
Clinical, biological, and histological evaluations of Mécano-Stimulation™
Upload
independentCategory
view
1download
0
Aquaculture 312 (2011) 117–125
Contents lists available at ScienceDirect
Aquaculture
j ourna l homepage: www.e lsev ie r.com/ locate /aqua-on l ine
Liver alterations induced by long term feeding on commercial diets in Atlantichalibut (Hippoglossus hippoglossus L.) females. Histological and biochemical aspects
Sylvie Bolla ⁎, Ove Nicolaisen, Anil AminFaculty of Biosciences and Aquaculture, Bodø University College, N-8049 Bodø, Norway
⁎ Corresponding author. Tel.: +47 75517451; fax: +E-mail address: [email protected] (S. Bolla).
0044-8486/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.aquaculture.2010.12.019
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 September 2010Received in revised form 8 December 2010Accepted 11 December 2010Available online 21 December 2010
Keywords:Brood stock nutritionAtlantic halibutLiver histologyLipid deposition
In the absence of a specific diet for Atlantic halibut, the commercial brood stocks are fed different andrandomly chosen diets, ranging from frozen fish to commercial dry feeds and premix. Since egg quality islargely dependent on the vitellogenin synthesised in the liver, hepatohistology of Atlantic halibut femalebrood stock and recruits was used to screen long-term effects of 3 diets commonly used in intensiveproduction, with livers of wild caught females as reference. Specific differences in liver histology wereobserved between the 3 different diets and the wild specimens. A severe macrovesicular steatosis wasobserved in females fed supplemented herring. The hepatomegaly and the profound histological changesobserved in these females address the functionality of the liver. Females fed a wet diet showed a moderatelipid and glycogen accumulation, while specimens on a dry marine diet presented an important glycogenaccumulation with lipid inclusions. Macrophage aggregates were observed in all specimens and the volumedensity was not significantly affected by the diet. Broad variations could be observed between individuals of asame group. Hemosiderin, melanin and ceroid deposition was limited in all specimens. The biochemicalanalyses confirm that diets had a significant effect on total lipid content. Both dry feed and herring diets gavesignificantly higher hepatic lipid contents than moist diets (P=0.0002 and P=0.0026) and natural diet(P=0.0026 and P=0.0223). Significant differences between diets were also found for the major PL classes.Dry feed and herring diets gave lower PL content than moist (Pb0.0001 and P=0.0064) and natural diet(Pb0.0001 and P=0.0105). Principal component analysis run on FAMES indicated that the diets separateroughly into 4 clusters. The main difference between wild specimens and females fed herring were observedin the content of long chain MUFA (C≥20) and accumulation could play a role in the development ofhepatosteatosis. Thus, herring or derivates are not recommended as brood stock diet. Liver can be used as abiomarker in brood stock nutrition studies for Atlantic halibut and possibly for other marine species as well.
47 75517410.
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The liver of vertebrates, a major organ in metabolism anddetoxification, can be affected by various biotic and xenobiotic factorsthat interfere with the normal hepatic function. Long-term exposureto stressors induces liver dysfunction that develops over time intomicroscopic changes and eventually macroscopic lesions due to lipidaccumulation in the hepatocytes. A liver affected by steatosis presentsas hepatomegaly and has a creamy colour compared with the healthyorgan. With the development of intensive production and the use offormulated diets, similar histological changes have been described inseveral domesticated species (Bac et al., 1983; Spisni et al., 1998).Metabolic dysfunction is a consequence of inadequate formulation ofthe artificial diets leading to the hepatic steatosis. The aetiology of themetabolic steatosis is not yet completely understood. However
several factors have been recognised in the development of thiscondition, among them lipid-rich diets (Morais et al., 2001), vitamin Edeficiency (Tocher et al., 2002) or fatty acid imbalance (Watanabe,1982). It should be noted that hepatic lipid accumulation is in somespecies a physiological adaptation associated with special develop-mental stages such as metamorphosis (Segner and Witt, 1990) orreproductive cycles (Lie et al., 1986; Craig et al., 2000). Thedevelopment of steatosis is considered to be a pathological condition(Spisni et al., 1998; Tucker et al., 2001).
In the natural environment, Atlantic halibut has intermediaryhepatic lipid content in the range 16–22% of wet liver weight (Lovernand Sharp, 1933) and fat is mainly deposited close to the fins and inthe muscle mass (Berge and Storebakken, 1991). A moderate increaseof the Liver Index has been reported by Haug and Gulliksen (1988)during inter-season followed by a decrease prior to the reproductiveseason, indicating that part of the liver reserves are exported to thegrowing gonads during vitellogenesis. Atlantic halibut is a newspecies in aquaculture, still under domestication, and the nutritionalrequirements at different developmental stages are not equally

118 S. Bolla et al. / Aquaculture 312 (2011) 117–125
documented. Grow out phase and early juvenile stages have receivedmost of the attention in nutrition research (Berge and Storebakken,1991; Helland and Grisdale-Helland, 1998; Hamre et al., 2003, 2005;Halten et al., 2005). Increased dietary lipid level from 8 to 20% hadapparently no effect on the lipid retention in carcass and liver (Bergeand Storebakken, 1991; Hamre et al., 2003). An increase in hepaticlipid content and increasing vacuolisation with increasing dietarylipid levels was, however, observed in halibut fed more than 22%dietary lipid (Halten et al., 2005; Martins et al., 2007). Macronutrientrequirements in females at reproductive age are poorly documented(Mazorra et al., 2003; Nordtvedt et al., 2003), mainly due to the sizeand limited numbers of brood fish available for nutritional studies.Nowadays, with the recruitment of brood fish of the F1 and F2generations, appropriate diets are crucial for retaining fecundity, eggquality and brood stock welfare, and thus for further development inproduction. In the absence of specific brood stock feed for Atlantichalibut, diets generally are chosen at random. Different diets, rangingfrom frozen fish to various commercial diets are fed to the commercialbrood stocks. Liver histology has been used to assess nutritional statusin wild juveniles (McFadzen et al., 1997) and effects of different dietformulations at early stages in reared juveniles (Bac et al., 1983; Spisniet al., 1998; Crespo et al., 2001). However histology has not beenapplied to evaluate influence of diet on liver structure of eggproducing females, even though egg quality is largely dependent onvitellogenin synthesised in the liver (Ng and Idler, 1983).
In the present study, liver histology of Atlantic halibut femalebrood stock and recruits was used to screen the effect of 3 dietscommonly used in intensive production. Livers of wild females caughtat the same period of the year, and thus in the same reproductivestage, were used as a reference. Hepatic lipid class and fatty acidcomposition are discussed in connection with the histologicalfeatures.
2. Materials and methods
2.1. Wild caught females
Wild Atlantic halibut (Hippoglossus hippoglossus L.) were caught bya professional fisherman using gillnets at a depth of approx. 150 mfrom the Salten fjord (67°16′N and 14°33′W) from December 7th to14th 2007. The 6 specimens were weighed and total length wasmeasured on board. The internal organs including gut and gonadswere kept cool during the short transportation to land andimmediately processed after reception. All stomachs and intestineswere inspected for feed content. The liver and gonads were weighedto the nearest g and inspected for colour, structure and the presence ofparasites. Samples for histology were taken at the periphery of thehepatic lobe and immediately fixated in cold buffered 4% formalde-hyde pH 7.4 at a volume ratio of approx. 1:10.
2.2. Halibut brood stock
The halibut brood stock and recruits at Bodø University CollegeResearch Station are kept in a separated section consisting of 3 landbased holding tanks of 10 m diameter, water depth of 2 m andcovered with light-proof tents. Netlon™ was used as bottomsubstrate. The tanks were supplied with 50 m deep seawater pumpedfrom the Salten fjord, filtrated and aerated before use. The flow ratewas adjusted to keep an oxygen level above 80% saturation in theoutlet, measured daily with an OxyGuard handy. Temperature overthe years was in the range 5.2 to 10.3 °C and salinity between 33.2 and33.8‰. Low light intensity was used for general illumination undernatural photoperiod. All fish were tagged with internal PIT tags andexternal identification by livestock ear-tags in the dorsal fin. The fishwere hand fed throughout the year to satiation every second day, 3times a week over a 3 year period before sampling. All fish were
weighed from weeks 34 to 37 in 2005. The specific growth rate wascalculated as: SGR=Ln(W1/W0)/d, with W1 total body weight atsampling, W0 live weight in 2005 and d number of days of the feedingperiod.
The tank fed supplemented herring had a stocking density of10.8 kg/m². The size of the females ranged from 43 to 98 kg with a sexratio ♀: ♂ of approx. 70:30. The diet consisted of headed and guttedfirst class herring (Domstein AS) with the body cavity filled withmoist feed prepared from the premix Fish Breed M, INVE, Belgium.The herrings were also supplemented with a special vitamin/mineralmixture for halibut brood stock provided by the National Institute ofNutrition and Seafood Research, Bergen, Norway.
In the tank fed a dry commercial feed, stocking density was 14 kg/m2 and sex ratio ♀: ♂ approx. 70:30. The immature recruits of thisholding tank were in the size range 3 to 7 kg. The commercial diet wasa regular dry feed for flatfish grow out (Europa Marin S, Skretting,Stavanger, Norway), pellet size 13 mm composed of 45% crudeprotein, 24% crude fat and 6–8% carbohydrates (NFE).
In the tank fed a moist diet, the stocking density was in the range12.5–15 kg/m2 with a sex ratio ♀: ♂ of approx. 20:80. The diet con-sisted of a commercial premix (Fish Breed M, INVE, Belgium) (62%crude protein and 16% crude fat) mixed with water and 5% additionalhigh quality Omega 3 fish oil 1812 TG (18% EPA and 12% DHA) fromDenomega Nutritional Oils AS, Leknes, Norway. The mixture wasformed into sausages (diameter 30 mm, length 150 mm). Brood stockmaintenance protocol was according to the Norwegian AquacultureAct.
2.3. Sampling procedures
Large specimens N60 kg were killed with an overdose of MS 222(Argent Laboratories, Redmond WA 98052, USA). Smaller fish werekilled by a sharp blow to the head. Body weight and total length wereregistered. The liver was weighed and samples for histology weretaken in the same way as for wild specimens. All stomachs andintestines were inspected for content.
Condition factor was calculated as K=100·TW/TL3.1293 and liverindex (LI) was calculated according to the formula for femalesLI=1000·LW/TL2.841 (Haug and Gulliksen, 1988) where LW is thewet liver weight, TW is the total weight, and TL total length.
2.4. Histology
Following fixation, the samples were dehydrated in alcohol ofincreasing concentration, embedded in paraffin wax and sectioned at3 μm thickness. Sections were stained with H&E to visualise thegeneral histological structure in the liver samples. PAS and Perl'smethodswere performed as subtractive staining according to routinesused in human pathology to detect the nature of inclusions, glycogenand iron stores respectively. Histological sections were observedunder an Olympus microscope BX51 equipped with CellB software.Images were stored in TIFF format. The lipid vacuolisation wasestimated by light microscopy and semi-quantitatively classified from1+ to 3+, from mild to severe according to the classification used inhuman pathology (Franzén et al., 2005).
2.5. Stereological analysis
The stereological analysis was performed with the help of thepublic domain image processing programme ImageJ (Rasband, W.S.,ImageJ, U. S. National Institutes of Health, Bethesda, Maryland, USA,http://rsb.info.nih.gov/ij/, 1997–2009). From each individual, 5 blockswere cut from the fixed liver samples. From each block, 5 sectionsseparated from each other by 80 μm were prepared and stained withPAS. On each section, 5 randomly chosen fields were photographedleading to a total of 125 fields in each specimen. A grid of 768 points

Table 1Biological data, total hepatic lipid and relative composition of the main lipid classes(% of lipid) for wild and domesticated females of Atlantic halibut fed different diets(mean±SEM). Within a row, different superscripts indicate significant differences,Pb0.05.
Wild(n=6)
Herring(n=3)
Dry feed(n=5)
Moist diet(n=6)
Length cm 161.3±6.1 175.3±3.3 94.6±4.3 122.4±4.5Weight kg 56.7±6.4 91.3±5.7 11.8±1.3 33.4±4.8
K 0.68±0.02 0.87±0.01 0.78±0.03 0.94±0.02LI 0.53±0.05 1.56±0.15 0.55±0.07 0.87±0.08
Total lipid% 18.21±2.26a 32.27±5.25b 33.96±2.49b 13.19±1.95a
PC% 7.66±0.68a 3.20±0.00b 3.23±0.19b 9.52±0.98a
PI% 1.24±0.68a Tr. Tr. 1.62±0.18a
PE% 4.27±0.49a 1.95±0.25b 1.63±0.12b 4.48 ±0.54a
Cholesterol% 3.32±0.31a 2.1 2.33±0.08a,b 4.08±0.37a,c
FFA% 2.92±0.13 12.70±3.30 2.62±0.11 2.73±0.33TGA% 52.83±3.59b 61.50±1.50a 69.83±0.95a 51.83±2.88b
Steryl esters% 26.33±0.99a 18.50±0.50a 19.67±1.33a 23.67±1.08a
119S. Bolla et al. / Aquaculture 312 (2011) 117–125
(Pr) was added to the pictures. The macrophage aggregates werestudied at ×200 magnification. Volume density was calculated as: Vv
(macrophages, liver)=(Ps/Pr)⁎100 as described by Weibel et al.,1969. The number of macrophage aggregates (n) on each field wasrecorded and the relative size of macrophage aggregates (RMS) wascalculated as the ratio Ps/n. The lipid vacuolisation was studied on atotal of 45 fields from each specimen at ×400 magnification. Threefields were photographed on each of the 5 blocks, and 3 fields on eachpicture were randomly analysed with a grid consisting of 150 points.The volume density for lipid vacuolisation Vv (lipid vacuoles, liver)was determined with the same formula as previously mentioned formacrophage aggregates.
2.6. Lipid classes and fatty acid analyses
The samples were stored at −40 °C under nitrogen atmosphereuntil analysis. Total lipid content was determined gravimetricallyafter extraction of fresh tissue with ehylacetate (NorwegianStandard NS9402.E). For fatty acid analyses, lipids were extractedaccording to Bligh and Dyer (1959) modified from freeze-driedduplicated samples. Fatty acid methyl esters were preparedaccording to Metcalfe et al. (1966) modified on wet material andC21:0 was used as intern standard. The lipids were first saponified in0.5 M sodium hydroxide in methanol for 15 min at 100 °C followedby methylation in 10% boron trifluoride in methanol for 5 min at100 °C. FAMEs were extracted with hexane and analysed on a gaschromatograph Agilent 6890 equipped with a fused silica capillarycolumn containing polysiloxane as stationary phase (0.1 mm ID,20 m, art no: Agilent 127-2222 DB-225), split/splitless injector andflame ionization detector. Hydrogen was used as carrier gas. Theinjector and detector temperatures were set to 260 °C and 250 °Crespectively and the column temperature was kept at 90 °C for4 min post injection. The column temperature was then increased to165 °C at a rate of 30 °C min−1, followed by an increase to 225 °Cwith a rate of 3 °C min−1 and kept then for 10 min. Peaks wereidentified by comparison with known standards (Nu-Chek Prep Inc,Elysian, MN, USA) and quantified by means of the internalnormalization of peaks areas standard C21:0. Data processing wasperformed with a Chemstation integrator (A0903). Phospholipidswere separated by HPTLC and characterized by densitometrydetection after development (Analyses purchased at NIFES, Bergen,Norway).
2.7. Statistics
Data are presented as means with standard error of the mean(SEM). Statistical analyses on biological measures, total lipid, lipidclasses and fatty acids were run on individual readings, whilehistological measures were run on mean values obtained from thecollective readings on each individual fish. Each response variablewas analysed separately, using General Linear Model (GLM)procedure, initially including diet as fixed factor and length as acovariate. Though, as there was no significant effect of length onany of the response variables, it was removed from all models.Assumptions of homoscedasticity and normality were checkedusing Levene's test and graphical examination of model residuals,respectively. For the response variables LI, K, RMS, TAG and Vv (lipidvacuoles), assumptions were initially met. Vv (macrophages) and PEwere log 10-transformed, and for Vv (macrophages), one extremeoutlier was removed. Following significant GLMs, Bonferroni posthoc tests were used to compare pairs of means between diets.Kruskall–Wallis test was used to analyse effect of diet on PC and PI,as transformations did not stabilize the between group variance.Fatty acid composition between diet classes was explored usingPrincipal Component Analysis (PCA), followed by GLMs for selectedfatty acid classes. Level of significance for all tests was set to Pb0.05.
GLM and correlation analysis was carried out using the statisticalpackage SPSS 15.0 for Windows (SPSS Inc., Chicago, IL, USA), andThe Unscrambler® Ver. 9.2 (Camo Software Inc.) was used for PCA.
3. Results
3.1. Biological data
Data on body length and weight, condition index and liver indexfor diet groups are given in Table 1. Large reproductive females hadthe lowest SGR, 0.017±0.01 and 0.027±0.04% a day for supple-mented herring and moist diet respectively. For dry feed, SGR was0.169±0.02% a day. Diet had significant effect on condition factor K(GLM, F3,16=25.13, Pb0.0001). Wild caught females had signifi-cantly lower K compared to females fed herring (P=0.0014) andwet feed (Pb0.0001). However, differences were not significantbetween wild fish and fish fed dry feed (P=0.1215).
The surface of liver of wild caught specimens had a uniform pinkcolour. The liver of domesticated fish presented some creamy patchesin dry and wet diets, while females fed on supplemented herring, hada uniformly yellowish coloured liver. Diet had significant effect on theliver index LI (GLM, F3,16=22.47, Pb0.0001). The LI of females fedsupplemented herring was significantly higher than for all other diets(Pb0.001). There were no significant differences between wildspecimens and females fed dry feed (P=1.000), while females fedmoist feed had significantly higher LI than wild specimens(P=0.0354).
3.2. Liver histology of wild caught Atlantic halibut
At lowmagnification the liver tissue showed a zonal distribution ofareas with low tissue density found around the portal system (Fig. 1a).At higher magnification these areas showed a higher degree ofvacuolisation with discrete, rounded macro and micro lipid vacuoles.The degree of vacuolisation varied between specimens, from 1+ to 2+
for 3 of the 6 females, while some areas with a vacuolisation degree upto 3+ could be detected around the portal system of the 3 otherspecimens. Females with a LI≤0.500 presented a grade of vacuolisa-tion from1+ to 2+. The volume density Vv (lipid vacuoles, liver) in thesample was 36.1±3.1% (range: 24.2–43.7%). The tubular parenchymaarrangement, the sinusoids and the spaces of Disse were evident inareas with low or intermediary degrees of vacuolisation. Aneffacement of these structures was observed in zones of highervacuolisation, giving a solid aspect to the hepatic parenchyma. Areaswith 1+ degree of vacuolisation always were PAS− while zones 2+
presented moderate PAS+ staining along with PAS− vacuoles. All
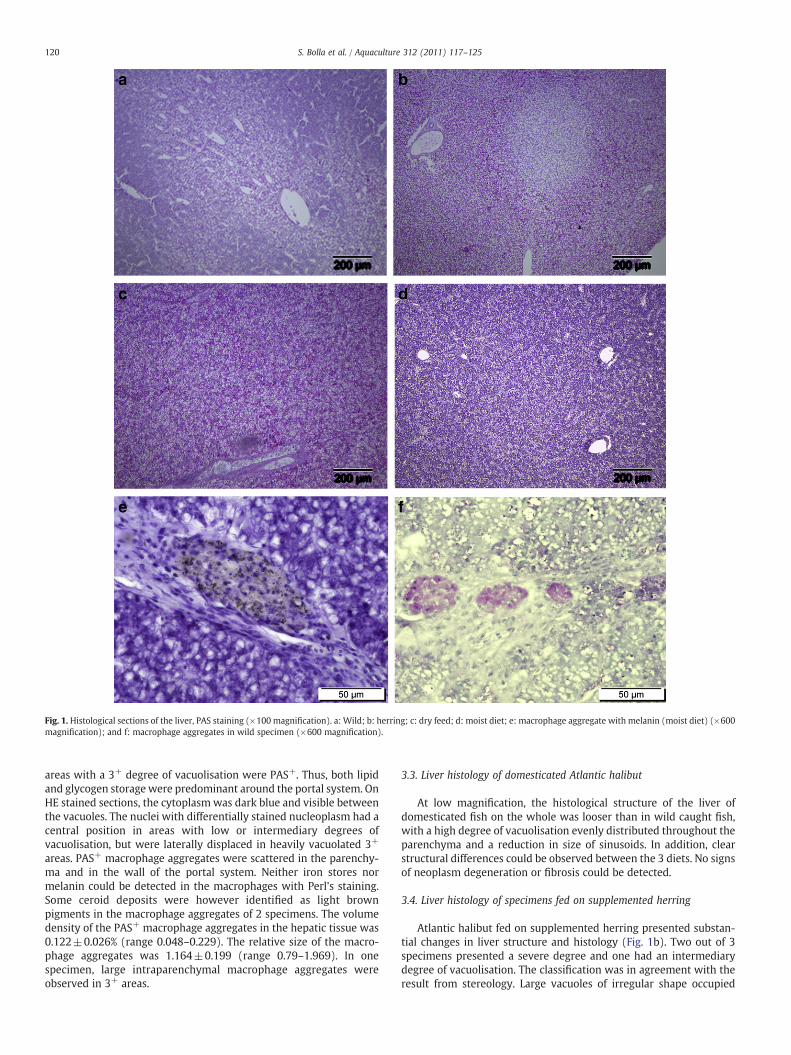
Fig. 1. Histological sections of the liver, PAS staining (×100 magnification). a: Wild; b: herring; c: dry feed; d: moist diet; e: macrophage aggregate with melanin (moist diet) (×600magnification); and f: macrophage aggregates in wild specimen (×600 magnification).
120 S. Bolla et al. / Aquaculture 312 (2011) 117–125
areas with a 3+ degree of vacuolisation were PAS+. Thus, both lipidand glycogen storage were predominant around the portal system. OnHE stained sections, the cytoplasm was dark blue and visible betweenthe vacuoles. The nuclei with differentially stained nucleoplasm had acentral position in areas with low or intermediary degrees ofvacuolisation, but were laterally displaced in heavily vacuolated 3+
areas. PAS+ macrophage aggregates were scattered in the parenchy-ma and in the wall of the portal system. Neither iron stores normelanin could be detected in the macrophages with Perl's staining.Some ceroid deposits were however identified as light brownpigments in the macrophage aggregates of 2 specimens. The volumedensity of the PAS+ macrophage aggregates in the hepatic tissue was0.122±0.026% (range 0.048–0.229). The relative size of the macro-phage aggregates was 1.164±0.199 (range 0.79–1.969). In onespecimen, large intraparenchymal macrophage aggregates wereobserved in 3+ areas.
3.3. Liver histology of domesticated Atlantic halibut
At low magnification, the histological structure of the liver ofdomesticated fish on the whole was looser than in wild caught fish,with a high degree of vacuolisation evenly distributed throughout theparenchyma and a reduction in size of sinusoids. In addition, clearstructural differences could be observed between the 3 diets. No signsof neoplasm degeneration or fibrosis could be detected.
3.4. Liver histology of specimens fed on supplemented herring
Atlantic halibut fed on supplemented herring presented substan-tial changes in liver structure and histology (Fig. 1b). Two out of 3specimens presented a severe degree and one had an intermediarydegree of vacuolisation. The classification was in agreement with theresult from stereology. Large vacuoles of irregular shape occupied
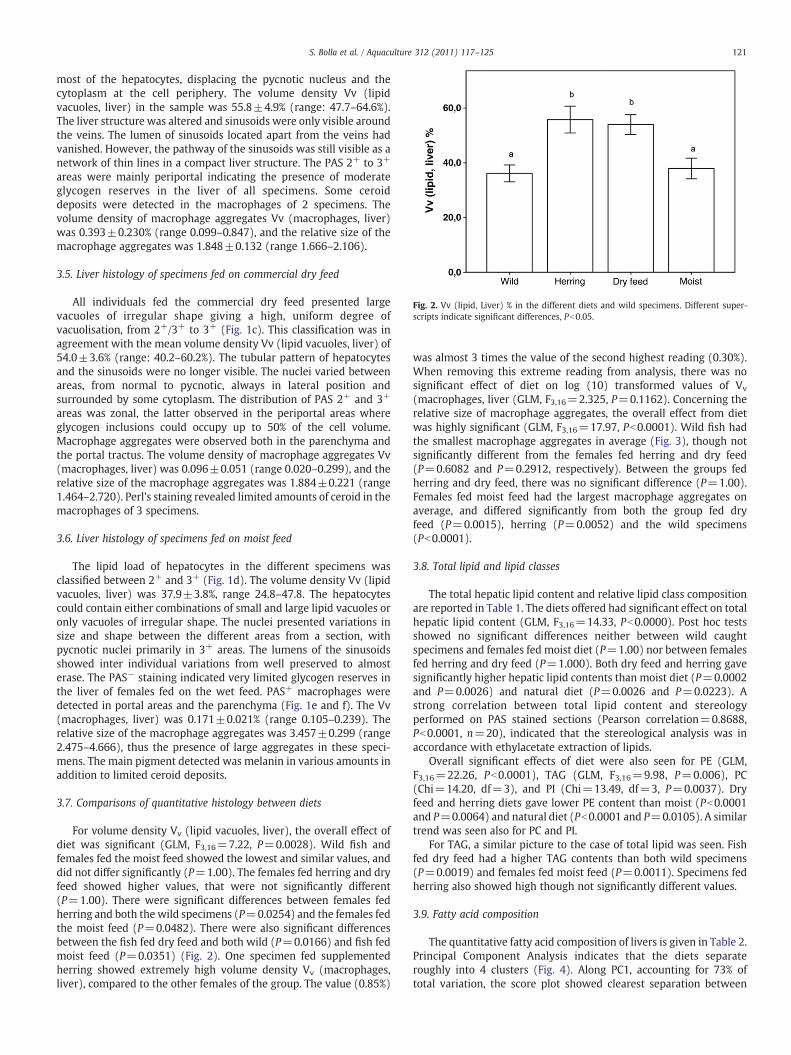
Fig. 2. Vv (lipid, Liver) % in the different diets and wild specimens. Different super-scripts indicate significant differences, Pb0.05.
121S. Bolla et al. / Aquaculture 312 (2011) 117–125
most of the hepatocytes, displacing the pycnotic nucleus and thecytoplasm at the cell periphery. The volume density Vv (lipidvacuoles, liver) in the sample was 55.8±4.9% (range: 47.7–64.6%).The liver structure was altered and sinusoids were only visible aroundthe veins. The lumen of sinusoids located apart from the veins hadvanished. However, the pathway of the sinusoids was still visible as anetwork of thin lines in a compact liver structure. The PAS 2+ to 3+
areas were mainly periportal indicating the presence of moderateglycogen reserves in the liver of all specimens. Some ceroiddeposits were detected in the macrophages of 2 specimens. Thevolume density of macrophage aggregates Vv (macrophages, liver)was 0.393±0.230% (range 0.099–0.847), and the relative size of themacrophage aggregates was 1.848±0.132 (range 1.666–2.106).
3.5. Liver histology of specimens fed on commercial dry feed
All individuals fed the commercial dry feed presented largevacuoles of irregular shape giving a high, uniform degree ofvacuolisation, from 2+/3+ to 3+ (Fig. 1c). This classification was inagreement with the mean volume density Vv (lipid vacuoles, liver) of54.0±3.6% (range: 40.2–60.2%). The tubular pattern of hepatocytesand the sinusoids were no longer visible. The nuclei varied betweenareas, from normal to pycnotic, always in lateral position andsurrounded by some cytoplasm. The distribution of PAS 2+ and 3+
areas was zonal, the latter observed in the periportal areas whereglycogen inclusions could occupy up to 50% of the cell volume.Macrophage aggregates were observed both in the parenchyma andthe portal tractus. The volume density of macrophage aggregates Vv(macrophages, liver) was 0.096±0.051 (range 0.020–0.299), and therelative size of the macrophage aggregates was 1.884±0.221 (range1.464–2.720). Perl's staining revealed limited amounts of ceroid in themacrophages of 3 specimens.
3.6. Liver histology of specimens fed on moist feed
The lipid load of hepatocytes in the different specimens wasclassified between 2+ and 3+ (Fig. 1d). The volume density Vv (lipidvacuoles, liver) was 37.9±3.8%, range 24.8–47.8. The hepatocytescould contain either combinations of small and large lipid vacuoles oronly vacuoles of irregular shape. The nuclei presented variations insize and shape between the different areas from a section, withpycnotic nuclei primarily in 3+ areas. The lumens of the sinusoidsshowed inter individual variations from well preserved to almosterase. The PAS− staining indicated very limited glycogen reserves inthe liver of females fed on the wet feed. PAS+ macrophages weredetected in portal areas and the parenchyma (Fig. 1e and f). The Vv(macrophages, liver) was 0.171±0.021% (range 0.105–0.239). Therelative size of the macrophage aggregates was 3.457±0.299 (range2.475–4.666), thus the presence of large aggregates in these speci-mens. The main pigment detected was melanin in various amounts inaddition to limited ceroid deposits.
3.7. Comparisons of quantitative histology between diets
For volume density Vv (lipid vacuoles, liver), the overall effect ofdiet was significant (GLM, F3,16=7.22, P=0.0028). Wild fish andfemales fed the moist feed showed the lowest and similar values, anddid not differ significantly (P=1.00). The females fed herring and dryfeed showed higher values, that were not significantly different(P=1.00). There were significant differences between females fedherring and both the wild specimens (P=0.0254) and the females fedthe moist feed (P=0.0482). There were also significant differencesbetween the fish fed dry feed and both wild (P=0.0166) and fish fedmoist feed (P=0.0351) (Fig. 2). One specimen fed supplementedherring showed extremely high volume density Vv (macrophages,liver), compared to the other females of the group. The value (0.85%)
was almost 3 times the value of the second highest reading (0.30%).When removing this extreme reading from analysis, there was nosignificant effect of diet on log (10) transformed values of Vv
(macrophages, liver (GLM, F3,16=2.325, P=0.1162). Concerning therelative size of macrophage aggregates, the overall effect from dietwas highly significant (GLM, F3,16=17.97, Pb0.0001). Wild fish hadthe smallest macrophage aggregates in average (Fig. 3), though notsignificantly different from the females fed herring and dry feed(P=0.6082 and P=0.2912, respectively). Between the groups fedherring and dry feed, there was no significant difference (P=1.00).Females fed moist feed had the largest macrophage aggregates onaverage, and differed significantly from both the group fed dryfeed (P=0.0015), herring (P=0.0052) and the wild specimens(Pb0.0001).
3.8. Total lipid and lipid classes
The total hepatic lipid content and relative lipid class compositionare reported in Table 1. The diets offered had significant effect on totalhepatic lipid content (GLM, F3,16=14.33, Pb0.0000). Post hoc testsshowed no significant differences neither between wild caughtspecimens and females fed moist diet (P=1.00) nor between femalesfed herring and dry feed (P=1.000). Both dry feed and herring gavesignificantly higher hepatic lipid contents than moist diet (P=0.0002and P=0.0026) and natural diet (P=0.0026 and P=0.0223). Astrong correlation between total lipid content and stereologyperformed on PAS stained sections (Pearson correlation=0.8688,Pb0.0001, n=20), indicated that the stereological analysis was inaccordance with ethylacetate extraction of lipids.
Overall significant effects of diet were also seen for PE (GLM,F3,16=22.26, Pb0.0001), TAG (GLM, F3,16=9.98, P=0.006), PC(Chi=14.20, df=3), and PI (Chi=13.49, df=3, P=0.0037). Dryfeed and herring diets gave lower PE content than moist (Pb0.0001and P=0.0064) and natural diet (Pb0.0001 and P=0.0105). A similartrend was seen also for PC and PI.
For TAG, a similar picture to the case of total lipid was seen. Fishfed dry feed had a higher TAG contents than both wild specimens(P=0.0019) and females fed moist feed (P=0.0011). Specimens fedherring also showed high though not significantly different values.
3.9. Fatty acid composition
The quantitative fatty acid composition of livers is given in Table 2.Principal Component Analysis indicates that the diets separateroughly into 4 clusters (Fig. 4). Along PC1, accounting for 73% oftotal variation, the score plot showed clearest separation between
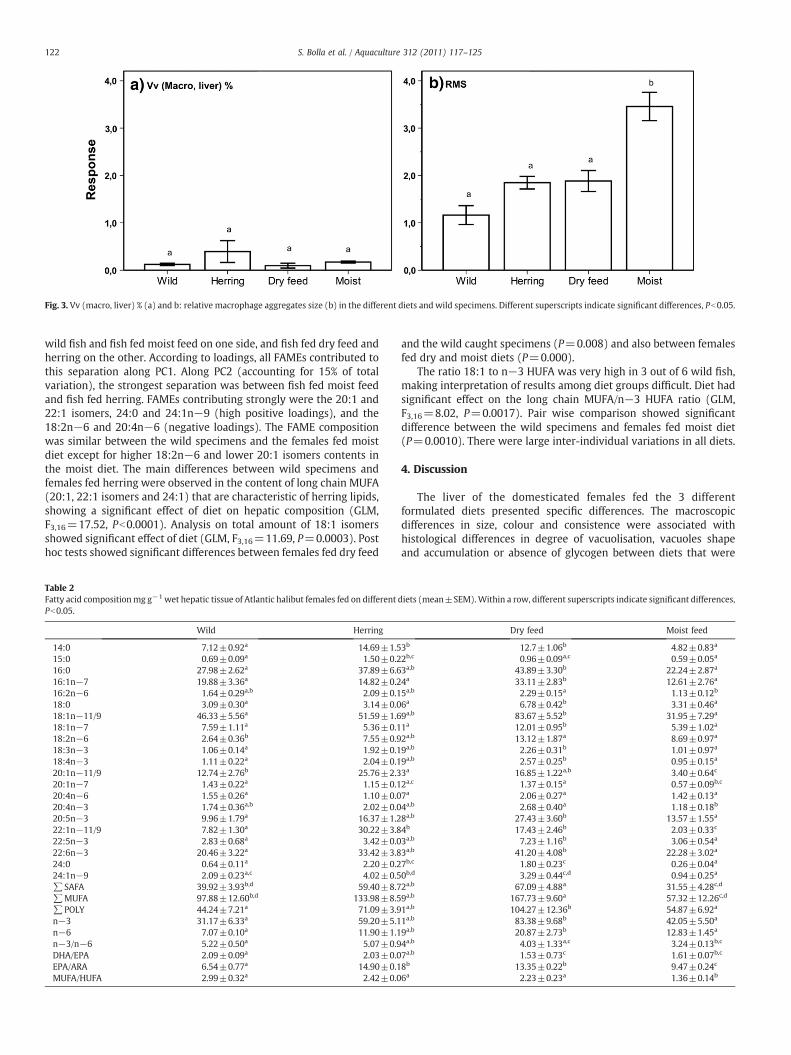
Fig. 3. Vv (macro, liver) % (a) and b: relative macrophage aggregates size (b) in the different diets and wild specimens. Different superscripts indicate significant differences, Pb0.05.
122 S. Bolla et al. / Aquaculture 312 (2011) 117–125
wild fish and fish fed moist feed on one side, and fish fed dry feed andherring on the other. According to loadings, all FAMEs contributed tothis separation along PC1. Along PC2 (accounting for 15% of totalvariation), the strongest separation was between fish fed moist feedand fish fed herring. FAMEs contributing strongly were the 20:1 and22:1 isomers, 24:0 and 24:1n−9 (high positive loadings), and the18:2n−6 and 20:4n−6 (negative loadings). The FAME compositionwas similar between the wild specimens and the females fed moistdiet except for higher 18:2n−6 and lower 20:1 isomers contents inthe moist diet. The main differences between wild specimens andfemales fed herring were observed in the content of long chain MUFA(20:1, 22:1 isomers and 24:1) that are characteristic of herring lipids,showing a significant effect of diet on hepatic composition (GLM,F3,16=17.52, Pb0.0001). Analysis on total amount of 18:1 isomersshowed significant effect of diet (GLM, F3,16=11.69, P=0.0003). Posthoc tests showed significant differences between females fed dry feed
Table 2Fatty acid compositionmg g−1 wet hepatic tissue of Atlantic halibut females fed on differentPb0.05.
Wild Herring
14:0 7.12±0.92a 14.69±1.515:0 0.69±0.09a 1.50±0.216:0 27.98±2.62a 37.89±6.616:1n−7 19.88±3.36a 14.82±0.216:2n−6 1.64±0.29a,b 2.09±0.118:0 3.09±0.30a 3.14±0.018:1n−11/9 46.33±5.56a 51.59±1.618:1n−7 7.59±1.11a 5.36±0.118:2n−6 2.64±0.36b 7.55±0.918:3n−3 1.06±0.14a 1.92±0.118:4n−3 1.11±0.22a 2.04±0.120:1n−11/9 12.74±2.76b 25.76±2.320:1n−7 1.43±0.22a 1.15±0.120:4n−6 1.55±0.26a 1.10±0.020:4n−3 1.74±0.36a,b 2.02±0.020:5n−3 9.96±1.79a 16.37±1.222:1n−11/9 7.82±1.30a 30.22±3.822:5n−3 2.83±0.68a 3.42±0.022:6n−3 20.46±3.22a 33.42±3.824:0 0.64±0.11a 2.20±0.224:1n−9 2.09±0.23a,c 4.02±0.5∑SAFA 39.92±3.93b,d 59.40±8.7∑MUFA 97.88±12.60b,d 133.98±8.5∑POLY 44.24±7.21a 71.09±3.9n−3 31.17±6.33a 59.20±5.1n−6 7.07±0.10a 11.90±1.1n−3/n−6 5.22±0.50a 5.07±0.9DHA/EPA 2.09±0.09a 2.03±0.0EPA/ARA 6.54±0.77a 14.90±0.1MUFA/HUFA 2.99±0.32a 2.42±0.0
and the wild caught specimens (P=0.008) and also between femalesfed dry and moist diets (P=0.000).
The ratio 18:1 to n−3 HUFA was very high in 3 out of 6 wild fish,making interpretation of results among diet groups difficult. Diet hadsignificant effect on the long chain MUFA/n−3 HUFA ratio (GLM,F3,16=8.02, P=0.0017). Pair wise comparison showed significantdifference between the wild specimens and females fed moist diet(P=0.0010). There were large inter-individual variations in all diets.
4. Discussion
The liver of the domesticated females fed the 3 differentformulated diets presented specific differences. The macroscopicdifferences in size, colour and consistence were associated withhistological differences in degree of vacuolisation, vacuoles shapeand accumulation or absence of glycogen between diets that were
diets (mean±SEM).Within a row, different superscripts indicate significant differences,
Dry feed Moist feed
3b 12.7±1.06b 4.82±0.83a
2b,c 0.96±0.09a,c 0.59±0.05a
3a,b 43.89±3.30b 22.24±2.87a
4a 33.11±2.83b 12.61±2.76a
5a,b 2.29±0.15a 1.13±0.12b
6a 6.78±0.42b 3.31±0.46a
9a,b 83.67±5.52b 31.95±7.29a
1a 12.01±0.95b 5.39±1.02a
2a,b 13.12±1.87a 8.69±0.97a
9a,b 2.26±0.31b 1.01±0.97a
9a,b 2.57±0.25b 0.95±0.15a
3a 16.85±1.22a,b 3.40±0.64c
2a,c 1.37±0.15a 0.57±0.09b,c
7a 2.06±0.27a 1.42±0.13a
4a,b 2.68±0.40a 1.18±0.18b
8a,b 27.43±3.60b 13.57±1.55a
4b 17.43±2.46b 2.03±0.33c
3a,b 7.23±1.16b 3.06±0.54a
3a,b 41.20±4.08b 22.28±3.02a
7b,c 1.80±0.23c 0.26±0.04a
0b,d 3.29±0.44c,d 0.94±0.25a
2a,b 67.09±4.88a 31.55±4.28c,d
9a,b 167.73±9.60a 57.32±12.26c,d
1a,b 104.27±12.36b 54.87±6.92a
1a,b 83.38±9.68b 42.05±5.50a
9a,b 20.87±2.73b 12.83±1.45a
4a,b 4.03±1.33a,c 3.24±0.13b,c
7a,b 1.53±0.73c 1.61±0.07b,c
8b 13.35±0.22b 9.47±0.24c
6a 2.23±0.23a 1.36±0.14b

Fig. 4. Scores and loadings plot of Principal Component Analysis performed on the quantitative fatty acid composition of livers of Atlantic halibut females fed on different diets (●:moist; ■: dry; ▲: herring) and wild specimens (○).
123S. Bolla et al. / Aquaculture 312 (2011) 117–125
confirmed by the lipid analyses. Profound alterations were noted withsupplemented herring and dry diet. The herring diet induced a severemacrovesicular steatosis that addresses the functionality of the liversince all hepatic lesions are considered detrimental for health invertebrates. The limited ceroid deposition in macrophages isconsistent with low amounts of oxidised dietary lipids. Females fedmoist diet presented an intermediary and zonal lipid accumulationwith limited glycogen reserves while dry diet increased both lipid andglycogen inclusions.
Macrophage aggregates were found in all specimens. A broadindividual variation was observed in all groups. Average volumedensity was not significantly affected by diet, but relative macrophagesize showed a different pattern with clear diet effects. Differenceswere not significant; however fish fed moist diet presentedsignificantly larger macrophage aggregates than all other groups.The pigment load was limited in all specimens. Pigment accumulationin macrophages is under influence of many factors, such as aging(Brown and George, 1985), pollution (Wolke, 1992), health status(Jordanova et al., 2008), nutritional status and general stress underculture conditions (Montero et al., 1999; Caballero et al., 2004) as wellas tissue damages (Broeg, 2010). Seasonal variations related toreproduction with a significant increase at postspawning stage arealso reported in Ohrid trout (Jordanova et al., 2008).
As expected, SGR ranges decreasedwith increasing size. The SGR inthe given temperature range could not be evaluated due to a lack ofinformation on growth rates of fish above 10 kg, which is out of thecommercial production range. The wild specimens had lowercondition factor K than all three groups of captive fish, althoughdifferences between wild fish and the immature recruits fedcommercial dry feed were not significant. This is consistent withprevious studies showing that wild specimens are in general leanerthan fish fed ad. lib. in captivity (Grigorakis et al., 2002). The mean LIof wild specimens was 0.52 and in the range reported by Haug andGulliksen (1988) for fish at similar late maturation stage. The LI ofimmature females fed dry diet thus not different from LI of wildcaught specimens, was higher than the mean value of about 0.32 forwild immature females (Haug and Gulliksen, 1988). Both conditionfactor and LI varied considerably between groups fed different dietsunder similar rearing conditions, indicating that these changes arerelated to nutrition.
The hepatic structure of Atlantic halibut is of tubular form andtherefore of the same type as the other pleuronectiformes studied(Akiyoshi and Inoue, 2004). Glycogen storage around the sinusoids inwild caught specimens was limited. Thus the micro and macro-
vacuolisation in the hepatic tissue was mainly the result of lipidaccumulation and consistent with intermediary hepatic lipid content(Lovern et al., 1933). This zonal distribution of lipidwas assumed to bethe normal histological structure in female halibut liver towards theend of vitellogenesis, as no previous record on the hepatic histology isavailable in this species. Histological studies in flatfishes generallyaddress to pathological changes induced by environmental stressors.Due to the bottom dwelling existence, these fishes are speciallyexposed and develop various liver lesions from fat accumulation totumours (Feist et al., 2004). In flounder (Platichthys flesus) (Vethaakand Wester, 1996) and in English sole (Pleuronectes vetulus) (Myerset al., 1994) from polluted areas a similar hepatic structure to the oneobserved in wild caught specimens has been classified as storagedisorder. In contrary to the sedentary flat fish species, Atlantic halibutmigrates over long distances in relation to feeding and spawning(Devold, 1939). Nothing in our study however indicates that the wildcaught specimens have been directly exposed to environmentalstressors. However, since Atlantic halibut is a long-lived carnivorousfish, xenobiotics accumulate through the natural preys. Julshamn et al.(2007) reported dioxin and dioxin-like PCBs contents exceeding theupper limit of 8.0 ng/kg wet weight defined by the EU in halibut over120 kg, while fish between 30 and 35 kg showed values identical toother fat fishes. These data refer to a limited number of specimens sofar, and no data are available for the weight range of the present study.
The wild specimens and females fed the moist diet showedsignificantly lower hepatic lipid content than females fed herring orthe dry diet. The 2D stereological analysis was consistent with hepaticlipid contents. The application of dry diet resulted in elevated oleicacid content as described by Spisni et al. (1998) for gilthead sea breamand sea bass. These authors related the hepatic steatosis to an increasein 18:1/n−3 HUFA ratio. No such relationship could be found in thepresent study. Besides the difference in lipid class composition in thefemales fed supplemented herring, hepatic lipids presented highercontents of the 20:1 and 22:1 isomers. Long-chain MUFA showedsignificant increase in females fed herring, while MUFA/n−3 HUFAratio did not changed. These results indicate thatMUFA content ratherthan their ratio to n−3 HUFA plays a role in the development ofhepatosteatosis in Atlantic halibut. A comparison between wild andreared gilthead sea bream (Grigorakis et al., 2002) showed a doublingof the 18:1n−9 content and a significant accumulation of 20:1 and22:1 isomers in the muscle of farmed fish. These fatty acids originatefrom the fatty alcohol moieties of wax esters of calanoid copepodsfrom high latitude and are incorporated at significant levels in thelipids of fishes grazing on the calanoid copepods (Pascal and Ackman,

124 S. Bolla et al. / Aquaculture 312 (2011) 117–125
1976). In warm marine environment, only traces of the long chainmonounsaturated fatty acids are found in the natural diet (Spisniet al., 1998), themuscle in gilthead sea bream (Grigorakis et al., 2002),muscle and liver of female broodstocks of white seabream, Diplodussargus (Cejas et al., 2004). Cold water species naturally presentmoderate amounts of long chain MUFA. The long-chain MUFA areoxidised in the peroxisomes. For most of the fishes studied the fattyacyl CoA oxidase activity is lower compared with mammalian liver(Moyes et al., 1991; Crockett and Sidell, 1993). The poor efficiency inperoxisomal activity in fish combined with high dietary MUFA resultin accumulation of these fatty acids in the enterocytes. Environmentalfactors (temperature, season, and salinity), biotic factors (species, age,gender, and feeding habits) (Rochas et al., 2003) as well as nutritionalstatus (Leaver et al., 2005) strongly affect peroxisomal activity infishes. Since fishes are a largely heterogeneous group regardinghabitat and physiological adaptations, cautious interpretation isnecessary between different species. Peroxisomal activity in Atlantichalibut and the presumed role of the long-chain MUFA in thedevelopment of hepatic steatosis has to be investigated. The long-term effect of herring diet in the present study contrasts with theminor changes in HIS and fatty acid composition reported by Martinset al., 2007 in short-term experiments conducted on juveniles for13 weeks. Hepatic lipids and total body lipids increased withincreasing dietary lipids from 14 to 22%, while long-chain MUFAcontent remains similar in all treatments.
The profound histological changes observed in Atlantic halibutliver after long-term feeding on herring, indicate that herring andderivatives should not be used as ingredients in brood stock dietbefore more documentation on their effect on liver functions isavailable. Besides, herring is rarely found in the stomach content ofAtlantic halibut (Lovern and Sharp, 1933). The recruits should beselected early and offered an improved diet to minimise the hepaticglycogen accumulation observedwhen using regular marine grow outdiet.
The liver is commonly used as a biomarker in environmentalstudies, and lesions induced by xenobiotic substances have beendescribed and classified. No similar classification is available eventhough effects of nutrition on liver histology are well documented.The present results clearly show specific features induced by 3 dietson Atlantic halibut brood stock compared with wild caught speci-mens. A deeper understanding of the histological and metaboliccharacteristics of livers from wild specimens would provide a usefultool for the development of appropriate diets for marine fishes.
Acknowledgments
This study was funded by the Norwegian Research Council project176622. The authors are grateful to Olaf Pedersen and his boat RandiAnita for kindly providing the livers and data of the wild caughthalibut, Heidi Ludviksen for preparing the histological sections andEllen Sirnes for fatty acid analyses. SB designed the experiment,performed stereology and wrote the article. ON performed thestatistical analyses and wrote the manuscript. AA was involved inthe interpretation of the histological sections and commented on themanuscript.
References
Akiyoshi, H., Inoue, A., 2004. Comparative histological study of teleost livers in relationto phylogeny. Zool. Sci. 21, 841–850.
Bac, N., Biagianti, S., Bruslé, B., 1983. Etude cytologique ultrastructurale des anomalieshépatiques du loup, de la daurade et de l'anguille, induites par une alimentationartificielle. Bases Biol. Aquacult., Montpellier 1983, IFREMER. Actes Colloques 1,473–484.
Berge, G.M., Storebakken, T., 1991. Effect of dietary fat level on weight gain, digestibilityand fillet composition of Atlantic halibut. Aquaculture 99, 331–338.
Bligh, E.C., Dyer, W.J., 1959. A rapid method of total lipid extraction and purification.Can. J. Biochem. Physiol. 37, 911–927.
Broeg, K., 2010. The activity of macrophage aggregates in the liver of flounder(Platichthys flesus) and wrasse (Symphodus melops) is associated with tissuedamage. Mar. Environ. Res. 69 Suppl, S14–16. http://hdl.handle.net/10013/epic.34230.
Brown, C.L., George, C.T., 1985. Age-dependent accumulation of macrophage aggregatesin the yellow perch Perca flavescens Mitchill. J. Fish Dis. 8, 135–138.
Caballero, M.J., Izquierdo, M.S., Kjørsvik, E., Fernàndez, A.J., Rosenlund, G., 2004.Histological alterations in the liver of sea bream, Sparus aurata L., caused by short-or long-term feeding with vegetable oils. Recovery of normal morphology afterfeeding fish oil as the sole lipid source. J. Fish Dis. 27, 531–541.
Cejas, J.R., Almansa, E., Jérez, S., Bolaños, A., Samper, M., Lorenzo, A., 2004. Lipid andfatty acid composition of muscle and liver from wild and captive female broodstocksof white seabream Diplodus sargus. Comp. Biochem. Physiol., B 138, 91–102.
Craig, S.R., McKenzie, D.S., Jones, G., Delbert, M., Gatlin III, D.M., 2000. Seasonal changesin reproductive condition and body composition of free-ranging red drum,Sciaenops ocellatus. Aquaculture 190, 89–102.
Crespo, S., Marín de Mateo, M., Santamaría, C.A., Sala, R., Grau, A., Pastor, E., 2001.Histopathological observations during larval rearing of common dentex Dentexdentex L. (Sparidae). Aquaculture 192, 121–132.
Crockett, E.L., Sidell, B.D., 1993. Peroxisomal beta-oxidation is a significant pathway forcatabolism of fatty acids in a marine teleost. Am. J. Physiol. Regul. Integr. Comp.Physiol. 264, R1004–R1009.
Devold, F., 1939. Kveiteundersøkelsene i 1938. FiskDir. Skr. Ser. HavUnders. 6, 85–96.Feist S.W., Lang T., Stentiford G.D., Köhler A., 2004. Biological effects of contaminants:
use of liver pathology of the European flatfish dab (Limanda limanda L.) andflounder (Platichthys flesus L.) for monitoring. ICES Techniques in MarineEnvironmental Sciences, No. 38. 42 pp. ISSN 0903-2606 ISBN 87-7482-024-9.
Franzén, L.E., Ekstedt, M., Kechagias, S., Bodin, L., 2005. Semiquantitative evaluationoverestimates the degree of steatosis in liver biopsies: a comparison to stereologialpoint counting. Mod. Pathol. 18, 912–916.
Grigorakis, K., Alexis, M.N., Taylor, K.D.A., Hole, M., 2002. Comparison of wild andcultivated gilthead sea bream (Sparus autata); composition, appearance andseasonal variations. Int. J. Food Sci. Technol. 37, 477–484.
Halten, B., Grisdale-Helland, B., Helland, S., 2005. Growth, feed utilization and bodycomposition in two size groups of Atlantic halibut (Hippoglossus hippoglossus) feddiets differing in protein and carbohydrate content. Aquaculture 249, 401–408.
Hamre, K., Øfsti, A., Næss, T., Nortvedt, R., Holm, J.C., 2003. Macronutrient compositionin formulated diets for Atlantic halibut (Hippoglossus hippoglossus, L.) juveniles — amultivariate approach. Aquaculture 227, 233–244.
Hamre, K., Baeverfjord, G., Harboe, T., 2005. Macronutrient composition of formulateddiets for Atlantic halibut (Hippoglossus hippoglossus, L.) juveniles, II: protein/lipidlevels at low carbohydrate. Aquaculture 244, 283–291.
Haug, T., Gulliksen, B., 1988. Variations in liver and body condition during gonaddevelopment of Atlantic halibut, Hippoglossus hippoglossus (L.). FiskDir. Skr.HavUnders. 18, 351–363.
Helland, S.J., Grisdale-Helland, B., 1998. Growth, feed utilization and body compositionof juvenile Atlantic halibut (Hippoglossus hippoglossus) fed diets differing in theratio between the macronutrients. Aquaculture 166, 49–56.
Jordanova, M., Rebok, K., Miteva, N., Rocha, E., 2008. A qualitative and quantitativestudy of the hepatic pigmented macrophage aggregates during the breeding cycleoh Ohrid trout Salmo letnica Kar. (Teleostei, salmonidae). Microsopy Res. Tech. 71,822–830.
Julshamn, K., Øygard, J., Måge, A., 2007. Dioksiner, dioksinlignende PCB og andre PCBer ifiskevarer og konsumferdige fiskeoljer, bromerte flammehemmere og andre nyemiljøgifter i sjømat og tungmetaller i sjømat. Nasjonalt Institutt for Ernærings-ogSjømatforskning, Bergen, Norway. http://www.nifes.no/publikasjoner/rapporter/index.php?page_id=&article_id=2849&lang_id=1.
Leaver, M.J., Boukouvala, E., Antonopoulou, E., Diez, A., Favre-Krey, L., Tariq, Ezaz M.,Bautista, J.M., Tocher, D.R., Krey, G., 2005. Three peroxisome proliferator-activatedreceptor isotypes from each of two species of marine fish. Endocrinology 146,3150–3162.
Lie, Ø., Lied, E., Lambertsen, G., 1986. Liver retention of fat and of fatty acids in cod(Gadus morhua) fed different oils. Aquaculture 59, 187–196.
Lovern, J.A., Sharp, J.G., 1933. CXCVIII. The diet of the halibut and intensity of feeding, inrelation to the vitamin S potency of liver oil. Biochem. J. 27, 1471–1472.
Lovern, J.A., Edisbury, J.R., Morton, R.A., 1933. CXCVII. Variations in vitamin A content iffish-liver oils, with particular reference to seasonal fluctuations in the potency ofhalibut-liver oil. Biochem. J. 27, 1461–1469.
Martins, D.A., Valente, L.M.P., Lall, S.P., 2007. Effects of dietary lipid level on growth andlipid utilization by juvenile Atlantic halibut (Hippoglossus hippoglossus, L.).Aquaculture 263, 150–158.
Mazorra, C., Bruce, M., Bell, J.G., Davie, A., Alorend, E., Jordan, N., Rees, J., Papanikos, N.,Porter, M., Bromage, N., 2003. Dietary lipid enhancement of brood stockreproductive performance and egg and larval quality in Atlantic halibut(Hippoglossus hippoglossus). Aquaculture 227, 21–33.
McFadzen, I.R.B., Coombs, H., Halliday, N.C., 1997. Histological indices of the nutritionalcondition of sardine, Sardina pilchardus (Walbaum) larvae off the north coast ofSpain. J. Exp. Mar. Biol. Ecol. 212, 239–258.
Metcalfe, L.D., Schimtz, A.A., Pelka, J.R., 1966. Rapid preparation of fatty acid esters fromlipids for gas chromatography. Anal. Chem. 38, 514–515.
Montero, D., Blazer, V.S., Socorro, J., Izqierdo, M.S., 1999. Dietary and culture influencesonmacrophage aggregate parameters in gilthead seabream Sparus aurata juveniles.Aquaculture 179, 523–534.
Morais, S., Bell, J.G., Robertson, D.A., William, J.R., Morris, P.C., 2001. Protein/lipid ratiosin extruded diets for Atlantic cod (Gadus morhua L.): effects on growth, feedutilisation, muscle composition and liver histology. Aquaculture 203, 101–119.

125S. Bolla et al. / Aquaculture 312 (2011) 117–125
Moyes, C.D., Suarez, R.K., Brown, G.S., Hochachka, P.W., 1991. Peroxisomal beta-oxidation: insights from comparative biochemistry. J. Exp. Zool. 260, 267–273.
Myers, M.S., Stehr, C.S., Olson, O.P., Johnston, L.L., McCain, B.B., Chan, S.L., Varanasi, U.,1994. Relationships between toxicopathic hepatic lesions and exposure to chemicalcontaminants in English sole (Pleuronectes vetulus), starry flounder (Platichthysstellatus), and white croaker (Genyonemus lineatus) from selected marine sites onthe Pacific Coast, USA. Environ. Health Perspect. 102, 200–215.
Ng, T.B., Idler, D.R., 1983. Yolk formation and differentiation in teleost fishes. In: Hoar,W.S., Randall, D.J., Donaldson, E.M. (Eds.), Fish Physiology, Vol. 9. Academic Press,New York, pp. 373–404.
Nordtvedt R., Mangor-Jensem A., Waagbø R., Norberg B., 2003. Ernæringsbetingeteggkvalitet hos kveite, Hippoglossus hippoglossus. Sluttrapport Nordisk MinisterrådNMR prosjektnr. 661045-90929. http://www.norden.org/no/publikasjoner/publi-kasjoner/2003-415/at_download/publicationfile.
Pascal, J.C., Ackman, R.G., 1976. Long chain monoethylenic alcohol and acid isomers inlipids of copepods and capelin. Chem. Phys. Lipids 16, 219–223.
Rochas, M.J., Rocha, E., Resende, A.D., Lobo-da-Cunha, A., 2003. Measurement ofperoxisomal enzyme activity in the liver of brown trout (Salmo trutta), usingspectrophotometric methods. BMC Biochem. 4, 2 http://www.biomedcentral.com/1471-209/4/2.
Segner, H., Witt, U., 1990. Weaning experiments with turbot (Scophthalmus maximus):electron microscopic study of liver. Mar. Biol. 105, 353–361.
Spisni, E., Tugnoli, M., Ponticelli, A., Mordenti, T., Tomasi, V., 1998. Hepatic steatosis inartificially fed marine teleosts. J. Fish Dis. 21, 177–184.
Tocher, D.R., Mourente, G., van der Eecken, A., Evjemo, J.O., Diaz, E., Bell, J.G., Guerden, I.,Lavens, P., Olsen, Y., 2002. Effects of dietary vitamin E on antioxidant defencemechanisms of juvenile turbot (Scophthalmus maximus L.), halibut (Hippoglossushippoglossus L.) and sea bream (Sparus aurata L.). Aquac. Nutr. 8, 195–207.
Tucker Jr., J.W., Lellis, W.A., Vermeer, G.K., 2001. The correlation between liver drymatter and liver lipid in cultured juvenile red drum fed various diets. North Am. J.Aqua. 63, 38–44.
Vethaak, A.D., Wester, P.W., 1996. Diseases in flounder Platichthys flesus in Dutchcoastal and estuarine waters, with particular reference to environmental stressfactors. II Liver histopathology. Dis. Aquat. Org. 26, 99–116.
Watanabe, T., 1982. Lipid nutrition in fish. Comp. Biochem. Physiol. 73B, 3–15.Weibel, E.R., Kistler, G.S., Scherle, W.F., 1969. Practical stereological methods for
morphometric cytology. J. Cell Biol. 30, 23–30.Wolke, R.E., 1992. Piscine macrophage aggregates: a review. Annu. Rev. Fish Dis.
91–108.
Copyright © 2022 FDOKUMEN




















