
A Pivotal Prefix Based Filtering Algorithm for String Similarity ...

ORIGINAL ARTICLE
B-1 cells are pivotal for in vivo inflammatory giant cell formation
Cristina Stewart Bittencourt Bogsan, Ronni Romulo Novaes e Brito, Maiko da Cruz Palos, Renato
Arruda Mortara, Sandro Rogerio Almeida, Jose Daniel Lopes and Mario Mariano
Department of Microbiology, Immunology and Parasitology, Federal University of Sao Paulo, Sao Paulo, Brazil
Received for publication:
15 October 2003
Accepted for publication:
18 March 2005
Correspondence:
Prof Mario Mariano
Rua Botucatu 862
4th floor, 04023-9000 Sao Paulo
Brazil
Tel.: +55 11 5496073
Fax: +55 11 55723328
E-mail: [email protected]
Abstract
The mechanisms that govern giant cell (GC) formation in inflammatory, neoplastic
and physiologic conditions are far from being understood. Here, we demonstrate that
B-1 cells are essential for foreign-body GC formation in the mouse. GCs were analysed
on the surface of glass cover slips implanted into the subcutaneous tissue of the
animals. It was demonstrated that GCs are almost absent on cover slips implanted
into the subcutaneous tissue of BALB/c or CBA/N X-linked immunodeficient mice. As
these animals do not have B-1 cells in the peritoneal cavity, they were reconstituted
with B-1 cells obtained from cultures of adherent mouse peritoneal cells. Results
showed that in B-1-reconstituted animals, the number of GCs on the implant surface
surpassed the values obtained with preparations from wild animals. In animals select-
ively irradiated (pleural and peritoneal cavities) to deplete these cavities of B-1 cells,
GCs were also not formed. Enriched suspensions of B-1 cells grown in culture were
labelled with [3H]-tymidine and injected into the peritoneal cavity of naive mice before
implantation of glass cover slips. After 4 days, about 17% of mononuclear cells had
their nuclei labelled, and almost 70% of GCs had one or more of their nuclei labelled
when analysed by histoautoradiographic technique. A few GCs expressed an immuno-
globulin M when analysed by immunostaining and confocal microscopy. Overall, these
data demonstrate that B-1 cells are pivotal in the mechanisms of foreign-body GC
formation in the mouse.
Keywords
B-1 cells, inflammatory giant cell, macrophages
Although scientific reports found in the older literature
describing multinucleated cells are contemporaneous with
the emerging concept of Virchow’s cellular theory, the origin,
properties and fate of giant cells (GCs) are still poorly under-
stood. As reviewed by Harris (1970), the first description of
cell multinucleation was in 1838 with Muller who first
observed such cells in tumours of vertebrates. Later, in
1855, Rokitansky referred their presence in tuberculous tis-
sue. In 1858, Robin reported their presence among bone
marrow cells, and Virchow and Langhans showed that they
were present in a variety of normal, inflammatory and neo-
plastic lesions in 1858 and 1868, respectively. Although at
that times the biological significance of multinucleation was
unexplained, Metchnikoff (1968) hypothesized that fusion of
phagocitic cells to form ‘plasmodia’ was a characteristic cel-
lular defence mechanism and suggested, on a mechanistic
basis, that fusion of macrophages would result in a more
efficient cellular element to cope with parasites. Although
Int. J. Exp. Path. (2005), 86, 257–265
� 2005 Blackwell Publishing Ltd 257

provocative, his hypothesis has not yet been fully confirmed
or refuted. However, GCs have the capacity to phagocytose
particles via CR1 and Fc receptors (Mariano & Spector 1974;
Papadimitriou & Walters 1979). This phagocytic capacity
was less effective when compared with that of inflammatory
macrophages (Papadimitriou et al. 1975), but it remains to be
clarified whether or not these cells are endowed with bacter-
icidal ability.
GCs are a hallmark histological feature of some types of
chronic inflammation and neoplasia. They occur in various
conditions where the pathological agent is known, e.g.
bacterial, viral, parasitic and fungal infections as well as
where it is not known, e.g. sarcoidosis and rheumatoid
arthritis. GCs are formed in foreign-body lesions induced
by asbestos, carbon, silica and sutures (Papadimitriou &
Walters 1979; Chambers & Spector 1982; Anderson 2000).
They are observed at the tissue–material interface of med-
ical devices implanted in soft and hard tissue and remain
for the lifetime of the device implantation (Anderson 2000).
Foreign-body implantation in man (Keymeulen &
Dillemans 2004) and animals (Brand et al. 1977) can
induce the development of sarcomas. It was also demon-
strated that GCs can undergo aberrant mitosis which
results in chromosome pulverization and micro and poly-
ploid nuclei formation (Mariano & Spector 1974). The
possible relationship between these two phenomena has
not been addressed in the literature.
About 30 years ago, experimental evidences suggested, as
Metchnikoff predicted, that GCs or polykaria are formed in
foreign-body lesions induced in mice by fusion of inflamma-
tory mononuclear cells (Gillman & Wright 1966; Mariano &
Spector 1974). However, since then there has been little
advance in knowledge about the mechanism of the fusion
process itself. It is not clear how mononuclear cell fusion is
induced in vivo, and whether different mechanisms are
involved in different pathological conditions.
A number of possible mechanisms have been discussed in
the literature about how the formation of multinucleated
GCs might be induced in vitro and in vivo. In vitro, these
include the use of conditioned medium (Postlethwaite et al.
1982; Kreipe et al. 1988; Most et al. 1990), several different
cytokines (Nagasawa et al. 1987; Weinberg et al. 1987;
McInnes & Rennick 1988; Enelow et al. 1992a, 1992b;
Fais et al. 1994, 1997; Kazazi et al. 1994; McNally &
Anderson 1995), the addition of lectins alone or in combin-
ation with interferon-g (Chambers 1978; Takashima et al.
1993) and the addition of anti-macrophage antibodies or
phorbol myristate acetate or a combination thereof
(Chambers 1978; McInnes & Rennick 1988; Hassan et al.
1989). Despite all these efforts to understand the
mechanisms of multinucleation, it is not known why GCs
are spontaneously formed during long-term culture of differ-
ent cell types (Most et al. 1997).
Few studies have addressed the question of whether in vivo
multinucleate GCs originate from fusion of macrophages or
other cell types (Gillman & Wright 1966; Spector & Lykke
1966; Mariano & Spector 1974; Chambers 1977; Most et al.
1997). Some time ago, Mariano & Spector (1974) showed
experimental evidence that these cells are formed in foreign-
body lesions by fusion of two cell types called by the authors
old and young or newly arrived ‘macrophages’. Based on the
demonstration that structural chromosome alterations occur
in inflammatory cells in the cover slip model, these authors
hypothesized that the ‘newly arrived cells’, by recognizing
genetically altered cells, would result in cell fusion with
consequent GC formation. Chambers (1977) argued this
hypothesis further with evidence that GCs are formed when
more than one macrophage tempts to phagocytose a single
particle resulting in cell membrane fusion. Nevertheless,
neither the nature of the two cell populations involved in
GC formation nor the concomitant phagocytosis hypothesis
proposed by Chambers (1977) have been fully validated or
refuted.
We have recently demonstrated that B-1b cells – a subtype
of B cell lineage which express surface markers such as
Mac-1, IgM, IgD and B220 promiscuously (Iacomini &
Imanishi-Kari 1992) and found predominantly in pleural
and peritoneal cavities – have a peculiar morphology
(Abrahao et al. 2003), down-regulate macrophage functions
in vitro via IL-10 (Popi et al. 2004) and proliferate in long-
term cultures of adherent mouse peritoneal cells. Further,
these cells can be modulated towards a novel type of mono-
nuclear phagocyte as demonstrated by Borrello et al. (2001),
secrete IgM in vitro and can migrate from the peritoneal
cavity to a non-specific inflammatory lesion induced in
mice (Almeida et al. 2001). Based on these observations
and using the X-linked immunodeficient (Xid) mice, a
mouse strain that have Bruton’s tyrosine kinase mutation
and lacks B-1 cells (Scher 1982; Hardy et al. 1983), we
investigated whether B-1 cells might be involved in foreign-
body GC formation. Following this hypothesis, here we
present evidences that B-1 cells do indeed participate in GC
formation in a foreign-body model of inflammation.
Materials and methods
Animals
BALB/c, BALB/c Xid, CBA/J and CBA/N Xid mice, 8–12-
week-old, were used. Mice were obtained from animal
258 C. S. B. Bogsan et al
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265

facilities of the Department of Immunology, University of Sao
Paulo and UNICAMP, Sao Paulo, Brazil.
B1-b-cell cultures
B1-b cells were produced according to the method proposed
by Almeida et al. (2001). Briefly, the peritoneal cavities of
normal BALB/c mice were washed with 6 ml of RPMI 1640
medium (Sigma Chemical Co., St Louis, MO, USA). Cells
were cultured at 37 �C in a humidified atmosphere containing
5% CO2 in RPMI 1640 medium supplemented with 10% of
foetal calf serum (FCS) in 75 cm2 culture vessels (BD Falcon,
Bedford, MA, USA). After 1 h, the supernatant of these cul-
tures were discarded, and the adherent cells were harvested
and re-suspended in RPMI 1640 medium supplemented with
10% FCS. After 7 days of culture without changing the med-
ium, about 90% of the cells in the supernatant of the cultures
were characterized as B1-b cells by flow-cytometry analysis.
The cover slip model
Round glass cover slips (13 mm) were implanted into the
subcutaneous tissue of mice from different experimental
groups as described (Ryan & Spector 1970; Mariano &
Spector 1974). After 4 days of implantation, cover slips were
removed and the cells on their surface fixed in 3.5% formalin
for 1 h, stained with haematoxilin-eosin or used for histoau-
dioradiographic analysis. Using a 20x objective, the mean of
the total number of multinucleated cells/microscopic field
obtained from 10 microscopic fields per cover slip was
recorded. The number of nuclei per GC was also evaluated.
A minimum of five animals with two implanted cover slips in
each was used to constitute the experimental groups.
Confocal microscopy
Cover slips were implanted in normal BALB/c mice, removed
after 4 days and the cells on their surface prepared for the
detection of immunoglobulin M (IgM) and F4/80 (Santa Cruz
Biotechnology, Santa Cruz, CA, USA) using immunostaining
and confocal microscopy as described previously (Abrahao
et al. 2003).
Flow cytometry
Two-colour flow cytometry was used to determine the pheno-
type of peritoneal cells collected from normal, irradiated and
reconstituted mice. Cells were collected by peritoneal lavage
with phosphate-buffered saline (PBS). After centrifugation,
cells were washed with PBS plus 10% FCS, and 10 000 events
were stained with monoclonal antibodies (mAbs) directed to
mouse CD45/B220 and CD11b (Mac-1) (Pharmingen, San
Diego, CA, USA). Analysis was performed using a fluorescein
isothiocyanate-labelled Mac-1, as well as biotin-conjugated
B220 mAbs (Pharmingen, San Diego, CA, USA). Cells were
analysed in a FACS ConvertTM.Software, Facscalibur (Becton
Dickinson, Franklin Lakes, NJ, USA).
Selective body irradiation
The thoracic and abdominal regions of mice were irradiated
with 900 rad using the technique proposed by Novaes and
Brito et al. (unpublished data). Briefly, the animals were held
in a lead device that protects their fore and hind limbs from
irradiation
Cell transfer experiments
Adherent peritoneal cells collected from the peritoneal cavity
of BALB/c mice were cultured for 7 days at 37 �C and pulsed,
for 16 h, with [3H]-tymidine (1 mCi/ml). Free-floating cells
were harvested, washed three times in cold PBS, and aliquots
were adjusted to have 5 · 106 cells in 0.5 ml of PBS. These
aliquots were injected into the peritoneal cavity of naive
BALB/c mice. One hour after cell inoculation, two pockets
were opened in the subcutaneous tissue of the dorsal region of
the animals into which a round glass cover slip was implanted.
Cover slips were removed after 4 days, fixed in Bouin’s fixa-
tive for 2 h, mounted in a glass slide and prepared for his-
toautoradiographic analysis. The slides were coated with K5
autoradiographical emulsion (Ilford Ltd, Ilford, UK) by the
dipping method (Kopriwa & Leblond 1962). After exposure
at 4 �C for 30 days, the radio autographs were developed with
D19b developer (Ilford) fixed and stained with haematoxylin
and eosin.
Reconstitution of CBA/N (Xid) mice with B-1 cells
CBA/N mice were reconstituted with 2.5 · 106 B-1 cells
obtained from cultures of adherent peritoneal cells from
CBA/J mice. Cells were injected by intraperitoneal route,
and 1 week later, groups of CBA/J, CBA/N and CBA/N recon-
stituted animals were implanted with glass cover slips. Cover
slips were removed after 4 days, and the cells on their surface
stained with haematoxylin-eosin and mounted for micro-
scopic examination. The absence of B-1 cells in Xid mice
and irradiated mice and their presence in the peritoneal cavity
of reconstituted animals was monitored by flow cytometry.
B-1 cells and inflammatory giant cell formation 259
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265
Statistical analysis
Comparisons among groups were made using ordinary ANOVA.
Statistical significance was assumed at P < 0.05.
Results
B-1 cells are almost absent in the peritoneal cavity of
irradiated mice but present in reconstituted animals
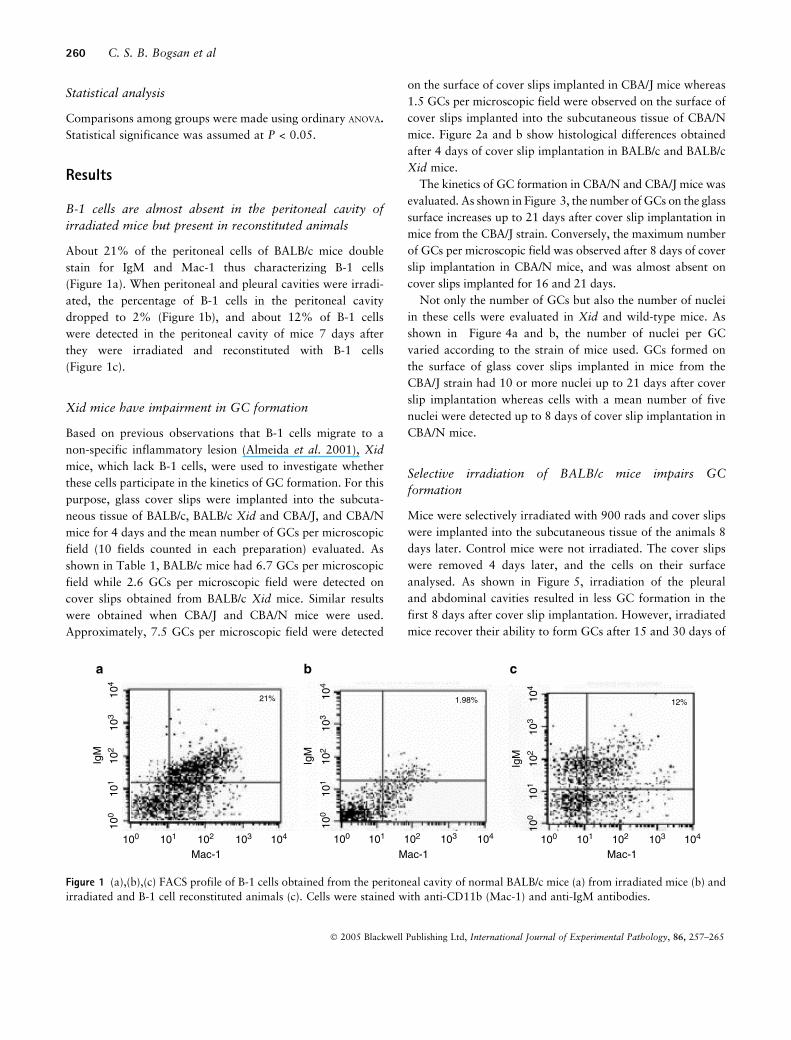
About 21% of the peritoneal cells of BALB/c mice double
stain for IgM and Mac-1 thus characterizing B-1 cells
(Figure 1a). When peritoneal and pleural cavities were irradi-
ated, the percentage of B-1 cells in the peritoneal cavity
dropped to 2% (Figure 1b), and about 12% of B-1 cells
were detected in the peritoneal cavity of mice 7 days after
they were irradiated and reconstituted with B-1 cells
(Figure 1c).
Xid mice have impairment in GC formation
Based on previous observations that B-1 cells migrate to a
non-specific inflammatory lesion (Almeida et al. 2001), Xid
mice, which lack B-1 cells, were used to investigate whether
these cells participate in the kinetics of GC formation. For this
purpose, glass cover slips were implanted into the subcuta-
neous tissue of BALB/c, BALB/c Xid and CBA/J, and CBA/N
mice for 4 days and the mean number of GCs per microscopic
field (10 fields counted in each preparation) evaluated. As
shown in Table 1, BALB/c mice had 6.7 GCs per microscopic
field while 2.6 GCs per microscopic field were detected on
cover slips obtained from BALB/c Xid mice. Similar results
were obtained when CBA/J and CBA/N mice were used.
Approximately, 7.5 GCs per microscopic field were detected
on the surface of cover slips implanted in CBA/J mice whereas
1.5 GCs per microscopic field were observed on the surface of
cover slips implanted into the subcutaneous tissue of CBA/N
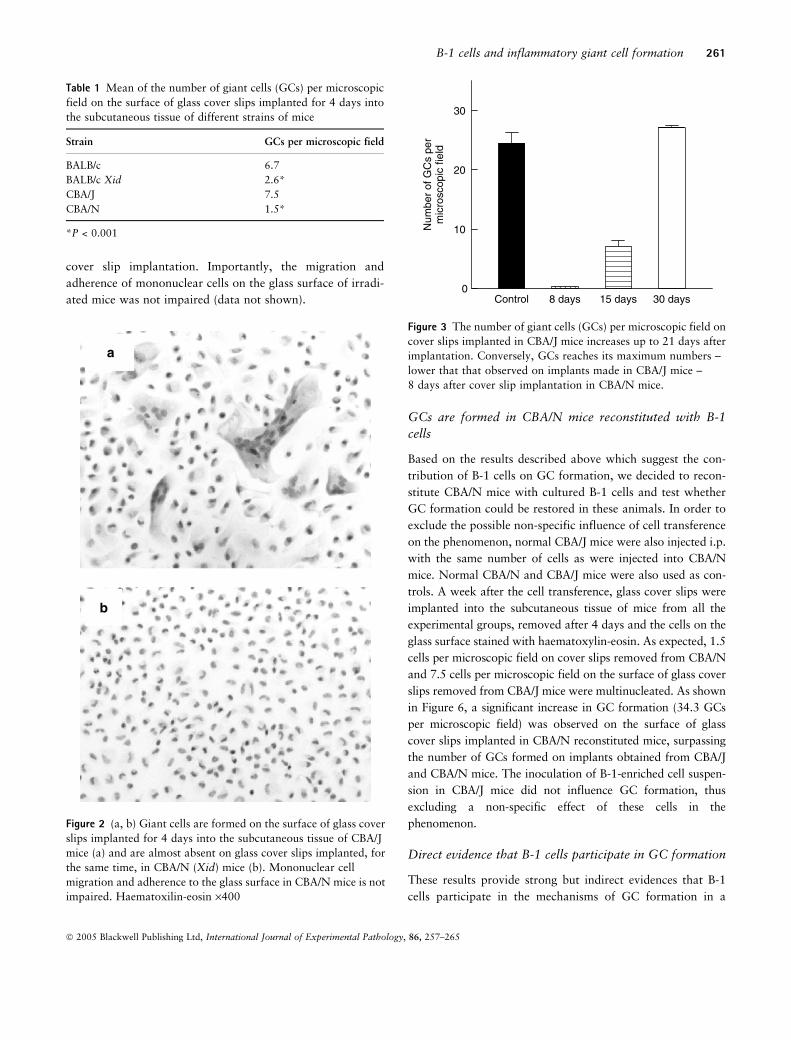
mice. Figure 2a and b show histological differences obtained
after 4 days of cover slip implantation in BALB/c and BALB/c
Xid mice.
The kinetics of GC formation in CBA/N and CBA/J mice was
evaluated. As shown in Figure 3, the number ofGCs on the glass
surface increases up to 21 days after cover slip implantation in
mice from the CBA/J strain. Conversely, the maximum number
of GCs per microscopic field was observed after 8 days of cover
slip implantation in CBA/N mice, and was almost absent on
cover slips implanted for 16 and 21 days.
Not only the number of GCs but also the number of nuclei
in these cells were evaluated in Xid and wild-type mice. As
shown in Figure 4a and b, the number of nuclei per GC
varied according to the strain of mice used. GCs formed on
the surface of glass cover slips implanted in mice from the
CBA/J strain had 10 or more nuclei up to 21 days after cover
slip implantation whereas cells with a mean number of five
nuclei were detected up to 8 days of cover slip implantation in
CBA/N mice.
Selective irradiation of BALB/c mice impairs GC
formation
Mice were selectively irradiated with 900 rads and cover slips
were implanted into the subcutaneous tissue of the animals 8
days later. Control mice were not irradiated. The cover slips
were removed 4 days later, and the cells on their surface
analysed. As shown in Figure 5, irradiation of the pleural
and abdominal cavities resulted in less GC formation in the
first 8 days after cover slip implantation. However, irradiated
mice recover their ability to form GCs after 15 and 30 days of
100 101 102 103 104 100 101 102 103 104 100 101 102 103 104
104
103
102
101
100
104
103
102
101
100
104
103
102
101
100
21% 1.98% 12%
a b c
Mac-1Mac-1Mac-1
IgM
IgM
IgM
Figure 1 (a),(b),(c) FACS profile of B-1 cells obtained from the peritoneal cavity of normal BALB/c mice (a) from irradiated mice (b) and
irradiated and B-1 cell reconstituted animals (c). Cells were stained with anti-CD11b (Mac-1) and anti-IgM antibodies.
260 C. S. B. Bogsan et al
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265
cover slip implantation. Importantly, the migration and
adherence of mononuclear cells on the glass surface of irradi-
ated mice was not impaired (data not shown).
GCs are formed in CBA/N mice reconstituted with B-1
cells
Based on the results described above which suggest the con-
tribution of B-1 cells on GC formation, we decided to recon-
stitute CBA/N mice with cultured B-1 cells and test whether
GC formation could be restored in these animals. In order to
exclude the possible non-specific influence of cell transference
on the phenomenon, normal CBA/J mice were also injected i.p.
with the same number of cells as were injected into CBA/N
mice. Normal CBA/N and CBA/J mice were also used as con-
trols. A week after the cell transference, glass cover slips were
implanted into the subcutaneous tissue of mice from all the
experimental groups, removed after 4 days and the cells on the
glass surface stained with haematoxylin-eosin. As expected, 1.5
cells per microscopic field on cover slips removed from CBA/N
and 7.5 cells per microscopic field on the surface of glass cover
slips removed from CBA/J mice were multinucleated. As shown
in Figure 6, a significant increase in GC formation (34.3 GCs
per microscopic field) was observed on the surface of glass
cover slips implanted in CBA/N reconstituted mice, surpassing
the number of GCs formed on implants obtained from CBA/J
and CBA/N mice. The inoculation of B-1-enriched cell suspen-
sion in CBA/J mice did not influence GC formation, thus
excluding a non-specific effect of these cells in the
phenomenon.
Direct evidence that B-1 cells participate in GC formation
These results provide strong but indirect evidences that B-1
cells participate in the mechanisms of GC formation in a
a
b
Figure 2 (a, b) Giant cells are formed on the surface of glass cover
slips implanted for 4 days into the subcutaneous tissue of CBA/J
mice (a) and are almost absent on glass cover slips implanted, for
the same time, in CBA/N (Xid) mice (b). Mononuclear cell
migration and adherence to the glass surface in CBA/N mice is not
impaired. Haematoxilin-eosin ·400
Num
ber
of G
Cs
per
mic
rosc
opic
fiel
d
Control 8 days 15 days 30 days0
10
20
30
Figure 3 The number of giant cells (GCs) per microscopic field on
cover slips implanted in CBA/J mice increases up to 21 days after
implantation. Conversely, GCs reaches its maximum numbers –
lower that that observed on implants made in CBA/J mice –
8 days after cover slip implantation in CBA/N mice.
Table 1 Mean of the number of giant cells (GCs) per microscopic
field on the surface of glass cover slips implanted for 4 days into
the subcutaneous tissue of different strains of mice
Strain GCs per microscopic field
BALB/c 6.7
BALB/c Xid 2.6*
CBA/J 7.5
CBA/N 1.5*
*P < 0.001
B-1 cells and inflammatory giant cell formation 261
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265

foreign-body-induced lesion. Based on previous observations
that B-1b cells migrate from the peritoneal cavity to a non-
specific inflammatory lesion (Almeida et al. 2001), we inves-
tigated whether B-1 cells could fuse to form GCs. For this
purpose, B-1 cells labelled with [3H]-thymidine in vitro were
injected intraperitoneally into BALB/c mice followed by the
implantation of glass cover slips into the subcutaneous tissue
of the animals. The cover slips were removed after 4 days and
prepared for autoradiographic analysis. Results showed that
about 70% of multinuclear GCs had at least one labelled
nucleus, and 17% of the mononuclear adherent cells were
labelled (Figure 7).
IgM is expressed by GCs
As shown in Figure 8, the cytoplasm of a multinucleated cell
on the surface of glass cover slip implanted for 4 days in BALB/
c mice immunostained for both IgM in the cytoplasm and F4/
80 in the outer cell membrane. This pattern of reaction was not
constantly observed. The positive reaction was only obtained
when cells were treated with detergent to permeabilize the cell
membrane, indicating an intracellular location of IgM.
Num
ber
of G
Cs
per
mic
rosc
opic
fiel
d
2–5 6–10 n > 100
10
20
a
b
4 days
8 days
16 days
21 days
Number of nuclei per GC
Num
ber
of G
Cs
per
mic
rosc
opic
fiel
d
2–5 6–10 n > 100.0
2.5
5.0
7.5
10.0
4 days
8 days
16 days
21 days
Figure 4 (a,b) The number of nuclei per giant cell was
evaluated. Figure 4a shows that cells with 10 or more nuclei
were observed on the surface of glass cover slips implanted for up
to 21 days into the subcutaneous tissue of mice from the CBA/J
strain. However, cells with no more then five nuclei were
observed on the surface of cover slips implanted for 8 days into
the subcutaneous tissue of CBA/N mice.
Num
ber
of G
Cs
per
mic
rosc
opic
fiel
d
Control 8 days 15 days 30 days0
10
20
30
Figure 5 The selective irradiation of CBA/J mice induces
impairment in giant cell formation up to 8 days of cover slip
implantation. This effect is progressively reversed after 15 and 30
days of irradiation.
Num
ber
of G
Cs
per
mic
rosc
opic
fiel
d
Control B-1 transferred0
10
20
30
40 CBA/J
CBA/N*
Figure 6 CBA/N mice were reconstituted with B-1 cells obtained
from cultures of adherent CBA/J peritoneal cells. CBA/J mice
were also injected with the same cell suspension. Results show
that transference of B-1 cells to CBA/J mice does not influence the
number of giant cells (GCs) formed on the surface of implanted
cover slips. Conversely, the cell transference to CBA/N mice
restored their ability to form GCs.
262 C. S. B. Bogsan et al
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265

Discussion
The origin, properties and fate of foreign body and other types
of GCs are fragmentary. Here, we bring direct evidences that
GCs induced by the implantation of glass cover slips into the
subcutaneous tissue of mice is dependent on the participation
of B-1 cells.
The Xid (Amsbaugh et al. 1972; Scher 1982) share features
with the severe immunodeficiency linked with the X chromo-
some that occurs in man. In Xid mice, the number of B-2 cells
(conventional B cells) is only slightly reduced. Here, we con-
firm that these mice lack or have only small numbers of cells
(Riggs et al. 2003) with the characteristics of B-1b cells,
expressing IgMlo, IgDhi, MAC-1 and B-220 antigens (Scher
1982; Hardy et al. 1983). Another subset of B-1 cells, the B-1a
cells have all the characteristics of B-1b cells but are also
CD5+. B-1a cells are also absent in Xid mice (Iacomini &
Imanishi-Kari 1992).
The hypothesis that GCs are formed by the concurrence of
two populations of macrophages one ‘old’ and another newly
arrived from circulation, as proposed by Mariano & Spector
(1974), and the recent demonstration by our group that B-1
cells exit coelomatic cavities and migrate to an inflammatory
site where they can transform into a novel type of mono-
nuclear phagocyte not related to the monocyte-derived macro-
phage (Almeida et al. 2001), led us to investigate the possible
role that B-1 cells might play in GC formation.
In a first set of experiments, we analysed the kinetic of GC
formation in wild and Xid mice. Results clearly show that Xid
mice have impairment in GC formation. In addition, there are
also fewer nuclei in GCs formed in Xid mice when compared
with preparations obtained from wild mice. Further indirect
evidence of the participation of B-1 cells in the phenomenon
was obtained with the irradiation and cell transfer experiments.
B1 cells are radiosensitive (Almeida et al. 2001), and when the
peritoneal and pleural cavities of the animals were irradiated,
GC formation was drastically reduced. Further, the reconstitu-
tion of Xid mice with B-1 cells from adherent peritoneal cells
fromwildmice resulted in an increase in the number of GCs that
surpassed that observed in control groups.
The transfer experiments with [3H]-thymidine-labelled B-1
cells and the immunostaining for IgM expression analysed by
confocal microscopy brought further evidence that B-1 cells
participate in the mechanisms of GC formation in this model.
The transfer experiments with [3H]-thymidine-labelled B-1
cells showed clearly that not only mononuclear cells with a
macrophage-like morphology but also a large proportion of
GCs had labelled nuclei thus indicating that B-1 cells migrate
from the peritoneal cavity to the inflammatory milieu. Almost
all the GCs in these preparations had from one to five labelled
Figure 8 Cells on the surface of glass cover slips implanted for
4 days into the subcutaneous tissue of BALB/c mice were
labelled with monoclonal antibodies anti-F4/80 (red), and
immunoglobulin (IgM) (green) and DAPI (blue) and analysed
by confocal microscopy. A typical foreign-body giant cell (GC)
expressing IgM in the cytoplasm and F4/80 in the outer
membrane is shown. F4/80+ mononuclear cells are in intimate
contact with the GC. Arrows indicate mononuclear cells
expressing IgM.
Figure 7 [3H]-thymidine was added to 7-day cultures of
adherent peritoneal cells from BALB/c mice. Labelled cells
were collected after 16 h and transferred to the peritoneal
cavity of naive BALB/c mice. One hour after cell transfer-
ence, glass cover slips were implanted into the subcutaneous
tissue of the animals and removed after 4 days for auto-
radiographic analysis. Silver grains are observed on the
nuclei of a typical Langerhans giant cell (GC). Haematoxilin-
eosin ·1000
B-1 cells and inflammatory giant cell formation 263
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265

nuclei. The non-specific labelling of these cells is difficult to be
admitted, because the B-1 cells were labelled in vitro, thor-
oughly washed before transference and analysed after 4 days.
The demonstration that GCs have IgM in their cytoplasm
also supports the view that B-1 cells participate in the genesis
of these cells although endocytosis of IgM should be excluded.
This hypothesis can be ruled out on the basis that B-1 cells
secret large amounts of IgM in vivo (Stall et al. 1992) and in
vitro (Almeida et al. 2001). Second, it has been demonstrated
in our laboratory that these cells have a promiscuous expres-
sion of myeloid and lymphoid transcription factors, loosing
the expression of lymphoid factors as they differentiate into a
mononuclear phagocyte. As analysed by immunostaining and
confocal microscopy, B-1 cells loose IgM expression in the
course of differentiation keeping the expression of myeloid
markers (Popi et al. 2004). The same pattern of immunostain-
ing for IgM was observed in cells on the cover slip surface.
Although most of the mono and multinucleated cells stained
for myeloid markers (Mac-1, F4/80) the expression of both
IgM/F4/80 was detected in a fewer number of cell. If endo-
cytosed, it should be expected IgM to be present in the cyto-
plasm of the majority of cells on the glass surface.
Although these data demonstrate the participation of B-1
cells in GC formation, it remains to be seen whether GCs
are solely B-1 derived or whether they result from fusion of
B-1 derived mononuclear phagocytes and monocyte-derived
macrophages. This question is an issue under investigation
in our laboratory. It also remains to be demonstrated
whether GC formation in different pathologies are depend-
ent on B-1 cell participation. If so, it opens the possibility
not only to better understand the biologic significance of
the intriguing phenomenon of phagocyte multinucleation in
vivo but also to investigate the participation of adaptive
immunocompetent cells in determining the course of non-
immune inflammation.
Acknowledgements
This paper is dedicated to the memory of W.G. Spector. We
are indebted to Prof Telma Zorn for the preparation of the
historadiograms. We are supported by grant from Fundacao
de Amparo a Pesquisa do Estado de Sao Paulo (FAPESP), no
99/11144–4
References
Abrahao T.B., Freymuller E., Mortara R.A., Lopes J.D.,MarianoM.
(2003) Morphological characterization of mouse B-1 cells.
Immunobiology. 208, 401–411.
Almeida S.R., Aroeira L.S., Frymuller E. et al. (2001) Mouse B-1
cell-derived mononuclear phagocyte, a novel cellular compo-
nent of acute non-specific inflammatory exudate. Int.
Immunol. 13, 1193–1201.
Amsbaugh D.F., Hansen C.T., Prescott B., Stashak P.W.,
Barthold D.R., Baker P.J. (1972) Genetic control of the anti-
body response to type 3 pneumococcal polysaccharide in mice.
I. Evidence that an X-linked gene plays a decisive role in
determining responsiveness. J. Exp. Med. 136, 931–949.
Anderson J.M. (2000) Multinucleated giant cells. Curr. Opin.
Hematol. 7, 40–47.
BorrelloM.A., Palis J., Phipps R.P. (2001) The relationship of CD5+
B lymphocytes to macrophages: insights from normal bipheno-
typic b/macrophage cells. Int. Rev. Immunol. 20, 137–155.
Brand I., Buoen L.C., Brand K.G. (1977) Foreign-body tumors of
mice: strain and sex differences in latency and incidence.
J. Natl. Cancer. Inst. 58, 1443–1447.
Chambers T.J. (1977) Failure of altered macrophage surface to
lead to the formation of polykaryons. J. Pathol. 122, 185–189.
Chambers T.J. (1978) Multinucleate giant cells. J. Pathol 126,
125–148.
Chambers T.J. & Spector W.G. (1982) Inflammatory giant cells.
Immunobiology 161, 283–289.
Enelow R.I., Sullivan G.W., Carper H.T., Mandell G.L. (1992a)
Cytokine-induced human multinucleated giant cells have
enhanced candidacidal activity and oxidative capacity com-
pared with macrophages. J. Infect. Dis. 166, 664–668.
Enelow R.I., Sullivan G.W., Carper H.T., Mandell G.L. (1992b)
Induction of multinucleated giant cell formation from in vitro
culture of human monocytes with interleukin-3 and interferon-
gamma: comparison with other stimulating factors. Am. J.
Respir. Cell. Mol. Biol. 6, 57–62.
Fais S., Burgio V.L., Capobianchi M.R., Gessani S., Pallone F.,
Belardelli F. (1997) The biological relevance of polykaryons in
the immune response. Immunol. Today. 18, 522–527.
Fais S., Burgio V.L., Silvestri M., CapobianchiM.R., Pacchiarotti A.,
Pallone F. (1994) Multinucleated giant cells generation
induced by interferon-gamma. Changes in the expression and
distribution of the intercellular adhesion molecule-1 during
macrophages fusion and multinucleated giant cell formation.
Lab. Invest. 71, 737–744.
Gillman T. & Wright L.J. (1966) Probable in vivo origin of multi-
nucleated giant cells from circulating mononuclears. Nature.
209, 263–265.
Hardy R.R., Hayakawa K., Parks D.R., Herzenberg L.A. (1983)
Demonstration of B-cell maturation in X-linked immunodefi-
cient mice by simultaneous three-colour immunofluorescence.
Nature. 306, 270–272.
Harris H. (1970) Cell Fusion. pp. 108. Ipswich: Oxford
University Press.
Hassan N.F., Kamani N., Meszaros M.M., Douglas S.D. (1989)
Induction of multinucleated giant cell formation from human
264 C. S. B. Bogsan et al
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265

blood-derived monocytes by phorbol myristate acetate in in
vitro culture. J. Immunol. 143, 2179–2184.
Iacomini J. & Imanishi-Kari T. (1992) The effect of an immuno-
globulin mu transgene on B cell maturation. Eur. J. Immunol.
22, 745–751.
Kazazi F., Chang J., LopeZ. A., Vadas M., Cunningham A.L.
(1994) Interleukin 4 and human immunodeficiency virus sti-
mulate LFA-1-ICAM-1-mediated aggregation of monocytes
and subsequent giant cell formation. J. Gen. Virol. 75 (10),
2795–2802.
Keymeulen K. & Dillemans B. (2004) Epitheloid angiosarcoma of
the splenic capsula as a result of foreign body tumorigenesis. A
case report. Acta. Chir. Belg. 104, 217–220.
Kopriwa B. & Leblond C. (1962) Improvements in the coating
technique of radioautography. J. Histochem. Cytochem. 10,
269–284.
Kreipe H., Radzun H.J., Rudolph P. et al (1988) Multinucleated
giant cells generated in vitro. Terminally differentiated macro-
phages with down-regulated c-fms expression. Am. J. Pathol.
130, 232–243.
Mariano M. & Spector W.G. (1974) The formation and proper-
ties of macrophage polykaryons (inflammatory giant cells).
J. Pathol. 113, 1–19.
McInnes A. & Rennick D.M. (1988) Interleukin 4 induces cul-
tured monocytes/macrophages to form giant multinucleated
cells. J. Exp. Med. 167, 598–611.
McNally A.K. & Anderson J.M. (1995) Interleukin-4 induces
foreign body giant cells from human monocytes/macrophages.
Differential lymphokine regulation of macrophage fusion leads
to morphological variants of multinucleated giant cells. Am. J.
Pathol. 147, 1487–1499.
Metchnikoff E. (1968) Lectures on the Comparative Pathology of
Inflammation. New York: Dover Publications, Inc.
Most J., Neumayer H.P., Dierich M.P. (1990) Cytokine-induced gen-
eration of multinucleated giant cells in vitro requires interferon-
gamma and expression of LFA-1. Eur. J. Immunol. 20, 1661–1667.
Most J., Spotl L., Mayr G., Gasser A., Sarti A., Dierich M.P.
(1997) Formation of multinucleated giant cells in vitro is
dependent on the stage of monocyte to macrophage matur-
ation. Blood. 89, 662–671.
Nagasawa H., Miyaura C., Abe E., Suda T., Horiguchi M. (1987)
Fusion and activation of human alveolar macrophages induced
by recombinant interferon-gamma and their suppression by
dexamethasone. Am. Rev. Respir. Dis. 136, 916–921.
Papadimitriou J.M. & Robertson T.A., Walters M.N. (1975) An
analysis of the phagocytic potential of multinucleate foreign
body giant cells. Am. J. Pathol. 78, 343–358.
Papadimitriou J.M. & Walters M.N. (1979) Macrophage poly-
karya. CRC. Crit. Rev. Toxicol. 6, 211–255.
Popi A.F., Lopes J.D., Mariano M. (2004) Interleukin-10 secreted
by B-1 cells modulates the phagocytic activity of murine macro-
phages in vitro. Immunology. 113, 348–354.
Postlethwaite A.E., Jackson B.K., Beachey E.H., Kang A.H.
(1982) Formation of multinucleated giant cells from human
monocyte precursors. Mediation by a soluble protein from
antigen-and mitogen-stimulated lymphocytes. J. Exp. Med.
155, 168–178.
Riggs J., Howell K., Matechin B., Matlack R., Pennello A.,
Chiasson R. (2003) X-chromosome-linked immune-deficient
mice have B-1b cells. Immunology. 108, 440–451.
Ryan G.B. & Spector W.G. (1970) Macrophage turnover in
inflamed connective tissue. Proc. R. Soc. Lond. B Biol. Sci.
174, 269–292.
Scher I. (1982) The CBA/N mouse strain: an experimental model
illustrating the influence of the X-chromosome on immunity.
Adv. Immunol. 33, 1–71.
Spector W.G. & Lykke A.W. (1966) The cellular evolution of
inflammatory granulomata. J. Pathol. Bacteriol. 92, 163–167.
Stall A.M., Adams S., Herzenberg L.A., Kantor A.B. (1992)
Characteristics and development of the murine B-1b (Ly-1 B
sister) cell population. Ann. N Y Acad. Sci. 651, 33–43.
Takashima T., Ohnishi K., Tsuyuguchi I., Kishimoto S.
(1993) Differential regulation of formation of multi-
nucleated giant cells from concanavalin A-stimulated human
blood monocytes by IFN-gamma and IL-4. J. Immunol. 150,
3002–3010.
Weinberg J.B., Hobbs M.M., Misukonis M.A. (1987)
Recombinant human gamma-interferon induces human mono-
cyte polykaryon formation. Proc. Natl. Acad. Sci. USA. 81,
4554–4557.
B-1 cells and inflammatory giant cell formation 265
� 2005 Blackwell Publishing Ltd, International Journal of Experimental Pathology, 86, 257–265
Copyright © 2022 FDOKUMEN




















