
Bahig El-Deeb, Hesham Elhariry, Abdulla Altalhi, Sabry Hassan and Youssuf Gherbawy (2013). Isolation...
16
This article was downloaded by: [Professor Bahig El-Deeb] On: 01 August 2013, At: 17:25 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Food Biotechnology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/lfbt20 Isolation and Characterization of Endophytic Bacilli Bacterium from Maize Grains Able to Detoxify Aflatoxin B1 Bahig El-Deeb a b , Abdulla Altalhi a , Ghada Khiralla a c , Sabry Hassan a d & Youssuf Gherbawy a e a Department of Biology, Faculty of Science , Taif University , Taif , Saudi Arabia b Botany Department, Faculty of Science , Sohag University , Sohag , Egypt c National Organization for Drug Control and Research , Giza , Egypt d Microbiology Department, Faculty of Veterinary Medicine , South Valley University , Qena , Egypt e Botany Department, Faculty of Science , South Valley University , Qena , Egypt To cite this article: Bahig El-Deeb , Abdulla Altalhi , Ghada Khiralla , Sabry Hassan & Youssuf Gherbawy (2013) Isolation and Characterization of Endophytic Bacilli Bacterium from Maize Grains Able to Detoxify Aflatoxin B1, Food Biotechnology, 27:3, 199-212, DOI: 10.1080/08905436.2013.811083 To link to this article: http://dx.doi.org/10.1080/08905436.2013.811083 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Bahig El-Deeb, Hesham Elhariry, Abdulla Altalhi, Sabry Hassan and Youssuf Gherbawy (2013). Isolation...

This article was downloaded by: [Professor Bahig El-Deeb]On: 01 August 2013, At: 17:25Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Food BiotechnologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/lfbt20
Isolation and Characterization ofEndophytic Bacilli Bacterium from MaizeGrains Able to Detoxify Aflatoxin B1Bahig El-Deeb a b , Abdulla Altalhi a , Ghada Khiralla a c , SabryHassan a d & Youssuf Gherbawy a ea Department of Biology, Faculty of Science , Taif University , Taif ,Saudi Arabiab Botany Department, Faculty of Science , Sohag University , Sohag ,Egyptc National Organization for Drug Control and Research , Giza , Egyptd Microbiology Department, Faculty of Veterinary Medicine , SouthValley University , Qena , Egypte Botany Department, Faculty of Science , South Valley University ,Qena , Egypt
To cite this article: Bahig El-Deeb , Abdulla Altalhi , Ghada Khiralla , Sabry Hassan &Youssuf Gherbawy (2013) Isolation and Characterization of Endophytic Bacilli Bacteriumfrom Maize Grains Able to Detoxify Aflatoxin B1, Food Biotechnology, 27:3, 199-212, DOI:10.1080/08905436.2013.811083
To link to this article: http://dx.doi.org/10.1080/08905436.2013.811083
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,

systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

Food Biotechnology, 27:199–212, 2013Copyright © Taylor & Francis Group, LLCISSN: 0890-5436 print / 1532-4249 onlineDOI: 10.1080/08905436.2013.811083
Isolation and Characterizationof Endophytic Bacilli Bacteriumfrom Maize Grains Able toDetoxify Aflatoxin B1
Bahig El-Deeb1,2, Abdulla Altalhi1, Ghada Khiralla1,3, SabryHassan1,4, and Youssuf Gherbawy1,5
1Department of Biology, Faculty of Science, Taif University, Taif, Saudi Arabia2Botany Department, Faculty of Science, Sohag University, Sohag, Egypt3National Organization for Drug Control and Research, Giza, Egypt4Microbiology Department, Faculty of Veterinary Medicine, South Valley University,Qena, Egypt5Botany Department, Faculty of Science, South Valley University, Qena, Egypt
Aflatoxins are commonly found in cereals worldwide and bring significant threats to thefood industry and animal food production. Among a collection of aflatoxin-degradativeendophytic bacteria isolated from grains of corn plant, the strain TUBF1 was selectedbased on its high-ability to utilize aflatoxin B1 (AFB1) (10 µg/mL) as the sole carbonsource. The morphological, physiological, and phylogenetic studies indicated that strainTUBF1 belonged to the Bacillus sp. group. HPLC was used to determine the reduc-tion in AFB1 concentrations. Bacillus sp. TUBF1 had the strongest ability to detoxifytoxin, where the degradation percentages of AFB1 was 81.5% and 100% after 48 and72 h, respectively. The degradation of AFB1 was mainly in the culture supernatantof TUBF1 rather than its cells. About 90% of AFB1 was degraded within the first12 h and reduced to the undetectable level after 24 h. The supernatant was charac-terized by considerable activity at wide range of temperatures (10–40◦C) with optimalactivity at 32◦C and pH 6.5–7.5. Biosafety assessment test indicated that the crudeenzymes had the high ability to detoxify AFB1. In conclusion, this is the first report onAFB1 detoxification using an endophytic Bacillus isolated from grains.
Key Words: endophytic bacteria; Bacillus sp.; aflatoxins B1; detoxification
INTRODUCTION
Aflatoxin B1 (AFB1) is one of the mycotoxins produced by some strains ofAspergillus flavus, Aspergillus nomius and Aspergillus parasiticus (Deiner
Address correspondence to Bahig El-Deeb, Department of Biology, Faculty of Science,Taif University, Taif, 888, Saudi Arabia; E-mail: [email protected]
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

200 B. El-Deeb et al.
et al., 1987; Kurtzman et al., 1987; Cotty and Bhatnagar, 1994). This toxinhas been reported to be carcinogenic, teratogenic, and hepatotoxic (Eatonand Groopman, 1994; Guengerich et al., 1996). It poses a serious threatto both livestock productivity and human health and leads to economiclosses each year. Controlling AFB1 in food and feed has been a challengesince the toxins are markedly stable under different environmental condi-tions. Different physical and chemical methods have been developed andtested for removing AFB1. However, these methods have many restrictions,such as nutritional loss, sensory quality reduction, and high cost of equip-ment (Kabak et al., 2006). Therefore, biological approaches, such as theuse of microorganisms and enzymes, to convert the toxins into non or lesstoxic compounds, have the potential to serve as an alternative for aflatoxinsremoval (Altalhi and El-Deeb, 2009). Previous reports indicated that somefungi such as Aspergillus niger, A. parasiticus, Trichoderma viride, Mucorambiguus have been shown significant AFB1 degradation abilities (Mann andRehm, 1976; Tsubouchi et al., 1983; Huynh et al., 1984; Line et al., 1994).However, some of these fungi with degradation potential may also produceAFB1 under varying conditions (Huynh et al., 1984). Removal of aflatoxinsby bacteria has been given much attention in recent years. Several bacte-rial isolates were known to effectively degrade AFB1 such as Flavobacteriumaurantiacum (Ciegler et al., 1966; Line and Brackett, 1995; Smiley andDraughon, 2000; Teniola et al., 2005; Alberts et al., 2006), Mycobacteriumfluoranthenivorans (Hormisch et al., 2004; Teniola et al., 2005), Rhodococcuserythropolis (Teniola et al., 2005; Alberts et al., 2006), Bacillus licheni-formis (Petchkongkaew et al., 2008), and Stenotrophomonas maltophilia (Guanet al., 2008). Lillehoj et al. (1971) have also demonstrated the potential ofFl. aurantiacum to remove AFM1 from naturally contaminated milk. Thedetoxification by specific enzymes avoids the drawback of using the microor-ganism, which may, in addition to its degradation activity, change flavor orimpair the nutritional value and acceptability of the product (Shapira, 2004).A novel aflatoxin degradation enzyme had been isolated and purified fromPleurotus ostreatus by Motomura et al. (2003). AFB1 was treated with culturesupernatants from 19 mushroom strains. The supernatant from P. ostrea-tus showed aflatoxin-degradation activity, whereas other strains had no oronly weak activity. Zhao et al. (2010) demonstrated that aflatoxin degradationenzyme from Myxococcus fulvus can effectively remove aflatoxin B1, G1, andM1 in solution.
This research was conducted to demonstrate the ability of Bacillus sp.TUBF1, isolated from contaminated maize grains to degrade AFB1 in liquidcultures. Degradation of AFB1 using cell-free supernatant as a source of crudeenzymes was investigated to determine the optimal temperature and pH of theAFB1 degradative enzyme(s).
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

Isolation of Endophytic Bacilli Bacterium 201
MATERIAL AND METHOD
Chemicals and Seed SourcesAflatoxin B1 (AFB1) was purchased from Sigma Chemical Company
(Sigma, St Louis, Mo., USA). All other chemicals were at least of analytical-reagent. AFB1 was dissolved in Dimethyl sulfoxide (DMSO; 1 mg/mL) andused as a standard stock solution. Corn seeds were purchased from markets inTaif, Saudi Arabia. Fresh and stored samples from different market locationswere obtained.
Enrichment and Degradation Studies
Seed Surface Sterilization
Seed surface sterilization for endophytic bacterial isolation was carried outas described by Dietmar et al. (2008), with slight modifications. To soften seedand revitalize endophytic populations, seeds were soaked in autoclaved dis-tilled water for 48 h, drained, and washed in 0.1% Triton X-100 detergent for3 min with shaking. Then the detergent was drained and the seeds washedwith 3% sodium hypochlorite for 3 min. The seeds were drained and rinsedwith sterilized distilled water before being washed for 10 min in 95% ethanol.After that, seeds were drained and rinsed three times with sterilized distilledwater. To check for surface sterility, five seeds per treatment were momentarilyplaced on nutrient agar plates and incubated for 10 d at 25◦C.
To extract bacteria from surface sterilized samples, 15 seeds were groundin an autoclaved mortar. One ml of 50 mM Na2HPO4 buffer was added per 1 gdry weight of ground seeds. This mixture was used for further work.
Enrichment Culture
Enrichment culture method was used for isolation of endophytic bacteriafrom corn seeds as described by Kang et al. (2012), with some modification.Seed grounded mixtures were allowed to settle for 15 min. One mL of eachsupernatant was used to inoculate 100 mL of Minimal salt medium (MSMmedium) composed of 0.1% NH4NO3, 0.025% K2HPO4, 0.025% MgS04.7H2O,0.0001% FeSO4.7H2O and 0.5% yeast extract (Difco Laboratories, Detroit,Mich., USA). This medium was supplemented with 10 µg AFB1/mL. TheAFB1 solution that suspended in DMSO was filtered through a Millex®
HV PVDF 0.45µm, Millipore, and added to the media after sterilization.Cultivation was carried out on a rotary shaker at 30◦C for 7 d and at agita-tion speed of 150 rpm. After one week of incubation, a subenrichment culturewas made with a 1% inoculum of the previous enrichment culture and trans-ferred onto MSM medium containing AFB1 as described above. Cultures were
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

202 B. El-Deeb et al.
fully grown at the time of each transfer. After four weeks, the final enrich-ment culture was spread onto MSM agar plates containing 10 µg/mL AFB1asa sole source of carbon and energy and incubated at 30oC. The bacteria able toform colonies on plates were regarded as AFB1-assimilating bacteria and coulduse AFB1 as a sole carbon and energy. Isolates were maintained by monthlytransfers onto slants containing AFB1 and, after growth, stored at 4oC. Whendesired, cells from the slants were inoculated into 10 mL of Luria-Bertani broth(LB) (Sambrook et al., 1989) in 50-mL Erlenmeyer flasks, grown aerobically,and used for characterization and growth studies.
Genotypic Characterization of the AFB1 Degrading StrainStandard tests were performed for identification of the studied strain
in accordance with Bergey’s Manual of Determinative Bacteriology (Holtet al., 1994). Cell form and size, gram staining, spore formation, motil-ity, colony pigmentation, and production of UV-fluorescent pigments weretested. The strain was also identified by determination of 16S rRNA genesequences. The genomic DNA of studied bacteria was extracted (Govindarajanet al., 2007), and 16S rDNA was amplified in polymerase chain reac-tion (PCR) using the genomic DNA as template and bacterial univer-sal primers, 27 F (5′-GAGTTTGATCACTGGCTCAG-3′) and 1492 R (5′-TACGGCTACCTTGTTACGACTT-3′) (Byers et al., 1998). Briefly, a 25-µL reac-tion mixture contained 1.25 U RedTaq polymerase (Sigma Chemical Co.),0.2 mM dNTPs, 2.5 mM MgCl2 (Sigma), 10 pmol of each primer, 2.5 µL of 10×reaction buffer (Sigma), and 1 µg of template DNA. Aliquots of PCR reactionproducts were electrophoresed in 1% agarose containing 10 µg·ml—1 ethidiumbromide. The sequence of the strain TUBF1 was blasted against all nucleotidesequences in the European Nucleotide Archive (ENA). TUBF1 and other DNAsequences of genebank were aligned with CLUSTAL Software.
Growth Kinetics of Bacillus sp. and Degradation of AFB1 in LiquidCulturesBacterial strain TUBF1 was cultivated in 5 mL of LB medium at 30◦C
with reciprocal shaking (120 rpm), and 100 µL of the culture was trans-ferred to 100 mL of the MSM medium amended with AFB1 (10 µg/mL) and/
or glucose (1g/100 mL). The following two controls were included: (a) MSMmedium with no cells and (b) MSM medium containing autoclaved Bacilluscells. After incubation, the cells were removed by centrifugation (10000 xg for5 min), the AFB1 was extracted from the supernatants and quantified by high-performance liquid chromatography (HPLC) as described in a later section.Bacterial cell numbers were determined at 24 h intervals up to 144 h, and asample was withdrawn from flasks for analysis of cell numbers and residues
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

Isolation of Endophytic Bacilli Bacterium 203
of AFB1. At each sampling period, serial dilutions were prepared with sterile0.8% saline solution and spread on LB agar plates. Plates were incubated at30◦C for 48 h.
Extraction and Quantification of Residual AFB1Preparation of sample was done by extracting culture supernatants with
ethyl acetate and removing the solvent on a rotary evaporator. The residue wasdissolved with 1 mL acetonitrile, filtered (Millex®HV PVDF 0.45µm, Millipore)and analyzed by HPLC. The HPLC method for AFB1 analysis was modifiedfrom Chan et al. (2004). The HPLC apparatus consisted of a solvent deliverysystem with both fluorescence (λex=364 nm; λem=440 nm) and UV detectors(λ=225 nm and 362 nm). The spectra range is from 200 to 500 nm. The ana-lytical column used was a 150x4.6 mm Uptisphere 5 µm C18 ODB fitted witha guard column of 10x4 mm. The column temperature was 25◦C. An aliquot ofsample (80 µL) was injected using an autoinjector (BIO-TEK, Milan, Italy).The mobile phase was 0.1% phosphoric acid (A) and methanol/acetonitrile(50:50) (B) delivered at flow rate of 1 mL/min for 30 min. The sample waseluted with a linear gradient for 30 min. AFB1 and its derivatives weredetected by comparing the elution time and maximum absorption of UV withthe standards.
Degradation of AFB1 by Culture Supernatants of Bacillus sp. TUBF1Bacterial strain was cultivated in 5 mL of LB medium at 30◦C with recip-
rocal shaking (120 rpm), and 100 µL of the culture was transferred to 100 mLof the same medium. After cultivation at 30◦C with shaking at 120 rpm for48 h, the cells were harvested by centrifugation (5000 xg, 10 min, 4◦C) and theextracellular fraction used for AFB1 degradation. The reaction mixture wasperformed in 2-mL-Eppendorf tubes in a final volume of 1,000 µL and an ini-tial AFB1 concentration of 10 µg/mL. This involved the addition of 10 µL ofAFB1 stock solution to 740 µL culture supernatant and 250 µL of 0.1M sodiumacetate buffer (pH 7.0). The mixture was incubated in the dark at 30◦C withoutshaking for different time intervals; samples were centrifuged at 10000 xg for5 min, filtered through Millex®HV PVDF 0.45µm, Millipore, and placed in vialsfor subsequent analysis. All experiments were carried out in triplicates. Eachexperiment was terminated by the addition of 500 µL of HPLC grade chloro-form for extraction of the remaining AFB1 and analyzed by HPLC as describedpreviously.
Enzyme PropertiesThe effect of temperature on the supernatants (crude enzyme(s)) activity
was estimated by the crude enzyme assay method, lightly modified, in the
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

204 B. El-Deeb et al.
temperature range from 10◦C, 20◦C, 30◦C, and 40◦C over a period of 20 h atpH 7.0. The supernatant was added to the reaction medium after the mixtureof buffer and substrate solutions had reached the reaction temperature. Thesame procedure was used to evaluate the effect of pH on the enzyme activity at37◦C, using 100 mM phosphate buffer for pH range from 5.0 to 7.5 and 50 mMTris-HCl buffer for pH between 8.0 and 8.8. All experiments were carried out intriplicates. Each experiment was terminated by the addition of 750 µL chloro-form for extraction of the remaining AFB1 and analysis by HPLC. To determinethe effect of temperature, the reaction mixtures was incubated at 20◦C, 30◦C,37◦C, and 45◦C for 72 h. Controls were set at the above temperatures by usingMSM medium.
Toxicity AssessmentThe toxicity of AFB1 and its degradative products were compared by bioas-
say against Artemia salina based on the method of Harwig and Scott (1971).In the bioassays, groups of 10–20 larvae were exposed to a range of concentra-tions of different AFB1 in a 96-well microtitration plates. Stock concentrationof toxin was prepared as described above. The final concentration of AFB1 was10 µg/mL, which was approximately 90% lethal concentration (LC90) for thelarvae. The AFB1 solutions were then treated with crude enzyme. Groups withonly toxins solution in wells served as negative controls, and those with onlycrude enzyme served as positive control. All treatments were in triplicate andall experiments were repeated three times with plates in the growth chamberat 28◦C and mortality evaluated at 28◦C. Complete immobility or paralysis wastaken as indication for neurotoxic activity towards the insect larvae.
Statistical AnalysisData were analyzed as a completely randomized single factor design by
ANOVA using the general linear models procedure in SAS. Significant F testsat the 0.05 levels of probability were reported. When a significant F-valuewas detected, Duncan’s Multiple Range Test was used to determine significantdifferences among means.
RESULTS AND DISCUSSION
Five strains of endophytic bacteria were isolated from contaminated corn seedwith AFB1. All of them were able to degrade AFB1 with different activity lev-els. One strain was selected due to its high removal efficiency of AFB1 (about80%) and was used for further studies and designated as TUBF1. Therefore,contaminated samples with aflatoxins, such as animal feces, decayed barks,
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

Isolation of Endophytic Bacilli Bacterium 205
soils, and cereal grains, were chosen as sources for selection of microorganismsthat degrade AFB1 (Völkl et al., 2004).
Taxonomic Characterization of the AFB1 Degrading StrainThe isolate TUBF1 was gram-positive bacterium, on the basis of mor-
phological, physiological, and biochemical characteristics (data not shown)and comparative analysis of the 16s rRNA gene sequence (720 bp) withalready available database, which showed that the TUBF1 was close to themembers of the genus Bacillus sp. (identity percentage 99%). The sequencewas deposited at GenBank (Bacillus sp. TUBF1, Accession no. HF546395).Although many authors have reported that Lactobacillus and Rhodococcus aremainly responsible for the detoxification of AFB1 A (El-Nezami et al., 1998;Teniola et al., 2005), numerous published work showed that many speciesapart from Lactobacillus and Rhodococcus can degrade aflatoxins, includingBacillus spp. (Petchkongkaew et al., 2008; Gao et al., 2011; Farzaneh et al.,2012). However, this is the first report indicating that endophytic bacterium inthis genus isolated from grain possesses function in mycotoxin degradation.
Growth KineticsThe growth curve of Bacillus sp. TUBF1 in MSM amended with glucose
or AFB1 is shown in Fig 1. The initial inoculums of Bacillus sp. TUBF1 wasapproximately at Log 5 CFU/mL. Comparing the growth curve of strainTUBF1 in growth medium, cell numbers of Bacillus sp. TUBF1 in growthmedium with an initial AFB1 of 10 µg/mL reached the maximum 8.2 CFU/mL,whereas cell numbers of TUBF1 in presence of glucose reached the maxi-mum 8.8 CFU/mL at 24 h. Moreover, cells increased after AFB1 was degraded(Fig. 1), indicating growth was directly proportional to AFB1 assimilation as asole source of carbon and energy. Degradation and assimilation of aflatoxins asa sole source of carbon and energy by different bacteria have been documented(Teniola et al., 2005; Alberts et al., 2006; Petchkongkaew et al., 2008; Gao et al.,2011; Farzaneh et al., 2012). With an initial AFB1 concentration of 10 µg/mL, ahigh reduction of AFB1 was observed when incubated in the presence of bacte-rial cells (Fig. 1). By contrast, no significant reduction took place in the absenceof bacterial cells. Similar results were obtained elsewhere. Teniola et al. (2005)reported a high reduction of AFB1 was observed during incubation in thepresence of Rhodococcus erythropolis cells. Hormisch et al. (2004) indicatedthat liquid cultures of Mycobacterium strain FA4 could reduce AFB1 level by70–80% within 36 h and completely degrade AFB1 in 72 h. In comparison, mostof the lactic acid bacteria that were able to bind AFB1 could rapidly removethe toxin from liquid; however, they would release AFB1 to some extent inthe prolonged incubation period (Peltonen et al., 2000; El-Nezami et al., 1998).
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

206 B. El-Deeb et al.
Figure 1: Bacterial degradation of AFB1 (10 µg/mL) by Bacillus sp. strain TUBF1 1 in liquid mineralmedium (MSM) at 30 oC and 150 rpm. Aflatoxin B1 concentrations were determined by HPLCanalysis of organic extracts from cell culture and uninoculated control. Growth is shown asincrease in the cell count in presence of glucose (10 mg/mL) only/or AFB1(10 µg/mL).
In the present study, the continuous increase in detoxification by Bacillus spTUBF1 with time indicated that binding might not play important role in theAFB1 reduction.
Degradation of AFB1 by Culture Supernatant of TUBF1AFB1 degradation by bacterial supernatant over a period of 24 h was
followed, and different levels of degradation were observed (Fig. 2). These
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

Isolation of Endophytic Bacilli Bacterium 207
Figure 2: Aflatoxin B1 (10 µg/mL) degradation kinetics observed over a period of 24 usingsupernatant of Bacillus sp. TUBF1. The values are means of three replicates and their standarderrors. Means with different letters are significantly different according to Duncan’s MultipleRange Test (P < 0.05).
results were similar to the observation demonstrated by (Mann and Rehm,1976; Smiley and Draughon, 2000). They concluded that the degradationof AFB1 could be enzymatic. Similar results were also obtained elsewhere.Alberts et al. (2006) reported that AFB1 was effectively degraded by extracellu-lar extracts from R. erythropolis liquid cultures. A novel aflatoxin degradationenzyme had been isolated and purified from Pleurotus ostreatus by Motomuraet al. (2003).
Effect of Temperature on the AFB1 Degradation by CultureSupernatant of TUBF1The effect of different temperatures on the loss of AFB1 by cell-free extracts
of TUBF1 strain was also evaluated (Fig. 3). Similar results were obtained byTeniola et al. (2005), who reported that AFB1 degradation by cell extracts of R.erythropolis and M. fluoranthenivorans were about the same between 10 and40◦C (> 90%). They proposed either that the enzymes in the extracts had awide temperature range of activity or that other factors were involved in thedegradation.
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3
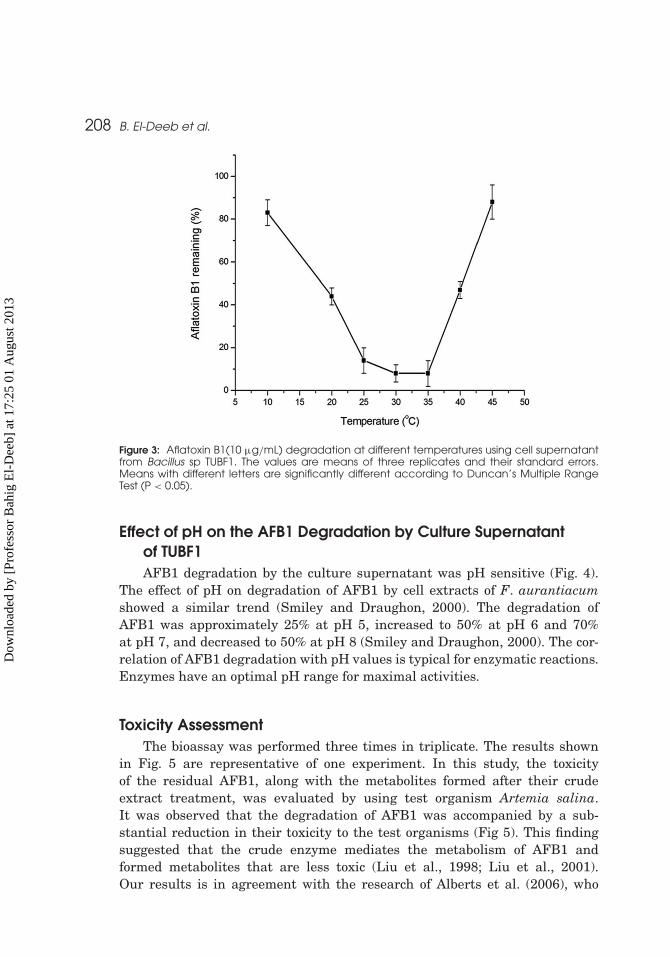
208 B. El-Deeb et al.
Figure 3: Aflatoxin B1(10 µg/mL) degradation at different temperatures using cell supernatantfrom Bacillus sp TUBF1. The values are means of three replicates and their standard errors.Means with different letters are significantly different according to Duncan’s Multiple RangeTest (P < 0.05).
Effect of pH on the AFB1 Degradation by Culture Supernatantof TUBF1AFB1 degradation by the culture supernatant was pH sensitive (Fig. 4).
The effect of pH on degradation of AFB1 by cell extracts of F. aurantiacumshowed a similar trend (Smiley and Draughon, 2000). The degradation ofAFB1 was approximately 25% at pH 5, increased to 50% at pH 6 and 70%at pH 7, and decreased to 50% at pH 8 (Smiley and Draughon, 2000). The cor-relation of AFB1 degradation with pH values is typical for enzymatic reactions.Enzymes have an optimal pH range for maximal activities.
Toxicity AssessmentThe bioassay was performed three times in triplicate. The results shown
in Fig. 5 are representative of one experiment. In this study, the toxicityof the residual AFB1, along with the metabolites formed after their crudeextract treatment, was evaluated by using test organism Artemia salina.It was observed that the degradation of AFB1 was accompanied by a sub-stantial reduction in their toxicity to the test organisms (Fig 5). This findingsuggested that the crude enzyme mediates the metabolism of AFB1 andformed metabolites that are less toxic (Liu et al., 1998; Liu et al., 2001).Our results is in agreement with the research of Alberts et al. (2006), who
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3
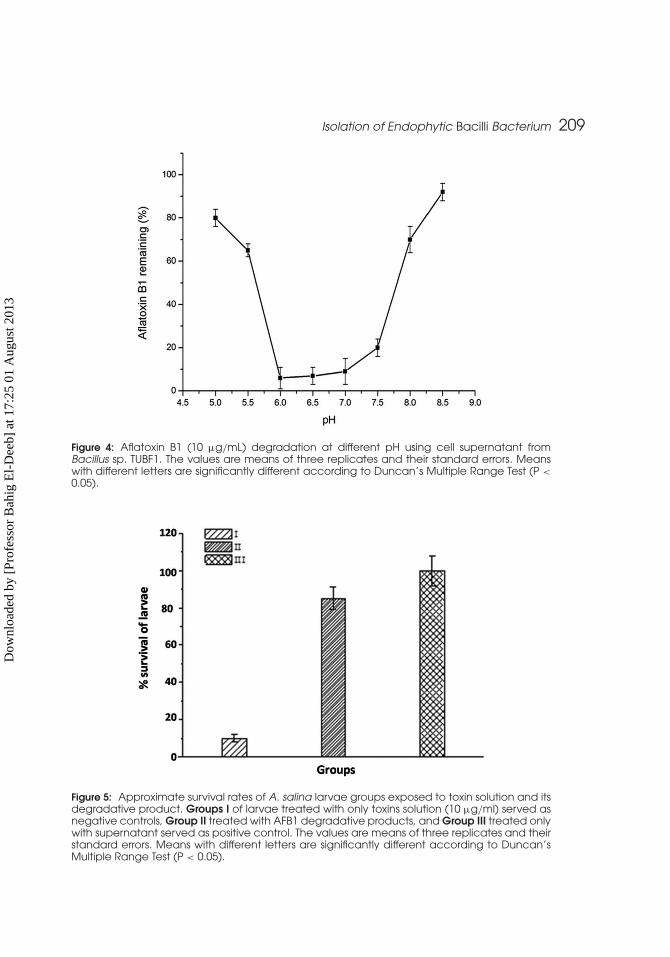
Isolation of Endophytic Bacilli Bacterium 209
Figure 4: Aflatoxin B1 (10 µg/mL) degradation at different pH using cell supernatant fromBacillus sp. TUBF1. The values are means of three replicates and their standard errors. Meanswith different letters are significantly different according to Duncan’s Multiple Range Test (P <
0.05).
Figure 5: Approximate survival rates of A. salina larvae groups exposed to toxin solution and itsdegradative product. Groups I of larvae treated with only toxins solution (10 µg/ml) served asnegative controls, Group II treated with AFB1 degradative products, and Group III treated onlywith supernatant served as positive control. The values are means of three replicates and theirstandard errors. Means with different letters are significantly different according to Duncan’sMultiple Range Test (P < 0.05).
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

210 B. El-Deeb et al.
reported that AFB1 was effectively degraded by extracellular extracts fromR. erythropolis liquid cultures, with only 3.2% residual AFB1 after 72 h.A novel aflatoxin degradation enzyme had been isolated and purified fromPleurotus ostreatus by Motomura et al. (2003). AFB1 was treated with culturesupernatants from 19 mushroom strains. The supernatant from P. ostreatusshowed aflatoxin-degradation activity, whereas other strains had no or onlyweak activity.
CONCLUSION
The high degradation rate and wide temperature and pH range for degrada-tion by Bacillus sp. TUBF1 supernatants indicate a potential and promisingapplication for the degradation of AFB1 in the foods and feeds processing.
ACKNOWLEDGMENT
This research was supported by the Research Program of Taif University, SaudiArabia (Project 1- 432- 689).
REFERENCES
Alberts, J.F., Engelbrecht, Y., Steyn, P.S., Holzapfel, W.H., van Zyl, W.H. (2006).Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures. Int.J. Food. Microbiol. 109:121–126.
Altalhi, D., El-Deeb, B. (2009). Localization of zearalenone detoxification gene(s) inpZEA-1 plasmid of Pseudomonas putida ZEA-1 and expressed in Escherichia coli.J. Hazard Mater. 161:1166–1172.
Byers, H.K., Stackebrandt, E., Hayward, C., Blackall, L.L. (1998). Molecular investiga-tion of a microbial mat associated with the great artesian basin. FEMS Microbiol.Ecol. 25:391–403.
Chan, D., MacDonald, S.J., Boughtflower, V., Brereton, P. (2004). Simultaneousdetermination of aflatoxins and ochratoxin A in food using a fully automatedimmunoaffinity column clean-up and liquid chromatography–fluorescence detec-tion. J. Chromatog. 1059:13–16.
Ciegler, A., Lillehoj, E.B., Peterson, R.E., Hall, H.H. (1966). Microbial detoxification ofaflatoxin. Appl. Microbiol. 14:934–938.
Cotty, P.J., Bhatnagar, D. (1994). Variability among atoxigenic Aspergillus flavusstrains in ability to prevent aflatoxin contamination and production of aflatoxinbiosynthetic pathway enzymes. Appl. Environ. Microbiol. 60:2248–2251.
Deiner, U.L., Cole, R.J., Sanders, T.H., Payne, G.A., Lee, L.S., Klich, M.A. (1987).Epidemiology of aflatoxin formation by Aspergillus flavus. Annu. Rev. Phytopathol.25:240–270.
Dietmar, B., Patricia, G., Munoz-Rojas, J., Estrella, D., Moreno-Morilla, S., Sanchez, L.,Ramos, J.L. (2008). Rhizormediation of lindane by root colonizing Sphingomonas.Microb. Biotechnol. 1:87–93.
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

Isolation of Endophytic Bacilli Bacterium 211
Eaton, D.L., Groopman, J.D. (1994). The Toxicology of Aflatoxins. Human Health,Veterinary and Agricultural Significance. San Diego, CA: Academic Press.
El-Nezami, H., Kankaanpaa, P., Salminen, S., Ahokas, J. (1998). Ability of dairy strainsof lactic acid bacteria to bind a common food carcinogen, Aflatoxin B1. Food Chem.Toxicol. 36:326–361.
Farzaneh, M., Shi, Z.-Q., Ghassempour, A., Sedaghat, N., Ahmadzadeh, M., Mirabolfath,M., Nikkhah, M.J. (2012). Aflatoxin B1 degradation by Bacillus subtilisUTBSP1 isolated from pistachio nuts of Iran. Food Control. 23:100–106.
Gao, X., Ma, Q., Zhao, L., Lei, Y. Shan, Y. (2011). Isolation of Bacillus subtilis: screeningfor aflatoxins B1, M1, and G1 detoxification. Euro. Food Res. Technol. 232:957–962.
Govindarajan, M., Kwon, S., Weon, H. (2007). Isolation, molecular characterization andgrowth-promoting activities of endophytic sugarcane diazotroph Klebsiella sp. GR9.World. Microbial. Biotechnol. 23:997–1006.
Guan, S., Ji, C., Zhou, T., Li, J., Ma, Q., Niu, T. (2008). Aflatoxin B(1) degradationby Stenotrophomonas maltophilia and other microbes selected using coumarinmedium. Int. J. Mol. Sci. 9:1489–1503.
Guengerich, F.P., Johnson, W.W., Veng, Y.F., Shimade, T. (1996). Involvementof cytochrome P450, glutathione S-tranferase and epoxide hydrolase in themetabolism of Aflatoxin B1 and relevance to risk of human liver cancer. Environ.Health Perspect. 104:557–562.
Harwig, J., Scott, P.M. (1971). Brine shrimp (Artemia salina L.) larvae as a screeningsystem for fungal toxins. Appl. Microbiol. 21:1011–1016.
Holt, J.G., Krieg, N.R., Sneath, P.H.A., Staley, J.T., Williams, S.T. (1994). Bergey’sManual of Determinative Bacteriology, 9th ed. Baltimore, MD: Williams & Wilkins.
Hormisch, D., Brost, I., Kohring, G.W., Giffhorn, F., Kroppenstedt, R.M., Stackebrandt,E., Färber, P., Holzapfel, W.H. (2004). Mycobacterium fluoranthenivorans sp. nov.,a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of aformer coal gas plant. Syst. Appl. Microbiol. 27:653–660.
Huynh, V.L., Gerdes, R.G., Lloyd, A.B. (1984). Synthesis and degradation of aflatoxinsby A. parasiticus. II: comparative toxicity and mutagenicity of aflatoxin B1 and itsautolytic breakdown products. Aust. J. Biol. Sci. 37:123–129.
Kabak, B., Dobson, A.D.W., Var, I. (2006). Strategies to prevent mycotoxin contamina-tion of food and animal feed: a review. Crit. Rev. Food Sci. Nutr. 46:593–619.
Kang, J.W., Khan, Z., Doty, S.L. (2012). Biodegradation of trichloroethylene by anendophyte of hybrid poplar. Appl. Environ. Microbiol. 78:3504–3507.
Kurtzman, C.P., Horn, B.W., Hesseltine, C.W. (1987). Aspergillus nomius, a newaflatoxin producing species related to Aspergillus flavus and Aspergillus tamarii.Antoine van Leeuwenhoek. 53:147–158.
Lillehoj, E.B., Stubblefield, R.D., Shannon, G.M., Shotwell, O.L. (1971). AflatoxinM1 removal from aqueous solutions by Flavobacterium aurantiacum. Mycopathol.Mycol. Appl. 45:259–266.
Line, J.E., Brackett, R.E. (1995). Factors affecting aflatoxin B1 removal byFlavobacterium aurantiacum. J. Food Prot. 58:91–94.
Line, J.E., Brackett, R.E., Wilkinson, R.E. (1994). Evidence for degradation of aflatoxinB1 by Flavobacterium aurantiacum. J. Food Prot. 57:788–791.
Liu, D.L., Yao, D.S., Liang, R., Ma, L., Cheng, W.Q., Gu, L.Q. (1998). Detoxification ofaflatoxin B1 by enzymes isolated from Armillariella tabescens. Food. Chem. Toxicol.36:563–574.
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3

212 B. El-Deeb et al.
Liu, D.L., Yao, D.S., Liang, Y.Q., Zhou, T.H., Song, Y.P., Zhao, L., Ma, L. (2001).Production, purification, and characterization of an intracellular aflatoxin-detoxifizyme from Armillariella tabescens (E-20). Food. Chem. Toxicol. 39:461–466.
Mann, R., Rehm, H.J. (1976). Degradation products from aflatoxin B1 byCorynebacterium rubrum, Aspergillus niger, Trichoderma viride and Mucorambiguus. Eur. J. Appl. Microbiol. 2:297–306.
Motomura, M., Toyomasu, T., Mizuno, K., Shinozawa, T. (2003). Purification and charac-terization of an aflatoxin degradation enzyme from Pleurotus ostreatus. Microbiol.Res. 158:237–242.
Peltonen, K., El-Nezami, H., Salminen, S., Ahokas, J. (2000). Binding of aflatoxin B1 byprobiotic bacteria. J. Sci. Food Agric. 80:1942–1945.
Petchkongkaew, A., Taillandier, P., Gasaluck, P., Lebrihi, A. (2008). Isolation of Bacillusspp. from Thai fermented soybean (Thua-nao): screening for aflatoxin B1 andochratoxin A detoxification. J. Applied Microbiol. 104:1495–1502.
Sambrook, J., Fritsch, E., Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual,2nd ed. New York: Cold Spring Harbor Laboratory Press, pp. 304–324, 363–371.
Shapira, R. (2004). Detection and control. In: Magan, N., Olsen, M., eds. Mycotoxins inFood. Boca Raton, FL: CRC Press, p. 207.
Smiley, R.D., Draughon, F.A. (2000). Preliminary evidence that degradation of aflatoxinB1 by Flavobacteria aurantiacum is enzymatic. J. Food Prot. 63:415–418.
Teniola, O.D., Addo, P.A., Brost, I.M., Färber, P., Jany, K.D., Alberts, J.F., van Zyl,W.H., Steyn, P.S., Holzapfel, W.H. (2005). Degradation of aflatoxin B1 by cell-freeextracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp.nov. DSM44556T. Int. J. Food Microbiol. 105:111–117.
Tsubouchi, H., Yamamoto, K., Hisada, K., Sakabr, Y. (1983). Degradation of aflatoxinsby A. niger and aflatoxin nonproducing A. flavus. Shakuhin Eiseigaku Zasshi24:113–119.
Völkl, A., Vogler, B., Schollenberger, M., Karlovsky, P. (2004). Microbial detoxificationof mycotoxin deoxynivalenol. J. Basic Microbiol. 44:147–156.
Zhao, L.H., Guan, S., Gao, X., Ma, Q.G., Lei, Y.P., Bai, X.M., Ji, C. (2010). Purificationand characteristics of an aflatoxin degradation enzyme from Myxococcus fulvusANSM068. J. Appl. Microbiol. 110:147–155.
Dow
nloa
ded
by [
Prof
esso
r B
ahig
El-
Dee
b] a
t 17:
25 0
1 A
ugus
t 201
3




















