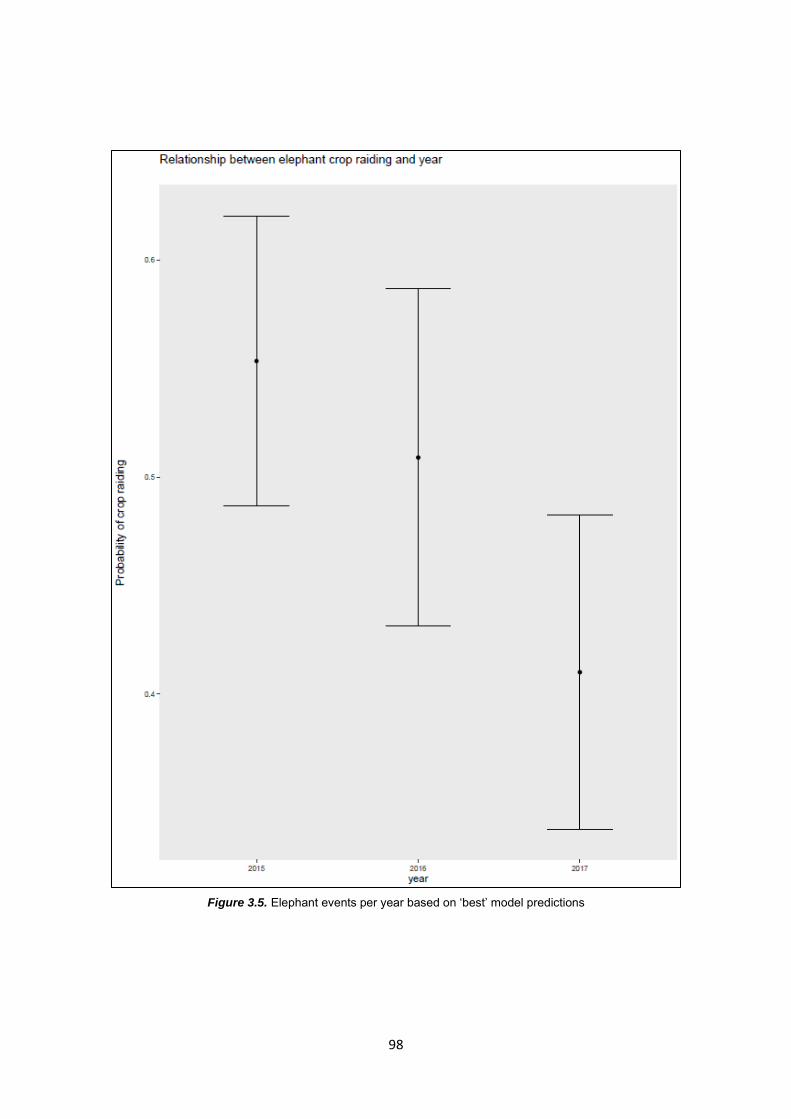
98
Figure 3.5. Elephant events per year based on ‘best’ model predictions
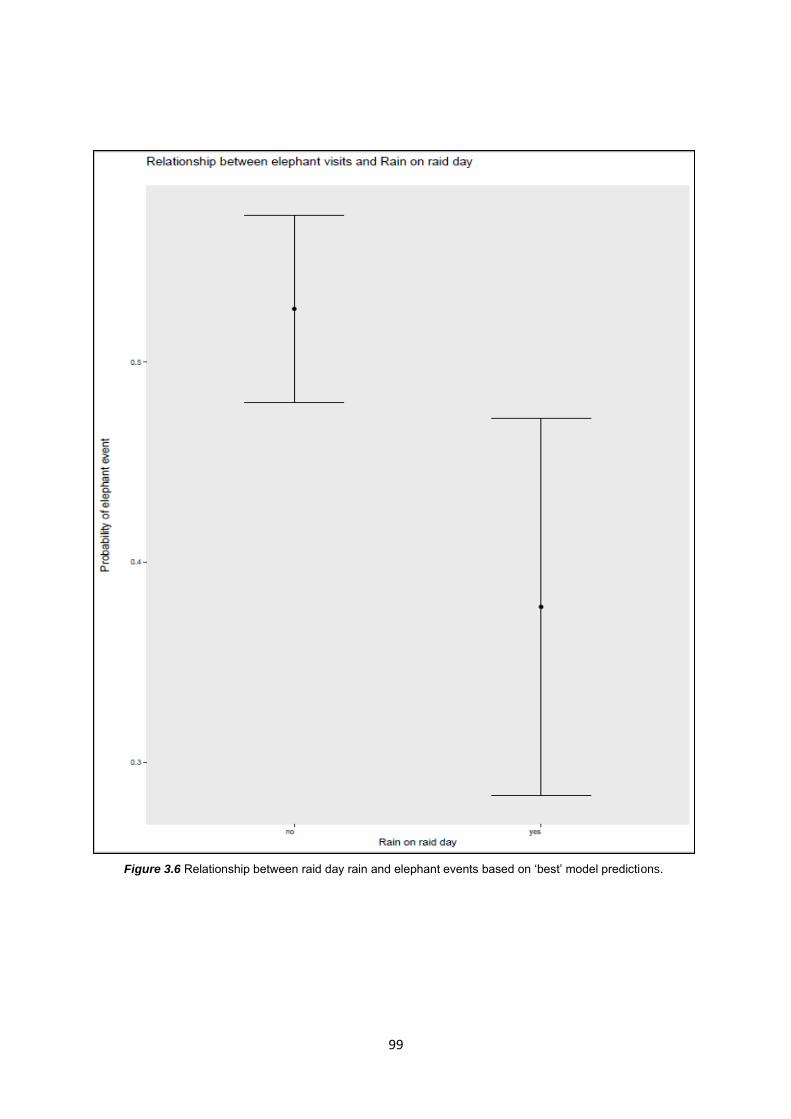
99
Figure 3.6 Relationship between raid day rain and elephant events based on ‘best’ model predictions.

100
3.4.3 Factors influencing within-site variations in elephant events
A total of 437 elephant events were reported by individual households. Frequency of elephant events
varied widely per household: at one end of the spectrum only two elephant events were reported by
the Rathnasena family (H31) yet at the other end, the Sumanawathi family (H35) sighted elephants on
their property on 63 occasions. Overall, the mean (± SE) count of elephant events over the entire study
period per household was 16.18 (±2.89).
I selected location in village (north/south), proximity to road, total number of adults in household, and
mean annual income as likely predictors influencing the frequency of elephant visits per household in
Dewagiriya Village. Specifically, I hypothesised that: participants who lived in the south of the village
and whose property did not border the main road would be subject to more frequent elephant events;
as would households with fewer adult members and lower mean annual income, as these households
would likely be less well-equipped to invest in elephant deterrents.
To account for all possible combinations of the predictor variables, 15 models were built and
compared. The top-ranked models comprised two models with Δ i <2, and eight within the full 95% CI
set (Table 3.3). I fitted each of these models in the 95% confidence set separately and selected all
influential variables to include in the final ‘best’ model. The best model indicated that only location
and proximity to road significantly influenced the frequency of elephant events per household.
Households located to the south of the main road in Dewagiriya Village were visited by elephants
significantly more often than households located to the north. Participants whose properties bordered
the main road were significantly less likely to experience high levels of elephant events. This is likely
because properties in the south of the village, and particularly those set back from the main road,
were in closest proximity to the water tanks and forest habitat, which are both valuable resources to
elephants. Overall, results showed 46.4% of the variance in frequency of elephant visits to explained
by the ‘best’ model when only fixed effects were considered (R₂(marginal) = 0.463), and 80% of the
variance was explained when fixed and random effects were accounted for (R₂(conditional) = 0.798).
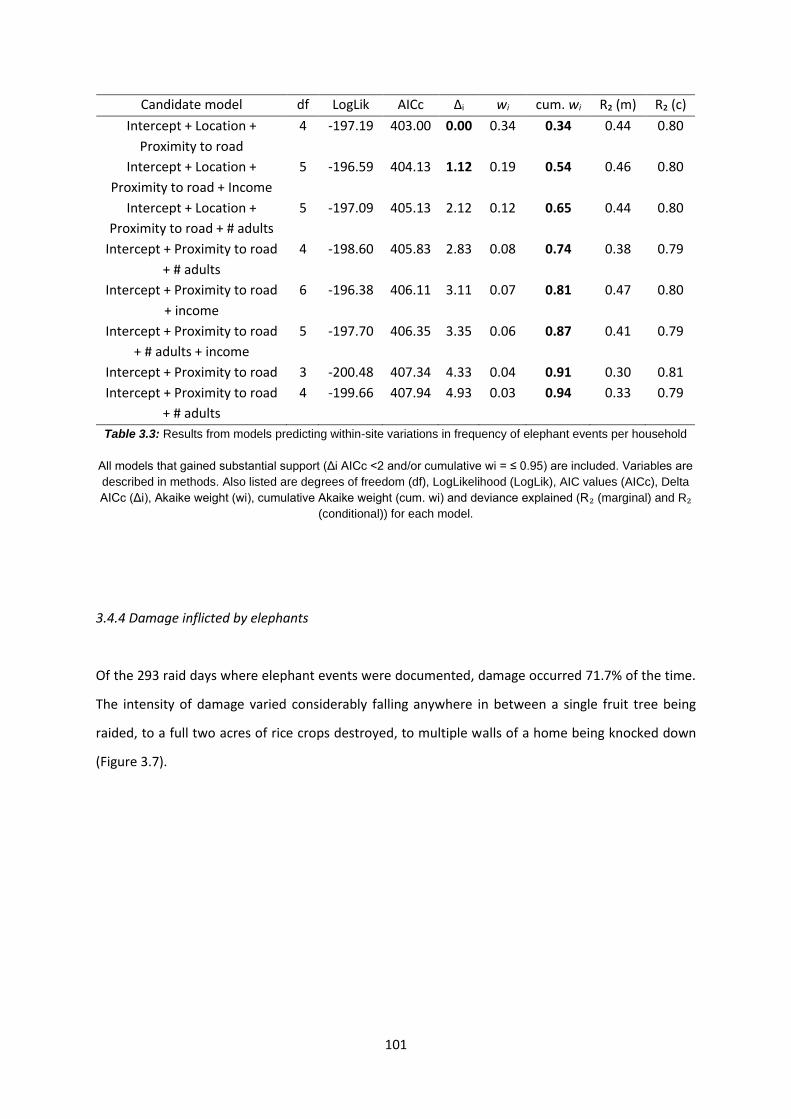
101
3.4.4 Damage inflicted by elephants
Of the 293 raid days where elephant events were documented, damage occurred 71.7% of the time.
The intensity of damage varied considerably falling anywhere in between a single fruit tree being
raided, to a full two acres of rice crops destroyed, to multiple walls of a home being knocked down
(Figure 3.7).
Candidate model df LogLik AICc Δi wi cum. wi R₂ (m) R₂ (c)
Intercept + Location +
Proximity to road
4 -197.19 403.00 0.00 0.34 0.34 0.44 0.80
Intercept + Location +
Proximity to road + Income
5 -196.59 404.13 1.12 0.19 0.54 0.46 0.80
Intercept + Location +
Proximity to road + # adults
5 -197.09 405.13 2.12 0.12 0.65 0.44 0.80
Intercept + Proximity to road
+ # adults
4 -198.60 405.83 2.83 0.08 0.74 0.38 0.79
Intercept + Proximity to road
+ income
6 -196.38 406.11 3.11 0.07 0.81 0.47 0.80
Intercept + Proximity to road
+ # adults + income
5 -197.70 406.35 3.35 0.06 0.87 0.41 0.79
Intercept + Proximity to road 3 -200.48 407.34 4.33 0.04 0.91 0.30 0.81
Intercept + Proximity to road
+ # adults
4 -199.66 407.94 4.93 0.03 0.94 0.33 0.79
Table 3.3: Results from models predicting within-site variations in frequency of elephant events per household
All models that gained substantial support (Δi AICc <2 and/or cumulative wi = ≤ 0.95) are included. Variables are
described in methods. Also listed are degrees of freedom (df), LogLikelihood (LogLik), AIC values (AICc), Delta
AICc (Δi), Akaike weight (wi), cumulative Akaike weight (cum. wi) and deviance explained (R₂ (marginal) and R₂
(conditional)) for each model.

102
Figure 3.7 Elephant damage in Dewagiriya Village. Top Left: elephant footprint damage in the rice field. Top
Right: Elephants visited the Thilanarathne family during the night, knocking down part of the wall while they
were inside. Bottom: elephant damage to banana trees at the Ghanawathi home garden.
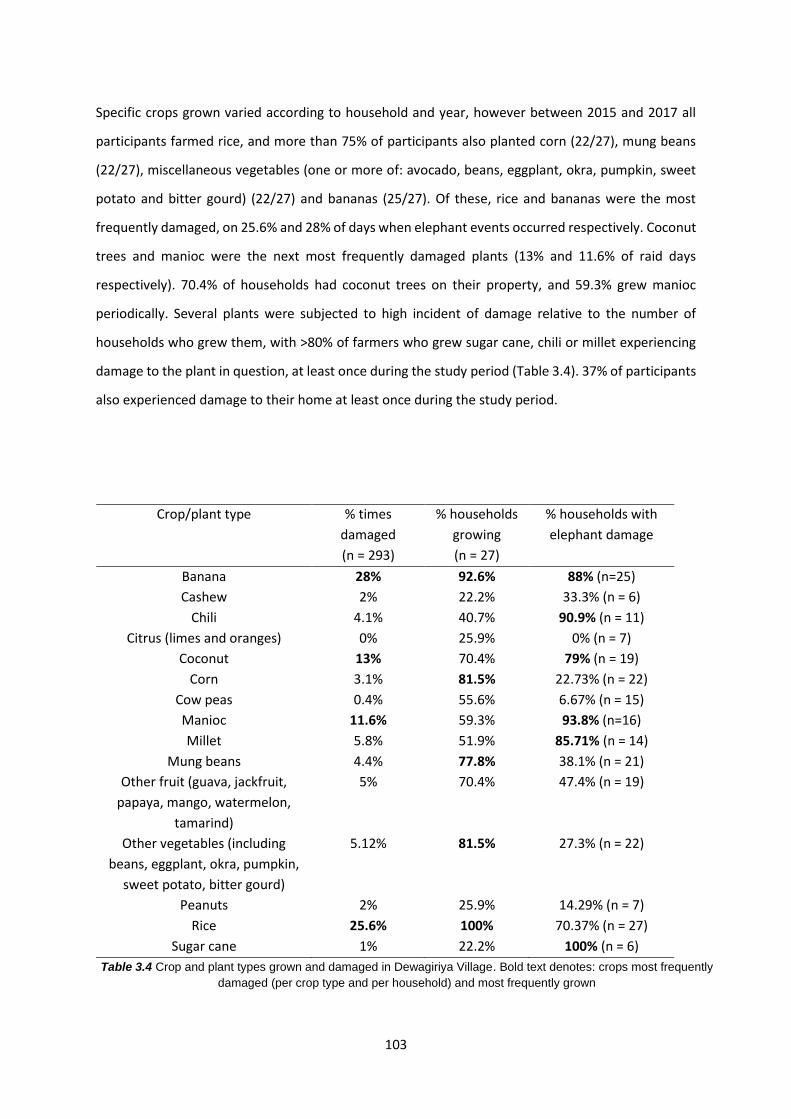
103
Specific crops grown varied according to household and year, however between 2015 and 2017 all
participants farmed rice, and more than 75% of participants also planted corn (22/27), mung beans
(22/27), miscellaneous vegetables (one or more of: avocado, beans, eggplant, okra, pumpkin, sweet
potato and bitter gourd) (22/27) and bananas (25/27). Of these, rice and bananas were the most
frequently damaged, on 25.6% and 28% of days when elephant events occurred respectively. Coconut
trees and manioc were the next most frequently damaged plants (13% and 11.6% of raid days
respectively). 70.4% of households had coconut trees on their property, and 59.3% grew manioc
periodically. Several plants were subjected to high incident of damage relative to the number of
households who grew them, with >80% of farmers who grew sugar cane, chili or millet experiencing
damage to the plant in question, at least once during the study period (Table 3.4). 37% of participants
also experienced damage to their home at least once during the study period.
Crop/plant type % times
damaged
(n = 293)
% households
growing
(n = 27)
% households with
elephant damage
Banana 28% 92.6% 88% (n=25)
Cashew 2% 22.2% 33.3% (n = 6)
Chili 4.1% 40.7% 90.9% (n = 11)
Citrus (limes and oranges) 0% 25.9% 0% (n = 7)
Coconut 13% 70.4% 79% (n = 19)
Corn 3.1% 81.5% 22.73% (n = 22)
Cow peas 0.4% 55.6% 6.67% (n = 15)
Manioc 11.6% 59.3% 93.8% (n=16)
Millet 5.8% 51.9% 85.71% (n = 14)
Mung beans 4.4% 77.8% 38.1% (n = 21)
Other fruit (guava, jackfruit,
papaya, mango, watermelon,
tamarind)
5% 70.4% 47.4% (n = 19)
Other vegetables (including
beans, eggplant, okra, pumpkin,
sweet potato, bitter gourd)
5.12% 81.5% 27.3% (n = 22)
Peanuts 2% 25.9% 14.29% (n = 7)
Rice 25.6% 100% 70.37% (n = 27)
Sugar cane 1% 22.2% 100% (n = 6)
Table 3.4 Crop and plant types grown and damaged in Dewagiriya Village. Bold text denotes: crops most frequently
damaged (per crop type and per household) and most frequently grown
The 2nd column shows how often each crop/plant type was damaged as a % of total raid days. Bold text represents the 4 most commonly damaged. The 3rd column shows how many of the participating households grew each crop/plant at least once between 2015 and 2017. The 4th column shows the % of households who experienced damage to the given crop/plant at least once between 2015 and 2017 (excluding households who did not grow the crop/plant). For the 3rd and 4th columns, bold text represents scores of ≥75%).

104
As with presence or absence of elephant events, I hypothesised that certain factors would influence
whether damage was inflicted when elephant events occurred. Again, I selected month, year, moon
phase, raid day rain, and previous month rain as my predictor variables, expecting monthly and annual
fluctuations in damage frequency, more damage to occur during new moon phases and when the
previous months rainfall was high, and less frequent damage on nights of actual rainfall.
To test for possible relationships between the predictor variables and presence/absence of damage
occurring on nights of elephant events, I incorporated all possible combinations of the predictor
variables to test 22 models. The resulting model selection comprised two models with Δ i <2. These
models, along with four others, comprised the 95% CI set (Table 3.5). I fitted each of these models
separately and selected all influential variables to include in the final ‘best’ model.
Results indicated that previous month rain and year were important predictors of elephant damage.
Specifically, elephant damage increased in frequency when the previous month rain was high but
decreased significantly in 2017 compared to other years. These results were represented in both the
prediction models and observed data and are likely a reflection of increased crop growth following
high rainfall, and reduced crop availability in Dewagiriya Village in 2017 due to poor availability of
water. Still, only 4.2% of the overall variation was explained by this ‘best’ model (R² = 0.042).
Candidate model df LogLik AICc Δi wi cum. wi D2
Intercept + Prev month rain + Year 4 -150.31 308.78 0.00 0.43 0.43 0.04
Intercept + Prev month rain + Raid day
rain + Year
5 -150.02 310.26 1.48 0.21 0.64 0.04
Intercept + Year 3 -152.82 311.73 2.95 0.10 0.74 0.03
Intercept + Prev month rain 2 -153.94 311.92 3.14 0.09 0.83 0.02
Intercept + Raid day rain + Year 4 -151.94 312.04 3.26 0.09 0.91 0.03
Intercept + Prev month rain + Raid day
rain
3 -153.71 313.52 4.74 0.04 0.95 0.02
Table 3.5 Results from models predicting likelihood of damage during elephant events in Dewagiriya Village.
All models that gained substantial support (Δi AICc <2 and/or cumulative wi = ≤ 0.95) are included. Variables are
described in methods. Also listed are degrees of freedom (df), LogLikelihood (LogLik), AIC values (AICc), Delta
AICc (Δi), Akaike weight (wi), cumulative Akaike weight (cum. wi) and deviance explained (D2) for each model.
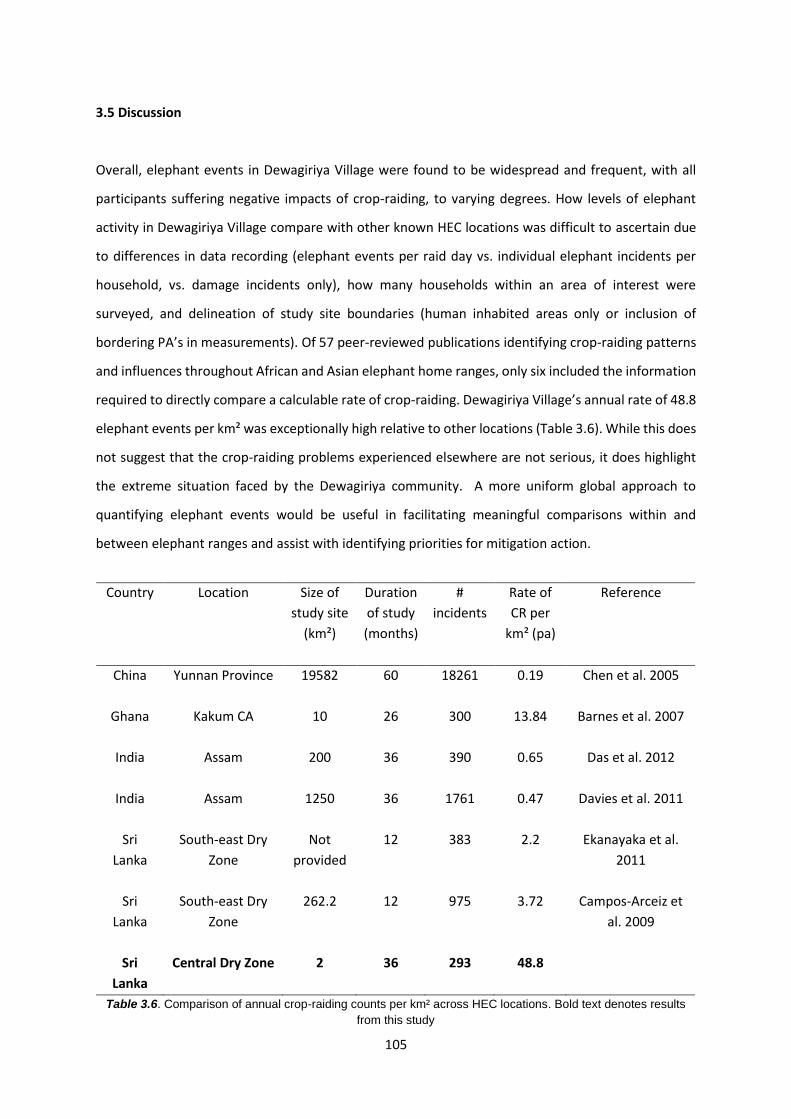
105
3.5 Discussion
Overall, elephant events in Dewagiriya Village were found to be widespread and frequent, with all
participants suffering negative impacts of crop-raiding, to varying degrees. How levels of elephant
activity in Dewagiriya Village compare with other known HEC locations was difficult to ascertain due
to differences in data recording (elephant events per raid day vs. individual elephant incidents per
household, vs. damage incidents only), how many households within an area of interest were
surveyed, and delineation of study site boundaries (human inhabited areas only or inclusion of
bordering PA’s in measurements). Of 57 peer-reviewed publications identifying crop-raiding patterns
and influences throughout African and Asian elephant home ranges, only six included the information
required to directly compare a calculable rate of crop-raiding. Dewagiriya Village’s annual rate of 48.8
elephant events per km² was exceptionally high relative to other locations (Table 3.6). While this does
not suggest that the crop-raiding problems experienced elsewhere are not serious, it does highlight
the extreme situation faced by the Dewagiriya community. A more uniform global approach to
quantifying elephant events would be useful in facilitating meaningful comparisons within and
between elephant ranges and assist with identifying priorities for mitigation action.
Country Location Size of
study site
(km²)
Duration
of study
(months)
#
incidents
Rate of
CR per
km² (pa)
Reference
China Yunnan Province 19582 60 18261 0.19 Chen et al. 2005
Ghana Kakum CA
10 26 300 13.84 Barnes et al. 2007
India Assam 200 36 390 0.65 Das et al. 2012
India Assam 1250 36 1761 0.47 Davies et al. 2011
Sri
Lanka
South-east Dry
Zone
Not
provided
12 383 2.2 Ekanayaka et al.
2011
Sri
Lanka
South-east Dry
Zone
262.2 12 975 3.72 Campos-Arceiz et
al. 2009
Sri
Lanka
Central Dry Zone 2 36 293 48.8
Table 3.6. Comparison of annual crop-raiding counts per km² across HEC locations. Bold text denotes results
from this study

106
3.5.1 Characteristics of crop-raiding elephants
In concurrence with common characteristics of elephants at most crop-raided locations (see: Africa:
Graham et al. 2010; Sitati et al. 2003, Asia: Aziz et al. 2016; Borah and Bhuyan 2016; Thaufeek et al.
2014; Das et al. 2012; Haturusinghe and Weerakoon 2012; Nath et al. 2013; Campos-Arceiz et al.
2009), it was found that solitary elephants or small groups most commonly utilised crop-raiding as a
foraging strategy in Dewagiriya Village. Globally, it is typical for male elephants to dominate crop-
raiding activity (Africa: Chiyo et al. 2012; Graham et al. 2010, Asia: Borah and Bhuyan 2016; Nath et
al. 2013; Das et al. 2012; Haturusinghe and Weerakoon 2012; Ekanayake et al. 2011; Fernando et al.
2005; Wikramanayake et al. 2003; Sukumar 1988), and again, results from Dewagiriya appear to
concur with this general trend. Of 155 events where sex was determined, on only six occasions were
females involved, indicating a strong male-bias. This is further supported by camera trap evidence of
nine individually identified males in or on the periphery of Dewagiriya, all bearing wounds and scars
consistent with human inflicted injuries (pers. comm). It should be noted though, that for 47% of
elephant events, community members could not accurately identify sex, and that in general people
were more confident identifying adult males than differentiating between younger male and adult
female elephants. Thus, it is possible that more females than reported were involved in crop-raids,
and female-led herds are known to raid intermittently in areas of south-east (Fernando et al. 2005),
north-central (Wikramanayake et al. 2003), and north-west Sri Lanka (Haturusinghe and Weerakoon
2012). Still, it seems unlikely that raiding by females is anything bar a rare event in Dewagiriya Village,
although further research using structured night-time observations and/or mitochondrial DNA
analysis on elephant faeces would be beneficial for confirmation.
It has been suggested that male and female elephants differ in their propensity to take risks, and that
crop-raiding is one such behaviour where this difference is apparent (Ekanayaka et al. 2011; Sukumar
and Gadgil 1988). Despite having equal access to cultivated fields, male elephants are typically more
likely to adopt crop-raiding as a high-risk, high-gain strategy, benefitting from bountiful supplies of
nutritionally high foods, while females prefer to forage in the relative safety of wild habitat rather
than exposing themselves and their young to the potential risk of human interaction (de Silva et al.
2013; Haturusinghe and Weerakoon 2012; Fernando et al. 2011). Furthermore, studies in Asia
(Ekanayaka et al. 2011) found that non-raiding female herds obtained ample resources from wild
fodder without needing to delve into cultivated fields as some of their male counterparts were doing,

107
indicating that neither nutritional benefits nor access fully explain the motivation to crop-raid.
Although outside the scope of this study, I saw no obvious distinction between body condition of adult
females and crop-raiding males, or indication that ample nutrition could not be obtained without
raiding crops (K. Butler, pers. obs). Further research combining an assessment of body condition and
nutritional intake, with detailed behavioural studies of daily activities of crop-raiding and non-crop-
raiding individuals, would go a long way to understanding the motivational states of crop-raiding
elephants, and perhaps expose another layer of characteristics and influences driving crop-raiding
behaviour. However, this requires a long-term commitment and is fraught with difficulties in forested
habitat, so in the meantime mitigation planning based on current evidence and more efficient
assessments of crop-raiding activity must go on.
3.5.2 Influences of crop-raiding behaviour
Investigating broad influences of crop-raiding behaviour in Dewagiriya Village, including timing of
raids, targeted crop types, and seasonal variations, I found no wild deviations from the norms
identified throughout both Asia and Africa. In all locations where crop-raiding is a major HEC issue,
elephants utilise this foraging strategy nocturnally, and it is extremely rare to see elephants in villages
during the day (Wilkie and Douglas-Hamilton 2018; Das et al. 2014; von Gerhardt et al. 2014; Nath et
al. 2013; Graham et al. 2010; Santaipillai et al. 2010; Campos-Arceiz et al. 2009; Jackson et al. 2008;
Sitati et al. 2003). Unsurprisingly, Dewagiriya Village is no exception and although very occasionally
elephants were seen at water tanks in the late afternoon, only once throughout the three-year study
did an elephant enter a field during daylight. Furthermore, elephant crop-raiding occurred most
frequently between 9 pm and 3 am, the hours of darkness where outdoor human activity is virtually
non-existent. Although most studies do not specify times of raiding beyond ‘nocturnal’, similar peaks
in specific raiding times when people are generally inside and asleep have been identified in Kenya
(Sitati et al. 2003) and Sri Lanka (Santiapillai et al. 2010).
‘Awareness’ and ‘forward-thinking’ in animals are difficult to prove conclusively, but still many
prominent researchers believe elephants possess an awareness of, and ability to assess, the potential
danger posed by interactions with humans (Wilkie and Douglas-Hamilton 2019; Gunn et al. 2013;
Graham et al. 2009; Barnes et al. 2006). Foraging on crops under the dark cover of night, and
specifically adjusting nocturnal foraging to hours of lowest human activity, is likely a risk-avoidance

108
strategy that some elephants adopt to minimise the risk of human encounters, while still benefiting
from access to highly nutritious cultivated foods (Gunn et al. 2013; Graham et al. 2009; Barnes et al.
2006). Patterns identified in Dewagiriya Village, coupled with farmer reports and anecdotal evidence
from camera trap footage of elephants resting and foraging in the peripheral forest during the day,
concur with these risk-minimisation theories.
In Ghana (Barnes 2007) and Tanzania (Gunn et al. 2013) it appears elephants have taken risk-
minimisation strategies one step further. Not only do elephants raid nocturnally, they are also
influenced by the light availability of the moon, and crop-raid more frequently during new lunar
phases when nights are at their darkest. This would presumably reduce the likelihood of being
detected by humans even further. In Dewagiriya Village however, moon phase was not found to be
an influencing factor on crop-raiding activity. Perhaps the extremely high frequency of elephant events
precluded influence of moonlight, or perhaps other factors were simply more influential.
It was interesting to note that in Dewagiriya Village, elephants were less likely to crop-raid on nights
when rain fell. I could not find any other studies considering the effects of daily rainfall on crop-raiding,
so it is impossible to know if this is typical or atypical. I can only hypothesise that elephants utilising
habitat in and around Dewagiriya Village, preferred to remain under the relative shelter of the forest
during rain. While certainly not the be-all and end-all of understanding crop-raiding patterns,
identifying very specific influences of crop-raiding behaviour such as lunar phase or nightly rain, could
at least help farmers plan mitigation by knowing when crop-guarding efforts need to be intensified or
could reasonably be relaxed.
In most locations, the crop most targeted by elephants is the crop most frequently grown.
Unfortunately, this generally means the crop that is most valued by farmers, for subsistence or income
generation or both. In many areas of Asia this crop is rice (also called paddy) (e.g. Patil and Patil 2017;
Borah and Bhuyan 2016; Palei and Singh 2016; Nath et al. 2013; Sahu and Das 2012; Davies et al. 2011;
Ekanayaka et al. 2011; Webber et al. 2011; Santiapillai et al. 2010; Sarkar and Roskaft 2010; Campos-
Arceiz et al. 2009), while in Africa maize is often the most heavily impacted (e.g. Amwata and Mganga
2014; Guerbois et al. 2012; Graham et al. 2010; Sitati and Walpole 2006; Chiyo et al. 2005). Commonly
grown fruits, such as banana (Africa and Asia) or coconut (Asia only), are also highly susceptible to
elephant raids (e.g. bananas: Berliani et al. 2016; Chen et al. 2016; Harich et al. 2013; Nath et al. 2013;

109
Sahu and Das 2012; Ekanayaka et al. 2011; Webber et al. 2011; Chiyo et al. 2005; Sitati et al. 2003,
bananas and coconut: Patil and Patil 2017; Borah and Bhuyan 2016; Palei and Singh 2016; Santaipillai
et al. 2010; Campos-Arceiz et al. 2009). Dewagiriya Village was no different, with rice and bananas the
most frequently damaged crops. This is no surprise given that rice was grown annually by all
participants and bi-annually by farmers who had year-round access to water, and bananas were a
common feature in all home gardens. Coconut trees, also grown by almost every family, and both
manioc and millet which some farmers grow as an alternative to rice during the ‘short rains’, were
also frequently raided. Overall, 25 cultivated crop types were identified as being damaged by
elephants in Dewagiriya Village, including trampling of unpalatable crops such as chili.
Admittedly, the assessment of crop types damaged in this study is simplistic, fulfilling only the aim of
identifying which crops were damaged most frequently. Given the extremely high level of elephant
activity and the variety of crops grown, collecting detailed information on timing of plant growth,
proportion of crop damage relative to plot size, and identifying economic losses based on growth stage
of plants, was beyond the capacity of this study. However, there is no denying that this information
would be valuable in determining precise economic losses caused by elephant presence for the overall
community, and individual households, and giving further depth to impact assessments. It should also
be noted that some households in Dewagiriya, refrain from growing certain crops that were previously
valuable to them, due to the risk of elephant depredation.
A strategy often put forward to mitigate human-elephant conflict is to change cultivation types (e.g.
Berliani et al. 2016; Palei and Singh 2016; Allendorf et al. 2015), or to plant buffer zones of unpalatable
crops (e.g. Gross et al. 2017; Allendorf et al. 2015; Gross et al. 2015; Chiyo et al. 2005). For
communities, such as Dewagiriya, where the main crop is the one relied upon for subsistence, a major
shift in cultivation type is highly unlikely to be practical or supported by the community. In Dewagiriya,
planting buffer zones of unpalatable crops would also be difficult, given that rice fields and gardens
are already relatively small, and the risk of trampling would be high. Still, including small plots of high
value unpalatable crops close to the house or within small fenced areas might be worthwhile, as it
could both reduce the proportion of crops in the home garden attractive to elephants and generate
income through market sales. Several types of medicinal and aromatic plants have already proved
promising as small-scale alternative crops in both Africa (Gross et al. 2015) and Asia (Gross et al. 2017).
It would be well worth investigating options to help rural communities on a site-specific basis.

110
However when designing trials or proposing changes, it needs to be remembered that in villages such
as Dewagiriya, most families have very little disposable income, and people would either need to be
supported through initial trials, or scientific evidence of success should be strong before asking
farmers to invest their own money in planting alternative unpalatable crops.
Finally, although elephants were present in Dewagiriya all throughout the year, notable peaks and
troughs in activity existed. In most locations, such fluctuations are influenced by seasonal rainfall and
subsequent crop growth and harvests (Borah and Bhuyan 2016; Djagoun et al. 2016; Palei and Singh
2016; Das et al. 2014; Nath et al. 2013; Das et al. 2012; Gubbi 2012; Haturusinghe and Weerakoon
2012; Santiapillai et al. 2010; Campos-Arceiz et al. 2009; Jackson et al. 2008; Sitati and Walpole 2006;
Chiyo et al. 2005; Sukumar 1990; Sukumar 1988), however in Dewagiriya Village there was no clear
influence of rainfall or season on crop-raiding. Elephant events were typically low during January and
February despite this being part of the ‘long rain’ season, with paddy fields abundant and on their way
to maturity. It could be that abundance of natural fodder outside of human habitations during this
time, or increased crop-guarding by farmers as rice fields begin to flourish, impacted a reduction in
elephant visits. However, these factors were not investigated during the study and are therefore
speculations, warranting further investigation. Elephant events were also low in October, which is
before the commencement of the ‘long rains’ and towards the end of the dry season when only the
hardiest of plants grow. The lack of clear patterns relating to rainfall or season was likely a product of
unusual weather, with rainfall in 2016 and early 2017 well below average. As a result, many farmers
altered their planting patterns, with some forgoing planting of rice, manioc or millet during the ‘short
rains’ and many postponing and/or planting smaller plots of rice than usual during the ‘long rains’. To
confidently determine if influences of rainfall or season impact elephant activity in Dewagiriya Village,
on-going long-term monitoring of elephant events measured against rainfall and crop-growth are
recommended.
3.5.3 Within-site variations in elephant events
Developing an understanding of the characteristics and influences driving crop-raiding at a particular
village or location, is a useful tool for facilitating meaningful comparisons across HEC sites, for
developing mitigation techniques, and for identifying times or circumstances where crop-guarding
efforts need to be most concentrated. Still, crop-raiding is rarely uniform throughout a village, and

111
identifying within-site nuances and their potential influences, should also be a priority in crop-raiding
studies. This can help to determine within-village hotspots of elephant activity where mitigation
efforts should be focused, or to identify families requiring additional support.
In Dewagiriya Village, households located to the south of the ‘main road’ (a 2.5 metre wide dirt track
running east to west through the village) were much more susceptible to crop raids then those to the
north, and this was further amplified for families whose properties did not border the road. The main
surrounding forest habitat for elephants, plus four large water tanks, are all located to the south of
Dewagiriya, therefore it makes sense that those closest to elephant refuge bear the brunt of elephant
activity. Houses located closer to the road are to some degree buffered by properties behind their
own and closer to the village edge, meaning that there is more chance that elephants will be detected
by their neighbours before reaching those properties. This ‘buffer’ of properties between the forest
and fields and homes bordering the road, is probably more influential than the road itself, although
the increased human activity as people traverse the village using the road, coupled with a slightly
higher density of dwellings may also contribute to reduced elephant activity.
This proximity to elephant habitat or refuge as a predictor of crop-raiding incidence, holds sway in
both large-scale studies investigating elephant events over multiple villages (e.g. Santiapillai et al.
2010) and at the micro-scale. Although only a few studies have investigated specific within-site
variations, an increased vulnerability of households on the outskirts of villages is consistent, not only
when considering crop-raiding by elephants (REF) but for other species involved in crop-raiding such
as baboons (Hill 2000). This goes back to the ‘risk-minimisation’ theory, as logically elephants
entertain a somewhat smaller risk by raiding in fields closest to their escape route (back to the forest),
then venturing into the midst of a village where they are likely to encounter more people. Even in a
very small village such as Dewagiriya, where distances between properties on the periphery and
properties adjacent to the main road can be as little as 500 m, and only a narrow road divides the
north and south, there was an obvious increase in elephant events closest to the forest.
It is supposed that socio-economic factors can be a predictor of vulnerability to crop-raiding, however
this was not the case in Dewagiriya Village. Although households with less annual income might be
less equipped to invest in elephant deterrents, they are also less likely to have multiple rice growing
seasons per year, and typically have smaller crop plots and home gardens, so less area for elephants

112
to damage. Furthermore, I hypothesised that households with less adults might be more susceptible
to crop-raids, having less manpower to defend their crops but this was not the case, perhaps because
neighbours often join together to guard against elephants, especially if occupants of the household
are elderly or a single adult living alone.
3.6 Conclusion
This study of localised human-elephant conflict in a rural Sri Lankan village, provides a three-year
snapshot of the frequent crop-raiding experienced by a small farming community heavily reliant on
crop production and home-grown vegetables for income generation and subsistence. Previously
unquantified, we now have a clearer idea of how elephant activity in Dewagiriya Village fits into the
bigger picture of HEC in Sri Lanka, and indeed other locations of Asia and Africa experiencing similar
issues. Ultimately, the similarities across sites (e.g. nocturnal foraging strategy primarily undertaken
by males in small groups and targeting commonly grown crops) are greater than the differences (e.g.
extremely high frequency of elephant events and no clear seasonal patterns), although this is not to
downplay the importance of identifying site-specific nuances to develop appropriate mitigation
strategies.
Overall, our data show that elephant crop-raiding occurs year-round and that although farmers would
be wise to increase crop-guarding efforts when their most valuable crops are at stake, there is no time
of year when they can afford to lapse with vigilance. As elephants raid more often on nights of no
rainfall, it would pay to increase guarding efforts in clear conditions. If communal guarding or
defending of crops was considered, concentrating mitigation efforts in the south area of the village,
and particularly along the fields bordering the water tanks and forest should be of highest priority, as
preventing elephant entry here would likely benefit most of the community. Further study specifically
investigating a variety of appropriate mitigation techniques is essential to help this community.
Importantly, data collected during studies such as this should contribute not only to the greater
discussion of human-elephant conflict and its drivers and consequences but should be of practical use
to the communities involved. It is hoped that this systematically produced profile of crop-raiding in
Dewagiriya, can be used by the community to demonstrate their plight and need for assistance to local
authorities, and also within the community to assist with mitigation planning.

113
3.7 References
Barnes, RFW, Hema, EM, Nandjui, A, Manford, M, Dubuire, UF, Danquah, EKA & Boafo, Y 2005, 'Risk
of crop-raiding by elephants around the Kakum Conservation Area, Ghana', Pachyderm, vol. 39, pp.
19-25.
Barnes, RFW, Dubiure, UF, Danquah, E, Boafo, Y, Nandjui, A, Hema, EM & Manford, M 2006, 'Crop-
raiding elephants and the moon', African Journal of Ecology, vol. 45, pp. 112-115.
Baruch-Mordo, S, Breck, SW, Wilson, KR & Theobald, DM 2008, 'Spatiotemporal distribution of black
bear-human conflicts in Colorado, USA', Journal of Wildlife Management, vol. 72 (8), pp.1853 – 1862.
Campos-Arceiz, A, Takatsuki, S, Ekanayaka SKK & Hasegawa, T 2009, ‘The human-elephant conflict in
Southeastern Sri Lanka: Type of damage, seasonal patterns, and sexual differences in the raiding
behaviour of elephants’, Gajah, vol. 31, pp. 5 – 14.
Chakraborty, K & Mondal, J 2013, 'Perceptions and patterns of human-elephant conflict at Barjora
block of Bankura district in West Bengal, India: insights for mitigation and management',
Environmental Development and Sustainability, vol. 15, pp. 547-565.
Chen, Y, Marino, J, Chen, Y, Tao, Q, Sullivan, CD, Shi, K & MacDonald, DW 2016, 'Predicting hotspots
of human-elephant conflict to inform mitigation strategies in Xishuangbanna, Southwest China', PLoS
ONE, vol. 11(9): e0162035.doi:10.1371/journal.pone.0162035.
Choudhury, A 2004, 'Human-elephant conflicts in Northeast India', Human Dimensions of Wildlife, vol.
9(4), pp. 261-270.
Das, BJ, Lahkar, BP & Talukdar, BK 2012, 'Increasing trend of human elephant conflict in Golaghat
District, Assam, India: Issues and concerns', Gajah, vol. 37, pp. 34-37.
Das, BJ, Saikia, BN, Baruah, KK, Bora, A & Bora, M 2014, 'Nutritional evaluation of fodder, its preference
and crop-raiding by wild Asian elephant (Elephas maximus) in Sonitpur District of Assam,

114
Davies, TE, Wilson, S, Hazarika, N, Chakraborty, J, Das,D, Hodgson, DJ & Zimmerman, A 2011,
'Effectiveness of intervention methods against crop-raiding elephants', Conservation Letters, doi:
10.1111/j.1755-263X.2011.00182.x
Dickman, AJ 2010, ‘Complexities of conflict: the importance of considering social factors for effectively
resolving human-wildlife conflict’, Animal Conservation, vol. 12, pp. 459 – 466.
de Silva, M & de Silva, PK 2007, The Sri Lankan elephant: Its evolution, ecology and conservation, WHT
Publications, Colombo.
Ekanayaka, SSK, Campos-Arceiz, A, Rupasinghe, M, Pastorini, J & Fernando, P 2011, 'Patterns of crop-
raiding by Asian elephants in a human-dominated landscape in southeastern Sri Lanka', Gajah, vol. 34,
vol. 20-25.
Fernando, P, Wikramanayake, E, Weerakoon, D, Jayasinghe, LKA, Gunawardene, M & Janaka HK 2005,
‘Perceptions and patterns of human-elephant conflict in old and new settlements in Sri Lanka: insights
for mitigation and management’, Biodiversity and Conservation, vol. 14, pp. 2465 – 2481.
Fernando, P, Jayewardene, J, Prasad, T, Hendavitharana, W & Pastorini, J 2011, ‘Current status of Asian
Elephants in Sri Lanka’, Gajah, vol. 25, pp. 93 – 103.
Graham, MD, Douglas-Hamilton, I, Adams, WM & Lee, PC 2009, 'The movement of African elephants
in a human-dominated land-use mosaic', Animal Conservation, vol. 12, pp. 445-455.
Graham, MD, Notter, B, Adams, WM, Lee, PC & Ochieng, TN 2010, ‘Patterns of crop-raiding by
elephants, Loxodonta Africana, in Laikipia, Kenya, and the management of human-elephant conflict’,
Systematics and Biodiversity, vol. 8, no. 4, pp. 435 – 445.
Gunn, J, Hawkins, D, Barnes, RFW, Mofulu, F, Grant, RA & Norton, GW 2013, 'The influence of lunar
cycles on crop-raiding elephants: evidence for risk avoidance', African Journal of Ecology, vol. 52, pp.
129-137.

115
Harisch, FK, Treydte, AC, Sauerborn, J & Owusu, EH 2013, ‘People and wildlife: conflicts arising around
the Bia Conservation Area in Ghana’, Journal for Nature Conservation, vol. 21, no. 5, pp. 342 – 249.
Haturusinghe, HS & Weerakoon, DK 2012, ‘Crop raiding behaviour of elephants in the Northwestern
region of Sri Lanka’, Gajah, vol. 36, pp. 26 – 31.
Hedges, S, Tyson, MJ, Sitompul, AF, Kinnaird, M, Gunaryadi, D & Aslan 2005, 'Distribution, status and
conservation needs of Asian elephants (Elephas maximus) in Lampung Province, Sumatra, Indonesia',
Biological Conservation, vol. 124, pp. 139-146.
Hill, CM 2004, ‘Farmers perspectives of conflict at the wildlife-agriculture boundary: some lessons
learned from African subsistence farmers’, Human Dimensions of Wildlife, vol. 9, pp. 279 – 286.
Ikanda, D & Packer, C 2008, 'Ritual vs. retaliatory killing of African lions in Ngorongoro conservation
area, Tanzania', Endangered Species Resources, vol. 6, pp. 67-74.
Kansky, R, Kidd, M & Knight, AT 2016, 'A wildlife tolerance model and case study for understanding
human wildlife conflicts', Biological Conservation, vol. 201, pp. 137-145.
Leimgruber, P, Gagnon, JB, Wemmer, C, Kelly, DS, Songer, MA & Selig, ER 2003, ‘Fragmentation of
Asia’s remaining wildlands: implications for Asian elephant conservation’, Animal Conservation, vol. 6,
pp. 347 – 359.
Loe, J & Roskaft, E 2004, 'Large carnivores and human safety – a review', Ambio, vol. 33 (6), pp. 283 –
288.
Naughton-Treves, L, Mena, JL, Treves, A, Alvarez, N & Radeloff, VC 2003; Wildlife survival beyond park
boundaries: the impact of slash-and-burn agriculture and hunting on mammals in Tambopata, Peru',
Conservation Biology, vol. 17(4), pp. 1106-1117.
Njiman, V & Nekaris, KA 2010, 'Testing a model for predicting primate crop-raiding using crop-and-
farm-specific risk values', Applied Animal Behaviour Science, vol. 127, pp. 125-129.

116
O'Connell-Rodwell, CE, Rodwell, T, Rice, M & Hart, LA 2000, 'Living with the modern conservation
paradigm: can agricultural communities co-exist with elephants? A five-year case study in East Caprivi,
Namibia', Biological Conservation, vol. 93, pp. 381-391.
Packer, C 2005, 'Lion attacks on humans in Tanzania', Nature, vol. 436 (18), pp. 927 – 928.
Parker, GE, Osborn, FV, Hoare, RE and Niskanen, LS 2007, Human-elephant conflict mitigation: a
training course for community-based approaches in Africa. Participant's manual, Elephant Pepper
Development Trust, Livingstone Zambia and IUCN/SSC AfESG, Nairobi, Kenya.
Perera, BMAO 2009, ‘The Human-Elephant Conflict: A review of current status and mitigation
methods’, Gajah, vol. 30, pp. 41 – 52.
Perez, E & Pacheco, LF 2006, 'Damage by large mammals to subsistence crops within a protected area
in a montane forest of Bolivia', Crop Protection, vol. 25, pp. 933-939.
Puyravaud, JP, Gubbi, S, Poornesha, HC & Davidar, P 2019, ‘Deforestation increases frequency of
incidents with elephants (Elephas maximus)’, Tropical Conservation Science, vol. 12:
https://doi.org/10.1177/194008291986595.
Saraswat, R, Sinha, A & Radhakrishna, S 2015, 'A god becomes a pest? Human-rhesus macaque
interactions in Himachal Pradesh, northern India', European Journal of Wildlife Resources, vol. 61, pp.
435 – 443.
Sitati, NW, Walpole, MJ, Smith, RJ & Leader-Williams, N 2003, ‘Predicting spatial aspects of human-
elephant conflict’, Journal of Applied Ecology, vol. 40, pp. 667 – 677.
Sitati, NW, Walpole, MJ & Leader-Williams, N 2005, 'Factors affecting susceptibility of farms to crop-
raiding by African elephants: using a predictive model to mitigate conflict', Journal of Applied Ecology,
vol. 42, pp. 1175-1182.

117
Sitati, NW & Walpole, MJ 2006, ‘Assessing farm-based measures for mitigation human-elephant
conflict in Transmara District, Kenya’, Oryx, vol. 40, no. 3, pp. 279 – 286.
Studsrod, JE & Wegge, P 1995, 'Park-people relationships: the case of damage caused by park animals
around the Royal Bardia National Park, Nepal', Environmental Conservation, vol. 22(2), pp. 133 – 142.
Sukumar, R 2003, The living elephants: evolutionary ecology, behaviour and conservation, Oxford
University Press, New York.
Sukumar, R 2006, 'A brief review of the status, distribution and biology of wild Asian elephants,
Elephas maximus', International Zoo Yearbook, vol. 40, pp. 1 – 8.
Thapa, S 2010, 'Effectiveness of crop protection methods against wildlife damage: a case study of two
villages at Bardia National Park, Nepal', Crop Protection, vol. 29, pp. 1297 – 1304.
von Gerhardt, K, Niekerk, AV, Kidd, M, Samways, M & Hanks, J 2014, 'The role of elephant Loxodonta
africana pathways as a spatial variable in crop-raiding location', Oryx, vol. 48(3), pp. 436 – 444.
Wikramanayake, ED, Hathurusinghe, HS, Janaka, HK, Jayasinghe, LKA, Fernando, P, Weerakoon, DK &
Gunawardene, MD 2004, 'The human-elephant conflict in Sri Lanka: lessons for mitigation,
management, and conservation from traditional land-use patterns', in: (ed) Jayewardene, J
Endangered Elephants: Past, present and future, Proceedings of the symposium for human-elephant
relationships and conflicts, Sri Lanka, September 2003. Biodiversity and Conservation Trust, Colombo,
pp. 164-169.
Wilson, ML, Hauser, MD & Wrangham, RW 2007, ‘Chimpanzees (Pan troglodytes) modify grouping
and vocal behaviour in response to location-specific risk’, Behaviour, vol. 144, pp. 1621 – 1653.
Woodroffe, R, Thirgood, S & Rabinowitz, A 2005, 'The impact of human-wildlife conflict on natural
systems', IN: (eds) Woodroffe, R, Thirgood, S & Rabinowitz, A, People and Wildlife: Conflict or
Coexistence? Cambridge University Press, UK.

118
Chapter 4: Beehive fences as a human-elephant conflict mitigation tool for farmers in rural Sri Lanka

119
4.1 Abstract
As crop-raiding continues to threaten both elephant conservation and the livelihoods of farmers living
in proximity to elephants, the need for practical and effective mitigation tools becomes increasingly
important. In recent years, interest in community-based mitigation methods has grown, and there has
been a push towards increased implementation of eco-deterrents such as planting unpalatable crops
or using chili-ropes or briquettes. Beehive fencing is another deterrent showing promise in Africa
however it’s applicability as an Asian elephant deterrent has not been thoroughly explored. In this
study, we initiated a three-year trial, using ten beehive fences to protect home gardens in Dewagiriya,
a small village in Sri Lanka vulnerable to crop-raiding. We found a significant decrease in elephant
incursions into home gardens protected by a beehive fence compared to those without. However,
beekeeping was challenging with colonies frequently absconding and honey harvests low.
Furthermore, despite being relatively inexpensive the costs of establishing a beehive fence were
prohibitive to the Dewagiriya community which is probably reflective of many rural communities in
Sri Lanka. Despite high initial interest in trial participation, farmer motivation fluctuated throughout
the trial, and only 40% of farmers planned to continue using their fences long-term. Further trials in
areas of Asia, preferably with communities already familiar with beekeeping, would be beneficial to
determining its usefulness as a practical, community-based mitigation tool for rural communities
experiencing problems with crop-raiding.
4.2 Introduction
Social, economic and conservation issues resulting from human-elephant conflict (HEC), and
specifically crop-raiding, have been documented in scientific and popular mediums for several
decades (examples: Hoffmeier-Karimi and Schulte 2014; Baskaran et al. 2013; Chakraborty et al. 2013;
Ranasinghe et al. 2010a; 2010b; Bandara and Tisdell 2003; Osborn and Parker 2002; de Silva 1998;
Sukumar 1990). Management plans focusing on deterrent or mitigation methods have been devised,
implemented and assessed with varying levels of success, for various locations (examples: Gunaryadi
et al. 2017; Davies et al. 2011; Perera 2009; Fernando et al. 2008; Graham and Ochieng 2008; Sitati
and Walpole 2006; Dublin and Hoare 2004; Osborn and Parker 2003). However, overall it does not
appear that the problem is alleviating. In fact, the common consensus is that crop-raiding is only
getting worse (Fernando et al. 2011). It is generally accepted that anthropogenically influenced

120
changes to local environments are at the root cause of HEC (Amwata and Mganga 2014; Chakraborty
et al. 2013; Fernando et al. 2011; Campos-Arceiz et al. 2009; Santiapillai et al. 2006; Fernando et al.
2005; Choudhury 2004; Sitati et al. 2003) however the humans who bear the brunt of the problem are
usually not those responsible for the underlying causes (Barua et al. 2013; Santaipillai et al. 2010; Ogra
2008; de Silva 1998). Not surprisingly, communities in HEC zones are increasingly frustrated by the
ongoing challenges presented by living in proximity to wild elephants. Many people feel disillusioned
both by the perceived unwillingness of local authorities to assist (Nsonsi et al. 2018; Mabelunga et al.
2016; Chakraborty and Mondal 2013), and a feeling of helplessness at their own oft-limited capacity
to protect their livelihoods and families (Jasmine et al. 2015; Nath et al. 2015; Bandara and Tisdell
2003).
The question of how best to manage HEC is one with no easy answers. With crop-raiding presenting
challenges in almost every elephant range country, site-specific differences mean there is no one-size-
fits-all solution and most researchers agree that developing a ‘toolbox’ of various deterrents to be
used in combination, or rotated depending on site specific characteristics, would be most appropriate
(van de Water and Matteson 2018; King et al. 2017; Pozo et al. 2017; Hoare 2015; Karidozo and Osborn
2015; Hedges and Gunaryadi et al. 2009; Fernando et al. 2008; Dublin and Hoare 2004; Osborn and
Parker 2003). Innumerable factors can influence the likelihood of a deterrent method being successful
including characteristics of the local elephant population, lay-out of villages and farms, crops grown,
climate, local cultures and traditions, availability of outside assistance, and socio-economics
(examples: Barnes et al. 2015; Chakraborty and Mondal 2013; Gubbi 2012; Webber et al. 2011;
Graham et al. 2010; Campos-Arceiz et al. 2009; Chiyo et al. 2005; Fernando et al. 2005; Sitati et al.
2003). HEC is often seen as an ‘in-trend’ topic for wildlife researchers, attracting international
attention and donor funding, yet whether any meaningful dint in the overall problem has been made
is arguable. This is not for lack of trying or compassion but rather a reflection on the challenging and
changing nature of crop-raiding.
4.2.1 Human-elephant conflict mitigation
Tried and tested methods range from sophisticated government interventions to basic deterrents
making use of common household objects. At the upper end of the scale, methods are often large-
scale, implemented with assistance of government authorities and involve translocating or killing

121
problem individuals or attempting to confine elephants to PA’s using electric fencing (van Eden et al.
2016; Ponnusamy et al. 2016; Fernando et al. 2011; Kioko 2008; Santiapillai et al. 2006; Nelson et al.
2003). When well looked after, electric fencing can be quite effective however it is expensive, requires
frequent maintenance, and does not always account for the natural movement patterns of elephants
or their ecological requirements (Fernando et al. 2011; Perera 2009; Santaipillai 1996). Less success
has been demonstrated by translocations or culling, and serious ethical implications exist for both
(Fernando et al. 2012; Santiapillai 1996).
At the lower end of the scale, and often referred to as ‘traditional’ or ‘farm-based’ methods, are
measures farmers implement themselves. Common techniques include banging on pots or tin drums
and shouting, throwing firecrackers, throwing sticks or clothes doused in petrol and set alight, shining
bright torches, or hanging reflective objects or materials from wire strands (examples: van de Water
and Matteson 2018; Haturusinghe and Weerakoon 2012; Fernando et al. 2011; Hedges and Gunaryadi
2009; Perera 2009; Sitati and Walpole 2006; Thaufeek et al. 2004). Although these methods can be
effective at chasing elephants away, most farmers find elephants habituate over time, with bolder
elephants sometimes ignoring these efforts completely (Fernando et al. 2011; Sitati and Walpole 2006;
Thaufeek et al. 2004; Osborn and Parker 2003). Furthermore, farmers are required to be in close
proximity to the elephants to execute the deterrents. Given that most crop-raiding occurs at night
(Das et al. 2014; von Gerhardt et al. 2014; Graham et al. 2010; Campos-Arceiz et al. 2009;
Wikramanayake et al. 2004; Bandara and Tisdell 2004), and that methods are designed to agitate or
frighten the elephants, the use of ‘traditional’ methods often places farmers in situations fraught with
danger (Madden 2004). Additionally, psychological costs are often incurred due to the stress and
fatigue caused by living in a constant state of high alert and spending long hours guarding crops at
night after working in fields during the day (Barua et al. 2013; Hartter et al. 2011; Ogra 2008; Sukumar
2006; Thirgood et al. 2005; Hill 2004; Hoare 2000).
Out of frustration and desperation, farmers are increasingly resorting to often-illegal methods that
inflict substantial harm or injury to elephants (Haturusinghe and Weerakoon 2012; Santiapiallai et al.
2010; Bandara and Tisdell 2002). Some farmers will lace favoured foods of elephants with poison,
which can have harmful effects on numerous species who may come into contact with it (Haturusinghe
and Weerakoon 2012; Bandara and Tisdell 2002). In Sri Lanka, explosive devices (locally called hakka-
pattas) have been placed in vegetables along the edges of fields, designed to detonate as an elephant

122
bites into the vegetable (Haturusinghe and Weerakoon 2012). Shooting elephants, either directly or
by setting trap-guns, is also quite common, as evidenced by the prevalence of elephants bearing
wounds consistent with gunshot injuries (Haturusinghe and Weerakoon 2012; Bandara and Tisdell
2002). Exactly how frequently such methods occur, or how many people employ them, is difficult to
quantify as farmers very rarely admit to using illegal techniques for fear of repercussions (Bandara and
Tisdell 2002). Some scientists purport that elephants who are exposed to violence or harm at the
hands of humans, are in turn responding more aggressively to humans, thus perpetuating a cycle of
conflict (Fernando et al. 2011; Perera 2009).
In the last two decades, a shift towards community-based eco-deterrents has become more prevalent.
Examples include chili fences, chili-dung briquettes, bio-fences such as Palmyra trees, and buffer zones
of unpalatable crops (examples: van de Water and Matteson et al. 2018; Gross et al. 2017; Changa et
al. 2016; Gross et al. 2016; Karidozo et al. 2015; Baishya et al. 2012; Hedges and Gunaryadi 2009;
Fernando et al. 2008; Graham and Ochieng 2008; Sitati and Walpole 2006; Nelson et al. 2003). Some
have the advantage of exposing approaching elephants to a negative consequence. For example, chili-
based deterrents contain the capsaicin compound which is an irritant to the sensitive eyes, trunk and
mouth of elephants (Changa et al. 2016; Karidozo and Osborn 2015; Hedges and Gunaryadi 2009).
Some may provide an alternative or additional source of income. In Nepal, several species of medicinal
and aromatic plants (e.g. chamomile, coriander, basil, turmeric, lemon grass and citronella) that are
of high market value have been identified as unpalatable to elephants, and their potential as either
strategically planted buffers or alternative crops are being investigated (Gross et al. 2015). Typically,
methods such as these are employed with the assistance of local or international NGO’s however the
premise is that after set-up and any initial training required, farmers will be able to maintain and
expand the deterrents independently (Changa et al. 2016; Karidozo and Osborn 2015; King et al. 2011;
2009). As is the norm with HEC mitigation, each method comes with challenges and advantages, and
success levels are variable (Pozo et al. 2017; Gross et al. 2017; 2016; Baishya et al. 2012; Hedges and
Gunaryadi 2009).
4.2.2 Elephant interactions with bees and beehive fences- Africa
A relatively new community-based eco-deterrent currently generating a lot of interest worldwide is
the beehive fence. Put simply, a beehive fence consists of a series of beehives hung from posts, with

123
each hive spaced about 10 m apart and connected with a single wire. These connected hives are
strategically positioned around an area to be protected from elephants. Should an elephant attempt
to enter the protected area by walking in between beehives, he/she will hit the connecting wire,
causing the hives to swing and disturb the bee colony inside, inciting the bees to exit the hives and
swarm at the elephant (King et al. 2011; 2010; 2009) (Figure 4.1 and 4.2). Beehive fences have the
potential advantage of producing both a direct negative consequence to elephants (bee stings) and
an opportunity for additional income generation through sales of honey and wax products (Scheijen
et al. 2018; King et al. 2017; 2011; 2009; Nair and Jayson 2016).
Figure 4.1. Diagram of a beehive fence © Morgan Tipper
Figure 4.2. A beehive fence © Kylie M. Butler.

124
Understandably, the concept of the thick-skinned elephant being afraid of the comparatively tiny bee
can initially seem absurd. However, in Africa at least, this fear has long been expressed through
folklore and rock art (King 2010). Of more direct relevance are the anecdotes relayed by local farmers
living in proximity to elephants, mostly involving elephants running away from bee swarms after
accidentally disturbing hives (King 2010; King et al. 2009). Scientific evidence supports anecdotal
evidence with field experiments showing that African elephants avoid resting under, or feeding from,
trees containing beehives (Cook et al. 2017; Ngama et al. 2016; Vollrath and Douglas-Hamilton 2002).
Audio playback experiments exposing African elephants to the sounds of disturbed African honeybees
caused elephants to increase vigilance behaviours, to explore the immediate surroundings using
olfaction, to head shake or dust, and to walk quickly or run away from the source of the sound (King
et al. 2007). African elephants also produced a specific rumble vocalisation thought to warn nearby
conspecifics of a threat in the area, and conspecifics responded to audio playback of the rumbles with
similar behavioural responses as when exposed to actual bee sounds (King et al. 2010; 2007).
Capitalising on this apparent fear of bees, Dr. Lucy King developed the beehive fence as a HEC
mitigation tool (King 2010). Encouraging results from a pilot study in Kenya demonstrated a reduction
in crop damage on farms with beehive fences (King et al. 2009) and inspired more trials in other
locations in Kenya (King et al. 2017) and further afield within Africa (Scheijen et al. 2017). At one
location in Kenya, beehive fences showed a capacity to deter up to 80% of crop-raids from small-scale
farms (King et al. 2017). Results from a linear fence in Tanzania aiming to prevent elephants from
entering farmlands bordering PA’s also demonstrated success (Scheijen et al. 2017). That farmers
expressed positive perceptions of participating in trials and additional members requested
involvement was another promising indicator (King et al. 2009). Trials also elicited several successful
honey harvests from which farmers could keep the honey for personal use or sell it at local markets
(King et al. 2017; 2011; 2009).
4.2.3 Elephant interactions with bees and beehive fences- Asia
Positive signs of deterrent success using beehive fences in Africa, has prompted much speculation
regarding its potential applicability as an elephant deterrent in Asia. Despite many obvious similarities
between the HEC situations in Africa and Asia, and particularly crop raiding on small-scale farms, HEC
mitigation tools are not always transferrable. In the example of beehive fences, it is important to

125
recognise that both the Asian elephant and Asian honey bee (Apis cerana indica) species differ from
their African counterparts, and to consider what impact this may have on beehive fence deterrent
success.
While in Africa it is generally accepted that elephants will avoid bees (Cook et al. 2017; Vollrath and
Douglas-Hamilton 2002) less evidence, either anecdotal or scientific, exists in Asia to support the
notion that Asian elephants might also display fear or avoidance in response to the presence of bees.
In northern Thailand, a normally placid captive elephant was observed trumpeting and attempting to
run before being brought under control by her mahout while resting under a tree containing a
disturbed beehive (pers. obs. 2012). However, none of the local elephant owners or mahouts had ever
heard of, or witnessed, elephants being afraid of bees (pers. comm. 2012). In Sri Lanka, informal
conversations with residents at the study site of Dewagiriya Village did not reveal any historical
knowledge of elephants avoiding bees, despite people being familiar with encountering both wild
elephants and wild beehives in the forests (pers. comm. 2014). Still, playback experiments in Sri Lanka
exposing wild Asian elephants to audio of disturbed Asian honey bees, showed elephants responded
with avoidance and exploratory behaviours (King et al. 2017), suggesting there may be potential for
the use of bees as a deterrent.
In Asia, and at our study site in Sri Lanka specifically, two characteristics stand out as potential limiting
factors for the success of beehive fencing. Firstly, the Asian honey bee is comparatively less aggressive
than the African honey bee (Apis mellifera scutellata) (Punchihewa 1994). Also, of the three honey
bee species found in Sri Lanka, Apis cerana is the only one to nest in enclosed cavities (Punchihewa
1994), rendering it the only species possible for use in a beehive fence. However, this honey bee is
sensitive to disturbance, swarming or absconding easily and thus being difficult to attain bountiful
honey harvests from (Koeniger and Koeniger 1994). On the one hand, this behavioural characteristic
could be advantageous for deterrent effect, in that easily disturbed bees might respond aggressively
towards elephants. However frequent absconding could be problematic for overall fence
management and beekeeping.
At present, little is known about the potential of beehive fencing in Asia. To the best of my knowledge,
the only peer-reviewed study is from a small trial in India using 20 beehives to block elephant
pathways into crop fields (Nair and Jayson 2016). Although results were positive, the study is limited

126
by a relatively short nine-month study period and a small sample size with ten of 14 elephant
approaches instigated by the same elephant (Nair and Jayson 2016). The fence design also differed
from the original African beehive fence, as the beehives were placed to block path entrances (Nair and
Jayson 2016) rather than restricting access to crops by surrounding a plot or garden (King et al. 2017;
Scheijen et al. 2017; King et al. 2009; 2007).
Other indicators of beehive fence success in Asia stem from anecdotal newspaper reports (Dutta 2013;
Manoj 2012) and one short three-month trial where a small beehive fence was used to block elephant
access at one corner of a five-acre paddy field (Shri et al. 2013). Whilst encouraging, accurate
evaluations of the deterrent effect of beehive fencing for Asian elephants, require longer-term studies
with systematically collected data. Additionally, cost analyses and identifying farmer willingness to
adopt this method are essential to an overall assessment of the potential role of beehive fencing as
an Asian elephant HEC mitigation tool.
Here we aimed to address this knowledge gap by establishing a trial site of ten beehive fences in a
small village in Sri Lanka heavily impacted by crop-raiding, to assess the potential of beehive fencing
using Asian honey bees as an Asian elephant deterrent. Specifically, we aimed to:
− quantify and compare the frequency of elephant approaches to experimental and control
gardens, and the frequency with which elephants enter experimental and control gardens
− determine the financial costs of establishing and maintaining a beehive fence, and identify
common maintenance issues or challenges
− identify farmer willingness to adopt beehive fences as an elephant deterrent
− discuss overall feasibility of beehive fencing as an Asian HEC mitigation tool based on findings
from this trial
4.3 Methods
4.3.1 Study Site
This study was conducted in the small rural community of Dewagiriya Village (07°33.928' N,
080°56.866' E), located in the dry zone of Central Sri Lanka. Spanning an area of approximately 2 km²
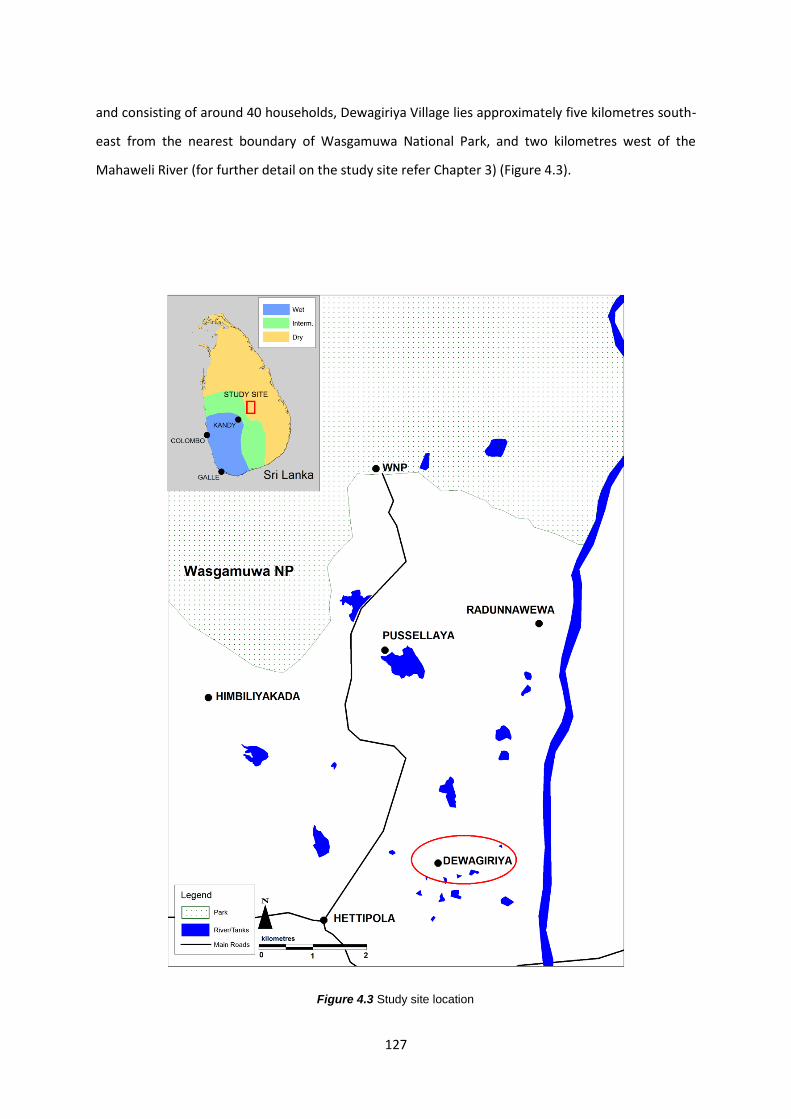
127
and consisting of around 40 households, Dewagiriya Village lies approximately five kilometres south-
east from the nearest boundary of Wasgamuwa National Park, and two kilometres west of the
Mahaweli River (for further detail on the study site refer Chapter 3) (Figure 4.3).
Figure 4.3 Study site location

128
The community of Dewagiriya Village rely primarily on rice farming for subsistence, as a staple of their
diet and/or to generate income. The average household size is 3.21 adults (± SE 0.24) and the average
annual income per household is 131 500 LKR (± SE 18 662, range: 24 000 to 339 000) or 747 USD (± SE
107, range: 136 to 1928). The community is heavily impacted by HEC in the form of crop-raiding, and
all households are negatively affected to varying degrees. Households located to the south of the main
road, which are closest to the water tanks and forest, typically experience the most frequent elephant
visits and crop damage (see Chapter 3). Elephants not only raid crops growing in fields, but they visit
home gardens where most families have banana and coconut trees and plant small fruit and vegetable
plots. Structural damage to houses as elephants attempt to access stored rice inside is a further
consequence. Elephant activity in proximity to homes is a serious safety concern for the Dewagiriya
community.
Beekeeping is not traditionally practised in Dewagiriya Village, however some households place clay
pots in their gardens to attract wild bees. Some community members, mainly older men, have
experience harvesting honey from wild hives in the forest however this is not commonly practiced
anymore. Beekeeping in its modern form using Langstroth’s hives was not practiced by any community
members prior to this study.
4.3.2 Introducing the concept of beehive fencing and participant selection for the beehive fence trial
The concept of using beehive fences to deter elephants from entering and foraging in crop fields or
home gardens was introduced during two community meetings. Prior to this, no community members
were aware of this HEC mitigation strategy nor did anyone have experience or knowledge of elephants
being afraid of bees. All adults in the community were encouraged to attend the meetings and were
notified one week prior by local community leader Mr. B.K.G. Jayasena. Meetings were held in June
and July 2014 and were attended in full by 20 and 17 adults respectively. At both meetings, additional
community members joined for shorter periods in between tending their fields. Dialogue was
translated from English to Sinhalese and vice versa on the spot by local bilingual assistants.

129
At the first meeting local HEC issues, patterns of crop-raiding, and deterrent methods currently used
were discussed. The basic premise of the beehive fence deterrent was explained, and I showed
footage of African elephants responding with avoidance to playback audio of disturbed African honey
bees and photos of beehive fences used in Africa (courtesy of Dr. King, pers. comm). Most attendees
expressed surprise or disbelief that elephants displayed fear or avoidance of bees but thought that if
beehive fences were demonstrating effectiveness in Africa that it would be worth trying in Asia also.
Most of the attendees expressed interest in learning beekeeping skills due to the medicinal properties
of honey and its market value.
At the second meeting, the merits of protecting crop fields versus smaller home garden areas were
discussed. Trials in Africa tend to use the beehive fences to surround crop plots of one to two acres,
however the layout of rice farming in Dewagiriya Village makes it difficult to delineate rice fields into
similar sized plots. Although each family farms between one to three acres, the fields of multiple
families are often contiguous making the overall crop field much larger. Given that elephants also
frequently raid home gardens where various fruits, grains or vegetables grow year-round, and that
home garden boundaries are more clearly defined, the community suggested that home gardens form
the focus of the beehive fence trial. Attendees then identified 18 households in the village which they
deemed to be most affected by crop-raiding and most in need of assistance to defend their gardens
(Table 4.1).
Local field assistant Mr. S. Herath and I visited each listed household individually to assess their
suitability for participation in the beehive fence trial. Having already established that crop-raiding was
a threat to livelihoods and/or safety, suitability was based on the following criteria: a) fruits, grains or
vegetables must be grown in the garden for at least part of each year; b) at least one adult must live
at the home year-round; c) at least one adult must be willing to complete ‘elephant event’ data sheets
and/or communicate verbally details of ‘elephant events’ on at least a bi-monthly basis; d) at least
one adult must be willing to learn beekeeping and actively participate in fence establishment and
maintenance

130
Household name House code Location description
UG Gunapala H01 South of road, west side of village, near water tank
*UG Sobana H02 South of road, west side of village, near water tank
*AG Jaysinghe H06 South of road, west side of village
UG Sirisena H07 South of road, west side of village
GMG Dharmadasa H12 South of road, west side of village
*AG Ariyadasa H20 South of road, mid-village
*PGN Somathilaka H22 South of road but adjoining, mid-village
MG Jothirathne H26 South of road, mid-village
UA Ariyasena H27 South of road, mid-village
BG Sirisena H28 North of road, east side of village
*BKG Jayasena H30 South of road but adjoining, east side of village
UA Rathnasena H31 North of road, east side of village
*WG Dharmadasa H32 South of road, east side of village, near water tank
*UG Guneris H33 North of road, east side of village
*WG Karunadasa H34 South of road but adjoining, east side of village
*BG Sumanawathi H35 South of road, east side of village, near water tank
*LG Ghanawathi H36 South of road, far-east of village, near water tank
UGSK Thilanarathne H37 South of road but adjoining, far-east of village
Table 4.1 List of households experiencing the most negative impacts of crop-raiding in Dewagiriya Village as
determined by 17 community meeting attendees. * next to household name denotes selection for participation in
the beehive fence study as an experimental subject.
In the spirit of a true collaborative community HEC mitigation initiative, and to foster a sense of
ownership and responsibility of individual fences, community preferences were taken into
consideration when selecting the ten households to participate in the experimental beehive fence
trial. Although an ideal experimental design would see beehive fences distributed evenly throughout
the village, the community was insistent that those most affected by crop-raiding and in need of the
most help, should be the experimental participants. Each experimental home garden was paired with
a control home garden of approximately the same size and growing similar fruits, grains or vegetables.
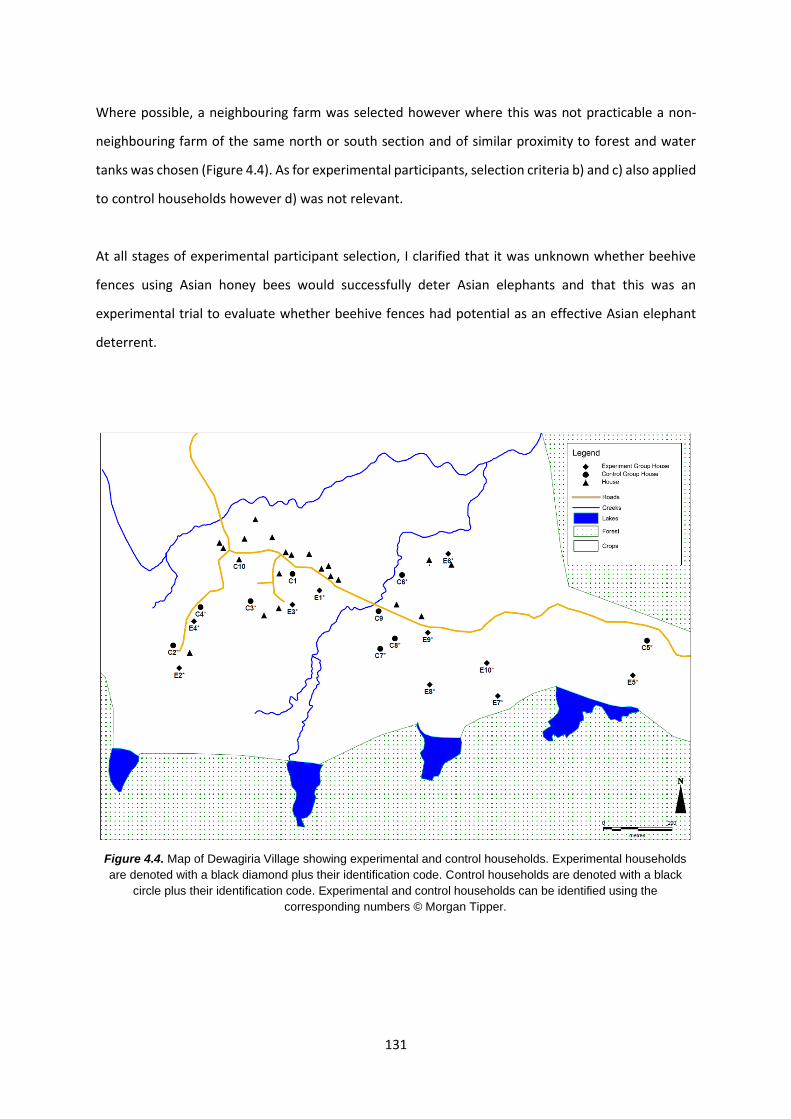
131
Where possible, a neighbouring farm was selected however where this was not practicable a non-
neighbouring farm of the same north or south section and of similar proximity to forest and water
tanks was chosen (Figure 4.4). As for experimental participants, selection criteria b) and c) also applied
to control households however d) was not relevant.
At all stages of experimental participant selection, I clarified that it was unknown whether beehive
fences using Asian honey bees would successfully deter Asian elephants and that this was an
experimental trial to evaluate whether beehive fences had potential as an effective Asian elephant
deterrent.
Figure 4.4. Map of Dewagiria Village showing experimental and control households. Experimental households
are denoted with a black diamond plus their identification code. Control households are denoted with a black
circle plus their identification code. Experimental and control households can be identified using the
corresponding numbers © Morgan Tipper.

132
4.3.3 Establishment of beehive fences
The beehive fence design used in Dewagiriya Village followed the basic protocol outlined in the
‘Beehive Fence Construction Manual’ (King 2014) and trialled in Kenya (King et al. 2017; 2011; 2009),
with some alterations made to better suit the Sri Lankan environment and make use of locally available
materials. The beehives used were ten-frame Langstroth hives with a single honey super on top (Figure
4.5). Langstroth hives were favoured over traditional log or clay pot hives as they were easier to hang
and more ‘user friendly’ in terms of monitoring hive health and harvesting honey. Also, they were
readily available for purchase from Sri Lankan beekeepers and relatively inexpensive (14.50 USD/1 900
LKR). Initially, hive parts were held together using coconut rope however during strong wind or rain
hive compartments tended to slip. To better secure hive parts, we sourced ‘z’ clips from Australia
which were far more effective, particularly during the monsoon (figure 4.6).
Hooks were attached to the sides of beehives and fencing wire was used to hang the hives from strong
wooden posts at a height of approximately 1.5 m. Each post was between 2.5 and 2.8 m length and
was treated with old engine oil, as an alternative to insecticides, for protection against termites prior
to fence building (figure 4.7).
LEFT: Figure 4.5 Langstroth beehive used in the beehive fence and secured with coconut rope ©
Kylie M. Butler. RIGHT: Figure 4.6. Langstroth beehive with the ‘z’ clips used to secure hive parts ©
Kylie M. Butler

133
Posts were embedded in 0.75 m holes with stones and soil packed around the base. Each beehive was
hung between two posts spaced approximately 3 m apart. Neighbouring beehives were approximately
8 m distance away and were connected to one another using single strands of strong, taut fencing
wire. Above each beehive, a shade roof was hung (figure 4.8).
Figure 4.7. Painting fence posts with old engine oil to protect from termites –
researcher Kylie M. Butler and field assistant Supun Herath © Morgan Tipper.
Figure 4.8. A beehive fence at the Jayasena household. © Kylie M. Butler

134
Over the course of the trial four types of shade roof were used: a) banana leaf on a stick frame; b)
dried grass on a stick frame; c) thin plywood with waterproof paint; and d) thick plywood with
waterproof paint (figure 4.9). Finally, although most beehive fences in Africa alternate ‘dummy
beehives’ (wooden boards cut to the same dimensions and painted the same colour as beehives) and
real beehives, it was decided that only real beehives would be used to help the beehive fences achieve
the strongest deterrent effect possible.
A total of ten beehive fences were built between October 2014 and September 2015. With the
assistance of Mr. Herath, I worked closely with each household to design individual fence layouts
taking into consideration: a) the direction elephants typically approached; b) any natural barriers such
as thick or tall vegetation; c) the boundaries of their home garden; and d) location of seasonal
vegetable or crop plots that might make it difficult to access the fence for maintenance or beekeeping.
As a result, the exact shape and size of each fence varied somewhat (Figure 4.10), with the smallest
Figure 4.9. Beehive fence shade roofs. Top Right: Mr. Somathilaka standing by his beehive fence with a traditional
woven banana leaf shade roof. Top Left: A ‘rice’ grass shade roof on the Guneris family’s beehive fence. Bottom
Right: A thin plywood shade roof warping on the Ariyadasa family’s fence. Bottom Left: A thick plywood shade roof
on the Somathilaka family’s fence. © Kylie M. Butler
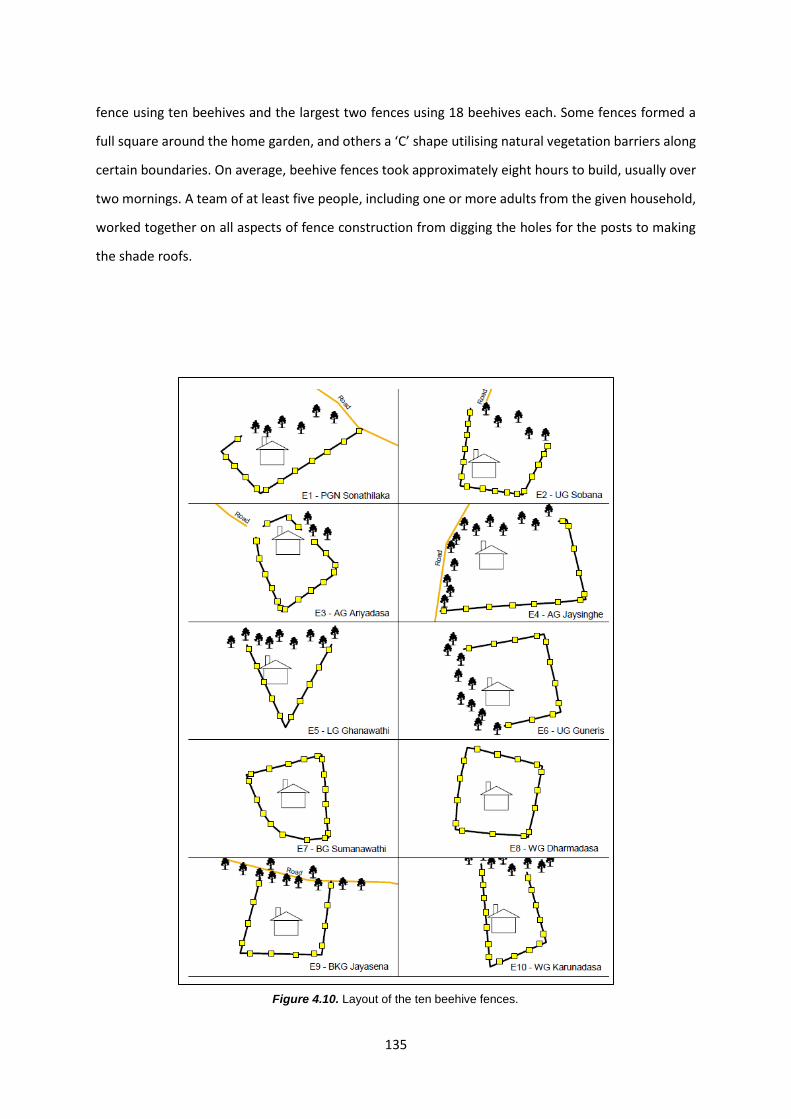
135
fence using ten beehives and the largest two fences using 18 beehives each. Some fences formed a
full square around the home garden, and others a ‘C’ shape utilising natural vegetation barriers along
certain boundaries. On average, beehive fences took approximately eight hours to build, usually over
two mornings. A team of at least five people, including one or more adults from the given household,
worked together on all aspects of fence construction from digging the holes for the posts to making
the shade roofs.
Figure 4.10. Layout of the ten beehive fences.
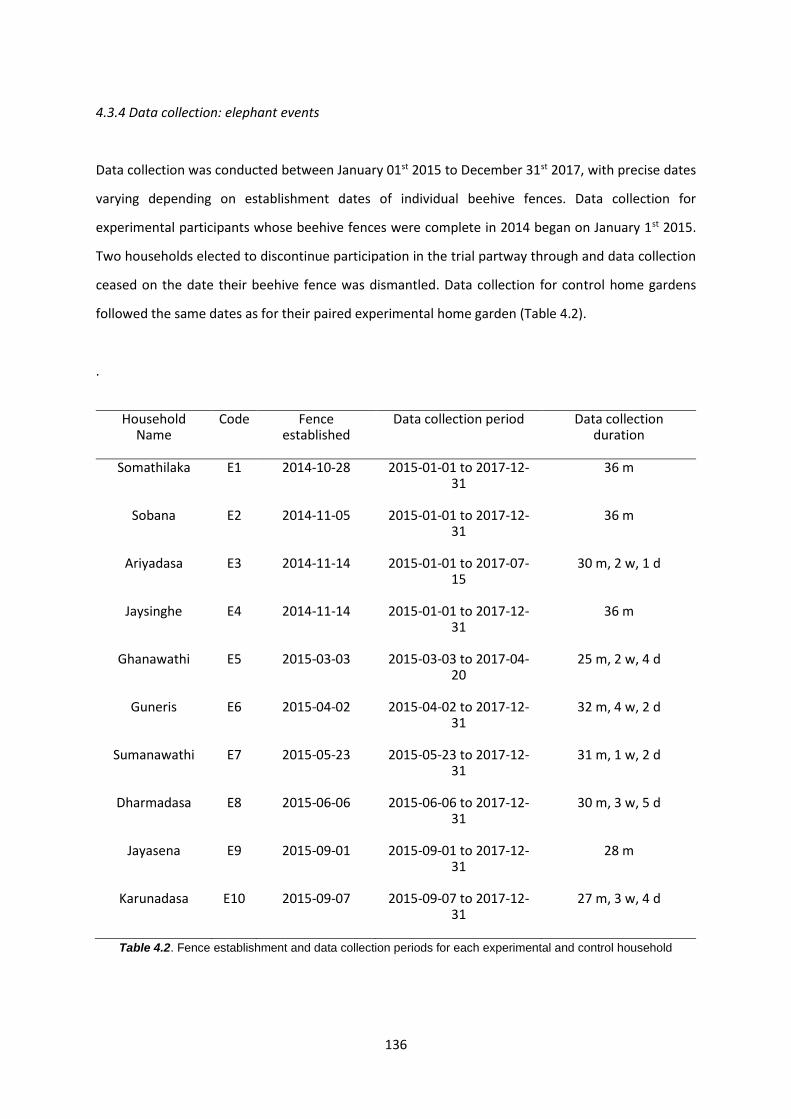
136
4.3.4 Data collection: elephant events
Data collection was conducted between January 01st 2015 to December 31st 2017, with precise dates
varying depending on establishment dates of individual beehive fences. Data collection for
experimental participants whose beehive fences were complete in 2014 began on January 1st 2015.
Two households elected to discontinue participation in the trial partway through and data collection
ceased on the date their beehive fence was dismantled. Data collection for control home gardens
followed the same dates as for their paired experimental home garden (Table 4.2).
.
Household Name
Code Fence established
Data collection period Data collection duration
Somathilaka E1 2014-10-28 2015-01-01 to 2017-12-31
36 m
Sobana E2 2014-11-05 2015-01-01 to 2017-12-31
36 m
Ariyadasa E3 2014-11-14 2015-01-01 to 2017-07-15
30 m, 2 w, 1 d
Jaysinghe E4 2014-11-14 2015-01-01 to 2017-12-31
36 m
Ghanawathi E5 2015-03-03 2015-03-03 to 2017-04-20
25 m, 2 w, 4 d
Guneris E6 2015-04-02 2015-04-02 to 2017-12-31
32 m, 4 w, 2 d
Sumanawathi E7 2015-05-23 2015-05-23 to 2017-12-31
31 m, 1 w, 2 d
Dharmadasa E8 2015-06-06 2015-06-06 to 2017-12-31
30 m, 3 w, 5 d
Jayasena E9 2015-09-01 2015-09-01 to 2017-12-31
28 m
Karunadasa E10 2015-09-07 2015-09-07 to 2017-12-31
27 m, 3 w, 4 d
Table 4.2. Fence establishment and data collection periods for each experimental and control household

137
Following the same protocol as previously detailed in ‘Chapter 3’, data collection utilised resident
reports of ‘elephant events’ collected during at least bi-monthly home visits to each experimental and
control household. However, here an ‘elephant event’ was defined as any sighting of one or more
elephants within 50 m of the home garden, as well as any successful crop-raids or property damage
that occurred within the home garden boundary. The same data sheets were used and again,
participants preferred to relay information verbally during visits rather than using the provided data
sheets, and this preference was accommodated.
4.3.5 Fence maintenance
At least one adult per experimental household was trained in basic fence maintenance. This involved
a bi-monthly walk around the beehive fence perimeter to ensure the following: a) that wires
connecting neighbouring hives were attached; b) posts were upright and strong; c) hives were hanging
approximately 1.5. m from the ground and not askew, and that shade roofs were not touching the
hives; d) empty hives were free of pests or debris; e) shade roofs were not damaged or falling apart;
and f) the area underneath beehives was clear of tall vegetation (Figure 4.11). It was recommended
that small tasks (e.g. reattaching connecting wires and straightening hives) were completed during the
check, while larger tasks (e.g. replacing fence posts or making new shade roofs) were scheduled for
completion as soon as possible.
However, although participants were requested to complete fence checks regularly, with support
available for any maintenance requiring new materials, most households failed to do this. As accurate
assessment of beehive fence deterrence required fences to be in good condition, our field team
performed fence checks at least monthly, asking farmers to assist when they were home. All
maintenance issues and tasks completed were documented on data sheets detailing the date,
description of maintenance issue, and whether it had been rectified.

138
Figure 4.11. Fence maintenance issues. Top Right: Researcher Kylie M. Butler, field assistant Supun Herath
and Mr. Sumanawathi raising the shade roofs. Top Left: Mr. Somathilaka straightens a crooked beehive. Centre:
Mr. Guneris repaints his post with engine oil to protect against termites. Bottom Right: A mouse escapes from a
beehive. Bottom Left: Chew damage from mice in an occupied beehive. All photos © Kylie M. Butler except Top
Right © Morgan Tipper

139
4.3.6 Beekeeping
At least one adult from each experimental household was trained in basic beekeeping skills including:
safe conduct around bees, colony inspection to assess strength and health, feeding sugar water,
attracting wild bees, transferring and splitting colonies, and honey extraction. Initial training
throughout 2015 consisted of home visits with a local beekeeping expert. In 2016 and 2017, follow-
up workshops and home visits were conducted by both international and local beekeepers (Table 4.3).
On-call support from our field team was available throughout the study period. Each experimental
household was provided with protective equipment including 2 x beekeeping face veils and hats,
beekeeping gloves and a smoker. Multiple beekeeping jackets were distributed throughout the village
for farmers to share when working with more active or aggressive hives.
Participants were encouraged to actively assist with occupied hive inspections at least quarterly.
Records of colony introductions or abscondments, hive checks and honey harvests were documented
using simple data sheets.
Dates Beekeeper Training type Skills covered
2015-03-16 and 2015-03-17
Dr. R.W.K. Punchihewa
Home visits Hive inspection, identifying comb parts, extracting pests, feeding sugar
water
2015-05-11, 2015-05-12 and
2015-05-25
Dr. R.W.K. Punchihewa
Home visits Introducing colonies, smoking bees and hive inspections
2015-11-24 and 2015-11-25
Dr. R.W.K Punchihewa
Home visits Checking hive health, feeding sugar water
2016-10-03 to 2016-10-07
Dr. D. Gormley O-Brien and
Miss E. Collins
Home visits and group workshop
Hive inspections and identifying issues (pests, weak colonies), feeding bees,
working with aggressive colonies
2017-03-01 and 2017-03-02
Mr. F. Ryde Home visits and group workshop
Attracting wild swarms, splitting colonies, transferring colonies from
clay pots, basic honey harvesting
2017-05-31 and 2017-06-01
Mr. F. Ryde Home visits and group workshop
Splitting and transferring colonies, honey harvesting
Table 4.3 Beekeeping training and workshop record

140
4.3.7 Cost analysis
In order to determine the overall costs of building and maintaining a beehive fence, and to assess its
affordability for small-scale rural farmers in Sri Lanka, all expenses incurred were recorded. This
included all fencing materials, basic tools, basic beekeeping protective equipment, colonies and colony
delivery, and replacement materials required for maintenance. A minimum cost was then calculated
based on initial set-up requirements for a 50% hive occupation rate.
4.3.8 Farmer acceptance and willingness to participate in using beehive fencing
Farmer acceptance of, and willingness to participate in, using beehive fencing to deter elephants was
assessed through one-on-one social research interviews, informal conversations, and researcher
observations. One-on-one interviews were conducted at the participants home with a local translator.
These involved a series of statements ranked by Likert scale, relating to the perceived effectiveness
of, and interest in continuing to use, beehive fencing. Informal conversations, translated by field
assistant Mr. Herath, occurred while visiting households for data collection or working together on
fence maintenance or beekeeping tasks. These conversations enabled a free-flowing exchange of
ideas and opinions in a relaxed setting. Finally, researcher observations were incorporated into the
overall assessment, as verbally expressed enthusiasm for beehive fencing by participants was not
always in-line with on-ground action.
It must be acknowledged these modes of data collection are largely subjective and reliant on
anecdotal information. However, given the aim of implementing a genuine participatory community-
based deterrent, information gained through frequent interactions with participants in a relaxed
setting has particular relevance. Still it is acknowledged that such qualitative data should be
interpreted with some caution.
4.3.9 Statistical analysis
To identify whether a difference in frequency of elephant events occurred between experimental
(beehive fences present) and control (beehive fences absent) farms, a paired-sample t-test was used.
To assess whether elephants were less likely to enter a family’s home garden area when a beehive

141
fence was employed, I used Pearson’s chi-square test of independence (χ²) with Yate’s continuity
correction.
To assess whether the frequency with which elephants breached a fence perimeter was influenced by
the amount of hives overall, or specifically the number of occupied hives, I used a generalised linear
mixed model (GLMM), assuming a poisson distribution. The number of hives present, and the number
of occupied hives, were included as predictor variables, with the farm location fitted as a random
intercept to allow for the repeated-measure comparison of fences containing different levels of hive
occupations at different times. Pearson correlations were used to assess collinearity between
predictors before inclusion in the model. Model fit was determined using the marginal and conditional
r-squared (Nakagawa and Schielzeth 2013).
Statistical analyses were conducted with the R Studio Statistical Package 3.5.1 (R core team 2018).
Generalised Linear Mixed Models (GLMM’s) were constructed using the package ‘lme4’ (Bates et al.
2014), with model predictions generated using the ‘effects’ package (Fox 2003). All visual
representations were constructed using ‘ggplot2’ (Wickham 2016). Results were considered
significant when p <0.05.
4.4 Results
4.4.1 Elephant events and deterrent effect of beehive fences
A total of 176 elephant events were reported by experimental (64%, n = 122) and control (36%, n =
64) households, with elephant events occurring significantly more frequently at experimental home
gardens (M = 11.2, ± SE = 1.8, n = 122) compared to control home gardens (M = 6.4, ± SE = 0.90, t(9) =
2.71, p = 0.02) (Figure 4.12).
The likelihood of elephants physically entering home gardens also differed. Here, elephants entered
control home gardens on 83% (n = 64) of approaches compared to only 43% (n = 122) for experimental
home gardens. This shows a significant increase in the likelihood of elephants breaching the home
garden boundary of unfenced control gardens (M = 85.66%, ± SE 4.21%) compared to the beehive
fenced perimeter of experimental farms ((M = 37.55%, ± SE 6.69%) (χ² (1, N = 176) = 24.98, p <0.01)
(figure 4.13).
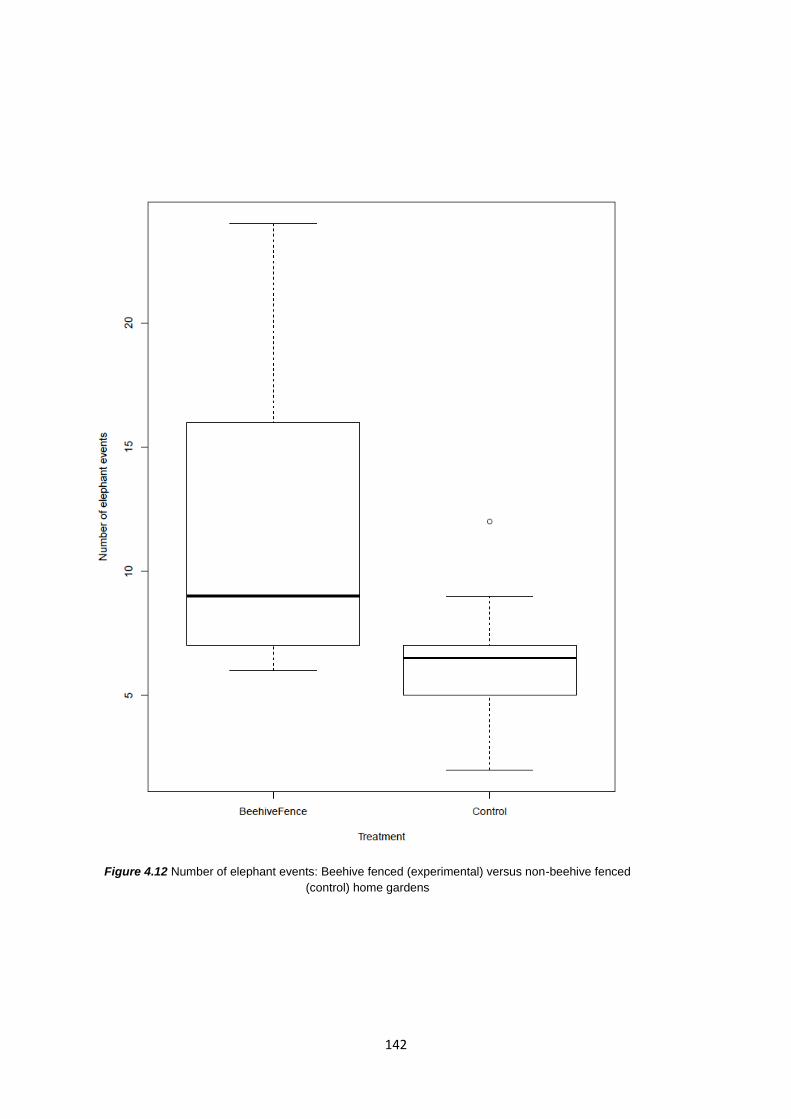
142
Figure 4.12 Number of elephant events: Beehive fenced (experimental) versus non-beehive fenced
(control) home gardens
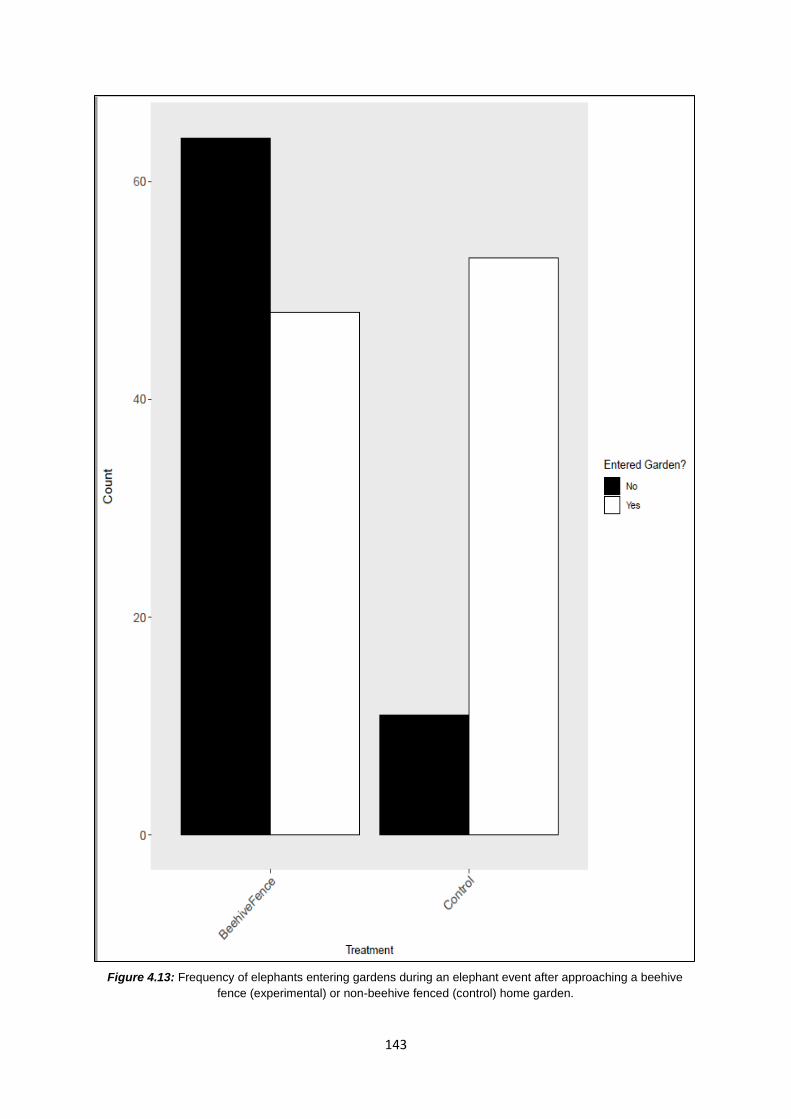
143
Figure 4.13: Frequency of elephants entering gardens during an elephant event after approaching a beehive
fence (experimental) or non-beehive fenced (control) home garden.
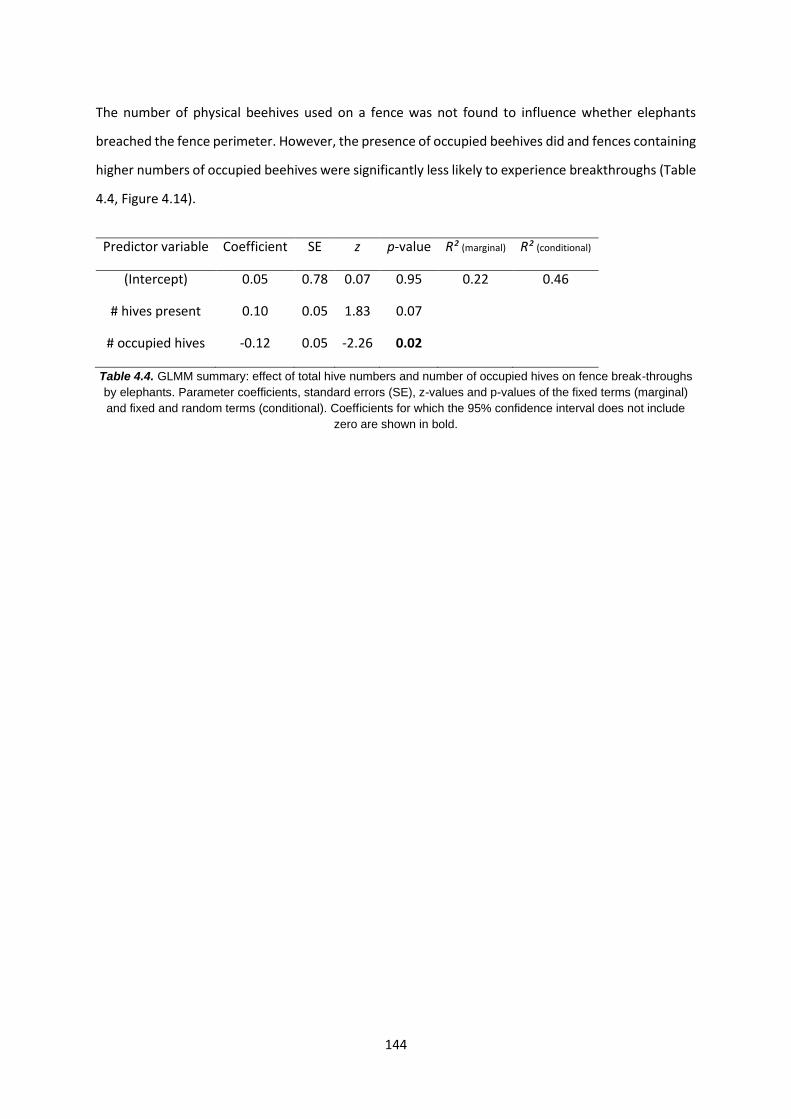
144
The number of physical beehives used on a fence was not found to influence whether elephants
breached the fence perimeter. However, the presence of occupied beehives did and fences containing
higher numbers of occupied beehives were significantly less likely to experience breakthroughs (Table
4.4, Figure 4.14).
Predictor variable Coefficient SE z p-value R² (marginal) R² (conditional)
(Intercept) 0.05 0.78 0.07 0.95 0.22 0.46
# hives present 0.10 0.05 1.83 0.07
# occupied hives -0.12 0.05 -2.26 0.02
Table 4.4. GLMM summary: effect of total hive numbers and number of occupied hives on fence break-throughs
by elephants. Parameter coefficients, standard errors (SE), z-values and p-values of the fixed terms (marginal)
and fixed and random terms (conditional). Coefficients for which the 95% confidence interval does not include
zero are shown in bold.
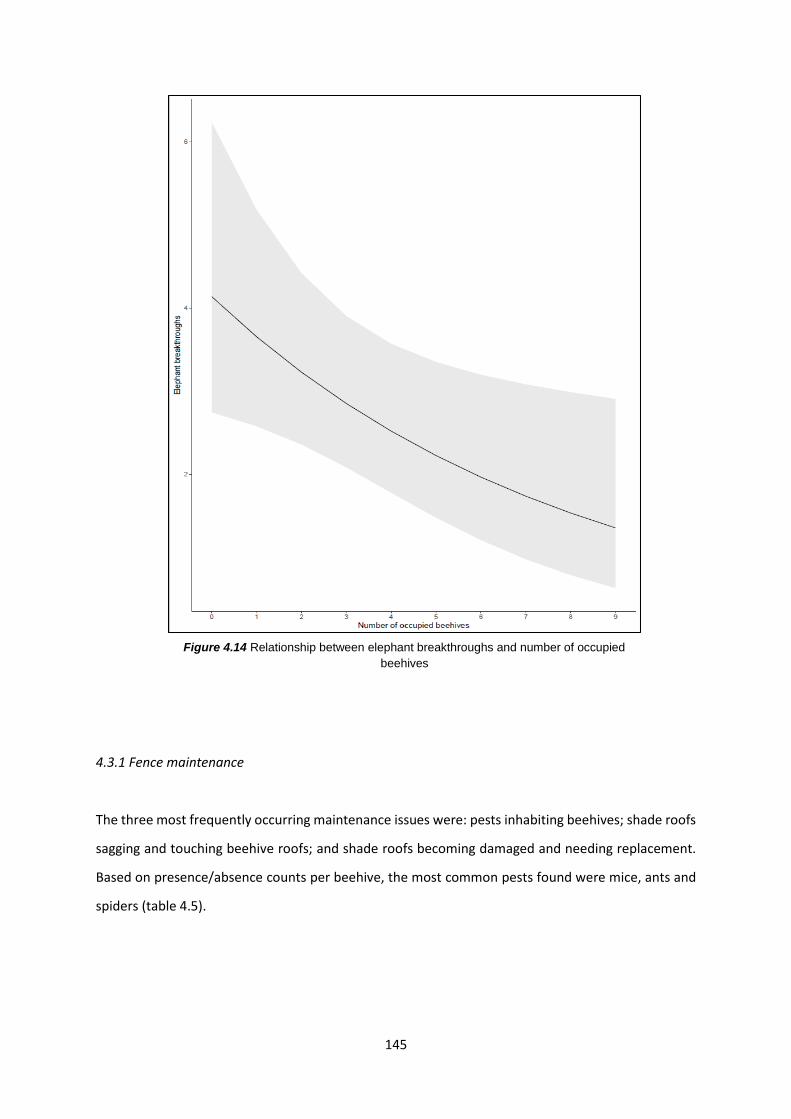
145
4.3.1 Fence maintenance
The three most frequently occurring maintenance issues were: pests inhabiting beehives; shade roofs
sagging and touching beehive roofs; and shade roofs becoming damaged and needing replacement.
Based on presence/absence counts per beehive, the most common pests found were mice, ants and
spiders (table 4.5).
Figure 4.14 Relationship between elephant breakthroughs and number of occupied
beehives

146
Year Ants Mice Lizards Frogs Spiders Wasps Snakes Other
2015 55 52 19 11 25 0 2 5
2016 55 86 15 3 21 5 0 15
2017 31 58 8 3 11 6 6 3
Total 141 192 42 14 57 11 8 23
Table 4.5. Presence/absence counts of different pests found in beehives. Bold text denotes the three most
frequently occurring pests per year and overall. The ‘Other’ category includes cockroaches, beetles, palm
squirrels and a bird. Whilst the number of beehives present throughout the study varied according to fence
establishment dates and due to two fences being dismantled in 2017, all hives present were checked monthly for
each of the listed pests. Thus, the table gives an indication of which pests most frequently occurred each year of
the study, as well as overall.
The shade roofs required raising and straightening at least monthly. This involved tightening the wires
around the cross bars, took less than a minute per hive and could be done quite easily by one person.
Shade roofs were frequently damaged by wind or rain and two or more shade roofs per fence required
replacing on 60 occasions. In September 2015 and March 2016, all original grass or banana leaf shade
roofs needed replacing. By August 2016, all grass roofs had been replaced with thin plywood however
these warped during the rainy season. Thick plywood roofs were installed instead in February 2017,
which were more durable but still vulnerable to cracking in the heat.
Other maintenance issues included: farmers leaving connecting wires unattached, overgrown
vegetation underneath hives, beehives hanging askew, damaged beehives or hive parts, and damaged
or weak posts. Fire-damage occurred accidentally at four fences when farmers burnt the land adjacent
to the fence in preparation for planting. All fence posts were replaced at least once during the three-
year period, as a result of termite damage or general weathering.
All households experienced the same types of fence maintenance issues, however those who checked
their fences regularly and rectified issues promptly spent less time on maintenance overall.
Quantifying precise durations of time spent on maintenance was difficult given the different sizes of
fences, capacities of households, and specific maintenance tasks required. However, based on our
experiences assisting with maintenance and information from participants, a fence check including
rectification of basic maintenance issues took between one to two and a half hours with two people,

147
and was required bi-monthly. Larger maintenance issues, such as replacing shade roofs or posts,
needed to be addressed approximately every three months, and took between four to eight hours
with a team of four people.
4.4.3 Beekeeping and honey harvests
The total number of beehives across all ten fences was 142 and of these, 76 (53.5%) hives were
occupied at least once. Overall, 100 colonies occupied beehives. Of these, 23 occupations occurred
naturally when wild honey bees entered the fence hives, and a further three colonies were transferred
into fence hives after being found in clay pots. Despite efforts to attract wild colonies by placing clay
pots and traditional log hives in gardens, wild honeybee occupation rates were low. Thus, most hive
occupations resulted from purchased colonies which were sourced from Kandy or Colombo then
transported to Dewagiriya Village where they were transferred into fence hives. Overall, 74 colonies
were introduced this way.
Of the 100 colonies, 63 absconded before the end of the trial. Excluding failed introductions, where
the colony absconded within seven days of hive transfer, the mean duration of occupation by colonies
was 201 days ((SE ± 26.5). The number of hives occupied at least once, the number and mode of colony
introductions, and the number of abscondments varied per fence (Table 4.6).
All households experienced periods where no active hives were present on their fences, particularly
following fence establishment until the first wild occupation occurred or purchased colonies could be
sourced. Additionally, four fences experienced periods where the number of absconded colonies
exceeded introductions and active hive presence reverted back to 0%. Overall, the mean ‘active hive’
rate per fence was 17.8%. Active hive rates were somewhat biased by an uneven distribution of
purchased colonies. Given the difficulties associated with sourcing and transporting healthy colonies,
and the expense of purchase, more colonies were allocated to beehive fences where participants
demonstrated active involvement in fence maintenance and beekeeping, rather than equally dividing
colonies amongst participants (Table 4.7).
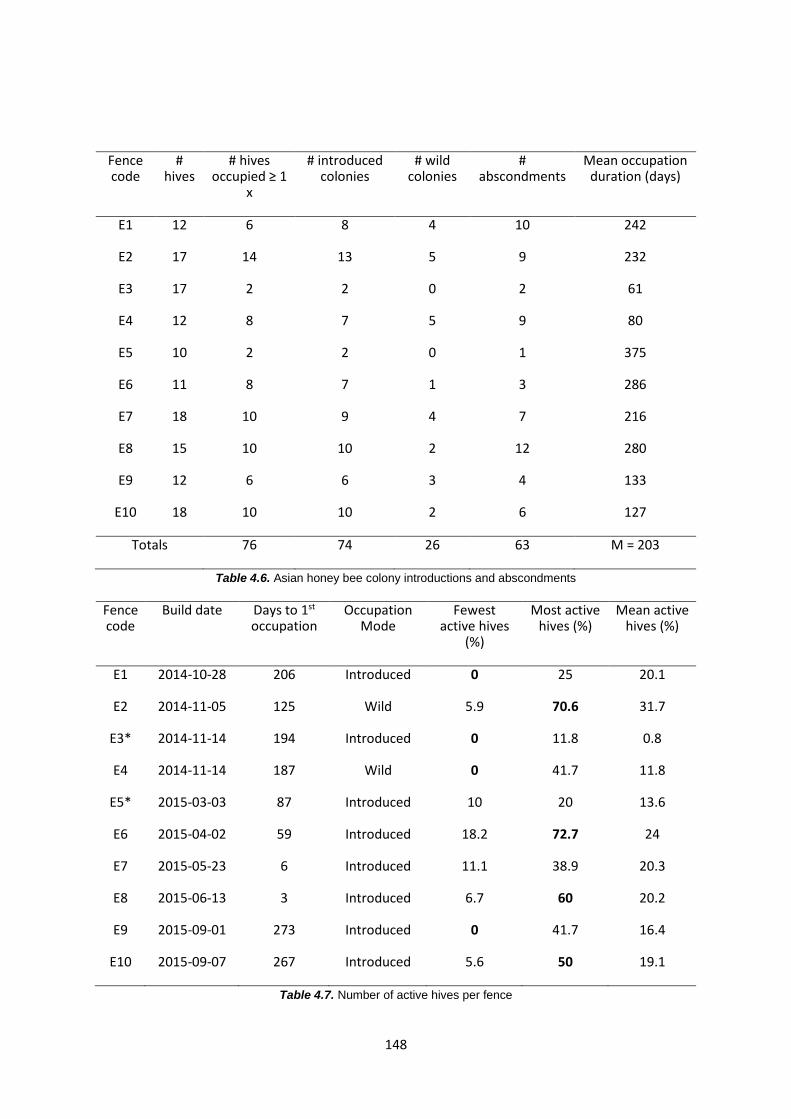
148
Fence code
# hives
# hives occupied ≥ 1
x
# introduced colonies
# wild colonies
# abscondments
Mean occupation duration (days)
E1 12 6 8 4 10 242
E2 17 14 13 5 9 232
E3 17 2 2 0 2 61
E4 12 8 7 5 9 80
E5 10 2 2 0 1 375
E6 11 8 7 1 3 286
E7 18 10 9 4 7 216
E8 15 10 10 2 12 280
E9 12 6 6 3 4 133
E10 18 10 10 2 6 127
Totals 76 74 26 63 M = 203
Table 4.6. Asian honey bee colony introductions and abscondments
Fence code
Build date Days to 1st occupation
Occupation Mode
Fewest active hives
(%)
Most active hives (%)
Mean active hives (%)
E1 2014-10-28 206 Introduced 0 25 20.1
E2 2014-11-05 125 Wild 5.9 70.6 31.7
E3* 2014-11-14 194 Introduced 0 11.8 0.8
E4 2014-11-14 187 Wild 0 41.7 11.8
E5* 2015-03-03 87 Introduced 10 20 13.6
E6 2015-04-02 59 Introduced 18.2 72.7 24
E7 2015-05-23 6 Introduced 11.1 38.9 20.3
E8 2015-06-13 3 Introduced 6.7 60 20.2
E9 2015-09-01 273 Introduced 0 41.7 16.4
E10 2015-09-07 267 Introduced 5.6 50 19.1
Table 4.7. Number of active hives per fence

149
Honey production was low and only six households harvested any honey throughout the trial, totalling
approximately seven litres. The most honey harvested from any individual beehive fence was two
litres and no honey at all was harvested from four fences (table 4.8).
Fence code Honey harvested (L) Month/year
E1 0.5 Mar 2017
E2 0.5 Feb 2017
E3 0
E4 0
E5 0.5 Mar 2017
E6 2 Mar 2017, Sep 2017, (1 L each)
E7 1.0 Mar 2016
E8 0.5 Mar 2017
E9 0
E10 2 Mar 2017
Total 7
Table 4.8 Honey harvesting records per household
Common challenges impacting beekeeping across all experimental fences included: lack of adequate
shade, insufficient food and water resources for bees, pests entering hives (including ants and mice);
and wax moth invasions. Although it is difficult to quantify the precise effects of the aforementioned
challenges, it is reasonable to assume that each contributed to the low rate of wild occupations, the
high instance of colony abscondments, and poor honey production.
4.4.5 Cost of beehive fences
The initial set-up cost of each fence varied somewhat depending on precise fence layout. Efforts were
made to minimise costs by using local materials and labour.
Estimates for the minimum include: beehives (15 USD, 2 626 LKR each), posts (3 USD, 525 LKR each),
cross bars (0.50 USD, 87 LKR each), shade roofs (3.50 USD, 612 LKR each), strong wire for hanging and
connecting beehives (3.50 USD, 612 LKR, p/kg), thin wire for hanging shade roofs (1.35 USD, 236 LKR,
p/kg), and ‘z’ clips to keep moveable hive parts secure (1 USD, 175 LRK, each) (Table 4.9).
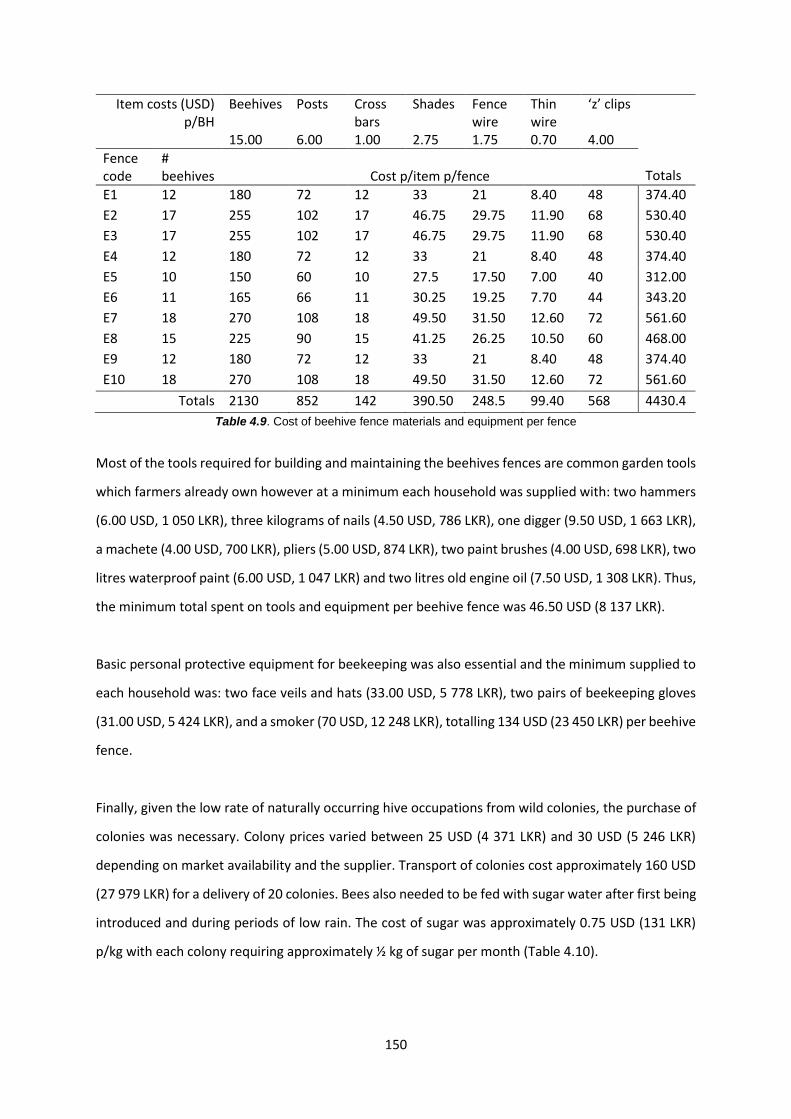
150
Item costs (USD) p/BH
Beehives 15.00
Posts 6.00
Cross bars 1.00
Shades 2.75
Fence wire 1.75
Thin wire 0.70
‘z’ clips 4.00
Totals
Fence code
# beehives
Cost p/item p/fence
E1 12 180 72 12 33 21 8.40 48 374.40
E2 17 255 102 17 46.75 29.75 11.90 68 530.40
E3 17 255 102 17 46.75 29.75 11.90 68 530.40
E4 12 180 72 12 33 21 8.40 48 374.40
E5 10 150 60 10 27.5 17.50 7.00 40 312.00
E6 11 165 66 11 30.25 19.25 7.70 44 343.20
E7 18 270 108 18 49.50 31.50 12.60 72 561.60
E8 15 225 90 15 41.25 26.25 10.50 60 468.00
E9 12 180 72 12 33 21 8.40 48 374.40
E10 18 270 108 18 49.50 31.50 12.60 72 561.60
Totals 2130 852 142 390.50 248.5 99.40 568 4430.4
Table 4.9. Cost of beehive fence materials and equipment per fence
Most of the tools required for building and maintaining the beehives fences are common garden tools
which farmers already own however at a minimum each household was supplied with: two hammers
(6.00 USD, 1 050 LKR), three kilograms of nails (4.50 USD, 786 LKR), one digger (9.50 USD, 1 663 LKR),
a machete (4.00 USD, 700 LKR), pliers (5.00 USD, 874 LKR), two paint brushes (4.00 USD, 698 LKR), two
litres waterproof paint (6.00 USD, 1 047 LKR) and two litres old engine oil (7.50 USD, 1 308 LKR). Thus,
the minimum total spent on tools and equipment per beehive fence was 46.50 USD (8 137 LKR).
Basic personal protective equipment for beekeeping was also essential and the minimum supplied to
each household was: two face veils and hats (33.00 USD, 5 778 LKR), two pairs of beekeeping gloves
(31.00 USD, 5 424 LKR), and a smoker (70 USD, 12 248 LKR), totalling 134 USD (23 450 LKR) per beehive
fence.
Finally, given the low rate of naturally occurring hive occupations from wild colonies, the purchase of
colonies was necessary. Colony prices varied between 25 USD (4 371 LKR) and 30 USD (5 246 LKR)
depending on market availability and the supplier. Transport of colonies cost approximately 160 USD
(27 979 LKR) for a delivery of 20 colonies. Bees also needed to be fed with sugar water after first being
introduced and during periods of low rain. The cost of sugar was approximately 0.75 USD (131 LKR)
p/kg with each colony requiring approximately ½ kg of sugar per month (Table 4.10).

151
Colonies
Transport
Sugar
Cost estimate USD 27.50 ea 160 p/20 colonies
0.75 p/kg
Fence code # colonies for 50% occupation
Cost per item per fence
Totals
E1 6 165 48 18 231 E2 9 247.50 72 27 346.5 E3 9 247.50 72 27 346.5 E4 6 165 48 18 231 E5 5 137.50 40 15 192.5 E6 6 165 48 18 231 E7 9 247.50 72 27 346.5 E8 8 220 64 24 308 E9 6 165 48 18 231 E10 9 247.50 72 27 346.5
Totals 2007.5 584 219 2810.5 Table 4.10 Cost estimates for colony establishment and feeding per fence
Fence code BHF materials Tools/equipment PPE Colonies (incl. transport and sugar)
Totals (USD)
E1 374.40 46.50 134.00 231.00 785.90 E2 530.40 46.50 134.00 346.50 1057.40 E3 530.40 46.50 134.00 346.50 1057.40 E4 374.40 46.50 134.00 231.00 785.90 E5 312.00 46.50 134.00 192.50 685.00 E6 343.20 46.50 134.00 231.00 754.70 E7 561.50 46.50 134.00 346.50 1088.50 E8 468.00 46.50 134.00 308.00 956.50 E9 374.40 46.50 134.00 231.00 785.90 E10 561.60 46.50 134.00 346.50 1088.50
Totals (USD) 4430.40 465.00 1340.00 2810.50 9045.90 Table 4.11. Cost estimate for fencing materials, tools/equipment and colonies overall (per fence)
Accounting for all the expenses detailed above, the cost of establishing the physical beehive fence,
introducing colonies to reach a 50% hive occupation target, supplying households with protective
beekeeping equipment and feeding sugar water to bees ranged between 685.00 USD (119 875 LKR)
and 1088.50 USD (190 488 LKR), with a mean cost of 904.57 (±SE 50.58) (158 299 LKR, ±SE 8 851)
(Table 4.11). Replacement of posts and plywood shade roofs when necessary (estimated 18-month
intervals if well looked after) are additional on-going expenses that need to be considered.

152
Additionally, if bee colonies abscond and need to be re-introduced from suppliers, this adds
substantially to ongoing costs.
4.4.6 Farmer acceptance and willingness to participate
Enthusiasm for trialling beehive fences as a HEC mitigation tool was expressed during initial
community meetings and further demonstrated by over half of the households in Dewagiriya Village
requesting to participate. Still, people were unsure whether Asian honey bees would actually have a
deterrent effect on elephants, and at least part of people’s enthusiasm in the trial could be attributed
to the prospect of harvesting honey for personal use or income generation.
One-on-one interviews conducted in October 2017 with all ten experimental households, revealed
that only one found their beehive fence to be ‘very’ effective, while a further five households stated
the beehive fence was ‘somewhat’ effective. Five households believed elephants were deterred by
the physical structure of the fence as the connecting wires resembled those on an electric fence,
rather than being deterred by the presence of bees. Only two households felt that Asian elephants
were afraid of Asian honey bees.
Participants from all households said they had the theoretical knowledge to look after their fences but
were limited by physical skills or resources, resulting in two households withdrawing from the trial in
2017. Of the remaining eight households, only four said they ‘definitely’ planned to continue using
their beehive fences. The other four felt that without on-going in-field support they did not have
sufficient beekeeping skills. No household had the financial means to replace colonies or broken fence
parts such as posts or hives. However, all eight remaining households stated they wanted to continue
beekeeping for honey production, despite low honey yields during the trial.
Researcher observations concurred with expressed challenges in the capacity to independently look
after fences but also found motivation to be an issue. By the end of 2017, only three households
consistently kept up with basic maintenance requirements and beekeeping. Of the remaining five
households, two were limited in their ability to care for their fences due to being physically incapable
or lacking time. The remaining three households appeared to simply not put in the minimum effort
required. Overall, fluctuations in motivation were evident throughout the trial. Peaks were observed

153
after initial fence establishment and following colony introductions. Troughs were observed in the
period between building the fences and introducing the first colonies, and when hive occupations
were low. Farmers were much more active when working alongside our team, or when consistently
reminded what to do.
4.5 Discussion
Given that crop-raiding has been a recognised HEC issue for decades (Fernando et al. 2005; de Silva
1998; Santiapillai 1996), it is no surprise that a variety of deterrent techniques have been trialled and
that many of them have demonstrated success in deterring elephants (Baishya et al. 2012; Gunaryadi
et al. 2017; Davies et al. 2011; Thapa 2010; Perera 2009). It’s then fair to pose the question: why does
HEC, and specifically crop-raiding, remain such a major issue negatively impacting both elephant and
human communities?
One reason could be that most studies either focus specifically on, or publish information primarily
about, the success of the deterrent effect by quantifying elephant visits or damage incurred (Nair and
Jayson 2016; Baishya et al. 2012; Gross et al. 2017; Gross et al. 2016; Santiapillai 1996). Of course, if
the goal is to prevent elephants from entering a certain location or damaging certain crops, the
capacity of the given deterrent to do this, is a major factor in its success, and an important first step.
However, numerous factors including labour intensiveness, cost-effectiveness, and stakeholder
acceptance, also influence the success of a deterrent in a practical context. More and more studies
are incorporating an increasingly holistic approach into the evaluation of HEC mitigation techniques
(Gunaryadi et al. 2017; King et al. 2017; Changa et al. 2016; Dickman 2010; Graham and Ochieng 2008;
Sitati and Walpole 2006). It is hoped that this will be benefit management planning by facilitating
identification of appropriate deterrent techniques for various HEC locations, that are effective not
only in theory but in practice.
We incorporated a holistic approach in the overall evaluation of our three-year beehive fence trial in
Dewagiriya. The capacity of the beehive fences to deter elephants from people’s home gardens,
maintenance and beekeeping challenges, cost-effectiveness, and farmer motivation were all
important factors in determining whether beehive fencing had potential to help the local community
alleviate their problems with elephants. Taking these factors together, we identified some positive

154
indicators of success but also highlighted several challenges that need to be considered carefully if this
technique is to be recommended as an Asian HEC mitigation tool.
If deterrent effect is the first indicator of success, the beehive fence trial in Dewagiriya ticked this
initial box. Results showed evidence that the beehive fence had the capacity to deter Asian elephants
from areas within the fence perimeters. Significantly less elephant incursions occurred in home
gardens surrounded by beehive fences compared to those without. The deterrent effect also appeared
to strengthen with increased hive occupations, which is a promising indicator that elephants were
deterred by the presence of bees rather than simply the physical fence structure.
Still, elephants entered home gardens of beehive fenced properties on 43% of approaches. Although
this is a vast improvement on the 83% of elephant entries that occurred at non-fenced gardens, the
question remains whether this is a sufficient reduction to warrant wider implementation of beehive
fences throughout the village. Elephants often cause considerable damage during even a single crop-
raiding event to crop or vegetables plots, fruit trees, or even houses. A much higher deterrent rate
would be required before community members could relax their vigilance and be confident the
beehive fences could keep them and their gardens safe.
No comparable beehive fence trials exist in Asia, from which to judge whether a 57% deterrent rate
indicates good, bad or average performance. Our methods were based on research trials located in
Kenya, with African honey bees and African elephants. Here, a deterrent success rate of approximately
80% was observed, and this is a clear positive benefit to crop protection (King et al. 2017). Ideally, we
would have liked to have seen a similar success rate in Dewagiriya Village however biological
differences in the aggression levels of the two bee species was always a concern, with the Asian honey
bees comparatively less aggressive (Punchihewa 1994). Based on responses to audio playback
experiments, Asian elephants do elicit avoidance and exploratory responses to the perceived presence
of bees however with less intensity than African elephants (King et al. 2018). It is therefore not
surprising that the deterrent effect observed in beehive fence trials across the two continents
mirrored this pattern. Expanding on the anecdotal newspaper reports (Dutta 2013; Manoj 2012) and
short-term trial in India (Nair and Jayson 2016) by establishing further beehive fence trials following a
similar protocol to King et al. (2011; 2017) and using a multi-factor evaluation process would provide

155
a basis for comparison and help make an informed decision about the worthiness of pursuing beehive
fences using Asian honey bees as a deterrent technique.
The lower levels of aggression weren’t the only challenge presented by bees in Dewagiriya Village.
Natural colonies were difficult to attract into fence hives. On advice from local beekeeping experts,
traditional log and clay-pot hives were strategically placed near homes to attract wild swarms, and
bees wax smeared on hive entrances. Still, only 26 of 100 hive occupations occurred naturally,
rendering the purchase of colonies from Kandy or Colombo necessary, and adding additional expense
to the project budget. Colony health was often weak, indicating that local food sources were
insufficient, even during the rain seasons. We had only ten fences in the village each with between
ten and 18 beehives, yet the mean hive occupation rate per fence was only 17.8% which was far below
our target of 50% occupation rates per fence. This begs the question as to what the saturation point
for supporting bee colonies in Dewagiriya Village would be, and had we already exceeded it? Further
research investigating what level of hive occupation is required to consistently deter elephants, and
how the local environment could be improved to better support bee health, would assist in
determining how feasible it would be to expand beehive fences in Dewagiriya Village.
Whether a deterrent is financially viable for a community is another important consideration. The
beehive fence is described as a low-cost, low-maintenance community based-deterrent (King et al.
2011) and compared to large-scale techniques such as electric fencing, expenses are relatively low.
Still, HEC disproportionately affects poorer communities and it is vital that costs are considered in the
context of the economic status of the community in which it is being proposed. In Dewagiriya Village,
the mean cost of building a beehive fence and introducing a 50% rate of hive occupations exceeded
the mean annual household income (904 USD versus 747 USD). Other studies suggest that economic
benefits from honey production and other bee products could help off-set costs (King et al. 2017).
However, in Dewagiriya Village honey production was low and no economic benefits were generated
from this. The most honey any single household extracted over the three-year period was 2 kg’s and
while farmers enjoyed the small quantities of honey, it was nowhere near a level required to
contribute financially. Realistically, outside funding to support the establishment of beehive fences
would be essential for most communities requiring one.

156
The last factor incorporated into our evaluation was farmer motivation and willingness to participate.
The largely qualitative and subjective nature of assessing people’s motivations makes it difficult to
assess, and even more difficult to compare across studies. Still, it is too important to discount. For
community-based mitigation, regardless of how strong the deterrent effect of a particular technique
is, if farmers have no interest in actively using the technique, it will undoubtedly fail.
In Dewagiriya Village, the motivation of participants ebbed and flowed. Farmers verbally expressed
enthusiasm for the project however this did not always translate into active participation on-ground.
During important milestones, such as establishing the first fences and introducing colonies, members
of all households were actively involved. However, during lulls in hive occupations or when people
were busy planting or harvesting their fields, beehive fences were often neglected. To ensure fences
were well-maintained so we could properly assess deterrent effect, our field team often did
maintenance work alone. This perhaps led to expectations that households did not have to take
responsibility for their own fences. Farmers who were enthusiastic about beekeeping took the best
care of their fences. Overall, only two households were capable of looking after their fences
independently, and two households withdrew from the trial in 2017. When asked if they wanted to
continue using their fences after the three-year trial period, only four households said yes. Most felt
they could not manage their bees without assistance, and that costs of even small repairs would
unaffordable.
When assessing the feasibility of using chili-based deterrents to combat crop-raiding in Indonesia,
Hedges and Gunaryadi (2009) found that participation rates were higher if farmers had confidence in
the effectiveness of the proposed deterrent technique. In Dewagiriya Village, people seemed hopeful
that the beehive fences would help deter elephants but also expressed scepticism about whether it
would work. All participating households said they were motivated by the potential benefits of
harvesting honey. King et al. (2017) found that Kenyan farmers tended to fence maintenance issues
promptly as they recognised the tangible economic benefits from honey yields. Although not explicitly
stated, it is likely low honey production negatively impacted motivation to maintain fences.
Another potential issue with farmer motivation is that engendering a sense of ownership and
responsibility for HEC mitigation with a community can require a major shift in thinking, especially if
they are used to relying on outside assistance (Osborn and Parker 2003). Although the Dewagiriya

157
community felt that outside assistance was lacking, all participants felt that the Wildlife Department
should be responsible for keeping elephants away from their village (researcher pers. comm). Most
people felt that they lacked the financial or physical capacity to deter elephants from their homes,
and, for farmers who struggled with their fences, introducing a technique requiring the development
of new skills (beekeeping) may not have generated a sense of ‘empowerment’ and instead validated
the challenging nature of HEC mitigation.
4.6 Conclusion
Across all elephant range countries, numerous communities struggle to cope with crop-raiding
elephants, and the challenge of identifying feasible and effective elephant deterrents is real. Since it
was first developed and trialled in Kenya, beehive fencing has attracted interest as a practical eco-
deterrent that can be implemented at the community level. Despite obvious differences in the biology
and behaviour of the African and Asian elephant and honey bee species, interest in using beehive
fencing in Asia has been strong. Our trial provided the first long-term opportunity to evaluate the
practical application of beehive fencing as a deterrent for Asian elephants, incorporating not only
deterrent effect but also beekeeping challenges, cost effectiveness, and farmer motivation into future
recommendations.
Despite promising signs of being a relatively effective deterrent with a > 50% success rate at keeping
elephants from entering home gardens, it is unlikely that beehive fences could contribute
meaningfully to mitigating HEC at a community level in Dewagiriya Village. The difficulties in attracting
natural colonies, poor honey production and apparent lack of food resources for bees were consistent
over the study period, suggesting the saturation rate for supporting viable colonies in Dewagiriya
Village is low. Economically, farmers were unable to meet even small expenses such as replacing
damaged posts or hives, and financial support would be required for both initial set-up and ongoing
maintenance. Finally, although farmers seemed to enjoy participating in the project, motivation or
capacity to independently look after their fences was low, and it is unlikely that most of the existing
fences will continue to be used following this trial. Given the promising signs of deterrent success, it
would be worth exploring the potential of beehive fencing as an Asian elephant deterrent tool further.
However, future trials would benefit from careful selection of HEC location more amenable to
beekeeping.

158
4.7 References
Amwata, DA & Mganga, KZ 2014, 'The African elephant and food security in Africa: experiences from
Baringo District, Kenya', Pachyderm, vol. 55, pp. 22 – 28.
Bandara, R & Tisdell, C 2003, 'Comparison of rural and urban attitudes to the conservation of Asian
elephants in Sri Lanka: empirical evidence', Biological Conservation, vol. 110, pp. 327 – 342.
Bandara, R & Tisdell, C 2002, 'Asian elephants as agricultural pests: economics of control and compensation in Sri Lanka', Natural Resources Journal, vol. 42(3), pp. 491-519.
Baishya, HK, Dey, S, Sarmah, A, Sharma, A, Gogoi, S, Aziz, T, Ghose, D & Williams, AC 2012, 'Use of chilli
fences to deter Asian elephants – a pilot study', Gajah, vol. 36, pp. 11 – 13.
Barnes, RFW, Danquah, E, Hema, EM, Dubiure, UF, Manford, M, Nandjui, A & Boafo, Y 2015,
'Retrospective versus prospective designs for studies of crop-raiding by elephants in Kakum, Ghana',
Pachyderm, vol. 56, pp. 44 – 50.
Barua, M, Bhagwat, SA & Jadhav, S 2013, 'The hidden dimensions of human-wildlife conflict: health
impacts, opportunity and transaction costs', Biological Conservation, vol. 157, pp. 309 – 316.
Campos-Arceiz, A, Takatsuki, S, Ekanayaka, SKK & Hasegawa, T 2009, 'The human-elephant conflict in
southeastern Sri Lanka: type of damage, seasonal patterns, and sexual differences in the raiding
behaviour of elephants', Gajah, vol. 31, pp. 5 – 14.
Chakraborty, K & Mondal, J 2013, 'Perceptions and patterns of human-elephant conflict at Barjora
block of Bankura district in West Bengal, India: insights for mitigation and management',
Environmental Development and Sustainability, vol. 15, pp. 547 – 565.
Chang’a, A, de Souza, N, Muya, J, Keyyu, J, Mwakatobe, A, Malugu, L, Ndossi, HP, Konuche, J, Omondi,
R, Mpinge, A, Hahn, N, Palminteri, S and Olson, D 2016, 'Scaling-up the use of chilli fences for reducing
human-elephant conflict across landscapes in Tanzania', Tropical Conservation Science, vol. 9, no. 2,
pp. 921 – 930.

159
Chiyo, PI, Cochrane, EP, Naughton, L & Basuta, GI 2005, 'Temporal patterns of crop raiding by
elephants: a response to changes in forage quality or crop availability?', African Journal of Ecology,
vol. 43, pp. 48 – 55.
Choudhury, A 2004, 'Human-elephant conflicts in northeast India', Human Dimensions of Wildlife, vol.
9, no. 4, pp. 261 – 270.
Cook, RM, Witkowski, ETF, Helm, CV, Henley, MD & Parrini, F 2017, 'Recent exposure to African
elephants after a century of exclusion: rapid accumulation of marula tree impact and mortality, and
poor regeneration', Forest Ecology and Management, vol. 401, pp. 107 – 116.
Das, BJ, Saikia, BN, Baruah, KK, Bora, A & Bora, M 2014, 'Nutritional evaluation of fodder, its preference
and crop-raiding by wild Asian elephant (Elephas maximus) in Sonitpur District of Assam, India',
Veterinary World, vol. 7(12), pp. 1082-1089.
Davies, TE, Wilson, S, Hazarika, N, Chakrabaty, J, Das, D, Hodgson, DJ & Zimmerman, A
2011,'Effectiveness of intervention methods against crop-raiding elephants', Conservation Letters,
doi: 10.1111/j.1755-263X.2011.00182.x
de Silva, M 1998, 'Status and conservation of the elephant (Elephas maximus) and the alleviation of
man-elephant conflict in Sri Lanka', Gajah, vol. 19, pp. 1 – 68.
Dickman, AJ 2010, 'Complexities of conflict: the importance of considering social factors for effectively
resolving human-wildlife conflict', Animal Conservation, vol. 13, pp. 458 - 466.
Dublin, HT & Hoare, RE 2004, 'Searching for solutions: the evolution of an integrated approach to
understanding and mitigating human-elephant conflict in Africa', Human Dimensions of Wildlife, vol.
9, no. 4, pp. 271 – 278.
Dutta, A 2013, ‘Bees, acrid smoke from chillis keep elephants away and reduce conflict’, The Times of
India, June 24, 2013.

160
Fernando, P, Leimgruber, P, Prasad, T & Pastorini, J 2012, ‘Problem-elephant translocation:
translocating the problem and the elephant?’, PLoS ONE, vol. 7, no. 12, e50917,
https://doi.org/10.1371/journal.pone.0050917.
Fernando, P, Jayewardene, J, Prasad, T, Hendavitharana, W & Pastorini, J 2011, ‘Current status of Asian
Elephants in Sri Lanka’, Gajah, vol. 25, pp. 93 – 103.
Fernando, P, Kumar, MA, Williams, AC, Wikramanayake, E, Aziz, T & Singh, SM 2008, Review of human-
elephant conflict mitigation measures practiced in South Asia, AREAS Technical Support Document,
World Wide Fund for Nature.
Fernando, P, Wikramanayake, E, Weerakoon, D, Jayasinghe, LKA, Gunawardene, M & Janaka, HK 2005,
'Perceptions and patterns of human-elephant conflict in old and new settlements in Sri Lanka: insights
for conservation and management', Biodiversity and Conservation, vol. 14, pp. 2465 – 2481.
Graham, MD, Notter, B, Adams, WM, Lee, PC & Ochieng, TN 2010, 'Patterns of crop-raiding by
elephants, Loxodonta africana, in Laikipia, Kenya, and the management of human-elephant conflict',
Systematics and Biodiversity, vol. 8, no. 4, pp. 435 – 445.
Graham, MD & Ochieng, T 2008, 'Uptake and performance of farm-based measures for reducing crop-
raiding by elephants Loxodonta Africana among smallholder farms in Laikipia District, Kenya', Oryx,
vol. 42, pp. 76 – 82.
Gross, EM, Drouet-Hoguet, N, Subedi, N & Gross, J 2017, 'The potential of medicinal and aromatic
plants (MAPs) to reduce crop damages by Asian elephants (Elephas maximus)', Crop Protection, vol.
100, pp. 29 – 37.
Gross, EM, Mc Robb, R and Gross, J 2016, 'Cultivating alternative crops reduces crop losses due to
African elephants' Journal of Pest Science, vol. 89, no. 2, pp. 495 – 506.
Gubbi, S 2012, 'Patterns and correlates of human-elephant conflict around a south Indian reserve',
Biological Conservation, vol. 148, pp. 88 – 95.

161
Gunaryadi, D, Sugiyo, Hedges, S 2017, 'Community-based human-elephant conflict mitigation: the
value of an evidence-based approach in promoting the uptake of effective methods', PLoS ONE, vol.
12, no. 5, e0173742.
Hartter, J, Goldman, A & Southworth, J 2011, 'Responses by households to resource scarcity and
human-wildlife conflict: issues of fortress conservation and the surrounding agricultural landscape',
Journal for Nature Conservation, vol. 19, pp. 79 – 86.
Haturusinghe, HS & Weerakoon, DK 2012, ‘Crop raiding behaviour of elephants in the Northwestern
region of Sri Lanka’, Gajah, vol. 36, pp. 26 – 31.
Hedges, S & Gunaryadi, D 2009, 'Reducing human-elephant conflict: do chillies help deter elephants
from entering crop fields?', Oryx, vol. 44, no. 1, pp. 139 – 146.
Hoare, R 2015, ‘Lessons from 20 years of human-elephant conflict mitigation in Africa’, Human
Dimensions of Wildlife, vol. 20, no. 4, pp. 289 – 295.
Hoare, R 2000, 'African elephants and humans in conflict: the outlook for co-existence', Oryx, vol. 34,
pp. 34 – 38.
Hoffmeier-Karimi, RR & Schulte, BA 2014, 'Assessing perceived and documented crop damage in a
Tanzanian village impacted by human-elephant conflict (HEC)', Pachyderm, vol. 56, pp. 51 – 60.
Jasmine, B, Ghose, D and Das, SK 2015, 'An attitude assessment of human-elephant conflict in a critical
wildlife corridor within the Terai Arc landscape, India', Journal of Threatened Taxa, vol. 7, no. 2, pp.
6843 – 6852.
Karidozo, M & Osborn, FV 2015, 'Community based conflict mitigation trials: results of field tests of
chilli as an elephant deterrent', Biodiversity and Endangered Species, vol. 3 no. 1,
http://dx.doi.org/10.4172/2332-2543.1000144.
King, L, Pardo, M, Weerathunga, S, Kumara, TV, Jayasena, N, Soltis, J & de Silva, S 2018, ‘Wild Sri Lankan
elephants retreat from the sound of disturbed Asian honey bees’, Current Biology, vol. 28, pp 64 – 65.

162
King, LE, Lala, F, Nzumu, H, Mwambingu, E & Douglas-Hamilton, I 2017, 'Beehive fences as a
multidimensional conflict-mitigation tool for farmers coexisting with elephants', Conservation Practice
and Policy, vol. 31, no. 4, pp. 743 – 752.
King, LE 2014, Beehive fence construction manual: a step by step guide to building a protective beehive
fence to deter crop-raiding elephants from farm land, Elephants and Bees Project, Save the Elephants,
Nairobi.
King, LE 2010, The interaction between the African elephant (Loxodonta africana africana) and the
African honey bee (Apis mellifera scutellate) and its potential application as an elephant deterrent,
PhD Thesis, University of Oxford, UK.
King, LE, Soltis, J, Douglas-Hamilton, I, Savage, A & Vollrath, F 2010, 'Bee threat elicits alarm call in
African elephants', PLoS ONE, vol. 5, no. 4, e10346. doi:10.1371/journal.pone0010346.
King, LE, Lawrence, A, Douglas-Hamilton, I & Vollrath, F 2009, ‘Beehive fence deters crop-raiding
elephants’, African Journal of Ecology, vol. 47, pp. 131 – 137.
King, LE, Douglas-Hamilton, I & Vollrath, F 2007, 'African elephants run from the sounds of disturbed
bees', Current Biology, vol. 17, no. 19, R832.
Kioko, J, Muruthi, P, Omondi, P & Chiyo, PI 2008, 'The performance of electric fences as elephant
barriers in Amboseli, Kenya', South African Journal of Wildlife Research, vol. 38(1), pp. 52-58.
Koeniger, N & Koeniger, G 1994, Honeybees of Asia, IN: Ed (RWK Punchihewa), Beekeeping for honey
production in Sri Lanka, Sri Lanka Department of Agriculture, Peradeniya, Sri Lanka.
Mabelunga, T, Kumar, DAV, Gayathri, A & Krishnan, A 2016, 'Influence of elephant-human interactions
on agrarian communities in the Bengaluru-Bannerghatta Landscape – a perspective study', Gajah, vol.
45, pp. 28 – 32.
Madden, F 2004, 'Creating coexistence between humans and wildlife: global perspectives on local
efforts to address human-wildlife conflict', Human Dimensions of Wildlife, vol. 9, no. 4, pp. 247 – 257.

163
Manoj, EM 2012, ‘Jumbos get a sting of defeat’, The Hindu Newspaper, February 25, 2012.
Nath, NK, Lahkar, BP, Dutta, SK and Das, JP 2015, 'Human-elephant conflict around Manas National
Park, India: local people's attitudes, expectations and perceptions', Gajah, vol. 42, pp. 15 – 21.
Nair, RP and Jayson, EA 2016, 'Effectiveness of beehive fences to deter crop raiding elephants in
Kerala, India', International Research Journal of Natural and Applied Sciences, vol. 3, no. 1, pp. 14 – 19.
Nelson, A, Bidwell, P and Sillero-Zubiri 2003, 'A review of human-elephant conflict mitigation
strategies', People and Wildlife Initiative, Wildlife Conservation Research Unit, Oxford University.
Nsonsi, F, Heymans, J, Diamouangana, J, Mavinga, FB and Breuer, T 2018, 'Perceived human-elephant
conflict and its impact for elephant conservation in northern Congo', African Journal of Ecology, vol.
56, pp. 208 – 215.
Ogra, MV 2008, 'Human-wildlife conflict and gender in protected area borderlands: a case study of
costs, perceptions and vulnerabilities from Uttarakhand (Uttaranchal), India', Geoforum, vol. 39, pp.
1408 – 1422.
Osborn, FV & Parker, GE 2003, 'Towards an integrated approach for reducing the conflict between
elephants and people: a review of current research', Oryx, vol. 37, no. 1, doi.
10.1017/90030605302000000.
Perera, BMAO 2009, 'The human-elephant conflict: a review of current status and mitigation
methods', Gajah, vol. 1, pp. 341 – 344.
Pozo, RA, Coulson, T, McCulloch, G, Strongza, A & Songhurst, A 2017, 'Chilli-briquettes modify the
temporal behaviours of elephants, but not their numbers', Oryx, doi: 10.1017/S0030605317001235.
Ponnusamy, V, Chackrapani, P, Lim, TW, Saaban, S and Campos-Arceiz, A 2016, 'Farmers' perceptions
and attitudes towards government constructed electric fences in Peninsular Malaysia', Gajah, vol. 45,
pp. 4 – 11.

164
Punchihewa, RWK 1994, Beekeeping for honey production in Sri Lanka, Sri Lanka Department of
Agriculture, Peradeniya, Sri Lanka.
Ranasinghe, D, Ratnapala, S, Seneviratne, L, Schwammer, H, Schwammer, G & Perera, BH 2010a, Saga
of the elephant in Sri Lanka: extracts from Loris Volume 1 (1936) to Volume 10 (1964), YN Print
Graphics, Nedimala, Sri Lanka.
Ranasinghe, D, Ratnapala, S, Seneviratne, L, Schwammer, H, Schwammer, G & Perera, BH 2010b, Saga
of the elephant in Sri Lanka: extracts from Loris Volume 11 (1965) to Volume 20 (1993), YN Print
Graphics, Nedimala, Sri Lanka.
Santiapillai, C, Wijeyamohan, S, Bandara, G, Athurupana, R, Dissanayake, N & Read, B 2010, 'An
assessment of the human-elephant conflict in Sri Lanka', Ceylon Journal of Biological Sciences, vol. 39,
no. 1, pp. 21 – 33.
Santiapillai, C, Fernando, P and Gunewardene, M 2006, 'A strategy for the conservation of the Asian
elephant in Sri Lanka', Gajah, vol. 25, pp. 91 – 102.
Santiapillai, C 1996, 'Mitigation of human-elephant conflicts in Sri Lanka', Gajah, vol. 15, pp. 1 – 7.
Scheijen, CPJ, Richards, SA, Smit, J, Jones, T & Nowak, K 2018, 'Efficacy of beehive fences as barriers
to African elephants: a case study in Tanzania', Oryx, doi: 10.1017/S0030605317001727.
Shri, KP, Gunasekaran, C, Salahudeen, M, Asaikkutti, A & Deepa, AA 2013, ‘Assessment of response of
elephants to beehives placed in a paddy field’, Gajah, vol. 38, pp. 42 – 43.
Sitati, NW & Walpole, MJ 2006, 'Assessing farm-based measures for mitigation of human-elephant
conflict in Transmara District, Kenya', Oryx, vol. 40, no. 3, pp. 279 – 286.
Sitati, NW, Walpole, MJ, Smith, RJ & Leader-Williams, N 2003, 'Predicting spatial aspects of human-
elephant conflict', Journal of Applied Ecology, vol. 40, pp. 667 – 677.
Sukumar, R 2006, 'A brief review of the status, distribution and biology of wild Asian elephants Elephas
maximus', International Zoo Yearbook, vol. 40, pp. 1 – 8.

165
Sukumar, R 1990, 'Ecology of the Asian elephant in southern India. II. Feeding habits and crop raiding
patterns', Journal of Tropical Ecology, vol. 6, pp. 33 – 53.
Thapa, S 2010, 'Effectiveness of crop protection methods against wildlife damage: a case study of two
villages at Bardia National Park, Nepal', Crop Protection, vol. 29, pp. 1297 – 1304.
Thirgood, S, Woodroffe, R and Rabinowitz, A 2005, 'The impact of human-wildlife conflict on human
lives and livelihoods', IN: (eds: Woodroffe R, Thirgood S and Rabinowitz A) People and wildlife: conflict
or coexistence? Cambridge, Cambridge University Press, pp. 13 – 26.
van de Water, A and Matteson, K 2018, 'Human-elephant conflict in western Thailand: socio-economic
drivers and potential mitigation strategies', PLoS ONE, vol. 13, no. 6, e0194736.
van Eden, M, Ellis, E and Bruyere, BL 2016, 'The influence of human-elephant conflict on electric fence
management and perception among different rural communities in Laikipia County, Kenya', Human
Dimensions of Wildlife, vol. 21, no. 4, pp. 283 – 296.
Vollrath, F & Douglas-Hamilton, I 2002, 'African bees to control African elephants',
Naturwissenschaften, vol. 89, pp. 508 – 511.
von Gerhardt, K, Niekerk, AV, Kidd, M, Samways, M & Hanks, J 2014, 'The role of elephant Loxodonta
africana pathways as a spatial variable in crop-raiding location', Oryx, vol. 48(3), pp. 436 – 444.
Webber, CE, Sereivathana, T, Maltby, MP and Lee, PC 2011, 'Elephant crop-raiding and human-
elephant conflict in Cambodia: crop selection and seasonal timing of raids', Oryx, vol. 45, no. 2, pp.
243 – 251.
Wikramanayake, ED, Hathurusinghe, HS, Janaka, HK, Jayasinghe, LKA, Fernando, P, Weerakoon, DK &
Gunawardene, MD 2004, 'The human-elephant conflict in Sri Lanka: lessons for mitigation,
management, and conservation from traditional land-use patterns', in: (ed) Jayewardene, J
Endangered Elephants: Past, present and future, Proceedings of the symposium for human-elephant
relationships and conflicts, Sri Lanka, September 2003. Biodiversity and Conservation Trust, Colombo,
pp. 164-169.

166
Chapter 5: Discussion

167
5.1 Introduction
The research presented here constitutes a multi-faceted investigation, where we attempt to better
understand the behaviour of a local elephant population in north-central Sri Lanka, the patterns of
crop-raiding, and to work closely with a local community heavily impacted by HEC to evaluate a newly
introduced elephant deterrent tool. To be more specific, we investigated the general behaviour
patterns of elephants utilising habitat representing two different levels of anthropogenic disturbance
or ‘risk’ to elephants (Wasgamuwa National Park: low risk; Weheragala Tank: medium risk), in the
presence and absence of an immediate disturbance stimuli. We also monitored crop-raiding activity
over a three-year period to produce a comprehensive profile of the crop-raiding patterns and
predictors occurring in Dewagiriya Village, a small community heavily impacted by HEC. Finally, in
collaboration with farmers from the Dewagiriya community, we established, monitored and evaluated
the practical feasibility of an elephant deterrent new to the area: the use of beehive fences to protect
home gardens.
In this chapter, I begin with a brief summary of research findings. I then discuss methodological and
project challenges. Finally, I suggest future research directions that I believe would help further our
understanding of complex HEC situations and complement the development of site-specific mitigation
planning to benefit both elephants and humans.
5.2 Summary of results
By scoring the duration of general behavioural activities (feeding, locomotion, standing still) and
frequency of specific ‘reactive’ behaviours (e.g. head-shaking, dusting, backing away) from video
footage obtained during direct field observations, we were able to compare the behaviour of male
and female elephants inhabiting areas within Wasgamuwa National Park (a low-risk zone) and at
Weheragala Tank (a medium-risk zone). We were also able to identify whether responses to
immediate disturbances in the environment (e.g. approach of vehicles, pedestrians or dogs) were
consistent across the two risk zones.
We found no evidence to suggest that general behaviour patterns of elephants differed according to
the perceived level of anthropogenic risk (medium or low) represented by their location. Not

168
surprisingly, elephants spent the majority of their time feeding, whether they were observed in
Wasgamuwa National Park or at Weheragala Tank. However, the presence of an immediate
anthropogenic risk or disturbance, did cause elephants to alter their behaviour, with this effect
stronger at Weheragala Tank. At both locations, the frequency of ‘reactive behaviours’ (typically
smelling or assuming the ‘vigilance’ posture) increased. However, at Weheragala Tank elephants of
both sexes spent less time feeding during or following a disturbance event. This is in line with a study
in India where elephants also decreased feeding time as levels of human disturbance increased
(Srinivassaiah et al. 2012). In the presence of a disturbance, female elephants also increased the time
they spent in locomotion, often displaying avoidance behaviour as they interrupted feeding to retreat
back to the relative safety of forest cover.
Farmer-collected data was used to document sightings of elephants within 50 m of their property,
including date, time, number of elephants, sex of elephants, and damage incurred. Crop-raiding
patterns and characteristics were measured from these data. Our results show that Dewagiriya Village
experiences exceptionally high levels of crop-raiding, especially when compared to other locations
within both Africa and Asia, based on a review of available literature (examples: Das et al. 2012; Davies
et al. 2011; Ekanayaka et al. 2011; Campos-Arceiz et al. 2009; Barnes et al. 2007; Chen et al. 2005).
Several characteristics of crop-raiding in Dewagiriya were typical of most crop-raiding situations: In
particular, our study showed that almost all crop-raiding occurred at night and evidence strongly
suggested that male elephants were the predominant crop-raiders, acting either individually or in
small bachelor groups (for comparison see examples: Thaufeek et al. 2014; Das et al. 2014; Graham et
al. 2010; Campos-Arceiz et al. 2009; Bandara and Tisdell 2003). Elephants raided a wide variety of
crops, with rice, bananas and coconuts the most frequently targeted, presumably at least in part
because they were the most widely available (for comparison see examples: Haturusinghe and
Weerakoon 2012; Ekanayaka et al. 2011; Campos-Arceiz et al. 2009; Wikramanayake et al. 2004).
Dewagiriya differs from most other locations (examples: Gubbi 2012; Webber et al. 2011; Campose-
Arceiz et al. 2009; Fernando et al. 2005), in that no clear seasonal patterns were identified. Elephants
raided year-round and no overt influence of season or rainfall was apparent, aside from a reduction
in crop-raiding events when it had rained during that 24-hour period. Clear within-site variations in
crop-raiding were evident, with homes and fields located furthest from the road and closest to the
water tanks and forested habitat, bearing the highest burden of elephant events and crop-raids.

169
During our three-year beehive fence trial we worked closely with the local community to decide on
experimental participants, design fence layouts, build and monitor the fences, and record data on
elephant events occurring within 50 m of home garden boundaries for experimental (beehive fence)
and control (no beehive fence) properties. Results provided evidence that beehive fences were
effective at reducing elephant incursions into home gardens relative to control home gardens.
However, there was still a 43% rate of elephants breaching fence perimeters on approach. Still, the
deterrence effect strengthened with increased hive occupations, indicating that the presence of bees
was influential in preventing elephants from entering the fences. Had we been able to reach our 50%
hive occupation target on more fences, less breakthroughs may have occurred. That said, beekeeping
was challenging and attracting wild colonies into our fence hives was largely unsuccessful. Colony
health was often weak and as a result honey production was low and no economic benefits were seen.
Farmer motivation fluctuated and only four experimental participants planned to continue using their
fences after the trial ended.
5.3 Methodological challenges
5.3.1 Behavioural assessments
Studying the behaviour of wild animals in a natural setting can be fraught with challenges, the most
obvious being an inability to account for all confounding variables. Numerous factors, ranging from
subtle variations in environmental conditions (Griffin 2007) to the personalities of individuals within
the group (Hollander et al. 2008; Freeman et al. 2007) can influence behaviour. In an uncontrolled
environment, such as our study sites of Wasgamuwa National Park and Weheragala Tank, we were
unable to select specific individuals for observation, resulting in unintentional bias towards inclusion
of individuals who were ‘bold’ enough to enter the observation space in the presence of our research
vehicle. It is entirely possible that more elephants from the same group or family as those observed
were present during observations but did not leave the relative safety of forest cover. Another
potential issue is that we had little or no knowledge of events that happened previously that day (or
earlier) that may have influenced behaviour outside of our parameters of interest. Another constraint
was our reliance on opportunistic disturbance, and we had no control over what particular disturbance
occurred or which elephants were present at the time. This limited our sample size of study subjects
and the contexts in which they could be observed. Still captive studies cannot be expected to
accurately replicate the ‘wild’ environment, and natural studies are important. Comparing responses

170
of captive elephants during exposure to anthropogenically induced disturbances would have its own
ethical considerations, and relevance to a ‘real life’ setting would be limited. We dealt with these
constraints by randomly selecting study subjects and disturbance events and acknowledge that
conclusions rely on a small degree of speculation.
In this study we made the decision to include only a ‘low’ and ‘medium’ risk zone (Wasgamuwa
National Park and Weheragala Tank respectively). Including a ‘high’ risk zone would have added an
extra layer of complexity to our study, allowing us to compare elephant behaviour at two ‘extremes’
of anthropogenic disturbance, whilst still investigating potential impacts of ‘medium’ level
disturbance. Dewagiriya Village, where the crop-raiding profile and beehive fence trial were
conducted, seemed an obvious candidate for a ‘high’ risk zone. Elephants were known to inhabit the
forest on the periphery of Dewagiriya Village during the day and move towards the village to drink
from the water tanks and crop-raid in the evenings. However, the thick forest habitat was accessible
only on foot and no clearings suitable for video recording were available. Observing elephants at night
would have reduced comparability across risk zones as all other observations were conducted
between 3 pm and 7 pm. If differences in behaviour had been observed, it would have been difficult
to tease apart the influence of time on behaviour versus the influence of heightened anthropogenic
risk. Furthermore, community members did not want our team conducting observations in Dewagiriya
Village at night, ostensibly (and reasonably) for our own safety but I speculate that certain farmers
within the community did not want outsiders observing their personal interactions with elephants as
illegal methods are sometimes used to chase them out of the village.
I would encourage researchers at other HEC locations to investigate the behaviour of elephants in
‘medium’ risk zones. To take Sri Lanka as an example, approximately 65% of suitable elephant habitat
is located outside of protected boundaries (Santiapillai et al. 2006), and most elephants do venture
outside of PA’s to utilise other suitable habitat (Weerakoon et al. 2004). Elephants tend to visit villages
(or high risk zones) only to crop raid which means a lot of time is spent in areas akin to our ‘medium’
risk zone. In Dewagiriya Village, humans and elephants both utilise water and forest resources, and
use the paths and roads to travel from village to village. Knowledge of the general behaviour patterns
of elephants, and their responses to human presence or disturbance (whether on foot or in vehicles)
can help facilitate safe interactions between the two species, outside of the intense conflict situations
occurring nightly when elephants come to feed on crops.

171
5.3.2 Crop-raiding activity
To identify crop-raiding activity in Dewagiriya Village, we relied heavily on information from farmers.
We tried to make this as systematic as possible, but most families did not fill out their data sheets,
and we found it more accurate to visit bi-monthly and transcribe information on-the-spot. Calendars
were distributed, so people could at least mark the date of elephant events to ensure none were
forgotten but only some families used this. Still, an elephant event was a memorable occasion, despite
the frequency with which they occurred, and generally people were keen to provide reports. To the
best of our knowledge, elephant events were reported with accuracy, and reports from neighbouring
households typically corroborated. Even allowing for some degree of exaggeration, the frequency of
elephant events in Dewagiriya Village is undeniably high, and a major social concern for the
community.
Admittedly, our study had a large focus on frequency of elephant events and quantifying precise
measurements of crop damage was outside our scope. Further research quantifying crop damage
relative to overall size of crop plots and the stage of growth when damaged/impacts on crop yields
would facilitate economic impact assessments of crop-raids. We simply did not have enough time or
manpower to go into this detail. It was also obvious that even small crop losses were significant to
individual households.
5.3.3 Beehive fence trials
Taking a concept designed for another species of bees and another species of elephants (King et al.
2011) was always going to be challenging but given the success shown in Africa and that beekeeping
can produce additional benefits (e.g. honey production), we felt it was an idea well worth attempting.
However, although we found indicators of success at preventing elephants from entering home
gardens, the deterrent rate was not as strong as hoped. This combined with the challenges of
beekeeping, fence costs, and low farmer motivation perhaps suggests that the observed positives do
not outweigh the challenges, at least for Dewagiriya Village.
To rigorously test the efficacy of beehive fences as an elephant deterrent, we would ideally leave
occupied fences as the only line of defence. However, in the real-life context of Dewagiriya Village it
would not have been fair to ask farmers to eliminate use of all traditional methods. Elephants can

172
easily destroy numerous crops or plants in a single visit, plus allowing elephants to approach when
family members are inside their homes is a huge risk to human safety (Fernando et al. 2005). Instead
we asked farmers to list other deterrents used and found the same methods used at both
experimental and control households. This typically involved shouting, throwing firecrackers or
thunder-flashes, and shining bright flashlights in the direction of elephants. However, we were unable
to measure farmer vigilance and it’s possible that households who guarded more closely or had more
manpower were better able to defend their property regardless of whether they had a beehive fence,
or how many occupied hives were present.
The only way to potentially combat this methodological challenge, would be to plant a series of crop
plots outside of the village using project funds, and surround these with beehive fences. This would
negate the risk to farmer livelihoods and personal safety. However, finding an unplanted and
unforested area large enough to support multiple beehive fences (and getting permission to use it)
would be difficult. Furthermore, because elephants are known to use a combination of olfactory,
audio and visual cues to distinguish threats (Bates et al. 2007; Thuppil and Coss 2013), such a
fabricated environment would fail to fully represent the in-village crop-raiding scenario where human
influences such as lights, sounds and smell are always present. Perhaps the best way to answer the
question of whether beehive fencing is a useful Asian elephant deterrent is in the exact context where
the complexities and variables involved in mitigation conflict exist.
It should also be noted that in the lifespan of either a human or elephant, 3 years is a relatively short
time. Although we consider this a sufficient time span to provide indicators of the likelihood of beehive
fence success, a much longer-term study is recommended to gain a true sense of its worthiness in an
overall human-elephant conflict toolkit. Over time elephants could habituate to the presence of bees
and find more ways to breach the deterrent, as they have been known to do with other mitigation
techniques (examples: Fernando et al. 2011; Osborn and Parker 2003). Alternatively, elephants could
learn more about the potential negative consequences of disturbing beehives and be less likely to
break-through. Farmer willingness to participate could also change over time. If beekeeping skills
improved, and honey harvests were higher, this could be a major incentive to maintain fences in good
condition.

173
Although we did our best to ensure the community were empowered with the skills necessary to look
after their fences and bee colonies, after three years only two households could manage their fences
and beekeeping independently. Two households did not have the time and were not confident with
beekeeping, and therefore withdrew from the trial. The other households fell somewhere in the
middle, needing some active support and reminding of maintenance tasks but also capable of doing
most things alone. Another limitation to farmers continuing with the deterrent is that no households
were in a financial position to afford to replace damaged posts or beehives, or to introduce new
colonies if their colonies absconded. Overall, only four households planned to continue using their
beehive fences post-trial. We had hoped three years would be a sufficient period to properly establish
the beehive fence project and transfer management to the community, but this did not eventuate.
Efforts were made to connect with local organisations who could continue to provide small-scale
practical and financial support in fence maintenance and beekeeping, following our three-year trial
period. However, despite expressed enthusiasm and training provided, to the best of our knowledge
this has not been implemented.
5.4 Closing statements
This study has contributed to the general body of knowledge on elephant behaviour in an
anthropogenically influenced context, and specifically on patterns of Asian elephant crop-raiding in a
high HEC zone, and the potential of beehive fencing as an Asian elephant deterrent. We show that at
our particular study site, elephants do make behavioural choices based on the presence of
anthropogenic disturbance in their immediate environment; that crop-raiding in Dewagiriya Village is
characterised by similar sex-biases and nocturnal timing as most locations but lacks clear seasonal
patterns; and that beehive fencing shows some positive indicators of deterrent success however is
limited by beekeeping challenges, cost-effectiveness, and farmer motivation.
There is no doubt that the Dewagiriya community is struggling to cope with HEC and given personal
observations from other nearby villages and the available HEC literature spanning elephant range
countries, it is fair to say that too many communities are struggling. The conversations about crop-
raiding and mitigation often seem to go in circles, although it is commonly agreed that genuine
collaboration between stakeholders (government authorities, NGO’s, wildlife managers,
conservationists, and local communities) is necessary to enact meaningful change. However,

174
translating this theoretical realisation into on-ground action is difficult. Still, we need to continue to
do the best we can to help individuals and communities, and to generate information that might be
of use to others. For elephant conservation to succeed, cooperation rather than pushing individual
agendas, and free sharing of research findings (positive or negative results) is one of the most powerful
weapons we have. Finally, we must remember that the well-being and survival of people who share
their land with elephants is equally as important as the charismatic species we are trying so
desperately to protect.

175
5.5 References
Bandara, R & Tisdell, C 2003, 'Comparison of rural and urban attitudes to the conservation of Asian
elephants in Sri Lanka: empirical evidence', Biological Conservation, vol. 110, pp. 327 – 342.
Bates, LA, Sayialel, KN, Njiraini, NW, Moss, CJ, Poole, JH and Byrne, RW 2007, 'Elephants classify human
ethnic groups by odor and garment colour', Current Biology, vol. 17, no. 22,
doi:10.1016/j.cub.2007.09.060.
Campos-Arceiz, A, Takatsuki, S, Ekanayaka, SKK & Hasegawa, T 2009, 'The human-elephant conflict in
southeastern Sri Lanka: type of damage, seasonal patterns, and sexual differences in the raiding
behaviour of elephants', Gajah, vol. 31, pp. 5 – 14.
Chen, Y, Marino, J, Chen, Y, Tao, Q, Sullivan, CD, Shi, K & MacDonald, DW 2016, 'Predicting hotspots
of human-elephant conflict to inform mitigation strategies in Xishuangbanna, Southwest China', PLoS
ONE, vol. 11(9): e0162035.doi:10.1371/journal.pone.0162035.
Das, BJ, Saikia, BN, Baruah, KK, Bora, A & Bora, M 2014, 'Nutritional evaluation of fodder, its preference
and crop-raiding by wild Asian elephant (Elephas maximus) in Sonitpur District of Assam, India',
Veterinary World, vol. 7(12), pp. 1082-1089.
Davies, TE, Wilson, S, Hazarika, N, Chakrabaty, J, Das, D, Hodgson, DJ & Zimmerman, A
2011,'Effectiveness of intervention methods against crop-raiding elephants', Conservation Letters,
doi: 10.1111/j.1755-263X.2011.00182.x
Ekanayaka, SKK, Campos-Arceiz, A, Rupasinghe, M, Pastorini, J & Fernando, P 2011, 'Patterns of crop
raiding by Asian elephants in a human-dominated landscape in southeastern Sri Lanka', Gajah, vol. 34,
pp. 20 -25.
Fernando, P, Jayewardene, J, Prasad, T, Hendavitharana, W & Pastorini, J 2011, ‘Current status of Asian
Elephants in Sri Lanka’, Gajah, vol. 25, pp. 93 – 103.

176
Fernando, P, Wikramanayake, E, Weerakoon, D, Jayasinghe, LKA, Gunawardene, M & Janaka, HK 2005,
'Perceptions and patterns of human-elephant conflict in old and new settlements in Sri Lanka: insights
for conservation and management', Biodiversity and Conservation, vol. 14, pp. 2465 – 2481.
Freeman, HD, Brosnan, SF, Hopper, LM, Lambeth, SP, Schapiro, SJ & Gosling, SD 2013, 'Developing a
comprehensive and comparative questionnaire for measuring personality in chimpanzees using a
simultaneous top-down/bottom-up design', American Journal of Primatology, vol. 75, pp. 1042 –
1053.
Graham, MD, Notter, B, Adams, WM, Lee, PC & Ochieng, TN 2010, 'Patterns of crop-raiding by
elephants, Loxodonta africana, in Laikipia, Kenya, and the management of human-elephant conflict',
Systematics and Biodiversity, vol. 8, no. 4, pp. 435 – 445.
Griffin, SC, Valois, T, Taper, ML & Mills, LS 2007, 'Effects of tourists on behaviour and demography of
Olympic marmots', Conservation Biology, vol. 21, no. 4, pp. 1070-1081.
Gubbi, S 2012, 'Patterns and correlates of human-elephant conflict around a south Indian reserve',
Biological Conservation, vol. 148, pp. 88 – 95.
Haturusinghe, HS & Weerakoon, DK 2012, ‘Crop raiding behaviour of elephants in the Northwestern
region of Sri Lanka’, Gajah, vol. 36, pp. 26 – 31.
Hollander, FA, van Overveld, T, Tokka, I & Matthysen, E 2008, ‘Personality and nest defence in the
great tit (Parsus major)’, Ethology, vol. 114, pp. 405 – 412.
King, LE, Lala, F, Nzumu, H, Mwambingu, E & Douglas-Hamilton, I 2017, 'Beehive fences as a
multidimensional conflict-mitigation tool for farmers coexisting with elephants', Conservation Practice
and Policy, vol. 31, no. 4, pp. 743 – 752.
King, LE, Douglas-Hamilton, I & Vollrath, F 2011, 'Beehive fences as effective deterrents for crop-
raiding elephants: field trials in northern Kenya', African Journal of Ecology, doi: 10.1111/j.1365-
2028.2011.01275.x.

177
Osborn, FV & Parker, GE 2003, 'Towards an integrated approach for reducing the conflict between
elephants and people: a review of current research', Oryx, vol. 37, no. 1, doi.
10.1017/90030605302000000.
Santiapillai, C, Fernando, P and Gunewardene, M 2006, 'A strategy for the conservation of the Asian
elephant in Sri Lanka', Gajah, vol. 25, pp. 91 – 102.
Srinivasaiah, NM, Anand, VD, Vaidyanathan, S & Sinha, A 2012, 'Usual populations, unusual
individuals: insights into the behaviour and management of Asian elephants in fragmented
landscapes', PLoS ONE, vol. 7, no. 8, e42571. doi: 10.1371/journal.pone.0042571.
Thaufeek, UL, Padmalal, UKGK and Fernando, P 2014, 'Land use and human-elephant conflict in the
Sigiriya Sanctuary, Sri Lanka', Gajah, vol. 40, pp. 26 – 30.
Thuppil, V and Coss RG 2013, 'Wild Asian elephants distinguish aggressive tiger and leopard growls
according to perceived danger', Biology Letters, Vol. 9, 20130518.
http://dx.doi.org/10.1098/rsbl.2013.0518.
Webber, CE, Sereivathana, T, Maltby, MP and Lee, PC 2011, 'Elephant crop-raiding and human-
elephant conflict in Cambodia: crop selection and seasonal timing of raids', Oryx, vol. 45, no. 2, pp.
243 – 251.
Weerakoon, DK, Gunawardene, MD, Janaka, HK, Jayasinghe, LKA, Perera, RAR, Fernando, P and
Wikramanayake, E 2004, 'Ranging behaviour and habitat use of elephants in Sri Lanka', in: (ed)
Jayewardene, J Endangered Elephants: Past, present and future, Proceedings of the symposium for
human-elephant relationships and conflicts, Sri Lanka, September 2003. Biodiversity and Conservation
Trust, Colombo, pp. 68 - 70.
Wikramanayake, ED, Hathurusinghe, HS, Janaka, HK, Jayasinghe, LKA, Fernando, P, Weerakoon, DK &
Gunawardene, MD 2004, 'The human-elephant conflict in Sri Lanka: lessons for mitigation,
management, and conservation from traditional land-use patterns', in: (ed) Jayewardene, J
Endangered Elephants: Past, present and future, Proceedings of the symposium for human-elephant
relationships and conflicts, Sri Lanka, September 2003. Biodiversity and Conservation Trust, Colombo,
pp. 164-169.

178
Copyright © 2022 FDOKUMEN




















