![Twee stijgbeugels, eene steng en beenderen opgegraven in Zwinderen, Klenckerheugte 15 nr. 1 (2015), 2-7. [Title in english: Two stirrups, a stick and bones excavated in Zwinderen]](https://static.fdokumen.com/doc/165x107/632313b4050768990e10246f/twee-stijgbeugels-eene-steng-en-beenderen-opgegraven-in-zwinderen-klenckerheugte.jpg)
Biochemical and physical correlates of DNA contamination in archaeological human bones and teeth...
9
Biochemical and physical correlates of DNA contamination in archaeological human bones and teeth excavated at Matera, Italy M. Thomas P. Gilbert a, * , Lars Rudbeck b , Eske Willerslev a,c , Anders J. Hansen c,d , Colin Smith e,1 , Kirsty E.H. Penkman e,2 , Kurt Prangenberg e,f , Christina M. Nielsen-Marsh e , Miranda E. Jans g , Paul Arthur h , Niels Lynnerup i , Gordon Turner-Walker j,3 , Martin Biddle k , Birthe Kjølbye-Biddle k , Matthew J. Collins e,g,2 a Henry Wellcome Ancient Biomolecules Centre, Department of Zoology, University of Oxford, South Parks Road, Oxford OX1 3PS, UK b Research Laboratory, Institute of Forensic Medicine, University of Copenhagen, Frederik V Vej 11, DK-2100 Copenhagen, Denmark c Department of Evolutionary Biology, Zoological Institute, University of Copenhagen, 5 Universitetsparken, DK-2100 Copenhagen Ø, Denmark d Department of Forensic Genetics, University of Copenhagen, Frederik V’s Vej 11, DK-2100 Copenhagen, Denmark e Fossil Fuels and Environmental Geochemistry, NRG, Drummond Building, University of Newcastle, Newcastle upon Tyne NE1 7RU, UK f Institut fu ¨r Geowissenschaften, Sigwartstrasse 10, 72076 Tu ¨bingen, Germany g Institute for Geo and Bioarchaeology, Vrije Universiteit Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam, Holland h Dipartimento di Beni Culturali, Via D. Birago, 64, University of Lecce, 73100 Lecce, Italy i Laboratory of Biological Anthropology, Institute of Forensic Medicine, University of Copenhagen, Frederik V Vej 11, DK-2100 Copenhagen, Denmark j Institutt for arkeologi og kulturhistorie, NTNU, Vitenskapsmuseet, 7491 Trondheim, Norway k Hertford College, University of Oxford, Oxford OX1 3BW, UK Received 28 October 2004; received in revised form 29 October 2004 Abstract The majority of ancient DNA studies on human specimens have utilised teeth and bone as a source of genetic material. In this study the levels of endogenous contamination (i.e. present within the sample prior to sampling for the DNA analysis) are assessed within human bone and teeth specimens sampled from the cemetery of Santa Lucia alle Malve, Matera, Italy. This site is of exceptional interest, because the samples have been assayed for 18 measures of biochemical and physical preservation, and it is the only one identified in a study of more than 107 animal and 154 human bones from 43 sites across Europe, where a significant number of human bones was well preserved. The findings demonstrate several important issues: (a) although teeth are more resilient * Corresponding author. Ecology and Evolutionary Biology, The University of Arizona, 1041 East Lowell Street, Tucson, AZ 85721, USA. Tel.: C1 520 621 4881; fax: C1 520 621 9190. E-mail address: [email protected] (M. Thomas P. Gilbert). 1 Present address: Department of Palaeobiology, Museo Nacional de Ciencias Naturales (CSIC), C/Jose´ Gutie´ rrez Abascal, 228006 Madrid, Spain. 2 Present address: BioArch, The King’s Manor, University of York, York YO1 7EP, UK. 3 Present address: Institute for Cultural Heritage Conservation, National Yunlin University of Science and Technology, 123 University Road Sec. 3, Touliu, Yunlin 640, Taiwan. 0305-4403/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.jas.2004.12.008 Journal of Archaeological Science 32 (2005) 785e793 http://www.elsevier.com/locate/jas
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Biochemical and physical correlates of DNA contamination in archaeological human bones and teeth...

Journal of Archaeological Science 32 (2005) 785e793
http://www.elsevier.com/locate/jas
Biochemical and physical correlates of DNAcontamination in archaeological human bones and
teeth excavated at Matera, Italy
M. Thomas P. Gilberta,*, Lars Rudbeckb, Eske Willersleva,c,Anders J. Hansenc,d, Colin Smithe,1, Kirsty E.H. Penkmane,2,
Kurt Prangenberge,f, Christina M. Nielsen-Marshe, Miranda E. Jansg,Paul Arthurh, Niels Lynnerupi, Gordon Turner-Walkerj,3, Martin Biddlek,
Birthe Kjølbye-Biddlek, Matthew J. Collinse,g,2
aHenry Wellcome Ancient Biomolecules Centre, Department of Zoology, University of Oxford,
South Parks Road, Oxford OX1 3PS, UKbResearch Laboratory, Institute of Forensic Medicine, University of Copenhagen, Frederik V Vej 11,
DK-2100 Copenhagen, DenmarkcDepartment of Evolutionary Biology, Zoological Institute, University of Copenhagen, 5 Universitetsparken,
DK-2100 Copenhagen Ø, DenmarkdDepartment of Forensic Genetics, University of Copenhagen, Frederik V’s Vej 11, DK-2100 Copenhagen, Denmark
eFossil Fuels and Environmental Geochemistry, NRG, Drummond Building, University of Newcastle,
Newcastle upon Tyne NE1 7RU, UKfInstitut fur Geowissenschaften, Sigwartstrasse 10, 72076 Tubingen, Germany
gInstitute for Geo and Bioarchaeology, Vrije Universiteit Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam, HollandhDipartimento di Beni Culturali, Via D. Birago, 64, University of Lecce, 73100 Lecce, Italy
iLaboratory of Biological Anthropology, Institute of Forensic Medicine, University of Copenhagen,
Frederik V Vej 11, DK-2100 Copenhagen, DenmarkjInstitutt for arkeologi og kulturhistorie, NTNU, Vitenskapsmuseet, 7491 Trondheim, Norway
kHertford College, University of Oxford, Oxford OX1 3BW, UK
Received 28 October 2004; received in revised form 29 October 2004
Abstract
The majority of ancient DNA studies on human specimens have utilised teeth and bone as a source of genetic material. In this
study the levels of endogenous contamination (i.e. present within the sample prior to sampling for the DNA analysis) are assessedwithin human bone and teeth specimens sampled from the cemetery of Santa Lucia alle Malve, Matera, Italy. This site is ofexceptional interest, because the samples have been assayed for 18 measures of biochemical and physical preservation, and it is the
only one identified in a study of more than 107 animal and 154 human bones from 43 sites across Europe, where a significantnumber of human bones was well preserved. The findings demonstrate several important issues: (a) although teeth are more resilient
* Corresponding author. Ecology and Evolutionary Biology, The University of Arizona, 1041 East Lowell Street, Tucson, AZ 85721, USA. Tel.:
C1 520 621 4881; fax: C1 520 621 9190.
E-mail address: [email protected] (M. Thomas P. Gilbert).1 Present address: Department of Palaeobiology, Museo Nacional de Ciencias Naturales (CSIC), C/Jose Gutierrez Abascal, 228006 Madrid, Spain.2 Present address: BioArch, The King’s Manor, University of York, York YO1 7EP, UK.3 Present address: Institute for Cultural Heritage Conservation, National Yunlin University of Science and Technology, 123 University Road Sec.
3, Touliu, Yunlin 640, Taiwan.
0305-4403/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jas.2004.12.008

786 M. Thomas P. Gilbert et al. / Journal of Archaeological Science 32 (2005) 785e793
to contamination than bone, both are readily contaminated (presumably through handling or washing), and (b) once contaminated
in this way, both are difficult (if not impossible) to decontaminate. Furthermore, although assessed on bone samples, several of thespecific biochemical and physical characteristics that describe overall sample preservation, levels of microbial attack and relatedincreases in sample porosity directly correlate with the presence of observable contamination in both bone and teeth samples from
individual samples. While we can only speculate on the cause of this relationship, we posit that they provide useful guides for theassessment of whether samples are likely to be contaminated or not.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Ancient DNA; Biopreservation; Bone; Contamination; Diagenesis; Human; Teeth
1. Introduction
Bones and teeth are normally the longest lastingphysical evidence of human or animal presence at anarchaeological site, and are also the most widely usedsources of samples for ancient DNA (aDNA) studies.Post hoc explanations of their suitability as a sourceof ancient DNA have identified retarded rates ofdecomposition, arising from adsorption of DNA tohydroxyapatite [30], low water content [22], ‘mummifi-cation’ of individual cells [4,5] and physical exclusion ofmicrobes and external contaminants [22]. Recently, anawareness of sample handling as a source of contami-nation has led researchers to investigate teeth as anaDNA source. One hypothetical benefit is protectionconferred by enamel [32]. Additionally, although histo-logical studies identify higher numbers of DNAcontaining cells per unit area of bone than teeth [12],several studies have reported better DNA yields in teeththan bone [29,32].
Richards et al. [36] have argued convincingly thatcontamination, not DNA preservation, is the greatestproblem facing the field, although the two are evidentlylinked e sparse, damaged endogenous DNA is less likelyto be amplified than modern contamination. Although itis known that teeth and bone may become contaminatedprior to aDNA extraction [36], current techniques usedto decontaminate specimens e the application of bleach,exposure to UV light, and grinding or shot-blasting ereflect a belief that contamination is concentrated in theouter surface of the material. Protocols designed to limitcontamination stress the prevention of contact betweensamples and previously amplified DNA (amplicons)[11,18]. Nevertheless, even when strict protocols arefollowed contaminants are frequently observed. Forexample, human DNA has been reported from cave bear[21], 500-year-old pig samples [36] and 109 out of 168relatively recent fox teeth [43]. More seriously, severalstudies report significant numbers of human remainscontaminated with multiple human sequences [19,27].Obviously, decontamination methods are not 100%efficient, and contamination remains a serious threat tothe validity of ancient DNA studies, particularly onhuman templates.
In this study we have investigated the presence andpersistence of contamination in teeth and femur samplescollected from human skeletons excavated at thecemetery of Santa Lucia alle Malve, Matera, Italy [8].Bone samples from the specimens have been assayed for18 measures of biochemical and physical preservation,allowing these parameters to be correlated with modernDNA contamination in both bones and teeth.
2. Materials and methods
Twenty-six teeth and eight femur samples were takenfrom 13 individuals excavated at the cemetery above thecave-church of Santa Lucia alle Malve, Matera, Italy [8](Table 1). The pH of the soil at the site ranges from 8.0to 8.3. Carbon-14 dating indicates that the samples dateto approximately the late 14th century (Clare Owen,Oxford RLAHA, pers. comm.). In a study thatinvestigated more than 107 animal and 154 humanbones from 41 sites across Europe (spanning fourclimatic regimes e Mediterranean, Continental, Mari-time (coastal) and Subarctic, and dating from 250 to5950 Y.B.P. [24]), this site was the only one in whicha significant number of human bones (7 of 14) were wellpreserved [23e25,38] (Table 1). Additionally, as part ofthis study the extent of the combined asparagines/aspartic acid (Asx) racemization was determined in onetooth from each individual using the extraction methodof Poinar et al. [35] and the analytical method ofKaufman and Manley [26]. Furthermore, all have beenstored together, and have undergone similar amounts ofhuman handling post-excavation.
2.1. Ancient DNA extraction and amplification
DNA was extracted from samples following strictancient DNA protocols in order to prevent samplecontamination with previously amplified DNA [15].Importantly, several different extractions (2e3) wereperformed per individual from distinct parts of the body(i.e. different teeth plus femur) to help identify both the‘endogenous’ DNA sequence and any contaminants.One extraction blank was used for every four samples to
Bulk
density
Skeletal
density
Total
Hg
Hg!
100 nm
Hg! 100
nme6 mm
HgO
6 mm
Asx
2.1 2.6 0.187 0.133 0.055 �0.001 0.06
2.1 2.6 0.187 0.133 0.055 �0.001 0.07
2 2.2 0.085 0.024 0.021 0.04 0.09
2 2.2 0.085 0.024 0.021 0.04 0.08
1.6 2 0.154 0.05 0.034 0.07 0.04
1.6 2 0.154 0.05 0.034 0.07 0.05
1.8 2.5 0.237 0.059 0.097 0.081 0.05
1.8 2.5 0.237 0.059 0.097 0.081 0.05
1.5 2.2 0.312 0.07 0.194 0.048 0.06
1.5 2.2 0.312 0.07 0.194 0.048 0.06
1.6 2.2 0.268 0.077 0.163 0.028 0.07
1.6 2.2 0.268 0.077 0.163 0.028 0.07
1.4 2.3 0.375 0.089 0.266 0.02 0.08
1.4 2.3 0.375 0.089 0.266 0.02 0.06
1.9 2.1 0.083 0.023 0.032 0.028 0.06
1.9 2.1 0.083 0.023 0.032 0.028 0.06
1.6 2.2 0.221 0.045 0.159 0.017 0.06
1.6 2.2 0.221 0.045 0.159 0.017 0.07
1.8 2.1 0.123 0.031 0.065 0.027 0.05
1.8 2.1 0.123 0.031 0.065 0.027 0.05
1.5 2.3 0.334 0.08 0.223 0.031 0.06
0.8 1.1 0.229 0.039 0.167 0.023 0.06
1.4 2.3 0.387 0.07 0.24 0.077 0.06
1.4 2.3 0.387 0.07 0.24 0.077 0.07
1.8 2.5 0.237 0.059 0.097 0.081 n/a
1.5 2.2 0.312 0.07 0.194 0.048 n/a
1.6 2.2 0.268 0.077 0.163 0.028 n/a
1.4 2.3 0.375 0.089 0.266 0.02 n/a
1.9 2.1 0.083 0.023 0.032 0.028 n/a
1.6 2.2 0.221 0.0449 0.1592 0.0171 n/a
1.8 2.1 0.123 0.031 0.065 0.027 n/a
1.4 2.3 0.387 0.07 0.24 0.077 n/a
0.01 0.20 0.07 0.82 !0.01 0.25 0.75
Asx which was directly assessed on teeth specimens). For
es from each individual where 0=minimum, 5=maximum
itudinal fungal attack; Budded, presence (1) or absence (0) of
red as the percentage of osteons with at least one radial crack;
, bone infra-red splitting factor, measuring phosphate oxygen
cluding all pores (density in a vacuum on 0); Skeletal Density,
100 nm, pore volume in the pore range of less than 100 nm as
6 mm, pore volume in the pore range of greater than 6 mm as
tion induced by hydrolysis (6 M HCl, 6 h at 110 �C [35] was
s used to investigate whether any correlation exists between
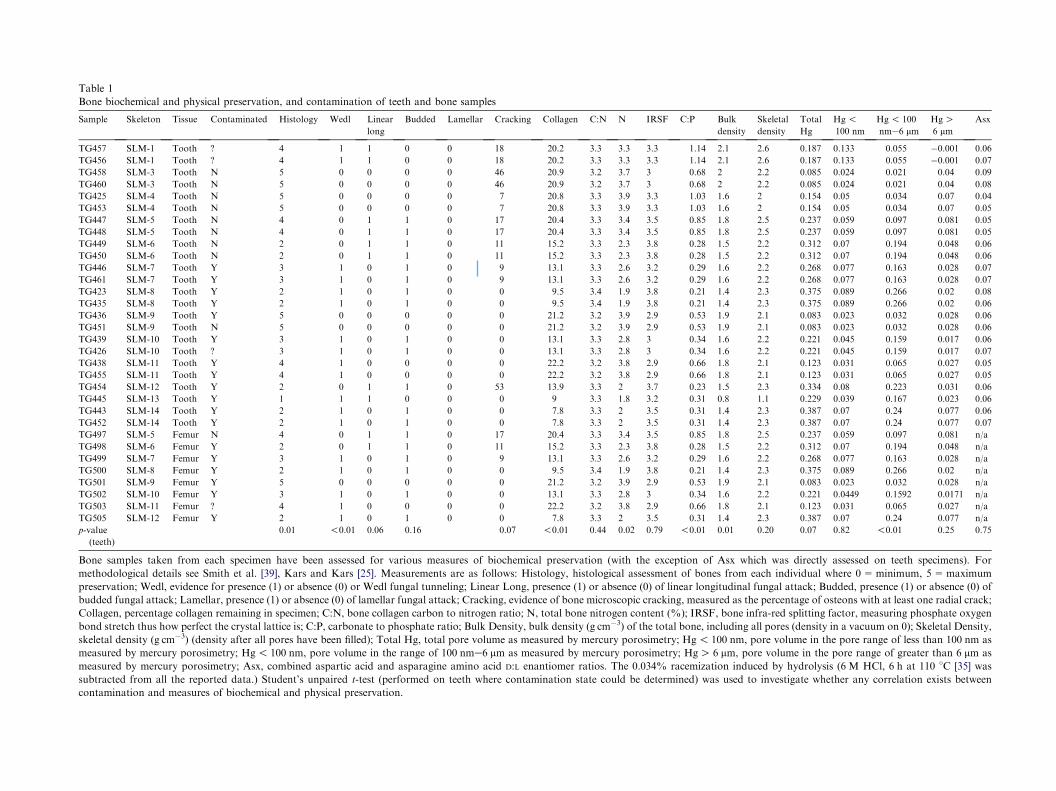
Table 1
Bone biochemical and physical preservation, and contamination of teeth and bone samples
Sample Skeleton Tissue Contaminated Histology Wedl Linear
long
Budded Lamellar Cracking Collagen C:N N IRSF C:P
TG457 SLM-1 Tooth ? 4 1 1 0 0 18 20.2 3.3 3.3 3.3 1.14
TG456 SLM-1 Tooth ? 4 1 1 0 0 18 20.2 3.3 3.3 3.3 1.14
TG458 SLM-3 Tooth N 5 0 0 0 0 46 20.9 3.2 3.7 3 0.68
TG460 SLM-3 Tooth N 5 0 0 0 0 46 20.9 3.2 3.7 3 0.68
TG425 SLM-4 Tooth N 5 0 0 0 0 7 20.8 3.3 3.9 3.3 1.03
TG453 SLM-4 Tooth N 5 0 0 0 0 7 20.8 3.3 3.9 3.3 1.03
TG447 SLM-5 Tooth N 4 0 1 1 0 17 20.4 3.3 3.4 3.5 0.85
TG448 SLM-5 Tooth N 4 0 1 1 0 17 20.4 3.3 3.4 3.5 0.85
TG449 SLM-6 Tooth N 2 0 1 1 0 11 15.2 3.3 2.3 3.8 0.28
TG450 SLM-6 Tooth N 2 0 1 1 0 11 15.2 3.3 2.3 3.8 0.28
TG446 SLM-7 Tooth Y 3 1 0 1 0 9 13.1 3.3 2.6 3.2 0.29
TG461 SLM-7 Tooth Y 3 1 0 1 0 9 13.1 3.3 2.6 3.2 0.29
TG423 SLM-8 Tooth Y 2 1 0 1 0 0 9.5 3.4 1.9 3.8 0.21
TG435 SLM-8 Tooth Y 2 1 0 1 0 0 9.5 3.4 1.9 3.8 0.21
TG436 SLM-9 Tooth Y 5 0 0 0 0 0 21.2 3.2 3.9 2.9 0.53
TG451 SLM-9 Tooth N 5 0 0 0 0 0 21.2 3.2 3.9 2.9 0.53
TG439 SLM-10 Tooth Y 3 1 0 1 0 0 13.1 3.3 2.8 3 0.34
TG426 SLM-10 Tooth ? 3 1 0 1 0 0 13.1 3.3 2.8 3 0.34
TG438 SLM-11 Tooth Y 4 1 0 0 0 0 22.2 3.2 3.8 2.9 0.66
TG455 SLM-11 Tooth Y 4 1 0 0 0 0 22.2 3.2 3.8 2.9 0.66
TG454 SLM-12 Tooth Y 2 0 1 1 0 53 13.9 3.3 2 3.7 0.23
TG445 SLM-13 Tooth Y 1 1 1 0 0 0 9 3.3 1.8 3.2 0.31
TG443 SLM-14 Tooth Y 2 1 0 1 0 0 7.8 3.3 2 3.5 0.31
TG452 SLM-14 Tooth Y 2 1 0 1 0 0 7.8 3.3 2 3.5 0.31
TG497 SLM-5 Femur N 4 0 1 1 0 17 20.4 3.3 3.4 3.5 0.85
TG498 SLM-6 Femur Y 2 0 1 1 0 11 15.2 3.3 2.3 3.8 0.28
TG499 SLM-7 Femur Y 3 1 0 1 0 9 13.1 3.3 2.6 3.2 0.29
TG500 SLM-8 Femur Y 2 1 0 1 0 0 9.5 3.4 1.9 3.8 0.21
TG501 SLM-9 Femur Y 5 0 0 0 0 0 21.2 3.2 3.9 2.9 0.53
TG502 SLM-10 Femur Y 3 1 0 1 0 0 13.1 3.3 2.8 3 0.34
TG503 SLM-11 Femur ? 4 1 0 0 0 0 22.2 3.2 3.8 2.9 0.66
TG505 SLM-12 Femur Y 2 1 0 1 0 0 7.8 3.3 2 3.5 0.31
p-value
(teeth)
0.01 !0.01 0.06 0.16 0.07 !0.01 0.44 0.02 0.79 !0.01
Bone samples taken from each specimen have been assessed for various measures of biochemical preservation (with the exception of
methodological details see Smith et al. [39], Kars and Kars [25]. Measurements are as follows: Histology, histological assessment of bon
preservation; Wedl, evidence for presence (1) or absence (0) or Wedl fungal tunneling; Linear Long, presence (1) or absence (0) of linear long
budded fungal attack; Lamellar, presence (1) or absence (0) of lamellar fungal attack; Cracking, evidence of bone microscopic cracking, measu
Collagen, percentage collagen remaining in specimen; C:N, bone collagen carbon to nitrogen ratio; N, total bone nitrogen content (%); IRSF
bond stretch thus how perfect the crystal lattice is; C:P, carbonate to phosphate ratio; Bulk Density, bulk density (g cm�3) of the total bone, in
skeletal density (g cm�3) (density after all pores have been filled); Total Hg, total pore volume as measured by mercury porosimetry; Hg!measured by mercury porosimetry; Hg! 100 nm, pore volume in the range of 100 nme6 mm as measured by mercury porosimetry; HgOmeasured by mercury porosimetry; Asx, combined aspartic acid and asparagine amino acid D:L enantiomer ratios. The 0.034% racemiza
subtracted from all the reported data.) Student’s unpaired t-test (performed on teeth where contamination state could be determined) wa
contamination and measures of biochemical and physical preservation.

788 M. Thomas P. Gilbert et al. / Journal of Archaeological Science 32 (2005) 785e793
monitor contaminants entering during the DNA extrac-tion. DNA extractions from bone used 0.2 g bonepowder, collected as in Barnes et al. [3]. Unlessotherwise stated, all DNA was extracted from teethfollowing Gilbert et al. [15]. All PCRs were performedon each sample at least twice, using the polymeraseenzyme Platinum Taq Hi-Fidelity (invitrogen), and in-corporating one PCR blank to every three samples.PCRs for human mitochondrial DNA (mtDNA) usedprimers L16209-H16356 [19] following Gilbert et al. [15].We have previously demonstrated that these primers areexceedingly sensitive to low levels of template DNA,thus are unlikely to generate false negative data [16].DNA extracts that did not yield PCR products werescreened for the presence of PCR inhibitors (a commonphenomena in aDNA studies [33]) through ‘spiking’PCRs containing amplifiable DNA with an equalvolume (1 ml) of the potential inhibitor, and monitoringany reduction in PCR success [15]. All amplified humanPCR products were cloned. Up to 12 colonies weresequenced per cloned PCR (325 total, average 10.2clones per PCR), thus providing between 16 and 35cloned sequences across all independent extracts of eachindividual skeleton (average 25 per skeleton). Forfurther details refer to supplemental information.
Endogenous and contaminant samples were identi-fied as described in the supplemental information. Inbrief, cloned PCR products were sequenced andcompared with the Cambridge Reference Sequence [2]to identify sample-specific motifs, both within the clonesfrom each extract, and between different extracts fromeach individual. Under an assumption that samplescontain authentic, uncontaminated DNA, it is expectedthat all cloned sequences will yield the same sequence(Fig. 1A). However, due to small amounts of post-mortem DNA damage (for example, hydrolytic de-amination of cytosine to uracil, providing isolated C/Tand G/A mutations, or hydrolytic deamination ofadenine to hypoxanthine, providing isolated A/G andT/C mutations), aDNA sequences often exhibit smallamounts of variation around a consensus sequence(Fig. 1B). If however the sample is uncontaminated, allsequences will contain shared motifs that identify themas originating from one original source of DNA(Fig. 1A and B). However, should a sample becontaminated with non-endogenous sources of DNA,in most situations sequences that do not share motifswill be observed among the clones (Fig. 1C).
This method has two potential weaknesses. Firstly, ifa sample contains no endogenous DNA, but has beencontaminated with one source of exogenous DNA(which possibly due to the time-lapse since contamina-tion may even contain some evidence of damage-drivenmiscoding lesions), the results will appear authentic.However, as all samples investigated in this study havebeen handled by multiple individuals, there is no
plausible reason why only one DNA source will berepresented among contaminant sequences. A moreproblematic issue arises when samples have beencontaminated by mtDNA sequences that are identical,or very similar (e.g. differing by 1e2 bp) to that of thesample. Based on the diversity observed among WesternEuropean mtDNA sequences (c.f. [37]), this is unlikelyto be an issue unless the sequence is exceedinglycommon, as in the case of basal haplogroup Hsequences (those that are identical to, or differ bya single step from, the Cambridge Reference Sequence(CRS) over the region of interest, and observed atfrequencies of up to 60% in Western Europe [41]). Insuch cases the lack of distinct motifs in the sequencemakes it exceedingly difficult to distinguish contaminantsequences from original, damaged sequences (Fig. 1D).Therefore, such samples were left out of the analysis toprevent misidentification of contaminated samples. Forfurther details on this method and why we believe it toenable us to differentiate between authentic andcontaminant DNA sequences, we refer readers to thearguments presented elsewhere [16] and to the in-formation presented in the supplemental information.
2.2. Assessment of bone preservation correlateswith contamination
Human bones suffer common patterns of alterationwith microscopic focal destruction resulting in areas ofdense mineralization surrounding pores with diametersat 600 nm and 1.2 mm microbial alteration (‘‘m’’porosity, 600 nme1.5 mm [24,42]). We believe that thisdiagenetic feature is a characteristic of the rapidputrefaction of interred corpses by blood borne gutbacteria as suggested by Bell et al. [5]. In order tosimplify inter-sample variation, the mid-shaft of thefemur (as a large and commonly preserved element) wasassessed to determine the extent of diagenetic alteration.At the Matera site, only half of the sampled individualshad the putrefaction alteration which characterized 74%of all human skeletons in the European study. Theremaining femurs were remarkably well preserved,partly it would appear because of secondary mineralprecipitation. Matera was the only site of 50 studiedwith a significant number of well-preserved humanbones [25]. We still remain ignorant of why Matera is sounusual in this respect, the burial in limestone cuts, andextreme seasonal variation in temperature and moisture,may both be implicated. The detailed record of bonepreservation of the Matera samples, the dipolar patternof preservation and the similar treatment of the bonessince excavation presents an ideal opportunity toinvestigate correlations between contamination andpreservation indices.
Unpublished data shows that bone is more proneto secondary contamination than dentine (M.T.P.G,

789M. Thomas P. Gilbert et al. / Journal of Archaeological Science 32 (2005) 785e793
1.A201 210 220 230 240 250 260caagcaagtacagcaatcaaccctcaactatcacacatcaactgcaactccaaagccacccctc
A ......................t................................t........B ......................t................................t........C ......................t................................t........D ......................t................................t........E ......................t................................t........F ......................t................................t........G ......................t................................t........H ......................t................................t........
1.B201 210 220 230 240 250 260caagcaagtacagcaatcaaccctcaactatcacacatcaactgcaactccaaagccacccctc
A ......................t...g............................t........B ......................t................................t........C ......................t................................t........D ..........t....g......t................................t........E ....a.................t................................t........F .......................................................t........G ......................t................................t........H ......................t................................t........
1.C201 210 220 230 240 250 260caagcaagtacagcaatcaaccctcaactatcacacatcaactgcaactccaaagccacccctc
A ......................t...........g....................t........B ......................t................................t........C ........c.......................................................D ......................t................................t......c.E ........c.......................................................F ......................t................................t........G ........c.......................................................H ........c.......................................................
1.D201 210 220 230 240 250 260caagcaagtacagcaatcaaccctcaactatcacacatcaactgcaactccaaagccacccctc
A .......................................................t........B ......................t.........................................C .......................................................t........D ................................................................E ................................................................F .......................................................t........G ................................................................H .....................................g..........................
Fig. 1. The identification of contaminated sequences from a cloned PCR product. (A) Cloned PCR products from uncontaminated, undamaged
DNA extracts show no sequence variation. (B) Uncontaminated, but damaged PCR products show some sequence variation, but also conserved
sequence motifs among the clones. (C) Cloned PCR products containing 1 old, damaged DNA source, plus a modern, contaminant source of DNA.
(D) In some situations it is not possible to differentiate whether variation among cloned sequences is due to contamination or post mortem damage.
For full details on figure refer to main text. For each decision applied to samples in this study refer to supplemental information.
unpublished data), as the enamel partially protects thedentine from allochthnaous DNA. We, therefore, choseto attempt to extract DNA from the dentine, in additionto extractions from the mid-shaft femur samples,reasoning that although we were unable to conduct thesame suite of analyses on the dentine, the femur datashould give an insight into the deterioration of the wholeskeleton.
3. Results
All extraction and PCR blanks were consistentlynegative throughout the study, indicating that the
results are unlikely to derive from contaminants in theextraction or PCR processes. Furthermore, the extent ofracemization of aspartic acid and alanine in the sampleswas below the threshold over which DNA is unlikely tosurvive (following [35]).
3.1. Preservation and contamination ofMatera teeth and bones
Nine Matera teeth samples contained only one DNAsequence, and appear to be uncontaminated, while 12teeth extracts contained multiple sequences and wereidentified as contaminated (Table 1). The majority of
790 M. Thomas P. Gilbert et al. / Journal of Archaeological Science 32 (2005) 785e793
contaminated samples appear to contain more than twosources of DNA, indicating either contamination onmultiple occasions, or that the samples have beenexposed to multiple sources of DNA in one go (forexample, through washing in dirty water). Additionallyin most cases, the multiple teeth and bone specimensfrom the same skeleton share at least some of thecontaminant sequences. A further three teeth could notbe accurately identified as contaminated, as it could notbe resolved whether observed sequence variation wasdue to damage-driven miscoding lesions or the presenceof multiple DNA sources. DNA could not be amplifiedfrom the remaining two teeth e one each from SLM-12and SLM-13 (data not shown). For full sequence detailssee supplemental information.
There are several clear patterns between DNAcontamination in teeth and the preservation statedetermined for bone samples from the same individuals.For p-values calculated on the comparisons of bio-chemical and physical measurements between contam-inated and uncontaminated samples (using Student’sunpaired t-test) refer to Table 1. In particular, alluncontaminated samples (except for teeth from skeletonSLM-6) lack evidence of organic degradation (Histology( p=0.01), collagen content ( p! 0.01), nitrogen con-tent ( p=0.02)), but have high mineral carbonate:phos-phate ratios, indicative of secondary carbonateprecipitation ( p! 0.01). Similarly, they all lack evi-dence of fungal attack (identified through a lack of Wedltunneling, p! 0.01). Most interesting, the presence ofhuman contaminant DNA in teeth correlates with theincreased porosity in the bones of the same individual( p! 0.01), the pores having a diameter that containsthe range of those indicative of microbial alteration (so-called ‘m’ porosity ranging from 600 nm to 1.5 mm [42]).
Despite the rigorous bleach and UV cleaning meth-ods, all Matera bone samples contained multiple DNAsequences, with the exception of samples Tg497, ex-tracted from skeletons SLM-5. Preservation data [38] onthis sample identifies it as exhibiting similar values forhistology, porosity, collagen content and mineralogy asmodern bone, with no evidence of microbial alteration(Table 1). Perhaps the most intriguing aspect of thissample is its very high carbonate:phosphate ratios in themineral, indicating the presence of secondary carbonate.Amino acid racemization (AAR) values tended to belower in uncontaminated samples with the exception ofskeleton SLM-03, although all values were tightly ranged(0.07e0.13). However even in this sample there was noevidence from D:L ratios of Ala, Glx or Ser of measur-able (peptidoglycan derived) microbial biomass (c.f. [34],data not shown). The bone from skeleton SLM-03 hadthe lowest porosity of any bone from Matera, so it ispossible that the elevated AAR value is due to the lowerrate of leaching of soluble (highly racemized gelatin), asspeculated by Collins et al. [10].
4. Discussion
The results of this study highlight a correlationbetween sample bone and tooth contamination, andoverall bone preservation and porosity e in particularthat characteristic of microbial attack [42]. It haspreviously been speculated [42] that putrefaction ofintact inhumed corpses is the dominant factor control-ling ‘m’ porosity linked to microbial attack. Conse-quently (butchered) animal bone is less likely to putrefy,thus likely to have lower ‘m’ porosity and more likely tohave potential for reliable aDNA recovery than buriedhuman remains. Furthermore the data indicate that aspreviously hypothesized (c.f. [13]), bone is moresusceptible to contamination than teeth.
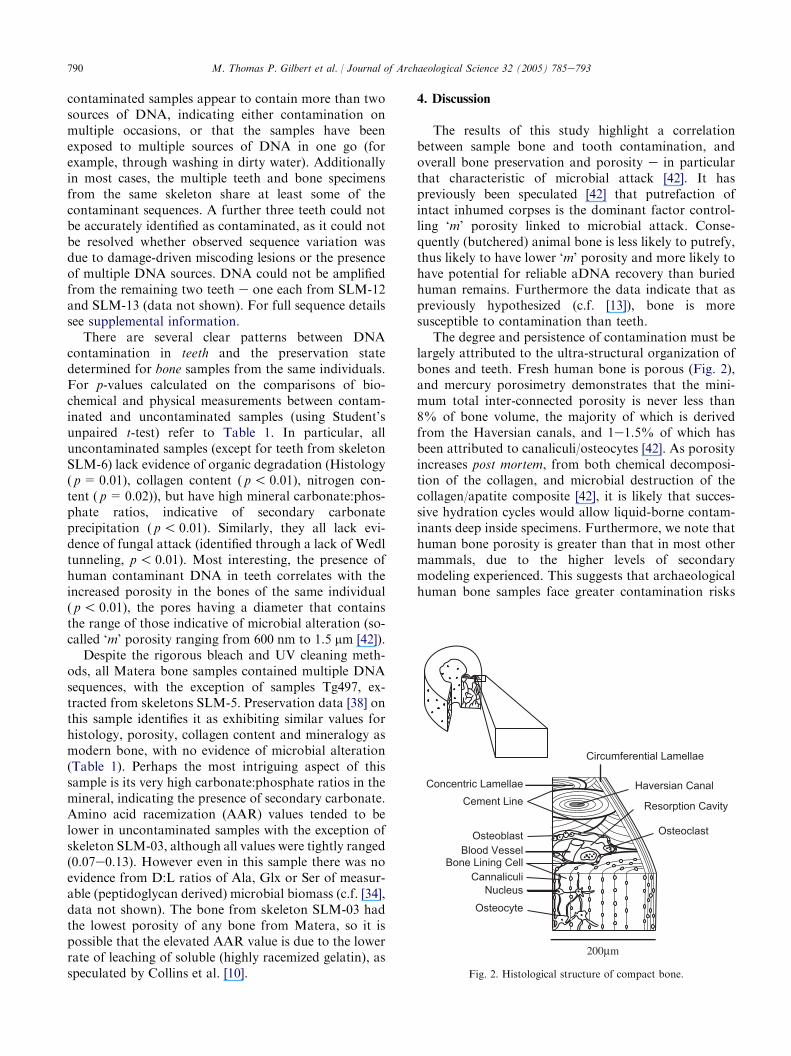
The degree and persistence of contamination must belargely attributed to the ultra-structural organization ofbones and teeth. Fresh human bone is porous (Fig. 2),and mercury porosimetry demonstrates that the mini-mum total inter-connected porosity is never less than8% of bone volume, the majority of which is derivedfrom the Haversian canals, and 1e1.5% of which hasbeen attributed to canaliculi/osteocytes [42]. As porosityincreases post mortem, from both chemical decomposi-tion of the collagen, and microbial destruction of thecollagen/apatite composite [42], it is likely that succes-sive hydration cycles would allow liquid-borne contam-inants deep inside specimens. Furthermore, we note thathuman bone porosity is greater than that in most othermammals, due to the higher levels of secondarymodeling experienced. This suggests that archaeologicalhuman bone samples face greater contamination risks
Haversian CanalCement Line
Circumferential Lamellae
Osteocyte
OsteoclastOsteoblast
CannaliculiNucleus
Bone Lining CellBlood Vessel
Resorption Cavity
Concentric Lamellae
Fig. 2. Histological structure of compact bone.

791M. Thomas P. Gilbert et al. / Journal of Archaeological Science 32 (2005) 785e793
(due to fluid uptake) than associated-animal remains,thus implies that the use of such remains as a ‘contam-ination test’ (e.g. [11]) may be inadequate. Similarproblems are also likely to affect teeth e although toothdentine is devoid of Haversian canals, and the enamelencasing tooth crowns is ideal for preventing permeationof contaminants [13], tooth roots are surprisinglyporous (Fig. 3). In particular, the pulp cavity is directlyconnected to the exterior of the root by numerous dentaltubules [12]. Therefore, once a tooth is washed or theroot is directly handled, the impervious nature of enamelis of no help in limiting contamination, which mayexplain the multiple sources of DNA found in mostteeth in this study.
In view of the correlation observed between boneporosity, and tooth and bone contamination in suchwell-preserved samples, it seems plausible that wide-spread contamination may be present in archaeologicalspecimens that were excavated in the past. For example,in many cases freshly excavated samples were cleaned indirty water that was not changed between samples. Inaddition, one tool of choice to aid cleansing was old andused human toothbrushes. Thus these would haveprovided an immediate source of human and bacterialDNA. However it is surprising that in our experiments,soaking samples in bleach does not degrade contami-nant DNA. One possibility is that bleach is preventedfrom fully penetrating samples by deposits (potentiallyderiving from sediment in water used to wash samples)that are laid down within the porous regions of thesamples as samples dry over time. Alternatively somedrying may seal pore interconnectivity through collaps-ing collagen bundles or drying fronts of other dissolvedspecies. Indirect evidence for this comes from onemeasure of preservation that inversely correlates withcontamination e ‘cracking’ e which Hedges [20] has
Enamel
Dental Tubules
Dentine
Root End Opening
Root
Crown
Pulp cavity
Supporting Ligaments
Jaw Bone
Cementum
Accessory Canal
Fig. 3. Histological structure of a human tooth. Dental tubules are
shown larger than actual size.
hypothesized results from bone swelling, due to mineraldeposition within Haversian canals.
If the above hypothesis is true, then it is possible thatthe disruption of internal blockages through techniqueswhich powder the samples prior to cleansing warrantfurther investigation. An alternative approach to con-sider is partial demineralization of the bone or teeth withEDTA prior to the bleaching. This may increase bothaccessibility to contaminant DNA through the action ofEDTA removing diagenetic calcium and iron salts, andthe action of bleach destroying collapsed regions ofcollagen. A final method that is also worth investigatingis the application of bleach to bone under a vacuum.
4.1. Prescreening samples for contamination
Why a correlation between bone preservation indicesand tooth contamination exists is a difficult question.We speculate that the fundamental biochemical andstructural similarities between teeth and bone [12] resultin similar patterns of degradation. However, regardlessof the explanation, the data clearly indicate thatcorrelations exist, and these provide an important aidto identifying suitable specimens for aDNA analysis.Such indices would save considerable time, whilelimiting destructive sampling, but may help criticallyassess previous ancient human DNA results. Overallthese will be of most use if they help provide ataphonomic context in which DNA preservation andcontamination can be understood.
Porosity, in particular that caused by microbialalteration of the bone, appears to be the most helpfulmeasure. Whilst this is not the only factor, techniquessuch as microscopy or mercury porosimetry which assaythis [42] will provide a useful guide to the contaminationload of a sample (other factors that will presumablydestroy DNA but not lead to microbial porosity includeaccelerated destruction of the organic phase as observedat Apigliano, Italy [38]). An alternative technique nowroutinely used on ancient samples prior to DNAanalysis that may also be of use is amino acidracemization analyses ([35], though see [10]) thoughwhether a significant correlation exists between racemi-zation and porosity requires further investigation.However, given the presence of microbial destructioneven in bones with apparently exceptional preservationfrom Matera, perhaps the best approach is to select forbones which have not derived from interred corpses. Wenote in passing that because animals are rarely interredas intact corpses they display much less evidence ofporosity attributed to putrefaction [24]. In addition toa potential associated impact on that may reduce theoverall rate of DNA degradation, this may explain whythey are proving a more promising substrate for aDNAstudy than human material.

792 M. Thomas P. Gilbert et al. / Journal of Archaeological Science 32 (2005) 785e793
5. Conclusions
The results of this study highlight serious inadequa-cies in techniques used to decontaminate archaeologicalsamples (in particular those that rely on bone) fromcontaminant DNA, and have implications for anyaDNA study where contamination is a risk. Importantly,this also includes studies of pathogen DNA in ancientsamples, which may be at risk from DNA originatingfrom environmental organisms [14], as well as studies onanimal bones that have been treated with preservativesand glues containing animal DNA [31]. Furthermore, theresults suggest that, if such contamination persists within‘cleaned’ samples, even the adoption of authenticitycriteria (e.g. [11]) such as independent replication cannotguarantee the retrieval of authentic DNA sequences.
The simplest remedy for human DNA contaminationis the controlled excavation of human samples, prevent-ing direct handling and washing of samples that are tobe used for aDNA analyses. Unfortunately as mostsamples cannot be freshly excavated, and many impor-tant specimens were collected in the past, the majority islikely to be already contaminated. Thus further studiesare required to investigate the possibility of newdecontamination techniques on such specimens. Inaddition other sources of aDNA that may be moreresilient to contamination such as hair need to beinvestigated [16].
The findings of this study also suggest that a criticalreader must consider three key points before gaugingthe reliability of results from previous human aDNAstudies. Firstly, the preservation of the samples. Forexample, Clisson et al. [9] report human DNA retrievalfrom a O2000 year old specimen from Kazakhstan. Theexcellent preservation reported suggests that contami-nation is not an issue. Secondly, sequence authenticity.For example, although not well preserved, the distinctnature of Neanderthal sequences helps suggest authen-ticity of results [28]. Thirdly, and most importantly,knowledge of the history of the sample handling prior tothe analysis is critical. For example, in the importantearly Australian ‘Mungo man’ study [1], although theresearchers took full anti-contamination precautions, itwas not mentioned that the sample had been excavatedin the 1970s and handled numerous times since (thuslikely to be contaminated), contained ‘negligible organicpreservation’ [17], and is considered too fragmentary tosex reliably [7,40] e in short, of very poor preservation.Without such information, it is very difficult to commentobjectively on the reliability of results.
Acknowledgements
The impetus behind this work was a series of in-comprehensible aDNA results [15] on human bone from
Repton, England [6]. We thank Dott.ssa Maria LuisaNava (Archaeological Superintendent of Basilicata)permission to sample, and Dott.sse Brunella Bruno andErminia Lapadula for assistance in sampling. M.T.P.G.thanks the Wellcome Trust for funding his research(061610/Z/00/Z). E.W. and A.J.H. thank Villum Rass-mussen Fonden for funding their research. Diageneticparameters of the bone samples were analyzed as part ofthe ‘Degradation of Bone as an Indicator in the De-terioration of the European Archaeological Property’project funded by the EU (ENV4-CT98-0712).
Appendix A. Supplementary information
Supplementary information for this manuscript canbe downloaded at doi: 10.1016/j.jas.2004.12.008.
References
[1] G. Adcock, E. Dennis, S. Easteal, G. Huttley, L. Jermelin,
W. Peacock, A. Thorne, Mitochondrial DNA sequences in
ancient Australians: implications for modern human origins,
Proc. Natl. Acad. Sci. USA 98 (2001) 537e542.
[2] S. Anderson, A. Bankier, B. Arrell, M. de Bruijn, A. Coulson,
J. Drouin, I. Eperon, D. Nierlich, B. Roe, F. Sanger, P. Schreier,
A. Smith, R. Staden, I. Young, Sequence and organisation of the
human mitochondrial genome, Nature 290 (1981) 457e465.
[3] I. Barnes, P. Matheus, B. Shapiro, D. Jensen, A. Cooper,
Dynamics of mammal population extinctions in Eastern Beringia
during the last glaciation, Science 295 (2002) 2267e2270.
[4] L.S. Bell, G. Cox, J. Sealy, Determining isotopic life history
trajectories using bone density fractionation and stable isotope
measurements: a new approach, Am. J. Phys. Anthropol. 116
(2001) 66e79.
[5] L.S. Bell, M.F. Skinner, S. Jones, The speed of post mortem
change to the human skeleton and its taphonomic significance,
Forensic Sci. Int. 82 (1996) 129e140.
[6] M. Biddle, B. Kjølbye-Biddle, Repton and the ‘‘great heathen
army’’, 873e4, in: J. Graham-Campbell (Ed.), Vikings and the
Danelaw: Select Papers from the Proceedings of the Thirteenth
Viking Congress, Oxbow Books, Oxford, 2001, pp. 45e60.
[7] P. Brown, Australian pleistocene variation and the sex of Lake
Mungo 3, J. Hum. Evol. 38 (2000) 743e749.
[8] Excavations atMatera were directed by Paul Arthur. See B. Bruno,
Archeologia nei Sassi diMatera, in: S. Patitucci Uggeri (Ed.), Scavi
Medievali in Italia 1996e1999, Herder, Rome, 2001, pp. 137e148.
[9] I. Clisson, C. Keyser, H.P. Francfort, E. Crubezy, Z. Samashev,
B. Ludes, Genetic analysis of human remains from a double
inhumation in a frozen kurgan in Kazakhstan (Berel site, Early
3rd Century BC), Int. J. Legal Med. 116 (2002) 304e308.
[10] M.J. Collins, E.R. Waite, A.C.T. van Duin, Predicting protein
decomposition: the case of aspartic-acid racemization kinetics,
Philos. Trans. R. Soc. Lond. B 354 (1999) 51e64.
[11] A. Cooper, H. Poinar, Ancient DNA: do it right or not at all,
Science 289 (2000) 1139.
[12] J. Currey, Bones: structure and mechanics, Princeton University
Press, Princeton, 2002.
[13] M. Drancourt, G. Aboudharam, M. Signoli, O. Dutour,
D. Raoult, Detection of 400-year-old Yersinia pestis DNA in

793M. Thomas P. Gilbert et al. / Journal of Archaeological Science 32 (2005) 785e793
human dental pulp: an approach to the diagnosis of ancient
septicemia, Proc. Natl. Acad. Sci. USA 95 (1998) 12637e12640.
[14] M.T.P. Gilbert, J. Cuccui, W. White, N. Lynnerup, R.W. Titball,
A. Cooper, M.B. Prentice, Absence of Y. pestis-specific DNA in
human teeth from five European excavations of putative plague
victims, Microbiology 150 (2004) 341e354.
[15] M.T.P. Gilbert, E. Willerslev, A.J. Hansen, L. Rudbeck,
I. Barnes, N. Lynnerup, A. Cooper, Distribution patterns of post
mortem damage in human mitochondrial DNA, Am. J. Hum.
Genet. 72 (2003) 32e47.
[16] M.T.P. Gilbert, A.S. Wilson, M. Bunce, A.J. Hansen,
E. Willerslev, B. Shapiro, T.F.G. Higham, M.P. Richards,
T.C. O’Connell, D.J. Tobin, R.C. Janaway, A. Cooper, Ancient
mitochondrial DNA from hair, Curr. Biol. 14 (2004) R463e
R464.
[17] R. Gillespie, R.G. Roberts, On the reliability of age estimates for
human remains at LakeMungo, J. Hum. Evol. 38 (2000) 727e732.
[18] O. Handt, M. Hoss, M. Krings, S. Paabo, Ancient DNA:
methodological challenges, Experientia 50 (1994) 524e529.[19] O. Handt, M. Krings, R. Ward, S. Paabo, The retrieval of ancient
human DNA sequences, Am. J. Hum. Genet. 59 (1996) 368e376.
[20] R.E.M. Hedges, Bone diagenesis: an overview of processes,
Archaeometry 44 (2002) 319e328.
[21] M. Hofreiter, D. Serre, H.N. Poinar, M. Kuch, S. Paabo, Ancient
DNA, Nat. Rev. Genet. 2 (2001) 353e358.
[22] S. Hummel, B. Hermann, General aspects of sample preparation,
in: B. Hermann, S. Hummel (Eds.), Ancient DNA, Springer
Verlag, New York, 1994, pp. 59e68.
[23] M.M.E. Jans, H. Kars, C.M. Nielsen-Marsh, C.I. Smith,
A.G. Nord, P. Arthur, N. Earl, In situ preservation of
archaeological bone: a histological study within a multidisciplin-
ary approach, Archaeometry 44 (2002) 343e352.
[24] M.M.E. Jans, C.M. Nielsen-Marsh, C.I. Smith, M.J. Collins,
H. Kars, Characterisation of microbial attack on archaeological
bone, J. Archeol. Sci. 31 (2004) 87e95.
[25] E.A.K. Kars, H. Kars, The degradation of bone as an indicator
for the deterioration of the European archaeological property,
Final Report, Rijksdienst voor het Oudheidkundig Bodemonder-
zoek, Amersfoort, 2003.
[26] D.S. Kaufman, W.F. Manley, A new procedure for determining
DL amino acid ratios in fossils using reverse phase liquid
chromatography, Quatern. Sci. Rev. 17 (1998) 987e1000.[27] C.J. Kolman, N. Tuross, Ancient DNA analysis of human
populations, Am. J. Phys. Anthropol. 111 (2000) 5e23.
[28] M. Krings, A. Stone, R. Schmitz, H. Krainitzki, M. Stoneking,
S. Paabo, Neanderthal DNA sequences and the origin of modern
humans, Cell 90 (1997) 19e30.
[29] G. Kurosaki, T. Matsushita, S. Ueda, Individual DNA identifi-
cation from ancient human remains, Am. J. Hum. Genet. 53
(1993) 638e643.
[30] T. Lindahl, Instability and decay of the primary structure of
DNA, Nature 362 (1993) 709e715.
[31] G.J. Nicholson, J. Tomiuk, A. Czarnetzki, L. Bachmann,
C. Pusch, Detection of bone glue treatment as a major source
of contamination in ancient DNA analyses, Am. J. Phys.
Anthropol. 118 (2002) 117e120.
[32] H. Oota, N. Saitou, T. Matsushita, S. Ueda, A genetic study of
2000 year old human remains from Japan using mitochondrial
DNA sequences, Am. J. Phys. Anthropol. 98 (1995) 133e145.
[33] S. Paabo, R. Higuchi, A. Wilson, Ancient DNA and the
polymerase chain reaction, J. Biol. Chem. 264 (1989) 9709e9712.
[34] A.G.U. Pedersen, T.R. Thomsen, B.A. Lomstein, N.O.G.
Jorgensen, Bacterial influence on amino acid enantiomerization in
a coastal marine sediment, Limnol. Oceanogr. 46 (2001) 1358e1369.
[35] H. Poinar, M. Hoss, J. Bada, S. Paabo, Amino acid racemization
and the preservation of ancient DNA, Science 272 (1996) 864e866.
[36] M. Richards, B. Sykes, R. Hedges, Authenticating DNA
extracted from ancient skeletal remains, J. Archeol. Sci. 22
(1995) 291e299.[37] M. Richards, V. Macaulay, E. Hickey, E. Vega, B. Sykes,
V. Guida, C. Rengo, D. Sellitto, F. Cruciani, T. Kivisild,
R. Villems, M. Thomas, S. Rychkov, O. Rychkov, Y. Rychkov,
M. Golge, D. Dimitrov, E. Hill, D. Bradley, V. Romano, F. Calı,
G. Vona, A. Demaine, S. Papiha, C. Triantaphyllidis,
G. Stefanescu, J. Hatina, M. Belledi, A. Di Rienzo,
A. Novelletto, A. Oppenheim, S. Nørby, S. Santachiara-Bener-
ecetti, R. Scozzari, A. Torroni, H.-J. Bandelt, Tracing European
founder lineages in the Near Eastern mtDNA pool, Am. J. Hum.
Genet. 67 (2000) 1251e1276.
[38] Smith, Modelling diagenesis in archaeological bone, PhD thesis,
University of Newcastle, 2002.
[39] C.I. Smith, C.M. Nielsen-Marsh, M.M.E. Jans, P. Arthur,
A.G. Nord, M.J. Collins, The strange case of Apligliano:early
‘fossilization’ of medieval bone in southern Italy, Archaeometry
44 (2002) 405e416.
[40] A. Thorne, D. Curnoe, Sex and significance of Lake Mungo 3:
reply to Brown ‘‘Australian Pleistocene variation and the sex of
Lake Mungo 3’’, J. Hum. Evol. 39 (2000) 587e600.[41] A. Torroni, H.-J. Bandelt, L. D’Urbano, P. Lahermo, P. Moral,
D. Sellitto, C. Rengo, P. Forster, M.-L. Savontaus, B. Bonne-
Tamir, R. Scozzari, MtDNA analysis reveals a major late
paleolithic population expansion from southwestern to north-
eastern Europe, Am. J. Hum. Genet. 62 (1998) 1137e1152.
[42] G. Turner-Walker, C.M. Nielsen-Marsh, U. Syversen, H. Kars,
M.J. Collins, Sub-micron spongiform porosity is the major ultra-
structural alteration occurring in archaeological bone, Int. J.
Osteoarcheol. 12 (2002) 407e414.
[43] P. Wandeler, S. Smith, P.A. Morin, R.A. Pettifor, S.M. Funk,
Patterns of nuclear DNA degeneration over time e a case study in
historic teeth samples, Mol. Ecol. 12 (2003) 1087e1093.




















