
Biogeochemistry of a tropical river affected by human activities in its catchment: Brantas River...
12
Biogeochemistry of a tropical river affected by human activities in its catchment: Brantas River estuary and coastal waters of Madura Strait, Java, Indonesia T.C. Jennerjahn a, ) , V. Ittekkot a , S. Klo¨pper a , Seno Adi b , Sutopo Purwo Nugroho b , Nana Sudiana b , Anyuta Yusmal b , Prihartanto b , B. Gaye-Haake c a Center for Tropical Marine Ecology, Fahrenheitstrasse 6, D-28359 Bremen, Germany b Directorate of Technology for Land Resources and Region Management, Agency for the Assessment and Application of Technology, Jl. M.H. Thamrin 8, 10340 Jakarta, Indonesia c Institute of Biogeochemistry and Marine Chemistry, University of Hamburg, Bundesstrasse 55, D-20146 Hamburg, Germany Received 5 November 2003; accepted 23 February 2004 Abstract On a global scale tropical SE Asia is critical in terms of fluvial nutrient and sediment input into the ocean as well as in terms of human modifications of the coastal zone altering these inputs. In order to obtain information on the biogeochemistry of the Brantas River estuary and adjacent coastal waters of the Madura Strait in eastern Java, Indonesia, a densely-populated area which is strongly affected by human activities in the river catchment, we investigated water, suspended matter and sediments from estuarine and coastal waters and plants and soils from the catchment collected during the dry season. Water samples were analyzed for dissolved nutrients and phytoplankton abundance and composition. Suspended matter, sediment, plant and soil samples were analyzed for carbon, nitrogen, amino acids and stable carbon and nitrogen isotopes. Nutrient concentrations were high in the river and then decreased rapidly seaward. Runoff from agricultural soils may be a major nutrient source. Phytoplankton was dominated by diatoms and most abundant in the estuarine mixing zone. A fairly high phytoplankton abundance despite nitrate-depletion and an N/P ratio !4 in coastal waters indicates additional nitrogen sources. Biogeochemical characteristics and stable isotopes indicate the autochthonous origin of suspended organic matter (OM) during the dry season. Major part of the fluvial input appears to be trapped in the inner estuary. Similar information obtained from the analysis of sediments suggests seasonal differences in the quantity and origin of suspended sediments and OM transported by the river with high inputs into coastal waters during the rainy season, the period of peak river discharge. The amino acid composition in combination with stable isotopes indicates agricultural soils to be a major OM source during that time. A high proportion of amino acid-bound nitrogen in suspended matter and sediments and the presumed sources of dissolved nitrogen other than nitrate to sustain primary productivity in the coastal zone suggest that organic nitrogen may play a more important role for coastal food webs and the nitrogen cycle in tropical regions than previously thought. Ó 2004 Elsevier Ltd. All rights reserved. Keywords: biogeochemistry; rivers; stable isotopes; amino acids; nutrients; Indonesia 1. Introduction Rivers are connecting land and ocean and nourish the continents as well as the coastal ocean and the people living in the respective areas. Natural and anthropogenic environmental changes of the past decades led and will lead to substantial changes of these ecosystems and the biogeochemical cycles of the coastal ocean. Investiga- tions of the past decade have shown that small and mid- sized rivers play a vital role in this respect as they are carrying a sediment load one to two orders of magnitude larger than that of the major river systems (Milliman and Syvitski, 1992). However, particularly in the second half of the 20th century, the diversion and damming of ) Corresponding author. E-mail address: [email protected] (T.C. Jennerjahn). Estuarine, Coastal and Shelf Science 60 (2004) 503e514 www.elsevier.com/locate/ECSS 0272-7714/$ - see front matter Ó 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.ecss.2004.02.008
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Biogeochemistry of a tropical river affected by human activities in its catchment: Brantas River...

Estuarine, Coastal and Shelf Science 60 (2004) 503e514
www.elsevier.com/locate/ECSS
Biogeochemistry of a tropical river affected by humanactivities in its catchment: Brantas River estuary
and coastal waters of Madura Strait, Java, Indonesia
T.C. Jennerjahna,), V. Ittekkota, S. Kloppera, Seno Adib, Sutopo Purwo Nugrohob,Nana Sudianab, Anyuta Yusmalb, Prihartantob, B. Gaye-Haakec
aCenter for Tropical Marine Ecology, Fahrenheitstrasse 6, D-28359 Bremen, GermanybDirectorate of Technology for Land Resources and Region Management, Agency for the Assessment and Application of Technology,
Jl. M.H. Thamrin 8, 10340 Jakarta, IndonesiacInstitute of Biogeochemistry and Marine Chemistry, University of Hamburg, Bundesstrasse 55, D-20146 Hamburg, Germany
Received 5 November 2003; accepted 23 February 2004
Abstract
On a global scale tropical SE Asia is critical in terms of fluvial nutrient and sediment input into the ocean as well as in terms ofhuman modifications of the coastal zone altering these inputs. In order to obtain information on the biogeochemistry of the Brantas
River estuary and adjacent coastal waters of the Madura Strait in eastern Java, Indonesia, a densely-populated area which isstrongly affected by human activities in the river catchment, we investigated water, suspended matter and sediments from estuarineand coastal waters and plants and soils from the catchment collected during the dry season. Water samples were analyzed fordissolved nutrients and phytoplankton abundance and composition. Suspended matter, sediment, plant and soil samples were
analyzed for carbon, nitrogen, amino acids and stable carbon and nitrogen isotopes. Nutrient concentrations were high in the riverand then decreased rapidly seaward. Runoff from agricultural soils may be a major nutrient source. Phytoplankton was dominatedby diatoms and most abundant in the estuarine mixing zone. A fairly high phytoplankton abundance despite nitrate-depletion and
an N/P ratio !4 in coastal waters indicates additional nitrogen sources. Biogeochemical characteristics and stable isotopes indicatethe autochthonous origin of suspended organic matter (OM) during the dry season. Major part of the fluvial input appears to betrapped in the inner estuary. Similar information obtained from the analysis of sediments suggests seasonal differences in the
quantity and origin of suspended sediments and OM transported by the river with high inputs into coastal waters during the rainyseason, the period of peak river discharge. The amino acid composition in combination with stable isotopes indicates agriculturalsoils to be a major OM source during that time. A high proportion of amino acid-bound nitrogen in suspended matter and
sediments and the presumed sources of dissolved nitrogen other than nitrate to sustain primary productivity in the coastal zonesuggest that organic nitrogen may play a more important role for coastal food webs and the nitrogen cycle in tropical regions thanpreviously thought.� 2004 Elsevier Ltd. All rights reserved.
Keywords: biogeochemistry; rivers; stable isotopes; amino acids; nutrients; Indonesia
1. Introduction
Rivers are connecting land and ocean and nourish thecontinents as well as the coastal ocean and the peopleliving in the respective areas. Natural and anthropogenic
) Corresponding author.
E-mail address: [email protected] (T.C. Jennerjahn).
0272-7714/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.ecss.2004.02.008
environmental changes of the past decades led and willlead to substantial changes of these ecosystems and thebiogeochemical cycles of the coastal ocean. Investiga-tions of the past decade have shown that small and mid-sized rivers play a vital role in this respect as they arecarrying a sediment load one to two orders of magnitudelarger than that of the major river systems (Millimanand Syvitski, 1992). However, particularly in the secondhalf of the 20th century, the diversion and damming of

504 T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
rivers has led to an overall reduction of sediment inputsinto the ocean (Milliman, 1997). In the same time globalriverine nitrogen and phosphorus inputs into the oceanhave trebled due to human activities whereas the inputof dissolved silicate stemming from natural sources wassignificantly reduced. Changed ratios of the essentialnutrients Si, N and P entail changes of the planktoncommunity and the biogeochemical cycles (e.g. Conleyet al., 1993; Humborg et al., 1997; Meybeck, 1998;Rabalais, 1999; Smith et al., 2003).
Because of regional climate, geology and human acti-vities this is particularly important in the indoasian/western Pacific region where about one-fifth of theannual water input and nearly half of the global annualsediment input into the ocean are introduced (Millimanet al., 1995). Moreover, SE Asia is among the regionswith the strongest human modifications of the coastalzone worldwide (Elvidge et al., 1997) implying thathuman activities there have the most vigorous effects onthe ecology and the elemental cycles of coastal ecosys-tems. However, anthropogenically induced environmen-tal changes can have contrasting effects. Deforestation,extensive agriculture, fertilizer and pesticide use, wastedischarge, river diversion and destabilization of riverbeds lead to increased nutrient and sediment loads ofrivers. Damming of the rivers has an opposite effect.
This paper presents results of biogeochemical inves-tigations in the Brantas River estuary and the adjacentcoastal waters of the Madura Strait in eastern Java,Indonesia. It is a mid-sized river that is under strongecological pressure due to intensive human activities inits catchment.
2. Material and methods
2.1. Study area
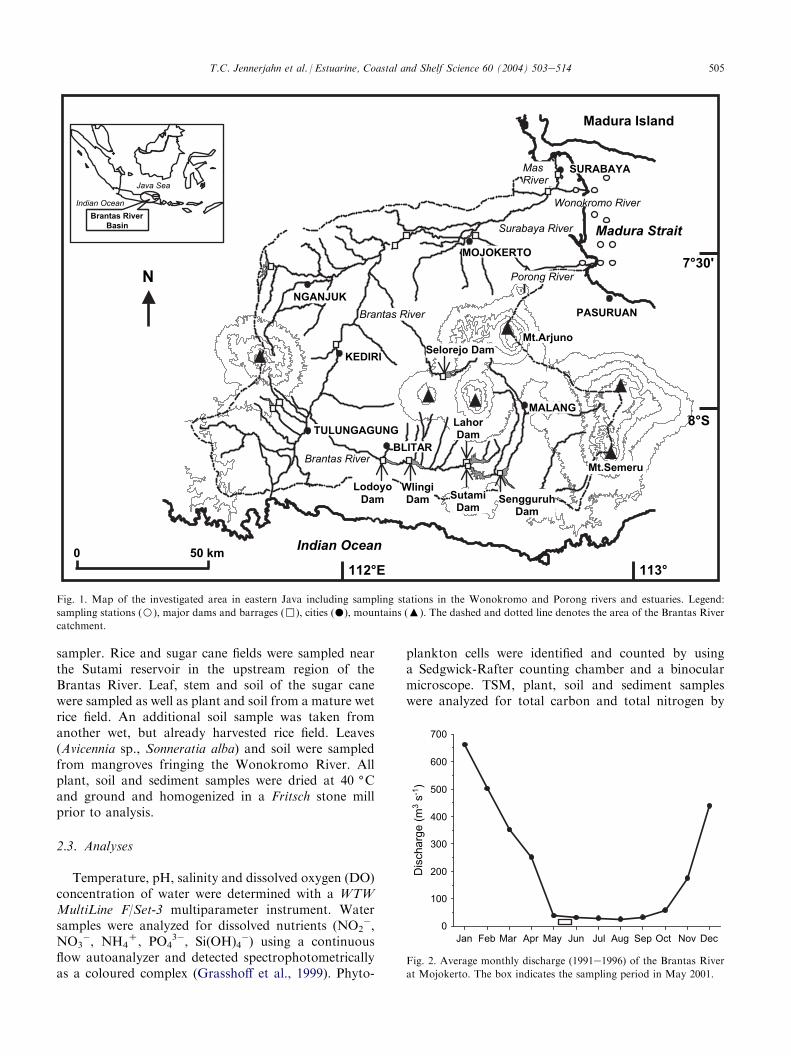
The Brantas with a length of 320 km and a catchmentarea of 11,050 km2 (extending approx. from 111(30#E to112(45#E and 7(20#S to 8(20#S) is the second largestriver of Java. It originates near the volcano Arjuno anddiverts into three branches in the coastal lowlands(Fig. 1). The Porong and the Wonokromo are the twomajor branches discharging into Madura Strait while thesmaller Mas River discharges into the narrowest part ofMadura Strait north of Surabaya after passing the city.The climate is dominated by the monsoons, but incontrast to the western part of the island eastern Javaexperiences only one wet season during the monthsNovembereApril with an average annual rainfall of2220 mm. This is also the period of peak river dischargewhich has been on the order of 217 m3 s�1 on an annualaverage during the period of 1991e1996 (Fig. 2).Approximately 40 km before Surabaya City the riverthen branches in two, the northeastward flowing
Surabaya River and the eastward-directed Porong Riverbeing the major transporting agent for water and sedi-ment. During the wet season when almost 80% of thewater supplied by the Brantas is diverted to the Porongits average discharge can be about 600 m3 s�1 which mayrise to 1200 m3 s�1 in extremely wet years (Hoekstraet al., 1989). During El Nino years, however, dischargecan be much lower due to a decrease in precipitation.Approximately 10 km before Surabaya City the riverthen branches in two, the Kali Mas with a low volume ofwater passing Surabaya City and the eastward-directedWonokromo River which discharges into Madura Strait30 km north of the Porong River. Due to high sedimentloads, particularly during the wet season, the Porong hasa strongly prograding delta. During the dry season flowis mainly diverted to the town of Surabaya and dischargeof the Porong is extremely low (Hoekstra, 1989). TheMadura Strait is characterized by a mixed diurnalesemidiurnal tide in the micro- to mesotidal range(Hoekstra et al., 1989).
Approximately 16 million people live in and on theresources of the Brantas area. With the dramaticallyincreasing population in the 20th century human acti-vities have altered Java’s landscape and ecology. Majordams and reservoirs, respectively, have been built in the1970s and 80s, mainly for the purposes of electricalpower generation, irrigation and flood control in theBrantas catchment. Other human activities like defor-estation, intensive agriculture (mainly rice cultivation),urban and industrial waste disposal, the formation oflevees and sand mining in the downstream portion, riverdiversion and conversion of the estuarine mangroves toaquaculture ponds alter the flow regime and the amountand composition of substances transported by the riverand discharged into Madura Strait.
2.2. Sampling and sample preparation
The Wonokromo and Porong estuaries were sampledfor water, phytoplankton, suspended matter and sedi-ments along a salinity gradient starting from zerosalinity at high tide in May 2001 during the dry seasonwhen river discharge is low. Water samples for nutrientanalysis were filtered through single use membranefilters into prewashed PE bottles, preserved with mercurychloride solution (20 g l�1) and kept frozen until anal-ysis. Plankton samples were obtained by filtering 20 l ofsurface water (0e0.5 m water depth) through a net of35 mm mesh size in 100 ml PE bottles. Samples werepreserved with formol solution and stored in the darkuntil analysis. Water samples for total suspended matter(TSM) filtration were taken in PE tanks, cooled andstored in the dark until filtration. Samples were filteredover precombusted (5 h, 450 (C) Whatman GF/F filtersand dried at 40 (C. Surface sediment samples (uppercm) for biogeochemical analysis were taken by a grab
505T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
Java Sea
Indian Ocean
Mt.Arjuno
Mt.Semeru
Madura Strait
Indian Ocean
Brantas River
Brantas River
Surabaya River
Mas
River
Porong River
Lodoyo
Dam
Selorejo Dam
Wlingi
DamSutami
Dam
Lahor
Dam
Sengguruh
Dam
MALANG
KEDIRI
TULUNGAGUNG
BLITAR
PASURUAN
NGANJUK
MOJOKERTO
Brantas River
Basin
Madura Island
0 50 km
N
8°S
7°30'
113°112°E
SURABAYA
Wonokromo River
Fig. 1. Map of the investigated area in eastern Java including sampling stations in the Wonokromo and Porong rivers and estuaries. Legend:
sampling stations (B), major dams and barrages (,), cities (�), mountains (:). The dashed and dotted line denotes the area of the Brantas River
catchment.
sampler. Rice and sugar cane fields were sampled nearthe Sutami reservoir in the upstream region of theBrantas River. Leaf, stem and soil of the sugar canewere sampled as well as plant and soil from a mature wetrice field. An additional soil sample was taken fromanother wet, but already harvested rice field. Leaves(Avicennia sp., Sonneratia alba) and soil were sampledfrom mangroves fringing the Wonokromo River. Allplant, soil and sediment samples were dried at 40 (Cand ground and homogenized in a Fritsch stone millprior to analysis.
2.3. Analyses
Temperature, pH, salinity and dissolved oxygen (DO)concentration of water were determined with a WTWMultiLine F/Set-3 multiparameter instrument. Watersamples were analyzed for dissolved nutrients (NO2
�,NO3
�, NH4C, PO4
3�, Si(OH)4�) using a continuous
flow autoanalyzer and detected spectrophotometricallyas a coloured complex (Grasshoff et al., 1999). Phyto-
plankton cells were identified and counted by usinga Sedgwick-Rafter counting chamber and a binocularmicroscope. TSM, plant, soil and sediment sampleswere analyzed for total carbon and total nitrogen by
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Dis
char
ge (m
3 s-1
)
0
100
200
300
400
500
600
700
Fig. 2. Average monthly discharge (1991e1996) of the Brantas River
at Mojokerto. The box indicates the sampling period in May 2001.

506 T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
high-temperature combustion in a Carlo Erba NA 2100elemental analyzer (Verardo et al., 1990). Organic carbonwas determined similarly after removal of carbonate byacidification with 1 N HCl and subsequent drying at40 (C. Inorganic carbon was calculated by subtraction oforganic from total carbon. Biogenic opal was determinedphotometrically as silicomolybdate complex followinga modified procedure of Mortlock and Froelich (1989).
Total hydrolyzable amino acids (AA) and totalhydrolyzable hexosamines (HA) were analyzed witha Pharmacia LKB Alpha Plus 4151 Amino Acid Analyzerafter hydrolysis with 6 N HCl for 22 h at 110 (C.Hexosamine concentrations were multiplied by a factorof 1.4 to compensate for losses during hydrolysis(Muller et al., 1986). The amino acids and hexosamineswere separated on a cation exchange resin and detectedfluorimetrically (Roth and Hampai, 1973). A detaileddescription of the procedure is given by Jennerjahn andIttekkot (1999).
The nitrogen isotope composition (d15N) was de-termined in a Finnigan Delta Plus gas isotope ratio massspectrometer after high temperature combustion in aFlash 1112 EA elemental analyzer. d15N is given as &-deviation from the nitrogen isotope composition of at-mospheric air. The carbon isotope composition (d13Corg)was determined similarly after removal of carbonate byadding 1 N HCl and subsequent drying at 40 (C. d13Corg
is given as &-deviation from the carbon isotope com-position of the PDB standard. Salinity was measured inpractical salinity units.
3. Results and discussion
The stations and respective samples have beensubdivided into three groups (river, mixing zone, coastalwaters) according to their location in the investigatedarea and variations of salinity.
3.1. Nutrient and phytoplankton compositionand sources during the dry season
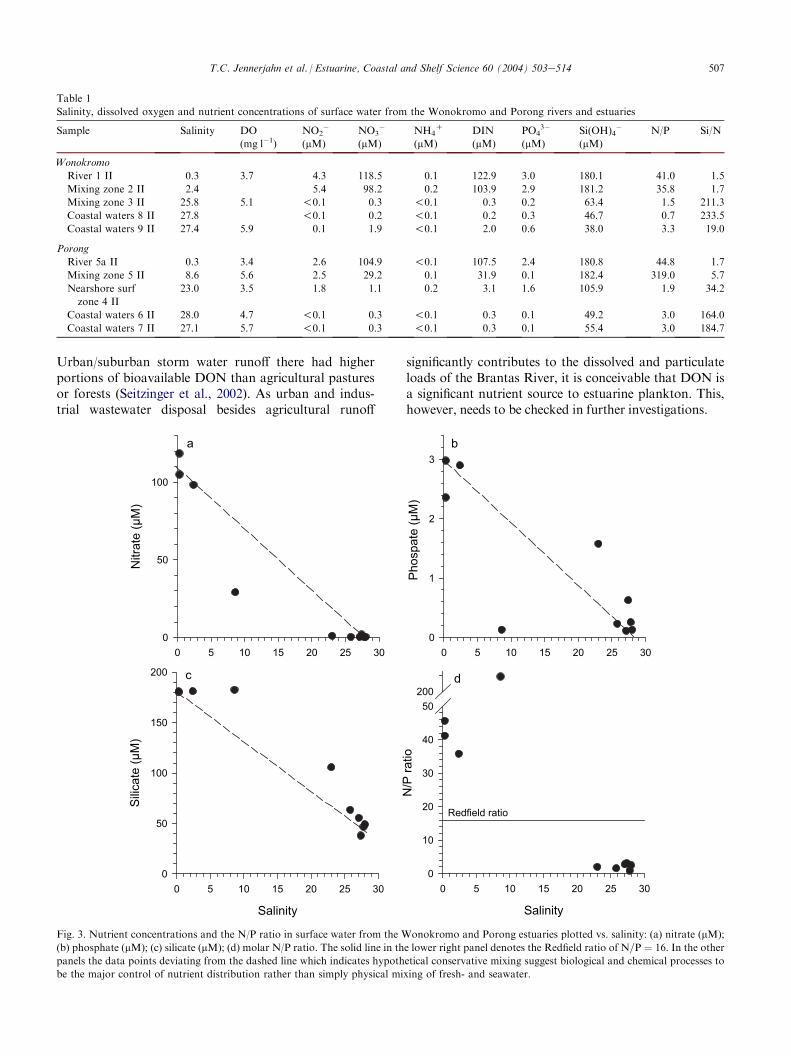
In general, the concentrations of dissolved nutrientsare high in the riverine stations with salinities !10(Table 1, Fig. 3). Dissolved inorganic nitrogen (DIN)mainly consists of nitrate with concentrations of up to119 mM in the rivers and rapidly decreases to extremelylow concentrations with increasing salinity. Nitrite con-centrations range between 2 and 5 mM in low salinitywater and decrease seaward. Ammonium is present intraces only at all stations. Phosphate ranges between 2and 3 mM in low salinity water and decreases to traceamounts seaward. Although also decreasing seaward,silicate concentrations are high at all stations. Un-expectedly high values at station 4 II which is located in
the nearshore surf zone may result from resuspension ofsediments and recycling of OM and nutrients.
The phytoplankton concentration increases from afew thousand cells at the freshwater end of the estuariesto maximum numbers of around 250! 103 cells l�1 inthe mixing zone and then rapidly decreases seaward.Subdivided into five groups the phytoplankton compo-sition is dominated by diatoms (Baciallariophyceae)except for Wonokromo stations 1 II and 2 II wherecyanobacteria (Cyanophyceae) and chlorophyta (Chlor-ophyceae) are the dominant groups (Table 2). Asignificant contribution of benthic diatoms to the phy-toplankton corroborates the presumed resuspension ofsediments at station 4 II.
Increased phytoplankton abundance dominated bydiatoms at the freshwatereseawater boundary suggestsincreased productivity in the mixing zone leading to theobserved depletion of nitrate and phosphate anda change in the nutrient mix. The N/P ratio is rapidlydropping from much larger than the Redfield ratio inthe rivers to much lower in the outer estuary (Fig. 3).Despite the high N/P ratio, however, elevated phosphateand high TSM concentrations indicate that primaryproductivity in the river appears to be lightdrather thanphosphorus-limited due to high turbidity. High nitrateconcentrations and the associated high N/P ratio mayresult mainly from the combined effects of extensivefertilizer use in agriculture in the river catchment and thelow flow during the dry season. The strong non-lineardecrease of nitrate and phosphate with increasingsalinity indicates rapid uptake by phytoplankton ratherthan simply dilution with seawater being the majorcontrol for low nutrient concentrations in coastal waters(Fig. 3).
Interestingly, the phytoplankton with 40e70! 103
cells l�1 is fairly abundant in the coastal zone althoughDIN is almost zero and the N/P ratio around 3 or lesswhich is much lower than the Redfield ratio of 16 (Fig. 3).The nitrogen required to sustain this phytoplankton stocktherefore must come from other sources. Because of theinsignificance of Cyanophyceae, fixation of atmosphericnitrogen can be excluded as a source. Benthic recyclingmay provide DIN which is consumed immediatelykeeping its concentration low. However, densities andbiomass of macrobenthic fauna in the investigated areahave been observed to be low (Erftemeijer and Swennen,1990). Hence dissolved organic nitrogen (DON) may bean additional source to sustain productivity in DIN-depleted coastal waters. While amino acids are knownto be utilized by phytoplankton by direct uptake, othercompounds of the DON pool, like urea, activate micro-bial food webs which then by conversion to inorganicN species indirectly support phytoplankton growth (Berget al., 2001; Hasegawa et al., 2001). Investigations fromtemperate regions have shown that DON may be an im-portant contributor to the pool of bioavailable nitrogen.
507T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
Table 1
Salinity, dissolved oxygen and nutrient concentrations of surface water from the Wonokromo and Porong rivers and estuaries
Sample Salinity DO
(mg l�1)
NO2�
(mM)
NO3�
(mM)
NH4C
(mM)
DIN
(mM)
PO43�
(mM)
Si(OH)4�
(mM)
N/P Si/N
Wonokromo
River 1 II 0.3 3.7 4.3 118.5 0.1 122.9 3.0 180.1 41.0 1.5
Mixing zone 2 II 2.4 5.4 98.2 0.2 103.9 2.9 181.2 35.8 1.7
Mixing zone 3 II 25.8 5.1 !0.1 0.3 !0.1 0.3 0.2 63.4 1.5 211.3
Coastal waters 8 II 27.8 !0.1 0.2 !0.1 0.2 0.3 46.7 0.7 233.5
Coastal waters 9 II 27.4 5.9 0.1 1.9 !0.1 2.0 0.6 38.0 3.3 19.0
Porong
River 5a II 0.3 3.4 2.6 104.9 !0.1 107.5 2.4 180.8 44.8 1.7
Mixing zone 5 II 8.6 5.6 2.5 29.2 0.1 31.9 0.1 182.4 319.0 5.7
Nearshore surf
zone 4 II
23.0 3.5 1.8 1.1 0.2 3.1 1.6 105.9 1.9 34.2
Coastal waters 6 II 28.0 4.7 !0.1 0.3 !0.1 0.3 0.1 49.2 3.0 164.0
Coastal waters 7 II 27.1 5.7 !0.1 0.3 !0.1 0.3 0.1 55.4 3.0 184.7
Urban/suburban storm water runoff there had higherportions of bioavailable DON than agricultural pasturesor forests (Seitzinger et al., 2002). As urban and indus-trial wastewater disposal besides agricultural runoff
significantly contributes to the dissolved and particulateloads of the Brantas River, it is conceivable that DON isa significant nutrient source to estuarine plankton. This,however, needs to be checked in further investigations.
0 5 10 15 20 25 30
Nitr
ate
(µM
)
0
50
100
0 5 10 15 20 25 30
Phos
pate
(µM
)
0
1
2
3
Salinity
0 5 10 15 20 25 30
Silic
ate
(µM
)
0
50
100
150
200
Salinity
0 5 10 15 20 25 30
N/P
ratio
0
10
20
30
40
50200
Redfield ratio
a
dc
b
Fig. 3. Nutrient concentrations and the N/P ratio in surface water from the Wonokromo and Porong estuaries plotted vs. salinity: (a) nitrate (mM);
(b) phosphate (mM); (c) silicate (mM); (d) molar N/P ratio. The solid line in the lower right panel denotes the Redfield ratio of N=P ¼ 16. In the other
panels the data points deviating from the dashed line which indicates hypothetical conservative mixing suggest biological and chemical processes to
be the major control of nutrient distribution rather than simply physical mixing of fresh- and seawater.
508 T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
Table 2
Phytoplankton community composition in surface water of the Wonokromo and Porong rivers and estuaries
Sample Bacillariophyceae Chlorophyceae Dinophyceae Euglenophyceae Cyanophyceae
(103 cells l�1) (%) (103 cells l�1) (%) (103 cells l�1) (%) (103 cells l�1) (%) (103 cells l�1) (%)
Wonokromo
River 1 II 0.07 2.5 0.58 19.8 0 0 0.08 2.9 2.17 74.8
Mixing zone 2 II 28.78 53.1 20.89 38.5 0.39 0.7 0 0 4.19 7.7
Mixing zone 3 II 214.95 99.8 0 0 0.54 0.3 0 0 0 0
Coastal waters 8 II 67.73 99.8 0 0 0.17 0.3 0 0 0 0
Coastal waters 9 II 77.85 99.9 0 0 0.03 0.1 0 0 0 0
Porong
River 5a II 6.35 87.6 0.83 11.4 0 0 0 0 0.07 1.0
Mixing zone 5 II 248.54 100.0 0 0 0 0 0 0 0 0
Nearshore surf
zone 4 II
277.90 99.0 0 0 2.81 1.00 0 0 0 0
Coastal waters 6 II 63.00 96.8 0 0 1.84 2.8 0 0 0.23 0.4
Coastal waters 7 II 41.06 97.2 0 0 1.17 2.8 0 0 0 0
3.2. Sources and reactivity of suspendedparticulate OM during the dry season
TSM contents vary between 7.2 and 70.7 mg l�1 inthe rivers and coastal waters with an exceptionally highvalue of 5392.0 mg l�1 due to resuspension of sedimentsin the nearshore surf zone at station 4 II (s.a.).Generally, maximum TSM contents were found in themixing zone of freshwater and seawater, the so-calledturbidity maximum zone (Abril et al., 2002). Minimum
TSM contents were found in coastal waters off thePorong (Table 3). Except for the resuspended materialat station 4 II, Corg and N contents of TSM are in therange of 4.0e12.0% and 0.4e1.3%, respectively, withmaximum concentrations found in the outer estuary.Corg and N contents are generally higher in the Porongestuary. The C/N ratio displays little variation and isalmost in the range of marine OM except for station 4 IIwhere a contribution of resuspended sedimentary OMraises the C/N ratio to 38.5. Its variability is lower in the
Table 3
Biogeochemical characteristics of suspended matter and sediments from the Wonokromo and Porong rivers and estuaries
Sample Salinity TSM
(mg l�1)
Biogenic
opal (%)
Corg
(%)
N
(%)
C/N
by wt.
d13Corg
(&)
d15N
(&)
AA
(mg g�1)
HA
(mg g�1)
AA-C%
(%)
AA-N%
(%)
RI
Wonokromo TSM
River 1 II 0.3 55.2 4.2 0.5 8.8 �24.7 �2.3 16.0 1.0 16.8 47.0 13.0
Mixing zone 2 II 2.4 24.8 4.9 0.8 6.4 �25.4 0.9 33.7 2.5 30.3 61.4 8.1
Mixing zone 3 II 25.8 70.7 4.0 0.4 9.5 �19.6 5.3 17.1 1.1 18.8 54.1 11.2
Coastal waters 8 II 27.8 28.0 4.7 0.6 7.5 �19.6 6.0 25.5 2.2 23.8 55.3 8.9
Coastal waters 9 II 27.4 17.4 8.1 0.9 9.5 �19.2 7.3 54.2 4.1 29.7 88.1 29.1
Wonokromo sediments
River 1 II 2.5 0.9 !0.1 23.3 �22.0 3.8 1.7 0.5 7.8 57.7 1.6
Mixing zone 2 II 3.1 1.2 0.1 20.7 �23.3 4.0 2.9 0.6 9.8 64.2 2.6
Mixing zone 3 II 3.3 1.1 0.1 17.5 �21.3 5.1 3.9 0.5 16.0 87.5 4.3
Coastal waters 8 II 3.2 0.7 !0.1 17.8 �21.1 4.8 1.4 0.6 8.2 47.0 3.0
Coastal waters 9 II 2.9 0.4 !0.1 21.5 �21.8 5.3 0.8 0.5 7.9 53.5 3.5
Porong TSM
River 5a II 0.3 11.4 10.1 0.8 12.7 �28.9 5.7 73.8 3.4 32.7 100.0 14.9
Mixing zone 5 II 8.6 52.8 6.6 1.3 5.2 �24.5 10.0 61.5 3.8 41.8 65.9 16.2
Nearshore surf zone 4 II 23.0 5392.0 0.8 !0.1 38.5 �21.4 3.1 4.0 0.5 22.5 100.0 4.6
Coastal waters 6 II 28.0 8.7 8.4 1.0 8.3 �19.7 6.5 51.3 4.7 27.4 69.0 21.3
Coastal waters 7 II 27.1 7.2 12.0 1.1 10.7 �20.9 6.4 59.6 5.0 22.2 73.1 23.8
Porong sediments
River 5a II 2.0 0.5 !0.1 25.0 �24.8 5.6
Mixing zone 5 II 0.1 !0.1 17.5 �22.0
Nearshore surf zone 4 II 3.1 1.2 0.1 17.6 �21.7 5.6 4.0 0.6 13.9 76.5 3.3
Coastal waters 6 II 2.7 1.1 0.1 22.2 �22.0 5.1 3.1 0.2 8.2 47.0 2.5
Coastal waters 7 II 2.9 0.9 0.1 15.5 �20.9 5.2 3.0 0.1 7.9 53.5 2.8
The RI (reactivity index) is the molar ratio of the aromatic amino acids tyrosine+ phenylalanine vs. the non-protein amino acids b-alanine+
g-aminobutyric acid. High RI values indicate the freshness of the OM (Jennerjahn & Ittekkot, 1997).

509T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
Wonokromo (mean 8.3) than in the Porong River (mean9.2, excluding station 4 II).
AA and HA concentrations vary between 16.0e73.8 mg g�1 and 1.0e5.0 mg g�1, respectively, with ex-ceptionally lower values at station 4 II (Table 3). Theseare in the range of suspensions from coastal and openocean waters (California coast 27e52 mg g�1, equatorialPacific 33e50 mg g�1; Siezen and Mague, 1978), but atthe lower end of concentrations found in suspendedmatter from the Potomac estuary (60e129 mg g�1;Sigleo and Shultz, 1993). The portion of N accountedfor by amino acids (AA-N%) varying between 47.0%and 100.0%, however, is significantly higher than inmajor world rivers (average 43%; Ittekkot and Zhang,1989) and in Brazilian coastal waters (average 53.9%;Jennerjahn and Ittekkot, 1999). Also, the reactivityindex (RI) of OM varying between 4.6 and 29.1 is higherthan in plankton and suspended matter from variouscoastal regions (Jennerjahn and Ittekkot, 1997). Theseresults indicate that major part of the suspendedparticulate N is contributed by labile organic com-pounds and that suspended OM is highly reactive, henceof high nutritional value during the dry season.
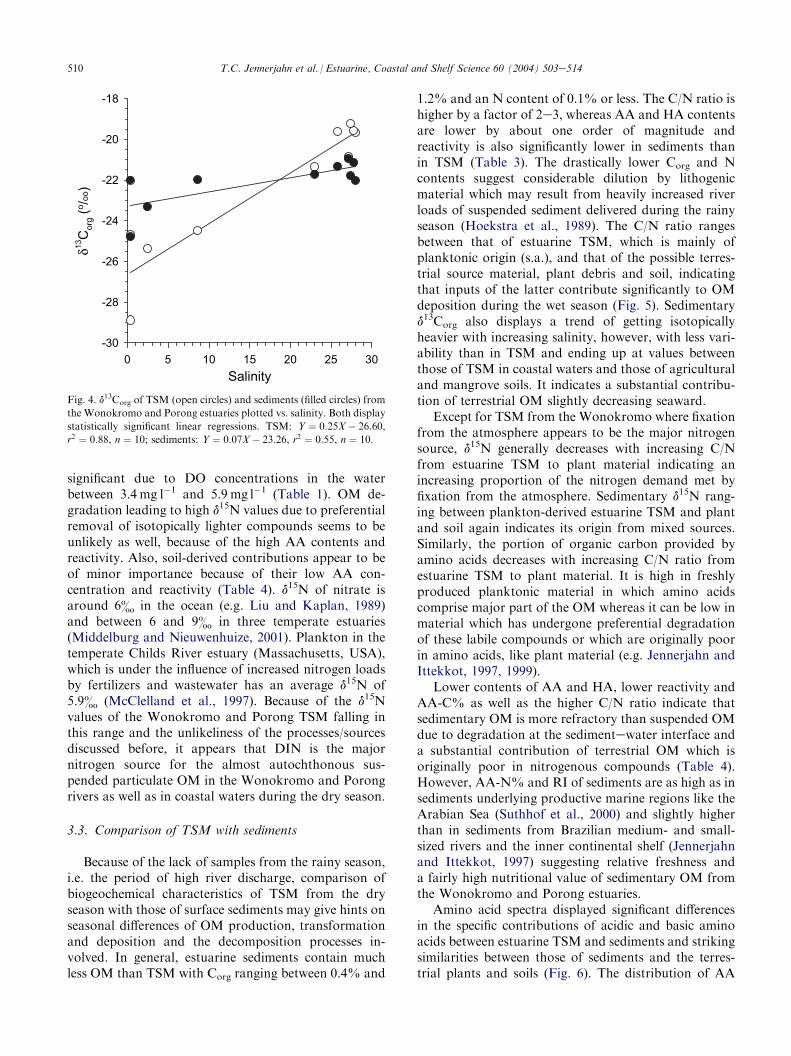
d13Corg increases significantly from values of �25&to �29& in the rivers to values around �20& towardsthe sea which are typical for marine plankton in lowlatitudes (e.g. Fischer, 1991). The isotopically lightervalues in the rivers may result from a mixture of OMfrom different sources. According to the differences incarbon source and uptake, terrestrial plants of the C3(�25& to �30&) and C4 and CAM type (�10& to�18&) and marine plankton (�18& to �22& intropical regions; e.g. Fischer, 1991) usually displaydistinctly different isotopic signatures making d13Corg
a suitable tool for determining the source of OM in themarine environment. However, the typical values forfreshwater plankton being in the range of �24& to�30& hamper the use of d13Corg as a source indicator atthe freshwatereseawater boundary. Freshwater plank-
ton is usually more enriched in the light isotope thanmarine plankton due to large amounts of isotopicallylight CO2 of organic origin reaching the water body withgroundwater (Galimov, 1985). In two upstream reser-voir lakes of the Brantas River, the Sutami and Selorejoreservoirs, d13Corg values between �25& and �30&were determined in dry season TSM which almostexclusively consisted of planktonic OM (Jennerjahn,unpubl. data). Besides autochthonous freshwater plank-ton plant debris and soil from the major agriculturalcrops in the Brantas River catchment, rice and sugarcane, and from the mangroves which cover the wholeestuarine region are potential allochthonous OMsources. Sugar cane, a C4 plant, and its soil are isotopi-cally heavier than estuarine TSM and thus appear not tobe a significant OM source (Table 4). Although d13Corg
of rice and mangrove plant and soil is in the same rangeas that of estuarine TSM, their distinctly higher C/Nratio renders them an unlikely OM source as well. Astatistically significant correlation of d13Corg of estuarineTSM with salinity indicates that the distribution ofparticulate organic matter is mainly controlled byphysical mixing of river and coastal waters (Fig. 4).
d15N displays a wide range from �2.3& to 10.0& inestuarine TSM samples. d15N can be affected by anumber of processes like the degree of nutrient utiliza-tion in the water column, nitrification or denitrification,organic matter diagenesis or material inputs fromdifferent sources (e.g. Liu and Kaplan, 1989; Wadaand Hattori, 1990; Schafer and Ittekkot, 1993; Holmeset al., 1998; Ostrom et al., 1998; Carpenter et al., 1999;Voss et al., 2001; Stewart et al., 2002). Negative or nearto zero d15N of TSM indicates N2-fixation to be a majornitrogen source at Wonokromo stations 1 II and 2 II.This is in good agreement with the phytoplanktoncomposition to a large extent consisting of nitrogen-fixing cyanobacteria. Other processes and/or sourcesappear to be responsible for the higher d15N of TSMat other stations. Denitrification is unlikely to be
Table 4
Biogeochemical characteristics of plants and soils from major agricultural crops near the Sutami reservoir and the mangroves bordering the
Wonokromo estuary
Sample Biogenic
opal (%)
Corg
(%)
N
(%)
C/N d13Corg
(&)
d15N
(&)
AA
(mg g�1)
HA
(mg g�1)
AA-C%
(%)
AA-N%
(%)
RI
Near Sutami reservoir
Rice plant 4.7 34.3 1.2 29.2 �28.1 1.2 52.3 0.2 6.1 100.0 6.2
Rice fresh soil 2.5 1.9 0.1 20.7 �26.0 2.0 5.3 0.8 22.5 79.7 5.0
Rice harvested soil 9.1 1.2 0.1 22.1 �23.4 2.9 0.8 0.2 3.0 23.4 1.8
Sugar cane leaf 2.2 40.2 0.7 54.5 �12.0 �2.7 63.9 0.7 7.0 100.0 4.9
Sugar cane stem 1.4 38.5 0.1 294.6 �12.6 �0.8 9.0 0.2 1.0 92.2 2.1
Sugar cane soil 2.3 1.4 0.1 20.6 �20.0 3.7 2.6 0.8 8.3 51.2 1.1
Wonokromo estuary
Leaf Avicennia 0 38.5 1.6 24.5 �31.0 2.9 64.7 0.5 7.6 56.5 27.9
Leaf Sonneratia alba 0 38.5 0.7 58.9 �29.3 2.1 36.6 0.4 4.3 76.4 66.4
Mangrove soil 2.5 1.4 0.1 19.5 �25.1 3.7 3.9 0.5 12.0 78.4 4.9
510 T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
significant due to DO concentrations in the waterbetween 3.4 mg l�1 and 5.9 mg l�1 (Table 1). OM de-gradation leading to high d15N values due to preferentialremoval of isotopically lighter compounds seems to beunlikely as well, because of the high AA contents andreactivity. Also, soil-derived contributions appear to beof minor importance because of their low AA con-centration and reactivity (Table 4). d15N of nitrate isaround 6& in the ocean (e.g. Liu and Kaplan, 1989)and between 6 and 9& in three temperate estuaries(Middelburg and Nieuwenhuize, 2001). Plankton in thetemperate Childs River estuary (Massachusetts, USA),which is under the influence of increased nitrogen loadsby fertilizers and wastewater has an average d15N of5.9& (McClelland et al., 1997). Because of the d15Nvalues of the Wonokromo and Porong TSM falling inthis range and the unlikeliness of the processes/sourcesdiscussed before, it appears that DIN is the majornitrogen source for the almost autochthonous sus-pended particulate OM in the Wonokromo and Porongrivers as well as in coastal waters during the dry season.
3.3. Comparison of TSM with sediments
Because of the lack of samples from the rainy season,i.e. the period of high river discharge, comparison ofbiogeochemical characteristics of TSM from the dryseason with those of surface sediments may give hints onseasonal differences of OM production, transformationand deposition and the decomposition processes in-volved. In general, estuarine sediments contain muchless OM than TSM with Corg ranging between 0.4% and
Salinity0 5 10 15 20 25 30
δ13C
org
( o /oo)
-30
-28
-26
-24
-22
-20
-18
Fig. 4. d13Corg of TSM (open circles) and sediments (filled circles) from
the Wonokromo and Porong estuaries plotted vs. salinity. Both display
statistically significant linear regressions. TSM: Y ¼ 0:25X� 26:60,
r2 ¼ 0:88, n ¼ 10; sediments: Y ¼ 0:07X� 23:26, r2 ¼ 0:55, n ¼ 10.
1.2% and an N content of 0.1% or less. The C/N ratio ishigher by a factor of 2e3, whereas AA and HA contentsare lower by about one order of magnitude andreactivity is also significantly lower in sediments thanin TSM (Table 3). The drastically lower Corg and Ncontents suggest considerable dilution by lithogenicmaterial which may result from heavily increased riverloads of suspended sediment delivered during the rainyseason (Hoekstra et al., 1989). The C/N ratio rangesbetween that of estuarine TSM, which is mainly ofplanktonic origin (s.a.), and that of the possible terres-trial source material, plant debris and soil, indicatingthat inputs of the latter contribute significantly to OMdeposition during the wet season (Fig. 5). Sedimentaryd13Corg also displays a trend of getting isotopicallyheavier with increasing salinity, however, with less vari-ability than in TSM and ending up at values betweenthose of TSM in coastal waters and those of agriculturaland mangrove soils. It indicates a substantial contribu-tion of terrestrial OM slightly decreasing seaward.
Except for TSM from the Wonokromo where fixationfrom the atmosphere appears to be the major nitrogensource, d15N generally decreases with increasing C/Nfrom estuarine TSM to plant material indicating anincreasing proportion of the nitrogen demand met byfixation from the atmosphere. Sedimentary d15N rang-ing between plankton-derived estuarine TSM and plantand soil again indicates its origin from mixed sources.Similarly, the portion of organic carbon provided byamino acids decreases with increasing C/N ratio fromestuarine TSM to plant material. It is high in freshlyproduced planktonic material in which amino acidscomprise major part of the OM whereas it can be low inmaterial which has undergone preferential degradationof these labile compounds or which are originally poorin amino acids, like plant material (e.g. Jennerjahn andIttekkot, 1997, 1999).
Lower contents of AA and HA, lower reactivity andAA-C% as well as the higher C/N ratio indicate thatsedimentary OM is more refractory than suspended OMdue to degradation at the sedimentewater interface anda substantial contribution of terrestrial OM which isoriginally poor in nitrogenous compounds (Table 4).However, AA-N% and RI of sediments are as high as insediments underlying productive marine regions like theArabian Sea (Suthhof et al., 2000) and slightly higherthan in sediments from Brazilian medium- and small-sized rivers and the inner continental shelf (Jennerjahnand Ittekkot, 1997) suggesting relative freshness anda fairly high nutritional value of sedimentary OM fromthe Wonokromo and Porong estuaries.
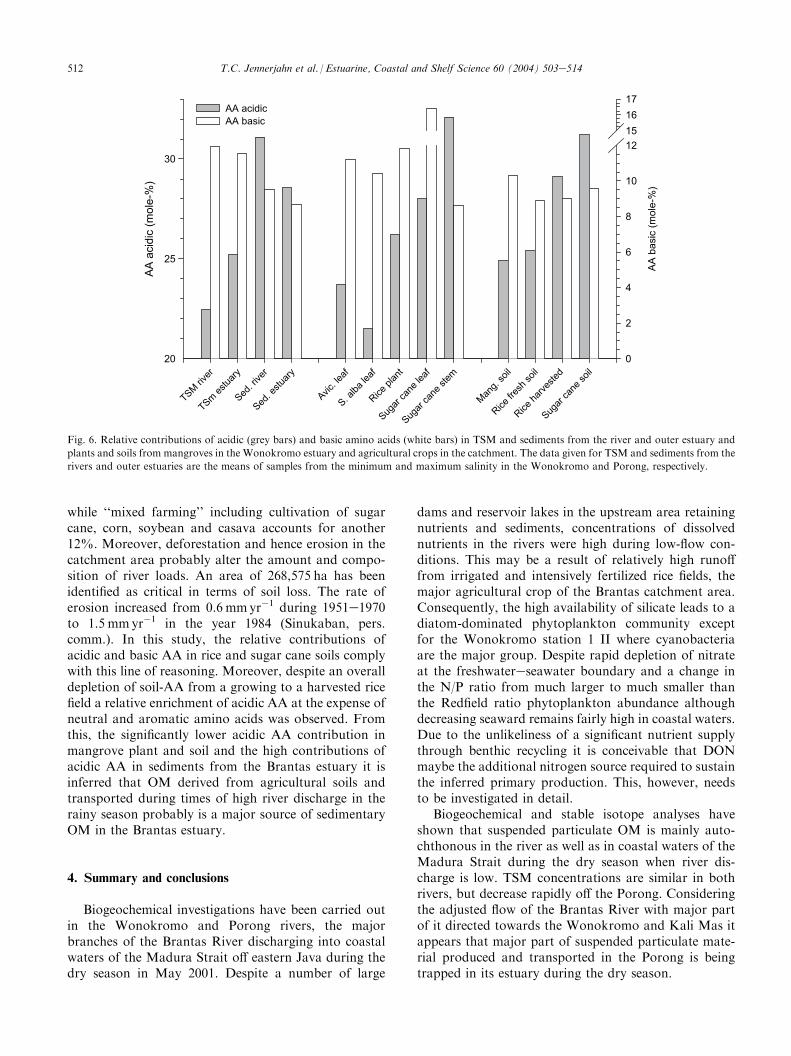
Amino acid spectra displayed significant differencesin the specific contributions of acidic and basic aminoacids between estuarine TSM and sediments and strikingsimilarities between those of sediments and the terres-trial plants and soils (Fig. 6). The distribution of AA
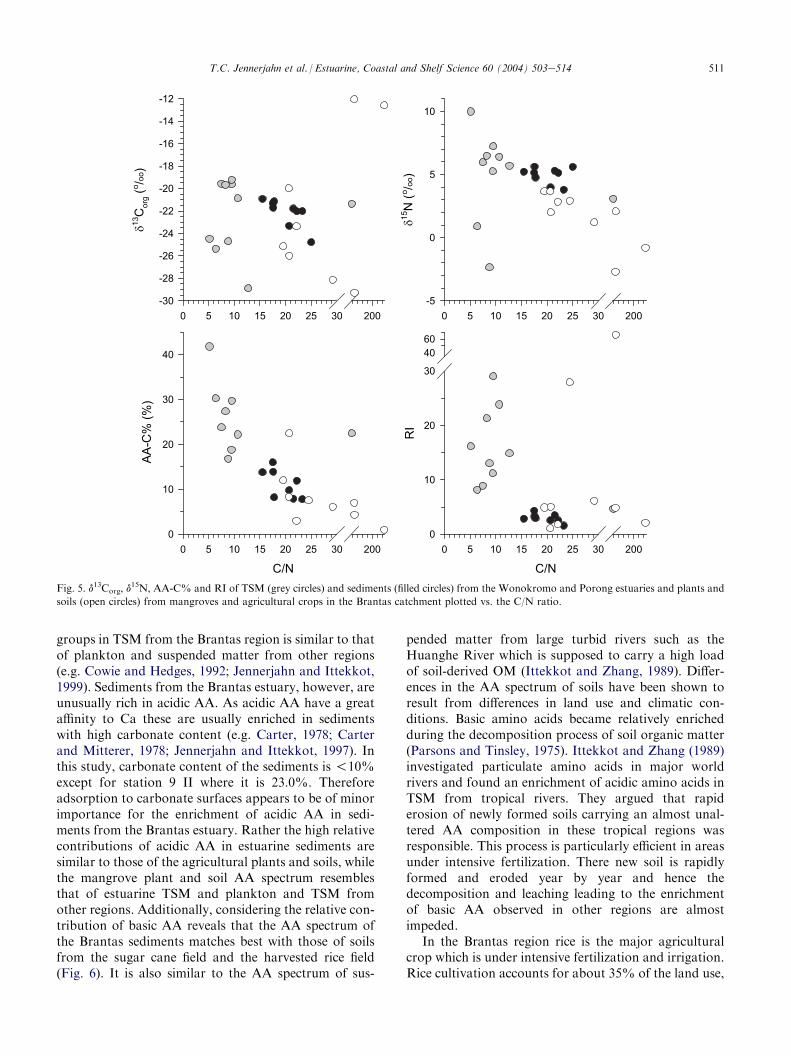
511T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
0 5 10 15 20 25 30 200
δ13C
org
( o /oo)
-30
-28
-26
-24
-22
-20
-18
-16
-14
-12
C/N0 5 10 15 20 25 30 200
AA-C
% (%
)
0
10
20
30
40
C/N0 5 10 15 20 25 30 200
RI
0
10
20
30
4060
0 5 10 15 20 25 30 200
δ15N
(o /oo)
-5
0
5
10
Fig. 5. d13Corg, d15N, AA-C% and RI of TSM (grey circles) and sediments (filled circles) from the Wonokromo and Porong estuaries and plants and
soils (open circles) from mangroves and agricultural crops in the Brantas catchment plotted vs. the C/N ratio.
groups in TSM from the Brantas region is similar to thatof plankton and suspended matter from other regions(e.g. Cowie and Hedges, 1992; Jennerjahn and Ittekkot,1999). Sediments from the Brantas estuary, however, areunusually rich in acidic AA. As acidic AA have a greataffinity to Ca these are usually enriched in sedimentswith high carbonate content (e.g. Carter, 1978; Carterand Mitterer, 1978; Jennerjahn and Ittekkot, 1997). Inthis study, carbonate content of the sediments is !10%except for station 9 II where it is 23.0%. Thereforeadsorption to carbonate surfaces appears to be of minorimportance for the enrichment of acidic AA in sedi-ments from the Brantas estuary. Rather the high relativecontributions of acidic AA in estuarine sediments aresimilar to those of the agricultural plants and soils, whilethe mangrove plant and soil AA spectrum resemblesthat of estuarine TSM and plankton and TSM fromother regions. Additionally, considering the relative con-tribution of basic AA reveals that the AA spectrum ofthe Brantas sediments matches best with those of soilsfrom the sugar cane field and the harvested rice field(Fig. 6). It is also similar to the AA spectrum of sus-
pended matter from large turbid rivers such as theHuanghe River which is supposed to carry a high loadof soil-derived OM (Ittekkot and Zhang, 1989). Differ-ences in the AA spectrum of soils have been shown toresult from differences in land use and climatic con-ditions. Basic amino acids became relatively enrichedduring the decomposition process of soil organic matter(Parsons and Tinsley, 1975). Ittekkot and Zhang (1989)investigated particulate amino acids in major worldrivers and found an enrichment of acidic amino acids inTSM from tropical rivers. They argued that rapiderosion of newly formed soils carrying an almost unal-tered AA composition in these tropical regions wasresponsible. This process is particularly efficient in areasunder intensive fertilization. There new soil is rapidlyformed and eroded year by year and hence thedecomposition and leaching leading to the enrichmentof basic AA observed in other regions are almostimpeded.
In the Brantas region rice is the major agriculturalcrop which is under intensive fertilization and irrigation.Rice cultivation accounts for about 35% of the land use,
512 T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
TSM river
TSm estua
ry
Sed. ri
ver
Sed. e
stuary
Avic. le
af
S. alba
leaf
Rice pl
ant
Sugar
cane
leaf
Sugar
cane
stem
Mang.
soil
Rice fre
sh so
il
Rice ha
rveste
d
Sugar
cane
soil
AA a
cidi
c (m
ole-
%)
20
25
30
AA b
asic
(mol
e-%
)
0
2
4
6
8
10
12151617
AA acidicAA basic
Fig. 6. Relative contributions of acidic (grey bars) and basic amino acids (white bars) in TSM and sediments from the river and outer estuary and
plants and soils from mangroves in the Wonokromo estuary and agricultural crops in the catchment. The data given for TSM and sediments from the
rivers and outer estuaries are the means of samples from the minimum and maximum salinity in the Wonokromo and Porong, respectively.
while ‘‘mixed farming’’ including cultivation of sugarcane, corn, soybean and casava accounts for another12%. Moreover, deforestation and hence erosion in thecatchment area probably alter the amount and compo-sition of river loads. An area of 268,575 ha has beenidentified as critical in terms of soil loss. The rate oferosion increased from 0.6 mmyr�1 during 1951e1970to 1.5 mmyr�1 in the year 1984 (Sinukaban, pers.comm.). In this study, the relative contributions ofacidic and basic AA in rice and sugar cane soils complywith this line of reasoning. Moreover, despite an overalldepletion of soil-AA from a growing to a harvested ricefield a relative enrichment of acidic AA at the expense ofneutral and aromatic amino acids was observed. Fromthis, the significantly lower acidic AA contribution inmangrove plant and soil and the high contributions ofacidic AA in sediments from the Brantas estuary it isinferred that OM derived from agricultural soils andtransported during times of high river discharge in therainy season probably is a major source of sedimentaryOM in the Brantas estuary.
4. Summary and conclusions
Biogeochemical investigations have been carried outin the Wonokromo and Porong rivers, the majorbranches of the Brantas River discharging into coastalwaters of the Madura Strait off eastern Java during thedry season in May 2001. Despite a number of large
dams and reservoir lakes in the upstream area retainingnutrients and sediments, concentrations of dissolvednutrients in the rivers were high during low-flow con-ditions. This may be a result of relatively high runofffrom irrigated and intensively fertilized rice fields, themajor agricultural crop of the Brantas catchment area.Consequently, the high availability of silicate leads to adiatom-dominated phytoplankton community exceptfor the Wonokromo station 1 II where cyanobacteriaare the major group. Despite rapid depletion of nitrateat the freshwatereseawater boundary and a change inthe N/P ratio from much larger to much smaller thanthe Redfield ratio phytoplankton abundance althoughdecreasing seaward remains fairly high in coastal waters.Due to the unlikeliness of a significant nutrient supplythrough benthic recycling it is conceivable that DONmaybe the additional nitrogen source required to sustainthe inferred primary production. This, however, needsto be investigated in detail.
Biogeochemical and stable isotope analyses haveshown that suspended particulate OM is mainly auto-chthonous in the river as well as in coastal waters of theMadura Strait during the dry season when river dis-charge is low. TSM concentrations are similar in bothrivers, but decrease rapidly off the Porong. Consideringthe adjusted flow of the Brantas River with major partof it directed towards the Wonokromo and Kali Mas itappears that major part of suspended particulate mate-rial produced and transported in the Porong is beingtrapped in its estuary during the dry season.

513T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
Biogeochemical characteristics of sedimentary OM,however, reveal an increased proportion of allochtho-nous material probably derived from upstream agricul-tural soils and the mangroves bordering the estuaries.Apparently, major part of this material particularly richin amino acid bound nitrogen must have been depositedduring the wet season. Also, the anthropogenic alter-ation of the flow regime of the river with almost cuttingoff discharge from the Porong during the dry season anddiverting about 80% of total flow through it during therainy season amplify the seasonal variation of OMproduction and deposition in coastal waters which, inturn, may affect the coastal food web and habitatstructure.
The Porong is known to have a huge sediment loadduring the wet season which resulted in the formation ofa major delta during the last century (Hoekstra, 1989;Hoekstra et al., 1989). This and the significant differ-ences in biogeochemical characteristics of dry seasonTSM and surface sediments suggest stronger fluvial ex-port of dissolved and particulate substances during therainy season which, however, needs to be investigated infurther studies.
The observed high amount and reactivity of aminoacid-bound nitrogen in TSM and sediments from theBrantaseMadura Strait region and the potential rele-vance of DON for the nutrition of river and coastalwaters suggest that organic nitrogen may play a moreimportant role for coastal food webs and the nitrogencycle in tropical regions than previously thought,particularly under conditions of high agricultural runoffand urban waste disposal.
Acknowledgements
This project is being carried out in the frame of theGermaneIndonesian cooperation in the field of marinescience. We thank Didik Agus Wijanarko and his staffat the Water Quality Laboratory in Surabaya forsupport in the field, the Jasatirta Public Corporationfor support in the field and in compiling discharge data,Mark Wunsch and Hermann Behling for help andexpertise during the expedition and Matthias Birkichtand Dorothee Dasbach for laboratory work. Financialsupport by the German Federal Ministry for Educationand Science (Grant No. 03F0301A) and the IndonesianAgency for the Assessment and Application of Tech-nology is gratefully acknowledged.
References
Abril, G., Nogueira, M., Etcheber, H., Cabecxadas, G., Lemaire, E.,
Brogueira, M.J., 2002. Behaviour of organic carbon in nine
contrasting European estuaries. Estuarine, Coastal and Shelf
Science 254, 241e262.
Berg, G.M., Gilbert, P.M., Jørgensen, N.O.G., Balode, M., Purina, I.,
2001. Variability in inorganic and organic nitrogen uptake
associated with riverine nutrient input in the Gulf of Riga, Baltic
Sea. Estuaries 24, 204e214.
Carpenter, E.J., Montoya, J.P., Burns, J., Mulholland, M.R.,
Subramanian, A., Capone, D.G., 1999. Extensive bloom of a N2-
fixing diatom/cyanobacterial association in the tropical Atlantic
Ocean. Marine Ecology Progress Series 185, 273e283.
Carter, P.W., 1978. Adsorption of amino acid-containing organic
matter by calcite and quartz. Geochimica et Cosmochimica Acta
42, 1239e1242.
Carter, P.W., Mitterer, R.M., 1978. Amino acid composition of
organic matter associated with carbonate and non-carbonate sedi-
ments. Geochimica et Cosmochimica Acta 42, 1231e1238.Conley, D.J., Schelske, C.L., Stoermer, E.F., 1993. Modification of the
biogeochemical cycle of silica with eutrophication. Marine Ecology
Progress Series 101, 179e192.Cowie, G.L., Hedges, J.I., 1992. Sources and reactivities of amino
acids in a coastal marine environment. Limnology and Oceanog-
raphy 37, 703e724.
Elvidge, C.D., Baugh, K.E., Kihn, E.A., Kroehl, H.W., Davis, E.R.,
1997. Mapping city lights with nighttime data from the DMSP
operational linescan system. Photogrammetric Engineering &
Remote Sensing 63, 727e734.
Erftemeijer, P., Swennen, C., 1990. Densities and biomass of macro-
benthic fauna of some intertidal areas in Java, Indonesia.
Wallaceana 59, 60, 1e6.
Fischer, G., 1991. Stable carbon isotope ratios of plankton carbon and
sinking organic matter from the Atlantic sector of the Southern
Ocean. Marine Chemistry 35, 581e596.
Galimov, E.M., 1985. The Biological Fractionation of Isotopes.
Academic Press, Orlando, 261 pp.
Grasshoff, K., Kremling, K., Ehrhardt, M. (Eds.), 1999. Methods of
Seawater Analysis, third ed. Wiley-VCH, Weinheim, 600 pp.
Hasegawa, T., Koike, I., Mukai, H., 2001. Release of dissolved organic
nitrogen by a planktonic community in Akkeshi Bay. Aquatic
Microbial Ecology 24, 99e107.
Hoekstra, P., 1989. Hydrodynamics and depositional processes of the
Solo and Porong Deltas, East Java, Indonesia. Proceedings of the
KNGMG Symposium ‘‘Coastal Lowlands, Geology and Geo-
technology’’ 1987. Kluwer, Dordrecht, pp. 161e173.
Hoekstra, P., Nolting, R.F., van der Sloot, H.A., 1989. Supply and
dispersion of water and suspended matter of the rivers Solo and
Brantas into the coastal waters of east Java, Indonesia. Nether-
lands Journal of Sea Research 23, 501e515.
Holmes, M.E., Muller, P.J., Schneider, R.R., Segl, M., Wefer, G.,
1998. Spatial variations in euphotic zone nitrate utilization based
on d15N in surface sediments. Geo-Marine Letters 18, 58e65.
Humborg, C., Ittekkot, V., Cociasu, A., Bodungen, B.v., 1997. Effect
of Danube River dam on Black Sea biogeochemistry. Nature 386,
385e388.
Ittekkot, V., Zhang, S., 1989. Pattern of particulate nitrogen transport
in world rivers. Global Biogeochemical Cycles 3, 383e391.
Jennerjahn, T.C., Ittekkot, V., 1997. Organic matter in sediments
in the mangrove areas and adjacent continental margins of
Brazil: I. Amino acids and hexosamines. Oceanologica Acta 20,
359e369.
Jennerjahn, T.C., Ittekkot, V., 1999. Changes in organic matter from
surface waters to continental slope sediments off the Sao Francisco
River, eastern Brazil. Marine Geology 161, 129e140.
Liu, K.-K., Kaplan, I.R., 1989. The eastern tropical Pacific as a source
of 15N-enriched nitrate in seawater off southern California.
Limnology and Oceanography 34, 820e830.
McClelland, J.W., Valiela, I., Michener, R.H., 1997. Nitrogen-stable
isotope signatures in estuarine food webs: a record of increasing

514 T.C. Jennerjahn et al. / Estuarine, Coastal and Shelf Science 60 (2004) 503e514
urbanization in coastal watersheds. Limnology and Oceanography
42, 930e937.
Meybeck, M., 1998. The IGBP water group: a response to a growing
global concern. Global Change Newsletters 36, 8e12.Middelburg, J.J., Nieuwenhuize, J., 2001. Nitrogen isotope tracing of
dissolved inorganic nitrogen behaviour in tidal estuaries. Estuarine,
Coastal and Shelf Science 53, 385e391.
Milliman, J.D., 1997. Blessed dams or damned dams? Nature 386,
325e327.
Milliman, J.D., Syvitski, J.P.M., 1992. Geomorphic/tectonic control
of sediment discharge to the ocean: the importance of small
mountainous rivers. Journal of Geology 100, 525e544.Milliman, J.D., Rutkowski, C., Meybeck, M., 1995. River discharge to
the sea: a global river index. LOICZ Report, Texel, 125 pp.
Mortlock, R.A., Froelich, P.N., 1989. A simple method for the rapid
determination of biogenic opal in pelagic marine sediments. Deep-
Sea Research 36, 1415e1426.
Muller, P.J., Suess, E., Ungerer, C.A., 1986. Amino acids and amino
sugars of surface particulate and sediment trap material from
waters of the Scotia Sea. Deep-Sea Research 33, 819e838.
Ostrom, N.E., Knoke, K.E., Hedin, L.O., Robertson, G.P., Smucker,
A.J.M., 1998. Temporal trends in nitrogen isotope values of nitrate
leaching from an agricultural soil. Chemical Geology 146, 219e227.
Parsons, J.W., Tinsley, J., 1975. Nitrogenous substances. In: Giesek-
ing, J.E. (Ed.), Organic Components. Soil Components, vol. 1.
Springer-Verlag, New York, pp. 263e304.
Rabalais, N., 1999. Changes in Mississippi River nutrient fluxes and
consequences for the northern Gulf of Mexico coastal ecosystem.
LOICZ Newsletter 13, 1e4.
Roth, M., Hampai, A., 1973. Column chromatography of amino
acids with fluorescence detection. Journal of Chromatography 83,
353e356.
Schafer, P., Ittekkot, V., 1993. Seasonal variability of 15N in settling
particles in the Arabian Sea and its paleogeochemical significance.
Naturwissenschaften 80, 511e513.
Seitzinger, S.P., Sanders, R.W., Styles, R.V., 2002. Bioavailability
of DON from natural and anthropogenic sources to estuarine
plankton. Limnology and Oceanography 47, 353e366.
Siezen, R.J., Mague, T.H., 1978. Amino acids in suspended particulate
matter from oceanic and coastal waters of the Pacific. Marine
Chemistry 6, 215e231.
Sigleo, A.C., Shultz, D.J., 1993. Amino acid composition of suspended
particles, sediment-trap material, and benthic sediment in the
Potomac estuary. Estuaries 16, 405e415.Smith, S.V., Swaney, D.P., Talaue-McManus, L., Bartley, J.D.,
Sandhei, P.T., McLaughlin, C.J., Dupra, V.C., Crossland, C.J.,
Buddemeier, R.W., Maxwell, B.A., Wulff, F., 2003. Humans,
hydrology, and the distribution of inorganic nitrogen loading to
the ocean. BioScience 53, 235e245.
Stewart, G.R., Aidar, M.P.M., Joly, C.A., Schmidt, S., 2002. Impact
of point source pollution on nitrogen isotope signatures (d15N) of
vegetation in SE Brazil. Oecologia 131, 468e472.
Suthhof, A., Jennerjahn, T.C., Schafer, P., Ittekkot, V., 2000. Nature
of organic matter in surface sediments from the Pakistan conti-
nental margin and the deep Arabian Sea: amino acids. Deep-Sea
Research II 47, 329e351.
Verardo, D.J., Froelich, P.N., McIntyre, A., 1990. Determination of
organic carbon and nitrogen in marine sediments using the Carlo
Erba NA-1500 Analyzer. Deep-Sea Research 37, 157e165.Voss, M., Dippner, J.W., Montoya, J.P., 2001. Nitrogen isotope
patterns in the oxygen-deficient waters of the Eastern Tropical
North Pacific Ocean. Deep-Sea Research I 48, 1905e1921.Wada, E., Hattori, A., 1990. Nitrogen in the Sea: Forms, Abundances,
and Rate Processes. CRC Press, Boca Raton, 224 pp.




















