
The species of Cortinarius, section Bovini, associated with conifers in northern Europe
Upload
uni-goettingenCategory
view
0download
0
ORIGINAL PAPER
Broadleaf seedling responses to warmer temperatures ‘‘chilled’’by late frost that favors conifers
Nicholas Fisichelli • Torsten Vor • Christian Ammer
Received: 29 November 2012 / Revised: 17 January 2014 / Accepted: 21 January 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract Climate change includes not only shifts in
mean conditions but also changes in the frequency and
timing of extreme weather events. Tree seedlings, as the
potential future overstory, are responding to the selective
pressures of both mean and extreme conditions. We
investigated how increases in mean temperature and the
occurrence of late spring frosts affect emergence, devel-
opment, growth, and survival of 13 native and non-native
broadleaf and conifer tree species common in central
Europe. Three temperature levels (ambient, ?3, and
?6 �C) and three spring frost treatments (control, late, and
very late) were applied. Development responses of first-
year seedlings to warmer temperatures were similar in
direction and magnitude for broadleaf and conifer species.
Stem size also increased with rising mean temperature for
most species, though broadleaf species had maximal height
advantage over conifer species in the warmest treatment.
Sensitivity to frost differed sharply between the broadleaf
and conifer groups. Broadleaf survival and stem length
exhibited strong reductions due to frost events while
conifer species only showed minor decreases in survival.
Importantly, more rapid development and earlier leaf-out
in response to warmer temperatures were associated with
increased mortality from frost for broadleaf species but
decreased mortality for conifer species. This research
suggests that compositional shifts in the direction of spe-
cies favored by increasing mean temperatures may be
slowed by extreme events, and thus, the occurrence and
impacts of such weather events must be acknowledged and
incorporated into research and forest planning.
Keywords Broadleaf seedling � Climate change � Conifer
seedling � Extreme weather events � Phenology � Spring
frost � Survival
Introduction
Forest management in the face of multiple global change
agents must apply flexible approaches to foster healthy and
productive forests (Millar et al. 2007; Bolte et al. 2009;
Sohn et al. 2013). Diversifying monospecific stands,
notably conifer stands expected to be threatened by
droughts and more virulent pests, is one approach to dif-
fusing risk (Ammer et al. 2008; Knoke et al. 2008; Kolling
et al. 2009; Overbeck and Schmidt 2012). However,
determining which species may be most successful under
future climatic conditions is not clear. There are not only
uncertainties in the sensitivity of various species to
changing climate (for European beech, see, e.g., Leuschner
et al. 2001; Rennenberg et al. 2004; Ammer et al. 2005),
but also uncertainties in the magnitude of mean tempera-
ture increases and shifts in the seasonal timing and fre-
quency of extreme events (Jentsch et al. 2007). For
example, increasing mean temperatures and a longer
growing season are likely to favor species best able to
Communicated by G. Brazaitis.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10342-014-0786-6) contains supplementarymaterial, which is available to authorized users.
N. Fisichelli (&) � T. Vor � C. Ammer
Department of Silviculture and Forest Ecology of the Temperate
Zones, Georg-August-Universitat Gottingen, Busgenweg 1,
37077 Gottingen, Germany
e-mail: [email protected]
N. Fisichelli
Natural Resource Stewardship and Science, U.S. National Park
Service, Fort Collins, CO, USA
123
Eur J Forest Res
DOI 10.1007/s10342-014-0786-6

maximize productivity under the changed conditions,
whereas increased climate variability such as late spring
frost events may select for species with more conservative
growth strategies (Hufkens et al. 2012). Research is needed
to understand how co-occurring broadleaf and conifer tree
species respond to changes in climate means and extremes
at the earliest stages of development and thus which spe-
cies may thrive under future conditions.
Spring average temperatures are increasing in many
regions; however, this does not necessarily mean that late
spring frost dates are retreating at the same rate or that the
risk of frost damage is decreasing (Meehl et al. 2000; Gu
et al. 2008; Inouye 2008). Changes in the frequency and
timing of late spring frosts will depend as much on changes
in variance as in climate means (Rigby and Porporato
2008). For example, a 1 �C change in the standard devia-
tion of the temperature distribution will influence the fre-
quency of late spring frosts more than a 1 �C change in
mean temperature (Meehl et al. 2000). Recent late spring
frost events in eastern North America have impacted for-
ests across large regions (Gu et al. 2008; Augspurger 2009;
Hufkens et al. 2012). Interspecific differences in growth
strategies and susceptibility to damage resulted in shifts in
competitive dynamics and an advantage for species less
impacted by spring frost (Hufkens et al. 2012). In European
forests, the potential ecological importance of late spring
frosts is exemplified by Fagus sylvatica, which due to late
spring frost events is precluded from areas with otherwise
sufficiently high growing season temperatures (Peters
1992).
Development responses to warming in spring may alter
risk of frost damage. Plant species face a trade-off between
maximizing growing season length and protecting against
frost damage (Saxe et al. 2001). Investments in cold tol-
erance, for example, by conifer species, reduce growth
rates, and frost avoidance in the form of late-onset spring
growth may confer a competitive advantage to neighboring
stems that emerged at an earlier time (Hannerz et al. 1999;
Loehle 1998). Conversely, early-onset growth of frost-
sensitive tissue, such as by some broadleaf species, could
lead to greater frost damage (Cannell and Smith 1986;
Inouye 2000). However, more rapid development beyond
the most frost-sensitive stages, especially in a warmer
world, could reduce risk of frost damage for some species
(Coursolle et al. 1998; Kreyling et al. 2012).
In this study, we examined how potential future climatic
conditions, including warmer temperatures and late spring
frost events, impact first-year seedling emergence, devel-
opment, growth, and survival of tree species commonly
grown in central Europe. We hypothesized that warming
temperatures would lead to earlier emergence, more rapid
development, and increased growth of seedlings and that
responses would be greater for broadleaf than conifer
species. Conversely, we hypothesized that late spring frost
events would favor conifer species over broadleaf species,
due to higher survival rates. Furthermore, we expected that
the combination of increased temperatures and very late
frost would lead to the greatest impacts on plant perfor-
mance. These data will inform managers on how future
variations in climate may impact first-year seedling per-
formance and potentially the direction and rate of forest
change.
Methods
A growth chamber experiment was conducted in the spring
and summer of 2012 at Georg-August-Universitat Gottin-
gen, Germany, and included a fully crossed two-way fac-
torial design with temperature (three levels: ambient, ?3,
and ?6 �C) and frost (three levels: control, late, and very
late) treatments. Seeds from 22 tree species commonly
found growing in central Europe were collected in the fall
of 2011 in central and northern Germany (Lower Saxony)
and processed and cold stratified (generally at 3 �C for
60–90 days) by a professional nursery. Seeds were planted
in 5-l plastic pots (17 9 17 9 17 cm) filled with 2,000 g
of potting material (‘‘Einheitserde SP VM’’ peat, clay, and
sand mix, pH = 5.8, nitrogen concentration 0.2 %). Of the
original 22 tree species, only 13 had sufficient emergence
rates ([*7 %) to permit analyses (Table 1). This included
most of the common native and non-native broadleaf and
conifer species present in central European forests. The
number of replicate pots per warming treatment was gen-
erally five (range 4–9), though the number of seeds and
pots per species varied with seed availability and seed size
(e.g., 882 seeds in 18 pots for Picea abies and 432 seeds in
27 pots for Fagus sylvatica; see Table 1).
The experiment was conducted within climate-con-
trolled growth chambers (BBC, Brown Boveri-York,
Hamburg, Germany). Temperature levels (day/night: 15/8,
18/11, and 21/14 �C) were based on late spring/early
summer values (Gottingen, Germany 1970–2000) and
potential temperatures by the end of the twenty-first cen-
tury (EEA 2012). All growth chambers began with a day/
night temperature of 10/6 �C, and temperatures were
increased incrementally over 14 days to reach experiment
levels. The first day that maximum treatment temperatures
were reached was considered day 1 of the experiment. Pots
were watered two to three times per week to field capacity,
with frequency increasing with rising temperatures.
Humidity in all chambers was kept at 60 % during the day
and 75 % at night. Light availability within the chambers
was similar to moderately open forest understory condi-
tions (PAR * 150 lmols/m2/s), and day length began with
12 h and increased to 16 h. As only three growth chambers
Eur J Forest Res
123

were available for the study, pot location within chamber
was randomized weekly and pots were rotated among
chambers every 2 weeks to account for any within- or
among-chamber-treatment effects. Two replications of the
late spring frost treatment occurred on days 27 and 28 and
two replicates of the very late frost on days 46 and 47. Pots
receiving the frost treatment were moved to a frost cham-
ber in the evening and cooled at a rate of 3 �C per hour,
held at -5 �C for 2 h, and then warmed again at a rate of
3 �C per hour. Pots were packed in loose potting soil to
insulate the roots from freezing, and temperatures 2 cm
below the soil surface remained above 0 �C during all frost
treatments.
Pots were surveyed twice per week to determine the
percentage of seedlings emerging (emergence is defined as
the epicotyl breaking the soil surface) and development
stage (emerging, 1st leaf fully expanded). For conifers, the
first leaf fully expanded stage was reached when the first
true needles were [1 cm in length. Survival of all stems
was monitored for 19 days after each frost treatment. Total
stem length was measured on day 62.
Seedling responses to experimental treatments and
interactions were analyzed with linear mixed-effects
models and pot as a random effect (i.e., random intercept to
account for autocorrelation of stems within the same pot) in
R (v. 2.12; R Development Core Team 2008). Emergence
day, leaf-out day, and stem length responses were assessed
with linear mixed-effects models and a Gaussian error
distribution using the ‘‘nlme’’ package (Pinheiro et al.
2009). As the first frost was not applied until after the
fastest-developing species in the ?6 �C treatment level had
leafed out, emergence and leaf-out analyses only included
the temperature treatment. Comparisons of factor levels
(Tukey’s test) were carried out using the ‘‘multcomp’’
package (Hothorn et al. 2008). The percentage of seedlings
emerging and survival (of aboveground tissue) post-frost
were assessed with generalized linear mixed-effects mod-
els and a binomial error distribution using the ‘‘lme4’’
package (Bates et al. 2011). Likelihood ratio tests were
utilized to formally compare emergence percentage and
survival models and test whether temperature and frost
improved model performance over simpler models with
each variable individually removed (Crawley 2007). Sur-
vival post-frost was also compared among development
stages. All figures shown include the same eight species,
four broadleaves and four conifers, which had the largest
sample sizes and are representative of responses for each
functional group.
Results
Emergence
The number of species responding to warming treatments
increased with development stage (Table 2). The percent-
age of seedlings emerging varied with temperature treat-
ment for five out of 13 species (see Electronic
Supplementary Material, Online Resource 1 for all pair-
wise comparisons). Acer pseudoplatanus was the only
species to exhibit a decrease in the percentage of seedlings
emerging with increasing temperature, from 58 ± 2 %
(mean ± se) at ?6 �C to 73 ± 2 % at ambient temperature
(z = -2.86, p = 0.01). The only other broadleaf species
with an emergence percentage shift was Juglans regia,
where the ?6 �C treatment emergence was 69 ± 8 % and
ambient temperature emergence was 40 ± 3 % (z = 2.71,
p = 0.02). Three conifer species exhibited increased
emergence with warming treatments. For Abies alba,
emergence in the ?3 and ?6 �C treatments (both
9 ± 3 %) was significantly higher than under ambient
temperature (3 ± 0.5 %; z = 2.82, p = 0.01; z = 2.70,
p = 0.02, respectively). Similar effects were found for
Picea abies, with the ?3 and ?6 �C treatments (70 ± 3
and 75 ± 4 %, respectively) having higher emergence
rates than with ambient conditions (53 ± 3 %; z = 3.46,
p = 0.01; z = 4.43, p = 0.001, respectively). Finally, for
Table 1 Broadleaf and conifer tree species commonly found in
central European forests and included in this study
Species Seeds/
pot
Pots Total
seeds
Emergence
(%)
Native
range
Broadleaf species
Acer
pseudoplatanus
25 15 375 65.1 Europe
Fagus sylvatica 16 27 432 68.1 Europe
Juglans regia 9 15 135 57.0 Europe
Quercus
petraea
6 12 72 40.3 Europe
Quercus robur 9 15 135 37.8 Europe
Robinia
pseudoacacia
36 15 540 18.7 Eastern
North
America
Conifer species
Abies alba 49 15 735 7.1 Europe
Abies grandis 25 15 375 14.7 Western
North
America
Larix decidua 49 15 735 40.1 Europe
Larix
kaempferi
49 15 735 65.0 Japan
Picea abies 49 18 882 65.6 Europe
Pinus sylvestris 49 15 735 87.9 Europe
Pseudotsuga
menziesii
49 15 735 11.7 Western
North
America
Eur J Forest Res
123

Pseudotsuga menziesii, more seedlings emerged in the
?6 �C treatment (19 ± 3 %) than under ambient temper-
ature (5 ± 1 %, z = 4.45, p = 0.002).
The amount of time required for emergence (emergence
day) decreased with increasing temperature for nine out of
13 species, and no species showed a later emergence date at
a higher temperature (Table 2, Online Resource 2). Fur-
thermore, trends were similar among early- and late-
emerging species, with three out of four earliest and three
out of four latest species exhibiting significant responses to
temperature. For the earliest species, Robinia pseudoacacia,
seedlings in the ?6 �C treatment (day 1 ± 0.5; mean ± se)
emerged significantly earlier than in the ?3 �C treatment
(day 6 ± 0.3; z = -3.46, p = 0.002) and ambient treatment
(day 9 ± 2; z = -5.08, p \ 0.0001). Pinus sylvestris stems
in the ambient treatment emerged on average on day
10.9 ± 0.8, ?3 �C on day 8.1 ± 0.8, and ?6 �C on day
5.7 ± 0.5, with all three treatments significantly different
from one another (p \ 0.02). For Fagus sylvatica, seedlings
emerged at similar times in the ?3 and ?6 �C treatments
(days 9 ± 0.5 and 8 ± 0.8, respectively) and both were
significantly earlier than under ambient conditions (day
13 ± 0.7, p \ 0.0001 in both comparisons).
Late-emerging species included the three largest seeded,
hypogeous species, Juglans regia, Quercus petraea, and
Quercus robur, and the conifer Pseudotsuga menziesii. For J.
regia, emergence was significantly later in the ambient
treatment (day 65 ± 9) than in ?3 �C (day 40 ± 2; z =
-3.98, p = 0.0002) and ?6 �C treatments (day 30 ± 3;
z = -5.58, p \ 0.0001). Stems in the ?6 �C treatment
emerged over 19 days earlier than under ambient tempera-
ture for both Q. robur and P. menziesii (p value \0.02 for
both species). Quercus petraea emerged on average on day
56 ± 5, the earliest Q. robur stems (in the ?6 �C treatment)
on day 46 ± 3, and the earliest P. menziesii stems (?6 �C
treatment) on day 61 ± 1. Thus, these three species
emerged, on average, after the very late frost treatment.
Leaf-out
The date of first leaf fully expanded (leaf-out day) shifted
to an earlier time with heating for all species (Table 2;
Fig. 1, the eight species shown in figures had the largest
samples and were representative of variation in responses
for each functional group). Warming of ?6 �C resulted in
significantly reduced leaf-out time compared with ambient
for all 13 species (all p values\0.01) (Online Resource 3);
for example, reductions in development time for broadleaf
species included 14.1 ± 2 days for A. pseudoplatanus
(z = -6.74, p \ 0.0001), 19 ± 2 days for F. sylvatica
(z = -8.45, p \ 0.0001), 28.5 ± 4 days for J. regia
(z = -7.21, p \ 0.0001), and 32 ± 2 days for R. pseudo-
acacia (z = -13.70, p \ 0.0001) (Fig. 1a–d). Similar
responses were found for conifer species, with the ?6 �C
warming causing leaf-out to be earlier than in the ambient
treatment by 19 ± 6 days for Abies grandis (z = -3.44,
p = 0.002), 17 ± 2 days for Larix decidua (z = -6.98,
p \ 0.0001), 22 ± 2 days for P. abies (z = -7.21,
Table 2 ANOVA results of seedling emergence percentage, emergence day, and leaf-out day responses to the fixed effect of temperature (3
levels: ambient, ?3, and ?6 �C)
Species Response: Emergence (%) Emergence day Leaf-out day
den df Trend v2 p Trend F p Trend F p
Acer pseudoplatanus 12 2 8.40 0.01 0 2.76 0.14 / 26.30 <0.0001
Fagus sylvatica 24 0 0.24 0.89 / 17.01 <0.0001 / 39.13 <0.0001
Juglans regia 12 1 8.15 0.02 / 15.95 0.0004 / 26.52 0.0001
Quercus petraea 9 0 4.39 0.11 0 2.26 0.16 / 7.10 0.02
Quercus robur 12 0 1.08 0.58 / 6.89 0.01 / 15.86 0.0006
Robinia pseudoacacia 12 0 0.10 0.95 / 13.44 0.0009 / 95.05 <0.0001
Abies alba 12 1 8.94 0.01 0 2.30 0.14 / 5.59 0.04
Abies grandis 10 0 1.96 0.38 3.93 0.05 / 7.31 0.01
Larix decidua 12 0 5.40 0.07 0 2.28 0.14 / 26.42 <0.0001
Larix kaempferi 12 0 3.52 0.17 / 31.13 <0.0001 / 154.20 <0.0001
Picea abies 15 1 19.80 <0.0001 / 19.83 0.0001 / 57.23 <0.0001
Pinus sylvestris 12 0 3.19 0.20 / 17.54 0.0003 / 242.41 <0.0001
Pseudotsuga menziesii 12 1 15.47 0.000 / 5.74 0.02 / 6.82 0.01
Symbols -/? denote decreases/increases in emergence percentage with increasing temperature; / denotes earlier emergence/leaf-out with
increasing temperature. ‘‘den df’’ is denominator degrees of freedom. Emergence day and leaf-out day responses are from linear mixed-effects
models
Values in bold are significant at p \ 0.05
Eur J Forest Res
123

p \ 0.0001), and 23 ± 1 days for P. sylvestris (z =
-21.74, p \ 0.0001) (Fig. 1e–h). Temperature differences
of 3 �C caused significant shifts (p \ 0.05) in leaf-out day
for eight out of 13 species when comparing ambient and
?3 �C treatments and also eight out of 13 species when
comparing ?3 and ?6 �C treatments (Online Resource 3).
Growth and survival
Emergence and development responses to temperature treat-
ments shown above were generally similar across broadleaf
and conifer groups, whereas survival and growth responses to
temperature and frost treatments differed more strongly
between these functional groups. Survival of aboveground
tissue varied significantly with temperature for one species
and with frost treatment for eight species (Table 3; Fig. 2).
Similar to the reduction in emergence, A. pseudoplatanus
exhibited a small, though significant reduction in survival with
increasing temperature. Mean survival in the ?3 �C
(86 ± 7 %) and ?6 �C treatments (86 ± 7 %) was lower
than the ambient treatment (97 ± 3 %).
In response to frost, all four broadleaf species had
strongly reduced survival, with the very late event gen-
erally resulting in the greatest mortality (Fig. 2a–d).
Survival for 19 days after the very late frost varied from a
high of 61 ± 9 % for A. pseudoplatanus to a low of
8.5 ± 5 % for J. regia. Conversely, survival was gener-
ally much higher for conifers than for broadleaf species
Fig. 1 Leaf-out day responses to mean temperature for four broadleaf (a–d) and four conifer (e–h) tree species. See Table 2 for ANOVA test
output. Means and standard errors based on pot-level data
Table 3 ANOVA results of
seedling survival responses to
the fixed effects of temperature
and frost
Analyses were run using
generalized linear mixed-effects
models, a binomial error
distribution, and pot as a
random effect. Temperature by
frost interaction was not
significant (see text).
Differences in post-frost
survival by development stage
(emerging vs. leafed-out) were
also tested. Empty cells indicate
insufficient sample sizes, such
as for Q. petraea, Q. robur, and
P. menziesii, which primarily
emerged after frost treatments
Values in bold are significant at
p \ 0.05
Species Survival (prop) Temperature Frost Post-frost survival by
development stage
v2 p v2 p v2 p
Acer pseudoplatanus 0.86 12.45 0.002 68.13 <0.0001 29.14 <0.0001
Fagus sylvatica 0.63 2.73 0.26 176.0 <0.0001 0.84 0.36
Juglans regia 0.62 0.13 0.94 71.99 <0.0001 8.22 0.004
Quercus petraea 1.00
Quercus robur 0.95
Robinia pseudoacacia 0.72 3.82 0.15 55.05 <0.0001 0.04 0.85
Abies alba 0.97
Abies grandis 0.83 4.89 0.09 15.59 0.0004 1.81 0.18
Larix decidua 0.96 0.01 0.99 7.68 0.02 7.10 0.008
Larix kaempferi 0.95 0.01 0.99 9.68 0.008 10.83 0.001
Picea abies 0.95 2.26 0.32 135.3 <0.0001 1.70 0.19
Pinus sylvestris 0.98 1.40 0.50 3.87 0.14 19.02 <0.0001
Pseudotsuga menziesii 1.00
Eur J Forest Res
123
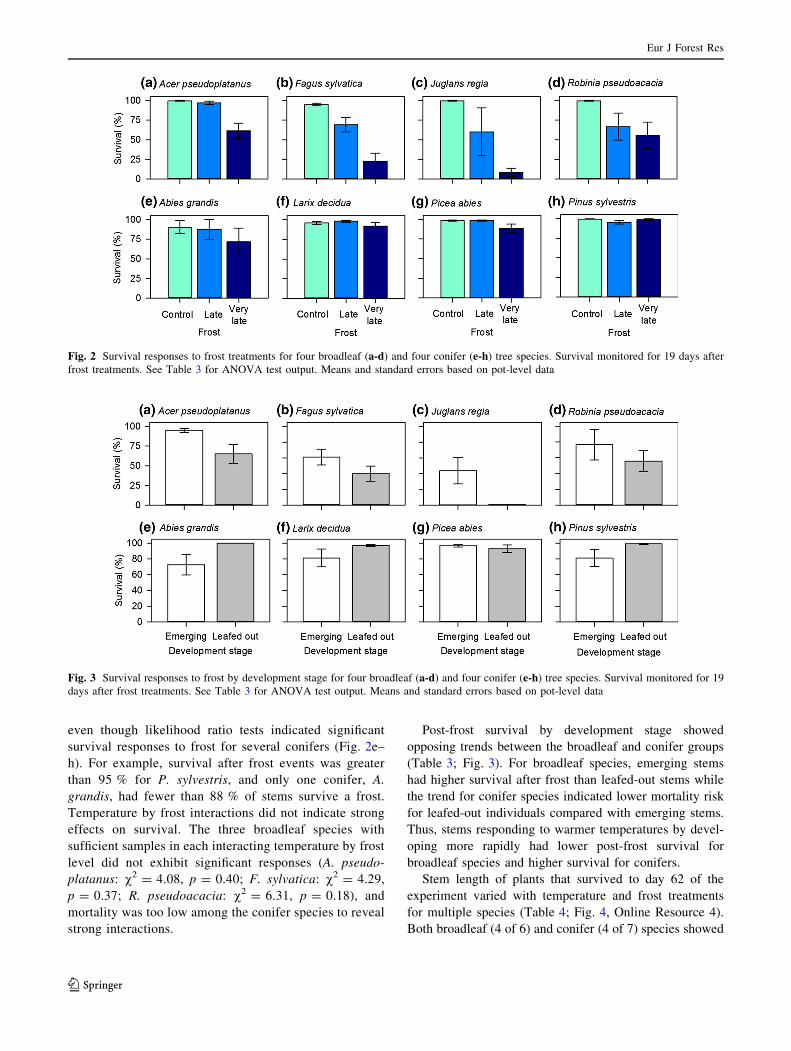
even though likelihood ratio tests indicated significant
survival responses to frost for several conifers (Fig. 2e–
h). For example, survival after frost events was greater
than 95 % for P. sylvestris, and only one conifer, A.
grandis, had fewer than 88 % of stems survive a frost.
Temperature by frost interactions did not indicate strong
effects on survival. The three broadleaf species with
sufficient samples in each interacting temperature by frost
level did not exhibit significant responses (A. pseudo-
platanus: v2 = 4.08, p = 0.40; F. sylvatica: v2 = 4.29,
p = 0.37; R. pseudoacacia: v2 = 6.31, p = 0.18), and
mortality was too low among the conifer species to reveal
strong interactions.
Post-frost survival by development stage showed
opposing trends between the broadleaf and conifer groups
(Table 3; Fig. 3). For broadleaf species, emerging stems
had higher survival after frost than leafed-out stems while
the trend for conifer species indicated lower mortality risk
for leafed-out individuals compared with emerging stems.
Thus, stems responding to warmer temperatures by devel-
oping more rapidly had lower post-frost survival for
broadleaf species and higher survival for conifers.
Stem length of plants that survived to day 62 of the
experiment varied with temperature and frost treatments
for multiple species (Table 4; Fig. 4, Online Resource 4).
Both broadleaf (4 of 6) and conifer (4 of 7) species showed
Fig. 2 Survival responses to frost treatments for four broadleaf (a-d) and four conifer (e-h) tree species. Survival monitored for 19 days after
frost treatments. See Table 3 for ANOVA test output. Means and standard errors based on pot-level data
Fig. 3 Survival responses to frost by development stage for four broadleaf (a-d) and four conifer (e-h) tree species. Survival monitored for 19
days after frost treatments. See Table 3 for ANOVA test output. Means and standard errors based on pot-level data
Eur J Forest Res
123

significantly greater size (p \ 0.05) with increased tem-
perature. For A. pseudoplatanus, F. sylvatica, and J. regia,
a ?6 �C warming significantly increased stem length by
40–60 % over the ambient treatment (Fig. 4a–c). Broadleaf
R. pseudoacacia had the largest increase in stem length
with a ?6 �C warming, almost 500 % (Fig. 4d, note the
y-axis scale). When significant, stem length differences due
to a ?6 �C warming for conifer species were similar to the
first three broadleaf species above, with increases of
25–128 % (Fig. 4e–h).
Unlike warming, frost treatments had much larger
impacts on stem lengths of broadleaf than conifer species
(Table 4; Fig. 4, Online Resource 4). Frost caused stem
length decreases of up to 80 % for broadleaf species, while
no conifer species exhibited a difference in stem length by
frost treatment. Thus, size advantage for broadleaf species
over conifers was greatest under the combination of the
warmest temperatures and lack of frost occurrence. High
mortality and subsequent small sample sizes at the pot level
reduced the ability to detect temperature by frost interac-
tions, although stem-level data suggest a trend toward very
late frosts resulting in greater decreases in size at warmer
temperatures (e.g., F. sylvatica and R. pseudoacacia,
Fig. 4b, d).
Discussion
Our results show that increased mean temperatures and late
frost events impact emergence, development, growth, and
Table 4 ANOVA results of
seedling stem length responses
to the fixed effects of
temperature, frost, and the
interaction
Analyses were run using linear
mixed-effects models, a
Gaussian error distribution, and
pot as a random effect. Empty
cells indicate sample sizes that
were too small to carry out
statistical tests. ‘‘den df’’ is
denominator degrees of freedom
Values in bold are significant at
p \ 0.05
Species n den df Temperature Frost Temp 9 frost
F p F p F p
Acer pseudoplatanus 207 6 11.42 0.009 5.94 0.04 1.37 0.35
Fagus sylvatica 189 15 6.01 0.01 11.74 0.0009 1.55 0.24
Juglans regia 55 8 14.17 0.002 23.10 0.0005
Quercus petraea 18 2 2.29 0.30 0.63 0.61
Quercus robur 31 3 3.35 0.17 1.05 0.45
Robinia pseudoacacia 66 4 29.03 0.004 6.04 0.06 3.67 0.12
Abies alba 42 4 1.62 0.31 1.20 0.39 4.47 0.09
Abies grandis 46 5 0.55 0.61 0.47 0.65 3.12 0.12
Larix decidua 176 6 26.56 0.001 3.09 0.12 0.95 0.49
Larix kaempferi 209 6 48.53 0.0002 0.11 0.89 0.88 0.53
Picea abies 228 9 60.07 <0.0001 2.08 0.18 2.83 0.09
Pinus sylvestris 193 6 36.94 0.0004 0.02 0.98 1.80 0.25
Pseudotsuga menziesii 25 5 4.17 0.09 0.80 0.50
Fig. 4 Stem length responses to temperature and frost treatments for four broadleaf (a–d) and four conifer (e–h) tree species. Note the different
y-axis scaling for (d) R. pseudoacacia. See Table 4 for ANOVA test output. Means and standard errors based on stem-level data
Eur J Forest Res
123

survival of tree seedlings at early stages of development.
Emergence and leaf-out responses to warmer temperatures
were generally similar in direction and magnitude for first-
year broadleaf and conifer seedlings. Both groups also
increased size with temperature, though height advantage
of broadleaf species over conifers increased with temper-
ature. Sensitivity to spring frost differed sharply between
these two functional groups, with broadleaf survival and
stem length exhibiting strong reductions due to frost events
and conifer species showing only minor decreases in sur-
vival. Furthermore, more rapid development in response to
warmer temperatures was associated with increased mor-
tality from frost for broadleaf species but decreased mor-
tality for conifer species. This research indicates that the
magnitude of warming and frequency and timing of future
frost events may in part determine the relative performance
of broadleaf and conifer seedlings in central European
forests.
Seedling establishment, growth, and ascension into lar-
ger size classes are bottleneck phases of population growth
(Walck et al. 2011), and small differences in relative per-
formance may be sufficient to tip competitive dynamics in
favor of species or provenances better adapted to current
climatic conditions. Frost-tolerant conifers were unable to
keep pace with the growth rates of broadleaf species,
especially under the warmest mean temperatures (Loehle
1998). In addition to obvious physiological differences
between functional groups, there was also wide variation in
emergence responses within the broadleaf group that could
shift competitive dynamics (Hufkens et al. 2012). The three
broadleaf species with the largest seeds, J. regia, Q. pet-
raea, and Q. robur, on average emerged late, and thus,
many individuals avoided frost damage. In contrast, the
small-seeded R. pseudoacacia emerged very early and
grew very rapidly in the warmest conditions; however, this
aggressive growth strategy was heavily penalized by cli-
mate variability, with large decreases in stem size and
survival after frost. Though not examined here, genetic
diversity within species, such as from multiple prove-
nances, may also result in intraspecific differences in
responses to climate change (Kreyling et al. 2012).
Our experiment looked at responses to means and
extremes in a single year, and multiple extreme events (or
the lack of such events) over multiple years would likely
have compounding effects and result in greater fitness
differences between species. Extreme weather events such
as late frosts can also impact older tree regeneration as well
as overstory trees and overall ecosystem productivity. For
example, broadleaf saplings typically flush their leaves
earlier than overstory trees and produce a large portion of
their photosynthetic carbon gain during this period of high
light availability, and stems forced to refoliate after a frost
will miss this high-light window and may show greater
mortality than was found for first-year seedlings in this
study (Augspurger 2009). Additionally, this research
assessed aboveground vegetative tissue, and frost is typi-
cally more damaging to reproductive tissue (Sakai and
Weiser 1973) and can cause widespread reproductive
failure for a species in a given year (Inouye 2000), illus-
trating that frosts can also impact the relative numbers of
seeds and seedlings in subsequent years after reproduction.
Other manifestations of increased climatic variability
will also impact tree species. Similar to frosts during the
growing season, warm temperature spells during the dor-
mant season can stress broadleaf and conifer trees and lead
to widespread regional forest declines (Bourque et al. 2005;
Lazarus et al. 2006). Increased temperature variability has
been linked to summer heat waves in Europe (Schar et al.
2004), and the combination of late spring frosts and sum-
mer droughts could result in major declines in productivity
and higher mortality for many tree species (Gu et al. 2008).
A single late frost event, by itself, can reduce gross annual
ecosystem productivity by 10 % or more for the subsequent
growing season (Hufkens et al. 2012).
Climate change will also interact with many other fac-
tors to determine the relative success of tree species (Fis-
ichelli et al. 2013). For example, earthworms may interact
with warming temperatures to exacerbate drought condi-
tions (Eisenhauer et al. 2012), while browse pressure by
deer is likely to inhibit growth responses to warming
temperatures (Fisichelli et al. 2012). Increased atmospheric
CO2 concentrations, such as the levels predicted by the end
of the century, lead to heightened frost sensitivity, so
similar frost events under future atmospheric conditions
may result in greater damage and ecosystem productivity
losses (Lutze et al. 2002; Gu et al. 2008). Frost can also
interact with other factors such as browse pressure to
modify growth and mortality (Buckley et al. 1998).
Key attributes close to nature forestry such as mixing
tree species, reliance on natural regeneration, and keeping
shelter or retention trees for decades (Grassi et al. 2003,
Pommerening and Murphy 2004) may help to decrease the
risk of stand loss due to extreme events. First, as shown in
this study, tree species respond differently to such events,
supporting the idea of mixtures being more resistant to
disturbances than monocultures (Pretzsch 2003; Griess
et al. 2012). Second, natural selective pressures at these
early stages seem to foster adaptation to changed climate
conditions (Bilela et al. 2012). Third, silviculture practices
such as retaining overstory trees for their insulating effects
can reduce occurrence and impact of extreme events and
encourage the growth of tree species better adapted to a
warmer world (Buckley et al. 1998).
Understanding which tree species will be most viable
under likely future conditions requires detailed ecophysi-
ological knowledge of species-level responses (Kimball
Eur J Forest Res
123

et al. 2010; Walck et al. 2011) as well as fine temporal and
spatial scale predictions of climatic conditions. Given the
multiple global change factors that must be taken into
account and imprecise forecasts of future conditions, forest
managers must hedge their bets and make decisions that
will be robust under the broadest swath of potential future
scenarios (Lawler 2009; Kolling et al. 2009). Composi-
tional shifts toward species favored by increasing mean
temperatures are likely to be slowed by extreme events,
and thus, the occurrence and impacts of extreme events
must be acknowledged and incorporated into research and
forest planning.
Acknowledgments Financial support was provided by the U.S.
Fulbright Program. Thanks to M. Unger and H. Coners for laboratory
assistance.
References
Ammer C, Albrecht L, Borchert H et al (2005) Zur Zukunft der Buche
(Fagus sylvatica L.) in Mitteleuropa-Kritische Anmerkungen zu
einem Beitrag von Rennenberg et al.(2004). All Forst-u J Ztg
176:4
Ammer C, Bickel E, Kolling C (2008) Converting Norway spruce
stands with beech-a review of arguments and techniques. Aus J
For Sci 125:3–26
Augspurger CK (2009) Spring 2007 warmth and frost: phenology,
damage and refoliation in a temperate deciduous forest. Funct
Ecol 23:1031–1039
Bates D, Maechler M, Bolker B (2011) lme4: linear mixed-effects
models using S4 classes. R package version 0.999375-1.
R Foundation for Statistical Computing, Vienna, Austria
Bilela S, Dounavi A, Fussi B et al (2012) Natural regeneration of
Fagus sylvatica L. adapts with maturation to warmer and drier
microclimatic conditions. For Ecol Manag 275:60–67
Bolte A, Ammer C, Lof M et al (2009) Adaptive forest management
in central Europe: climate change impacts, strategies and
integrative concept. Scand J For Res 24:473–482
Bourque CPA, Cox RM, Allen DJ et al (2005) Spatial extent of winter
thaw events in eastern North America: historical weather records in
relation to yellow birch decline. Glob Change Biol 11:1477–1492
Buckley DS, Sharik TL, Isebrands J (1998) Regeneration of northern
red oak: positive and negative effects of competitor removal.
Ecology 79:65–78
Cannell M, Smith R (1986) Climatic warming, spring budburst and
forest damage on trees. J Appl Ecol 177–191
Coursolle C, Bigras F, Margolis H (1998) Frost tolerance and
hardening capacity during the germination and early develop-
mental stages of four white spruce (Picea glauca) provenances.
Can J Bot 76:122–129
Crawley MJ (2007) The R book. Wiley, USA
Eisenhauer N, Fisichelli NA, Frelich LE et al (2012) Interactive
effects of global warming and ‘global worming’ on the initial
establishment of native and exotic herbaceous plant species.
Oikos 121:1121–1133
European Environmental Agency (EEA) Global and European
temperature (CSI 012)—Assessment June 2012
Fisichelli NA, Frelich LE, Reich PB (2012) Sapling growth responses
to warmer temperatures ‘cooled’ by browse pressure. Glob
Change Biol 18:3455–3463
Fisichelli NA, Frelich LE, Reich PB (2013) Climate and interrelated
tree regeneration drivers in mixed temperate-boreal forests.
Landsc Ecol 28:149–159
Grassi G, Minotta G, Giannini R et al (2003) The structural dynamics
of managed uneven-aged conifer stands in the Italian eastern
Alps. For Ecol Manag 185:225–237
Griess VC, Acevedo R, Hartl F et al (2012) Does mixing tree species
enhance stand resistance against natural hazards? A case study
for spruce. For Ecol Manag 267:284–296
Gu L, Hanson PJ, Mac Post W et al (2008) The 2007 eastern US
spring freeze: increased cold damage in a warming world?
Bioscience 58:253–262
Hannerz M, Aitken SN, King JN et al (1999) Effects of genetic
selection for growth on frost hardiness in western hemlock. Can
J For Res 29:509–516
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in
general parametric models. Biometrical J 50:346–363
Hufkens K, Friedl MA, Keenan TF et al (2012) Ecological impacts of
a widespread frost event following early spring leaf-out. Glob
Change Biol 18:2365–2377
Inouye D (2000) The ecological and evolutionary significance of frost
in the context of climate change. Ecol Lett 3:457–463
Inouye DW (2008) Effects of climate change on phenology, frost
damage, and floral abundance of montane wildflowers. Ecology
89:353–362
Jentsch A, Kreyling J, Beierkuhnlein C (2007) A new generation of
climate-change experiments: events, not trends. Front Ecol
Environ 5:365–374
Kimball S, Angert AL, Huxman TE et al (2010) Contemporary
climate change in the Sonoran Desert favors cold-adapted
species. Glob Change Biol 16:1555–1565
Knoke T, Ammer C, Stimm B et al (2008) Admixing broadleaved to
coniferous tree species: a review on yield, ecological stability
and economics. Eur J For Res 127:89–101
Kolling C, Knoke T, Schall P et al (2009) Uberlegungen zum Risiko
des Fichtenanbaus in Deutschland vor dem Hintergrund des
Klimawandels. Forstarchiv 80:42–54
Kreyling J, Thiel D, Nagy L et al (2012) Late frost sensitivity of
juvenile Fagus sylvatica L. differs between southern Germany
and Bulgaria and depends on preceding air temperature. Eur J
For Res 131:717–725
Lawler JJ (2009) Climate change adaptation strategies for resource
management and conservation planning. Ann NY Acad Sci
1162:79–98
Lazarus BE, Schaberg PG, Hawley GJ et al (2006) Landscape-scale
spatial patterns of winter injury to red spruce foliage in a year of
heavy region-wide injury. Can J For Res 36:142–152
Leuschner C, Backes K, Hertel D et al (2001) Drought responses at
leaf, stem and fine root levels of competitive Fagus sylvatica L.
and Quercus petraea (Matt.) Liebl. trees in dry and wet years.
For Ecol Manag 149:33–46
Loehle C (1998) Height growth rate tradeoffs determine northern and
southern range limits for trees. J Biogeogr 25:735–742
Lutze J, Roden J, Holly C et al (2002) Elevated atmospheric [CO2]
promotes frost damage in evergreen tree seedlings. Plant, Cell
Environ 21:631–635
Meehl GA, Karl T, Easterling DR et al (2000) An introduction to
trends in extreme weather and climate events: observations,
socioeconomic impacts, terrestrial ecological impacts, and
model projections. Bull Am Meteorol Soc 81:413–416
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and
forests of the future: managing in the face of uncertainty. Ecol
Appl 17:2145–2151
Overbeck M, Schmidt M (2012) Modelling infestation risk of Norway
spruce by Ips typographus (L.) in the Lower Saxon Harz
Mountains (Germany). For Ecol Manag 266:115–125
Eur J Forest Res
123

Peters R. (1992) Ecology of beech forests in the northern hemisphere.
Dissertation. Landbouwuniversiteit Wageningen, The Netherlands
Pinheiro J, Bates D, DebRoy S et al (2009) nlme: linear and nonlinear
mixed effects models, R package version 3.1-96
Pommerening A, Murphy S (2004) A review of the history,
definitions and methods of continuous cover forestry with
special attention to afforestation and restocking. Forestry
77:27–44
Pretzsch H (2003) Diversitat und Produktivitat von Waldern. Allg For
Jgdztg 174:88–98
R Development Core Team (2008) R: a language and environment for
statistical computing. R Foundation for Statistical Computing,
Vienna, Austria. (http://www.R-project.org)
Rennenberg H, Seiler W, Matyssek R et al (2004) Die Buche (Fagus
sylvatica L.)–ein Waldbaum ohne Zukunft im sudlichen Mitte-
leuropa. Allg For Jgdztg 175:210–224
Rigby J, Porporato A (2008) Spring frost risk in a changing climate.
Geophys Res Lett 35:L12703
Sakai A, Weiser CJ (1973) Freezing resistance of trees in North
America with reference to tree regions. Ecology 54:118–126
Saxe H, Cannell MGR, Johnsen Ø et al (2001) Tree and forest
functioning in response to global warming. New Phytol
149:369–399
Schar C, Vidale PL, Luthi D et al (2004) The role of increasing
temperature variability in European summer heatwaves. Nature
427:332–336
Sohn J, Gebhardt T, Ammer C et al (2013) Mitigation of drought by
thinning: short-term and long-term effects on growth and
physiological performance of Norway spruce (Picea abies).
For Ecol Manage 308:188–197
Walck JL, Hidayati SN, Dixon KW et al (2011) Climate change and
plant regeneration from seed. Glob Change Biol 17:2145–2161
Eur J Forest Res
123
Copyright © 2022 FDOKUMEN




















