
Biochemical composition and size distribution of organic matter at the Pacific and Atlantic...
15
Biochemical composition and size distribution of organic matter at the Pacific and Atlantic time-series stations Karl Kaiser ⁎, Ronald Benner University of South Carolina, Marine Science Program, Columbia, SC 29208, United States abstract article info Article history: Received 9 June 2008 Received in revised form 19 November 2008 Accepted 8 December 2008 Available online 24 December 2008 Keywords: Marine detrital organic matter Amino acids Amino sugars Neutral sugars Size distribution Size-reactivity-composition continuum Time-series stations BATS and HOT Amino acids, neutral sugars and amino sugars were analyzed to investigate the chemical composition and diagenetic processing of suspended particulate organic matter (POM, N100 nm), high-molecular-weight dissolved organic matter (HMW DOM, 1–100 nm) and low-molecular-weight dissolved organic matter (LMW DOM, b1 nm) at the time-series stations near Bermuda (BATS) and Hawaii (HOT). Differences between BATS and HOT were principally related to location-specific biogeochemical processes and water mass ventilation ages. Concentrations of amino acids, neutral sugar and amino sugars in unfiltered seawater sharply declined with depth at both stations, indicating an upper ocean source and rapid turnover of these components. The size distribution of organic matter was heavily skewed to smaller molecular sizes. Depth comparisons showed that larger size classes of organic matter were more efficiently removed than smaller size classes. Carbon-normalized yields of amino acids, neutral sugars and amino sugars decreased rapidly with depth and molecular size. Together these biochemicals accounted for 55% of organic carbon in surface POM but only 2% of the organic carbon in LMW DOM in deep water. Chemical compositions showed distinct differences between organic matter size classes indicating the extent of diagenetic processing increased with decreasing molecular size. These findings are consistent with the size-reactivity continuum model for organic matter in which bioreactivity decreases with decreasing molecular size and diagenetic processes lead to the formation of smaller components that are resistant to biodegradation. The data also provided evidence for a size-composition continuum. Carbon-normalized yields of amino acids, neutral sugars and amino sugars were sensitive indicators of diagenetic alterations. Diagenetic indicators based on amino acid compositions revealed distinct patterns for the North Pacific and Sargasso Sea possibly indicating the influence of varying sources or diagenetic processing. © 2008 Elsevier B.V. All rights reserved. 1. Introduction One of the primary goals of the US Joint Global Ocean Flux Study Program (JGOFS) was to better understand the cycling of organic carbon and associated bioreactive elements in the ocean. Long-term time-series stations established near Bermuda (BATS) and Hawaii (HOT) led to numerous significant discoveries of the ocean's role in the global carbon cycle and its response to anthropogenic perturbations (Karl and Michaels, 1996; Karl, 1999; Steinberg et al., 2001). JGOFS studies demonstrated the importance of dissolved organic matter (DOM) for the microbial food web and carbon export flux (Carlson et al., 1994; Karl, 1999), but few studies explored the sources, chemical composition and diagenetic processing of DOM at BATS and HOT. Studies of sinking particles in the surface ocean indicate that amino acids, carbohydrates, and lipids account for most of the organic carbon. By the time sinking particles reach the sediment interface in the deep ocean these biochemicals become minor components of organic matter (Wakeham et al., 1997; Hedges et al., 2001). In marine DOM, common biochemicals that dominate living biomass always comprise much less than half of the organic carbon and nitrogen (Benner, 2002). Comparisons reveal that the composition of sinking particles is dominated by amino acids (Wakeham et al., 1997; Hedges et al., 2001), whereas carbohydrates are the most abundant characterizable bio- chemicals in DOM (McCarthy et al., 1996; Benner, 2002). The remaining unrecognizable components exhibit functional groups typical of biopolymers in living organisms, but they are not identified by compound-specific analyses (Benner et al., 1992; McCarthy et al., 1997; Hedges et al., 2000a; Sannigrahi et al., 2005). The lack of knowledge about molecular composition greatly reduces our understanding of the sources and reaction potential of marine organic matter. Given that about 50% of oceanic primary production is routed through the dissolved organic matter (DOM) pool, most marine organic matter is rapidly consumed. The fraction that escapes immediate remineralization remains in the ocean and cycles on timescales of months to millennia (Nagata, 2000; Carlson, 2002). Numerous studies have shown that substrate quality and composition play an important role in the recycling of organic matter. Nitrogen- and phosphorus- containing organic compounds are typically more quickly remineralized Marine Chemistry 113 (2009) 63–77 ⁎ Corresponding author. Tel.: +1 803 777 1017; fax: +1 803 777 4002. E-mail address: [email protected] (K. Kaiser). 0304-4203/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.marchem.2008.12.004 Contents lists available at ScienceDirect Marine Chemistry journal homepage: www.elsevier.com/locate/marchem
Transcript of Biochemical composition and size distribution of organic matter at the Pacific and Atlantic...

Marine Chemistry 113 (2009) 63–77
Contents lists available at ScienceDirect
Marine Chemistry
j ourna l homepage: www.e lsev ie r.com/ locate /marchem
Biochemical composition and size distribution of organic matter at the Pacific andAtlantic time-series stations
Karl Kaiser ⁎, Ronald BennerUniversity of South Carolina, Marine Science Program, Columbia, SC 29208, United States
⁎ Corresponding author. Tel.: +1 803 777 1017; fax: +1E-mail address: [email protected] (K. Kaiser).
0304-4203/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.marchem.2008.12.004
a b s t r a c t
a r t i c l e i n f oArticle history:
Amino acids, neutral sugars Received 9 June 2008Received in revised form 19 November 2008Accepted 8 December 2008Available online 24 December 2008Keywords:Marine detrital organic matterAmino acidsAmino sugarsNeutral sugarsSize distributionSize-reactivity-composition continuumTime-series stations BATS and HOT
and amino sugars were analyzed to investigate the chemical composition anddiageneticprocessingof suspendedparticulate organicmatter (POM, N100nm), high-molecular-weight dissolvedorganicmatter (HMWDOM,1–100 nm) and low-molecular-weight dissolved organicmatter (LMWDOM, b1 nm)at the time-series stations near Bermuda (BATS) and Hawaii (HOT). Differences between BATS and HOT wereprincipally related to location-specific biogeochemical processes andwatermass ventilation ages. Concentrationsof amino acids, neutral sugar and amino sugars in unfiltered seawater sharply declined with depth at bothstations, indicating an upper ocean source and rapid turnover of these components. The size distribution oforganicmatterwas heavily skewed to smallermolecular sizes. Depth comparisons showed that larger size classesof organic matter were more efficiently removed than smaller size classes. Carbon-normalized yields of aminoacids, neutral sugars and amino sugars decreased rapidly with depth and molecular size. Together thesebiochemicals accounted for 55% of organic carbon in surface POMbut only 2% of the organic carbon in LMWDOMin deepwater. Chemical compositions showed distinct differences between organicmatter size classes indicatingthe extent of diagenetic processing increased with decreasing molecular size. These findings are consistent withthe size-reactivity continuum model for organic matter in which bioreactivity decreases with decreasingmolecular size and diagenetic processes lead to the formation of smaller components that are resistant tobiodegradation. The data also provided evidence for a size-composition continuum. Carbon-normalized yields ofamino acids, neutral sugars and amino sugars were sensitive indicators of diagenetic alterations. Diageneticindicators based on amino acid compositions revealed distinct patterns for the North Pacific and Sargasso Seapossibly indicating the influence of varying sources or diagenetic processing.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
One of the primary goals of the US Joint Global Ocean Flux StudyProgram (JGOFS) was to better understand the cycling of organic carbonand associatedbioreactive elements in the ocean. Long-term time-seriesstations established near Bermuda (BATS) and Hawaii (HOT) led tonumerous significant discoveries of the ocean's role in the global carboncycle and its response to anthropogenic perturbations (Karl andMichaels, 1996; Karl, 1999; Steinberg et al., 2001). JGOFS studiesdemonstrated the importance of dissolved organic matter (DOM) forthemicrobial foodweb and carbon export flux (Carlson et al.,1994; Karl,1999), but few studies explored the sources, chemical composition anddiagenetic processing of DOM at BATS and HOT.
Studies of sinking particles in the surface ocean indicate that aminoacids, carbohydrates, and lipids account for most of the organic carbon.By the time sinking particles reach the sediment interface in the deepocean these biochemicals becomeminor components of organic matter
803 777 4002.
l rights reserved.
(Wakeham et al., 1997; Hedges et al., 2001). In marine DOM, commonbiochemicals that dominate living biomass always comprise much lessthan half of the organic carbon and nitrogen (Benner, 2002).Comparisons reveal that the composition of sinking particles isdominated by amino acids (Wakeham et al., 1997; Hedges et al., 2001),whereas carbohydrates are the most abundant characterizable bio-chemicals in DOM (McCarthy et al., 1996; Benner, 2002). The remainingunrecognizable components exhibit functional groups typical ofbiopolymers in living organisms, but they are not identified bycompound-specific analyses (Benner et al., 1992; McCarthy et al., 1997;Hedges et al., 2000a; Sannigrahi et al., 2005). The lack of knowledgeabout molecular composition greatly reduces our understanding of thesources and reaction potential of marine organic matter.
Given that about 50% of oceanic primary production is routedthrough the dissolved organic matter (DOM) pool, most marine organicmatter is rapidly consumed. The fraction that escapes immediateremineralization remains in the ocean and cycles on timescales ofmonths to millennia (Nagata, 2000; Carlson, 2002). Numerous studieshave shown that substrate quality and composition play an importantrole in the recycling of organic matter. Nitrogen- and phosphorus-containingorganic compounds are typicallymore quickly remineralized

Table 1Concentrations of total organic carbon (TOC) in unfiltered seawater and concentrationsof particulate organic carbon (POC) and high-molecular-weight dissolved organiccarbon (HMW DOC) recovered by ultrafiltration at Stations HOT and BATS
Location Depth TOC POC HMW DOC
(m) (µmol L−1) (µmol L−1) (µmol L−1)
HOT 20 83 1.0 16.680 83 1.1 17.8110 78 2.4 17.6150 63 0.7 13.5200 62 0.3 9.5250 51 0.4 7.3300 53 0.4 8.4500 40 0.7 7.4750 41 0.5 7.7
2000 36 0.3 5.94000 38 0.7 5.8
BATS 20 70 0.3 15.9100 60 0.2 12.1350 53 0.3 11.0500 54 0.1 nda
1360 46 0.1 12.52000 46 0.1 12.13250 43 0.1 10.74300 45 0.2 12.3
a nd, not determined.
64 K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
than other organic molecules. Carbohydrates and amino acids arepreferentially utilized during microbial decomposition of marineorganic matter (Skoog and Benner, 1997; Lee et al., 2000; Amon et al.,2001) and serve as useful indicators of diagenetic history (Cowie andHedges, 1994; Dauwe andMiddelburg,1998). Size-fractionation experi-ments lead Amon and Benner (1994, 1996) to postulate a size-reactivitycontinuum model, which provides a framework for understanding thecycling of organicmatter in the ocean. In this conceptual framework, thebioreactivity of organic matter decreases along a continuum of larger tosmaller sizes. The model implies that degradation reactions lead to theformation of low-molecular-weight (LMW) compounds that areresistant to biodegradation. Several studies have confirmed such asize–reactivity relationship of organic matter in the ocean (Harvey andMannino, 2001; Hama et al., 2004; Zou et al., 2004; Loh et al., 2004).
In this study, we present a comprehensive analysis of majorbiochemicals (amino acids, neutral sugars, and amino sugars) in oceanprofiles at the time-series stations BATS and HOT. In addition, thesebiochemicals were analyzed in various size fractions of marine organicmatter to investigate its chemical composition and diagenetic proces-sing. The results confirm a relationship between size and reactivityanalogous to that proposed by Amon and Benner (1994, 1996) and alsoindicate a size-composition continuum. Furthermore, variousdiageneticindicators were evaluated for suspended particulate organic matter(POM) and DOM in the ocean. The results indicate that carbon-normalizedyieldsofmajor biomolecules are themost reliable diageneticindicators, whereas indicators based on amino acid compositions maybe sensitive to sources or variable microbial processing.
2. Materials and methods
2.1. Sample collection and isolation
Samples were collected at the HOT site (Station ALOHA) and atBATS during cruises on the RV Kai-ma-kai O'Kanaloa in November1999 and on the R/V Cape Hatteras in June 2001 with Niskin bottlesmounted on a rosette with a CTD. Samples for total organic carbon(TOC), total hydrolyzable amino acids (THAA), total hydrolyzableamino sugars (THAS), and total hydrolyzable neutral sugars (THNS)samples were directly collected from Niskin bottles into acid-washed60 mL HDPE bottles and immediately frozen (20 °C) until analysis.Different size fractions of organic matter were isolated by ultrafiltra-tion aboard ship as described by Benner et al. (1997). Water wasdrained from Niskin bottles with acid-washed silicone tubing andfiltered through a Nitex screen (60 µm) to remove large particle. Theprocessed volumes were 1000 L at HOT and 600 L at BATS. The POMfraction (0.1–60 µm) was collected using an Amicon DC10L systemwith a polysulfone hollow-fiber membrane (H5MP01). The remainingfiltrate was continuously fed into an Amicon DC30L system with ninepolysulfonemembranes (S10N1) with a nominal pore size of 1 nm anda molecular weight cutoff of 1000 Da. High-molecular-weight (HMW)retentates (1–100 nm) were desalted by diafiltration with deionizedwater and frozen (20 °C). Carbon loss during diafiltration is typicallyless than 5% at the high concentration factors used in this study(Benner et al., 1997). The LMW filtrate (b1 nm) was subsampled foranalyses of dissolved organic carbon (DOC) and biochemicals. TheHMW fraction was dried in a Savant Speed-Vac evaporator uponreturn to the laboratory.
2.2. Analyses
Concentrations of total and dissolved organic carbon (TOC andDOC) were determined using a high-temperature-combustionmethod and a Shimadzu TOC 5000 or Shimadzu TOC-V analyzer.The organic carbon and nitrogen contents of dried POM and HMWDOM samples were determined after vapor-phase acidification using aCarlo Erba 1108 CHN analyzer.
Amino acids were analyzed as o-phthaldialdehyde (OPA) derivatives(Lindroth and Mopper, 1979) after hydrolysis in 6 M HCl on a HP1100liquid chromatography system. Samples were hydrolyzed either with amicrowave-assisted vapor phase technique (Kaiser and Benner, 2005) orwith 6 M HCl in sealed ampoules for 20 h at 110 °C (Henrichs, 1991). ALicrospher 100RP18 (5 µm, 4.5×250mm) columnwith guard columnat20 °C and a flow rate of 0.9 ml min−1 was used for separation. Aminoacids were separated using a linear gradient starting with 77% sodiumacetate (29 mmol L−1, pH=6.18) and 23% methanol to 29% methanol at4 min, 44% methanol at 20 min and 77% methanol at 33 min. Excitationand emission wavelengths for the detection of fluorescent OPAderivatives were set at 330 and 450 nm, respectively. Asparagine andglutamine were deaminated during the hydrolysis and were quantifiedas aspartic acid (Asx) and glutamic acid (Glx). Amino acid concentrationswere compared with results from D- and L-amino acid analyses (Kaiserand Benner, 2008) performed on identical samples and concentrationswere typically within ±8%. Glutamic acid (Glx) and arginine (Arg) weregenerally 2-fold lower with the D/L-amino acid method due to a co-eluting peakwith Glx andArg. Therefore, concentrations forGlx andArgdetermined by the D/L-amino acid method were used here.
Concentrations of galactosamine (GalN), mannosamine (ManN)and glucosamine (GlcN), were determined by high-performanceanion-exchange chromatography coupled to a pulsed amperometricdetector (PAD, Kaiser and Benner, 2000). Samples were neutralizedwith a self-absorbed ion retardation resin after hydrolysis in 3 M HCl(5 h, 100 °C) and frozen until analysis. Samples were desalted by solid-phase extraction using Biorad's AG50 X8 resin in the Na+-form beforechromatographic separation. GalN, ManN, and GlcN were separatedisocratically on a Dionex PA20 anion-exchange column with 2 mMNaOH at a flow rate of 1 ml min−1.
Neutral sugars were analyzed according to Skoog and Benner (1997)with modifications. Briefly, samples were hydrolyzed in 1.2 mol L−1
sulfuric acid and neutralized with a self-absorbed ion retardation resin(Kaiser and Benner, 2000). After desalting with a mixture of cation andanion exchange resins, neutral sugars were isocratically separated with25 mM NaOH on a PA 1 column in a Dionex 500 system with a PAD.Detector settings were analogous to Skoog and Benner (1997).
All analyses were performed in duplicate or triplicate. Precision forTOC and DOC analyses was 5–11%. Precision for all biochemicalmeasurements was 2–8% in the 50–600 nmol L−1 range and 4–15% inthe 2–50 nmol L−1 range.
65K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
2.3. Mass balance calculations
At HOT, concentrations of biochemicals and TOC were determinedin samples of unfiltered seawater, POM, HMW DOM, and LMW DOM.At BATS, concentrations of biochemicals and TOC were measured insamples of unfiltered seawater, POM, and HMW DOM. Calculations atHOT showed that the sum of neutral sugar and amino sugarconcentrations determined in POM, HMW DOM and LMW DOMwere within ±1–23% of concentrations measured in unfilteredseawater. For amino acids, the sum of POM, HMW DOM, and LMWDOM concentrations was 11–95% higher than those in unfilteredseawater concentrations. This indicated that some LMWDOM isolateswere likely contaminatedwith free amino acids during the handling ofthese large volumes of water. Therefore, reported concentrations ofbiochemicals in LMW DOM were calculated by subtracting POM andHMW DOM concentrations from concentrations determined inunfiltered seawater.
3. Results
3.1. Organic carbon concentration and distribution
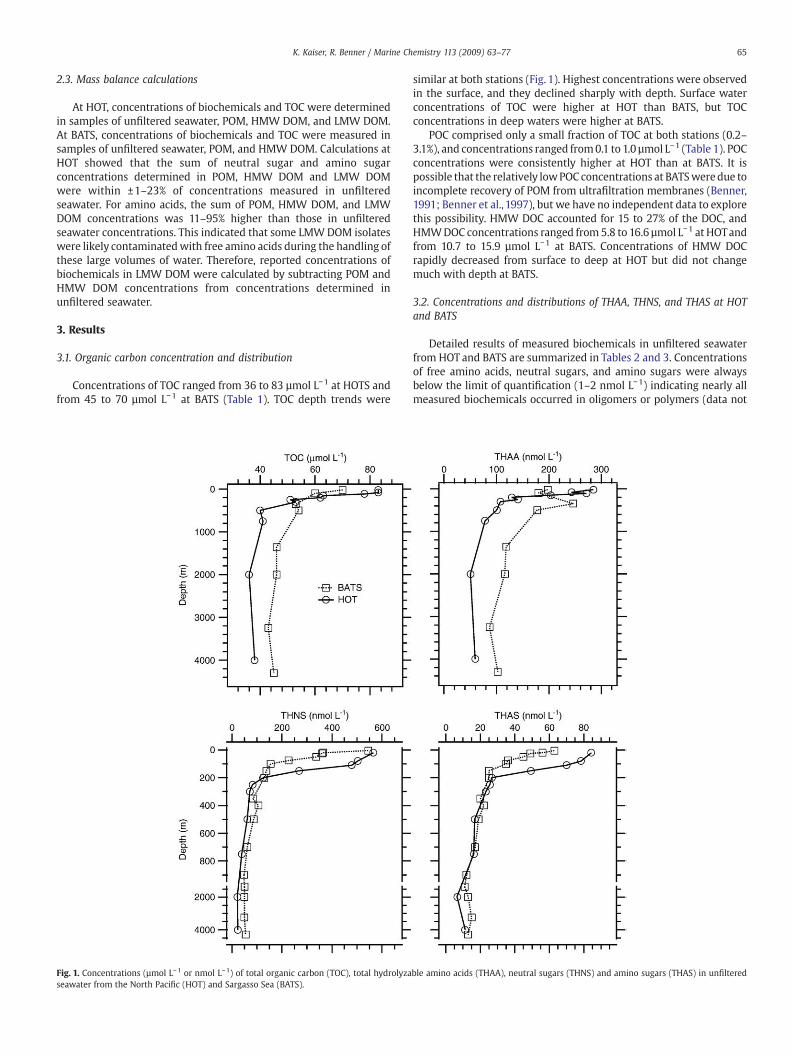
Concentrations of TOC ranged from 36 to 83 µmol L−1 at HOTS andfrom 45 to 70 µmol L−1 at BATS (Table 1). TOC depth trends were
Fig. 1. Concentrations (µmol L–1 or nmol L–1) of total organic carbon (TOC), total hydrolyzaseawater from the North Pacific (HOT) and Sargasso Sea (BATS).
similar at both stations (Fig. 1). Highest concentrations were observedin the surface, and they declined sharply with depth. Surface waterconcentrations of TOC were higher at HOT than BATS, but TOCconcentrations in deep waters were higher at BATS.
POC comprised only a small fraction of TOC at both stations (0.2–3.1%), and concentrations ranged from0.1 to 1.0 µmol L−1 (Table 1). POCconcentrations were consistently higher at HOT than at BATS. It ispossible that the relatively lowPOC concentrations at BATSweredue toincomplete recovery of POM from ultrafiltration membranes (Benner,1991; Benner et al., 1997), but we have no independent data to explorethis possibility. HMW DOC accounted for 15 to 27% of the DOC, andHMWDOC concentrations ranged from 5.8 to 16.6 µmol L−1 at HOTandfrom 10.7 to 15.9 µmol L−1 at BATS. Concentrations of HMW DOCrapidly decreased from surface to deep at HOT but did not changemuch with depth at BATS.
3.2. Concentrations and distributions of THAA, THNS, and THAS at HOTand BATS
Detailed results of measured biochemicals in unfiltered seawaterfrom HOT and BATS are summarized in Tables 2 and 3. Concentrationsof free amino acids, neutral sugars, and amino sugars were alwaysbelow the limit of quantification (1–2 nmol L−1) indicating nearly allmeasured biochemicals occurred in oligomers or polymers (data not
ble amino acids (THAA), neutral sugars (THNS) and amino sugars (THAS) in unfiltered
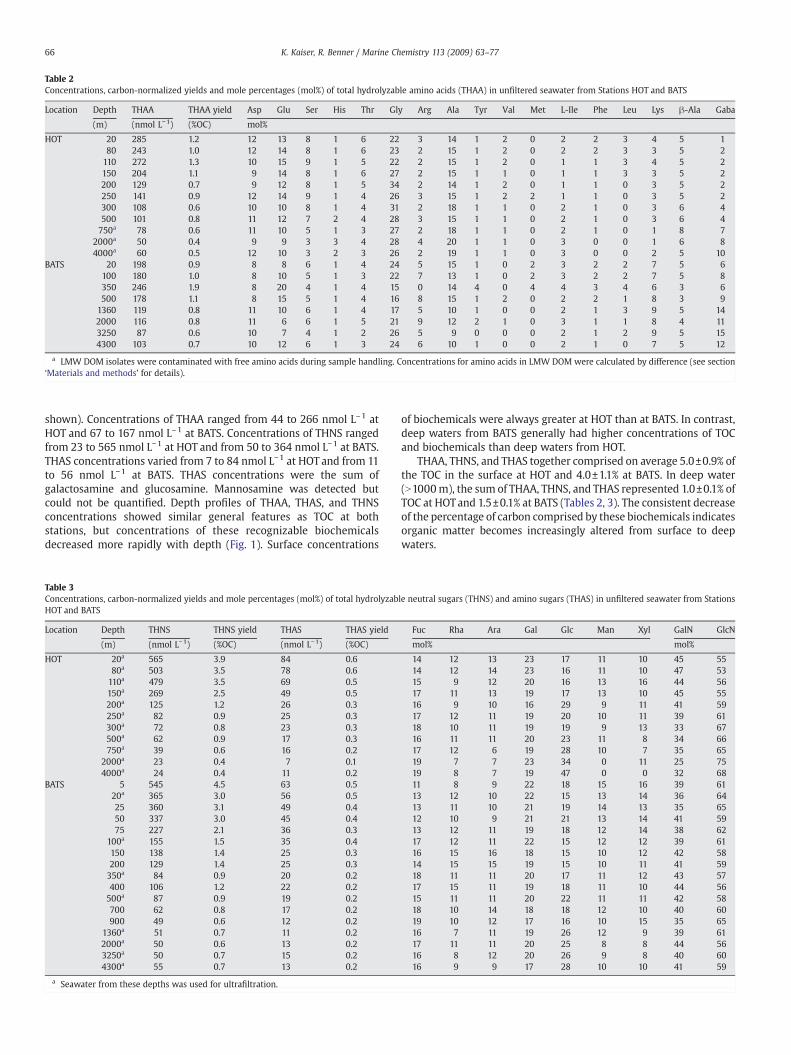
Table 2Concentrations, carbon-normalized yields and mole percentages (mol%) of total hydrolyzable amino acids (THAA) in unfiltered seawater from Stations HOT and BATS
Location Depth THAA THAA yield Asp Glu Ser His Thr Gly Arg Ala Tyr Val Met L-Ile Phe Leu Lys β-Ala Gaba
(m) (nmol L−1) (%OC) mol%
HOT 20 285 1.2 12 13 8 1 6 22 3 14 1 2 0 2 2 3 4 5 180 243 1.0 12 14 8 1 6 23 2 15 1 2 0 2 2 3 3 5 2110 272 1.3 10 15 9 1 5 22 2 15 1 2 0 1 1 3 4 5 2150 204 1.1 9 14 8 1 6 27 2 15 1 1 0 1 1 3 3 5 2200 129 0.7 9 12 8 1 5 34 2 14 1 2 0 1 1 0 3 5 2250 141 0.9 12 14 9 1 4 26 3 15 1 2 2 1 1 0 3 5 2300 108 0.6 10 10 8 1 4 31 2 18 1 1 0 2 1 0 3 6 4500 101 0.8 11 12 7 2 4 28 3 15 1 1 0 2 1 0 3 6 4750a 78 0.6 11 10 5 1 3 27 2 18 1 1 0 2 1 0 1 8 7
2000a 50 0.4 9 9 3 3 4 28 4 20 1 1 0 3 0 0 1 6 84000a 60 0.5 12 10 3 2 3 26 2 19 1 1 0 3 0 0 2 5 10
BATS 20 198 0.9 8 8 6 1 4 24 5 15 1 0 2 3 2 2 7 5 6100 180 1.0 8 10 5 1 3 22 7 13 1 0 2 3 2 2 7 5 8350 246 1.9 8 20 4 1 4 15 0 14 4 0 4 4 3 4 6 3 6500 178 1.1 8 15 5 1 4 16 8 15 1 2 0 2 2 1 8 3 91360 119 0.8 11 10 6 1 4 17 5 10 1 0 0 2 1 3 9 5 142000 116 0.8 11 6 6 1 5 21 9 12 2 1 0 3 1 1 8 4 113250 87 0.6 10 7 4 1 2 26 5 9 0 0 0 2 1 2 9 5 154300 103 0.7 10 12 6 1 3 24 6 10 1 0 0 2 1 0 7 5 12
a LMW DOM isolates were contaminated with free amino acids during sample handling. Concentrations for amino acids in LMW DOM were calculated by difference (see section‘Materials and methods’ for details).
66 K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
shown). Concentrations of THAA ranged from 44 to 266 nmol L–1 atHOT and 67 to 167 nmol L–1 at BATS. Concentrations of THNS rangedfrom 23 to 565 nmol L−1 at HOT and from 50 to 364 nmol L−1 at BATS.THAS concentrations varied from 7 to 84 nmol L−1 at HOT and from 11to 56 nmol L−1 at BATS. THAS concentrations were the sum ofgalactosamine and glucosamine. Mannosamine was detected butcould not be quantified. Depth profiles of THAA, THAS, and THNSconcentrations showed similar general features as TOC at bothstations, but concentrations of these recognizable biochemicalsdecreased more rapidly with depth (Fig. 1). Surface concentrations
Table 3Concentrations, carbon-normalized yields and mole percentages (mol%) of total hydrolyzabHOT and BATS
Location Depth THNS THNS yield THAS THAS yield
(m) (nmol L−1) (%OC) (nmol L−1) (%OC)
HOT 20a 565 3.9 84 0.680a 503 3.5 78 0.6110a 479 3.5 69 0.5150a 269 2.5 49 0.5200a 125 1.2 26 0.3250a 82 0.9 25 0.3300a 72 0.8 23 0.3500a 62 0.9 17 0.3750a 39 0.6 16 0.2
2000a 23 0.4 7 0.14000a 24 0.4 11 0.2
BATS 5 545 4.5 63 0.520a 365 3.0 56 0.525 360 3.1 49 0.450 337 3.0 45 0.475 227 2.1 36 0.3
100a 155 1.5 35 0.4150 138 1.4 25 0.3200 129 1.4 25 0.3350a 84 0.9 20 0.2400 106 1.2 22 0.2500a 87 0.9 19 0.2700 62 0.8 17 0.2900 49 0.6 12 0.2
1360a 51 0.7 11 0.22000a 50 0.6 13 0.23250a 50 0.7 15 0.24300a 55 0.7 13 0.2
a Seawater from these depths was used for ultrafiltration.
of biochemicals were always greater at HOT than at BATS. In contrast,deep waters from BATS generally had higher concentrations of TOCand biochemicals than deep waters from HOT.
THAA, THNS, and THAS together comprised on average 5.0±0.9% ofthe TOC in the surface at HOT and 4.0±1.1% at BATS. In deep water(N1000m), the sum of THAA, THNS, and THAS represented 1.0±0.1% ofTOC at HOTand 1.5±0.1% at BATS (Tables 2, 3). The consistent decreaseof the percentage of carbon comprised by these biochemicals indicatesorganic matter becomes increasingly altered from surface to deepwaters.
le neutral sugars (THNS) and amino sugars (THAS) in unfiltered seawater from Stations
Fuc Rha Ara Gal Glc Man Xyl GalN GlcN
mol% mol%
14 12 13 23 17 11 10 45 5514 12 14 23 16 11 10 47 5315 9 12 20 16 13 16 44 5617 11 13 19 17 13 10 45 5516 9 10 16 29 9 11 41 5917 12 11 19 20 10 11 39 6118 10 11 19 19 9 13 33 6716 11 11 20 23 11 8 34 6617 12 6 19 28 10 7 35 6519 7 7 23 34 0 11 25 7519 8 7 19 47 0 0 32 6811 8 9 22 18 15 16 39 6113 12 10 22 15 13 14 36 6413 11 10 21 19 14 13 35 6512 10 9 21 21 13 14 41 5913 12 11 19 18 12 14 38 6217 12 11 22 15 12 12 39 6116 15 16 18 15 10 12 42 5814 15 15 19 15 10 11 41 5918 11 11 20 17 11 12 43 5717 15 11 19 18 11 10 44 5615 11 11 20 22 11 11 42 5818 10 14 18 18 12 10 40 6019 10 12 17 16 10 15 35 6516 7 11 19 26 12 9 39 6117 11 11 20 25 8 8 44 5616 8 12 20 26 9 8 40 6016 9 9 17 28 10 10 41 59
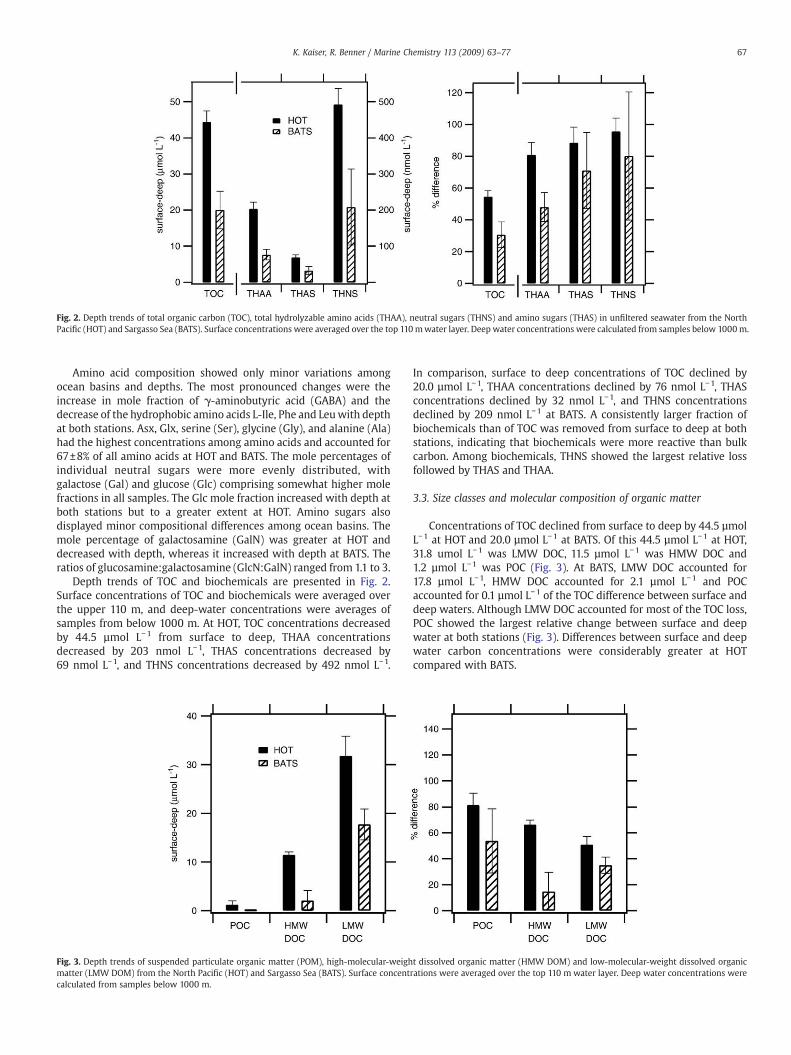
Fig. 2. Depth trends of total organic carbon (TOC), total hydrolyzable amino acids (THAA), neutral sugars (THNS) and amino sugars (THAS) in unfiltered seawater from the NorthPacific (HOT) and Sargasso Sea (BATS). Surface concentrations were averaged over the top 110mwater layer. Deep water concentrations were calculated from samples below 1000 m.
67K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
Amino acid composition showed only minor variations amongocean basins and depths. The most pronounced changes were theincrease in mole fraction of γ-aminobutyric acid (GABA) and thedecrease of the hydrophobic amino acids L-Ile, Phe and Leuwith depthat both stations. Asx, Glx, serine (Ser), glycine (Gly), and alanine (Ala)had the highest concentrations among amino acids and accounted for67±8% of all amino acids at HOT and BATS. The mole percentages ofindividual neutral sugars were more evenly distributed, withgalactose (Gal) and glucose (Glc) comprising somewhat higher molefractions in all samples. The Glc mole fraction increased with depth atboth stations but to a greater extent at HOT. Amino sugars alsodisplayed minor compositional differences among ocean basins. Themole percentage of galactosamine (GalN) was greater at HOT anddecreased with depth, whereas it increased with depth at BATS. Theratios of glucosamine:galactosamine (GlcN:GalN) ranged from 1.1 to 3.
Depth trends of TOC and biochemicals are presented in Fig. 2.Surface concentrations of TOC and biochemicals were averaged overthe upper 110 m, and deep-water concentrations were averages ofsamples from below 1000 m. At HOT, TOC concentrations decreasedby 44.5 µmol L−1 from surface to deep, THAA concentrationsdecreased by 203 nmol L−1, THAS concentrations decreased by69 nmol L−1, and THNS concentrations decreased by 492 nmol L−1.
Fig. 3. Depth trends of suspended particulate organic matter (POM), high-molecular-weighmatter (LMW DOM) from the North Pacific (HOT) and Sargasso Sea (BATS). Surface concentcalculated from samples below 1000 m.
In comparison, surface to deep concentrations of TOC declined by20.0 µmol L−1, THAA concentrations declined by 76 nmol L−1, THASconcentrations declined by 32 nmol L−1, and THNS concentrationsdeclined by 209 nmol L−1 at BATS. A consistently larger fraction ofbiochemicals than of TOC was removed from surface to deep at bothstations, indicating that biochemicals were more reactive than bulkcarbon. Among biochemicals, THNS showed the largest relative lossfollowed by THAS and THAA.
3.3. Size classes and molecular composition of organic matter
Concentrations of TOC declined from surface to deep by 44.5 µmolL−1 at HOT and 20.0 µmol L−1 at BATS. Of this 44.5 µmol L−1 at HOT,31.8 umol L−1 was LMW DOC, 11.5 µmol L−1 was HMW DOC and1.2 µmol L−1 was POC (Fig. 3). At BATS, LMW DOC accounted for17.8 µmol L−1, HMW DOC accounted for 2.1 µmol L−1 and POCaccounted for 0.1 µmol L−1 of the TOC difference between surface anddeep waters. Although LMW DOC accounted for most of the TOC loss,POC showed the largest relative change between surface and deepwater at both stations (Fig. 3). Differences between surface and deepwater carbon concentrations were considerably greater at HOTcompared with BATS.
t dissolved organic matter (HMW DOM) and low-molecular-weight dissolved organicrations were averaged over the top 110 m water layer. Deep water concentrations were
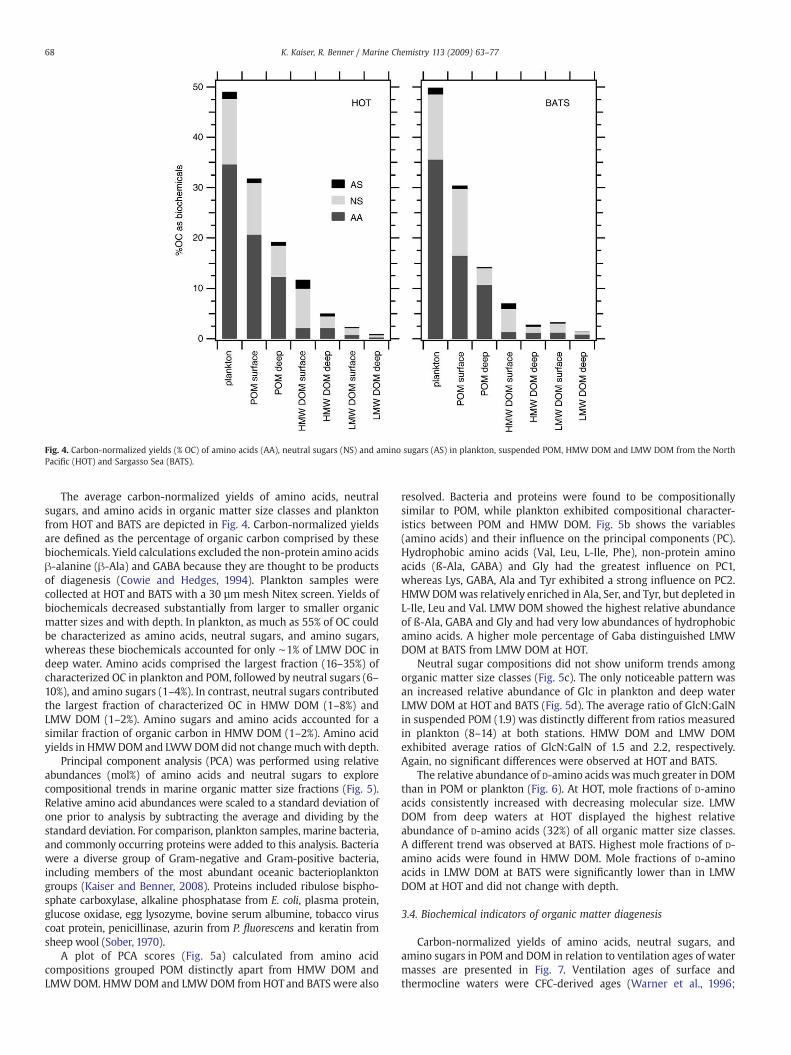
Fig. 4. Carbon-normalized yields (% OC) of amino acids (AA), neutral sugars (NS) and amino sugars (AS) in plankton, suspended POM, HMW DOM and LMW DOM from the NorthPacific (HOT) and Sargasso Sea (BATS).
68 K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
The average carbon-normalized yields of amino acids, neutralsugars, and amino acids in organic matter size classes and planktonfrom HOT and BATS are depicted in Fig. 4. Carbon-normalized yieldsare defined as the percentage of organic carbon comprised by thesebiochemicals. Yield calculations excluded the non-protein amino acidsβ-alanine (β-Ala) and GABA because they are thought to be productsof diagenesis (Cowie and Hedges, 1994). Plankton samples werecollected at HOT and BATS with a 30 µm mesh Nitex screen. Yields ofbiochemicals decreased substantially from larger to smaller organicmatter sizes and with depth. In plankton, as much as 55% of OC couldbe characterized as amino acids, neutral sugars, and amino sugars,whereas these biochemicals accounted for only ∼1% of LMW DOC indeep water. Amino acids comprised the largest fraction (16–35%) ofcharacterized OC in plankton and POM, followed by neutral sugars (6–10%), and amino sugars (1–4%). In contrast, neutral sugars contributedthe largest fraction of characterized OC in HMW DOM (1–8%) andLMW DOM (1–2%). Amino sugars and amino acids accounted for asimilar fraction of organic carbon in HMW DOM (1–2%). Amino acidyields in HMWDOM and LWWDOMdid not changemuchwith depth.
Principal component analysis (PCA) was performed using relativeabundances (mol%) of amino acids and neutral sugars to explorecompositional trends in marine organic matter size fractions (Fig. 5).Relative amino acid abundances were scaled to a standard deviation ofone prior to analysis by subtracting the average and dividing by thestandard deviation. For comparison, plankton samples, marine bacteria,and commonly occurring proteins were added to this analysis. Bacteriawere a diverse group of Gram-negative and Gram-positive bacteria,including members of the most abundant oceanic bacterioplanktongroups (Kaiser and Benner, 2008). Proteins included ribulose bispho-sphate carboxylase, alkaline phosphatase from E. coli, plasma protein,glucose oxidase, egg lysozyme, bovine serum albumine, tobacco viruscoat protein, penicillinase, azurin from P. fluorescens and keratin fromsheep wool (Sober, 1970).
A plot of PCA scores (Fig. 5a) calculated from amino acidcompositions grouped POM distinctly apart from HMW DOM andLMWDOM. HMWDOM and LMWDOM fromHOTand BATS were also
resolved. Bacteria and proteins were found to be compositionallysimilar to POM, while plankton exhibited compositional character-istics between POM and HMW DOM. Fig. 5b shows the variables(amino acids) and their influence on the principal components (PC).Hydrophobic amino acids (Val, Leu, L-Ile, Phe), non-protein aminoacids (ß-Ala, GABA) and Gly had the greatest influence on PC1,whereas Lys, GABA, Ala and Tyr exhibited a strong influence on PC2.HMWDOMwas relatively enriched in Ala, Ser, and Tyr, but depleted inL-Ile, Leu and Val. LMW DOM showed the highest relative abundanceof ß-Ala, GABA and Gly and had very low abundances of hydrophobicamino acids. A higher mole percentage of Gaba distinguished LMWDOM at BATS from LMW DOM at HOT.
Neutral sugar compositions did not show uniform trends amongorganic matter size classes (Fig. 5c). The only noticeable pattern wasan increased relative abundance of Glc in plankton and deep waterLMW DOM at HOT and BATS (Fig. 5d). The average ratio of GlcN:GalNin suspended POM (1.9) was distinctly different from ratios measuredin plankton (8–14) at both stations. HMW DOM and LMW DOMexhibited average ratios of GlcN:GalN of 1.5 and 2.2, respectively.Again, no significant differences were observed at HOT and BATS.
The relative abundance of D-amino acids wasmuch greater in DOMthan in POM or plankton (Fig. 6). At HOT, mole fractions of D-aminoacids consistently increased with decreasing molecular size. LMWDOM from deep waters at HOT displayed the highest relativeabundance of D-amino acids (32%) of all organic matter size classes.A different trend was observed at BATS. Highest mole fractions of D-amino acids were found in HMW DOM. Mole fractions of D-aminoacids in LMW DOM at BATS were significantly lower than in LMWDOM at HOT and did not change with depth.
3.4. Biochemical indicators of organic matter diagenesis
Carbon-normalized yields of amino acids, neutral sugars, andamino sugars in POM and DOM in relation to ventilation ages of watermasses are presented in Fig. 7. Ventilation ages of surface andthermocline waters were CFC-derived ages (Warner et al., 1996;
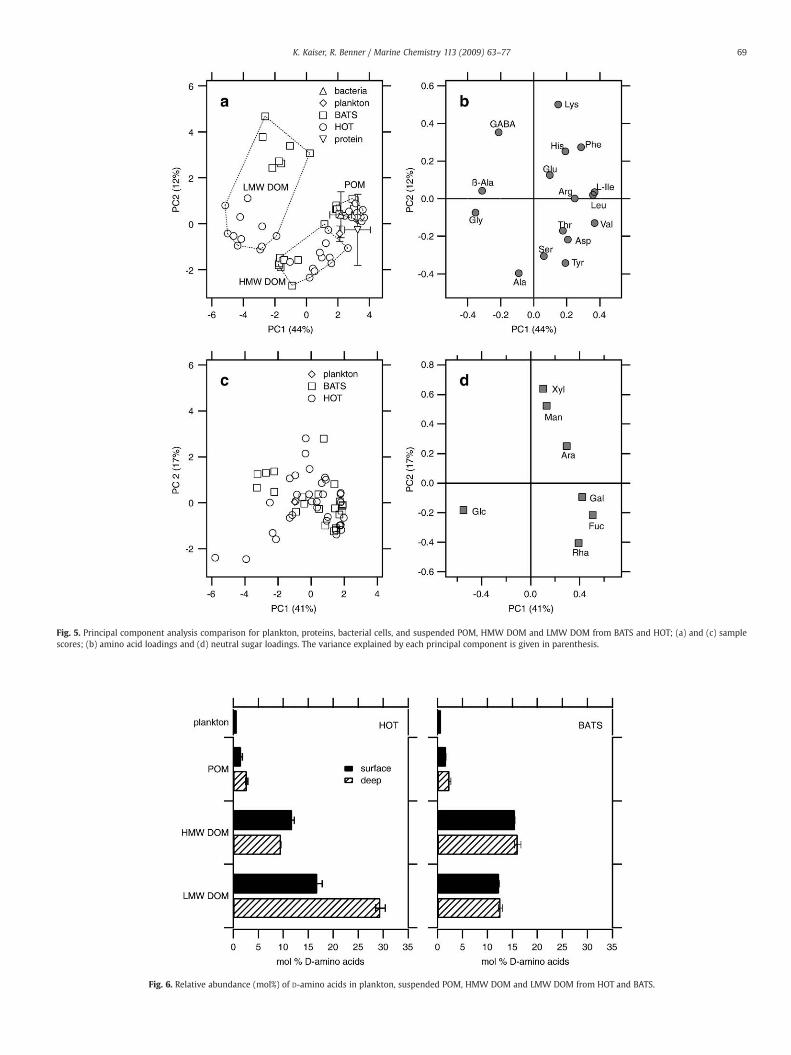
Fig. 5. Principal component analysis comparison for plankton, proteins, bacterial cells, and suspended POM, HMW DOM and LMW DOM from BATS and HOT; (a) and (c) samplescores; (b) amino acid loadings and (d) neutral sugar loadings. The variance explained by each principal component is given in parenthesis.
Fig. 6. Relative abundance (mol%) of D-amino acids in plankton, suspended POM, HMW DOM and LMW DOM from HOT and BATS.
69K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
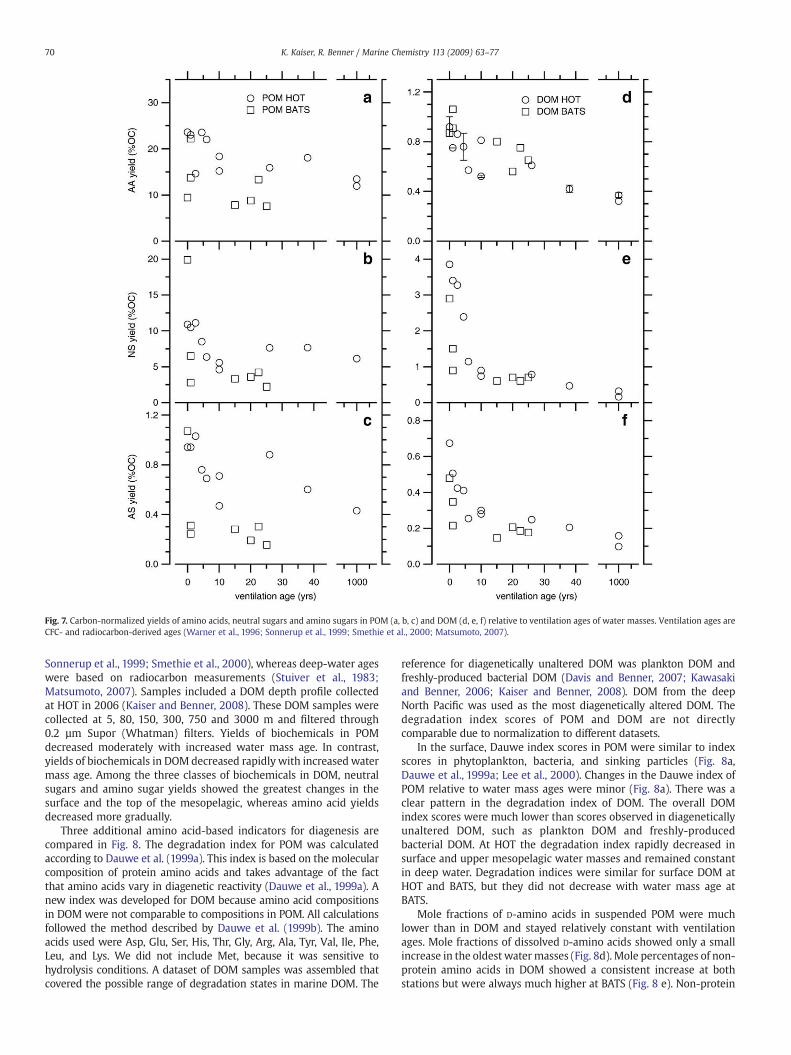
Fig. 7. Carbon-normalized yields of amino acids, neutral sugars and amino sugars in POM (a, b, c) and DOM (d, e, f) relative to ventilation ages of water masses. Ventilation ages areCFC- and radiocarbon-derived ages (Warner et al., 1996; Sonnerup et al., 1999; Smethie et al., 2000; Matsumoto, 2007).
70 K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
Sonnerup et al., 1999; Smethie et al., 2000), whereas deep-water ageswere based on radiocarbon measurements (Stuiver et al., 1983;Matsumoto, 2007). Samples included a DOM depth profile collectedat HOT in 2006 (Kaiser and Benner, 2008). These DOM samples werecollected at 5, 80, 150, 300, 750 and 3000 m and filtered through0.2 µm Supor (Whatman) filters. Yields of biochemicals in POMdecreased moderately with increased water mass age. In contrast,yields of biochemicals in DOM decreased rapidly with increased watermass age. Among the three classes of biochemicals in DOM, neutralsugars and amino sugar yields showed the greatest changes in thesurface and the top of the mesopelagic, whereas amino acid yieldsdecreased more gradually.
Three additional amino acid-based indicators for diagenesis arecompared in Fig. 8. The degradation index for POM was calculatedaccording to Dauwe et al. (1999a). This index is based on the molecularcomposition of protein amino acids and takes advantage of the factthat amino acids vary in diagenetic reactivity (Dauwe et al., 1999a). Anew index was developed for DOM because amino acid compositionsin DOM were not comparable to compositions in POM. All calculationsfollowed the method described by Dauwe et al. (1999b). The aminoacids used were Asp, Glu, Ser, His, Thr, Gly, Arg, Ala, Tyr, Val, Ile, Phe,Leu, and Lys. We did not include Met, because it was sensitive tohydrolysis conditions. A dataset of DOM samples was assembled thatcovered the possible range of degradation states in marine DOM. The
reference for diagenetically unaltered DOM was plankton DOM andfreshly-produced bacterial DOM (Davis and Benner, 2007; Kawasakiand Benner, 2006; Kaiser and Benner, 2008). DOM from the deepNorth Pacific was used as the most diagenetically altered DOM. Thedegradation index scores of POM and DOM are not directlycomparable due to normalization to different datasets.
In the surface, Dauwe index scores in POM were similar to indexscores in phytoplankton, bacteria, and sinking particles (Fig. 8a,Dauwe et al., 1999a; Lee et al., 2000). Changes in the Dauwe index ofPOM relative to water mass ages were minor (Fig. 8a). There was aclear pattern in the degradation index of DOM. The overall DOMindex scores were much lower than scores observed in diageneticallyunaltered DOM, such as plankton DOM and freshly-producedbacterial DOM. At HOT the degradation index rapidly decreased insurface and upper mesopelagic water masses and remained constantin deep water. Degradation indices were similar for surface DOM atHOT and BATS, but they did not decrease with water mass age atBATS.
Mole fractions of D-amino acids in suspended POM were muchlower than in DOM and stayed relatively constant with ventilationages. Mole fractions of dissolved D-amino acids showed only a smallincrease in the oldest water masses (Fig. 8d). Mole percentages of non-protein amino acids in DOM showed a consistent increase at bothstations but were always much higher at BATS (Fig. 8 e). Non-protein
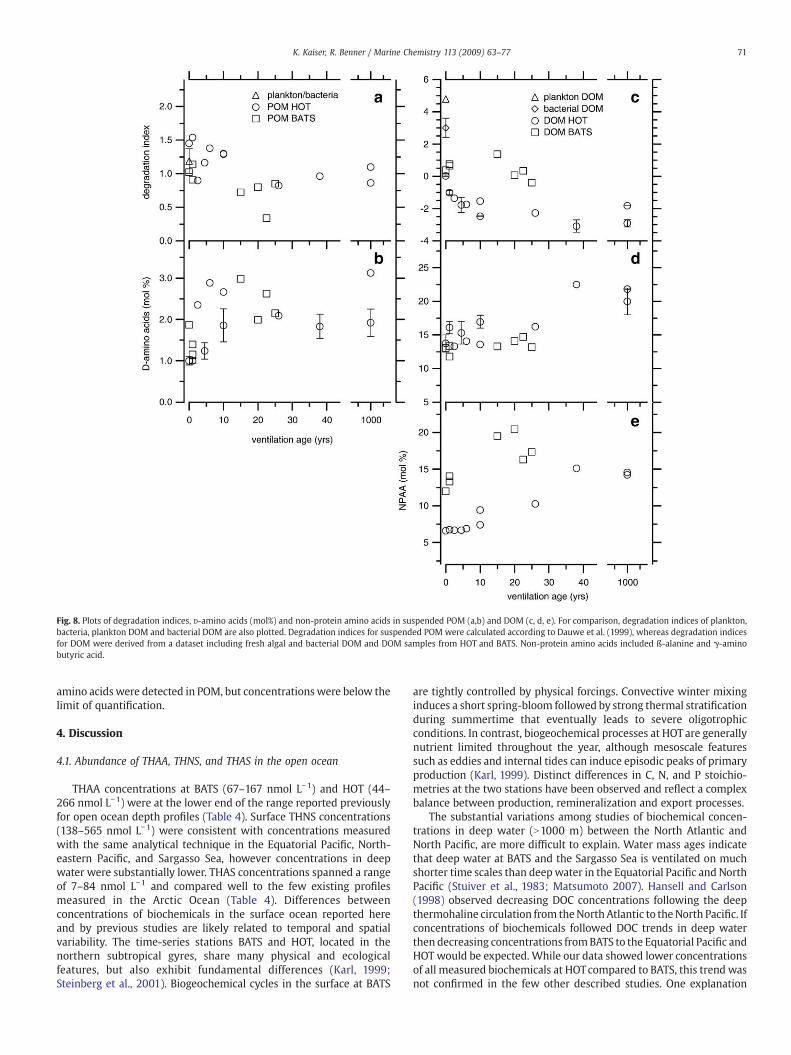
Fig. 8. Plots of degradation indices, D-amino acids (mol%) and non-protein amino acids in suspended POM (a,b) and DOM (c, d, e). For comparison, degradation indices of plankton,bacteria, plankton DOM and bacterial DOM are also plotted. Degradation indices for suspended POM were calculated according to Dauwe et al. (1999), whereas degradation indicesfor DOM were derived from a dataset including fresh algal and bacterial DOM and DOM samples from HOT and BATS. Non-protein amino acids included ß-alanine and γ-aminobutyric acid.
71K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
amino acids were detected in POM, but concentrationswere below thelimit of quantification.
4. Discussion
4.1. Abundance of THAA, THNS, and THAS in the open ocean
THAA concentrations at BATS (67–167 nmol L−1) and HOT (44–266 nmol L−1) were at the lower end of the range reported previouslyfor open ocean depth profiles (Table 4). Surface THNS concentrations(138–565 nmol L−1) were consistent with concentrations measuredwith the same analytical technique in the Equatorial Pacific, North-eastern Pacific, and Sargasso Sea, however concentrations in deepwater were substantially lower. THAS concentrations spanned a rangeof 7–84 nmol L−1 and compared well to the few existing profilesmeasured in the Arctic Ocean (Table 4). Differences betweenconcentrations of biochemicals in the surface ocean reported hereand by previous studies are likely related to temporal and spatialvariability. The time-series stations BATS and HOT, located in thenorthern subtropical gyres, share many physical and ecologicalfeatures, but also exhibit fundamental differences (Karl, 1999;Steinberg et al., 2001). Biogeochemical cycles in the surface at BATS
are tightly controlled by physical forcings. Convective winter mixinginduces a short spring-bloom followed by strong thermal stratificationduring summertime that eventually leads to severe oligotrophicconditions. In contrast, biogeochemical processes at HOT are generallynutrient limited throughout the year, although mesoscale featuressuch as eddies and internal tides can induce episodic peaks of primaryproduction (Karl, 1999). Distinct differences in C, N, and P stoichio-metries at the two stations have been observed and reflect a complexbalance between production, remineralization and export processes.
The substantial variations among studies of biochemical concen-trations in deep water (N1000 m) between the North Atlantic andNorth Pacific, are more difficult to explain. Water mass ages indicatethat deep water at BATS and the Sargasso Sea is ventilated on muchshorter time scales than deepwater in the Equatorial Pacific and NorthPacific (Stuiver et al., 1983; Matsumoto 2007). Hansell and Carlson(1998) observed decreasing DOC concentrations following the deepthermohaline circulation from theNorthAtlantic to theNorth Pacific. Ifconcentrations of biochemicals followed DOC trends in deep waterthen decreasing concentrations fromBATS to the Equatorial Pacific andHOT would be expected. While our data showed lower concentrationsof all measured biochemicals at HOTcompared to BATS, this trend wasnot confirmed in the few other described studies. One explanation
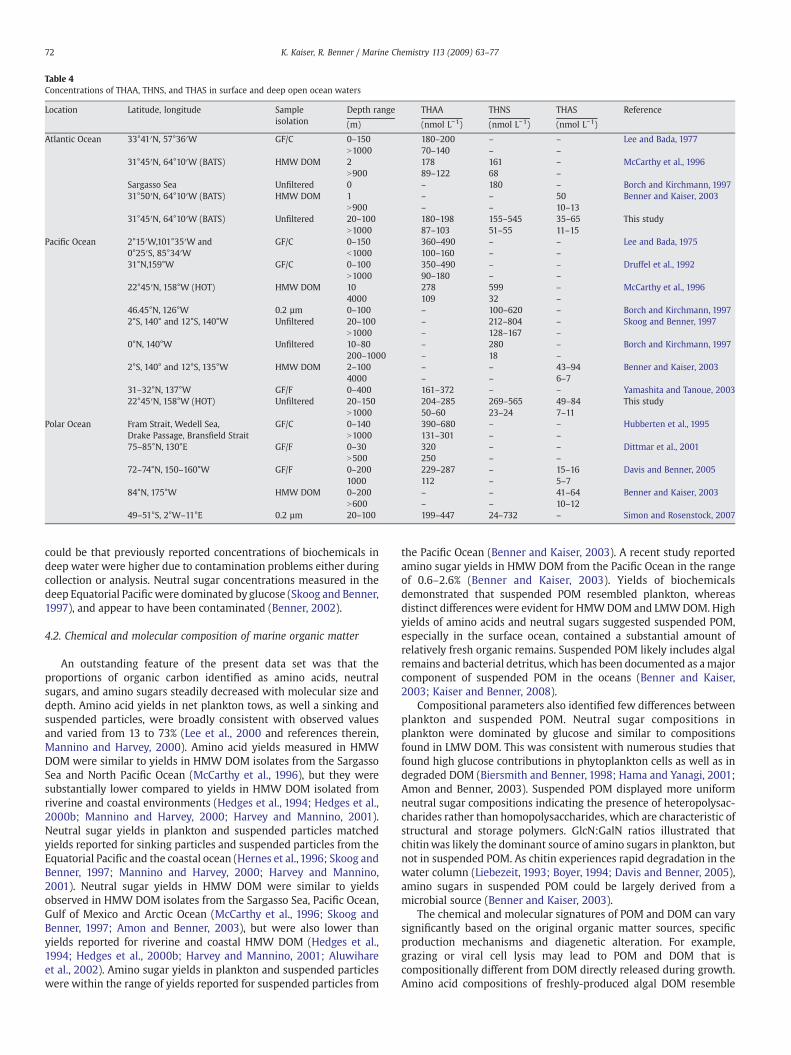
Table 4Concentrations of THAA, THNS, and THAS in surface and deep open ocean waters
Location Latitude, longitude Sampleisolation
Depth range THAA THNS THAS Reference
(m) (nmol L−1) (nmol L−1) (nmol L−1)
Atlantic Ocean 33°41′N, 57°36′W GF/C 0–150 180–200 – – Lee and Bada, 1977N1000 70–140 – –
31°45′N, 64°10′W (BATS) HMW DOM 2 178 161 – McCarthy et al., 1996N900 89–122 68 –
Sargasso Sea Unfiltered 0 – 180 – Borch and Kirchmann, 199731°50′N, 64°10′W (BATS) HMW DOM 1 – – 50 Benner and Kaiser, 2003
N900 – – 10–1331°45′N, 64°10′W (BATS) Unfiltered 20–100 180–198 155–545 35–65 This study
N1000 87–103 51–55 11–15Pacific Ocean 2°15′W,101°35′W and GF/C 0–150 360–490 – – Lee and Bada, 1975
0°25′S, 85°34′W b1000 100–160 – –
31°N,159°W GF/C 0–100 350–490 – – Druffel et al., 1992N1000 90–180 – –
22°45′N, 158°W (HOT) HMW DOM 10 278 599 – McCarthy et al., 19964000 109 32 –
46.45°N, 126°W 0.2 µm 0–100 – 100–620 – Borch and Kirchmann, 19972°S, 140° and 12°S, 140°W Unfiltered 20–100 – 212–804 – Skoog and Benner, 1997
N1000 – 128–167 –
0°N, 140°W Unfiltered 10–80 – 280 – Borch and Kirchmann, 1997200–1000 – 18 –
2°S, 140° and 12°S, 135°W HMW DOM 2–100 – – 43–94 Benner and Kaiser, 20034000 – – 6–7
31–32°N, 137°W GF/F 0–400 161–372 – – Yamashita and Tanoue, 200322°45′N, 158°W (HOT) Unfiltered 20–150 204–285 269–565 49–84 This study
N1000 50–60 23–24 7–11Polar Ocean Fram Strait, Wedell Sea, GF/C 0–140 390–680 – – Hubberten et al., 1995
Drake Passage, Bransfield Strait N1000 131–301 – –
75–85°N, 130°E GF/F 0–30 320 – – Dittmar et al., 2001N500 250 – –
72–74°N, 150–160°W GF/F 0–200 229–287 – 15–16 Davis and Benner, 20051000 112 – 5–7
84°N, 175°W HMW DOM 0–200 – – 41–64 Benner and Kaiser, 2003N600 – – 10–12
49–51°S, 2°W–11°E 0.2 µm 20–100 199–447 24–732 – Simon and Rosenstock, 2007
72 K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
could be that previously reported concentrations of biochemicals indeep water were higher due to contamination problems either duringcollection or analysis. Neutral sugar concentrations measured in thedeep Equatorial Pacific were dominated by glucose (Skoog and Benner,1997), and appear to have been contaminated (Benner, 2002).
4.2. Chemical and molecular composition of marine organic matter
An outstanding feature of the present data set was that theproportions of organic carbon identified as amino acids, neutralsugars, and amino sugars steadily decreased with molecular size anddepth. Amino acid yields in net plankton tows, as well a sinking andsuspended particles, were broadly consistent with observed valuesand varied from 13 to 73% (Lee et al., 2000 and references therein,Mannino and Harvey, 2000). Amino acid yields measured in HMWDOM were similar to yields in HMW DOM isolates from the SargassoSea and North Pacific Ocean (McCarthy et al., 1996), but they weresubstantially lower compared to yields in HMW DOM isolated fromriverine and coastal environments (Hedges et al., 1994; Hedges et al.,2000b; Mannino and Harvey, 2000; Harvey and Mannino, 2001).Neutral sugar yields in plankton and suspended particles matchedyields reported for sinking particles and suspended particles from theEquatorial Pacific and the coastal ocean (Hernes et al., 1996; Skoog andBenner, 1997; Mannino and Harvey, 2000; Harvey and Mannino,2001). Neutral sugar yields in HMW DOM were similar to yieldsobserved in HMW DOM isolates from the Sargasso Sea, Pacific Ocean,Gulf of Mexico and Arctic Ocean (McCarthy et al., 1996; Skoog andBenner, 1997; Amon and Benner, 2003), but were also lower thanyields reported for riverine and coastal HMW DOM (Hedges et al.,1994; Hedges et al., 2000b; Harvey and Mannino, 2001; Aluwihareet al., 2002). Amino sugar yields in plankton and suspended particleswere within the range of yields reported for suspended particles from
the Pacific Ocean (Benner and Kaiser, 2003). A recent study reportedamino sugar yields in HMW DOM from the Pacific Ocean in the rangeof 0.6–2.6% (Benner and Kaiser, 2003). Yields of biochemicalsdemonstrated that suspended POM resembled plankton, whereasdistinct differences were evident for HMWDOM and LMWDOM. Highyields of amino acids and neutral sugars suggested suspended POM,especially in the surface ocean, contained a substantial amount ofrelatively fresh organic remains. Suspended POM likely includes algalremains and bacterial detritus, which has been documented as amajorcomponent of suspended POM in the oceans (Benner and Kaiser,2003; Kaiser and Benner, 2008).
Compositional parameters also identified few differences betweenplankton and suspended POM. Neutral sugar compositions inplankton were dominated by glucose and similar to compositionsfound in LMW DOM. This was consistent with numerous studies thatfound high glucose contributions in phytoplankton cells as well as indegraded DOM (Biersmith and Benner, 1998; Hama and Yanagi, 2001;Amon and Benner, 2003). Suspended POM displayed more uniformneutral sugar compositions indicating the presence of heteropolysac-charides rather than homopolysaccharides, which are characteristic ofstructural and storage polymers. GlcN:GalN ratios illustrated thatchitinwas likely the dominant source of amino sugars in plankton, butnot in suspended POM. As chitin experiences rapid degradation in thewater column (Liebezeit, 1993; Boyer, 1994; Davis and Benner, 2005),amino sugars in suspended POM could be largely derived from amicrobial source (Benner and Kaiser, 2003).
The chemical and molecular signatures of POM and DOM can varysignificantly based on the original organic matter sources, specificproduction mechanisms and diagenetic alteration. For example,grazing or viral cell lysis may lead to POM and DOM that iscompositionally different from DOM directly released during growth.Amino acid compositions of freshly-produced algal DOM resemble

73K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
oceanic DOM rather than cell structural organic matter (Davis andBenner, 2005). Phytoplankton blooms are known to produce largeamounts of carbohydrate-rich DOM (Ittekkot et al., 1981; Biddandaand Benner, 1997). Several studies have shown that phytoplanktoncultures release carbohydrate-rich HMW DOM during growth that iscompositionally similar to surface ocean HMW DOM (Biersmith andBenner, 1998; Hama and Yanagi, 2001; Aluwihare et al., 2002).
Extensive heterotrophic processing of DOM fractions was reflectedin the low yields of biochemicals. Amino acids and neutral sugarsgenerally comprise a large portion of bioavailable organic matter andare preferentially consumed by microorganisms (Cowie and Hedges,1994; Davis and Benner, 2007). Microbial alterationswere also evidentfrom the relative increases of non-protein amino acids and D-aminoacids with smaller molecular sizes. The non-protein amino acids GABAand ß-Ala are generally considered to be decarboxylation products ofGlu and Asp, respectively (Cowie and Hedges, 1994). Interestingly,both non-protein amino acids showed highest relative contributionsin LMW DOM at BATS although the bulk radiocarbon age of DOM atBATS is much younger than at HOT (Williams and Druffel, 1987;Druffel et al., 1992). Multiple degradation pathways may help explainthese trends or perhaps there was an additional biotic source of theseamino acids at BATS. If the production of GABA and ß-Alapreferentially occurs in the very early stages of diagenesis and LMWDOM reflects the most degraded DOM, then further degradationmight slowly remove these amino acids again. Eukaryotic organismsand bacteria are known to synthesize free GABA (Koolman and Roehm,2005), but no biotic sources for covalently bound non-protein aminoacids have yet been identified.
The bacterial imprint in DOM compositions was reflected by therelatively high mole fractions of D-amino acids in HMW DOM andLMW DOM. Several reports have demonstrated that bacterial organicmatter is a major component of non-living organic matter in theoceans (McCarthy et al., 1998; Dittmar et al., 2001; Kaiser and Benner,2008). High mole fractions of D-amino acids do not necessarilyindicate advanced diagenesis, as heterotrophic bacteria releasebioreactive DOM during normal growth that is preferentially enrichedin D-amino acids compared to their cellular composition (Kawasakiand Benner, 2006; Kaiser and Benner, 2008). This reflects the selectiveincorporation of specific bacterial macromolecules into bacterialDOM. There are also two distinct sources of D-amino acids in theoceans that could affect their size distribution. Bacterivorous grazingcan lead to production of POM and HMWDOM enrichedwith D-aminoacids, whereas direct release of bacterial DOM results in D-amino acidrich LMW DOM (Kawasaki and Benner, 2006).
Mole fractions of D-amino acids in HMW DOM were fairly similaramong stations and also fairly comparable to LMW DOM at BATS andsurface LMW DOM at HOT. Only LMW DOM in the deep North Pacificshowed much higher mole percentages of D-amino acids than anyother DOM size fraction. This trend appeared to reflect extensivealteration of deepwater DOMduring circulation in the ocean conveyor.
Other observed compositional changes included the pronouncedincrease in the mole fraction of glycine with depth and with decreas-ing molecular size. Increases of glycine contributions in sinkingparticles and sedimentary organic matter have been attributed to theincorporation of diatom cell walls (Dauwe and Middelburg, 1998).Diatom cell walls are also enriched in serine and threonine (Heckyet al., 1973) but this was not observed in LMW DOM, which displayedthe highest glycine content. The preservation of glycinemay be relatedto an intrinsic structural property of glycine-rich macromolecules.Nguyen and Harvey (1997) found that selective glycine preservationwas a common pattern during phytoplankton decay under oxic andanoxic conditions.
In addition to the changes in Gly mole fraction, aromatic aminoacids rapidly disappeared from larger to smaller molecular sizes andwith depth. This could be related to the fact that aromatic amino acidspreferentially occur in the cell plasma (Hecky et al., 1973). Aromatic
amino acids contain a chromophore and phototransformations couldbe another explanation for their relatively rapid removal (Mopper etal., 1991; Benner and Biddanda, 1998). However, the largest changesoccurred below the euphotic zone indicating that phototransforma-tions play a minor role in removal of these amino acids.
Neutral sugars showed surprisingly small compositional changesbetween suspended POM and DOM size classes. The only pronouncedshift was a higher mole fraction of glucose in LMW DOM. This featurehas beenobserved in several other studies and appears to be a commondiagenetic trend (Skoog and Benner,1997; Amon andBenner, 2003 andreferences therein). The incorporation of glucose-rich polymers inrecalcitrant organic matter remains enigmatic but may be related tostructural characteristics. Other reported trends, such as increasingmole percentages of deoxy-sugars (rhamnose and arabinose) withprogressing diagenesis in marine and terrigenous POM (Cowie andHedges, 1984; Ittekkot and Arian, 1986; Hedges et al., 1994; Herneset al., 1996; Benner and Opsahl, 2001; Amon and Benner, 2003) couldnot be confirmedwith the present data set.Meon andKirchman (2001)documented that carbohydrate compositions of DOM released byphytoplankton blooms immediately shifted to compositions observedin marine DOM. This indicates that diagenetic changes are rapidlyimprinted in carbohydrate compositions and further degradationreactions do not alter neutral sugar compositions substantially.
GalN and GlcN were the dominant amino sugars in all organicmatter size fractions. They occurred in fairly constant proportions insuspended POM, HMWDOM and LMWDOM, (1.5–2.2). This indicatedthat chitin was only a minor source of glucosamine and amino sugarscould be of bacterial origin (Benner and Kaiser, 2003). Similar GalN:GlcN ratios in all size fractions also provided evidence for a commonmacromolecular origin of these amino sugars. Another interestingfeature was that the majority of amino sugars occurred in HMWDOM.Numerous macromolecular sources exist for amino sugars but only afew show proportions of GalN and GlcN that match ratios found inorganic matter size fractions. These are the oligosaccharides of O-linked glycoproteins and lipopolysaccharides, which are importantconstituents of Gram-negative cell membranes. Glycoproteins with O-linked oligosaccharides are ubiquitous constituents of eukaryotic cells(Schachter and Brockhausen, 1992) and recent reports have indicatedtheir occurrence in archaeal and gram-positive bacterial cell surfaces(Schaffer and Messner, 2001). However, until now no evidence existsindicating glycoproteins occur in Gram-negative bacteria (Schafferand Messner, 2001).
Bacterial lipids indicative of Gram-negative lipopolysaccharidesare a distinctive feature of marine DOM (Wakeham et al., 2003).Cadieux et al. (1983) documented that Gram-negative bacteriaspontaneously release dissolved lipopolysaccharides during normalgrowth. Further, ratios of GalN and GlcN are very similar in DOMderived from bacteria and marine DOM (Ogawa et al., 2001; Kawasakiand Benner, 2006). It is possible that lipopolysaccharides andoligosaccharides from O-linked glycoproteins originating from eukar-yotic and prokaryotic organisms are the main sources for aminosugars in marine organic matter. Multiple sources of amino sugars areconsistent with observed reactivity patterns. Benner and Kaiser(2003) found that amino sugars were recycled more efficiently inthe upper water column than amino acids. Experiments studying thecomposition and reactivity of bacterial DOMdemonstrated that aminosugars could persist for long periods of time (Ogawa et al., 2001;Kawasaki and Benner, 2006).
4.3. Comparison of compound-specific analyses with bulk chemicalanalyses
The fraction of organic matter that could be characterized bycompound-specific analyses rapidly decreased with depth andmolecular size. Based on this observation one could hypothesizethat diagenetic reactions alter the original source biochemicals and

74 K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
prevent their detection by compound-specific analyses. Diagenesiscould also lead to the preservation of macromolecular structures thatare resistant to hydrolytic conditions or form heteropolycondensatesduring hydrolysis (Hedges et al., 2000a).
Analyses by nuclear magnetic resonance spectroscopy (NMR) andmass spectrometry are very informative because these techniques stillallow the detection of chemical bonds indicative of specific compoundgroups even if they are removed from the analytical window of a wet-chemistry technique. Solid-state 13C-NMR of HMW DOM from thePacific Ocean indicated that carbohydrates comprised between 25–50% of the carbon (Benner et al., 1992; Sannigrahi et al., 2005). Thecolorimetric MBTH technique measured 15–22% of the carbon incarbohydrates in the same HMW DOM samples (Pakulski and Benner,1992). In comparison, concentrations of neutral sugars and aminosugars in HMW DOM from HOT and BATS determined by compound-specific analyses accounted for only 3–12% of the organic carbon.Certain carbohydrates, such as ribose, deoxyribose and fructose aresusceptible to acid degradation and this could explain part of thedifferences between NMR, colorimetric and compound-specificcarbohydrate estimates. Both colorimetric and compound-specificanalyses use strong acid to hydrolyze covalently-linked organicmatter. Methylated carbohydrates and acidic carbohydrates (e.g.uronic acids and sulfated carbohydrates) are also important groupsof carbohydrates but they were not characterized in the present study.Based on NMR, mass spectrometry analyses and periodate oxidationexperiments, methylated sugars appear to be ubiquitous carbohy-drates in natural organic matter, but it is difficult to relate thesefindings to quantitative estimates (Boon et al., 1998; Panagiotopouloset al., 2007; Quan and Repeta, 2007). A recent study by Hung et al.(2001) reported only minor contributions of uronic acids tocarbohydrates of estuarine waters. Acid-hydrolysis efficiently cleavesof the sulfate group in sulfated carbohydrates and these carbohydratesare measured as their neutral analog (Kaiser and Benner, 2000). Thecolorimetric MBTH method is sensitive to all vicinal diols that arelocated at the terminal end, which could result in overestimates ofcarbohydrate concentrations.
Hydrolysis-induced condensation reactions could also lead tolower recoveries of carbohydrates in compound-specific analysescompared to spectroscopic techniques. Maillard-type reactions readilyform complex heteropolycondensates from amino acids and carbohy-drates, however formation is strongly inhibited at a pH below 4(March, 1992). Carbohydrates could be converted to furfural deriva-tives by dehydration and concurrent elimination reactions. Further, itis possible that carbohydrates form aldols and their dehydrationproducts during acid hydrolysis. The initial step in this reaction is theformation of ß-hydroxy aldehydes that are subject to dehydration andfurther condensation (March, 1992).
Another possibility is that the hydrolytic conditions employed incompound-specific analyses are not sufficient to break the covalentbonds in hydrolysis-resistant macromolecules. 15N-NMR analyses ofHMWDOM from the Equatorial Pacific Ocean estimated that 75–94% ofthe nitrogen was in amide structures (McCarthy et al., 1997). The mostcommon biopolymers with amide nitrogen are proteins, peptides, andmacromolecules containing N-acetylated-amino-sugar, such as chitinandpeptidoglycan. In comparison, compound-specific analyses ofHMWDOM in this study found only 12–21% of the nitrogen in amino sugarsand amino acids. Aluwihare et al. (2005) suggestedmost amides presentin HMW DOM resist acid hydrolysis and that compound-specificanalyses significantly underestimate amino sugar concentrations. Thisconclusionwasbased on linking the abundance of acetic acid exclusivelyto N-acetylated amino polysaccharides. However, acetylation is also acommon post-translational modification of proteins and the acetyl-functionality is found in various biomolecules in addition to aminosugars (Wold, 1981; Martinez et al., 2000), indicating the acetic acidreleased from HMW DOM during acid hydrolysis is as likely to beassociated with peptides and proteins as it is with amino sugars. The
bacterial cell wall polymer peptidoglycan was thought to constitute amajor component of DOM and its relative resistance to acid hydrolysismade it an ideal candidate for compoundsmissed in compound-specificanalyses (McCarthy et al., 1998; Hedges et al., 2000a). However, recentfindings indicate that peptidoglycan is only aminor component of DOM(Kaiser and Benner, 2008). Closing the gap between compound-specificanalyses and bulk chemical characterizations is critical to gain insightsabout the sources and diagenetic processing of organic matter in theocean.
4.4. Size continuum of marine organic matter and reactivity
Previous studies of diagenetic processes and carbon cycling in theocean suggested a distinct relationship between size and biologicalreactivity of organic matter. Amon and Benner (1994, 1996) found thatbacterial utilization of HMW DOM was typically greater than LMWDOM, indicating diagenetic processes lead to the production ofsmaller molecules that are more resistant to bacterial degradation.Several other studies documented a similar relationship betweenreactivity and size for POM and DOM (Mannino and Harvey, 2000;Harvey and Mannino, 2001; Hama et al., 2004; Zou et al., 2004;Seitzinger et al., 2005). Furthermore, Loh et al. (2004) observed adistinct size-age continuum based on radiocarbon ages of isolatedorganic matter fractions from HOT and BATS.
Our findings not only support the size-reactivity continuummodelproposed by Amon and Benner (1994, 1996) but also provide evidencefor a size-composition continuummodel (Benner, 2002). The relation-ship between size, reactivity, and chemical composition has severalimplications. One is that it provides a framework for understandingthe cycling of marine organic matter. Detrital organic matter in theocean exists in a continuum of sizes with variable biochemicalcompositions that are remineralized on different timescales. Highyields of characterizable biochemicals generally relate to unalteredorganic remains with high bioreactivity. Finally, enzymatic degrada-tion reactions are a major mechanism for the formation of smallerorganic compounds with reduced bioreactivity.
Second, these conceptual models also provide clues about theformation of refractory organic matter in the ocean. As organic matteris degraded and decreases in size and bioreactivity, its molecularsignature is altered and lost. In this context, membrane encapsulationappears to be a minor mechanism for the preservation of organicmatter in the ocean (Nagata et al., 1998). The direct production ofintrinsically refractory LMW also seems to be a minor pathway(Brophy and Carlson, 1989). Several hypotheses offer potentialexplanations for the formation of small refractory organic compounds.Ogawa et al. (2001) speculated that nonspecific enzyme reactionscould lead to secondary products that are resistant to degradation.Products of such enzymatic degradations may not resemble thestructure of original biochemicals thereby greatly reducing enzymaticrecognition and further biodegradation. Alternatively, photochemicaldecomposition reactions may result in the formation of structurallycomplex LMW compounds that are highly resistant to degradation(Benner and Biddanda, 1998). Numerous studies have shown thatphotochemical reactions lead to decreases in the molecular weightand size of DOM (Opsahl and Benner, 1998; Mopper and Kieber, 2002).
Size itself can also affect the bioreactivity of individual organicmatter size fractions. Colloidal organic matter, which is part of HMWDOM, is much less accessible to bacteria than particles larger than afew µm because it occupies a minimum between 2 different transportregimes (Kepkay, 1994; Wells and Goldberg, 1993). Brownian motiondominates transport of smaller colloids to bacteria, while largerparticles are primarily transported to bacteria by turbulent shear(Kepkay, 1994).
Another important observation is that the relative diageneticreactivities of the three classes of biochemicals analyzed in POM,HMWDOM and LMWDOM are distinctly different. Neutral and amino

75K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
sugars always showed much greater relative losses with depth in allsize fractions than amino acids. These findings suggest that neutralsugars and amino sugars are considerably more bioreactive inmesopelagic waters than amino acids in all organic matter sizefractions. On the contrary, in sinking particles the relative reactivity ofamino acids was found to be greater than that of neutral sugars(Wakeham et al., 1997). One explanation for the low reactivity ofamino acids in suspended POM and DOM is that as nitrogen-bearingcompounds they are already significantly degraded. Previous studieshave noted that amino acids are important components of labileorganic matter pool and are consumed rapidly in surface waters(Coffin, 1989; Keil and Kirchman, 1993; Cherrier and Bauer, 2004;Davis and Benner, 2007). The high bioreactivity of neutral and aminosugars suggests that these biochemicals fuel much of the hetero-trophic metabolism in the mesopelagic ocean.
Although LMW DOM comprises the most diagenetically altered andrefractory organicmatter in the ocean, it still contains a small fraction ofcharacterizable biochemicals. The relative stability of these biochemicalsis intriguing because they must be subject to intense remineralizationpressure. Additional studies are necessary to investigate the mechan-isms that control the cycling of LMW DOM in the ocean.
4.5. Biochemical indicators of organic matter diagenesis
Compound-specific analyses have tremendous potential to investi-gate the diagenetic alterations of organic mixtures in aquatic environ-ments. Cowie andHedges (1994) showed that carbon-normalized yieldsof amino acids and carbohydrates sensitively recorded the extent ofdegradation of natural organic matter. The relative compositions ofamino acids are useful for identifying diagenetic trends in sedimentaryorganic matter (Dauwe and Middelburg, 1998). Although all of thesecompound-specific indicators effectively documented the relativediagenetic state of organic matter, they were sensitive to differentstages of degradation (Cowie and Hedges, 1994; Davis et al., in press).Davis et al. (in press) have demonstrated that amino acid yields bestdescribed the very early stages of DOMdiagenesis, whereas non-proteinaminoacidsweremost sensitive in the later stages. Anamino-acid basedcompositional parameter functionedmost reliably on annual to decadaltime scales of organic matter diagenesis.
The present data provided a unique opportunity to evaluate theperformance of various diagenetic indicators for marine POM andDOM from different ocean basins and over long time scales. Theaverage age of deep water DOC at HOT was reported to be close to6000 years (Williams and Druffel, 1987) and thus likely represents themost degraded and altered organic matter in the ocean. All molecularindicators were related to ventilation ages of water masses derivedfrom CFC- and radiocarbon measurements (Warner et al., 1996;Sonnerup et al., 1999; Smethie et al., 2000; Matsumoto, 2007) toprovide a temporal scale for evaluating molecular indicators ofdiagenesis. Carbon-normalized yields of amino acids, neutral sugars,and amino sugars provided the most consistent information on thediagenetic state of POM and DOM. Sharp declines in yields relative toventilation ages occurred in DOM indicating the selective removal ofthese components during degradation. Yields of all biochemicalschanged more moderately in POM but still indicated progressivediagenetic alterations within older water masses. In DOM, yields ofbiochemicals also appeared to be sensitive to different stages ofalterations. Yields of neutral sugars and amino sugars declined mostrapidly in surface and upper mesopelagic water masses that wereventilated on time scales of 1–10 years. In contrast, yields of aminoacids decreased gradually over a time span of about 1000 years.
Amino-acid based compositional parameters in DOM displayeddistinct differences between ocean basins. TheDOMdegradation indexat HOT indicated diagenetic alterations in a similar fashion as yields ofbiochemicals. In contrast, the degradation index showed minordiagenetic alterations with increasing ventilation ages at BATS.
Furthermore, mole fractions of non-protein amino acids were clearlydifferent between HOTand BATS for similar ventilation ages. However,the pattern of mole fractions of non-protein amino acids relative toventilation ages was similar at both stations and consistent withincreasing diagenetic alterations. It is possible that amino acidcompositions are sensitive to variable heterotrophic processing,perhaps caused by different microbial communities. Alternatively,these indicators could be compromised by varying DOM sources. Asalready discussed, BATS and HOT show distinctly different seasonaland biogeochemical cycles in surfacewaters. Deepwater at BATS formsin the North Atlantic where it entrains a significant amount of Arctic-derived terrigenous DOM (Benner et al., 2005). The dominantcomponent of North Pacific deep water at HOT is Antarctic BottomWater (Matsumoto, 2007). Overall, these variable patterns limit theapplication of diagenetic indicators based on amino acid compositions.
D-amino acids (mol%) were evaluated as an additional diageneticindicator for POM and DOM. Laboratory experiments showed D-aminoacids and bacterial DOM are less bioreactive than algal DOM (Jorgensenet al., 1999; Amon et al., 2001) implying microbial alteration reactionscould lead to refractory DOM enriched in D-amino acids. D-amino acids(mol%) did not increasemuchwith ventilation ages in POM. In DOM, theonly change of D-amino acids (mol%) was observed after a ventilationage of 40 years. Surface DOM already contains fairly high amounts of D-amino acids, although fresh algal DOM only contributes L-amino acids.Previous studies have shown similar degradation rates of D- and L-aminoacids in estuarine waters (Jorgensen et al., 2003; Middleboe andJorgensen, 2006) and North Atlantic Deep Water (Teira et al., 2006). Inaddition, bacterially derived D- and L-amino acids appear to have similarreactivities (Kawasaki and Benner, 2006; Kaiser and Benner, 2008). Thissuggests D-amino acid (mol%) signatures are rapidly imprinted in theambient marine DOM pool and latter diagenetic reactions have a minorinfluence on the relative contribution of D-amino acids to DOM. Thesefindings indicate that D-amino acids (mol%) may only be an effectiveindicator of the early stages of diagenesis.
5. Conclusions
Although a dominant fraction of detrital organicmatter in the oceansstill eludes detailed chemical characterization, compound-specificanalyses of common biochemicals can provide important insightsabout its decomposition and preservation. Compositional informationindicated variable sources for suspended POM and DOM and that theextent of diagenetic alterations increases with decreasing molecularsize. Carbon-normalized yields of amino acids, neutral sugars, andamino sugars sharply decreased from suspended POM to HMW DOMand LMW DOM. As much as 55% of OC could be characterized as thesebiochemicals in suspended POM from the surface, but only 2% of the OCin LMW DOM from the deep North Pacific. The results of this studyfirmly support a size-reactivity and a size-composition continuum forthe diagenetic processing of organic matter in the ocean. Microbialalterations lead to the production of smaller compounds that are moreresistant to biodegradation and not recognizable by commonmolecularcharacterizations. This study also provided anopportunity to investigatethe usefulness of various diagenetic indicators of organic matteralteration. Carbon-normalized yields of amino acids, neutral sugarsandamino sugars proved to be themost reliablediagenetic indicators forsuspended POMandDOM. Indicators basedon amino acid compositionsindicatedmixed results suggesting that theymay be sensitive to variablesources or diagenetic processing.
Acknowledgement
We gratefully acknowledge Xiaoqun Wang for help with carbohy-drate analyses and Nobuyuki Kawasaki for help with total organiccarbon analyses. We thank the officers, scientists and crew aboard theKai-ma-kai O'Kanaloa and Cape Hatteras for assistance with sampling.

76 K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
We thank Ellery Ingall for providing plankton samples and twoanonymous reviewers for helpful comments on an earlier draft of thismanuscript. This work was supported by NSF grant OCE-0080782.
References
Aluwihare, L.I., Repeta, D.J., Chen, R.F., 2002. Chemical composition and cycling ofdissolved organic matter in the Mid-Atlantic Bight. Deep-Sea Res., Part II, Top. Stud.Oceanogr. 49 (20), 4421–4437.
Aluwihare, L.I., Repeta, D.J., Pantoja, S., Johnson, C.G., 2005. Two chemically distinctpools of organic nitrogen accumulate in the ocean. Science 308 (5724), 1007–1010.
Amon, R.M.W., Benner, R., 1994. Rapid-cycling of high-molecular-weight dissolvedorganic-matter in the ocean. Nature 369 (6481), 549–552.
Amon, R.M.W., Benner, R., 2003. Combine neutral sugars as indicators of the diageneticstate of dissolved organic matter in the Arctic Ocean. Deep-Sea Res., Part I,Oceanogr. Res. Pap. 50, 151–169.
Amon, R.M.W., Benner, R., 1996. Bacterial utilization of different size classes of dissolvedorganic matter. Limnol. Oceanogr. 41 (1), 41–51.
Amon, R.M.W., Fitznar, H.P., Benner, R., 2001. Linkages among the bioreactivity, chemicalcomposition, and diagenetic state of marine dissolved organic matter. Limnol.Oceanogr. 46 (2), 287–297.
Benner, R., 1991. Ultrafiltration for the concentration of bacteria, viruses, and dissolvedorganic matter. In: Hurd, D.C., Spencer, D.W. (Eds.), Marine Particles: Analysis andCharacterization. Geophys. Monogr., 63. Am. Geophys. Union, Washington, DC,pp. 181–185.
Benner, R., 2002. Chemical composition and reactivity. In: Hansell, D.A., Carlson, C.A.(Eds.), Biogeochemistry of Marine Dissolved Organic Matter. Academic Press,Amsterdam, pp. 59–85.
Benner, R., Biddanda, B., 1998. Photochemical transformations of surface and deepmarinedissolved organic matter: effects on bacterial growth. Limnol. Oceanogr. 43 (6),1373–1378.
Benner, R., Opsahl, S., 2001. Molecular indicators of the sources and transformations ofdissolved organic matter in the Mississippi river plume. Org. Geochem. 32 (4),597–611.
Benner, R., Kaiser, K., 2003. Abundance of amino sugars and peptidoglycan in marineparticulate and dissolved organic matter. Limnol. Oceanogr. 48 (1), 118–128.
Benner, R., Pakulski, J.D., McCarthy, M., Hedges, J.I., Hatcher, P.G., 1992. Bulk chemicalcharacteristics of dissolved organic-matter in the ocean. Science 255 (5051),1561–1564.
Benner, R., Biddanda, B., Black, B., McCarthy, M., 1997. Abundance, size distribution, andstable carbon and nitrogen isotopic compositions of marine organic matter isolatedby tangential-flow ultrafiltration. Mar. Chem. 57 (3–4), 243–263.
Benner, R., Louchouarn, P., Amon, R.M.W., 2005. Terrigenous dissolved organic matter inthe Arctic Ocean and its transport to surface and deep waters of the North Atlantic.Global Biogeochem. Cycles 19 (2).
Biddanda, B., Benner, R., 1997. Carbon, nitrogen, and carbohydrate fluxes during theproduction of particulate and dissolved organic matter by marine phytoplankton.Limnol. Oceanogr. 42 (3), 506–518.
Biersmith, A., Benner, R., 1998. Carbohydrates in phytoplankton and freshly produceddissolved organic matter. Mar. Chem. 63 (1–2), 131–144.
Boon, J.J., Klap, V.A., Eglinton, T.I., 1998. Molecular characterization of microgram amountsof oceanic colloidal organicmatter by direct temperature-resolved ammonia chemicalionization mass spectrometry. Org. Geochem. 29 (5–7), 1051–1061.
Borch, N.H., Kirchmann, D.L., 1997. Concentration and composition of dissolvedcombined neutral sugars (polysaccharides) in seawater determined by HPLC-PAD.Mar. Chem. 57 (1–2), 85–95.
Boyer, J.N., 1994. Aerobic and anaerobic degradation andmineralization of 14C-chitin bywater column and sediment inocula of the York River estuary, Virginia. Appl.Environ. Microbiol. 60, 174–179.
Brophy, J.E., Carlson, D.J., 1989. Production of biologically refractory dissolved organiccarbon by natural seawater microbial populations. Deep-Sea Res. 36, 497–507.
Cadieux, J.E., Kuzio, J., Milazzo, F.H., Kropinski, A.M., 1983. Spontaneous release oflipopolysaccharide by Pseudomonas aeruginosa. J. Bacteriol. 155 (2), 817–825.
Carlson, C.A., 2002. Production and removal processes. In: Hansell, D.A., Carlson, C.A.(Eds.), Biogeochemistry of Marine Dissolved Organic Matter. Academic Press,Amsterdam, pp. 91–139.
Carlson, C.A., Ducklow, H.W., Michaels, A.F., 1994. Annual flux of dissolved organic-carbon from the euphotic zone in the northwestern Sargasso Sea. Nature 371 (6496),405–408.
Cherrier, J., Bauer, J.E., 2004. Bacterial utilization of transient plankton-deriveddissolved organic carbon and nitrogen inputs in surface ocean waters. Aquat.Microb. Ecol. 35 (3), 229–241.
Coffin, R.B., 1989. Bacterial uptake of dissolved free and combined amino acids inestuarine waters. Limnol. Oceanogr. 34 (3), 531–542.
Cowie, G.L., Hedges, J.I., 1984. Carbohydrate sources in a coastal marine environment.Geochim. Cosmochim. Acta 48, 2075–2087.
Cowie, G.L., Hedges, J.I., 1994. Biochemical indicators of diagenetic alteration in naturalorganic-matter mixtures. Nature 369 (6478), 304–307.
Dauwe, B., Middelburg, J.J., 1998. Amino acids and hexosamines as indicators of organicmatter degradation state inNorth Sea sediments. Limnol. Oceanogr. 43 (5), 782–798.
Dauwe, B., Middelburg, J.J., Van Rijswijk, P., Sinke, J., Herman, P.M.J., Heip, C.H.R., 1999a.Enzymatically hydrolyzable amino acids in North Sea sediments and their possibleimplication for sediment nutritional values. J. Mar. Res. 57 (1), 109–134.
Dauwe, B., Middleburg, J.J., Herman, P.M.J., Heip, C.H.R., 1999b. Linking diageneticalteration of amino acids and bulk organic matter reactivity. Limnol. Oceanogr. 44,1809–1814.
Davis, J., Benner, R., 2005. Seasonal trends in the abundance, composition andbioavailability of particulate and dissolved organic matter in the Chukchi/BeaufortSeas andwestern CanadaBasin. Deep-SeaRes., Part II, Top. Stud. Oceanogr. 52 (24–26),3396–3410.
Davis, J., Benner, R., 2007. Quantitative estimates of labile and semi-labile dissolvedorganic carbon in the western Arctic Ocean: a molecular approach. Limnol.Oceanogr. 52 (6), 2434–2444.
Davis, J., Kaiser, K., Benner, R., in press. Amino acid and amino sugar yields andcompositions as indicators of dissolved organic matter diagenesis, Org. Geochem.doi:10.1016/j.orggeochem.2008.12.003.
Dittmar, T., Fitznar, H.P., Kattner, G., 2001. Origin and biogeochemical cycling of organicnitrogen in the eastern Arctic Ocean as evident from D- and L-amino acids. Geochim.Cosmochim. Acta 65 (22), 4103–4114.
Druffel, E.R.M.,Williams, P.M.,Bauer, J.E., Ertel, J.R.,1992.Cyclingofdissolvedandparticulateorganic-matter in the open ocean. J. Geophys. Res.[Ocean] 97 (C10), 15639–15659.
Hama, T., Yanagi, K., 2001. Production and neutral aldose composition of dissolvedcarbohydrates excreted by natural marine phytoplankton populations. Limnol.Oceanogr. 46 (8), 1945–1955.
Hama, T., Yanagi, K., Hama, J., 2004. Decrease in molecular weight of photosyntheticproducts ofmarine phytoplankton during early diagenesis. Limnol. Oceanogr. 49 (2),471–481.
Hansell, D.A., Carlson, C.A., 1998. Deep-ocean gradients in the concentration ofdissolved organic carbon. Nature 395 (6699), 263–266.
Harvey, H.R., Mannino, A., 2001. The chemical composition and cycling of particulateandmacromolecular dissolved organic matter in temperate estuaries as revealed bymolecular organic tracers. Org. Geochem. 32 (4), 527–542.
Hecky, R.E., Mopper, K., Kilham, P., Degens, E.T., 1973. The amino acid and sugarcomposition of diatom cell-walls. Mar. Biol. 19, 51–60.
Hedges, J.I., Cowie, G.L., Richey, J.E., Quay, P.D., Benner, R., Strom, M., Forsberg, B.R., 1994.Origins and processing of organic-matter in the Amazon River as indicated bycarbohydrates and amino-acids. Limnol. Oceanogr. 39 (4), 743–761.
Hedges, J.I., Eglinton, G., Hatcher, P.G., Kirchman, D.L., Arnosti, C., Derenne, S., Evershed,R.P., Kogel-Knabner, I., de Leeuw, J.W., Littke, R., Michaelis, W., Rullkotter, J., 2000a.Themolecularly-uncharacterized component of nonliving organic matter in naturalenvironments. Org. Geochem. 31 (10), 945–958.
Hedges, J.I., Mayorga, E., Tsamakis, E., McClain, M.E., Aufdenkampe, A., Quay, P., Richey, J.E.,Benner, R., Opsahl, S., Black, B., Pimentel, T., Quintanilla, J., Maurice, L., 2000b. Organicmatter in Bolivian tributaries of the Amazon River: a comparison to the lowermainstream. Limnol. Oceanogr. 45 (7), 1449–1466.
Hedges, J.I., Baldock, J.A., Gelinas, Y., Lee, C., Peterson, M., Wakeham, S.G., 2001. Evidencefor non-selective preservation of organic matter in sinking marine particles. Nature49 (15), 801–804.
Henrichs, S.M., 1991. Methods of sample handling and analysis for dissolved andparticulate amino acids and carbohydrates in seawater. In: Hurd, D.C., Spencer, D.W.(Eds.), Marine Particles: Analysis and Characterization. Geophysical MonographSeries. American Geophysical Union, Washington, pp. 139–149.
Hernes, P.J., Hedges, J.I., Peterson, M.L., Wakeham, S.G., Lee, C., 1996. Neutralcarbohydrate geochemistry of particulate material in the central equatorial Pacific.Deep-Sea Res., Part II, Top. Stud. Oceanogr. 43 (4–6), 1181–1204.
Hung, C.C., Tang, D.G., Warnken, K.W., Santschi, P.H., 2001. Distributions of carbohydrates,including uronic acids, in estuarine waters of Galveston Bay. Mar. Chem. 73 (3–4),305–318.
Ittekkot, V., Brockmann, U., Michaelis, W., Degens, E.T., 1981. Dissolved free andcombined carbohydrates during a phytoplankton bloom in the northern North-Sea.Mar. Ecol. Prog. Ser. 4 (3), 299–305.
Ittekkot, V., Arian, R., 1986. Nature of particulate organic matter in the river Indus,Pakistan. Geochim. Cosmochim. Acta 50, 1643–1653.
Hubberten, U., Lara, R.J., Kattner, G., 1995. Refractory organic-compounds in polarwaters— relationship between humic substances and amino-acids in the Arctic andAntarctic. J. Mar. Res. 53 (1), 137–149.
Jorgensen, N.O.G., Tranvik, L.J., Berg, G.M., 1999. Occurrence and bacterial cycling ofdissolved nitrogen in the Gulf of Riga, the Baltic Sea. Mar. Ecol. Prog. Ser. 191, 1–18.
Jorgensen, N.O.G., Stepanaukas, R., Pedersen, A.G.U., Hansen, M., Nybroe, O., 2003.Occurrence and degradation of peptidoglycan in aquatic environments. FEMSMicrobiol. Ecol. 46 (3), 269–280.
Kaiser, K., Benner, R., 2000. Determination of amino sugars in environmental sampleswith high salt content by high performance anion exchange chromatography andpulsed amperometric detection. Anal. Chem. 72 (11), 2566–2572.
Kaiser, K., Benner, R., 2005. Hydrolysis induced racemization of amino acids. Limnol.Oceanogr.-Methods 3, 318–325.
Kaiser, K., Benner, R., 2008. Major bacterial contribution to the ocean reservoir ofdetrital organic carbon and nitrogen. Limnol. Oceanogr. 53 (1), 99–112.
Karl, D.M., 1999. A sea of change: biogeochemical variability in the North PacificSubtropical Gyre. Ecosystems 2 (3), 181–214.
Karl, D.M., Michaels, A.F., 1996. Ocean time-series: results from the Hawaii and Bermudaresearch programs. Deep-Sea Res., Part II, Top. Stud. Oceanogr. 43.
Kawasaki, N., Benner, R., 2006. Bacterial release of dissolved organic matter during cellgrowth and decline: molecular origin and composition. Limnol. Oceanogr. 51 (5),2170–2180.
Keil, R.G., Kirchman, D.L., 1993. Dissolved combined amino-acids — chemical form andutilization by marine-bacteria. Limnol. Oceanogr. 38 (6), 1256–1270.
Kepkay, P.E., 1994. Particle aggregation and the biological reactivity of colloids. Mar.Ecol. Prog. Ser. 109, 293–304.

77K. Kaiser, R. Benner / Marine Chemistry 113 (2009) 63–77
Koolman, J., Roehme, K.-H., 2005. Color Atlas of Biochemistry, second ed. Thieme,Stuttgart.
Lee, C.L., Bada, J., 1975. Amino acids in equatorial Pacific Ocean water. Earth Planet. Sci.Lett. 26, 61–68.
Lee, C., Bada, J.L., 1977. Dissolved amino acids in the equatorial Pacific, the Sargasso Sea,and Biscayne Bay. Limnol. Oceanogr. 22 (3), 502–510.
Lee, C., Wakeham, S.G., Hedges, J.I., 2000. Composition and flux of particulate aminoacids and chloropigments in equatorial Pacific seawater and sediments. Deep-SeaRes., Part I, Oceanogr. Res. Pap. 47 (8), 1535–1568.
Liebezeit, G., 1993. Amino sugars in Bransfield Strait and Weddell Sea sediments.Senkenb. Marit. 23 (1/3), 29–35.
Lindroth, P., Mopper, K., 1979. High performance liquid chromatographic determinationof subpicomole amounts of amino acids by precolumn fluorescence derivatizationwith o-phthaldialdehyde. Anal. Chem. 51, 1667–1674.
Loh, A.N., Bauer, J.E., Druffel, E.R.M., 2004. Variable ageing and storage of dissolvedorganic components in the open ocean. Nature 430 (7002), 877–881.
Mannino, A., Harvey, H.R., 2000. Biochemical composition of particles and dissolvedorganic matter along an estuarine gradient: sources and implications for DOMreactivity. Limnol. Oceanogr. 45 (4), 775–788.
March, J., 1992. Advanced Organic Chemistry, fourth ed. JohnWiley and Sons, New York.Martinez, J.S., Zhang, G.P., Holt, P.D., Jung, H.T., Carrano, C.J., Haygood,M.G., Butler, A., 2000.
Self-assembling amphiphilic siderophores from marine bacteria. Science 287 (5456),1245–1247.
Matsumoto, K., 2007. Radiocarbon-based circulation age of theworld oceans. J. Geophys.Res.[Ocean] 112 (C9).
Meon, B., Kirchman, D.L., 2001. Dynamics and molecular composition of dissolvedorganic material during experimental phytoplankton blooms. Mar. Chem. 75 (3),185–199.
McCarthy, M.D., Hedges, J.I., Benner, R., 1996. Major biochemical composition ofdissolved high molecular weight organic matter in seawater. Mar. Chem. 55 (3–4),281–297.
McCarthy, M.D., Pratum, T., Hedges, J.I., Benner, R., 1997. Chemical composition ofdissolved organic nitrogen in the ocean. Nature 390 (6656), 150–154.
McCarthy, M.D., Hedges, J.I., Benner, R., 1998. Major bacterial contribution to marinedissolved organic nitrogen. Science 281 (5374), 231–234.
Middelboe, M., Jorgensen, N.O.G., 2006. Viral lysis of bacteia: an important source ofdissolved amino acids and cell wall compounds. J. Mar. Biol. Assoc. U.K. 86, 605–612.
Mopper, K., Kieber, D.J., 2002. Photochemistry of carbon, sulfur, nitrogen andphosphorus. In: Hansell, D.A., Carlson, C.A. (Eds.), Biogeochemistry of MarineDissolved Organic Matter. Academic Press, Amsterdam, pp. 59–85.
Mopper, K., Zhou, X.L., Kieber, R.J., Kieber, D.J., Sikorski, R.J., Jones, R.D., 1991.Photochemical degradation of dissolved organic-carbon and its impact on theoceanic carbon-cycle. Nature 353 (6339), 60–62.
Nagata, T., Fukuda, R., Koike, I., Kogure, K., Kirchman, D.L., 1998. Degradation by bacteriaof membrane and soluble protein in seawater. Aquat. Microb. Ecol. 14, 29–37.
Nagata, T., 2000. Productionmechanisms of dissolved organic matter. In: Kirchman, D.L.(Ed.), Microbial ecology of the oceans. Wiley-Liss, New York, pp. 121–152.
Nguyen, R.T., Harvey, H.R., 1997. Protein and amino acid cycling during phytoplanktondecomposition in oxic and anoxic waters. Org. Geochem. 27 (3–4), 115–128.
Panagiotopoulos, C., Repeta, D.J., Johnson, C.G., 2007. Characterization of methyl sugars,3-deoxysugars and methyl deoxysugars in marine high molecular weight dissolvedorganic matter. Org. Geochem. 38 (6), 884–896.
Pakulski, J.D., Benner, R., 1992. An improvedmethod for the hydrolysis andMBTH analysisof dissolved and particulate carbohydrates in seawater. Mar. Chem. 40 (3–4),143–160.
Ogawa, H., Amagai, Y., Koike, I., Kaiser, K., Benner, R., 2001. Production of refractorydissolved organic matter by bacteria. Science 292 (5518), 917–920.
Opsahl, S., Benner, R., 1998. Photochemical reactivity of dissolved lignin in river andocean waters. Limnol. Oceanogr. 43 (6), 1297–1304.
Quan, T.M., Repeta, D.J., 2007. Periodate oxidation of marine high molecular weightdissolved organic matter: evidence for a major contribution from 6-deoxy- andmethyl sugars. Mar. Chem. 105 (3–4), 183–193.
Sannigrahi, P., Ingall, E.D., Benner, R., 2005. Cycling of dissolved and particulate organicmatter at station Aloha: insights from C-13 NMR spectroscopy coupled withelemental, isotopic and molecular analyses. Deep-Sea Res., Part I, Oceanogr. Res.Pap. 52 (8), 1429–1444.
Schachter, H., Brockhausen, I., 1992. Glycoconjugates: Composition, Structure, andFunction. Dekker, New York, pp. 263–332.
Schaffer, C., Messner, P., 2001. Glycobiology of surface layer proteins. Biochimie 83 (7),591–599.
Seitzinger, S.P., Hartnett, H., Lauck, R., Mazurek, M., Minegishi, T., Spyres, G., Styles, R.,2005. Molecular-level chemical characterization and bioavailability of dissolvedorganic matter in stream water using electrospray-ionization mass spectrometry.Limnol. Oceanogr. 50 (1), 1–12.
Simon, M., Rosenstock, B., 2007. Different coupling of dissolved amino acid, protein, andcarbohydrate turnover to heterotrophic picoplankton production in the SouthernOcean in austral summer and fall. Limnol. Oceanogr. 52 (1), 85–95.
Skoog, A., Benner, R., 1997. Aldoses in various size fractions of marine organic matter:implications for carbon cycling. Limnol. Oceanogr. 42 (8), 1803–1813.
Smethie, W.M., Fine, R.A., Putzka, A., Jones, E.P., 2000. Tracing the flow of North AtlanticDeepWaterusing chlorofluorocarbons. J.Geophys. Res.[Ocean]105(C6),14297–14323.
Sober, H.A., 1970. Handbook for Molecular Biology, second ed. The Chemical Rubber Co.,Cleveland.
Sonnerup, R.E., Quay, P.D., Bullister, J.L., 1999. Thermocline ventilation and oxygenutilization rates in the subtropical North Pacific based on CFC distributions duringWOCE. Deep-Sea Res., Part I, Oceanogr. Res. Pap. 46 (5), 777–805.
Steinberg, D.K., Carlson, C.A., Bates, N.R., Johnson, R.J., Michaels, A.F., Knap, A.H., 2001.Overview of the US JGOFS Bermuda Atlantic Time-series Study (BATS): a decade-scale look at ocean biology and biogeochemistry. Deep-Sea Res., Part II, Top. Stud.Oceanogr. 48 (8–9), 1405–1447.
Stuiver, M., Quay, P.D., Ostlund, H.G., 1983. Abyssal water C-14 distribution and the ageof the world oceans. Science 219 (4586), 849–851.
Teira, E., Lebaron, P., van Aken, H., Herndl, G.J., 2006. Distribution and activity of Bacteriaand Archaea in the deepwatermasses of the North Atlantic. Limnol. Oceanogr. 51 (5),2131–2144.
Wakeham, S.G., Lee, C., Hedges, J.I., Hernes, P.J., Peterson, M.L., 1997. Molecularindicators of diagenetic status inmarine organic matter. Geochim. Cosmochim. Acta61 (24), 5363–5369.
Wakeham, S.G., Pease, T.K., Benner, R., 2003. Hydroxy fatty acids in marine dissolvedorganic matter as indicators of bacterial membrane material. Org. Geochem. 34 (6),857–868.
Warner, M.J., Bullister, J.L., Wisegarver, D.P., Gammon, R.H., Weiss, R.F., 1996. Basin-widedistributionsof chlorofluorocarbonsCFC-11andCFC-12 in thenorthPacific: 1985–1989.J. Geophys. Res.[Ocean] 101 (C9), 20525–20542.
Wells, M.L., Goldberg, E.D., 1993. Colloid aggregation in seawater. Mar. Chem. 41,353–358.
Williams, P.M., Druffel, E.R.M., 1987. Radiocarbon in dissolved organic-matter in thecentral north Pacific-Ocean. Nature 330 (6145), 246–248.
Wold, F., 1981. In vivo chemical modification of proteins. Annu. Rev. Biochem. 50,783–814.
Yamashita, Y., Tanoue, E., 2003. Distribution and alteration of amino acids in bulk DOMalong a transect from bay to oceanic waters. Mar. Chem. 82 (3–4), 145–160.
Zou, L., Wang, X.C., Callahan, J., Culp, R.A., Chen, R.F., Altabet, M.A., Sun, M.Y., 2004.Bacterial roles in the formation of high-molecular-weight dissolved organic matterin estuarine and coastal waters: evidence from lipids and the compound-specificisotopic ratios. Limnol. Oceanogr. 49 (1), 297–302.




















