
Biotechnology and Crop Improvement
65
Journal of Crop Improvement, 24:153–217, 2010 Copyright © Taylor & Francis Group, LLC ISSN: 1542-7528 print/1542-7535 online DOI: 10.1080/15427520903584555 Biotechnology and Crop Improvement SATBIR S. GOSAL, SHABIR H. WANI, and MANJIT S. KANG Punjab Agricultural University, Ludhiana, India Plant biotechnology, a major component of agricultural biotech- nology, deals with various aspects of plant tissue culture, genetic transformation, and molecular biology techniques. Tissue cul- ture methods offer a rich scope for creation, conservation, and utilization of genetic variability for the improvement of field, fruit, vegetable, and forest crops, and medicinal/aromatic plants. Micropropagation technology ensures true to type, rapid and mass multiplication of plants that possesses special significance in veg- etatively propagated plant species. This technology has witnessed a huge expansion globally, with an estimated global market of 15 billion US$/annum for tissue-culture products. Some basic techniques of tissue culture, such as anther/microspore culture, somaclonal variation, embryo culture, and somatic hybridiza- tion, are being exploited to generate useful genetic variability for obtaining incremental improvement in commercial cultivars. Production of secondary metabolites, such as food flavors, food colors, dyes, perfumes, drugs, and scented oils used in aromather- apy, through cell cultures and hairy root cultures, are leading examples of molecular farming. Cryopreservation of germplasm at the cell/tissue/organ levels, in liquid nitrogen at −196 ◦ C, is highly rewarding for establishing germplasm banks, especially for vegetatively propagated crops and rare, endangered plant species. During the past 15 years, remarkable achievements have been made in the production, characterization, field evalua- tion, and release of transgenic varieties/hybrids in several crops. Transgenic varieties/hybrids of maize, cotton, soybean, potato, tomato, and papaya are now being commercially grown on about 134 million hectares spread across 25 countries. Research in genomics allows high-resolution genetic analysis for physical mapping and positional gene cloning of useful genes for crop Address correspondence to Dr. Satbir S. Gosal, School of Agricultural Biotechnology, Punjab Agricultural University, Ludhiana-141004, India. E-mail: [email protected] 153
Transcript of Biotechnology and Crop Improvement

Journal of Crop Improvement, 24:153–217, 2010Copyright © Taylor & Francis Group, LLCISSN: 1542-7528 print/1542-7535 onlineDOI: 10.1080/15427520903584555
Biotechnology and Crop Improvement
SATBIR S. GOSAL, SHABIR H. WANI, and MANJIT S. KANGPunjab Agricultural University, Ludhiana, India
Plant biotechnology, a major component of agricultural biotech-nology, deals with various aspects of plant tissue culture, genetictransformation, and molecular biology techniques. Tissue cul-ture methods offer a rich scope for creation, conservation, andutilization of genetic variability for the improvement of field,fruit, vegetable, and forest crops, and medicinal/aromatic plants.Micropropagation technology ensures true to type, rapid and massmultiplication of plants that possesses special significance in veg-etatively propagated plant species. This technology has witnesseda huge expansion globally, with an estimated global market of15 billion US$/annum for tissue-culture products. Some basictechniques of tissue culture, such as anther/microspore culture,somaclonal variation, embryo culture, and somatic hybridiza-tion, are being exploited to generate useful genetic variabilityfor obtaining incremental improvement in commercial cultivars.Production of secondary metabolites, such as food flavors, foodcolors, dyes, perfumes, drugs, and scented oils used in aromather-apy, through cell cultures and hairy root cultures, are leadingexamples of molecular farming. Cryopreservation of germplasmat the cell/tissue/organ levels, in liquid nitrogen at −196◦C, ishighly rewarding for establishing germplasm banks, especiallyfor vegetatively propagated crops and rare, endangered plantspecies. During the past 15 years, remarkable achievements havebeen made in the production, characterization, field evalua-tion, and release of transgenic varieties/hybrids in several crops.Transgenic varieties/hybrids of maize, cotton, soybean, potato,tomato, and papaya are now being commercially grown onabout 134 million hectares spread across 25 countries. Researchin genomics allows high-resolution genetic analysis for physicalmapping and positional gene cloning of useful genes for crop
Address correspondence to Dr. Satbir S. Gosal, School of Agricultural Biotechnology,Punjab Agricultural University, Ludhiana-141004, India. E-mail: [email protected]
153

154 S. S. Gosal et al.
improvement. Molecular (DNA) markers help in precise character-ization of germplasm, construction of saturated linkage maps, andDNA fingerprinting of crop varieties. Molecular markers are nowincreasingly being used for marker-assisted gene pyramiding andalien gene introgression. Current research, involving large-scaleDNA sequencing, microarrays, and robotics, is heading towardsgene revolution and nanobiotechnology.
KEYWORDS biotechnology, tissue culture, micropropagation,haploids, genetic transformation, transgenic crops, gene-cloning,molecular markers, crop improvement
INTRODUCTION
Biotechnology in a broad sense has been practiced for centuries for curdmaking, food preservation, pickle making, and fermentation. However,biotechnology received a boost during the 1970s with the discovery ofrestriction enzymes, which led to the development of a variety of gene tech-nologies and is, thus, considered the greatest scientific revolution of the 20th
century. Biotechnology, in a true sense, deals with changing and improv-ing, more efficiently than traditional technologies can, the characteristics ofan organism at cellular and molecular levels for the benefit of mankind.Depending upon the biological system (organism) involved, the field ofbiotechnology may be divided into plant biotechnology, microbial biotech-nology, animal biotechnology, and human biotechnology. Plant biotechnol-ogy deals with cell and tissue culture, genetic transformation, gene cloning,DNA markers, and other molecular approaches. Unlike conventional plantbreeding, the biotechnological techniques for genetic modifications largelyoperate at organ, tissue, cell, protoplast, and molecular levels. These inno-vative techniques are considered an adjunct to the conventional methods forefficient and precision plant breeding (Kang et al. 2007).
TISSUE CULTURE
Plant tissue culture broadly refers to the in vitro cultivation of plants,seeds, and plant parts (tissues, organs, embryos, single cells, protoplasts) onnutrient media under closely controlled and aseptic conditions. It includesseveral specialized areas, such as induction of callus and plant regeneration,micropropagation, somatic embryogenesis, somaclonal variation, meristemculture, anther culture, embryo culture, protoplast culture, cryopreserva-tion, and production of secondary metabolites. Tissue-culture methods hold

Biotechnology & Crop Improvement 155
significant promise for creation, conservation, and utilization of genetic vari-ability for improvement of a wide variety of crop plants. Among these,micropropagation, somaclonal variation, and embryo, anther and protoplastculture have direct applications in crop improvement.
Microporpagation of Plants
Micropropagation of plants is one of the best and most successful exam-ples of the commercial application of tissue-culture technology. It entails invitro propagation of plants from very small plant parts (0.2-10 mm) in thelaboratory, followed by their establishment in soil under greenhouse condi-tions. An estimated more than 500 million plants belonging to different plantspecies are now being annually produced through micropropagation in theworld. Micropropagation industry is environment-friendly and requires verylittle raw material in the form of chemicals. This technology possesses thefollowing definite advantages:
1. True-to-type plants produced, i.e., identical to donor (in vitro cloning).2. Selected plant species can be multiplied anywhere in the world.3. Rapid and mass multiplication (1 to 10/cycle of 2 weeks each) of
elite clones/varieties otherwise difficult to multiply through conventionalmethods.
4. Independent of seasonal and raw material constraints.5. Production of disease-free plants.6. Elite plant material for national/international exchange of germplasm,
avoiding the risk of pathogens and insects.7. Micropropagated, field-grown plants give higher yield and exhibit better
quality.
Micropropagation involves four stages: 1) establishment of asepticcultures; 2) shoot bud/shoot multiplication; 3) induction of rooting andhardening; and 4) transfer of plantlets to soil. Micropropagation meth-ods for important field, ornamental, fruit, medicinal, aromatic, and forestplant species are now well established (Table 1). During genetic transfor-mation, this technique helps to increase the plant number of elite eventsfor precise characterization and efficient transfer of regenerated plants togreenhouse.
Likewise, during anther/pollen culture, micropropagation of regener-ants helps to minimize the risk of losing any genotype during hardeningand transfer to soil. Now scores of multimillion-dollar industries around theworld propagate a variety of plant species through tissue culture, whichallows environment-friendly industries to flourish. The clean planting mate-rial can certainly improve the yield potentials of vegetatively propagated

156 S. S. Gosal et al.
crops like sugarcane, potato, banana, strawberry, sweet potato, cassava, andornamental species. It is likely that automation of multiplication systems willbe commercially feasible within the next few years. Improvement of somaticembryogenesis (Gill, Gill, & Gosal 2004), coupled with embryo desiccationand encapsulation technology, may lead to the utilization of artificial seedsfor mass cloning of plants. Micropropagation possesses special significancein the following areas:
1. Production of high quality, disease-free, super elite planting material forfurther seed production, especially in the vegetatively propagated plantspecies.
2. Rapid spread of new varieties of vegetatively propagated crops like sug-arcane, potato, poplar, medicinal/aromatic plants (Gosal & Gill 2004) forcrop diversification.
3. Mass production of ornamental plants otherwise difficult to multiplythrough conventional methods for domestic and international markets.
4. Rejuvenation of old varieties/clones of vegetatively propagated crops forimproving their yield and quality.
5. Mass cloning of rootstocks in horticultural plants like citrus, peach, andapple.
6. Mass cloning of genetically superior (plus trees), cross-pollinated, andseed-propagated trees like eucalyptus.
7. Multiplication of male-sterile lines for hybrid seed production or themultiplication of F1 hybrids and transgenic clones/varieties.
8. Interstate/international exchange of germplasm without the risk ofpathogens and insects.
9. This technology possesses tremendous potential for making environmentclean and green.
Somaclonal Variation
Success of a plant-breeding program largely depends upon the nature andextent of genetic variability available in the germplasm. In this regard,somaclonal variation, i.e., the variation among callus-derived plants, is apotent force for broadening the genetic base, more particularly in the vege-tatively propagated species (Evans & Sharp 1986). Using in vitro selection,many million cells/protoplasts can be screened against various biotic andabiotic stress factors in a single petri dish, which is more efficient as com-pared with field screening that requires more time and space (Kaur et al.2001). Several interesting and potentially useful traits have been recoveredusing this method in sugarcane, potato, tomato, maize, rice, alfalfa, rape-seed, and mustard (Table 1). Recovery of novel variants that either do notexist or are rare in the natural gene pool—for example, atrazine resistancein maize, glyphosate resistance in tobacco, improved lysine and methionine

Biotechnology & Crop Improvement 157
contents in cereals, increased seedling vigor in lettuce, jointless pedicelsin tomato, and Fusarium resistance in alfalfa—is of much significance.Genetic, cytogenetic, and molecular evidences for increased recombinationfrequency through cell cuture have now been provided (Larkin et al. 1993).Tissue culturing of wide-hybrids also helps in breaking undesirable link-ages and achieving introgression from alien sources. Several new varietieshave been developed through somaclonal variation in tomato, sugarcane,potato, celery, brassica, and sorghum. This simple and cost-effective tech-nique possesses a huge potential for the improvement of apomictic andvegetatively propagated plant species and, of course, seed-propagated cropplants with narrow genetic base (Table 1). In India, a somaclonal variantof a medicinal plant, Citronella java, has been released as a commercialvariety, ‘B-3,’ which gives higher yield and oil content than the originalvariety. Likewise, ‘Pusa Jai Kishan’ is a variety of B. juncea released as asomaclonal variant of ‘Varuna’ variety. However, under several situations,low plant-regeneration ability and the lack of correspondence in expres-sion of the trait in field-grown plants are the major problems (Karp 1995).The occurrence of somaclonal variation has been described in a wide vari-ety of crop plants for various traits (Table 1), including potato (Das et al.2000), sugarcane (Leal et al. 1994; Doule 2006; Jalaja et al. 2006; Singh et al.2008; Sengar et al. 2009), banana (Hwang & Ko 2004; James et al. 2007;Oh et al. 2007b), Prunus persica (Hammerschlag & Ognjanov 1990), applerootstocks (Rosati & Predieri 1990), and Heliconia bihai (Rodrigues 2008).Several somaclones have been released as improved cultivars.
Anther/Pollen Culture for Production of Haploids/Doubled Haploids
In self-pollinated crops, an inordinately long period is required to assem-ble desirable gene combinations from different sources in homozygousform. Generally, it takes 8-10 years to develop stable, homozygous andready-to-use materials from a fresh cross of two or more parental lines. Incross-pollinated crops, because of inbreeding depression, it becomes dif-ficult to develop vigorous inbreds for hybrid seed production programs.In this regard, haploids possessing a gametic chromosome number arevery useful for producing instant homozygous true-breeding lines. In addi-tion, haploids constitute an important material for induction and selectionof mutants, particularly for recessive genes. In conventional breeding, theearly segregating-generation populations involve variation attributable toboth additive and non-additive genetic effects (Khush & Virk 2002), whereasdoubled haploid (DH) lines exhibit variation only of additive genetic nature,including additive x additive type of epistatis, which can be easily fixedthrough a single cycle of selection. The elimination of dominance effectsleads to high narrow-sense heritability, and availability of sufficient seed ofeach DH line allows for replicated testing. Thus, in contrast to relatively

158 S. S. Gosal et al.
large segregating populations in conventional genetic studies, fewer DHlines are required for the purpose of selection of desired recombinants. Forinstance, in rice about 150 DH lines derived from F1, instead of 4,000-5,000F2 plants, are sufficient for selecting desirable genotypes. Production of hap-loids has also been exploited during wide hybridization for the developmentof addition and substitution lines.
Anther/pollen culture is an attractive alternative for developing hap-loids (sporophytes with gametophytic chromosome number). One of thevery popular methods for production of haploids is anther or microsporeculture. Incubation of cultures under optimum conditions leads to growthof microspores into sporophytes. The following parameters have been rec-ognized as particularly important for successful anther/microspore culture:(i) growth conditions of donor plant, (ii) genotype of donor plant, (iii)pretreatment of anthers, (iv) developmental stage of anthers/microspores,(v) composition of culture medium, and (vi) physical conditions during cul-ture growth. Anthers are cultured in liquid or on semisolid agar medium (Gillet al. 2003), where they may directly give rise to embryoids or may lead tocallus formation before differentiation. The embryoids develop into haploidplantlets or doubled haploids in some crops (because of spontaneous dou-bling of chromosomes during callus proliferation). Haploids may be treatedwith colchicine to obtain fertile, doubled haploid homozygous plants forfield testing and selection. Production of haploids/doubled haploids throughanther culture from F1 rice plants results in true-breeding plants in lessthan one year, which otherwise takes 7 to 8 generations through conven-tional methods (Gosal et al. 1996). Several cultivars are either in test orhave been released in rice, wheat, maize, rapeseed, and mustard in China,Canada, Denmark, the United States and France (Guzman & Zapata-Arias2000; see Table 1). But in several instances, poor androgenesis, occurrenceof mixoploids, and albino plants have been the recurring problems.
Attempts have also been made to produce haploids in grain-legumeslike pigeon pea (Bajaj, Singh, & Gosal 1980) and vegetable crops, includingtomato (Segui-Simarro & Nuez 2007). In the case of tomato, both game-tophytic and sporophytic calli were produced from cultured anthers. Invitro-induced disturbance of cytokinesis and subsequent fusion of daugh-ter nuclei may be the reason for mixoploidy and genome doubling duringtetrad compartmentalization and callus proliferation (Segui-Simarro & Nuez2007). After the first report on androgenesis (Guha & Maheshwari 1964) fol-lowing this approach, haploids have been produced in more than 247 plantspecies and hybrids belonging to 38 genera and 34 families of dicots andmonocots (http://www.biotechnology4u.com/). Hundreds of varieties havebeen developed through anther culture in rice, brassica, barley, and wheat.The doubled-haploid approach is also being used for the rapid developmentof QTL-mapping populations, construction of genetic linkage maps for traitsof interest, and rapid fixing of transgenes.

Biotechnology & Crop Improvement 159
TABLE 1 Plant Tissue and Protoplast Culture Techniques in Relation to Crop Improvement
Technique Plant material Remarks Reference
Micropropagation Banana Musa spp.(‘Philippine,’ ‘Lacatan,’‘Grande Naine’)Plantain(‘Pelipita’ and ‘Saba’).
Rapidly multiplying culturesfrom excised shoot tips.
Cronauer &Krikorian,1984; Damascoet al. 1997
Strawberry Fragariaananassa
Efficient method for massproduction of plantingmaterial.
Chopra, Dhaliwal& Gosal 1993
Eucalyptus tereticornis In vitro clonal propagationthrough nodal segments.
Gill, Gill, & Gosal1994
Rice Oryza sativa L. var.Jaya
Clonal propagation of indicarice through proliferationof axillary shoots.
Sandhu et al.1995
Dalbergia sissoo Roxb. Rapid in vitro propagationfrom mature trees.
Gill, Gill, & Gosal1997
Banana Musa spp cv.Grande Naine
Sunlight for micropropagationsystems as a way ofreducing tissue culturecosts.
Kodym & Zapata1999
Potato Solanum tuberosum In vitro propagation throughmicrotubers.
Gopal et al. 1998
Dianthus caryophyllusL.(carnation) cv. scania
In vitro propagation throughaxillary shoot proliferation.
Sooch et al. 1998
Sugarcane Saccharumofficinarum L.
Micropropagation protocolfor mass plant production.
Gosal et al.1998;Gill, Malhotra,& Gosal 2006;Sood et al.2006; Jalajaet al. 2006;Jalaja,Neelamathi, &Sreenivasan2008
Poplar P. deltoides Protocol for mass productionof two important clonesviz., G3 and G48 throughinduced shootdifferentiation of leaf, stemand root explants collectedfrom adult trees.
Chaturvedi et al.2004
Brahmi Bacopa monnieri Efficient protocol formicropropagation throughaxillary shoot proliferation.
Gill, Gupta, &Gosal 2004
Rice Oryza sativa L.var.‘Mocoi FCA‘ItapéP.A.‘Fortuna INTA,‘EMBRAPA-7- Taim,’ and‘CT 6919,’ ‘BR IRGA409’)
Clonal propagation protocolusing shoot tip cultures,and the genetic stability ofthe micropropagated plantswas verified by isozymeanalysis.
Medina et al.2004
(Continued)

160 S. S. Gosal et al.
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Potato Solanumtuberosum L.
A protocol for production ofpre-basic potato materialby micro cuttings, obtainedfrom plants with a shortperiod of acclimatization.
Pereira & Fortes2004
Neem Azadirachta indica Macro and micro propagationprotocols.
Gill, Chauhan, &Gosal 2006
Potato (Solanumtuberosum L.) Nif,Clone 122, Agria andResy
The highest microtubernumber (2.8), microtuberyield (278.1 mg) and singlemicrotuber weight (92.2mg) were obtained in theMS medium containing 2.0mg BAP/l and 60 g sucrose.
Ozturk & Yldrm2006
Potato (Solanumtuberosum L.) cv.Marfona
Use of continuous and semicontinuous bioreactors andtheir functions at shootmultiplication and microtuberization of potato.
Ebadi,Iranbakhsh, &Khaniki 2007
Banana Protocol formicropropagation of Musasapientum using shootmeristems.
Kalimuthu,Saravanakumar,& Senthikumar2007
Grapevine (Vitis. vinifera) Best shoot development forthe initial culture ofrootstock VR043-43 in vitrousing nodal segments andbest micro cuttingsmultiplication using e QLmedium.
Machado et al.2007
Strawberry Fragariaananassa
Mass propagation viameristem tip culture.
Mohamed 2007
Sugarcane Saccharumofficinarum L.cv. CoS99259
Spacing of 90cm × 60 cmwas the most suitable fortransplantingtissue-cultured plantlets.
Ramanand et al.2007
Banana Musa spp. Inflorescence apices werefound more suitable forrapid in vitro propagation.
Resmi & Nair2007
Grapevine (Vitis vinifera)cv. ‘Bidaneh Sefid’
Micropropagation protocolfor quick multiplication.
Salami et al. 2007
Sugarcane Saccharumofficinarum L.cv. Co86032
Sets obtained frommicropropagated plantletsresulted in higher seedyields.
Salokhe 2007
Aloe vera Micropropagated plantsexhibited elevated levels ofbioactive compounds.
Thind, Jain, &Gosal 2007
(Continued)

Biotechnology & Crop Improvement 161
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Chrysanthemumcinerariifolium (Trev.)
Rapid propagationtechnology was establishedand optimized in vitro.
Liu & Gao 2007
Banana Musa spp. cv.‘Cavendish Dwarf’ and‘Valery’
Efficient medium for clonalmass propagation (MS + 30gl-1 of sucrose,N-phenyl-N- 1, 2,3-thidiazol 5-yl Urea(0.5 mgl-1) and IAA(2 mgl-1).
Farahani et al.2008
Grape vine Vitis viniferaL. cv. Perlette
Clonal propagation of grapesfor increasing plantmaterial for cultivation.
Jaskani et al.2008
Sugarcane Saccharumofficinarum L
No variation was detectedamong the regeneratedplants of a particularvariety on the basis ofRAPD markers and theprofiles of micropropagated clones werecomparable to those of therespective donor plants.
Lal et al. 2008
Potato Successful micropropagationwas achieved using nodalsegments as explants.
Badoni &Chauhan 2009
Scoparia dulcis A suitable protocol wasestablished throughmultiple shoot inductionfrom nodal segment andshoot tip explants of thisimportant medicinal herb.
Rashid et al. 2009
Somaclonalvariation
Prunus persica cvs.Sunhigh, Red haven
Somaclones S156 and S122resistant to leaf spot,moderately resistant tocanker.
Hammerschlag &Ognjanov 1990
Apple rootstocks (M26,MM106
S-2 (M26) performed betteragainst Phytophthoracactorum.
Rosati & Predieri1990
Banana Giant Cavendisha 10 somaclones. GCTCV215-1released for commercialplanting.
Hwang & Ko,1992; 2004
Apple cv. Greensleeves 16 somaclones. 21% lesssymptoms thanGreensleeves against fireblight strain T.
Donovan et al.1994
(Continued)

162 S. S. Gosal et al.
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Sugarcane Saccharumofficinarum L.
Results confirmed thesuperiority of twosomaclones, one resistantand one tolerant to eyespotdisease.
Leal et al. 1994
Potato Solanumtuberosum L.
Somaclones for heat tolerance Das et al. 2000
Potato Solanum tuberosumL cv. Desiree
Somaclones IBP-10, IBP-27and IBP-30, infected withAlternaria solani andStreptomyces scabiei,exhibited higher resistanceto the pathogen, comparedto the susceptible cultivarDesiree.
Veitia-Rodriguezet al. 2002
Actinidia deliciosa cv.Tamuri
5 somaclones tolerant to NaCl(85.5 mM)
Caboni et al.2003
Potato cv. Desiree, Tomatocv. Amalia leaves,Soyabean cv. William 82Coffee cv. Robusta
Monomorphism was detectedin tomato, coffee, andsoyabean, indicating thegenetic stability of thecrops. Potato callusesshowed variations in theelectrophoretic patterns ofperoxidase and esteraseisoenzymes, indicatingsomaclonal variation in thecrop.
Lara et al. 2003
Oryza sativa L. cv. CICA-8 Four somaclones showedsignificantly higher degreeof partial resistance whencompared with the parentcultivar CICA-8.
Araujo & Prabhu2004
Durum wheat (Triticumdurum Desf.) cv.(Selbera, Sebou, andKyperounda)
Somaclonal variation thusappears to induce a widerange of modificationsamong individualcomponents ofdrought-resistancemechanisms.
Bajji et al. 2004
Sugarcane Saccharumofficinarum L.cv.CP-43/33
The somaclones performedbetter than the sourceplant.
Khan et al. 2004
Maize (Zea mays) Somaclones thus derivedwere tolerant to NaCl.
Zheng et al. 2004
(Continued)

Biotechnology & Crop Improvement 163
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Bread wheat (Triticumaestivum) cv. (Sakha 8,Sakha 69, Giza 157, Giza160, Lerma Rojo 64, andTobari 66)
Somaclones were superior totheir original cultivars.
Ahmed &Abdelkareem2005
Wheat (Triticum aestivum)cv. Sakha 61
Twenty-one out of the 23somaclones outperformedthe original cultivar Sakha61 in terms of leaf rustresistance and grain yield.
Sabry et al. 2005
Oryza sativa L. Somaclones were obtainedfrom anther culture ofhybrid combinations INCALP-10/C4 153,Amistad-82/C4 153, INCALP-10, as well as fromAmistad-82.
Cristo, Gonzalez,& Perez 2006
Sugarcane Saccharumofficinarum L.
Six tissue culture-derivedsugarcane somaclonesTC-434, TC-435, TC-436,TC-237, and TC-045 fromCoC671 and somaclonenumber TC-338 fromCo7219 were evaluated.Somaclone TC-435 gavehigher cane yield at 12months crop age over CoC671. Somaclone TC-435 hadsignificantly higher millablecane height and number ofinternodes than that ofdonor parent CoC671.
Doule 2006
Sugarcane Saccharumofficinarum L.
A new sugarcane variety, Co94012, was released in thename of Phule Savitri forcultivation in Maharashtra,India, for Pre and Suruseasons. It is an early,sugar-rich, high-yieldingvariety with high CCS yield.Co 94012 is a somaclonalvariant of CoC 671, withbetter sucrose content andmoderate resistance to redrot (Glomerellatucumanensis) and smutdiseases (Ustilagoscitaminea). This is the first
Jalaja et al. 2006
(Continued)

164 S. S. Gosal et al.
TABLE 1 (Continued)
Technique Plant material Remarks Reference
sugarcane variety to bereleased in India throughthe use of somaclonalvariation.
Alfalfa Medicago sativa An increase of variability wasnoted in importantquantitative and qualitativetraits compared with theinitial cultivars, includingproductivity of the aboveground mass and seeds,resistance to fungaldiseases, and winterhardiness.
Rozhanskaya2006
Oryza sativa L. cv.‘Pokkali’
The grain yield wassignificantly high insomaclones ‘BTS 11-1,’‘BTS 28,’ ‘BTS 24,’ ‘BTS9-2(S),’ ‘BTS-17(S),’ ‘BTS10-2,’ and ‘BTS 11-7,’ whichexhibited higher flag leafarea and moderate leafarea index compared toother somaclones.
Elanchezhian &Mandal 2007
Olive cv. ‘Frangivento’ Somaclonal variation couldbe found in olive plantsregenerated throughsomatic embryogenesis;this appears in matureplants in the field.
Leva &Petruccelli2007
Banana In banana cultivars (Musa xacuminata, Musa xbalbisiana), somaclonalvariation can be useful inselecting for clones withimproved agronomiccharacteristics.
James et al. 2007;Oh et al. 2007b
Dieffenbachia cvCamouflage, Camille,Star Bright.
Potential for new cultivardevelopment by selectingcallus-derived somaclonalvariants of dieffenbachiawas demonstrated.
Shen et al. 2007
Sugarcane Saccharumofficinarum L.
Development of somaclonesresistant to red rot diseaseusing in vitro and fieldselection.
Singh et al.2008,Sengaret al. 2009
(Continued)

Biotechnology & Crop Improvement 165
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Anther culture/
HaploidProduction
Wheat Doubled haploid wheatvariety ‘Florin’ wasdeveloped.
De Buyser et al.1986
Wheat A single 2,4-D treatmentgiven to spikes one dayafter pollination withmaize, enabled embryos tobe recovered from all 19varieties.
Laurie &Reymondie1991
Rice Revised medium was used forincreasing anther cultureefficacy and improvedfeasibility of using doubledhaploids in genetic andbreeding research withindica rice.
Raina & Zapata1997
Rice ‘Bicol’ first F1 antherculture-derived line froman indica/indica cross insaline-prone areas.
Senadhira et al.2002
Rice An improved method forpollen culture in rice.
Sarao et al. 2003;Grewal, Gill, &Gosal 2006b
Durum Wheat Dicamba and 2,4-D, was bestfor improving the yield ofhaploid plants of durumwheat through crosses withmaize.
García-llamas,Martín, &Ballesteros2004
Maize Zea mays L. Embryogenic induction ofmicrospores within anthersin in vitro conditions wasthe best when combinationof cold treatment, TIBA(0.1 mg l-1) in media andcolchicine (0.02% duringfirst 3 days of culture) wasapplied.
Obert &Barnabas 2004
Citrus (Citrus clementina) Anther culture as a rapid andattractive method ofobtaining new triploidvarieties in clementine.
Germanà et al.2005
Citrus (Citrus clementina) Influence of light quality onanther culture of Citrusclementina Hort. ex Tan.,cultivar Nules wasreported.
Antonietta et al.2005
(Continued)

166 S. S. Gosal et al.
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Maize Zea mays L. 15, 10, 10, and 3 fertiledoubled-haploid plantswere obtained in culturestreated with paraquat,t-BHP, methioninecombined with riboflavin,and menadione,respectively.
Ambrus et al.2006
Wheat Simplified wheat x maizehaploid productionprotocol that is 100%effective across all breadwheat cultivars, generatingdata means of 25% forembryo excision, 90% to95% for plantletregeneration, and between95% to 100% for doubledhaploid (2n = 6x = 42,AABBDD) outputs.
Mujeeb-Kaziet al. 2006
Maize Zea mays L. Maize haploid plants by invitro culture of pollinatedovaries.
Tang et al. 2006
Durum Wheat Triticumdurum
Novel pretreatmentcombining mannitol 0.3Mand cold for seven dayshad a strong effect on thenumber of embryosproduced and regeneratedgreen plants; 11.55 greenplants were produced per100,000 microspores.
Labbani, deBuyser, &Picard 2007
Tomato Lycopersiconesculentum L.
Embryogenesis and plantregeneration by in vitroculture of isolatedmicrospores and wholeanthers of tomato.
Segui-Simarro &Nuez 2007
Rice The 9 DH lines could providethe basic materials forbreeding on Dian-typehybrid rice with both goodquality and high blastresistance in the future.
Zhahg-Yi et al.2008
Maize Doubled haploids should beinduced from F2 plantsrather than from F1 plants.
Bernardo 2009
(Continued)

Biotechnology & Crop Improvement 167
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Cryopreservation Rice Oryza sativa L. Rice plants (cv. Taipei 309)were regenerated fromdifferent cryopreservedcalli, and cryopreservationwas declared to be areliable way to storetransformation competentrice lines.
Moukadiri &Cornejo 1996
Potato Solanumtuberosum L
Shoot tips of in vitro grownpotato (Solanumtuberosum L.) plants werecryopreserved by amodified vitrificationmethod.
Halmagyi et al.2004
Potato Solanumtuberosum L
In vitro plants of potatocultivars ‘Superior’ and‘Atlantic’ were coldacclimated, and axillarybuds were precultured,osmoprotected, exposed toPVS-2 solution, plungedinto liquid nitrogen,thawed, and finally plantedin the regenerationmedium. Aftercryopreservation, vitrifiedshoot tips resumed growthwithin a week.
Zhao et al. 2005b
Rubus spp. (blackberry andraspberry)
Encapsulation-dehydrationand PVS2-vitrificationcryopreservation protocolswere found successful forpreserving diverse Rubusgermplasm (Shoot tips)
Gupta & Reed2006
Arabidopsis Arabidopsis can besuccessfully cryopreservedusing either plantvitrification solution 2(PVS2) or plant vitrificationsolution 3 (PVS3) ascryoprotectants prior torapidly cooling shoot tipsin liquid nitrogen (LN).
Volk et al. 2007
Potato Solanum tuberosumL. cv. Dejima and(STN13)
Protocol was tested with 12selected cultivated varietiesand wild species and thesurvival percentagesobtained ranged between64.0% and 94.4%.
Yoon-Ju et al.2007
(Continued)

168 S. S. Gosal et al.
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Citrus Up to 98.1% of the plantsobtained bycryopreservation were freefrom HLB bacterium, ascompared with a sanitationrate of 25.3% yielded byconventional meristem tipculture.
Ding et al. 2008
Protoplast fusion Potato S. brevidens xS. tuberosum
Somatic hybrids wereproduced by electrofusion.
Fish, Karp, &Jones 1988
Brassica B. juncea, B.nigra and B. carinata XB. napus.
Resistance to Phoma lingamwas expressed in allsymmetric hybrids, and in19 of 24 toxin-selectedasymmetric hybrids.
Sjödin et al. 1989
Potato S. brevidens xS. tuberosum
Twenty hybrids testedexpressed a high level ofresistance to PVY.
Rokka et al. 1994
Brassica Brassica napus xBrassica oleracea
Inoculations Xanthomonascampestris pv campestrisidentified four somatichybrids with highresistance.
Hansen et al.1995
Brassica Brassica oleraceax Brassica rapa
Disease assays showed thatmost somatic hybrids hadlower disease severityratings against bacterial softrot.
Ren, Dickson, &Earle 2000
Citrus Citrus sinensis L.Osbeck x C.volkameriana Pasquale,C. reticulata Blanco
Somatic hybrids combinedcharacteristics from bothsources and have potentialfor tolerance to blight andcitrus tristeza virus (CTV).
Mendes et al.2001
Banana ‘Maçã’(Musa AABgroup) x Lidi’ Musa sp.AA group.
Somatic hybrids wereidentified by using RAPDmarkers.
Matsumoto,Vilarinhos, &Oka 2002
Citrus Mandarin xpummelo, Sweet orangex pummelo
Somatic hybrids wereconfirmed by leafmorphology, ploidyanalysis via flow cytometry,and RAPD analysis. Somatichybrids are beingpropagated by tissueculture for furtherevaluation of diseaseresistance and horticulturalperformance in field trials.
Ananthakrishnanet al. 2006
(Continued)

Biotechnology & Crop Improvement 169
TABLE 1 (Continued)
Technique Plant material Remarks Reference
Potato Aminca-Cardinal xCardinal-Nicola
Complete resistance to PVYwas noted for one somatichybrid line (CN2). All otherhybrids also showedimproved tolerance toPythium aphanidermatuminfection during tuberstorage or after plantinoculation.
Nouri-Ellouzet al. 2006
Potato Solanum tuberosumx Solanum tuberosum
Tetraploid intraspecificsomatic hybrids between16 different diploidbreeding lines of Solanumtuberosum L. wereproduced by PEG-inducedfusion.
Przetakiewiczet al. 2007
Brassica Brassica oleraceax Brassica rapa
Calli were screened by RAPDanalysis for their hybridcharacter. This is the firstreport about a hybridformation between twohaploid protoplasts.
Liu et al. 2007a
Brassica napus(2n = 38) XOrychophragmusviolaceus (2n = 24)Brassicanapus(2n = 38) xOrychophragmusviolaceus (2n = 24)
Symmetric fusions ofmesophyll protoplasts andsubsequent development ofB. napus- O. violaceouschromosome addition lines.
Zhao et al. 2008
Embryo/ Ovule Culture
During wide hybridization, when parents are genetically diverse, endospermdegeneration leads to embryo abortion and the failure of the cross. Undersuch situations, embryo culture/rescue is a practical approach to obtaininterspecific and intergeneric hybrids (Gosal & Bajaj 1983). Embryo cul-ture is also used to produce haploids from wide crosses (Bains et al. 1995;Verma et al. 1999). Embryo culture has been successfully used to transferdesirable genes from wild relatives into cultivated varieties of several fieldand vegetable crops (Kaur, Satija, & Gosal 2002). Ovule culture in grapes(Singh, Brar, & Gosal 1991) aids in developing hybrids, even in seedlessgrapes.

170 S. S. Gosal et al.
Protoplast Culture and Somatic Hybridization
Protoplast culture and somatic cell hybridization involving fusion of proto-plasts from different plant species are important approaches for combiningcharacteristics from otherwise sexually incompatible species. Further, cybrids(cytoplasmic hybrids) and organelle recombinants, not possible through con-ventional methods, can also be developed (Hinnisdaels et al. 1988). Sincethe production of the first somatic hybrid between Nicotiana glauca andN. langsdorfii in 1970, numerous intraspecific, interspecific, and intergenericsomatic hybrids have been produced. Somatic hybrids can be classified intotwo categories, viz., symmetric and asymmetric. Symmetric hybrids carry thecomplete sets of chromosomes from both the parents, whereas asymmetrichybrids possess the full chromosome complement of only one parent andpartial genome of the second parent.
Earlier efforts were to combine full genomes from both the parents andto develop symmetric hybrids among closely related and cross compatiblespecies, where somatic hybrids resembled sexual hybrids. For instance, thesomatic hybrids between Brassica campestris and B. oleracea resembled theB. napus. Later, researchers thought to produce novel hybrids by fusingprotoplasts from genetically distant species. However, the somatic hybridsthus produced, particularly among the phylogenetically distant species, haveexhibited somatic incompatibility, genetic instability, and sterility. It is obvi-ous that such problematic somatic hybrids cannot be incorporated intobreeding programs. Therefore, the interest has moved from creation of novelhybrids to the production of cybrids, chromosome transfer, and gene intro-gression. It is well known that alloplasmic association leads to male sterilityas a consequence of interactions between nuclear and mitochondrial ele-ments. Male sterility has been developed by fusing protoplasts of Nicotianatabacum with X-irradiated protoplasts of N. africana. Likewise, male sterilityhas been transferred from Raphanus sativus into Brassica napus. Moreover,resistance to herbicide triazine has been combined with male sterility byfusing Brassica napus protoplasts from a male-sterile line with B. napusprotoplasts from a triazine-resistant parent. Cytoplasmic genetic male sterilityhas also been successfully transferred into rice.
Resistance to some diseases like potato leaf roll virus, PVX, and PVYhave been incorporated into Solanum tuberosum from Solanum brevidensand Solanum phureja through protoplast fusion (Rokka et al. 1994, Nouri-Ellouz et al. 2006). Likewise, resistance against Phoma lingam disease inBrassica species (Sjödin & Glimelius 1989) and tristeza virus (CTV) in cit-rus (Mendes et al. 2001) was developed through protoplast fusion. Hybridsproduced following protoplast fusion among hybrids of Iris fulva (4x) xIris laevigata; because Iris fulva has unique brown flowers, this trait couldbe very useful for flower color improvement in Iris laevigata, which lacksthis color (Inoue et al. 2006). The protoplast-to-plant system developed in

Biotechnology & Crop Improvement 171
basmati rice (Jain et al. 1995; Kaur et al. 1999) can be exploited for single-step transfer of male sterility from one line to another for production ofhybrid rice. Pollen protoplasts have also been fused using pollen proto-plasts of Brassica oleracea var. italica and haploid mesophyll protoplasts ofBrassica rapa (Liu et al. 2007 a). Calli were screened by random amplifiedpolymorphic DNA analysis for their hybrid character.
Cryopreservation and In Vitro Germplasm Storage
The aim of germplasm conservation is to ensure the ready availability ofuseful germplasm. In seed-propagated crops, seed is extensively used forconservation of germplasm using conventional methods. However, in vege-tatively propagated species where conventional storage techniques are used,it is very difficult to store germplasm for long durations. The conservation ofplant parts in vitro has a number of advantages over in vivo conservation,e.g., in vitro techniques allow conservation of plant species in danger ofbeing extinct. In vitro storage of vegetativly propagated plants can result ingreat savings in storage space and time, and sterile plants that cannot bereproduced generatively can be maintained in vitro. Complete plants havebeen successfully regenerated from tissues cryopreserved at −196◦C in liquidN2 in several crops for several months to years (Ford, Jones, & Van Staden2000; Panis, Swennen, &. Engelmann 2001; Halmagyi et al. 2004; Gupta &Reed 2006; Ding et al. 2008). This method is now being practically used atseveral national and international germplasm banks.
Successful cryopreservation of plant shoot tips is dependent uponeffective desiccation through osmotic or physical processes. Cryoprotectivetreatments, which favor survival of small, meristematic cells and young leafcells, are most likely to produce high survival rates after liquid-nitrogenexposure. Further, microscopy techniques have been used to determinethe extent of cellular damage and plasmolysis that occurs in pepper-mint (Mentha piperita) shoot tips during the process of cryopreservation,using cryoprotectant plant vitrification solution 2 (PVS2) (30% glycerol,15% dimethyl sulfoxide, 15% ethylene glycol, 0.4 M sucrose) prior toliquid-nitrogen exposure (Volk & Casperson, 2007). Arabidopsis, which isincreasingly being used in genomic studies, can be successfully cryopre-served using either PVS2 or PVS3 as cryoprotectants prior to rapidly coolingshoot tips in liquid nitrogen (LN). PVS3 contains 50% glycerol as comparedto PVS2 that contains 30% glycerol. PVS3 was less injurious than PVS2. Allof the shoot tips regrew after LN exposure when cryoprotected with PVS3for 60 min at 22◦C (Towill, Bonnart, & Volk 2006). The high levels of shootformation after LN exposure of Arabidopsis shoot tips make this a desirablesystem in which molecular tools can be used to examine how alterations inbiochemical, metabolic, and developmental processes affect regrowth aftercryoprotective treatments.

172 S. S. Gosal et al.
In Vitro Production of Secondary Metabolites
Several plant species are known to produce secondary metabolites in vivoor in vitro. These metabolites do not perform vital physiological functions,but some act as potential predators and attract pollinators. Further, thesemetabolites act as a valuable source of a vast array of chemical compounds,including fragrances, flavors, natural sweeteners, and industrial feed stocks.Cultured cells/organs produce a wide range of secondary products. Mainlythree approaches have been followed: (i) the rapid growth of cell sus-pension cultures in large volumes, (ii) immobilization of plant cells, and(iii) growing hairy root cultures in vitro. There are several advantages ofthe cell-culture systems over the conventional cultivation for production ofsecondary metabolites, e.g., 1) independence from various environmentalfactors; 2) any cell of the plant could be multiplied to yield specific metabo-lite; and 3) a culture of cells may prove suitable in cases where plants aredifficult or expensive to grow in the field because of their long life cycles.Since the 1970s when the possibility of producing useful secondary productsin plant cell cultures was first recognized, considerable progress has beenmade, and a number of plant species have been found to produce secondaryproducts, such as shikonin, diosgenin, caffeine, glutathione, capsaicin, andanthraquinone (Havkin-Frenkel, Dorn, & Leustek 1997; Varindra et al. 1997;Fischer et al. 1999). Large-scale production of such compounds (molecularfarming) is becoming increasingly popular with the industry where somephysical and chemical conditions for growth and product formation havebeen optimized.
GENETIC TRANSFORMATION AND MOLECULAR APPROACHES:PLANT GENETIC ENGINEERING
During the past 15 years, the combined use of recombinant DNA technology,gene transfer methods, and tissue-culture techniques has led to the effi-cient transformation and production of transgenics in a wide variety of cropplants (Chahal & Gosal 2002). In fact, transgenesis has emerged as an addi-tional tool to carry out single-gene breeding or transgenic breeding of crops.Unlike conventional breeding, only the cloned gene(s) of agronomic impor-tance is/are being introduced without co-transfer of other undesirable genesfrom the donor. The recipient genotype is least disturbed, which eliminatesthe need for repeated backcrosses. Above all, the transformation methodprovides access to a greater gene pool as the gene(s) may come fromviruses, bacteria, fungi, insects, animals, human beings, unrelated plants,and even from chemical synthesis in the laboratory. Various gene-transfermethods (Christou 1994; Sudhakar et al. 1998; Maqbool & Christou 1999;Gosal & Gosal 2000; Ahmed et al. 2002) have been developed for genetic

Biotechnology & Crop Improvement 173
transformation of plants. Rapid and remarkable achievements have beenmade in the production, characterization, and field evaluation of transgenicplants in several field crop, and fruit and forest plant species. However, themajor interest has been in the introduction of cloned gene(s) of agronomicimportance into commercial cultivars for their incremental improvement.Using different gene-transfer methods and strategies, transgenics carryinguseful agronomic traits have been developed and released in several crops.
Engineering for Herbicide Resistance
There have been two approaches to develop herbicide-resistant transgenicplants.
TRANSFER OF GENES WHOSE ENZYME PRODUCTS DETOXIFY THE HERBICIDE
(DETOXIFICATION)
In this approach, the introduced gene produces an enzyme that degrades theherbicide sprayed on the plant. For instance, introduction of the bar gene,cloned from bacteria Streptomyces hygroscopicus into plants, makes themresistant to herbicides. The bar gene produces an enzyme, phosphinothricinacetyl transferase (PAT), which degrades phosphinothricin into a non-toxicacetylated form. Plants engineered with the bar gene were found to growin phosphinothricin at levels 4-10 times higher than those found in normalfield applications (Gordon-Kamm et al. 1990; Choi et al. 2007; Shizukawa& Mii 2008). Likewise, bxn gene of Klebsiella ozaenae, which producesnitrilase enzyme, imparts resistance to plants against herbicide bromoxynil.Other genes, including tfdA for 2, 4-D tolerance and GST gene for atrazinetolerance, have also been used (Table 2).
TRANSFER OF GENES WHOSE ENZYME PRODUCT BECOMES INSENSITIVE TO
HERBICIDE (TARGET MODIFICATION)
Under this approach, a mutated gene is introduced into the plant, whichproduces a modified enzyme in the transgenic plant that is not recognizedby the herbicide. Hence, the herbicide cannot kill the plant. For instance,a mutated aroA gene from Salmonella typhimurium has been used fordeveloping tolerance to glyphosate herbicide. The target site of glyphosateis a chloroplast enzyme 5-enol pyruvylshikimic acid 3-phosphate synthase(EPSPS). Introduction of the mutated aroA gene produces modified EPSPS,not recognizable to glyphosate. Likewise, sulphonylurea and imidazolinoneherbicides inhibit acetolactate synthase (ALS) chloroplast protein. Toleranceto these herbicides has been achieved by engineering the expression of themutated ALS gene derived from plants (Table 2).

174 S. S. Gosal et al.
TABLE 2 Important Transgenes/Transgene Products being Used for Engineering Crop PlantsPossessing Herbicide Resistance
Trnasgene (s) Source Plant species Target herbicide Reference
Bar Streptomyceshygroscopicus
Nicotianatabaccum,Solanumtuberosum,Lycopersiconesculentum
Phosphinothricin andbialaphos
De Block et al.1987
Bar Streptomyceshygroscopicus
Zea mays Glufosinate, bialaphos Gordon-Kammet al. 1990
Bxn Klebsiellaozaenae
Nicotianatabaccum
Bromoxynil(3,5-dibromo-4-hydroxybenzonitrile)
Stalker,Mcbride, &Malyj 1998
Bxn Klebsiellaozaenae
Clover Bromoxynil(3,5-dibromo-4-hydroxybenzonitrile)
Dear et al. 2003
GAT – Arabidopsis,tobacco, maize
Glyphosate Castle et al.2004
Bar Streptomyceshygroscopicus
Sweet potato Glufosinate Choi et al. 2007
CYP1A1,CYP2B6,CYP2C19
Homo sapiens Oryza sativa Metolachlor Kawahigashiet al. 2008
MxPPO Myxococcusxanthus
Tall fescue Oxyfluorfen, acifluorfen Lee et al. 2008
Bar Streptomyceshygroscopicus
Nierembergiarepens
Bialaphos Shizukawa et al.2008
Engineering for Insect Resistance
There have been two approaches to develop insect-resistant transgenicplants by transferring insect-control-protein genes.
INTRODUCTION OF BACTERIAL GENES
Bacillus thuringiensis synthesizes an insecticidal crystal (cry) protein, whichresides in the inclusion bodies produced by the Bacillus during sporula-tion. Upon ingestion by insect larvae, this crystal protein is solublized in thealkaline environment of the midgut of the insect and processed by midgutproteases to produce a protease-resistant polypeptide toxic to the insect.Lepidopteran-specific Bt gene from Bacillus thuringiensis subsp. kurstakihas been widely and successfully used in tobacco, tomato, potato, cotton,rice, and maize to develop resistance against several lepidopteran insectpests (Table 3) (DeMaagd, Bosch, & Stiekema 1999; Gosal et al. 2003; Bashiret al. 2005; Wei et al. 2006; Cao, Shelton, & Earle 2008; Jafari et al. 2009).The use of redesigned synthetic Bt gene has also been made in some of
Biotechnology & Crop Improvement 175
these crops and, in several instances, the synthetic versions have exhibitedup to 500-fold increases in the expression (Brandy et al. 2001). Insect-resistant transgenic varieties/hybrids of several crops have been releasedfor commercial cultivation in many countries (James 2009).
INTRODUCTION OF PLANT GENE(S)
Several insecticidal proteins of plant origin, such as lectins, amylaseinhibitors, and protease inhibitors, can retard insect growth and develop-ment when ingested by insects at high doses (Jongsma & Boulter 1997;Tamayo et al. 2000). Some genes like CpTi, PIN-1, PIN 11, αA-1, and GNAhave been cloned from different plants and are being used in the genetic-transformation programs aimed at developing insect resistance in plants (Buet al. 2007; Maheswaran et al. 2007; McCafferty, Moore, & Zhu 2008). A listof transgenes used for developing insect-resistant transgenic plants is givenin Table 3.
TABLE 3 Important Transgenes/Transgene Products being Used for Engineering Crop PlantsPossessing Insect Resistance
Transgene (s) Source Plant species Target pest Reference
Cry genesCry9C Bacillus
thuringiensisZea mays Ostrinia nubilalis
(Hübner)Jansens et al.
1997Cry1Ab Bacillus
thuringiensisOryza sativa Chilo suppressalis,
Scirpophaga incertulas,Cnaphalocrocismedinalis,Herpitogrammalicarisalis, Sesamiainferens, Narangaanescens, Mycalesisgotama, Hesperiidae,Parnara guttata
Shu et al. 2000
Cry1Ac Bacillusthuringiensis
Oryza sativa Yellow stem borer Gosal et al.2003;Grewal,Gill, & Gosal2006a
Cry1Ac Bacillusthuringiensis
Pinus taeda Dendrolimuspunctatus Walker,CrypyotheleaformosicolaStaud
Tang & Tian2003
Cry1Ac +CpT1
Bacillusthuringiensis,Vignaunguiculata
Oryza sativa Chilo suppressalis Zhang et al.2004
Cry1Ac +cry2A
Bacillusthuringiensis
Oryza sativa Lepidoptera Bashir et al.2005
(Continued)

176 S. S. Gosal et al.
TABLE 3 (Continued)
Transgene (s) Source Plant species Target pest Reference
Cry2A Bacillusthuringiensis
Oryza sativa Lepidoptera Chen et al. 2005
Cry1AbCry1Acfusion
Bacillusthuringiensis
Oryza sativa Lepidoptera Huang et al.2005
Cry1c Bacillusthuringiensis
Oryza sativa Lepidoptera Wei et al. 2006
Cry1Ac +Cry1c
Bacillusthuringiensis
Brassicajuncea
Plutella xylostella Cao, Shelton &Earle 2008
Cry1Ac,Cry1Ie
Bacillusthuringiensis
Nicotianatabacum
Helicoverpa armigera Lian et al. 2008
Cry1Ab Bacillusthuringiensis
Beta vulgaris Spodoptera littoralis Jafari et al. 2009
Cry1Ac +Cry2Ab
Bacillusthuringiensis
Gossypiumhirsutum L.
Helicoverpa armigera L. Gao et al. 2009
GNA genesGNA Galanthus
nivalis.Oryza sativa Nilaparvata lugens Xiaofen et al.
2001GNA Galanthus
nivalis.Zea mays Rhopalosiphum maidis
FitchWang et al.,
2005GNA Galanthus
nivalis.Carica papaya Tetranychus
cinnabarinusMcCafferty,
Moore, & Zhu2008
GNA Galanthusnivalis.
Saccharumofficinarum
Ceratovacuna lanigeraZehnther
Zhangsun et al.2008
CpTI genesCpTI Vigna
unguiculataOryza sativa Chilo. Spp. Xu et al. 1996
CpTI Vignaunguiculata
Triticumaestivum
Sitotroga cerealellaOlivier
Bi et al. 2006
OthersICP Bacillus
thuringiensisOryza sativa Scirpophaga incertulas Nayak et al.
1997PINII-2x Solanum
tuberosumOryza sativa Chilo suppressalis Walker Bu et al. 2007
PI Nicotianatabaccum
Malus Spp. Epiphyas postvittiana Maheswaranet al. 2007
OCII Oryza sativa Medicagosativa
Phytodecta fornicataBrüggemann
Ninkovic et al.2007
Aprotinin Bos Taurus Saccharumofficinarum
Scirpophaga excerptalis Christy et al.2009
Engineering for Disease Resistance
VIRUS RESISTANCE
The genetic engineering of virus-resistant plants has exploited new genesderived from viruses themselves in a concept referred to as ‘pathogen-derived resistance’ (PDR). Some such genes are discussed below.

Biotechnology & Crop Improvement 177
Coat-protein-mediated resistance (CP-MR). Introduction of viral coat-protein gene into plants makes the plants resistant to the virus from whichthe gene for the CP was derived. This was first demonstrated for TMV intobacco (Powel-Abel et al. 1986). Subsequently, virus-resistant transgenicshave been developed in tomato, melon, rice, papaya, potato, and sug-arbeet (Table 4) (Fuchs & Gonsalves 2007). A variety of yellow squashcalled ‘Freedom 11’ has been released in the United States. Several vari-eties of potato, cucumber, and tomato, in which CP-MR has been used, areunder field evaluation. Transgenic papaya resistant to papaya ring-spot virus(PRSV) has been developed and is being commercially grown in the UnitedStates (Ferreira et al. 2002).
TABLE 4 Genetic Engineering for Disease Resistance in Plants
Transgene Source Target species Pathogen Reference
BacterialDiseases
T4 Lysozyme Human Tobacco ErysipheCichoracearum,Pseudomonassyringae pv.tabaci
Nakajima,Muranaka, &Ishige 1994
NPR1 Arabidopsisthaliana
Arabidopsisthaliana
Pseudomonassyringae andPeronosporaparasitica
Cao, Li, & Dong1998
Pto Lycopersiconpimpinellifolium
Lycopersiconesculentum
Pseudomonassyringae pv. tomato
Tang et al. 1999
chly Chicken Potato Erwinia carotovorasubsp, atroseptica
Serrano et al.2000
Synthetic D4E1 Cecropia (insect) Populus tremula,Populus alba
Agrobacteriumtumefaciens,Xanthomonaspopuli pv. Populi.,Hypoxylonmammatum
Mentag et al.2003
NH1 (NPR1) Oryza sativa Oryza sativa Xanthomonas oryzaepv. oryzae
Chern et al.2005
Rxo1 (NBS-LRR) Zea mays Oryza sativa Xanthomonas oryzaepv. oryzicola
Zhao et al.2005a
Xa21 (NBS-LRR) Oryza sativa Oryza sativa Xanthomonas oryzaepv. oryzae
Wang et al.2007; Zhaiet al. 2002
Fungal DiseasesPR-3(I) French Bean Tobacco
RapeseedRhizoctonia solani Broglie et al.
1991Vst1 (Stilbene
(resveratrol)synthase)
Vitis vinifera Nicotianatabacum
Botrytis cinerea Hain et al. 1993
(Continued)

178 S. S. Gosal et al.
TABLE 4 (Continued)
Transgene Source Target species Pathogen Reference
Aglul, RCH10 Alfalfa, Rice Nicotianatabacum
Cercospora nicotianae Zhu et al. 1994
PR-3(I) Rice Rice Rhizoctonia solani Lin, Anuratha, &Datta 1995
Aglul, RCH10 Alfalfa, Rice Alfalfa Phytophthoramegasperma f. sp.medicaginis (Pmm)
Masoud,Mojtaba, &Behzad 1996
PR-3(I) Rice Cucumber Botrytis cinerea Tabei, Kitade, &Nishizawa1998
Vst1 (Stilbene(resveratrol)synthase) pss(pinosylvinsynthase)
Vitis vinifera,Pinus sylvestris
Hordeumvulgare,Triticumaestivum
Botrytis cinerea;Puccinia reconditaf.sp. tritici &Stagonospora(Septoria) nodorum
Leckband &Lorz 1998;Serebriakova,Oldach, &Lorz 2005
RCC2 Rice Grape vine Uncinula necator,Elisinoe ampelina
Yamamoto,Iketani, &Ieki 2000
Synthetic D4E1 Cecropia (insect) Nicotianatabacum
Colletotrichumdestructivum
Cary et al. 2000
AiiA Bacillus Solanumtuberosum
Pectobacterium(Erwinia)carotovora
Dong et al. 2001
RC7 chitinasePR-3
Oryza sativa Oryza sativa Rhizoctonia solani. Datta et al. 2001
gf-2.8 (oxala-teoxidase)
Triticum aestivum Glycine max Sclerotiniasclerotiorum
Cober et al.2003;
Cry1Ab (Bttoxin)
Bacillusthuringiensis
Zea mays Fusarium spp Clements et al.2003;Hammondet al. 2004
Rpi-blb2(NB-LRR)
Solanumbulbocastanum
Solanumtuberosum
Phytophthorainfestans
Van derVossenet al.2003, 2005
Vf (Cf) Malus floribunda Malus domestica Venturia inaequalis Belfanti et al.2004
gf-2.8 (oxalateoxidase)
Triticum aestivum Populuseuramericana
Septoria musiva Liang et al. 2004
Rchit Rice Pigeon pea Kumar et al.2004
9f-2.8 (oxalateoxidase)&TaPERO(peroxidase)
Triticum aestivum Triticumaestivum
Blumeria graminisf.sp. tritici
Altpeter et al.2005
Chit French Bean Cotton Verticillium dahliae Masoud et al.2005
Chi11(chitinase)Tlp (PR-4)
Oryza sativa Oryza sativa Rhizoctonia solani Kalpana et al.2006
(Continued)
Biotechnology & Crop Improvement 179
TABLE 4 (Continued)
Transgene Source Target species Pathogen Reference
NPR1 Arabidopsisthaliana
Triticumaestivum
Fusariumgraminearum
Makandar et al.2006
KP4 Virus infectingUstilago maydis
Triticumaestivum
Tilletia caries Schlaich et al.2006;
Gfzhd101 Clonostachys rosea Zea mays Fusariumgraminearum
Igawa et al.2007
Synthetic D4E1 Cecropia (insect) Gossypiumhirsutum
Thielaviopsis basicola Rajasekaranet al. 2007
PFLP Pepper Tomato R. Solanacearum Huang et al.2007
Chi 18(chitinase)
Solanumtuberosum
Raphanus sativusLinn
Rhizoctonia solani Yang et al. 2009
Viral DiseasesREP (TYLCV) Tomato yellow leaf
curl virus(TYLCV)
Lycopersiconesculentum
TYLCV Yang et al. 2004
SyntheticsequencegeneratingmiRNA basedon P69TYMV) andHC-Pro(TuMV)
Turnip yellowmosaic virus(TYMV) & turnipmosaic Virus(TuMV)
Arabidopsisthaliana
TYMV & TuMV Niu et al. 2006
PSRV coatprotein gene
Papaya ringspotvirus (PRSV)
Carica papaya PRSV Fuchs &Gonsalves2007
C1 (TYLCV) Tomato YellowLeaf Curl Virus(TYLCV)
Lycopersiconesculentum
TYLCV Fuentes et al.2006
ORF2b (PLRV) Potato leaf-rollvirus (PLRV)
Solanumtuberosum
PLRV Vazquez,Asurmend, &Hopp 2001
Suppressor 2b(CMV)
Cucumber mosaicvirus (CMV)
Nicotianatabacum
CMV Qu, Ye, & Fang2007
RNAi Technology. Antiviral RNAi technology has been successfullyused for viral disease management in human cell lines (Bitko & Barik2001; Gitlin, Karelsky, & Andino 2002; Jacque, Triques, & Stevenson 2002;Novina et al. 2002). The effectiveness of the technology in generating virus-resistant plants was first reported for PVY in potato harboring vectors forsimultaneous expression of both sense and antisense transcripts of thehelper-component proteinase gene. Many examples of extreme virus resis-tance and post transcriptional gene silencing of endogenous or reportergenes have been described in transgenic plants containing sense or antisensetransgenes. In these cases of either cosuppression or antisense suppression,there appears to be induction of a surveillance system within the plant

180 S. S. Gosal et al.
that specifically degrades both the transgene and target RNAs. Transformingplants with virus or reporter gene constructs that produce RNAs capable ofduplex formation confered virus immunity or gene silencing on the plants.It has been accomplished by using transcripts from one sense gene and oneantisense gene colocated in the plant genome, a single transcript that hasself-complementarity, or sense and antisense transcripts from genes broughttogether by crossing (Waterhouse, Graham, & Wang 1998). Likewise, RNAi-mediated silencing of African cassava mosaic virus (ACMV) resulted in a 99%reduction in Rep transcripts and a 66% reduction in viral DNA (Vanitharani,Chellappan, & Fauquet 2003). The siRNA was able to silence a closelyrelated strain of ACMV but not a more distantly related virus. More than40 viral suppressors have been identified in plant viruses (Ruiz-Ferrer &Voinnet 2007).
Qu, Ye, and Fang (2007) used a different amiRNA vector to target the2 b viral suppressor of the Cucumber mosaic virus (CMV)—a suppressorthat interacted with and blocked the slicer activity of AGO1. This suppres-sor has been shown to confer resistance to CMV infection in transgenictobacco. A strong correlation between virus resistance and the expressionlevel of the 2 b-specific amiRNA was shown in individual plant lines. Itis evident from the above-mentioned reports that the RNA components,such as single-strand template RNA, dsRNA, and/or siRNA of the silenc-ing pathways, are the preferred targets of most viral suppressors. However,plant viruses are known to have evolved a counter-silencing mechanism byencoding proteins that can overcome such resistance (Li & Ding 2006; Díaz-Pendón & Ding 2008). These suppressors of gene silencing, often involvedin viral pathogenicity, mediate synergism among plant viruses and resultin the induction of more severe disease than that caused by individualviruses. Simultaneous silencing of such diverse plant viruses can be achievedby designing hairpin structures that can target a distinct virus in a singleconstruct (Díaz-Pendón & Ding 2008).
Fungal Resistance
Genetic engineering for fungal resistance has been limited. But several newadvances in this area now present an optimistic outlook. Many reports(Makandar et al., 2006; Huang et al. 2007; Yang et al. 2009) show positiveresults relative to transgenic plants expressing genes for disease resistance(Table 4).
ANTIFUNGAL PROTEIN-MEDIATED RESISTANCE
Introduction of chitinase gene in tobacco and rice has been shown toenhance fungal resistance in plants (Lee & Raikel 1995; Nishizawa et al.

Biotechnology & Crop Improvement 181
1999). Chitinase enzyme degrades the major constituents of the fungal cellwall (chitin and a-1,3 glucan). Coexpression of chitinase and glucanasegenes in tobacco and tomato plants confers a higher level of resistance thanthat imparted by either gene alone. Use of genes for ribosome-inactivatingproteins (RIP), along with chitinase, has also shown synergistic effects.A radish gene encoding antifungal protein 2 (Rs-AFP2) was expressed intransgenic tobacco, and resistance to Alternaria longipes was observed(Broekaert et al. 1995). Other pathogenesis-related proteins/peptides includeosmotin, thionins, and lectins (Florack & Stiekema 1994).
ANTIFUNGAL COMPOUND-MEDIATED RESISTANCE
The low molecular weight compounds, such as phytoalexins, possess antimi-crobial properties and have been implicated in imparting plant resistance tofungal and bacterial pathogens (Leckband & Lorz 1998). Expression of a stil-bene synthase gene from grapevine in tobacco resulted in the production ofa new phytoalexin (resveratrol), which enhanced resistance to infection byBotrytis cinerea. Active oxygen species (AOS), including hydrogen peroxide,also play an important role in plant defense responses to pathogen infection(Wu et al. 1995). Transgenic potato plants expressing an H2O2-generatingfungal gene for glucose oxidase were found to have elevated levels of H2O2
and enhanced levels of resistance both to fungal and bacterial pathogens—particularly to the Verticillium wilt pathogen. Further, overexpression ofdefense-response genes in transgenic plants has provided enhanced resis-tance to a variety of fungal pathogens (Muehlbauer & Bushnell 2003). Forexample, transgenic wheat lines carrying a barley-seed class II chitinaseexhibited enhanced resistance to powdery mildew (Bliffeld et al. 1999;Oldach, Becker, and Lorz 2001). Varying levels of resistance toward powderymildew were observed in transgenic wheat lines carrying a barley chitinaseor a barley β-1,3-glucanase (Bieri, Potrykus, & Futterer 2003). With respectto fusarium head blight (FHB), a transgenic wheat line carrying a rice tlpand a line carrying a combination of a wheat β-1, 3-glucanase and chiti-nase exhibited delayed symptoms of FHB in greenhouse trials (Chen et al.1999; Anand et al. 2003). In addition, transgenic Arabidopsis plants carryingan overexpressed Arabidopsis thionin have exhibited increased resistance toF. oxysporum (Epple, Apel, & Bohlmann 1997). Transgenic wheat express-ing the Arabidopsis NPR1 gene, a gene that regulates defense responses, wasshown to exhibit a high level of resistance to FHB in greenhouse evaluations(Makandar et al. 2006).
RNA-mediated gene silencing (RNA silencing) is being tried as a reversetool for gene targeting in fungi. Homology-based gene silencing induced bytransgenes (co-suppression), antisense RNA, or dsRNA has been demon-strated in many plant pathogenic fungi, including Cladosporium fulvum(Hamada & Spanu 1998), Magnaporthae oryzae (Kadotani et al. 2003),

182 S. S. Gosal et al.
Venturia inaequalis (Fitzgerald, Kha, & Plummer 2004), Neurospora crassa(Goldoni et al. 2004), Aspergillus nidulans (Hammond & Keller 2005), andFusarium graminearum (Nakayashiki et al. 2005). Fitzgerald, Kha, andPlummer (2004), using hairpin-vector technology, have been able to trig-ger simultaneous high-frequency silencing of a green fluorescent protein(GFP) transgene and an endogenous trihydroxynaphthalene reductase gene(THN ) in V. inaequalis. The GFP transgene acted as an easily detectable vis-ible marker, while the trihydroxynaphthalene reductase gene (THN ) playeda role in melanin biosynthesis.
Nakayashiki and colleagues (2005) developed a protocol for silenc-ing the mpg1 and polyketide synthase-like genes. The mpg1 gene is ahydrophobin gene essential for pathogenicity, as it acts as a cellular relayfor adhesion and trigger for the development of appressorium (Talbot et al.1996). Nakayashiki and colleagues (2005) were successful in silencing theabove-mentioned genes to varying degrees by pSilent-1-based vectors in70%–90% of the transformants. Ten to fifteen percent of the silenced trans-formants exhibited almost “null phenotype.” This vector was also efficientlyable to silence a GFP reporter in another ascomycete fungus, Colletotrichumlagenarium.
Bacterial Resistance
Genetic engineering for bacterial resistance has been met with relatively lit-tle success. The expression of a bacteriophage T4 lysozyme in transgenicpotato tubers led to increased resistance to Erwinia carotovora. In addition,the expression of barley a-thionin gene significantly enhanced the resistanceof transgenic tobacco to Pseudomonas syringae. Advances in the cloningof several new bacterial resistance genes (Tables 4, 6) should provide bet-ter understanding of plant-bacterial interactions. Further, RNAi technologyhas been tried to control bacterial diseases. Escobar and colleagues (2001)developed a crown gall disease-management strategy that targets the pro-cess of tumorigensis (gall formation) by initiating RNAi of the iaaM andipt oncogenes. Expression of these genes is a prerequisite for wild-typetumor formation. Transgenic Arabidopsis thaliana and Lycopersicon escu-lentum transformed with RNAi constructs that targeted iaaM and ipt gene(s)showed resistance to the crown gall disease. Transgenic plants generatedthrough this technology contained a modified version of these two bacte-rial gene(s); this was the first report on managing a major bacterial diseasethrough RNAi. The extra genes recognize the bacterial gene and effectivelyshut down the expression of the corresponding bacterial gene during infec-tion, thus preventing the spread of infection. The invading bacteria could notmake the hormones needed to cause tumors; plants deficient in the silenc-ing mechanism were hyper-susceptible to A. tumefaciens (Dunoyer et al.

Biotechnology & Crop Improvement 183
2007). Successful infection relies on a potent anti-silencing state establishedin tumors, whereby siRNA synthesis is specifically inhibited. The proce-dure can be exploited to develop broad-spectrum resistance in ornamentaland horticultural plants susceptible to crown gall. This approach can beadvocated for the effective management of those pathogens that multiplyvery rapidly and cause tumor formation, e.g., Albugo candida, Synchytriumendobioticum, and Erwinia amylovora. The natsiRNA (nat-siRNAATGB2)was strongly induced in Arabidopsis upon infection by Pseudomonassyringae pv. Tomato; it down-regulated a PPRL gene that encodes a neg-ative regulator of the RPS2 disease-resistance pathway. As a result, theinduction of nat-siRNAATGB2 increased the RPS2-mediated, race-specificresistance against P. syringae pv. tomato in Arabidopsis (Katiyar-Agarwalet al. 2006b).
Recently, the accumulation of a new class of sRNA, 30 to 40 nucleotidesin length, designated as long-siRNAs (lsiRNAs), was found to be associatedwith P. syringae infection. One of these lsiRNAs, AtlsiRNA-1, contributed toplant bacterial resistance by silencing AtRAP, a negative regulator of plantdefense (Katiyar-Agarwal et al. 2007). A Pseudomonas bacterial flagellin-derived peptide was found to induce the accumulation of miR393 inArabidopsis. The miR393 negatively regulated mRNAs of F-box auxin recep-tors, causing increased resistance to P. syringae, and the overexpression ofmiR393 was shown to cause a five-fold reduction in plant’s bacterial titer(Navarro et al. 2006).
Engineering for Male Sterility
The exploitation of heterosis (hybrid vigor) through the use of hybrid vari-eties is one of the major achievements of conventional plant breeding.However, in many crops, an efficient and economical method of producinghybrid seed is not available. To overcome these difficulties, genetic engi-neering of male sterility (Williams 1995) and its restoration have emergedas tangible options for the development of male sterile and restorer linesfor hybrid seed production. There are several biotechnological approachesto develop male sterile lines, but the barnase-barstar genes have beenused with greater success. The barnase gene, from the bacterium Bacillusamyloliquefaciens, encodes the enzyme barnase (ribonuclease), which isproduced in the transgenic plant/line during the development of the anthers.Barnase destroys the tapetal tissues, preventing pollen production and con-ferring male sterility. The introduction and expression of the barstar gene,also from Bacillus amyloliquefaciens, into another plant/line result in thedevelopment of a restorer line. The hybrid plants derived from crosses ofmale sterile and restorer lines are fully fertile. This system has been com-mercially exploited in maize and oilseed rape (Denis et al. 1993; Bisht et al.

184 S. S. Gosal et al.
2004, Ray et al. 2007). This method can be extended to other crops for theproduction of hybrid seeds.
Engineering for Improved Nutritional Quality
There is now a growing interest in improving the nutritional quality offood crops. Genes cloned from plants and microbes are being introducedinto crop plants to enhance their nutrional value. For instance, introduc-tion of provitamin A and β carotene genes have resulted in the productionof ‘golden rice’ (Burkhardt et al. 1997; Ye et al. 2000; Beyer et al. 2002).Likewise, high protein ‘phaseolin’ and AmA1 genes have been introducedto hetereologous systems such as tobacco and potato. Intoduction of AmA1gene into potato (Chakraborty, Chakraborty, & Datta 2000; Randhawa, Singh,& Sharma 2009) has improved its yield, protein content, and quality. Vitamin-producing transgenic plants have also been developed (Herbers 2003)and emphasis is being laid on multigene engineering (Daniell & Dhingra,2002). The main objective of these crops is to add value to agri-foods(Newell-McGloughlin 2008).
Engineering for Molecular Farming/Pharming
An additional goal for the development of transgenic plants is the useof living systems for the production of metabolites at the industrial scale,e.g., specialty chemicals, antibodies, pharmaceuticals, edible vaccines, etc.The cell’s metabolic networks that evolved in nature are not optimizedfor industrial production of these metabolites. So, the performance ofmetabolic pathways is manipulated so that metabolites are over-produced.The introduction of heterologous genes and regulatory elements in theliving systems is commonly called ‘metabolic engineering,’ There are anumber of reports dealing with the production of specialty chemicals, bio-pharmaceuticals, and edible vaccines that can be stored and distributed asseeds, tubers, or fruits (Fischer & Emans 2000; Gidding et al. 2000, Daniell,Streatfield, & Wycoff 2001; Daniell & Dhingra 2002; Bonetta 2002; Maliga2003). Solulin is a recombinant, soluble derivative of human thrombomod-ulin (Weisel et al. 1996). To evaluate the production of the pharmaceuticalprotein in plants, expression vectors were generated using four differentN-terminal signal peptides. Immunoblot analysis of transiently transformedtobacco leaves showed that intact ‘solulin’ could be detected using threeof these signal peptides (Schinkel et al. 2005). Furthermore, transgenictobacco plants and BY2 cells producing solulin were generated. It hasbeen shown that plants and plant cell cultures can be used as alterna-tive systems for the production of an active recombinant thrombomodulinderivative.
Biotechnology & Crop Improvement 185
Engineering for Abiotic Stress Tolerance
The transfer of cloned genes has resulted in transgenics tolerant to someabiotic stresses (Gosal, Wani, & Kang 2009; Table 5). For instance, for frostprotection, an antifreeze protein gene from fish has been transferred intotomato and tobacco. Likewise, a gene coding for glycerol-3-phosphate acyl-transferase from Arabidopsis has been transferred to tobacco to enhancecold tolerance in tobacco. Hal2 gene is being tried for developing salt tol-erance. P5CS from Vigna aconitifolia was introduced through the ‘particlegun’ method of gene transfer into Saccharam officinarum under the action
TABLE 5 Plant Genetic Engineering for Abiotic Stress Tolerance
Gene and source Plant species Remarks Reference
P5CS (Pyrroline-5-carboxylatesynthetase) fromVigna aconitifolia
N. tabacum Transgenic plants produced 10-18fold more proline than controlplants. Overproduction ofpraline also enhanced rootbiomass and flowerdevelopment in transgenicplants.
Kishore et al.1995
codA fromA. globiformis
Oryza sativa Transgenic plants had high levelsof glycinebetaine and grewfaster as compared to wild typeson removal of stress.
Sakamoto,Murata, &Murata 1998
bet A and bet B fromE. coli
N. tabacum Transgenic plants showedincreased tolerance to salt stressas measured by biomassproduction of green housegrown plants.
Holmstromet al. 2000
HVA1 from Hordeumvulgare
Triticumaestivum
Transgenic plants showedimproved growth characteristicsin response to soil-water deficits.Field trials showed that HVA1gene had the potential to conferdrought-stress protection ontransgenic spring wheat.
Sivamani et al.2000;Bahieldinet al. 2005
CBF1 from A. thaliana Lycopersiconesculentum
Transgenic tomato plants weremore resistant to water-deficitstress than the wild-type plants.
Hsieh et al.2002
mtlD from E. coli Triticumaestivum
Ectopic expression of the mtlDgene for the biosynthesis ofmannitol in wheat improvestolerance to water stress andsalinity
Abebe et al.2003
DREB 1A fromA. thaliana
Triticumaestivum
Transgenic plants expressingDREB1A gene demonstratedsubstantial resistance to waterstress under greenhouseconditions.
Pellegrineschiet al. 2004
(Continued)

186 S. S. Gosal et al.
TABLE 5 (Continued)
Gene and source Plant species Remarks Reference
CBF3/ DREB 1A fromA. thaliana
Oryza sativa Tolerance to drought and highsalinity without growthretardation or any phenotypicalteration.
Oh et al. 2005
DREB1A fromA. thaliana
Solanumtuberosum
Transgenic plants showedsignificant tolerance againstsalinity stress (1M NaCl).
Behnam et al.2007
GmTP55 from Glycinemax
A. thalianaandN. tabacum
Ectopic expression of GmTP55 inboth Arabidopsis and tobaccoconferred tolerance to salinityduring germination and to waterdeficit during plant growth.Antiquitin may be involved inadaptive responses mediated bya physiologically relevantdetoxification pathway in plants.
Rodrigues et al.2006
CMO(cholinemonooxygenase)from Spinaciaolearceae
Oryza sativa Transgenic plants were tolerant tosalt and temperature stress atseedling stage. CMO expressingrice plants were not effective foraccumulation of glycinebetaineand improvement ofproductivity.
Shirasawa,Takabe, &Kishitani 2006
P5CS from Vignaaconitifolia
Saccharumofficinarum
Stress-inducible prolineaccumulation in transgenicsugarcane plants underwater-deficit stress acts as acomponent of antioxidativedefense system rather than as anosmotic adjustment mediator.
Molinari et al.2007
TPS1-TPS-2 fromE. coli
A. thaliana No morphological growthalterations were observed inlines over-expressing theTPS1-TPS2 construct, while theplants over expressing the TPS1alone under the control of 35Spromoter had abnormal growth,color, and shape.
Miranda et al.2007a
DREB1A fromA. thaliana
Oryza sativa Transgenic rice resulted in anincrease in tolerance to drought,high salinity, andlow-temperature stresses withoutstunting growth.
Oh et al. 2007a
DREB1A fromA. thaliana
N.tabaccum Expression was up-regulated byAtDREB1A in transgenic plantsunder normal conditions andfurther enhanced by high salt,
Cong et al. 2008
(Continued)

Biotechnology & Crop Improvement 187
TABLE 5 (Continued)
Gene and source Plant species Remarks Reference
which at least partiallycontributed to the high salttolerance inAtDREB1A-expressing plants.
AtMYB44 fromArabidopsis
Arabidopsis Transgenic plants exhibitedreduced rate of water loss, asmeasured by the fresh-weightloss of detached shoots, andremarkably enhanced toleranceto drought and salt stresscompared to wild-type plants.
Jung et al. 2008
of AIPC promoter. Results indicated that stress-inducible proline accumu-lation in transgenic sugarcane plants under water-deficit stress acted as acomponent of antioxidative defense system rather than as an osmotic adjust-ment mediator (Molinari et al. 2007). TPS1-TPS2 fusion gene construct wasintroduced into Arabidopsis through Agrobacterium-mediated gene trans-fer under the control of CaMV 35S or stress-regulated Cd 29A promoter.No morphological growth alterations were observed in lines overexpress-ing the TPS1-TPS2 construct, whereas the plants overexpressing the TPS1alone under the control of 35S promoter showed abnormal growth, color,and shape (Miranda et al. 2007a). Thus, it can be concluded that engineer-ing trehalose metabolism in plants can substantially increase their capacityto tolerate abiotic stresses. Late embryogenesis abundant (LEA) proteins aremainly low molecular weight (10–30 kDa) proteins involved in protectinghigher plants from damage caused by environmental stresses, especiallydrought (dehydration) (Hong, Zong-Suo, & Ming-An 2005). OsLEA 3-1 hasbeen transformed into rice through Agrobacterium-mediated gene transfermethod under the control of different promoters. Constitutive and stress-inducible expression of Os LEA 3-1 under the control of CaMV 35S andHVA1-like promoter, respectively, resulted in transgenic rice plants showingincreased tolerance to drought under field conditions (Xiao et al. 2007).
The gene regulating protein factors that regulate gene expressionand signal transduction and function under stress responses may be use-ful for improving the abiotic stress tolerance in plants. These genescomprise regulatory proteins, i.e., transcription factors (bZip, MYC, MYB,DREB, NACs, NAM, ATAF and CUC), protein kinases (MAP kinase, CDPkinase, receptor protein kinase, ribosomal protein kinase, and transcriptionregulation protein kinase), and proteinases (phosphoesterases and phos-pholipase) (Katiyar-Agarwal et al. 2006a). Transgenic overexpression ofHvCBF4 from barley in rice resulted in increased tolerance to drought,

188 S. S. Gosal et al.
high salinity, and low-temperature stresses without stunting plant growth.Using the 60 K Rice Whole Genome Microarrays, 15 rice genes wereidentified that were activated by HvCBF4. When compared with 12 targetrice genes of CBF3/DREB1A, five genes were common to both HvCBF4and CBF3/DREB1A, and 10 and 7 genes were specific to HvCBF4 andCBF3/DREB1A, respectively (Oh et al. 2007a). Overexpression of stress-responsive gene SNAC1 (stress-responsive NAC 1) significantly enhanceddrought resistance in transgenic rice (22%–34% higher seed-set than control)in the field under severe drought stress conditions at the reproductive stagewhile showing no phenotypic changes or yield penalty. The transgenic riceexhibited significant improvement in drought resistance and salt toleranceat the vegetative stage. Compared with wild type, the transgenic rice wasmore sensitive to abscisic acid and lost water more slowly by closing morestomatal pores, yet displayed no significant difference in the rate of photo-synthesis. DNA chip analysis revealed that a large number of stress-relatedgenes were up-regulated in the SNAC1-overexpressing rice plants. SNAC1holds promise in improving drought and salinity tolerance in rice (Hu et al.2006). Several transcription factors have been used to develop transgenicplants tolerant to abiotic stresses (Pellegrineshci et al. 2004; Oh et al. 2005;Cong et al. 2008; Jung et al. 2008).
Engineering for Improved Post-Harvest Preservation
The ripening of fleshy fruits involves changes in color, texture, and fla-vor. This process is mediated through the action of enzymes. The activitiesof many of the key enzymes are regulated by the controlled expressionof ‘ripening genes.’ In climacteric fruits, such as tomato, the expressionof ripening genes is stimulated by ethylene, which functions as a ripen-ing hormone. Many ripening genes have now been cloned, includingpolyglacturonase and pectinesterase involved in cell wall softening; phy-totene synthase, required for carotenoid synthesis; and ACC synthase andACC oxidase that catalyse the production of ethylene. A number of tech-niques are now available for overexpression of particular genes in fruits tochange their physiological and biochemical properties. The techniques ofgene silencing either using antisense genes or sense-supression have beenused successfully to reduce or inactivate the expression of specific genesand determine their function. Ripening of fruits and senescence in flow-ers can be delayed by antisense expression of genes involved in pectinmetabolism or ethylene biosysnthesis. Genetically engineered tomatoes pos-sess lower activity of the ripening enzyme, polygalacturonase, which delaysripening. Transgenic tomato variety ‘Flavr Savr’ possessing longer shelf lifewas released in United States. Reduction in ethylene synthesis has beenshown to improve quality and storage life of tomato and melon. Fruit ripen-ing was delayed by controlling ethylene production in apple (Yao et al.

Biotechnology & Crop Improvement 189
1995; Hrazdina et al. 2000), grapes (Mehlonbacher 1995), and citrus (Wonget al. 2001). Further, these approaches are now being used to improve flow-ering plants by changing plant architecture, flower color, and shape, andto improve vase-life of cut flowers to meet consumer demand. Transgeniccarnations and geraniums that produce less ethylene exhibit delayed petalsenescence. Attempts are now being made to develop marker gene-freetransgenic plants carrying only the gene(s) of agronomic importance toaddress biosafety issues (Puchta 2003).
GENE CLONING
With the availability of a variety of cloning vectors, useful genes can now beisolated, cloned, and characterized. The discovery of the reverse transcrip-tase enzyme made it possible to isolate genes for specific proteins or geneshaving tissue-specific expression. Nowadays, PCR cloning has become rou-tine especially for those genes whose base sequences are known. Severalgenes, particularly for disease resistance, have been isolated and cloned(Table 6) from genomic or cDNA libraries using heterologus or synthetricprobes. Using such genes, suitable gene constructs carrying marker gene(s),promoter, and terminator sequences are developed for plant transformation.
MOLECULAR MARKERS
Development and utilization of molecular markers to detect differencesin the DNA of individual plants has many applications of value to cropimprovement. These differences are known as molecular markers becausethey are often associated with specific genes and act as ‘signposts’ to thosegenes. Several types of molecular markers that have been developed andare being used in plants include: restriction fragment-length polymorphism(RFLP), amplified fragment-length polymorphism (AFLP), randomly ampli-fied polymorphic DNA (RAPD), sequence-tagged sites (STS), expressedsequence tags (ESTs), simple sequence repeats (SSRs) or microsatellites,sequence-characterized amplified regions (SCARs), and single nucleotidepolymorphisms (SNPs)(Paterson, Tanksley, & Sorrels 1991; Hoisington,Listman, & Morris 1998; Joshi, Ranjekar, & Gupta 1999). Such markers,closely linked to genes of interest, can be used to select indirectly for thedesirable allele, which represents the simplest form of marker-assisted selec-tion (MAS) and is now being exploited to accelerate backcross breedingand pyramid several desirable alleles (Singh et al. 2001). Using molecularmarkers, genotypes can be distinguished at plant, tissue, or even cellularlevel. These molecular markers are not stage-specific. Selection of a markerflanking a gene of interest allows selection for the presence (or absence) of
190 S. S. Gosal et al.
TABLE 6 List of Cloned/Fine Mapped Important QTL for Disease Resistance
Gene Host Pathogen Reference
Xa1 Rice Xanthomonas oryzae pv.Oryzae
Yoshimura et al.1998
RPM1 Arabidopsis Pseudomonas syringae Boyes, Nam, &Dangl 1998
Pm24 Wheat Blumeria graminis f. sp. Tritici Huang et al. 2000Ve Tomato Verticillium dahliae Kawchuk et al. 2001Lr10 Wheat Puccinia triticina Feuillet et al. 2003Pm5e Wheat Blumeria graminis f. sp. tritici Huang et al. 2003Pm3 Wheat Blumeria graminis f. sp. tritici Huang et al. 2004PmPS5A,
PmPS5BTriticum
carthlicumBlumeria graminis f. sp. tritici Zhu et al. 2005
Pi-d2 Rice Magnaporthae oryzae Chen et al. 2006Pm34 Aegilops
tauschii cossBlumeria graminis f. sp. tritici Miranda et al. 2006
PmY201 andPmY212
Aegilopstauschii Coss
Blumeria graminis f. sp. tritici Sun et al. 2006
Pi9 Rice Magnaporthae oryzae Qu et al. 2006NCA6 Pm T. monococcum Blumeria graminis f. sp. tritici Miranda et al. 2007cPm35 Aegilops tauschii Blumeria graminis f. sp. tritici Miranda et al. 2007bPi40(t) Rice Magnaporthae oryzae Jeung et al. 2007Pi36 Rice Magnaporthae oryzae Liu et al. 2007bPi-ta Rice Magnaporthae oryzae Huang et al. 2008Snn1 Wheat Stagonospora nodorum Reddy et al. 2008Lr 60(LrW2) Wheat Puccinia triticina Hiebert et al. 2008Pi20(t) Rice Magnaporthae oryzae Li et al. 2008Pik-h Rice Magnaporthae oryzae Xu et al. 2008Pm36 Triticum turgidum
var. dicoccoidesBlumeria graminis f. sp. tritici Blanco et al. 2008
Pm37 Triticumtimopheevii
Blumeria graminis f. sp. tritici Perugini et al. 2008
Pm23 Wheat Blumeria graminis f. sp. tritici Hao et al. 2008Lr34/Yr18 Wheat (Puccinia triticina) and
(P. striiformis f. sp. tritici)Spielmeyer et al.
2008YrC591 Wheat Puccinia striiformis f. sp.
TriticiLi et al. 2009
Pm43 Thinopyrumintermedium
Blumeria graminis f. sp. Tritici He et al. 2009
a gene in new progeny. Thus, molecular markers can be used to follow anynumber of genes during the breeding program (Paran & Michelmore 1993).DNA markers are now extensively being used for gene mapping/tagging(Vikal et al. 2004; Gill et al. 2007; Kuraparthy et al. 2007a, b; Singh et al.2007; Gill et al. 2008; Lagudah et al. 2009; Li, Niu, & Chen 2009; Tsilo,Chao, & Jin 2009). Further, these can also be used for dissecting poly-genic traits into their Mendelian components or quantitative trait loci (Luo &Kearsey 1992; Yamasaki et al. 1999; Table 7). Likewise, molecular markersare also extensively used to probe the level of genetic diversity among dif-ferent varieties and related species. The applications of such evaluations aremany, including DNA profiling, fingerprinting for patenting and IPR issues,
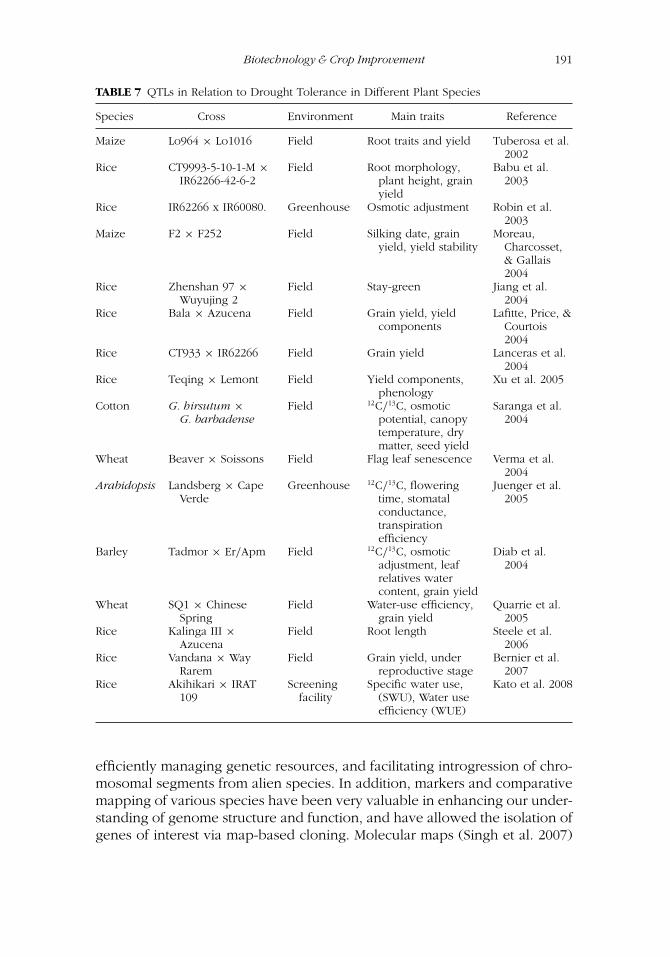
Biotechnology & Crop Improvement 191
TABLE 7 QTLs in Relation to Drought Tolerance in Different Plant Species
Species Cross Environment Main traits Reference
Maize Lo964 × Lo1016 Field Root traits and yield Tuberosa et al.2002
Rice CT9993-5-10-1-M ×IR62266-42-6-2
Field Root morphology,plant height, grainyield
Babu et al.2003
Rice IR62266 x IR60080. Greenhouse Osmotic adjustment Robin et al.2003
Maize F2 × F252 Field Silking date, grainyield, yield stability
Moreau,Charcosset,& Gallais2004
Rice Zhenshan 97 ×Wuyujing 2
Field Stay-green Jiang et al.2004
Rice Bala × Azucena Field Grain yield, yieldcomponents
Lafitte, Price, &Courtois2004
Rice CT933 × IR62266 Field Grain yield Lanceras et al.2004
Rice Teqing × Lemont Field Yield components,phenology
Xu et al. 2005
Cotton G. hirsutum ×G. barbadense
Field 12C/13C, osmoticpotential, canopytemperature, drymatter, seed yield
Saranga et al.2004
Wheat Beaver × Soissons Field Flag leaf senescence Verma et al.2004
Arabidopsis Landsberg × CapeVerde
Greenhouse 12C/13C, floweringtime, stomatalconductance,transpirationefficiency
Juenger et al.2005
Barley Tadmor × Er/Apm Field 12C/13C, osmoticadjustment, leafrelatives watercontent, grain yield
Diab et al.2004
Wheat SQ1 × ChineseSpring
Field Water-use efficiency,grain yield
Quarrie et al.2005
Rice Kalinga III ×Azucena
Field Root length Steele et al.2006
Rice Vandana × WayRarem
Field Grain yield, underreproductive stage
Bernier et al.2007
Rice Akihikari × IRAT109
Screeningfacility
Specific water use,(SWU), Water useefficiency (WUE)
Kato et al. 2008
efficiently managing genetic resources, and facilitating introgression of chro-mosomal segments from alien species. In addition, markers and comparativemapping of various species have been very valuable in enhancing our under-standing of genome structure and function, and have allowed the isolation ofgenes of interest via map-based cloning. Molecular maps (Singh et al. 2007)

192 S. S. Gosal et al.
have been developed for several crop plants such as tomato, wheat, andrice. Molecular markers have facilitated the dissection of quantitative traitsinto their simple genetic components (Tanksley 1993; Pelleschi et al. 2006;Bernier et al. 2007; Kato et al. 2008). Further, these markers assist selectionand pyramiding of QTL alleles through MAS (Ribaut et al. 2004; Chhunejaet al. 2007; Neeraja et al. 2007; Ribaut & Ragot 2007). A novel upland ricevariety, Birsa Vikas Dhan 111 (PY 84), has recently been released in theIndian state of Jharkhand (Steele 2009). It was developed using marker-assisted backcrossing, with selection for multiple quantitative trait loci (QTL)for improved root growth to improve its performance under drought condi-tions. It is an early-maturing, drought-tolerant and high-yielding variety withgood grain quality suitable for direct-seeding in uplands and transplantingin medium lands of eastern India. It outyields its recurrent parent by 10%under rainfed conditions.
CONCLUSIONS
Innovative approaches of cellular and molecular biotechnology haveemerged as a valuable adjunct to supplement and complement the con-ventional methods for precise and efficient breeding of a wide varietyof crop plants. Biotechnological approaches are now increasingly beingused for creation, conservation, characterization, and utilization of geneticvariability for germplasm enhancement. Micropropagation ensures true-to-type and rapid multiplication for bulk production of super-elite plantingmaterial, especially of the vegetatively propagated plant species. This tech-nology has witnessed a huge expansion globally, with an estimated globalmarket of US$15 billion/annum for tissue-culture products. Techniquesof anther/microspore culture, somaclonal variation, embryo culture, andsomatic hybridization are being exploited to obtain incremental improve-ments in plants. Production of secondary metabolites, such as foodflavors, food colors, dyes, perfumes, drugs, and scented oils used in aro-matherapy, through cell culture/hairy root culture are leading examplesof molecular farming/pharming. Cryopreservation of germplasm at thecellular/tissue/organ level in liquid nitrogen at −196◦C is highly rewardingfor establishing germplasm banks, especially in the case of vegetatively prop-agated and rare, endangered plant species. Several genes, particularly forinsect and disease resistance, have been isolated and cloned from differentorganisms using genomic or cDNA libraries, heterologus or synthetic probes,transposon tagging, promoter- enhancer traps, subtractive hybridization, andmap-based cloning.
Several vector and vectorless methods have been developed for genetictransformation of plants. Agrobacterium and ‘particle gun’ methods havebecome routine and are now being widely used for efficient production of

Biotechnology & Crop Improvement 193
transgenic plants. In fact, transgenesis has emerged as an additional toolto carry out single-gene breeding or transgenic breeding of crop plants.Unlike conventional breeding, only the cloned genes of agronomic impor-tance are introduced without co-transfer of other, undesirable genes fromthe donor. The transformation method provides access to a larger genepool as the gene(s) can be acquired from viruses, bacteria, fungi, insects,animals, human beings, unrelated plants, and even from chemical synthe-sis in the laboratory. Remarkable achievements have been made in theproduction, characterization, and field evaluation of transgenic plants in sev-eral field, fruit, vegetable, ornamental and forest crops. Transgenic varietiesbelonging to 10 crops are now being commercially grown across an area ofabout 134 million hectares spread across 25 countries. Genetically modifiedfoods, such as tomato containing high-lycopene flavonols as antioxidants,cavity-fighting apples, golden rice with enhanced vitamin A, golden brassicawith pro-vitamin A, canola rich in vitamin E, proteinaceous potatoes, ediblevaccines, decaffeinated tea and coffee, and nicotine-free tobacco are lead-ing examples of genetically engineered crops. Several types of molecularmarkers, such as RFLPs, AFLPs, RAPDs, STS, ESTs, SSRs or microsatellites,SCARs, SNPs, and insertions/deletions (INDELS) are being used in allelemining, developing saturated linkage maps, marker-assissted breeding, andgene pyramiding. Biotechnolgy has made tremendous a contribution towardincreasing crop yields; improving nutritional quality; enabling crops to beraised under adverse conditions; increasing shelf life of fruits, vegetablesand flowers; and developing resistance to pests and diseases.
REFERENCES
Abebe, T., A.C. Guenzi, B. Martin, and J.C. Cushman. 2003. Tolerance of mannitol-accumulating transgenic wheat to water stress and salinity. Plant Physiol.131:748–1755.
Ahmed, A., S.B. Maqbool, S. Riazudin, and M.B. Sticklen. 2002. Expression of syn-thetic Cry1Ab and Cry1Ac genes in basmati rice (Oryza sativa L.) variety 370 viaAgrobacterium-mediated transformation for the control of the European cornborer (Ostrinia nubilalis ) In Vitro Cell. Dev. Biol. Plant 38:213–220.
Ahmed, K.Z., and A.A. Abdelkareem. 2005. Somaclonal variation in bread wheat(Triticum aestivum L.). II. Field performance of somaclones. Cereal Res.Commu. 33:485–492.
Altpeter, F., A. Varshney, O. Abderhalden, D. Douchkov, C. Sautter, J. Kumlehn, R.Dudler, and P. Schweizer. 2005. Stable expression of a defense-related genein wheat epidermis under transcriptional control of a novel promoter conferspathogen resistance. Plant Mol. Biol. 57:271–283.
Ambrus, H., E. Darko, L. Szabo, F. Bakos, Z. Kiraly, and B. Barnabas. 2006. In vitromicrospore selection in maize anther culture with oxidative-stress stimulators.Protoplasma 228:87–94.

194 S. S. Gosal et al.
Anand, A., T. Zhou, H.N. Trick, B.S. Gill, W.W. Bockus, and S. Muthukrishnan.2003. Greenhouse and field testing of transgenic wheat plants stably expressinggenes for thaumatin-like protein, chitinase and glucanase against Fusariumgraminearum. J. Exp. Bot. 54:1101–1111.
Antonietta, G.M., C. Benedetta, I. Calogero, and M. Rosario. 2005. The effect of lightquality on anther culture of Citrus clementina Hort. ex Tan. Acta Physiol. Plant.27:717–721.
Araujo, L.G., and A.S. Prabhu. 2004. Partial resistance to blast in somaclones of ricecultivar CICA-8. Fitopatologia Brasileira 29:394–398.
Ananthakrishnan, G., M. Calovic, P. Serrano, and J.W. Grosser. 2006. Productionof additional allotetraploid somatic hybrids combining mandarins and sweetorange with pre-selected pummelos as potential candidates to replace sourorange rootstock. In Vitro Cell. Dev. Biol. Plant 42:367–371.
Babu, C.R., B.D. Nguyen, V. Chamarerk, P. Shanmugasundaram, P. Chezhian,P. Juyaprakash, S.K. Ganesh, A. Palchamy, S. Sadasivam, and S. Sarkarung.2003. Genetic analysis of drought resistance in rice by molecular mark-ers: Association between secondary traits and field performance. Crop Sci.43:1457–1469.
Badoni, A., and J.S. Chauhan. 2009. Single node callus culture: improvement formicropropagation of Solanum tuberosum (cv. Kufri Himalini) Nat. Sci. 7:99–103.
Bahieldin, A., H.T. Mahfouz, H.F. Eissa, O.M. Saleh, A.M. Ramadan, I.A. Ahmed, W.E.Dyer, H.A. El-Itriby, and M.A. Madkour. 2005. Field evaluation of transgenicwheat plants stably expressing the HVA1 gene for drought tolerance. Physiol.Planta. 123:421–427.
Bains, N.S., J. Singh, Ravi, and S.S. Gosal. 1995. Production of wheat haploid throughembryo rescue from wheat x maize crosses. Curr. Sci. 69:621–623.
Bajaj, Y.P.S., H. Singh, and S.S. Gosal. 1980. Haploid embryogenesis in anther cultureof pigeonpea (Cajanus cajan). Theor. Appl. Genet. 58:157–159.
Bajji, M., P. Bertin, S. Lutts, and J.M. Kinet. 2004. Evaluation of drought resistance-related traits in durum wheat somaclonal lines selected in vitro. Aust. J. Exp.Agri. 44:27–35.
Bashir, K., T. Husnain, T. Fatima, N. Riaz, R. Makhdoom, and S. Riazuddin. 2005.Novel Indica basmati line (B-370) expressing two unrelated genes of Bacillusthuringiensis is highly resistant to two lepidopteran insects in the field. CropProt. 24:870–879.
Behnam, B., A. Kikuchi, F. Celebi-Toprak, S. Yamanaka, M. Kasuga, K. Yamaguchi-Shinozaki, and K.N. Watanabe. 2007. The Arabidopsis DREB1A gene driven bythe stress-inducible rd29A promoter increases salt-stress tolerance in proportionto its copy number in tetrasomic tetraploid potato (Solanum tuberosum). PlantCell Rep. 26 :1275–1282.
Belfanti, E., D.E. Silfverberg, S. Tartarini, A. Patocchi, M. Barbieri, J. Zhu,B.A. Vinatzer, L. Gianfranceschi, C. Gessler, and S. Sansavini. 2004. The HcrVf2gene from a wild apple confers scab resistance to a transgenic cultivated variety.Proc. Nat. Acad. Sci. 101:886–890.
Bernardo, R. 2009. Should maize doubled haploids be induced among F1 or F2plants? Theor. Appl. Genet. 119:255–262.

Biotechnology & Crop Improvement 195
Bernier, J., A. Kumar, V. Ramaiah, D. Spaner, and G. Atlin. 2007. A large-effect QTLfor grain yield under reproductive-stage drought stress in upland rice. Crop Sci.47:505–516.
Beyer, P., S. Al-Babili, X. Ye, P. Lucca, P. Schaub, R. Welsch, and I. Potrykus.2002. Golden rice: Introducing the β-carotene biosynthesis pathway into riceendosperm by genetic engineering to defeat vitamin A deficiency. J. Nutr.132:506–510.
Bi, R.M., H.Y. Jia, D.S. Feng, and H.G. Wang. 2006. Production and analysis oftransgenic wheat (Triticum aestivum L.) with improved insect resistance by theintroduction of cowpea trypsin inhibitor gene. Euphytica 151:351–360.
Bieri, S., I. Potrykus, and J. Futterer. 2003. Effects of combined expression of anti-fungal barley seed proteins in transgenic wheat on powdery mildew infection.Mol. Breed. 11:37–48.
Bisht, N.C., A.Jagannath, V. Gupta, P.K. Burma, and D. Pental. 2004. A two gene–two promoter system for enhanced expression of a restorer gene (Barstar) anddevelopment of improved fertility restorer lines for hybrid seed production incrop plants. Mol. Breed. 14:129–144.
Bitko, V., and S. Barik. 2001. Phenotypic silencing of cytoplasmic genes withsequence specific double stranded short interfering RNA and its applicationsin the reverse genetics of wild type negative strand RNA virus. BMC Microbiol.1:34–44.
Blanco, A., A. Gadaleta, A. Cenci, A.V. Carluccio, A.M.M. Abdelbacki, andR. Simeone. 2008. Molecular mapping of the novel powdery mildew resistancegene Pm36 introgressed from Triticum turgidum var. dicoccoides in durumwheat. Theor. Appl. Genet. 117:135–142.
Bliffeld, M., J. Mundy, I. Potrykus, and J. Futterer. 1999. Genetic engineering ofwheat for increased resistance to powdery mildew disease. Theor. Appl. Genet.98:1079–1086.
Bonetta, L. 2002. Edible vaccines: Not quite ready for prime time. Nat. Med. 8:94–97.Boyes, D.C., J. Nam, and J.L. Dangl. 1998. The Arabidopsis thaliana RPM1 dis-
ease resistance gene product is a peripheral plasma membrane protein that isdegraded coincident with the hypersensitive response. Proc. Natl. Acad. Sci.95:15849–15854.
Brandy, D.C., M. William, L. Seung-Bum, M. Michael, and D. Henry. 2001.Overexpression of the Bt cry2Aa2 operon in chloroplasts leads to formationof insecticidal crystals. Nat. Biotech. 19:71–77.
Broekaert, W.F., F.R.G. Terras, B.P.A. Cammue, and R.W. Osborn. 1995. Plantdefensins: novel antimicrobial peptides as components of the host defensesystem Plant Physiol. 108:1353–1358.
Broglie, K., I. Chet, M. Holliday, et al. 1991. Transgenic plants with enhancedresistance to the fungal pathogen Rhizoctonia solani. Sci. 254:1194–1197.
Bu, Q.Y., L. Wu, S.H. Yang, and J.M. Wan. 2007. Cloning of a potato proteinaseinhibitor gene pinii-2x from diploid potato (Solanum phurejia l.) and transgenicinvestigation of its potential to confer insect resistance in rice. J . Integr. PlantBiol. 48:732–739.
Burkhardt, P.K., P. Beyer, J. Wünn, A. Klöti, G. Armstrong, M. Schledz, J.L. Von, andI. Potrykus.1997. Transgenic rice (Oryza sativa) endosperm expressing daffodil

196 S. S. Gosal et al.
(Narcissus pseudonarcissus) phytoene synthase accumulates phytoene, a keyintermediate of provitamin A biosynthesis. Plant J. 11:1071–1078.
Caboni, E., S. Anselmi, E. Donato, and F. Manes. 2003. In vitro selection ofActinidia deliciosa clones tolerance to NaCl and their molecular and in vivoecophysiological characterisation. Acta Hort. 618:77–83.
Cao, H., X. Li, and X.N. Dong. 1998. Generation of broadspectrum disease resis-tance by overexpression of an essential regulatory gene in systemic acquiredresistance. Proc. Nat. Acad. Sci. 95:6531–6536.
Cao, J., A.M. Shelton, and E.D. Earle. 2008. Sequential transformation to pyramidtwo Bt genes in vegetable Indian mustard (Brassica juncea L.) and its potentialfor control of diamondback moth larvae. Plant Cell Rep. 27:479–487.
Cary, J.W., K. Rajasekaran, J.M. Jaynes, and T.E. Cleveland. 2000. Transgenic expres-sion of a gene encoding a synthetic antimicrobial peptide results in inhibitionof fungal growth in vitro and in planta. Plant Sci. 154:171–181.
Castle, L.A., D.L. Sieh, R. Gorton, P.A. Patten, Y.H. Chen, S. Bertain, H.J. Cho,N. Duck, J.Wong, D. Liu, and M.W. Lassner. 2004. Discovery and directedevolution of a glyphosate tolerance gene. Sci. 304:1151–1154.
Chahal, G.S., and S.S. Gosal. 2002. Principles and procedures of plant breeding:biotechnological and conventional approaches. New Dehli: Narosa Publishing.
Chakraborty, S., N. Chakraborty, and A. Datta. 2000. Increased nutritive valueof transgenic potato by expressing a nonallergenic seed albumin gene fromAmaranthus hypochondriacus. Proc. Natl. Acad. Sci. 97:3724–3729.
Chapman, E.J., A.I. Prokhnevsky, K. Gopinath, V.V. Dolja, and J.C. Carrington. 2004.Viral RNA silencing suppressors inhibit the micro-RNA pathway at an interphasestep. Genes Dev. 18:1179–1186.
Chaturvedi, H.C., A.K. Sharma, B.Q. Agha, M. Jain, and M. Sharma. 2004. Productionof cloned trees of Populus deltoides through in vitro regeneration of shoots fromleaf, stem and root explants and their field culivation. Ind. J. Biotech. 3:203–208.
Chen, W.P., P.D. Chen, D.J. Liu, R. Kynast, B. Friebe, R. Velazhahan,S. Muthukrishnan, and B.S. Gill. 1999. Development of wheat scab symp-toms is delayed in transgenic wheat plants that constitutively express a ricethaumatin-like protein gene. Theor. Appl. Genet. 99:755–760.
Chen, H., W. Tang, C. Xu, X. Li, Y. Lin, and Q. Zhang. 2005. Transgenic Indicarice plants harboring a synthetic cry2A gene of Bacillus thuringiensis exhibitenhanced resistance against lepidopteran rice pests. Theor. Appl. Genet. 111:1330–1337.
Chen, X., J. Shang, D. Chen, C. Lei, Y. Zou, W. Zhai, G, Liu, J. Xu, Z. Ling, G. Cao,B. Ma, Y. Wang, X. Zhao, S. Li, and L. Zhu. 2006. A B-lectin receptor kinasegene conferring rice blast resistance. Plant J . 46:794–804.
Chern, M.S., H.A. Fitzgerald, P.E. Canlas, D.A. Navarre, and P.C. Ronald. 2005.Overexpression of a rice NPR1 homolog leads to constitutive activation ofdefense response and hypersensitivity to light. Mol. Plant- Microbe Interactions18:511–520.
Chhuneja, P., S. Kaur, R.K. Goel, M. Aghaee-Sarbarzeh, M. Prashar, andH.S. Dhaliwal. 2007. Transfer of leaf rust and stripe rust resistance from Aegilopsumbellulata Zhuk. to bread wheat (Triticum aestivum L). Genet. Resour. CropEvol. 55:849–859.

Biotechnology & Crop Improvement 197
Choi, H.J., T. Chandrasekhar, H.Y. Lee, and K.M. Kim. 2007. Production of herbicide-resistant transgenic sweet potato plants through Agrobacterium tumefaciensmethod. Plant Cell, Tissue and Organ Cult. 91:235–242.
Chopra, H.R., H.S. Dhaliwal, and S.S. Gosal. 1993. Micropropagation: An efficientmethod to multiply strawberries. The Punjab Vegetable Grower 28:9–11.
Christou, P. 1994. In Particle bombardment technology for gene transfer, eds.N.S. Yang and P. Christou, 71–99. Oxford: Oxford Univ Press.
Christy, L.A., S. Arvinth, M. Saravanakumar, M. Kanchana, N. Mukunthan, J. Srikanth,G. Thomas, and N. Subramonian. 2009. Engineering sugarcane cultivars withbovine pancreatic trypsin inhibitor (aprotinin) gene for protection against topborer (Scirpophaga excerptalis Walker) Plant Cell Rep. 28:175–184.
Clements, M.J., K.W. Campbell., C.M. Maragos, C. Pilcher, J.M. Headrick, J.K. Pataky,and D.G. White. 2003. Influence of Cry1Ab protein and hybrid genotype onfumonisin contamination and fusarium ear rot of corn. Crop Sci. 43:1283–1293.
Cober, E.R., S. Rioux, I. Rajcan, P.A. Donaldson, and D.H. Simmonds. 2003. Partialresistance to white mold in a transgenic soybean line. Crop Sci. 43:92–95.
Cong, L., H.C. Zheng. Y.X. Zhang, and T.Y. Chai. 2008. Arabidopsis DREB1A confershigh salinity tolerance and regulates the expression of GA dioxygenases inTobacco. Plant Sci. 174:156–164.
Cristo, E., M.C. Gonzalez, and A.V. Perez. 2006. Obtaining somaclones derived fromrice (Oryza sativa L.) plants through anther culture of hybrids and varieties.Cultivos Tropic. 27:35–39.
Cronauer, S.S, and A.D. Krikorian. 1984. Multiplication of Musa from excised stemtips. Ann. Bot. 53:321–28.
Damasco, O.P., M.K. Smith., I.D. Godwin., S.W. Adkins., R.M. Smillie, andS.E. Hetherington. 1997. Micropropagated dwarf off-type Cavendish bananas(Musa spp., AAA) show improved tolerance to suboptimal temperatures. Aust.J. Agric. Res. 48:377–84.
Daniell, H., S.J. Streatfield, and K. Wycoff. 2001. Medical molecular farming:Production of antibodies, biopharmaceuticals, and edible vaccines in plants.Trends Plant Biol. 6:219–26.
Daniell, H., and A. Dhingra. 2002. Multigene engineering: Dawn of an exciting newera in biotechnology. Curr. Opin. Biotechnol. 13:136–141.
Das, A., S.S. Gosal, J.S. Sidhu, and H.S. Dhaliwal. 2000. Induction of mutationsfor heat tolerance in potato by using in vitro culture and radiation. Euphytica114:205–209.
Datta K., J. Tu, N. Oliva, I. Ona, R Velazhahana, T.W. Mew, S. Muthukrishnand, andS.K. Datta. 2001. Enhanced resistance to sheath blight by constitutive expressionof infection-related rice chitinase in transgenic elite Indica rice cultivars. PlantSci. 160:405–414.
De Block, M., J. Botterman, M. Vandewiele, J. Dockx, C. Thoen, V. Gossele,N.R. Movva, C. Thompson, M.V. Montagu, and J. Leemans. 1987. Engineeringherbicide resistance in plants by expression of a detoxifying enzyme. EMBO. J .6:2513–2518.
De Buyser, J., Y. Henry, P. Lonnet, R. Hertzog, and A. Hespel. 1986. ‘Florin’: Adoubled haploid wheat variety developed by the anther culture method. PlantBreed. 98:53–56.

198 S. S. Gosal et al.
Dear, B.S., G.A. Sandral, D. Spencer, M.R. I. Khan, and T.J.V. Higgins. 2003. Thetolerance of three transgenic subterranean clover (Trifolium subterraneum L.)lines with the bxn gene to herbicides containing brmoxynil. Aust. J. Agric. Res.54:203–210.
DeMaagd, R.A., D. Bosch, and W. Stiekema. 1999. Bacillus thuringiensis toxin-mediated insect resistance in plants. Trends Plant Sci. 4:9–13.
Denis, M., R. Delourme, J.P. Gourret, C. Mariani, and M. Renard.1993. Expressionof engineered nuclear male sterility in Brassica napus: Genetics, morphology,cytology and sensitivity to temperature. Plant Physiol. 101:1295–1304.
Diab, A.A., B. Teulat-Merah, D. This, N.Z. Ozturk, D. Benscher, and M.E. Sorrells.2004. Identification of drought-inducible genes and differentially expressedsequence tags in barley. Theor. Appl. Genet. 109:1417–1425.
Díaz-Pendón, J.A., and S.W. Ding. 2008. Direct and indirect roles of viral suppressorsof RNA silencing in pathogenesis. Ann. Rev. Phytopathol. 46:303–326.
Ding, F., S. Jin, N. Hong, Y. Zhong, Q. Cao, G. Yi, and G. Wang. 2008. Vitrification-cryopreservation, an efficient method for eliminating Candidatus Liberobacterasiaticus, the citrus Huanglongbing pathogen, from in vitro adult shoot tips.Plant Cell Rep. 27:241–250.
Dong, Y.H., L. Wang, J.L. Xu, H.B. Zhang, X.F. Zhang, and L.H. Zhang.2001. Quenching quorum-sensing-dependent bacterial infection by an N-acylhomoserine lactonase. Nature 411:813–817.
Donovan, A.M., R. Morgan, V. Piagnani, M.S. Ridout, D.J. James, and C.M.E. Garrett.1994. Assessment of somaclonal variation in apple. I. Resistance to fire blightpathogen Erwinia amylora. J. Hort. Sci. 69:105–13.
Doule, R.B. 2006. Cane yield and quality characters of some promising somaclonalvariants of sugarcane. Sugar Tech. 8:191–193.
Dunoyer, P., C. Himber, V. Ruiz-Ferrer, A. Alioua, and O. Voinnet. 2007. Intra- andintercellular RNA interference in Arabidopsis thaliana requires componentsof the microRNA and heterochromatic silencing pathways. Nature Genet. 39:848–856.
Ebadi, M., A. Iranbakhsh, and G.B. Khaniki. 2007. Shoot micropropagation andmicrotuberization in potato (Solanum tuberosum L.) by the semi-continuousbioreactor. Pak. J . Biol. Sci. 10:861–867.
Elanchezhian, R., and A.B. Mandal. 2007. Growth analysis of somaclones generatedfrom a salt tolerant traditional ‘Pokkali’ rice (Oryza sativa). Indian J. Agric. Sci.77:184–187.
Epple, P., K. Apel, and H. Bohlmann. 1997. Overexpression of an endogenousthionin enhances resistance of Arabidopsis against Fusarium oxysporum. PlantCell 9: 509–520.
Escobar, M.A., E.L. Civerolo, K.R. Summerfelt, and A.M. Dandekar. 2001. RNAi-mediated oncogene silencing confers ressitance to crown gall tumorigenesis.Proc. Natl. Acad. Sci. 98:13437–13442.
Evans, D.A., and W.R. Sharp. 1986. Somaclonal variation in agriculture.Biotechnology 4:528–532.
Farahani, F., H. Aminpoor, M. Sheidai, Z. Noormohammadi, and M.H. Mazinani.2008. An improved system for in vitro propagation of banana (Musaacuminata L.) cultivars. Asian J. Plant Sci. 7:116–118.

Biotechnology & Crop Improvement 199
Ferreira, S.A., K.Y. Pitz, R. Manshardt, F. Zee, M.M. Fitch, and D. Gonsalves. 2002.Virus coat protein transgenic papaya provides practical control of papayaringspot virus in Hawaii. Plant Disease 86:101–105.
Feuillet, C., S. Travella, N. Stein, L. Albar, A. Nublat, and B. Keller. 2003. Map-basedisolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat(Triticum aestivum L.) genome. Proc. Natl. Acad. Sci. 100:15253–15258.
Fischer, R., N. Emans, F. Schuster, S. Hellwig, and J. Drossard. 1999. Towards molec-ular farming in the future: using plant-cell-suspension cultures as bioreactorsBiotechnol. Appl. Biochem. 30:109–112.
Fischer, R, and N. Emans. 2000. Molecular farming of pharmaceutical proteinsTransgenic Res. 9:279–299.
Fish, N., A. Karp, and M.G.K. Jones. 1988. Production of somatic hybrids byelectrofusion in Solanum. Theor. Appl. Genet. 76:260–266.
Fitzgerald, A., J.A.V. Kha, and K.M. Plummer. 2004. Simultaneous silencing of multi-ple genes in the apple scab fungus Venturia inaequalis, by expression of RNAwith chimeric inverted repeats. Fungal Genet. Biol. 41:963–971.
Florack, D.E.A., and W.J. Stiekema. 1994. Thionins: properties, possible biologicalroles and mechanisms of action Plant Mol. Biol. 26:25–37.
Ford, C.S., N.B. Jones, and J. Van Staden. 2000. Optimization of a working cryop-reservation protocol for Pinus patula embryogenic tissue. In Vitro Cell. Dev.Biol. Plant 36:366–399.
Fuchs, M., and D. Gonsalves. 2007. Safety of virus-resistant transgenic plants twodecades after their introduction: Lessons from realistic field risk assessmentstudies. Ann. Rev. Phytopathology 45:173–202.
Fuentes, A., P.L. Ramos, E. Fiallo, D. Callard, Y. Sanchez, R. Peral, R. Rodríguez,and M. Pujol. 2006. Intron-hairpin RNA derived from replication associatedprotein C1 gene confers immunity to tomato yellow leaf curl virus infection intransgenic tomato plants. Transgenic Res. 15:291–304.
Gao, Y., W. Kongming, G. Fred, and S. Zhicheng. 2009. Cry2Ab tolerance responseof Helicoverpa armigera (Lepidoptera: Noctuidae) populations from Cry1Accotton planting region. J. Econ. Entomol. 102:1217–1223.
García-llamas, C., A. Martín, and J. Ballesteros. 2004. Differences among auxin treat-ments on haploid production in durum wheat × maize crosses. Plant Cell Rep.23:46–49.
Germanà, M.A., B. Chiancone, O. Lain, and R. Testolin. 2005. Anther culture in Citrusclementina: a way to regenerate tri-haploids. Aust. J. Agric. Res. 56:839–845.
Gidding, S.G., G. Allison, D. Brooks, and A. Carter. 2000. Transgenic plants asfactories for biopharmaceuticals. Nature Biotech. 18:1151–1155.
Gill, B.S., L. Huang, V. Kuraparthy, W.J. Raupp, D.L. Wilson, and B. Friebe. 2008.Alien genetic resources for wheat leaf rust resistance, cytogenetic transfer, andmolecular analysis. Aust. J. Agric. Res. 59:197–205.
Gill, B.S., W. Li, S. Sood, V. Kuraparthy, B. Friebe, K.J. Simons, Z. Zhang, andJ.D. Faris. 2007. Genetics and genomics of wheat domestication-driven evolu-tion. Israel J. Plant Sciences 55:223–229.
Gill, R.I.S., S.S. Gill, and S.S. Gosal. 1994. Vegetative propagation of eucalyptustereticornis Sm. through tissue culture. Plant Tissue Culture 4:59–67.

200 S. S. Gosal et al.
Gill, S.S., R.I.S. Gill, and S.S. Gosal. 1997. Rapid propagation of Dalbergia sisso frommature trees through tissue culture. Plant Tissue Culture 7:13–19.
Gill, R., N. Kaur, A.S. Sindhu, T.S. Bharaj, and S.S. Gosal. 2003. In Advances in RiceGenetics, eds. G.S. Khush, D.S. Brar, and B. Hardy, 503–505. Manila, Phillipines:Internatinal Rice Research Publications.
Gill, N.K., R. Gill, and S.S. Gosal. 2004. Factors enhancing somatic embryogene-sis and plant regeneration in sugarcane (Saccharum officinarum L.). IndianJ. Biotechnol. 3:119–123.
Gill, R., P.K. Gupta, and S.S. Gosal. 2004. Brahmi: Efficient protocol for microprop-agation. Indian Medicinal and Aromatic Plants Today. May/June:3–5.
Gill, M.K., S.K. Chauhan, and S.S. Gosal. 2006. Macro-and Micro-propagation ofAzadirachta indica. Indian Forester 132:1159–1168.
Gill, R., P.K. Malhotra, and S.S. Gosal. 2006. Direct plant regeneration from culturedyoung leaf segments of sugarcane. Plant Cell, Tissue Organ Culture 84:227–31.
Gitlin, L., S. Karelsky, and R. Andino. 2002. Short interference confers intracellularantiviral immunity in human cells. Nature 4:418–430.
Goldoni, M., G. Azzalin, G. Macino, and C. Cogoni. 2004. Efficient gene silencing byexpression of double stranded RNA in Neurospora crassa. Fungal Genet. Biol.41:1016–1024.
Gopal, J., J.L. Minocha, and S.S. Gosal. 1998. Variability in responsive potatogenotypes to in vitro propagation. J. Indian Potato Assoc. 25:119–124.
Gordon-Kamm, W.J., T.M. Spencer, M.L. Mangano, T.R. Adams, R.J. Daines,W.G. Start, J.V. O’Brien, S.A. Chambers, W.R. Adams, Jr., N.G. Willetts,T.B. Rice, C.J. Mackey, R.W. Krueger, A.P. Kausch, and P.G. Lemaux. 1990.Transformation of maize cells and regeneration of fertile transgenic plants. PlantCell. 2:603–618.
Gosal, S.S., and Y.P.S. Bajaj. 1983. Interspecific hybridization between Vigna mungoand Vigna radiata through embryo culture. Euphytica 32:129–137.
Gosal, S.S., A.S. Sindhu, J.S. Sandhu, R. Gill, G.S. Khehra, G.S. Sidhu, and H.S.Dhaliwal. 1996. Haploidy in rice. In In vitro Haploid Production in HigherPlants, eds. S.M. Jain S.K. Sopory and R.E. Veilleux, 1–37. Netherlands: KluwerAcademic.
Gosal, S.S., K.S. Thind, and H.S. Dhaliwal. 1998. Micropropagation of sugarcane: Anefficient protocol for commercial plant production. Crop Improv. 25:1–5.
Gosal, S.S, and S.K. Gosal. 2000. Genetic transformation and production of trans-genic plants. In Plant biotechnology: Recent advances, Ed. P.C. Trivedi, 29–40.New Delhi: Panima Publishers.
Gosal, S.S., R. Gill, A.S. Sindhu, H.S. Dhaliwal, and P. Christou. 2003. Introducingthe Cry1A(c) gene into basmati rice and transmitting transgenes to R3 progeny.In Advances in rice genetics, eds. G.S. Khush, D.S. Brar, and B. Hardy, 558–560.Proc. Fourth Rice Genetics Symposium, 22-27 October 2000. Phillipines: IRRI.
Gosal, S.S., and R. Gill. 2004. Different biotechnological options for improv-ing medicinal and aromatic plants. Indian Med. Aromat. Plant Today.May/June:19–21.
Gosal, S.S., S.H. Wani, and M.S. Kang. 2009. Biotechnology and drought tolerance.J. Crop Improv. 23:19–54.

Biotechnology & Crop Improvement 201
Grewal, D.K., R. Gill, and S.S. Gosal. 2006a. Genetic engineering of Oryza sativa byparticle bombardment. Biol. Plant. 50:311–314.
Grewal, D.K., R. Gill, and S.S. Gosal. 2006b. Role of cysteine in enhancing andro-genesis and regeneration of indica rice (Oryza sativa L.). Plant Growth Regul.49:43–47.
Guha, S., and S.C. Maheshwari. 1964. In vitro production of embryos from anthersof Datura. Nature 204:497.
Gupta, S., and B.M. Reed. 2006. Cryopreservation of shoot tips of blackberry andraspberry by encapsulation-dehydration and vitrification. Cryo Letters 27:29–42.
Guzmán, M., and F.J. Zapata-Arias. 2000. Increasing anther culture efficiency in rice(Oryza sativa L.) using anthers from ratooned plants. Plant Sci. 151:107–114.
Hain, R., H.J. Reif, E. Krause, R. Langebartels, H. Kindl, B. Vornam, W. Wiese,E. Schmelzer, P. Schreier., R. Stöcker, and K. Stenzel. 1993. Disease resis-tance results from foreign phytoalexin expression in a novel plant. Nature 361:153–156.
Halmagyi, A., C. Deliu, A. Coste, M. Keul, O. Cheregi, and V. Cristea. 2004.Vitrification of potato shoot tips for germplasm cryopreservation. ContributiiBotanice. 39:187–193.
Hamada, W., and P.D. Spanu. 1998. Co-suppression of the hydrophobin gene Hcf-1is correlated with antisense RNA biosynthesis in Cladosporium fulvum. Mol.Gen. Genet. 259:630–638.
Hammerschlag., F.A, and V. Ognjanov. 1990. Somaclonal variation in peach: screen-ing for resistance to Xanthomonas camperstris pv. pruni and Pseudomonassyringae pv. syringae. Acta Hort. 280:403–408.
Hammond, B.G., K.W. Campbell, C.D. Pilcher, T.A. Degooyer, A.E. Robinson,B.L. McMillen, S.M. Spangler, S.G. Riordan, L.G. Rice, and J.L. Richard. 2004.Lower fumonisin mycotoxin levels in the grain of Bt corn grown in the UnitedStates in 2000–2002. J Agric. Food Chem. 52:1390–1397.
Hammond, T.M., and N.P. Keller. 2005. RNA silencing in Aspergillus nidulans isindependent of RNA-dependent RNA polymerase. Genetics 169:607–617.
Hansen, L.N., and E.D. Earle. 1995. Transfer of resistance to Xanthomonascampestris pv campestris into Brassica oleracea L. by protoplast fusion. Theor.Appl. Genet. 9:1293–1300.
Hao, Y., A. Liu, Y. Wang, D. Feng, J. Gao, X. Li, S. Liu, and H. Wang. 2008. Pm23:A new allele of Pm4 located on chromosome 2AL in wheat. Theor. Appl. Genet.117:1205–1212.
Havkin-Frenkel, D., R. Dorn, and T. Leustek. 1997. Plant tissue culture for productionof secondary metabolites. Food Technol. 51:56–61.
He, R., Z. Chang, Z. Yang, Z. Yuan, H. Zhan, X. Zhang, and J. Liu. 2009. Inheritanceand mapping of powdery mildew resistance gene Pm43 introgressed fromThinopyrum intermedium into wheat. Theor. Appl. Genet. 118:1173–1180.
Herbers, K. 2003. Vitamin production in transgenic plants. Plant Physiol. 160:821–829.
Hiebert, C.W., J.B. Thomas, B.D. McCallum, and D.J. Somers. 2008. Genetic mappingof the wheat leaf rust resistance gene Lr60 (LrW2). Crop Sci. 48:1020–1026.
Hinnisdaels, S., I. Negrutiv, M. Jacobs, and V. Sidorov. 1988. Plant somatic cellhybridizations: evaluations and perspectives. IAPTC Newsletter 55:2–10.

202 S. S. Gosal et al.
Hoisington, D., G.M. Listman, and M.L. Morris. 1998. Varietal development: Appliedbiotechnology. In Maize seed industries in developing countries, ed M.L. Morris,77–102. Boulder, CO: Lynne Rienner Publishers/CIMMYT.
Holmstrom, K.O., S. Somersalo, A. Mandal, T.E. Palva, and W. Bjorn. 2000. Improvedtolerance to salinity and low temperature in transgenic tobacco producingglycinebetaine. J. Exp. Bot. 51:177–85.
Hong, B.S., L. Zong-Suo, and S. Ming-An. 2005. LEA proteins in higher plants:Structure, function, gene expression and regulation. Colloids Surf. B: Biointerf .45:131–135.
Hrazdina, G., E. Kirs, C. Rosenfield, J.L.Norelli, and H.S. Aldwinckle. 2000. Downregulation of ethylene production in apples. Use of agriculturally impor-tant genes in biotechnology. Proceedings of the NATO-Advanced Research-Workshop, Szeged, Hungary, 17-21 Oct, 2000, 26-31.
Hsieh, T.H., J.T. Lee, Y.Y. Charng, and M.T. Chan. 2002. Tomato plants ectopicallyexpressing Arabidopsis CBF1 show enhanced resistance to water deficit stress.Plant Physiol. 130:618–626.
Hu, H., M. Dai, J. Yao, B. Xiao, X. Li, Q. Zhang, and L. Xiong. 2006. Overexpressinga NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistanceand salt tolerance in rice. Proc. Natl. Acad. Sci. 103:12987–12992.
Huang, X.Q., S.L.K. Hsam, F.J.Zeller, G. Wenzel, and V. Mohler. 2000. Molecularmapping of wheat powdery mildew resistance gene Pm24 and markervalidation for molecular breeding. Theor. Appl. Genet. 10:407–414.
Huang, X.Q., L.X. Wang, M.X. Xu, and M.S. Röder. 2003. Microsatellite mappingof the powdery mildew resistance gene Pm5e in common wheat (Triticumaestivum L.). Theor. Appl. Genet. 106:858–865.
Huang, X.Q., S.L.K. Hsam, V. Mohler, M.S. Röder, and F. Zeller. 2004. Genetic map-ping of three alleles at the Pm3 locus conferring powdery mildew resistance incommon wheat (Triticum aestivum L.). Genome 47:1130–1136.
Huang, J.K., R.F. Hu, S. Rozelle, and C. Pray. 2005. Insect-resistant GM ricein farmers’ fields: Assessing productivity and health effects in China. Sci.308:688–690.
Huang, H.E., C.A. Liu, M.J. Lee, C.G. Kuo, H.M. Chen, M.J. Ger, Y.C. Tsai,Y.R. Chen, M.K. Lin, and T.Y. Feng. 2007. Resistance enhancement of trans-genic tomato to bacterial pathogens by the heterologous expression of sweetpepper ferredoxin-I protein. Phytopathology 97:900–906.
Huang, C.L., S.Y. Hwang, Y.C. Chiang, and T.P. Lin. 2008. Molecular evolution ofthe Pi-ta gene resistant to rice blast in wild rice (Oryza rufipogon). Genetics179:1527–1538.
Hwang, S.C., and W.H. Ko. 1992. Breeding for resistance to Fusarium wiltof Cavendish banana by using tissue culture method. Special Publication-Taichung District Agricultural Improvement Station 29:229–237.
Hwang, S.C., and W.H. Ko. 2004. Cavendish banana cultivars resistant to Fusariumwilt acquired through somaclonal variation in Taiwan. Plant Disease. 88:580–588.
Igawa, T., N. Takahashi-Ando, N. Ochiai, S. Ohsato, T. Shimizu, T. Kudo,I. Yamaguchi, and M. Kimura. 2007. Reduced contamination by the Fusarium

Biotechnology & Crop Improvement 203
mycotoxin zearalenone in maize kernels through genetic modification with adetoxification gene. Appl. Environ. Microbiol. 73:1622–1629.
Inoue, K., T. Kato, A. Nobukunib, H. Kunitake, and T. Yabuya. 2006.Characterization of tetraploid plants regenerated via protoplast culture of Irisfulva and their crossability with Japanese irises Scientia Hort. 110:334–335.
Jacque, J.M., K. Triques, and M. Stevenson. 2002. Modulation of HIV-1 replicationby RNA interference. Nature 418:435–438.
Jafari, M., P. Norouzi, M.A. Malboobi, B. Ghareyazie, M. Valizadeh, S.A. Mohammadi,and M. Mousavi. 2009. Enhanced resistance to a lepidopteran pest in transgenicsugar beet plants expressing synthetic cry1Ab gene. Euphytica 165:333–334.
Jain, R.K., G.S. Khehra, S.H. Lee, N.W. Blackhall, R. Merchant, M.R. Davey,J.B. Power, E.C. Cocking, and S.S. Gosal. 1995. An improved procedure forplant regeneration from indica and Japonica rice protoplasts. Plant Cell Rep.14:515–519.
Jalaja, N.C., T.V. Sreenivasan, S.M. Pawar, P.G. Bhoi, and R.M. Garker. 2006. Co94012: A new sugarcane variety through somaclonal variation. Sugar Tech.8:132–136.
Jalaja, N.C., D. Neelamathi, and T.V. Sreenivasan. 2008. Micropropagation for qual-ity seed production in sugarcane in Asia and the Pacific. Food and AgricultureOrganization of the United Nations, Rome; Asia–Pacific Consortium onAgricultural Biotechnology, New Delhi; Asia-Pacific Association of AgriculturalResearch Institutions, Bangkok, i-x, 46.
James, A.C., S. Peraza-Echeverria, L. Peraza-Echeverria, and V.A. Herrera-Valenci.2007. Variation in micropropagated plants. Acta Hort. 748:55–63.
James, C. 2009. Global status of commercialized biotech/GM crops: ISAAA Brief No.41. Ithaca, New York: ISAAA.
Jansens, S., A. Vliet, C. Dickburt, L. Buysse, C. Piens, B. Saey, A.D. Wulf, andV. Gosselé. 1997. Transgenic corn expressing a Cry9C insecticidal protein fromBacillus thuringiensis protected from European corn borer damage. Crop Sci.37:1616.
Jaskani, M.J., H. Abbas, R. Sultana, M.M. Khan, M. Qasim, and I.A. Khan. 2008. Effectof growth hormones on micropropagation of Vitis vinifera L. cv. Perlette. Pak.J. Bot. 40:105–109.
Jeung, J.U., B.R. Kim, Y.C. Cho, S.S. Han, H.P. Moon, Y.T. Lee, and K.K. Jena.2007. A novel gene, Pi40(t), linked to the DNA markers derived from NBS-LRRmotifs confers broad spectrum of blast resistance in rice. Theor. Appl. Genet.115:1163–1177.
Jiang, G.H., Y.Q. He, C.G. Xu, X.H. Li, and Q. Zhang. 2004. The genetic basis ofstay-green in rice analyzed in a population of doubled haploid lines derivedfrom an Indica X Japonica cross. Theor. Appl. Genet. 108(4): 688–699.
Jongsma, M.A., and C.J. Boulter.1997. The adaptation of insects to plant proteaseinhibitors. J. Insect Physiol. 43:885–895.
Joshi, S.P., P.K. Ranjekar, and V.S. Gupta. 1999. Molecular markers in plant genomeanalysis. Curr. Sci. 77:230–240.
Juenger, T.E., J.K. Mckay, N. Hausmann, J.J.B. Keurentjes, S. Sen, K.A. Stowe,T.E. Dawson, E.L. Simms, and J.H. Richards. 2005. Identification and charac-terization of QTL underlying whole plant physiology in Arabidopsis thaliana:

204 S. S. Gosal et al.
d13C, stomatal conductance and transpiration efficiency. Plant Cell Environ.28:697–708.
Jung, C., J.S. Seo, S.W. Han, Y.J. Koo, C.H. Kim, S.I. Song, B.H. Nahm, Y.D. Choi,and J.J. Cheong. 2008. Overexpression of AtMYB44 enhances stomatal closureto confer abiotic stress tolerance in transgenic Arabidopsis. Plant Physiol. 146:623–635.
Kadotani, N., H. Nakayashiki, Y. Tosa, and S. Mayama. 2003. RNA silencing inthe pathogenic fungus Magnaporthe oryzae. Mol. Plant-Microbe Interaction16:769–776.
Kalimuthu, K., M. Saravanakumar, and R. Senthikumar. 2007. In vitro micropropa-gation of Musa sapientum L. (Cavendish Dwarf). Afr. J. Biotech. 6:1106–09.
Kalpana, K., S. Maruthasalam, T. Rajesh, K. Poovannan, K.K. Kumar, E. Kokiladevi,J.A.J. Raja, D. Sudhakar, R. Velazhahan, R. Samiyappan, andP. Balasubramanian. 2006. Engineering sheath blight resistance in elite Indicarice cultivars using genes encoding defense proteins. Plant Sci. 170:203–215.
Kang, M.S., P.K. Subudhi, N. Baisakh, and P.M. Priyadarshan. 2007. Crop breed-ing methodologies: Classic and modern. In Breeding major food staples, eds.M.S. Kang and P.M. Priyadarshan, 5–40. Hoboken, NJ: Blackwell.
Karp, A .1995. Somaclonal variation as a tool for crop improvement. Euphytica85:295–302.
Katiyar-Agarwal, P., P. Agarwal, M.K. Reddy, and S.K. Sopory. 2006a. Role of DREBtranscription factors in abiotic and biotic stress tolerance in plants. Plant CellRep. 25:1263–1274.
Katiyar-Agarwal, S., R. Morgan, D. Dahlbeck, O. Borsani, J.A. Villegas, J. Zhu,B.J. Staskawicz, and H. Jin. 2006b. A pathogen-inducible endogenous siRNAin plant immunity. Proc. Natl. Acad. Sci. 103:47–52.
Katiyar-Agarwal, S., S. Gao, A. Vivian-Smith, and H. Jin. 2007. A novel class ofbacteria-induced small RNAs in Arabidopsis. Genes Develop. 21:3123–3134.
Kato, Y., S. Hirotsu, K. Nemoto, and J. Yamagishi. 2008. Identification of QTLcontrolling rice drought tolerance at seedling stage in hydroponic culture.Euphytica 160:423–430.
Kaur, D., R. Gill, A.S. Sindhu, and S.S. Gosal. 1999. Protoplast culture and plantregeneration in basmati rice. Crop Improv. 26:14–18.
Kaur, A., S.S. Gosal, R. Gill, and K.S. Thind. 2001. Induction of plant regenerationand somaclonal variation for some agronomic traits in sugarcane (Saccharumofficinarum L.). Crop Improv. 28:167–172.
Kaur, J., C.K. Satija, and S.S. Gosal. 2002. In vitro synthesis of white grained primaryhexaploid triticales. Plant Tissue Cult. 12:1–9.
Kawahigashi, H., S. Hirose, H. Ohkawa, and Y. Ohkawa. 2008. Transgenic riceplants expressing human P450 genes involved in xenobiotic metabolism forphytoremediation. J. Mol. Microbiol. Biotechnol. 15:212–219.
Kawchuk, L.M., J. Hachey, D.R. Lynch, F. Kulcsar, G. Rooijen, D.R. Waterer,A. Robertson, E. Kokko, R. Byers, R.J. Howard, R. Fischer, and D. Prufer. 2001.Tomato Ve disease resistance genes encode cell surface-like receptors. Proc.Natl. Acad. Sci. 98:6511–6515.
Khan, S.J., M.A. Khan, H.K. Ahmad, R.D. Khan, and YUSUF-Zafar. 2004. Somaclonalvariation in sugarcane through tissue culture and subsequent screening for salttolerance. Asian J. Pl. Sci. 3:330–334.

Biotechnology & Crop Improvement 205
Khush, G.S., and P.S. Virk. 2002. Rice improvement: Past, present and future. In Cropimproment challenges in the twenty-first century, ed. MANJIT S. Kang, 17–42.New York: Haworth Press.
Kishore, P.B.K., Z. Hong, G.H. Miao, C.A.A. Hu, and D.P.S. Verma. 1995.Overexpressionof �-1 pyrroline-5-carboxylate synthetase increases proline pro-duction and confers osmotolerance in transgenic plants. Plant Physiol. 108:1387–1394.
Kodym, A., and F.J. Zapata-Arias. 1999. Natural light as an alternative light sourcefor the in vitro culture of banana (Musa acuminata cv. ‘Grande Naine’). PlantCell, Tissue and Organ Cult. 55:141–45.
Kumar, S.M., B.K. Kumar, K.K. Sharma, and P. Devi. 2004. Genetic transformationof pigeonpea with rice chitinase gene. Plant Breed. 123:485–489.
Kuraparthy, V., P. Chhuneja, H.S. Dhaliwal, Satinder Kaur, R.L. Bowden, and B.S.Gill. 2007a. Characterization and mapping of cryptic alien introgression fromAegilops geniculata with novel leaf rust and stripe rust resistance genes Lr57and Yr40 in wheat. Theor. Appl. Genet. 114:1379–1389.
Kuraparthy, V., S. Sood, P. Chhuneja, H.S. Dhaliwal, and B.S.Gill. 2007b.Identification and mapping of tiller inhibition gene (tin3) in wheat. Theor. Appl.Genet. 114:285–294.
Labbani, Z., J. de Buyser, and E. Picard. 2007. Effect of mannitol pretreatment toimprove green plant regeneration on isolated microspore culture in Triticumturgidum ssp. durum cv. ‘Jennah Khetifa.’ Plant Breed. 126:565–568.
Lafitte, H.R., A.H. Price, and B. Courtois. 2004. Yield response to water deficit in anupland rice mapping population: associations among traits and genetic markers.Theor. Appl. Genet. 109:1237–1246.
Lagudah, E.S., S.G. Krattinger, S. Herrera-Foessel, R.P. Singh, J. Huerta-Espino,W. Spielmeyer, G. Brown-Guedira, L.L. Selter, and B. Keller. 2009. Gene-specifcmarkers for the wheat gene Lr34/Yr18/Pm38, which confers resistance tomultiple fungal pathogens. Theor. Appl. Genet. 119:889–898.
Lal, M., R.K. Singh, S. Srivastava, N. Singh, S.P. Singh, and M.L. Sharma. 2008.RAPD marker based analysis of micropropagated plantlets of sugarcane forearly evaluation of genetic fidelity. Sugar Tech. 10:99–103.
Lanceras, J.C., G.P. Pantuwan, B. Jongdee, and T. Toojinda. 2004. Quantitativetrait loci associated with drought tolerance at reproductive stage in rice. PlantPhysiol. 135:1–16.
Lara, R.M., M. lorido, D. Plana, O. More, M.E. Gonzalez, M. Alvarez, andM.M. Hernandez. 2003. Isoenzymatic analysis for detecting in vitro variabilityand/or stability of economically important crops. Cultivos-Tropic. 24:39–47.
Larkin, P.J., Y. Li, L.H. Spindler, G.J. Tanner and P.M. Banks. 1993. Disease resistance,cell culture and somatic recombination. Acta Hort. 336:341–346.
Laurie, D.A., and S. Reymondie. 1991. High frequencies of fertilization and haploidseedling production in crosses between commercial hexaploid wheat varietiesand maize. Plant Breed. 106:182–189.
Leal, M.R., R.H. Maribona, A. Ruiz, S. Korneva, E. Canales, T.D. Dinkova,F. Izquierdo, O. Goto, and D. Rizo. 1994. Somaclonal variation as a sourceof resistance to eyespot disease of sugarcane. Plant Breed. 115:37–42.

206 S. S. Gosal et al.
Leckband, G., and H. Lorz. 1998. Transformation and expression of a stilbenesynthase gene of Vitis vinifera L. in barley and wheat for increased fungalresistance. Theor. Appl. Genet. 96:1004–1012.
Lee, H.I., and N.V. Raikel. 1995 Prohevein is poorly processed but shows enhancedresistance to a chitin-binding fungus in transgenic tomato plants. Braz. J. Med.Biol. Res. 28:743–750.
Lee, K.W., N. Ahsan, S.H. Lee, D.G. Lee, K.H. Kim, I. Alam, S.Y. Kwon, J.S. Kim,K. Back, S.S. Lee, and B.H. Lee. 2008. Responses of MxPPO overexpressingtransgenic tall fescue plants to two diphenyl-ether herbicides, oxyfluorfen andacifluorfen. Acta Physiol. Plant. 30:745–754.
Leva, A.R., and R. Petruccelli. 2007. Field performance of olive somaclones derivedfrom somatic embryos. Acta Horti. 748:181–189.
Li, F., and S.W. Ding. 2006. Virus counter defense: diverse strategies for evading theRNA silencing immunity. Ann. Rev. Microbiol. 60:503–531.
Li, W., C. Lei, Z. Cheng, Y. Jia., D. Huang, J. Wang, J. Wang, X. Zhang, N. Su,X. Guo, H. Zhai, and J. Wan. 2008. Identification of SSR markers for a broad-spectrum blast resistance gene Pi20(t) for marker-assisted breeding. Mol. Breed.22:141–149.
Li, Y., Y.C. Niu, and X.M. Chen. 2009. Mapping a stripe rust resistance gene YrC591in wheat variety C591 with SSR and AFLP markers. Theor. Appl. Genet. 118:339–346.
Lian, Y., Z.W. Jia, K.L. He, Y.J. Liu, F.P. Song, B.M. Wang, and G.Y. Wang. 2008.Transgenic tobacco plants expressing synthetic Cry1Ac and Cry1Ie genes aremore toxic to cotton bollworm than those containing one gene. Chin. Sci.Bulletin. 53:1381–1387.
Liang, H., C.A. Maynard, R.D. Allen, and W.A. Powell. 2004. Increased Septoriamusiva resistance in transgenic hybrid poplar leaves expressing a wheat oxalateoxidase gene. Plant Mol. Biol. 45:619–629.
Lin, W., C.S. Anuratha, and K. Datta. 1995. Genetic engineering of rice for resistanceto sheath blight. Biotechnol. 13:686–691.
Liu, F., U. Ryschka, F. Marthe, E. Klocke, G. Schumann, and H. Zhao. 2007a. Cultureand fusion of pollen protoplasts of Brassica oleracea L. var. italica with haploidmesophyll protoplasts of B. rapa L. ssp. pekinensis. Protoplasma 231:89–97.
Liu, X., F. Lin, L. Wang, and Q. Pan. 2007b. The in silico map-based cloning of Pi36,a rice CC-NBS-LRR 1 gene which confers race-specific resistance to the blastfungus. Genetics 176:2541–2549.
Liu, Z., and S. Gao. 2007. Micropropagation and induction of autotetraploid plantsof Chrysanthemum cinerariifolium (Trev.) Vis. In Vitro Cell. Dev. Biol. Plant43:404–408.
Luo, Z., and W. Kearsey. 1992. Interval mapping of quantitative trait loci in an F2
population. Heredity 69:236–242.Machado, M.P., L.A. Biasi, M. Ritter, L.L.F. Ribas, H.S. Koehler, and F. Zanette. 2007.
Culture media in micropropagation of grapevine rootstock ‘VR043-43’ (Vitisvinifera x Vitis rotundifolia). Ciencia-Rural. 37:277–280.
Maheswaran, G., L. Pridmore, P. Franz, and M.A. Anderson. 2007. A proteinaseinhibitor from Nicotiana alata inhibits the normal development of light-brown

Biotechnology & Crop Improvement 207
apple moth, Epiphyas postvittana in transgenic apple plants. Plant Cell Rep.26:773–782.
Makandar, R., J.S. Essig, M.A. Schapaugh, H.N. Trick, and J. Shah. 2006. Geneticallyengineered resistance to Fusarium head blight in wheat by expression ofArabidopsis NPR1. Mol. Plant Microbe Interact. 19:123–129.
Maliga, P. 2003. Progress towards commercialization of plastid transformationtechnology. Trends Biotechnol. 21:20–28.
Maqbool, S.B., and P. Christou. 1999. Multiple traits of agronomic importancein transgenic Indica rice plants: analysis of transgene integration patterns,expression levels and stability. Plant Mol. Breed. 5:471–480.
Masoud, S.A., Q. Zhu, C. Lamb, and R.A. Dixon. 1996. Constitutive expression ofan inducible β-1,3-Glucanase in alfalfa reduces disease severity caused by theoomycete pathogen Phytophthora megasperma f. sp. medicaginis, but does notreduce severity of chitin- containing fungi. Transgenic Res. 5:313–323.
Masoud, T., M. Mojtaba, and G. Behzad. 2005. Agrobacterium -mediated transfor-mation of cotton (Gossypium hirsutum) using a heterologous bean chitinasegene. Plant Cell, Tissue Organ Cult. 83:83–96.
Matsumoto, K., A.D. Vilarinhos, and S. Oka. 2002. Somatic hybridization byelectrofusion of banana protoplasts. Euphytica 125:317–324.
McCafferty, H.R.K., P.H. Moore, and Y.J. Zhu. 2008. Papaya transformed with theGalanthus nivalis GNA gene produces a biologically active lectin with spidermite control activity. Plant Sci.175:385–393.
Medina, R., M. Faloci, M.A. Marassi, and L.A. Mroginski. 2004. Genetic stability inrice micropropagation. Bio Cell 28:13–20.
Mehlenbacher, S.A. 1995. Classical and molecular approaches to breeding fruit andnut crops for disease resistance. Hort Science 30:466–477.
Mendes, B.M.J., F.A. Mourão Filho, P.M. Farias, and V.A. Benedito. 2001. Citrussomatic hybridization with potential for improved blight and CTV resistance. InVitro Cell. Dev. Biol. Plant 37:490–495.
Mentag, R., M. Luckevich., M.J. Morency, and A. Seguin. 2003. Bacterial diseaseresistance of transgenic hybrid poplar expressing the synthetic antimicrobialpeptide D4E1. Tree Physiol. 23:405–411.
Miranda, L.M., J.P. Murphy, S. Leath, and D. Marshall. 2006. Pm34: A new powderymildew resistance gene transferred from Aegilops tauschii Coss. to commonwheat (Triticum aestivum L.). Theor. Appl. Genet. 113:1497–1504.
Miranda, J.A., N. Avonce, R. Suárez, J.M. Thevelein, P.V. Dijck, and G. Iturriaga.2007a. A bifunctional TPS-TPP enzyme from yeast confers tolerance to mul-tiple and extreme abiotic-stress conditions in transgenic Arabidopsis. Planta226:1411–1421.
Miranda L.M., J.P. Murphy, D. Marshall, C. Cowger, and S. Leath. 2007b.Chromosomal location of Pm35, a novel Aegilops tauschii derived powderymildew resistance gene introgressed into common wheat (Triticum aestivumL.). Theor. Appl. Genet. 114:1451–1456.
Miranda, L.M., L. Peruginia, G. Srni, G.B. Guedira, D. Marshall, S. Leath, andJ.P. Murphy. 2007c. Genetic mapping of a Triticum monococcum-derivedpowdery mildew resistance gene in common wheat. Crop Sci. 47:2323–2329.
Mohamed, A.E. 2007. Somaclonal variation in micro-propagated strawberry detectedat the molecular level. Inter. J. Agricul. Biol. 9:721–725.

208 S. S. Gosal et al.
Molinari, H.B.C., C.J. Marura, E. Darosb, M.K. Freitas de Camposa, J.F.R. Portela deCarvalhoa, J.C.B. Filhob, L.F.P. Pereirac, and L.G.E. Vieira. 2007. Evaluation ofthe stress-inducible production of proline in transgenic sugarcane (Saccharumspp.) osmotic adjustment, chlorophyll fluorescence and oxidative stress.Physiol. Planta. 130:218–229.
Moreau, L., A. Charcosset, and A. Gallais. 2004. Use of trial clustering to study QTL3environment effects for grain yield and related traits in maize. Theor. Appl.Genet. 110:92–105.
Moukadiri, O., and M.J. Cornejo. 1996. Freezing stress response in progenies of riceplants regenerated from cryopreserved cells. Biotechnologia Aplicada 13:31.
Muehlbauer, G.J., and W.R. Bushnell. 2003. Transgenic approaches to resistance.In Fusarium head blight of wheat and barley, eds. K.J. Leonard and W.R.Bushnell,. St. Paul, MN: American Phytopathological Society Press.
Mujeeb-Kazi, A., A. Gul, J. Ahmed, and J.I. Mirza. 2006. A simplified and effectiveprotocol for production of bread wheat haploids (n = 3x = 21, ABD) withsome application areas in wheat improvement. Pak. J . Bot. 38:393–406.
Nakajima, H., T. Muranaka, and F. Ishige. 1994. Fungal and bacterial diseaseresistance in transgenic plants expressing human lysozyme. Plant Cell Rep.16:674–679.
Nakayashiki, H., S. Hanada, B.Q. Nguyen, N. Kadotani, Y. Tosa, and S. Mayama.2005. RNA silencing as a tool for exploring gene function in ascomycete fungi.Fungal Genet. Biol. 42:275–283.
Navarro, L., P. Dunoyer, F. Jay, B. Arnold, N. Dharmasini, M. Estelle, O.Vionnet,and J.D. Jones. 2006. A plant miRNA contributes to antibacterial resistance byrepressing auxin signalling. Sci. 312:436–439.
Nayak, P., D. Basu, S. Das, A. Basu, D. Ghosh, N.A. Ramakrishnan, M. Ghosh, andS.K. Sen. 1997. Transgenic elite indica rice plants expressing CryIAc ∂-endotoxinof Bacillus thuringiensis are resistant against yellow stem borer (Scirpophagaincertulas) Proc. Natl. Acad. Sci. 94:2111–2116.
Neeraja, C.N., R. Maghirang-Rodriguez, A. Pamplona, S. Heuer, B.C.Y. Collard, E.M.Septiningsih, and G. Vergara et al. 2007. A marker-assisted backcross approachfor developing submergence-tolerant rice cultivars. Theor. Appl. Genet. 115:767–776.
Newell-McGloughlin, M. 2008. Nutritionally improved agricultural crops. PlantPhysiol. 147:939–953.
Ninkovic, S., J. Miljuš-Dukic, S. Radovic, V. Maksimovic, J. Lazarevic, B. Vinterhalter,M. Neškovic, and A. Smigocki. 2007. Phytodecta fornicata Brüggemann resis-tance mediated by Oryzacystatin II proteinase inhibitor transgene. Plant Cell,Tissue Organ Cult. 91:289–294.
Nishizawa, Y.Z., K. Nishio, M. Nakazono, E. Soma, M.U. Nakajima and T. Hibi. 1999.Enhanced resistance to blast (Magnaporthe grisea) in transgenic Japonica riceby constitutive expression of rice chitinase. Theor. Appl. Genet. 99:383–390.
Niu, Q.W., S.S. Lin, J.L. Reyes, K.C. Chen, H.W. Hu, S.D. Yeh, and N.H. Chua. 2006.Expression of artificial micro- RNA in transgenic Arabidopsis thaliana confersvirus resistance. Nat. Biotech. 24:1420–1428.
Nouri-Ellouz, O., R. Gargouri-Bouzid, D. Sihachakr, M.A. Triki, G. Ducreux,N. Driraf, and L. Lakhoua. 2006. Production of potato intraspecific somatic

Biotechnology & Crop Improvement 209
hybrids with improved tolerance to PVY and Pythium aphanidermatum. J.Plant Physiol.163:1321–1332.
Novina, C.D., M.F. Murray, D.M. Dykxhoorn, P.J. Beresford, J. Riess, S.K. Lee,R.G. Collman, J. Lieberman, P. Shanker, and P.A. Sharp. 2002. siRNA-directedinhibition of HIV-1 infection. Nature Mediterranean 8:681–686.
Obert, B., and B. Barnabas. 2004. Colchicine induced embryogenesis in maize. PlantCell Tissue Organ Cult. 77:283–285.
Oh, S.J., S.I. Song, Y.C. Kim, H.J. Jang, S.Y. Kim, M. Kim, Y.K. Kim, B.H. Nahm,and J.K. Kim. 2005. Arabidopsis CBF3/DREB1A and ABF3 in transgenic riceincreased tolerance to abiotic stress without stunting growth. Plant Physiol.138:341–351.
Oh, S.J., C.W. Kwon, D.W. Choi, S.I. Song and J.K. Kim. 2007a. Expression of barleyHvCBF4 enhances tolerance to abiotic stress in transgenic rice. Plant Biotech.J. 5:646–656.
Oh, T.J., M.A. Cullis, K. Kunert, I. Engelborghs, R. Swennen, and C.A. Cullis. 2007b.Genomic changes associated with somaclonal variation in banana (Musa spp.)Physiol. Plant. 129:766–774.
Oldach, K.H., D. Becker, and H. Lorz. 2001. Heterologous expression of genesmediating enhanced fungal resistance in transgenic wheat. Mol. Plant MicrobeInteract. 14:832–838.
Ozturk, G., and Z. Yldrm. 2006. The effect of various nutrient media on microtuberproduction in potato (Solanum tuberosum L.) under in vitro conditions. Turk.J. Field Crops 11(2): 38–45.
Panis, B., R. Swennen, and F. Engelmann. 2001. Cryopreservation of plantgermplasm Acta Horti. 560:79–86.
Paran, I., and R.W. Michelmore. 1993. Development of reliable PCR-based markerslinked to downy mildew resistance genes in lettuce. Theor. Appl. Genet. 85:985–993.
Paterson, A.H., S.D. Tanksley, and M.E. Sorrels. 1991. DNA markers in plantimprovement. Adv. Agron. 46:39–90.
Pellegrineschi, A., M. Reynolds, M. Pacheco, R.M. Brito, R. Almeraya, K. Yamaguchi-Shinozaki, and D. Hoisington. 2004. Stress-induced expression in wheat ofthe Arabidopsis thaliana DREB1A gene delays water stress symptoms undergreenhouse conditions Genome 47:493–500.
Pelleschi, S., A. Leonardi, J.P. Rocher, G. Cornic, D. de Vienne, C. Thévenot, andJ.L. Prioul. 2006. Analysis of the relationships between growth, photosynthe-sis and carbohydrate metabolism using quantitative trait loci (QTLs) in youngmaize plants submitted to water deprivation. Mol. Breed. 17:21–39.
Pereira, J.E.S., and G.R. Fortes. 2004. Production of pre-basic potato plants bycuttings obtained from micropropagated plants. Horti. Brasil. 22:186–192.
Perugini, L.D., J.P. Murphy, D. Marshall, and G.B. Guedira. 2008. Pm37, a newbroadly effective powdery mildew resistance gene from Triticum timopheevii.Theor. Appl. Genet. 116:417–425.
Powell-Abel, P., R.S. Nelson, B. De, N. Hoffman, S.G. Rogers, R.T. Fraley, and R.N.Beachy. 1986. Delay of disease development in transgenic plants that expressthe tobacco mosaic virus coat protein gene. Sci. 232:738–743
Puchta, H. 2003. Marker free transgenic plants. Plant Cell Tissue Organ Cult. 74:123–134.

210 S. S. Gosal et al.
Przetakiewicz, J., A. Nadolska-Orczyk, D. Kuc, and W. Orczyk. 2007. Tetraploidsomatic hybrids of potato (Solanum tuberosum L.) obtained from diploidbreeding lines. Cell. Mol. Biol. Lett. 12:253–267.
Qu, B., G. Liu, B. Zhou, M. Bellizzi, L. Zeng, L. Dai, B. Han, and G.L. Wang. 2006.The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-bindingsite-leucine-rich repeat protein and is a member of a multigene family in rice.Genetics 172:1901–1914.
Qu, J., J. Ye, and R. Fang. 2007. Artificial miRNA-mediated virus resistance in plants.J . Virology. 81:6690–6699.
Quarrie, S. A., A. Steed, C. Calestani, A. Semikhodskii, C. Lebreton, C. Chinoy,and N. Steele. 2005. A high-density genetic map of hexaploid wheat (Triticumaestivum L.) from the cross Chinese Spring SQ1 and its use to compare QTLsfor grain yield across a range of environments. Theor. Appl. Genet. 110:865–880.
Raina, S.K., and F.J. Zapata. 1997. Enhanced anther culture efficiency of Indicarice (Oryza sativa L.) through modification of the culture media. Plant Breed.116:305–315.
Rajasekaran, K., J.W. Cary, J.M. Jaynes, and T.E. Cleveland. 2007. Disease resis-tance conferred by the expression of a gene encoding a synthetic peptidein transgenic cotton (Gossypium hirsutum L.) plants. Plant Biotechnol. J .3:545–554.
Ramanand., N. Singh., N. Kureel, M. Lal, and S.B. Singh. 2007. Effect of transplantingspacing on growth and yield of tissue culture raised crop of sugarcane. IndianSugar 57:33–36.
Randhawa G. J., M. Singh, and R. Sharma. 2009. Duplex, triplex and quadruplexPCR for molecular characterization of genetically modified potato with betterprotein quality. Curr. Sci. 97:21–23.
Rashid, M.H., M. Khalekuzzaman, M.F. Hasan, R. Das, M.S. Hossain, andS. Mahabbat-E-Khoda. 2009. Establishment of an efficient method for micro-propagation of an important medicinal herb (Scoparia dulcis L.) from shoottips and nodal segments. Int. J. Sustain. Crop Prod. 4:5–9.
Ray, K., N.C. Bisht, D. Pental, and P.K.Burma. 2007. Development of bar-nase/barstar transgenics for hybrid seed production in Indian oilseed mustard(Brassica juncea L. Czern & Coss) using a mutant acetolactate synthase geneconferring resistance to imidazolinone-based herbicide ‘Pursuit.’ Curr. Sci.93:1390–1396.
Reddy, L., T.L. Friesen, S.W. Meinhardt, S. Chao, and J.D. Faris. 2008. Genomic anal-ysis of the Snn1 locus on wheat chromosome arm 1BS and the identification ofcandidate genes. Plant Genome 1:55–66.
Ren, J.P., M.H. Dickson, and E.D. Earle. 2000. Improved resistance to bacterial softrot by protoplast fusion between Brassica rapa and B. oleracea. Theor. Appl.Genet. 100:810–819.
Resmi, L., and A.S. Nair. 2007. Plantlet production from male inflorescence tipsof Musa acuminata cultivars from South India. Plant Cell, Tissue Organ Cult.88:333–338.
Ribaut, J.M., M. Bänziger, T. Setter, G. Edmeades, and D. Hoisington. 2004. Geneticdissection of drought tolerance in maize: A case study. In Physiology andbiotechnology integration for plant breeding, eds. N. Nguyen and A. Blum,571–611. New York: Marcel Dekker.

Biotechnology & Crop Improvement 211
Ribaut, J.M., and M. Ragot. 2007. Marker-assisted selection to improve droughtadaptation in maize: The backcross approach, perspectives, limitations, andalternatives. J. Exp. Bot. 58:351–360.
Robin, S., M.S. Pathan, B. Courtois, R. Lafitte, S. Carandang, S. Lanceras, M. Amante,H.T. Nguyen, and Z. Li. 2003. Mapping osmotic adjustment in an advancedback-cross inbred population of rice. Theor. Appl. Genet. 107:1288–1296.
Rodrigues, S.M., M.O. Andrade, A.P.S. Gomes, F.M. DaMatta, M.C. Baracat-Pereira,and E.P.B. Fontes. 2006. Arabidopsis and tobacco plants ectopically expressingthe soybean antiquitin-like ALDH7 gene display enhanced tolerance to drought,salinity, and oxidative stress. J. Exp. Bot. 55:301–308.
Rodrigues, P.H.V. 2008. Somaclonal variation in micropropagated Heliconia bihaicv. Lobster claw I plantlets (Heliconiaceae). Sci. Agric. 65:681–684.
Rokka, V.M., Y.S. Xu, J. Kankila, A. Kuusela, S. Pulli, and E. Pehu. 1994. Identificationof somatic hybrids of dihaploid Solanum tuberosum lines and S. brevidens byspecies specific RAPD patterns and assessment of disease resistance of thehybrids. Euphytica 80:207–217.
Rosati, P., and S. Predieri. 1990. In vitro selection of apple rootstock somacloneswith Phytophthora cactorum culture filtrate. Acta Hort. 280:409–413.
Rozhanskaya, O.A. 2006. Quantitative variability in populations of alfalfa somaclonesand mutants. Russian Agric. Sci. 2:4–7.
Ruiz-Ferrer, V., and O. Voinnet. 2007. Viral suppression of RNA silencing: 2b winsthe golden fleece by defeating argonaute. Bioassays 29:319–323.
Sabry, S.R.S., A.M. Moussa, A.M. Menshawy, and H.S. El-Borhami. 2005.Regeneration of leaf rust (Puccinia recondita) resistant high-yielding wheat(Triticum aestivum L.) somaclones from embryogenic callus of Sakha 61cultivar. Bull-Faculty Agri. Cairo Univ. 56(3):469–481.
Sakamoto, A.A., A. Murata, and N. Murata. 1998. Metabolic engineering of rice lead-ing to biosynthesis of glycinebetaine and tolerance to salt and cold. Plant Mol.Biol. 38:1011–1019.
Salami, S.A., A. Ebadi, Z. Zamani, and M. Ghasemi. 2007. In vitro mass propa-gation of grapevine (Vitis vinifera L.) from fragmented shoot tip via directorganogenesis. Iran. J. Agrl. Sci. 37:1141–1149.
Salokhe, S.S. 2007. Performance of micropropagated sugarcane seed. Indian Sugar57:23–27.
Sandhu, J.S., S.S. Gosal, M.S. Gill, and H.S. Dhaliwal. 1995. Micropropagation ofIndica rice through proliferation of axillary shoots. Euphytica. 81:139–142.
Saranga, Y., C.X. Jiang, R.J. Wright, D. Yakir, and D.H. Paterson. 2004. Geneticdissection of cotton physiological responses to arid conditions and theirinterrelationships with productivity. Plant Cell Environ. 27:263–277.
Sarao, N.K., M.S. Gill, R. Gill, T.S. Bharaj, and S.S. Gosal. 2003. An improved methodfor pollen culture in rice. Oryza 40:77–79.
Schinkel, H., A. Schiermeyer, R. Soeur, R. Fischer, and S. Schillberg. 2005. Productionof an active recombinant thrombomodulin derivative in transgenic tobaccoplants and suspension cells. Transgen Res. 14:251–259.
Schlaich, T., B.M. Urbaniak, N. Malgras, E. Ehler, C. Birrer, L. Meier, andC. Sautter. 2006. Increased field resistance to Tilletia caries provided by aspecific antifungal virus gene in genetically engineered wheat. Plant Biotechnol.J . 4:63–75.

212 S. S. Gosal et al.
Seguı-Simarro, J.M, and F. Nuez. 2007. Embryogenesis induction, caulogenesis, andplant regeneration by in vitro culture of tomato isolated microspores and wholeanthers. J. Expt. Bot. 58:1119–1132.
Senadhira, D., F.J. Zapata-Arias, G.B. Gregoric, M.S. Alejar, H.C. de la Cruz,T.F. Padolina, and A.M. Galvez. 2002. Development of the first salt-tolerantrice cultivar through Indica/Indica anther culture. Field crop Res. 76:103–110.
Sengar, A.S., K.S. Thind, B. Kumar, P. Mittal, and S.S. Gosal. 2009. In vitro selection atcellular level for red rot resistance in sugarcane (Saccharum sp.). Plant GrowthRegul. 58:2001–2009.
Serebriakova, L., K.H. Oldach, and H. Lorz. 2005. Expression of transgenic stilbenesynthases in wheat causes the accumulation of unknown stilbene derivativeswith antifungal activity. J. Plant Physiol. 162:985–1002.
Serrano, C., P. Arce-Johnson, H. Tortes, M. Gebauer, M. Gutierrez, M. Moreno,X. Jordana, A. Venegas, J. Kalazich, and L. Holuigue. 2000. Expression of theChicken lysozyme gene in potato enhances resistance to infection by ErwiniaCarotovora subsp. Atroseptica. Amer. J. Potato Res. 77:191–199.
Shen, X.L., J.J. Chen, M.E. Kane, and R.J. Henny. 2007. Assessment of somaclonalvariation in dieffenbachia plants regenerated through indirect shoot organogen-esis. Plant Cell, Tissue and Organ Cult. 91:21–27.
Shirasawa, K., T. Takabe, and S. Kishitani. 2006. Accumulation of glycinebetainein rice plants that overexpress choline monooxygenase from spinach andevaluation of their tolerance to abiotic stress. Ann. Bot. 98:565–571.
Shizukawa, Y, and M. Mii. 2008. Production of bialaphos-resistant Nierembergiarepens by electroporation. Plant Biotechnology Reports 2:219–226.
Shu, Q., G. Ye, H. Cui, X. Cheng, Y. Xiang, D. Wu, M. Gao, C. Hu, R. Sardana,and I. Altosaar. 2000. Transgenic rice plants with a synthetic cry1Ab genefrom Bacillus thuringiensis were highly resistant to eight lepidopteran rice pestspecies. Mol. Breed. 6:433–439.
Singh, Z.S., J.S. Brar, and S.S. Gosal. 1991. Ovule culture of seedless grapes (Vitisvinifera L.) cv. Perlette. Acta Horticulture 300:325–329.
Singh Kuldeep, M. Ghai, M. Garg, P. Chhuneja, P. Kaur, T. Schnurbusch, B. Keller,and H.S. Dhaliwal. 2007. An integrated molecular linkage map of diploid wheatusing a Triticum boeoticum x T monococcum RIL population. Theor. Appl.Genet. 115:301–312.
Singh, S., J.S. Sidhu, N.Huang, Y.Vikal, Z. Li, D.S. Brar, H.S. Dhaliwal, andG.S. Khush. 2001. Pyramiding three bacterial blight resistance genes (xa5, xa13and Xa21) using marker-assisted selection into Indica rice cultivar PR106. Theor.Appl. Genet. 102:1011–1015.
Singh, G., S.K. Sandhu, M. Meeta, K. Singh, R. Gill, and S.S. Gosal. 2008. In vitroinduction and characterization of somaclonal variation for red rot and otheragronomic traits in sugarcane. Euphytica 160:35–47.
Sivamani, E., A. Bahieldin, J.M. Wraith, T. Al-Niemi, W.E. Dyer, T.H.D. Ho, andR. Wu. 2000. Improved biomass productivity and water use efficiency underwater deficit conditions in transgenic wheat constitutively expressing the barleyHVA1 gene. Plant Sci. 155:1–9.

Biotechnology & Crop Improvement 213
Sjödin, C., and K. Glimelius. 1989. Transfer of resistance against Phoma lingamto Brassica napus by asymmetric somatic hybridization combined with toxinselection. Theor. Appl. Genet.78:513–520.
Sooch, M., J.S. Arora, K. Singh, and S.S Gosal. 1998. Studies on in vitro propagatonof carnation (Dianthus caryophyllus L.). Plant Tissue Culture. 8:61–68.
Sood, N., K. Piyush, R.K. Srivastava, and S.S. Gosal. 2006. Comparative stud-ies on field performance of micropropagated and conventionally propagatedsugarcane plants. Plant Tissue Culture Biotechnol. 16:25–29.
Spielmeyer, W., R. P. Singh, H. McFadden, C.R. Wellings, J. Huerta-Espino, X. Kong,R. Appels, and E.S. Lagudah. 2008. Fine scale genetic and physical mappingusing interstitial deletion mutants of Lr34/ Yr18: A disease resistance locuseffective against multiple pathogens in wheat. Theor. Appl. Genet. 116:481–490.
Stalker, D.M., K.E. Mcbride, and L.D. Malyj. 1998. Herbicide resistance in transgenicplants expressing a bacterial detoxification gene. Science 242:419–423.
Steele, K.A. 2009. Novel upland rice variety bred using marker-assisted selectionand client-oriented breeding released in Jharkhand, India. Plant Breeding News200. May 1, 2009.
Steele, K.A., A.H. Price, H.E. Shashidhar, and J.R. Witcombe. 2006. Marker-assistedselection to introgress rice QTLs controlling root traits into an Indian uplandrice variety. Theor. Appl. Genet. 112:208–221.
Sudhakar, D., L.T. Duc, B.B. Bong, P. Tinjuangjun, S.B. Maqbool, M. Valdez,R. Jefferson, and P. Christou. 1998. An efficient rice transformation systemutilizing mature seed-derived explants and a portable, inexpensive particlebombardment device. Transgenic Res. 7:289–294.
Sun, X.L., D. Liu, H.Q. Zhang, N.X. Huo, R.H.Z. Zhou, and J.Z. Jia. 2006.Identification and mapping of two new genes conferring resistance to pow-dery mildew from Aegilops tauschii (coss.) Schmal. J. Integrative Plant Biology48:1204–1210.
Tabei, Y., S. Kitade, and Y. Nishizawa. 1998. Transgenic cucumber plants harboring arice chitinase gene exhibit enhanced resistance to gray mold (Botrytis cinerea).Plant Cell Rep.17:159–164.
Talbot, N.J., M.J. Kershaw, G.E. Wakley, O.M.H. de Vries, J.G.H. Wessels, andJ.E. Hamer. 1996. MPG1 encodes a fungal hydrophobin involved in surfaceinteractions during infection-related development of Magnaporthe grisea. PlantCell 8:985–999.
Tamayo, M.C., M. Rufat, J.M. Bravo, and S. San. 2000. Accumulation of a maizeproteinase inhibitor in response to wounding and insect feeding, and char-acterization of its activity toward digestive proteinases of Spodoptera littoralislarvae. Planta 211:62–71.
Tang, X.Y., M.T. Xie, Y.J. Kim, J.M. Zhou, D.F. Klessig, and G.B. Martin. 1999.Overexpression of Pto activates defense responses and confers broad resis-tance. Plant Cell 11:15–29.
Tang, W., and Y. Tian. 2003. Transgenic loblolly pine (Pinus taeda L.) plants express-ing a modified delta-endotoxin gene of Bacillus thuringiensis with enhancedresistance to Dendrolimus punctatus and Crypyothelea formosicola. J. Exp. Bot.54:835–44.

214 S. S. Gosal et al.
Tang, F., Y. Tao, T. Zhao, and G. Wang. 2006. In vitro production of haploid anddoubled haploid plants from pollinated ovaries of maize (Zea mays) Plant Cell,Tissue and Organ Culture 84:233–237.
Tanksley, S.D. 1993. Mapping polygenes. Annu. Rev. Genet. 27:205–233.Thind, S.K., N. Jain, and S.S. Gosal. 2007. Aloe vera L. clones produced in vitro
and under natural conditions vary in carbohydrate accumulation. J . Pl. Sci. Res.23:41–45.
Tsilo, T. J., S. Chao, and Y. Jin. 2009. Identification and validation of SSR markerslinked to the stem rust resistance gene Sr6 on the short arm of chromosome 2Din wheat. Theor. Appl. Genet. 118:515–524.
Towill, L.E., R. Bonnart, and G.M. Volk. 2006. Cryopreservation of Arabidopsisthaliana shoot tips. Cryo Letters 27:353–60.
Tuberosa, R., S. Salvi, M.C. Sanguineti, P. Landi, M. MacCaferri, and S. Conti. 2002.Mapping QTLs regulating morpho-physiological traits and yield: case stud-ies, shortcomings and perspectives in drought-stressed maize. Ann. Bot. 89:941–963.
Van der Vossen, E., A. Sikkema, B.T.L. Hekkert, J. Gros, P. Stevens, M. Muskens,D. Wouters, A. Pereira, W. Stiekema, and S. Allefs. 2003. An ancient R genefrom the wild potato species Solanum bulbocastanum confers broad-spectrumresistance to Phytophthora infestans in cultivated potato and tomato. Plant J .36:867–882.
Van der Vossen, E., J. Gros, A. Sikkema, M. Muskens, D. Wouters, P. Wolters,A. Pereira, and S. Allefs. 2005. The Rpi-blb2 gene from Solanum bulbocas-tanum is a Mi-1 gene homolog conferring broad-spectrum late blight resistancein potato. Plant J. 44:208–222.
Vanitharani, R., P. Chellappan, and C.M. Fauquet. 2003. Short interfering RNA-mediated interference of gene expression and viral DNA accumulation incultured plant cells. Proc. Natl. Acad. Sci. 100:9632–9636.
Varindra, R. Randhawa, K.L. Bajaj, and S.S. Gosal. 1997. Capsaicin accumulationin callus culture and whole fruits of Capsicum annum. Plant Tissue Culture.7:47–51.
Vazquez R.C., S. Asurmend, and H.E. Hopp. 2001. Transgenic resistance in potatoplants expressing potato leafroll virus (PLRV) replicase gene sequences is RNA-mediated and suggests the involvement of post-transcriptional gene silencing.Archives Virol. 146:1337–1353.
Veitia-Rodriguez, N., J. Francisco-Cardoso, J.N. Perez, L. Garcia-Rodriguez,I. Bermudez-Caraballosos, L. Garcia-Rodriguez, Y. Padron-Montesinos,P. Orellana-Perez, C. Romero-Quintana, and N. Hernandez. 2002. Evaluationsin field of somaclones of Irish potatoes (Solanum tuberosum Lin.) of the varietyDesiree obtained by somaclonal variation and in vitro mutagenesis. Biotecnol.Veget. 2:21–26.
Verma, V., N.S. Bains, G.S. Mangat, G.S. Nanda, S.S. Gosal, and K. Singh. 1999. Maizegenotypes show striking differences for induction and regeneration of haploidwheat embryos in the wheat x maize system. Crop Sci. 39:1722–1727.
Verma, V., M.J. Foulkes, A.J. Worland, R. Sylvester-Bradley, P.D.S Caligari, andJ.W. Snape. 2004. Mapping quantitative trait loci for flag leaf senescence

Biotechnology & Crop Improvement 215
as a yield determinant in winter wheat under optimal and drought-stressedenvironments. Euphytica 135:255–263.
Vikal, Y., P. Chhuneja, Singh Rippy and H.S. Dhaliwal. 2004. Tagging of an Aegilopsspeltoides derived leaf rust resistance gene Lr28 with a microsatellite marker inwheat. J. Plant Biochem. & Biotech. 13:47–49.
Volk, G.M., and A.M. Casperson. 2007. Plasmolysis and recovery of different celltypes in cryoprotected shoot tips of Mentha X piperita. Protoplasma 231:215–226.
Wang, G.L., W.Y. Song, D.L. Ruan, S. Sideris, and P.C. Ronald. 2007. The clonedgene, Xa21, confers resistance to multiple Xanthomonas oryzae pv. oryzaeisolates in transgenic plants. Mol. Plant-Microbe Interac. 9:855.
Wang, Z., K. Zhang, X. Sun, K. Tang, and J. Zhang. 2005. Enhancement of resistanceto aphids by introducing the snowdrop lectin gene gna into maize plants. J.Biosciences 30:627–638.
Waterhouse, P.M., M.W. Graham, and M.B. Wang. 1998. Virus resistance and genesilencing in plants can be induced by simultaneous expression of sense andantisense RNA. Proc. Natl. Acad. Sci. 95:13959–13964.
Wei, T., H. Chen, C.G. Xu, X.H. Li, Y.J. Lin, and Q.F. Zhang. 2006. Development ofinsect resistant transgenic Indica rice with a synthetic cry1C gene. Mol. Breed.18:1–10.
Weisel, J.W., C. Nagaswami, T.A. Young, and D.R. Light. 1996. The shape ofthrombomodulin and interactions with thrombin as determined by electronmicroscopy. J. Biol. Chem. 271:31485–31490.
Williams, M.E. 1995. Genetic engineering for pollination control. Trends Biotechnol.13:344–349.
Wong, W.S., G.G. Li, W. Ning, Z.F. Xu, W.L.W. Hsiao, L.Y. Zhang, and N. Li.2001. Repression of chilling induced ACC accumulation in transgenic citrusby over production of antisense 1-aminocyclopropane-1-carboxylate synthaseRNA. Plant Science 161:969–977.
Wu, G.B., J. Shortt, E.B. Lawerence, E.B. Levine, K.C. Fitzsimmons, and D.M. Shah.1995. Disease resistance conferred by expression of a gene encoding H2O2-generating glucose oxidase in transgenic potato plants. Plant Cell 7:1357–1368.
Xiao, B., Y. Huang, N. Tang, and L. Xiong. 2007. Over expression of LEA genein rice improves drought resistance under field conditions. Theor. Appl. Genet.115:35–46.
Xiaofen, S., K. Tang, B. Wan, H. Qi, and X. Lu. 2001. Transgenic rice homozygouslines expressing GNA showed enhanced resistance to rice brown plant hopperChin. Sci. Bullet. 46:1698–1703.
Xu, D., Q. Xue, D. McElroy, Y. Mawal, V. A. Hilder, and R. Wu. 1996. Constitutiveexpression of a cowpea trypsin inhibitor gene, CpTi, in transgenic rice plantsconfers resistance to two major rice insect pests. Mol. Breed. 2:167–173.
Xu, J.L., H.R. Lafitte, Y.M. Gao, B.Y. Fu, R. Torres, and Z.K. Li. 2005. QTLs fordrought escape and tolerance identified in a set of random introgression linesof rice. Theor. Appl. Genet. 11:1642–1650.
Xu, X., N. Hayashi, C.T. Wang, H. Kato, T. Fujimura, and S. Kawasaki. 2008. Efficientauthentic fine mapping of the rice blast resistance gene Pik-h in the Pik cluster,using new Pik-h-differentiating isolates. Mol. Breed. 22:289–299.

216 S. S. Gosal et al.
Yamamoto, T., H. Iketani, and H. Ieki. 2000. Transgenic grapevine plants expressinga rice chitinase with enhanced resistance to fungal pathogens. Plant Cell Rep.15: 639–646.
Yamasaki, M., H. Tsunematsu, A. Yoshimura, N Iwata, and H. Yasmi. 1999.Quantitative Trait Locus mapping of ovicidal response in rice (Oryza sativaL.) against whitebacked planthopper (Sogatella furcifera Horváth) Crop Sci.39:1178–1183.
Yang, Y., T.A. Sherwood, C.P. Patte, E. Hiebert, and J.E. Polston. 2004. Use oftomato yellow leaf curl virus (TYLCV) rep gene to engineer TYLCV resistancein tomato. Phytopathology 94:490–496.
Yang, C.Y., Y.C. Hoa, J.C. Panga, S.S. Huang, and J.S.M. Tschen. 2009. Cloning andexpression of an antifungal chitinase gene of a novel Bacillus subtilis isolatefrom Taiwan potato field. Bioresource Technology 100:1454–1458.
Yao, J.L., D. Cohen. R. Atkinson, K. Richardson, and B. Morris. 1995. Regenerationof transgenic plants from the commercial apple cultivar Royal Gala. Plant CellRep. 4:407–412.
Ye, X., S. Al-Babili, A. Klöti, J. Zhang, P. Lucca, P. Beyer, and I. Potrykus. 2000.Engineering provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 287:303–305.
Yoshimura, S., U. Yamanouchi, Y. Katayose, S. Toki, Z.X. Wang, I. kono, N. Kurata,M. Yano, N. Iwata, and T. Sasaki.1998. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc. Natl. Acad.Sci. 95:1663–1668.
Yoon-Ju, W., H. Kim-Haeng, Gi. Cho-Eun, C. Ko-Ho, S. Hwang-Hae, E. Park-Young, and F. Engelmann. 2007. Cryopreservation of cultivated and wild potatovarieties by droplet vitrification procedure. Acta Hort. 760:203–208.
Zhahg-Yi, Y., K. Hong-Ru, W. Zhahg-Jin, Y. Li-Zheng, and C. Zeng-Qian. 2008. Highquality and blast resistance DH lines via anther culture. Southwest-China J.Agrl-Sci. 21:75–79.
Zhai, W.X., W.M. Wang, Y.L. Zhou, X.B. Li, X.W. Zheng, Q. Zhang, G. Wang, andL. Zhu. 2002. Breeding bacterial blight-resistant hybrid rice with the clonedbacterial blight resistance gene Xa21. Mol. Breed. 8:285–293.
Zhang, Y.J., H.Y. Zhao, K.M. Wu, K.J. Zhao, Y.F Peng, and Y.Y. Guo. 2004.Expression of Cry1Ac protein in cry1Ac/CpTI transgenic rice and its resis-tance in different developmental stages to Chilo suppressalis. Chin. J . Agri.Biotechnol. 1:149–153.
Zhangsun, D., S. Luo, R. Chen, and K. Tang. 2008. Improved Agrobacterium-mediated genetic transformation of GNA transgenic sugarcane. Biologia.62:386–393.
Zhao, B., X. Lin, J. Poland, H. Trick, J. Leach, and S. Hulbert. 2005a. A maizeresistance gene functions against bacterial streak disease in rice. Proc. Natl.Acad. Sci. 102: 15383–15388.
Zhao, M.A., Y.Z. Xhu, S.P. Dhital, D.M. Khu, Y.S. Song, M.Y. Wang, and H.T. Lim.2005b. An efficient cryopreservation procedure for potato (Solanum tubero-sum L.) utilizing the new ice blocking agent, Supercool X1000. Plant Cell Rep.24:477–481.

Biotechnology & Crop Improvement 217
Zhao, Z., T. Hu, X.H. Ge, X. Du, L. Ding, and Z. Li. 2008. Production andcharacterization of intergeneric somatic hybrids between Brassica napus andOrychophragmus violaceus and their backcrossing progenies. Plant Cell Rep.27:1611–1621.
Zheng, X., M. Wei-Xiao, Y. Ji-Liang, and M. Hu-Yan. 2004. In vitro selection of NaCl-tolerant variants of maize and analysis of salt tolerance. J. Henan Agrl. Univ.38:139–143.
Zhu, Q., E.A. Maher, S. Masoud, R.A. Dixon, and C.J. Lamb. 1994. Enhanced pro-tection against fungal attack by constitutive coexpression of chitinase andglucanase genes in transgenic tobacco. Biotechnol. 12:807–812.
Zhu, Z.D., R.H. Zhou, X.Y. Kong, Y.C. Dong, and J.Z. Jia. 2005. Microsatellitemarkers linked to two genes conferring resistance to powdery mildew incommon wheat introgressed from Triticum carthlicum accession PS5. Genome48:585–590.




















