
AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA
616
AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA i
Transcript of AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
i

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
ii

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
iii
Antártica: nuestra ventana para ver el mundo
Como todos sabemos, hace unos 20.000 años el planeta vivió el Último Máximo Glacial. La Patagonia y gran parte de los Andes se cubrieron de hielo, Argentina y Brasil tuvieron climas templados: América entera fue como un soplo blanco y prolongado del polo sur. Hoy, pasados 20.000 años, la Antártica parece cubrirnos nuevamente, pero esta vez con la clara voz de la ciencia. Se cumplen ya siete versiones de este encuentro de investigación polar latinoamericana, que naciera casi como la reunión de unos pocos locos interesados en la Antártica y que es ahora un congreso y con toda propiedad. Los títulos de los simposios que forman este congreso (Sistemas de observación antárticos terrestres y costeros; Genómica de microorganismos antárticos: avances y desafíos; Ecofisiología vegetal; Glaciología y Cambio Climático; Nuevos avances en el estudio de las aves antárticas; Evolución de la biota del Meso-Cenozoico; y Océano Austral) demuestran el alto nivel y especificidad que hemos alcanzado. Si a esto agregamos el número de trabajos presentados (más de 160) y la excelencia de las conferencias, podemos afirmar que el Congreso Latinoamericano de Ciencia Antártica tiene una calidad difícil de igualar. Es paradójico que un lugar tan lejano como la Antártica sirva para congregar a personas de países tan diversos como Rusia, China, Corea del Sur, con quienes los países latinoamericanos suelen compartir en la isla Rey Jorge. Igual de curioso es que tengamos la misma blanca excusa para reunirnos como estamos haciendo ahora en la hermosa ciudad de La Serena. Junto al Congreso Latinoamericano tienen lugar otras reuniones que vale la pena nombrar: Reunión de la Red Latinoamericana de Microbiología Antártica, Reunión de la Red Latinoamericana de Investigación en Plantas Antárticas y Mesa Redonda de APECS Latinoamérica. También quisiera destacar que junto a este congreso, hemos organizado la XXIV Reunión de Administradores de Programas Antárticos Latinoamericanos (RAPAL), para que los diversos operadores tengan la oportunidad de interiorizarse en los temas y avances científicos polares y así cuenten con mayores antecedentes al momento de tomar decisiones que digan relación con el desarrollo de la actividad antártica futura. Esta confluencia nos lleva a decir que la Antártica es nuestra ventana para ver el mundo. En realidad, no sólo para verlo, sino que para participar en él. Fenómenos como el agujero de ozono ayer y el calentamiento global hoy, han puesto los ojos del mundo en el Continente Blanco y nuestros países están contribuyendo, en una forma reconocida internacionalmente, al avance del conocimiento científico en estas problemáticas. La vocación antártica de nuestros países nace, sin duda, de la cercanía que tenemos con dicho continente. Pero este es sólo un punto de partida. El desafío siguiente es asegurar que lo que hacemos en esas altas latitudes sea algo significativo para los ciudadanos de nuestros respectivos países (a fin de cuentas, son los principales financistas de nuestras tareas) y también para la comunidad internacional. El nivel de este congreso nos dice que vamos por buen camino, porque si establecemos reglas claras, que estimulen la competencia y el respeto por estándares reconocidos por la comunidad científica, entonces, los resultados serán de provecho no sólo para cada una de nuestras naciones, sino también para el amplio foro polar y científico. Es una enorme alegría que en este congreso se premie el Espíritu Antártico del Dr. Francisco Hervé. Con “Pancho” hemos compartido muchas veces y en todas hemos aprendido algo. Su trayectoria ha sido

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
iv
ejemplar en cada uno de los aspectos de la vida académica: como científico, como profesor y como divulgador, además de ser una excelente persona. La geología y las ciencias de la Tierra en la Antártica no tendrían el desarrollo que tienen en Chile sin el aporte fundamental y prolongado del Dr. Hervé. Finalmente, agradecemos el apoyo del Centro de Estudios Avanzados en Zonas Áridas (CEAZA), que ha sumado sus esfuerzos con el Instituto Antártico Chileno (INACH) para hacer posible esta reunión. También agradecemos el auspicio de la Corporación de Fomento de la Producción (Corfo) y el apoyo de las empresas Ivens, Bask, Maptek, Antarctica XXI y DAP.
Dr. José Retamales Espinoza Director Instituto Antártico Chileno

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
v
Premio al Espíritu Antártico 2013 FRANCISCO HERVÉ ALLAMAND
Geólogo de la Universidad de Chile (1965), especializado en petrología metamórfica. Doctor de
3er Ciclo de la Universidad de París (Francia, 1968) y Doctor en Ciencias de la Universidad de Hokkaido (Japón, 1974). Es profesor titular de las universidades de Chile y Andrés Bello. En 1985, año en que fue instaurado, recibió el Premio Nacional de Geología; en 1996, el Premio Cátedra Presidencial en Ciencias. Es miembro de la Academia Chilena de Ciencias, donde ocupa el sillón N° 36. Con más de 450 publicaciones científicas, este “chico de Providencia” ha incursionando también en la narrativa y es autor de “Soy Jemmy Button, el salvaje”, una biografía novelada sobre la increíble historia de este famoso yámana y su lado no-salvaje (2003).
Su primer viaje a la Antártica fue en el verano de 1963-64. Alentado por el profesor Humberto Fuenzalida, junto a su compañero Roberto Araya estudió las rocas de las islas Shetland del Sur cuando las investigaciones chilenas eran escasas. Aún conserva un pingüino papúa de aquel primer viaje.
En la temporada siguiente, fue parte de la primera expedición organizada por el Instituto Antártico Chileno y pudo comprobar, como conclusión de su memoria de título, que los glaciares del área estaban disminuyendo. Desde entonces, de a poco le han ido hablando las rocas y hoy lidera un proyecto que busca comprender en profundidad las conexiones geológicas entre Sudamérica y la Antártica.
El espíritu inquisitivo y colaborativo de Francisco Hervé lo ha vinculado a numerosos grupos de investigación nacionales y extranjeros, siendo incesante en la generación de nuevos conocimientos y en el uso de nuevas tecnologías. Ha formado varias generaciones de geólogos chilenos y, pese a las adversidades, ha conseguido que un buen puñado de ellos se dedique a la investigación polar. En este sentido, su labor pedagógica y divulgativa es extraordinaria, logrando desarrollar la capacidad de exponer en forma precisa y amena ante diversos públicos, académicos y no académicos, las complejidades de las investigaciones geológicas. Más aún, ha contribuido a consolidar la imagen de la Antártica como un lugar fundamental para analizar los procesos de la Tierra.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
vi
Índice
Gómez-Fuentes, et al. DISEÑO, CONSTRUCCIÓN Y OPERACIÓN DE UN SISTEMA DE TRATAMIENTO DE AGUAS RESIDUALES EN BAHÍA FILDES Y SU INFLUENCIA EN EL MEDIO AMBIENTE. (Design, construction and operation of a wastewater treatment system in Fildes Bay and its influence on the environment)
1
Chiarandini, et al. CARACTERIZACION AMBIENTAL PARA EL USO SOSTENIDO DEL AGUA POTABLE EN BASE ESPERANZA - NORTE DE LA PENINSULA ANTARTICA ARGENTINA (Environmental characterization for sustainable use of water at Hope Bay- Antarctic Peninsula North – Argentina)
5
Mónica Puentes, et al. CONSTRUCCIÓN DEL MAPA DE CORROSIÓN ATMOSFÉRICA DE CHILE: ESTACION ANTARTICA (Map construction of atmospheric corrosion of Chile: Antarctic station)
9
Acuña-Rodríguez IS, et al.TENDENCIAS DE LA INVESTIGACIÓN ECOLOGÍCA ANTÁRTICA DURANTE EL ÚLTIMO SIGLO (Trends in Antarctic ecological research over the last century)
12
Lemmertz, et al. LOS LÍMITES Y LAS LIMITACIONES DE LA CIENCIA: CONSIDERACIONES SOBRE LA AUTONOMIA DE LOS INVESTIGADORES BRASILEROS EN ANTÁRTICA (The limits and limitations of science: autonomy considerations of Brazilian researchers in Antarctica)
16
Pellizari, et al.ELUCIDANDO OS MECANISMOS MOLECULARES DE ADAPTAÇÃO DA BACTÉRIA EXIGUOBACTERIUM ANTARCTICUM (Dilucidando los mecanismos moleculares de adaptación de la bactéria Exiguobacterium antarcticum)
20
García-Sancho, L.INFLUENCIA DE LOS FACTORES CLIMÁTICOS EN LA DIVERSIDAD Y FUNCIONAMIENTO DE LAS COMUNIDADES VEGETALES ANTÁRTICAS. POSIBLE RESPUESTA EN UN ESCENARIO DE CALENTAMIENTO GLOBAL(Influence of climatic factors on the diversity and functioning of Antarctic plant communities. Possible answer in a global warming scenario)
21
Bergstrom, D.M. COLLAPSE OF A SUB-ANTARCTIC ALPINE FELDMARK ECOSYSTEM (El colapso de un ecosistema alpino subantártico de feldmark)
21
Cavieres, et al.CARACTERÍSTICAS ECOFISIOLÓGICAS DE LAS PLANTAS VASCULARES ANTÁRTICAS: CERTEZAS, INCERTEZAS E IMPLICANCIAS PARA LAS CONSECUENCIAS DEL CAMBIO CLIMÁTICO (Ecophysiological characteristics of the Antarctic vascular plants: certainties, uncertainties and implications for consequences of climate change)
22
Callejas, et al.ANÁLISIS DE LA ESTRUCTURA DE CONSORCIOS MICROBIANOS TERRESTRES DE LA PENÍNSULA DE FILDES (ANTÁRTIDA MARÍTIMA) MEDIANTE PIROSECUENCIACIÓN (Analysis by pyrosequencing of microbial structure in terrestrial mat consortia of Fildes Peninsula, maritime Antarctica)
26
Ferrés, et al. USO DE LA METAGENÓMICA FUNCIONAL PARA LA IDENTIFICACIÓN DE ENZIMAS DE POTENCIAL INTERÉS BIOTECNOLÓGICO PRESENTES EN LA PENÍNSULA ANTARTICA (Functional metagenomics as a tool for the identification of biotechnological interesting enzymes in the Antarctic Peninsula)
29
Rodríguez-Marconi, et al.CARACTERIZACIÓN TAXONÓMICA Y FUNCIONAL DE LAS COMUNIDADES NANOEUCARIONTES FOTOSINTÉTICAS SOMETIDAS A DISTINTOS REGÍMENES DE LUZ EN BAHÍA FILDES, ANTÁRTICA, MEDIANTE SECUENCIACIÓN MASIVA (Taxonomic and functional characterization of photosynthetic nanoeukariotic community subjected to different light regimes in Fildes Bay, Antarctica, through massive sequencing technology)
33
Ratti, et al. DETERMINACIÓN DE LA TAXONOMÍA Y DIVERSIDAD DE POBLACIONES MICROBIANAS EN SUELOS ANTÁRTICOS A TRAVÉS DE DGGE Y SECUENCIACIÓN 454 (Determination of the taxonomy and diversity of soil microbial populations based on DGGE fingerprinting and 454 sequencing)
37
Corcuera, et al. ECOFISIOLOGÍA DE PLANTAS ANTÁRTICAS: UN NUEVO PORTAL WEB (Ecophysiology of Antarctic Plants: a new web page)
40
Reyes, et al.PATRONES DE ESTRUCTURACION POBLACIONAL A DOS ESCALAS GEOGRAFICAS DEL ALGA ROJA, GIGARTINA SKOTTSBERGII (Patterns of populations structure of a red algae, Gigartina skottsbergii at two geographic scale)
41

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
vii
Mansilla , et al.PRIMER REGISTRO DE VARIACION DE LA ACTIVIDAD FOTOSINTÉTICA EN Desmarestia menziesii J. AGARDH (DESMARESTIALES, PHAEOPHYCEAE) A TRAVES DE UN GRADIENTE LATITUDINAL EN LA PENÍNSULA ANTÁRTICA (First record of changes in photosynthetic activity of Desmarestia menziesii J. Agardh throughout a latitudinal gradient in the Antarctic peninsula)
45
Klagges, et al.LAS POBLACIONES DE COLOBANTHUS QUITENSIS PRESENTAN DIFERENCIACIONES MORFO- FISIOLÓGICAS QUE PODRÍAN EVIDENCIAR LA FORMACIÓN DE ECOTIPO EN SU HÁBITAT (Colobanthus quitensis populations have morpho-physiological differentiations which might evidence a habitat ecotype formation)
49
Gutiérrez, et al.EFECTO DE LA DESCARGA DE AGUA DULCE SOBRE LA ESTRUCTURA DE LA COMUNIDAD MICROBIANA EN EL FIORDO ADYACENTE AL GLACIAR JORGE MONTT, PATAGONIA CHILENA(Effect of freshwater flux on microbial community structure in the fjord adjacent to glacier Jorge Montt, Chilean Patagonia)
53
Franco, et al. ESTRUCTURA DE LAS COMUNIDADES MICROBIANAS PRESENTES EN SEDIMENTOS MARINOS COSTEROS EN TRES VERANOS AUSTRALES DE LA BAHIA DEL ALMIRANTADO, ISLA REY JORGE, PENINSULA ANTARTICA (Microbial community structure found in coastal marine sediments during three austral summers in Admiralty Bay, king George Island)
57
Antelo, et al. PRESENCIA DE INTEGRONES CLASE I EN COMUNIDADES BACTERIANAS TERRESTRES DE LA ISLA REY JORGE (Detection of integrons (intI gene) in terrestrial samples collected in King George island)
59
Ramírez, et al.DIVERSIDAD BACTERIANA EN SUELOS ANTÁRTICOS DESDE DIVERSOS ASENTAMIENTOS ANIMALES (Bacterial diversity in Antarctic soils from different animal settlements)
63
Cebuhar , et al.ESTRÉS OXIDATIVO ENPINGÜINOS PAPÚA Y ADELIA EN ZONAS DE BAJO Y ALTO IMPACTO ANTRÓPICO DE BAHIA ESPERANZA (Oxidative stress inGentoo and Adelie penguins in areas of low and high anthropogenic impact at Hope Bay)
67
Di Fonzo, et al. ESTRÉS OXIDATIVO DETERMINADO DURANTE TRES VERANOS CONSECUTIVOS EN SANGRE DE PINGÜINOS ANTÁRTICOS (Blood oxidative stress determination of Antarctic penguins in three consecutive summers)
71
González, , et al. PROTEINAS DE ESTRÉS EN INVERTEBRADOS MARINOS ANTARTICOS: UNA VISIÓN MOLECULAR Y FISIOLOGICA (Stress proteins in Antarctic marine invertebrates: a molecular and physiological view)
75
Jara, et al. NIVELES DE CONTAMINANTES ORGANOCLORADOS PERSISTENTES Y CONCENTRACIÓN DE PORFIRINAS PARA EVALUAR EXPOSICIÓN EN PINGÜINOS PYGOSCELIS PRESENTES EN LA PENÍNSULA ANTÁRTICA (Organochlorine contaminants and porphyrins levels for of the exposure assessment in Pygoscelis penguins present in the Antarctic Peninsula)
78
Canário, et al. MERCURIO EN LA ISLA DE DECEPCIÓN, ANTARTIDA – UN ESTUDO INTEGRADO (Mercury in Deception Island, Antarctica: An integrated study)
82
González, et al. AREAS LIBRES DE HIELO Y DISTRIBUCION DE Sanionia uncinata (Hedw.)Loeske EN LAS ISLAS SHETLAND DEL SUR, ANTARTICA. (Ice-free areas and distribution of Sanionia uncinata (Hedw.)Loeske on the South Shetland Islands, Antarctica)
85
Michel, et al. STOCK DE CARBONO ORGÁNICO EN EL SUELO DE LA PENÍNSULA FILDES Y LA ISLA ARDLEY (Soil Organic Carbon Stock of Fildes Peninsula and Ardley Island)
89
Salamene, et al. EL ANÁLISIS EXPLORATORIO DE LOS DATOS DE TEMPERATURA DEL SUELO EN SEIS SITIOS DE MONITOREO DE LA PENÍNSULA KELLER, ISLA REY JORGE (Exploratory data analysis of soil temperature at six sites monitoring the Keller Peninsula, King George Island)
93
Rojas, et al. ACTIVIDAD ANTICONGELANTE DE PÉPTIDOS SINTÉTICOS DE MOTIVOS SECUENCIALES DE PROTEINAS ANTICONGELANTES (Antifreeze activity of synthesized peptides with sequential motifs of natural antifreeze proteins)
97
Contreras, et al. CARACTERIZACIÓN MOLECULAR Y ANÁLISIS DE LA PRODUCCIÓN DE METABOLITOS CON POTENCIAL BIOTECNOLÓGICO DE PROBABLES CEPAS DE X. DENDRORHOUS AISLADAS DESDE EL TERRITORIO ANTARTICO CHILENO (Molecular characterization and metabolite with biotechnological potential production analysis of Xanthophyllomyces
101

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
viii
dendrorhous likely strains isolated from the Chilean Antarctic Territory)
Rabbia, et al. ACTIVIDAD DE DIFERENTES ANTIBIOTICOS SOBRE CEPAS DE Escherichia coli AISLADAS DESDE MUESTRAS DE AGUA DE BAHÍA FILDES EN ISLA REY JORGE, ANTÁRTICA.(Activity of different antibiotics against Escherichia coli strains isolated from water samples from Fildes Bay in King George Island, Antarctica)
105
Plaza, et al. BIOSÍNTESIS DE NANOPARTÍCULAS FLUORESCENTES MEDIANTE BACTERIAS PSICROTOLERANTES DEL GÉNERO PSYCHROBACTER SPP.: VENTAJAS COMPARATIVAS DEL CONTINENTE ANTÁRTICO (Biosynthesis of fluorescent nanoparticles using psychrotolerant bacteria genus Psychrobacter spp.: Comparative advantages of Antarctic continent)
109
Órdenes-Aenishanslins, et al. FABRICACIÓN DE CELDAS SOLARES DEL TIPO DYE-SENSITIZED SOLAR CELL (DSSC) MEDIANTE LA INCORPORACIÓN DE PIGMENTOS PROVENIENTES DE BACTERIAS ANTÁRTICAS (Solar cell manufacturing type dye-sensitized solar cell (DSSC) by the incorporation of pigments obtained from Antarctic bacteria)
113
Díaz-Vásquez, et al. AISLAMIENTO Y CARACTERIZACIÓN DE BACTERIAS RESISTENTES A MERCURIO Y TELURITO PROVENIENTES DEL TERRITORIO ANTÁRTICO CHILENO (Isolation and characterization of mercury- and tellurite-resistant bacteria from the Chilean Antarctic Territory)
117
Muñoz-Villagrán, et al. CARACTERIZACIÓN DE LA RESPUESTA A TELURITO EN BACTERIAS HIPER-RESISTENTES AISLADAS DEL TERRITORIO ANTÁRTICO CHILENO (Characterization of the tellurite response in hyper-resistant bacteria from the Chilean Antarctic Territory)
121
Valdivia, et al. IDENTIFICACIÓN Y CARACTERIZACIÓN DE UNA SHEWANELLA ANTARTICA REDUCTORA DE TELURITO Y COBRE (Identification and characterization of a tellurite- and copper-reducing Antarctic Shewanella)
125
Simões, DESAFIOS DA PESQUISA GLACIOLÓGICA E CLIMÁTICA ANTÁRTICA: UM OLHAR SUL-AMERICANO (Challenges of the Antarctic glaciological and climatic research: A South American point of view)
129
MacDonell, et al. LA IMPORTANCIA HIDROLÓGICA DE "CRYOCONITE HOLES" SOBRE UN GLACIAR EN ANTÁRTICA (The hydrological significance of cryoconite holes on an Antarctic glacier)
130
Bezada, et al. EVIDENCIAS GEOMORFOLOGICAS Y ESTRATIGRAFICAS DEL RETROCESO ACTUAL DEL GLACIAR QUITO Y DE UNA GLACIACION TERCIARIA EN LA ISLA GREENWICH, ARCHIPIELAGO SHETLAND DEL SUR, CONTINENTE ANTÁRTICO. (Geomorphologic and stratigraphic evidence of the current retreat of Quito Glacier and a Tertiary glaciations on Greenwich Island, South Shetland islands, Antarctica)
134
Goyanes, et al. ESTADO TÉRMICO Y ESPESOR DE CAPA ACTIVA EN EL COLLADO IRIZAR (ISLA DECEPCIÓN, ANTÁRTIDA): PERIODO 2009-2013. (Thermal state and active layer thickness in Collado Irizar, Deception Island, Antarctica: 2009-2013 period)
137
Carmona, et al. ESTACIONALIDAD EN LOS DATOS DE GRAVIMETRÍA Y ALTIMETRÍA RADAR EN LA ANTÁRTICA Y VARIABILIDAD CLIMÁTICA ASOCIADA. APLICACIÓN A LA PENÍNSULA ANTÁRTICA. PRIMERA PARTE: RESULTADOS GRAVIMETRÍA SATELITAL (Seasonality in gravimetry and radar altimetry data in the Antarctic and associated climate variability. Application to the Antarctic Peninsula. Part I: Satellite gravimetry results)
140
Chacón, et al. DIFIEREN LOS PATRONES DE CICLAJE DE P ENTRE LAS ÁREAS LIBRES DE HIELO Y LAS FRONTERAS DE LOS GLACIARES EN LA REGIÓN DE LA MARÍTIMA ANTÁRTICA? (Do P cycling patterns differ between ice-free areas and glacial boundaries in the Maritime Antarctic region?)
145
Jaña, et al. ESTUDIO DE LAS PROPIEDADES DE LA NIEVE MEDIANTE DATOS SATELITALES EN GLACIAR UNION, HERITAGE RANGE, WEST ANTARCTICA (Snow property study based on remote sensing data at Union Glacier, Heritage Range, West Antarctica)
149
Pętlicki, et al. DISTRIBUCIÓN DE TAMAÑO Y TIEMPO DE OCURRENCIA DE EVENTOS DE DESPRENDIMIENTO. INFORMACIÓN OBTENIDA DE LA COMBINACIÓN DE DATOS DE VIDEO Y DE ESCANEO A TRAVÉS DE LASER TERRESTRE (Calving events size and time distribution from combined Terrestrial Laser Scanning and video data)
153
Zamora, et al. CAMPAÑAS GLACIOLOGICAS DEL CECs Y ALE A UNION GLACIER 2008-2010, ANTÁRTICA OCCIDENTAL (CECs and ALE glaciological surveys between 2008 to 2010 to Union Glacier, West Antarctica)
155

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
ix
Castro, et al: HIBRIDACIÓN ENTRE Catharacta maccormicki Y Catharacta lonnbergi EN LA ISLA REY GEORGE (ANTÁRTIDA), DETECTADA POR EL ENFOQUE GENÉTICO EN PEQUEÑA ESCALA. (Hybridization between Catharacta maccormicki and Catharacta lonnbergi in King George Island (Antarctic) as detected by genetic approach in small scale)
159
Orgeira, et al. RESPUESTAS DE AVES MARINAS PELÁGICAS DEL MAR DEL SCOTIA, ANTÁRTIDA, ANTE FLUCTUACIONES CLIMÁTICAS (Responses of pelagic seabirds of the Scotia Sea, Antarctica, to climatic fluctuations)
162
Valenzuela-Guerra, et al.IMPLICANCIAS DE LA VARIACIÓN MORFOLÓGICA A NIVEL GEOGRÁFICO ENTRE LAS COLONIAS DE PINGÜINOS PAPUA (PYGOSCELIS PAPUA) PARA EL USO DE MEDIDAS MORFOMÉTRICAS COMO MÉTODO DE SEXAJE (Implications of geographic morphological variation among colonies of Gentoo penguin (Pygoscelis papua) using morphometric measurements as a method of sex determination)
166
Costa, et al. REPRODUCTIVE BIOLOGY AND BEHAVIOR, NON-INVASIVE ANALYSES OF MERCURY AND STRESS ECOLOGY OF ANTARCTIC SKUAS (CATHARACTA SPP.) (Biología reproductiva y comportamiento, análisis no invasivo de mercurio y ecología de estrés de la skúa antártica (Catharacta spp.))
170
Leppe, et al. PUENTES FINICRETÁCICOS ENTRE PATAGONIA Y ANTÁRTICA: EVIDENCIAS DESDE LA PALEOBIOLOGÍA(End-Cretaceous bridges between Patagonia and Antarctica: evidences from the paleobiology)
171
Nishida, et al. RESTOS VEGETALES PERMINERALIZADOS: UNA NUEVA VENTANA A LA VEGETACIÓN PASADO DE PATAGONIA Y ANTÁRTICA (Permineralized plant debris: A novel window to the past vegetation of Patagonia and Antarctica)
175
Reguero, et al. LAND VERTEBRATES BIOGEOGRAPHY OF WEST ANTARCTICA/SOUTH AMERICA AND THE FINAL BREAK-UP OF GONDWANA (Biogeografía de vertebrados terrestres de Antártida oeste/Sudamérica y la ruptura final de Gondwana)
179
Ortiz, et al. REGISTRO DE LOS PLESIOSAURIOS MÁS AUSTRALES DE AMÉRICA: SISTEMÁTICA, PALEOAMBIENTE Y ESTRATIGRAFÍA (Record of plesiosaurs southernmost of America: Systematics, paleoenvironment and stratigraphy)
182
Ruiz Barlett, et al. VARIABILIDAD TEMPORAL DE MASAS DE AGUA EN EL MAR DE LA FLOTA (Temporal variability of water masses in Bransfield Strait)
186
Bastos, et al. VARIAÇÃO DOS DADOS HIDROQUÍMICOS E BIOLÓGICOS EM ÁGUAS DE SUPERFÍCIE DO ESTREITO DE BRANSFIELD (ANTÁRTICA), CONSIDERANDO DOIS DIFERENTES ESTÁGIOS DO PERÍODO DE VERÃO AUSTRAL EM 2011 (Variation of hydrochemical and biological data in Bransfield Strait surface waters (Antarctica), considering two different stages of austral summer period in 2011)
190
Moreno, et al. CARACTERIZACIÓN ESPACIAL DE LA COMPOSICIÓN TAXONÓMICA Y FUNCIONAL DE COMUNIDADES FITOPLANCTÓNICAS EN BAHÍA FILDES, ISLA REY JORGE, ANTÁRTICA (Spatial characterization of taxonomic and functional composition of phytoplanktonic communities in Fildes Bay, King George Island, Antarctica.)
194
Sanhueza, et al. COMPOSICIÓN DE LA COMUNIDAD BACTERIANA MARINA EN EL OCÉANO ÁRTICO (Marine bacterial community composition at the Arctic Ocean)
198
Garcia, et al. DINÁMICA ESPACIAL DEL MESOZOOPLANCTON Y SU RELACIÓN CON EL DERRETIMIENTO GLACIARIO EN CALETA POTTER, ISLA 25 DE MAYO (SHETLANDS DEL SUR, ANTÁRTIDA) (Spatial dynamics of mesozooplankton in Potter Cove, King George Island (South Shetlands, Antarctica) and its relation to glacial melting)
202
Valdivia, et al. SCALE-DEPENDENT SPATIAL VARIATION IN DIVERSITY AND STRUCTURE OF ROCKY ANTARCTIC COMMUNITIES (Escalas de variación espacial en diversidad y estructura de comunidades antárticas rocosas)
206
Lee, et al. LOS ENSAMBLES MEIOBENTONICOS INTERMAREALES DE LAS ISLAS SHETLAND DEL SUR, COSTA DANCO Y BAHÍA MARGARITA (The intertidal meiofaunal assemblages of the South Shetland Islands, Danco Coast and Margarita Bay)
210

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
x
Gutiérrez, DIETA DEL PEZ HARPAGIFER ANTARCTICUS NIBELYN, 1947 (NOTOTHENIOIDEI: HARPAGIFERIDAE), DE POZAS INTERMAREALES DE ISLA REY JORGE: POSIBLE PERSISTENCIA TEMPORAL EN UN OCÉANO AUSTRAL CALENTÁNDOSE (Diet of the fish Harpagifer antarcticus Nibelyn, 1947 (Notothenioidei: Harpagiferidae), from interdidal pools of King George Island: possible temporal persistence in a warming Southern Ocean)
214
Dias, et al. ESTRUCTURA DE LA COMUNIDAD BACTERIANA DE SUELOS ANTÁRTICOS CONTAMINADOS CON HIDROCARBUROS DURANTE LA BIOESTIMULACIÓN CON DIFERENTES NUTRIENTES Y UN SURFACTANTE (Bacterial community structure of an Antarctic hydrocarbon-contaminated soil during the biostimulation with different nutrients and a surfactant)
218
Vera, et al. EVIDENCIA DE ACTIVIDAD SURFACTANTE EN MICROORGANISMOS PSICRÓFILOS: UNA HERRAMIENTA BIOTECNOLÓGICA PARA LA BIORREMEDIACIÓN DE HIDROCARBUROS (Biosurfactant activity evidence in Psychrophiles Microorganisms: A Biotechnology Tool for Hydrocarbon Biorremediation)
222
Guerra, CAMBIO CLIMÁTICO O CONDICIONES AMBIENTALES EXTREMAS: ADAPTACIÓN DEL EPILITON EN LAS ISLAS GREENWICH Y REY JORGE, ISLAS SHETLAND DEL SUR, ANTÁRTICA (Climate change or extreme environmental conditions: epilithon adaptation in the Greenwich and King George Islands, South Shetland islands – Antarctica)
226
Verdugo, et al. DISTRIBUCIÓN DE COMPUESTOS TRAZA CON EFECTO CLIMÁTICO Y ACTIVIDAD MICROBIANA EN EL OCÉANO ÁRTICO DURANTE EL VERANO BOREAL, RESULTADOS PRELIMINARES DEL CRUCERO LOMROG III (Distribution of trace compound with climate effects and the microbial activity in the Arctic Ocean during boreal summer, preliminary results from LOMROG III Cruise)
229
Mark, et al. LA BÚSQUEDA DEL MEJOR LUGAR PARA OBSERVACIONES ASTRONÓMICAS EN EL ANTÁRTICA OESTE (The Search for the Best Astronomical Observations Site in Western Antarctica)
233
Manograsso, , et al. GEOQUÍMICA Y MINERALOGÍA DE LOS SEDIMENTOS DE PENÍNSULA Y CALETA POTTER, ISLA 25 DE MAYO (KGI): SUS IMPLICANCIAS EN LOS AMBIENTES SEDIMENTARIOS ACTUALES DE ANTÁRTIDA (Geochemistry and mineralogy of Potter Cove and Potter Peninsula sediments, 25 de Mayo (KGI) Island: their implications on current sedimentary environments of Antarctica)
237
Moura, et al.MINERALOGÍA DE LOS SUELOS DE LA PENINUSLA BYERS, ISLA LIVINGSTON (Mineralogy of soils peninsula Byers, Livingston Island)
241
Rondanelli-Reyes, et al. REGISTRO POLÍNICO DE MUESTRAS DE SUELO SUPERFICIAL EN ANTÁRTICA MARÍTIMA (Pollen record of superficial soil samples in the maritime Antarctic)
245
Arata, et al. DESARROLLO DE UN SISTEMA DE AREAS MARINAS PROTEGIDAS EN EL SECTOR OCCIDENTAL DE LA PENINSULA ANTARTICA Y ARCO DEL SCOTIA (Development of a system of Marine Protected Areas in the West Antarctic Peninsula and Scotia Arc)
249
Costa, SOOS- SOUTHERN OCEAN OBSERVING SYSTEM: MISSION AND OBJECTIVES (SOOS - Sistema de Observaciones del Océano Austral: misión y objetivos)
252
Montiel, et al. RESISTENCIA VERSUS ELASTICIDAD ECOLÓGICA DE LA COMUNIDAD BENTÓNICA ANTÁRTICA: UNA SIMULACIÓN EXPERIMENTAL EN LA PLATAFORMA CONTINENTAL DEL MAR DE WEDDELL (Ecological resistance versus resilience of the Antarctic benthic community: a simulation experiment in the Weddell Sea Continental Shelf)
254
Arriagada, et al. ANÁLISIS DEL COLAPSO Y FALTA DE RECUPERACIÓN EN DOS NOTOTÉNIDOS ANTÁRTICOS USANDO UNA PERSPECTIVA ECOSISTÉMICA (Analyzing the collapse and lack of recovery in two Antarctic nototheniid stocks using an ecosystem approach)
258
Rivadeneira, et al. DETERMINANTES HISTÓRICO-EVOLUTIVOS DEL ENANISMO EN BIVALVOS ANTÁRTICOS MODERNOS(Historic-evolutionary determinants of the dwarfism in modern Antarctic bivalves)
262
Nielsen, EL “ATAJO ANTÁRTICO” – UNA POSIBLE EXPLICACIÓN PARA EL INCREMENTO DEL INTERCAMBIO FAUNÍSTICO EN EL CENOZÓICO MEDIO ENTRE NUEVA ZELANDA Y SUDAMÉRICA (The Antarctic Shortcut – a possible explanation for increased mid-Cenozoic faunal exchange between New Zealand and South America)
266

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
xi
González-Wevar, et al. PATTERNS OF GENETIC DIVERSITY AND STRUCTURE, AS WELL AS DNA-BASED MODELLING ANALYSES SUPPORTS THE EXISTENCE OF GLACIAL REFUGIA IN SOUTH GEORGIA ISLAND FOR THE ANTARCTIC LIMPET DURING THE LAST GLACIAL MAXIMA (Patrones de diversidad y estructura genética, así como análisis de modelamiento basados en secuencias de DNA, apoyan la existencia de refugio glacial en Isla Georgia del Sur para la lapa Antártica durante el Último Máximo Glacial)
267
Poulin, et al. ESTRUCTURA GENÉTICA DEL ERIZO INCUBANTE ABATUS AGASSIZII EN LA BAHÍA FILDES, ISLA REY JORGE (Fine-scale genetic structure in the Antarctic brooding sea urchin Abatus agassizii in Fildes Bay, King George Islands)
271
Alcántara-Hernández, et al. CARACTERIZACIÓN Y COMPARACIÓN DE COMUNIDADES DESNITRICANTES EN TAPETES MICROBIANOS DE LA ISLA DEL REY JORGE (ANTÁRTIDA MARÍTIMA)(Characterization and comparison of denitrifying bacterial communities in microbial mats from King George Island, maritime Antarctica)
274
Arredondo, et al. DETECCIÓN MOLECULAR DE CRYPTOSPORIDIUM PARVUM EN PYGOSCELIS PAPUA EN BASE PRESIDENTE GABRIEL GONZÁLEZ VIDELA, ANTÁRTICA (64°49’S; 62°51’O) (Molecular Detection of Cryptosporidium parvum from Pygoscelis papua from Base Presidente Gabriel González Videla, Antártica (64°49’ S; 62°51’W))
278
Bruzzone, et al. EVALUACIÓN DE UN MÉTODO ANALÍTICO PARA SU IMPLEMENTACIÓN EN LA BÚSQUEDA DE MICROORGANISMOS ANTÁRTICOS PRODUCTORES DE ÁCIDOS GRASOS POLIINSATURADOS (Evaluation of an analytical method for use in detecting polyunsaturated fatty acids in Antarctic microorganisms)
282
Cabrera, et al. PYROCOCCUS: UN NUEVO MICROORGANISMO ANTÁRTICO CON ACTIVIDAD NITRILASA (Pyrococcus: A novel antarctic microorganism with nitrilase activity)
286
Cid, et al. AISLAMIENTO Y CARACTERIZACIÓN DE ACTINOBACTERIAS ANTÁRTICAS (Isolation and Characterization of Antarctic Actinobacterias)
288
Cornejo, et al. PURIFICACIÓN PARCIAL DE LA ACTIVIDAD TELURITO REDUCTASA DEL AISLADO ANTÁRTICO PSEUDOMONAS SP. BNF22(Partial purification of the tellurite reductase activity from the Antarctic isolate Pseudomonas sp. BNF22)
292
Cunachi, et al. DETECCIÓN DEL GÉNERO HELICOBACTER EN HECES Y TEJIDOS DE ANIMALES ANTÁRTICOS (Detection of Helicobacter genus in feces and tissues of Antarctic animals)
296
Dennett, et al. UNA NUEVA NITRILASA PROVENIENTE DE MICROORGANISMOS HIPERTERMOFILOS ANTARTICOS(Novel nitrilase from Antarctic hyperthermophilic microorganisms)
300
Ferrés, et al. BÚSQUEDA, IDENTIFICACIÓN Y CARACTERIZACIÓN DE DIVERSAS ACTIVIDADES ENZIMÁTICAS A PARTIR DE UNA COLECCIÓN DE BACTERIAS CULTIVABLES AISLADAS DE LA ANTÁRTIDA (Search, identification and characterization of enzymatic activities in a collection of culturable bacteria isolated from Antarctic samples)
303
Franzosi, et al. VARIABILIDAD INTERANUAL DE LA ABUNDANCIA DE PICOCIANOBACTERIAS EN EL PASAJE DRAKE E INMEDIACIONES DE LA PENINSULA ANTARTICA (Internannual variability of picocyanobacteria abundance in the Drake Passage and waters surrounding the Antarctic Peninsula)
306
Gallardo-Cerda, et al. BACTERIAS RIZOSFÉRICAS DE Deschampsia antárctica CON ACTIVIDAD ANTIBACTERIANA (Rhizosferic bacteria of Deschampsia antarctica with antibacterial activity)
309
González, et al. ESTUDIO DE LA BIOMASA DE MICROORGANISMOS FOTOSINTÉTICOS EN DIFERENTES AMBIENTES EN LAS ISLAS GREENWICH, DEE Y BARRIENTOS, ANTÁRTIDA (Study of photosynthetic microorganisms in different environments in Greenwich, Dee and Barrientos Islands, Antarctica)
312
Hernández, et al. ESTRUCTURA Y DINÁMICA ESTACIONAL DEL BACTERIOPLANCTON EN AGUAS ANTÁRTICAS CERCANAS A LA CALETA POTTER, ISLA 25 DE MAYO, ISLAS SHETLAND DEL SUR (Structure and seasonal dynamics of the bacterioplankton in Antarctic’s sea water near Potter cove, Isla 25 de Mayo, South Shetlands Islands)
316
Jorcín, et al. HONGOS FILAMENTOSOS DE LA ANTÁRTIDA Y SU POTENCIAL EN LA PRODUCCIÓN DE METABOLITOS BIOACTIVOS (Antarctic fungi and their potential in the production of bioactive metabolites)
320

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
xii
Lagos, et al. DIVERSIDAD DE LAS COMUNIDADES DE ACTINOBACTERIAS EN ECOSISTEMAS ANTÁRTICOS (Actinobacteria community diversity in Antarctic ecosystems)
323
Lavín, et al. ACTIVIDAD ANTIMICROBIANA DE COMPUESTOS PROTEICOS Y NO PROTEICOS PROVENIENTES DE DOS CEPAS DE STREPTOMYCES SP. AISLADA DESDE ANTÁRTICA (Antimicrobial activity of proteinaceous and nonproteinaceous compounds from two strains of Streptomyces sp. isolated from Antarctic)
326
Lavín, et al. CEPA ANTÁRTICA DE BACILLUS SP. CON ACTIVIDAD EXTRACELULAR DE TIPO AGAROLÍTICA Y ALGINATOLIASA (Antarctic strain of Bacillus sp. with extracellular agarolitic and alginate-lyase activities)
329
Leyton, et al. ACTIVIDAD INHIBITORIA DE LA BACTERIA ANTÁRTICA PSEUDOMONAS FRAGI EN LA FORMACIÓN DE BIOPELÍCULAS DE FLAVOBACTERIUM PSYCHROPHILUM (Inhibitory activity of the antarctic bacterium Pseudomonas fragi on biofilm formation of Flavobacterium psychrophilum)
333
Monserrate-Maggi, et al. IDENTIFICACIÓN Y BIOPROSPECCIÓN DE LEVADURAS ANTÁRTICAS (Identification and bioprospecting of antarctic yeasts)
335
Morales, et al. EVALUACIÓN DE LEVADURAS MARINAS PROVENIENTES DE LA ANTARTIDA COMO PRODUCTORAS DE ENZIMAS DE INTERÉS INDUSTRIAL Y ÁCIDOS GRASOS POLI-INSATURADOS (Evaluation of marine yeast from Antarctica as producers of enzymes and polyunsaturated fatty acids)
337
Pavlov, et al. BIOPROSPECCIÓN DE BACTERIAS ANTÁRTICAS PARA EL DESCUBRIMIENTO DE NUEVOS COMPUESTOS PEPTÍDICOS CON ACTIVIDAD ANTIMICROBIANA. (Bioprospection of Antarctic bacteria for finding new peptidic compounds with antimicrobial activity)
341
Pereira, et al. PURIFICATION OF AN ALKALINE PHOSPHATASE FROM AN ANTARCTIC BACTERIUM (Purificación de una fosfatasa alcalina desde una bacteria Antártica)
345
Pérez, et al. EL EFECTO DEL GUANO SOBRE LA FIJACIÓN BIOLÓGICA DEL NITRÓGENO EN LAS COSTRAS BIOLÓGICAS EN LA PENÍNSULA DE ARDLEY, ISLA REY JORGE. (The effect of guano on N fixation of biological soil crusts in Ardley Peninsula, King George Island)
348
Pozo, et al. RESPUESTA A DAÑO OXIDATIVO DE CEPAS HIPER RESISTENTES A TELURITO PROVENIENTES DEL TERRITORIO ANTÁRTICO CHILENO. (Response to oxidative damage of tellurite hiper-resistant strains from the Chilean Antarctic territory)
351
Rodríguez, et al. EFECTO DEL MERCURIO EN LA POTENCIACIÓN DE LA RESISTENCIA A TELURITO EN BACTERIAS AISLADAS DEL TERRITORIO ANTÁRTICO CHILENO (Effect of mercury on the potentiation of tellurite resistance in bacteria isolated from the Chilean Antarctic territory)
355
Soto, et al. CARACTERIZACIÓN METABÓLICA Y DE ESTRUCTURA DE MEMBRANA DE AISLADOS BACTERIANOS ANTÁRTICOS CAPACES DE SINTETIZAR QUANTUM DOTS (Metabolic and Membrane Structure Characterization of Antarctic Bacterial Strain Able to Synthesize Quantum Dots)
359
Torres, et al. EFECTO DE EXTRACTO DE LÍQUENES EN LA FORMACIÓN DE BIOPELÍCULAS DEL PATÓGENO DE PECES VIBRIO ANGUILLARUM (Lichens extract effect in the biofilms formation of fish pathogen Vibrio anguillarum)
363
Urtubia, et al. MICROFLORA BACTERIANA CULTIVABLE ASOCIADA AL TRACTO DIGESTIVO DEL BACALAO DE PROFUNDIDAD (Culturable microbiota along the digestive tract of Patagonian toothfish)
367
Vargas, et al. BIOPROSPECCIÓN DE MICROORGANISMOS ANTÁRTICOS Y SUS ENZIMAS EXTREMÓFILAS. (Bioprospecting of Antarctic microorganisms and their extremophilic enzymes)
371
Viteri, et al. BACTERIAS ANTARTICAS EXHIBEN ACTIVIDAD ANTIBACTERIANA CONTRA MICROORGANISMOS PATOGENOS (Antarctic bacteria exhibits antibacterial activity against pathogens)
374
Zúñiga, et al. ESPECIFICIDAD Y SELECTIVIDAD DE LOS MICOBIONTES DE LÍQUENES DEL GÉNERO Peltigera DE LAS ISLAS DE TIERRA DEL FUEGO, NAVARINO Y DECEPCIÓN (Mycobiont Specificity and Selectivity of Peltigera lichens from Tierra del Fuego, Navarino and Deception Islands)
377
Martinez Alvarez, et al. BUSQUEDA DE MICROORGANISMOS CON POTENCIAL BIOTECNOLOGICO A PARTIR DE SUELOS ANTARTICOS CONTAMINADOS CON HIDROCARBUROS (Looking for microorganisms with biotechnological potential from an Antarctic hydrocarbon-contaminated soil)
381
Martínez, et al. LEVADURAS OLEAGINOSAS ANTARTICAS COMO FUENTE DE TRIGLICERIDOS 385

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
xiii
PARA LA PRODUCCIÓN DE BIODISEL (Oleaginous yeasts as raw material for biodiesel production) Godoy, et al. ESTUDIOS PRELIMINARES SOBRE METEORIZACION BIOGENICA EN CRONOSECUENCIAS DE SUELOS EN PENINSULA FILDES, ANTARCTICA (Preliminary studies about the biogenic weathering of soil chronosequences in Fildes Península, Antarctica)
389
Rojas, et al. RELACIÓN FLEXIBILIDAD TÉRMICA DE ESTRUCTURA SECUNDARIA Y ACTIVIDAD ANTICONGELANTE DE HOMOPÉPTIDOS DE PROLINA, LISINA Y ARGININA (Relationship between thermal flexibility of secondary structure and antifreeze activity of proline, lysine and arginine homopeptides)
393
Alvarez, et al. HISTORIA DE LA ARQUITECTURA EN LA ANTÁRTIDA: UNA PROPUESTA DE CLASIFICACIÓN(Architectural History in Antarctica: a proposal for classification)
397
Alvarez, et al. NUEVOS EDIFICIOS EN ANTÁRTIDA: LA BUSQUEDA POR SISTEMAS CONSTRUCTIVOS MÁS EFICIENTES (New buildings in Antarctica: in search of building systems efficient)
401
Braga, et al. DUAS SITUAÇÕES CONTRASTANTES DE INÍCIO DE VERÃO NO ESTREITO DE BRANSFIELD (Two contrasting situations at the beginning of summer in the Bransfield Strait region)
405
Costa, et al. A ASSOCIAÇÃO DE PESQUISADORES POLARES EM INÍCIO DE CARREIRA NO BRASIL (APECS-BRASIL): AVANÇANDO E INOVANDO PELO PROGRESSO DA CIÊNCIA (The Association of Polar Early Career Scientists in Brazil (APECS-Brazil): advancing and innovating to the science progress)
408
Peña, et al. DISEÑO DE EDIFICACIONES CON ALTA EFICIENCIA ENERGÉTICA Y UTILIZACIÓN DE ENERGÍAS RENOVABLES PARA LA ESTACIÓN PEDRO VICENTE MALDONADO DEL INSTITUTO NACIONAL ANTÁRTICO ECUATORIANO (Design of buildings with high energy efficiency and utilization of renewable energy for the Pedro Vicente Maldonado Station from the National Ecuadorian Institute.)
410
Bastos, et al. EVIDENCIA DE LA PRESENÇA DE PAPUACEDRUS (CUPRESSACEAE, CALLITROIDEA) EN EL CRETÁCICO TARDIO DE LA PENÍNSULA ANTÁRTICA (Fossil evidence to the presence of Papuacedrus – Cupressaceae, Callitroidea - in the Upper Cretaceous of northern Antarctic Peninsula)
414
Bozo, et al. ESTUDIO DE LA MADERA DE Araucaria araucana (Molina) K. Koch, VARIABILIDAD Y COMPARACIÓN CON MADERAS FOSILES AFINES (wood study of araucaria araucana (molina) k. koch, variability and comparison with related fossil woods)
418
Manfroi, et al. ANÁLISIS DE CARBÓN VEGETAL FÓSIL EN DEPÓSITOS CRETÁCICOS DE LA PENÍNSULA ANTÁRTICA (Análise de carvão vegetal fóssil em depósitos cretáceos da península Antártica)
422
Mansilla-Vera, et al. LABORATORIO DE PALEOBIOLOGÍA DE ANTÁRTICA Y PATAGONIA,
INSTITUTO ANTÁRTICO CHILENO, PUNTA ARENAS, CHILE. (Antarctic and Patagonian Paleobiology
Laboratory at the Chilean Antarctic Institute, Punta Arenas, Chile)
425
Suwalsky, et al. EFECTOS DE EXTRACTOS ACUOSOS DEL LIQUEN U. aurantiaco Y MUSGO W. sarmentosa PROVENIENTES DE LA ANTARTICA SOBRE ERITROCITOS HUMANOS Y MODELOS MOLECULARES DE SUS MEMBRANAS (Effects of the aqueous extracts of the Antarctic species U. aurantiaco (lichen) and W. sarmentosa (moss) on human erythrocytes and molecular models of their membranes)
429
Arancibia, et al. EFECTO DEL GRADIENTE DE TEMPERATURA EN LA TASA FOTOSINTETICA DE SPHAGNUM Y PROYECCIONES COMO COLONIZADOR DE LA PENÍNSULA ANTÁRTICA E ISLAS (Effect of a temperature gradient on the photosynthesis rate of Sphagnum and its projections as a colonizer in the Antarctic Peninsula and islands)
432
Cuba-Díaz, et al. Colobanthus quitensis DE LA MARISMA, UNA NUEVA POBLACIÓN PARA LA COLECCIÓN GENÉTICA DE LA ESPECIE (Colobanthus quitensis from Marisma, a new population for specie’s genetic collection)
435
Fuentes, et al. CARACTERIZACIÓN MORFOLÓGICA Y FENOLOGÍA DE Juncus bufonius L CULTIVADO EN CONDICIONES CONTROLADAS (Morphological characterization and phenology of Juncus bufonius L. growing in controlled conditions)
439
Velásquez, et al. IDENTIFICACION Y ANALISIS DE EXPRESION DE G3PD, FtsH Y SOD, TRES GENES REGULADOS POR FRIO EN Deschampsia antarctica Desv. (Identification and expression analysis of G3PD, FtsH and SOD, three genes regulated by low temperature in Deschampsia antarctica Desv.)
443

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
xiv
Alfonso, et al. VARIACIÓN DEL ALBEDO ESPECTRAL DE LA NIEVE EN PUNTA FORT WILLIAMS, ISLA GREENWICH, ANTÁRTIDA MARINA (Variation of the snow spectral albedo in Fort William Point, Greenwich Island, marine Antartic)
447
E. Johnson, et al. THE POTENTIAL OF TERRASAR-X INTERFEROMETRY ON UNION GLACIER (El potencial de la interferometría Terrasar X en glaciar Unión)
451
Petsch, et al. ANÁLISIS DE MODELOS DIGITALES DE ELEVACIÓN PARA APLICACIÓN EN LA PENÍNSULA ANTÁRTICA (Analysis of digital elevation models for application in the Antarctic Peninsula)
452
Rodrigo, et al. PLUMAS DE SEDIMENTOS GLACIARES EN PEQUEÑAS BAHÍAS Y FIORDOS DE LA COSTA DE DANCO, PENÍNSULA ANTÁRTICA (Glacial sediment plumes in small bays and fjords of the Danco Coast, Antarctic Peninsula)
456
Di Fonzo, et al. QUÍMICA SANGUÍNEA DE DOS ESPECIES DE PINGÜINOS ANTÁRTICOS EN ÁREAS CON ALTO Y BAJO IMPACTO AMBIENTAL (Plasma and serum biochemistry from two different species of Antarctic penguins in areas with low and high environmental impact)
460
Lagos, et al. EFECTO DE LA RADIACIÓN ULTRAVIOLETA SOBRE LA RAZÓN RNA: DNA DE COPÉPODOS ANTÁRTICOS Y DE LA ZONA CENTRAL DE CHILE (Effects of the ultraviolet radiation on the RNA: DNA ratio of Antarctic and central Chile zone copepods)
464
Rovirosa, et al. METABOLITOS SECUNDARIOS BIO-ACTIVOS DE ORGANISMOS MARINOS DE LA ANTÁRTICA (Bio-active secondary metabolites from Antarctic marine organisms)
467
Seixas, et al. VIGILANCIA DE VIRUS INFLUENZA EN AVES MARINAS DE ELEPHANT ISLAND, ANTARTIDA, DE 2010 a 2012(Surveillance of Influenza virus in seabirds from Elephant Island, Antarctica, from 2010 to 2012)
470
Souza, et al. MENSURANDO OS NÍVEIS DE ESTRESSE EM PINGUIM-ANTÁRTICO (PYGOSCELIS ANTARCTICA): UMA COMPARAÇÃO ENTRE AS FASES DE REPRODUÇÃO E MUDA (Measuring stress levels in chinstrap penguins (Pygoscelis antarctica): a comparison between nesting and molting stages)
473
Sáez, et al. CAMBIOS MICROCLIMÁTICOS INDUCIDOS POR OTC Y SUS CONSECUENCIAS EN EL CRECIMIENTO DE PLANTAS ANTÁRTICAS (Microclimatic changes induced by OTC and their consequences on antarctic growth plants)
475
Sanhueza, et al. CAMBIOS MORFO-FISIOLÓGICOS EN POBLACIONES DE Colobanthus quitensis A LO LARGO DE UN GRADIENTE LATITUDINAL ANTARTICO-PATAGÓNICO (Morphophysiological changes in Colobanthus quitensis populations along an antarctic-patagonian latitudinal gradient)
479
Abdala, et al. EL PROGRAMA CIENTÍFICO Y AMBIENTAL ANTÁRTICO URUGUAYO (The Uruguayan Scientific and environmental program)
483
Calisto-Ulloa, et al.MONITOREO AMBIENTAL EN BASES ANTÁRTICAS: UNA HERRAMIENTA PARA LA TOMA DE DECISIONES Y LA MEJORA DEL DESEMPEÑO AMBIENTAL (Environmental monitoring in Antarctic Stations: a tool for decision-making and improvement environmental performance)
485
Chiang, et al. BIOACUMULACION DE CONTAMINANTES ORGANICOS PERSISTENTES EN PINGUINOS PYGOSCELIS: UNA MIRADA A LOS PROCESOS DE BIOMAGNIFICACION EN PENINSULA ANTARTICA (Bioaccumulation of Persistent Organic Pollutants in Pygoscelis penguins: an insight to biomagnification processes in the Antarctic Peninsula)
489
Espejo, et al. CONCENTRACIÓN DE METALES TRAZA EN EXCRETAS DE DOS ESPECIES DE PINGÜINOS EN DIFERENTES LOCALIDADES DE LA ANTÁRTICA (Concentration of trace metals in excrements of two species of penguins from different locations of the Antarctica)
492
Gran-Scheuch, et al. CARACTERIZACIÓN QUÍMICA Y BIOLÓGICA DE SUELOS CONTAMINADOS CON DIESEL PROVENIENTES DE LA ISLA REY JORGE, ANTÁRTICA (Chemical and biological characterization of diesel contaminated soils obtained from Isla Rey Jorge, Antártica)
496
Mattatal, et al. ESTUDIO INMUNO-HEMATOLÓGICO DE EFECTOS ATRIBUIBLES A CONTAMINANTES ORGÁNICOS PERSISTENTES EN PINGUINOS Pygoscelis DE LA PENÍNSULA ANTÁRTICA (Immunohaematological study on effects atributable to persistant organic pollutants in Pygoscelis penguins in Antarctic Peninsula)
500
Pereira, et al. MERCURY CONCENTRATION IN FEATHERS OF PHALACROCORAX ATRICEPS FROM KING GEORGE ISLAND, SOUTH SHETLAND ARCHIPELAGO, ANTARCTICA (Concentración de
503

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
xv
mercurio en plumas de Phalacrocorax atriceps de isla Rey Jorge, archipiélago Shetland del Sur, Antártica) Perfetti-Bolaño, et al. PLOMO EN SUELOS DE COLONIAS DE PINGÜINOS PYGOSCELIDOS EN LA PENINSULA ARDLEY, ANTARTICA (Lead in colonies of Pygoscelid penguins in Ardley Peninsula, Antarctica)
504
Silva, et al. FEATHERS AND SOIL AS INDICATORS OF ENVIRONMENTAL CONTAMINATION (Plumas y suelo como indicadores de contaminación ambiental)
508
Suarez, et al. PROPIEDADES FÍSICAS Y COMPOSICIÓN ELEMENTAL DE LOS AEROSOLES EN LA ESTACIÓN ANTÁRTICA PERUANA MACHU PICCHU (Physical properties and elemental composition of aerosols in Machu Picchu Peruvian Antarctic Station)
509
Aguayo-Lobo, LOS VARAMIENTOS DE LOS CETÁCEOS ¿SON UNA CAUSA DE MUERTE NATURAL? (The cetaceans strandings, Are they a natural a cause of death?)
510
Aldea, et al. DIVERSIDAD DE POLIPLACÓFOROS ANTÁRTICOS CON ESPECIAL ÉNFASIS EN NUEVOS REGISTROS DE LAS CAMPAÑAS BENTART (Diversity of Antarctic Polyplacophorans with special reference to new records from BENTART cruises)
514
Caiafa, et al. BIOGEOGRAFÍA DE LÍQUENES ANTÁRTICOS: UN ESTUDIO A PEQUEÑA ESCALA (Biogeography of Antarctic lichens: a small-scale study)
518
Correa, et al. BIOLOGÍAY ESTRUCTURA POBLACIONAL DEL KRILL (Euphausia superba) EN EL ESTRECHO DE BRANSFIELD DURANTE EL VERANO DEL 2013 (EXPEDICIÓN ANTAR XXI – PERÚ) (Biology and population structure of krill (Euphausia superba) in the Bransfield Strait during summer of 2013(ANTAR XXI Expedition - Perú)
522
Egas, et al. VARIACIONES DE LAS COMUNIDADES EUCARIONTES PICO, NANO Y MICROFITOPLANCTÓNICOS EN BAHIA FILDES, ISLA REY JORGE, ANTARTICA, DURANTE EL VERANO 2011 Y 2012 (Variations of pico-, nano- and micro- phytoplanktonic eukaryotic communities in Fildes Bay, King George Island, Antarctic, during summer 2011 and 2012)
525
Haro, et al. PRIMERA COMPARACIÓN ISOTÓPICA DE LAS ÁREAS DE ALIMENTACIÓN DE LA BALLENA JOROBADA (Megaptera novaeangliae) EN AGUAS SUBÁNTARTICAS DE SUDAMÉRICA Y PENÍNSULA ANTÁRTICA (First isotopic comparison of feeding grounds of humpback whale (Megaptera novaeangliae) from the South American subantarctic waters and Antarctic Peninsula)
527
Leiva, et al. ESPECIFICIDAD Y SELECTIVIDAD EN LÍQUENES TRIPARTITOS DEL GÉNERO Placopsis DESDE DOS SITIOS DE LAS ISLAS SHETLAND DEL SUR (Specificity and selectivity in tripartite lichens of the genus Placopsis from two sites in the South Shetland Islands)
531
Loza, et al. ESTIMACION DE LA EDAD ABSOLUTA EN FOCAS CANGREJERAS LOBODON CARCINOPHAGA MOMIFICADAS DE LA PENÍNSULA ANTÁRTICA. (Absolute age determination of mummified crabeater seals Lobodon carcinophaga from Antarctic Peninsula)
535
Márquez, et al. VALOR CALÓRICO DE LAS PRINCIPALES PRESAS DE UN PREDADOR GENERALISTA, LA FOCA LEOPARDO HYDRURGA LEPTONYX, Y LOS REQUERIMIENTOS ENERGÉTICOS DE SUS DIFERENTES GRUPOS ETARIOS (Caloric value of main prey of a generalist predator, the leopard seal Hydrurga leptonyx and the energetic requirements of their different age groups)
539
Olivares, et al. VARIACIÓN EN LA DISTRIBUCIÓN DE MEIOFAUNA INTERMAREAL, UNA COMPARACIÓN ENTRE LA ANTÁRTICA Y LA REGIÓN DE MAGALLANES (Variation in meiofaunal distribution in intertidal beaches, a comparison between Antarctica and the region of Magallanes)
543
Orgeira, SOBRE LA NECESIDAD DE ACTUALIZAR LAS DISTRIBUCIONES DE LAS AVES MARINAS PELÁGICAS EN OCÉANO ATLÁNTICO SUR Y ANTÁRTIDA (The need to update the seabird pelagic distribution in South Atlantic Ocean and Antarctica)
547
Ramírez, et al. LIQUENOBIOTA EN LA ESTACIÓN CIENTÍFICA ANTÁRTICA MACHU PICCHU (Lichen biota at the Antarctic research station Machu Picchu)
551
Silva, et al. DISTRIBUCIÓN Y DENSIDAD INCIDENTAL DE AVES MARINAS EN UN CRUCERO DEL ATLÁNTICO SUR OCÉANO ANTÁRTICO (Distribution and density of seabirds counting in a cruise from South Atlantic to Antarctic Ocean)
553
Díaz, et al. ANÁLISIS PRELIMINAR DE LA DIVERSIDAD GENETICA Y FILOGEOGRAFÍA DEL CARACOL ENDÉMICO NEOBUCCINUM EATONI EN EL OCÉANO AUSTRAL (Preliminary analysis of diversity genetics and phylogeography of the endemic snail Neobuccinum eatoni in the southern ocean)
554

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
xvi
González-Wevar, et al. PATRONES DE DIVERSIDAD GENÉTICA Y CONECTIVIDAD EN POBLACIONES NATURALES DEL Puye (Galaxias maculatus) EN PATAGONIA (Patterns of genetic Diversity and connectivity in natural populations of el Puye (Galaxias maculatus) in Patagonia)
558
Mura-Jornet, et al. NUEVOS MARCADORES NUCLEARES PARA PINGÜINOS PYGOSCELIS (New DNA nuclear markers for Pygoscelis penguins)
563
Cherene, et al. DISTRIBUCIÓN DE METALES, GRANULOMETRÍA Y COMPOSICIÓN ELEMENTAL E ISOTÓPICA EN SEDIMENTOS SUPERFICIALES DE LA BAHIA DE ALMIRANTADO Y DEL ESTRECHO DE BRANSFIELD - PENÍNSULA ANTÁRTICA (Distribution of metals, granulometry and isotopic and elementary composition in surface sediments of Admiralty Bay and Bransfield Strait - Antarctic Peninsula)
567
Pinedo, et al. DISTRIBUCION VERTICAL DEL ZOOPLANCTON ANTARTICO EN LOS ALREDEDORES DE LA ISLA ELEFANTE. VERANO AUSTRAL 2007. PERU ANTAR XVII (Vertical distribution of Antarctic zooplankton around Elephant Island. Austral summer 2007. Peru ANTAR XVII)
572
Salas-Berrios, et al. FEEDING HABITS AND DIET OVERLAP IN MARINE FISH LARVAE FROM THE PERIANTARCTIC MAGELLAN REGION (Hábitos alimentarios y sobreposición trófica en larvas de peces marinos de la región periantártica de Magallanes)
576
Salazar, et al. EVALUACIÓN HIDROACÚSTICA DEL KRILL (Euphausia superba) EN EL ESTRECHO DE BRANSFIELD Y ALREDEDORES DE LA ISLA GIBBS DURANTE LA XXI CAMPAÑA ANTÁRTICA PERUANA 2013 (Hidroacustic survey of krill (Euphausia superba) in Bransfield and Gibbs Island during XXI Peruvian Antartic Campaign 2013)
580
Sierra, et al. EFECTO DEL VIENTO Y LA MAREA EN LAS PROPIEDADES FISICO-QUIMICAS DE CALETA POTTER (Wind and tide effects on physical and chemical properties of Potter Cove)
584
Uribe, et al. COMPARACIÓN DE LA RESPUESTA FOTOSINTÉTICA A LA VARIACIÓN DE LA INTENSIDAD DE LUZ DE DIATOMEAS BENTÓNICAS ANTÁRTICAS DE LOS GÉNEROS NAVICULA Y NITZSCHIA (Comparison of the photosynthetic response to light intensity variation of benthic Antarctic diatoms from the genus Navicula and Nitzschia)
588
Cárdenas, et al. DESCRIPCION DEL MITOGENOMA EN Nacella concinna Y COMPARACION CON OTRAS NACELLAS SUBANTARTICAS. (The mitogenome of Nacella concinna and the comparison with others Subantartic Nacellids)
592

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
RESÚMENES EXTENDIDOS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
1
DISEÑO, CONSTRUCCIÓN Y OPERACIÓN DE UN SISTEMA DE TRATAMIENTO DE AGUAS RESIDUALES EN BAHÍA FILDES Y SU INFLUENCIA EN EL MEDIO AMBIENTE. (Design, construction and operation of a wastewater treatment system in Fildes Bay and its influence on
the environment)
Gómez-Fuentes, C.1, Calisto-Ulloa, N.1
1CIMAA, Centro de Investigación y Monitoreo Ambiental Antártico, Departamento de Ingeniería Química, Universidad de Magallanes, Punta Arenas, CHILE. E-mail: [email protected] INTRODUCCIÓN La mayoría de las bases antárticas están situadas en las áreas costeras libres de hielo, las cuales son especialmente sensibles a las perturbaciones humanas y representan alrededor de un 2% de la superficie total del continente (Gröndahl et al. 2009). Estas bases suelen contar con sistemas de tratamientos de aguas residuales con descargas al mar, de acuerdo a los requerimientos de Anexo III del Protocolo al Tratado Antártico sobre la protección del medio ambiente 1. La descarga de estos efluentes representan el único tipo de residuo que se permite liberar al medio ambiente antártico y constituye la única actividad que podría estar introduciendo intencionalmente microorganismos fecales en el medio (Aronson et al. 2011). Bahía Fildes, como puerta de entrada a la Antártica desde el continente americano, cuenta con una gran cantidad de instalaciones y es una de las zonas con alta presión antropogénica. En temporada estival, con el aumento de población científica, del personal logístico y los turistas, las exigencias sobre los sistemas de tratamiento de aguas residuales aumenta significativamente, lo cual puede traducirse en un impacto mayor en el medio costero (Gómez-Fuentes & Calisto-Ulloa 2013). En 2009 se decidió construir una planta de tratamiento de aguas residuales conjunta para la Base Profesor Julio Escudero y la Estación Marítima Bahía Fildes, aprovechando la cercanía de ambas bases y como una forma de optimizar los recursos disponibles, disminuyendo el número de instalaciones extra. Este trabajo presenta una visión general del diseño, las principales consideraciones teóricas y los resultados que se obtuvieron en los monitoreos ambientales en las cercanías del punto de descarga. ESTUDIO, DISEÑO Y CONSTRUCCIÓN DE UN SISTEMA DE TRATAMIENTO Previo a la construcción de la actual planta de tratamiento conjunta de la Estación Marítima Bahía Fildes y la Base Profesor Julio Escudero, cada una de las bases tenía su planta de tratamiento de aguas residuales compacta, con un diseño estándar de aireación prolongada. Las bases ejercían un pobre o nulo control de la calidad de sus efluentes tratados. Diagnósticos preliminares mostraban que estos sistemas tenían problemas tanto en la formación del lodo activado como en la decantación secundaria, necesaria para la clarificación final y posterior desinfección del efluente. El año 2009, INACH y la Armada de Chile, aprovechando que la Estación Marítima iniciaba la ampliación de su estructura, llegan al acuerdo de construir una nueva planta de tratamiento diseñada de acuerdo a las necesidades de ambas instalaciones, descartando el uso de plantas de tratamiento compactas, debido al pobre rendimiento obtenido con anterioridad. La construcción de un sistema de tratamiento conjunto presenta una serie de ventajas, tales como evitar los periodos de puesta en marcha, donde los rendimientos de depuración son mínimos, evitar la reintroducción de lodos al sistema de tratamiento cada inicio de temporada (necesidad previa de la base
1 Secretaria del Tratado Antártico (1998). Anexo III al Protocolo al Tratado Antártico Sobre Protección del Medio Ambiente Eliminación y Tratamiento de Residuos.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
2
Escudero, cuya ocupación es estacional), disminuir las necesidades de control y el costo asociado a mano de obra y analítica, obtención de economías de escala, tanto en la construcción como en la operación y, finalmente, un diseño que puede adaptarse a las limitaciones físicas de espacios disponibles y a las características de producción de agua residual de las bases. Para determinar los parámetros de diseño se sostuvieron reuniones con ambas instituciones, donde se consideraron planes de expansión, hábitos de trabajo y tecnologías de minimización del consumo de agua instaladas. Finalmente se llegó a los siguientes parámetros de diseño:
Parámetro Valor
Unidades Observaciones Base Escudero
Estación Marítima Bahía Fildes
Población 401 122 personas 1. Ocupación estacional 2. Ocupación continua
Consumos de aguas 1303 220 L hab-1 d-1 3. Considera medidas de ahorro de agua
Factor Punta 4,84 4. Considera la mezcla de ambas instalaciones
Concentración de agua residual 250 200 mg DBO5 L-1
60 60 mg NTK L-1 230 190 mg SS L-1
Tabla 1. Características del afluente para el diseño de la planta de tratamiento de aguas residuales. Table 1. Influent wastewater characteristics to design the wastewater treatment plant.
Para la evaluación de las tecnologías a implementar, se tomaron en consideración los costos económicos, los costos de mantenimiento, las condiciones de operación y los requerimientos de espacio. Finalmente, se seleccionó como tecnología de depuración un sistema convencional de aireación extendida, alternativa que se impuso a reactores de membrana, biodiscos y SBR (Sequencing Biological Contactors). Entre las ventajas de esta tecnología están tener un alto de grado de flexibilidad, operación relativamente simple, baja producción de lodos y mínimos requerimientos de entrenamiento del personal responsable. La línea de tratamiento propuesta se muestra en la figura 1. Como primer paso se incluye un estanque pulmón, para suavizar las puntas de caudal, las cuales en pequeñas poblaciones alcanzan valores que pueden comprometer el funcionamiento del sistema biológico (Metcalf 2003). Posteriormente se incluye un desarenador, balsas de aireación y decantadores secundarios. El sistema finaliza con la desinfección del efluente mediante luz UV, sistema que no deja residuos en el efluente tratado, a diferencia de la cloración, pero cuya efectividad está supeditada al buen funcionamiento de los tratamientos previos, ya que su eficiencia es muy sensible a la presencia de sólidos suspendidos. Para responder adecuadamente a los cambios de población, el sistema se diseñó en forma modular, contando con tres líneas de tratamiento, las cuales pueden actuar independientemente o como una sola unidad de tratamiento. Esta configuración además presenta la ventaja de que, en época invernal, las balsas de aireación no utilizadas pueden servir como digestor aerobio de lodos, facilitando el traslado de lodos al continente al inicio de la siguiente temporada. Finalmente, esta configuración ayuda a disminuir el problema del tiempo de respuesta frente a aumentos repentinos de población, ya que la línea activa actúa como planta de inoculación a las líneas que entran en operación, a la vez que éstas actúan como tanques de almacenamiento hasta alcanzar su máxima capacidad de operación. Otros ingenieros ya habían utilizado la configuración modular; por ejemplo, la planta de tratamiento de la Base McMurdo utiliza una lógica de diseño similar (Law et al. 2006).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
3
Figura 1. Esquema de la planta de tratamiento de aguas residuales.
Figure 1. Schematic flow of the designed wastewater treatment plant.
RESULTADO DE LOS MONITOREOS EN EL MEDIO AMBIENTE La figura 2 muestra los conteos de coliformes totales en la costa alrededor del punto de descarga (punto C en el gráfico). Los resultados muestran que, previo a la construcción de la nueva planta de tratamiento (año 2009), en la costa habían sobre 5000 UCF/100 ml de coliformes totales, con una influencia hasta los 50 metros de la descarga, donde el conteo disminuía hasta aproximadamente 1000 UFC/100 ml. Debe notarse que la legislación chilena permite, para los vertidos al mar, un máximo de 1000 UFC/100 ml de coliformes fecales. Estos resultados son concordantes con los publicados por otros autores que han detectado altas concentraciones de coliformes y microorganismos entéricos cerca de las descargas de aguas servidas (Bruni et al. 1997; Delille & Gleizon 2003; Hughes 2003).
Figura 2. Muestreos de coliformes totales en la costa alrededor del punto de vertido.
Figure 2. Total coliform samples on the coast around the discharge point. El efecto en el medio ambiente de la instalación de la nueva planta de tratamiento, medido el 2010, mostró una inmediata reducción de las coliformes a valores cercanos a cero en las cercanías de la descarga. Este efecto se mantiene el año 2011. La causa de esta reducción no sólo se debe a un mejor control el sistema de tratamiento, sino también al cambio en el sistema de disposición de los efluentes tratados, los cuales ya no son incorporados directamente al mar, sino que son infiltrados en la costa, lo que entrega un grado adicional de depuración. Efectos similares de la instalación de una nueva planta de tratamiento han sido observados por otros autores (Hughes 2004). El año 2012 sólo un punto mostró un conteo alto, mientras que en el año 2013, se vuelve a observar un impacto negativo en la costa. Respecto de las causas para estos resultados, se pueden identificar varios factores: aumento de las poblaciones en las bases por sobre los límites de diseño; falta de un mantenimiento adecuado de las instalaciones y falta de capacitación del personal. Por último, la posible saturación de la zona de vertido o las modificaciones físicas de la playa pueden haber limitado el efecto filtrante del medio. En la actualidad, la planta se encuentra operativa, en la temporada 2013-2014 se realizarán actividades de mantención e incorporación de biomasa fijada a soporte, con el objetivo de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
4
responder a los aumentos de población, sin la necesidad de cambiar el sistema completo; adicionalmente, se mantendrá y mejorará el sistema de monitoreo ambiental y de control del sistema. CONCLUSIONES El diseño de plantas para bases antárticas requiere de la consideración de factores únicos dados por las características extremas de las condiciones de operación: lejanía, dificultad de control del sistema, muy pequeñas poblaciones, aumentos repentinos de usuarios, etc. Los resultados parecen indicar que un factor a tener en cuenta es la forma de verter el efluente en el medio, ya que sistemas de infiltración parecen proveer de mayor seguridad en la operación. No obstante los resultados del año 2013, el modelo de asociatividad presentado parece ser la mejor forma de enfrentar el tratamiento de residuos líquidos, lo que debería favorecer la respuesta de las instituciones frente a los resultados de los monitoreos ambientales con el objetivo de mantener un alto nivel de desempeño ambiental. AGRADECIMIENTOS Al INACH por apoyo logístico. Proyectos de INACH: T_09-08 y MA_01-12 (CIMAA) BIBLIOGRAFÍA Aronson R. B., Thatje S., McClintock J. B. and Hughes K. A. 2011. Anthropogenic impacts on marine ecosystems in Antarctica. Ann N Y Acad Sci 1223(1): 82-107. Bruni V., Maugeri T. L. and Monticelli L. 1997. Faecal pollution indicators in the Terra Nova Bay (Ross Sea, Antarctica). Marine Pollution Bulletin 34(11): 908-12. Delille D. and Gleizon F. 2003. Distribution of enteric bacteria in Antarctic seawater surrounding the Port-aux-Francais permanent station (Kerguelen Island). Mar Pollut Bull 46(9): 1179-83. Gómez-Fuentes C. and Calisto-Ulloa N. 2013. Quantification of marine pollution near Antarctic Station. In: XI SCAR Biology Symposium, Barcelona, España. Gröndahl F., Sidenmark J. and Thomsen A. 2009. Survey of waste water disposal practices at Antarctic research stations. Polar Research 28(2): 298-306. Hughes K. A. 2003. Influence of seasonal environmental variables on the distribution of presumptive fecal Coliforms around an Antarctic research station. Appl Environ Microbiol 69(8): 4884-91. Hughes K. A. 2004. Reducing sewage pollution in the Antarctic marine environment using a sewage treatment plant. Mar Pollut Bull 49(9-10), 850-3. Law K. P., Kulchawik R. J., Zenz D. R. and Bouchard T. B. 2006. Overview of the Design, Construction, and Operation of the McMurdo Wastewater Treatment Facility in Antarctica. Proceedings of the Water Environment Federation 2006(12): 1439-48. Metcalf E. 2003. Inc., Wastewater Engineering, Treatment and Reuse. New York: McGraw-Hill.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
5
CARACTERIZACION AMBIENTAL PARA EL USO SOSTENIDO DEL AGUA POTABLE EN BASE ESPERANZA (NORTE DE LA PENINSULA ANTARTICA ARGENTINA)
(Environmental characterization for sustainable use of water at Hope Bay- Antarctic Peninsula North – Argentina)
Chiarandini, J.P.1, Marcomini, S.2 y Santillana, S.1, 2
1Instituto Antártico Argentino, Balcarce 290, C1010AAZ, Buenos Aires, Argentina. E-mail: [email protected] 2 Depto. de Geología, Fac. de Cs. Exactas y Naturales, U. de Buenos Aires, Ciudad Universitaria, Buenos Aires, Argentina. RESUMEN El retroceso de los flujos de hielo (ice stream) ha dado origen a lagunas proglaciarias que son empleadas para abastecimiento de agua potable, como es el caso de lo que ocurre en base Esperanza. Estas lagunas, presentan una evolución geomorfológica característica que les imparte características geológicas, físico-químicas y biológicas muy variables en periodos cortos de tiempo (decenios). Esta variabilidad hace que las fuentes de abastecimiento de agua para las bases tengan características efímeras y en consecuencia produzcan problemas para la toma de agua potable para la población. El presente trabajo tiene como objeto caracterizar y monitorear la evolución geomorfológica, química y ambiental de estas lagunas con el fin de evaluar su durabilidad para ser empleadas como fuente de abastecimiento de agua potable permanente para la base Esperanza. METODOLOGÍA Y ÁREA DE ESTUDIO Se realizó una caracterización geomorfológica del área de estudio considerando imágenes satelitales de los años 2001, 2005, 2006 y 2012 correspondientes al Google Earth. Los trabajos de campo y toma de muestras fueron efectuados durante los meses de enero y febrero durante la campaña de verano-2013. Se tomaron muestras de agua de la actual toma de agua potable, de los tanques de almacenamiento, del efluente cloacal crudo y del vertido del mismo. Se midieron parámetros físico- químicos in situ (temperatura, conductividad, pH, turbiedad, salinidad y oxígeno disuelto) a través de una ecosonda multiparametrica HORIBA. Las muestras destinadas para análisis cualitativo de fitoplancton fueron tomadas de la orilla de la laguna y colocadas en botellas de plástico de 500 mL de capacidad y fijadas con solución de formol al 4% para la observación directa al microscopio óptico. La zona de estudio corresponde a Base Esperanza (63°23’ S, 56°59’ O) localizada en el extremo nor-oriental de la Península Antártica. Se encuentra rodeada por los glaciares Buenos Aires, Pirámide, Flora y Kenney. Las temperaturas medias anuales oscilan entre los -5 y 3,3°C, siendo las medias del invierno entre -8 y -12°C y las del verano de 1 a -2,5 °C. Las precipitaciones medias anuales no superan los 620 mm. Los vientos dominantes son del sudoeste, alcanzando los 200 km/h. Las condiciones ambientales son típicas zonas frías con permafrost rico en hielo (Ermolin 2005). En la Península Tabarin, afloran rocas ígneas del arco magmático de la península Antártica y estratos mesozoicos de la cuenca de Larsen (Del Valle et al. 2001). En el área se reconocen el basamento metasedimentario asignado a la Formación Hope Bay, (Grupo Trinity Península) del Permo-Triásico, los sedimentos continentales de laFm. Mount Flora, (Gr. BotanyBay) del Jurásico Inferior y rocas volcánicas silícicas subaéreas de la Fm. Kenney Glacier del Jurásico Medio (Gr. Volcánico AntarcticPeninsula) (Montes et al., 2004). El afloramiento más extendido en el área de estudio corresponde a la Formación Bahía Hope que forma una secuencia bien estratificada básicamente monoclinal con inclinación hacia el O-NO.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
6
RESULTADOS Las lagunas proglaciares se forman por endicamiento de agua de ablación al frente de las lenguas glaciarias en retroceso. Por lo general estas lagunas están indicadas por morenas terminales o bedrockbump que, luego que la lengua glaciaria retrocede, tienden a romperse generando fenómenos de flujo al frente de los mismos. Por este motivo constituyen morfologías temporales y fuentes de abastecimiento de aguas superficiales temporales, ya que tenderán en su evolución geomorfológica a desaparecer por ruptura o erosión de las morenas terminales o filtración del talik. La antigua explotación de agua para abastecimiento de la Base Esperanza se realizaba de la laguna Boeckella. Esta laguna se aloja sobre una depresión cuya base se encuentra sobre till correspondiente a morenas basales y está bordeada por morenas terminales y laterales correspondientes al retroceso del frente del glaciar Pirámide. El cuerpo de agua presenta un modelo térmico típico de los glaciares antárticos fríos, regulado por las bajas temperaturas de la zona y por el ingreso del agua proveniente de la ablación de los glaciares circundantes. El nivel del agua del lago aumenta durante el verano en relación a los periodos de ablación del hielo. En épocas de deshielo se pueden distinguir dos horizontes térmicos, el superior hasta los 2,5 m y uno más profundo con temperaturas de fondo de 2,3 a 3,5 C.(Ermolín 2005). La profundidad máxima de la laguna es de 5,8 m, la cuenca tiene una superficie de 0,3 km2, mientras que el lago ocupa 0,07 km2, el volumen de agua es de 1,4x105 m3 y el cambio estival del nivel de 0,4 a 0,5 m (Pose e Izaguirre 1997). En el año 2001 se rompe la morena frontal que contiene al lago y comienza a disminuir el nivel del agua. Por tal motivo, se construye un dique de hormigón que recuperó temporalmente el nivel del lago, y disminuyó los procesos de filtración. Entre Febrero y Marzo de año 2002 se construye un albardón artificial con áridos provenientes de una terraza fluvioglacial, de 32 m de largo, 3 m de ancho, y 1,5 a 2 m de altura sobre el nivel medio del lago. Sobre dicho albardónse colocaron 14 sifones de convección líquida a aproximadamente 2 m de distancia cada uno, endicados a una profundidad de 4 m, con el objetivo de elevar el techo del permafrost para impermeabilizar con el hielo los áridos rellenados en el sector del acuífero freático (Ermolín 2005). Luego de los 2 primeros años se observa una recuperación, con formación del núcleo impermeable del dique bajo el efecto de la cementación del hielo en el terraplén (Ermolín 2005). Posteriormente, las aguas superficiales de ablación vuelven a cortar el dique de tierra con la consecuente pérdida del agua de la laguna. Por ese motivo se deja de utilizar la laguna Boeckella para la captación de agua (año 2007) y se traslada la captación a una laguna ubicada hacia el sur, la cual es también una laguna proglaciaria con características semejantes. Los parámetros físico químicos del agua obtenidos para esta laguna son semejantes a los reconocidos para la laguna Boeckella en el año 1999 (Izaguirre et al., 2003) por ser ambas lagunas formadas por retroceso del glaciar Pirámide y sedimentos semejantes. En la tabla 1 se comparan los valores con la laguna de captación actual.
Boeckella (1999) Captación actual (2013) Temperatura © 0- 4 0.2- 0.8 pH 6.5-6.79 6.20- 6.44 Conductividad (µS/cm-1) 0.025- 0.067 0.020- 0.038
Oxígeno disuelto (mg/L-1) 7.7-12.4 mg/L-1 10.84- 12.20 Turbiedad (NTU) ------ 50-55
Tabla 1. Características físico-química de la laguna Boeckella y la laguna actual. Table 1.Physico-chemistry characterization of the Boeckella Lagoon and the actually lagoon
En Base Esperanza las aguas residuales se canalizan a través de un entubamiento de aproximadamente 4 m de longitud y se acumulan en dos tanques domiciliarios de capacidad de 700 L cada uno. En uno ellos se colectan los efluentes cloacales (baño), y en el otro el resto de los efluentes domiciliarios (aguas de lavado y cocina). Estos líquidos son bombeados a la Planta de Tratamiento de la base, una vez que se

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
7
realiza cloacas en cada establecimiento los días lunes, miércoles y viernes. Luego de 15 minutos de bombeo en cada establecimiento, todos los efluentes se colectan en una pileta que se encuentra ubicada en la parte inferior de la planta de tratamiento y a partir de allí son bombeados, por medio de un ducto de desagote, directamente a la caleta Cabaña. En la tabla 2 se presentan las características físico-químicas que se midieron in situ, correspondientes al efluente cloacal crudo (directamente en el caño de descarga), en el área de descarga (antes y después del vertido). Esta zona comprende una distancia de aproximadamente 50 m donde está ubicado el vertido del efluente cloacal propiamente dicho. El líquido cloacal es fresco y de color gris característico, olor propio de la materia fecal, debido a la descomposición de algunos productos o residuos alimenticios, con altos valores de turbiedad debido al conteniendo de materia en suspensión y con déficit permanente de oxígeno disuelto generando un estrés aeróbico a los organismos presentes. Los valores presentados en la tabla 2 son característicos de un efluente cloacal crudo. En referencia a la descarga, antes y después del vertido, los parámetros medidos no registraron cambios significativos, excepto la turbiedad que aumentó después de la descarga debido al vuelco del efluente que tiene alta carga de material en suspensión, pero a medida que transcurre el tiempo, se produce el efecto de dilución, característico de un vertido al mar. Otro parámetro que se vio levemente modificado, fue la salinidad. Antes del vertido tiene un valor propio al agua de mar, y después del vertido se observa una disminución que resulta propia debido al vuelco de agua dulce a la zona costera. Por el caudal del efluente cloacal descargado en la zona, el cambio registrado resulta ser insignificante.
Efluente cloacal crudo Descarga(antes del vertido) Descarga(después del vertido) Temperatura © 8.3- 8.7 1.4- 3.8 1.5- 3.9 pH 7.18- 8.12 7.95- 8.04 7.44- 8.04 Conductividad (mS/cm-1) 1.71- 2.78 54.4- 54.7 47.3- 50.2 Oxígeno disuelto (mg/L-1) 3.50- 3.80 15.2- 15.9 12.34- 15.2 Turbiedad (NTU) 590- 935 0 61-100 Salinidad (‰) 0.08- 0.13 3.34- 3.35 2.93- 3.34
Tabla 2. Características físico-química del efluente cloacal crudo y la descarga. Table 2.Physico-chemisty characterization of the sewage effluent and discharge.
Los valores que se presentan en la tabla 3 corresponden a algunos puntos claves que fueron estudiados en la base. Como se puede observar, se encontró una diferencia significativa en cuanto al pH de la casa 12 en relación a los otros puntos. Esto es debido a que el tanque de almacenamiento de agua para consumo de esa casa fue el único al que se le realizó una limpieza al comienzo del periodo de hibernación. Distintos procesos químicos y biológicos se llevan a cabo en el nicho de los tanques de agua que dan lugar a las variaciones de pH generando medios alcalinos o ácidos vinculados a distintas reacciones que realizan los microorganismos, principalmente los aerobios debido a que los valores de OD encontrados son aptos para el desarrollo de este tipo de organismo.
Tabla 3. Característica
s físic
o-química de algunos puntos claves en la red de distribución del agua potable en Base Esperanza. Table 3.Physico-chemistry characterization of some key points in the distribution network of drinking water in Hope Bay.
Según la reglamentación del ANMAT el pH puede variar de 6.5 a 8.5 para el agua potable, mientras que en el agua mineralizada la variación permitida es de 4.5 a 9.00 (2007). El pH obtenido para el agua de
Captación actual Tanque Casa 4 Tanque Casa 12 Tanque Enfermería Tanque Casino
Temperatura © 0.2- 0.8 12-14 12- 14 12- 14 14- 16 pH 6.20- 6.44 4.3- 4.5 6.2- 6.4 4.3- 4.6 4.3- 4.5 Conductividad (mS/cm-1) 0.020- 0.038 0.03- 0.06 0.01- 0.03 0.01- 0.03 0.03- 0.06 Oxígeno disuelto (mg/L-1) 10.84- 12.20 16.0- 16.7 16.2- 16.7 16.2- 16.7 16.0- 16.7 Turbiedad (NTU) 50-55 S/D S/D S/D S/D

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
8
captación de la laguna está entre 6.22 a 6.44; mientras que el pH medido del agua de los tanques de almacenamiento fue variable dependiendo de si el mismo había tenido limpieza o no. CONCLUSIONES La evolución geomorfológica de las lagunas de captación de agua son fuentes efímeras por tratarse de lagunas proglaciarias endicadas por morenas frontales y no pueden considerarse como fuentes permanentes de abastecimiento de agua potable para la Base Esperanza. Por tal motivo se recomienda reforzar periódicamente el sector frontal de la laguna con albardones artificiales, en aquellos sectores que puedan sufrir cortes por canalizaciones fluvioglaciarias para mantener el nivel de la laguna. El material de relleno puede provenir de till o terrazas fluvioglaciares adyacentes y es recomendable que contenga una matriz arcillosa. Los barros de decantación sedimentados en el fondo de los tanques han favorecido el proceso de acidificación del agua, razón por la cual se recomienza realizar el vaciado y limpieza de los tanques anualmente. Luego del análisis del vertido del efluente cloacal crudo se recomienda mejorar y/o controlar el tratamiento en la planta correspondiente. AGRADECIMIENTOS Al IAA, al Lic. Horacio Tassone y Lic. Gabriel Blanco. A la Base Esperanza, a la Tte. 1° Vanesa Pía y a los Dres. Pablo Nykotyszac y Pablo Dankiewicz por su colaboración y guía en el trabajo de campo. REFERENCIAS BIBLIOGRÁFICAS Cook, A., Fox, JA., Vaughan, DG & JG. Ferrigno. 2005. Retreating Glacier Fronts on the Antarctic Peninsula over the Past Half-Century.Science, 308 (5721) 541-544. Shepherd, A., Wingham, D., Payne, T., Skvarca, P. 2003. Larsen Ice Shelf Has Progressively Thinned. Science 302, 856. Warren, C. and M. Aniya. 1999. The caving glaciers of southern South America. GlobalPlanet. Change, 22, 59–77. Warren, C., Benn, CD., V. Winchester and S. Harrison. 2001. Buoyancy-driven lacustrine calving, Glacier Nef, Chilean Patagonia. J. Glaciol., 47(156), 135–146. Warren, C.R., D.R. Greene and N.F. Glasser. 1995. Glaciar Upsala, Patagonia:rapid calving retreat in fresh water. Ann. Glaciol., 21, 311-316. Vaughan, D.G., Marshall, G.J., Connelly, W.M., King, J.C., and Mulvaney, R., 2001. Devil in the detail. Science, 2001. 293(5536): p. 1777-1779. Vaughan, D.G., and C.S.M. Doake. 1996. Recent atmospheric warming and retreat of ice shelves on the Antarctic Peninsula. Nature 379: 328-331. Ermolín, E.. 2005. Dique en permafrost antártico con termosifones de convección líquida (lago Boeckella, bahía Esperanza). ASAGAI, 21, 1-26, Buenos Aires. Paciullo, F., Ribeiro, A., Andreis, R.R., and Trouw, R.A.J. 2002. Facies associations in the Permian-Triassic Hope Bay Formation, Antarctic Peninsula. Antarctica at the close of a millennium. Royal Soc. New Zealand Bull., 35:175–183. Rott, H., Stuefer, M. and A. Siegel. 1998. Mass fluxes and dynamics of Moreno Glacier, Southern Patagonia Icefield. Geophys. Res. Lett., 25(9), 1407–1410. Montes, M., Martín-Serrano, A. y R. del Valle. 2004. Mapa geológico de la costa Austral de Bahía Esperanza y el Monte Flora, Península Antártica. En prensa. Pose, M. e I. Izaguirre. 1997. Sucesión estival de las poblaciones fitiplantónicas en el lago Boeckella (Bahía Esperanza, Antártida). Contribución del Instituto Antártico Argentino 449, 27. Skvarca, P., Raup, B. and H. de Angelis. 2003. Recent behaviour of GlaciarUpsala, a fast-flowing calving glacier in LagoArgentino, southern Patagonia. Ann. Glaciol., 36,184–188. Skvarca, P., De Angelis, H. Naruse, R., Warren, CR.and M. Aniya.2002. Calving rates infresh water: new data from southern Patagonia. Ann. Glaciol., 34, 379–384.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
9
CONSTRUCCIÓN DEL MAPA DE CORROSIÓN ATMOSFÉRICA DE CHILE: ESTACION ANTARTICA
(Map construction of atmospheric corrosion of Chile: Antarctic station)
Mónica Puentes1, Rosa Vera1, Raquel Araya1, Paula Rojas2, Ana María Carvajal3 1Instituto de Química, Facultad de Ciencias, Pontificia Universidad Católica de Valparaíso, Av. Universidad 330, Placilla, Valparaíso, Chile. [email protected] 2Escuela de Ingeniería Mecánica, Facultad de Ingeniería, Pontificia Universidad Católica de Valparaíso, Av. Los Carrera 01567, Quilpué, Chile. 3Escuela de Construcción Civil, Pontificia Universidad Católica de Chile, Avda. Libertador Bernardo O’Higgins 340, Santiago, Chile. RESUMEN La corrosión atmosférica es un problema de gran importancia para diversos ámbitos de la vida cotidiana, y en especial para la industria, la minería y la construcción. Es por ello que se hace necesario realizar un mapa de corrosión atmosférica de Chile como una herramienta para uso público. En la Antártica Chilena, al poseer bases militares, es de gran interés tener datos de cómo la corrosión afecta a los metales y aleaciones, para poder luego realizar una mejor elección de éste o un mejor recubrimiento. Para realizar los ensayos se colocaron muestras de cobre, acero al carbono, aluminio y acero galvanizado, de 10 x 10 x 0,4 cm en la Base Julio Escudero la que se encuentra ubicada en Lat. 62ºS y Long. 58ºW, clasificada de acuerdo a normas ISO 9223 a 9226 como, S2 y P0, cuyo contenido de iones cloruros y dióxido de azufre es de 94,12 mg.m-2d-1 y 9.28 mg.m-2d-1, respectivamente. Por otra parte, las variables meteorológicas de humedad relativa, temperatura, cantidad de agua caída, dirección y velocidad de los vientos se midieron mensualmente. Las diferentes muestras metálicas se evalúan por medidas de pérdida de masa cada 3 meses por un período de 3 años. Para la identificación de los productos de corrosión se utiliza la técnica de difracción de rayos X, y en la caracterización morfológica de éstos se emplea microscopía electrónica de barrido (MEB). La determinación de Cloruro solubles encontrados sobre la superficie metálica de cada material, se realizó mediante el método de Volhard. Los resultados preliminares muestran que la velocidad de corrosión alcanza una clasificación C4 para el caso del cobre, acero al carbono y aluminio y C2 para el acero galvanizado. Los productos de corrosión que presenta el cobre a los 24 meses de exposición son principalmente de color verde (paratacamita) y para el acero al carbono de color rojizo de características poco adherentes y presencia de exfoliación. En general la cara posterior de las muestras se encuentra con un mayor grado de corrosión ya que la cara expuesta al medio ha permanecido gran parte del período de exposición cubierta con nieve, especialmente para aluminio. INTRODUCCION La corrosión atmosférica comprende el ataque a los metales expuestos al aire y es el tipo de corrosión más común en los metales [1-2]. Es un tema de preocupación mundial debido a su importancia para la vida útil de los equipos y la durabilidad de los materiales estructurales. Por lo mismo es importante tener datos de corrosividad para distintas zonas de Chile, ya que se puede hacer una adecuada selección del material a utilizar o se puede mejorar el recubrimiento o la protección que requiera el metal seleccionado y es por esto que se decidió realizar un mapa de corrosión de Chile en donde existe una gran variedad de climas, entre ellos el polar en la Antártica en donde se encuentran bases militares y diversas construcciones por lo que es importante conocer la corrosividad de un lugar con características meteorológicas tan distintas [3-5].

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
10
PROCEDIMIENTO EXPERIMENTAL El material utilizado fueron placas de cobre y aluminio, ambas al 99,9% de pureza, acero al carbono y acero galvanizado con 98,5% de zinc, 1% de hierro y 0,5% de aluminio. Las dimensiones de las muestras fueron de 100 mm x 100 mm x 4mm las cuales fueron desengrasadas en acetona y secadas con aire frío a presión. Luego las muestras fueron instaladas en bastidores con un ángulo de inclinación de 45º con respecto a la horizontal. La estación atmosférica se ubica en la Base Julio Escudero (figura 1), la que se encuentra ubicada en Lat. 62ºS y Long. 58ºW, a 100 metros del mar y 10 metros sobre el nivel del mar. La evaluación mensual de los parámetros climáticos y ambientales y el uso de las normas ISO 9223 permiten clasificar la agresividad de las atmósferas de las estaciones [6]. Según la norma ISO 9223, que clasifica la agresividad de las atmósferas considerando el tiempo de humidificación (τ o TDH) y la deposición de los contaminantes en el ambiente (salinidad, S y compuestos sulfurados, P), a la estación Antártica le correspondería una clasificación de τ5, S2, P0, propio de un ambiente marino con un índice de agresividad de corrosión C3 ó C4. El tiempo de humidificación (τ o TDH) se determina en base al número de horas que la muestra se encuentra expuesta a una Humedad Relativa (HR) igual o superior a 80 % y a una temperatura del aire igual o superior a 0ºC. La velocidad de corrosión se determinó cada 3 meses mediante pérdida de masa, según norma ASTM G1-03 [7], y las variables ambientales, fueron evaluadas mensualmente empleando norma ISO 9223.
Figura 1. Estación de corrosión en Base Julio Escudero.
Figure 1. Base Julio Escudero corrosion station.
RESULTADOS Y DISCUSIÓN De acuerdo a los resultados obtenidos, la estación Antártica se clasifica de acuerdo a normas ISO 9223 como, S2 y P0, cuyo contenido de iones cloruro y dióxido de azufre es de 94,12 mg.m-2d-1 y 9,28 mg.m-
2d-1, respectivamente y el valor del tiempo de humidificación es 64,5% que corresponde a τ5 clasificándose la estación C5 lo cual es muy agresiva. La Tabla 1 presenta la velocidad de corrosión promedio de cada metal después de un año de exposición y la clasificación según la norma ISO 9223. Se aprecia que, para el acero al carbono, cobre y aluminio, la estación Antártica es agresiva. Para el caso del acero galvanizado la clasificación es baja. Para el

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
11
acero al carbono y el cobre la clasificación de velocidad por pérdida de masa es mayor y esto se debe a los productos de corrosión formados sobre las superficies de los metales. En el caso del acero al carbono, al ser los productos de corrosión menos adherentes y de un mayor espesor, se pierde más masa mientras que, en el caso del cobre, aunque el producto de corrosión es adherente y protector, y de acuerdo a la difracción de rayos X realizada a los productos de corrosión, se tiene que el porcentaje de cobre que hay luego de 1 año de exposición es menor al 90%. En el aluminio la velocidad de corrosión es mayor ya que éste metal sufre corrosión generalizada por la parte expuesta a la atmósfera y corrosión por picadura por la parte posterior de la muestra en estudio, éste tipo de corrosión es una de las más agresivas. La clasificación de la velocidad de corrosión a los 2 años de exposición no cambia a excepción del acero al carbono que baja a C3 dado que tiene un espesor mayor de producto de corrosión que actúa en el tiempo como barrera al medio expuesto.
Metal V. corrosión (µm/año) Clasificación Acero al carbono 66,5 C4
Cobre 2,3 C4 Acero galvanizado 0,62 C2
Aluminio 3,7 C4 Tabla 1: Clasificación de velocidad de corrosión según norma ISO 9223.
Table 1: Corrosion rate classification according to ISO 9223.
CONCLUSIONES En la estación Antártica se obtuvo un alto índice de corrosividad dado que los niveles de cloruro son altos por la localización de la estación de estudio que se encuentra a 100 metros del mar, sumándose el tiempo de humidificación que es ligeramente alto lo que favorece una rápida corrosión del material. Los productos de corrosión formados para todos los materiales contienen en su composición cloruro y difieren en su espesor, adherencia y en el grado de protección. AGRADECIMIENTOS INNOVA CORFO, INACH y Dirección de Investigación de la PUCV. REFERENCIAS Syed, S. 2012. Atmospheric corrosion of materials; Emirates Journal for Engineering Research, 11 (1), 1-24. Melchers, R.E. 2008. A new interpretation of the corrosion loss processes for weathering steels in marine atmospheres. Corrosion Science, 50: 3446-3454. Vera, R., Delgado, D., Araya, D., Puentes, M., Rojas, P., Guerrero, I., Cabrera, G., Erazo, S. y A.M. Carvajal. 2012. Construcción de mapas de corrosión atmosférica de Chile. Resultados preliminares. Revista Latinoamericana de Metalurgia y Materiales, 32(2): 269-276. Morcillo, M., Chico, B., de la Fuente, D., Almeida, E., Joseph, G., Rivero, S. & B. Rosales. 2004. Atmospheric corrosion of reference metals in Antarctic sites, Cold Region Science and Technology 40:165-178. Vera, R., Puentes, M., Araya, R., Rojas, P., y A.M. Carvajal. 2012. Mapa de corrosión atmosférica de Chile. Resultados después de 1 año de exposición. Revista de la Construcción, 12(22), 61-72, 2012. Revista ISI. ISO 9223, Corrosion of metals and alloys, Classification of corrosivity of atmospheres, ISO, Geneva, 1991. ASTM G1-03, Standard Practice for preparing, cleaning and evaluating corrosion test specimens.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
12
TENDENCIAS DE LA INVESTIGACIÓN ECOLOGÍCA ANTÁRTICA DURANTE EL ÚLTIMO SIGLO
(Trends in Antarctic ecological research over the last century)
Acuña-Rodríguez IS1,2, Carrasco-Urra F3, Stotz GC4, Salgado-Luarte C1, Ríos RS1, Gianoli E1,3 y Molina-Montenegro MA2
1Departamento de Biología, Universidad de La Serena, La Serena, Chile. [email protected] 2Centro de Estudios Avanzados en Zonas Áridas (CEAZA), Facultad de Ciencias del Mar, Universidad Católica del Norte,
Coquimbo, Chile. 3Departamento de Botánica, Universidad de Concepción, Concepción, Chile. 4Biological Sciences Department, University of Alberta, Edmonton, Canada. INTRODUCCIÓN La investigación Antártica es por lo general muy costosa y demandante de recursos, pero entre las diversas temáticas, en éste continente la investigación ecológica debería ser prioritaria; tanto por su relevancia para el mantenimiento de las dinámicas de los ecosistemas Antárticos, así como para su correcto monitoreo en el marco de los cambio globales actualmente experimentados (Aronson et al., 2011). Las condiciones geofísicas Antárticas posicionan al continente en el extremo de varios gradientes ambientales globales, dando forma a ecosistemas tan únicos como desconocidos (Chown & Convey, 2007; Wall et al., 2011). Bajo éstas condiciones, los efectos del cambio global se están observando con mayor intensidad en este continente y sus ecosistemas (Bergstrom et al., 2006; Aronson et al., 2011); por lo tanto, un profundo conocimiento de los mismos es imperativo para hacer frente al desafío que esta tendencia nos presenta. Es en este sentido donde la ecología Antártica se constituye en un área de investigación prioritaria, y no solamente para sus propios ecosistemas dado que la actual investigación de las respuestas ecológicas en la Antártica , nos brindarán las claves para entender los cambios globales en curso (Aronson et al., 2011). Pero, ¿cuál es el alcance del actual conocimiento sobre la ecología Antártica?, ¿cómo se ha enfocado dicha investigación históricamente? y ¿quiénes están contribuyendo con este conocimiento? METODOLOGIA Hemos abordado estas preguntas mediante una amplia revisión bibliográfica y un análisis temático y socioeconómico general de todos los documentos relacionados con la ecología Antártica publicados durante más de un siglo (base de datos ISI-Web, 1904-2010). Se indagó por los artículos que contaran con alguna de las palabras clave: antartic, antarctica, antartica o antartida; encontrando 3184 artículos en las categorías “Ecology”, “Biodiversity & Conservation” y “Evolutionary Biology”. Se incluyeron estas categorías ya que consideramos que agrupan el núcleo teórico de la ecología. Adicionalmente se verificó que el reporte de un documento en la búsqueda no fuera consecuencia de un nombre científico o una zona subantártica. Cada artículo se clasificó según el reino de la especie estudiada, el nivel de organización biológica analizado, el año de publicación y el origen del documento, definido este último por las naciones representadas en las afiliaciones de los autores. RESULTADOS Encontramos grandes diferencias en las temáticas de la investigación ecológica Antártica, siendo la más evidente el dominio del reino animal, el cual es de lejos el reino más estudiado (fig. 1a). Como era esperado para un ecosistema con una reciente historia de investigación, la mayoría de sus publicaciones se vinculan con el nivel biótico del organismo, siendo estas principalmente descripciones taxonómicas.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
13
Adicionalmente, un claro déficit de publicaciones se observa a nivel poblacional y ecosistémico, incluso en Animalia (Figs. 1bc).
Figura 1. Tendencia histórica de las publicaciones sobre ecología Antártica por; reino (a), y nivel de organización biológica
(b). Aporte relativo de cada nivel biótico al total de publicaciones en cada reino (c). Figure 1. Historical trend of publications on Antarctic ecology by; kingdom (a), and level of biological organization (b).
Relative contribution of each to total biotic level publications in each realm (c).
Históricamente las publicaciones se han originado en países desarrollados (Tabla 1), sin embargo, los países emergentes están aumentando su participación en los últimos tiempos (Fig. 2). Esto cobra particular importancia en el marco general de las publicaciones ISI sobre ecología, donde la fracción de aquellas asociadas al continente Antártico se ha mantenido constante durante los últimos 20 años (Fig. 3).
Tabla 1. Participa
ción regional en las
publicaciones sobre
ecología Antártica 1904-2010. Pub.Cont. (publicación continua) hace referencia al año desde el cual la región publica al menos un
artículo por año. La tasa de publicación anual (Pub. Ann.) se calculó como la media geométrica de artículos por año. Table 1. Regional participation in the 1904-2010 Antarctic ecology publications. Pub.Cont. (continuous publication) refers to
the year from which the region published at least one article per year. The annual rate (Pub. Ann.) Was calculated as the geometric mean of items per year.
Región Países Publicaciones Pub. Cont. Pub. Ann. (±SD) Europa 25 1869 (51.6%) 1975 21.8 (± 5.1) America del Norte 2 680 (18.8%) 1980 7.1 (± 4.3) Oceania 3 512 (14.1%) 1986 9.4 (± 3.2) Latinoamérica 10 265 (7.3%) 1990 7.5 (± 2.7) Asia 13 230 (6.3%) 1988 5.4 (± 2.8) Africa 2 70 (1.9%) 2005 2.5 (± 2.2)
a
c
b

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
14
Figura 2. Tendencia histórica relativa de las publicaciones antárticas agrupadas por naciones según su estatus
socioeconómico, con base en el índice de desarrollo humano HDI: desarrolladas: HDI >0.8; emergentes: 0.5 < HDI < 0.8 y no-desarrolladas: HDI< 0.5 (UNDP, 2011). La línea continua es la tendencia para el periodo 1991-2011 (20 años); la línea
punteada marca el periodo 2001-2011 (10 años). Coeficientes de regresión significativos (±SD): países desarrollados1990-2010 = -0.47 ±0.12, r2
(adj) = 0.42, p=<0.001; países emergentes1990-2010 = 0.53 ±0.09, r2(adj) = 0.59, p=<0.001; países desarrollados2000-
2010 = -1.16 ±0.24, r2(adj) = 0.68, p=<0.001; países emergentes2000-2010 = 0.92 ±0.20, r2
(adj) = 0.66, p=0.001). Para los países no-desarrollados no se obtuvieron coeficientes significativos.
Figure 2. Historical trend of publications on Antarctic grouped by nations according to their socioeconomic status, based on the Human Development Index HDI: developed: HDI> 0.8; emerging: 0.5 <HDI <0.8 and non-developed: HDI <0.5 (UNDP, 2011 ). The solid line is the trend for the period 1991-2011 (20 years), the dotted line marks the period 2001-2011 (10 years). Significant regression coefficients (± SD): developed countries1990-2010 = -0.47 ± 0.12, r2 (adj) = 0.42, p = <0.001; emerging
countries1990-2010= 0.53 ± 0.09, r2 (adj) = 0.59, p = <0.001; developed countries 2000-2010 = -1.16 ± 0.24, r2 (adj) = 0.68, p = <0.001; emerging countries 2000-2010= 0.92 ± 0.20, r2 (adj) = 0.66, p = 0.001). For non-developed countries did not obtain
significant coefficients.
Figura 3: Proporción anual de publicaciones ecológicas ISI dedicadas a la Antártica (1991-2011). El set de publicaciones para cada año se obtuvo del total de artículos para el año correspondiente reportados en las categorías “Ecology”, “Biodiversity &
Conservation” y “Evolutionary Biology” de la ISI-Web of Science. Figure 3: Annual proportion of ISI ecological publications dedicated to Antarctica (1991-2011). The set of publications for
each year was obtained from the total for the year reported in the "Ecology", "Biodiversity & Conservation" and "Evolutionary Biology" subject of the ISI-Web of Science.
DISCUSIÓN Y CONCUSIONES Un claro desbalance se puede observar en la tendencia taxonómica de las investigaciones ecológicas en este continente; muy poca investigación se ha realizado (de manera comparativa) fuera del reino animal, y aunque algunos reinos presenten una baja riqueza de especies en la Antártica (Bergstrom et al., 2006), sus representantes se encuentran experimentando los mismos extremos ecológicos de esta zona del

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
15
mundo, por lo cual son per se de gran interés para la ciencia. De igual manera, los estudios poblacionales y ecosistémicos son en general más escasos, aún en el reino animal. Es llamativo esté déficit ya que son estos niveles donde se observa con mayor eficacia los cambios en procesos ecológicos vinculados con la distribución de los individuos en el espacio (dinámicas poblacionales, demografía, migraciones, etc.) y sus respuestas a procesos regionales (ciclos de nutrientes y cambios en el paisaje). Las naciones desarrolladas han generado históricamente la mayor parte de las publicaciones sobre ecología Antártica, pero llama la atención como su participación relativa ha disminuido durante los últimos años (fig. 2). Y si bien es una reducción leve (aunque significativa), es muy importante tanto en el contexto general de las publicaciones ecológicas ISI, donde la proporción de artículos de ecología Antártica se ha mantenido constante durante los últimos años (fig. 3); como desde la urgente necesidad de construir el conocimiento ecológico mínimo suficiente para abarcar la complejidad de un continente poco conocido, que cambia de manera acelerada. Es acá donde el aumento relativo de la participación de los países emergentes como fuente de investigación ecológica Antártica cobra gran relevancia. En este grupo destacan por Latinoamérica: Argentina, Brasil y Chile; quienes en conjunto son responsables por más del 95% de los artículos de la región (Stotz, et al., 2013) Existe una compleja, pero necesaria tarea de comparar el esfuerzo de investigación entre los países presentes en el continente Antártico, debido a que éste es un territorio bajo administración multinacional. El nivel de injerencia de las naciones en las decisiones que sobre éste continente se toman, depende en parte de los niveles de investigación sustancial que cada nación lleva a cabo (Conference on Antarctica, 1959; art. IX[2]). Entre la diversa investigación potencial en este territorio, las temáticas ecológicas deben ser prioridad debido a la urgente necesidad de este conocimiento, tanto para monitorear el acelerado cambio de estos ecosistemas ante los procesos de calentamiento global, como para dilucidar las mejores estrategias para la futura administración y conservación del continente Antártico. Como en otros aspectos de la sociedad, la integración regional (de información y logística), será determinante para el posicionamiento de Latinoamérica dentro de la investigación ecológica en la Antártica en el futuro próximo. AGRADECIMIENTOS Proyecto INACH G-22-11. BIBLIOGRAFÍA Aronson RB, Thatje S, McClintock JB y Hughes KA. 2011. Anthropogenic impacts on marine ecosystems in Antarctica. Annals of the New York Academy of Sciences. 1223: 82–107. Bergstrom DM, Convey P y Huiskes AHL (eds.). 2006. Trends in Antarctic terrestrial and limnetic ecosystems: Antarctica as a global indicator. Springer, Dordrecht. 360p. Conference on Antárctica. 1959. The Antarctic Treaty. Washington DC. Chown SL y Convey P. 2007. Spatial and temporal variability across life’s hierarchies in the terrestrial Antarctic. Philosophical Transactions of the Royal Society of London B. 362: 2307–2331. Stotz GC, Salgado-Luarte C, Rios RS, Acuña-Rodríguez IS, Carrasco-Urra F, Molina-Montenegro MA y Gianoli E. 2013. Trends in Antarctic ecological research in Latin American countries. Polar Research. En prensa. United Nations Development Programme [UNDP]. 2011. Human Development Report 2011, Sustainability and Equity: A Better Future for All. UNDP. Wall DH, Lyons WB, Chown SL, Convey P, Howard-Williams C, Quesada A y Vincent WF. 2011. Long-term ecosystem networks to record change: an international imperative. Antarctic Science. 23: 209–209.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
16
LOS LÍMITES Y LAS LIMITACIONES DE LA CIENCIA: CONSIDERACIONES SOBRE LA AUTONOMIA DE LOS INVESTIGADORES BRASILEROS EN ANTÁRTICA.
(The limits and limitations of science: autonomy considerations of Brazilian researchers in Antarctica)
Lemmertz, H.1, Almeida, M. L.1 1Programa de Pós Graduação em Sociologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, Rio Grande do Sul, Brasil. [email protected] [email protected] O presente trabalho visa compreender as limitações e os limites da produção científica brasileira atual a partir da consideração de um caso específico, qual seja, o das pesquisas realizadas na Antártica no contexto do Programa Antártico Brasileiro – PROANTAR. Para buscar compreender esta questão, o foco será analisar os diversos procedimentos e instâncias fomentadores ou limitadores da produção científica realizada por pesquisadores envolvidos nesse processo de produção de conhecimento no continente antártico, bem como a própria opinião de alguns desses pesquisadores em relação ao programa como um todo e à sua própria condição em particular. Mais especificamente, a proposta deste trabalho consiste em tentar conhecer e analisar os agentes que influenciam na produção científica antártica. O objetivo geral deste trabalho é mapear o grau de autonomia do PROANTAR a partir da identificação dos atores e esferas de valor que estão por trás de sua implementação (determinações sociais) e como os “portadores dos valores da ciência”, ou seja, os cientistas posicionam-se diante do perfil das pesquisas incentivadas pelo programa. Como objetivos específicos são propostas as seguintes questões: conhecer os atores que interagem na pesquisa antártica brasileira, compreender as esferas que influenciam na determinação das pesquisas realizadas através do PROANTAR, compreender a visão dos pesquisadores em relação a sua condição profissional e de agente de produção de conhecimento e verificar a posição brasileira em relação à produção de conhecimento em cooperação internacional; também é objetivo deste trabalho a apresentação das transformações que o PROANTAR vem sofrendo ao longo de sua existência. No decorrer de mais de duas décadas, o Programa desenvolveu-se em termos de amplitude de inserção antártica e em seus temas de pesquisa, abrangendo temas bastante diversos, como atualmente as pesquisas na área de ciências humanas. Precisamente em virtude desse processo de progressiva intensificação das atividades lá realizadas, é necessário compreender os procedimentos, atores e interesses que estão por trás da proposição dessas linhas de pesquisa incentivadas pelo PROANTAR, o que permite inclusive compreender e analisar qual a autonomia real dos pesquisadores na realização das pesquisas. Uma das questões de fundo a ser respondida por esta pesquisa é quais são os interesses dos pesquisadores que estão inseridos no Programa Antártico Brasileiro. Em outros termos, queremos encontrar elementos que nos permitam identificar se há uma coerência entre o interesse científico manifesto oficialmente no programa e os interesses dos diversos agentes que estão envolvidos em sua realização, seja enquanto fomentadores (que determinam as linhas de pesquisa, a verba disponibilizada, etc.), seja enquanto executores (aqueles envolvidos diretamente nas expedições, tais como os pesquisadores e mesmo os militares). A realização desta investigação tem por objetivo compreender como se dá a relação entre ciência e demais esferas de valor, e, dada a singularidade do Programa Antártico Brasileiro, faz-se adequado utilizá-lo como objeto capaz de elucidar esta questão. A busca por objetos de estudos cada vez mais complexos exige que os cientistas se desloquem para realizar suas pesquisas de campo. É necessário compreender os fatores externos à pesquisa que influem em seu processo de realização e

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
17
em seus resultados. É importante também verificar como os distintos atores envolvidos estão relacionados, a partir do discurso formado pelas vivências em seu contexto de investigação. Esta pesquisa se insere no contexto de uma discussão que está dentre as mais centrais do mundo contemporâneo, e que diz respeito ao significado e às condições de possibilidade de autonomia da ciência. Para realizar esta investigação buscaremos amparo na teoria produzida por autores vinculados à Sociologia do Conhecimento, os quais analisam em sua grande maioria a questão das influências que afetam a produção de conhecimento. O tema perpassa a produção cientifica de vários autores que tratam da questão do conhecimento, e é consenso que a produção de conhecimento é sempre influenciada, em maior ou menor grau, por fatores sociais. Max Weber trata da produção de conhecimento, assumindo que existem influências que agem sobre as esferas de valor, sendo a ciência uma destas esferas que opera segundo uma lógica própria, assim como as demais esferas: a política, a econômica, a estética e a erótica (Weber 1979). Weber não considerava que esta autonomia seria objetivizada, visto que admitia que influências externas às esferas agissem sobre estas, essencialmente porque quem produz o conhecimento são os homens e, por isso, a ciência estaria atrelada ao sistema de valores deste indivíduo que a constrói (Weber 1992). Ainda que se assuma a existência da influência do sistema de valores do pesquisador, isso não eliminaria a autonomia do campo cientifico. Segundo a teoria weberiana sobre a ciência na modernidade, a autonomia da produção científica é colocada em xeque quando a influência exercida sobre a ciência advém de outras esferas de valor, pois aí são lógicas diferentes que estarão sendo sobrepostas, interferindo na autonomia da própria esfera. Ou seja, se é inevitável que cada cientista imprima seu próprio leque de convicções no momento em que escolhe que conjunto de questões espera desvendar, algo muito diferente se passa quando se trata da interferência de esferas de valor que são regidas por diferentes princípios. Aliás, Weber não apenas faz o diagnóstico da autonomização das esferas de valor como uma característica intrínseca – e constitutiva – da modernidade (Weber 1979) como ainda alertou para os problemas que poderiam resultar de um situação em que a lógica de uma esfera fosse transposta a uma outra, como ele exemplifica nos dois ensaios em que preconiza a importância de manter política e ciência como duas vocações distintas (Weber 2004) Além de Weber diversos outros autores têm trabalhado a relação entre interferências mútuas entre sistemas valorativos distintos e os impactos disso para a produção do conhecimento, como Karl Mannheim, Gerard Fourez, Edgar Morin e, mais recentemente, Pierre Bourdieu. Todos esses autores contribuíram para construir o quadro teórico que operou como pano de fundo para a presente investigação, apresentando visões complementares que permitiram delinear com maior precisão as implicações sociais e sociológicas do modo como opera o PROANTAR. Embora influenciado por Weber em certos aspectos, Karl Mannheim (Mannheim, 1952) aprofunda da questão da produção de conhecimento e os interesses que influenciam sobre este processo, desde a escolha de determinadas temáticas a serem pesquisadas até a influência que a produção de um novo conhecimento pode gerar sobre um conhecimento existente. Ainda em relação a essa mesma temática, Gerard Fourez (Fourez, 1995) analisa a diferenças de interesses que a própria comunidade científica pode apresentar, de acordo com suas temáticas de pesquisa e formação acadêmica, apesar de possuírem um objetivo em comum, o de produzir conhecimento. Além disso, o autor pontua a existência da influência de grupos associados a cientistas, sejam eles dos mais diversos segmentos e interesses, que muitas vezes subsidiarão a produção científica. Em uma perspectiva um pouco diferente, Edgar Morin (Morin, 1989) corrobora a afirmação de que o conhecimento não é puro, mas sim permeado por lógicas externas. O autor considera que há uma instrumentalização da ciência, uma vez que esta fornece subsídios para transformar a sociedade e, por outro lado, a sociedade influencia na produção científica, sendo a ciência um meio de acessar espaços

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
18
regidos pela mesma lógica, como é o caso da Antártida, em que é somente através da produção de conhecimento que os países justificam suas bases no continente antártico. Para finalizar o quadro dos principais autores que nos fornecem um aporte teórico, temos Pierre Bourdieu (Bourdieu, 2004) que parece buscar inspiração no conceito de esferas de valor de Weber para formular seu conceito de campo, que será definido por Bourdieu como “microcosmos relativamente autônomos”. Abordaremos o conceito de campo científico, o qual possui uma lógica própria de produção, mas que sofre influências advindas de outros campos. Este trabalho esteve pautado por um contínuo entrelaçamento entre teoria e realidade empírica e para emergir nesta realidade foram realizadas entrevistas semi¬estruturadas com pesquisadores que pertencem ao PROANTAR e que já estiveram na Antártida. Além das entrevistas, foram analisados documentos referentes ao Programa, assim como, material divulgado pela mídia. Foi feito também análise da observação realizada em congressos científicos voltados para o tema antártico. É possível afirmar, do ponto de vista dos pesquisadores entrevistados, que a ciência realizada no âmbito do Programa Antártico Brasileiro é produzida pelos cientistas de maneira autônoma. No decorrer das três décadas de PROANTAR, não só aumentou a autonomia dos cientistas, como também houve uma ampliação das linhas de pesquisa contempladas pelo Programa, que inicialmente estava direcionada basicamente às investigações relativas à biodiversidade do continente e que hoje contemplam linhas de pesquisa mais abrangentes, que incluem geologia, geoquímica, estudos climáticos e atmosféricos e de impacto ambiental no continente antártico e Oceano Austral, além de aspectos tecnológicos, culturais e sócio-econômicos, incluídos no Programa recentemente. A hipótese da qual partimos inicialmente, alicerçada sobre a teoria weberiana da separação das esferas de valor, era a de que a produção científica gerada dentro do Programa Antártico Brasileiro fosse fortemente influenciada por outras esferas, como, por exemplo, a esfera política e econômica o que, segundo acreditávamos inicialmente, seria um fator limitador à plena autonomia da esfera científica. No decorrer da pesquisa, com a análise das entrevistas e documentos, tendo sempre como fio condutor a teoria weberiana, bem como os outros referenciais teóricos discutidos no primeiro capítulo, chegou-se a conclusão de que autonomia da esfera científica frente às demais esferas é maior do que se supunha inicialmente, e, mais do que isso, é possível diagnosticar um processo de progressiva autonomização. Evidentemente, não se trata de afirmar a existência de uma autonomia absoluta, na medida em que no contexto do PROANTAR há diferentes esferas coexistentes, isto é, que não existem de forma isolada, mas sim, que há uma constante influência de esferas entre si mesmas. Da mesma forma que Max Weber os demais autores que constituíram o embasamento teórico desta investigação corroboram para a afirmação de que a ciência produzida é autônoma, na medida em que segue uma lógica própria de produção, mas que não está imune à influência de determinações advindas de outras esferas. No caso do PROANTAR, são determinações oriundas das instituições que gerenciam o Programa, sendo elas de cunho financeiro e logístico, ou seja, determinações econômicas e, ainda, determinações que partem dos militares, grupo que trabalha junto aos pesquisadores antárticos, tanto na formulação e gerenciamento do Programa, quanto no apoio às expedições à Antártica. AGRADECIMIENTOS Agradezco a los investigadores del PROANTAR que me ayudaron en esa investigación. REFERENCIAS Azambuja, P. 1980. Antártida: história e geopolítica. Porto Alegre: Corag. Bourdieu, P. 2004. Os usos sociais da ciência: por uma sociologia clínica do campo científico. São Paulo: Editora UNESP. Coelho, A.P. 1983. Nos Confins dos Três Mares... a Antártida. Rio de Janeiro: Biblioteca do Exército. Colacrai, M. 2004. El Ártico y la Antártida em las relaciones internacionales. Porto Alegre: Editora da UFRGS.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
19
Ferreira, F., Rodrigues G. 2009. O sistema do tratado da Antártica: evolução do regime e seu impacto na política externa brasileira. Brasília: Fundação Alexandre de Gusmão. Figueira, P. de A. 2005.Nascimento da ciência moderna: Descartes. Campo Grande: Editora UNIDERP. Fourez, G.A. 1995.construção das ciências : introdução à filosofia e à ética das ciências. São Paulo : Ed. da UNESP. Freitas, M.E. de. 2012. Lições organizacionais vindas da Antártica. P. 915-937. Rio de Janeiro: RAP - Revista de Administração Pública, vol. 46, n° 4. Gandra, R., Madruga. 2009. O Brasil e a Antártida: ciência e geopolítica., P. 65-74. Belo Horizonte: Revista Geografias, julho-dezembro. Mannheim, K. 1952. Ideologia e Utopia: introdução à sociologia do conhecimento. Porto Alegre: Globo. Mannheim, K. 1952. O Problema de uma Sociologia do Conhecimento. In: Bertelli, Palmeira & Velho. Sociologia do Conhecimento. Rio de Janeiro: Zahar. Mannheim, K. 1974. Sociologia da Cultura. São Paulo, Perspectiva. Morin, E. 1996. Ciência com Consciência. Rio de Janeiro: Editora Bertrand Brasil. Morin, E. 1989.Para uma Sociologia do Conhecimento. Lisboa: Revista Sociologia, Problemas e Práticas, n° 6, p. 135 à 146. Mucchielli, L. 1998. La Découverte du Social: naissance de la sociologie en France (1870 – 1914). Paris: La Decouverte. Vieira, F Brum. 2006. O Tratado da Antártica: Perspectivas Territorialista e Internacionalista, p. 49-82. São Paulo: Cadernos PROLAM/USP, ano 5, vol. 2, Weber M. 1992. A “Objetividade” do conhecimento nas Ciências Sociais. In: Metodologia das ciências sociais. P. 107-154. Parte I. São Paulo: Cortez. Weber M. 2004. Ciência e política: duas vocações. São Paulo: Martin Claret. Weber M. 1979. “Rejeições Religiosas do Mundo e suas Direções” In: Ensaios de Sociologia. Rio de Janeiro, Zahar. Whimster, S. 2009. Weber. Porto Alegre: Artmed.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
20
ELUCIDANDO OS MECANISMOS MOLECULARES DE ADAPTAÇÃO DA BACTÉRIA EXIGUOBACTERIUM ANTARCTICUM
(Dilucidando los mecanismos moleculares de adaptación de la bactéria Exiguobacterium antarcticum)
Pellizari, VH3 e Baraúna, RA1; Carneiro, AR1; Dall’Agnol, HPM2; Nóbrega, F3; Ramos, RTJ1; de Sá, PG1; Nunes, CIP2; Carepo, MSP2; Azevedo, V4; Schneider, MPC1; Silva, A1
1Laboratório de Polimorfismo de DNA, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, Brasil. [email protected] 2REQUIMTE/CQFB, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Lisboa, Portugal. 3Laboratório de Ecologia de Micro-organismos, Instituto de Oceanografia, Universidade de São Paulo, São Paulo, Brasil. 4Laboratório de Genética Celular e Molecular, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belo Horizonte, Brasil. RESUMO Exiguobacterium antarcticum B7 é um extremófilo capaz de crescer em temperaturas que variam de -3°C à 42°C. Esta bactéria foi isolada a partir de um biofilme formado no sedimento do Lago Ginger, Ilha Rei George, Península Antártica. O genoma de E. antarcticum B7 foi obtido através de seqüenciamento com a plataforma SOLiD bem como o transcriptoma pela técnica de RNA-seq a fim de caracterizar a expressão gênica da bactéria a 0°C e 37°C em conjunto com as técnicas proteômicas 2D-DIGE e espectrometria de massas. Os resultados obtidos apresentaram o perfil molecular de adaptação da E. antarcticum B7 (2.815.863 pb) ao frio e demonstram a alta sintenia gênica com a bactéria psicrofílica E. sibiricum 255-15. No transcriptoma foram identificados 51 sRNA, 4 genes falso-positivos que foram deletados do genoma e 5 anotados por não terem sido preditos na anotação automática. A montagem de novo do transcriptoma identificou 6 novos transcritos não representados anteriormente. Foi confirmada a expressão de 88,20% (RPKM ≥ 10) das CDSs preditas e 38,73% dos genes foram diferencialmente expressos a 0°C, sendo 593 induzidos (fold change ≥ 2, p-value < 0,01) e 481 reprimidos (fold change ≤ -2, p-value < 0,01). Vários genes característicos da adaptação bacteriana ao frio foram diferencialmente expressos, como proteínas de choque frio, RNA helicases e a chaperona Trigger Factor. A abordagem transcriptomica confirmou 9 das 13 proteínas diferenciais do proteoma. A proteína de choque frio Csp1 apresentou um valor de volume relativo 32 vezes maior em 0°C nos géis DIGE. Uma hiper-expressão acima daquela observada pelo fold change calculado para o mesmo gene (somente 6 vezes mais expresso). Isto demonstra que a concentração de mRNA e proteína são diferentes no citosol da célula e que provavelmente outros mecanismos de regulação pós-transcricional estão envolvidos neste processo. As chaperonas e chaperoninas DnaK, HtpG, GroL, GroS e GrpE foram todas menos expressas no frio, evidenciando a importância da proteína Csp1 para a manutenção da homeostase celular. Os resultados obtidos apresentam os atributos genômicos da E. antarcticum B7 e as modificações na expressão que ocorrem para sua adaptação ao frio, as quais diferem daquelas observadas para bactérias psicrofílicas.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
21
INFLUENCIA DE LOS FACTORES CLIMÁTICOS EN LA DIVERSIDAD Y FUNCIONAMIENTO DE LAS COMUNIDADES VEGETALES ANTÁRTICAS. POSIBLE
RESPUESTA EN UN ESCENARIO DE CALENTAMIENTO GLOBAL (Influence of climatic factors on the diversity and functioning of Antarctic plant communities. Possible
answer in a global warming scenario)
García-Sancho, L.1 1Departamento de Biología Vegetal II, Facultad de Farmacia, Universidad Complutense de Madrid. [email protected] RESUMEN En la comunidad científica existe un interés creciente por conocer qué factores controlan la distribución actual de la vegetación en la Antártida, puesto que tanto la biodiversidad como la productividad vegetal podrían ser usadas como bioindicadores de cambio climático. Avances recientes en el conocimiento del comportamiento ecofisiológico y en la diversidad de la vegetación permiten utilizar la temperatura como factor discriminante. Se ha llevado a cabo una regresión lineal entre la temperatura media, a partir de datos macroclimáticos, y el número de especies de líquenes, musgos y hepáticas en un gradiente latitudinal desde 62ºS a 86ºS. A partir de esta regresión la vegetación Antártica puede dividirse en dos zonas: La zona microambiental se sitúa en una latitud superior a los 72ºS y en ella la biodiversidad (incluyendo riqueza en especies, cobertura y crecimiento) aparece desacoplada con respecto al macroclima y en su lugar está determinada por la coincidencia en localidades concretas de calor, disponibilidad de agua líquida, exposición favorable y protección al viento. La zona macroambiental, al norte de 72ºS, está fuertemente y positivamente correlacionada con la temperatura media, seguramente debido a la mayor disponibilidad de agua líquida, mayor duración del periodo de actividad y mayor precipitación, lo cual permite un aumento creciente de la productividad a medida que disminuye la latitud. Un posible calentamiento del clima en la Antártida daría lugar a un desplazamiento hacia el sur de la frontera entre estas dos zonas. Se discute además la relación entre la productividad, medida como crecimiento anual, y la biodiversidad vegetal en dos regiones extremas de la Antártida, las islas Shetland del Sur y los Valles Secos.
COLLAPSE OF A SUB-ANTARCTIC ALPINE FELDMARK ECOSYSTEM (El colapso de un ecosistema alpino subantártico de feldmark)
Bergstrom, D.M.1
1Australian Antarctic Division, Department of the Sustainability, Environment, Water, Population and Communities, 203 Channel Highway, Kingston, Tas 7050, Australia, ABSTRACT Detrimental changes in the alpine feldmark on Macquarie Island, a sub-Antarctic World Heritage property, were observed for the first time in December 2008. Many individuals of the endemic keystone cushion plant (Azorella macquariensis Orchard) did not emerge from winter senescence. Many bryophytes have also died. Such widespread dieback has not previously been observed for the island in 100 years of biological exploration. I describe this widespread occurrence, our exploration for cause and management response of this event.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
22
CARACTERÍSTICAS ECOFISIOLÓGICAS DE LAS PLANTAS VASCULARES ANTÁRTICAS: CERTEZAS, INCERTEZAS E IMPLICANCIAS PARA LAS
CONSECUENCIAS DEL CAMBIO CLIMÁTICO. (Ecophysiological characteristics of the Antarctic vascular plants: certainties, uncertainties and
implications for consequences of climate change)
Cavieres, L.A.1, Sanhueza, C.1, Sáez, P.2, Bravo, L.A.3
1ECOBIOSIS, Departamento de Botánica, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción. Barrio Universitario s/n, Concepción. Casilla 160-C. Concepción, Chile. [email protected]. 2Laboratorio Cultivo de Tejidos Vegetales, Centro de Biotecnología, Departamento de Silvicultura, Facultad de Ciencias Forestales, Universidad de Concepción, Casilla 160-C, Concepción, Chile. 3Laboratorio de Fisiología y Biología Molecular Vegetal, Instituto de Agroindustria, Departamento de Ciencias Agronómicas y Recursos Naturales, Facultad de Ciencias Agropecuarias y Forestales, Universidad de La Frontera. Center of Plant, Soil Interaction and Natural Resources Biotechnology, Scientific and Technological Bioresource Nucleus. Universidad de La Frontera, Casilla 54-D, Temuco, Chile. INTRODUCCIÓN De acuerdo al paradigma Darwiniano, los rasgos y/o características ecofisiológicas de una especie deberían estar relacionadas con las características bióticas y abióticas de su ambiente a fin de obtener el máximo "fitness" posible. Bajo esta mirada, las plantas que habitan la Antártica, deberían poseer una serie de rasgos y características ecofisiológicas que permiten su subsistencia en las condiciones extremas ambientales que caracterizan a estos ambientes. Deschampsia antarctica y Colobanthus quitensis son las únicas plantas vasculares que habitan naturalmente La Península Antártica. Si bien se han realizado una serie de estudios y revisiones sobre algunos aspectos de la ecofisiología de estas plantas vasculares antárticas (e.g. Xiong et al. 1999, 2000, Alberdi et al. 2001), hasta ahora no se ha sistematizado la información disponible en función de cómo sus características fisiológicas y la variación (o ausencia de variación) de estas a lo largo de gradientes ambientales nos pueden dar pistas sobre las consecuencias del cambio climático sobre estas plantas. Por otro lado, aunque se han realizado estudios experimentales de cambios ambientales en terreno (e.g. experimentos de aumento de la temperatura ambiental con open-top chambers (OTC) o calentadores Infra-rojos) (e.g. Day et al. 1999, 2008), pocos de ellos han estado centrados en la evaluación de los efectos de los cambios climáticos sobre las características ecofisiológicas de las plantas vasculares antárticas, y de cómo estas características ecofisiológicas determinan a su vez las respuestas observadas. A través del proyecto Anillo de Investigación en Ciencia Antártica en Ecofisiología de las plantas Vasculares nos hemos propuesto evaluar los efectos del cambio climático (i.e. aumento de la temperatura) sobre las respuestas ecofisiológicas de D. antarctica y C. quitensis tanto in situ como en condiciones de laboratorio. En este trabajo analizamos la información disponible sobre características morfo-fisiológicas de D. antarctica y C. quitensis en distintas condiciones ambientales tanto en el antártica como fuera de ella. A su vez mostramos algunos resultados preliminares sobre las respuestas en algunas características ecofisiológicas de estas al aumento de la temperatura in situ con el objetivo de comprender las posibles consecuencias del cambio climático sobre estas especies. MATERIALES Y MÉTODOS En Diciembre de 2012 se instalaron cámaras de calentamiento pasivo (Open-Top Chambers) en dos poblaciones de D. antarctica y C. quitensis creciendo en Isla Rey Jorge, en las cercanías de la base polaca Arctowski (62° 09' S, 58°28' W). Posteriormente, en Febrero de 2013 se registró las

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
23
modificaciones microclimáticas que generan las OTC en relación a: temperatura del aire y la superficie del suelo, temperatura foliar, radiación fotosintéticamente activa (PAR) y humedad de suelo. Adicionalmente, se tomaron muestras de suelo, dentro y fuera de OTC para realizar análisis nutricional. Tanto dentro como fuera de OTC se caracterizó la respuesta fotosintética de D. antarctica y el desempeño fotoquímico a diferentes intensidades lumínicas en D. antarctica y C. quitensis. RESULTADOS A partir del trabajo pionero de David Moore (1970), se comenzó a analizar y reportar la gran variación morfológica que presentan C. quitensis y D. antarctica, tanto a lo largo de todo su rango geográfico de distribución, como dentro de la antártica (e.g. Chwedorzewska et al. 2005, Giełwanowska et al. 2005). Al analizar plantas de C. quitensis colectadas en Punta Arenas (50°16’S; 75°15’W), en las cercanía de la Base Polaca Arctowski, Isla Rey Jorge (62° 09' S; 58°28' W) e Isla Lagotellerie en Bahía Margarita (67° 52’ S; 68° 42' W), encontramos que hacia las poblaciones más polares, existe una disminución del área de la sección transversal y longitud de las hojas (ver resumen de Sanhueza et al. para más detalles). No obstante, esta variación morfológica, análisis de las características fotosintéticas de estas poblaciones muestran muy poca variación. Por ejemplo, no se encontraron diferencias en la curva de respuesta fotosintética a la temperatura entre las 3 procedencias (Fig. 1).
Temperatura (ºC)
-5 0 5 10 15 20 25 30 35 40
A (
um
ol m
-2 s
-1)
0
2
4
6
8
10
12
14
16
18
ArctowskiPunta Arenas Lagotellerie
Figura 1: Tasa de fotosíntesis (A) a diferentes temperaturas en plantas de Colobanthus quitensis provenientes de diferentes localidades (Punta Arenas, Estación Arctowski e Isla Lagotellerie) y que han estado 3 meses en condiciones de laboratorio. Figure 1: Photosynthetic rate (A) at different temperatures in Colobanthus quitensis plants from different localities (Punta Arenas, Arctowski station and Isla Lagotellerie) under laboratory conditions during 3 months. Edward & Smith (1988) reportaron que en plantas de D. antarctica y C. quitensis mantenidas en invernaderos a 8º y 15ºC la temperatura a la cual se alcanza la máxima fotosíntesis era de 13ºC en D. antarctica y 19ºC en C. quitensis. Interesantemente, no observaron variaciones en este parámetro entre poblaciones ubicadas entre los 54º y 67º de latitud sur en ambos casos. En nuestras estimaciones para C. quitensis el óptimo fue de 17ºC, y como mencionamos no varió entre poblaciones. Xiong et al. (1999) reporta que la temperatura a la cual se produce la máxima fotosíntesis en condiciones de campo es 10ºC para D. antarctica y 14ºC para C. quitensis y que sus puntos de compensación a alta temperatura se alcanzan a temperaturas relativamente bajas (22º y 26ºC, respectivamente), sugiriéndose una alta

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
24
sensibilidad al aumento de la temperatura. Sin embargo, tanto Edward & Smith (1988) como en nuestro estudio encontramos dichos valores están por sobre los 30ºC. Las plantas pueden modificar sus curvas de respuesta ambiental conforme el ambiente se modifica. Así, Xiong et al. (2000) reportan que en condiciones de laboratorio mientras C. quitensis muestra cierta plasticidad en los óptimos de temperatura para la máxima fotosíntesis según la temperatura a la que crece (de 8.4ºC cuando crece a 7ºC a 11.5ºC cuando crece a 20ºC), resultado similar a lo reportado por Sierra-Almeida et al. (2007) (de 17ºC cuando crece a 4ºC a 18.6ºC cuando crece a 15ºC). En el estudio de Xiong et al. (2000) D. antarctica no muestra cambios (ca. 10ºC, similar a lo encontrado por Bystrzejewska-Piotrowska & Urban (2009)). Por lo tanto, por un lado parece poco probable que el aumento de la temperatura afecte la respuesta fotosintética de estas especies. Por otro lado, los altos valores de temperatura para realizar el máximo de fotosíntesis en relación a las temperaturas habituales de la antártica sugieren que el calentamiento podría tener efectos positivos. Mediciones realizadas este verano en plantas de D. antarctica creciendo dentro y fuera de OTC sugieren que efectivamente el calentamiento tiene efectos positivos sobre la fotosíntesis (Fig. 2), y en consecuencia sobre el crecimiento (ver resumen de Sanhueza et al.).
OTC EA
Fo
tosí
nte
sis
(um
ol m
-2 s
-1)
0
2
4
6
8
10
4°C 12°C
Fig. 2. Tasa de fotosíntesis en plantas de Deschampsia antarctica creciendo dentro y fuera de OTC en las cercanías de la
estación polaca Arctowski. Las barras de distinto color indican mediciones realizadas a diferentes temperaturas. Fig.2. Photosynthetic rate (A) in Deschampsia antarctica plants growing inside and outside of OTC around of the Arctowski Polish
station. Bars with different colors indicate measurements conducted at different temperatures. DISCUSIÓN Otros estudios donde se ha aumentado artificialmente la temperatura ambiental muestran que el calentamiento favorece el contenido de carbono en D. antarctica (Day et al. 2008), lo que se podría relacionar con una mayor fotosíntesis. No obstante, tanto en el experimento de Day et al. (2008), como en nuestro estudio en Arctowski, el calentamiento tiene efectos directos sobre los nutrientes. Por ejemplo, nitrógeno y materia orgánica incrementaron significativamente en los dos sitios con OTC instalados en Arctowski (ver resumen Sáez et al. para mayor detalle). Por lo tanto, surge la interrogante de en que medida son las mayores temperaturas, los mayores niveles de nutrientes o el efecto sinérgico de ambas variables lo que está detrás de las respuestas observadas en las plantas vasculares antárticas. D. antactica y C. quitensis se distribuyen preferentemente en sitios más ricos en nutrientes en la Isla Rey Jorge. No obstante, otros atributos del suelo, tales como pH bajo, mal drenaje del suelo y longitud de la cobertura de nieve en la estación de crecimiento son factores aún más críticos que la disponibilidad de nitrógeno y fosforo para la sobrevivencia de las plantas (Park et al. 2012). Esto apunta a que estas

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
25
plantas serían altamente tolerantes al déficit nutricional del suelo y por lo tanto muy eficientes en la absorción de nutrientes. Por ejemplo, se ha reportado que estas dos plantas vasculares poseen una asociación con hongos endófitos (Upson et al. 2008). Se ha postulado que estos hongos endófitos podrían mineralizar péptidos y aminoácidos en la rizósfera (Upson et al. 2009), esto además implica que las poblaciones de estas dos especies son capaces de modificar las características químicas del suelo. Algunos resultados refrendan este postulado en que C. quitensis y D. antarctica significativamente cambian la dinámica temporal de la mineralización de N y su inmovilización, ejerciendo una fuerte influencia sobre el reciclaje de nitrógeno y carbono (Roberts et al. 2009). Por lo tanto, ambas especies poseen una serie de adaptaciones al ambiente particular en que habitan. El cambio climático está modificando estas condiciones y por ello existen múltiples interrogantes respecto a ¿cómo responderán las plantas al aumento de la temperatura? ¿cómo se modificarán los atributos de las plantas en el escenario de cambio climático? ¿será esta expansión de las poblaciones sostenible en el tiempo? AGRADECIMIENTOS Proyecto PIA ART 1102. REFERENCIAS BIBIOGRÁFICAS Alberdi, M. et al. 2001. Ecophysiology of Antarctic vascular plants. Physiologia Plantarum 115: 479-486. Bystrzejewska-Piotrowska, G. & Urban, P.L. 2009. Tufted hairgrass (Deschampsia caespitosa) exhibits a lower photosynthetic plasticity than Antractic hairgrass (D. antarctica). Journal of Integrative Plant Biology 51: 593-603. Chwedorzewska, K.J. et al. 2005. High anatomical and low genetic diversity in Deschampsia antarctica from King George Island, the Antarctic. Polish Polar Research 29: 377-386. Day, T.A. et al. 1999. Growth and reproduction of Antarctic vascular plant in response to warming and UV-B radiation reductions in the field. Oecologia 119: 24-35. Day, T.A. et al. 2008. Warming increases aboveground plant biomass and C stock in vascular-plant-dominated Antarctic tundra. Global Change Biology 14: 1827-1843. Edward, J.A. & Smith, R.I.1988. Photosynthesis and respiration of Colobanthus quitensis and Deschampsia antarctica from the maritime antarctic. British Antarctic Survey Bulletin 81: 43-63. Giełwanowska, I. et al. 2005. Anatomical features and ultrastructure of Deschampsia antarctica leaves from different growing habitats. Annals of Botany 96: 1109-1119. Moore, D.M. 1970. Studies in Colobanthus quitensis and Deschampsia antarctica II. British Antarctic Survey Bulletin 23: 63-80. Park J.S. et al. 2012. Influence of soil properties on the distribution of Deschampsia antarctica on King George Island, Maritime Antarctica Polar Biology 35:1703-1711. Roberts P. et al. 2009 Vegetation cover regulates the quantity, quality and temporal dynamics of dissolved organic carbon and nitrogen in Antarctic soils. Polar Biology 32: 999-1008. Upson R. et al. 2008. Root-Fungal Associations of Colobanthus quitensis and Deschampsia antarctica in the Maritime and Subantarctic Arctic Antarctic and Alpine Research 40: 592-599. Upson R. et al. 2009. Nitrogen form influences the response of Deschampsia antarctica to dark septate root endophytes. Mycorrhiza 20:1-11. Xiong, F.S. et al. 1999. Photosynthetic temperature responses of Antarctic vascular plants Colobanthus quitensis and Deschampsia antarctica. Physiologia Plantarum 106: 272-286. Xiong, F.S. et al. 2000. Photosynthetic and respiratory acclimation and growth response of Antarctica vascular plants to contrasting temperature regimes. American Journal of Botany 87: 700-710.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
26
ANÁLISIS DE LA ESTRUCTURA DE CONSORCIOS MICROBIANOS TERRESTRES DE LA PENÍNSULA DE FILDES (ANTÁRTIDA MARÍTIMA) MEDIANTE PIROSECUENCIACIÓN.
(Analysis by pyrosequencing of microbial structure in terrestrial mat consortia of Fildes Peninsula, maritime Antarctica)
Callejas, C.1, Batista, S.1 y Etchebehere, C2.
1 Molecular Microbiology Unit, IIBCE, Montevideo, 11600, Uruguay. [email protected] 2 Laboratory of Microbial Ecology, IIBCE, Montevideo, Uruguay
INTRODUCTION King George Island (KGI), the largest of South Shetland archipelago, is located at the north-western region of Antarctic Peninsula. The southernmost part of the island, the Fildes Peninsula, becomes essentially ice-free during the Austral summer. During this period, development and succession of microbial mats routinely arise at isolated inland glacial seepages and other freshwater systems (Vogt & Braun, 2004). Microbial mats can be described as “thick biofilms” where closely related clusters of microorganisms attached to a surface and often embedded in extra cellular polymeric substances (EPS), embracing a plethora of metabolic pathways and phylotypes (de los Ríos et al., 2004). Recently, some studies have analyzed the molecular diversity in terrestrial microbial communities present in the Fildes Peninsula (Texeira et al., 2010, Callejas et al., 2011). Climate change is affecting the Antarctic Peninsula. The impacts on this region include between others warming trend, glacial retreat, exposure of new land areas without ice cover and changes in biota distribution (Folland et al., 2001). For this reason, it is important to know more about microbial diversity and succession and also how these systems are affected by environmental changes.
AIMS To describe the diversity of bacteria (including cyanobacteria) and archaea in terrestrial microbial mats from King George/ 25 de Mayo Island. To study the changes in the structure and diversity of the microbial communities of the mats during the summer. METHODOLOGY We estimated the composition and diversity of two microbial mats collected in Fildes Peninsula, during the summer of 2011. We used two methodologies: terminal- restriction fragment length polymorphism (T-RFLPs) and 454 pyrosequencing of 16S ribosomal RNA (rRNA) gene, amplified from the whole environmental DNA.
RESULTS AND DISCUSSION We analyzed two mats sampled three times during by the summer: in December 2010 (beginning of Austral summer), February and March 2011. We analyzed roughly 34000 sequences of the V4 region of the 16S rRNA gene of the samples collected during December, February and March. Sequence analysis indicated that mats present in this extreme habitat could be considered as diverse ecosystems, since we could estimate 2313 OTUs using 3% cutoff (Table 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
27
Sample # seq # obs Shannon (H) Coverage (%) B Jan 4200 780 8.1 81 B Feb 4200 667 7.4 84 B Mar 4200 682 7.5 84 C Jan 4200 743 8 82 C Feb 4200 757 7.5 82 C Mar 4200 821 7.7 80
Tabla 1. Datos de la pirosecuenciación mostrando el número de secuencias (# seq), el número de OTUs estimado con un 3% de distancia (# obs), el valor del índice de Shannon (H) y la cobertura (Coverage) de la diversidad en porcentaje, para cada
muestra. Table 1. Pyrosequencing data showing the number of sequences (# seq) obtained, number of OTUs estimated with 3%
distance cutoff (# obs), Shannon index value (H) and the coverage of the diversity estimated in percentage (Coverage) for each sample.
The phyla present in the samples were similar to those described for soil samples and other Antarctic terrestrial environments (Texeira et al, 2010, Makhalanyane et al., 2013). Most abundant phyla were Verrucomicrobia (24-30%), Proteobacteria (17-26%), Alphaproteobacteria (10-18%), Cyanobacteria (9-17%), Gemmatimonadetes (4-10%) (Figure 1).Phyla abundances were similar between samples, while some differences were evident at the Order level over time. Unicellular Cyanobacteria (Chroccocales) abundance increased during the summer, becoming dominant at the end of the season. On the other hand, Stramenopiles (algae, diatoms) decreased with time, in both samples analyzed. A very low abundance from Archaea was detected.
Figura 1. Abundancia relativa de las diez OTUs más abundantes en cada muestra analizada.
Figure 1. Relative abundances of the ten most dominant OTUs in each sample analyzed. CONCLUSIONS This is one of the first studies focused on Antarctic microbial mats using pyrosequencing data analysis. Our results show that microbial communities in different Antarctic mats have a conserved diversity at phylum-level and a greater diversity of OTUs, particular and characteristic of each site.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
28
ACKNOWLEDGMENTS This project was funded by Instituto Antártico Uruguayo (IAU) and Instituto de Investigaciones Biológicas Clemente Estable (IIBCE).
BIBLIOGRAPHY Callejas, C., Gill, P.R., Catalán, A.I., Azziz, G., Castro-Sowinski, S., and. Batista, S. 2011. Phylotype diversity in a benthic cyanobacterial mat community on King George Island, maritime Antarctica. World J Microbiol Biotechnol 27(6):1507-1512. DOI 10.1007/s11274-010-0578- de los Ríos, A., Ascaso, C., Wierzchos, J., Fernández-Valiente, E., andQuesada, A. 2004. Microstructural characterization of cyanobacterial mats from the McMurdo Ice Shelf, Antarctica. Appl Environ Microbiol 70:569-580. Folland &T.R. Karl 2001. Observed climate variability and change. En: Climate change 2001: the scientific basis (ed. J. T. Houghton, Y. Ding, D. J. Griggs, M. Noguer, P. J. van den Linden, X. Dai, K. Maskell & C. A. Johnson), Cambridge, UK: Cambridge University Press. pp. 99-182. Makhalanyane TP, Valverde A, Birkeland NK, Cary SC, Marla Tuffin I, Cowan DA. 2013. Evidence for successional development in Antarctic hypolithic bacterial communities. ISME J. Jun 13. doi: 10.1038/ismej.2013.94. Teixeira, L., Peixoto, R.L., Cury, J.C., Sul, W.S., Pellizari, V.H., Tiedje, J., Rosado, A.S. 2010. Bacterial diversity in rhizosphere soil from Antarctic vascular plants of Admiralty Bay, maritime Antarctica ISME J 4:989-1001. Vogt, S. and Braun,M. 2004. Influence of glaciers and snow cover on terrestrial and marine ecosystems as revealed by remotely-sensed data. Pesqui Antár Bras 4:105-118.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
29
USO DE LA METAGENÓMICA FUNCIONAL PARA LA IDENTIFICACIÓN DE ENZIMAS DE POTENCIAL INTERÉS BIOTECNOLÓGICO PRESENTES EN LA PENÍNSULA ANTARTICA
(Functional metagenomics as a tool for the identification of biotechnological interesting enzymes in the Antarctic Peninsula)
Ferrés, I.1, Amarelle, V., Noya, F. y Fabiano, E.
1Departamento de Bioquímica y Genómica Microbianas, Instituto de Investigaciones Biológicas Clemente Estable, Av. Italia 3318, Montevideo, Uruguay. E-mail: [email protected] INTRODUCCIÓN El empleo de enzimas en procesos químicos, ofrece numerosas ventajas dado que presentan gran especificidad sobre los sustratos en que actúan, funcionan a temperaturas y pH moderados, se derivan de recursos renovables y son biodegradables (Cherry y Fidantsef 2003). Esto ha llevado a que su utilización en la industria sea cada vez más generalizada, aunque deben poder competir con los procesos tradicionales manteniendo un bajo costo de producción. Una de las maneras de lograr una disminución en los costos es adaptar los procesos para ser realizados a bajas temperaturas, evitando así la necesidad de invertir energía en las etapas de calentamiento (Cavicchioli et al. 2002). Esto ha llevado a industrias de diversas ramas de la economía a investigar el potencial uso de enzimas adaptadas al frío. Por otra parte, el estudio de enzimas adaptadas al frío es de gran interés académico ya que presentan particularidades estructurales y bioquímicas únicas que le permiten trabajar en esas condiciones extremas de temperatura (Gerday et al. 2000). Los ambientes fríos son el hábitat de microorganismos especialmente adaptados a vivir en esas condiciones, denominados microorganismos psicrófilos. Para ello, estos microorganismos han desarrollado varias adaptaciones fisiológicas. A nivel enzimático, la síntesis de enzimas adaptadas a funcionar a bajas temperaturas parece ser el principal mecanismo utilizado por los microorganismos psicrófilos (Collinset al. 2008). Algunas características de las enzimas adaptadas al frío incluyen alta actividad específica a bajas y moderadas temperaturas, y la capacidad de ser inactivadas fácilmente con un incremento en la temperatura (Gerday et al. 2000). Estas propiedades pueden ser muy útiles en diversas aplicaciones biotecnológicas. Los métodos tradicionalmente utilizados para la búsqueda de nuevos biocatalizadores dependen del aislamiento, cultivo y posterior búsqueda de los productos de interés. Se estima que una fracción muy pequeña (menor al 1%) de los microorganismos presentes en un determinado ambiente pueden ser recuperados mediante las técnicas tradicionales de cultivo (Giovannoni, S. J., et al. 1990). Esto conlleva a que las colecciones de cultivos existentes sub-representan la diversidad genética y el potencial biotecnológico de estos microorganismos. Para avanzar en el conocimiento de ese 99% restante de especies aún no cultivables se ha desarrollado una aproximación metodológica denominada metagenómica, o genómica colectiva. La metagenómica permite el estudio de una población presente en una muestra ambiental sin necesidad de conocer la composición de la misma. Consiste en la extracción del ADN genómico total de la comunidad microbiana allí presente, el cual es utilizado para la construcción de genotecas en bacterias de uso corriente en el laboratorio. Se logra de esta manera tener representados gran cantidad de los genomas de las especies que integran la comunidad sin el requisito previo de aislar y cultivar los microorganismos en el laboratorio. Una manera de analizar la metagenoteca consiste en detectar los clones portadores de una determinada función, esta estrategia es conocida como metagenómica funcional. Esta aproximación tiene la ventaja

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
30
de no sesgar la búsqueda a partir del conocimiento previo de enzimas ya conocidas. El principal inconveniente es que los genes buscados deberán transcribirse y traducirse correctamente en la bacteria hospedera elegida para construir la genoteca. Asimismo, las proteínas sintetizadas en el nuevo hospedero deberán ser activas y eventualmente exportarse para que sea posible detectar su función (Cowan et al. 2005). Con la finalidad de buscar nuevas enzimas adaptadas al frio potencialmente aplicables en biotecnología, nos propusimos la utilización de una aproximación metagenómica funcional a partir de una muestra de agua tomada de un charco de deshielo en la península Antártica. MATERIALES Y MÉTODOS Se partió de una muestra tomada en la campaña antártica ANTARKOS XXIX, en enero de 2013. La muestra consistió en una toma de agua de un charco de deshielo con presencia de crecimiento de algas y musgos, de pH 9.0 tomada en el sitio S 63° 13´ 12,8¨; W 057° 13´5,4´´. La muestra se mantuvo en hielo unas horas, luego de lo cual se extrajo el ADN metagenómico a partir de 300 l de muestra empleando el kit PowerSoil MOBIO para extracción de ADN de suelo. El ADN metagenómico se almacenó en frío hasta su procesamiento en el laboratorio. Se comprobó la calidad y el tamaño del ADN extraído mediante electroforesis, y se cuantificó espectrofotométricamente. Para la construcción de la biblioteca metagenómica se utilizó el kit Copy ControlTM Fosmid Library Production de Epicentre Biotechnologies. Se repararon los extremos del ADN metagenómico de manera de poder clonarlos en el vector pCCFOS1. Este vector contiene una resistencia a cloranfenicol (CmR) como marcador de selección y sitios cos para ser reconocido y empaquetado por el fago Lambda. La mezcla de ligación fue empaquetada en el fago Lambda y posteriormente se infectó la cepa EPI300T1R de Escherichia coli. Se seleccionaron aquellos clones con resistencia a Cm y se ordenaron en placas de 96 pocillos en conjuntos de aproximadamente 30 colonias por pocillo. Para identificar los clones de la biblioteca portadores de funciones de interés, se crecieron en medio sólido en presencia de diferentes sustratos que permitieran detectar las siguientes actividades: lipolítica, celulasa, hemicelulasa, peroxidasa, manganeso peroxidasa y lacasa. También se evaluó la producción de compuestos antimicrobianos y la capacidad de utilización de celulosa microcristalina (avicel) como única fuente de carbono. En una primera instancia se incubaron las placas a 30ºC durante 24 horas, de manera de permitir el crecimiento de la cepa hospedera. Dado que la mayoría de los microorganismos capaces de crecer en frío son psicrotolerantes y se desarrollan correctamente a 25ºC, es esperable que las enzimas codificadas por el ADN metagenómico también funcionen a 25ºC. Entonces, transcurridas las 24 horas a 30ºC, se incubaron las placas a 25ºC para favorecer la expresión de genes provenientes de bacterias psicrófilas. A partir de los conjuntos que presentaron actividades positivas se aisló el clon que codifica para dicha actividad enzimática. Posteriormente se extrajo el fósmido de los clones positivos y se reintrodujo en la cepa hospedera nuevamente, de manera de verificar que la función estaba asociada a la presencia del fósmido y no a otros factores externos. Para determinar si las actividades enzimáticas con más de un clon positivo eran resultado de la presencia del mismo fósmido en todos los casos, se compararon los perfiles de restricción de dichos fósmidos. RESULTADOS A partir de ADN metagenómico de una muestra ambiental colectada en la península Antártica se construyó una biblioteca metagenómica de aproximadamente 20 mil clones. Mediante el análisis funcional de la biblioteca se detectaron 50 clones con actividad lipolítica, 2 clones con actividad

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
31
celulolítica y 2 clones capaces de crecer con avicel como única fuente de carbono. No se detectaron clones con actividad lacasa o peroxidasa, ni tampoco clones que produjeran compuestos antibacterianos. Interesantemente, encontramos 3 clones que presentaron una coloración marrón-rojiza en los medios reportados para detección de actividad manganeso peroxidasa. Dado que las placas contienen manganeso, el fenotipo observado podría corresponder a la presencia de actividad manganeso oxidasa, que induce la formación de óxido de manganeso el cual es insoluble y forma precipitados marrones-rojizos. Se seleccionaron 3 clones con actividad lipolítica promisoria (halos de hidrólisis más grande en menos tiempo de incubación) y los 3 clones con posible actividad manganeso oxidasa para continuar este estudio. Se determinó mediante la comparación de los perfiles de restricción que los 3 clones con actividad lipolítica correspondían a distintos fósmidos, mientras que los 3 clones con actividad manganeso oxidasa correspondían al mismo fósmido. Se comprobó que la función observada estaba codificada en el fósmido ya que al transferirlo por electroporación a otra cepa de E. coli, ésta adquiría la capacidad de expresar la función correspondiente. DISCUSIÓN Y CONCLUSIONES En este trabajo se generó una biblioteca metagenómica de aproximadamente 20 mil clones a partir de ADN metagenómico extraído de una muestra ambiental obtenida en la península Antártica. Considerando que cada clon consta de aproximadamente 40kb de información proveniente de ADN metagenómico, la información total contenida en la biblioteca corresponde a 0,8 Gb. Mediante una aproximación metagenómica funcional pudimos detectar la presencia de varias actividades enzimáticas en la biblioteca. Actualmente continuamos profundizando en la determinación de la secuencia nucleotídica y la caracterización bioquímica de dichas enzimas. El suelo antártico es rico en microorganismos que pueden representar un reservorio aún poco explorado de enzimas adaptadas al frío, con potencial aplicación biotecnológica. En este trabajo demostramos que mediante la utilización de una aproximación metagenómica funcional es posible identificar diversas enzimas de interés. Si bien no es un abordaje sencillo y tiene varias limitaciones, al ser una aproximación independiente de cultivo permite abarcar de forma representativa la información contenida en una muestra ambiental. Si bien en este trabajo se evaluaron solamente algunas actividades enzimáticas, la biblioteca metagenómica construida puede ser utilizada en un futuro para la búsqueda de otras actividades. Una biblioteca metagenómica es en sí misma un recurso que puede ser ampliamente explotado ya que constituye una fuente de información muy amplia. Podemos concluir que la metagenómica funcional posee un gran potencial como herramienta para la identificación de nuevas funciones enzimáticas que puedan tener una aplicación directa en la industria o en la investigación científica. AGRADECIMIENTOS Instituto Antártico Uruguayo (IAU), PEDECIBA Biología-Química REFERENCIAS BIBLIOGRÁFICAS Cavicchioli, R., Siddiqui, K. S., Andrews, D. y Sowers, K. R. 2002. Low-temperature extremophiles and their applications. Curr Opin Biotechnol 13(3): 253-261. Collins, T., Roulling, F., Piette, F., Marx, J. C., Feller, G., Gerday C. y D’Amico, S. 2008. Fundamentals of Cold-Adapted Enzymes. Heidelberg, Springer. Cowan, D., Meyer, Q., Stafford, W., Muyanga, S., Cameron, R. y Wittwer, P. 2005. Metagenomic gene discovery: past, present and future. Trends Biotechnol 23(6): 321-329. Cherry, J. R. y Fidantsef, A. L. 2003. Directed evolution of industrial enzymes: an update. Curr Opin Biotechnol 14(4): 438-

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
32
443. Gerday, C., Aittaleb, M., Bentahir, M., Chessa, J. P., Claverie, P., Collins, T., D'Amico, S., Dumont, J., Garsoux, G., Georlette, D., Hoyoux, A., Lonhienne, T., Meuwis, M. A. y Feller, G. 2000. Cold-adapted enzymes: from fundamentals to biotechnology. Trends Biotechnol 18(3): 103-107. Giovannoni, S. J., Britschgi, T. B., Moyer, C. L. y Field, K. G. 1990. Genetic diversity in Sargasso Sea bacterioplankton. Nature 345(6270): 60-63.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
33
CARACTERIZACIÓN TAXONÓMICA Y FUNCIONAL DE LAS COMUNIDADES NANOEUCARIONTES FOTOSINTÉTICAS SOMETIDAS A DISTINTOS REGÍMENES DE
LUZ EN BAHÍA FILDES, ANTÁRTICA, MEDIANTE SECUENCIACIÓN MASIVA. (Taxonomic and functional characterization of photosynthetic nanoeukariotic community subjected to different light regimes
in Fildes Bay, Antarctica, through massive sequencing technology)
Rodríguez-Marconi, S.1, Moreno, M.1, Sánchez, C.1, Rojas, M.1, Egas, C.2, De la Iglesia, R.2 &Trefault, N.1
1Centro de Genómica y Bioinformática, Universidad Mayor. Camino La Pirámide #5750, Santiago, Chile. 2Departamento de Genética Molecular y Microbiología, Pontificia Universidad Católica de Chile, Alameda #340, Santiago, Chile. E-mail: [email protected] RESUMEN Los microorganismos son una de las entidades biológicas más importantes en los ecosistemas marinos, formando parte de la base de las tramas tróficas oceánicas, y siendo un elemento esencial en los ciclos biogeoquímicos a nivel global. Dentro de ellos, los microorganismos marinos fotosintéticos, algunos de los cuales constituyen el fitoplancton, son un componente fundamental, ya que son capaces de transformar el CO2 atmosférico en materia orgánica utilizando la luz como fuente de energía. Es por esta razón que la luz es un factor ambiental de gran relevancia para este tipo de microorganismos, pues las fluctuaciones de luz podrían generar variaciones en la estructura y composición de las comunidades fitoplanctónicas. En este sentido, la Antártica es un lugar que presenta grandes variaciones en intensidad y duración de luz, o radiación fotosintéticamente activa (PAR) durante el día, lo que la convierte en un ambiente ideal para el estudio del efecto de las fluctuaciones en los regímenes de luz sobre las comunidades fitoplanctónicas marinas. Uno de los efectos que podría tener una variación en la intensidad de luz sobre la columna de agua sería la generación de florecimientos de microalgas, que consisten en un rápido incremento en la abundancia de determinados taxas que componen el fitoplancton. En este contexto, un antecedente clave para la realización de este trabajo fue el registro, obtenido mediante cuantificación por citometría de flujo, de un incremento en la abundancia de nano eucariontes fotosintéticos (3 a 20µm de tamaño)) durante un pulso de surgencia registrado en Bahía Fildes, Isla Rey Jorge, Antártica, durante el verano del 2012. Una aproximación que ha tomado importancia para el estudio de microorganismos es la secuenciación masiva. Las metodologías de secuenciación que han sido desarrolladas en los últimos tiempos han permitido describir de manera mucho más precisa las comunidades de microorganismos en distintos ambientes, debido a la gran cantidad de información que logran entregar. Por ejemplo, han dilucidado componentes de la llamada “biósfera poco abundante”, gracias a elevada cobertura que alcanzan (Bartram et al., 2011). Por otra parte, a diferencia del uso de marcadores genéticos, los metagenomas analizan la información genética completa de una comunidad, lo que permite hacer una descripción del potencial funcional esta (Dinsdale et al., 2008). MATERIALES Y MÉTODOS Este trabajo tiene como objetivo realizar una caracterización taxonómica y funcional de la comunidad eucarionte fotosintética nanoplanctónica expuesta a distintas intensidades de PAR mediante el uso de secuenciación masiva. Este estudio se llevó a cabo usando secuenciación de alto rendimiento MiSeq (Illumina). Se utilizaron muestras de agua tomadas en Bahía Fildes, Isla Rey Jorge, Antártica, superficialmente, y a una profundidad correspondiente al 10% del PAR medido en superficie. Se realizó una filtración secuencial

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
34
para separar las fracciones nanoplanctónica pequeña (3 a 12µm) y grande (12 a 20µm), para generar un total de 4 librerías. Los datos obtenidos, luego de un proceso de filtración por calidad y trimming, para eliminar extremos de baja calidad, fueron subidos al servidor de análisis metagenómicos MG-RAST (Meyer et al., 2008). Este servidor permite realizar control de calidad, anotación automatizada y análisis de muestras provenientes de secuenciación shotgun de comunidades microbianas, además de estandarizaciones y comparaciones entre metagenomas. Por otra parte, se realizaron análisis in house a través de un pipeline de análisis de datos metagenómicos, que incluye el filtrado, ensamblaje, anotación y comparación, mediante programas bioinformáticos específicos para cada etapa. RESULTADOS El análisis de los datos indica que existen diferencias de distribución de secuencias correspondientes a bacterias y eucariontes en las dos fracciones de tamaño analizadas. De esta forma, se observa que la fracción nanoplanctónica pequeña (3 a 12µm) presenta cerca de un 20% menos de secuencias correspondientes a organismos eucariontes que la fracción nanoplanctónica de mayor tamaño (12 a 20µm). Enfocando el análisis hacia las secuencias correspondientes al dominio eucarionte, se encuentra, para todas las muestras, una dominancia de secuencias correspondientes al Phylum Bacillariofita, conocidas comúnmente como diatomeas. Esto es concordante con el aumento de nanoeucariontes fotosintéticos medido por citometría de flujo para las mismas muestras, y con el pulso de surgencia ocurrido en este periodo. Además concuerda con el hecho que las diatomeas son usualmente responsables del inicio de florecimientos fitoplanctónicos en zonas costeras con altos contenidos de nutrientes y escasa estratificación, que parece ser el caso de Bahía Fildes. Si se realiza una comparación a nivel taxonómico entre las distintas fracciones y regímenes de luz, se encuentra dentro de la fracción nanoplanctónica de 12 a 20µm la presencia de la clase Bacillarioficeae que sólo se ve en la zona superficial y no en la profundidad correspondiente al 10% del PAR registrado en superficie. Por otra parte, encontramos que esta misma clase sí se encuentra en profundidad al analizar la fracción nanoplanctónica pequeña (Figura 1)
Figura 1: Comparación taxonómica de las muestras correspondientes a la fracción nanoplanctónica mayor (12 - 20µm)
correspondiente a ambas profundidades estudiadas y la fracción nanoplanctónica menor (3 - 12µm) de la profundidad del 10% del PAR superficial. Los datos de abundancia se encuentran estadarizados y normalizados entre 0 y 1, en que 1

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
35
corresponde al tipo de secuencia más abundante. El análisis fue llevado a cabo utilizando la fuente de anotación M5NR, con un valor máximo de E-value de 1e-5, un porcentaje mínimo de identidad de 80% y un largo mínimo de alineamiento de 60
pb. Figure 1: Taxonomical comparison between samples from the 12 - 20µm nanoplanktonic fraction from both depths and 3-
12µm fraction from the 10% superficial PAR depth. The sequence abundance data are normalized and standardized between 0 and 1, with 1 being the most abundant. The source of annotation is M5NR,with a cutoff E-value of 1e-5, 80% minimum
identity and 60bp minimum alignment. En cuanto a comparaciones funcionales, según la anotación basada en KEGG Orthology (KO), se puede observar una mayor abundancia de secuencias asociadas a pigmentos antena (relacionados con la fotosíntesis), en la fracción nanoplanctónica de mayor tamaño que se encuentra en profundidad. Por otro lado, si se comparan las dos fracciones nanoplanctónicas a la profundidad del 10% del PAR superficial, utilizando la anotación KO además de una basada en subsistemas, se encuentra que la fracción de mayor tamaño contiene más secuencias asociadas a fotosíntesis que la pequeña. Interesantemente según la clasificación KO, sólo en la fracción pequeña se encuentran secuencias asociadas a fijación de carbono en organismos fotosintéticos (Figura 2).
Figura 2: comparación funcional de las muestras correspondientes a la fracción nanoplanctónica mayor (12 - 20µm)
correspondiente a ambas profundidades estudiadas y la fracción nanoplanctónica menor (3 - 12 µm). Los datos de abundancia de secuenciar se encuentran estadarizados y normalizados entre 0 y 1, en que 1 corresponde al tipo de secuencia más
abundante. Se muestran análisis en base a las fuentes de anotación KEGG Orthology, con un valor máximo de E-value de 1e-5, un porcentaje mínimo de identidad de 80% y un largo mínimo de alineamiento de 60 pb. Figure 2: Functional comparison
between samples from the 12 - 20µm nanoplanktonic fraction from both depths and 3-12µm fraction from the 10% superficial PAR depth. The sequence abundance data are normalized and standardized between 0 and 1, with 1 being the
most abundant. The source of annotation is KEGG orthology, with a cutoff E-value of 1e-5, 80% minimum identity and 60bp minimum alignment.
En resumen, se encuentra una dominancia de secuencias asociadas al Phylum Bacillariophyta, lo que concuerda con mediciones de abundancia de nanoeucariontes fotosintéticos y con la ocurrencia de un

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
36
pulso de surgencia durante el periodo de muestreo. Los resultados preliminares de comparación taxonómica y funcional indican que existen diferencias asociadas tanto a las dos fracciones de tamaño nanoplanctónicas como a intensidad de luz a la que se encontraban sometidos los microorganismos al momento del muestreo. En conjunto estos resultados contribuyen al conocimiento de la ecología y diversidad de microorganismos marinos en ambientes extremos como la Antártica, abarcando tanto su composición taxonómica como su potencial metabólico, lo que integrado a estudios fisicoquímicos y de abundancia de microorganismos en la zona aporta con una descripción completa de la zona de estudio.
AGRADECIMIENTOS Proyecto INACH T16-10 REFERENCIAS Bartram, AK., Lynch, M. D. J., Stearns, J. C., Moreno-Hagelsieb, G. y. Neufeld, J.D., 2011. Generation of Multimillion-Sequence 16S rRNA Gene Libraries from Complex Microbial Communities by Assembling Paired-End Illumina Reads. Applied Environmental Microbioogy 77 (11):3846-3852 Dinsdale, E., Edwards, R., Hall, D., Angly, F., Breitbart, M., Brulc, J. M., Furlan, M., Desnues C., Haynes, M., L. Li, L. McDaniel, L. ,Moran, M.A., Nelson, K. E., Nilsson, C., Olson, R. Paul, J., Rodriguez Brito, Ruan, B. Y. , Swan, B. K., Stevens, R., Valentine, D. L., Vega Thurber, R., Wegley, L., White, B. A. y Rohwer, F., 2008. Functional metagenomic profiling of nine biomes. Nature, 452(7187): 629–32. Meyer, F., Paarmann, D., D'Souza, M., Olson, R., Glass, E.M., Kubal, M. Paczian, T., Rodriguez, A., Stevens, R., Wilke, A., Wilkening, J. y Edwards, R.A., 2008. The metagenomics RAST server – a public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinformatics 9:386.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
37
DETERMINACIÓN DE LA TAXONOMÍA Y DIVERSIDAD DE POBLACIONES MICROBIANAS EN SUELOS ANTÁRTICOS A TRAVÉS DE DGGE Y SECUENCIACIÓN 454
(Determination of the taxonomy and diversity of soil microbial populations based on DGGE fingerprinting and 454 sequencing)
Ratti, M.F. 1, Monserrate, B.L., Vargas, J.D., Catagua, D.J., van Bruggen, A.H.C.2, Liao, H. L.3,
Cevallos-Cevallos, J.M. y Peralta, E.L. 1Centro de Investigaciones Biotecnológicas del Ecuador – Escuela Superior Politécnica del Litoral (CIBE-ESPOL). Km 30.5 Vía Perimetral, Campus Gustavo Galindo, Edif. PROTAL, planta alta. [email protected] 2 Planth Pathology Department, University of Florida, Gainesville, FL. 3 Biology Department. Duke University, Durham, NC. RESUMEN La diversidad microbiana en suelos ha interesado a la comunidad científica por su importancia en el ciclo de macromoléculas, en la ecología y la bioprospección. Como resultado de las expediciones a la Antártida se ha concluido una baja diversidad comparada con muestras mesófilas; atribuyéndose esto a las condiciones ambientales extremas, ciclos de congelamiento y descongelamiento y a la escasez de nutrientes y humedad (Pierce 2012). Sin embargo, la mayoría de estos estudios han considerado sólo la cantidad de microorganismos cultivables presentes en dichas muestras. Los análisis moleculares, a diferencias de las técnicas dependientes de cultivo, proveen datos más precisos y en menor tiempo. Además, las aproximaciones obtenidas por medio de metagenómica, nos dan más luces en la investigación a nivel poblacional, aplicándose diversas tecnologías como análisis filogenéticos (Babaloa 2009), estudios de especies con características novedosas (Lo Giudice 2007, Murray 1998, Wietz 2008, Yakimov 1999), análisis de expresión genética (Yergeau 2012), entre otros. En este estudio, se aplicó técnicas dependientes de ADN (PCR, DGGE fingerprinting y secuenciación 454) para el estudio de muestras de suelo colectado durante los veranos de los años 2010, 2011 y 2012 en las islas antárticas Greenwich, Dee y Barrientos. Con los resultados de estos ensayos, se determinó los principales grupos de hongos, levaduras y bacterias presentes, así como la distribución y diversidad entre las muestras analizadas. La recolección de las muestras se realizó en los mismos puntos para los tres años considerados. En la figura 1 se muestran los sitios de recolección.
Figure1: Sitios de muestreo.
Figure 1: Sampling sites. El procesamiento de estas muestras incluyó la siembra de microorganismos en medios de amplio espectro, establecimiento de la colección antártica CIBE-ESPOL y la identificación molecular de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
38
individuos predominantes. Para este último paso, se extrajo ADN individual, se amplificó por PCR las zonas semiconservadas usadas por excelencia para el análisis taxonómico (16SrRNA para bacterias e ITS para hongos y levaduras) y se secuenció los amplicones obtenidos para su comparación con la bases de datos de NCBI. La metodología utilizada incluye diferentes tipos de extracción (Dellaporta 1983) y protocolos de PCR diseñados por Woese et al. (1977) y Gardes (1993). Los taxones predominantes de hongos fueron Mortierella pulchella, M. fymbricystis, M. parvispora, Antarctomyces psychrotrophicus, Geomyces spp., Variscosporium elodeae. En el caso de levaduras, se encontró principalmente Cryptoccocus gasrticus, C. gilvescens y Rhodotorula glacialis. Mientras que las bacterias pertenecieron en su mayoría a Pseudomonas antarctica., P. fluorescens, Arthrobacter ramosus, y Rhodococcus sp.
Nombre Secuencia Aplicación ITS 1 TCCGTAGGTGAACCTGCG
Identificación de aislados (hongos y levaduras)
ITS 5 GGAAGTAAAAGTCGTAACAAGG
ITS 4 TCCTCCGCTTATTGATATGC
968F AACGCGAAGAACCTTAC Identificación de aislados (bacterias) R1378 CGGTGTGTACAAGGCCCGGGAACG
27F AGAGTTTGATCMTGGCTCAG
DGGE 1492R TACGGCTACCTTGTTACGACTT
968F-GC CGCCCGGGGCGCGCCCCGGGCGGGGCGGGAACGCGAAGAACCTTAC
4F968 CGTATCGCCTCCCTCGCGCCATCAGACGAGTGCGT*AACGCGAAGAACCTTAC
454 41378R CTATGCGCCTTGCCAGCCCGCTCAGCGTGTCTCTA*CGGTGTGTACAAGGCCCGGGAACG
4ITS1 CGTATCGCCTCCCTCGCGCCATCAGACGAGTGCGT*TCCGTAGGTGAACCTGCG
4ITS4 CTATGCGCCTTGCCAGCCCGCTCAGCGTGTCTCTA*TCCTCCGCTTATTGATATGC
Tabla 1: Iniciadores usados en las distintas técnicas aplicadas para el presente estudio. (*) indica región variable en el diseño de iniciadores para secuenciación 454 (Life Sciences Corp. 2011).
Table 1: Primer pairs used in each of the techniques applied in this study. (*) mean variable region in the design of 454 primers (Life Sciences Corp. 2011).
A nivel de poblaciones, se aisló el ADN total de las muestras por métodos comerciales y se amplificaron las regiones anteriormente mencionadas con protocoles específicos para las subsiguientes aplicaciones (DGGE y 454). Se realizó electroforesis en geles de gradiente desnaturalizante (DGGE) en las distintas poblaciones, cuyos datos fueron usados para determinación de diversidad (Índice de Shannon – Wiener “H”). Como resultado, la muestra más diversa perteneció a la zona de guano de pingüino (H: 258.11) y la de menor diversidad se localizó cerca al deshielo del glacial Quito (H: 146.61). Con los datos obtenidos por medio de secuenciación 454, analizados con el software Quiime (Sutton 2013), se conoció las principales familias de bacterias: Lactobacillaceae, Xanthomonadaceae, Chlorobiaceae, Comamonadaceae, Rhodobacteraceae, Pseudomonadaceae, Micrococcaceae, Moraxellaceae y Oxalobacteraceae. Además se realizó Análisis de Componentes Principales (PCA), el cual reveló agrupamiento de muestras acorde al año de muestreo. Las Unidades Taxonómicas Operacionales (OTUs) responsables de este agrupamientos fueron Psychromonas and Microcoleus, así como las OTUs de las familias

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
39
Rhodobacteraceae y Chthoniobacteraceae. Otros OTUs que influyen en esta distribución no han sido caracterizados aún. Muestras de sitios especiales, mostrando presencia de biofilm (BF) y contaminación de hidrocarburos (HC), formaron un grupo separado en el PCA. Los OTUs principales en HC fueron Acidimicrobiales y Actinobacteriales. En la muestra BF se encontraron mayormente Acidobacteria. Hubo además un alto porcentaje de familias no caracterizadas, indicando un alto potencial para exploración de nuevos grupos taxonómicos que pueden ser de interés científico e industrial. AGRADECIMIENTOS Los autores agradecen el auspicio de ESPOL, INAE y SENESCYT para el desarrollo de esta investigación. REFERENCIAS BIBLIOGRÁFICAS 454 Life Sciences Corp. 2011. 454 Sequencing System Guidelines for Amplicon Experimental Design. Babalola, O. O., Kirby, B. M., Le Roes-Hill, M., Cook, A. E., Cary, S. C., Burton, S. G., et al. 2009. Phylogenetic analysis of actinobacterial populations associated with Antarctic Dry Valley mineral soils. Environmental Microbiology 11(3):566-76. Dellaporta, S. L., Wood, J, y Hicks, J. B. 1983. A DNA minipreparation: version II. Plant Molecular Biology Reports 1:19-21. Gardes, M., y Bruns, T. D. 1993. ITS primers with enhanced specificity for basidiomycetes--application to the identification of mycorrhizae and rusts. Molecular Ecology 2(2):113-8. Lo Giudice, A., Bruni, V., y Michaud, L. 2007. Characterization of Antarctic psychrotrophic bacteria with antibacterial activities against terrestrial microorganisms. Journal of Basic Microbiology, Dec;47(6):496-505. Murray, A. E., Preston, C. M., Massana, R., Taylor, L. T., Blakis, A., Wu, K., et al. 1998. Seasonal and Spatial Variability of Bacterial and Archaeal Assemblages in the Coastal Waters near Anvers Island, Antarctica. Applied and Environmental Microbiology, 64(7):2585-95. Pearce, D., Newsham, K., Thorne, M., Calvo-Bado, L., Krsek, M., Laskaris, P., Hodson, A. y Wellington, E. 2012. Metagenomic analysis of a southern maritime Antarctic soil. Front. Microbiol., doi: 10.3389/fmicb.2012.00403. Sutton, N. B., Maphosa, F., Morillo, J. A., Al-Soud, W. A., Langenhoff, A., Grotenhuis, T., Rijnaarts, H., y Smidt, H. 2013. Impact of Long-Term Diesel Contamination on Soil Microbial Community Structure. Applied Environmental Microbiology. 79:2 619-630. Wietz, M., Månsson, M., Bowman, J. S., Blom, N., N. Y y Gram, L. 2012. Wide Distribution of Closely Related, Antibiotic-Producing Arthrobacter Strains throughout the Arctic Ocean. Applied and Environmental Microbiology, 78(6):2039-42. Woese, C.R., Fox, G. E. 1977. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proceedings of the National Academy of Sciences, 74(11):5088-90. Yakimov MM, Giuliano L, Bruni V, Scarfi S, Golyshin PN. 1999. Characterization of antarctic hydrocarbon-degrading bacteria capable of producing bioemulsifiers. The new microbiologica, Jul;22(3):249-56. Yergeau, E., Sanschagrin, S., Beaumier, D. y Greer, C. W. 2012. Metagenomic Analysis of the Bioremediation of Diesel-Contaminated Canadian High Arctic Soils. PLoS ONE, 7(1):e30058.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
40
ECOFISIOLOGÍA DE PLANTAS ANTÁRTICAS: UN NUEVO PORTAL WEB (Ecophysiology of Antarctic Plants: a new web page)
Corcuera LJ1, Sáez P2 y Bravo LA3
1Departamento de Botánica, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile 2Departamento de Silvicultura, Facultad de Ciencias Forestales, Universidad de Concepción, Concepción, Chile;
3Facultad de Ciencias Agropecuarias y Forestales, Universidad de La Frontera, Temuco, Chile email: [email protected]; [email protected]; [email protected] El proyecto Anillo de Investigación en Ciencia Antártica, Programa de Investigación Asociativa CONICYT-INACH ART1102 es una iniciativa de investigación multidisciplinaria, tri-institucional, cuyo objetivo es esclarecer las bases biológicas que explican la expansión de las plantas vasculares en Antártica Marítima en el actual escenario de calentamiento global. El Proyecto está oficialmente administrado por la Universidad de La Frontera, donde participan León A. Bravo (Director), Mirén Alberdi (Sub-directora) y Marjorie Reyes (Investigadora Asociada). Además, incluye un grupo de investigadores de la Universidad de Concepción: Luis Corcuera (Investigador Principal) Lohengrin Cavieres (Investigador Asociado), Angela Sierra y Patricia Saez (Investigadoras Asociadas) y Nicole Trafault del Centro de Genómica y Bioinformática de la Universidad Mayor. Este grupo de investigadores nacionales cuenta con la colaboración de investigadores de tres instituciones internacionales (Marilyn Ball, Australian National University; Norman Hüner, University of Western Ontario; Leopoldo García Sancho, Universidad Complutense de Madrid). La línea principal de investigación es la ecofisiología de las plantas antárticas. La aproximación de este grupo de investigadores es netamente experimental, se someterá a calentamiento a las plantas directamente en terreno utilizando unas cámaras denominadas OTC (del inglés open top chambers). Además, se realizarán estudios en cámaras de cultivo y biotrón. Principalmente se busca comprender si la temperatura en forma directa modifica el balance de carbono aumentando la productividad y crecimiento de las plantas o alternativamente si el aumento de temperatura ejerce un efecto indirecto. Por ejemplo, modificando las comunidades bacterianas y la actividad de estas en el reciclaje de nutrientes, aumentando la disponibilidad de nutrientes del suelo y con ello favoreciendo la nutrición y el crecimiento de las plantas. El Anillo de Investigación en Ecofisiología de Plantas Antárticas ha construido un nuevo portal web ubicado en la dirección: www.antarcticplantscience.cl en castellano e inglés. El portal contará con información relevante acerca del grupo de investigación, sus principales contribuciones a la Ecofisiología de las plantas vasculares antárticas y las actividades de difusión hacia la comunidad educativa y público en general. Tendremos un boletín electrónico que será despachado periódicamente por por e-mail y un muro de Facebook en la dirección Antarctic Plant. Esperamos de esta manera, dar a conocer nuestro quehacer a todo aquel que esté interesado en seguirnos y colaborar en la consecución de los propósitos de este interesante proyecto. Además de la investigación, nos interesa contagiar de un espíritu científico a los jóvenes de la comunidad educativa y al público en general. Agradecimientos: proyecto Anillo de Investigación en Ciencia Antártica, Programa de Investigación Asociativa CONICYT-INACH ART1102.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
41
PATRONES DE ESTRUCTURACION POBLACIONAL A DOS ESCALAS GEOGRAFICAS DEL ALGA ROJA, GIGARTINA SKOTTSBERGII.
(Patterns of populations structure of a red algae, Gigartina skottsbergii at two geographic scale)
Reyes, J1, Mancilla, A2., Faugeron, S3. y M-L. Guillemin1
1Instituto de Cs. Ambientales y Evolutivas, Universidad Austral de Chile, Valdivia, Chile. [email protected] 2Depto. de Cs. y Rec. Naturales, Facultad de Ciencias, Universidad de Magallanes, Punta Arenas, Chile. 3Departamento de Ecología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Santiago, Chile. INTRODUCCIÓN La distribución de las especies y de los linajes genéticos está estrechamente relacionada con eventos históricos y características ecológicas contemporáneas. Eventos de vicarianza, extinción y dispersión explicarían en gran parte los patrones de distribución de las especies (Avise 2009). El Hemisferio Sur ha tenido una compleja historia geológica y climática que afectó los patrones de distribución de muchos organismos. La historia evolutiva de algunas especies probablemente comenzó en el Cretácico inferior-medio hace aproximadamente unos 157-130 Ma (Crame 1999). Este periodo corresponde a la separación del súper-continente Gondwana, que posteriormente siguió con la apertura del Paso Drake y la separación geográfica de la península Antártica y del continente Sudamericano hace 40 millones de años (Scher y Martin, 2006). Durante este periodo se formó también la Corriente Circumpolar Antártica (CCA) que representa hoy en día el principal factor de aislamiento físico entre Antártica y las masas continentales cercanas y que además actúa como barrera fisiológica para la mayoría de las especies marinas (Thatje 2005). Otros eventos históricos climáticos importantes fueron los periodos glaciares y en particular el ultimo máximo glacial (Hewitt 2000). Durante este periodo las especies se vieron obligadas a restringir a zonas libres de hielo, llamados refugios glaciares (Hewitt 2000). En Chile, durante el UMG, la capa de hielo se extendió aproximadamente desde Chiloé (42ºS) hasta el Cabo de Hornos (56ºS) para la zona costera (Hulton 2002). Se ha demostrado que durante el UMG, muchas especies terrestres del centro-sur y sur de Chile y Argentina tuvieron una distribución restringida en refugios glaciares (Villagrán et al. 2004). En antártica también se comprobó la existencia de extinciones masivas de muchas poblaciones durante los periodos glaciares, sin embargo estudios recientes proveen evidencia de profunda divergencia entre especies antárticas y patagónicas (Fraser 2012; Gònzalez-wevar et al., 2012), indicando la existencia de refugios glaciares en Antártica. Gigartina skottsbergii es una macroalga roja haploide-diploide, perteneciente al orden de las Gigartinales. Es una especie submareal presente en Sur de Sudamérica, en las islas sub-Antárticas y en la península Antártica. Su distribución es altamente parchosa, con poblaciones generalmente de tamaño menor a 20km2 (Ramirez y Santelices 1991) y su capacidad de migración es muy baja. Se escogió a esta especie como modelo biológico no solo por su distribución antártica y sub-antártica, sino también porque pertenece a un orden antiguo de algas rojas cuyo origen está claramente identificado en el continente antártico, en la época en que este estaba unido a Australia (continente Australasia) y a Sudamérica (Hommersand et al. 1995).En nuestro estudio se infirió la historia evolutiva de Gigartina skottsbergii, desde la separación geográfica de las poblaciones continentales y antárticas, utilizando el marcador molecular cox 23. MATERIALES Y MÉTODOS Se utilizaron muestras de Gigartina skottsbergii de un total de 15 localidades pertenecientes a la costa Chilena, Malvinas e islas Antárticas (Fig. 1D). El DNA fue extraído usando el método de Saunders (1993), modificado por Faugeron et al. (2001).El marcador utilizado fue el cox2-3 que corresponde a una región mitocondrial no-codificante intergénica de 400 pb aproximadamente. Para la amplificación

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
42
se usaron los partidores pseudo-universales diseñados por Zuccarello (1999).Las reacciones de PCR se realizaron en un termociclador PerkinElmer Gene Amp PCR system 9700 (Applied Biosystems, Foster City, USA). Las muestras amplificadas de cada individuo fueron purificadas con el kit comercial UltraCleanTM (MO BIO Laboratories, Carlsbad, USA). Las muestras purificadas se enviaron a secuenciar a Macrogen Inc. (Seoul, Corea del Sur). Se utilizaron los programas Chromas v. 2.33 (McCarthy 1997) para visualizar las secuencias y Mega v 5 (Tamura et al. 2011) para alinearlas.Se construyó una red de haplotipos con el programa NETWORK v 4.510 (Bandelt et al. 1999) usando solo las muestras de la costa Chilena y Malvinas. Para estas mismas muestras, se calcularon los índices D de Tajima (Tajima 1989) y Fsde Fu (Fu 1997) para comprobar si estas poblaciones se alejan del equilibrio mutación/deriva mediante el programa ARLEQUIN V.3.5 (Excoffier y Lischer 2010). Para detectar la presencia de discontinuidades genéticas entre poblaciones de la costa chilena y de las Islas Malvinas se uso una aproximación Bayesiana de agrupamiento, desarrollada en el programa de genética del paisaje GENELAND v.3.14 (Guillot et al., 2005). Se realizó 50.000.000 iteraciones de Monte Carlo de cadena de Markov(MCMC) con un modelo de frecuencia correlacionado.Para los análisis filogenéticos se construyó un árbol de máximo verisimilitud con el programa Tree finder (Jobb et al 2004). El mejor modelo se escogió utilizando el criterio AIC (Akaike information criterion).Además se construyó un árbol de inferencia bayesiana mediante el programa MrBayes v3.2 (Huelsenbeck & Ronquist, 2001) empleando un método de MCMC. Se usó como grupo externo especies del orden de las Gigartinales cercanas que se encuentran en GENBANK. RESULTADOS Y DISCUSIÓN Para el marcador cox2-3 (350 pb) se secuenció un total de 215 individuos y se obtuvieron 166 secuencias para las poblaciones del continente incluyendo a islas Malvinas y 49 para las poblaciones de la región antártica (Fig. 1D).El árbol de máximo-verosimilitud muestra claramente la presencia de tres linajes de haplotipos genéticamente muy diferentes y sostenidos por altos valores de bootstrap(Fig 1A) para G. skottsbergii y este resultado concuerda con resultados obtenidos a través de marcador molecular rbcL (marcador cloroplastico, datos no mostrados). El primer grupo (Linaje1) corresponde a todos los individuos de la costa chilena continental y Malvinas, el segundo grupo (Linaje 2) corresponde 41 individuos de todas las islas antárticas muestreadas (HAN, PRA, AIT y ORK) y el tercer grupo (Linaje 3) comprende seis individuos de ORK y dos de HAN. La alta diferenciación entre poblaciones antárticas/subantárticas y las poblaciones del continente suramericano podría estar relacionada con el aislamiento geográfico de estas dos regiones desde la apertura del Paso Drake y la formación del CCA. Para los 166 individuos pertenecientes al continente (Linaje 1), existe un haplotipo compartido en todas las poblaciones (H1) que representa el 81% del total de muestras para el continente. La red de haplotipos para estos individuos presenta una típica forma en “estrella” característica de las poblaciones en expansión (Figura 1B) y los resultados de los índices de D de Tajima y Fs de Fu arrojaron valores negativas y significativas (-2,012 y -1,258, respectivamente) apoyando la idea de la existencia de un crecimiento poblacional para las poblaciones Chilenas. Este cambio demográfico podría estar relacionado a eventos post-glaciares de recolonización, un patrón que ya ha sido observado en otras especies de algas en la costa de Patagonia (Fraseret al. 2009, Montecinos et al. 2012). Los análisis espaciales de estructuración para poblaciones de la costa chilena y Malvinas en G. skottsbergii, basado en el programa GENELAND infirieron la existencia de cuatro grupos de poblaciones genéticamente distintas (1) Ancud, Isla Clarens, Bahia Chilota, Isla London, Bahia Orange y Puerto Toro; (2) Malvinas; (3) Bahia Inutil y Calbuco y (4) Puerto Aguirre y Bahia Low (Fig. 1C). La fuerte estructuracion existente entre las poblaciones del continente podria estar relacionado con la baja capacidad de dispersion del organismo en estudio y la existencia de poblaciones sumamente parchozas.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
43
Estos resultados apoyan a trabajos anteriores basados en marcadores nucleares (Faugeron et al. 2004, Martínez et al. 2005).
Figura 1: A) Árbol Filogenético para el marcador cox 2-3 (123 pb) en cada rama se muestran los valores de Maximo Verosimilitud y los valores de Inferencia Bayesiana. B) Red de haplotipos. para las 166 secuencias de G. skottsbergii
obtenidas en poblaciones de la Costa Chilena y Malvinas. Cada círculo representa un haplotipo y su tamaño es proporcional a su frecuencia en el set de datos. La distancia entre los haplotipos es proporcional a la distancia en pasos mutacionales. El
cuadrado negro indica una deleción de 12 pb. C) Mapa de probabilidad posterior entregado por el programa GENLAND de los individuos muestreados de la costa chilena y Malvinas, las zonas mas claras corresponde a la mayor probabilidad que una localidad pertenezca a un cluster, los numeros corresponden a las poblaciones que estan indicados entre parentesis en D. D)
Localidades muestreadas para la Costa Chilena, Malvinas e Islas Antarticas, con N= número de individuos secuenciados para este estudio.
Figure 1: A) Cox 2-3 (123pb) maximum likelihood (ML) phylogenetic tree, the boostrap values for the ML reconstruction and the support values for the bayesian inference are shown above each branch. B) Haplotype network including 166
sequences of G. skotssbergii from the Chilean coast and Falkland Islands. Each circle represents a different haplotype, circle size is proportional to the haplotype frequency in the data set and distance among haplotypes is proportional to the distance in
mutational steps. The black square represents a deletion of 12 pb. C) Map of posterior probability calculated in the GENELAND program using individuals from the Chilean coast and Falkland Islands. For each cluster, clear areas represent localities that are more likely to belong to this cluster. Each locality correspond to a black dot and the number noted close by
correspond to the one noted between parenthesis in D. D) Names of localities that where sampled along the Chilean coast, Falkland Islands and Antarctic Islands. N=Number individuals sequenced.
CONCLUSIONES Estos resultados muestran que los patrones de estructuración genética para G. skottsbergii en el cono sur podrían estar relacionados con el aislamiento geográfico entre las regiones muestreadas. Por otro lado, el patrón de crecimiento demográfico de las poblaciones patagónicas es posiblemente debido una recolonización postglacial en el sur de Chile. AGRADECIMIENTOS Proyecto INACH regular T16-11. REFERENCIAS BIBLIOGRÁFICAS Avise, J. C. 2009. Phylogeography: retrospect and prospect. Journal of Biogeography, 36:3–15

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
44
Bandelt, H.J., Forster P. y Röhl, A. 1999.Median-Joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol., 16:37-48. Crame, J.A. 1999. An evolutionary perspective on marine faunal connections between southernmost South America and Antarctica. Scientia Marina, 63(1):1–14. Excoffier, L. y Lisher, H. 2010. Arlequin suite ver 3.5. A new series of programs to perform population genetics analyses under Linux and Windows. Molecular EcologyResources,10:56. Faugeron, S., Martinez, E.A., Correa, J.A., Cárdenas, L., Destombe, C. y Valero, M. 2004. Reduced genetic diversity and increased population differentiation in peripheral and overharvested populations of Gigartina skottsbergii(Rhodophyta, Gigartinales) in southern Chile. Journal of Phycology. 40: 454–462. Fraser, C.I., Nikula, R..Ruzzante, D.E y Waters J.M. 2012. Poleward bound: biological impacts of Southern Hemisphere glaciation Trends in Ecology & Evolution. 27(8)462-471 Fraser, C.I., Hay, C.H., Spencer, H.G. y Waters, J.M. 2009. Genetic and morphological analyses of the southern bull kelp Durvillaea Antarctica (Durvillaeales) in New Zealand reveal cryptic species. Journal of Phycology, 45:436-443. Fu Y.X. 1997. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics, 147:915-925. González-Wevar, C., Hüne, M., Cañete, J., Mansilla, A., Nakano, T. y Poulin, E. 2012. Towards a model of postglacial biogeography in shallow marine species along the Patagonian Province: lessons from the limpet Nacella magellanica (Gmelin, 1791). BMC Evolutionary Biology. 12: 139 Guillot, G., Mortier, F. y Estoup, A. 2005. Geneland: a program for landscape genetics. Molec. Ecol. Notes. 5: 712-715. Huelsenbeck, J.P. y Ronquist, F. 2001. MRBAYES: Bayesian inference of phylogeny. Bioinformatics, 17:754–755. Hewitt G.M. 2000.The genetic legacy of the Quaternary ice ages. Nature. 405:907-913. Hommersand, M.H., Fredericq, S., y Freshwater, D.W. 1995. Phylogenetic systematics and biogeography of the Gigartinaceae (Gigartinales, Rhodophyta) based on sequence analysis of rbcL. Botanica Marina, 37:193-203. Hulton, N.R.J., Purves, R.S., McCulloch, R.D., Sugden, D.E. y Bentley, M.J. 2002. The Last Glacial Maximum and deglaciation in southern South America. Quaternary Science Reviews, 21:233-241. Martínez, E.A., Correa, J.A., Faugeron, S., Mansilla, A., Ávila, M. y Camus, P. 2005. Levantamiento demográfico y genético del alga roja GigartinaskottsbergiiSetchell et Gardner (Rhodophyta, Gigartinacea) a lo largo de su rango de distribución en el Pacífico Sur. Ciencia y Tecnología del Mar 28 (1): 63-74. Montecinos, A., Broitman, B.R., Faugeron, S., Haye, P.A., Tellier, F. y Guillemin, M-L. 2012. Species replacement along a linear coastal habitat: phylogeography and speciation in the red alga Mazzaella laminarioides along the South East Pacific. BMC Evolutionary Biology, 12(1), 97. McCarthy, C. 1997. Chromas Version 1.41. Griffith University, Brisbane, Queensland Ramírez, M.E., Santelices, B. 1991.Catálogo de las algas marinas bentónicas de la costa temperada del Pacífico de Sudamérica. Monografías Biológicas, 5:1-437. Scher, H.D. y Martin, E.E. 2006. Timing and climatic consequences of the opening of Drake Passage. Science 312(5772), 428-430. Thatje, S., Hillenbrand, C-D y Larter, R. 2005. On the origin of Antarctic marine benthic community structure. Trends in Ecology and Evolution, (10).534-540. Tajima, F. 1989. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics, 123:585-595. Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. y Kumar, S. 2011. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution, 28:2731-2739. Villagrán, C., León, A. y Roig, F.A. 2004. Paleodistribución del alerce y ciprés de las Guaitecas durante períodos interestadiales de la Glaciación Llanquihue: provincias de Llanquihue y Chiloé, Región de Los Lagos, Chile. Revista Geológica de Chile, 31:133-151. Zuccarello, G.C., Sandercock, B. y West, J.A. 2002. Diversity within red algal species: Variation in world-wide samples of Spyridia filamentosa (Ceramiaceae) and Murrayella periclados (Rhodomelaceae) using DNA markers and breeding studies. European Journal of Phycology 37: 403–417.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
45
PRIMER REGISTRO DE VARIACION DE LA ACTIVIDAD FOTOSINTÉTICA EN Desmarestia menziesii J. AGARDH (DESMARESTIALES, PHAEOPHYCEAE) A TRAVES DE
UN GRADIENTE LATITUDINAL EN LA PENÍNSULA ANTÁRTICA (First record of changes in photosynthetic activity of Desmarestia menziesii J. Agardh throughout a
latitudinal gradient in the Antarctic peninsula)
Mansilla A1,2, Ojeda J1,2*, Marambio J1, Rosenfeld S1,2, Murcia S1, Rendoll J1, Mendez F1 & Yokoya NS3
1Laboratorio de Macroalgas Antárticas y Subantárticas (LMAS), Casilla 113-D, Universidad de Magallanes, Punta Arenas, Chile.*[email protected] 2Instituto de Ecología & Biodiversidad, Casilla 653, Santiago, Chile. 3Instituto de Botânica, Secretaria de Estado do Meio Ambiente, São Paulo, C. Postal 68041, 04045-972, SP, Brasil. INTRODUCCIÓN La riqueza de especies de macroalgas marinas antárticas está representada por alrededor de 117 especies (Wiencke & Clayton 2002), y se caracteriza por el alto grado de endemismo de aproximadamente 33% de las especies (Wulff et al. 2009). En términos ecológicos las costas rocosas a lo largo de la península Antártica occidental están cubiertos por extensos grupos de macroalgas submareales, proporcionando el 70% de la cobertura del sustrato (Wulff et al. 2009), esta riqueza y biomasa disminuyen al aumentar la latitud (DeLaca y Lipps 1976). Comparativamente la península Antártica se diferencia de zonas templadas y árticas por la ausencia del orden Laminariales, no obstante, este nicho ecológico es ocupado por especies endémicas del orden de las Desmarestiales (Wiencke & Clayton 2002). Para este grupo, se encuentran referidas tres especies de Desmarestia: D. antarctica Moe & Silva, D. anceps Montagne y D. menziesii J. Agardh (Ramirez 2010). En zonas costeras rocosas del continente antártico D. menziesii es una de las especies que aportan mayor biomasa al ensamble de macroalgas, siendo su principal distribución batimétrica bajo la abrasión del hielo marino hasta unos 10 metros de profundidad (Wulff et al. 2009). En términos de distribución latitudinal los principales registros de alta biomasa están en las islas Shetland del Sur (e.g., Quartino et al. 2005). No obstante, a medida que aumenta la latitud las referencias poblacionales disminuyen, por ejemplo, Bowden (2005) en Rothera (67º34,5’) registra la presencia de algunos individuos de D. antarctica, pero no de D. menziesii. El presente estudio registra la presencia de poblaciones de D. menziesii alrededor de isla Adelaida (e.g., base Carvajal 67º46’). Para estas altas latitudes como la zona sur de isla Adelaida, el principal fenómeno descrito es la disminución de la riqueza de especies de macroalgas asociado a la reducida disponibilidad de luz (Moe y DeLaca 1976). La variación estacional en la duración del día en los ambientes polares es el principal factor que regula la actividad fotosintética de las algas marinas (Gómez 2001). En D. menziesii se ha descrito que las fases reproductivas tienen diferentes características fotosintéticas; las micro-fases, gametofitos y estadios tempranos de los esporofitos están mejor adaptados para usar niveles más bajos de luz que las plantas adultas, debido al mayor contenido de pigmentos, eficiencia fotosintética y muy bajos requerimientos de intensidad luminosa (Gómez 2001). Considerando que existe una variación latitudinal en las horas luz en la península antártica y que D. menziesii presenta estrategias estacionales fotosintéticas para persistir en zonas polares, la interrogante que surge es: ¿Si los individuos ya establecidos en altas latitudes presentan diferencias en la actividad fotosintética en comparación con individuos que habitan en latitudes menores de la península Antártica? Por lo tanto, el objetivo del presente estudio fue evaluar parámetros de la actividad fotosintética como la

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
46
tasa de transporte de electrones máxima ETR max, eficiencia fotosintética α y curvas de luz (fotosíntesis versus irradianza) evaluando posibles diferencias fotosintéticas latitudinales. MATERIALES Y MÉTODOS Las muestras de D. menziesii fueron colectadas durante la Expedición Antártica (ECA 49-INACH), realizada en febrero del 2013, donde fueron muestreados 13 sitios: i) bahía Fildes (S 62º12’; W 58º54’), ii) base O`Higgins (S 63º19’; W 57º53’), iii) base Prat (S 62º27’; W 59º44’), iv) isla Decepción (S 62º59’; W 60º35’), v) punta Spring (S 64º17’; W 61º05’), vi) islas Argentinas (S 65º14’; W 64º15’), vii) base Rothera (S 67º33’; W 68º07’), viii) base Carvajal (S 67º46’; W 68º52’), comprendidos entre el Archipiélago de las Shetland del Sur hasta Bahía Margarita (dentro del círculo polar antártico). Para la colecta de las muestras se realizó un buceo autónomo, entre los 5 y 7 metros de profundidad, extrayendo 7 individuos de D. menziesii por lugar. Posteriormente, se realizaron medidas de variables fotosintéticas utilizando el Diving-PAM (Fluorómetro de clorofila portable). Se desarrollaron curvas de luz usando 9 niveles de irradianza (desde 0 a 551 µmol fotones m-2 s-1), generando la base para la obtención de la tasa de transporte de electrones relativa (rETR) y el cálculo de los parámetros fotosintéticos mediante la ecuación de Platt et al. (1980): ETR máxima (ETR max) y eficiencia fotosintética (αETR). Finalmente para determinar diferencias estadísticas entre los sitios de estudios se aplicó un análisis de varianza de una vía (ANOVA) y el test Tukey post hoc. Todos los análisis se basaron en un 95% de confianza (p<0,05), para ello se utilizó el software STATISTICA 7.0. RESULTADOS Las comparaciones fotosintéticas de D. menziesii basados en la curva de luz obtenida para cada sitio (ver Figura 1), evidenciaron diferencias significativas en la tasa de transporte de electrones máxima (rETR max) (p<0,05) (ver Figura 2). Se observaron dos grupos: el primero conformado por altos valores de rETR max provenientes desde B. Fildes, B. O’Higgins, B. Prat, I. Decepción y Pta. Spring (Figura 2). El mayor valor promedio de rETR max se registró en Punta Spring con 20,9±7,1 µmol fotones m-2 s-1 y el mínimo de este grupo en isla Decepción con 17±4,2 µmol fotones m-2 s-1.
Figura 1: Curvas de luz (P-I) de la tasa de transporte de electrones relativa (rETR) versus curva de irradiancia (PAR) de
Desmarestia menziesii. Las flechas negras indican los sitios muestreados para D. menziesii. Las barras indican el nivel de desviación (n = 7, para los valores de rETR).
Figure 1: Curves of light (P-I) of the relative electron transport rate (rETR) vs., curves of irradiance (PAR) in Desmarestia menziesii. The black arrows indicates those sampled sites for D. menziesii. Barrs indicated the desviation level (n = 7, for
rETR values). El segundo grupo estuvo conformado por valores bajos de rETR max provenientes de sitios de altas latitudes como islas Argentinas, base Rothera y base Carvajal (Figura 2). El mayor valor promedio de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
47
rETR max de 10,9±3 µmol fotones m-2 s-1 y el mínimo de 9,4±1,7 µmol fotones m-2 s-1 se registró en el sitio Islas Argentinas. La eficiencia fotosintética (αETR) presentó diferencias significativas (p<0,05), específicamente en el sector de I. Argentinas comparándolas con B. Prat, I. Decepción y Pta. Spring. El menor αETR promedio se registró en I. Argentinas con 0,37±0,08 µmol fotones m-2 s-1 y el máximo αETR se observó en I. Decepción con 0,52±0,03 µmol fotones m-2 s-1.
Figura 2: Tasa de transporte de electrones máxima (ETR max) de Desmarestia menziesii por cada sitio. Se muestra el
promedio y la desviación estándar con un n= 7 individuos por sitio. Las letras distintas indican diferencias significativas entre los sitios estudiados.
Figure 2: Maximum electron transport rate (ETR max) of Desmarestia menziesii per each site. Its showed the average and standard desviation (n = 7 individuals per site). Different letters indicate significant differences among assessed sites.
DISCUSIÓN Las bajas demandas de luz para el crecimiento y fotosíntesis de las macroalgas, no solo posibilitan la sobrevivencia de macroalgas en escenarios luminosos estacionalmente cambiantes, sino que también posibilitan la expansión de sus límites de distribución batimétrica, como por ejemplo en D. menziesii (Gómez 2001). A su vez, esta especie es denominada una macroalga “season anticipator” estrategia que se basa en la capacidad de su aparato fotosintético para hacer uso de la irradiancia disponible en el aumento de horas luz disponible a finales de invierno y primavera (Gómez 2001). Probablemente estos mismos mecanismos eco-fisiológicos posibilitan la ocurrencia de poblaciones de D. menziesii en altas latitudes y permiten la colonización de nuevos sustratos desprovistos de hielo. Este estudio constituye la primera referencia de mediciones de parámetros fotosintéticos en un gradiente latitudinal en el continente antártico y donde se concluye que la actividad fotosintética medida por el parámetro de rETR max presenta una variación latitudinal disminuyendo hacia altas latitudes en la Península Antártica.
AGRADECIMIENTOS Proyecto FONDECYT 1110875, INACH, Armada de Chile y CONICYT (MEC 80100021). REFERENCIAS BIBLIOGRÁFICAS Bowden DA. 2005. Quantitative characterization of shallow marine benthic assemblages at Ryder Bay, Adelaide Island, Antarctica. Marine Biology 146: 1235–1249.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
48
DeLaca TE & JH Lipps. 1976. Shallow-water marine associations, Antarctic Peninsula. Antarctic Journal of the United States. 11: 12–20. Gómez I. 2001. Ecophysiology of Antarctic macroalgae: effects of environmental light conditions on photosynthetic metabolism. Revista Chilena de Historia Natural 74: 251-271. Moe RL & DeLaca TE. 1976. Occurrence of macroscopic algae along the Antarctic Peninsula. Antarctic Journal of the United States. 11: 20–24. Platt T, Gallegos CL & Harrison WG. 1980. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. Journal of Marine Research 38: 687-701. Quartino ML, Boraso de Zaixso AL & Zaixso H. 2005. Biological and environmental characterization of marine macroalgal assemblages in Potter Cove, South Shetland Islands, Antarctica. Botanica Marina 48: 187–197. Ramírez ME. 2010. Flora marina bentónica de la región austral de Sudamérica y la Antártica. Anales del Instituto de la Patagonia, Chile 38: 57-71. Wiencke C & Clayton MN. 2002. Antarctic seaweeds. In: (J.W. Wa¨ gele and J. Sieg, eds) Synopses of the Antarctic benthos 9. A.R.G. Gantner Verlag KG, Ruggell, Liechtenstein. pp. 1–239. Wulff A, Iken K, Quartino ML, Al-Handal A, Wiencke C & Clayton MN. 2009. Biodiversity, biogeography and zonation of marine benthic micro- and macroalgae in the Arctic and Antarctic. Botanica Marina 52: 491-507.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
49
LAS POBLACIONES DE COLOBANTHUS QUITENSIS PRESENTAN DIFERENCIACIONES MORFO- FISIOLÓGICAS QUE PODRÍAN EVIDENCIAR LA FORMACIÓN DE ECOTIPO
EN SU HÁBITAT. (Colobanthus quitensis populations have morpho-physiological differentiations which might evidence a
habitat ecotype formation)
Klagges, M.1,2 , Cordero, C.3, Cuba-Díaz M.1,2.
1Laboratorio de Biotecnología y Estudios Ambientales. 2Departamento de Ciencias y Tecnología Vegetal, Escuela de Ciencias y Tecnologías, Campus Los Ángeles, Universidad de Concepción, Los Ángeles, Chile. 3Laboratorio de Genética Molecular Vegetal, INTA, Santiago, Chile. e-mail: [email protected].
INTRODUCCIÓN Colobanthus quitensis (Kunth) Bartl. (Cariophylaceae) es una de las dos especies de plantas vasculares nativas que colonizaron las islas de la Antártida marítima. En condiciones naturales su reproducción es asexual y también puede autofecundarse; muestra una considerable variabilidad morfológica a lo largo de su amplia distribución, que se extiende desde México (17⁰N) al sur de la Península Antártica (68⁰S) y desde 0 a 4200 msnm (Moore, 1970; Smith, 2003). En el territorio antártico, esta especie, se encuentra expuesta a bajas temperaturas todo el año, fuertes vientos, radiación elevada, déficit hídrico, aerosoles de agua salada, disponibilidad limitante de nutrientes y cobertura de nieve durante prácticamente todo el año. Todo esto ha llevado a que estas plantas desarrollen mecanismos moleculares, bioquímicos y fisiológicos que les han permitido la sobrevivencia en su hábitat extremo (Cuba, 2011). En un estudio de diferenciación ecotípica en morfología y resistencia al frío en poblaciones de Colobanthus quitensis realizado por Gianoli et al. (2004), se ha logrado establecer dos ecotipos entre las poblaciones, Antártica y Andina, correspondientes a los sectores de la Isla Rey Jorge y el cerro La Parva (región metropolitana), respectivamente. En esta investigación se aplicaron diferentes marcadores morfo-fisiológicos para determinar la existencia de variaciones fenotípicas existentes entre cinco poblaciones incluyendo tres nuevas poblaciones a las ya analizadas en estudios anteriores.
MATERIALES Y MÉTODOS Las poblaciones analizadas corresponden a dos Andinas (La Parva (pPar) y Conguillio (pC)), una Costera de Punta Artenas (Marisma (pPA) y dos insulares (Arctowski (pA) y Punta Hannah (pH)). Dentro de las poblaciones continentales se encuentra la población del ecotipo andino colectada desde el cerro La Parva (pPar), ubicada bajo las coordenadas 33⁰19’S; 70⁰16’W a una altitud de 3600 m.s.n.m, la población pPA colectada desde Punta Arenas en la zona Marisma ubicada a 1 m.s.n.m bajo las coordenadas 52⁰22’S; 70⁰58’W y la población pC encontrada a 2575 m.s.n.m en el Parque Nacional Conguillio, región de la Araucanía, bajo las coordenadas 38⁰36’S; 71⁰36’W. En cuanto a las poblaciones insulares, ambas son provenientes de la Antártica Marítima. Una de ellas (pA) fue colectada desde la Isla Rey Jorge (62⁰09’S; 58⁰28’W), específicamente en una zona cercana a la estación de Arctowski ubicada a una altitud aproximada entre los 3-23 m.s.n.m, y la población correspondiente a Punta Hannah (pH) ubicada a 62⁰39’S; 60⁰37’W, ubicada en las isla Livingstone a 10 m.s.n.m. El material vegetal colectado desde terreno fue reproducido vegetativamente bajo condiciones de jardín común, donde las cinco poblaciones fueron separadas en la cámara de crecimiento con el objetivo de prevenir riesgos de retrocruzamiento. Las plantas fueron depositadas en potes plásticos con una mezcla de suelo:turba:vermiculita 3:2:1, y se mantuvieron a en cámara de crecimiento a una temperatura de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
50
15±2⁰C, con una densidad de flujo fotónico de 100-120 µmol m-2 s-1, bajo fotoperíodo correspondiente a 16/8 horas y humedad relativa entre 75-80%. Para realizar el análisis morfológico se utilizó un tamaño muestral de 150 plantas de Colobanthus quitensis de las distintas poblaciones; 30 de cada una, con este tamaño muestral se midió 7 parámetros cuantitativos, correspondientes a: ancho de hoja, largo de hoja, longitud de raíz principal, longitud del área foliar, forma de la hoja, diámetro del cuello y número de hojas. Para los 6 primeros parámetros se utilizó un calliper de resolución 0.5 mm. Además se realizaron 4 mediciones restantes: peso fresco (FW), el cual fue determinado utilizando una balanza analítica (Radwad) y el peso seco (DW), determinado después de secar las muestras por 4 días a 37⁰C hasta obtener un peso constante, posteriormente con estos datos se midió el contenido hídrico (WC), obtenido con la fórmula: WC= 100(FW-DW)/FW. Y finalmente se obtuvo el área foliar utilizando el software Sigmascan 5.0. Los datos obtenidos fueron analizados estadísticamente a través de un ANOVA de una vía, utilizando el software STATISTICA versión 7.0, seguido de un test de Tukey para detectar diferencias significativas entre las poblaciones. Finalmente, los valores medios de los diferentes parámetros de las cinco poblaciones fueron transformados a distancias Euclidianas utilizando el programa Statistica versión 7.0.
RESULTADOS En la Tabla 1, se indican los resultados obtenidos del análisis de varianza realizado a las once variables en las cinco poblaciones estudiadas, donde se muestra que existen diferencias significativas en la morfología y algunos parámetros fisiológicos de las plantas provenientes de las distintas localidades.
Parámetros Morfo-Fiosiolóficos pPar pA pPA pH pC
Número de hojas 17,33 ± 3,07ab
17,40 ± 1,81a
13,23 ± 1,89bc
11,83 ± 2,25c
16,37 ± 3,03ac
Largo de hoja (mm) 15,15 ± 5,43ab
14,21 ± 1,77a
21,24 ± 4,60bc
11,50 ± 1,17a
26,46 ± 3,39c
Ancho de hoja (mm) 1,10 ± 0,17ab
1,24 ± 0,09a
1,09 ± 0,19ab
1,12 ± 0,11a
0,91 ± 0,07b
Longitud de raíz (mm) 46,54 ± 14,27ab
41,34 ± 9,91a
51,16 ± 13,59bc
31,43 ± 14,46d
58,88 ± 9,68c
Longitud de área foliar (mm) 28,85 ± 8,04ab
22,50 ± 3,54a
37,14 ± 8,77bd
17,07 ± 2,02c
41,17 ± 4,42d
Diámetro de cuello (mm) 1,24 ± 0,33ab
1,37 ± 0,27ab
1,27 ± 0,22ab
1,22 ± 0,15a
1,63 ± 0,23c
Forma de hoja (largo/ancho) 13,61 ± 3,89ab
11,50 ± 1,61a
19,66 ± 4,04b
10,33 ± 1,27a
28,97 ± 2,95c
Área foliar (cm2) 0,26 ± 0,14bc
0,17 ± 0,03ab
0,26 ± 0,08c
0,11 ± 0,02a
0,14 ± 0,04a
Peso fresco (g) 0,44 ± 0,21c
0,13 ± 0,02ab
0,19 ± 0,04b 0,11 ± 0,03a 0,16 ± 0,06
ab
Peso seco (g) 0,04 ± 0,02b
0,01 ± 0,003a
0,02 ± 0,005ac
0,02 ± 0,003a
0,03 ± 0,01bc
Contenido hídrico (%) 91,66 ± 1,71a
88,72 ± 2,09ab
89,67 ± 3,71ab
85,05 ± 2,50bc
82,24 ± 1,55c
TABLA 1. Media, desviación estándar y letras de significancia (p < 0,05) de los parámetros morfo-fisiológicos evaluados en
las cinco poblaciones de Colobanthus quitensis. TABLE 1. Mean standard deviation and letters significance (p < 0,05) of morpho-physiological parameters evaluated in the
five Colobanthus quitensis populations.
De acuerdo con los datos obtenidos, pC presentó los mayores valores medios en largo de hoja (26,46 mm), longitud de raíz (58,88 mm), longitud de área foliar (41,17 mm), diámetro de cuello (1,63 mm) y forma de hoja (28,97), sin embargo, el ancho de hoja (0,91 mm) y contenido hídrico (82,24%) presentaron los valores medios mínimos en las cinco poblaciones estudiadas. Los valores mÁs bajos fueron obtenidos desde la población proveniente de Punta Hannah (pH), la cual presentó el menor largo de hoja (11,50 mm), longitud de raíz (31,43 mm), longitud de área foliar (17,07 mm), diámetro de cuello (1,22 mm), forma de hoja (10,33), área foliar (0,11 cm2) y peso fresco (0,11 g). A partir del análisis de distancias Euclidianas, con los valores obtenidos es posible establecer una relación o similitud morfológica entre las poblaciones, tal como se muestra en la fig. 1, donde el primer

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
51
cluster (o agrupamiento) esta establecido entre las poblaciones pPar y pPA, con una distancia Euclídea estimada en 12,0. El siguiente agrupamiento esta representado por las poblaciones pA y pH con una distancia Euclídea estimada en 12,2, y finalmente mucho más lejana a ambos agrupamientos de similitud se encuentra la población de pC, la cual presenta una distancia mayor con la población pH (44,0) y menor distancia con la población pPA (15,8).
T ree Diagram for 5 Variab les
Sing le Linkage
Eucl idean distances
11,5 12,0 12,5 13,0 13,5 14,0 14,5 15,0 15,5 16,0
L inkage Distance
pC
pH
pA
pPa
pPar
Figura 1. Agrupamiento de caracteres morfo-fisiológicos (cluster), utilizando distancias Euclidianas y acoplamiento completo
(complete linkage) como criterio de agrupamiento. Figure 1. Morpho-physiological characters cluster using Euclidean distances and complete linkage criterion.
DISCUSIÓN Se pudo comprobar alta variabilidad entre las poblaciones estudiadas, lo que coincide con lo informado por Moore (1970), de la existencia de una amplia variabilidad de la especie a través de su amplia distribución. De la misma manera Cordero (2012), observó una amplia variabilidad morfológica y genética al analizar estas poblaciones exceptuando pC. Esta variabilidad podría ser atribuida a una selección continua relacionada con la fragmentación geográfica del hábitat de la especie lo que fuerza un proceso de selección continua de los caracteres, lo cual puede ser adaptativo al geo-ecotipo, caracteres que se mantienen distintivos aún en condiciones de jardín común. De acuerdo a lo observado en Cordero (2012) y Cordero et al. (2012), se observó que pH fue la población con las menores medidas en la mayoría de los parámetros estudiados, a diferencia de los resultados obtenidos por estos mismos autores en esta estudio, la población pC se caracterizó por presentar la mayores medias, al menos en la mayoría de los parámetros medidos, excepto por el ancho de hojas y contenido hídrico, los que pueden estar directamente relacionados entre sí y con el hecho de que entre las 3 poblaciones con caracteres de mayor media fue la que menor número de hojas presentó (Tabla 1). Esta diferencias llevaron a que esta población, se mantuviera agrupada completamente separada del resto de las poblaciones continentales. La agrupación entre poblaciones insulares y las otras dos continentales (pPA y pPar), coincide con lo informado por Cordero (2012). En estudios previos se ha observado que las poblaciones de C. quitensis con distribución más al norte, al comparar individuos provenientes de la Parva (pPar) con individuos de la Antártica insular (pA), presentan los individuos de mayor tamaño, tanto en caracteres de morfología externa como de su micromorfología (Gianali et al., 2004; Bazcuñán-Godoy et al., 2010). Estos resultados son consistentes con los observados por Cordero (2012). Más estudios son requeridos, incluyendo estudios de variabilidad genética, donde se corrobore la diferenciación mostrada por la población pC. Cordero (2012) al realizar un test de Mantel donde incluyó variables genéticas y morfológicas, así como geográficas (distribución y altitud), observó que las variaciones morfológicas de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
52
las diferentes poblaciones, al parecer, tienen mayor relación con el ambiente que con la variabilidad genética de la especie, diferencias que se mantienen aún en condiciones de jardín común.
CONCLUSIONES Existe una amplia variabilidad morfológica entre poblaciones de C. quitensis, donde predomina que las poblaciones más al norte de su distribución presentan mayor tamaño. Existe una clara agrupación entre poblaciones insulares (Antárticas) y poblaciones continentales, exceptuando la población pC que se separa de ambos grupos.
AGRADECIMIENTOS Proyecto INACH T_03-09, a F. Ruíz y F. Ruminot por propagar material vegetal.
REFERENCIAS BIBLIOGRÁFICAS Bascuñán-Godoy, L., García-Plazaola, J., Bravo, L., y Corcuera, L. (2010). Leaf functional and micro-morphological photoprotective attributes in two ecotypes of Colobanthus quitensis from the Andes and Maritime Antarctic. Polar Biology 33:885–896. Cordero, C. 2012. Caracterización y análisis de variabilidad morfológica y genética en poblaciones de Colobanthus quitensis (Kunth) Bartl. (Caryophyllaceae). Memoria para optar el título de Ingeniero en Biotecnología Vegetal. Universidad de Concepción, Los Ángeles, Chile 52 pp. Cordero, C., Klagges, M., Opazo, O., Cuba-Díaz, M. 2012. Morphologial and genetic analysis suggests a low genertic flow presumably as a possible isolation by distance in Colobanthus quitensis (Kunth.) Bartl. populations. VII Reunión Biología Vegetal, Programa y Resúmenes p45. Cuba, M. (2011). El clavelito antártico y los mecanismos que lo protegen del frio polar. Boletín antártico chileno. Vol. 30. Nº 1. Gianoli, E., Zuñiga-Feest, A., Reyes-Diaz, M., Bravo, L., y Corcuera, L. (2004). Ecotypic differentiation in morphology and cold resistance in populations of Colobanthus quitensis from the Andes of central Chile and the maritime Antarctic. Arctic, Antarctic and Alpine Research 36: 470-475. Lewis-Smith R. I. L. (2003). The enigma of Colobanthus quitensis and Deschampsia antartica. In Huiskes, A . H. L., Gieskes, W. W. C., Rozema, J., Schoro, R. M. L., van der Vies, S. M., and Wolff, W. J. (eds.), Antarctic Biology in a Global Context. Leiden: Backhuys Publishers 2: 34-239. Moore, D.M. (1970). Studies in Colobanthus quitensis (Kunth) Bartl. and Deschampsia antarctica Desv. II. Taxonomy, distribution and relationships. Br Br Antarct Surv Bull 23: 63-80.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
53
EFECTO DE LA DESCARGA DE AGUA DULCE SOBRE LA ESTRUCTURA DE LA COMUNIDAD MICROBIANA EN EL FIORDO ADYACENTE AL GLACIAR JORGE
MONTT, PATAGONIA CHILENA (Effect of freshwater flux on microbial community structure in the fjord adjacent to glacier Jorge Montt,
Chilean Patagonia)
Gutiérrez, M.H.1, Galand, P.2, Pantoja, S. 1,3, Moffat, C.1,3
1Programa COPAS Sur- Austral, Universidad de Concepción, Casilla 160-C, Concepción, [email protected] 2UPMC University Paris 06 and CNRS, UMR 8222, LECOB, Observatoire Océanologique, F-66650, Banyuls/Mer, France 3Departamento de Oceanografía, Universidad de Concepción, Casilla 160-C, Concepción RESUMEN En las últimas décadas se ha evidenciado una acelerada pérdida en la cobertura de hielo en algunos glaciares Patagónicos (Rivera et al. 2012). Entre éstos, el glaciar Jorge Montt (uno de los más extensos del Campo de Hielo Patagónico Sur), ha experimentado un retroceso aproximado de 19.5 km desde 1898 a 2011 (Rivera et al. 2012). Esto trajo como consecuencia la apertura de una cuenca que alcanza en su parte más profunda hasta 400 m. Este sistema de fiordo, separado del Canal Baker por una elevación del fondo que resulta en profundidades de ~30m, está sujeto a la entrada de agua dulce en superficie (0-10m) y la entrada de agua más marina en aguas subsuperficiales y profundas (>10m-400m). Esto le confiere un patrón de circulación tipo estuarino, característico del sistema de fiordos y canales patagónicos (Silva et al. 1998). El significativo aporte de agua proveniente del glaciar tiene consecuencias no solo sobre las características hidrográficas y el patrón de circulación, sino también sobre las características químicas y biológicas del ecosistema adyacente. En este contexto, es sabido que el derretimiento glaciar puede representar un importante origen de materia orgánica (Stibal et al. 2012), así como de componentes microbianos (Bidle et al. 2007), por lo que puede impactar directamente la estructura y funcionamiento de la comunidad microbiana que habitan los ecosistemas aledaños. En ambientes fríos, se considera que los microorganismos dominan la biomasa y diversidad, así como la actividad metabólica (Morgan-Kiss et al. 2006; Miteva 2008). En particular, en ambientes supraglaciares los microorganismos participan activamente en la síntesis y procesamiento de materia orgánica, siendo responsables del intercambio de CO2 con la atmósfera y de la liberación de materia orgánica (Stibal et al. 2012) y posiblemente de componentes microbianos viables hacia los ecosistemas adyacentes (Bidle et al. 2007). En este contexto, en este trabajo describimos por primera vez la diversidad molecular de microorganismos (bacterias, arqueas y hongos) en un fiordo Patagónico y evidenciamos cambios espaciales y temporales en la estructura de la comunidad microbiana a lo largo de un gradiente salino influenciado por la descarga de agua dulce proveniente de un glaciar. El estudio fue realizado en el fiordo adyacente al glaciar Jorge Montt, ubicado en el extremo norte del Campo de Hielo Patagónico Sur (48º20’S; 73º30’ W). Durante el muestreo, realizado durante Otoño y fin de invierno de 2012, se colectaron muestras de agua superficial y de fondo a lo largo de un gradiente desde la cabeza a la boca del fiordo, y sedimentos superficiales dentro y fuera de la cuenca adyacente al glaciar. Alícuotas de 1 L de agua fueron filtradas por filtros de membrana estériles, los filtros y el sedimento superficial fueron almacenados a -20ºC. Adicionalmente, se realizó una caracterización de la condiciones hidrográficas y se obtuvo muestras para determinar la concentración de nutrientes, clorofila-a y abundancia microbiana. El ADN extraído del material particulado y el sedimento superficial fue enviado a Research and Testing Laboratory (Lobbock, TX, USA) para ser sometido a pirosecuenciación para el análisis de diversidad microbiana (“tag-encoded titanium amplicon pyrosequencing”; Dowd et al. 2008). La región 16S del rADN de arqueas y bacterias fue procesada utilizando los partidores 341F (5=-

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
54
GYG CASCAGKCGMGAAW-3=) y 958R (5=-GGA CTA CVS GGG TAT CTA AT-3=), y 28F (5=-GAG TTT GAT CNT GGC TCA G-3=) y 519R (5=-GTN TTA CNG CGG CKG CTG-3=), respectivamente. Para hongos, la región ITS (internal transcribed spacer) del gen 18S del rRNA fue analizada utilizando los partidores ITS1F (5=-CTT GGT CAT TTA GAG GAA GTA A-3=) y ITS4R (5=-TCC TCC GCT TAT TGA TAT GC-3=). El análisis bioinformático fue realizado utilizando una combinación del software pyrotagger (www.pyrotagger.jgi-psf.org) y el paquete de software Qiime (Caporaso et al. 2010). Nuestros resultados mostraron una columna de agua fuertemente estratificada durante ambos periodos, con una capa superficial de agua fría (0-4 ºC) y baja salinidad (<25 psu) de mayor espesor durante el otoño de 2012 y en la zona más cercana al frente del glaciar. Bajo la termoclina/haloclina se observa un aumento gradual de la temperatura y la salinidad hasta valores alrededor de los 8 ºC y >30 psu en aguas de fondo dentro de la cuenca del fiordo. Adicionalmente, se observan diferencias en las condiciones hidrográficas en el fiordo adyacente al glaciar y en el Canal Baker, lo que resulta en un gradiente de influencia del glaciar desde la cabeza a la boca del fiordo. Variaciones espaciales y temporales fueron también observadas en la concentración de nutrientes y clorofila-a y en la abundancia de microorganismos. Se detectó un total de 1443, 164 and 298 unidades operacionales taxonómicas (OTU, por su sigla en inglés) de bacterias, arqueas y hongos, respectivamente. En aguas al interior del fiordo, se observó una mayor abundancia de OTUs de bacteria en aguas de fondo que en aguas superficiales, mientras que para hongos la mayor riqueza de OTUs fue observada en aguas superficiales durante ambos periodos. A lo largo del gradiente horizontal en aguas superficiales, un mayor número de OTUs de bacteria fue observado en el Canal Baker. Contrario a esto, para hongos se observó un mayor número de OTUs en la zona cercana al frente del glaciar. Para estos dos grupos, la abundancia de OTUs fue más alta durante otoño que invierno, mientras que para arqueas las mayores abundancias de OTUs fueron detectadas durante invierno. La composición de la comunidad microbiana estuvo dominada por bacterias del grupo Proteobacteria y Cianobacteria, aqueas del grupo Thaumarchaeota y representantes de hongos basidiomicetes y ascomicetes. Diferencias fueron observadas en la composición de la comunidad microbiana entre aguas superficiales y de fondo, así como entre otoño y fines de invierno. El análisis de similitud realizado combinando OTUs de bacteria, arqueas y hongos evidenció la presencia de diferentes ensambles microbianos (Figura 1) representados por los siguientes grupos: i) una comunidad de microorganismos en aguas superficiales durante otoño, compuesta principalmente por cianobacterias, proteobacterias y actinobacterias (M1); ii) una comunidad en aguas de fondo durante invierno compuesta en su mayoría por arqueas, hongos basidiomicetes y representantes de proteobacterias (M2); iii) una comunidad en aguas profundas durante otoño dominada por hongos ascomicetes (M3); y iv) una comunidad en aguas superficiales durante invierno dominada por arqueas y basidiomicetes (M4). Resultados similares fueron obtenidos cuando se analizan las comunidades de bacterias y hongos individualmente.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
55
Figura 1. Análisis de cluster para el total de OTUs (bacterias, arqueas y hongos) y “heatmap” mostrando la distribución de los
principales OTUs en cada uno de los grupos definidos. Distribución de los principales grupos de microorganismos en cada una de las muestras analizadas (panel inferior). Ap2-3m corresponde a la muestra tomada en abril en la estación 2 (1 indica
más cercanía al glaciar) a 2 metros. Figure 1. Cluster analysis for the total OTUs (bacteria, archaea and fungi) and heatmap with the main OTUs in the defined
groups. Distribution of the main groups of microorganisms in each sample analyzed (bottom panel). Ap2-3m indicates sample from April at station 2 (1 is closets to the glacier) at 2 meters.
Nuestros resultados muestran diferentes comunidades de microorganismos habitando aguas superficiales y profundas, así como fluctuaciones estacionales en la estructura de la comunidad microbiana en el fiordo adyacente al glaciar Jorge Montt. Estas variaciones estuvieron asociadas a las características hidrográficas de la columna de agua, en donde la fuerte estratificación representaría una barrera para la mezcla de aguas superficiales y profundas. Sin embargo, es altamente probable que factores tales como cambios en las condiciones de luz y la disponibilidad de materia orgánica, así como cambios en la intensidad de la descarga de agua dulce proveniente del glaciar y el recambio de agua profunda al

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
56
interior de la cuenca también tengan influencia en estas variaciones. El análisis de la identidad taxonómica de los OTUs predominantes, muestra un alto grado de similitud con microorganismos identificados en otros ambientes de bajas temperaturas, lo que evidencia la prevalencia de una comunidad microbiana adaptada a las condiciones extremas observadas en este tipo de ecosistemas. Adicionalmente, diferencias observadas en la composición de microorganismos detectada al interior del fiordo y en las aguas del Canal Baker, sugieren que algunos componentes microbianos podrían estar siendo liberados como resultado de la descarga de agua de deshielo al ecosistema adyacente. Nuestros resultados sugieren que en un escenario de cambio climático, el aporte sostenido de agua dulce producto del retroceso glaciar podría representar una fuente de alteración significativa para el ecosistema de fiordos y canales Patagónicos, en donde la entrada de materia orgánica y componentes microbianos alóctonos podrían tener un fuerte impacto sobre los flujos de carbono y los ciclos biogeoquímicos. AGRADECIMIENTOS Al Laboratorio Internacional Asociado LIA MORFUN y Programa COPAS Sur-Austral y al proyecto Fondecyt 11110515. REFERENCIAS BIBLIOGRÁFICAS Bidle, K.D., Lee, S., Marchant, D.R. y Falkowsky, P.G. 2007. Fossil genes and microbes in the oldest ice on Earth. Proceding of the Natural Academy od Sciences, 104(33): 13455-13460 Dowd, S.E., Callaway, T.R., Wolcott, R.D., Sun, Y., McKeehan, T., Hagevoort, R.G. y Edrington, T.S. 2008. Evaluation of the bacterial diversity in the feces of cattle using16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiology, 8:125 Caporaso, J.G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F.D., Costello, E.K., Fierer, N., Pena, A.G., Goodrich, J.K. Gordon, J.I., Huttley, G.A., Kelley, S.T., Knights, D., Koenig, J.E., Ley, R.E., Lozupone, C.A., McDonald, D., Muegge, B.D., Pirrung, M., Reeder, J., Sevinsky, J.R., Turnbaugh, P.J., Walters, W.A., Widmann, J., Yatsunenko, T., Zaneveld, J., y Knight, R. 2010. QIIME allows analysis of high-throughput community sequencing data. Nature Methods; doi:10.1038/nmeth.f.303 Miteva, V.I. 2008. Bacteria in snow and glacier ice. En: Psychrophiles: from biodiversity to biotechnology. R. Margesin, F. Schinner, J.-C. Marx, C. Gerday (eds), Springer-Verlad , pp.31-50 Morgan-Kiss, R.M., Priscu, J.C., Pocock, T., Gudynaite-Savitch, L. Y Huner, N.P.A. 2006. Adaptation and aclimation of photosynthetic microorganisms to permanently cold enviroments. Microbiology and molecular biology reviews, 70(1): 22-252 Rivera, A., Corroipio, J., Bravo, C. y Cisternas, S. 2012. Glaciar Jorge Montt (Chilean Patagonia) dynamics derived from potos obtained by fixed cameras and satellite image feature tracking. Annals of Glaciology, 53(60): 147-155 Silva, N., C. Calvete & H. A. Sievers. 1998. Masas de agua y circulación general para algunos canales australes chilenos entre Puerto Montt y laguna San Rafael (Crucero CIMAR-Fiordo 1). Ciencia y Tecnología del Mar, 21: 17-48 Stibal, M., Sabacká, M. y Zarsky, J. 2012. Biological processes on glacier and ice sheet surfaces. Nature Geosciencies, 5: 771-774

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
57
ESTRUCTURA DE LAS COMUNIDADES MICROBIANAS PRESENTES EN SEDIMENTOS MARINOS COSTEROS EN TRES VERANOS AUSTRALES DE LA BAHIA DEL
ALMIRANTADO, ISLA REY JORGE, PENINSULA ANTARTICA (Microbial community structure found in coastal marine sediments during three austral summers
in Admiralty Bay, king George Island)
Franco, D.C1; Duarte, R. T1, Bendia, A.G1; Pellizari, V.H1*
1 Laboratório de Ecologia Microbiana, Instituto Oceanográfico, Universidade de São Paulo, Praça do Oceanográfico 191 CEP05588001, São Paulo, SP, Brasil. Email: [email protected] INTRODUÇÃO A diversidade microbiana na Península Antártica tem sido explorada em diferentes ecossistemas incluindo ambientes como solos, sedimentos, lagos, fontes hidrotermais, vulcões, geleiras, permafrost, etc. Pesquisas têm demonstrado que a fração de micro-organismos na biosfera bêntica pode constituir entre um décimo e um terço da biomassa total da terra e cerca de 70% da biomassa procariota total (PARKES et al, 1994; WHITMAN et al, 1998) A abundancia de células microbianas em sedimentos marinhos é correlacionada com as taxas de sedimentação e a distancia da costa. Densidade celular nos sedimentos superficiais aumenta em direção às margens continentais e declinam em direção ao mar aberto (KALLMEYER et al. 2012) Os sedimentos marinhos e sua microbiota apresentam papel fundamental na região Antártica, uma vez que atuam como indicadores das alterações ambientais que ocorrem em resposta a diferentes processos na coluna d’água. Vários estudos foram realizados em sedimentos marinhos de diferentes locais do planeta (Sedimentos marinhos dos oceanos ártico, atlântico, pacifico, assim como dos mares báltico e mediterrâneo) com o objetivo determinar a composição das comunidades microbianas, empregando técnicas de sequenciamento de nova geração, obtendo dados que permitem fazer analises filogenéticas e funcionais dos processos biogeoqumicos. As sequencias do domínio Bacteria descritas nesses trabalhos pertencem na sua grande maioria ao filo Proteobacteria, especialmente Gamma-Proteobacteria e Delta-Proteobacteria. Por sua vez, as sequencias do domínio Archaea pertencem ao grupo Euryarchaeota. (HAMDAN et al. 2012; ZINGER et al. 2011; LYRA et at. 2013; LI et al. 2009; DANOVARO et al. 2010; SOGIN et al. 2006). Nesse contexto, o objetivo deste trabalho foi estudar, através de sequenciamento de última geração, a variação da estrutura das comunidades microbianas presentes nos sedimentos marinhos costeiros da Baía do Almirantado, Ilha Rei George.
METODOLOGIA Amostras de sedimentos marinhos de quatro pontos (Botany Point, Ullman Point, Estação Antártica Comandante Ferraz e Refugio 2), compreendendo a Enseada Martel e a Enseada MacKellar, foram coletadas por meio do amostrador tipo Van Veen e congeladas a -20º C para análise posterior no Brasil. O DNA total das amostras foi extraído utilizando PowerSoil DNA Isolation Kit (MoBio, USA) e depois amplificado com os iniciadores contendo barcodes U519F e U1068R , que correspondem às regiões V4-V6 do gene 16S rRNA de E. coli. Os amplicons foram sequenciados utilizando 454 GS FLX Pyrosequencing (Roche)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
58
RESULTADOS Aproximadamente 150.000 sequencias de tamanhos variados foram obtidas (~90-570 pb). Sequencias de alta qualidade e não-quiméricas foram analisadas por meio do RDP classifier mostrando alta abundância de bactérias em relação a arquéias. Os filos mais abundantes incluem Proteobacteria, Firmicutes, Actinobacteria, Verrucomicrobia, Bacteroidetes e Planctomycetes. DISCUSSÃO E CONCLUSÕES PARCIAIS Análises preliminares demonstram uma mudança no perfil das comunidades em cada um dos pontos ao longo do tempo, sugerindo que diferentes aportes de substratos orgânicos e inorgânicos podem ter favorecido o estabelecimento de novos organismos na matriz sedimentar. Diversos relatos sobre a caracterização de sedimentos na Enseada MacKellar e Martel revelaram nos primeiros centímetros da coluna sedimentar, anoxia e abundância de pirita e calcopirita assim como uma alta concentração de matéria orgânica (SHAEFER et al, 2004), podendo apresentar condições ideais para o crescimento dos organismos com metabolismo heterotrófico fermentativo.
AGRADECIMENTOS CAPES, CNPq-PROANTAR, INCT-APA BIBLIOGRAFIA Danovaro, R.; Company, J. B.; Corinaldesi, C.; D’onghia, G.; Galil, B.; Gambi, C.; Gooday, A. J.; Lampadariou, N.; Luna, G. M.; Morigi, C.; Olu, K.; Polymenakou, P.; Ramirez-Llodra, E.; Sabbatini, A.; Sardà, F.; Sibuet, M. Y Tselepides, A. Deep-Sea biodiversity in the Mediterranean Sea: The known, the unknown, and the unknowable. In: PLoS ONE, 5(8): e11832. doi:10.1371/journal.pone.0011832, 2010. Hamdan, L. J.; Sikaroodi, M. Y Gillevet, P. M. Bacterial community composition and diversity in methane charged sediments revealed by Multitag Pyrosequencing. Geomicrobiology Journal, 29: 340-351, 2012 Kallmeyer, J.; Pockalny, R.; Adhikari, R. R.; Smith, D. C. Y D’hondt, S. Global distribution of microbial abundance and biomass in subseafloor sediment. Proceedings of the National Academy of Sciences USA, 109(40):16213–16216, 2012. Li, H.; Yu, Y.; Luo, W.; Zeng, Y. Y Chen, B. Bacterial diversity in surface sediments from the Pacific Arctic Ocean. Extremophiles : life under extreme conditions 13, 233–46, 2009. Lyra, C.; Sinkko, H.; Rantanen, M.; Paulin, L. Y Kotilainen, A. Sediment bacterial communities reflect the history of a Sea Basin. PLoS ONE 8(1) e54326. doi:10.1371/journal.pone.0054326, 2013. Parkes, R. J.; Cragg, B. A.; Bale, S. J.; Getliff, J. M.; Goodman, K.; Rochelle, P. A.; Fry, J. C.; Weightman, A. J. Y Harvey, S. M; Deep bacterial biosphere in Pacific Ocean sediments. Nature, 371:410–413, 1994. Schaefer, Cegr.; Dias, L.E.; Campos, L.S.; Albuquerque Filho M.R.; Costa L.M. Y Borges Júnior, M. Monitoramento ambiental em sedimentos costeiros da Baía do Almirantado:granulometria,teores de macronutrientes e metais biodisponíveis. In: Schaefer Cegr, Francelino Mr, Simas Fnb, Albuquerque Filho Mr (eds) Ecossistemas costeiros e monitoramento ambiental da Antártica Marítima, Baía do Almirantado, Ilha Rei George. NEPUT Universidade Federal de Viçosa,Viçosa, 192pp (2004) Sogin, M., et al. Microbial diversity in the deep sea and the underexplored "rare biosphere”. Proceedings of the National Academy of Sciences USA, 103(32), 2006. Whitman, W. B.; Coleman, D. C. Y Wiebe, W. J. Prokaryotes: the unseen majority. Proceedings of the National Academy of Sciences USA, 95:6578–6583,1998. Zinger, L.; Amaral-Zettler, L. A.; Fuhrman, J. A.; Horner-Devine, M. C.; Huse, S. M.; et al. Global Patterns of Bacterial Beta-Diversity in Seafloor and Seawater Ecosystems. PLoS ONE, 6(9): e24570. doi:10.1371/journal.pone.0024570, 2011

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
59
PRESENCIA DE INTEGRONES CLASE I EN COMUNIDADES BACTERIANAS TERRESTRES DE LA ISLA REY JORGE.
(Detection of integrons (intI gene) in terrestrial samples collected in King George island)
Antelo V.1, Batista S1.
1Unidad de Microbiología Molecular. Instituto de Investigaciones Biológicas Clemente Estable. Av. Italia 3318.Montevideo 11600. Uruguay. E-mail: [email protected]@gmail.com INTRODUCCIÓN Los integrones son elementos genéticos capaces de incorporar y expresar marcos de lectura abiertos (ORFs) exógenos (genes cassette) (Stokes et al., 1989). Están compuestos por tres elementos funcionales necesarios para la captura, escisión y expresión de genes cassette. El gen intI codifica para la enzima integrasa que cataliza la inserción de los genes cassette, un sitio de recombinación primario attI donde ocurre la inserción de estos genes y un promotor Pc que dirige la transcripción de los genes incorporados (Hall y Collis, 1995) (Figura 1). Estos elementos pueden adquirir determinantes de resistencia, virulencia y otras funciones metabólicas que le confieren a la bacteria hospedera una amplia versatilidad para adaptarse a nuevas condiciones de supervivencia (Di Conza et al., 2010). La relevancia de los integrones estuvo históricamente asociada a la resistencia a antibióticos por organismos patógenos. Sin embargo, la incorporación de nuevas estrategias de trabajo en ecología microbiana y el desarrollo de la secuenciación en gran escala permitió establecer que los integrones están ampliamente distribuidos en la naturaleza y no están confinados únicamente a bacterias patógenas y/o multi-resistentes, sino que forman parte de los mecanismos microbianos de adaptación al ambiente (Stokes et al., 2006; Xu et al., 2007).
Figura 1: Estructura básica de un integrón. Los genes cassette se insertan en el sitio de recombinación attI. El promotor Pc
controla la expresión de los genes insertados. Figure 1: Basic organization of an integron, showing the circular gene cassette inserted at the specific attI site, downstream of
promoter Pc. OBJETIVO Identificar y analizar la diversidad de genes que codifican para integrasas clase 1 mediante métodos dependientes e independientes del cultivo. MATERIALES Y MÉTODOS Muestras de suelo-sedimento y matas microbianas fueron colectadas en distintos sitios de la Península Fildes, durante las campañas estivales del 2008 y 2010. Una porción de cada muestra se suspendió en PBS 1x. Estas suspensiones se sembraron en placas de petri con medio LB, para generar una colección de organismos heterótrofos capaces de crecer a 5ºC y a 25ºC. El perfil de resistencia a antibióticos de los aislamientos se determinó en medio solido LB, utilizando concentraciones de 50 y 100 µg/ml. Para los ensayos con Trimetoprim se utilizó medio Mueller-Hinton agar.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
60
El ADN total y plasmídico de los aislamientos fue purificado usando DNeasy Blood & Tissue Kit (Qiagen, Germany) y Gene Plasmid Midprep Elute® HP (Sigma-Aldrich Co, MO) respectivamente. El ADN ambiental fue purificado mediante el método descripto previamente por Griffiths et al., (2000). La identidad de los aislamientos fue efectuada mediante análisis de secuencia de parte del gen 16S ARNr amplificado mediante PCR con los primers 27f y 1492r (Lane, 1991). Para determinar la presencia de genes integrasa clase 1 (intI), el ADN genómico de 32 aislamientos TrimR y el ADN ambiental total de 25 muestras, fue purificado y empleado como templado en reacciones de PCR utilizando los oligonucléotidos descriptos en el trabajo de Mazel et al. (2000) (intI1_fwd e intI1_rev). Para la construcción de la biblioteca de amplicones se utilizó el Kit de clonado TOPO TA Cloning®(Invitrogen, USA) de acuerdo al protocolo indicado por el fabricante. Para la clonación se utilizaron células competentes de Escherichia coli TOP10. Los productos fueron ligados con el vector pCR®2.1-TOPO® de Invitrogen. La secuenciación de ADN de los fragmentos amplificados y regiones adyacentes se realizó en el servicio de secuenciación del Instituto Pasteur (Montevideo, Uruguay) y en el Instituto Pasteur de París (Francia). Para el análisis de las secuencias se usaron los programas Blastn y Blastx http:/blast.ncbi.nlm.nih.gov/Blast.cgi) (Zhang et al., 2000). RESULTADOS Estrategia dependiente del cultivo Se obtuvo una colección de 250 aislamientos bacterianos capaces de crecer a 5ºC y 25°C. Setenta de ellos exhibieron altos niveles de resistencia a aminoglicósidos, β-lactámicos y trimetoprim. Se determinó la presencia de integrones clase l (intI1) en seis aislamientos TmR. Las secuencias nucleotídicas de los aislamientos fueron muy similares entre sí y las secuencias aminoacídicas deducidas exhibieron un 99% de identidad con las de integrasas de Acinetobacter baumanii (ZP07241546), Klebsiella pneumoniae (WP004162648) y Vibrio cholerae (EGS66892). En todos estos aislamientos, intI1 se localizó en un plásmido conjugativo con una organización genética similar a la descripta en los plásmidos pKOX105 de Klebsiella oxytoca, y p12 de Klebsiella pneumoniae 12, todas cepas de origen clínico (Carattoli et al., 2010; Gootz et al., 2009). Próximo al sitio attI del integrón, se encontró el gen dfrA14, conservado, que codificaría para la enzima dihidrofolato reductasa, que confiere resistencia a trimetoprim. El análisis de secuencia de parte del gen 16S ARNr indicó que todos los aislamientos positivos para el gen integrasa intI1 pertenecían al género Enterobacter.
Aislamiento Amp Km Tc Gm Cm Tm Nal Strp Spc 100 100 10 20 50 50 100 100 50
BA13 + - + - + + - + - CDTR5 + + - - + + - + -
IA12 + - - - + + - + - HTP19. + - - - + + - + -
TR3 + + - - + + + + - GC4 + - - - + + - + - CA4 + - - - + + - + - CN11 + - - - + + - + -
Tabla 1: Perfil de Resistencia antibiótica en aislamientos Trimr IntI+ Table 1: Antibiotic profile of Trimr IntI+ isolates.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
61
Estrategia independiente del cultivo Se construyó una biblioteca de 200 amplicones del gen intI1, habiéndose generado hasta el presente 29 secuencias de alta calidad, a partir de la muestra HTP19 (sitio vecino a Half-Three-Point). El 76% (n=22) de estas secuencias fueron idénticas a las secuencias intI de los seis aislamientos TrimR mencionados previamente. Siete secuencias de la biblioteca (intI) fueron distintas a las anteriormente mencionadas, aunque exhibieron similitud con las secuencias aminoacídicas de genes integrasas de bacterias no cultivables (Nemergut et al., 2004, Rodríguez-Minguela et al., 2009) (Figura 2). El análisis de las secuencias aminoacídicas obtenidas a partir de los clones de la biblioteca permitió identificar las regiones Box 1, Box 2, PatchII y Patch III en la mayoría de ellas. Además, las secuencias de los siete clones mencionados previamente, contenían un dominio adicional, previamente descrito por Rodríguez-Minguela et al. (2009), lo que sugiere que las mismas podrían corresponder a integrasas de una clase distinta a las ya descritas. Figura 2: Análisis filogenético utilizando el método de Neighbour-Joining basado en secuencias parciales IntI obtenidas en la Península Fildes. El porcentaje de réplicas (1000 bootstrap) > 50% se muestran en las ramas. Como grupo externo se utilizó
la secuencia XerC de E.coli FVEC 141. Figure 2: Consensus Neighbour-Joining tree constructed with partial IntI sequences from Fildes Peninsula. The tree was rooted using XerC from E. coli FVEC1412.Values represent the percentage of 1,000 bootstrap replica that supported the
branching order. Values of bootstrap support below 50% are not shown.
32
87
16
7
13
11
29
30
Klebsiella pneumoniae WP_004170180
AFR43808 integrase class 1
WP001222787 Acinetobacter baumanii
WP001979661 Vibrio cholerae
WP_019592869 Thioalkalivibrio sp. ALE18.
AF314190_1 integrase intI7
WP_004998506 Nitrococcus mobilis
WP_007808350 Rhodanobacter spathiphylli.
31
AF314191_1 integrase intI6
YP007081555 Pleurocapsa sp. PCC 7327
WP006562399 Oscillochloris tricoides
WP_009769460 XerD Oscillatoriales JSC-12.
BAM 62456 integron integrase unculture bacterium
ABD62687 uncultured bacterium.
8
BAM62447 integron integrase
83
YP_002763042 Gemmatimonas aurantiaca T-27.
AAP37603 integrase (intI)
26
28
27
57
xerC Escherichia coli FVEC1412.
73
65
100
99
100
0.1

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
62
CONCLUSIONES Se detectaron integrones clase 1 asociados a determinantes de resistencia a antibióticos y localizados en plásmidos conjugativos en enterobacterias psicrotolerantes, con una organización genética similar a la encontrada en cepas de origen clínico. Los resultados primarios obtenidos a partir del análisis metagenómico del gen intI permitieron establecer la presencia de genes intI idénticos a los caracterizados en los aislamientos TmR mencionados previamente, También se identificaron otros genes integrasa que pertenecerían a otro grupo diferente al de las integrasas de clase 1 descriptas clásicamente. AGRADECIMIENTOS Al Instituto Antártico Uruguayo, a la Agencia Nacional de Investigación e Innovación (ANII) y al Programa de Desarrollo de las Ciencias Básicas (PEDEClBA) REFERENCIAS Carattoli, A. Aschbacher, R..March A, Larcher C., Livermore D.M y Woodford N.. 2010. Complete nucleotide sequence of the IncN plasmid pKOX105 encoding VIM-1, QnrS1 and SHV-12 proteins in Enterobacteriaceae from Bolzano, Italy compared with IncN plasmids encoding KPC enzymes in the USA. Journal Antimicrobial and Chemotherapy 65 (10): 2070-2075. Di Conza, J.A. y Gutkind G.O. 2010. Integrones: los coleccionistas de genes. Revista Argentina de Microbiología 42: 63-78. Gootz, T.D, Lescoe M.K, Dib-Hajj F., Dougherty B.A., He W., Della-Latta P. y Huard R.C. 2009. Genetic organization of transposase regions surrounding blaKPC carbapenemase genes on plasmids from Klebsiella strains isolated in a New York City hospital. Antimicrobial Agents and Chemotherapy 53 (5): 1998-2004. Griffiths, R.I., Whiteley AS, O’Donnell A.G. y Bailey M.J. 2000. Rapid method for coextraction of DNA and RNA from natural environments for analysis of ribosomal DNA and rRNA-based microbial community composition. Applied Environmental Microbiology 66 (12): 5488-5491. Hall, R.M y Collis C.M. 1995. Mobile gene cassette and integrons: capture and spread of genes by site-specific recombination. Micro Review. Molecular Microbiology 15(4): 593-600. Lane. D.J 1991. 16S/23S rRNA sequencing. Nucleic Acid Techniques in Bacterial Systematics, eds Stackebrandt E, Goodfellow M (Wiley, New York), pp 115–175. Mazel, D. Dychincho B., Webb V. y Davies J. 2000. Antibiotic resistance in the ECOR collection: integrons and identification of a novel aad gene. Antimicrobial Agents and Chemotherapy 44 (6) 1568-1574. Nemergut D.R, Martin A.P y Schmidt S.K 2004. Integron diversity in heavy metal contaminated mine tailings and inferences about integron evolution. Applied and Environmental Microbiology 70 (2): 1160-1168. Rodriguez-Minguela C.M, Apajalahti J.H.A, Chai B. Cole J.R y Tiedje J.M. 2009. Worldwide prevalence of class 2 integrase outside the clinical setting is associated with human impact. Applied and Environmental Microbiology 75(15):5100-5110. Stokes, H.W. y Hall R.M.1989. A novel family of potentially mobile DNA elements encoding site-specific gene-integration functions: integrons. Molecular Microbiology 3: 1669-1683. Stokes, H.W., Nesbo C.L Holley M., Bahl M.I., Gillings M.R. y Boucher Y. 2006. Class 1 Integrons potentially predating the association with Tn402-Like transposition genes are present in a sediment microbial community. Journal of Bacteriology. 1888 (16): 5722-5730. Xu, H. J. Davies, y Miao V. 2007. Molecular characterization of class 3 integrons from Delftia spp. Journal of Bacteriology 189 (17): 6276-6283. Zhang Z. Schwartz S., Wagner L. y Miller W. 2000. A greedy algorithm for aligning DNA sequences. Journal Computational Biology 7(1-2): 203-14.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
63
DIVERSIDAD BACTERIANA EN SUELOS ANTÁRTICOS DESDE DIVERSOS ASENTAMIENTOS ANIMALES
(Bacterial diversity in Antarctic soils from different animal settlements)
Ramírez, L.1, Farías, F., Carú, M.1, Orlando, J.1
1Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile. [email protected] INTRODUCCIÓN En las áreas libres de hielo en la Antártida existen diferentes colonias de animales marinos. Tanto aves como mamíferos juegan un rol importante en el ciclaje de nitrógeno y carbono transfiriendo nutrientes desde el ambiente marino al ambiente terrestre (Tatur, 2002; Zdanowski y col., 2005; Cannone y col., 2008). Los suelos ricos en deposiciones animales generalmente poseen un alto contenido de carbono orgánico, nitrógeno y fósforo total, siendo éstos mayores en suelo con guano de ave en comparación con los suelos de asentamientos de mamíferos (Chong y col., 2009; Zhu y col., 2009). Estas condiciones sustentan una alta diversidad y actividad microbiana y la diferencia en la composición química podría diferenciar las comunidades bacterianas presentes en los diferentes ambientes. En este contexto, se han explorado diversos factores edáficos que podrían afectar el comportamiento y la estructura de los microorganismos como el pH (Fierer y Jackson, 2006), la disponibilidad de amonio (Avrahami y col., 2002) y la relación C/N (Zhang y col., 2006), entre otros. En este trabajo se estudió la diversidad de la comunidad bacteriana en diferentes asentamientos animales en la Isla Livingston, Islas Shetland del Sur. En particular, se analizaron asentamientos de aves (pingüinos y gaviotas) y de mamíferos (lobos y elefantes marinos) presentes en el Cabo Shirreff (ZAEP 149). Además, se caracterizaron las propiedades fisicoquímicas de los diferentes suelos y se relacionó la diversidad bacteriana con variables como cantidad de nitrógeno disponible, contenido hídrico, materia orgánica y pH. MATERIALES Y MÉTODOS Recolección de las muestras de suelo. El sitio de recolección de las muestras de suelo se ubicó en el Cabo Shirreff (62°27’30"S, 60°47’17"O), Isla Livingston, Islas Shetland del Sur, Antártida. En esta zona se han descrito diferentes tipos de asentamientos animales (ZAEP 149), entre ellos pingüinos (Pygoscelis antarctica y P. papua), elefantes marinos (Mirounga leonina), lobos marinos (Arctocephalus gazella), y aves como la gaviota cocinera (Larus dominicanus). De cada uno de estos asentamientos animales se tomaron 3 muestras de suelo a una profundidad de 10 cm y se nombraron “Pingüineras Barbijo”, “Pingüineras Papúa”, “Elefanteras”, “Loberas” y “Pajareras”, respectivamente. Además, se recolectaron 3 muestras de suelo “Controles” sin presencia de asentamientos animales y sin impacto visible de deposiciones de animales. Las muestras posteriormente se almacenaron a 4°C hasta su análisis. Medición de variables edáficas. El contenido hídrico (CH) del suelo se determinó gravimétricamente (Steubing y col., 2002). Brevemente, aproximadamente 1 g de suelo (peso húmedo; PH) se secó a 65°C durante 24 h para determinar el peso seco (PS) y el contenido hídrico se calculó según la fórmula: CH = PH-PS/PS*100. El contenido de materia orgánica (MO) se determinó a partir de las muestras secas, las cuales se calcinaron a 400ºC durante 16 h y se determinó el peso calcinado (PC) (Steubing y col., 2002). El porcentaje de MO se calculó según la fórmula: MO = PS-PC/PC*100. El pH se determinó potenciométricamente a partir de extractos en agua (Steubing y col., 2002). El contenido de nitrato (µg NO3
- / g PS) se determinó colorimétricamente (Yang y col., 1998), mientras que el contenido de amonio (µg NH4
+ / g PS) se determinó utilizando un electrodo ion selectivo.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
64
Extracción de DNA y amplificación por PCR. A partir de 0,25 g de cada muestra de suelo se extrajo el DNA empleando el kit “Power Soil DNA Isolation” (MoBio Lab., Inc.). Posteriormente, se amplificó la región variable V1-V3 del gen del rRNA 16S utilizando los partidores A17, marcado en su extremo 5’ con el fluoróforo FAM (6-carboxyfluoresceína), y 519R (Kumar y col., 2011). Perfiles de TRFLP (polimorfismo en el largo de los fragmentos de restricción terminal). Los amplicones de la región variable del gen del rRNA 16S marcados con fluorescencia se purificaron mediante el kit “UltraClean PCR Clean-up” (MoBio Lab., Inc.). Posteriormente, 20-50 ng de los amplicones se digirieron con 20 U de la enzima AluI (Fermentas) durante 12 h a 37ºC. Para cada muestra se generó un electroferograma donde se determinó el número de fragmentos de restricción terminal (TRFs) de cada muestra (riqueza) y la fluorescencia de cada TRF (abundancia). Análisis de los perfiles de T-RFLP y su relación con las variables edáficas. La diversidad de TRFs de cada una de las muestras, como medida de la diversidad bacteriana, se determinó mediante el índice de diversidad de Shannon (Shannon, 1948). Para evaluar la influencia de las variables edáficas sobre la diversidad bacteriana, se realizó un análisis de correspondencia canónica (CCA; Ter Braak, 1986) utilizando el programa PAST v2.16 (Hammer y col., 2001). Análisis estadístico. Para establecer si hubo diferencias significativas entre los parámetros edáficos y el índice de diversidad de los diferentes asentamientos animales, se realizó un análisis de varianza (ANOVA) de una vía y un post-test de Tukey en el programa GraphPad v.5.00 (GraphPad Software, www.graphpad.com). RESULTADOS Y DISCUSIÓN Del análisis de los TRFs de las muestras de cada uno de los asentamientos, se determinó que las comunidades bacterianas poseen una diversidad similar entre ellas, concordante con lo encontrado por Aislabie y col. (2009). Sin embargo, se observó una menor diversidad de la comunidad bacteriana, aunque no significativa, en las muestras provenientes de ambas pingüineras (P. antarctica y P. papua), en las cuales a su vez se determinó un alto contenido de nitrógeno disponible y una mayor cantidad de materia orgánica (Tabla 1), lo que concuerda con lo encontrado en estudios previos (Chong y col., 2009, Zhu y col., 2009, Kim y col., 2012). Tabla 1. Valores de los índices de diversidad y los factores edáficos. Table 1. Values of diversity index and edaphic factors.
pH
CH
MO
Amonio
Nitrato
H'
Pajareras 4,8 ± 0,2 a 14,2 ± 3,0 a 9,4 ± 3,2 a 104,9 ± 70,0 a 1034,0 ± 370,7 abc 2,47 ± 0,12 a
Elefanteras 5,6 ± 0,7 a 11,3 ± 4,1 a 6,8 ± 1,7 a 662,3 ± 299,0 a 1330,0 ± 631,7 abc 2,50 ± 0,12 a
Loberas 4,0 ± 0,1 a 6,8 ± 3,9 a 13,3 ± 4,2 a 31,9 ± 17,2 a 261,1 ± 164,2 b 2,57 ± 0,15 a
Pingüineras Barbijo 4,8 ± 0,4 a 20,5 ± 5,0 a 21,1 ± 3,4 a 967,7 ± 258,3 b 2061,0 ± 428,0 c 1,73 ± 0,28 a
Pingüineras Papúa 4,7 ± 0,5 a 19,2 ± 2,6 a 32,4 ± 15,1 a 477,9 ± 78,5 ab 2142,0 ± 319,6 c 1,83 ± 0,28 a
Controles 4,9 ± 0,2 a 17,4 ± 2,8 a 9,7 ± 6,1 a 8,9 ± 4,4 a 219,8 ± 23,5 ab 2,60 ± 0,10 a
Se muestran los valores ± error estándar. Los valores seguidos de letras iguales en una columna no son diferentes significativamente de acuerdo al ANOVA de una vía seguido del post-test de Tukey. CH: contenido hídrico, MO: materia orgánica, H’: índice de diversidad de Shannon. Values ± standard errors are shown. Values followed of the same letters in a column are not significantly different according to one-way ANOVA followed by Tukey's post-test. CH: water content, MO: organic matter, H': Shannon diversity index.
Lo observado podría deberse a que las altas concentraciones de nitrógeno formarían hábitats específicos en donde sólo colonizarían algunos microorganismos altamente adaptados. Ramírez y col. (2012) describieron que en suelos suplementados con altas cantidades de nitrógeno disminuía la biomasa y la actividad microbiana, existiendo una mayor abundancia de Actinobacterias y Firmicutes mientras que disminuía la presencia de Acidobacterias y Verrucomicrobia. Además, Zhu y col. (2009) compararon

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
65
suelo antártico influenciado por guano de pingüinos con suelo de asentamientos de focas y encontraron que en los primeros existía una mayor emisión de N2O, CH4 y CO2, lo que sugiere que poblaciones específicas como las bacterias desnitrificadoras podrían estar altamente representadas. En el caso de las comunidades bacterianas presentes en los suelos de los demás asentamientos, se puede observar que existe una mayor diversidad bacteriana a pesar de contener una menor cantidad de materia orgánica y nitrógeno disponible, siendo esta tendencia más notoria en el caso de los suelos desde las loberas y los controles. Cabe destacar que el hecho de que las loberas sean más similares a los controles puede deberse al comportamiento menos sedentario de estos mamíferos en comparación al resto. En el análisis de correspondencia canónica (Figura 1), en el que ambos ejes explican más de un 65% de la varianza (eje 1 42,8%; eje 2 24,8%), se puede observar una separación de las muestras desde los diferentes asentamientos animales, siendo las muestras de suelo asociadas a las pingüineras las más diferentes según la distribución a lo largo del eje 1. Las variables que principalmente explican la separación de las muestras son la cantidad de nitrógeno, en forma de nitrato (71,4%) y amonio (74,1%), y la materia orgánica (MO; 73,5%).
Figura 1. Análisis de correspondencia canónica de las comunidades bacterianas de los diferentes asentamientos animales. MO (materia orgánica); CH (contenido hídrico). (Canonical correspondence analysis of the bacterial communities from the different animal settlements. MO (organic matter), CH (water content)). Las variables abióticas como el pH y el CH no tienen una gran influencia sobre la diversidad de las comunidades bacterianas encontradas, a pesar de que el pH es considerado a nivel global el mejor descriptor de la diversidad y riqueza de ensambles microbianos (Fierer y Jackson, 2006). Estos resultados confirman los datos obtenidos en el análisis de la diversidad de las comunidades bacterianas y además apoyan lo encontrado por Ganzert y col. (2011), en el cual se determinó que los parámetros que mayormente influyen en la estructura de las comunidades bacterianas en ambientes antárticos, son las cantidades de carbono y de nitrógeno. CONCLUSIONES Se puede predecir que en ambientes extremos con altos niveles de nitrógeno existirá una tendencia a una menor diversidad de microorganismos, actuando el nitrógeno como un factor de selección de bacterias metabólicamente adaptadas para sobrevivir en estas condiciones. Sin embargo, se requiere de técnicas más resolutivas para determinar cuáles son estos organismos y qué rol desempeñan en este ambiente.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
66
Además, la evidencia sugiere que variables como el pH y el CH no influirían mayormente en la diversidad microbiana presente en los diferentes asentamientos de animales antárticos evaluados. AGRADECIMIENTOS INACH - ECA 48 (Proyecto F_02-10). REFERENCIAS BIBLIOGRÁFICAS Aislabie, J., Jordan, S., Ayton, J., Klassen, J. L., Barker, G. M., y Turner S. 2009. Bacterial diversity associated with ornithogenic soil of the Ross Sea region, Antarctica. Canadian Journal of Microbiology, 55(1):21-36. Avrahami, S., Conrad, R. y Braker, G. 2002. Effect of soil ammonium concentration on N2O release and on the community structure of ammonia oxidizers and denitrifiers. App. and Environmental Microbiology. 68(11):5685-5692. Cannone, N., Wagner, D., Hubberten, H. W. y Guglielmin, M. 2008. Biotic and abiotic factors influencing soil properties across a latitudinal gradient in Victoria Land, Antarctica. Geoderma, 144(1-2):50-65. Chong, C. W., Dunn, M. J., Convey, P., Tan, G. Y. A., Wong, R. C. S., y Tan, I. K. P. 2009. Environmental influences on bacterial diversity of soils on Signy Island, maritime Antarctic. Polar Biology, 32(11):1571-1582. Fierer, N. y Jackson, R. 2006. The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America, 103(3):626-631. Ganzert, L., Lipski, A., Hubberten, H. W. y Wagner, D. 2011. The impact of different soil parameters on the community structure of dominant bacteria from nine different soils located on Livingston Island, South Shetland Archipelago, Antarctica. FEMS Microbiology Ecology, 76(3):476-491. Hammer, Ø., Harper, D. y Ryan, P. 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4:9. Kim, O. S., Chae, N., Lim, H. S., Cho, A., Kim, J. H., Hong, S. G., y Oh, J. 2012. Bacterial diversity in ornithogenic soils compared to mineral soils on King George Island, Antarctica. Journal of Microbiology, 50(6):1081-1085. Kumar, P., Brooker, M., Dowd, S., y Camerlengo, T. 2011. Target region selection is a critical determinant of community fingerprints generated by 16S pyrosequencing. PLoS ONE, 6(6): e20956. Ramírez, K., Craine, J. M. y Fierer, N. 2012. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Global Change Biology, 18(6):1918-1927. Steubing, L., Godoy, R. y Alberdi, M. 2002. Métodos de Ecología Vegetal. 1º Edición. Editorial Universitaria, Santiago, Chile. Tatur, A. 2002. Ornithogenic ecosystems in the Maritime Antarctic - formation, development and disintegration. In: Beyer, L., Bölter, M. (Eds.), Geoecology of Antarctic ice-free coastal landscapes. Ecological Studies 154, Springer, Berlin, pp. 161-184. Ter Braak, C. J. F. 1986. Canonical Correspondence Analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology, 67(5):1167-1179. Yang, J. E., Skogley, E. O., Schaff , B. E. y Kim J. J. 1998. Simple spectrophotometric determination of nitrate in water, resin and soil extracts. Soil Science Society of America, 62(4):1108-1115. ZAEP 149. 2005. Plan de Gestión de la Zona Antártica Especialmente Protegida Nº 149: Cabo Shirreff e Isla San Telmo, Isla Livingston, Islas Shetland del Sur. XXVIII Reunión Consultiva del Tratado Antártico. Zdanowski, M. K., Zmunda, M. J. y Zwolska, I. 2005. Bacterical role in the decomposition of marine-derived material (penguin guano) in the terrestrial maritime Antarctic. Soil Biology and Biochemistry, 37(3):581-595. Zhang, Y., Li D., Wang H., Xiao Q., y Liu X. 2006. The diversity of denitrifying bacteria in the alpine meadow soil of Sanjiangyuan natural reserve in Tibet Plateau. Chinese Science Bulletin, 51(10):1245-1254. Zhu R., Liu Y., Ma, E., Sun, J., Xu, H. y Sun L. 2009. Nutrient compositions and potential greenhouse gas production in penguin guano, ornithogenic soils and seal colony soils in coastal Antarctica. Antartic Science, 21(5):427-438.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
67
ESTRÉS OXIDATIVO ENPINGÜINOS PAPÚA Y ADELIA EN ZONAS DE BAJO Y ALTO IMPACTO ANTRÓPICO DE BAHIA ESPERANZA
(Oxidative stress inGentoo and Adelie penguins in areas of low and high anthropogenic impact at Hope Bay)
Cebuhar J. D. 1, Di Fonzo, C. I. 1, Libertelli, M. 1, Zappala C. M. 1y M. Ansaldo1,2
1Instituto Antártico Argentino, Dirección Nacional del Antártico. Cerrito 1248, (C1010AAZ) Ciudad Autónoma de Buenos Aires, Argentina. 2Laboratorio de Ecotoxicología. Universidad Argentina JF Kennedy. Sarmiento 4564, (C1197AAR) Ciudad Autónoma de Buenos Aires, Argentina. Email: [email protected], [email protected] INTRODUCCIÓN Los pingüinos antárticos se encuentran expuestos a condiciones ambientales severas, temperaturas extremadamente bajas y elevada irradiación UV, que pueden incrementar el estrés oxidativo. La generación de oxi-radicales endógenos puede ser influenciada por diferentes factores ambientales y biológicos. En general, la eficiencia basal del sistema antioxidante refleja la presión normal de las especies reactivas del oxígeno (ROS) a las que están expuestos los organismos (Corsolini y otros, 2001). Se ha observado que los pingüinos poseen una mayor capacidad para atrapar los radicales del oxígeno, que el analizado en otras especies de aves antárticas. Esta mayor resistencia puede sugerir que los pingüinos están naturalmente expuestos a una mayor presión basal de ROS en comparación con las otras aves antárticas estudiadas (Corsolini y otros, 2001). Las defensas antioxidantes juegan un rol adaptativo crucial para las especies que rutinariamente experimentan amplias variaciones en el acceso a oxígeno debido al buceo o a patrones de respiración en condiciones de apnea por metabolismo limitado (hibernación) (Storey, 1996). En bahía Esperanza habitan poblaciones de Pygoscelis adeliae (Adelia) y Pygoscelis papua (Papúa), las que se encuentran distribuidas en parches, cerca y lejos de la Base Antártica Esperanza. Allí, Acero y col. (1991) identificaron zonas de alto impacto con: alto ruido de helicópteros, residuos sólidos, derrames de combustibles, desembarco de turistas y circulación de vehículos y/o personas; y otras de bajo impacto, es decir, sin esas perturbaciones. El aumento de actividades antrópicas en Antártida ha dado lugar a interacciones muy frecuentes entre personas y fauna antártica, pudiéndose generar efectos adversos en la fisiología y comportamiento de especies sensibles como los pingüinos (Culik y col., 1990, Giesse 1998). La exposición a diversas sustancias ambientales tóxicas, incluyendo metales pesados, hidrocarburos aromáticos (policíclicos y polihalogenados), pesticidas y micotoxinas, puede implicar una cascada de reacciones que incluya la producción de ROS y, por ende, generar estrés oxidativo (Bagchi, 2012). Por tal motivo resulta imprescindible la realización de estudios de monitoreo que indiquen el estado antioxidante en relación alas actividades antrópicas y la contaminación asociada a la que están expuestos los pingüinos. De este modo, se podrá determinar cuáles serían los mejores biomarcadores a la hora de evaluar el estado funcional de cada organismo. Con el objetivo de determinar el estado fisiológico de las poblaciones de Adelia y Papúa que habitan zonas de alto y bajo impacto antrópico de Bahía Esperanza, se determinó la actividad de enzimas y metabolitos que participan de las defensas antioxidantes sanguíneas. MATERIALES Y MÉTODOS Durante el verano 2012-2013, en dos áreas bien diferenciadas dependiendo del impacto antrópico generado por las actividades de la Base Antártica Esperanza (Bahía Esperanza, Península Antártica (63°23′54″S 56°59′46″O)), se tomaron muestras de sangre de P. adeliae y P. papua.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
68
Las muestras, de 15 pichones y 15 adultos de cada especie, fueron extraídas con jeringas heparinizadas. En el laboratorio de la Base se centrifugaron las muestras (1500g x 10 min) para separar los eritrocitos del plasma. Tanto las muestras de eritrocitos como de plasma se conservaron en frío hasta su análisis en el laboratorio de Ecofisiología y Ecotoxicología del Instituto Antártico Argentino (Buenos Aires). Se determinó en eritrocitos la actividad de enzimas antioxidantes: superóxido dismutasa (SOD), catalasa (Cat), glutatión S-transferasa (GST) y glutatión peroxidasa (GPX), así como los niveles deglutatión reducido (GSH), de oxidación de lípidos (Tbars) y de de proteínas (OP). Para el análisis de estas muestras se utilizaron técnicas espectrofotométricas Ansaldo y col (2007). Para el análisis estadístico de los resultados se empleó el modelo lineal general y mixto, teniendo en cuenta dos factores: tipo de impacto y edad. El mejor modelo se obtuvo por comparación de los IC de Akaike y Schwartz (InfoStat, 2012). Se utilizó como post test para las comparaciones entre los grupos la prueba DGC (Di Rienzo et al., 2002). RESULTADOS
0
6
12
18
aa
b
c
Tbar
s (m
mol
/ml h
emol
isad
o)
BAJO ALTO BAJO ALTO0.0
0.3
0.6
0.9
1.2 a
b
b b
Tipo de impacto
Tbar
s (m
mol
/ml p
lasm
a)
a)
b)
0.0
2.5
5.0
7.5
10.0
Adulto
Pichóna a
b
aG
ST
(mU
/mg
prot
eína
.min
)
BAJO ALTO BAJO ALTO0
2
4
6
8
aa
b
c
Tipo de impactoGP
X (
pmol
es N
AD
PH
/min
.mgp
rote
ina)
c)
d)
Figura 1. P. papua: niveles de Tbars en a) hemolisado y b) plasma; actividad de c) GST y d) Gpx vs tipo de impacto (BAJO
y ALTO) en pichones y adultos. Los valores están expresados como media ± error estándar. Diferentes letras indican diferencias significativas (p< 0,05).
Figure 1. P. papua: a) hemolysate and b) plasma TBARS levels; c) GST and d) Gpx activity vs type of impact (LOW and HIGH) in chicks and adults. Values are expressed as mean ± standard error. Different letters indicate significant differences
(p <0.05). Figura 2. P. papua: Actividad de SOD vs tipo de impacto (BAJO y ALTO). Los valores están expresados como media ± error estándar. Diferentes letras indican diferencias significativas (p< 0,05). Figure 2. P. papua: SOD activity vs LOW and HIGH impact zones. Values are expressed as mean ± standard error. Different letters indicate significant differences (p <0.05).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
69
BAJO ALTO BAJO ALTO0
4
8
12a a
ba
Adulto
Pichón
Tipo de impacto
Tba
rs (
mm
ol/m
lhe
mol
isad
o)
BAJO ALTO0
10
20
30
40
a
b
Tipo de impacto
OP
(nm
oles
DN
PH
/mg
prot
)
a)
b)
BAJO ALTO0
3
6
9
ab
Tipo de impactoG
ST
(m
U/m
gpr
oteí
na.m
in)
BAJO ALTO0
1
2
3
4
5
6
a
b
Tipo de impacto
Cat
(nm
oles
/mg
prot
eína
.min
)
c)
d)
Figura 3. P. adeliae: niveles de Tbars en a) hemolisado vs tipo de impacto (BAJO y ALTO) en pichones y adultos. b)
Niveles de OP; c) actividad de Cat y d) actividad de GSTen eritrocitosvs tipo de impacto (BAJO y ALTO). Los valores están expresados como media ± error estándar. Diferentes letras indican diferencias significativas (p< 0,05).
Figure 3. P. adeliae: a) hemolysate TBARS levels vs type of impact (LOW and HIGH) in chicks and adults. b) OP levels; c) Catactivity; and d) GST activity from erythrocytes vs LOW and HIGH impact zones. Values are expressed as mean ±
standard error. Different letters indicate significant differences (p <0.05). Los pichones de Papúa registraron en eritrocitos valores de Cat y OP significativamente (p<0.003) más bajos que en los adultos. A su vez, los adultos exhibieron niveles plasmáticos de SOD, GSH y OP significativamente menores (p<0.003) que los pichones. En cambio los pichones de Adelia tuvieron niveles de SOD significativamente menores (p<0.04) que los adultos. Para esta especie, además, los niveles de GST, GPX (p<0.0001) y GSH (p<0.02) fueron significativamente mayores en los pichones. En las zonas de alto impacto, se registraron para los eritrocitos de Papúa adultos aumentos significativos de los niveles de Tbars (p<0.0001) así como de la actividad de GST (p<0.005) y de Gpx (p<0.0001) (Fig. 1). En cuanto a lo determinado en pingüinos Adelia se registraron altos niveles de Tbars (p<0.0001) solo en adultos e incremento de actividad de GST (p<0.04) en ambos estadíos (Fig. 3). En cambio, se vieron disminuidos para las zonas impactadas tanto la actividad de Cat (p<0.0003) como los niveles de OP (p<0.01) en Adelia (Fig. 3), mientras que en Papúa fueron los niveles de Tbars (p<0.0001) en plasma de pichones (Fig. 1) y la actividad de SOD (p<0.006) (Fig. 2).
CONCLUSIONES Tanto P. adeliae y P. papua se encuentran igualmente afectados por la contaminación asociada a las actividades antrópicas en cuanto a las defensas antioxidantes evaluadas. El presente trabajo proporciona, además de la determinación de valores basales de los parámetros relacionados al estrés oxidativo, información sobre el estado metabólico-sanitario de estas poblaciones de pingüinos y un punto de referencia para monitoreos futuros y/o comparaciones con otras poblaciones.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
70
Por último, en virtud de nuestras observaciones, consideramos que debe prestarse mayor atención a la clara determinación de las alteraciones fisiológicas de los pingüinos, ya que allí subyace la respuesta real al fenómeno de impacto antrópico. AGRADECIMIENTOS Este proyecto fue realizado con el subsidio PICTO 0091 (FONCyTe IAA-DNA) y la Universidad Argentina JF Kennedy. Se agradece especialmente a la dotación de la Base Antártica Esperanza. REFERENCIAS BIBLIOGRÁFICAS Acero, J. y col. 1996. Revisión ambiental de las actividades antárticas en Bahía Esperanza, Península Antártica. Instituto Antártico Argentino. Publicación especial N°26. Ansaldo, M. Sacristán, H. y Wider, E. 2007. Comp. Biochem. Physiol., Part C 146: 118–123. Bagchi, M. Zafra-Stone, S. Bagchi, D. y Patel, S. 2012. Oxidative stress and chemical toxicity. Veterinary Toxicology: Basic and Clinical Principles, 426. Corsolini, S. Nigro, M. Olmastroni, S. Focardi S. y Regoli. F. 2001. Susceptibility to oxidative stress in Adélie and emperor penguin. Polar Biol 24:365-368. Culik, B. y col. 1990. The Effect of Disturbance on the Heart Rate and Behaviour of Adélie Penguins (Pygoscelis adeliae) During the Breeding Season. Antarctic Ecosystems, pp 177-182 Di Rienzo, J. Guzman, A. y Casanoves, F. 2002. A Multiple Comparisons Method based On the Distribution of the Root Node Distance of a Binary Tree Obtained by Average Linkage of the Matrix of Euclidean Distances between Treatment Means. JABES 7(2), 129-142. Giese, M. 1998. Guidelines for people approaching breeding groups of Adélie penguins (Pygoscelis adeliae).Polar Record, 34, pp 287-292. InsfoStat 2012. InfoStat versión 2012. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Di Rienzo J.A., Casanoves F., Balzarini M.G., Gonzalez L., Tablada M., Robledo C.W. Storey, J. 1996. Oxidative stress: animal adaptations in nature. Braz. J. Biol. Res. 29:1715-1733

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
71
ESTRÉS OXIDATIVO DETERMINADO DURANTE TRES VERANOS CONSECUTIVOS EN SANGRE DE PINGÜINOS ANTÁRTICOS
(Blood oxidative stress determination of Antarctic penguins in three consecutive summers)
Di Fonzo, C. I.1, Rosa, G.A.1, Archuby, D.1 yM. Ansaldo1,2
1Instituto Antártico Argentino, Dirección Nacional del Antártico. Cerrito 1248, (C1010AAZ) Ciudad Autónoma de Buenos Aires, Argentina. [email protected], [email protected] 2Laboratorio de Ecotoxicología. Universidad Argentina JF Kennedy. Sarmiento 4564, (C1197AAR) Ciudad Autónoma de Buenos Aires, Argentina. INTRODUCCIÓN La mayoría de los organismos terrestres basan su metabolismo en la reducción de la molécula de oxígeno para obtener energía. De manera natural se generan en este proceso especies reactivas del oxígeno (ROS) perjudiciales para la salud de los organismos pero que, también de forma natural, son desactivadas o atrapadas por sus defensas antioxidantes. Un cambio, en el equilibrio en el estado estacionario entre las ROS y los compuestos antioxidantes, dará lugar a estrés oxidativo (Halliwell y Gutteridge, 2007). En general, el estrés oxidativo es generado por: 1) el aumento en la producción de pro oxidantes, 2) la disminución de los niveles de antioxidantes, 3) la depleción de los cofactores esenciales dietarios (Se, Mg, Zn) necesarios para la actividad de enzimas antioxidantes, o 4) fallas en los sistemas de reparación o reemplazo (Constantini y col., 2008). La generación de ROS endógenos, puede ser influenciada por diferentes factores ambientales y biológicos. En general, la eficiencia basal del sistema antioxidante refleja la presión normal de las ROS a la que están expuestos los organismos (Corsolini y col., 2001). Constantini y Dell´ Omo (2006) y Rubolini y col. (2006) reportaron la existencia de diferencias en la capacidad antioxidante en plasma y suero en distintas especies de aves, hecho que fue relacionado a componentes ambientales como la calidad de la dieta y componentes genéticos. En particular, dentro de las aves antárticas, se ha observado que los pingüinos poseen una mayor capacidad de atrapamiento de las especies reactivas de oxígeno. Esta mayor resistencia puede sugerir que los pingüinos están naturalmente expuestos a una mayor presión basal de ROS en comparación con otras aves antárticas (Corsolini y col., 2001). Asimismo, los pingüinos poseen defensas antioxidantes que juegan un rol crucial adaptativo, dado que habitualmente experimentan amplias variaciones en el acceso a oxígeno debido al buceo (Storey, 1996). Aunque los pingüinos Barbijo, Papúa y Adelia, comparten el mismo género: Pygoscelis, poseen diferentes historias de vida y preferencias de hábitat. Adelia es una especie realmente Antártica, distribuida alrededor de todo el continente y en áreas donde el hielo marino se extiende durante todo el invierno. Barbijo y Papúa, por su lado, tienden a evitar zonas con hielo marino persistente, pues evolucionaron en hábitats sub-antárticos donde las condiciones son de mar abierto con escaso hielo (McClintock, 2008). Las 3 especies de pingüinos se alimentan principalmente de krill y en menor medida de peces (Libertelli y col., 2010). Existe una estrecha correspondencia entre el aumento de temperatura debida al cambio climático y la variación en cantidad y espesor del hielo marino que se forma en el invierno austral (Stammerjohn y col., 2008). Los cambios en las características del hielo marino tienen profundas consecuencias en el ecosistema antártico, entre ellas, la alteración del ambiente físico necesario para mantener la población de krill (principalmente Euphausia superba) (Atkinson y col., 2004). El éxito reproductivo del krill correlaciona con la extensión y duración de los hielos marinos (Loeb, 1996), es decir, el kril no puede completar su ciclo de vida y reproducirse satisfactoriamente sin hielo marino (McClintock, 2008). Considerando la importancia del krill, como la principal presa que habita el océano Austral y en la que se sustenta la red trófica antártica, es fácil suponer que la disminución de la abundancia de este crustáceo

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
72
afectará a sus depredadores directos, que incluyen a las tres especies de pingüinos antes mencionadas (McClintock, 2008). Una alteración en la disponibilidad y variedad del alimento en el ecosistema trae aparejado mayor tiempo en la búsqueda de la presa y/o la explotación de presas alternativas por parte de los consumidores, con la alteración de su metabolismo (Croll y col., 2006; Barbosa, 2011) y la consecuente variación del estado antioxidante del medio interno. El objetivo de este trabajo es determinar y comparar el estado antioxidante sanguíneo en tres especies de pingüinos antárticos P. antarctica, P. papua y P. adeliae durante tres veranos consecutivos.
MATERIALES Y MÉTODOS Durante tres veranos consecutivos (2009, 2010 y 2011) se realizaron monitoreos en las colonias de pingüinos antárticos: Pygoscelis papua (Papúa) y Pygoscelis adeliae (Adelia) de Península Potter y Pygoscelis antarctica (Barbijo), de Punta Narebsky (Península Barton), en la isla 25 de Mayo, Shetland del Sur, Antártida. Mediante punción de la vena braquial con jeringas heparinizadas, se tomaron muestras de sangre en pingüinos adultos. En el laboratorio de la Base Carlini se centrifugaron las muestras de sangre (1500g x 10 min) para separar los eritrocitos del plasma. Ambas fracciones fueron congeladas de inmediato (-20ºC) para su posterior análisis en el laboratorio de Ecofisiología y Ecotoxicología del Instituto Antártico Argentino (Buenos Aires). Se determinó espectrofotométricamente la actividad de enzimas antioxidantes como catalasa (Cat), glutatión S-transferasa (GST) y glutatión peroxidasa (GPX), así como los niveles de glutatión reducido (GSH), de lípidos oxidados (Tbars) y de proteínas oxidadas (OP) (Ansaldo y otros, 2007). Se realizó el Análisis de Función Discriminante (AFD) para: (1) describir las diferencias entre las especies durante los 3 veranos, en función de las seis variables registradas, y (2) determinar cuáles de dichas variables contribuyen más a la separación entre los grupos formados. Por otro lado, se realizó un Análisis de Componentes Principales (ACP) con las mismas seis variables para comparar e integrar los resultados con el AFD. Con el fin de evaluar si existen diferencias significativas entre los nueve grupos (especie*año) con respecto al primer componente principal (CP1) retenido, se realizó un ANOVA de un factor (considerando el factor cruzado especie*año) y como variable respuesta al CP1. Se realizaron contrastes a posteriori de Tukey para ver cuáles son los grupos que difieren significativamente entre sí.
RESULTADOS A partir del Análisis de Función Discriminante (AFD) se observa el ordenamiento de las especies a lo largo de los dos ejes canónicos retenidos. Ambas funciones discriminantes explicaron el 79% de la varianza total (Fig. 1). El 94% del los individuos fueron clasificados correctamente en cada grupo (especie/año). En el Análisis de Componentes Principales, con dos ejes (54% de la variabilidad total), se pudieron discriminar cuatro grupos diferentes (Fig. 2). El ANOVA de un factor del CP1 reveló diferencias significativas entre los grupos especies*año (p-valor < 0,0001). Por medio de los contrastes de Tukey, se encontraron diferencias significativas entre cuatro grupos: (1) Adelia y Barbijo 2010, (2) Papúa 2010, (3) Papúa 2009 y Adelia 2011, y por último (4) el resto de las combinaciones especie*año, que no pudieron ser diferenciados completamente.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
73
Figura 1. Biplot de los ejes canónicos extraídos del AFD y porcentajes de la varianza total explicada por cada función
discriminante. Los vectores representan las seis variables medidas: Tbars, OP, Cat, GST, GPX y GSH. Figure 1. Biplot of canonical axes extracted from the discriminant function analysis and percentage of total variance
explained by each discriminant function. The vectors represent the six measured variables: TBARS, OP, Cat, GST, GPX and GSH.
Figura2. Biplot de los dos primeros componentes extraídos del ACP y porcentajes de la varianza total explicada por cada componente. Los vectores representan las seis variables medidas: Tbars, OP, Cat, GST, GPX y GSH.
Figure 2. Biplot of the first two components extracted from the principal component analysis and percentage of total variance explained by each component. The vectors represent the six measured variables: TBARS, OP, Cat, GST, GPX and GSH.
DISCUSIÓN Se encontraron diferencias entre distintos grupos (especie*año) a partir de los distintos análisis realizados. A partir del AFD se distinguieron cuatro grupos: Barbijo y Papúa 2010; Barbijo 2011; Adelia 2010; y el resto (Adelia 2009 y 2011, Barbijo 2009, y Papúa 2009 y 2011). El primer grupo, Barbijo y Papúa 2010, se caracterizó por altos valores de Tbars y de GSH, esto significa que experimentan una alta oxidación de lípidos, lo que podría ser interpretado como indicador de aumento de estrés. Durante el 2010, se incrementó el consumo de peces en la dieta de Papúa respecto del año anterior (Coria y col., comunicación personal), causa que pudiera haber originado dicho aumento en Tbars. No existen registros de dieta de Barbijo, sin embargo dado que se comporta de manera similar a Papúa, podríamos presuponer que también aumentó su consumo de peces. El 2010 fue un año anómalamente frío, lo que podría haber contribuido al aumento de krill. No obstante, como fue mencionado antes, se incrementó el consumo de peces en la dieta de Papúa (y posiblemente de Barbijo). Esto sugeriría la existencia de otros factores que pudieron estar afectando el comportamiento alimenticio de estas dos especies durante ese período. Por otro lado, Adelia 2010 registró valores más bajos de Tbars respecto a Barbijo y Papúa, lo que indicaría un menor nivel de estrés para dicha especie. Adelia es una especie verdaderamente antártica, mejor adaptada a temperaturas muy bajas que las otras dos especies que evolucionaron en ambiente subantártico. Además, Adelia 2010 presentó altos valores de GSH, lo que mostró ser la principal respuesta antioxidante sanguínea de esta especie para ese año. Por su parte, Barbijo 2011 se caracterizó por altos valores de actividad GPX, bajos niveles de GSH y de Tbars. Estos resultados son difíciles de interpretar dado que se hubiese esperado que este grupo no se encuentre diferenciado del cuarto grupo (Adelia 2009 y 2011, Barbijo 2009, y Papúa 2009 y 2011) ya que las condiciones ambientales no fueron atípicas como las del 2010. Por lo tanto, se requeriría de información complementaria de la dieta de Barbijo en el 2011 para poder interpretar estos resultados. El ACP refuerza los resultados obtenidos con el AFD, distinguiendo los mismos cuatro grupos principales. Al analizar la nueva variable CP1 (que indicaría grado de estrés en relación a Tbars, OP y GSH) con
2 1

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
74
ANOVA de un factor (especie*año), se encontraron diferencias significativas (p-valor < 0,0001) entre los nueve grupos predefinidos (tres especies en tres años). Por medio de los contrastes de Tukey se distinguieron significativamente al menos cuatro grupos: (1) Adelia y Barbijo 2010, (2) Papúa 2010, (3) Papúa 2009 y Adelia 2011, y por último (4) el resto de las combinaciones especie*año, que no pudo diferenciarse completamente. En el AFD Adelia 2010 se encontró separado como grupo de Barbijo y Papúa 2010 (con cualquiera de los dos ejes principales, Fig. 1) lo que no se evidencia en el ANOVA del CP1, ya que se necesita el CP2 para separar estos dos grupos (Fig. 2). No se pudo realizar el ANOVA del CP2 por no cumplirse los supuestos. En contraposición al AFD y al ACP (observando los dos ejes en cada análisis), el ANOVA del CP1 separó a Papúa 2010 de Adelia y Barbijo 2010. En conclusión, el análisis de las variables de estrés oxidativo determinadas permitió diferenciar grupos de especies*año.
AGRADECIMIENTOS Este proyecto fue realizado con el subsidio PICTO 0091 (FONCyT y IAA-DNA). Se agradece especialmente a la dotación del Ejército Argentino de la Base Antártica Carlini.
REFERENCIAS BIBLIOGRAFICAS Ansaldo, M. Sacristán, H. y Wider, E. 2007. Does starvation influence the antioxidant status of the digestive gland of Nacella concinna in experimental conditions? Comparative Biochemistry and Physiology, Part C 146: 118–123. Atkinson, A. y col. 2004. Long-term decrease in krill stocks and increase in salps within the Southern Ocean. Nature Vol. 432: 100-103. Barbosa, A. 2011. Efectos del cambio climático sobre pingüinos antárticos. Ecosistemas 20 (1): 33-41. Constantini, D. y Dell´ Omo, G. 2006. Environmental and genetic components of oxidative stress in wild kestrel nestlings Falco tinnunculus. J. Comp. Physiol. B, 176, 575-579 Constantini, D. 2008 Oxidative stress in ecology and evolution: lessons from avian studies. Ecology letters, 11:1238-1251 Corsolini, S. Nigro, M. Olmastroni, S. Focardi, S.y Regoli, F. 2001 Susceptibility to oxidative stress in Adélie and emperor penguin. Polar Biol 24:365-368. Croll, D. y col. 2006. Effects of variability in prey abundance on reproduction and foraging in chinstrap penguins (Pygoscelis antarctica). Halliwell, B. y Gutteridge, J. 2007. Free Radicals in Biology and Medicine, 4th ed. Oxford University Press, New York. Libertelli, M. Juares, M. Santos, M. Favero, M. y Coria, N. 2010. Variation in the diet of Gentoo penguin at Cierva Point and Esperanza Bay (Antarctic Peninsula) and Stranger Point (South Shetland Islands), Antarctica. SCAR XXXI Open Science Conference, Bs As, Argentina. Loeb, V. y col. 1996. Effects of sea-ice extent and krill or salp dominance on the Antarctic food web. Nature 387, 897 – 900. McClintock, J. Ducklow, H. y Fraser, W. 2008. Ecological responses to climate change on the Antarctic Peninsula. American Scientist 96:302-310. Rubolini, D. y col, 2006. Early maternal, genetic and environmental components of antioxidant protection, morphology and immunity of yellow legged gull (Larus michahellis) chicks. J. Evol. Biol., 19, 1571-1584 Stammerjohn S. y col, 2008. Sea ice in the western Antarctic Peninsula region: Spatio-temporal variability from ecological and climate change perspectives. Deep-Sea Research II 55:2041-2058.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
75
PROTEINAS DE ESTRÉS EN INVERTEBRADOS MARINOS ANTARTICOS: UNA VISIÓN MOLECULAR Y FISIOLOGICA
(Stress proteins in Antarctic marine invertebrates: a molecular and physiological view).
González, M.1, Perez-Troncoso, C.1, Gilbert, J. 2, Muñoz, M.2, Mercado, L.3, Bethke, J.3 y K, Paschke4 1Laboratorio de Biorrecursos Antárticos, Instituto Antártico Chileno. 2Escuela Politécnica del Litoral, Guayaquil. Ecuador/Instituto Antártico Ecuatoriano (INAE) 3Instituto de Biología. Pontificia Universidad Católica de Valparaíso. 4Laboratorio de Ecofisiología de Crustáceos, Instituto de Acuicultura, Universidad Austral de Chile. E-mail: [email protected] INTRODUCCIÓN Actualmente se ha reportado una tendencia del calentamiento entre 0.2° y 1°C en la temperatura superficial del Océano Austral de la península Antártica. Los modelos predictivos prevén que la temperatura media del mar aumentará alrededor de 3°C para el año 2100 (Ericson et al 2012), lo que provocaría que organismos marinos antárticos estén expuestos más allá de sus límites de supervivencia y comiencen a perder críticamente sus funciones biológicas. En este contexto se hace necesario poder contar con biomarcadores a corto plazo respecto de la salud o mecanismos de resistencia que son necesarios para poder comprender y evaluar los mecanismos que afectan la biodiversidad marina Antártica.Es así como proteínas que son inducibles podrían ser usados como bioindicadores de respuesta al estrés. Normalmente la exposición de un organismo a temperaturas elevadas induce una rápida y transitoria expresión de proteínas de estrés térmico (HSP). Sin embargo, en especies marinas antárticas esta situación no es constante debido a la perdida, en algunos casos, de la capacidad de inducción de este gen producto de la estabilidad térmica de las aguas antárticas (Clark et al, 2008). De esta manera nos hemos centrado en la expresión de dos proteínas la HSP70 y la metalotioneína (MT), que podrían tener características de inducibles frente a factores ambientales que pueden gatillar su expresión. La expresión de estas proteínas se ha evaluado en el erizo Antártico (Sterechinus neumayeri), en el isópodo (Glyptonotus antarcticus) y en otro crustáceo (Branchinecta gaini) que a diferencia de los otros dos anteriores habita en lagos oligotróficos de la Península Antártica. Se han realizado ensayos de inducción utilizando bacterias como un factor biótico y temperatura como factor abiótico. MATERIALES Y MÉTODOS Organismos y experimentos de estrés Se recolectaron erizos antárticos (S. neumayeri) e isópodos (G. antarcticus) por buceo autónomo, mientras que camarón hada (B. gaini) se obtuvo desde la laguna Jade en la península Fildes (Isla Rey Jorge, Antártica). Se dividieron en grupos y se sometieron a estrés térmico de 3, 5 y 10°C durante 48 horas. Se obtuvieron muestras de tejido en diferentes intervalos de tiempo a 1, 24 y 48 horas. Los grupos controles para cada especie se mantuvieron a 0.5 ± 0.5 °C. Evaluación de la expresión de MT y HSP70 Se aisló ARN total de tejido por medio del método de TRIZOL (Invitrogen). Se sintetizó ADNc a partir de 1µg de ARN siguiendo el protocolo de la transcriptasa inversa de Invitrogen. Utilizando partidores para MT (MTFW: CACCATGCCTGATGTCAAGT; MT RV: TGTCTGGCTTGGAGCATGTTG) y HSP70 (FW QHSP70: GCAGAGGCATACCTTGGAAA; RV QHSP70- M1: AACACCAGCATCCTTCGT) se amplificaron amplicones de 292 y 99 pb, respectivamente. La expresión de Hsp70 y MT fue normalizada con respecto al gen control 28S de S.neumayeri. La

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
76
expresión de la proteína HSP70 fue evaluada en el isópodo por medio de western blot. Actualmente se están analizando muestras de B. gaini para poder amplificar estos mismos genes.
Figura 1. Evaluación por qPCR de la expresión de HSP70 y MT en el tracto digestivo de S. neumayeri a 3°C. Figure 1. qPCR evaluation of the expression of HSP70 and MT in the digestive tract of S. neumayeri at 3°C.
Figura 2. Variaciones en la concentración de proteínas durante el estrés térmico en la hemolinfa de G. antarcticus
(A) y el líquido celómico de S. neumayeri (B). Figure 2. Variations in the concentration of protein during heat stress in the hemolymph of G. antarcticus (A) and
coelomic fluid of S. neumayeri (B).
Figura 3.Incremento en el consumo de oxígeno después de estrés térmico a 3 y 6 °C en G. antarcticus.
Figure 3. Increase in oxygen consumption after thermal stress at 3 and 6 ° C in G. antarcticus Consumo de oxígeno y concentración de proteínas en S. neumayeri y G. antarcticus La tasa respiratoria se midió en S. neumayeri a través de la tasa de oxígeno consumido después de 1h de estrés térmico a 5 y 10°C. Para el caso de G. antarcticus este se sometió a 3 y 6°C. El consumo de oxigeno fue medido antes y después de la incubación por medio de un sensor óptico conectado a un

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
77
oxigenómetro (Microx MX3 AOT, Alemania). Los datos fueron analizados con un ANOVA de un factor para determinar los diferencias significativa entre tratamiento, los cuales fueron considerados estadísticamente significante con un P˂0.05. Para cada cuantificación se obtuvo tres replicas, estos fueron presentados como promedio con sus respectivas desviaciones estándar. Adicionalmente para cada especie se cuantificó la concentración de proteínas en el líquido celómico y la hemolinfa con el reactivo BIOQUANT basado en el método de Bradford (Merck). RESULTADOS Y DISCUSIÓN Diversos factores pueden gatillar la expresión de genes responsables de la respuesta al estrés. Para el caso de las proteínas de estrés térmico hemos determinado que para S. neumayeri la expresión de esta proteína no es inducible por calor en los celomocitos, que corresponden a un tipo particular de células inmunitarias. Ampliándose a validar esta expresión en otros tejidos hemos evaluado esta expresión en el tejido del tracto intestinal. Para el caso del erizo un estrés térmico a 3°C no induce la expresión del gen, sin embargo la expresión de la metalotioneína aumenta considerablemente después de una hora. La expresión de transcriptos de ARN de HSP70 confirmaría que este factor no induciría la expresión y sería similar al reportado para otras especies Antárticas. Por otra parte, la MT sería inducible por calor en estrés de corto plazo que podría explicarse por la posibilidad de que el promotor de MT pudiera estar regulada por factores de estrés térmico (Van Cleef-Toedt et al., 2001). El estrés térmico a nivel de proteínas circulantes en los líquidos para estos dos invertebrados presentaría una tendencia a la disminución después de estrés térmico a 5° y 10 °C. Para el caso de G. antarcticus también se aprecia una disminución, sin embargo esta muestra una tendencia a aumentar después de 24 horas. Es probable que el aumento de temperatura en el corto plazo produzca una disminución en la actividad transcripcional que provoque a su vez una disminución de las proteínas circulantes. Por otra parte, el consumo de oxígeno sería dependiente de la temperatura. Respecto del gasto metabólico que representa el hacer frente al aumento de la temperatura, ambas especies presentan un aumento considerable en consumo de oxígeno que indicaría la activación de la maquinaria celular y molecular. La activación de ciertos genes implicados en la mantención y protección de la maquinaria celular podrían aportar a este aumento del metabolismo. En esta misma dirección trataremos de evidenciar la expresión de proteínas de estrés para crustáceos como B. gaini presentes en laguna, que presentan variaciones mayores en su régimen térmico. Durante este año se comenzarán a realizar los estudios de transcriptoma de estas tres especies que entregarán la posibilidad de obtener secuencias completas de diversas HSP u otras proteínas de estrés. AGRADECIMIENTOS Proyectos Fondecyt 11090265 y 1131001. Apoyo INAE e INACH. REFERENCIAS BIBLIOGRAFICAS Clark, M. S., Fraser, K. P. y Peck, L.S. 2008. Lack of an HSP70 heat shock response in two Antarctic marine invertebrates. Polar Biol. 31 (9): 1059-1065. Ericson, J. A., Ho, M. A., Miskelly, A., King, C. K., Virtue, P., Tilbrook, B. y Byrne, M. 2012. Combined effects of two ocean change stressors, warming and acidification, on fertilization and early development of the Antarctic echinoid Sterechinus neumayeri. Polar Biology 35:1027–1034. Van Cleef-Toedt, K, Kaplan, L. y Crivello, J. 2000. Killifish metallothionein messenger RNA expression following temperature perturbation and cadmium exposure. Cell Stress Chaperones,6: 351–359.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
78
NIVELES DE CONTAMINANTES ORGANOCLORADOS PERSISTENTES Y CONCENTRACIÓN DE PORFIRINAS PARA EVALUAR EXPOSICIÓN EN PINGÜINOS
PYGOSCELIS PRESENTES EN LA PENÍNSULA ANTÁRTICA (Organochlorine contaminants and porphyrins levels for of the exposure assessment in Pygoscelis
penguins present in the Antarctic Peninsula)
Jara S. 1, Sánchez-Hernández J.C.2, Chiang G.3, Barra R. 1 1Laboratorio de Biomarcadores, Centro EULA, Universidad de Concepción-Chile. [email protected] 2Laboratorio de Ecotoxicología, Universidad de Castilla, Toledo–España. 3Canadian Rivers Institute, Department Biology, University of New Brunswick, Saint John NB, Canada. E-mail: INTRODUCCION La Antártica es considerada el continente más remoto del planeta, pero a pesar de esto no queda exenta del impacto antrópico a escalas local (bases científicas, turismo) y global (circulación global de contaminantes). Investigaciones demuestran la presencia de Contaminantes Orgánicos Persistentes (COPs) en la biota antártica, principalmente en aquellas especies que se sitúan en eslabones superiores de la red trófica. Esto ha conllevado a un incremento considerable de estudios que examinan la distribución y exposición de estas sustancias en los ecosistemas antárticos, por lo cual las porfirinas son consideradas un buen biomarcador para determinar exposición a este tipo de contaminantes. El objetivo de este trabajo fue determinar niveles de contaminantes organoclorados, PCBs y porfirinas, en muestras de heces de tres especies de pingüinos Pygoscelis presentes en la Antártica y validar la aplicación del método QuEChERS para el análisis de pesticidas organoclorados y PCBs en este tipo de matriz. La detección de los analitos de interés y su cuantificación se realizó en cromatografía gaseosa, con detector de captura de electrones (GC-ECD); las porfirinas fueron obtenidas mediante una digestión ácida y fueron medidas por fluorescencia. Se detectaron concentraciones importantes de PCBs (138, 153 y 180), así como pesticidas organoclorados. Estos resultados preliminares demuestran una exposición considerable de COPs. Así mismo, se sugiere el uso de matrices no invasivas como las heces para realizar monitoreo de los COPs y análisis de biomarcadores en poblaciones de pingüinos antárticos. La Antártica es el continente más remoto del hemisferio sur, posee un clima extremo propio de un desierto frío y posee una población humana no nativa. Como muchas otras áreas remotas del planeta, la Antártida no escapa al efecto de las actividades humanas locales y globales (Bargagli 2008). La presencia de contaminantes organoclorados semivolátiles y persistentes ha aportado nueva información sobre el transporte global atmosférico de estos contaminantes (Geiz et al., 2008) Se ha descrito la presencia de contaminantes orgánicos persistentes (COPs) en la Antártica desde la década de los 60 en la biota (Risebrough R & G Carmignani, 1972), atmósfera (Bidleman1993), agua (Bícego et al, 1996) y en sedimentos (Montone et al, 2001). Las aves marinas son consideradas especies centinelas de cambios y perturbaciones ambientales (Boersma P. 2008), debido a que se encuentran en la parte superior de la red trófica, presentan ciclos de vida largos y, además, dan respuestas sutiles con respecto a la presencia de contaminantes, por lo cual son especies que ayudan a determinar el estado de salud del ecosistema en que habitan (Joshi et al,2013) Los pingüinos antárticos se encuentran ampliamente distribuidos en la Antártica, representan el 75% de la biomasa aviar de este continente, son especies altamente especializadas, ya que se apoyan en sus reservas de grasa para poder mitigar los fríos invernales, periodos de escasez de alimento y periodos de incubación. Por lo cual reflejan los cambios asociados a la presencia de contaminantes orgánicos (Roosens et al, 2008, Taniguchi et al, 2009) y juegan una importante función dentro del ecosistema antártico, siendo capaces de entregar un escenario claro del impacto de los COPs en ecosistemas remotos (Inomata et al, 1996). Las muestras de heces han

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
79
demostrado ser una de las mejores matrices no destructivas para medir contaminantes orgánicos, gracias al mínimo de intervención humana existente en la toma de muestra, además que demuestra lo que el ave ha consumido y no ha asimilado (Yin et al, 2008,Sun et al, 2006, Sun et al, 2004). Las porfirinas son utilizadas como biomarcadores para medir el grado de exposición a contaminantes como metales pesados y COPs en aves y mamíferos; son un grupo de metabolitos que participan en la síntesis de muchas sustancias importantes (hemoglobina y citocromos) (Caisni et al, 2003); productos oxidados como la uroporfirina y la cropoporfirina (derivados de la síntesis del grupo hemo) no son tóxicos a niveles normales, pero en presencia de contaminantes tienden a acumularse en exceso pueden afectar el hígado y/o la médula ósea (Lim 1991, Leonzio et al, 1996). El procedimiento QuEChERS (Quick, Easy, Cheap, Effective, Rugged and Safe) es un método desarrollado originalmente para la extracción de una amplia gama de pesticidas en muestras de frutas y hortalizas (Anastassiades et al, 2006). Por su rapidez de análisis, bajo costo y alto rendimiento, este método ha resultado muy popular en la química ambiental, siendo mejorado ligeramente para el análisis de pesticidas en otras matrices ambientales (suelo, peces y grasa humana) (Norli et al, 2011, Castillo et al, 2011). El objetivo de este trabajo fue determinar niveles de contaminantes organoclorados, PCBs y porfirinas en muestras de heces de tres especies de pingüinos Pygoscelis presentes en la Antártica y validar la aplicación del método QuEChERS para el análisis de pesticidas organoclorados y PCBs en este tipo de matriz biológica. MATERIALES Y METODOS Las muestras fueron obtenidas en la expedición Antártica chilena del año 2011. Las muestras de pingüino Adelia (Pygoscelis adeliae) y pingüino Barbijo (P. antarctica) fueron recolectadas en Isla Kopaitic (63°19'01"S 57°54'29"O) y las muestras de Pingüino Papua (P. papua) fueron obtenidas en Península Ardley (62º13’S; 58º54’O). Se obtuvieron 10 muestras de cada especie. Para la extracción de contaminantes se realizó la metodología de QuEChERS descrita por Asensio-Ramos (2010), utilizada para suelos. El extracto final (1 ml) fue almacenado en un vial ámbar, llevado a seco bajo corriente de nitrógeno y luego se le agregó el estándar interno (PCNB 5 ppb), resuspendiéndolo en un volumen final de 1 mL con n-hexano. Las muestras fueron inyectadas en un cromatógrafo de gases con detector de captura de electrones (GC-ECD) Perkin Elmer Autosystem, serie 9000. La separación de los analitos de interés se realizó utilizando una columna capilar PTE-5 (30 m x 0,25 mm de diámetro interno y 0,25 µm de espesor de fase estacionaria), y helio como gas de carga. Las temperaturas del inyector y del ECD fueron de 240°C y 360°C, respectivamente, mientras que el programa de temperaturas de la columna fue el siguiente: 100ºC por 10 minutos; se utiliza una gradiente de 5ºC/min para llegar a una temperatura de 280ºC por 12 minutos. Tiempo total de corrida de 58 minutos. El análisis de COPs en las heces de pingüinos fue acompañado de un riguroso programa de control de calidad. Éste consistió en el análisis repetitivo de muestras blanco (ausente de heces) y muestras de heces dopadas con concentraciones conocidas de los analitos de interés. Los límites de detección del equipo son de 0,002µg/L. La extracción de porfirinas se determinó de acuerdo a Lockwood et al. (1985). La cuantificación de las porfirinas se realizo a través de análisis fluorimétrico, como se describe en Casini et al. (2001). Para análisis estadísticos de los datos se utilizó el software Sigmaplot versión 11. RESULTADOS Como resultados podemos observar que contaminantes organoclorados y PCBs presentan mayores concentraciones en P. papua y P. antarctica sobre los niveles encontrados en P. adeliae (diferencias significativas estadísticamente p > 0,01). En cuanto a las concentraciones de porfirinas, demuestran menores concentraciones de porfirinas en P. antarctica (1,067-2,868 nmol/g feca p.s) sobre las concentraciones encontradas en P. adeliae (1,696-3,665 nmol/g feca p.s) y P. papua (1,093-3,518 nmol/g feca p.s).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
80
PCB 138 PCB 153 PCB 180
Con
cen
trac
ión
PC
Bs
[pg/
g pe
so h
úm
edo
]
0
10
20
30
40
50 P. papua P. antarcticaP. adeliae
Fig. 1. Valores medios y error estándar de concentración de contaminantes organoclorados y PCBs (congéneres 138, 153 y
180) en pingüinos Pygoscelis. Fig. 1. Mean values and standard error of concentration of organochlorine contaminants and PCBs (congeners 138, 153 and
180) in Pygoscelis penguins.
P. antarctica P. adeliae P. papua
Con
cent
raci
ón d
e p
orfir
ina
s (n
mol
g-1
fec
a p.
s)
0
2
4
6
8
CoproporfirinaUroporfirinaProtoporfirinaPorfirinas totales
Fig. 2. Valores medios y error estándar de concentración de porfirinas en pingüinos Pygoscelis presentes en la Antártica.
Fig. 2. Mean values and standard error of porphyrins concentration in Pygoscelis penguin present in Antarctic. Material biológico no destructivo, como excrementos obtenidos de pingüinos antárticos, indica algún tipo de contaminación, la que puede estar asociada por contaminantes orgánicos presentes en estas especies antárticas, en un nivel considerado como preocupante debido a las concentraciones encontradas. Se observa una alta correlación entre los contaminantes y los niveles de porfirinas encontrados en este estudio. Las porfirinas son biomarcador útil para la evaluación de la exposición a contaminantes en los pingüinos. Metodología como QuEChERs -inicialmente desarrollado para la extracción de pesticidas en productos hortofrutícolas- resulta una herramienta analítica potente en el análisis químico de COPs. Este estudio muestra que el método de extracción de contaminantes denominado QuEChERs resulta una herramienta analítica potente en el análisis químico de COPs. AGRADECIMIENTOS Investigación financiada por Instituto Antártico Chileno (proyecto INACH T 18-09) y CONICYT. REFERENCIAS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
81
Anastassiades M, Lehotay S, Stajnbaher D & Shenk F. 2003.Fast and easy multiresidue method employing acetonitrile extraction/partitioning and "dispersive solid-phase extraction" for the determination of pesticide residues in produce. J AOAC Int. 86(2):412-31. Asensio-Ramos M, Hernández-Borges J, Ravelo-Pérez L & Rodríguez-Delgado M. 2010. Evaluation of a modified QuEChERS method for the extraction of pesticides from agricultural, ornamental and forestal soils. Analytical and Bioanalytical Chemistry 396: 2307-2319. Bargagli R. 2008. Environmental contamination in Antarctic ecosystems. Sci. of the Total Environment 400, 212-226. Bícego M, Weber R & Ito R. 1996 Aromatic hydrocarbons on surface waters of Admiralty Bay, King George Island, Antarctica. Marine Pollution Bulletin 32, 549–553. Bidleman T, Walla M, Roura R, Carrs E & Schmidt S. 1993. Organochlorine Pesticides in the Atmosphere of the Southern Ocean and Antarctica, January– March, 1990. Marine Pollution Bulletin. 26. 258–262. Boersma P. 2008. Penguins as marine sentinels. Bioscience 58, 597-607. Casini, S.; Fossi, M.C.; Gavilan, J.F.; Barra, R.; Parra, O.; Leonzio, C. & Focardi, S. 2001. Porphyrin levels in excreta of sea birds of the Chilean coasts as nondestructive biomarker of exposure to environmental pollutants. Arch Environ Contam Toxicol 41, 65–72. Castillo M, González C & Miralles A. 2011. "An evaluation method for determination of non-polar pesticide residues in animal fat samples by using dispersive solid-phase extraction clean-up and GC-MS." Analytical and Bioanalytical Chemistry 400(5): 1315-1328. Geisz H, Dickhut R, Cochran M, Fraser W & Ducklow H. 2008. Melting glaciers: A probable source of DDT to the Antarctic marine ecosystem. Environmental Science & Technology 42, 3958-3962. Inomata O, Montone R, Lara W, Weber R & Toledo H. 1996. Tissue distribution of organochlorine residues – PCBs and pesticides – in Antarctic penguins. Antarctic Science 8, 253–255. Joshi M, Bakre P & Bhatnagar P. 2013. Avian guano: A non-destructive biomonitoring tool for organic pollutants in environment. Ecological Indicators 24: 284–286. Leonzio C, MC Fossi, S Casini. 1996. Porphyrins as biomarkers of methylmercury and PCBs exposure in experimental quail. BullEnviron Contam Toxicol 56, 244-250. Lim, C.K. 1991. Porphyrins. In: Hanai T (ed). Journal of Chromatography Library. Liquid chromatography in biomedical analysis. Elsevier, Amsterdam, Pp 209–229. Lockwood, W.H.; Poulus, V.; Rossi, E. & Curnow, D. 1985. Rapid procedure for fecal porphyrin assay. Clin Chem 31, 1163–1167. Montone R, S Taniguchi & R Weber 2001. Polychlorinated biphenyls in marine sediments of Admiralty Bay, King George Island, Antarctica. Marine Pollution 42 (7), 611–614. Norli H, A Christiansen & E Deribe 2011. "Application of QuEChERS method for extraction of selected persistent organic pollutants in fish tissue and analysis by gas chromatography mass spectrometry." Journal of Chromatography A 1218(41): 7234-7241. Risebrough R & G Carmignani. 1972. Chlorinated hydrocarbons in Antarctic birds. In: Parker, B.C. (Ed.), Proceedings of the Colloquium on Conservation Problems in Antarctica. Allen Press, Lawrence, Kansas 63–80. Roosens L, Van Den Brink N, Riddle M, Blust R, Neels H & Covaci A. 2008. Penguin colonies as secondary sources of contamination with persistent organic pollutants. J. Environ. Monit., 9, 822–825. Sun L, Liu X, Yin X, Zhu r, Zhou Q & Wang Y. 2004. A 1500-year record of Antarctic seal populations in response to climate change. Polar Biol. 27:495–501. Sun L, Yin X, Liu X, Zhu R, Pan C, Zhao Y, Liu F, Jiang S & Wang Y. 2006. Levels of Hexachlorocyclohexanes and Dichloro-Dipheny-Trichloroethanes in penguin droppings collected from Ardley Island, Maritime Antarctic. Human and Ecological Risk Assessment 12: 328-338. Taniguchi S, Montone R, Bícego M, Colabuono F, Weber R & Sericano J.2009. Chlorinated pesticides, polychlorinated biphenyls and polycyclic aromatic hydrocarbons in the fat tissue of seabirds from King George Island, Antarctica. Marine Pollution Bulletin 58: 129–133. Yin X, Xia L, Sun L, Luo H & Wang Y. 2008. Animal excrement: A potential biomonitor of heavy metal contamination in the marine environment. Sci. Total Environ. 399: 179-185.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
82
MERCURIO EN LA ISLA DE DECEPCIÓN, ANTARTIDA – UN ESTUDO INTEGRADO (Mercury in Deception Island, Antarctica: An integrated study)
Canário, J.1, Mão de Ferro, A.1, Mota, A.1, Mateus, M.2
1CQE, Instituto Superior Técnico, Technical University of Lisbon, Av. Rovisco Pais, 1049-001 Lisboa, Portugal. Email: [email protected] 2MARETEC, Instituto Superior Técnico, Universidade Técnica de Lisboa, Av. Rovisco Pais, 1049-001, Lisboa, Portugal Antarctica is seen as a remote and hostile place, symbol of a pristine environment untouched by human disturbances. However, the development of Antarctic research especially after the International Geophysical Year (1957-58) and tourism have resulted in sharp increase of pressure from human activities, leading to several pollution problems, mainly near scientific stations (e.g. Bargagli, 2008). Studies in King George Island (e.g. Santos et al., 2005) and Ross Island (e.g. Sheppard et al., 2000), indicate that the main sources of anthropogenic trace elements are due to fuel spills, waste disposal sites, sewage, paint residues and exhaust gases. However, there are important natural sources in Antarctica that have been also considered, namely in what concerns mercury (Hg). Comparatively to other locations in Antarctica, environmental studies in Deception Island (South Shetlands Archipelago) are scarce and are of great interest since it has potential sources of trace element related to the volcanic activity, as well as anthropogenic input due to its history, tourism and two scientific stations. Although Hg levels in Deception were previously analyzed in lichen specie Usnea antarctica by Bargagli et al. (1993), to the best of our knowledge this is the first integrated mercury study held in this volcanic island, in order to define sources, fluxes and pathways of mercury in this ecosystem. We also intend to investigate the speciation of Hg regarding one of its most important methylated form, MeHg, and to assess the role of particulate organic matter as well as the geochemistry nature of the sediment in the cycle of this important contaminant. In December 2011 a field campaign took place in Deception Island (Figure 1). Sediments, ice, water and suspended particulate matter were collected in Mecon River and in the Foster Bay during 8 consecutive days. During this period Hgº fluxes between fumarole soil/water and atmosphere were also measure in fumaroles at Fumarole Bay. Samples were processed in the laboratory of Gabriel de Castilla Spanish Antarctic Station and transported to Lisbon preserved in HNO3 or HCl (water) or dried (sediments/soils). In the laboratory all samples were analysed for total mercury (particulate – HgP – and dissolved HgD), methylmercury (MeHg) and other interpretative physico-chemical parameters (e.g. organic carbon, Al, Si, Fe). In addition, we have used a numerical model to reproduce the basic water circulation patterns inside the bay, and the water fluxes between the bay and the coastal area, so that we could: (1) identify the processes that explain the different distribution patterns of Hg inside the bay, and (2) have an idea on the exportation of this trace element to the open ocean.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
83
Figure 1: Sampling Sites in Deception Island Figura 1: Sitios de muestreo en isla Decepción.
In Mecon River total mercury concentrations in the upstream stations were lower compared to the downstream ones. Dissolved Hg levels varied from 0.54 ng L-1 in station #3 to 2.36 ng L-1 in #1 and particulate Hg concentrations from 0.012 µg g-1 to 1.60 µg g-1 at the same sampling sites. These results indicate an increase of Hg levels downstream suggesting that main source of Hg to the river may come from Port Foster’s water. Our results clearly showed that volcanic activity is the most important Hg source in Deception Island. Mercury levels in water and sediments sampled at two fumaroles were up to 10000 times higher than in the other sampling sites. Additionally, Hg fluxes (fumarole/air) were similar in both fumaroles (average: 12 ng Hg m-2 min-1) decreasing to 2 ng Hg m-2 min-1 (water/air) when the site was inundated by Port Foster water. At the same time of inundation dissolved Hg in the bay increase from 1 to 25 ng L-1) indicating the existence of a lixiviation of the Hg coming from the fumarole vents and the consecutive transport of that Hg into the bay. In this site dissolved methylmercury (MeHg) is below the detection limit in all samples, probably due to the very high temperature found in fumaroles (above 80ºC) probably inhibiting the methylation process. On the contrary, MeHg proportion (MeHg/HgT) was surprisingly high in saline waters of Port Foster (13 – 33 %), which suggests exceptional conditions for Hg methylation processes in the water column of Port Foster. A combination of factors such as the presence of important volcanic sources of inorganic Hg, lower light and temperature associated with high residence time of saline waters of Port Foster might have led to a higher concentration of Hg in those waters during winter which, in turn, will be

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
84
available for methylation processes during the warmer months. Additionally, the high residence time of water in the Bay may also contribute to create an inner pool of volcanic mercury and MeHg available for aquatic organisms. Moreover, the low capacity of sediments for Hg adsorption, (low carbon content and low surficial area (Si/Al>4)), may also contribute to keep mercury in solution and therefore increase its bioavailability. ACKNOWLEDGEMENTS The authors would like to thank to the Portuguese Polar Program (PROPOLAR) funded by the Fundação para a Ciência e Tecnologia and to the Spanish Polar Board. REFERENCES Bargagli R. 2008. Environmental contamination in Antarctic ecosystems, Science of The Total Environment, 400 (1-3): 212-226 Santos I. R., Silva-Filho E. V., Schaefer C. E. G. R., Albuquerque-Filho M. R., Campos L. S. 2005. Heavy metal contamination in coastal sediments and soils near the Brazilian Antarctic Station, King George Island, Marine Pollution Bulletin, 50 (2): 185-194. Sheppard D. S., Claridge G. G. C., Campbell I. B. 2000. Metal contamination of soils at Scott Base, Antarctica, Applied Geochemistry, 15 (4): 513-530.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
85
AREAS LIBRES DE HIELO Y DISTRIBUCION DE Sanionia uncinata (Hedw.)Loeske EN LAS ISLAS SHETLAND DEL SUR, ANTARTICA.
(Ice-free areas and distribution of Sanionia uncinata (Hedw.)Loeske on the South Shetland Islands, Antarctica)
González, I.1, Hebel, I.2 Jaña, R.3, 1
1Centro de Estudios del Cuaternario y Fuego Patagonia, CEQUA; 2Laboratorio de Biotecnología & Genética de Plantas, Centro Hortícola “Lothar Blunck”, Universidad de Magallanes; 3Instituto Antártico Chileno Email: [email protected] INTRODUCCIÓN El pulsante retroceso de los hielos, tiene una importancia ecológica fundamental, que radica en que las nuevas áreas libres de hielo, constituyen zonas susceptibles de ser colonizadas por especies vegetales. En Antártica y las Islas Shetland del Sur, los rasgos geomorfológicos en esas áreas libres de hielo evidencian procesos tales como: pulsos glaciares, rebotes isostáticos y transgresiones marinas, mostrando una serie de cambios a partir del Ultimo Máximo Glaciar (John, 1972; Serrano et al., 1998; Ingólfsson et al., 1998; Serrano, 2003; Serrano et al. 2004, Yoon et al., 2000). Estos cambios han permitido el establecimiento de diferentes especies vegetales, principalmente líquenes y musgos, a partir de condiciones más cálidas ocurridas a partir del Último Máximo Glacial (Premoli et al, 2010). Dentro de las especies vegetales se encuentra el musgo Saniona uncinata, hoy con una amplia distribución a lo largo de las Islas Shetland del Sur, pero, ¿Cuál ha sido su distribución en épocas pasadas? Para resolver esta pregunta, se propone reconstruir la probable localización geográfica de la especie usando dos herramientas: Modelo de Máxima Entropía de distribución potencial de especies MAXENT (Phillips et al., 2004, 2005) y SIG (Sistema de Información Geográficos QGIS y ARCMAP 10.1). MAXENT como herramienta de modelamiento de nichos ecológicos y con la cual se predice la distribución potencial de la especie en áreas libre de hielo y SIG para la espacializacion y rasterización de las variables compiladas en el modelo. La plataforma SIG también se utiliza para identificar espacialmente rangos de valores que estén por sobre el 51 % de probabilidad de predecir presencia, de las variables scenopoeticas incidentes en la predicción. Los resultados de MAXENT responden a una determinada temporalidad, condicionada por el momento de colecta de los datos de presencia (muestras) y por la manifestación temporal de las variables bióticas y/o abióticas. En el afán de reconstruir la distribución para otros momentos temporales del pasado, es necesario restringir la predicción, solamente al uso de algunas variables, identificadas como predictivas, pues no se cuenta con datos de presencia o ausencia de la especie. La Península Fildes se constituye en el contexto espacial sobre la cual se ha calibrado el modelo de distribución potencial de S. uncinata, para el presente. Como datos de presencia se utilizaron las colectas realizadas en el año 2010 (266 muestras) y 2011 (38 muestras) en Fildes. Las variables abióticas consideradas para cada una de las pruebas fueron construidas teniendo en cuenta las 19 variables de WORLD CLIM – Global Climate Data. Es así como se construye las variables espaciales de: temperatura (lapse rate), humedad suelo (índice topográfico de Humedad), pendiente, exposición y altura. Estas variables espaciales cumplen con tener formato raster, con resolución de grilla de 40m.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
86
Figura 1. Variables abióticas representadas en formato raster usadas en modelo MAXENT. De izquierda a derecha: a) altura, b) pendiente, c) exposición, d) temperatura. Península Fildes, Isla Rey Jorge, Antártica.
Figure 1. Abiotic variables represented in raster format used in MAXENT model. From left to right: a) altitude, b) slope, c) exposure, d) temperature. Fildes Peninsula, King George Island, Antarctica.
Las variables que tienen una participación significativa en la predicción, son las variables presentadas en la Figura 1. La bondad de ajuste para los datos de entrenamiento tienen un valor aceptable (parámetro AUC = 0,738 - valor máximo 1), siendo el valor máximo obtenido en las corridas del modelo realizadas hasta el momento. En términos del peso relativo de cada variable en la predicción, la temperatura y la altura en conjunto corresponden a un 75% del total del valor de la contribución a la predicción; luego les sigue la exposición y por último la pendiente. La humedad -derivada mediante el índice topográfico de humedad- contribuye con un poco menos del 3%. Además de los valores estadísticos que proporciona el modelo MAXENT para su validación, la predicción de la distribución potencial del musgo se comparó con una imagen satelital (QuickBird, febrero 26, 2010; combinación de bandas falso color), observándose correspondencia entre la distribución potencial de los valores de alta probabilidad de presencia con las zonas que actualmente presentan actividad clorofílica (figura 2), es decir hay zonas predichas efectivamente ocupadas por vegetación. También se observa, en menor medida, una predicción de la distribución en áreas que en la imagen no se advierten. Una explicación posible es que esto puede deberse a una combinación de causas: los valores calculados para las variables utilizadas, la correlación de las variables, el número de muestras disponibles y los ajustes propios del modelo en función de la resolución. Por otro lado, el modelo al definir variables scenopoeticas está mostrando zonas que cumplen con condiciones abióticas aptas para la especie S. uncinata, por su parte, la imagen de satélite deja en evidencia áreas que actualmente están colonizadas por especies. Los resultados preliminares permiten concluir que se ha identificado un conjunto de variables abióticas que contribuyen en forma significativa a la predicción, de acuerdo a los estimadores de sensibilidad del modelo. Esto se corrobora con la concordancia entre la predicción de distribución potencial y la localización observada en la imagen y los datos de terreno. Esto sugiere que sería posible reconstruir una distribución potencial de la especie S. uncinata en el pasado a partir de las condiciones abióticas de esos momentos, permitiendo estimar su distribución potencial a partir de la presencia actual.
a b c d

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
87
Figura 2. Distribución actual y potencial de S. uncinata en la Península Fildes, Antártica. a) La distribución potencial de S. uncinata resultado de MAXENT. b) Imagen satelital falso color. Los círculos negros muestran las coincidencias tanto en la
probabilidad de distribución como la presencia actual de vegetación. El círculo gris muestra un área en que el modelo predijo probabilidad de presencia, pero no se observa en la imagen falso color.
Figure 2. Current and potential distribution of S. uncinata in the Fildes Peninsula, Antarctica. a) The MAXENT result of potential distribution of S. uncinata. b) False color satellite image. The black circles show the similarities of both the
potential distribution and the actual presence of vegetation. The gray area shows the model predicted presence probability, but not observed in the false color image.
AGRADECIMIENTOS. Proyecto INACH código T_17-09 y al Centro de Estudios del Cuaternario, CEQUA. REFERENCIAS BIBLIOGRÁFICAS Elith, J. Phillips, S. Hastie, T. Dudík, En Chee, Y. and Yates, C. 2011. A statistical explanation of MaxEnt for ecologists. Diversity and Distributions, (Diversity Distrib.) 17: 43-57. Hall, B. 2007. “Late-Holocene advance of the Collins Ice Cap, King George Island, South Shetland Islands.” The Holocene 17(8): 1253-1258. Ingólfsson, Ó., Hjort, C., Berkman, P., Björck, S., Colhoun, E., Goodwin, I.D., Hall, B., Hirakawa, K., Melles, M., Möller, P. and Prentice, M. 1998. Antarctic glacial history since the Last Glacial Maximum: anoverview of the record on land. Antarctic Science, 10: 326-344. John, B.S. 1972. Evidence from the South Shetland Islands towards a glacial history of West Antarctica. In: Price, R. J. and Sugden, D.E. (Eds.). (1972). Polar geomorphology (No. 4). Institute of British Geographers. Phillips, S., Dudik, M. and Schapire, R. 2004. “A Maximum Entropy Approach to Species Distribution Modeling.”. Proceedings of the Twenty-First International Conference on Machine Learning. 655-662. Phillips, S., Anderson, R. and Schapire, R. 2005. Maximum entropy modeling of species geographic distribution. Ecological Modelling 190; 231-259.
a b

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
88
Premoli, A., Mathiasen, P. and Kitzberger, T. 2010. “Southern-most Nothofagus trees enduring ice ages: Genetic evidence and ecological niche retrodiction reveal high latitude (54°S) glacial refugia.” Palaeogeography, Palaeoclimatology, Palaeoecology 298 :247–256. Serrano, E. and Lopez-Martinez, J. 1998. Caracterización y distribución de las formas y los procesos periglaciares en las Islas Shetland del Sur (Antártida). Procesos Bio-Físicos Actuales en Medios Fríos. 181-204. Serrano, E. 2003. “Paisajes naturales y pisos geoecológicos en las áreas libres de hielo de la Antártida Marítima. (Islas Shetland del Sur).” Boletín A.G.E. 35: 5-32. Serrano, E. and López-Martínez, J. 2004 Morfogénesis periglaciar y deglaciación en las penínsulas Barton y Weaver (islas Shetland del Sur, Antártida). Bol. R. Soc. Esp. Hist. Nat. (Sec. Geol.), 99 (1-4): 131-140. López-Martínez, J., et al., 2012. Periglacial processes and landforms in the South Shetland Islands (northern Antarctic Peninsula region), Geomorphology, doi:10.1016/j.geomorph.2011.12.018. Yoon, H. I., Park, B. K., Kim, I. and Kim, D. 2000. “Glaciomarine sedimentation and itspalaeoceanographic implications along the fjord margins in the South Shetland Islands, Antarctica, during the last 6000 years.” Palaeogeography, Plaeoclimatology, Palaeoecology, 157: 189-211.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
89
STOCK DE CARBONO ORGÁNICO EN EL SUELO DE LA PENÍNSULA FILDES Y LA ISLA ARDLEY
(Soil Organic Carbon Stock of Fildes Peninsula and Ardley Island)
Michel R.F.M.1, Schaefer C.E.G.R2, Simas F.N.B. 2 and Bockheim J.G.3
1Fondazione Edmund Mach di San Michele all'Adige, Via E. Mach, S. Michele all'Adige, Italy. E-mail: [email protected] 2Departamento de Solos–Universidade Federal de Viçosa, Av. PH Rolfs, Viçosa, MG, Brasil. E-mail: [email protected], [email protected] 3Department of Soil Science University of Wisconsin - 1525 Observatory Drive Madison, WI, USA. E-mail: [email protected] INTRODUCTION Permafrost-affected soils (PAS) have been recognized as a important component of the global soil organic carbon poll (SOCP). In addition to occurring in cold landscapes of the higher latitudes, PAS are found at high elevations in the lower latitudes. Despite the low primary productivity of vegetation in such environments, the low decomposition rates of soil organic carbon (SOC) together with cryoturbation processes lead to great accumulation. These soils occupy 8 % of the global land area (Soil Survey Staff, 2010) being classified in the World Reference Base for Soil Resources (WRB) (ISSS, 2006) as Cryosols. These are the major soils occurring in ice free areas of Maritime Antarctica together with Inceptisols, Entisols and Gleysols, all with gelic properties (Bockheim et al. 1990, Simas et al. 2007, Francelino et al. 2011). The Antarctic and Sub-Antarctic Permafrost, Soils and Periglacial Environments Group (ANTPAS) launched a international project to address some of the current shortcomings and research needs. The Brazilian TERRANTAR working group, is part of this effort being responsible for the Maritime Antarctica (M.A.) and north part of the Antarctic Peninsula. During the Brazilian Antarctic Expeditions of 2008 and 2009, a detailed soil survey was carried out in Fildes Peninsula and Ardley Island as part of the Brazilian Antarctic Research Program. In this context, the objectives of the present work are to present and discuss analytical data for the main soil classes found in Fildes Peninsula and Ardley Island regarding SOC. MATERIAL AND METHODS King George Island (KGI) is the largest of the South Shetland’s Archipelago with approximately 1400 km2. Similarly to the rest of the archipelago, it is almost completely glaciated, having over 5 % of it’s area free of ice during the summer (Simões et al. 1999). Fildes Peninsula and Ardley Island are ice-free areas (latitude 62°08´ to 62°14´S and longitude 59°02´ to 58°51´W) situated in the southwest part of KGI, Fildes peninsula (29 km2) is the largest ice free area in the island and is connected to Ardley Island (1.2 km2 ) by an 400 m isthmus, exposed during low tide. The region experiences a sub-antarctic maritime climate with milder temperatures, strong winds and frequent weather changes. According to the Köppen climate classification, the south Shetlands islands have an ET climate, South Hemispheric Polar Oceanic (Köppen, 1936). Fildes Peninsula experiences a cold moist maritime climate characterized by mean annual air temperatures of -2°C and mean summer air temperatures above 0°C up to four months (Rakusa-Suszczewski et al. 1993, Wen et al. 1994), precipitation ranges between 350 and 500 mm per year, with rainfall occurring in the summer period (Øvstedal & Smith, 2001). Soil surveying and sampling were performed during the austral summer, between January and February of 2008 and 2009. The landscape is typical of most ice-free areas of the Shetlands Archipelago, where land exposure is mid Holocene (6000 YBP). Sampling pits were dug to the depth bedrock or permafrost (considering a control section of 1 m), followed by morphological description. Soil classification

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
90
followed the World Reference Base for Soil Resources classification system (ISSS, 2006). Total organic C was determined by wet combustion (Yeomans and Breemner, 1988). The content of > 0.2 mm particles in the soils, including cobbles and pebbles was obtained gravimetrically. C stocks were estimated according to Batjes (1996): Cst = (TOC)*(BD)*(LT)*(1 - % fragments>0.2 mm), where: Cst = carbon stock (Mg m-2); TOC = total organic carbon (gC gsoil-1), BD = bulk density (Mg m-3) and LT = layer thickness (m). C stocks were calculated for every soil horizon and averaged for the profile (considering the thickness of every soil horizon). RESULTS AND DISCUSSION Fildes Peninsula and Ardley Island are among the first sites in Maritime Antarctica exposed after the last glacial maximum (Birkenmajer, 1989), soils are well developed for Antarctic standards, profiles sustain weak to moderate structure. Arenosols and Cryosols (frequently turbed) are the most important soil classes, Leptosols, Gleysols and Cambisols also occur. Areas visited or colonized by the fauna (mainly avifauna) are extensive, where vegetation growth is diverse and vigorous. These sites represent the most important SOCP of the studied area. Soil depth range from 10 to 120 cm with average of 55 cm considering all 48 profiles. Soil texture ranged from sand to loam-clay, although coarser textures are more common, coarse sand content averaged 41 dag k-1 (±18.0 dag k-1), fine sand averaged 18 dag k-1 (±9.2 dag k-1), silt content averaged 27 dag k-1 (±13.0 dag k-1) and average clay content was 14 dag k-1 (±6.3 dag k-1). The levels of SOC were low, being higher for profiles located at lower parts of the landscape, sites visited or colonized by the fauna (Ornithogenic), specially at the western portion of the studied site, less exposed to the strong winds influenced by the Drake passage. Overall average for all horizons was 2.6 dag k-1 (max of 26.74 dag k-1, min of 0.09 dag k-1, standard deviation of ±4.0 dag k-1), soil carbon stock aver all the studied area was 0,0035 kg m-2, SOCP reached a total of 105.97 Mg. Soil mapping units containing Cryosols cover 1538.45 ha (51.34% of the ice free studied area, 2996.31 ha). Soil carbon content varied greatly due to the diversity of the profiles, with a minimum of 0.13 dag k-1
and a maximum of 5.20 dag k-1, the average carbon content was 1.09 dag k-1 (±1.3 dag k-1). The average soil carbon stock was 0.6 kg m-2 and the SOCP of the Cryosols was 55.12 Mg (49,91 Mg considering only Turbic Cryosols). Soil mapping units dominated by Arenosols, usually located at intermediate altitudes (maximum of 33 m), showed limited cryoturbation and little horizon differentiation. These sites are commonly visited or colonized by the fauna expressing moderate amounts of SOC. Overall average for all horizons was 0.46 dag k-1 (max of 0.56 dag k-1, min of 0.31 dag k-1, ±0.11 dag k-1). These soil units occupied 1113.39 ha (37.16% of the studied area), soil carbon stock averaged 0.30 kg m-2, and the SOCP totaled 26.70 Mg. Mapping units where Leptosols are predominant, englobing very shallow and very gravely soils, expressed varying contents of SOC, soils not colonized by the fauna had a average SOC around 0.52 dag k-1, when colonized SOC averaged 2.83 dag k-1. This class covers 100.4 ha (3.35% of the studied area), presented a average soil carbon stock of 0.56 kg m-2 and a total SOCP of 2.76 Mg. The Gleysols unit was identified in the thaw channels and outwash plains, this position in the landscape favors the accumulation of sediments and the preservation of SOC, average was 1.20 dag k-1; this soil class occupies 243.3 ha with average soil carbon stock of 0.87 kg m-2 and total SOCP of 21.38 Mg.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
91
Table 1: Soil class, Color, Texture, gravel content, Soil organic carbon and soil carbon stock of the sampled profiles Tabla 1: Tipo de suelo, textura, contenido de grava, carbón orgánico del suelo y stock de carbono del suelo en los perfiles muestreados.
ID Color SOC C Stock
Class g Kg-1 dag Kg-1 Kg m-2
Fildes Peninsula
1 Haplic Arenosol (Eutric, Gelic) 10 YR 4/3 loam-sand 31.4 0.56 0.2282
2 Haplic Arenosol (Eutric, Gelic) 10 YR 5/2 sand-loam 41.4 0.31 0.0837
3 Haplic Arenosol (Ornithic, Gelic) 7.5YR 7/2 sand 43.1 0.48 0.1564
4 Haplic Arenosol (Eutric, Gelic) 10Y 4/2 sand-loam 20.7 2.25 0.4663
5 Haplic Arenosol (Eutric, Gelic) 10YR 3/1 sand-loam 36.1 0.13 0.2996
6 Haplic Arenosols (“Biogenic”, Gelic) 2.5Y 3/3 sand-loam 20.2 3.08 0.2909
7 Protic Arenosol (Eutric, Gelic) 7.5YR 7/2 loam-sand 31.6 0.50 0.2282
8 Leptic Cambisol (Eutric, Gelic) 5Y 7/6 loam 37.4 0.31 0.1142
9 Leptic Cambisol (Ornithic, Gelic) 10YR 4/2 loam 27.4 10.38 1.3727
10 Epiturbic Cambic Cryosol (Eutric, Novic) 2.5Y 5/2 loam 35.0 0.54 0.1803
11 Haplic Cryosol (Ornithic, Arenic) 2.5Y 5/4 loam-sand 9.1 1.01 0.4057
12 Histic, Leptic, Cryosol (Reductaquic, Arenic) 10 YR 5/4 loam-sand 48.2 5.20 0.8303
13 Leptic Cryosols (Eutric, Oxyaquic) 2.5Y 4/2 loam 29.6 0.52 0.0585
14 Turbic Leptic Cryosol (“Patterned”, Eutric, Skeletic) 7.5YR 5/2 sand-loam 42.9 2.31 0.3839
15 Turbic Leptic Cryosol (Eutric, Reductaquic, Arenic) 7.5YR 6/3 loam-sand 55.1 0.17 0.3177
16 Turbic Cryosol (“Patterned”, Eutric) 7.5YR 7/2 loam 34.5 0.48 0.1841
17 Turbic Cambic Cryosol (“Patterned”, Calcaric, Siltic) 5Y 6/6 lom-silt 30.3 0.29 0.3252
18 Turbic Leptic Cryosol (“Patterned”, Eutric) 10 YR 6/2 loam-sand 48.7 0.89 0.6582
19 Turbic Leptic Cryosol (Eutric, Gelic) 10 YR 4/4 loam 37.7 0.55 0.2784
20 Turbic Cryosol (“Patterned”, Eutric, Arenic) 10 YR 5/4 loam-sand 32.8 0.88 0.4560
21 Turbic Cryosol (“Patterned”, Eutric, Arenic) 7.5YR 6/3 loam 39.1 0.64 0.1142
22 Turbic Cambic Cryosol (“Patterned”, Calcaric, Siltic) 2.5Y 7/6 loam-clay 15.9 0.38 1.0030
23 Turbic Cambic Cryosol (“Patterned”, Calcaric, Siltic) 2.5Y 7/6 loam-clay 14.3 0.26 0.4037
24 Turbic Cambic Cryosol (“Patterned”, Calcaric) 7.5 YR 5/4 loam 21.5 0.30 0.3978
25 Turbic Leptic Cryosol (Ornithic, Arenic) 7.5YR 4/3 sand-loam 19.8 3.13 0.3660
26 Turbic Cryosol (Eutric, Skeletic) 10YR 5/2 sand-loam 41.7 2.54 0.0839
27 Turbic Cryosol (“Patterned”, Eutric, Skeletic) 2.5Y 5/3 sand-loam 39.7 0.54 0.0935
28 Turbic Cryosol (“Patterned”, Eutric) 10YR 6/3 loam 38.6 1.63 0.5831
29 Haplic Gleysol (Colluvic, Gelic) 10 YR 5/2 loam-sand 39.2 1.20 0.8769
30 Haplic Leptosols (Eutric, Gelic) 7.5YR 5/2 sand-loam 44.7 0.67 0.4724
31 Lithic Haplic Leptosol (Eutric, Gelic) 2.5Y 3/2 sand-loam 18.2 0.53 0.0446
Ardley Island
32 Leptic Arenosol (Ornithic) 7.5YR 4/2 sand-loam 39.2 2.17 0.2594
33 Folic Leptic Cryosol (Ornithic, Arenic) 5Y 2.5/1 loam-sand 32.8 19.90 0.6186
34 Folic Leptic Cryosol (Ornithic, Reductaquic, Arenic) 10YR 4/3 loam-clay 33.0 15.67 0.6831
35 Haplic Cryosol (Ornithic, Reductaquic, Novic) 5Y 4/1 loam-sand 26.9 5.85 1.0362
36 Histic Leptic Cryosol (Ornithic, Reductaquic) 10YR 4/4 loam-sand 28.5 19.32 0.8528
37 Turbic Mollic Cryosol (Ornithic, Oxyaquic, Arenic) 2.5Y 5/3 loam-sand 35.0 9.46 1.5996
38 Turbic Histic Cryosol (Ornithic, Reductaquic) 2.5Y 7/2 loam-clay 33.9 5.93 1.8839
39 Turbic Molic Leptic Cryosol (Ornithic, Arenic) 10YR 6/3 loam-sand 40.5 9.68 1.1501
40 Turbic, Leptic Cryosol (Ornithic, Reductaquic, Arenic) 7.5YR 2.5/2 loam-sand 28.5 4.93 1.7082
41 Turbic Leptic Cryosol (“Patterned”, Ornithic) 2.5Y 6/2 loam-sand 37.1 15.25 0.6525
42 Turbic Hyperskeletic Cryosol (Ornithic) 10YR 4/2 loam-sand 41.8 26.74 0.5456
43 Turbic Folic Cryosol (Ornithic, Arenic) 10YR 5/4 loam-sand 37.4 2.99 0.5005
44 Turbic Leptic Cryosol (Eutric, Arenic) 2.5Y 7/4 loam 37.9 0.24 0.3486
45 Haplic Leptosol (Ornithic, Gelic) 2.5Y 7/4 sand 36.5 0.46 0.2495
46 Haplic Leptsol (Ornithic, Skeletic) 10YR 5/2 loam-sand 46.2 8.50 2.1064
47 Hyperskeletic Leptosol (Ornithic, Gelic) 2.5Y 3/3 sand 38.9 1.78 0.6951
48 Hyperskeletic Leptosol (Ornithic) 2.5Y 7/3 sand 35.5 7.67 0.5785

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
92
Table 2: Soil mapping units considering the World Reference Base for Soil Resources (ISSS, 2006), area, soil carbon stock and soil organic carbon poll for each unit.
Area C Stock C Stock SOCP
Tabla 2: Unidades de suelo mapeado, considerando el World Reference Base for Soil Resources (ISSS, 2006),área, stock de carbono en el suelo y carbono orgánico del suelo para cada unidad.
ha kg m-2 Mg ha-1 Mg
Haplic Arenosol (Eutric, Gelic) 63.48 0.4475 0.0447 2.84 Haplic Arenosol (Eutric, Gelic)+Lithic Haplic Leptosol (Eutric, Gelic) 358.42 0.2254 0.0225 8.08 Protic Arenosol (Eutric, Gelic) 691.49 0.2282 0.0228 15.78 Epiturbic Cambic Cryosol (Eutric, Novic) 66.80 0.1803 0.0180 1.20 Turbic Leptic Cryosol ("Patterned", Eutric) 119.33 0.3660 0.0366 4.37 Turbic Leptic Cryosol (Eutric) 207.61 0.3660 0.0366 7.60 Turbic Leptic Cryosol (Eutric, Arenic) 130.08 0.3486 0.0349 4.54 Turbic Leptic Cryosol (Eutric, Arenic)+Leptic Cambisol (Eutric, Gelic) 132.49 0.1367 0.0137 1.81 Turbic Leptic Cryosol (Eutric, Reductaquic, Arenic) 167.67 0.0935 0.0093 1.57 Turbic Leptic Cryosol (Ornithic, Arenic)+Haplic Leptosol (Ornithic, Gelic) 3.81 0.6508 0.0651 0.25 Turbic Leptic Cryosol (Ornithic, Reductaquic, Arenic)+Histic Leptic Cryosol (Ornithic, Reductaquic) 8.37 1.2805 0.1281 1.07 Turbic Molic Leptic Cryosol (Ornithic, Arenic)+Turbic Leptic Cryosol ("Patterned", Ornithic) 5.10 0.9013 0.0901 0.46 Turbic Mollic Cryosol (Ornithic, Oxyaquic, Arenic) 7.23 1.7417 0.1742 1.26 Leptic Cryosol (Eutric, Oxyaquic)+Histic Leptic Cryosol (Eutric, Reductaquic, Arenic) 117.23 0.4444 0.0444 5.21 Turbic Cambic Cryosol ("Patterned", Calcaric) 78.85 0.3252 0.0325 2.56 Turbic Cambic Cryosol ("Patterned", Calcaric, Siltic) 56.71 0.3508 0.0351 1.99 Turbic Cryosol ("Patterned", Eutric) 95.85 0.6632 0.0663 6.36 Turbic Cryosol ("Patterned", Eutric, Skeletic) 60.83 0.4560 0.0456 2.77 Turbic Cryosol ("Patterned", Eutric, Skeletic)+Turbic Leptic Cryosol (Eutric, Arenic) 267.15 0.3978 0.0398 10.63 Turbic Cryosol (Ornithic, Oxyaquic, Arenic) 6.89 1.7417 0.1742 1.20 Turbic Folic Cryosol (Ornithic, Arenic) 6.45 0.4246 0.0425 0.27 Haplic Gleysol (Colluvic, Gelic) 243.83 0.8769 0.0877 21.38 Haplic Leptosol (Eutric, Gelic)+Haplic Arenosol (Ornithic, Gelic) 91.20 0.2079 0.0208 1.90 Haplic Leptosol (Ornithic, Gelic, Skeletic) 9.45 0.9074 0.0907 0.86 Total 2996.31
105.95
CONCLUSIONS More than 75% of the soils of Fildes Peninsula and Ardley Island are cryoturbated and have moderate SOC, Ice-free areas are formed mainly by Cryosols and Arenosols and considerable variability was found in relation to soil taxon. Sites colonized by the fauna preserve the most important SOCP of the in ice-free areas at the studied sites. SOCP was also high for Gleysols, where accumulation and preservation of SOC is favored. The studied sites conserve a SOCP of 105.97 Mg over an area of 2996.31 ha. ACKNOWLEDGEMENTS This study was supported technically and logistically by the Brazilian Navy, MMA, and UFV; grants from CNPq. We also thank INACH for technical and logistical support during field activities, and AIRFORCE project conducted by FEM and Airborne Technologies. REFERENCES Bockheim, J. 1990. A review of pedogenic zonation in well-drained soils of the Southern Circumpolar Region. Quaternary Research, 34(1): 47-66. Simas, F., Schaefer, C., Melo, V., Albuquerque Filho, M., Michel, R. F. M., Pereira, V., et al. 2007. Ornithogenic cryosols from Maritime Antarctica: Phosphatization as a soil forming process. Geoderma, 138(3-4): 191-203. Francelino, M. R., Schaefer, Carlos Ernesto G.R., Simas, F. N. B., Filho, E. I. F., Souza, J. J. L. L. de, & Costa, L. M. da. 2011. Geomorphology and soils distribution under paraglacial conditions in an ice-free area of Admiralty Bay, King George Island, Antarctica. Catena, 85(3): 194-204.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
93
EL ANÁLISIS EXPLORATORIO DE LOS DATOS DE TEMPERATURA DEL SUELO EN SEIS SITIOS DE MONITOREO DE LA PENÍNSULA KELLER, ISLA REY JORGE
(Exploratory data analysis of soil temperature at six sites monitoring the Keller Peninsula, King George Island)
Salamene, S.1; Francelino, M. R.2; Lyra, G. B.2 e Schaefer, C.E.R.G.3
1Instituto Federal de Educação, Ciência e Tecnologia de São Paulo; Caraguatatuba, São Paulo, Brasil, [email protected]; 2Instituto de Floresta, Universidade Federal Rural do Rio de Janeiro, BR 465 km 07 Seropédica, Rio de Janeiro, Brasil, [email protected]; 3Departamento de Solos, Universidade Federal de Viçosa, MG, Brasil. [email protected] INTRODUCCIÓN Técnicas modernas desenvolvidas para obtenção e armazenamento automático de dados, empregadas nos sítios de monitoramento da camada ativa dos solos da Antártica, produziram grande quantidade de dados, que contribuem para o entendimento dos processos relacionados à temperatura do solo na região (Vieira et al., 2010). A análise exploratória de dados visa extrair as informações mais importantes a partir desses dados, além de orientar na aplicação de análises mais complexas e avançadas para o aprofundamento da pesquisa, dando suporte para conclusões mais confiáveis (LANDIM, 2011). Na Antártica, a temperatura do solo, por estar relacionada a vários atributos ambientais, sendo assim, pode constituir-se um eficiente indicador do efeito das mudanças climáticas na região (Beyer et al., 1999). Enquanto o clima influencia a temperatura do solo numa escala global, localmente a temperatura do solo é influenciada principalmente pela topografia e propriedades do solo (GRUBER & Haeberli, 2009). Em ambientes periglaciais, a altitude é inversamente proporcional à espessura da camada ativa. Assim, num gradiente altitudinal, o permafrost tende a estar mais próximo da superfície do solo nas regiões de maior altitude (Gruber & Haeberli, 2009). Outro fator determinante na temperatura do solo é a cobertura vegetal. Na Antártica, a cobertura vegetal protege o solo das variações térmicas, com efeito isolante ou tampão (Kejna & Laska 1999; Cannone & Guglielmin, 2009). Isso ocorre porque os processos de troca de energia entre o solo e o ar são mais diretos e efetivos em superfícies marcadas pela ausência de vegetação (Mendonça & Oliveira, 2007). A intensidade do isolamento térmico da vegetação irá depender da estrutura, densidade e espessura dessa cobertura no solo. O objetivo desse trabalho foi comparar os dados de diferentes sítios de monitoramento da camada ativa na península Keller, através de uma análise exploratória dos dados obtidos nos sítios entre os anos de 2008 e 2011. Buscou-se discutir as possíveis relações entre a temperatura do solo e outros fatores ambientais locais, como altitude, tipo de solo e presença de cobertura vegetal. MATERIALES Y MÉTODOS O estudo foi realizado na Península Keller, na Ilha Rei George, onde foram instalados seis sítios de monitoramento da camada ativa em locais distintos quanto às características de solos, altitude e cobertura vegetal. São eles: Cruzes, VLF (Very Low Frequency), Ponta Yellow, Platô Tyrrel, Ipanema e Refúgio II (Figura 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
94
Figura 1. Ubicación de los sitios de monitoreo en la península Keller.
Figure 1. Location of monitoring sites in Keller Peninsula. Para cada sítio foram instalados sensores de temperatura do solo (modelo 107 Temperature Probe, Campbell Scientific Inc.) em diferentes profundidades ao longo do perfil de solo, considerando a distinção entre horizontes e/ou camadas. Foram utilizados três sensores em Cruzes, VLF, Yellow e Ipanema, e quatro sensores em Tyrrel e Refúgio 2. Em quatro sítios (Yellow, Ipanema, Tyrrel e Refúgio) foi instalado também um sensor de umidade do solo (modelo CS616-L, Campbell Scientific Inc.) no último horizonte/camada do solo. Os sensores foram acoplados a data logger modelos CR10, CR21X e CR1000, todos da Campbell Scientific Inc. os quais foram programados para registrar os dados de temperatura e umidade do solo a cada hora. Foram calculadas as médias diárias da temperatura do solo nas diferentes profundidades de cada sítio de Keller. Posteriormente, realizou-se a análise de agrupamento, utilizando-se o método Ward (variância

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
95
mínima), tendo como medida de dessimilaridade a distância euclidiana, para identificar os sítios com variabilidade mensal homogênea quanto à temperatura a 10 cm de profundidade.
RESULTADOS Y DISCUSIÓN A análise de agrupamento (Figura 2) permitiu identificar dois grupos com padrões de variação homogênea de temperatura do solo a 0,10 m de profundidade, e um sítio com padrão independente. Observou-se variabilidade homogênea entre os sítios de VLF, Yellow e Refúgio, compondo o denominado de grupo 1, e entre os sítios de Tyrrel e Ipanema, que formaram o grupo 2. Cruzes apresentou um regime térmico da camada ativa significativamente diferente dos demais sítios.
figura 2. agrupación de los sitios de acuerdo a la temperatura del suelo promedio mensual en 10 cm de profundidad.
figure 2. grouping of sites according to monthly average soil temperature at 10 cm depth.
No grupo 1, a principal característica em comum dos três sítios é a altitude. VLF e Refúgio estão em altitudes próximas (49 e 45, respectivamente) e apresentam o mesmo tipo de solo (Cryosol lhitic). Yellow está numa altitude inferior (31 m) e seu solo (Leptosol gelic) é diferente dos demais sítios do seu grupo e, assim como VLF, está localizada na face leste de Keller. Uma característica ambiental que difere VLF dos demais sítios do seu grupo é a presença da cobertura vegetal, que nesse local é rara, composta predominante de tufos isolados de briófitas e liquens, enquanto no Yellow e Refúgio a vegetação é mais densa e diversificada, com presença inclusive de gramíneas. O grupo 2 é composto pelos sítios Tyrrel e Ipanema. Tyrrel está localizado na maior altitude (196m) em relação a todos os sítios e por isso esperava que esse local tivesse a menor temperatura do solo, onde é mais frio e ocorre maior deposição de neve. No entanto, esse sítio teve a temperatura similar à encontrada em Ipanema, com 89 m de altitude. Ipanema apresenta cobertura vegetal densa, com presença marcante de gramíneas, que funcionam como isolante térmico e que pode ter contribuído na variabilidade homogênea da temperatura do solo com o sítio Tyrrel, apesar da diferença altitudinal de 107 metros entre eles.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
96
Nos sítios de maiores altitudes, a camada de neve formada tende a ser mais espessa (Gruber & Haeberli, 2009) do que nos sítios de menor altitude. Como a neve funciona como isolante térmico, durante o inverno, o solo dos sítios de maior altitude transferem menos calor para a atmosfera em relação aos de menores altitudes, e assim, as temperaturas do solo mínima desses sítios é superior aos de menor altitude. No verão, a neve demora mais a derreter nos sítios de maior altitude, e quando derrete disponibilizam um maior volume de umidade para o solo, devido à camada de neve mais espessa. Esses processos contribuem para uma menor transferência de calor para o solo, além da maior umidade que contribui para o aumento do calor específico do solo, que associados resultam em menor aquecimento no verão e, dessa forma, temperaturas do solo máximas inferiores aos solos dos sítios de menor altitude. Cruzes, com 28m de altitude, não apresentou variabilidade homogênea com nenhum dos grupos formados e mostrou média de temperatura do solo mais elevada (-0,25 a -0,04º C). Isso pode ter ocorrido devido à influência antrópica no regime térmico desse sítio em função da sua proximidade com Estação Antártica Comandante Ferraz, além do fato desse sítio estar isolado por barreiras naturais, com a escarpa do Morro Flagstaff a oeste e uma grande moraina ao sul. A formação dos solos na Antártica, em função suas características ambientais extremas, ocorreram através de um processo lento. Por isso, a mínima perturbação humana pode acarretar numa resposta significativa nesses ambientais, principalmente nos solos mais jovens localizados nas regiões de menor altitude (BEYER et al., 1999). CONCLUSIONES A análise exploratória permitiu identificar os sítios com características homogêneas de temperatura do solo, formando diferentes grupos quanto ao regime térmico da camada ativa. O primeiro grupo foi formado por VLF, Yellow e Refugio, tendo como característica principal em comum a altitude, que varia de 31 a 49 m. Os sítios de Tyrrel e Ipanema compõem o grupo 2, que foi marcado pelo isolamento térmico em decorrência, respectivamente, da cobertura de neve e cobertura vegetal. Cruzes foi o sítio de Keller mais quente, com regime térmico da camada ativa diferente dos demais sítios, provavelmente em função de sua proximidade com a estação Ferraz.
AGRADECIMIENTOS Ao CNPq pela bolsa de doutorado e a Marinha Brasileira pelo apoio logístico.
REFERENCIAS BIBLIOGRÁFICAS Beyer, L.; Bockheim, J.G.; Campbell, I.B.; Claridge, G.G.C. 1999. Review Genesis, properties and sensitivity of Antarctic Gelisols. Antarctic Science, 11(4), 387–398. Cannone, N., & Guglielmin, M. 2009. Influence of vegetation on the ground thermal regime in continental Antarctica. Geoderma, 151(3-4), 215–223. Gruber, S. & Haeberli, W. 2009. Mountain Permafrost. In: MARGESIN, R. (ed.) Permafrost Soils. Berlin: Spring, 348p. Kejna, M. & Laska, K. 1999. Spatial differentiation of ground temperature in the region of Arctowski Station, King George Island, Antarctica in 1996. Polish Polar Research, v.20, n. 3, p.221-241. Landim, P.M.B. 2011. Análise estatística de dados geológicos multivariados. São Paulo: Oficina de Textos. 208p. Medonça, F. & Oliveira, I.M.D. 2007. Climatologia. São Paulo: Oficina de textos, 206p. Vieira, G.; Bockheim, J.; Guglielmin, M. et al. 2007. Thermal State of Permafrost and Active-layer Monitoring in the Antarctic: Advances During the International Polar Year 2007 – 2009. Permafrost and Periglacial Process, v. 21, p. 182–197.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
97
ACTIVIDAD ANTICONGELANTE DE PÉPTIDOS SINTÉTICOS DE MOTIVOS SECUENCIALES DE PROTEINAS ANTICONGELANTES
(Antifreeze activity of synthesized peptides with sequential motifs of natural antifreeze proteins) Rojas, R.E.1, Carvajal-Rondanelli, P.2,Marshall, S.H.3, Guzmán, F.4.
1Instituto de Química Pontificia Universidad Católica de Valparaíso (PUCV). [email protected] 2Escuela de Alimentos, PUCV 3Instituto de Biología PUCV 4Núcleo de Biotecnología Curauma (NBC), PUCV RESUMEN Las proteínas anticongelantes (AFPs) están presentes en peces que habitan las zonas polares, permitiéndoles sobrevivir en ambientes a temperaturas bajo cero. Esta propiedad física se considera de enorme importancia en la criopreservación de células, tejidos y organismos. Existen distintos tipos de estructura secundaria para las AFPs. En el presente estudio se sintetizó y caracterizó 4 péptidos lineales conteniendo los motivos Ala-Ala-Thr de los AFPs, reemplazando la Thr por Arg e incorporando residuos de Cys en el amino terminal. Los péptidos fueron sintetizados en fase solida mediante metodología Fmoc. Se demostró que la adición de cisteína (Cys) en el terminal amino de los péptidos incrementa la configuración β-turn, medida por Dicroismo Circular (CD), la cual incrementa la actividad anticongelante, efecto medido por Calorimetría Diferencial de Barrido (DSC). La actividad anticongelante se manifiesta a través de la inhibición de la recristalización del hielo de forma no coligativa, fenómeno denominada histéresis térmica (THA). Este trabajo genera lineamientos para crear estructuras análogas a AFGPs de síntesis más simple y de un menor costo, caracterizando modificaciones puntuales de péptidos y su potencial efecto sobre la inhibición del crecimiento de cristales de hielo durante el proceso de congelación. INTRODUCCIÓN La formación de cristales de hielo y su crecimiento desmedido generan una serie de problemas en la conservación de células, tejidos y fluidos biológicos. Una manera de prevenir tales cambios es a través del uso de moléculas supresoras del crecimiento de los cristales de hielo que son sintetizadas por los peces que viven en regiones polares. Las proteínas anticongelantes (AFPs) son un grupo de moléculas con el mejor potencial de inhibición de los cristales de hielo (Bang, 2013). El aislamiento de estas moléculas de peces polares ó la obtención a través de la síntesis química es de alto costo. La alternativa se enfoca en sintetizar moléculas de corto tamaño, de estructura simple y con un poder inhibitorio de cristalización similar o mejor al generado por los AFPs naturales. Este estúdio comparará las estructuras de péptidos lineales con motivos anticongenlantes de las AFPs, sustituciones de Arg en estos péptidos e inserciones de Cys en estas cadenas para determinar su efecto en la capacidadad anticongelante. MATERIALES Y MÉTODOS 2.1 Síntesis química y purificación de péptidos: Los péptidos (AAATAAATAAA, CAAATAAATAAA, AAARAARAAA y CAAARAAARAAA) fueron sintetizados mediante síntesis en fase sólida (Houghten RA., 1985), usando la metodología Fmoc con resina Rink amida con sustitución de 0,64 eq/gr, y aminoácidos protegidos con Fmoc/tBu en los casos pertinentes. Estos péptidos fueron clivados de la resina con una solución de ácido trifluoroacético (TFA), triisopropilsilano (TIS) y agua en razón 0.95, 0.025, 0.025 por un tiempo de 1 hora, y luego precipitados utilizando dietiléter . El crudo obtenido fue desalinizado por cromatografía de exclusión molecular, y los péptidos fueron analizados determinándose por RP-HPLC (Corporación Jasco, Tokio, Japón) una pureza del 95% y su masa

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
98
molecular fue confirmada mediante HPLC-MS y MALDI-TOF (Jofré et al., 2011). Los péptidos fueron liofilizados y almacenados hasta su uso. 2.2Espectroscopia de Dicroismo Circular (CD): Se empleó un equipo de Dicroismo Circular Jasco J-815 (Corporación Jasco, Tokio, Japón) equipado con un dispositivo de control de temperatura Peltier (Jasco CDF-426S/15).Los espectros fueron registrados a una velocidad de escaneo de 50 nm/min, con tiempo de respuesta de 4s, ancho de banda de 1 nm y 3 acumulaciones. Para el análisis se utilizaron 250µLde muestra de concentración 1,1mM en una cubeta rectangular de cuarzo con un paso óptico de 1mm; se corrigió la línea base y los datos fueron convertidos a elipticidad molar. 2.3 Calorimetría Diferencial de Barrido (DSC): Se empleó un equipo DSC 1 (Mettler Toledo, Suiza) con sensor HSS8 de sensibilidad ±1µWy un enfriador internoHuber TC45 (Huber, Alemania).Se utilizó un crisol de aluminio vacío de 40 µLsin pin para la calibración de la línea base; como control de calibración se utilizó indio y zinc antes de las mediciones de histéresis térmica (THA). Se empleó muestras de 10 µL de solución de péptido a distintas concentraciones (2, 10, 15 y 25mM) en crisoles de aluminio. La masa de las muestras fue registrada usando balanza analítica de precisión. Como referencia se utilizó un crisol vacío de aluminio. La medición de la muestra se realizó de acuerdo a la metodología descrita en la Tabla 1.
Paso T° Inicial (°C) T° Final (°C) Razón de cambio(°C/min) Tiempo(min)
1 10 -25 2 17,5 2 -25 10 1 35,0
Tabla 1: Metodología preliminar DSC. Table 1: Preliminary DSC Methodology.
Una vez determinado tanto el punto de fusión como la entalpia del proceso, se procede a elegir una temperatura de mantenimiento (THo), a fin de obtener una fusión parcial de la muestra superior al 95%, lo que permite tener el mínimo de puntos de nucleación y un efecto máximo del péptido sobre éstos, de acuerdo a la Tabla 2.La temperatura de inicio de la recristalización (To) y el área exotérmica del proceso fueron registradas para estimar la actividad anticongelante de los péptidos.El porcentaje de fusión de las muestras (%N) fue estimado con la siguiente fórmula: %N=[1 – (-∆Hf/∆Hm)] x 100%, donde ∆Hm es la entalpia de fusión y ∆Hfes la entalpia de recristalización de la muestra. La diferencia entre THo y To representa la histéresis térmica (THA).
Paso T° Inicial (°C) T° Final (°C) Razón de cambio(°C/min) Tiempo(min) 1 10 -25 3 17,5 2 -25 -10 3 35,0 3 -10 THo 1 4 THo THo 0 10,0 5 THo -10 1 6 -10 -25 3 5,0 7 -25 10 3 11,7
Tabla 2: Metodología de medición de actividad anticongelante mediante DSC Table 2: Methodology antifreeze activity measurement by DSC
RESULTADOS Y DISCUSIÓN El análisis de DC para los péptidos en distintos solventes a 15°C muestra que la adición de una cisteína en el extremo amino favorece la estructuración del péptido en los distintos solventes, siendo este efecto mayor en los péptidos que presentan arginina (Arg) en su secuencia que en los que poseen treonina (Thr).(Figura 1)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
99
Figura 1: Espectros CD de los péptidos en diferentes solventes. (A)Elpéptido AAARAAARAAA en aguay tampón muestra
una estructura de poliprolina II (PPII), la cual se transforma a estructura tipoβ-turnen TFE(30%). (B)El péptido AAATAAATAAA muestra una estructura de (PPII) de mayor intensidad a la anterior (A). (C) Comparación de los espectros
de los péptidos de (A) y (B) en solución tampón. (D) El péptido CAAARAAARAAA forma estructura tipo tipo β-turn en agua y solución tampón, la cual se acrecienta en TFE(30%). (E) El péptido CAAATAAATAAA presenta una estructura tipo
β-turnen agua y solución. (F) Comparación de los espectros de (A) y (D) en agua. Figure 1: CD spectra of analogue peptides at 15 °C in buffer, water, and TFE. (A) AAARAAARAAA shows a polyproline II (PPII) structure in water and buffer, but it changes to a type beta-turn in TFE (30%). (B) AAATAAATAAA shows a more
intense PPII structure than peptide in (A). (C) CD spectra comparison of peptides (A) and (B) in buffer.(D) CAAARAAARAAA forms a type beta-turn structure in water and buffer. (E) CAAATAAATAAA exists in a type beta-turn
in water and buffer. (F) CD spectra comparison of peptide (A) and (D) in water. Los datos de DSC para los péptidos son presentados en la figura 2 y corresponden a la histéresis máxima para estas muestras. Los péptidos sin cisteína presentan una baja histéresis térmica la cual no se incrementa a medida que se aumenta su concentración. En cambio los péptidos con incorporación de cisteína presentan un incremento gradual de la histéresis térmica a medida que se aumenta su concentración.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
100
Figura 2 Actividad anticongelante (histéresis térmica) para péptidos entre 2-25mM.
Figure 2: Antifreeze activity (thermal hysteresis) for peptides between 2-25 mM. CONCLUSIONES Este trabajo demuestra que los péptidos lineales con Arg análogos a los AFGPs presentan una estructura tipo β-turn más definida que su contraparte con residuos de Thr. La incorporación de cisteína a su secuencia incrementa la formación de una estructura tipo β-turn, en especial en los péptidos con sustituciones de Arg. La inhibición de la recristalización medida por DSC fue mayor en los péptidos que se incorporó Cys en su secuencia y efecto que se determinó cuando la fracción de hielo remanente fue inferior al 5%, mostrando un retraso en la recristalización comparado con los controles, demostrando una actividad anticongelante de forma no coligativa (histéresis térmica). AGRADECIMIENTOS Proyecto Fondecyt 110056. REFERENCIAS Bang J.,Lee J.,Murugan R.,Lee S.,Do H.,Koh H.,Shim H.,Kim H.y Kim H. 2013.Antifreeze Peptides and Glycopeptides, and Their Derivatives: Potential Uses in Biotechnology. Drugs,11(6), 2013-2041. Jofré C., Guzmán F., Cárdenas C., Albericio F. y Marshall S.H.2011. A natural peptide and its variants derived from the processing of infectious pancreatic necrosis virus (IPNV) displaying enhanced antimicrobial activity: A novel alternative for the control of bacterial diseases. Peptides, 328:852-858. Houghten RA. 1985. General method for the rapid solid-phase synthesis of large numbers of peptides: specificity of antigen-antibody interaction at the level of individual amino acids. Proceedings of the National Academy of Sciences of the United States of America, 82: 5131-5135.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
101
CARACTERIZACIÓN MOLECULAR Y ANÁLISIS DE LA PRODUCCIÓN DE METABOLITOS CON POTENCIAL BIOTECNOLÓGICO DE PROBABLES CEPAS DE X.
DENDRORHOUS AISLADAS DESDE EL TERRITORIO ANTARTICO CHILENO (Molecular characterization and metabolite with biotechnological potential production analysis of
Xanthophyllomyces dendrorhous likely strains isolated from the Chilean Antarctic Territory)
Contreras G.1, Barahona S.1, Baeza M.1, Cifuentes V.1, Alcaíno J.1
1Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile. Las Palmeras 3425, Ñuñoa, Santiago.
INTRODUCCIÓN La Antártica es un ambiente hostil para distintas formas de vida y algunos microorganismos han desarrollado diferentes adaptaciones para enfrentar este medio ambiente. En respuesta adaptativa al frío, se ha observado que en las membranas hay un aumento en el contenido de ácidos grasos polinsaturados (PUFAs) que regularían su fluidez (Robinson 2001), además también el ergosterol participaría en mantener la integridad de éstas (Zlatanov y col. 2001). Frente a la radiación ultravioleta, los microorganismos poseen variados mecanismos de fotoprotección, entre ellos, tienen moléculas capaces de absorber directamente la luz ultravioleta, como las micosporinas y/o moléculas antioxidantes como los carotenoides que disminuyen el estrés causado por la radiación (Roy 2000). Los PUFAs, el ergosterol, las micosporinas y los carotenoides son metabolitos de gran atractivo biotecnológico. Los PUFAs, como los omega-3 y omega-6, son esenciales y su suplementación previene enfermedades cardiovasculares, inflamatorias, entre otras (Simopoulos 2002). Por otra parte, el ergosterol al ser consumido y por efecto de la radiación UV, es convertido a ergocalciferol o vitamina D2 y su deficiencia provoca raquitismo en niños (Holick 2007). Con respecto a las micosporinas, éstas actuarían como filtros fotoprotectores de radiación UV (Carreto y col. 2011) lo que resulta promisorio para su utilización en la formulación de bloqueadores solares para la protección de la piel. Por otra parte, los carotenoides son antioxidantes y entre ellos destaca la astaxantina, cuya actividad antioxidante se ha estimado hasta 10 veces mayor que el β-caroteno (Miki 1991). Por otra parte, la astaxantina es la responsable del color rojo-anaranjado de una variedad de organismos marinos, la cual es adquirida a través de su alimento. En la salmonicultura se requiere para la pigmentación roja anaranjada de la carne del salmón, y los productores de salmones invierten sobre el 15% del costo total de producción en este pigmento (Higuera-Ciapara y col. 2006). La astaxantina es producida por pocos microorganismos y la levadura Xanthophyllomyces dendrorhous es uno de ellos. X. dendrorhous (estado asexual: Phaffia rhodozyma) ha sido aislada desde regiones frías del planeta, como Alaska, Rusia, Japón, entre otros (Phaff y col. 1972) y recientemente las regiones VIII, IX y X de Chile (datos no publicados). En la XLVI Expedición Científica Antártica (ECA) del año 2010, se tomaron muestras de tierra de la península de Barton de la Isla Rey Jorge de la Antártica Chilena, y desde ellas se encontraron 13 aislados con fenotipo similar a X. dendrorhous. Como respuesta adaptativa, se esperó que estos aislados posean moléculas fotoprotectoras, como los carotenoides y/o micosporinas. Además, de tener un alto contenido de PUFAs y un cambio en la cantidad de ergosterol en comparación a X. dendrorhous aisladas de otras regiones. El objetivo de esta investigación fue Caracterizar molecularmente las probables cepas de X. dendrorhous aisladas desde el territorio Sub-Antártico Chileno y analizar la producción de metabolitos con potencial biotecnológico.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
102
MATERIALES Y MÉTODOS Cepas y aislados utilizados: ANCH1-ANCH13: X. dendrorhous aisladas de muestras de tierra de la Isla Rey Jorge de la Antártica Chilena. UCD 67-385: Cepa de X. dendrorhous aislada de Japón y AVNH2: Cepa de X. dendrorhous aislada de hoja de Gevuina avellana (avellano) de VIII Región de Chile. Las cepas se cultivaron en medio completo YM y para evaluar el desarrollo de holobasidios éstas se cultivaron en medio DWR (Kucsera y col. 1998) y MMv (Retamales y col. 2002). La amplificación de las regiones ITS, IGS y D1/D2 se realizó según Reyes y col. 2004. Las secuencias obtenidas se compararon mediante BLAST en la base de datos de GeneBank. Análisis de micro/mini satélite-PCR se realizó según Libkind y col. 2003. La extracción de ergosterol, micosporinas y carotenoides, se realizaron según Shang y col. 2006, Volkmann y col. 2006 y An y col. 1989, respectivamente. Para el análisis de ácidos grasos (FA), se extrajo los lípidos totales según Bligh y col. 1959 y luego se evaluó su composición mediante cromatografía de gas en la Empresa Fundación Chile, GCL. RESULTADOS Caracterización molecular de los probables aislados de X. dendrorhous. Para identificar estas cepas como X. dendrorhous se amplificó y secuenció las regiones IGS, ITS y D1/D2 de los 13 aislados ANCHs. Las secuencias obtenidas fueron idénticas entre ellas y con la cepa nativa AVHN2. Estas secuencias se compararon con la base de datos GenBank y se confirmó que corresponden a X. dendrorhous/P. rhodozyma. Con el fin de discriminar si estos aislados son cepas distintas, se analizó mediante la técnica de fingerprinting de micro/mini satélite-PCR (Libkind y col. 2003). Al amplificar el DNA genómico de los 13 aislados ANCHs y de AVHN2 con los partidores M13, (GTG)5 y (GAC)5 por separado, se obtuvo igual patrón de bandeo, no así cuando se utilizó DNA de la cepa UCD 67-385 como molde. Por otra parte, se evaluó la presencia de elementos genéticos extracromosómicos. Los 13 aislados no contienen estos elementos, mientras que AVHN2 y UCD 67-385 si los poseen (Baeza y col. 2009). Con el objetivo de determinar si estos aislados corresponden a X. dendrorhous o al estado asexual, P. rhodozyma, se evaluó la presencia de estructuras sexuales (holobasidios con basidioesporas) de 3 de los aislados en placas de medios mínimo MMv y DWR. Los 3 aislados desarrollaron este tipo de estructuras por lo tanto, son homotálicas y corresponden a X. dendrorhous. Debido a que mediante estrategias moleculares no fue posible diferenciar estos aislados a nivel de cepas, éstos se agruparon de acuerdo a su fenotipo de pigmentación: roja, anaranjada o amarilla, y se escogió un representante de cada grupo: ANCH1, ANCH6 y ANCH8, respectivamente. Determinación de las condiciones óptimas de crecimiento y estudio de la tolerancia a la radiación UV de las cepas de X. dendrorhous aisladas desde la Antártica Chilena. En primer lugar se evaluó el crecimiento de ANCH1, ANCH6 y ANCH8 a 22, 15 y 10 °C en placas de medio YM. Para ello se fotografió las placas y se midió el área de la colonia en pixeles, durante 9 días. Los resultados revelan que ANCH1 y ANCH6 crecen mejor a 22°C, mientras que ANCH8 a 15°C. Luego, se comparó la cinética de crecimiento a 22°C en medio líquido. Los tiempos generacionales fueron de 5,75; 6,02 y 4,54 h, respectivamente. Se estudió la tolerancia a luz UV-B (310 nm) de los tres aislados en comparación con las cepas AVHN2 y UCD 67-385. Los aislados ANCH6 y ANCH8 presentaron un mayor porcentaje de sobrevida luego de 3 h de exposición: 71,2±0,7 % y 62,6±10,7 %, respectivamente. Mientras que UCD 67-385, ANCH1 y

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
103
AVHN2 tienen un porcentaje de sobrevida similar y menor (39,1±11,3 %; 46,7±12,7 % y 40,2±3,9 %, respectivamente). Análisis de producción de carotenoides, micosporina, ergosterol y FA de los aislados de X. dendrorhous de la Antártica Chilena. Se evaluó la producción de carotenoides, micosporinas y ergosterol y ácidos grasos (FA) en ANCH1, ANCH6 y ANCH8, y se comparó con las cepas AVHN2 y UCD 67-385. En cuanto a la producción de carotenoides, la composición de la cepa ANCH1 es similar a AVHN2 y UCD 67-385. Sin embargo, la cantidad de carotenoides totales es de hasta 6 veces mayor en ANCH1. Entre las cepas ANCH6 y ANCH8 no hay diferencias significativas en la cantidad de este tipo de pigmentos. Sin embargo, el carotenoide mayoritario es β-caroteno en oposición a las otras cepas, el cual es astaxantina (figura 1). En cuanto a la producción de micosporina, las cepas UCD 67-385 y AVHN2 producen la micosporina glutaminol glucósido. Contrario a lo esperado, en ANCH1, ANCH6 y ANCH8 no se encontraron este tipo de moléculas. En relación a la producción de ergosterol, las cepas ANCH1 y ANCH8 no presentan diferencias significativas con respecto a las cepas AVHN2 y UCD-67-385. Sin embargo, ANCH6 produce hasta 3 veces menos que las otras cepas. En cuanto a la producción de FA, análisis preliminares indican que ANCH8 posee 15,7% más de ácidos monoinsaturados y 6,7% más de ácidos polinsaturados (PUFAs) en relación a los ácidos grasos totales. DISCUSIÓN Y CONCLUSIONES La producción de carotenoides, y en consecuencia de astaxantina, de la cepa ANCH1 es de hasta 6 veces mayor en comparación a otras cepas silvestres, lo que la hace un buen candidato para realizar estrategias de mejoramiento genético y optimización de condiciones de cultivo para favorecer la síntesis de astaxantina. Las cepas de X. dendrorhous aisladas de la Antártica Chilena presentan una mayor tolerancia a la luz UV-B (ANCH6 y ANCH8). Sin embargo, no poseen MAAs y por lo tanto podrían poseer otros sistemas de fotoprotección aún no identificados.
Figura 1: Producción de carotenoides luego de 5 días de cultivo a 22 °C de las cepas de X. dendrorhous aisladas desde el
territorio sub-antártico chileno (ANCH1, ANCH6 y ANCH8), región del Biobío de Chile (AVHN2) y Japón (UCD 67-385). Figure 1:arotenoid production from 5 days-old yeast cultures grown at 22ºC of X. dendrorhous strains isolated from the
Chilean sub-Antarctic (ANCH1, ANCH6 y ANCH8), Biobío region in Chile (AVHN2) and Japan (UCD 67-385). AGRADECIMIENTOS Instituto Antártico Chileno, INACH. Proyectos M_01_11 (GC) y RG_07-12 (JA). REFERENCIAS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
104
An, G. H., D. B. Schuman, y cols. 1989. Isolation of Phaffia rhodozyma mutants with increased astaxanthin content. Applied and environmental microbiology, 55(1): 116. Baeza, M., M. Sanhueza, y cols. 2009. Polymorphism of viral dsRNA in Xanthophyllomyces dendrorhous strains isolated from different geographic areas. Virology Journal, 6(160): 1-7. Bligh, E. y W. J. Dyer. 1959. A rapid method of total lipid extraction and purification. Canadian journal of biochemistry and physiology, 37(8): 911-917. Carreto, J. I. y M. O. Carignan. 2011. Mycosporine-Like Amino Acids: Relevant Secondary Metabolites. Chemical and Ecological Aspects. Marine Drugs, 9(3): 387-446. Higuera-Ciapara, I., L. Felix-Valenzuela, y cols. 2006. Astaxanthin: a review of its chemistry and applications. Critical reviews in food science and nutrition, 46(2): 185-196. Holick, M. F. 2007. Vitamin D deficiency. New England Journal of Medicine, 357(3): 266-281. Kucsera, J., I. Pfeiffer, y cols. 1998. Homothallic life cycle in the diploid red yeast Xanthophyllomyces dendrorhous (Phaffia rhodozyma). Antonie van Leeuwenhoek, 73(2): 163-168. Libkind, D., S. Brizzio, y cols. 2003. Molecular characterization of carotenogenic yeasts from aquatic environments in Patagonia, Argentina. Antonie van Leeuwenhoek, 84(4): 313-322. Miki, W. 1991. Biological functions and activities of animal carotenoids. Pure Appl. Chem, 63(1): 141-146. Phaff, H., M. Miller, y cols. (1972). A comparative study of the yeast florae associated with trees on the Japanese islands and on the west coast of North America. Fourth International Fermentation Symposium Proceedings: Fermentation Technology Today, Society of Fermentation Technology, Osaka, Japan: 759-774 Retamales, P., G. Hermosilla, y cols. 2002. Development of the sexual reproductive cycle of Xanthophyllomyces dendrorhous. Journal of microbiological methods, 48(1): 87-93. Reyes, E., S. Barahona, y cols. 2004. Genetic polymorphism of clinical and environmental strains of Pichia anomala. Biol Res, 37(4 Suppl A): 747-57. Robinson, C. H. 2001. Cold adaptation in Arctic and Antarctic fungi. New phytologist, 151(2): 341-353. Roy, S. (2000). Strategies for the minimisation of UV-induced damage. The effects of UV radiation in the marine environment. S. D. S. de Mora, M. Verne, Cambridge Environmental Chemistry: 177-205. Shang, F., S. Wen, y cols. 2006. High-cell-density fermentation for ergosterol production by Saccharomyces cerevisiae. Journal of bioscience and bioengineering, 101(1): 38-41. Simopoulos, A. P. 2002. Omega-3 fatty acids in inflammation and autoimmune diseases. Journal of the American College of Nutrition, 21(6): 495. Volkmann, M. y A. A. Gorbushina. 2006. A broadly applicable method for extraction and characterization of mycosporines and mycosporine-like amino acids of terrestrial, marine and freshwater origin. FEMS Microbiology Letters, 255(2): 286-295. Zlatanov, M., K. Pavlova, y cols. 2001. Lipid composition of some yeast strains from Livingston Island, Antarctica. Folia microbiologica, 46(5): 402-406.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
105
ACTIVIDAD DE DIFERENTES ANTIBIOTICOS SOBRE CEPAS DE Escherichia coli AISLADAS DESDE MUESTRAS DE AGUA DE BAHÍA FILDES EN ISLA REY JORGE,
ANTÁRTICA. (Activity of different antibiotics against Escherichia coli strains isolated from water samples from Fildes
Bay in King George Island, Antarctica)
Rabbia, V.1, Jiménez, S.2, Bello, H.1, Domínguez, M.1, Molina, R.1, Vergara, L.3, Gómez-Fuentes, C.4, Calisto-Ulloa, N.4, González-Rocha, G.1
1Laboratorio de Investigación en Agentes Antibacterianos, Facultad de Ciencias Biológicas, U. de Concepción, Concepción. CHILE. 2Facultad de Ciencias Naturales y Oceanográficas, U. de Concepción, Concepción. CHILE. 3Facultad de Ciencia, Universidad San Sebastián, Concepción. CHILE. 4Departamento de Ingeniería Química, U. de Magallanes, Punta Arenas. CHILE. E-mail: [email protected] INTRODUCCIÓN Los antibióticos son sustancias químicas producidas naturalmente por microorganismos que inhiben el crecimiento de otros microorganismos (Davies, 2006). Estos compuestos han sido utilizados desde su descubrimiento hasta el presente, principalmente en humanos y animales, para tratar y prevenir infecciones bacterianas. Aunque la resistencia a antibióticos es un fenómeno natural (Martínez, 2012), su uso desmedido a nivel mundial ha generado ambientes con alta presión selectiva que favorecen la selección de bacterias resistentes (Dodd, 2012). El incremento acelerado de bacterias resistentes representa un peligro para los seres humanos y otros animales, concitando una gran preocupación, dado que la infección con un patógeno multiresistente o panresistente, aumenta la tasa de morbilidad y mortalidad (Allen y col., 2010). Se ha llegado a una situación en la que se cree estamos reentrando a la era pre-antibiótica, donde las infecciones bacterianas eran la principal causa de muerte. Otro problema asociado a la propagación de la resistencia es la pérdida de diversidad genética endémica del ambiente. El uso indiscriminado de antibióticos y la selección de patógenos resistentes han sido ligados, principalmente, a la actividad humana, por lo que en ambientes en que ésta se encuentra restringida, se espera baja frecuencia de bacterias resistentes. Este es el caso de la Antártica, continente sin población humana nativa y dónde la principal vía de entrada de microorganismos alóctonos es la migración de aves y la actividad humana que se desarrolla allí. Un buen indicador de la presencia de alguno de estos dos fenómenos en este ambiente remoto, son los indicadores de contaminación fecal (Stevens y col., 2003). Escherichia coli es el indicador de contaminación fecal más utilizado para evaluar calidad del agua potable y de aguas servidas tratadas (Stevens y col., 2003). Esta bacteria Gram negativa es habitante del tracto gastrointestinal de los mamíferos (Bonnedahl y col., 2008), donde puede adquirir genes de resistencia; por lo tanto, su perfil de resistencia es considerado un buen indicador del contacto que haya tenido con antibióticos en su hospedador. Se postula que la resistencia antibiótica en E. coli es mayor mientras mayor sea la cercanía con el hombre (Skurnik y col., 2006; Miller y col., 2009). Se ha informado que cepas de E. coli provenientes de animales antárticos carecen de resistencia a antibióticos (Miller y col., 2009); en cambio, en cepas aisladas de humanos de zonas urbanas, la resistencia se presenta, en promedio, en el 18% de las cepas (Skurnik y col., 2006; Tadesse y col., 2012). Por esta razón, la amplitud de los perfiles de resistencia y los tipos de antibióticos a los que las cepas son resistentes, permite teorizar sobre el probable origen de las cepas de E. coli en muestras de la Antártica. Debido a que Bahía Fildes tiene alta actividad antropogénica, es de interés estudiar la actividad de diferentes antibióticos sobre cepas de E. coli aisladas de agua de B. Fildes (Isla Rey Jorge, Islas Shetland

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
106
del Sur), resultados que podría ser comparados con la resistencia de microorganismos endémicos de Antártica y así determinar si existe transferencia génica a la población microbiana endémica. MATERIALES Y MÉTODOS Se trabajó con 119 cepas presuntivas de E. coli seleccionadas al azar desde placas de agar ChromoCult® Coliform Agar (Merck) utilizado para determinación de colimetría de muestras de agua, obtenidas de la Bahía Fildes durante la 49a Expedición Científica Antártica en enero de 2013. Se confirmó la identificación de las cepas mediante pruebas bioquímicas convencionales (escala bioquímica) (Edwards y Ewing, 1972). La actividad antibacteriana se determinó por el método de difusión en agar Müeller-Hinton (Oxoid) recomendado por el Clinical Laboratory Standards Institute (CLSI, 2012), utilizando los siguientes discos de antibióticos: ampicilina (AMP, 10 µg), cefalotina (CEP, 30 µg), cefotaxima (CTX, 30 µg), cefuroxima (CXM, 30 µg), ceftazidima (CAZ, 30 µg), cefepima (FEP, 30 µg), cefoxitina (FOX, 30 µg), estreptomicina (STR, 10 µg), kanamicina (KAN, 30 µg), gentamicina (GEN, 10 µg), amikacina (AMK, 30 µg), ácido nalidíxico (NAL, 30 µg), ciprofloxacina (CIP, 5 µg), tetraciclina (TET, 30 µg), cloranfenicol (CLO, 30 µg), sulfonamidas (SUL, 300 µg) y trimetoprim (TMP, 5 µg). Las cepas de E. coli fueron catalogadas como susceptible, intermedia o resistente según los estándares del CLSI (2012). Los antibióticos se clasificaron en los siguientes grupos: penicilinas (AMP), cefalosporinas (CEP, CTX, CXM, CAZ, FEP y FOX), aminoglucósidos (STR, KAN, GEN y AMK), quinolonas (NAL y CIP), tetraciclina (TET), cloranfenicol (CLO) y sulfonamida-trimetoprim (SUL y TMP). RESULTADOS De las 119 cepas presuntivas de E. coli, se confirmó mediante pruebas bioquímicas la identificación de E. coli en el 79,8 % de las cepas. Los resultados de la actividad antimicrobiana se presentan en la Figura 1. Se destaca el alto porcentaje de cepas no susceptibles (resistentes o intermedias) a STR (62,8 %), seguido por CEP (49,4 %) y TET (36,5 %). Todas las cepas fueron susceptibles a CLO. En la Figura 2 se observa el porcentaje de cepas no susceptibles a al menos un antibiótico de cada grupo, siendo los aminoglucósidos (69,4 %) y cefalosporinas (51,8 %) los grupos que presentaron los porcentajes más altos de cepas no susceptibles. En la Figura 3 se pueden observar los porcentajes de cepas que presentaron resistencia simultánea a distintos grupos de antibióticos. La mayoría de las cepas presentaron resistencia a un grupo de antibióticos (25,9 %), aunque un porcentaje levemente inferior se encontró para cepas resistentes a 2 y 3 grupos de antibióticos simultáneamente (24,7 %).
Figura 1: Perfil de susceptibilidad a los distintos antibióticos ensayados. En gris claro se observan las cepas susceptibles, en
gris oscuro las cepas intermedias y en negro las cepas resistentes. Figure 1:Susceptibility profiles to different antibiotics tested. In light gray: susceptible strains, in dark gray: intermediate
strains, and in black: resistant strains.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
107
Figura 2: Porcentaje de cepas no susceptibles a al menos un antibiótico de cada grupo. Figure 2: Percentage of non-susceptible strains to at least one antibiotic of each group.
Figura 3: Porcentaje de cepas no susceptibles a uno o más grupos de antibóticos. Figura 3: Percentage of non-susceptible strains to one or more antibiotic groups.
DISCUSIÓN Y CONCLUSIONES Para el análisis de la resistencia antibiótica, se consideraron “resistentes” todas aquellas cepas que según el CLSI (2012) fueron clasificadas como “intermedias” o “resistentes”. La resistencia a antibióticos ha mostrado ser baja si E. coli proviene de animales antárticos (Bonnedahl y col., 2008) y alta si tiene origen humano (Sayah y col., 2004). Debido a que las cepas de este trabajo fueron obtenidas en una bahía con alta actividad antropogénica, se sospecha que la mayoría de las cepas de E. coli provienen de humanos. Por otro lado, dado que las bases presentes en la bahía hospedan personas de distintos países, no es posible comparar la resistencia de este estudio con los perfiles de resistencia de una población o zona geográfica. La resistencia a estreptomicina (62,4%) y tetraciclina (36,5%) observada en este estudio se puede considerar elevada, incluso para cepas aisladas en plantas de tratamiento de agua, como también de animales de granja y humanos en presencia de presión selectiva (Sayah y col., 2004; Reinthaler y col., 2003). Estos resultados son consistentes con el estudio publicado por Hernández y col. (2012), quienes aislaron E. coli de muestras de agua distintas bahías asociadas a asentamientos humanos, entre ellas Bahía Fildes, y encontraron resistencia, principalmente, a ampicilina, tetraciclina, estreptomicina y trimetoprim - sulfametoxazol. A nivel mundial, la prevalencia de resistencia a ampicilina en individuos sanos es de 13% a 100% (Erb y col., 2007); mientras que los miembros de la familia Enterobacteriaceae aislados de animales antárticos, no presentan resistencia a ampicilina (Bonnedahl y col., 2008). En este estudio, se encontró que 17,4% de las cepas de E. coli es resistente a ampicilina, lo que apoya la hipótesis que estas cepas serían de origen humano. Por otra parte, se encontró susceptibilidad a cloranfenicol en 100% de las cepas y a ceftazidima, cefepima y gentamicina en más del 99%. Estos valores están dentro del rango de resistencia descrito en plantas de tratamiento de agua, animales de granja y en humanos en presencia de presión selectiva (Sayah y col., 2004; Reinthaler y col., 2003); y además, fue descrito en Antártica, para otras bacterias de la familia Enterobacteriaceae (Bonnedahl y col., 2008). El grupo de aminoglucósidos, fue el segundo grupo con mayor porcentaje de cepas no susceptibles a al menos un antibiótico del grupo (Figura 2). Esto se debe, principalmente, a la alta frecuencia de cepas resistentes a estreptomicina y, menos frecuentemente, a gentamicina. Por otro lado, la resistencia al grupo de las cefalosporinas se debe, principalmente, a la elevada frecuencia de cepas “no susceptibles” a cefalotina (49,4%). Hernández y col. (2012), pesquisaron el gen blaCTX-M (β-lactamasas de espectro extendido) en 4 cepas de E. coli aisladas en Bahía Fildes, sugiriendo que la resistencia a cefalosporinas de este estudio puede estar dada por presencia de este tipo de mecanismo de resistencia. La susceptibilidad y multiresistencia encontrada en las cepas de E. coli podría deberse a que éstas sean de origen antropogénico. Los resultados de este estudio dan cuenta de una persistencia de cepas resistentes

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
108
a antibióticos en el tiempo, dado que lo informado por Hernández y col. (2012) es en cepas aisladas en enero de 2011 y las muestras de este trabajo, en enero de 2013. Esta permanencia de la resistencia, podría estar dada por la periodicidad de descargas de aguas residuales a la bahía o bien, por la retención de la resistencia incluso sin presión selectiva en el medio. Resulta preocupante el perfil de resistencia encontrado en las cepas aisladas de Bahía Fildes, que en ciertos casos, se asemeja a los encontrados en plantas de tratamiento o sectores con alta presión selectiva. Este fenómeno plantea las interrogantes de si esta resistencia es transferible a la microbiota endémica y cuál es su impacto en el ecosistema antártico. Dado que existe muy poca información sobre resistencia antibiótica en cepas de E. coli aisladas de ambientes antárticos, es difícil establecer con certeza los factores que están contribuyendo, por lo tanto, es necesario continuar con estudios de resistencia antibiótica en cepas bacterianas en Antártica. AGRADECIMIENTOS Proyectos INACH RT_06-12 y MA-01-12, CIMAA-U. de Magallanes. REFERENCIAS BIBLIOGRÁFICAS Allen, H., Donato, J., Wang, H., Cloud-Hansen, K., Davies, J. y Handelsman, J. 2010. Call of the wild: antibiotic resistance genes in natural environments. Nature Review Microbiology, 8(4):251-259. Bonnedahl, J., Olsen, B., Waldenström, J., Broman, T., Jalava, J., Huovinen, P. y Österblad, M. 2008. Antibiotic susceptibility of faecal bacteria in Antarctic penguins. Polar Biology, 31(6):759-763. Clinical and Laboratory Standards Institute (CLSI). 2012. Performance Standards for Antimicrobial Susceptibility Testing Seventeenth Informational Supplement, M100-S17, 27. Davies, J. 2006. Are antibiotics naturally antibiotics? Journ Industrial Microbiology and Biotechnology, 33(7):496–499. Dodd, M. 2012. Potential impacts of disinfection processes on elimination and deactivation of antibiotic resistance genes during water and wastewater treatment. Journal of Environmental Monitoring, 14(1):1754-1771. Edwards, P. y Ewing. W. 1972. Differentiation of Enterobacteriaceae by Biochemical Reactions en Identification of Enterobacteriaceae, 3rd ed. Burgess Publishing Co., Minneapolis, Minn. Erb, A., Stürmer, T., Marre, R. y Brenner, H. 2007. Prevalence of antibiotic resistance in Escherichia coli: overview of geographical, temporal, and methodological variations. Eur Jour Clin Microb & Infect Dis, 26(2):83-90. Hernández, J., Stedt, J., Bonnedahl, J., Molin, Y., Drobni, M., Calisto-Ulloa, N., Gomez-Fuentes, C., Astorga, S., González-Acuña, D., Waldenström, J., Blomqvist, M. y Olsen, B. (2012). Human-Associated Extended-Spectrum beta-lactamase in the Antarctic. Applied and Environmental Microbiology, 78(6):2056-2058. Levine, M. 1918. Differentiation of B. coli and B. aerogenes on a Simplified Eosin-Methylene Blue Agar. The J of Infect Dis, 23(1):43-47. Martínez, J. 2012. Natural antibiotic resistance and contamination by antibiotic resistance determinants: the two ages in the evolution of resistance to antimicrobials. Front in Microb: Antimicrobials, Resistance and Chemotherapy, 3(1):1-3. Miller, R., Gammon, K. y Day, M. 2009. Antibiotic resistance among bacteria isolated from seawater and penguin fecal samples collected near Palmer Station, Antarctica. Canadian Journal of Microbiology, 55(3):37-45. Reinthaler, F., Posch, J., Feierl, G., Wüst, G.; Haas, D y Ruckenbauer, G., Mascher, F. y Marth, E. 2003. Antibiotic resistance of E. coli in sewage and sludge. Water Research, 37(8):1685-1690. Sayah, R., Kaneene, J., Johnson, Y. y Miller, R. 2005. Patterns of Antimicrobial Resistance Observed in Escherichia coli Isolates Obtained from Domestic and Wild-Animal Fecal Samples, Human Septage, and Surface Water. Applied and Environmental Microbiology, 71(9):1394-140. Skurnik, D., Ruimy, R., Andremont, A., Amorin, C., Rouquet, R., Ricard, B. y Denamur, E. 2006. Effect of human vicinity on antimicrobial resistance and integrons in animal faecal Escherichia coli. Journal of Antimicrobial Chemotheraphy, 57(6):1215-1219. Stevens, M., Ashbolt, N. y Cunliffe, D. 2003. Recommendations to change the use of coliforms as microbial indicators of drinking water quality. http://www.nhmrc.gov.au/_files_nhmrc/publications/attachments/eh32.pdf. Tadesse, D., Zhao, S., Tong, E., Ayers, S., Singh, A. y Bartholomew, M. 2012. Antimicrobial drug resistance in Escherichia coli from humans and food animals, United States, 1950–2002. Emerging Infectious Disease, 18(5):741-749.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
109
BIOSÍNTESIS DE NANOPARTÍCULAS FLUORESCENTES MEDIANTE BACTERIAS PSICROTOLERANTES DEL GÉNERO PSYCHROBACTER SPP.: VENTAJAS
COMPARATIVAS DEL CONTINENTE ANTÁRTICO (Biosynthesis of fluorescent nanoparticles using psychrotolerant bacteria genus Psychrobacter spp.:
Comparative advantages of Antarctic continent)
Plaza, D.O.1, Straub, Y.D.1, Gallardo, C.1, Bruna N.1, Monrás J.P.1 , Bravo D.2 y Pérez-Donoso JM1
1Laboratorio de BioNanotecnología y Microbiología, Centro de Bioinformática y Biología Integrativa (CBIB), Departamento de Ciencias Biológicas, Universidad Andrés Bello. Av. Republica # 239, Santiago, Chile. E-mail: [email protected] 2Laboratorio de Microbiología Oral, Facultad de Odontología, Universidad de Chile. Sergio Livingstone # 943, Santiago, Chile.
INTRODUCCIÓN Las nanopartículas (NPs) son estructuras metálicas (1-100 nanómetros) que poseen diferentes formas (esféricas, triangulares, barras, etc.) y propiedades únicas (químicas, físicas, ópticas, etc.) en comparación a materiales de mayor escala (Sau y Rogach, 2010). En las últimas décadas, las NPs fluorescentes llamadas “quantum dots” (QDs), han captado interés como marcadores biológicos (Chen y col., 2009; Idris y col., 2009), dispositivos opto-electrónicos (Gaponik y col., 1999) y celdas solares (Huynh y col., 2002), entre muchos otros. Los QDs están compuestos por átomos de Cd, Se, Te, In, As, entre otros, y podrían considerarse como fluoróforos ideales en comparación con sus pares orgánicos, ya que son más brillantes, más fotoestables y presentan múltiples colores que pueden ser excitados simultáneamente utilizando una única fuente de luz (Riegler y Nann, 2004; Liu y col., 2005; Jamieson y col., 2007; Resch-Genger y col., 2010; Wang y Zhu, 2011). La síntesis de QDs se realiza tradicionalmente mediante métodos químicos y físicos, generalmente nocivos para el medio ambiente debido al uso de químicos peligrosos, elevadas temperaturas y condiciones anaerobias. Además, estos métodos a menudo están limitados a la producción de pequeñas cantidades de NPs y en algunos casos se producen mezclas de ellas con pobre morfología (Mohanpuria y col., 2008; Rai y col., 2008; Sharma y col., 2009; Birla y col., 2009; Sau y Rogach, 2010). Actualmente, se trabaja en el desarrollo de procedimientos de síntesis de QDs de mayor rendimiento, bajo costo y sin químicos tóxicos, que permitan, además, obtener estructuras con tamaños y formas bien definidas (Maliszewska, 2011). El uso de sistemas biológicos para la producción de QDs con tamaños y formas controladas cuenta con ventajas como: la biomasa utilizada es más simple de manipular y puede ser liberada al medio ambiente, la síntesis puede ser llevada a cabo a temperatura y presión ambiental, pH neutro, sin uso de químicos tóxicos, menor costo y las NPs producidas presentan mayor biocompatibilidad (Kaushik y col., 2009; Rai y col., 2011; Suresh, 2012). En base a esto, existe un creciente interés en desarrollar procesos de síntesis más simples y amigables con el entorno, como aquellos que involucran sistemas biológicos, particularmente microorganismos. Nuestro grupo ha desarrollado un método de biosíntesis de QDs de CdTe mediante E. coli, donde se determinó que la bioproducción es afectada por la temperatura, el estado redox y las defensas antioxidantes intracelulares (Monrás y col., 2012). En relación a estos hechos, se ha reportado que la estabilidad de las especies reactivas del oxígeno (ROS) se incrementan a bajas temperaturas, probablemente debido a que los gases y radicales son más solubles y estables a bajas temperaturas (Piette y col., 2011). Por lo tanto, las defensas antioxidantes reforzadas son componentes evolutivos comunes en la adaptación a ambientes fríos y ricos en oxígeno (Piette y col., 2011; di Prisco y col., 2012). En este sentido, las regiones polares, conocidas por sus condiciones ambientales extremas, se caracterizan por bajas temperaturas persistentes con ciclos de congelación-descongelación, condiciones

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
110
extremas de luz (incluyendo una alta radiación UV) y una alta variabilidad en la disponibilidad de nutrientes y salinidad (Vincent, 2004). Específicamente, el continente Antártico ofrece un completo set de condiciones climáticas extremas, por lo cual constituye uno de los ambientes más duros del planeta. Diversos factores tales como temperaturas extremas, baja humedad atmosférica, baja disponibilidad de agua líquida, estrés osmótico y alta incidencia periódica de radiación, con largos períodos de completa oscuridad, contribuyen a que el continente sea inhóspito para el desarrollo de comunidades biológicas (Peck 2002a; Cowan y Tow, 2004; De Lorenzo, 2010). Esto sumado al aislamiento ambiental y geográfico del continente Antártico y la fuerte selección natural producida por las condiciones extremas, han generado una biota endémica altamente adaptada (Peck y col., 2005; Rogers, 2007). Por lo tanto, el aislamiento de bacterias especialmente adaptadas a ambientes extremos, utilizando mecanismos de defensa antioxidante especialmente adaptados, puede significar avanzar hacia el desarrollo de un área nueva de investigación para la bioproducción de QDs a partir de sales metálicas oxidantes como Cd+2 y TeO3
-2.
MATERIALES Y MÉTODOS A partir de muestras recolectadas de la Antártica, se aisló bacterias cultivables utilizando 8 diferentes medios de cultivo (LB, LB/10, LB 4% sal, Agar Marino, ATTC, PYT100, YM y R2A), a tres temperaturas de crecimiento (15, 20 y 37°C). Para potenciar la factibilidad de realizar biosíntesis, se aisló bacterias resistentes a Cd+2 y TeO3
-2 a partir de las 217 cepas en medio LB a 15 °C. La identificación de las bacterias resistentes se realizó mediante amplificación del 16S rRNA. Los niveles de resistencia a metales y a antibióticos fueron determinados mediante Concentración Mínima Inhibitoria (MIC) y el test de Kirby–Bauer con un set de 10 antibióticos comerciales (Ampicilina, Colistina, Fosfomicina, Gentamicina, Kanamicina, Lincomicina, Ácido Oxolínico, Polimixina, Rifampicina y Sulfonamida), respectivamente. La evaluación de la biosíntesis de QDs se llevó a cabo utilizando el protocolo desarrollado por Monrás y col. (2012). El efecto de la temperatura (10, 15 y 28 °C) sobre la biosíntesis, se evaluó exponiendo los pellets a luz UV (365 nm) cada 12 horas. RESULTADOS Y DISCUSIÓN Se logró aislar un total de 410 aislados bacterianos, de los cuales 235 fueron capaces de crecer a 15°C en diferentes medios de cultivo. A partir de estos, se creció un total de 217 en medio LB (medio de cultivo seleccionado). Se encontró 3 cepas psicrotolerantes pertenecientes al género Psychrobacter spp. (re-nombradas arbitrariamente: Psy1, Psy2 y Psy3) altamente resistentes a Cd+2 y TeO3
-2. Los valores obtenidos de MIC a las sales de metales fueron de 1.000 µg/mL para CdCl2, y para K2TeO3 entre 125 - 1000 µg/mL. Las cepas fueron resistentes a Fosfomicina, Lincomicina y Sulfonamida (excepto Psy 1). Este género comprende 34 especies reportadas (Azevedo y col., 2012), están formadas por bacilos Gram-negativo, no mótiles, aeróbicos y usualmente no pigmentados. Se caracterizan por su gran capacidad de adaptación al frío y a un amplio rango de concentración salina (Bowman, 2006). Se encuentran mayoritariamente en ambientes fríos, principalmente en la Antártica (Bowman y col., 1996; Shivaji y col., 2004; Shivaji y col., 2005). La alta resistencia a metales no ha sido reportada hasta el momento. Sin embargo, existe registro de algunas especies con alta actividad catalasa (Yumoto y col., 2010; Kimoto y col., 2012), así como plasmidios encontrados en especies aisladas del Ártico (Dziewit y cols., 2013) y de la Antártica (Tutino y col., 2000), algunos relacionados incluso con resistencia a antibióticos (Petrova y col., 2009). Estos antecedentes podrían apuntar hacia la adaptación a ambientes extremos por parte de este género. Los resultados de la biosíntesis muestran cómo los pellets bacterianos van cambiando de color en función del tiempo, pasando desde un color cercano al verde avanzando hacia el color rojo. Esto podría confirmar la presencia de QDs al interior de las células, debido al cambio del color de la fluorescencia

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
111
dependiente del tiempo (el color depende del tamaño de la nanopartícula), característica de este tipo de nanocristales, la cual los diferencia de los fluoróforos orgánicos tradicionales. Otra observación interesante, es que en algunas cepas se produce bastante reducción del telurito a teluro0, sugiriendo la presencia de reductasas capaces de funcionar a bajas temperaturas, esto podría favorecer la formación de QDs de CdS en desmedro de CdTe. Las diferentes cepas presentaron diferentes colores en las mismas condiciones de biosíntesis.
CONCLUSIONES Estos resultados dan indicios de la primera biosíntesis de QDs realizada por bacterias antárticas, a una temperatura inferior a 37˚C (única temperatura reportada a la fecha), con lo cual podría ser posible biosintetizar estas partículas a temperatura ambiente sin necesidad de equipamiento adicional. Estos resultados podrían a su vez, significar la obtención de nanopartículas con novedosas características y propiedades no descritas en la literatura hasta el momento. Sorprendentemente, los niveles de resistencia a las sales utilizadas en la biosíntesis fueron altos. Hasta la fecha no existen reportes en la literatura acerca de la resistencia a metales del género Psychrobacter, por lo cual también es la primera vez que se encuentran especies de este género con elevados valores de resistencia a los metales utilizados en la biosíntesis de QDs basados en Cadmio.
AGRADECIMIENTOS Programa Nacional Beca de estudios Magíster (CONICYT). Trabajo financiado por proyectos nacionales INACh (T-19_11) y FONDECYT (11110077). REFERENCIAS BIBLIOGRÁFICAS Azevedo, J. S. N. y Henriques, I. 2013. Molecular analysis of the diversity of genus Psychrobacter present within a temperate estuary. FEMS Microbiology Ecology. 84: 451–460. Birla, S. S., Tiwari, V. V., Gade, A. K., Ingle, A. P., Yadav, A. P., y Rai, M. K. 2009. Fabrication of silver nanoparticles by Phoma glomerata and its combined effect against Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus. Letter Applied Microbiology. 48: 173–179. Bowman, J. P., Cavanagh, J., Austin, J. J. y Sanderson, K. 1996. Novel Psychrobacter Species from Antarctic Ornithogenic Soils. International Journal of Systematic Bacteriology (26): 841–848. Bowman, J. P. 2006. The genus Psychrobacter. In The Prokaryotes (pp. 920-930). Springer New York. Cowan, D. A., y Tow, L. A. 2004. Endangered Antarctic environments. Annual Review Microbiology 58: 649–690. Chen, C. 2009. Quantum dots-based immunofluorescence technology for the quantitative determination of HER2 expression in breast cancer. Biomaterials; 30: 2912–2918. Di Prisco, G., Giordano, D., Russo, R. y Verde, C. 2012. The Challenges of Low Temperature in the Evolution of Bacteria, En G. di Prisco y C. Verde (eds.) Adaptation and Evolution in Marine Environments, Volume 1, From Pole to Pole, DOI: 10.1007/978-3-642-27352-0_10, Springer-Verlag Berlin Heidelberg Dziewit, L., Cegielski, A., Romaniuk, K., Uhrynowski, W., Szych, A., Niesiobedzki, P. y Zmuda-Baranowska, M. J. 2013. Plasmid diversity in arctic strains of Psychrobacter spp. Extremophiles : life under extreme conditions, 17(3): 433–44. Idris, N. M., Li, Z. Q., Ye, L., Sim, E. K. W., Mahendran, R. y Ho, P. C. L. 2009. Tracking transplanted cells in live Huynh, W. U., Dittmer, J.J., Alivisatos, A.P. 2002. Hybrid nanorod-polymer solar cells. Science; 295: 2425–7. Gaponik, N. P., Talapin, D. V., Rogach, A. L. 1999. A light-emitting device based on a CdTe nanocrystal/polyaniline composite. Physical Chemistry Chemical Physics.1:1787–9. Liu, T., Liu, B., Zhang, H. y Wang Y. 2005. The fluorescence bioassay platforms on quantum dots nanoparticles. Journal Fluorescence. 15: 729–733. Jamieson, T., Bakhshi, R., Petrova, D., Pocock, R., Imani, M. y Seifalian, A. M. 2007. Biological applications of quantum dots. Biomaterials. 28 (31):4717-32. Kaushik, N., Thakkar, M. S., Snehit, S., Mhatre, M. S., Rasesh, Y. y Parikh, M.S. 2010. Biological synthesis of metallic nanoparticles Nanomedicine: Nanotechnology, Biology, and Medicine. 6: 257 – 262. Kimoto, H., Yoshimune, K., Matsuyma, H. y Yumoto, I. 2012. Characterization of Catalase from Psychrotolerant Psychrobacter piscatorii T-3 Exhibiting High Catalase Activity. International Journal of Molecular Sciences, 13(2): 1733–1746.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
112
Maliszewska, I. 2011. Microbial Synthesis of Metal Nanoparticles En: Metal Nanoparticles in Microbiology. Springer-Verlag Berlin Heidelberg, Germany, pp 1-16. Mohanpuria, P. y Rana, Æ. N. K. 2008. Biosynthesis of nanoparticles : technological concepts and future applications. Journal of Nanoparticles Research 10, 507-517. Monrás, J. P., Díaz, V., Bravo, D., Montes, R. A., Chasteen, T. G., Osorio-Román, I. O. y Vásquez, C. C. 2012. Enhanced glutathione content allows the in vivo synthesis of fluorescent CdTe nanoparticles by Escherichia coli. PloS One, 7(11). Peck, L. S. 2002ª. Coping with change: stenothermy, physiological flexibility and environmental change in Antarctic seas. Proceedings of the 14th International Congress on Comparative Physiology. Peck, L. S., Clark, M, S., Clarke, A, Cockell, C. S., Convey, P., Detrich, H. W., Fraser, K. P., Johnston, I. A., Methe, B. A., Murray, A. E., Römisch, K. y Rogers, A. D. 2005. Genomics: applications to Antarctic ecosystems. Polar Biology 28: 351–365. Petrova, M., Gorlenko, Z. y Mindlin, S. 2009. Molecular structure and translocation of a multiple antibiotic resistance region of a Psychrobacter psychrophilus permafrost strain. FEMS Microbiology Letters, 296(2): 190–197. Piette, F., D’Amico, S., Mazzuchelli, G., Danchin A., Leprince P. y Feller G. 2011. Life in the cold: a proteomic study of cold-repressed proteins in the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125. Applied Environmental Microbiology, 77:3881–3883. Rai, M. K., Yadav, A. P. y Gade, A. K. 2011. Biogenic nanoparticles: An introduction to what they, how they are synthesized and their applications. In Metal Nanoparticles in Microbiology. Springer-Verlag Berlin Heidelberg, Germany, pp 1-16. Resch-Genger, U., Grabolle, M., Nitschke, R. y Nann, T. 2010. Nanocrystals and nanoparticles vs. molecular fluorescent labels as reporters for bioanalysis and the life sciences. A critical comparison. En: Advanced Fluorescence Reporters in Chemistry and Biology II. Springer Ser Fluoresc 9:40-41. Riegler, J. y Nann, T. 2004. Application of luminescent nanocrystals as labels for biological molecules. Anal Bioanalytical Chemistry. 379:913-919. Sharma, H. C., Crouch, J. H., Sharma, K. K., Seetharama, N. y Hash, C. T. 2002. Applications of biotechnology for crop improvement: prospects and constraints. Plant Science 163: 381-395. Shivaji, S., Reddy, G. S. N., Raghavan, P. U. M., Sarita, N. B. y Delille, D. 2004. Isolated from Fast Ice from Adelie Land , Antarctica, 635, 628–635. Shivaji, S., Reddy, G. S. N., Suresh, K., Gupta, P., Chintalapati, S., Schumann, P., Stackebrandt, E., 2005. Psychrobacter vallis sp. nov. and Psychrobacter aquaticus sp. nov., from Antarctica. International Journal of Systematic and Evolutionary Microbiology, 55, 757–762 Sau, T. K. y Rogach, A.L. 2010. Nonspherical noble metal nanoparticles: colloid-chemical synthesis and morphology control. Advanced Materials, 22(16):1781–1804. Tutino, M. L., Duilio, A., Moretti, M. A, Sannia, G. y Marino, G. 2000. A rolling-circle plasmid from Psychrobacter sp. TA144: evidence for a novel rep subfamily. Biochemical and Biophysical Research Communications, 274(2): 488–95. Vincent, W. F., Mueller, D. R y Bonilla, S. 2004. Ecosystems on ice: the microbial ecology of Markham Ice Shelf in the High Arctic. Cryobiology 48: 103-112. Wang, W., Zhu, J. J. 2011. Optical applications of quantum dots in biological system. Science China-Chemistry, 54: 1177-1184. Yumoto, I., Hirota, K., Kimoto, H., Nodasaka, Y., Matsuyama, H. y Yoshimune, K. 2010. Psychrobacter piscatorii sp. nov., a psychrotolerant bacterium exhibiting high catalase activity isolated from an oxidative environment. International Journal of Systematic and Evolutionary Microbiology, 60, 205–208.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
113
FABRICACIÓN DE CELDAS SOLARES DEL TIPO DYE-SENSITIZED SOLAR CELL (DSSC) MEDIANTE LA INCORPORACIÓN DE PIGMENTOS PROVENIENTES DE BACTERIAS
ANTÁRTICAS (Solar cell manufacturing type dye-sensitized solar cell (DSSC) by the incorporation of pigments
obtained from Antarctic bacteria)
Órdenes-Aenishanslins, N.1,2, Gran-Scheuch, A.1,2, Saona, L.A.1,2, Monrás, J.P.1,3 y Pérez-Donoso, J.M.1*
1Bionanotechnology and Microbiology Lab, Center for Bioinformatics and Integrative Biology (CBIB), Universidad Andrés Bello. República 239, Santiago, Chile. [email protected] 2Facultad de Ciencias Químicas y Farmacéuticas, Universidad de Chile. 3Facultad de Química y Biología, Universidad de Santiago de Chile.
INTRODUCCIÓN En el último tiempo, ha surgido con fuerza la necesidad de reemplazar el uso de combustibles fósiles para la generación de energía. Esto ha impulsado fuertemente la investigación y el desarrollo de las energías renovables no convencionales (ERNC), que surgen como alternativas sustentables para satisfacer la alta demanda energética mundial. En este sentido, nuestra investigación apunta a utilizar el recurso energético más abundante en la tierra: la energía solar, mediante el desarrollo de celdas fotovoltaicas empleando nuevos materiales. La forma más importante de aprovechar la energía proveniente del sol es transformarla directamente en energía eléctrica mediante sistemas fotovoltaicos. La tecnología fotovoltaica ofrece la ventaja de ser modular y escalable: desde pequeños sistemas para uso domiciliario hasta grandes instalaciones para fines industriales. Sin embargo, dentro de sus limitaciones se encuentra su elevadísimo costo de producción, difícil manufactura y gran cantidad de materiales requeridos, lo que ha repercutido en que su uso no se haya masificado aún. Paradójicamente, pese a que no hay emisión de contaminantes en la producción de energía eléctrica a partir de la energía solar, para producir la tecnología fotovoltaica se consume mucha energía, se emplean materiales de elevado costo y se emiten gran cantidad de compuestos tóxicos. La investigación mundial no ha estado ajena a esta problemática, lo que ha impulsado nuevas formas de recolectar la energía lumínica proveniente del sol, basadas en nuevos materiales y maneras de hacerlos interactuar, y si bien su eficiencia no supera a las celdas de silicio, hoy en día se está apuntando a abaratar los costos por sobre aumentar la eficiencia. En nuestro laboratorio trabajamos con un modelo de celda solar de bajo costo y alta eficiencia basada en moléculas sensibilizadoras capaces de cosechar fotones, unidas a semiconductores de band gap ancho (usualmente films de TiO2 coloidal) y un electrolito soporte. Este modelo de celda solar, conocida como Dye-Sensitized Solar Cell (DSSC) y desarrollada en el año 1991 por el químico suizo Michael Graetzel (Graetzel et al. 1991), abrió un nuevo campo de investigación y aplicación, que ha permitido el desarrollo y mejoramiento de nuevas celdas fotovoltaicas. De hecho, su modelo se ha empleado en la construcción de grandes paneles solares y significado la invención de una gran cantidad de patentes. A la fecha, se han reportado una gran cantidad de sensibilizadores para este tipo de celdas, como tintes orgánicos (Ito et al. 2008), compuestos de rutenio (Chen et al. 2009) y recientemente nanopartículas fluorescentes o Quantum Dots (Rühle et al. 2010). Sin embargo, nuestra investigación apunta a un nicho poco explorado, que es la utilización de microorganismos (principalmente antárticos) para la obtención, mediante biosíntesis, de materias primas necesarias para la fabricación sustentable de celdas fotovoltaicas. En este trabajo, mostramos algunos resultados donde pigmentos provenientes de bacterias

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
114
antárticas son utilizados como sensibilizadores en celdas del tipo DSSC, una alternativa prometedora y de bajo costo a las actuales tecnologías fotovoltaicas. Nuestro interés apunta, más que a aumentar la eficiencia en estas celdas, a desarrollar una tecnología cuyo proceso de fabricación sea mucho más verde o eco-friendly. MATERIALES Y MÉTODOS. Muestreo y aislamiento de microorganismos. Durante la expedición Antártica del año 2012 se obtuvo muestras de suelo proveniente de las cercanías de la base Presidente Eduardo Frei Montalva, en la Península Fildes de la Isla Rey Jorge en las Islas Shetland del Sur. A las muestras de suelo se les agregó agua destilada estéril y el sobrenadante fue diluido y plaqueado en medio LB agar y R2A agar. De las cepas aisladas, cinco fueron seleccionados pues evidenciaban alto grado de coloración. Extracción y purificación de pigmentos. Las cepas seleccionadas se crecieron en 50 mL de medio R2A durante 36 horas a 22 °C. Transcurrido este tiempo, el cultivo se centrifugó a 3260 x g durante 10 min y se desechó el sobrenadante. Luego, el pellet de células se resuspendió en 10 mL de agua destilada seguido por centrifugación a 3260 x g durante 10 min. Posteriormente, se desechó nuevamente el sobrenadante, esto para eliminar cualquier resto de medio de cultivo. Finalmente las células son tratadas con 1,5 mL de solución acetona:metanol (7:2, v/v) y sometidas a ultra sonido. La mezcla de solventes con el pigmento se extrae por ultracentrifugación del extracto celular a 21380 x g durante 10 min. De los pigmentos extraídos se seleccionó dos debido a su marcada coloración y elevada fotoestabilidad. Estos pigmentos fueron caracterizados mediante espectrofotometría de absorción. Fabricación de la DSSC. Para la fabricación de los electrodos de la celda se utilizó vidrios FTO (Fluorine-doped Tin Oxide) TEC15 de resistividad superficial 13 [Ω/sq], 85 % de transmitancia y tamaño 25 x 25 x 2 mm. Para el ánodo, a partir de una suspensión en ácido acético de nanopartículas de TiO2 se deposita un film de esta suspensión sobre el vidrio, mediante la técnica de spin-coating, a 2000 rpm por 20 segundos. Este electrodo de 225 cm2 se sometió a 450 °C por 30 minutos, para el sintering del material. Por otra parte, el cátodo o contra electrodo de platino se prepara a partir de una solución de H2PtCl6·6H2O en isopropanol. Se depositan 30 µL de la solución sobre un vidrio FTO mediante spin-coating y se calienta a 450 °C por 20 minutos. La sensibilización del film de TiO2 se realiza mediante la inmersión del ánodo en la solución que contiene el pigmento. El vidrio se mantiene sumergido en esta solución por 2 horas a temperatura ambiente y en oscuridad. Transcurrido ese tiempo, el electrodo se lava con etanol retirando el pigmento en exceso que no se unió al TiO2. Luego, ambos electrodos se empalman, y se añade la solución de electrolito. Se utiliza electrolito yoduro/triyoduro (I3
-/I-) compuesto por KI y I2 en etilenglicol. Finalmente la celda se sella quedando lista para las mediciones. La caracterización de la celda solar se realiza en condiciones de irradiancia y temperatura constante, utilizando una fuente de luz artificial (100 W de potencia) y una fuente de luz natural. RESULTADOS Espectros de absorbancia de pigmentos. La Fig.1 muestra los espectros de absorbancia de los pigmentos seleccionados.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
115
Figura 1: Espectros de absorbancia para dos pigmentos extraídos de bacterias antárticas.
Figure 1: Absorbance spectra for two pigments extracted from Antarctic bacteria. Se observa que ambos pigmentos tienen un amplio rango de absorción de la luz visible, lo que los hace buenos candidatos para su uso en celdas solares. Además, ambos pigmentos presentan distintos máximos de absorción, lo que es atribuible a que presentan estructuras químicas distintas. Mediciones de corriente-voltaje. A partir de los pigmentos seleccionados se construyen dos celdas cuyo funcionamiento general se resume en la Fig.2.
Figura 2: Esquema general de la celda solar sensibilizada con pigmentos.
Figure 2: Scheme of the pigments-sensitized solar cell.
El pigmento es capaz de absorber la luz incidente y producir electrones excitados que son transferidos al film de nanopartículas de TiO2 capaz de “recolectar” los electrones y transferirlos a un vidrio con la capacidad de conducir corriente eléctrica, a la vez que permite el paso de la radiación solar a través del sistema (vidrio FTO, por sus siglas en inglés de Fluorine-doped Tin Oxide). Esto constituye el fotoánodo de la celda. Del otro lado, encontramos el cátodo o contraelectrodo de platino. Todo este sistema se encuentra embebido en un electrolito redox, cuya recirculación en su estado oxidado-reducido permite restablecer los electrones perdidos por el sensibilizador, a la vez que sirve como una vía interna

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
116
para el paso de electrones desde el cátodo hacia el ánodo, completando así el circuito. Finalmente, la existencia de esta diferencia de potencial permite que, una vez conectados ambos electrodos, haya circulación de electrones y generación de corriente eléctrica. Las mediciones se realizan mediante un circuito estándar para determinar el voltaje de circuito abierto (VOC, máximo voltaje producido por la celda cuando la corriente es cero) y la corriente de corto circuito (ISC, máximo flujo de corriente que circula por la celda cuando el voltaje tiende a cero). Los resultados se resumen en la Tabla 1.
Tabla 1: Resultados de Corriente-Voltaje para las dos celdas diseñadas.
Table 1: Current-Voltage results for the two solar cells.
Con la celda sensibilizada con el Pigmento 1 se obtuvieron las mayores diferencias de potencial utilizando ambas fuentes de luz, sin embargo, en el caso de las corrientes este resultado se revierte; pues la celda con el Pigmento 2, pese a registrar menores voltajes, mantiene un flujo de corriente mucho mayor. Posiblemente esta diferencia se deba las características químicas y estructurales del pigmento o su grado de afinidad por el TiO2 que facilita en un caso la transferencia de electrones hacia el electrodo. DISCUSIÓN Y CONCLUSIONES Los resultados evidencian que los pigmentos extraídos de las bacterias antárticas efectivamente cosechan fotones y transfieren electrones al electrodo, en un proceso dependiente de la luz incidente. Las mediciones indican que los pigmentos tienen un buen desempeño en este tipo de celdas. En la actualidad, estamos trabajando en controlar algunas variables como la temperatura o condiciones del medio de cultivo que favorezcan la producción de estos pigmentos. Del mismo modo, estamos evaluando en algunas celdas el resultado que tendría mezclas de ellos, para así abarcar aún más el espectro solar. Por otro lado, como proyección del trabajo está la identificación de la estructura química de los pigmentos y modificar de algún modo su estabilidad o alguna propiedad óptica que pueda favorecer su uso en este tipo de celdas solares. AGRADECIMIENTOS Este trabajo es financiado por INACH T-19_11, FONDECYT 11110077 y ANILLO ACT 1107. REFERENCIAS Chen, C.Y., Wang, M., Li J-Ying, Pootrakulchote, N., Alibabaei, L., Ngoc-le C, et al. 2009. Highly Efficient Light-Harvesting Ruthenium Sensitizer for Thin-Film Dye-Sensitized Solar Cells. ACS Nano. 3 (10): 3103-3109. Graetzel, M. y O' Regan, B. 1991. A low-cost, high-efficiency solar cell based on dye-sensitized colloidal titanium dioxide films. Nature. 353 (6346): 737-740. Ito, S., Miura, H., Uchida, S., Takata, M., Sumioka, K., Liska, P., et al. 2008. High-conversion-efficiency organic dye-sensitized solar cells with a novel indoline dye. Chem. Commun. 44 (41): 5194-5196. Rühle, S., Shalom, M. y Zaban, A. 2010.Quantum Dot-Sensitized Solar Cells. Chemphyschem. 11 (11): 2290-2304.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
117
AISLAMIENTO Y CARACTERIZACIÓN DE BACTERIAS RESISTENTES A MERCURIO Y TELURITO PROVENIENTES DEL TERRITORIO ANTÁRTICO CHILENO
(Isolation and characterization of mercury- and tellurite-resistant bacteria from the Chilean Antarctic Territory)
Díaz-Vásquez W.A.1, 2, 3, Muñoz P.R.1, Rodríguez F.J.1 y C.C. Vásquez1
1 Laboratorio de Microbiología Molecular, Universidad de Santiago de Chile. [email protected] 2 Laboratorio de Microbiología Molecular y Biotecnología Ambiental, Universidad Técnica Federico Santa María. 3 Facultad de Ciencias de la Salud, Universidad San Sebastián. INTRODUCCIÓN La contaminación por metales pesados es una preocupación medioambiental importante. En Chile existen 74 minas de cobre y 28 de oro y plata que pueden generar desechos de derivados de teluro (11) y mercurio. En relación con este último, se ha visto que al calentar las amalgamas Hg/Au, se liberan vapores tóxicos de Hg que se depositan en el suelo. Además, se ha detectado altos niveles de mercurio en minas de Andacollo, región de Coquimbo y también en la Patagonia (13), provocando la contaminación de aguas y suelos. El mercurio presenta una elevada toxicidad y puede provocar enfermedades neurológicas (10) envenenamiento, e incluso la muerte. Este tóxico puede inactivar proteínas por interacción con tioles (SH), tioéteres e imidazol (17) y los compuestos órgano-mercuriales pueden bloquear la síntesis de proteínas y la incorporación de timidina al DNA (22). Emergencias medioambientales han sido descritas en Chile (8) y otros países. Bacterias resistentes a mercurio poseen generalmente el operón merRTPGCABDE, que codifica un sistema de reducción enzimática de Hg+2 a Hg0, que es posteriormente volatilizado (14). En este contexto destaca la reductasa mercúrica dependiente de NADPH (MerA) (12) que rompe enlaces mercurio-carbono otorgando resistencia a compuestos organomercuriales (15). Estos genes pueden encontrarse en plasmidios (19) y también en transposones (16). El teluro es obtenido como un subproducto de la refinación del Cu, Ni, Ag y Au, donde grandes cantidades de teluro y sus derivados son liberados al medio ambiente cada año. La evidencia de esta creciente contaminación ambiental se refleja en la gran cantidad de bacterias resistentes al telurito (TeO3
-2) clínicas (21) y ambientales como el Geobacillus stearothermophilus V (20), entre otras. El teluro se encuentra en la naturaleza en muy bajas concentraciones (~0,027 ppm) y no presenta una elevada toxicidad. Sin embargo, el telurito y el telurato son extremadamente tóxicos para bacterias Gram negativo (21). Parte de la toxicidad producida por telurito se relaciona con sus características como oxidante provocando la oxidación de tioles celulares (23), el desmantelamiento de centros de hierro-azufre [Fe-S] en enzimas (5) y el reemplazo erróneo de Te por S en proteínas (21), entre otros. En Escherichia coli se ha demostrado que el telurito produce estrés oxidativo incrementando los niveles de superóxido intracelulares (O2
.-) (18), especie reactiva de oxígeno (ROS) que también fue determinada in vitro cuando enzimas reducen telurito a teluro (Te0) en presencia de oxígeno y NAD(P)H (4). La resistencia a telurito ha sido asociada a la presencia de plasmidios (Ter), destacando los IncHI, IncHII y IncP (3, 24) entre otros. Diversas mecanismos han sido relacionados con la resistencia a telurito como la formación de intermediarios volátiles (9) y actividad telurito reductasa que exhiben enzimas como nitrato reductasas (1), catalasa (4), dihidrolipoamida deshidrogenasa (6), entre otras. Este trabajo muestra el proceso de aislamiento y selección de organismos psicrotolerantes del territorio antártico chileno que presentan resistencia a mercurio y telurito para ser utilizadas en futuros procesos de biorremediación a bajas temperaturas. Se ha demostrado que la resistencia a metales pesados y en especial a mercurio estaría asociada a la resistencia a una amplia gama de antibióticos lo cual se cual se correlacionaría con el

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
118
resultado de la cepa ATH-43 (2, 7), aunque no es el caso para la cepa ATH-62. Sin embargo, es necesario repetir este experimento en medio Mueller Hinton, el más utilizado en antibiogramas. MATERIALES Y MÉTODOS Muestras y aislamiento Las cepas fueron aisladas de muestras antárticas de tierra indicadas en la Tabla 1 (expedición ECA-48). La selección se realizó en placas con medio LB agar suplementado con HgCl2 40 μM y con K2TeO3 40 μM a 25 °C por 24 h. De 19 muestras analizadas, dos se utilizaron para los siguientes experimentos: la muestra 43 y la 62, las que serán referidas en adelante como ATH-43 y ATH-62, respectivamente. A los aislados se les realizó tinción Gram. Concentración mínima inhibitoria (MIC) de TeO3
-2 y HgCl2
Se creció pre-inóculos de 1 ml en medio LB con una agitación promedio de 150 rpm y 25 °C hasta saturación, ~24 horas después. El MIC fue determinado, por duplicado, en placas de 96 pocillos que contenían 200 μl de LB; en placas de 48 pocillos con un volumen de 1 ml y por último, en tubos de vidrio con un volumen de 3 ml. Cada inoculación se realizó aplicando un volumen 1:100 del pre-inóculo a la placa o tubo en que se realizó el ensayo. Las concentraciones de TeO3
-2 y HgCl2 utilizadas se ajustaron a la resistencia observada para cada aislado bacteriano. Curvas de crecimiento Las curvas de crecimiento fueron realizadas en el lector multimodal Tecan Infinite® m200 PRO a 25 °C y 150 rpm por 15 h. El crecimiento fue seguido como el cambio en la densidad óptica a 600nm cada 30 min. Los resultados corresponden al promedio de la medición obtenida en 9 puntos diferentes de cada pocillo, cuya desviación estándar no superó el valor 0,09 (no mostrado). Placas de 48 pocillos que contenían LB, LB + TeO3
-2 o LB + HgCl2 en un volumen de 1 ml fueron inoculadas con los aislados bacterianos. Las concentraciones utilizadas de cada tóxico para cada cepa corresponden a una concentración inferior al MIC. Se utilizó cloruro de mercurio 40 μM y telurito de potasio 20 μM. Antibiograma Las cepas ATH-43 y ATH-62 fueron crecidas en medio LB hasta fase exponencial (OD600 0,3-0,4) y sembradas en placas con medio LB agar. Posteriormente se agregó un multidisco de secreciones Gram negativo (Valtek). Todas las placas fueron crecidas a 25 °C por 14 h. El radio de los halos fue medido y las áreas calculadas para determinar si las cepas eran resistentes o susceptibles a los antibióticos mencionados. RESULTADOS Aislamiento, selección y MICs Con el método de selección utilizado se aisló dos cepas resistentes a telurito y a mercurio: ATH-43 y ATH-62. Ambas presentan una temperatura óptima de crecimiento de 25 °C y concentraciones mínimas inhibitorias (MIC) de mercurio 0,4 mM (ATH-43) y 0,2 mM (ATH-62). La MIC de telurito fue 32 μM para ambas cepas (Tabla 1). Estas concentraciones corresponden a 20 y 40 veces más que los MIC de Hg y de 8 veces más que el MIC de telurito para E. coli. La tinción Gram mostró que ambas cepas bacterianas son bacilos Gram negativo (Fig. 1). Curvas de crecimiento Los aislados ATH-43 y ATH-62 presentaron un retraso en el inicio de la fase de crecimiento exponencial en presencia de ambos tóxicos en relación al crecimiento en LB. Como se observa en la Fig. 2A, ATH-62 presenta un retraso mayor en presencia de HgCl2 que en presencia de TeO3
-2 respecto al

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
119
control; el retraso observado en el aislado ATH-43 para ambos metales es el mismo, describiendo en ambos casos una curva de crecimiento muy similar en toda su extensión (Fig. 2B). Resistencia a antibióticos Los resultados muestran que mientras ATH-62 es susceptible a varios antibióticos, por el contrario ATH-43 es resistente a agentes que interrumpen la síntesis de la pared celular incluyendo β-lactámicos como cefalosporinas de 1a, 2a y 3a generación y penicilinas (Tabla 2). Adicionalmente es resistente a antibióticos que bloquean la síntesis de proteínas, como aminoglicósidos y cloranfenicol. Sólo se observó susceptibilidad a un antibiótico de la familia de las quilononas, que bloquean la DNA girasa. RESULTADOS
Fig. 1. Tinción Gram para las cepas ATH-43 (A) y ATH-62 (B). Las imágenes fueron obtenidas utilizando un microscopio óptico con un aumento de 100X. (Fig. 1. Gram staining for strains ATH-43 (A) and ATH-62 (B). Images were taken using an optic microscope with a 100X lens.
Crecimiento ATH-62
0 5 10 15 200.0
0.5
1.0
1.5
2.0ControlHgCl2 40 uMTeO3-2 20 uM
Tiempo (h)
Densidad O
ptica (600 nm)
Crecimiento ATH-43
0 5 10 15 200.0
0.5
1.0
1.5
2.0ControlHgCl2 40 uMTeO3-2 20 uM
Tiempo (h)
Densidad O
ptica (600 nm)
A B
Figura 2: Curvas de crecimiento de cepas ATH-62 (A) y ATH-43 (B). En LB (círculos), en LB + HgCl2 40 μM (cuadrados) y
en LB + TeO3-2 20 μM (triángulos).
Figure 2: Growth curve for strains ATH-62 (A) and ATH-43 (B). In LB (circles), in LB + HgCl2 40 μM (squares) and in LB + TeO3
-2 20 μM (triangles).
Tabla 1: Características del muestreo, T (°C) óptima de
crecimiento, MICs. Table 1: Sampling features, optimal
growing T (°C), MICs.
Tabla 2: Resistencia a antibióticos cepas ATH-43 y ATH-62.
Table 2: Antibiotic resistance of strains ATH-43 and ATH-62.
R: Resistencia; S: Susceptible
AGRADECIMIENTOS Fondecyt # 1130362, Fondecyt
Postdoctorado # 1130362, Beca Apoyo a la realización de Tesis Doctoral 24121087. Vicerrectoría de Investigación y Postgrado, Universidad San Sebastián.
A B
Antibiótico Familia ATH-43 ATH-62
Cefotaxima (CTX 30µg) Cefalosporina 3ª R R
Amikacina (AK 30µg) Aminoglicósido R S
Gentamicina (GE 10µg) Aminoglicósido R S
Ciprofloxacino (CIP 5µg) Quilonona S S
Cefa lotina (CF 30µg) Cefalosporina 1ª R S
Amp-Sulbactam (SAM 10/10µg) B-lactámico R S
Ceftazidima (CAZ 30µg) Cefalosporina 3ª R S
Cefuroxima (CXM 30µg) Cefalosporina 2ª R R
Sulfatrimetoprim (SXT 25µg) Sulfonamida R R
Cloranfenicol (C 30µg) otros R R
Número de muestra Lugar Coordenadas Resistencia
T°C de crecimiento
MIC de Telurito(µM)
MIC de Mercurio (mM)
43Isla Greenwich, base
Prat 62° 30' S; 59° 39' W Te y Hg 25 32 0,4
62Isla Decepción, caleta péndulo 62° 55' S; 60° 37' W Te y Hg 25 32 0,2

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
120
REFERENCIAS Avazeri C., Turner R.J., Pommier J., Weiner J.H., Giordano G. y Vermeglio A. 1997. Tellurite reductase activity of nitrate reductase is responsible for the basal resistance of Escherichia coli to tellurite. Microbiology 143: 1181-1189 Barkay T. y Olson B.H.. 1986. Phenotypic and genotypic adaptation of aerobic heterotrophic sediment bacterial communities to mercury stress. Applied and Environmental Microbiology 52: 403-406 Bradley D.E. 1985. Detection of tellurite-resistance determinants in IncP plasmids. Journal of General Microbiology 131: 3135-3137 Calderón I.L., Arenas F.A., Pérez J.M., Fuentes D.E., Araya M.A. y Vásquez CC. 2006. Catalases are NAD(P)H-dependent tellurite reductases. PLoS ONE 1: e70 Calderón I.L., Elías A.O., Fuentes E.L., Pradenas G.A., Castro M.E. y Vásquez CC. 2009. Tellurite-mediated disabling of [4Fe-4S] clusters of Escherichia coli dehydratases. Microbiology 155: 1840-1846 Castro M.E., Molina R., Díaz W., Pichuantes S.E. y Vásquez C.C. 2008. The dihydrolipoamide dehydrogenase of Aeromonas caviae ST exhibits NADH-dependent tellurite reductase activity. Biochemical and Biophysical Research Communications 375: 91-94 Cervantes-Vega C., Chavez J., Cordova N.A., de la Mora P. y Amador Velasco J. 1986. Resistance to metals by Pseudomonas aeruginosa clinical isolates. Microbios 48: 159-163 Cortes S. y Fortt A. 2007. Mercury content in Chilean fish and estimated intake levels. Food Additives and Contaminants 24: 955-959 Chasteen T.G. y Bentley R. 2003. Biomethylation of selenium and tellurium: microorganisms and plants. Chemical Reviews 103: 1-25 D'Ltri P. y D'Ltri F. 1978. Mercury contamination: A human tragedy. Environmental Management 2: 3-16 Dittmer D.C. 2003. Tellurium. Chemical & Engineering News 81: 128 Fox B. y Walsh C.T. 1982. Mercuric reductase. Purification and characterization of a transposon-encoded flavoprotein containing an oxidation-reduction-active disulfide. Journal of Biological Chemistry 257: 2498-2503 Hermanns Y.M. y Biester H. 2013. Anthropogenic mercury signals in lake sediments from southernmost Patagonia, Chile. Science of the Total Environment 445-446: 126-135 Komura I., Funaba T. y Izaki K. 1971. Mechanism of mercuric chloride resistance in microorganisms. II. NADPH-dependent reduction of mercuric chloride and vaporization of mercury from mercuric chloride by a multiple drug resistant strain of Escherichia coli. The Journal of Biochemistry 70: 895-901 Misra T.K. 1992. Bacterial resistances to inorganic mercury salts and organomercurials. Plasmid 27: 4-16 Misra T.K., Brown N.L., Fritzinger D.C., Pridmore R.D., Barnes W.M. y Silver, S. 1984. Mercuric ion-resistance operons of plasmid R100 and transposon Tn501: the beginning of the operon including the regulatory region and the first two structural genes. Proceedings of the National Academy of Sciences USA 81: 5975-5979 Nieboer E. y Richardson D.H.S. 1980. The replacement of the nondescript term ‘heavy metals’ by a biologically and chemically significant classification of metal ions. Environmental Pollution Series B, Chemical and Physical 1: 3-26 Pérez J.M., Calderón I.L., Arenas F.A., Fuentes D.E., Pradenas G.A. y Vásquez CC 2007. Bacterial toxicity of potassium tellurite: unveiling an ancient enigma. PLoS ONE 2: e211 Summers A.O. y Silver S. 1972. Mercury resistance in a plasmid-bearing strain of Escherichia coli. Journal of Bacteriology 112: 1228-1236 Tantaleán J.C., Araya M.A., Saavedra C.P., Fuentes D.E., Pérez J.M. y Vásquez CC 2003. The Geobacillus stearothermophilus V iscS gene, encoding cysteine desulfurase, confers resistance to potassium tellurite in Escherichia coli K-12. Journal of Bacteriology 185: 5831-5837 Taylor D.E.. 1999. Bacterial tellurite resistance. Trends in Microbiology 7: 111-115 Thayer J.S. 1984. In organometallic compounds and living organisms. Academic Press eds. Orlando, Florida, USA, Turner R.J., Weiner J.H. y Taylor D.E. 1999. Tellurite-mediated thiol oxidation in Escherichia coli. Microbiology 145: 2549-2557 Whelan K.F. y Colleran E. 1992. Restriction endonuclease mapping of the HI2 incompatibility group plasmid R478. Journal of Bacteriology 174: 1197-1204

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
121
CARACTERIZACIÓN DE LA RESPUESTA A TELURITO EN BACTERIAS HIPER-RESISTENTES AISLADAS DEL TERRITORIO ANTÁRTICO CHILENO
(Characterization of the tellurite response in hyper-resistant bacteria from the Chilean Antarctic Territory)
Muñoz-Villagrán C.1, Pugin B.1, Pozo M.1, Cornejo F.1, Valdivia M.1, Díaz-Vásquez W.1,2,3., Arenas FA.1, CC. Vásquez 1
1 Laboratorio de Microbiología Molecular, Universidad de Santiago de Chile. [email protected] 2 Laboratorio de Microbiología Molecular y Biotecnología Ambiental, Universidad Técnica Federico Santa María. 3 Facultad de Ciencias de la Salud, Universidad San Sebastián. Los microorganismos pueden adaptarse a varias condiciones desfavorables como estrés salino, UV, metales, etc., dado que poseen variados determinantes genéticos que responden frente a estos tipos de estrés. La búsqueda de nuevos mecanismos de resistencia a metales, por ejemplo, presentes en organismos aislados desde hábitats extremos, como el territorio Antártico Chileno, es de gran interés para los investigadores ya que se puede identificar genes que podrían ser útiles en procesos de bioremediación y generación de nanopartículas, entre otros. Este conocimiento podría ser usado en la optimización de diversos procesos industriales para que sean más amigables con el medio ambiente. Se ha observado que microorganismos aislados desde el territorio Antártico Chileno son altamente resistentes a telurito, un oxianión derivado del elemento teluro (Te) que está presente en concentraciones traza en la naturaleza. El Te pertenece al grupo 16A de la Tabla Periódica junto a elementos esenciales para la vida como el oxígeno, azufre y selenio. Sin embargo, a la fecha no se ha encontrado un rol biológico para el teluro (Ba y cols., 2010). Aun cuando en su estado elemental el Te no es nocivo, los oxianiones telurito (TeO3
-2) y telurato (TeO4-2) son extremadamente tóxicos para la mayoría de los
organismos vivos, particularmente bacterias Gram negativo (Chasteen y cols., 2009). El teluro y sus derivados son comúnmente generados como subproducto de la refinación del cobre, oro y otros metales (Green, 2009) y además en la obtención de energía geotérmica. Entre los mecanismos generales de resistencia a metales se encuentra la adsorción, volatilización, liberación de compuestos quelantes al medio, bombas de eflujo, impermeabilidad, disminución en la incorporación de los tóxicos, destoxificación enzimática y secuestro intracelular (Srivastava y Kowshik, 2013; Sochor y cols., 2011). En el caso del telurito se ha determinado que los microorganismos pueden reducirlo enzimática o no enzimáticamente a un estado menos tóxico (Te0) (Chasteen y cols., 2009) con la concomitante producción de superóxido (Pérez y cols., 2007). Esta reducción se puede visualizar por la coloración negra que adquieren las células cuando crecen en presencia de este compuesto (Summers y Jacoby, 1977). Esta característica permitió aislar diversos microorganismos antárticos que crecen en altas concentraciones de telurito, los que se han identificado y su sensibilidad al tóxico ha sido determinada. Aun cuando todos reducen telurito, la idea es que ellos también presentan mecanismos alternativos (o diferentes estrategias) de resistencia que les permiten crecer en presencia de altas concentraciones de este tóxico. De este modo, en este trabajo se analizará, mediante distintas tecnologías, cómo estas células responden a telurito con el fin de caracterizar su respuesta. Para analizar el consumo de telurito por los diferentes aislados antárticos se diluyó cultivos saturados (1:100) con medio LB y se crecieron con agitación hasta una DO600 ~0,3. Luego se adicionó telurito a una concentración correspondiente a ¼ de la MIC de cada cepa. Se tomó alícuotas de 1 ml en el tiempo hasta 74 h, se centrifugaron a 8.000 x g por 3 min y el sobrenadante se utilizó para cuantificar telurito extracelular remanente reduciéndolo con NaBH4 (Molina y cols., 2010). La cuantificación de teluro

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
122
elemental se llevó a cabo por espectroscopía de emisión acoplada a espectroscopía óptica inducida por emisión de plasma (ICP - OES). Para ello, se trató previamente las cepas antárticas con telurito y se crecieron en LB hasta DO600 ~0,5-0,6 y se tomó 50 ml en cada punto evaluado. El pellet fue suspendido en 1 ml de buffer MES NaOH 50 mM pH 7,4. Se agregó 10% HNO3 para diluir las muestra 10 veces. El volumen total resultante fue utilizado para la determinación de teluro en un equipo Spectro CIROS Vision ICP-OES. Se utilizó teluro comercial para construir una curva de calibración. La viabilidad celular se determinó por citometría de flujo, creciendo las cepas antárticas hasta DO600 ~0,3-0,4. Luego se trataron con la concentración de telurito correspondiente con agitación por 75 min. Se centrifugaron a 8.000 x g por 5 min y el pellet se lavó dos veces con buffer Tris-HCl 50 mM pH 7,4. Finalmente se adicionó 3 µl de yoduro de propidio (1 mg/ml), se incubó por 5 min y se determinó con filtro PE. Como control positivo se incubó las células a 90 ºC por 10 min (Tanaka y cols. 2000). Los resultados obtenidos dan cuenta que las cepas aisladas presentan distintos niveles de resistencia a telurito, como reflejan las concentraciones mínimas inhibitorias (Tabla 1). Esto permitió clasificarlas como cepas más resistentes (MIC > 200 µg/ml) como BNF05, BNF08 y BNF20 y cepas menos resistentes (MIC < 200 µg/ml) como BNF01, BNF15 y BNF22.
Strains Most closely related species (%
similitary) Colony color
Morphology Gram staining
Optimal TºC
Td (h)
MIC (µg/ml)
BNF01 Sraphylococcus haemolyticus (99%) White Coccus Gram + 37ºC 0,65 225 BNF05 Staphylococcus sciuri (99%) Orange Coccus Gram + 37ºC 0,51 525 BNF08 Psychrobacter immobilis (99%) Colorless Rods Gram - 25ºC 1,65 275 BNF15 Acinetobacter haemolyticus (99%) Colorless Rods Gram - 25ºC 1,95 200 BNF20 Psychrobacter immobilis (99%) Colorless Rods Gram - 25ºC 1,69 575 BNF22 Pseudomonas lini (99%) Colorless Rods Gram - 25 1,34 35 E. coli -- Colorless Rods Gram - 37ºC 0,58 1
Tabla 1: Caracterización de cepas antárticas aisladas. E. coli BW25113 se usó como referencia. Tabla 1: Characterization of isolated antarctic strains. E. coli BW25113 was used as reference.
Para analizar la capacidad de estas cepas de consumir telurito, se trataron con concentraciones subletales del tóxico y se determinó a distintos tiempos cuanto de este compuesto quedaba en el sobrenadante. Como se indica en la Fig. 1A y 1B, las cepas presentan distintas capacidades para incorporar este oxianión. Esto se puede apreciar al comparar las pendientes de las curvas de cada cepa, así como el porcentaje residual de telurito en el sobrenadante después de 72 h (paréntesis en Fig. 1A y B). No se observa una correlación entre las cepas que resisten mayores concentraciones de telurito y una mayor incorporación de éste. Por ejemplo, la cepa BNF05 reduce más rápido y acumula más teluro intracelular (Fig. 1A y Fig. 1C). Sin embargo, no es la más resistente. Por otra parte, la cepa BNF20 tiene un mayor MIC pero no consume más que la cepa BNF08, que posee una menor resistencia. La cepa BNF15 que resiste 200 µg/ml de telurito no consume más que la BNF22, cuyo MIC es 35 µg/ml. Esto sugiere que estas cepas no presentan una respuesta universal a este tóxico. Los ensayos de viabilidad dan cuenta que todas las cepas tratadas con concentraciones subletales de telurito muestran un aumento en el porcentaje de células muertas en comparación al control (Fig. 2A), observándose ciertas diferencias entre cepas que son más sensibles al tratamiento como BNF15 y BNF22. Estos datos concuerdan con las curvas de crecimiento, ya estas mismas cepas son las que ven más afectado su crecimiento en comparación al control (no mostrado).
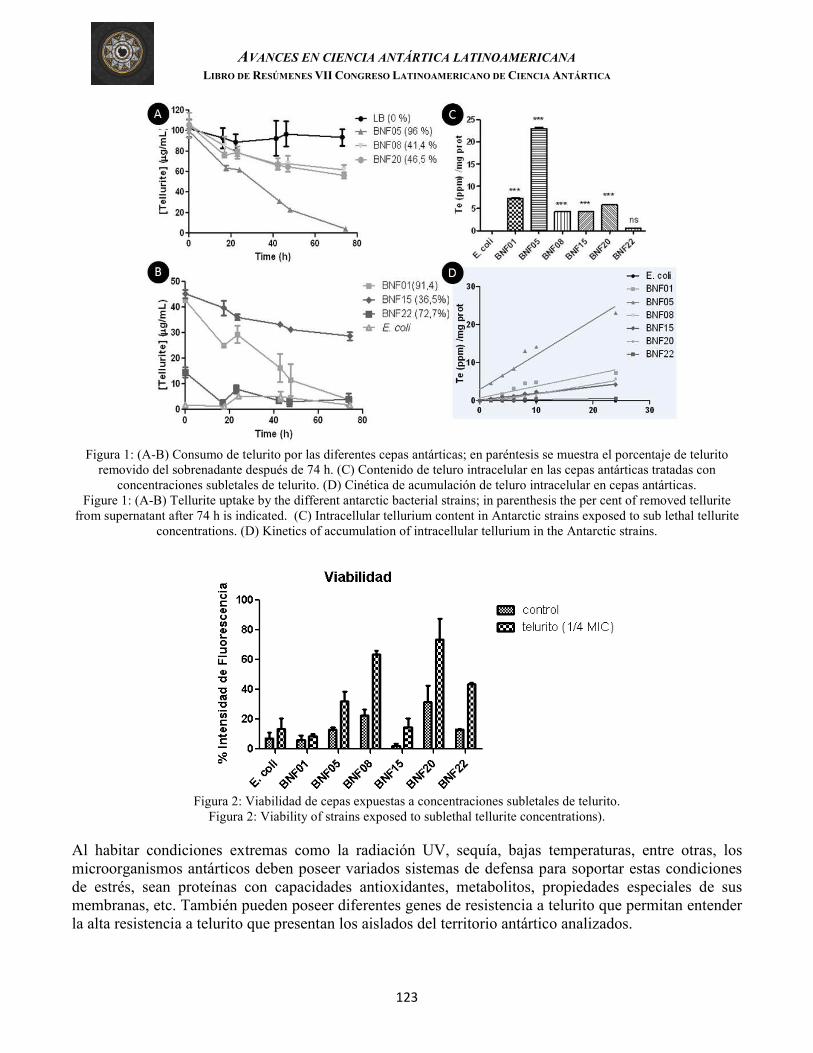
AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
123
Figura 1: (A-B) Consumo de telurito por las diferentes cepas antárticas; en paréntesis se muestra el porcentaje de telurito
removido del sobrenadante después de 74 h. (C) Contenido de teluro intracelular en las cepas antárticas tratadas con concentraciones subletales de telurito. (D) Cinética de acumulación de teluro intracelular en cepas antárticas.
Figure 1: (A-B) Tellurite uptake by the different antarctic bacterial strains; in parenthesis the per cent of removed tellurite from supernatant after 74 h is indicated. (C) Intracellular tellurium content in Antarctic strains exposed to sub lethal tellurite
concentrations. (D) Kinetics of accumulation of intracellular tellurium in the Antarctic strains.
Figura 2: Viabilidad de cepas expuestas a concentraciones subletales de telurito.
Figura 2: Viability of strains exposed to sublethal tellurite concentrations).
Al habitar condiciones extremas como la radiación UV, sequía, bajas temperaturas, entre otras, los microorganismos antárticos deben poseer variados sistemas de defensa para soportar estas condiciones de estrés, sean proteínas con capacidades antioxidantes, metabolitos, propiedades especiales de sus membranas, etc. También pueden poseer diferentes genes de resistencia a telurito que permitan entender la alta resistencia a telurito que presentan los aislados del territorio antártico analizados.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
124
Un mecanismo de resistencia bacteriana a este oxianión es su reducción a teluro elemental, fenómeno que ocurre en todas las cepas analizadas. Sin embargo, la eficiencia de reducción no es la misma entre ellas, ni tampoco existe una relación directamente proporcional con las cepas más resistentes (Fig.1 y Tabla 1). Esto se podría explicar dado que no todos los microorganismos poseen probablemente un mecanismo de resistencia global, sino que puede haber algunos que consumen menos telurito, o lo expulsan, o sus enzimas reductoras de telurito no son tan eficientes. Estas y otras interrogantes se estudian en el laboratorio con la finalidad de caracterizar los diferentes mecanismos de resistencia que poseen estos asilados antárticos. En conclusión, en este trabajo se determinó que las cepas aisladas del territorio antártico chileno son capaces de resistir altas concentraciones de telurito, siendo esta resistencia variable entre ellas. Aunque todas las cepas reducen telurito a teluro elemental, la eficiencia de la reacción varía entre ellas, lo cual dependería de los mecanismos intrínsecos de resistencia que posea cada especie. Agradecimientos Este trabajo fue financiado por el proyecto Fondecyt # 1130362 (CCV) y el proyecto Fondecyt Postdoctorado # 1130362 (FAA). REFERENCIAS Ba, L.A., Döring, M., Jamier, V. y Jacob, C. 2010. Tellurium: an element with great biological potency and potential. Org. Biomol. Chem. 8, 4203-4216. Chasteen, T.G., Fuentes, D.E., Tantaleán, J.C. y Vásquez, C.C. 2009. Tellurite: history, oxidative stress, and molecular mechanisms of resistance. FEMS Microbiol. Rev. 33, 820-832. Green, M. 2009. Estimates of Te and In prices from direct mining of known ores. Prog. Photovolt. Res. Appl. 17, 346-359. Molina, R.C., Burra, R., Pérez-Donoso, J.M., Elías, A.O., Muñoz, C., Montes, R.A., Chasteen, T.G. y Vásquez, C.C. (2010). Simple, fast, and sensitive method for quantification of tellurite in culture media. Appl. Environ. Microbiol. 76, 4901-4904. Pérez, J.M., Calderón, I.L., Arenas, F.A., Fuentes, D.E., Pradenas, G.A., Fuentes, E.L., Sandoval, J.M., Castro, M.E., Elías, A.O. y Vásquez, C.C. (2007). Baterial toxicity of potassium tellurite: unveiling an ancient enigma. PLoS ONE 2, e211. Sochor, J., Zitka, O., Hynek, D., Jilkova, E., Krejcova, L., Trnkova, L., Adam, V., Hubalek, J., Kynicky, J., Vrba, R. y Kizek, R. 2011. Bio-sensing of cadmium (II) ions using Staphylococcus aureus. Sensors 11, 10638-10663. Srivastava, P. y Kowshik, M., 2013. Mechanisms of metal resistance and homeostasis. In Haloarchaea Archaea: 732864. Summers, A. y Jacoby, G. 1977. Plasmid determined resistance to tellurium compounds. J. Bacteriol. 129, 276-281. Tanaka, Y., Yamaguchi, N. y Nasu, M. 2000. Viability of Escherichia coli O157:H7 in natural river water determined by the use of flow cytometry. J. Appl. Microbiol. 88, 228-236.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
125
IDENTIFICACIÓN Y CARACTERIZACIÓN DE UNA SHEWANELLA ANTARTICA REDUCTORA DE TELURITO Y COBRE
(Identification and characterization of a tellurite- and copper-reducing Antarctic Shewanella)
Valdivia, M.1, Muñoz, C. 1, Vargas, J.1, Pérez, J.M. 2, Vásquez, C. 1
1 Laboratorio Microbiología Molecular, Facultad de Química y Biología. Universidad de Santiago de Chile. Av. Libertador Bernardo O´Higgins #3363. Santiago. [email protected]
2Laboratorio de Bionanotecnología y Microbiología, Centro de Bioinformática y Biología Integrativa (CBIB), Facultad de Ciencias Biológicas, Universidad Andrés Bello. Republica 239. El estrés es una condición inevitable de la vida de todos los organismos. Esto se aplica a microorganismos que residen o se encuentran en todo tipo de ambientes, incluyendo algunos extremos como la Antártica (Gerday y Glansdorff, 2007). Los factores medioambientales más comunes que afectan a los microorganismos son temperatura, pH, limitación de nutrientes, presión, radiación y metales, entre otros (Storz y Hengge-Aronis, 2000). Sin embargo, los microorganismos presentan la habilidad de adaptarse rápidamente a cambios en su entorno. La regulación que modula esta adaptación es muy compleja y la mayor parte de los estudios sobre mecanismos de respuesta a estrés han utilizado como modelo Escherichia coli y Bacillus subtilis. Una bacteria ambiental que ha emergido en los últimos años corresponde a Shewanella, con forma de vara, Gram-negativo, aeróbica o anaeróbico facultativa, con flagelo polar y fácilmente cultivable en laboratorio (Yin y Gao, 2011). Mientras algunos aislados no han sido caracterizados, 52 especies han sido reconocidas a la fecha (Verma y cols., 2011). La principal característica de Shewanella es su habilidad para usar un amplio rango de aceptores de electrones durante la respiración anaeróbica incluyendo fumarato, nitrato, nitrito, tiosulfato, sulfuro, Fe(III), Mn(III), Cr(VI), U (VI), As (V), V(V) y otros (Fredrickson, 2008). Como resultado de esta propiedad, Shewanella ha llamado mucho la atención en bioremedación, biogeoquímica y bioelectricidad (Yin y Gao, 2011). Recientemente, los compuestos de teluro (Te) han sido utilizados ampliamente en la refinería del petróleo, paneles solares, óptica, producción de lentes y sensores (Kim y cols., 2012). Sin embargo, su uso ha incrementado la contaminación ambiental (Chasteen y cols., 2009). En el medio ambiente, el teluro se encuentra en su estado elemental Te(0) y no presenta toxicidad aparente. Sin embargo, el oxianión telurito (IV) es altamente tóxico para la mayoría de los organismos vivos, especialmente células procariotas con una concentración inhibitoria mínima de 4 µM. El cobre, por otra parte, es requerido para el crecimiento de todos los organismos vivos, aunque en altas concentraciones es tóxico (Toes y cols., 2008). Shewanella oneidensis MR-1 reduce telurito a teluro, con una acumulación intracelular y extracelular de nanocristales de Te(0) en la bacteria (Kim y cols., 2012). Además, esta bacteria expresa dos genes que participan en el transporte de cobre entre la membrana citoplasmática y la membrana externa (Toes y cols., 2008). A la fecha, esto es lo que se conoce sobre la reducción de telurito y cobre por esta bacteria. Se aisló bacterias de muestras obtenidas durante una expedición a la Antártica. Para seleccionar se utilizó el criterio de resistencia a telurito, el que a muy bajas concentraciones (E. coli por ejemplo presenta un MIC de 4 µM) es tóxico para los microorganismos. Se secuenció en gen del RNA16S de aquellas bacterias que resultaron resistentes al tóxico, el que se comparó en Ribosomal Database Project (base de datos de secuencias 16S) y se obtuvo que una de las bacterias que presentaban resistencia a telurito correspondía a Shewanella spp. El análisis bioquímico/microbiológico mostró que aun cuando la

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
126
cepa era similar a S. putrefaciens, se diferenciaba de esta en su habilidad para metabolizar sacarosa y arabinosa (no mostrado). La resistencia o sensibilidad a telurito y cobre de la Shewanella antártica se determinó por halos de inhibición del crecimiento. Para ello, las células fueron crecidas hasta una DO600 ~0,3 se diluyeron 1:2 con medio LB fresco y se aplicaron homogéneamente sobre la superficie de placas LB. Luego de secar brevemente, se colocó discos de papel filtro (6 mm) estéril sobre los que se depositó 10 µl de telurito de potasio 400 mM o 10 µl de sulfato de cobre 500 mM. Las placas se incubaron durante la noche a 25 ºC en presencia o ausencia de oxígeno. Dado que S. oneidensis MR-1 puede reducir telurito y otros metales, se ensayó una amplia variedad de metales no descritos anteriormente como aceptores de electrones en Shewanella (níquel, cadmio, cobre, aluminio, etc.). De todos los metales analizados, los que más llamaron la atención fueron telurito y cobre. Cuando la bacteria crece en condiciones aeróbicas se observa una alta sensibilidad al tóxico, mientras que en ausencia de oxígeno la reducción de telurito es total (Fig. 1).
Fig. 1. Zona de inhibición de crecimiento de Shewanella antártica en presencia de telurito. A) aerobiosis. B) anaerobiosis. (Growth inhibition zones of the antarctic Shewanella in the presence of tellurite. A) aerobic. B) anaerobic. Shewanella crece sin problema en medio LB. Para realizar experimentos más detallados, se probó distintos medios de crecimiento y fuentes de carbono. Sin embargo, la bacteria no creció en ninguna otra condición (no mostrado). La cepa se creció rutinariamente a partir de la dilución de un inóculo dejado la noche anterior. Una vez saturado, el cultivo se centrifugó a 6.000 x g por 10 min y el pellet se suspendió en tampón HEPES 10 mM, pH 7,0. A 1 ml de HEPES 10 mM que contenía telurito 100 µM, se agregó 10 µl de la Shewanella resuspendida. Cada tubo contuvo 10 mM de un dador de electrones apropiado. Se observó que el mejor dador era lactato seguido por piruvato, mientras que en citrato hubo una reducción muy baja, similar al control, en que no se agregó fuente de carbono (Fig. 2). El experimento se realizó en anaerobiosis.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
127
Figura: 2. Reducción de telurito en Shewanella antártica usando diferentes dadores de electrones. 1) sin fuente de carbono. 2)
lactato 10 mM. 3) piruvato 10 mM. 4) citrato 10 mM. Figure 2: Tellurite reduction by antarctic Shewanella using different electron donors. 1) without carbon source. 2) 10 mM
lactate. 3) 10 mM pyruvate. 4) 10 mM citrate).
Se realizó halos de inhibición del crecimiento con diversos metales, sin embargo, con único que se observó algo interesante fue con cobre. A las 24 h y en anaerobiosis se observó un halo metálico, fenómeno que no ocurre en aerobiosis (Fig. 3A). Para determinar si el fenómeno observado era producido por Shewanella antártica, se tomó una muestra del halo metálico y se creció en LB. Se volvió a repetir el experimento y se obtuvo el mismo resultado (no mostrado). Se puede observar mejor el halo metálico en la Fig. 3B. Experimentos adicionales donde se ha utilizado directamente concentrado de cobre extraído de Chuquicamata, han mostrado la generación de un halo metálico similar al de la figura 3B (no mostrado).
Figura 3: Zona de inhibición del crecimiento de Shewanella antártica por cobre. A) Shewanella crecida en aerobiosis
(izquierda) y en anaerobiosis (derecha). B) Halo metálico de cobre. Figure 3: Growth inhibition zones for antarctic Shewanella by copper. A) Shewanella grown in aerobic (left) or anaerobic
conditions (right). B) Metallic copper halo). En este estudio se determinó que la bacteria reduce telurito a muy altas concentraciones en condiciones de anaerobiosis, sugiriendo que la bacteria podría estar usando telurito como aceptor final de electrones. Sin embargo, esto es controversial debido a que si bien hay reducción de telurito, se desconoce como ocurre esta reducción. Según Kim y cols. (2012) habría reducción intracelular y extracelular de telurito en anaerobiosis. Por otro lado, Shewanella puede liberar flavinas al medio que utiliza como transportadores de electrones para reducir ciertos compuestos en el espacio extracelular (Marsili y cols., 2008), además de otros mecanismos para reducir otros aceptores de electrones. En el caso del cobre, no se conoce mucho sobre la reducción de cobre por Shewanella, de modo que esta sería una primera aproximación de una posible respiración de cobre por esta bacteria. Dado que la generación del halo metálico sólo ocurre en condiciones de carencia de oxígeno, la bacteria se vería obligada a utilizar cobre como aceptor final de electrones.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
128
En los últimos años, nuestro laboratorio se ha dedicado fuertemente a la reducción intracelular de telurito, descubriendo varias enzimas que reducen el tóxico. Se propone que la reducción de cobre y telurito en esta bacteria antártica ocurre en forma extracelular y mediado por flavinas, cuya demostración requiere de experimentos adicionales que se encuentran en ejecución en el laboratorio. AGRADECIMIENTOS Este trabajo fue financiado por el proyecto Fondecyt Regular # 1130362. REFERENCIAS Chasteen, T., Fuentes, D., Tantaleán, J. y Vásquez, C. 2009. Tellurite: history, oxidative stress, and molecular mechanisms of resistance. FEMS Microbiol. Rev. 33: 820–832. Fredrickson, J., Romine, M., Beliaev, A., Auchtung, J., Driscoll, M., Gardner, T., Nealson, K., Osterman, A., Pinchuk, G., Reed, J., Rodionov, D., Rodrigues, J., Saffarini, D., Serres, M., Spormann, A., Zhulin, I. y Tiedje, J. 2008.Towards environmental systems biology of Shewanella. Nat. Rev. Microbiol. 6: 592-603. Gerday, C. y Glansdorff, N. 2007. Physiology and biochemistry of extremophiles. American Society for Microbiology. Kim, D., Kanaly, R. y Hur, H. 2012. Biological accumulation of tellurium nanorod structures via reduction of tellurite by Shewanella oneidensis MR-1. Bioresour. Technol. 125: 127-131. Marsili, E., Baron, D., Shikhare, I., Coursolle, D., Gralnick, J. y Bond, D. 2008. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 105: 3968-3973. Storz, G. y Hengge-Aronis, R. 2000. Bacterial stress responses. American Society for Microbiology. Toes, A., Daleke, M., Kuenen, G. y Muyzer, G. 2008. Expression of copA and cusA in Shewanella during copper stress. Microbiology 154:2709–2718 Verma, P., Pandey, P., Gupta, A., Kim, H., Baik, K., Seong, C., Patole, M. y Shouche, Y. 2011. Shewanella indica sp. nov., isolated from the sediment of Arabian Sea. Int. J. Syst. Evol. Microbiol. 61: 2058-2064. Yin, J. y Gao, H. 2011. Stress responses of Shewanella. Int. J. Microbiol. 863623.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
129
DESAFIOS DA PESQUISA GLACIOLÓGICA E CLIMÁTICA ANTÁRTICA: UM OLHAR SUL-AMERICANO
(Challenges of the Antarctic glaciological and climatic research: A South American point of view)
Simões, J.C.1 1Instituto Nacional de Ciência e Tecnologia da Criosfera, Centro Polar e Climático, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, Brasil O conhecimento do papel da cobertura de gelo antártico no sistema climático global teve avanços fundamentais na última década, por exemplo, estudos de testemunhos de gelo proveram informações essenciais sobre a evolução da química atmosférica e da variabilidade do cinturão dos ventos de oeste. No entanto permanecem várias questões não respondidas sobre a relação Antártica-América do Sul. Por exemplo, sobre o papel da extensão do gelo marinho antártico na variabilidade climática da América do Sul, a determinação de trajetória de poluentes deste continente para o interior da Antártica, a ligação do registro paleoclimático dos dois continentes. Esta apresentação apresenta alguns desses desafios.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
130
LA IMPORTANCIA HIDROLÓGICA DE "CRYOCONITE HOLES" SOBRE UN GLACIAR EN ANTÁRTICA
(The hydrological significance of cryoconite holes on an Antarctic glacier)
MacDonell, S.1,2, Fitzsimons, S.2, and Sharp, M.3 1 Centro de Estudios Avanzados en Zonas Áridas, Campus Andrés Bello, Raúl Bitran s/n, La Serena, Chile. Email: [email protected] 2 Department of Geography, University of Otago, PO Box 56, Dunedin, New Zealand 3 Department of Earth and Atmospheric Sciences, University of Alberta, Edmonton, Alberta T6G 2E3, Canada INTRODUCTION Cryoconite holes are important water stores on the surface of cold glaciers, due to their ability to keep water as liquid beneath the ice surface, even where the surface ice is not at the melting point (MacDonell and Fitzsimons, 2008). They form when differential ablation from the preferential heating of sediment on the glacier surface causes a water-filled depression to form within the surface ice layer. Their development is largely restricted to cold glaciers in high latitude and high altitude regions (Takeuchi, 2002; Fountain et al., 2004; Anesio et al., 2007) where low melt rates are unable to wash sediment from the glacier surface. In arid regions, such as the McMurdo Dry Valleys, they are often ice-covered, and can remain isolated from the atmosphere and other holes for several years (Figure 1; Fountain et al., 2004; Bagshaw et al., 2007). Cryoconite holes have attracted a great deal of attention recently because they are often seen as simple, self-contained models which can be used to understand the nature of the interaction between physical hydrology and biogeochemical processes that take place on the surface of glaciers (Hodson et al., 2008; MacDonell and Fitzsimons, 2008). On arid and semi-arid glaciers, cryoconite holes can also play an important role in the production, routing and storage of surface and near-surface meltwater and they can also facilitate drainage from the glacier (Fountain et al., 2004; Tranter et al., 2005; MacDonell and Fitzsimons, 2012). One major limitation currently facing cryoconite hole researchers is the inability to understand connectivity between cryoconite holes. Without this understanding, it is difficult to account for processes such as: sediment cycling (MacDonell et al., 2013); meltwater delivery to the glacier outlet (Tranter et al., 2004); solute pathways within the glacier system (Hodson et al., 2005), or ecosystem stability (Foreman et al., 2007). This paper aims to understand how cryoconite holes may connect and disconnect from the wider hydrological system, and to what degree connections may vary spatially and temporally. To achieve these objectives we have undertaken a study of the distribution and biogeochemical characteristics of cryoconite holes on the surface of the Wright Lower Glacier (Figure 1), a small piedmont glacier in the McMurdo Dry Valleys, Antarctica. MATERIALS AND METHOD This study took a multifaceted approach that involved field survey of the distribution and development of cryoconite holes, measurement of the thermal properties of the glacier surface, sampling of water from cryoconite holes that was followed by analysis of the solute concentrations of the water samples.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
131
Figure 1: Map of the Wright Lower Glacier, McMurdo Dry Valleys, Antarctica, showing sampling locations. The inset photo
shows two cryoconite holes that are approximately 45 cm in diameter. Figura 1: Mapa del glaciar Wright Lower, McMurdo Dry Valleys, Antártica, que muestra los sitios de medición. La foto en el
recuadro muestra dos "cryoconite holes" los cuales tienen un diametro de 45 cm aproximadamente. At ten locations at 100 m intervals along the centreline of the Wright Lower Glacier (Figure 1, A-J), 25 m2 maps were made of cryoconite hole distribution and hole dimensions were recorded. Water samples were collected from five cryoconite holes at each of the ten sites, and ice samples were taken at 40 mm and 750 mm depth at each location. Water and ice samples were analysed for their anion and cation content at the Department of Geography, University of Otago. Chloride concentrations were used to approximate the length of cryoconite hole hydrological isolation following Fountain et al. (2004):
∆t =M (t)a× i
−h
dz
dt
−1
(1)
where: M(t) is the chloride concentration (µmol L-1) at a particular time t (years); a is the cross-sectional area of the hole (cm2); i is the average concentration of chloride (µmol L-1) in the surrounding ice, which is assumed to be constant through space and time; h is the depth of the hole (cm); and dz/dt is the melt rate at the base of the hole, which is assumed to be equal to the ablation rate of the ice surface. If Equation 1 gives a result of greater than zero, then that is the number of years that the hole has been isolated.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
132
RESULTS AND DISCUSSION Along the centreline of the Wright Lower Glacier, the average percentage area covered by cryoconite holes is 3.5%, and hole distribution depends on local topographic features (for example: no cryoconite holes were found on south-facing slopes), and the local ablation rate. Cryoconite hole diameter, ice cover thickness and water volume are positively correlated to distance from the terminus, elevation and slope, and negatively correlated with local ablation rate. Head space and total depth were also positively correlated with distance from the terminus and elevation, were negatively correlated with local ablation rate, but were not correlated with slope. The depth of water contained in a hole was not related to local surface properties. An average hole along the centreline contains 0.036 ± 0.03 m3 of water, and when this quantity is extrapolated across the planar surface of the glacier (an area of 31 km2), c40000 ± 30000 m3 of water is stored within, or delivered from, cryoconite holes. This accounts for approximately 27% of seasonal water yield from the planar region of the glacier. The amount of water draining from and stored within the planar surface is likely to vary throughout a melt season. Observations of cryoconite hole connectivity included: connections with surface fractures; shallow sub-surface channels; coalescence with other cryoconite holes; and intergranular drainage. It appeared that connections with cracks and other near-surface channels were largely ‘fortuitous’, that is a cryoconite hole had to form within a channelized zone to become connected to these networks. Observations of intergranular drainage during excavation of surface ice were widespread across the glacier surface, but it is not known how this process is limited spatially and temporally. By its nature, this process is unlikely to transport large quantities of water, but it does challenge the idea of cryoconite hole ‘isolation’. In order to assess the ratio of stored to transported water, an understanding of potential and real cryoconite hole connectivity is required. Using Equation 1 it was found that at least 20% of the measured holes on the planar surface (n = 50) were either connected to other holes, to intergranular passageways or to near-surface conduits. The isolation age suggests that the average age of the holes was four years, and that 40 of the 50 sampled holes were hydrologically isolated to a maximum age of 28 years at Flag ‘E’. The percentage of cryoconite holes that were connected in this study was considerably lower than that recorded by Fountain et al. (2004) on the Canada Glacier, McMurdo Dry Valleys. From a sample of 16 holes, they calculated that 56% of holes were connected. Fountain et al. (2004) postulated that holes were connected via surface and near-surface channels. In this study, all sampled holes were expected to be ‘isolated’ as holes which were obviously connected via conduits and cracks were avoided during sampling. Three holes were calculated to be ‘connected’ around flag ‘H’, however the connected holes were mostly located in the terminus region around flags ‘A’ and ‘B’ respectively. The preference of connectivity at the glacier terminus is possibly due to warmer air temperatures enabling higher rates of intergranular flow, and/or higher rates of ablation. These processes also mean that holes near the terminus are likely to be more transient, and hence younger, due to the higher likelihood of stripping events at lower elevations caused by relatively high air temperatures. However, as intergranular drainage was observed across the glacier surface, it is difficult to determine what causes a cryoconite hole to be ‘isolated’ or ‘connected’, as technically any hole experiencing intergranular drainage is by definition ‘connected’. It is possible that the result of Equation 1 does not give a binary ‘connected’ or ‘isolated’ signal, but instead can be used as a measure of connectedness to the drainage system.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
133
CONCLUSIONS Cryoconite holes on glacier surfaces in the McMurdo Dry Valleys are connected to the wider
drainage system in a myriad of ways. Within this study, we observed holes connected via discrete
channels, including through cracks, fractures, near-surface channels and open channels. We also
observed intergranular flow around cryoconite holes, which has not been recorded at other
McMurdo Dry Valleys sites. The possibility of passive connectivity has real implications for water
and solute transfer on cold glacier surfaces. According to the isolation time calculation, cryoconite hole connectivity is largely limited to areas
near the glacier terminus. However, the isolation age model should be used with caution, as it is
possibly better considered as an index of connectivity. Until detailed testing has been conducted,
the results should be treated with caution. ACKNOWLEDGEMENTS Financial assistance for SM was provided by a University of Otago Postgraduate Scholarship, and the Sir Robin Irvine Antarctica New Zealand Doctoral Scholarship. Financial support for SF was provided by a University of Otago Research Grant. Logistical support was provided by Antarctica New Zealand. REFERENCES Anesio AM, Mindl B, Laybourn-Parry J, Hodson AJ, Sattler B. 2007. Viral dynamics in cryoconite holes on a high Arctic glacier (Svalbard). Journal of Geophysical Research 112:G04S31. DOI: 10.1029/2006JG000350. Bagshaw EA, Tranter M, Fountain AG, Welch KA, Basagic H, Lyons WB. 2007. Biogeochemical evolution of cryoconite holes on Canada Glacier, Taylor Valley, Antarctica. Journal of Geophysical Research 112:G04S35. DOI: 10.1029/2007JG000442. Fountain AG, Tranter M, Nylen TH, Lewis KJ, Mueller DR. 2004. Evolution of cryoconite holes and their contribution to meltwater runoff from glaciers in the McMurdo Dry Valleys, Antarctica. Journal of Glaciology 50(168): 35–45. Hodson AJ, Mumford PN, Kohler J, Wynn PM. 2005. The High Arctic glacial ecosystem: new insights from nutrient budgets. Biogeochemistry 72: 233–56. Hodson AJ, Anesio AM, Tranter M, Fountain AG, Osborn AM, Priscu J, Laybourn-Parry J, Sattler B. 2008. Glacial ecosystems. Ecological Monographs 78: 41–67. MacDonell S, Fitzsimons S. 2008. The formation and hydrological significance of cryoconite holes. Progress in Physical Geography 32(6): 595–610. DOI: 10.1177/0309133308101382. MacDonell S, Fitzsimons S. 2012. Observations of cryoconite hole system processes on an Antarctic glacier, and implications for the wider glacier ecosystem. Revista Chilena de Historia Natural. 85: 393-407, MacDonell S, Fitzsimons S, Mölg T. 2012. Seasonal sediment fluxes forcing supraglacial melting on the Wright Lower Glacier, McMurdo Dry Valleys, Antarctica. Hydrological Processes. doi: 10.1002/hyp.9444. Takeuchi N. 2002. Optical characteristics of cryoconite surface dust on glaciers: the relationship between light absorbency and the property of organic matter contained in the cryoconite. Annals of Glaciology 34: 409–14. Tranter M, Fountain AG, Fritsen CH, Lyons WB, Priscu JC, Statham PJ, Welch KA. 2004. Extreme hydrochemical conditions in natural microcosms entombed within Antarctic ice. Hydrological Processes 18(2): 379–387. Tranter M, Fountain AG, Lyons WB, Nylen TH, Welch KA. 2005. The chemical composition of runoff from Canada Glacier, Antarctica: implications for glacier hydrology during a cool summer. Annals of Glaciology 40: 15-19.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
134
EVIDENCIAS GEOMORFOLOGICAS Y ESTRATIGRAFICAS DEL RETROCESO ACTUAL DEL GLACIAR QUITO Y DE UNA GLACIACION TERCIARIA EN LA ISLA GREENWICH,
ARCHIPIELAGO SHETLAND DEL SUR, CONTINENTE ANTÁRTICO. (Geomorphologic and stratigraphic evidence of the current retreat of Quito Glacier and a Tertiary
glaciations on Greenwich Island, South Shetland islands, Antarctica)
Bezada, M 1., Alfonso, J2., Carrera, J2., Terán, B3., Hernández, J.4
1 Universidad Pedagógica Experimental Libertador. Departamento de Ciencias de la Tierra, Laboratorio de Pedología y Cuaternario. Caracas, Venezuela, [email protected] 2 Instituto Venezolano de Investigaciones Científicas. Centro de Oceanología y Estudios Antárticos 3 Instituto Oceanográfico de la Armada, (INOCAR). Ecuador. 4 Universidad Central de Venezuela, Instituto Jardín Botánico de Caracas. Venezuela. INTRODUCCIÓN Por su ubicación geográfica en la parte más septentrional del continente Antártico, la isla Greenwich del archipiélago de las Shetlands del sur, presenta condiciones muy favorables para el estudio del retroceso glaciar desde el Último Máximo Glacial hasta el actual, inducido por el calentamiento global catalizado por los gases de efecto invernadero. En estas islas es donde mejor se han reportado los retrocesos glaciares antártico y por esta razón realizamos observaciones detalladas en el frente del glaciar Quito en la isla Greenwich. Adicionalmente, esta área de la Antártida, es muy importante, por preservar una importante información geológica y paleoambiental desde el Cenozoico, la cual está relacionada con la evolución climática de la Tierra, donde uno de los eventos más significativos fue la transición de condiciones cálidas y libre de hielo, que caracterizaron el Máximo Térmico del Inicio del Eoceno al paso progresivo a climas fríos y condiciones glaciales que predominan en la Antártida desde entonces. Este cambio de condiciones ha sido explicado entre otras causas por la evolución tectónica de la región que condujo a la creación de la Corriente Circumpolar Antártica y un enfriamiento gradual desde hace 34 millones de años, en el límite Eoceno-Oligoceno. Esta historia glacial del Terciario ha sido mayormente revelada a través de núcleos de sedimentos marinos profundos (Zachos et al 2001), los cuales a través del análisis de múltiples proxis, sedimentológicos, paleobotánicos, paleofaunales y a través del uso de isotopos de oxígeno han permitido hacer estas inferencias paleoclimáticas, que indican un enfriamiento terrestre y un crecimiento de las capa de hielo sobre el continente, marcarían el inicio de las glaciaciones del Terciario durante este intervalo de tiempo (Barron et al., 1991; Thompson 2004). De esta manera se ha podido definir en la estratigrafía del Cenozoico de la isla Rey Jorge, una secuencia de eventos glaciales e interglaciales, que representan al menos dos expansiones de capas de hielo durante el Oligoceno y el Mioceno correspondiente a las glaciaciones definidas como Polonez y Melville, con edades de 32-26 millones de años y 23-20 millones de años respectivamente (Birkenmajer, 2001., Troedson y Riding, 2002, Troedson y Smellie 2002). Por otra parte, la primera evidencia de un glaciar alpino Terciario en la Antártida occidental fue la presentada por Birkenmajer el al (2005), en la Formación Point Thomas en la bahía Almirantazgo de la isla Rey Jorge, donde un diamicton glacial es infra y suprayacido por sendas coladas de lavas, las cuales dieron una edad radiométrica de K-Ar., entre 45-41 millones de años, es decir, del Eoceno medio. Este hecho, nos motivó a buscar evidencias de glaciaciones del Terciario en el área estudiada. METODOLOGIA

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
135
Durante el verano austral del año 2013, se realizaron observaciones geomorfológicas, recolección de muestras litológicas y mediciones de campo para determinar el movimiento del frente del Glaciar Quito en la costa occidental de la Base Científica Antártica Pedro Vicente Maldonado (BCAPVM) de la República del Ecuador y para buscar evidencias geomorfológicas, en las áreas emergidas de glaciaciones del Terciario y del Cuaternario. La cronología del evento más antiguo se obtendrá mediante dataciones radiométricas Potasio Argón del basalto que suprayace al diacmicton glacial del Terciario. Los análisis petrográficos de litología contenida en los sedimentos glaciales del Terciario y el Cuaternario se realizaron mediante el análisis de secciones finas y una microscopia de luz polarizada. RESULTADOS Y DISCUSION Los datos obtenidos nos permitirán seguir monitoreando el retroceso del glaciar Quito, los cuales fueron iniciados por Santana y Dumont (2001), quienes a través de la comparación de fotos aéreas, mapas de 1999 y observaciones de campo determinaron que la línea de costa del eje del frente del glaciar Quito ha retrocedido 270 metros y 160 en las partes laterales en 11 años (1990-2001), lo cual representa un retroceso promedio anual del glaciar de 24,5 y 14,5 m., respectivamente. Nuestras observaciones de campo del eje frente glacial muestran que el retroceso reciente ocurre sobre una plataforma de abrasión solamente visible durante las horas de marea baja, durante la cual se observan remanentes erosionados de morrenas terminales recesionales. En este sector, como consecuencia de la erosión marina la base del frente glacial es abrupto, formando acantilados de hielo con pendiente cercana a los 90°, lo cual produce avalanchas eventuales de bloques, los más pequeños son transportados por la deriva litoral, la cual deja clastos de hielo en las costas adyacentes a la base y los bloques más grandes pasan a formar parte de la flota de icebergs observados en el área durante esta época del año. Hacia el occidente, a unos 5 km., de la BCAPVM, en el sector denominado Punta Ambato, el frente del glaciar, se ubica igualmente sobre una plataforma de abrasión, construida sobre la unidad inferior de basalto definida por Santana y Dumont (2001) como una unidad que presenta morfología erosionada y cortada por filones y domos. En este sector, la plataforma de abrasión esta emergida y protegida, en algunos sectores, por domos cuyas coladas buzan en sentido contrario del oleaje, por lo tanto la acción erosiva ha dejado de funcionar y en consecuencia el retroceso reciente del glaciar puede ser trazado fácilmente a través de múltiples morrenas que se han acumulado en sus etapas de estancamiento previo al retroceso anual. Las observaciones de campo nos permitieron definir 12 morrenas de retroceso, con altitudes entre 40 centímetros y 2 metros, con un espaciamiento variable entre 16 y 33 metros y un retroceso global para los últimos 12 años de 178 metros. Lo cual permitió determinar un retroceso promedio de 14,83 entre los años 2001 y 2013. Este valores indica que aunque el retroceso del frente glacial continua este ocurre en una forma más o menos uniforme durante los últimos 23 años, lo cual en parte, soporta la hipótesis de un incremento de la tasa de deshielo de Alley y Bindschadler (2001). Nuestras observaciones de campo también nos permitieron identificar las evidencias estratigráficas de un diamicton glacial, compuesto fundamentalmente por cantos rodados, gravas y gravilla, fuertemente cementado formando una tillita. La morfometría de los sedimentos es subredondeada a subangular y la litología es fundamentalmente de basalto, no presentando la variedad de rocas ígneas intrusivas que se pueden observar en los depósitos glaciales Cuaternarios, retrabajados por el oleaje en las costas adyacentes. El diacmicton, que estudiamos esta suprayacido por un flujo basáltico, masivo de estructura columnar, por el contrario el basamento infrayacente no pudo ser observado, ya que está cubierto por taludes de derrubio y quizás se encuentre algunos metros bajo tierra. Una gran limitante que encontramos es que los estudios geológicos de la isla de Greenwich son fundamentalmente de reconocimiento, siendo el estudio más completo el de Santana y Dumont (2001), quienes presentan un mapa geológico y descripciones petrográficas de los afloramientos, pero no presentan una cronología

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
136
radiométrica para las rocas precuaternarias; en consecuencia con base a las descripciones litológicas, estos investigadores hacen correlaciones directas con las formaciones que afloran en las islas ubicadas más al norte. Estos autores definieron dos unidades litológicas, compuestas por dos coladas de lava basáltica. La unidad inferior presenta una morfología erosional, cortada por filones y domos y una unidad superior constituida por un basalto masivo con estructura columnar, en algunas áreas esta unidad presenta sedimentos volcano sedimentarios con restos de madera silicificada. Santana y Dumont, (2001), denominaron estas unidades como Basalto Puyango y Basalto Fort Williams correlacionando ambas unidades como equivalente a la Formación Coopermine de edad Cretácico tardío con dataciones radiométricas (K-Kr) de 67,6 ± 4.5 Ma. Nosotros creemos que es pertinente realizar una petrografía detallada así como una datación radiométrica (K-Ar y/o Ar-Ar) de la colada basáltica que cubre el diamicton para determinar su edad mínima (estas dataciones actualmente se realizan en Canadá). La edad máxima del diamicton, por las razones explicadas, no podrá determinarse con exactitud, sin embargo fueron muestreados cantos basálticos no meteorizados del depósito, como una aproximación de la fuente de los sedimentos y por lo tanto podremos obtener una aproximación a la edad máxima de los sedimentos y en consecuencia la edad de la glaciación del Terciario que la origino. La presente investigación es un producto de la VI expedición Científica Venezolana al Continente Antártico, la cual se efectuó del 2 al 16 de febrero del año 2013 y forma parte del Proyecto Binacional de Investigación entre la República del Ecuador y la República Bolivariana de Venezuela firmado en el año 2010. AGRADECIMIENTOS Los autores agradecen al CN. José Olmedo, director de la BAEPVM, por el invaluable apoyo logístico prestado en la realización de esta investigación. REFERENCIAS BIBLIOGRAFICAS Alley, R. y R. Bindschadler. (2001). The west Antarctic ice sheet and sea level change. American Geophysical Union, Antarctic Research series. V. 77, pp 1-11. Barron, J., Larsen, B. y J. Baldauf. (1991). Evidences for Late Eocene to Early Oligocene Antarctic glaciations and observations on Late Neogene glacial history of Antarctica: result from Leg 119. En: Barron, J., Larsen, B. y J. Baldauf. (Eds) Proceeding of the Ocean Drilling Program, Scientific Results 119. pp. 869-891 Birkenmajer, K. (2001). Mesozoic and Cenozoic stratigraphic units in parts of the South Shetland Islands and Northern Antarctic Peninsula (as used by the Polish Antarctic Programmers). Studia Geologica Polonica. 118, pp 5-188. Birkenmajer,K., Gadzicki,A., Krajewski,A., Solecki, A., Tatur, A., y H, Yoon. (2005). First Cenozoic glaciers in West Antarctica. Polish Polar Research. Vol. 26, N°1, pp. 3-12 Thompson, M. (2004). Geological and Paleoenvironmental history of the Scotia Sea region as a base for biological interpretation. Deep-Sea Research II, 51, PP 14671487. Troedson, A., y J. Riding. (2002). Upper Oligocene to lowermost Miocene strata of King George Island, South Shetland Islands, Antarctica stratigraphic of the Antarctica Peninsula. Journal of Sedimentary Research. 72, pp. 510-523 Troedson, A., y J, Smellie. (2002). The Polonez Cove Formation of King George Island, Antarctica: stratigraphy, facies and implications for mid-Cenozoic cryosphere development. Sedimentology. 49, pp. 277-301. Santana, E. y J. Dumont. (2001). Geología de los alrededores de la Estación Ecuatoriana Pedro Vicente Maldonado (Isla Greenwich) e isla Dee, Península Antártica. Acta Antártica Ecuatoriana, PROANTEC, Ecuador Vol. 1. 25 págs. Zachos, J., Pagani, M., Thomas, E. y K, Billups. (2001). Trends, rhythms, and aberrations in global climate 65 Ma to present. Science, 292. Pp 686-693.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
137
ESTADO TÉRMICO Y ESPESOR DE CAPA ACTIVA EN EL COLLADO IRIZAR (ISLA DECEPCIÓN, ANTÁRTIDA): PERIODO 2009-2013.
(Thermal state and active layer thickness in Collado Irizar, Deception Island, Antarctica: 2009-2013 period)
Goyanes, G.1, Vieira, G.2 y Caselli, A.3
1Instituto de Estudios Andinos “Don Pablo Groeber” (UBA-CONICET), Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Intendente Güiraldes 2160 Pabellón II-Ciudad Universitaria, Buenos Aires, Argentina. [email protected] 2Instituto de Geografia e Ordenamento do Territorio, Centro de Estudos Geográficos, Universidade de Lisboa, Avenida Prof. Gama Pinto, Lisboa, Portugal. E-mail: [email protected] 3Universidad Nacional de Río Negro, 25 de Mayo 880-4º Piso, General Roca, Argentina. E-mail: [email protected] INTRODUCTION The Antarctic Peninsula is one of the regions in the World with a stronger climate warming trend, with an increase of ca. +2.5ºC in the mean annual air temperatures (MAAT) since 1950. While glaciers and ice-shelves are being monitored to evaluate the effects of climate change, permafrost, an important component of the Antarctic cryosphere has been largely neglected and only recently started to be monitored systematically (Vieira et al. 2010a, Bockheim et al. 2013). Permafrost is present beneath virtually all ice-free terrain, except at the lowest elevations of the maritime Antarctic and sub-Antarctic islands (Ramos et al. 2012). In addition, permafrost occurrence has been confirmed in the South Shetlands through both periglacial landform analysis, exposures, geophysics and boreholes. Climate at the South Shetland is cold-oceanic at sea level, with frequent summer rainfalls and a moderate annual temperature range. Mean annual air temperatures are close to -2 ºC at sea level and average relative humidity is very high (80-90%, Ramos et al. 2012). Deception Island (62º57’S; 60º38’W) is the south-westernmost island of the Bransfield Strait. The island represents the top of a complex volcanic structure characterized by a horseshoe-like shape with an opening in the south-east sector (Neptune Bellows) and a flooded caldera (Port Foster). The volcanic activity on the island developed in the last 0.2 Ma and the latest eruptions occurred in 1967, 1969 and 1970. The island has a distinct annual cycle of ground freezing (from autumn to mid-spring) and thawing (austral summer) (Ramos et al. 2012). The aim of this work is showing the first results about the thermal regime and the active layer thickness (ALT) evolution from the summer of 2008-2009 to the summer of 2012-2013 at Irizar CALM-S site in Deception Island. This site is part of The Global Terrestrial Network for Permafrost (GTN-P) and of the Circumpolar Active Layer Monitoring (CALM), initiated by the International Permafrost Association (IPA) to organize and manage a global network of permafrost observatories for detecting, monitoring and predicting climate change, and to monitor changes in active layer thickness and temperature. METHODOLOGY Surface boreholes down to 0.8 m depth for year-round temperature monitoring were installed in January 2009 in the Irizar CALM-S site (Vieira et al. 2010b). Ground temperatures are monitored at hourly intervals at 0.05, 0.10, 0.15, 0.20, 0.40 and 0.80 m depths using Hobo Pendant miniature loggers. I-button data loggers were installed just below the soil surface, at 0.02 m depth, close to the boreholes in order to measure ground surface temperature. I-buttons were inserted in 15x15x0.4 cm high diffusivity aluminium plates in order to better approach average ground temperatures.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
138
An air temperature Tinytag datalogger was installed 1.5 m above the ground in a standard radiation shelter next to BH2 to analyse the effect of meteorological factors on subsurface temperature at the borehole sites. The snowpack dynamics is characterized using temperature poles with i-buttons at different heights on wooden stakes (Lewkowicz 2008). The results provide snow thickness estimation but they are seemingly underestimated and they should be considered preliminary and only indicative. In addition ALT has been monitored (by mechanical probing following the CALM protocol – Brown et al. 2000) into a 100x100 grid (nodes every 10 m) near to the end of thaw season. It should be noted that the measurements were not made on the same days from one year to another. The site ground is composed by sandy-gravelly, poorly stratified pyroclasts. In this work, we analyse borehole, air, surface and ground temperatures, snowpack data and ALT from January 2009 to early January 2013. RESULTS AND DISCUSSION MAAT shows a slightly increased from -3.6 ºC to -3.2 ºC between 2009 and 2012 (Table 1). Maximum and minimum values of mean monthly air temperature (MMAT) have been achieved in 2011, reaching 6.4 ºC and -9.2 ºC respectively.
2009 2010 2011 2012 MAAT (ºC) -3.6 -2.3 -3.1 -3.2
Min. MMAT (ºC) -7.9 -5.2 -9.2 -6.8 Max. MMAT (ºC) 0.9 -0.2 6.4 1.9
Table 1: Statistics of the air temperature data measured from 2009 to 2012. Tabla 1: Estadísticas de la temperatura del aire medida desde el 2009 hasta el 2012.
Mean annual ground surface temperature (MAGST) in the site was always warmer than air temperature (Fig. 1) reaching -1.4 ºC in 2010, -2.9 ºC in 2011 and -2 ºC in 2012. The time series of monthly mean air and ground temperatures show winter decoupling of air and ground temperatures at this site. The magnitude of this separation is consistently larger during the winter than during the summer, except during 2011 winter where both temperatures were approximately equivalent. Snow season started in May in 2009 and 2010 (Fig. 2), reaching a stable-significant snow cover from July to November. During 2011 snow cover was more irregular and the maximum thickness was only reached in October-November. In 2012 snow cover only stabilized in September and snow melt occurred in early December. Borehole temperature regimes (Fig. 2) show that the thaw season took place from November to March with small interannual differences, while the freezing season took place from April to October. The freezing season reflects the control of snow cover on the ground temperature. Scarce snow in early winter in 2009 and 2011 together with the lower atmospheric temperatures were responsible for ground cooling events reaching -10 ºC at 0.6 m depth. On the contrary, the thicker and permanent snowpack during the winter-spring season in 2010 and 2012 prevented ground cooling and reduced spring warming effects, delaying the beginning of the thaw season. During the summers ground temperatures were 3 to 6 ºC and thaw reached 0.6 m. The borehole bottom (0.6-0.8 m) was frozen all year-round, showing the presence of the permafrost table. Mean average ALT in the CALM-S site was 0.67, 0.46, 0.55 and 0.65 m in 2009, 2010, 2011 and 2012 (Fig. 1), respectively, with a standard deviation of 0.06, 0.063, 0.06 and 0.043 m respectively. It should be noted that 2010 measurement was made one month earlier than the other years. Spatial distribution of thaw thickness inside the CALM-S site (Fig. 3) shows a similar pattern every summer (except in 2012), showing the lowest thickness at the northeastern, northwestern and southwestern sectors of the grid. The northwestern and northeastern sectors of the grid correspond to the lowest altitudes, with a concave shape, while the southeastern sector is the highest. Average thaw

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
139
thickness in 2009 was thicker (0.67 m) than on consecutive years and unfrozen ground inside the CALM-S grid was more extensive. A similar situation occurred in 2012, when average thaw thickness reached 0.65 m, but the ALT spatial distribution was more regular. The interannual and intrasite ALT evolution is controlled by snow cover. Thicker and longer lasting snow cover during the spring of 2010 delayed the beginning of the active layer thaw. On the contrary, in 2011, spring thaw of the active layer started earlier due to the absence of snow. Differences on thaw season onset influenced the maximum thickness of active layer and this is the main reason for shallower ALT in the summer of 2011 than in 2012. Besides, concave shapes on the CALM-S´ surface are sites with greater snow thickness that may persist during the summer, delaying active layer thaw. In early January 2011, extensive areas of Deception Island were still covered by snow, while they were snow free in January 2012.
Figure 1: (Left) Mean monthly air and ground temperature evolution from January 2009 to December 2012; (Right) ALT
annual evolutions from 2009 to 2012. Figura 1: (Izq.) Evolución de la temperatura media mensual del aire y del suelo desde Enero del 2009 hasta Diciembre del
2012; (Der.) Evolución anual del ALT desde el 2009 hasta el 2012. CONCLUSION The active layer thermal regime confirms top of permafrost occurrence at 0.6-0.8 m depth with an average temperature close to 0 ºC. Besides, borehole temperatures showed a good correlation with air temperatures when a thin snow cover occurred. The evolution of the active layer in the Irizar CALM-S site from 2009 to 2012 is mainly controlled by snow cover variability, which inside the grid is also dependent on microscale characteristics, such as slope and local topography. ALT and air temperature inter-annual variability and the short period with data do not allow for the identification of significant trends. Therefore, it is necessary to continue the monitoring in order to identify the presence of trends or pattern in order to understand their behaviour following climate change.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
140
Figure 2: Daily regime for air and ground temperatures and snow depth in the Irizar CALM-S site from 2009 to 2012.
Diagonal lines in the borehole thermal regime represent the areas with temperatures between -0.5 and 0.5 ºC. Figura 2: Régimen diario de temperaturas del aire y del suelo y, espesor de nieve en el sitio CALM-S Irizar, desde el 2009 hasta el 2012. Las líneas diagonales en el régimen térmico de la perforación representan las zonas que se encuentran con
temperaturas entre -0.5 y 0.5 º C.
Figure 3: Spatial distribution of ALT inside the CALM-S Irizar from 2009 to 2012.
Figura 3: Distribución espacial del ALT dentro del CALM-S Irizar, desde el 2009 hasta el 2012. ACKNOWLEDGEMENTS PERMANTAR-2, PROPOLAR (Portugal), Dirección Nacional del Antártico (Argentina). REFERENCES Bockheim, J., Vieira, G., Ramos, M., López-Martínez, J., Serrano, E., Guglielmin, M., Wilhelm, K. y Nieuwendam, A. 2013. Climate warming and permafrost dynamics in the Antarctic Peninsula region. Global and Planetary Change, 100, 215-223. Brown, J., Nelson, F.E., y Hilken, K.M. 2000. The Circumpolar Active Layer Monitoring (CALM) program research designs and initial results. Polar Geography, 3: 165-258. Lewkowicz, A. 2008. Evaluation of Miniature Temperature-loggers to Monitor Snowpack Evolution at Mountain Permafrost Sites, Northwestern Canada. Permafrost and Periglacial Processes, 19(3):323-331. Ramos, M., de Pablo, M.A., Sebastian, E., Armiens, C., Gómez-Elvira, J., 2012. Temperature gradient distribution in permafrost active layer, using a prototype of the ground temperature sensor (REMS-MSL) on Deception Island (Antarctica). Cold Regions Science and Technology, 72:23-32. Vieira, G., Bockheim, J., Guglielmin, M., Balks, M., Abramov, A., Boelhouwers, J., Cannone, N., Ganzert, L., Gilichinsky, Goryachkin, D., López-Martınez, D., Meiklejohn, J., Raffi, R., Ramos, M., Schaefer, C., Serrano, S., Simas, F., Sletten, R. y Wagner, D. 2010a. Thermal State of Permafrost and Active-layer Monitoring in the Antarctic: Advances During the International Polar Year 2007-2009. Permafrost and Periglacial Processes, 21:182-197. Vieira, G., Neves, M., Caselli, A., Ramos, M., Batista, V. y Bernardo, I. 2010b. A new CALM-S site at Irizar, Deception Island (Antarctic Peninsula region). First results. International Polar Year Oslo Open Science Conference, 8-12 June.
ESTACIONALIDAD EN LOS DATOS DE GRAVIMETRÍA Y ALTIMETRÍA RADAR EN LA ANTÁRTICA Y VARIABILIDAD CLIMÁTICA ASOCIADA. APLICACIÓN A LA PENÍNSULA

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
141
ANTÁRTICA. PRIMERA PARTE: RESULTADOS GRAVIMETRÍA SATELITAL (Seasonality in gravimetry and radar altimetry data in the Antarctic and associated climate variability.
Application to the Antarctic Peninsula. Part I: Satellite gravimetry results) Carmona, N. A 1, Abarca-del-Rio, R. 1, Remy. F. 2
1Departamento de Geofísica, Universidad de Concepción, Casilla 160-C, Concepción, Chile. [email protected], [email protected] 2LEGOS-CNES, 18 Av Ed Belin, 31400, Toulouse, Francia
El desarrollo de la teledetección satelital ha cumplido un rol fundamental en muchos temas afines a las geociencias y muy particularmente en lo que respeta la identificación de variables relacionadas con la variabilidad climática. Una de las novedosas utilizaciones que este avance ha permitido es el estudio de la variabilidad nival por medio de la altimetría de radar (RA). En los años 90, se lanzaron 2 satélites ERS1 y ERS2, presentando como similitud que ambos poseían una orbita de 35 días al igual que ENVISAT, primer satélite de 2 frecuencias (bandas S y Ku) lanzado en los años 2000. Nuestra propuesta se centra en esencialmente analizar la variabilidad de las señales de ERS-1 y ERS-2, seguidas por ENVISAT es decir sobre una duración de 22 años. En este trabajo extenderemos temporalmente el análisis de Horwath et al., (2012), Octubre 2002- Agosto 2009, realizado en el equipo del criología satelital del Centro Nacional de Estudios Espaciales (CNES), Francés. Caracterizaremos los diferentes modos temporales-espaciales de la cobertura altimétrica, identificando su asociación con la cobertura nival simulada por modelos de reanálisis atmosférico. Reconoceremos los diferentes modos de variabilidad climática y los asociaremos con firmas climáticas tropicales o de altas latitudes. Sin embargo, de manera de poder tener primero una visión más coherente de la variabilidad global de la capa de nieve y de la variabilidad climática en la Antártica trataremos inicialmente los datos de la misión gravimétrica GRACE (sitio web) que se extienden desde el año 2003 hasta nuestros días (2002-2013), sumado al estudio en paralelo de los reanálisis atmosféricos sobre la misma época. La finalidad de este análisis previo es de no solamente comprender los ciclos presentes en los datos gravimétricos asociados al cambio de masa, sino también, comprender como la variabilidad climática asociada a los grandes fenómenos como: El Niño - Oscilación Austral (ENOS), la oscilación Antártica (AAO) y la Oscilación Decenal Del Pacifico (PDO), y cambio climático influencian la variabilidad antártica de las masas de hielo. Cabe mencionar que la hipótesis de trabajo se centra en el análisis de estudios, que muestran que la variabilidad nival de la Antártica (Península Antártica) presenta una variabilidad estacional e interanual. Esta última esta asociada con la principales modos de variabilidad climática en la región, especialmente el Modo anular del Sur (SAM) asociado a la oscilación Antarctica (AAO) y el fenómeno de El Niño- Oscilación del Sur (ENOS) [Boening, Lebsock, Landerer, Stephens, 2012]. Así como también, la temperatura presenta una gran variabilidad interanual con una fuerte influencia del ENOS [Bromwich et al. 2000, Kwok and Comiso 2002, Bromwich et al. 2004], siendo el motivo por lo cual estas señales deben verse presentes en los datos altimétricos ERS-1, ERS-2, ENVISAT. Hasta el momento tales señales no han sido caracterizadas correctamente en esa base de datos. Sin embargo si han sido vistos en bases de datos satelitales complementarias, como son los datos GRACE [Sasgen et al., 2010]. Además de mostrar variaciones interanuales significativas más allá de una tendencia lineal o cuadrática. Un ejemplo es una anomalía positiva que se inició en el segundo semestre del año 2005 [Horwath and Dietrich, 2009; Velicogna 2009]. Parte de esta variabilidad interanual se ha relacionado con el balance superficial de masa [Sasgen et al., 2010]. En esta instancia se mostraran los objetivos, la metodología y los resultados del trabajo previo (análisis

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
142
de datos gravimétricos comparados con modelos de reanálisis atmosféricos sobre el periodo que cubre 2003-2012). Se trabajo con 2 tipos de datos: Datos de reanálisis atmosféricos y datos gravimétricos. En lo que respeta los datos atmosféricos se construyo una climatología de la zona, con los datos de reanálisis ERA – Interim, Por climatología se entiende la marcha común mes a mes durante un año usual. Este año usual consiste en construir medias (sobre los 10 años estudiados) para cada uno de los 12 meses del año. Esta nueva base de datos (ERA – Interim) posee mejoras en cuanto a calidad, al implementar nuevos modelos de procesos atmosféricos, que se traducen notablemente por el aumento de número de niveles de presión en altura (desde 23 en ERA-40 hasta 37 en ERA-Interim). Posee además una mejor resolución espacial de 0.7°. Esta resolución permitirá compararlos directamente con los datos GRACE (resolución 1°) así como efectuar la validación correspondiente con datos meteorológicos locales. Los datos gravimétricos que son analizados cubren el período 2003-2012. Esta misión puesta en marcha en el año 2002 [Schmidt et al., 2008], es capaz de detectar los cambios temporales del campo gravitatorio de la Tierra a través de sus efectos orbitales medidos de manera diferencial por un sistema de microondas que van entre los dos satélites en orbita. Cambios en el campo de gravedad temporal pueden entonces ser traducidos en dependencias geográficas equivalente al cambio de masa superficial [Wahr et al., 1998], expresadas en equivalente de agua o altura de hielo equivalente. De esta base de datos se extrae la zona Antártica (-60 a -90°N y 0 a 360° E). Sin embargo es necesario de saber que existen por lo menos 3 bases de datos oficiales, por lo cual primero es necesario realizar una inter- comparación entre las diferentes soluciones. Una vez esto realizado y la base de datos escogida como en el caso de los datos atmosféricos se realizan una climatología de la variabilidad. Sin embargo se compararan tanto las climatologías de los datos atmosféricos y los datos gravimétricos como la variabilidad interanual y de tendencia. Permitiéndonos así visualizar y en tanto estimar con claridad las variaciones presentes en una cierta cantidad de puntos. Notablemente alrededor de la península Antártica, la cual es prioridad en la segunda parte, altimetría radar, de este trabajo. En este presentación se presentará primero entonces la climatología de cada una de las variables atmosféricas analizadas, es decir de 3 variables atmosféricas en superficie, las cuales son: Temperatura, Precipitación al igual que Humedad (Precipitación – Evaporación), además de la variación de masa local tal cual detectada por el sensor gravimétrico. Es decir se presentaran las variaciones mes a mes de cada una de las variables de manera a poder explicar la variabilidad vista en los datos gravimétricos. Se presenta la estacionalidad (ciclo anual y semi-anual) en cada una de las variables. La estacionalidad sirve para comprender su influencia en la tendencia a largo tiempo, es decir estaciones y lugares donde hay aumento y disminución de la tasa de hielo sobre la Antártica. Queremos responder a la siguiente pregunta ¿En que estaciones del año las tendencias son mas marcadas? ¿Y de donde provienen estas tendencias? Es decir : ¿que variables climáticas están asociadas o influencian estas tendencias? En lo que sigue presentamos solo algunas imágenes de nuestros avances.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
143
Figura 1: Climatología 2003-2012, mes de mayo. Basada en datos ERA-interim y GRACE. Las variables son las siguientes: a) temperatura, b) precipitación, c) humedad superficial, por parte de los datos de reanálisis, d) espesor equivalente de agua
líquida por la misión GRACE. Figure 2: Climatology 2003-2012, May. Based on ERA-interim data and GRACE. The variables are the following: a)
temperature, b) precipitation, c) surface moisture by reanalysis data, d) liquid water equivalent thickness by the GRACE mission.
Por medio de la fig.1 es posible visualizar el comportamiento de 3 variables atmosféricas en superficie, las cuales son: a Temperatura, expresada en [°C], b Precipitación , expresada en [mm/day] al igual que c Humedad (Precipitación – Evaporación). En donde cabe destacar que en ellas las líneas negras representan los contornos de los valores de las variables, en el caso de temperatura es c/ 5°C y en las otras c/ 1 [mm/day] así como la línea de costa descargada de m_map. AGRADECIMIENTOS: Departamento de Geofisica (DGEO) de la Universidad de Concepción, por beca que permite esta

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
144
presentación. Centro Nacional de Estudios Espaciales (CNES), Francia por el acceso a diferentes bases de datos satélites. REFERENCIAS BIBLIOGRÁFICAS Boening, C.; Lebsock, M.; Landerer, F.; Stephens, G. Snowfall -driven mass change on the East Antarctic ice sheet. 2012. Geophys. Res. Letts., 39, L21501. Bromwich, D.; Rogers, A.; Kallberg, P.; Cullather, R.; White, J.; Kreutz, K. 2000. ECMWF analyses and reanalyses depiction of ENSO signal in Antarcticg precipitation. J. Climate, 13, 1406-1420. Bromwich, D.; Monaghan, A.; Guo, Z. 2004. Modeling the ENSO modulation of Antarctic climate in the late 1990s with Polar MM5. J. Climate , 17, 109-132. Horwarth, M.; Dietrich, R. 2009. Signal and error in mass change interferences from GRACE: the case of Antarctica. Geophys.J.Int., 177, 849 -864. Kwok, R., and J.C. Comiso. 2002. Southern Ocean climate and sea ice anomalies associated with the Southern Oscillation. J. Climate, 15, 487-501. Sasgen, I.; Dobslaw, H.;Martinec, Z.; Thomas, M. 2010. Satellite gravimetry observation of Antarctic snow accumulation related to ENSO. GRACE Earth and Planetary Science Letters 299 , 352–358. Schmidt, R.; Flechtner, F.; Meyer, U.; Neumayer, K-H.; Dahle, CH.; Konig, R.; Kusche, J. 2008. J.Hydrological Signals Observed by the GRACE Satellites. Surv Geophys, 29, 319:334. Velicogna, I. 2009. Increasing rates of ice mass loss from the Greenland and Antarctic ice sheets revealed by GRACE. Geophys. Res. Letts., 36, L19503. Wahr, J.; Molenaar, M.; y Bryan, F. 1998. Time variability of the Earth's gravity field: Hydrological and oceanic effects and their possible detection using GRACE. J. Geophys. Res. 103, 30 205–30 229.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
145
¿DIFIEREN LOS PATRONES DE CICLAJE DE P ENTRE LAS ÁREAS LIBRES DE HIELO Y LAS FRONTERAS DE LOS GLACIARES EN LA REGIÓN DE LA
MARÍTIMA ANTÁRTICA? (Do P cycling patterns differ between ice-free areas and glacial boundaries in the Maritime Antarctic
region?)
Chacón, N.*1,3, Ascanio M.1, Herrera, R.2, Benzo, D.1 , Flores, S.1 , Silva, S.J.3 y García, B.1
1Centro De Ecología, Instituto Venezolano de Investigaciones Científicas, Apdo. 20632. Caracas 1020-A, Venezuela. [email protected] 2Grupo de Evaluación y Restauración de Sistemas Agrícolas y Forestales, Universidad de Córdoba, España. 3Centro de Oeanología y Estudios Antárticos, Instituto Venezolano de Investigaciones Científicas, Apdo.20632. Caracas 1020-A, Venezuela.
INTRODUCCIÓN En la Marítima Antártica la expansión de las áreas libres de hielo debido al retroceso de los glaciares, es un resultado del calentamiento climático en esa región (Ugolini y Bockheim 2008). Durante el verano, estas áreas experimentan mayor temperatura que las zonas cercanas a los glaciares, lo cual les permite la formación de agua líquida (Chen y Blume 1999, 2000). Los procesos biogeoquímicos de los ecosistemas terrestres en la Antártica son principalmente influenciados por factores abióticos tales como la temperatura y disponibilidad de agua (Barret y col. 2006, 2009). Este estudio publicado recientemente (Chacón y col 2013) fue diseñado para determinar si el ciclaje biogeoquímico de fósforo (P) en el suelo difiere entre zonas adyacentes a los glaciares y áreas libres de hielo. Para ello, se estableció un sitio en la frontera del glacial Collins y otro en un área libre de hielo alrededor del Lago Uruguay. Ambos sitios están localizados en la Península de Fildes
MATERIALES Y MÉTODOS Área de estudio y colección de muestras El sitio de estudio estuvo localizado en las inmediaciones de la Base Científica Antártica Artigas (Base Cientıfica Antártica Artigas [BCAA]) en la Península de Fildes de la Isla Rey Jorge, la cual forma parte de las Islas Shetland del Sur, en la Marítima Antártica (61º54'-62º16S; Figura 1). Las muestras de suelo fueron colectadas de dos sitios en las cercanías de BCAA: uno en las fronteras del glaciar Collins (BG: 62º10'49.62“S, 58º54'34.75”W) y el otro en una área libre de hielo (IFA: 62º11'08.33”S, 58º54'33.02”W) cerca del Lago Uruguay. En cada sitio, se muestreó el horizonte mineral superficial (0-10 cm) en 15 puntos seleccionados de acuerdo a un diseño completamente aleatorio (n total = 30). Durante el muestreo, las temperaturas de campo registradas en los lagos adyacentes a los sitios de BG e IFA fueron 2,4ºC y 5,3 ºC, respectivamente.
Análisis químico Las fracciones de P en los suelos fueron obtenidas de acuerdo al método de Hedley y col. (1982), modificado por Tiessen y Moir (1993) y determinadas colorimetricamente por medio del método del molibdato-ácido ascórbico (Murphy y Riley 1962). Por su parte, las fracciones hierro (Fe) y aluminio (Al) fueron obtenidas a través de un método de disolución selectivo secuencial usado para estimar las formas de Fe y Al orgánicamente enlazadas, no cristalinas y cristalinas (Wieder y Lang 1986). Las concentraciones de ambos elementos en cada extracto fueron determinadas utilizando espectroscopia de absorción atómica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
146
Figura 1. Mapa de la Isla Rey Jorge mostrando la localización relativa de los sitios de estudio. BCCA = Base Científica Antártica Artigas, BG = frontera del glacial, IFA = área libre de hielo. Imagen digitalizada de la Carta 1111: Base Científica Antártica Artigas. Servicio de Oceanografía, Hidrografía y Meteorología de la Marina de Uruguay, 1991. Figure 1. Map of King George Island showing the relative locations of the studied sites. BCAA= Base Científica Antártica Artigas, BG = boundary of the glacier, IFA = ice-free area. Digitalized image from the Letter 1111: Artigas Antarctic Scientific Base. Service of Oceanography, Hydrography and Meteorology of the Navy of Uruguay 1991.
Análisis mineralógico La mineralogía del suelo fue obtenida a través de difracción de rayos X (DRX) utilizando radiación Cu-Kα. Cuatro muestras representativas de suelo de cada sitio de estudio fueron pulverizadas y prensadas en pastillas para el análisis. Las muestras fueron montadas sobre placas de vidrio y analizadas a partir de 5º hasta 90º 2θ con una velocidad de 2º 2θ por minuto. Análisis biológico y bioquímico Alícuotas de suelo fresco después de una incubación a 37º C fueron usadas para determinar la hidrólisis del flurescein diacetato (FD) y la actividad de la fosfatasa alcalina (FA), siguiendo los procedimientos de Schnürer y Rosswall (1982) y Tabatabai y Bremner (1969), respectivamente. En este estudio, la actividad de la FA fue medida como un índice de mineralización de P orgánico (Dick y Tabatabai 1993), mientras que la hidrólisis del FD fue usado como una medida de la actividad microbiana del suelo (Schnürer y Rosswall 1982; Green y col. 2006). Análisis estadístico Un análisis de varianza de una vía (ANOVA) fue usado para comparar las fracciones de P en el suelo, las formas de Fe y Al y las actividades microbiana y bioquímica. El test no paramétrico de Mann Whitney fue usado cuando los datos no cumplieron los supuestos para el ANOVA.
RESULTADOS Los resultados mostraron que el suelo del IFA tiene un mayor contenido de P adsorbido sobre los oxihidróxidos de Fe/Al (NaOH-Pi) y formas ocluidas (HCl (conc)-Pi) que el suelo del BG. Una tendencia opuesta fue observada para el P asociado al calcio (HCl(dil)-Pi ) (Tabla 1). En concordancia con esta distribución de P, el suelo de esta área mostró los más altos niveles de oxihidróxidos de Al (BG: 29 ± 3, IFA: 157 ± 12) y la presencia de minerales secundarios (hematita y el intergrado clorita-vermiculita-montmorillonita). Por su parte, la actividad de la FA y la hidrólisis del FD fueron también favorecidas en el IFA (Tabla 2).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
147
Fracción BG IFA
NaOH-Pi 2,96 ± 0,43b 24,30 ± 12,91a
HCl(dil)-Pi 385,89 ± 9.21a 281,75 ± 16,72b
HCl (conc)-Pi 26,80 ± 1,97b 39,15 ± 1,68a
Tabla 1 Valores de varias fracciones de P (mg kg-1) en los sitios de estudio. Valores medios ± 1ES seguidos por letras diferentes en las filas denotan diferencias significativas entre sitios. BG = frontera del glacial, IFA = área libre de hielo.
Table 1 Values of various fractions of P (mg kg-1) in the study sites. Mean values ± 1ES followed by different letters in rows denote significant differences between sites. BG = glacial border, IFA = ice-free area.
Propiedades del suelo Unidades BG IFA
AP µg p-NP g-1 soil h-1 2.92 ± 0.95b 13.67 ± 3.18a
FD µg fluorescein g-1 soil h-1 18.09 ± 2.06b 49.67 ± 16.87a
Tabla 2. Actividad de la fosfatasa alcalina (FA) e hidrólisis del fluorescein diacetato en los sitios de estudio. Valores medios ± 1ES seguidos por letras diferentes en las filas denotan diferencias significativas entre sitios. BG = frontera del glacial, IFA =
área libre de hielo. Table 2. Alkaline phosphatase activity (FA) and hydrolysis of fluorescein diacetate in the study sites. Mean values ± 1ES followed by different letters in rows denote significant differences between sites. BG = glacial border, IFA = ice-free area
DISCUSIÓN Y CONCLUSIÓN Los cambios en los reservorios de P inorgánico, el incremento en minerales secundarios y la mayor actividad de la FA e hidrólisis del FD en el suelo de la IFA, nos permite indicar que la disponibilidad de agua líquida en estos suelos permite la co-ocurrencia de una avanzada meteorización química y una mayor actividad biológica en este sitio. Ambos procesos tienen una fuerte influencia sobre el ciclaje biogeoquímico de P en las áreas libres de hielo. En un escenario de calentamiento climático sobre la Península Antártica, esta información puede contribuir a la comprensión de cómo el retroceso de los glaciares y consecuentemente el avance de las áreas libres de hielo puede estar alterando el ciclaje biogeoquímico de nutrientes esenciales en esta región. AGRADECIMIENTOS Programa Antártico Venezolano (Ministerio del Poder Popular para Ciencia, Tecnología e Innovación (MCTI)).
REFERENCIAS Barret, J.E., Virginia, R.A., Wall, D.H., Cary, S.C., Adams, B.J., Hacker, A.L. y Aislabie, J.M. 2006. Co-variation in soil biodiversity and biogeochemistry in northern and southern Victoria land, Antarctica. Antarctic Science, 18: 535-548. Barret, J.E., Gooseff, M.N. y Takacs-Vesbach, C. 2009. Spatial variation in soil active-layer geochemistry across hydrologic margins in polar desert ecosystems. Hydrology and Earth System Science, 13: 2358-2358 Chacón, N., Ascanio, M., Herrera, R., Benzo, D., Flores, S., Silva, S.J. y García B. 2013. Do P cyling patterns differ between ice-free areas and glacial boundaries in the Maritime Antarctic region?. Arctic, Antarctic and Alpine Research 45:190-200 Chen, B.J., y Blume, H.P. 1999. Study on the dynamics of soil moisture in an ice-free area of the Fildes Peninsula, King Geroge Island, the maritime Antarctica. Polarforschung, 66: 11-18. B. J. Chen y Blume, H.P. 2000. Element enrichment and migration within some soils on Fildes Peninsula (King George Island), the maritime Antarctica. Journal of Plant Nutrition and Soil Science, 163: 291-297. Dick, W.A. y Tabatabai, M.A.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
148
1993. Significance and potencial uses of soil enzymes. In Blaine, M.F. (ed). Soil Microbial Ecology, Applications in Agriculture and Environmental Management. Marcel Decker, New York, 95-127. Green, V.S., Stott, D.E. y Diack, M. 2006. Assay for fluorescein diacetate hydrolytic activity: optimization for soil samples. Soil Biology and Biochemistry, 38: 693-701. Hedley, M.J., Stewart, J.W.B. y Chauhan, B.S. 1982. Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Science Society of America Journal, 41: 970-976. Murphy, J. y Riley, J.P. 1962. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta, 27: 31-36. J. Schnürer y Rosswall, T. 1982. Fluorescein diacetate hydrolysis as measure of total microbial activity in soil and litter. Applied Environmental Microbiology, 43: 1256-1261. Tabatabai, M.A., y Bremner, J.M. 1969. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biology and Biochemistry, 1: 301-307. Tiessen, H. y Moir. J.O. 1993. Characterization of available P by sequential extraction. In Carter, M.R. (ed.), Soil sampling and methods of analysis. Special publication of the Canadian Society of Soil Science, Lewis, Boca Raton, 75-86. Ugolini, F.C. y Bockheim, J.G. 2008. Antarctic soils and soil formation in a changing environment: a review. Geoderma, 144: 1-8.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
149
ESTUDIO DE LAS PROPIEDADES DE LA NIEVE MEDIANTE DATOS SATELITALES EN GLACIAR UNION, HERITAGE RANGE, WEST ANTARCTICA
(Snow property study based on remote sensing data at Union Glacier, Heritage Range, West Antarctica)
Jaña, R.1, 2, Arigony-Neto, J.3, Medeiros, C.3, Simões, J.C.4, Borges-Fernandes, G.5, Vieira, R.5, Espinoza, J.M.A.3
1 Instituto Antártico Chileno, Plaza Gobernador Benjamín Muñoz Gamero 1055, CP 6200965, Punta Arenas, Chile, E-mail:
[email protected] 2 Fundación CEQUA, 21 de Mayo 1690, Punta Arenas, Chile 3 Laboratório de Monitoramento da Criosfera - LaCrio, Universidade Federal do Rio Grande – FURG, Instituto Nacional
Ciência e Tecnologia da Criosfera, Av. Itália, km 896201-900 - Rio Grande, RS - Brazil 4 Centro Polar e Climático Instituto de Geociências, Universidade Federal do Rio Grande do Sul (UFRGS), Instituto
Nacional de Ciência e Tecnologia da Criosfera, Av. Bento Gonçalves, 9500 Porto Alegre, RS - Brasil 5 Laboratório de Processos Sedimentares e Ambientais – Depto. de Geografia, Universidade Federal Fluminense, Instituto
Nacional de Ciência e Tecnologia da Criosfera, Rua José do Patrocínio 71, 28010-385, Campos dos Goytacazes, RJ, Brasil INTRODUCTION Several sites (Figure 1) in the area of Union Glacier (79°45.666’ S / 83º15.895’ W) are being studied by a joint Brazilian–Chilean program since December 2011, under the activities of the project “Clima de América del Sur y Antártica (CASA): buscando teleconecciones por medio de registros de testigos de hielo”. The area located at the eastern sector of Heritage Range in the Ellsworth Mountains, West Antarctica, is characterized by the feeding of several tributary glaciers that descend from surrounding Edson Hills, Buchanan Hills, Collier Hills and Enterprise Hills, to a major ice-shed flowing to Constellation Inlet at the base of Ronne-Filchner Ice Shelves system. At the head of Union Glacier the presence of an appropriate ice blue sector allows the regular landing of wheeled airplanes in a runway operated by ALE, a private tourism company, since 2007 and recently by Chilean Air Force (2012/2013). There, previous work has been carried out to measure mass balance by traditional methods and ice thickness by different RADAR instruments (Rivera et al., 2010). Consequently, using the opportunity to extend the record, cooperative work was established and the measuring of stakes along a cross section of the main glacier tongue was performed in 2011 (see Figure 1, grey color circles) to contribute with the running mass balance program (Rivera et al., 2012). Our research considers as a main goal the use of multi-sensor satellite data to extract information about glaciological parameters, snow properties and glacier dynamic in this area. For these purposes, different surveys across the area were designed to record ground truthing data to calibrate methods developed to extract features and parameters of mass balance, snow properties and ice dynamic. MATERIALS AND METHODS During the fieldwork carried out on December 2011 and January 2012, 07 Cosmo-SkyMed images were acquired with the Stripmap Image mode (3m spatial resolution, VV polarization). During that period of time a number of activities were carried out for ground truthing. These includes the surveying of geodetic datasets in selected surrounding features of spots to produce a large-scale absolute digital elevation model derived from ASTER satellite data and the surveying for mass balance and ice dynamic assessment based on dGPS and traditional measurements at Cryosphere glacier test site. To calibrate the methods for modeling the X-band backscattering of the snowpack on Union Glacier, we dug 07 snow pits with 2 meters deep and recorded the information as snow density and temperature each 10 cm,

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
150
limits and hardness of the snow layers, and size and shape of the snow grains. The snow pits were dug in sites on the glacier with different characteristics of the backscattering (Figure 2).
Figure 1. Map of Union Glacier area depicting location of ALE Camp (tent symbol), GPS reference station (square with dot), ground control points (triangles), snow pits (gray squares), ice dynamic/mass balance stakes on tributary Criosfera
Glacier (light color circles) and CECs’ ice dynamic/mass balance stakes on Union Glacier (grey color circles).
Figura 1. Mapa de Union Glacier mostrando ubicación de campamento ALE (símbolo tienda), estación base GPS (cuadrado con punto), puntos de control (triángulos), pozos nieve (cuadrados grises), balizas dinámica/balance de masa del hielo en glaciar Criosfera ( círculos color claro) y balizas dinámica/balance de masa del hielo del CECs en glaciar Union (círculos
grises).
Figure 2. Color composition of a cross-polarized COSMO-SkyMed image (R=VV, G=VH, B=VV-VH) acquired on
14/Jul/2011, used for selection of the snow pit sites (light color squares).
Figura 2. Composición de imagen polarización cruzada COSMO-SkyMed (R=VV, G=VH, B=VV-VH) adquirida el 14/Jul/2011, usada para selección de localización de pozos de nieve (cuadrados blancos).
Cosmo-Skymed imagery was calibrated and georreferenced using satellite ephemerides data. Backscattering values in dB were transformed to a linear scale. For modelling the backscattering at

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
151
Union Glacier, five empirical (Ulaby et al., 1986; Partington & Hannah, 1994; Ulaby, Siqueira et al., 1995, Arslan, 2000; Thakur et al., 2013) and five physical based models (Baghdadi et al. 2000; Nagler & Rott, 2000; Albert et al., 2005; Jinyang et al., 2010; Phan et al., in press) were tested. Empirical models calculate the backscattering of the targets by using the relationship between a set of SAR images and field data on snowpack characteristics. On the other hand, physically based models estimate the backscattering by modeling the physical interactions of the electromagnetic radiation with the target, considering the three different types of scattering: Rayleigh scattering, Mie scattering, and non-selective scattering. RESULTS Results of the empirical modelling of the snow backscattering at X-band show relatively accurate results when compared to backscattering measured by Cosmo-SkyMed (Figure 3). The empirical model proposed by Thakur et al. (2013) presented the best estimation of the backscattering, as showed by the coefficient of determination estimated for the analysis. However, the reduced number of snow pits used both in the model and to estimate the errors might have biased the assessment of accuracy of the model.
Figure 3. X-band backscattering of the snow at Union Glacier estimated by empirical models (B). (A) shows the standard
error and coefficient of determination of the estimation. Figura 3. Retrodispersión banda X de la nieve en Union Glacier estimada por modelos empíricos (B). (A) muestra el error
estándar y el coeficiente de determinación (R2) de la estimación.
Figure 4. X-band backscattering of the snow at Union Glacier estimated by physically based models (B). (A) shows the
standard error and coefficient of determination (R2) of the estimation Figura 4. Retrodispersión banda X de la nieve en Union Glacier estimada por modelos físicos (B). (A) muestra el error
estándar y el coeficiente de determinación (R2) de la estimación. In general, the results of the physically based models of X-band backscattering at Union Glacier (Figure 4) showed better results than that obtained by empirical models (Figure 3). The mean coefficient of

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
152
determination for these models was 87%, while the mean R2 for empirical models was 73%. The model proposed by Jinyang et al. (2008) produced the best results among the physical models. ACKNOWLEDGEMENTS This research has been funded by Brazilian Edital MCT/CNPq - Nº 23/2009 – Seleção pública de propostas para apoio a projetos de pesquisa, no âmbito do Programa Antártico Brasileiro - PROANTAR and is supported by the Brazilian Instituto Nacional de Ciência e Tecnologia da Criosfera (INCT da Criosfera), the Fundación CEQUA and the Chilean Antarctic Institute. We thank ALE Company Team for support and friendship on the field. REFERENCES Albert, M. D., T. E. Tan, H. T. Ewe, and H. T. Chuah, (2005). A theoretical and measurement study of sea ice and ice shelf in antarctica as electrically dense media. Journal of Electromagnetic Waves and Applications, 19 (14), 1973-1981. ALE/ERA (2011), Union Glacier Map 1:100.000, Ellsworth Mountains Series 4, ISBN 978-0-9552205-5-5. Arslan, A. N., Koskinen, J., Pulliainen, J., Hallikainen, M., (2000). A Semi empirical backscattering model snow using SAR data. Proceedings IGARS 2000: 1904-1906. Baghdadi, N., Gauthier, Y., Bernier, M., Fortin, J.P., (2000). Potential and limitations of RADARSAT SAR data for wet snow monitoring. IEEE T. Geoscience and Remote Sensing 38(1): 316-320. Jinyang, D., Shi, J., Rott, H., (2010). Comparison between a multi-scattering and multi-layer snow scattering model and its parameterized snow backscattering model Remote Sensing of Environment, 114, 1089 –1098. Nagler, T. and H. Rott, (2000). Retrieval of Wet Snow by Means of Multitemporal SAR Data. IEEE Trans. Geosc. Rem. Sens., 38 (2), 754 – 765. Partington, K.C. and Hannah, M., (1994). Modelling radar sea ice backscatter in support of ERS-1 SAR. EARSEL Adv. In Remote Sensing, 3(2), 9-23. Phan, X., Ferro-Famil, L., Gay, M., Durand, Y., Dumont, M., Allain, S., D’urso, G., (in press). Analysis of snowpack properties and structure from TerraSAR-X data, based on multilayer backscattering and snow evolution modeling approaches. International JSTARS. Rivera, A., R. Zamora, C. Rada, J. Walton and S. Proctor, (2010). Glaciological investigations on Union Glacier, Ellsworth Mountains, West Antarctica. Annals of Glaciology, 51(55), 91-96. Rivera, A., Zamora, R., Uribe, J., Jaña, R., Arigony-Neto, J., Carrion, D., Wendt, A., (2012). Ice Dynamic of Union Glacier, West Antarctica, IGS International Symposium Glaciers and Ice sheets in a Warming Climate, EEUU, Fairbanks, Alaska, Thakur, P. K., Garg, R. D., Aggarwal, S. P., Garg, P. K., Snehmani, and Shi, J. (2013). Snow density retrieval using SAR data: algorithm validation and applications in part of North Western Himalaya, The Cryosphere Discuss., 7, 1927-1960 Ulaby F.T., R.K. Moore and A.K. Fung, (1986). Microwave Remote Sensing: Active and Passive, Vol. III. Reading, MA: Addison-Wesley. Ulaby F.T., P. Siqueira, A. Nashashibi and K. Sarabandi, (1995). Semi-Empirical Model for Radar Backscattering from Snow at 35 and 94 GHz. Proceedings of IGARSS, 1499-1501.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
153
DISTRIBUCIÓN DE TAMAÑO Y TIEMPO DE OCURRENCIA DE EVENTOS DE DESPRENDIMIENTO. INFORMACIÓN OBTENIDA DE LA COMBINACIÓN DE DATOS DE
VIDEO Y DE ESCANEO A TRAVÉS DE LASER TERRESTRE (Calving events size and time distribution from combined Terrestrial Laser Scanning and video data)
Pętlicki M.1, Kinnard C.1, Abermann J.1, Zambra S.1
1 Centro de Estudios Avanzados en Zonas Áridas, Raúl Bitrán s/n, Colina El Pino, La Serena Chile, E-mail: [email protected] ABSTRACT The largest component of ablation of the world’s cryosphere (van der Veen, 2002) and in Antarctica (Bengtsson et al., 2011) is calving, which is the mechanical loss of ice from glaciers and ice shelves (Benn et al., 2007). Albeit its importance for regional mass balance and thus sea-level rise, calving and related dynamic processes are poorly represented in the state of the art ice-sheet models (Benn et al., 2007). One of the reasons for that is the difficulty to collect field data and thus quantify processes, as long-term studies of calving rates are generally based on remote sensing methods which have time resolution coarser than the processes involved (e.g. Mansell et al., 2012). There is a lack of detailed observations at the scale of single events. The method that has been among the most used is the time lapse photography (e.g. Kirkbride and Warren, 1997, O’Neel et al., 2003). It provides data in a relatively high spatial resolution and usually up to hourly sampling interval. However, it is unable to produce quantitative results as a stand-alone method (O’Neel et al., 2010). Up to now no work on the application of terrestrial laser scanning (TLS) to quantify the calving activity has been published (Chapuis, 2011), although it offers many advantages over other techniques, among others its speed, precision and high spatial resolution (Prokop, 2008). Calving from Air Force Glacier, Greenwich Island, Antarctica, is investigated using a combination of video monitoring, Terrestrial Laser Scanning (TLS) and nearby environmental data. We present a new and innovative approach to monitor calving events based on the calibration of a statistical calving model with combined TLS and video data. The occurrence, frequency, size- and volume distribution of calving events was quantified over a short period (days to weeks). For that, a series of repeated scans was taken to detect the total ice volume lost by calving over the time elapsed between individual scans. The quantitative split-up into individual events was done by analysis of the videos and measuring the number of pixels that a single ice-block covers on each video frame. In that way, the number of pixels could be related to the volume change as measured by TLS. In this way the size- and volume distribution of calving events can be deduced using only the video sequence. The combined TLS and video data allowed investigating the correlation between calving activity and meteorological and oceanographic variables. It has been shown that these relations are complex and not straightforward (Chapuis, 2011; O’Neel et al., 2003; O’Neel et al., 2010) but that they do exist (Vieli et al., 2001; Benn et al. 2007a). Our field measurements were conducted during the peak ablation season. Therefore our direct glaciological mass balance observations provide information on significant variations in surface melt. Furthermore, precipitation measurements allowed us to study the link between water input into the glacial system and observed variations in calving rates. The precise identification of individual calving events from combined video and TLS measurements and their linkage with the meteorological and oceanographic data will allow ranking the importance of the factors that drive calving.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
154
ACKNOWLEDGEMENTS We would like to thank INACH and the crew of Arturo Prat Station for the logistic support during field work in Antarctica. This work has been funded by INACH grant RT_12-12. REFERENCES Bengtsson, L. Koumoutsaris, S.and K. Hodges 2011, Large-Scale Surface Mass Balance of Ice Sheets from a Comprehensive Atmospheric Model, Surveys in geophysics:1–16.D. Benn, Warren, C., and Mottram, R. 2007. Calving processes and the dynamics of calving glaciers. Earth-Science Reviews, 82:143–179. Chapuis, A. 2011. What controls the calving of glaciers? From observations to predictions. (PhD thesis, Norwegian University of Life Sciences). Kirkbride, M.P., Warren, C. R. 1997. Calving processes at a grounded ice cliff. Ann.Glaciol., 24:116-121. Mansell, D.T., Luckman, A., Murray, T. 2012, Dynamics of tidewater surge-type glaciers in northwest Svalbard. J. Glaciol., 58:110-118. O’Neel, S., C. F. Larsen, N. Rupert, and R. Hansen 2010, Iceberg calving as a primary source of regional‐scale glacier‐generated seismicity in the St. Elias Mountains, Alaska, J. Geophys. Res., 115:F04034. O’Neel, S., K. A. Echelmeyer and R. J. Motyka. 2003, Short-term variations in calving of a tidewater glacier: LeConte Glacier, Alaska. J. Glaciol., 49(167) :587-598. Prokop, A. 2008, Assessing the applicability of terrestrial laser scanning for spatial snow depth measurements. Cold Regions Science and Technology, 54(3):155–163. van der Veen, C.J. 2002, Calving glaciers. Progress in Physical Geography, 26(1):96-122. Vieli, A., M. Funk and H. Blatter 2001, Flow dynamics of tidewater glaciers: a numerical modelling approach. J. Glaciol., 47(159):595-606.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
155
CAMPAÑAS GLACIOLOGICAS DEL CECs Y ALE A UNION GLACIER 2008-2010, ANTÁRTICA OCCIDENTAL
(CECs and ALE glaciological surveys between 2008 to 2010 to Union Glacier, West Antarctica)
Zamora, R.1, Uribe, J.1, Gacitúa, G.1, Rivera, A.1,2
1Centro de Estudios Científicos (CECs), Av. Prat 514, Valdivia, Chile, [email protected] 2Departamento de Geografía, Universidad de Chile, Santiago, Chile RESUMEN Durante los meses de noviembre y diciembre de los años 2008, 2009 y 2010 se realizaron mediciones glaciológicas en el Union glacier (79º46’S/83º24’W) en Antártica occidental. Estas expediciones fueron organizadas por el Centro de Estudios Científicos (CECs) de Valdivia y la empresa Antarctic Logistics and Expeditions (ALE). Las travesías se llevaron a cabo utilizando un tractor Camoplast BR-350, el cual remolcó trineos con carga, combustible y un módulo con equipos científicos. En este módulo se instaló un sistema que integra un radar de onda continua de 550 a 900 MHz para medir acumulación de nieve/neviza y uno de compresión de pulsos de 150 MHz para medir la topografía subglacial, desarrollados por el CECs. Ambos radares demostraron ser instrumentos adecuados para las mediciones de acumulación de nieve en los primeros cientos de metros de espesor y de la topografía subglacial a lo largo de 460 km de travesías, donde se alcanzó un espesor máximo de 2200 m. INTRODUCCIÓN El CECs y la empresa privada ALE realizaron travesías a Antártida Occidental durante los años 2008-2010, con el objetivo de estudiar Union glacier con métodos glaciológicos, geofísicos y geodésicos (Rivera y otros, 2010). Mediante el uso de radares fue posible caracterizar la acumulación de nieve, la estructura interna del hielo y la topografía subglacial en aproximadamente 460 km de travesías terrestres. La medición con radar consiste en un transmisor que envía pulsos de energía electromagnética hacia el interior del hielo y, a continuación, un receptor recibe la energía electromagnética reflejada desde el fondo subglacial y a partir del tiempo de retardo de la onda electromagnética, es posible determinar la profundidad en metros. Otras interfaces capaces de reflejar energía electromagnética, están ubicadas en la estructura interna del hielo y pueden relacionarse con zonas con cambios de densidad (nieve/neviza, grietas o estructura interna del hielo). Estos estudios se enmarcan en un programa más amplio de investigación que incluye numerosas mediciones en el área (Vaughan y otros, 2007), que tienen como objetivo evaluar la estabilidad de Antártica Occidental frente a posibles cambios climáticos. MÉTODOS Para la medición de las capas de nieve/neviza y el espesor de hielo, se desarrolló un sistema de radar que consiste en la integración de dos radares, uno de alta resolución para los primeros 300 m de nieve/neviza y otro de alta penetración de hasta 4 km de espesor. Esta integración permitió una importante reducción de costos en la construcción del sistema, como también la reducción del consumo de energía (< 80 W) y de peso (aproximadamente 40 Kg). Las mediciones se posicionaron con GPS. El radar de compresión de pulsos se utiliza para la medición de espesor de hielo. Este diseño permite usar transmisores de radio frecuencia (RF) de relativa baja potencia en lugar de usar generadores de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
156
impulso de alto voltaje. Para conseguir esto último, es necesario transmitir un pulso largo modulado (chirp), el cual incrementa la potencia promedio manteniendo una buena resolución vertical del radar. Un sistema digital genera las formas de onda de cada radar, mediante el uso de Síntesis Digital Directa (Direct Digital Synthesizer, DDS). En el caso del radar de compresión de pulsos, el sistema digital genera una forma de onda chirp de largo 10 us máx., que varía entre 145 y 165 MHz (radar de frecuencia central 155 MHz y ancho de banda 20 MHz). El uso de 20 MHz de ancho de banda permite obtener una resolución vertical en hielo de 4.2 m. Esta señal luego es amplificada hasta 200 W para ser radiada por la antena Yagi transmisora, de 12 dBi. Los ecos provenientes del hielo son captados por la antena Yagi receptora, para luego ser amplificados por el receptor, el cual posee dos canales: un canal de baja ganancia y otro canal de alta ganancia, lo que permite obtener un gran rango dinámico. Estas dos señales luego son convertidas a dominio digital mediante un sistema de adquisición (conversores análogo-digitales), posteriormente las señales capturadas son integradas para aumentar la relación señal/ruido (Signal to noise ratio, SNR).
Figura 1: 460 km de mediciones de acumulación de nieve y espesor de hielo en el Glaciar Union utilizando un radar de onda continua y compresión de pulsos.
Figure 1: 460 km of snow accumulation and ice thickness measurements in Union Glacier using a FMCW and pulse shirp radar.
En el caso de la medición de nieve/neviza, se diseñó un radar de frecuencia modulada – onda continua (Frequency Modulated – Continuous Wave, FMCW). Este radar posee una ancho de banda de 350 MHz (opera en la banda de 550 a 900 MHz), lo cual permite obtener una resolución de 0.5 m en espacio libre. En este caso, el generador de señal crea una señal chirp de 50 a 550 MHz, para luego en el transmisor FMCW, ser elevada en frecuencia hasta la banda de 550 – 900 MHz. Operando a estas frecuencias, es

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
157
posible usar antenas más pequeñas. Para las mediciones en terreno, se usaron antenas log-periódicas de 6 dBi que pueden operar en la banda de 0.2 a 2.0 GHz. La señal proveniente de los ecos recibidos por el sistema FMCW es recepcionada y convertida a una señal de Frecuencia Intermedia (IF), el cual posee un ancho de banda de 20 MHz máximo para un rango de medición de 480 m. RESULTADOS Las mediciones de espesor de hielo en Union glacier (Figura 1) permitieron obtener un espesor promedio de 1450 m (Figura 2). En la travesía desde Union glacier hacia el Plateau Antártico (Figura 1), muchos sectores mostraron una topografía subglacial rugosa y reflectores internos intermedios, en especial cuando se transitó por los glaciares Schanz, Schneider, Balish y Driscoll. El máximo espesor medido fue de 1600 m. Al compararse los espesores obtenidos en los cruces se obtuvo una diferencia menor a 20 m, lo que se estima corresponde al error de la medición. La tasa de captura horizontal de las mediciones fue de 12 m considerando una velocidad del tractor promedio de 8 km/h. Asimismo el radar VHF permitió detectar reflectores internos intermedios, ubicados desde varios cientos de metros debajo de la superficie hasta más de 1500 m de profundidad.
Figura 2: Perfil longitudinal del Glaciar Union medido con el radar de compresión de pulsos. El espesor máximo en esta sección es de 1500 m. En la figura 1 se muestra la ubicación espacial de este perfil (A-A’).
Figure 2: Union Glacier longitudinal profile measured with the pulse shirp radar. The maximum thickness in this section is 1,500 m. Figure 1 shows the spatial location of this section (A-A ').
Se realizaron además mediciones de acumulación de nieve con el radar FMCW, donde se procesaron 100 m de rango dinámico logrando detectar estratigrafía en nieve/neviza con una resolución menor a 1 m, lo que permitirá determinar la acumulación interanual cuando se considere el balance de masa superficial. Además, se detectó una detallada estratigrafía interna en gran parte de los perfiles, los que tienen una especial relevancia en las divisorias locales de glaciares lugares idóneos para extraer testigos de hielo. Estas divisorias tienen estructuras internas caracterizadas por ondulaciones típicas de flujos mínimos y divergentes, llamadas “Raymond Bumbs”, (Figura 3), los que permiten conocer cambios de largo plazo en el flujo de hielo.
CONCLUSIONES Los resultados obtenidos hasta ahora, han permitido generar una línea de base glaciológica del área, incluyendo el balance de masa, dinámica glaciar y cambios recientes. Esta línea de base permitirá

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
158
estudiar respuestas dinámicas del hielo, por ejemplo, en relación a cambios en la zona del grounding line (línea de flotación) local, que sólo está a 40 km de distancia del campamento base en Union glacier.
Figura 3: Radargrama medido utilizando el radar FMCW. Se puede ver un rango de 100 m de nieve/neviza a una resolución menor a 1 m. En la figura 1 se muestra la ubicación espacial de este perfil (B-B’). En este perfil se pueden ver isocronas o
capas de igual edad, cuyas inflexiones y espesores permiten estudiar cambios glaciológicos de mediano y largo plazo. Figure 3: Radargram measured using the FMCW radar. A total range of 100 m of snow / firn at a resolution of less than 1 m. Figure 1 shows the spatial location of this section (B-B '). Along this profile, it is possible to see isochronous. The inflections
and thickness of these layers allow studying mid and long term glaciological changes.
Los datos de espesor han permitido generar una topografía subglacial completa de la zona, con espesores máximos de 2200 m. Se ha detectado además un gran número de estructuras internas del hielo, que permitirán analizar cambios glaciológicos de largo plazo y variaciones decadales en la acumulación de nieve.
AGRADECIMIENTOS Las expediciones fueron apoyadas por la empresa privada Antarctic Logistic Expeditions (ALE). El CECs es financiado por el Gobierno de Chile a través de los Centros de Excelencia del Programa de Financiamiento Basal de la Comisión Nacional de Investigación Científica y Tecnológica de Chile (CONICYT). REFERENCIAS Rivera, A., R. Zamora, C. Rada, J. Walton and S. Proctor, 2010. Glaciological investigations on Union Glacier, Ellsworth Moun- tains, West Antarctica, Ann. Glaciol., 51(55), 91–96. Vaughan, D. G., A. Rivera, J. Woodward, H. F. J. Corr, J. Wendt and R. Zamora, 2007. Topographic and hydrological controls on Subglacial Lake Ellsworth, West Antarctica, Geophys. Res. Lett., 34, L18501.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
159
HIBRIDACIÓN ENTRE Catharacta maccormicki Y Catharacta lonnbergi EN LA ISLA REY GEORGE (ANTÁRTIDA), DETECTADA POR EL ENFOQUE GENÉTICO EN PEQUEÑA
ESCALA. (Hybridization between Catharacta maccormicki and Catharacta lonnbergi in King George Island
(Antarctic) as detected by genetic approach in small scale) Castro, L.1, Costa, E. 2, 3, Alves, M. 2. Torres, J. 3, Durigon, E. 4, Araujo, J.4, Morgante, J.1 y Dantas, G.
1,5 1Depto. de Genética e Biologia Evolutiva, Instituto de Biociências, Universidade de São Paulo (USP), São Paulo, Brazil; [email protected] 2Universidade do Estado do Rio de Janeiro (UERJ), Rio de Janeiro, RJ, Brazil; 3Universidade Federal do Rio de Janeiro (UFRJ), Rio de Janeiro, RJ, Brazil; 4Depto. de Microbiologia, Instituto de Ciências Biomédicas - USP, São Paulo, SP, Brazil; 5LBEM, Depto. de Biologia Geral, Instituto de Ciências Biologicas (ICB), Universidade Federal de Minas Gerais (UFMG), Belo Horizonte, MG, Brazil. The Skuas (Stercorariidae) are territorials and long-distance migratory seabirds, which occur in north and south temperate extension of the globe, being abundant in polar habitats. Due to uncertain taxonomic and complex phylogeny patterns, Stercorariidae has been studied by taxonomists and ecologists. In general, all Skuas species are classified into two groups: 1) genus Catharacta, covering the complex of the Southern Hemisphere, and genus Stercorarius, comprising species of the Northern Hemisphere. Catharacta has a recent origin related to the last period of the Antarctic glaciation. In this way, this genus seems to present incomplete divergence, favoring frequent cases of hybridization among species. The most frequent case of hybridization recorded for Stercorariidae is the observed Catharacta maccormicki (South Polar Skuas) and C. lonnbergi (Brown Skua) in Antarctic Peninsula. The hybrids generated by this hybridization are relatively frequent in the King George Island since the mid-twentieth century; however, the reported information about their characteristics, fitness, genetic frequency and genetic traits are limited. Given the importance of knowing the mixed offspring in order to understand the speciation process (which still uncertain for this genus), this study aimed to evaluate the genetic diversity and the phylogenetic relationship between C. maccormicki and C. lonnbergi by means of a genetic approach, using both mitochondrial (Cytochrome b gene) and nuclear (β-Fibrinogen) markers. Since nuclear β-Fibrinogen was not successfully amplified, based on mitochondrial gene analysis we found that the Skuas species included in this study show high haplotypic (0.672) and nucleotidic (0.00338) diversity, with an estimate of Theta per site (θ=4Nµ) equivalent to 0.0117. From the neutrality tests performed (R2, D´Tajima and Fu (Fs) we obtained significant results evidencing that the observed polymorphisms are not selectively neutral, but derived from imbalances from the expected between the neutral predilections (Table 2).
D Tajima Fu(Fs) R2
Total Cytb Haplotipes -2,08 P=0,10 -4,86 P=0,10 0,0728 P=0,016
C.maccormicki -1,237 p=0,03 -0,922 p=0,10 0,2259 p=0,271
C.lonnbergi -1,8399 p=0,01 -7,494 p=0,00 0,00472 p= 0,015
Hybrids -0,7864 p=0,32 -0,272 p=0,33 -0,272 p=0,33
Table 1: Neutrality tests generated from the Cytochrome b fragment: D Tajima, statistics Fu (Fs) Ramos-Onsins & Rozas (R2).
Tabla 1: Tests de neutralidad generados para Citocromo b: D de Tajima, Fu(Fs), y Ramos-Onsins y Rozas (R2).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
160
The generated Mismatch distribution (Figure 1), network (Figure 2) configured for the Skuas Cytochrome b gene and Mismatch distribution graph all together with parsimonious tree evidenced fast and growing populational expansion standards, in agreement with the deviations observed in the neutrality tests performed.
Figure1. Mismatch Distribution for the Cytochrome b gene of South Polar Skua (C.maccormicki), Brow Skua (C.lonnbergi) and hybrids from King George Island, Antarctic. The continuous traces elucidate the expected results and the dotted trace,
the observed. Figure1. Distribución Mismatch para el gen Citocromo b de las Skuas Antártidas (C.maccormicki), Skuas Marrón
(C.lonnbergi) e híbridos de la Isla Rey George, Antártida. Las continuo huellas dilucidar el resultado esperado y en punteado, los resultados observados.
Figure 2. Network configured for the Skuas Cytochrome b gene fragment to C.maccormicki (black circle), C.lonnbergi
(white circle) and hybrids (gray circle). King George Island, Antarctic. Figura 2. Network configurado para fragmento del Citocromo b de C.maccormicki (círculo negro), C.lonnbergi (círculo
blanco) e híbridos (círculo gris). Isla Rey Gorge, Antártida.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
161
Considering the results above, it can be concluded that the Skuas species analyzed in this study diverged in the recent past (20,000 to 30,000 years ago, during the Pleistocene), thus justifying the existence of many haplotypes shared by the species. This fact generated a parsimony tree with low bootstrap which, added to the other obtained results, gives rise to a scenario of frequent hybridization between C. maccormicki and C. lonnbergi. ACKNOWLEDGEMENTS National Council for Scientific and Technological Development (CNPq/MCT: 557049/2009-1; 48.4002/2011-2; 107951/1012-6) and Brazilian Antarctic Program (PROANTAR). REFERENCES Carneiro, A.P.B., Polito, M.J., Sander, M., Trivelpiece, W.Z. 2010. Abundance and spatial distribution of sympatrically breeding Catharacta spp. (Skuas) in Admiralty Bay, King George Island, Antarctica. Polar Biology, 33: 673–682. Costa, E.S. & Alves, M.A.S. 2012. Climatic changes, glacial retraction and the Skuas (Catharacta sp. - Stercorariidae) in Henneqin Point (King George Island, Antartic Peninsula). Pesquisa Antártica Brasileira 5: 163-170 ( Brasilian Antartic Research) ISSN 0103-4049. Ritz, M.S., Millar, C., Miller, G.D., Phillips, R.A., Ryan, P., Sternkopf, V., Liebers-Helbig, D., Ulrich Peter, H. 2008. Phylogeography of the southern skua complex—rapid colonization of the Southern Hemisphere during a glacial period and reticulate evolution. Molecular Phylogenetics and Evolution 49 292–303.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
162
RESPUESTAS DE AVES MARINAS PELÁGICAS DEL MAR DEL SCOTIA, ANTÁRTIDA, ANTE FLUCTUACIONES CLIMÁTICAS
(Responses of pelagic seabirds of the Scotia Sea, Antarctica, to climatic fluctuations) Orgeira, J. L.1, 2, Alderete, M.2 y Lencina, V.B.3
1 Departamento Biología de Predadores Tope, Instituto Antártico Argentino. Email: [email protected] 2 Cátedra Ecología General, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. 3Instituto de Investigaciones Estadísticas, Facultad de Ciencias Económicas, Universidad Nacional de Tucumán/CONICET. INTRODUCCIÓN En el continente antártico se han registrado incrementos de temperatura que se han intensificado desde la década del 50´s (Zazulie et al. 2010). Estos procesos tienen consecuencias directas sobre todo el ecosistema marino porque el derretimiento del hielo reduce el reclutamiento del krill y de toda la fauna planctónica asociada a éste (Flores et al. 2012). Desde fines de la década de 1980 el Instituto Antártico Argentino desarrolla un programa de monitoreo del ecosistema marino; uno de sus objetivos es identificar las especies más vulnerables a los cambios ambientales y su tipo de respuesta. Este trabajo analiza las variaciones de abundancia de seis especies de la comunidad de aves del Mar del Scotia comparando dos periodos que han presentado condiciones ambientales contrastantes: temporadas verano 1995 y 1996 y temporadas de verano 2010 a 2013, con una marcada fluctuación climática entre ambos períodos. MATERIALES Y MÉTODOS La información se obtuvo por medio de censos de aves a bordo de los buques rompehielos ARA Almirante Irizar (temporadas de verano de 1994 y 1995, n=303) y Puerto Deseado (temporadas de verano 2010 a 2013, n=733). La metodología empleada es la propuesta por SCAR (1981) que consiste de censos de aves de 10 min de duración donde se registraron riqueza y abundancias de diferentes especies de aves marinas y distintas variables ambientales. El área de estudio corresponde a la región marítima adyacente a las Is. Orcadas del Sur (Mar del Scotia) delimitado por el siguiente cuadrante: 58º a 63°.15´ S y 40º a 49ºW aproximadamente. Las especie estudiadas fueron Petrel damero Daption capense, Petrel plateado Fulmarus glacialoides, Petrel barba blanca Procellaria aequinoctialis, Prión antártico Pachyptila desolata, Petrel de las tormentas Oceanites oceanicus y Albatros de ceja negra Thalassarche melanophris. Se elaboró una curva de dominancia basada en el promedio de las abundancias relativas de cada especie en los censos para cada temporada. Estas curvas resultan del
ordenamiento en cada temporada de los siguientes promedios: 1
TmijT
i Tj T
pP
m=
=∑ , donde i jT
ijtjT
np
n+= es la
abundancia relativa de la especie i en el censo j de la temporada T, siendo jTn+ el total de individuos de
todas las especies registradas en el conteo j de la temporada T y Tm el número de censos con al menos un individuo observado. Esta curva es una modificación del modelo de dominancia-diversidad de Whittaker (Whittaker 1972), donde las especies se ordenaron de más a menos dominantes. Se compararon entre temporadas diferentes variables ambientales (temperatura de la superficie del mar, temperatura del aire, presión atmosférica, estado del mar, % de cobertura de hielo, % de hielo tipo escombros y % de hielo tipo tabular y/o témpanos). Se compararon los promedios en las variables cuantitativas entre las temporadas (test no paramétrico, K-W). Para las variables nominales, medidas como proporciones, se utilizó un test estadístico de comparación de proporciones. Se evaluó si había

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
163
diferencias significativas (p<0.05) entre la proporción de un tipo particular de hielo en una temporada con respecto a la temporada 1995. RESULTADOS En términos climáticos, las temporadas 2010 a 2013 (Periodo 2) han mostrado incrementos promedios de más de 2,5ºC en las temperaturas del aire y mar y caídas de hasta 15% en la cobertura de hielo respecto de las temporadas de verano 1995, 1996 (Periodo 1) (Tabla 1). En coincidencia con el cambio en las condiciones ambientales, se registraron variaciones de abundancia en las especies: el Petrel damero fue la especie dominante en el periodo 1, siendo reemplazada por el Prión antártico en el periodo 2. En la temporada 1995 el Petrel damero representaba el 50% del total de las abundancias registradas en esta temporada, mientras que en 2013 representó sólo 7%. El Prión antártico incrementó su abundancia relativa mientras que el Albatros de ceja negra y el Petrel barba blanca mostraron bajos valores de abundancia durante todas las temporadas analizadas (Fig. 1).
Tabla 1: Diferencias en las variables ambientales en el Mar del Scotia, Antártida, según los conteos de aves realizados.
Promedio ±±±± error estándar. %: porcentaje de conteos con la presencia de la variable. Letras distintas corresponden a diferencias significativas (p<0.05). P, presión atmosférica (hPa); TA, temperatura del aire (ºC); TSM, temperatura superficial del mar (ºC); EM, estado del mar (escala Beaufort); PACK, cobertura de hielo (%); ESC, presencia de hielos tipo escombros;
TAB, presencia de hielos tipo témpano/tabular. Table 1: Differences in environmental variables in the Scotia Sea, Antarctica. Average ±±±± standard error. %: percentage of
counts with the presence of the variable. Different letters means significative differences (p <0.05). P, air pressure (hPa), TA, air temperature (°C) TSM, sea surface temperature (°C), EM, sea state (Beaufort scale), PACK, ice cover (%), ESC, presence
of ice debris; TAB, presence of ice floe/tabular.
Figura 1. Curvas de dominancia para cada temporada. DCAP, Petrel damero; OOCE, Petrel de las tormentas; FGLA, Petrel plateado; PDES, Prión antártico; PAEQ, Petrel barba blanca; DMEL, Albatros de ceja negra.
Figure 1. Dominance curve model por each studied season. DCAP, Cape pigeon; OOCE, Wilson´s Storm Petrel: FGLA, Southern Fulmar; PDES, Antarctic Prion; PAEQ, White-chinned Petrel; DMEL, Black-browed Albatross.
VAR 1995 N=119
1996 N=184
2010 N=106
2011 N=252
2012 N=244
2013 N=131
P TA TSM EM PACK ESC TAB
970.98a±±±± 2.88 -0.09 a ±±±± 0.25 -0.54 a ±±±± 0.08 3.89 a ±±±± 1.31 9.13 a±±±± 12.2 25%a 6% a
987.10b±±±± 2.32 0.51 a ±±±± 0.2 -0.89 b±±±± 0.07 3.08 b ±±±± 1.25 1.98 b ±±±± 2.3 51 %b 54% b
990.07bc ±±±± 3.06 2.4bc ±±±± 0.26 1.22 c ±±±± 0.09 3.58 a ±±±± 1.41 2.14 c ±±±± 1.08 36 % a 41% b
983.66b ±±±± 1.98 2.95c±±±± 0.17 2.12 d±±±± 0.06 3.83 a ±±±± 1.53 0 6% b 17% b
994.89cd±±±± 2.01 2.06bd ±±±± 0.17 1.89 d±±±± 0.06 3.87 a ±±±± 1.35 0.05 c ±±±± 0.66 13% b 15% b
1004.83d±±±± 2.75 1.49d ±±±± 0.24 1.95 d ±±±± 0.08 3.60 a ±±±± 1.44 2.11 d ±±±± 6.20 18% a 22% b

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
164
DISCUSIÓN El incremento en las temperaturas y la reducción del hielo marino registrados en el periodo 2 forman parte de un proceso de calentamiento progresivo iniciado décadas atrás. Zazulie et al. (2010) han confirmado un incremento sostenido en la temperatura del aire estadísticamente significativo desde 1950 en las Is. Orcadas del Sur (el único sitio en Antártida con registros diarios de más de un siglo). Una de las consecuencias del calentamiento sobre los océanos es la reducción de la productividad primaria, porque se alteran los patrones de convección y advección y porque el derretimiento del hielo marino reduce el reclutamiento del krill junto con toda la fauna planctónica asociada al mismo (Zuur et al. 2009, Flores et al. 2012). Estos cambios se transmiten hacia la totalidad de la trama trófica incluidos los predadores tope como las aves marinas, las cuales resultan impactadas indirectamente a través de los cambios en la abundancia y disponibilidad de sus recursos tróficos. En nuestro estudio, las tres especies con mayor caída en abundancias tienen una dieta basada casi estrictamente en krill: Petrel damero, Petrel plateado y Petrel de las tormentas. El Petrel damero, de hecho, ha sido designada “especie indicadora” por la CCAMLR (Commission for the Conservation of Antarctic Marine Living Resources) y como tal se asume que refleja cambios en la disponibilidad de krill (CCMLR 1985). Por lo tanto, su estado tiene importantes implicancias ecológicas ya que sugiere cambios en la estructura del ecosistema del Mar del Scotia. En el periodo 2 el Petrel plateado ha mostrado alta proporción de ausencia en los censos pero también un destacado efecto de agregación, como la gran concentración de unos 800 individuos registrados en un solo censo el 27 de febrero de 2010, a los 60º 42’, 46 º58’. Junto al Petrel damero, el Petrel de las tormentas presentó una tendencia negativa en todas las temporadas. Sugerimos que los cambios de abundancia de las especies consumidoras de krill del periodo 2 se deben a i) la baja cobertura de hielo, que implica pobre reclutamiento de krill, lo que lleva a ii) disminuciones poblacionales por falla u omisión del proceso reproductivo ante condiciones ambientales adversas (como ha sido documentado por Jenouvrier et al. 2003 para el Petrel plateado); iii) cambios en las distribuciones de algunas especies por otras áreas marinas donde la presencia y disponibilidad de recursos es óptimo, como fue registrado por Péron et al (2010). La especie dominante del periodo 2, el Prión antártico, prefiere aguas libres de hielo y presenta una variada dieta basada en krill, otros crustáceos, peces y calamares (del Hoyo et al. 1992). Esta amplitud de nicho trófico junto con su independencia del hielo marino para su ciclo de vida, parecen haber minimizado el impacto de las fluctuaciones climáticas del periodo 2 sobre sus poblaciones, ya que fue la única especie con incrementos progresivos en todas las temporadas, desplazando al Petrel damero como especie dominante. El Albatros ceja negra y el Petrel barba blanca son especies subantárticas que visitan la región en busca de alimento. Sus abundancias interanuales han resultado variables, lo que sugiere cambios en sus hábitats entre una temporada y otra. De hecho, ambas especies fueron observadas alimentándose en campos de escombros de hielo (en compañía además del Petrel damero) en las temporadas 2011 a 2013. La presencia de estas especies alimentándose en zonas de campos de escombros es un interesante ejemplo de cómo las variables físicas empujan a los organismos a los límites de sus tolerancias fisiológicas llevándolas a un hábitat que no es su óptimo en la búsqueda de su alimento. Para nuestro estudio algunas especies han tenido caídas críticas en sus abundancias y otras han mostrado modificaciones de sus distribuciones en coincidencia con fluctuaciones climáticas, lo que sugiere cierta vulnerabilidad. Los análisis de los incrementos térmicos de Zazulie et al. (2010) muestran que debemos esperar más alteraciones en el ecosistema marino antártico. Por lo tanto, los futuros esfuerzos de monitoreos se enfatizarán sobre las especies más vulnerables a los cambios para evaluar su grado de impacto y proveer información para planes de conservación.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
165
REFERENCIAS BIBLIOGRAFICAS del Hoyo, J., Elliott, A. y Sargatal, J. 1992. Handbook of the birds of the World. Vol.1. (Ostrich to ducks). Barcelona: Linx Editions. CCMLR. 1985. Report of the fourth meeting of the Scientific Committee, Annex 7. Rep Ad hoc Working-Group on Ecosystem Monitoring. SC-CCAMLR-IV. Hobart, Australia. Flores, H., Atkinson, A., Kawaguchi, S., Krafft, B.A., Milinevsky, G., Nicol, S., Reiss, C., Tarling, G.A., Werner, R., Bravo-Rebolledo, E., Cirelli, V., Cuzin-Roudy, J., Fielding, S., Groeneveld, J.J., Haraldsson, M., Lombana, A., Marschoff, E., Meyer, B., Pakhomov, E.A., Rombolá, E., Schmidt, K., Siegel, V., Teschke, M., Tonkes, H., Toullec, J.Y., Trathan, P.N., Tremblay, N., Van de Putte, A.P., van Franeker, J.A. and Werner, T. 2012. Impact of climate change on Antarctic krill. Mar. Ecol. Prog. Ser. Vol. 458: 1-19. Jenouvrier, S., Barbraud, C. and Weimerskirch, H. 2003. Effects of climate variability on the temporal population dynamics of Southern fulmars. Journal of Animal Ecology 72, 576–587. Péron, C., Authier, M., Barbraud, C., Delord, K., Besson, D. and Weimerskirch, H. 2010. Interdecadal changes in at-sea distribution and abundance of subantarctic seabirds along a latitudinal gradient in the Southern Indian Ocean. S.C.A.R. 1981. BIOMASS Seabird Ecology. Post FIBEX Data Workshop. BIOMASS Report Series, 22. Zazulie, N., Rusticucci M. and Solomon, S. 2010. Changes in climate high Southern latitudes: a unique daily record at Orcadas spanning 1903-1908. American Meteorological Society 23: 189-196. Zuur, A.F., Barbraud, C., Ieno, E.N., Weimerskirch, H., Smith, G.M. and Walker, N. J. 2009. Estimating trends for Antarctic birds in relation to climate changes. Mixed effects models and extensions in Ecology with R. Statistics for Biology and Health. DOI 10.1007/978-0-387-87458-6_14. Spginger+Business Media. Whittaker, R.H. 1972. Evolution and measurement of species diversity. Taxon, 21: 213-251.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
166
IMPLICANCIAS DE LA VARIACIÓN MORFOLÓGICA A NIVEL GEOGRÁFICO ENTRE LAS COLONIAS DE PINGÜINOS PAPUA (PYGOSCELIS PAPUA) PARA EL USO DE
MEDIDAS MORFOMÉTRICAS COMO MÉTODO DE SEXAJE (Implications of geographic morphological variation among colonies of Gentoo penguin (Pygoscelis
papua) using morphometric measurements as a method of sex determination)
Valenzuela-Guerra P.1, Morales-Moraga D.1, González-Acuña D.2 y Vianna J.A.1
1Pontificia Universidad Católica de Chile, Departamento de Ecosistemas y Medio Ambiente, Vicuña Mackenna 4860, Macul, Santiago, Chile. E-mail : [email protected], [email protected] 2Universidad de Concepción, Facultad de Ciencias Veterinarias, Departamento de Ciencias Pecuarias, Chillán, Chile. INTRODUCCIÓN Los cambios fenotípicos pueden ocurrir en respuesta a condiciones ambientales locales, variación genética, evolución de poblaciones y la historia biogeográfica de una especie, lo que puede derivar en variación morfológica geográfica. Morfológicamente dos subespecies han sido descritas para Pingüinos papua (Pygoscelis papua), P. p. papua, J.R. Forster, 1781 y P. p. ellsworthii, Murphy, 1947 (Stonehouse 1970). Pese a lo antes mencionado, diferencias morfológicas entre poblaciones de las subespecies de pingüinos Papua no han sido investigadas. Por otra parte, determinar el sexo de los pingüinos es complicado, puesto que aparentemente estas aves son sexualmente monomórficas. Sin embargo, se ha observado cierto grado de dimorfismo sexual en diferentes especies de pingüino, permitiendo desarrollar funciones discriminantes, que utilizan datos morfométricos para diferenciar entre sexos. Esta técnica también se ha aplicado a Pygoscelis papua (Williams 1990; Renner y cols. 1998; Polito y cols. 2012), aunque estas funciones discriminantes, al ser utilizadas de forma general para determinar el sexo de los ambas subespecies, podría provocar errores de identificación debido a sus variaciones morfológicas geográficas. Por lo tanto, se deberían generar diferentes funciones discriminantes para cada subespecie, además de ser localidad específica. Para asegurar la correcta asignación del sexo de pingüinos Papua, utilizamos dos métodos moleculares de identificación sexual en individuos de diferentes localidades geográficas en la Península Antártica e Islas subantarticas. También evaluamos la variación morfométrica entre sexos y localidades. Finalmente, desarrollamos funciones discriminantes para determinar el sexo de Pygoscelis papua adultos usando medidas morfométricas respaldadas por sexaje molecular. MATERIALES Y MÉTODOS Se obtuvieron 300 muestras de sangre de Pygoscelis papua durante el verano del 2011, en 3 localidades cercanas a Bases Antárticas Chilenas: Profesor Julio Escudero (62°12'S-96 58°55'W) en Isla Ardley-Islas Shetland del Sur (A. Island), Bernardo O’Higgins (B. O’Higgins) (63°19'S-57°51'W) en Cabo Legoupil- Península Antártica y Gabriel González Videla (G.G.V.) (64°49'S-62°51'W) en Bahía Paraíso- Península Antárctica. Además, de cada individuo muestreado, se obtuvieron siete medidas morfométricas: Largo del pico (BL), Altura del pico (BD), Ancho del pico (BW), Largo total (TOL), Lago del ala (WL), Largo del tarso (TAL) y Largo de la Cola (TL) (Fig.1). Después de la extracción de ADN utilizamos los partidores P2/ P8 (Griffiths y cols. 1998) y PL/PR (Zhang y cols. 2012), para determinar el sexo de los individuos. Todas la reacciones fueron realizadas de acuerdo a Reis y cols. (2011). Los productos del PCR fueron visualizados por electroforesis en gel de agarosa al 3%. Conocido el sexo de 164 individuos adultos, se obtuvieron funciones discriminantes a

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
167
partir de análisis multivariante de las variables morfométricas, medidas mediante el procedimiento “backward stepwise regression” a partir del estadístico lambda de wilks. Se identificó para cada localidad y todas en conjunto, el orden de entrada de las variables que maximizaba las diferencias entre grupos y se escogieron las funciones discriminantes más robusta estadísticamente. La tasa de individuos correctamente sexados fue determinada mediante Jackniffe. También se realizó un análisis de componentes principales (PCA) para medir la cantidad de varianza aportada por cada variable en cada localidad y todas ellas combinadas, además de medir efectos de colinealidad a partir de la matriz de correlación intra-grupos (r>0.5).
Figura 1: Medidas tomadas a Pygoscelis papua. Largo del pico (BL), Altura del pico (BD), Ancho del pico (BW), Largo total
(TOL), Lago del ala (WL), Largo del tarso (TAL) y Largo de la Cola (TL). Figure 1:Measurements taken to Pygoscelis papua. Bill length (BL), bill depth (BD), bill width (BW), total length (TOL),
flipper length (WL), tarsus length (TAL) and tail length (TL). RESULTADOS Los dos pares de partidores utilizados mostraron los resultados esperados, dos bandas para las hembras y una para los machos de Pygoscelis papua. El éxito de amplificación vario entre ambos pares de partidores, P2/P8 determino el sexo del 89% de los individuos aproximadamente (266 de 300 individuos), mientras PL/PR determino el sexo del 81% de los individuos aproximadamente (244 de 300 individuos). Al comparar los resultados obtenidos por ambos pares de partidores, hubo discrepancia en el sexaje de 11 individuos. Considerando esto y el insuficiente dimorfismo entre juveniles y polluelos, solamente utilizamos las medidas morfométrica de 164 adultos para generar funciones discriminantes.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
168
En general, los machos fueron más grandes que las hembras para todas las variables, en los tres sitios de estudio. Al evaluar todas las variables en conjunto, seguimos observando la misma tendencia (Media TOL: 73.87 hembras y 76.34 machos; p-value< 0.001). Cuando comparamos todas las variables entre los tres sitios, encontramos que B. O’Higgins y A. Island no presentaban diferencias morfológicas significativas. Sin embargo, G.G.V fue significativamente diferente con B. O’Higgins y A. Island. Los individuos de G.G.V. fueron más pequeños en WL y TOL que en las otras localidades (t-test; p-value <0.05). Finalmente utilizando las funciones discriminantes desarrolladas, el sexo asignado a los individuos correctamente fue de: 90.25% para A. Island, 87.81% para B. O’Higgins y 82.93% para G.G.V. La función creada para todos los sitios en conjunto, obtuvo un 80% aproximadamente de éxito al asignar el sexo de los individuos. DISCUSIÓN P. p. ellsworthii se caracteriza por ser más pequeño que P. p. papua (Stonehouse 1970). Al comparar los datos morfométricos de tres colonias de P. p. ellsworthii observamos una tendencia similar. Los individuos de G.G.V. (la localidad más al sur) son más pequeños en comparación a los de las otras localidades, estas diferencias probablemente se deban a una limitada migración entre G.G.V. y las localidades más al norte. Las grandes distancias entre los archipiélagos que proporcionan terrenos para las colonias reproductivas pueden ser barreras geográficas, porque los pingüinos Papua son una especie no migratoria. Sin embargo, estudios demográficos recientes han mostrado el efecto del cambio climático en Pygoscelis papua, expandiendo su rango hacia mayores latitudes (Carlini y cols. 2009; Forcada y Trathan 2009). A pesar de esto, aun se observa una limitada migración entre la localidad más al Sur (G.G.V.) y las otras localidades, o bien de haber migración, esta es insuficiente para homogeneizar morfológicamente las poblaciones. Esto debe investigarse más a fondo, con estudios genéticos de poblaciones a través de diferentes latitudes, para evaluar el flujo génico hacia las localidades del sur. La variación morfológica a nivel geográfico, limita la aplicabilidad de funciones discriminantes para identificar el sexo de ambas subespecies de Pygoscelis papua (Renner y cols. 1998). Solo una función discriminante se ha obtenido para el sexaje de P. p. papua (Williams 1990), mientras que diferentes estudios para la identificación del sexo, utilizando medidas morfométrica, se han desarrollado para P. p. ellsworthii (Renner y cols. 1998; Polito y cols. 2012). Sin embargo, ninguno de estos estudios ha evaluado la variación morfológica entre las subespecies de pingüinos Papua. De acuerdo con las diferencias morfológicas que hemos encontrado entre los individuos de las diferentes localidades, las funciones discriminantes que hemos desarrollado en este estudio, para determinar el sexo de los individuos adultos de P. p. ellsworthii, solo deben ser utilizadas en las localidades para las cuales fueron creadas. A pesar de las diferencias morfológicas encontradas entre las localidades, desarrollamos una función discriminante general para la identificación del sexo de P. p. ellsworthii, la cual puede ser usada en diferentes localidades de la Península Antártica y las Islas Shetland del Sur con una bajo grado de confiabilidad (80% aproximadamente) en comparación con las funciones específicas desarrolladas para cada localidad (90.25% para A. Island, 87.81% para B. O’Higgins y 82.93% para G.G.V.). Sin embargo, el gran número de individuos (n=164) utilizados para desarrollar esta función general y un 80% de éxito al determinar el sexo de los individuos, son características aceptables para utilizar esta función discriminante en Pygoscelis papua (Kerry y cols. 1992). CONCLUSIÓN Las funciones discriminantes desarrolladas en este estudio, pueden proporcionar resultados inmediatos en estudios de campo, además, de ser utilizadas en diversos estudios que requieran el sexaje de los

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
169
individuos a lo largo de la Península Antártica, tales como incrementar el éxito reproductivo de las aves para desarrollar planes de conservación, de ser requeridos. De acuerdo con nuestros resultados y los de otros estudios la variación morfológica a nivel geográfico en diferentes especies de pingüinos limita la aplicabilidad de funciones discriminantes, por lo que recomendamos el uso de estas considerando la variación morfológica de los individuos a nivel geográfico. Por otra parte consideramos, que las variaciones morfológicas geográficas encontradas en Pygoscelis papua en este estudio son importantes para futuras investigaciones, tales como genética de poblaciones. AGRADECIMIENTOS INACH-G_06-11; INACH-T-27-10; INACH-MG_02-12. BIBLIOGRAFÍA Carlini, A.R., Coria, N.R., Santos, M.M., Negrete, J., Juares, M.A. y Daneri, G.A. 2009. Responses of Pygoscelis adeliae and P. papua populations to environmental changes at Isla 25 de Mayo (King George Island). Polar Biology, 32:1427–1433. Forcada, J. y Trathan, P.N. 2009. Penguin responses to climate change in the Southern Ocean. Global Change Biology, 15:1618–1630. Griffiths, R., Double, M.C., Orr, K. y Dawson, R.J.G. 1998. A DNA test to sex most birds. Molecular Ecology, 7: 1071–1075. Kerry, K.R., Agnew, D.J., Clarke, J.R. y Else, G.D. 1992. Use of morphometric parameters for the determination of sex of Adelie penguins. Wildlife Research, 19:657-664. Polito, M.J., Clucas, G.V., Hart, T. y Trivelpiece, W.Z. 2012. A simplified method of determining the sex of Pygoscelis penguins using bill measurements. Marine Ornithology, 40: 89–94 Reis, E.C., Aires, R.M., Moura, J.F., Matias, C.A., Tavares, M., Ott, P.H., Siciliano, S. y Lôbo- Hajdu, G. 2011. Molecular sexing of unusually large numbers of Spheniscus magellanicus (Spheniscidae) washed ashore along the Brazilian coast in 2008. Genetics and Molecular Research, 10:3731-3737. Renner, M., Valencia, J., Davis, L.S., Saez, D. y Cifuentes, O. 1998. Sexing of adult Gentoo Penguins in Antarctica using morphometrics. Colon Waterbirds, 21:444-449. Stonehouse, B. 1970. Geographic variation in Gentoo Penguins Pygoscelis papua. Ibis, 112:52–57. Williams, T.D. 1990. Annual variation in breeding biology of Gentoo Penguins, Pygoscelisp papua, at Bird Island, South Georgia. Journal of Zoology, 222: 247-258. Zhang, P., Han, J., Liu, Q., Zhang, J. y Zhang, X. 2012. Sex Identification of Four Penguin Species using Locus-Specific PCR. Zoo Biology 00:1-10.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
170
REPRODUCTIVE BIOLOGY AND BEHAVIOR, NON-INVASIVE ANALYSES OF MERCURY AND STRESS ECOLOGY OF ANTARCTIC SKUAS (CATHARACTA SPP.)
(Biología reproductiva y comportamiento, análisis no invasivo de mercurio y ecología de estrés de la skúa antártica (Catharacta spp.))
Costa, E.S.*, 1, 2 y Alves, M.A.S. 2
1Laboratório de Radioisótopos Eduardo Penna Franca, Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, RJ. CEP: 214941-900 [email protected] 2Laboratório de Ecologia de Aves, Instituto de Biologia Roberto Alcantara Gomes, Universidade do Estado do Rio de Janeiro, RJ. CEP 20550-011 ABSTRACT The general aim was to investigate aspects of reproductive biology, behavioral ecology, mercury contamination and stress levels of the Antarctic skuas, Catharacta maccormicki and C. lonnbergi. The thesis was divided into four chapters whose objectives were: 1) To study the reproductive biology and behavioral ecology using molecular techniques to complement information from both species, 2) To evaluate the use of vegetation in nests by skuas and the influence of vegetation in maintaining the temperature of eggs, in behavior of breeding pairs and in offspring’s survival, 3) To hold noninvasive studies of environmental pollution to assess contamination levels of mercury in feathers of chicks and adults, 4) To determine stress levels in the blood of adults and chicks in two conditions (immediately after arrest and after 30 minutes of restraint) as auxiliary tools in determining the environmental quality of the region. Field activities occurred mainly in Admiralty Bay (Antarctic Peninsula). The main results obtained were: 1) The use of weight and morphometric variables for the confirmation of sex by discriminant analysis was more efficient than use only observational data. 2) The use of morphometric and weight to classify the species was effective but it was not efficient for the separation between species and their hybrids. 3) The Skuas breeding most often in areas with medium and high vegetation cover in both regions analyzed and we also found a relationship between the occupation of the breeding areas and vegetation cover in the nests. 4) The vegetation in the nests influences the eggs’ temperature and the chicks’ survival for both species. 5) Adults of C. maccormicki showed the highest levels of mercury contamination and they can be excellent indicators of global contamination because they migrate to more remote areas and are exposed to global contamination by mercury. 6) Adults of C. lonnbergi and chicks of both species can be good indicators of local contamination by mercury because they suffer direct influence of contamination of the Antarctic environment. 7) Breeding adults showed basal levels of corticosterone significantly higher than no-breeding adults and there was no difference among corticosterone levels obtained for the young at different ages. 8) The corticosterone can be used as a measure to identify altered levels of in stress in seasonal situations (reproduction) and in non-predictable stress situations of stress, such as capture and restraint of individuals. This thesis presents novel results for the sampled species and for the study area and considerably increase knowledge on the ecology, reproductive biology, behavior, environmental contamination and stress ecology of Antarctic Skuas. ACKNOWLEDGEMENTS National Council for Scientific and Technological Development (CNPq/MCT: 557049/2009-1; 48.4002/2011-2), Brazilian Antarctic Program (PROANTAR), and Rio de Janeiro State Foundation of Support for Research (FAPERJ E-26/111.505/2010).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
171
PUENTES FINICRETÁCICOS ENTRE PATAGONIA Y ANTÁRTICA: EVIDENCIAS DESDE LA PALEOBIOLOGÍA
(End-Cretaceous bridges between Patagonia and Antarctica: evidences from the paleobiology) Leppe, M.1, Stinnesbeck, W.2, Mansilla, H.1, Ortiz, H.1,3, Manriquez, L.4, Gonzalez, E.5 y Vogt, M.2
1Laboratorio de Paleobiología de Patagonia y Antártica, Departamento Científico, Instituto Antártico Chileno, Plaza Muñoz Gamero 1055, Punta Arenas. [email protected] 2Institut für Geowissenschaften, Ruprecht-Karls-Universität Heidelberg, Alemania. 3Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile. 4Departamento Ciencias de la Tierra, Universidad de Concepción, Concepción, Chile. 5Departamento de Geología, Facultad de Ciencias Físicas y Matemáticas, Universidad de Chile. RESUMEN Las regiones australes albergan un registro único de los diversos procesos que tuvieron lugar antes y durante la apertura de una importante ruta de dispersión: el Paso de Drake, y del siguiente período de aislamiento del continente antártico hasta su posición actual. A pesar de los esfuerzos nacionales e internacionales durante el Año Polar Internacional 2007-2009, muchos de los aspectos fundamentales de las diferentes fuerzas motrices que conducen la evolución biológica y geológica de la Península Antártica (AP) y el sur de América del Sur (SSA), siguen siendo desconocidas. El presente trabajo discute la hipótesis que plantea la existencia de un puente terrestre a fines del Maastrichtiano, puente que habría tenido un efecto muy determinante sobre el origen y diversificación de lo que actualmente se conoce como “bosque valdiviano” o “bosques subantárticos del sur de Sudamérica”. INTRODUCCIÓN Los ecosistemas forestales en la Antártica han evolucionado principalmente en condiciones más cálidas que el presente, hasta el Neógeno, cuando una fuerte caída en la temperatura condiciona la existencia de flora arbórea (Cantrill 2001, Francis y Poole 2002). La estructura de la vegetación, sin embargo, sufrió cambios considerables, desde bosques dominados por coníferas y pteridofitas durante el Cretácico inferior (Falcon-Lang et al. 2001) a los bosques dominados por angiospermas hacia fines del Cretácico. Elementos comunes de las floras del Cretácico Superior de la AP están bien representadas en los diferentes tipos de bosques modernos chilenos (Dutra y Batten 2000, Poole et al. 2001, Cantrill y Poole 2002, Leppe et al. 2007). Depósitos del Cretácico Superior al Paleógeno tienen un registro masivo de la Antártica: de la cuenca James Ross (islas Seymour, James Ross y Vega), Grupo península Fildes (Rey Jorge y Nelson) e isla Adelaida, así como SSA: Cuenca de Magallanes (Cantrill 1997, 2000, Césari et al. 1999). Reconstrucciones paleoecológicas indican que modelos florísticos han evolucionado en el contexto de entornos de bajo perturbación volcánica intensa (Poole et al. 2001). Recientemente, se ha puesto en cuestión el modelo global de un fin del Cretácico con pulsos de calor durante los eventos Turoniano y frío durante el Maastrichtiano (Huber 1998). Las bajas temperaturas cercanas al final del Cretácico fueron nuevamente seguidas por un aumento de la temperatura u óptimo climático, durante el Paleoceno y Eoceno (Upchurch et al. 1998). Los taxa del lapso Jurásico Superior-Cretácico Inferior evolucionaron aislados en SSA y AP, y hasta el restablecimiento de un puente terrestre entre ambas masas de tierra durante el Turoniano. Durante el Campaniano-Maastrichtiano de AP dieron lugar condiciones propicias para la formación y diversificación de un nuevo modelo vegetacional, adaptado a las continuas perturbaciones por fuego (producto de un activo arco volcánico), a eventos esporádicos de enfriamiento (en un lapso

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
172
considerado de los más cálidos de la historia natural), a una estacionalidad muy marcada y al dominio de distintos taxa de la familia Nothofagaceae. La mezcla de estos elementos provenientes de AP con los remanentes de Patagonia Austral se pueden considerar predecesores de los modernos bosques templados de SSA. Recientemente, nuevos argumentos han ampliado el debate sobre los orígenes y la persistencia de esta biota templada del sur, sobre todo de la selva valdiviana, una hoja ancha y la ecorregión bosque mixto en la costa oeste del sur de Sudamérica, principalmente en Chile y el oeste de Argentina (Poole et al 2003, Povilauskas et al. 2008). Varios autores han señalado que la selva valdiviana es el equivalente más cercano a los bosques del Cretácico-Paleógeno superior de la Antártida (Francis y Poole 2002, Cantrill 2001, Poole et al. 2003), pero esta hipótesis se basaba únicamente en algunos taxones o localidades. La Región Antártica tiene una singular geografía e incluye una franja costera de Chile y el extremo sur de América del Sur, Antártica, islas subantárticas, y Nueva Zelanda. La Subregión Subantártica (sur de Chile, Patagonia, y Nueva Zelanda) posee una flora distintiva que comprende unos 50 géneros, entre los cuales Nothofagus es un elemento característico. Asociaciones vegetales de este tipo se reconocen ya en SA en depósitos de edad paleógena señalándose señaló el final del Paleógeno como el momento clave para el origen de los bosques del sur de América del Sur. Romero (1978) sugiere evolución florística de tres tipos de paleoflora de durante el Terciario: Neotropical, Mixta y Antártica. Por otra parte, de acuerdo con la hipótesis biogeográfica planteada por Schumithüsen (1956), los principales componentes de la flora de clima templado y los bosques subtropicales de Chile tendrían un origen común de las floras Neotropical y la Antártica palógena. En cambio Hinojosa y Villagrán (1997) se inclinan por postular que los conjuntos florísticos similares a los de los bosques templados del sur de Sudamérica se registraron durante el Oligoceno y el Mioceno. Una tercera hipótesis plantea que estos bosques serían un legado de los bosques subantárticos del Cretácico Superior-Paleógeno, apoyado por los registros paleobotánicos de la Península Antártica (Poole et al. 2001). El estudio de los orígenes de la biota del Hemisferio Sur ha prolífico en la producción numerosas explicaciones de los patrones observados en la actualidad. Sin embargo, existe un porcentaje importante de aproximaciones que usan a Nothofagus como género clave para reconstruir la compleja historia de Gondwana. El análisis paleobiológico del taxón arroja varias singularidades de gran significación biogeográfica en un escenario de fragmentación continental, como el del Meso-Cenozoico (Hill 2001). Por ejemplo, sus semillas tienen una baja capacidad de dispersión, su viabilidad de germinación es corta en el tiempo, sus semillas son intolerantes al agua de mar y sus granos de polen son anemocóricos, por lo que se pueden dispersar grandes distancias. Si a ello sumamos a una presencia prominente en los continentes del sur, una actual distribución disjunta y un excelente registro fósil, entonces el estudio integral de Nothofagus podría colaborar a reconstruir la intrincada historia geológica de SSA y AP. Recientemente se ha señalado a otros taxa, como Proteaceae, con características similares a las de Nothofagus, como potenciales sujetos para ayudar a testear los modelos planteados, pero además, varios autores han indicado que Podocarpaceae, Atherospermataceae (Poole y Francis 1999), Aquifoliaceae, Mataceae y Winteraceae (Poole y Francis 2000), son taxa que muestran una fuerte afinidad en la estructura y composición de los bosques templados del sur de Chile y Argentina (Dutra y Batten 2000, Poole et al 2001, Cantrill y Poole 2002, Leppe et al 2007) y tienen registro fósil antártico. El registro más antiguo de Nothofagaceae de AP data del Campaniano (~80 Ma) (Francis 1991, Dutra 1997, Dutra y Batten 2000, Dutra 2001, Dutra 2005). Otros antecedentes muestran que casi la mitad del polen de angiospermas del Maastrichtiano de la cuenca de James Ross es aparentemente endémica. Askin (1992) atribuye esta aparente exclusividad floral como un producto de diversas barreras geográficas, climáticas o biológicas. El alto grado de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
173
exclusividad las floras antárticas durante el Cretácico no apoya, al menos en el intervalo Campaniano-Maastrichtiano Inferior, la existencia de conexiones terrestres entre ambas masas de tierra. Los puentes de tierra entre la SSA y la AP se consideraría crucial para el flujo genético de las plantas y animales terrestres, así como para la determinación de las condiciones ambientales, debido a la ampliación de las condiciones oceánicas debido a los vientos del oeste (Elliot 1988, Hathway 2000). Estas condiciones climáticas, junto con el terreno montañoso como resultado de las lluvias orográficas, altos niveles de CO2 y la proximidad del arco volcánico, moderando las temperaturas extremas y la distribución de las precipitaciones durante todo el año, fueron las condiciones adecuadas para el rápido establecimiento y la expansión de tipo forestal valdiviano. Estas condiciones climáticas cambiaron en el Campaniano y Maastrichtiano superior inferior debido a la subducción continua y la agregación de tierra en el lado occidental de la AP, además de una disminución en los niveles atmosféricos globales de CO2. Estos cambios en las condiciones ambientales podrían explicar el patrón de anillo de crecimiento en la madera fósil de la parte occidental del istmo (Francis 1991). En el lado oriental, la cuenca James Ross estaba más protegida y los cambios son menos visibles. Recientemente, una campaña de un grupo de investigadores del Laboratorio de Paleobiología de Patagonia y Antártica del Instituto Antártico Chileno, del Institut für Geowissenschaften de la Universidad de Heidelberg y del Consejo de Monumentos Nacionales al valle del Río de Las Chinas, al norte de la Provincia de Última Esperanza, han reportado el hallazgo de los primeros dinosaurios (hadrosaurios y titanosaurios) de la Región de Magallanes, y subyaciendo a estos fósiles in situ, un nivel de 4,5 m de potencia con abundantes improntas fósiles de Taeniopteris, Sterculiaceae, al menos 10 morfos de angiospermas y dos tipos distintos de Nothofagus. Registros polínicos de Nothofagaceae no son raros para el Cretácico de Patagonia, atribuyéndose siempre su presencia a fuentes alóctonas, dada su baja representación en el recuento final de polen, pero el registro de más antiguo de improntas de hojas solo llegaba hasta el Paleógeno. Un análisis polínico de la secuencia arroja una edad Maastrichtiano Superior, correlacionándola con la Fm. Monte Chico o la Fm. Calafate en Patagonia Argentina, pero ninguna de las otras formaciones reporta improntas de Nothofagus. En base a los antecedentes presentados, postulamos la existencia de un puente terrestre a fines del Maastrichtiano, que permitió la primera entrada del bosque dominado por Nothofagus a SSA, sentando las bases para el establecimiento de la flora mixta.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
174
Figura 1: Flora maastrichtiana de Las Chinas. A: Sterculia, B: Taeniopteris, C y D: Nothofagus, E: Sitio de muestreo. Figure 1: Maastrichtian Flora from Las Chinas. A: Sterculia, B: Taeniopteris, C y D: Nothofagus, E: Sampling site.
AGRADECIMIENTOS Al personal de Estancia Cerro Guido, a INACH, proyectos FONDECYT 11080223, DAAD-CONICYT 259‐2010 y BMBF CHL 10A/09. REFERENCIAS Askin Ra. 1992. Late Cretaceous–early Tertiary Antarctic outcrop evidence for past vegetation and climates, in The Antarctic Paleoenvironment: A Perspective on Global Change: Part One, Antarct. Res. Ser., vol. 56, edited by J. P. Kennett and D. A. Warkne, pp. 61–73, AGU, Washington, D. C. Cantrill DJ. 1997. Hepatophytes from the Early Cretaceous of Alexander Island, Antarctica: Systematics and palaeoecology. International Journal of Plant Sciences 158: 476-488. Cantrill DJ. 2000. A Cretaceous (Aptian) flora from President Head, Snow Island, Antarctica. Palaeontographica Abteilung B-Palaophytologie 253: 153-191. Cantrill DJ. 2001. Early Oligocene Nothofagus from CRP-3, Antarctica: Implications for the vegetation history. Terra Antarctica (Germany) 8: 401-406. Cantrill DJ. & I Poole. 2002. Cretaceous to Tertiary patterns of diversity changes in the Antarctic Peninsula. In: Owen AW & JA Crame (eds) Palaeobiogeography and Biodiversity Change: A Comparison of the Ordovician and Mesozoic-Cenozoic Radiations, Geological. Society of London Special Publication, London 194:141-152. Cesari S, C Parica, M Remesal & F Salani. 1999. Palaeoflora del Cretácico Inferior de peninsula Byers, islas Shetland del Sur, Antártida. Ameghiniana 36: 3-22. Dutra TL. 1997. Composição e história da vegetação do Cretáceo e Terciário da ilha Rei George, Península Antártica. PhD. Thesis. Instituto Tecnológico em Micropaleontologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brasil. Dutra Tl. 2001. Paleoflora da ilha 25 de Mayo, Península Antártica: Contribuição à paleogeografi a, paleoclima e para a evolução de Nothofagus. Asoc. Paleon. Argentina, Publicación Especial 8: 29-37. Dutra TL. 2005. Part II: Leaf assemblages .taphonomy, paleoclimatology and paleogeography). In: Landman NH & DS Jones .eds) Topics in Geobiology Applied Stratigraphy Koutsoukos: 194-198. Springer, Netherland. Dutra TL & JD Batten. 2000. Upper Cretacoeus floras of King George Islands, West Antarctica, and their palaeoenviromental and phytogepgraphic implications. Cretaceous Research 21: 181-209. Falcon-Lang Hj, DJ. Cantrill & Gj Nichols. 2001. Biodiversity and terrestrial ecology of a mid-Cretaceous, high-latitude floodplain, Alexander Island, Antarctica. Journal of the Geological Society 158: 709-724. Francis J. 1991. Palaeoclimatic significance of Cretaceous-early Tertiary fossil forests of the Antarctic Peninsula. In: Thomson MRA, A Crame & JW Thomson .eds) Geological Evolution of Antarctica: 623-627. Cambridge University Press, New York.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
175
Francis J & I Poole. 2002. Cretaceous and early Tertiary climates of Antarctica: Evidence from fossil wood. Palaeogeography, Palaeoclimatology, Palaeoecology 182: 47-64. Hathway B. 2000. Continental rift to back-arc basin: Jurassic–Cretaceous stratigraphical and structural evolution of the Larsen Basin, Antarctic Peninsula. Journal of the Geological Society 157: 417-32. Huber BT. 1998. Tropical paradise at Cretaceous poles? Science 282: 2199-2200. Leppe M, W Michea, C Muñoz, S Palma-Heldt & F Fernandoy. 2007. Paleobotany of Livington Island: The fi rst report of a Cretaceous fossil flora from Hannah Point. In: Cooper AK & CR Raymond .ed) Antarctica: A keystone in a changing world: 145 National Academies Press (USA). Poole I, Rj Hunt & DJ. Cantrill. 2001. A fossil wood flora from King George Island: Ecological implications for an Antarctic Eocene vegetation. Annals of Botany 88: 33-54. Poole I, Amw Mennega & DJ. Cantrill. 2003. Valdivian ecosystems in the late Cretaceous and early Tertiary of Antarctica as evidenced from fossil wood. Review of Palaeobotany and Palynology 124: 9-27. Povilauskas L, V Barreda & S Marenssi. 2008. Polen y esporas de la Formación La Irene .Maastrichtiano), sudoeste de la provincia de Santa Cruz, Argentina: primeros resultados. Geobios 41: 819-831. Romero EJ. 1978. Paleoecología y paleofitogeografía de las tafofloras del Cenofitico de Argentina y áreas vecinas. Ameghiniana 15: 209-226. Upchurch Jr, Bl Otto-Bliesner & C Scotese. 1998. Vegetation-atmosphere interactions and their role in global warming during the latest Cretaceous. Philosophical Transactions of the Royal Society of London B 353: 97-112.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
176
RESTOS VEGETALES PERMINERALIZADOS: UNA NUEVA VENTANA A LA VEGETACIÓN PASADO DE PATAGONIA Y ANTÁRTICA
(Permineralized plant debris: A novel window to the past vegetation of Patagonia and Antarctica)
Nishida, H.1,2, Uemura, K.3, Terada, K.4, Yabe, A.3, Asakawa, T.5, Yamada, T.6, Kurita, Y.7, Rancusi,
M.8, Hinojosa, L.F.9, and M. Leppe10 1Department of Biological Sciences, Chuo University, 1-13-27 Kasuga, Bunkyo, Tokyo, 112-8551 Japan. E-mail: [email protected] 2Graduate School of Science, University of Tokyo, Tokyo 113-0033, Japan 3National Museum of Nature and Science, Tokyo, 169-0073 Japan 4Fukui Prefectural Dinosaur Museum, Fukui, 911-8601 Japan 5Faculty of Science, Chiba University, Chiba, 263-8522 Japan 6Faculty of Science, Kanazawa University, Kanazawa, Japan 920-1192 7Faculty of Science, Niigata University, Niigata, 950-2181 Japan 8Colegio Compañia de Maria, Santiago, Chile 9Chile Facultad de Ciencias, Universidad de Chile, Santiago, Chile 10Instituto Antártico Chileno-INACH, Punta Arenas ABSTRACT Permineralized plant fossil assemblages that are preserved in certain terrestrial or shallow marine deposits are rich source of not only plants but also of other co-existed biota or even ecosystems of the past. Two major preservation types have historically provided biological information of remarkable quality and quantity. Calcareous concretions represented, for example, by Carboniferous coal-balls and Cretaceous Hokkaido nodules from Japan are the one, and the silicified peat layer such as the famous Devonian Rhynie Chert, Permian Homevale Chert of Australia, and Eocene Princton Chert of Canada are the other. All these fossil types provide novel aspects of past vegetation and environment in contrast to those based on fossils of other preservation types. In Patagonia or even in South America, such materials have not been found or studied much, excepting larger and more attractive fossils such as wood. This work summarizes recent results obtained from our field research in south Chile and in Antarctica, which is aimed to collect permineralized plant remains, particularly focusing on small debri preserved in calcareous concretions of shallow marine origin or in silicified peat of terrestrial origin. The first plant-containing calcareous concretion from Patagonia was found at Riesco Island in 2003. Various array of terrestrial plants and fungi have been identified within acetate-cellulose peel sections obtained from only three nodules possibly derived from the Paleocene Chorillo Chico Formation (Nishida et al., 2006). Most interestingly the assemblage consists of a large amount of tiny plant debris including epiphytic fungi, mosses, and epiphytic filmy fern, which are not expected to be preserved in usual depositional condition (Fig. 1). Similar concretions have been collected from Riesco Island and other localities in Magallanes region in our later field investigation. A certain amount of nodules are confirmed to have originated from the Late Cretaceous Dorotea Formation based on associated ammonites.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
177
Fig. 1. A-O. Plantas permineralizadas de la Formación Chorrillo Chico. A. Cormo de Bryophyta. L1, L2: apéndices foliares. B,C. Conidioma de Coelomyceta que contiene una conidiospora (flecha). D. Esporangio de Lophosoria. Puntas de flecha indican borde marginal típico al género. E. Rizoma de Hymenophyllacea. F. Rizoma de Trichomanes radicans Sw., especies actuales para la comparación. G, H. Retoño y hoja de Cupressacea. I, Retoño de Podocarpacea y la hoja tipo-1. J. Hoja tipo-2 de Podocarpacea. K, Posible retoño ovulifero de Pteridosperma. L. Semilla del K ampliada. M-O. Fruta angiospérmica en forma de estrella. M, N. Imágenes por TC. O. Sección delgada. Escalas: A, D, 100 m. B, G-K, M, 1 mm. E, F, 0,5 mm. Fig. 1. A-O. Permineralized plants from the Chorrillo Chico Formation. A. Bryophyta cormus. L1, L2: leafy appendages. B,C. Coelomycetes conidioma containing a conidiospore (arrow). D. Sporangium of Lophosoria. Arrowheads indicate typical spore marginal rim. E. Hymenophyllaceae rhizome. F. Rhizome of extant Trichomanes radicans Sw. for comparison. G, H. Cupressoid shoot and leaf. I, Podocarpaceous shoot and leaf type-1. J. Podocarpaceous lear type-2. K,L. Possible Pteridosperm ovuliferous shoot. M-O. Star-shaped angiosperm fruit. M,N. CT images. O. Thin section. Scales: A, D, 100 µm. B, G-K, M, 1 mm. E,F, 0.5 mm.
Attempts to find similar calcareous concretions of the same time-span in northern part of Chile became successful at Cocholgue, north of Tomé, Concepción Province. Well-preserved plant debri have been found from the Maastrichtian Quiriquina Formation, also yielding petrified wood. It has been identified by now a wood rotting fungus, an epiphytic lycopod strobilus; fern rhizomes and rachises, including those of dennstaedtiaceous ferns; various conifer wood and leaves; and angiosperm remains (Fig.2, A-I). The assemblage shows components of humid warm temperate mixed forest that do not exist in Chile now. In 2011, a silicified peat layer was found for the first time in South America at Cocholgue (Fig. 2, J-L).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
178
Fig. 2. A-I. Plantas permineralizadas de la Formación Quiriquina. A-C. Estróbilo de Lycopodium Sección Huperzia. D.
Conidioma de hongo destruyendo la madera. E,F. Rizoma y raquis del helecho Dennstaedtioideo. G, H. Retoño y hoja de cupresácea. I. Retoño de Araucaria. J, K. Turba silicificada de la Formación Navidad. L. Rizoma de gleicheniácea. M. Vegetación actual pantanosa de la isla de Chiloé mostrando Gleichenia y el bosque valdiviano detrás. Escalas 1 mm.
Fig. 2. A-I. Permineralized plants from the Quiriquina Formation. A-C. Strobilus of Lycopodium Section Huperzia. D. Conidioma of wood rotting fungus. E,F. Dennstaedtioid fern rhizome and rachis. G, H. Cupressoid conifer shoot and leaf. I.
Araucaria shoot. J, K. Silicified peat from the Navidad Formation. L. Gleicheniaceous rhizome. M. Present bog vegetation in Chiloe Island showing Gleichenia and Valdivian forest behind. Scales 1 mm.
H.N. and M.L. tried to find comparable concretions in Antarctica in 2011. Because of bad weather, only one nodule containing a piece of wood was available, leaving future possibility. The permineralized debri assemblage can successively provide anatomy-based rich paleobiological information.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
179
ACKNOWLEDGEMENTS This work is supported by the Grant-in-Aid for Scientific Survey nos. 18405013, 21405012 and 24570112 from MEXT Japan to HN, and has been realized with the courtesy of Consejo de Monumentos Nacionales de Chile. Instituto Antartico Chileno (INACH) logistically supported the field work in Antarctica. REFERENCES Nishida, H., Uemura, K., Terada, K., Yamada, T., Rancusi, M. H., and Hinojosa, LF. 2006. Preliminary report on permineralized plant remains possibly from the Paleocene Chorrillo Chico Formation, Magallanes Region, Chile. In: Nishida, H. (ed.) Post-Cretaceous Floristic Changes in Southern Patagonia, Chile. Chuo University, Tokyo, pp. 11-27.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
180
LAND VERTEBRATES BIOGEOGRAPHY OF WEST ANTARCTICA/SOUTH AMERICA AND THE FINAL BREAK-UP OF GONDWANA
(Biogeografía de vertebrados terrestres de Antártida oeste/Sudamérica y la ruptura final de Gondwana)
Reguero, M.A.1,3, Abello, M.A.2, Gelfo, J.N.3, López, G.M.4, Candela, A.M.3, Santillana, S.N.5 and Marenssi, S.A.6
1CONICET - Instituto Antártico Argentino, Balcarce 290, C1064AAF Buenos Aires, Argentina. [email protected] 2CONICET - LASBE, Museo de La Plata, Paseo del Bosque s/n B1900FWA, La Plata, Argentina. [email protected] 3CONICET - División Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n B1900FWA, La Plata, Argentina. [email protected], [email protected] 4División Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n B1900FWA, La Plata, Argentina. [email protected] 5Instituto Antártico Argentino, Balcarce 290, C1064AAF Buenos Aires, Argentina and Universidad de Buenos Aires. [email protected] 6CONICET- Instituto Antártico Argentino, Balcarce 290, C1064AAF Buenos Aires, Argentina and Universidad de Buenos Aires. [email protected] ABSTRACT This contribution is part of a major biogeographic study in progress that involves terrestrial vertebrates of the lapse Late Cretaceous-Early Paleogene from South America and Antarctica. This review involves biogeographic reconstruction, which draws upon geological and geophysical data, plate tectonic reconstructions, cladistic biogeography, and the evaluation of stratigraphically calibrated phylogenetic analyses to document ghost lineages of select taxa. INTRODUCTION One of the current tenets of Southern Hemisphere biogeography is the notion that the distribution of land vertebrates reflects vicariance due to the break-up of Gondwana (Late Cretaceous/Early Paleogene) involving Antarctic Peninsula and South America continents. Knowledge of the generic morphology, phylogeny and evolution of some groups of these land vertebrates, i.e. Paleogene marsupials and South American native ungulates (SANU), has increased over the past decade (e.g., Bond et al. 2006, 2011; Goin et al. 2006, 2007; Reguero et al. 2002; Reguero and Marenssi 2010). SANU (including “Condylarthra, Litopterna, Notoungulata, Astrapotheria and Pyrotheria orders) were diverse, geographically widespread, stratigraphically long lived and largely terrestrial: they therefore provide an almost ‘ideal’ case study in Cenozoic biogeography. Despite this progress, the SANUs still pose challenge to biogeographers; particularly determination of its centre of origin. West Antarctica (James Ross Basin, Antarctic Peninsula) Paleogene terrestrial mammal fauna exhibits high levels of species endemism, yet, for certain groups, is relatively species-poor. Also taxonomically is imbalanced in that it is not typically “South American” (Reguero and Marenssi 2010), as might be expected if most of its ancestors dispersed over a land bridge across the isthmus between Antarctic Peninsula and the southern tip of South America or used several intervening islands in a stepping-stone chain facilitated by past lower sea levels (3). Although a few groups appear to be stranded relicts, resulting from Antarctic Peninsula and South America shared plate tectonic history with Gondwana, for most clades current evidence indicates that the basal stocks arrived before that South America and the

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
181
Antarctic Peninsula begun their geographic isolation, fact that enhance the probability of a biogeographic interpretation of vicariance model for these organisms.
South American native ungulates and the final break-up of Gondwana The history of dispersal and evolution of several Antarctic land mammals is closely related to the existence of a land bridge (Weddellian) between peninsular Antarctica and South America during the Early Paleogene. Reguero et al. (2013) summarized the phyletic implications of the gondwanathere, the eight marsupials, the litoptern, and the two astrapotheres of the La Meseta fauna (Seymour Island, Antarctic Peninsula) and concluded that their presence can be best accounted for via numerous dispersals from South America during an interval that ranged from approximately Late Cretaceous to late Paleocene.
Figure 1. Stratigraphically calibrated phylogeny and known Paleogene occurrences of select genus of Astrapotheria from South America and West Antarctica.
Based on phylogeny calibrated stratigraphy of two basal SANUs, Litopterna and Astrapotheria Gelfo et al. (2013) suggest that the Weddellian isthmus could have been functional until the Selandian (Middle Paleocene). A minimum divergence date of the Antarctic and Patagonia litopterns (61-58.7 Ma) and astrapoteres (62-60 Ma) at which their common ancestor were present on both land masses is established as at least Middle Paleocene, approximately 5 Ma earlier than their documented occurrences (Fig. 1). The most diverse SANU group in South America, Notoungulata, provides further paleogeographic information. The absence of Notoungulata in the Paleogene of Antarctica and in the middle Paleocene of Patagonia (Peligran Land Mammal Age) suggests that their radiation occurred after the final break-up of both continents (Early Eocene). In South America land mammals reach their greatest early Paleogene diversity at sites often inferred to be early Eocene in Patagonia (Las Flores Formation) and Brazil (Itaboraí; Gelfo et al. 2009, Woodburne et al. 2013). As summarized by Reguero and Marenssi (2010), the La Meseta fauna from Seymour (Marambio) island, Antarctic Peninsula was dominated by marsupials and litopterns, in contrast to the typical South American Eocene faunas, which were dominated by notoungulates and basal condylarths. The

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
182
endemism shown of the marsupials and litopterns in West Antarctica suggests prior isolation; for example, the litoptern Notiolofos is known from deposits dating to 54 Ma (Mörs et al. 2013), whereas the rest of the La Meseta fauna dates to 49–47 Ma. We conclude that (1) throughout the Early Paleogene there was a continuous causeway through Antarctica and South America; (2) mid- to large-sized, obligate terrestrial forms (e.g., litoptern sparnotheriodontids and astrapothere astrapotheriids) gained broad distribution across West Antarctica land mass prior to fragmentation and were isolated before the end of the Paleocene period with the interruption of the Weddellian isthmus, and (3) recent, stratigraphically calibrated phylogenies including large, terrestrial ungulates (SANU) taxa of South America and Antarctic Peninsula reveal ghost lineages that extended into the middle Paleocene, approximately 5 Ma earlier than their documented occurrences. REFERENCIAS Bond, M.A., Reguero, M.A., Vizcaíno, S.F. and Marenssi, S.A. 2006. A new “South American ungulate” (Mammalia: Litopterna) from the Eocene of the Antarctic Peninsula. In: Francis, J.E., Pirrie, D. & Crame, J.A (Eds), Cretaceous-Tertiary high-latitude palaeoenvironments, James Ross Basin, Antarctica. Geological Society of London, Special Publications 258: 163-176. Bond, M., Kramarz, A., Macphee, R. and Reguero, M. 2011. A new astrapothere (Mammalia, Meridiungulata) from La Meseta Formation, Seymour (Marambio) Island, and a reassessment of previous records of Antarctic astrapotheres. American Museum Novitates, N°3718, 16 pp. Chornogubsky, L., Goin, F.J. and Reguero, M.A., 2009. A reassessment of Antarctic polydolopid marsupials (Middle Eocene, La Meseta Formation). Antarctic Science 21(3):285-297. Gelfo, J.N., Goin, F.J., Woodburne, M.O. and de Muizon C., 2009. Biochronological relationships of the earliest South American Paleogene mammalian faunas. Palaeontology 52(1):251–69. Gelfo, J.N., López, G.M., Abello, M.A., Reguero, M.A., Santillana, S.N. and Marenssi, S.A., 2013. The record of Paleogene ungulates and the demise of the South America-Antarctica land connection. Goin, F.J., Reguero, M.A., Pascual, R., Von Koenigswald, W., Woodburne, M.O., Case, J.A., Vieytes, C., Marenssi, S.A. and Vizcaíno, S.F. 2006. First gondwanatherian mammal from Antarctica. In: Francis, J.E., Pirrie, D. & Crame, J.A. (Eds), Cretaceous-Tertiary high-latitude palaeoenvironments, James Ross Basin, Antarctica. Geological Society of London, Special Publications 258:135-144. Goin, F.J., Zimicz, N., Reguero, M.A., Santillana, S.N., Marenssi, S.A. and Moly, J.J. 2007. New Mammal from the Eocene of Antarctica, and the origins of the Microbiotheria. Revista de la Asociación Geológica Argentina, 62(4):597-603. Mörs, T., Gelfo, J.N., Reguero, M., Lorente, M. and López, G.M., 2013. The oldest known (early Eocene) mammal from Antarctica. Society of Vertebrate Paleontology 73rd Annual Meeting, Los Angeles, CA, USA. Reguero, M.A., Marenssi, S.A. and Santillana, S.N., 2002. Antarctic Peninsula and Patagonia Paleogene terrestrial environments: biotic and biogeographic relationships. Palaeogeography, Palaeoclimatology, Palaeoecology, 179:189-210. Reguero, M.A. and Marenssi, S.A., 2010. Paleogene climatic and biotic events in the terrestrial record of the Antarctic Peninsula: an overview. In: Madden, R., Carlini, A.A., Vucetich, M.G. and Kay, R. (Eds.). The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia. Cambridge University Press, pp. 383–97 Reguero M.A., Goin, F., Hospitaleche, C.A., Dutra, T. and Marenssi, S.A., 2013. Late Cretaceous/Paleogene West Antarctica Terrestrial Biota and Its Intercontinental Affinities. Dordrecht: Springer. 120 pp. Woodburne, M.O., Goin, F.J., Bond, M., Carlini, A.A., Gelfo, J.N., et al., 2013. Paleogene land mammal faunas of South America; a response to global climatic changes and indigenous floral diversity. J. Mamm. Evol. doi:10.1007/s10914-012-9222-1.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
183
REGISTRO DE LOS PLESIOSAURIOS MÁS AUSTRALES DE AMÉRICA: SISTEMÁTICA, PALEOAMBIENTE Y ESTRATIGRAFÍA.
(Record of plesiosaurs southernmost of America: Systematics, paleoenvironment and stratigraphy.)
Ortiz, H.1, Mansilla, H.2, Manríquez, L.3 y M. Leppe2
1Universidad de Concepción, Facultad de Ciencias Naturales y Oceanográficas, Concepción, Chile. [email protected] 2Laboratorio de Paleobiología de Patagonia y Antártica, Departamento Científico, Instituto Antártico Chileno. 3Departamento Ciencias de la Tierra, Universidad de Concepción, Concepción, Chile. INTRODUCCION Los plesiosaurios (Reptilia: Sauropterygia) se originaron entre el Triásico Superior hasta finales del límite K-T (Cretácico-Terciario), abarcando todos los continentes (Ketchum y Benson 2010). En el extremo suroeste de la Patagonia durante el Cretácico, se han registrado exclusivamente hallazgos de plesiosaurioideos (Gasparini et al. 2007), estando su distribución poco representada. Los primeros comentarios sobre la presencia de plesiosaurios, corresponden a restos mal asignados a Coelospondylus (Plesiosaurus) chilensis por Cecioni (1957) en el área del Río Don Guillermo (Puerto Natales). La errónea identificación, se debe a la utilización de caracteres morfológicos no diagnósticos para determinar la especie (Otero et al. 2010). Gasparini (1979) comenta sobre la presencia de plesiosaurios para el norte de Laguna Parrillar, Península de Brunswick. El 2009 se asigna la familia Elasmosauridae a un bloque rodado obtenido de la formación Dorotea, Puerto Natales (Otero et al. 2009). En el año 2011, se conoce la presencia de un plesiosaurio juvenil, en la localdad de Dumestre, Puerto Natales (Stinnesbeck, 2012 com. pers.). Finalmente (Ortiz et al. 2012) el infra-orden Plesioauria, en los estratos que rodean la Laguna Parrillar. El objetivo de este trabajo es identificar y asignar el nivel taxonómico (Familia o Genero) a los restos de plesiosaurios encontrados en la Reserva Nacional Laguna Parrillar e interpretar las asociaciones fosilíferas incluidas en los estratos que los acompañan; determinando la formación a los que pertenecen, y estableciendo su contexto paleoambiental y paleogeográfico. LOCALIDAD Y MARCO GEOLÓGICO La Reserva Nacional Laguna Parrillar está ubicada al centro de la península de Brunswick, 52 km. al sur de la ciudad de Punta Arenas, Chile (Figura 1). El área de estudio corresponde a 305m de afloramiento ubicado al costado noroeste de la Laguna Parrillar. Este consta de series estratificadas que corresponden a secuencias sedimentarias marinas y parálicas hacia el final del Cretácico Superior. A partir de la correlación de los estudios de Hollister (1943) y Farfán (1994), se puede inferir que la zona de estudio, específicamente donde se realizó la columna estratigráfica, correspondería a la parte final de la Formación Rocallosa (Figura 3). SISTEMATICA PALEONTOLÓGICA Reptilia Laurenti, 1768. Diapsida Osborn, 1903. Sauropterygia Owen, 1860.
Infra-Orden Plesiosauria de Blanville, 1835. Familia Elasmosauridae? Cope, 1969. Material: Figura 2, Los restos postcraneales corresponden a fragmentos del esqueleto paraxial (cinturas y extremidades incompletas incluidas en un bloque de arenisca fina calcárea de color gris, almacenado

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
184
en la Colección Paleontología de Antártica y Patagonia del Instituto Antartico Chileno como CPAP-0008. El estrato portador de los huesos fue asignado al tramo CLP-012 de la figura 3. Extremidad anterior derecha: Se conservaron huesos podiales y epipodiales compuestos por: 11 falanges articuladas, se diferencian 4 de 5 dígitos, dos de los 5 ulnare y 2 metacarpos sin identificación. Cintura pectoral: Se conservó la escápula izquierda incompleta. Se ha perdido parte de los bordes de la articulación escápula-coracoidea y la faceta glenoidea, ubicado en la parte proximal de la sínfisis de la escápula con el humero y coracoides. Desde la zona glenoidea hacia el proceso ventral y dorsal de la escápula, se diferencia una prolongación hacia la parte anterior generando una concavidad hacia la zona ventral.
Figura 1: Ubicación del área de estudio, indicando el centro de la península de Brunswick y al costado derecho la Laguna
parrillar y los cerros plegados que la rodean. Imagen obtenida a partir de la combinación de bandas 432 en ArcGis y Google maps.
Figure 1: Location of the study area, indicating the center of the Brunswick Peninsula and Laguna grilling right side and folded hills that surround it. Image obtained from the 432 band combination in ArcGIS and Google maps.
Comentarios: El ejemplar CPAP-0008 se asigna a la familia Elasmosauridae hasta nuevos hallazgos. A partir del análisis morfológico de las muestras de plesiosaurio y la comparación con la literatura, se postula el nivel taxonómico a que pertenece el ejemplar descrito es Familia Elasmosauridae?. El signo de interrogación (?) es producto de la ausencia de caracteres diagnósticos en una escápula que permitan confirmar una familia. La comparación a partir de una reconstrucción de la escápula CPAP-0008 con los elasmosaurios Alzadasaurus pumbertoni Welles 1949, Leurospondylus ultimus Brown 1913, Hydroterosaurus alexandrae Welles 1943 & Muraenosaurus stocki Welles 1943 y Polycotilidos Dolichorhyncus osborni Williston 1925, Leptocleidus Superstus Andrews 1922 y Eurycleidus Arcuatus Andrews 1922 del Campaniano Superior-Maastrichtiano arroja una mayor afinidad morfológica con la familia Elasmosauridae; sumado a que no existen registros de Polycotilydae en el extremo sur de América. Palinomorfos: Las asociaciones esporopolinicas encontradas en este estudio, son principalmente Pteridophytas (Gleicheniaceae, Lycopodiaceae, Osmundaceae, Schizaeaceae), seguida de polen de angiospermas como proteáceas. Los niveles fértiles donde se obtiene la presencia de palinomorfos, corresponden a LP-04 y LP-Bones (Figura 2). Las muestras palinológicas permanecen guardadas en la Palinoteca de la Colección Paleontológica de Antártica y Patagonia.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
185
Figura 2: CPAP-0008, Muestra de plesiosaurio indicando las falanges Ph; escapula Sc; Ulnares Uli y Ulii; metacarpos
indeterminados ib e ic. Figure 2: CPAP-0008, indicating plesiosaur Sample Ph phalanges, scapula Sc; ulnar Uli and Ulii; pasterns indeterminate ib
and ic. DISCUSIÓN Y CONCLUSIÓN El registro de la familia Elasmosauridae en la Reserva Nacional Laguna Parrillar son el registro más austral de América. La asignación estratigráfica de la formación Rocallosa está sujeta a errores, estos productos de la diferencia en la escala ya que Farfán (1994) trabaja con una escala regional en comparación con este estudio. El patrón de distribución de los plesiosaurios reafirma una conexión entre los mares del protopacífico y protoweddel hacia finales del Cretácico. La conexión entre la cuenca de Magallanes y James Ross, con el evento transgresivo de la Formación Quiriquina durante el Maastrichtiano, indican que la presencia de organismos similares en lugares separados geográficamente, fue producto del estrecho vínculo entre Sudamérica y Antártica. Las asociaciones fosilíferas y la estratigrafía del área de estudio, presentan características propias de un ambiente marino parálico, marginal, que estaría indicando la fase de somerización de la cuenca, a diferencia de lo que se encuentra en los niveles más basales asociados con condiciones más profundas. La alta presencia de esporas de Pteridofitas en relación al polen de angiospermas, indicaría un clima templado frio a cálido, con alta humedad; muy diferente a lo que es en la actualidad. Para la reconstrucción paleoambiental a partir de datos cuantitativos, se necesita un análisis palinológico más detallado, en donde los elementos florísticos nos entregarían a partir de la bioestratigrafía herramientas que nos permitan entender la evolución de la flora Austral-Antártica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
186
Figura 3: Correlación creada de la formación Rocallosa, indicando la litología, tramo, estructuras de sedimentación y fósiles.
En la zona izquierda de la imagen está ubicada la leyenda y la simbología. (Modificado de Farfán (1994) y los datos obtenidos en terreno.
Figure 3: Correlation Rocallosa formation created, showing the lithology, sedimentation structures and fossils. On the left of the image is located legend and symbology. (Modified from Farfán (1994) and field data. REFERENCIAS Cecioni, G. 1957. Etá dela flora de Cerro Guido e stratifrafia del Departamento Ultima Esperanza, provinvia di Magellano, Cile. Estratto dal Bollettino dela Societa Geologica Italiana. Vol 76:1-18. Chattereje, S. & Small, B. J. 1989. New plesiosaurs from the Upper Cretaceous of Antarctica; pp. 197-215 in J. A. Crame (ed.), Origins and Evolution of the Antarctic Biota. Special Publication of the Geological Society 47. Welles, S.P. 1952. A review of the North American Cretaceous elasmosaurs. University of California Publications in Geological Sciences. 29.46-144. O’Keefe, F. 2001. A Cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zool. Fennica 213: 1-63. Farfán, L. 1994. Geología estructural de la Península de Brunswick. Tesis para optar al Título de Geólogo, Universidad de Chile, Facultad de Ciencias Físicas y Matemáticas, Departamento de Geología, Santiago.143pp. Gasparini, Z., Fernández, M., de la Fuente, M. y Salgado, L. 2007. Reptiles marinos jurásicos y cretácicos de la Patagonia argentina: su aporte al conocimiento de la herpetofauna mesozoica. Asociación Paleontológica Argentina. Publicación Especial 11 Ameghiniana 50° aniversario: 125-136. Hollister, J. S., 1943. Preliminary report of three structures on the Brunswick Peninsula.Informe inédito, Archivo Técnico ENAP. Ketchum, H.F. & Benson, R.J.B.. 2010. Global interrelationships of plesioauria (Reptilia: Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses. Biological Reviews, 85, 361-392. Ortiz, H., Mansilla, H., Jujihara, T. & Leppe, M. 2012. Primer reporte de Plesiosauria (Reptilia: Sauropterygia) en estratos del Cretácico Superior en la Reserva Nacional Laguna Parrillar, Península de Brunswick, Región de Magallanes y Antártica Chilena. Libro de resúmenes II Simposio de Paleontología en Chile: 135-138. Otero, R., Suarez, M. & J. Le Roux. 2009. First record of Elasmosaurid Plesiosaurs (Sauropterygia: Plesiosauria) in upper levels of the Dorotea Formation, Late Cretaceous (Maastrichtian), Puerto Natales, Chilean Patagonia. Adean Geology 36 (2): 342-350. Otero, R.A., Soto-Acuña, S. & Rubilar-Rogers, D. 2010. Presence of Mauisaurus Hector (Plesiosauroidea, Elasmosauridae) in the Late Cretaceous (Maastrichtian) of central Chile. Acta Paleontologica Polonica 55, 361-364. Otero, R., Parham, J., Soto-Auña, S., Jimenez-Huidobro, P. & Rubilar-Rogers, D.2012. Marine Reptiles from Late Cretaceous (early Maastrichtian) deposits in Algarrobo, central Chile. Cretaceous Reseach 35: 124-132. Gasparini, Z. 1979. Comentarios críticos sobre los vertebrados mesozoicos de Chile. In Congreso Geológico Chileno, No 2, Actas 3: H15-H32. Arica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
187
VARIABILIDAD TEMPORAL DE MASAS DE AGUA EN EL MAR DE LA FLOTA
(Temporal variability of water masses in Bransfield Strait)
Ruiz Barlett, E.M.1, Tosonotto, G.V.1 y A.R. Piola2
1 Instituto Antártico Argentino, Balcarce 290, Ciudad de Buenos Aires, Argentina. E-mail: [email protected] 2 Servicio de Hidrografía Naval, Av. Montes de Oca 2124, Ciudad de Buenos Aires, Argentina. INTRODUCCION En el Mar de La Flota (MF) o Estrecho de Bransfield, que separa el archipiélago de las Islas Shetland del Sur de la Península Antártica (PA), se encuentran masas de agua de diferentes orígenes y características, influenciadas por la variabilidad temporal del clima en distintas escalas. Tres cuencas profundas, llamadas Cuenca Occidental (CO), Cuenca Central (CC) y Cuenca Oriental (CE), conforman las regiones más profundas en el MF. Las cuencas se profundizan hacia el noreste, siendo la máxima profundidad en la CO de ~1400 m, en la CC de ~2000 m y en la CE de ~2800 m. La CC está aislada de las otras dos por umbrales de ~1100 m. Al ser un mar semi-cerrado, donde el intercambio con las aguas circundantes es restringido, es particularmente propicio para analizar la variabilidad temporal de las masas de agua. El tiempo de residencia de las aguas del MF es del orden de los 30 años (Schlosser et al., 1988) mientras que el de las aguas de plataforma en el Mar de Weddell (MW) occidental es menor que un año (Hellmer et al., 2011). Estos tiempos extremadamente cortos para los estándares del océano profundo, permiten analizar la variabilidad temporal en una escala de tiempo mensurable (Wilson et al., 1999). Diversos trabajos realizados en el MF estudiaron la variabilidad de las propiedades hidrográficas en diferentes escalas temporales (Wilson et al., 1999; García y Mata, 2005 y otros). El clima de la PA Occidental registra el cambio más rápido observado en el Hemisferio Sur, a una tasa de (3.7 ± 1.6) ºC.siglo-1 (Vaughan et al., 2003). El calentamiento ha tenido profundas consecuencias en las especies marinas (Meredith y King, 2005) y para el campo de hielo de la PA, donde la mayoría de los glaciares (Cook et al., 2005) y la extensión de hielo marino (Meredith y King, 2005) han sufrido retrocesos durante los últimos 50 años. En este trabajo se analizará la variabilidad temporal de las propiedades termohalinas en las aguas de las cuencas profundas del MF, con el objetivo de observar variaciones interanuales asociadas a cambios en las fuentes de aguas y su posible relación con el cambio climático. MATERIALES Y METODOS Para analizar la variabilidad temporal de la temperatura del aire, el índice SAM y la extensión meridional del hielo marino, se utilizaron series temporales completas de medias mensuales entre 1980 y 2008. Los datos de temperatura del aire en las estaciones Belligshausen, Esperanza, Faraday y Marambio, se obtuvieron del British Antarctic Survey (BAS) y del Servicio Meteorológico Nacional (SMN); los datos de extensión meridional del hielo marino, de la base de datos del Australian Antarctic Data Centre (AADC) y los del índice del Modo Anular de Variabilidad del Hemisferio Sur (SAM; Southern Annular Mode), se hallan disponibles en la página de Internet del State Key Laboratory of Numerical Modeling for Atmospheric Sciences and Geophysical Fluid Dynamics (LASG). Los datos de temperatura y salinidad del agua se obtuvieron de la base histórica del National Oceanographic Data Center (NODC) y, por gentileza de M. Mata del Grupo de Oceanografía de Altas Latitudes (GOAL), se agregaron estaciones de las campañas BIOANTAR, ECOANTAR, GOAL 2003 y GOAL 2004. Se utilizaron datos de muestras discretas (entre 1926 y 2005) y casi continuas (CTDs, entre 1975 y 2004). El análisis de las masas de agua se restringe a profundidades mayores que 800 m (CO) y

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
188
1100 m (CC y CE). Para analizar la variabilidad temporal, los perfiles se agruparon en dos niveles (profundo y de fondo), de acuerdo a la profundidad de cada cuenca (los rangos utilizados se muestran en la Tabla 1). Las medias anuales de temperatura potencial y salinidad, se calcularon promediando las medias mensuales de los perfiles verticales obtenidos entre noviembre y enero (NDE). RESULTADOS Y DISCUSION Las medias anuales de temperatura potencial y salinidad muestran variabilidad interanual en las cuencas profundas del MF. Se aplicó una correlación lineal a los promedios anuales de CTDs de cada cuenca entre diciembre de 1980 y enero de 2004 (donde se encuentran la mayoría de los datos disponibles). Los valores medios, las tendencias y los coeficientes de correlación respectivos de las propiedades termohalinas para los 24 años estudiados, se presentan en la Tabla 1 (en negrita los valores significativos al nivel de confianza del 90%).
N° períodos
anuales Media (°C) (°C.año-1) r² Media (ups) (ups.año-1) r²
800-1000 6 -0.587 ± 0.098 0.0489 0.57 34.565 ± 0.009 0.0007 0.23
1000-1200 4 -0.663 ± 0.075 0.0282 0.41 34.557 ± 0.003 -0.0009 0.40
1100-1450 12 -1.570 ± 0.034 0.0015 0.02 34.565 ± 0.003 -0.0017 0.42
1450-1800 9 -1.608 ± 0.022 0.0022 0.08 34.572 ± 0.004 -0.0020 0.45
1100-1600 6 -1.094 ± 0.031 0.0033 0.16 34.558 ± 0.002 -0.0017 0.50
1600-2100 5 -1.144 ± 0.026 0.0061 0.37 34.562 ± 0.001 -0.0015 0.52
c/ 5 años
1100-1450 5 0.0072 0.03 -0.0070 0.851450-1800 5 0.0102 0.17 -0.0097 0.57
Central
Oriental
SalinidadTpot-0 Cuenca Rango (m)
Occidental
Central
Tabla 1. Medias, tendencias lineales y coeficientes de correlación de temperatura potencial y salinidad de las cuencas, para el
período NDE entre 1980 y 2004. En negrita los valores significativamente diferentes de cero a un α = 0.1 Table 1. Mean, linear trend and correlation coefficient of potential temperature and salinity of the basin, for NDJ between
1980 and 2004 period. In bold values significantly different from zero at α = 0.1. En las tres cuencas, se registró un aumento de temperatura y la CO mostró además un leve incremento de salinidad para profundidades menores que 1000 m, posiblemente asociados al calentamiento y la salinización que se han registrado en el CDW (Von Gyldenfeldt et al., 2002). Los gradientes verticales en las aguas más profundas que 1000 m en la CO difieren un orden de magnitud en comparación a los de las otras cuencas (Tabla 1). Esto podría explicarse por la mezcla de Agua Profunda Circumpolar modificada (mCDW) con Agua Profunda Cálida modificada (mWDW), lo cual resultaría en la intensificación del calentamiento y atenuación en la dilución. Las aguas dentro del nivel profundo en las CC y CE mostraron la misma tendencia significativa en decrecimiento de la salinidad (Tabla 1). La CC mostró una tendencia de dilución mayor en el nivel de fondo debido al aporte de Agua de Plataforma de Alta Salinidad (HSSW), que va disminuyendo su salinidad en las regiones de origen por la incorporación de agua de fusión. Los resultados en la CC son similares a los obtenidos por García y Mata (2005). La dilución en las aguas de plataforma, está asociada al desplazamiento del hielo marino hacia el sur y al aumento de precipitaciones sobre la plataforma continental del MW occidental, causados por el calentamiento climático en la PA desde mediados del siglo pasado (Hellmer et al., 2011). Un posible incremento en la contribución de Aguas de Plataforma de Baja Salinidad en las aguas de plataforma que se forman en el MW, también podría ser la causa de la dilución observada. El tiempo de residencia estimado para las aguas en las cuencas del MF es de 30 años (Schlosser et al., 1988). Así, las aguas más densas que ocupan el fondo de la CC, son las más antiguas. Los nuevos

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
189
aportes de HSSW que muestran la dilución de las aguas de plataforma, ocupan profundidades menores dentro de la cuenca, hasta que la mezcla posterior vaya modificando las propiedades de las aguas hasta el fondo. El calentamiento y la dilución fueron observados en la columna de agua de la CC a lo largo de los 24 años de análisis y, además, la pendiente en los perfiles verticales se ha intensificado a lo largo del mismo período (observado entre 1963 y 1995 por Wilson et al., 1999), lo que justificaría esta hipótesis. La temperatura del aire promediadas las cuatro estaciones meteorológicas (entre junio y agosto, JJA) presentó un incremento de 0.92 °C y el límite norte de extensión de hielo marino en 58 °W (entre julio y septiembre, JAS) experimentó un retroceso hacia el sur de 1.1 grados, entre los inviernos de 1980 a 2008. El coeficiente de correlación entre ambas variables fue significativo (r2 = 0.77). La variabilidad interanual de estos parámetros muestra un patrón cíclico en la extensión y duración, con períodos de mayor extensión cada cuatro a seis años, como ha sido señalado por algunos autores (Murphy et al., 1995). Las propiedades termohalinas en las cuencas (NDE) fueron correlacionadas con la temperatura del aire (JJA), la extensión de hielo marino (JAS) y el índice SAM (JJA), para el período 1980-2004. Los resultados obtenidos muestran correlaciones positivas y altas (y en general significativas) entre la temperatura del aire y la extensión de hielo marino con la temperatura del agua (Tabla 2). Los bajos valores en la correlación entre la temperatura potencial y la salinidad, sugieren que las variaciones observadas en salinidad, se han producido en las regiones donde se forman las masas de agua que llegan a las cuencas del MF.
Cuenca Rango (m) θ vs. S θ vs. Tair θ vs. HM θ vs. SAM S vs. Tair S vs. HM S vs. SAM
800-1000 0.63 0.77 0.91 0.85 0.81 0.79 0.421000-1200 0.39 0.48 0.58 0.77 -0.49 -0.46 -0.18
1100-1450 -0.26 0.62 0.69 0.36 -0.09 -0.06 -0.311450-1800 -0.27 0.74 0.73 0.57 -0.04 0.10 -0.52
1100-1600 0.43 0.75 0.74 0.57 0.30 0.23 0.101600-2100 0.19 0.84 0.91 0.53 0.30 0.31 -0.24
Occidental
Central
Oriental
Tabla 2. Coeficientes de correlación entre las propiedades termohalinas (NDE), temperatura del aire (JJA), extensión de hielo marino (JAS) e índice SAM (JJA), para el período 1980-2004. En negrita los valores significativamente diferentes de cero a
un α = 0.1 Table 2. Correlation coefficient between thermohaline properties (NDJ), air temperature (JJA), sea ice extent (JAS) and SAM
index (JJA), for 1980-2004 period. In bold values significantly different zero to α = 0.1. La variabilidad interanual en las aguas profundas de la CC está controlada por la presencia del HSSW y su variabilidad dentro de la cuenca. Las aguas profundas y de fondo en esta cuenca, presentaron períodos de calentamiento seguidos de enfriamiento, con ciclos de 4 a 6 años entre 1980 y 2004. Los mínimos de temperatura del aire y máximos en extensión de hielo marino hacia el norte se asocian a los mínimos de temperatura potencial observados esos años en las cuencas. Esta oscilación parece estar relacionada con la Onda Circumpolar Antártica (ACW), y por lo tanto, con las variaciones de temperatura del aire y la extensión de hielo marino. Los cambios en la temperatura del aire afectan la cobertura de hielo marino en el MW, produciendo cambios en la salinidad de las aguas de plataforma, con variaciones en períodos interanuales o más largos. La salinidad presentó anomalías positivas aproximadamente cada 5 años, a medida que se produjo la dilución de las aguas profundas en la CC. Anomalías positivas en el SAM, están relacionadas con la producción de aguas de plataforma en el MW (Timmerman et al., 2002). La tendencia positiva observada entre 1980 y 2008, si bien no es significativa, parecería estar relacionada con el calentamiento y la dilución de las aguas en las cuencas profundas. Sin embargo, los coeficientes de correlación entre el índice SAM y los parámetros termohalinos, fueron en general bajos y no significativos (Tabla 2). No se dispone de datos suficientes,

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
190
principalmente en las CO y CE, para aceptar o descartar la mayor influencia del SAM o de la ACW en forma definitiva. Dado el escaso número de observaciones de las aguas profundas y de fondo, estos resultados deben considerarse con precaución. CONCLUSIONES Los cambios en las propiedades termohalinas observados en las cuencas para el período 1980-2004, están asociados a modificaciones en las aguas circundantes al MF. Estos cambios podrían deberse al cambio climático. Las tres cuencas presentaron calentamiento y dilución (excepto las aguas más profundas de la CO, que mostraron salinización). Las variaciones en la CO fueron diferentes respecto a las otras cuencas por influencia del mCDW. La dilución más pronunciada, se observó en la CC por la influencia del HSSW. Las aguas en la CC, no sólo han presentado tendencias en los promedios anuales de las propiedades termohalinas, sino también cambios en la estructura vertical. La correlación de la temperatura del aire (JJA) con la extensión latitudinal de hielo marino (JAS) y con la temperatura del agua por debajo de los umbrales en las cuencas (NDE) para los 24 años estudiados, fue significativa. Por lo tanto, las variaciones interanuales observadas (en períodos de 4 a 6 años) parecen estar vinculadas a modos de variabilidad anual o de mayor período como la ACW. Los resultados obtenidos para la CC parecen ser más convincentes debido a la mayor resolución interanual de los datos disponibles. Una continuación en la realización de mediciones de las propiedades oceanográficas y la utilización de fondeos, es recomendable para poder confirmar la presencia de tendencias significativas y ondas interanuales en las cuencas del MF. AGRADECIMIENTOS Este estudio contó con datos provenientes del GOAL (Brasil) a través de M. Mata (FURG). REFERENCIAS BIBLIOGRAFICAS Cook, A.J., Fox, A.J., Vaughan, D.G. y Ferrigno, J.G. 2005. Retreating glacier fronts on the Antarctic Peninsula over the past half-century. Science, 308: 541-544. García, C.A.E y Mata, M.M. 2005. Deep and bottom water variability in the central basin of Bransfield Strait (Antarctica) over the 1980-2005 period. CLIVAR Exchanges, 10: 48-50. Hellmer, H.H., Huhn, O., Gomis, D. y Timmermann, R. 2011. On the freshening of the northwestern Weddell Sea continental shelf. Ocean Science, 7: 305-316. Meredith, M.P. y King, J.C. 2005. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophysical Research Letters, 32: L19604. Murphy, E.J., Clarke, A., Symon, C. y Priddle, J. 1995. Temporal variation in Antarctic sea-ice: analysis of long term fast-ice record from the South Orkney Islands. Deep-Sea Research, 42: 1045-1062. Schlosser, P., Suess, E., Bayer, R. y Rhein, M. 1988. 3He in the Bransfield Strait waters: indication for local injection from back-arc rifting. Deep-Sea Research, 34: 1919-1935. Timmerman, R., Hellmer, H.H. y Beckmann, A. 2002. Simulations of ice-ocean dynamics in the Weddell Sea Part II: interannual variability 1985-1993. Journal of Geophysical Research, 107: 101029/2000JC00742. Vaughan, D.G.; Marshall, G.J.; Connelley, W.M.; Parkinson, C.; Mulvaney, R.; Hodgson, D.A.; King, J.C.; Pudsey, C.J. y Turner, J. 2003. Recent rapid regional climate warming on the Antarctic Peninsula. Climate Change, 60: 243-274. Von Gyldenfeldt, A.-B., Fahrbach, E., García, M.A. y Shröder, M. 2002. Flow variability at the tip of the Antarctic Peninsula. Deep-Sea Research II, 49: 4743-4766. Wilson, C., Klinkhammer, G.P. y Chin, C.S. 1999. Hydrography within the Central and East Basin of the Bransfield Strait, Antarctica. Journal of Physical Oceanography: 465-479.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
191
VARIAÇÃO DOS DADOS HIDROQUÍMICOS E BIOLÓGICOS EM ÁGUAS DE SUPERFÍCIE DO ESTREITO DE BRANSFIELD (ANTÁRTICA), CONSIDERANDO DOIS DIFERENTES
ESTÁGIOS DO PERÍODO DE VERÃO AUSTRAL EM 2011 (Variation of hydrochemical and biological data in Bransfield Strait surface waters (Antarctica),
considering two different stages of austral summer period in 2011)
Bastos, A.T.C.C.1, Chiozzini, V.G.1, Scigliano, B.F.1 y E.S. Braga1 1Departamento de Oceanografia Física, Química e Geológica, Instituto Oceanográfico, Universidade de São Paulo, Praça do Oceanográfico, 191, São Paulo, SP, Brasil. E-mail: [email protected]; [email protected] INTRODUÇÃO O Oceano Austral se caracteriza como uma área HNLC (alta concentração de nutrientes e baixo teor de clorofila). A baixa produtividade primária pode ser atribuída a diversos fatores como: disponibilidade de micronutrientes, ao fotoperíodo, as baixas temperaturas, grazing, afundamento, dispersão por mistura vertical e outros processos advectivos. Entretanto, maiores estoques de fitoplâncton tem sido encontrados em áreas marginais ao gelo e em águas costeiras protegidas (Castro et al., 2002). O Estreito de Bransfield pode ser definido como uma zona onde há presença de águas de transição, ocupada majoritariamente por massas de água cujas propriedades são influenciadas pelos fluxos vindos dos mares adjacentes como o Mar de Bellinghausen, com águas relativamente mais quentes e menos salinas, e o Mar de Weddell, com águas mais frias e salinas (García et al., 2002). Diferenças sazonais interanuais na concentração de nutrientes e no teor de clorofila-a tem sido observadas no Oceano Austral, devido a dois fatores principais: a intensificação das características de inverno entre um período e outro e, a própria variação no verão austral, devido aos diferentes estágios de evolução da biota somada à hidrodinâmica no Estreito considerando o início e final do período, como é evidenciado neste trabalho. Este trabalho tem como objetivo avaliar a variação dos parâmetros hidroquímicos nas águas de superfície do Estreito de Bransfield em dois períodos distintos do verão austral, ou seja, março de 2011 (final de verão) e outubro de 2011 (início de verão). MATERIAL E MÉTODOS As amostras de água de superfície foram coletadas em 49 estações em março de 2011 e, em 17 estações em outubro de 2011, durante as campanhas OPERANTAR XXIX e XXX, respectivamente, a bordo do Navio de Apoio Oceanográfico Ary Rongel da Marinha do Brasil, no Estreito de Bransfield (Fig. 1), como parte do Projeto CARBO-THORIUM (CNPq) pertencente ao Programa Antártico Brasileiro (PROANTAR). A água foi coletada com auxílio de garrafas coletoras tipo Niskin, a temperatura da água foi obtida com termômetro de reversão protegido 2011. A salinidade foi determinada em salinômetro indutivo da marca Beckman, Mod RS-10. Os teores de fosfato e silicato foram determinados segundo o método descrito por Grasshoff et al. (1983). A clorofila-a, o material particulado em suspensão e a matéria orgânica em suspensão foram obtidos segundo a metodologia de Strickland & Parsons (1968).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
192
Figura 1 – Localização das estações de coleta de água de superfície no Estreito de Bransfield: a) OPERANTAR XXIX e b) OPERANTAR XXX.
Figure 1. Location of sampling stations for surface water in the Bransfield Strait: a) OPERANTAR XXIX and b) OPERANTAR XXX.
RESULTADOS E DISCUSSÃO Em março de 2011, a porção sul do Estreito apresentou temperaturas mais baixas, menores que 0,4°C, salinidades entre 34,15 a 34,50 e valores mais altos de silicato entre 44,0 µmol L-1 a 54,0 µmol L-1, mostrando uma possível intrusão de águas provenientes do Mar de Weddell; o fosfato variou de 1,20 µmol L-1 a 1,80 µmol L-1, o nitrato esteve entre 11,45 µmol L-1 e 30,85 µmol L-1 e a clorofila-a apresentou valores muito baixos, variando de concentrações menores que o limite de detecção do método a 4 mg m-3, indicando uma baixa produção primária, diminuindo a remoção de nutrientes. Em direção ao norte do Estreito, observou-se um aumento da temperatura e diminuição da salinidade associado à diminuição da concentração de fosfato e silicato e um aumento na clorofila-a, atingindo um valor de 10,0 mg m-3. No quadro geral, o material particulado em suspensão mostrou uma variação entre 13,40 mg L-1 na estação 51 e 32,00 mg L-1 na estação 21; a fração orgânica do material particulado em suspensão apresentou valores entre 2,9 mg.L-1 na estação 21 e 8,8 mg L-1 na estação 51. A maior fração de material orgânico (31%) foi observada em Março de 2011 na estão 13, em período onde foi estimada a maior produção primária em função dos valores de clorofila- a. Ainda na campanha de março de 2011 (no final do verão austral) foi possível observar a presença de águas do Mar de Bellingshausen, caracterizada por temperaturas positivas (de 0°C a 2°C) e salinidade variando de 33,00 a 34,20, e águas do Mar de Weddell, caracterizadas por temperaturas negativas e salinidades entre 34,1 e 34,6 (García et al. , 2002) (Fig. 2). As águas do Mar de Bellingshausen entram no Estreito através do Estreito de Gerlache, fazendo uma frente junto as Ilhas Shetlands do Sul (bacia ocidental), já as águas do Mar de Weddel, entram através do estreito de Boyd, permanecendo na bacia oriental. Na Bacia Central do Estreito observou-se a presença de um giro anticiclônico formando uma frente junto a Península Antártica como observado por Sangrá et al., (2011). Em outubro de 2011, os valores de temperatura superficial da água variaram de -2,1 °C a 0,6°C, característicos do começo do verão antártico. As maiores temperaturas foram encontradas na estação 1, mais ao norte, próxima à Passagem de Drake e na porção sul do Estreito, enquanto que os menores valores foram observados na região central do Estreito. A salinidade variou de 33,45 a 34,16; com distribuição semelhante a da temperatura: salinidades maiores nas estações mais ao norte e ao sul e menores na parte central do Estreito, sob maior influência do degelo. Os teores de silicato dissolvido variaram de 31,52 µmol L-1 a 86,95 µmol L-1, o nitrato apresentou valores entre 29,80 µmol L-1 e 8,80 µmol L-1. As concentrações de material particulado em suspensão (MPS) variaram de 12,40 mg L-1a 23,11 mg L-1 e a matéria orgânica em suspensão de 2,30 mg L-1 a 4,78 mg L-1. Os maiores valores foram
a) b)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
193
encontrados na parte mais ao sul do Estreito, nas estações próximas à Ilha Decepcion. As concentrações de clorofila-a foram baixas, variando de 0,19 mg m-3 a 1,73 mg m-3.
Figura 2 - Esquema feito com base na salinidade, mostrando da influência dos Mares de Bellingshausen e Weddell no
Estreito de Bransfield (Março de 2011). Figure 2. Scheme made based on salinity, showing the influence of the Bellingshausen and Weddell Seas in the Bransfield
Strait (March 2011). Tendo em conta os dados das águas de superfície obtidos no fim do verão austral (março) em 2011 e, início do verão seguinte, em outubro de 2011 (Tab. 1), foi observado que a temperatura superficial foi em média 1°C mais baixa em outubro, onde predominaram temperaturas abaixo de zero. No fim do verão (março), as temperaturas foram mais altas, atingindo um máximo de 2,6°C. A salinidade foi um pouco maior em março, mostrando maior influência da circulação, enquanto que em outubro, a média foi de 33,57, mostrando maior participação da água de degelo. A concentração média de silicato observada em março foi de 41,11 µmol L-1, enquanto em outubro foi de 62,21 µmol L-1. O teor médio de fosfato foi levemente superior em outubro (1,67 µmol L-1) que em março (1,34 µmol L-1). Assim, as maiores concentrações de silicato e fosfato observadas em outubro, estiveram associadas ao início de verão austral à presença de menores temperaturas e menores salinidades, indicando assim, uma maior influência de materiais continentais carreados junto com a água de degelo, que contribuiu para o aporte de nutrientes para as águas marinhas. O valor médio de clorofila-a que em outubro foi de 1,06 mg m-3, com um valor máximo de 1,73 mg m-3, enquanto em março observou-se um valor médio de 5,18 mg m-3, com máximo de 13,31 mg m-3. Este aumento da clorofila-a, associado à diminuição das concentrações de silicato e fosfato, sugere que no final do verão, o valor de clorofila reflete maior produtividade primária no período, devido à continuidade na disponibilidade de nutrientes, à incidência de luz e à temperatura média mais elevada, favorecendo o processo de fotossíntese. CONCLUSÃO Foi observado neste trabalho que há distinções entre a composição química e a produção biológica, bem como, a influência dor Mares de Bellingshausen e Weddel nas águas superficiais do Estreito de Bransfield, presentes em diferentes estágios do período de verão. O início do verão Antártico é marcado pelo princípio do processo de degelo, e a presença do aporte de materiais continentais e de água de degelo, que marcam a porção central do Estreito. No período de final de verão, os dados de temperatura

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
194
e salinidade estão mais elevados, marcando o final do período onde os valores de clorofila-a estão mais acentuados. As águas do Estreito contam com grande disponibilidade de nutrientes, em taxas suficientes à manutenção da produção primária, porém os fatores que a limitam são principalmente: a incidência de luz na camada eufótica e a temperatura.
Março 2011 Outubro 2011 Média Máximo Mínimo Média Máximo Mínimo
Temperatura da água (°C) 1,16 2,60 -0,20 -1,06 0,60 -2,10
Salinidade 34,15 34,54 33,70 33,57 34,16 33,45
Silicato (µM) 41,11 60,86 15,49 62,21 86,95 31,52
Fosfato (µM) 1,34 1,98 0,02 1,67 2,08 0,76
Nitrato (µM) 21,17 30,85 11,45 18,99 29,80 8,80
MPS (mg/L) 17,34 32,00 13,40 17,65 23,11 12,40
MO (mg/L) 4,59 2,95 8,83 3,66 4,78 2,30 Clorofila-a (mg m-3) 5,18 13,31 0,27 1,06 1,73 0,19
Tabela 2 - Tabela de dados hidroquímicos e biológicos, com valores máximos, mínimos e médios obtidos em março e outubro de 2011, em águas superficiais do Estreito de Bransfield (Antártica).
Table 2. Scheme made based on salinity, showing the influence of the Bellingshausen and Weddell Seas in the Bransfield Strait (March 2011).
AGRADECIMENTOS Agradecemos a PROANTAR e ao CNPq Proc 55 557125/2009-0, à tripulação do NApOc Ary Rongel da Marinha do Brasil e toda equipe LABNUT do Instituto Oceanográfico da USP. REFERÊNCIAS Castro, C.G., Ríos, A.F., Doval, M.D. and Pérez, F.F. 2002. Nutrient utilization and chlorophyll distribution in the Atlantic sector of the Southern Ocean during austral summer 1995-96. Deep-Sea Research II. 49:623-641. García, M. A., Castro, C. G., Ríos, A. F., Doval, M. D., Rosón, G., Gomis, D. and López, O. 2002. Water masses and distribution of physico-chemical properties in the Western Bransfield Strait and Gerlache Strait during Austral summer 1995 96. Deep Sea Research II. 49: 585 – 602. Grasshoff, K., Ehrardt, M., Kremling, K. 1983. Methods of seawater analysis.2nd rev. and extended ed. Weinheim: VerlagChemie. 419p. Sangrà, P., Gordoc, C., Hernández-Arencibia, M., Marrero-Díaz, A., Rodriguez-Santana, A., Stegmner, A., Martínez-Marrero, A., Pelegrí, J. L. and Pichon, T. 2011. The Bransfield current system. Deep-Sea Research I. 58 (4):390-402. Strickland, J. D. H. and Parsons, T. R. 1968. A practical handbook of seawater analyses. Ottawa: Fisheries Research Board of Canada. 311 p. (Bulletin.Fisheries Research Board of Canada, n. 167).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
195
CARACTERIZACIÓN ESPACIAL DE LA COMPOSICIÓN TAXONÓMICA Y FUNCIONAL DE COMUNIDADES FITOPLANCTÓNICAS EN BAHÍA FILDES, ISLA REY JORGE,
ANTÁRTICA.
(Spatial characterization of taxonomic and functional composition of phytoplanktonic communities in Fildes Bay, King George Island, Antarctica.)
Moreno M.1, Rojas R.1, Sánchez C.1, De la Iglesia, R.2 y N. Trefault1
1Centro de Genómica y Bioinformática, Universidad Mayor. Camino La Pirámide #5750, Santiago, Chile. [email protected] 2Departamento de Genética Molecular y Microbiología, Pontificia Universidad Católica de Chile, Alameda #340, Santiago, Chile. El ecosistema marino antártico constituye uno de los ambientes más extremos del planeta. Las fuertes influencias de masas de aguas dominadas por la corriente Circumpolar, los altos niveles de salinidad, las bajas temperaturas y largos periodos de estacionalidad, hacen de este ecosistema un lugar único para el estudio de las distintas comunidades microbianas. Una de las comunidades más importantes en todos los ecosistemas marinos, son los microorganismos fitoplanctónicos, ya que son un componente fundamental en procesos de producción primaria y en los ciclos bioenergéticos del planeta. Debido a que menos del 1% de los microorganismos ambientales son cultivables en condiciones de laboratorio, se han desarrollado diferentes estrategias para superar esta limitación. De esta forma, el uso de herramientas moleculares y de secuenciación tanto convencional como masiva ha permitido describir de manera más precisa y detallada una gran cantidad de comunidades microbianas y microorganismos, sin la necesidad de cultivarlos. En este contexto, el estudio de las comunidades microbianas fotosintéticas, pueden dividirse en dos grandes enfoques: el análisis bajo una mirada taxonómica (mediante marcadores moleculares asociados a genes ribosomales (X.Shi, Lepére et al. 2011) y desde una perspectiva funcional (por ejemplo empleando marcadores como el gen psbA que codifica para la subunidad D1 del fotosistema II (Man-Aharonovich et al. 2010)). Si bien se han realizado análisis de composición comunitaria de microorganismos fitoplanctónicos en la Antártica, poco se sabe sobre la diversidad taxonómica y funcional de estas comunidades en Bahía Fildes, Isla Rey Jorge, Antártica. Por otra parte, debido a que los sistemas polares son especialmente susceptibles a los cambios climáticos que ocurren a nivel global, es de gran interés el estudio de las comunidades fitoplanctónicas y el efecto de las cambiantes condiciones ambientales actuales. El objetivo de este trabajo es realizar una caracterización a nivel espacial de la composición taxonómica y funcional de la comunidad de microorganismos fitoplanctónicos presentes en Bahía Fildes, Isla Rey Jorge, Antártica. Este trabajo combina herramientas moleculares clásicas de análisis de diversidad microbiana junto con herramientas de secuenciación masiva. El estudio se realizó en Bahía Fildes, Isla Rey Jorge, Antártica, durante febrero de 2012. Se analizaron 17 estaciones distribuidas dentro de la Bahía (Figura 1). Para realizar el análisis taxonómico, se empleó como marcador molecular, el gen 16S ARNr cloroplastidial. Los análisis de diversidad se realizaron mediante la técnica de T-RFLP (Terminal Restriction Fragment Length Polymorphism), complementada mediante pirosecuenciación, usando la plataforma GS Junior 454 Life Sciences, Roche. Para llevar a cabo los análisis funcionales, se seleccionaron 3 estaciones, cuyas muestras de DNA fueron empleadas para realizar librería genética de clones para el gen psbA (que codifica para la subunidad I del fotosistema II).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
196
Figura 1. Ubicación de los puntos de muestreos para el análisis espacial de las comunidades de microorganismos
fotosintéticos eucariontes en Bahía Fildes, Isla Rey Jorge, Antártica. Figure 1. Location of sampling points for the spatial analysis of photosynthetic eukaryotes communities, in Fildes Bay, King
George Island, Antarctica.
Los resultados del análisis de diversidad y composición taxonómico de las comunidades de microorganismos fotosintéticos analizadas mediante la técnica de T-RFLP, se muestran en la Figura 2. El análisis de agrupamiento jerárquico (Fig. 2a), demostró que las comunidades analizadas presentaron una alta homogeneidad a nivel espacial. Esto es corroborado con el análisis NMDS (Fig. 2b). Por otra parte, los análisis de composición comunitaria basadas en el gen taxonómico 16S ARNr (Figura 3a) mostraron que todas las estaciones analizadas están dominadas por los phyla Heterokontophyta y Haptophyta y en menor proporción por el Phylum Cryptophyta. En este caso no se observó una variación considerable en la composición comunitaria entre las distintas estaciones, lo que concuerda con los resultados obtenidos de T-RFLP.
Figura 2. Análisis de las comunidades eucariontes fotosintéticos de Bahía Fildes, Isla Rey Jorge, Antártica en base a perfiles
de T-RFLP obtenidos de la digestión de productos de PCR del gen 16S rARN cloroplastidial con la enzima HaeIII. (a) Análisis de agrupamiento jerárquico y (b) Análisis de escalamiento no métrico multidimensional (NMDS) de las 17
estaciones de muestreo, nivel de estrés 0,13. Figure 2. Analysis photosynthetic eukaryotes communities Fildes Bay, King George Island, Antarctica based on T-RFLP
profiles obtained from digestion of PCR products of the 16S rRNA with the enzyme HaeIII cloroplastidial. (a) Hierarchical cluster analysis and (b) Analysis of non-metric multidimensional scaling (NMDS) of the 17 sampling stations, stress level
0.13.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
197
100
0
50
75
25
Ab
un
dan
cia
rela
tiva
(% s
ecu
enci
as d
el g
ene
s 16
S cl
oro
pla
stid
ial)
Estaciones
Heterokontophyta
Haptophyta
Cryptophyta
Clorophyta
100
0
50
75
25
Ab
un
da
nci
a r
ela
tiva
(% s
ecu
en
cias
de
l ge
ne
s 1
6S
clo
rop
last
idia
l)
ES
T.3
ES
T.7
ES
T.8
Estaciones
16s rRNA Cloroplastidial psbA
Figura 3 Composición comunitaria de los principales Phyla de microorganismos fitoplanctónicos obtenidos de los análisis taxonómicos y funcionales. La imagen muestra el porcentaje de los Phyla de microorganismos eucariontes fotosintéticos dominante analizados mediante pirosecuenciación del gen 16S rARN cloroplastidial (a) y secuencias obtenidas desde la
librería genética de clones para el gen psbA (b). En ambos casos se observa una dominancia de los phyla Heterokontophyta y Haptophyta.
Figure 3. Community Composition of the major phyla of phytoplankton organisms obtained from taxonomic and functional analysis. The picture shows the percentage of microorganisms Phyla dominant photosynthetic eukaryotes analyzed using
pyrosequencing of plastid 16S rRNA gene (a) and sequences obtained from the genetic library of clones for the gene psbA (b). In both cases there is a dominance of the phyla heterokonts and Haptophyta.
El resultado de los análisis de la composición comunitaria de los microgramos fitoplanctónicos empleando el gen funcional psbA (Figura 3b) mostró que no existen variaciones importantes en la composición comunitaria entre las estaciones analizadas. También mostró una dominancia de los Phyla Heterokontophyta y Haptophyta, similar a lo observado empleando el gen 16S ARNr cloroplastidial. Sin embargo, a diferencia de lo que se observa en el análisis taxonómico, Clorophyta es el Phylum que se encontraría en menor proporción. Esto puede explicarse en el enriquecimiento que tienen las bases de datos en secuencias de los distintos grupos y en que, aun cuando el gen psbA se ha utilizado en otros estudios de composición de comunidades de microorganismos fotosintéticos, las bases de datos todavía no tienen muchos representantes de este gen o de la proteína que codifica. Este sesgo es aún mayor cuando se consideran sólo secuencias provenientes de sistemas polares. En resumen, los resultados obtenidos permiten plantear que en Bahía Fildes, la comunidad fitoplanctónica presentó una alta homogeneidad espacial durante la época de muestreo. Esto podría estar explicado por masas de aguas que mantengan un régimen de mezcla constante en la zona. Por otra parte la dominancia de las comunidades fitoplanctónicas pertenecientes a los Phyla Heterokontophyta y Haptophyta puede responder a florecimientos de estos microorganismos, lo cual es propio de épocas de mayor intensidad lumínica (verano-primavera) (Kudela, Pitcher et al. 2005). En conjunto, estos resultados pretenden contribuir al conocimiento de las comunidades fitoplanctónicas en ambientes marinos antárticos, además de constituir un aporte al estudio de microorganismos en ambientes extremos. AGRADECIMIENTOS Agradecimientos al proyecto INACH T16-10.
REFERENCIAS BIBLIOGRÁFICAS
(b) (a)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
198
Man-Aharonovich, D., Philosof, A., Kirkup, B., Le Gall, F., Yogev, T., Berman-Frank, I., Polz, M. Vaulot, D. and Oded, B. 2010. Diversity of active marine picoeukaryotes in the Eastern Mediterranean Sea unveiled using photosystem-II psbA transcripts. The ISME journal, 4(8): 1044–1052 Kudela, R., Pitcher, G., Probyn, T., Figueiras, F., Moita, T. and Trainer, V. 2005. Harmful algal blooms in coastal upwelling systems. Ocaenography 18(2): 184-197. Shi, X., Lepére, C., Scanlan, D. and Vaulot, D. 2011. Plastid 16S rRNA Gene Diversity among Eukaryotic Picophytoplankton Sorted by Flow Cytometry from the South Pacific Ocean. PlosOne, 6(4): 1-13.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
199
COMPOSICIÓN DE LA COMUNIDAD BACTERIANA MARINA EN EL OCÉANO ÁRTICO
(Marine bacterial community composition at the Arctic Ocean)
Sanhueza, C.1, 2, Díez, B.1
1 Departamento de Genética Molecular y Microbiología, Facultad de Ciencias Biológicas. Pontificia Universidad Católica de Chile, Alameda #340, C.P 651 3677, Santiago, Chile. 2 Facultad de Ciencias Químicas y Farmacéuticas, Universidad de Chile, Sergio Livingstone Pohlhammer#1007, Independencia, Santiago, Chile. E-mail: [email protected] INTRODUCCIÓN Las regiones polares se consideran ambientes extremos desafiantes para la vida. Estas regiones representan ecosistemas con características ambientales únicas, como son las bajas temperaturas, y variaciones en los niveles de luz a lo largo del año. El estudio de la composición de la comunidad bacteriana en estos ecosistemas, y en particular del ambiente marino ha sido limitado. En este estudio hemos caracterizado y comparado la composición de la comunidad bacteriana presente en diferentes hábitats marinos del Océano Ártico en muestras colectadas en dos transectos diferentes durante los años 2002, 2005 y 2012. La diversidad y patrones de distribución (espacial y por hábitats) de los genes 16S rRNA de la comunidad bacteriana fue analizada mediante secuenciación masiva. El análisis demostró la ubiquidad de algunos grupos bacterianos presentes en las diferentes estaciones muestreadas y hábitats en los años investigados, así como también una composición diferencial representada por determinados grupos bacterianos asociados a un hábitat determinado. MATERIALES Y MÉTODOS En este estudio, se han analizado muestras recolectadas en diferentes ambientes (Tabla 1) a lo largo de tres transectos en el Océano Ártico (Figura 1). Las muestras fueron recolectadas en expediciones en el buque oceanográfico sueco ODEN en los años 2002 (Mayo), 2005 (Agosto) y 2012 (Agosto - Septiembre). Las muestras fueron tratadas de diferente manera en las expediciones 2002, 2005 respecto a las del 2012. En los años 2002 y 2005 la biomasa microbiana de aproximadamente 1 litro de agua de mar, nieve o hielo se colectó en filtros GF/F de fibra de vidrio (tamaño de poro de ~ 0,7 mm) utilizando una bomba peristáltica con una tasa de filtración de 50-100 ml min-1. Los filtros se congelaron inmediatamente en nitrógeno líquido y se almacenaron hasta su análisis a -80 ° C. En el caso del año 2012, las muestras se filtraron por distintos tamaño de poro (20, 8 y 0.2um) almacenándose las distintas fracciones para los posteriores análisis. En cada una de las expediciones, se han analizado parámetros fisicoquímicos y nutrientes disponibles en el medio (Tabla 1). Los ácidos nucleicos se extrajeron según protocolo Bauer et al 2007 y analizado el gen 16S rRNA mediante secuenciación masiva por metodología Illumina utilizando partidores para bacterias generales (PE_16S_V4_F y PE_16S_V4_R).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
200
RESULTADOS Los valores de nutrientes obtenidos demuestran que la zona muestreada en el Ártico es una zona de bajas concentraciones de nutrientes, o al menos es evidente la falta de nutrientes nitrogenados (Tabla 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
201
La determinación de la diversidad de bacterias en los distintos hábitats (nieve, hielo y agua) del ecosistema marino a lo largo de escalas espaciales y temporales (interanuales) del Ártico es representado en la figura 2. Este es el primer reporte con un número de muestras amplio que además cubre aguas abiertas de un área importante del Ártico, e incluye además diferentes ambientes marinos. Los grupos de bacterias más representados fueron las Proteobacterias (alfa- y gamma- principalmente) y los Bacteroidetes. Las gamma-proteobacteria (Oceanospirales, Alteromonadales, Methylococcales) fueron las que presentaron mayor abundancia relativa (tanto en 2005 como 2012), seguidas de las alfa-proteobacteria (sobre todo Rhodospirillales, Rickettsiales, Rhizobiales y Sphingomonadales). Miembros de Rhodospirillales y Sphingomonadales fueron los más representados en la salmuera del hielo. Las beta-proteobacterias (Rhodocyclales, Burkolderiales, y otras) están mejor representadas en las muestras de salmuera y a fracciones de tamaño menores (8 y 0.2 um). Por otro lado, los Bacteroidetes del grupo Flavobacteriales (75-100%) y Sphingobacteria (10-25%) con géneros conocidos para estos ambientes como Polaribacter y Psycroflexus fueron bien representados. El grupo de las Actinobacterias fue representado solo por miembros del grupo de los Actinomycetales.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
202
CONCLUSIÓN Y DISCUSIÓN El análisis demostró la ubiquidad de algunos grupos bacterianos presentes en las diferentes estaciones muestreadas y hábitats en los años investigados, así como también una composición diferencial representada por determinados grupos bacterianos asociados a un hábitat determinado. AGRADECIMIENTOS A INACH (Proyectos MT_01.12 y Regular 15-10) y a la Pontificia Universidad Católica de Chile.
REFERENCIAS BIBLIOGRÁFICAS Bano, N. y Hollibaugh, J.T. 2002. Phylogenetic composition of bacterioplankton assemblages from the Arctic Ocean. Applied and Environmental Microbiology 68 (2): 505-518. Bowman, J., Rasmussen, S.. Blom, N., Deming, JW., Rysgaard, S., y Sicheritz-Ponten, T. 2012. Microbial community structure of Arctic multiyear sea ice and surface seawater by 454 sequencing of the 16S RNA gene. ISME Journal 6 (1): 11-20. Collins, R. E., Rocap, G. y Deming, J. W. 2010. Persistance of bacterial and archaeal communities in the sea ice through an Arctic winter. Environmental Microbiology 12 (7): 1828-41.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
203
DINÁMICA ESPACIAL DEL MESOZOOPLANCTON Y SU RELACIÓN CON EL DERRETIMIENTO GLACIARIO EN CALETA POTTER, ISLA 25 DE MAYO (SHETLANDS
DEL SUR, ANTÁRTIDA)
(Spatial dynamics of mesozooplankton in Potter Cove, King George Island (South Shetlands, Antarctica) and its relation to glacial melting)
Garcia, M.D.*1,2, Hoffmeyer, M.S.1,2,3 y I.R. Schloss2,4,5
1Instituto Argentino de Oceanografía, CONICET, Bahía Blanca, Argentina. [email protected]
2CONICET, Buenos Aires, Argentina. 3Facultad Regional de Bahía Blanca, Universidad Tecnológica Nacional, Bahía Blanca, Argentina. 4Instituto Antártico Argentino, Buenos Aires, Argentina. 5Institut des sciences de la mer de Rimouski, Université du Québec à Rimouski, Rimouski (Québec), Canada. RESUMEN La isla 25 de Mayo es la más grande de las Shetland del Sur, ubicadas al noroeste del extremo de la Península Antártica (PA), una de las regiones de más rápido calentamiento de la Tierra (Turner et al., 2005). Los sistemas glaciarios del oeste de la PA responden en forma directa al calentamiento climático de las últimas décadas por medio de un acelerado retroceso en su extensión e incremento en la producción de agua de derretimiento. Este proceso, además de contribuir al incremento del nivel del mar (Rignot et al., 2005; Cook et al., 2005), influye directamente en la dinámica de la columna de agua, modificando la composición y variedad regional del fitoplancton (Schloss et al. 1997, 2002 y 2012) y afectando, en consecuencia, al zooplancton costero (Loeb et al., 1997; Atkinson et al., 2004; Moline et al., 2004). Los factores que regulan la distribución y composición del plancton en aguas antárticas presentan una gran variabilidad espacial y temporal, pudiendo variar significativamente aún dentro de escalas geográficas reducidas (Quetin et al., 1996; Hernández León et al., 1999). La mayor parte de los estudios sobre el zooplancton en la región de la PA, se han concentrado en el krill, especialmente Euphausia superba (Brinton et al., 1987; Loeb y Shulenberger, 1987; Siegel y Loeb, 1995, entre otros), siendo menos los que incluyeron a los distintos grupos mesozooplanctónicos en su conjunto, tales como Menshenina y Rakusa‐Suszczewski (1992), Park (1993), Fuentes (2006) y Marrari et al. (2011 a y b). El principal objetivo de este trabajo fue estudiar el impacto directo e indirecto del aporte de aguas de baja salinidad, así como del incremento en el número de partículas de origen terrestre que ingresa a las aguas costeras, ambos producto del derretimiento glaciario, sobre la dinámica espacial (horizontal y vertical) del mesozooplancton en Caleta Potter. El presente estudio se realizó entre diciembre 2010 y diciembre de 2011 en la Base Argentina Científica Jubany (actualmente Carlini) localizada en la margen oriental de esta caleta. Los muestreos se llevaron a cabo con una frecuencia mensual en invierno y quincenal en verano en dos sitios y dos profundidades diferentes: Sitio 1 (S1): en la zona interna de la caleta (cercana al glaciar), a 5 y 20 metros de profundidad. Sitio 2 (S2): en la zona externa de la caleta, a 10 y 30 metros de profundidad. En cada uno de los sitios de muestreo se realizaron mediciones de temperatura, salinidad y conductividad utilizando un CTD (SBE 19 plus) desde un bote tipo Zodiac. Además, en cada punto y profundidad se recolectaron muestras de la columna de agua con botella Niskin de 5 L con el fin de analizar el material particulado en suspensión (MPS) (Ferreyra, 1987 modificado de Banse et al., 1963) y la concentración de pigmentos fotosintéticos (Strickland y Parsons, 1972; Holm-Hansen et al, 1965). Para el muestreo de mesozooplancton se realizaron arrastres con redes de 80 µm de diámetro de poro, en cada sitio y en ambas profundidades. Todas las mediciones y muestras de plancton se obtuvieron en el mismo momento y en aquellas ocasiones en que la caleta se encontró congelada (junio-

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
204
octubre), la toma de muestras se realizó a partir de orificios cavados en el hielo. Los datos meteorológicos provienen de la estación meteorológica de la Fuerza Aérea Argentina ubicada en la Base Carlini. Se aplicaron test estadísticos utilizando software apropiados (Statistica y CANOCO). Nuestros datos sobre efectos del derretimiento del glaciar Fourcade sobre el sistema de la caleta corroboran lo informado por Schloss et al (1997, 2002 y 2012): aporte de agua dulce de baja temperatura y dilución salina en el sector interno de la caleta, arrastre de materia particulada en suspensión, situación que ocasiona estratificación de la columna de agua y gran turbidez en las capas superficiales. En comparación con S2, en S1 se observó en promedio, una menor temperatura y salinidad del agua, una turbidez significativamente mayor (p<0,05) (Fig. 1) y una concentración de Cla inferior. En ambos sitios, las muestras de agua superficial presentaron mayores valores de temperatura y de turbidez, y menores valores de salinidad. La Cla mostró en S1 mayores valores en profundidad y en S2 en superficie, mientras que la Cla <20 fue en ambos sitios mayor en superficie. El mesozooplancton en términos globales, estuvo representado en un 92,3% por copépodos, 5% por organismos meroplanctónicos, 2,4% por eufáusidos y 0,3% por individuos de otros grupos zooplanctónicos (ostrácodos, anfípodos, gasterópodos, quetognatos y salpas). El copépodo ciclopoideo Oithona similis fue la especie más representativa del mesozooplancton con una abundancia porcentual del 54,5%; otros copépodos con elevadas abundancias fueron Ctenocalanus citer, Calanus propinquus, Stephos longipes, Calanoides acutus y Metridia gerlachei. En S1 se encontraron mayores valores de densidad total de mesozooplancton (p<0.01), en comparación con S2 (Fig. 2).
Figura 1: Datos de Salinidad, Temperatura y Turbidez del agua en S1 y S2, relacionados al grado de congelamiento de la caleta. PRE CONGELAMIENTO (diciembre-mayo), CONGELAMIENTO (junio-octubre) y POST CONGELAMIENTO
(noviembre-diciembre) Figure 1: Data Salinity, Temperature and Turbidity of water in S1 and S2, related to the degree of freezing of the cove. PRE
FREEZING (December-May), FREEZING (June-October) and POST FREEZING (November-December)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
205
Figura 2: Abundancia de Oithona similis, larvas furcilia y mesozooplancton total en S1 y S2, relacionadas al grado de
congelamiento de la caleta. Figure 2: Abundance of Oithona similis, furcilia larvae and mesozooplankton total in S1 and S2, related to the degree of
freezing of the cove. En S1 los mayores valores de abundancia de copépodos y eufáusidos se encontraron en las muestras superficiales, inversamente a lo observado en S2. Estas condiciones explican la elevada MPS y baja biomasa fitoplanctónica (Cla nunca mayor de 3 µg/l) encontradas en superficie en S1. Los datos de abundancia mesozooplanctónica con dominancia de pequeñas especies tales como O. similis adaptadas a ambientes con alta turbidez y baja productividad (a lo largo de todo el año, particularmente en S1), coinciden con lo registrado en estudios previos en Caleta Potter (Elwers y Dahms, 1998; Fuentes, 2006; Fuentes et al, 2008). Estos copépodos y las larvas de krill antártico se encuentran en asociación directa con la comunidad microbiana de la parte inferior del hielo marino (Quetin y Ross, 2001). Esto último podría explicar la mayor abundancia de eufáusidos en S1 producto de la alta densidad de larvas furcilia observada en los momentos de mayor congelamiento de la caleta. Nuestros resultados demuestran así el rol fundamental del proceso de congelamiento-descongelamiento marino en la estructuración del mesozooplancton de Caleta Potter. Asimismo, este estudio representa un antecedente para contrastar a escala regional las alteraciones en la trama trófica pelágica producidas por efecto de la dinámica del hielo marino y del derretimiento glaciario. AGRADECIMIENTOS A la dotación de la Campaña Antártica de Invierno 2011 de Base Jubany. REFERENCIAS BIBLIOGRÁFICAS Atkinson, A., Siegel, V., Pakhomov, E., Rothery, P. 2004. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature vol 432. Brinton, E., Loeb, V. J., Macaulay, M. C., Shulenberger, E. 1987. Variability of Euphausia superba population near Elephant Island and the South Shetlands: 1981 vs. 1984. Polar Biology, 7: 345‐362. Cook, A.J., Fox, A.J., Vaughan, D.G., Ferrigno, J.G. 2005. Retreating glacier fronts on the Antarctic Peninsula over the past half century. Science 308:541–544.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
206
Elwers, K., Dahms, H. U. 1998. Species composition and seasonal population structure of Oithona similis (Copepoda, Cyclopoida). En: The Potter Cove Ecosystem – Sinopsis. Wiencke, C., G. Ferreyra, W. Arntz y C. Rinaldi (Eds.). Berichte zur Polarforschung, 299: 150‐155. Ferreyra, G. A. 1987. Etude spatio‐temporelle des transports particulaires vers une communaute benthique itertidale de substrat meuble (Estuaire Maritime du Saint‐ Laurent). Tesis de Maestria. Université du Québec à Rimouski. 98 pp. Fuentes, V. 2006. Estudio de la Comunidad Zooplanctónica de Caleta Potter y Bahía Guardia Nacional (Isla 25 de Mayo): su Rol en la Dinámica del Carbono en Zonas Costeras Antárticas. Tesis Doctoral. Universidad de Buenos Aires. 249 pp. Fuentes, V. L., Schnack-Schiel, S. B., Schloss, I. R., Esnal, G.G. 2008. Mesozooplankton of Potter Cove: Community composition and seasonal distribution in 2002 and 2003. In: The Antarctic ecosystem of Potter Cove, King-George Island (Isla 25 de Mayo). Synopsis of research performed 1999-2006 at the Dallmann Laboratory and Jubany Station. Edited by Wiencke, C., Ferreyra, G. A., Abele, D., Marenssi, S., pp 75-84. Hernández–León, S., Torre, S., Gómez, M. M., Montero, I., Almeida, C. 1999. Biomass and metabolism of zooplankton in the Bransfield Strait during austral spring. Polar Biology, 21: 214‐219. Holm Hansen, O., Lorenzen, C. J., Holmes, R. W., Strickland, J. D. H. 1965. Fluorometric determination of chlorophyll. J. Cons. Cons. Int. Explor. Mcr 30: 3-l 5. Loeb, V. J., Shulenberger, E. 1987. Vertical distribution and relations of Euphausids populations off Elephant Islands, March 1984. Polar Biology 7, 363‐373. Loeb, V., Siegel, V., Holm-Hansen, O., Hewitt, R., Fraser, W., Trivelpiece, W., Trivelpiece, S. 1997. Effects of sea-ice extent and krill or salp dominance on the Antarctic food web. Nature 387 (6636): 897–900. Marrari, M., Daly, K.L., Timonin, A., Semenova, T. 2011a. The zooplankton of Marguerite Bay, western Antarctic Peninsula. Part I: abundance, distribution, and population response to variability in environmental conditions. Deep-Sea Research II. 58: 1599-1613. Marrari, M., Daly, K.L., Timonin, A., Semenova, T. 2011 b. The Zooplankton of Marguerite Bay, Western Antarctic Peninsula. Part II: vertical distributions and habitat partitioning. Deep-Sea Research, this issue. 13:1614-1629. Menshenina, L., Rakusa‐Suszczewski, S. 1992. Zooplankton changes during the year in Admiralty Bay (February 1990 – January 1991). Polskie Archiwum Hydrobiologii, 39: 49‐ 58. Moline, M.A., Claustre, H., Frazer, T.K., Schofields, O., Vernet, M. 2004. Alteration of the food web along the Antarctic Peninsula in response to a regional warming trend. Global Change Biology 10 (12): 1973–80. Park, C., Wormuth, J. H. 1993. Distribution of zooplankton around Elephant Island during the austral summer of 1988, 1989 and 1990. Polar Biology: 13, 215‐225. Quetin, L. B., Ross, R. M., Frazar, T. K., Haberman, K. L. 1996. Factors affecting distribution and abundance of zooplankton, with an emphasis on Antarctic krill, Euphasia superba. Foundations for Ecological Research West of the Antarctic Peninsula, Antarctic Research Series, (17): 357‐371. Quetin, L. B. y Ross, R. M. 2001. Environmental variability ans its impact on the reproductive cycle of Antarctic krill. American Zoologist, 41: 74‐89. Rignot, E., Casassa, G., Gogineni, S., Kanagaratnam, P., Krabill, W., Pritchard, H., Rivera, A., Thomas, R., Turner, J., Vaughan, D. 2005. Recent ice loss from the Fleming and other glaciers, Wordie Bay, West Antarctic Peninsula. Geophysical Research Letters, 32(7), L07502. Schloss, I.R., Ferreyra, G.A., Curtosi, A., Klöser, H., Mercuri, G., Pinola, E. 1997. Factors governing phytoplankton and particulate matter variation in Potter Cove, King George Island Antarctica. In: Battaglia, B., Valencia, J., Walton, D.W.H. (Eds.), Antarctic Communities. Cambridge University Press, Cambridge, pp. 135–141. Schloss, I. R., Ferreyra, G. A., Ruiz-Pino, D. 2002. Phytoplankton biomass in Antarctic shelf zones: a conceptual model based on Potter Cove, King George Island. Journal of Marine Systems 36: 129– 143. Schloss, I. R., Abele, D., Moreau, S., Demers, S., Bers, A. V., González. O., Ferreyra, G. A. 2012. Response of phytoplankton dynamics to 19-year (1991–2009) climate trends in Potter Cove (Antarctica). Journal of Marine Systems 92:53–66. Siegel, V. y Loeb, V. J. 1995. Recruitment of Antarctic krill (Euphausia superba) and possible causes for its variability. Marine Ecology Progress Series, 123: 45‐56. Strickland, J. D. H. y Parsons, D. R. 1972. A practical handbook of seawater analysis. J. Fish. Res. Bd. Can. Bull., 167:1‐310. Turner, J., Colwell, S.R., Marshall, G.J., Lachlan-Cope, T.A., Carleton, A.M., Jones, P.D., Lagun, V., Reide, P.A., Iagovkinaf , S. 2005. Antarctic climate change during the last 50 years. Internatinal Journal of Climatology, 25:279–294.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
207
SCALE-DEPENDENT SPATIAL VARIATION IN DIVERSITY AND STRUCTURE OF ROCKY ANTARCTIC COMMUNITIES
(Escalas de variación espacial en diversidad y estructura de comunidades antárticas rocosas)
Valdivia, N.1, Díaz M.1, Holtheuer J.1, Garrido I.1, Huovinen P.1 y I. Gómez1
1Instituto de Ciencias Marinas y Limnológicas, Facultad de Ciencias, Universidad Austral de Chile, Valdivia, [email protected]. Processes that affect species distribution operate and generate variability at different spatial scales (Wiens 1989). Thus, ecological research has shifted from considering spatial variation in species assemblages as "noise" to emphasize spatial scales of ecological variation asimportant to understanding how species assemblages are structured (Fraschetti et al. 2005). This can be particularly relevant for highly dynamic systems, such as the Antarctic shores, where broad seasonal variations in environmental conditions and disturbance frequency (e.g. ice scouring) significantly influence the structure of benthic assemblages (Gutt et al. 2013). Herein, we report the results of a study done using rocky intertidal and subtidal systems from the Antarctic shores of King George Island, South Shetland Islands. Separately for each system (intertidal and subtidal), we tested the hypothesis that the variation in species diversity and structure depend on spatial scale. To test this prediction, we used in both systems a nested sampling design to quantify species abundances across the desired horizontal spatial scales in terms of differences in sampling interval. To explain the observed community patterns, we also analysed the variability in the abundance of the species with the highest contribution to the spatial variation in community structure. The study was conducted in Fildes Peninsula, King George Island (Fig. 1) during the austral summer 2013. For the intertidal and subtidal habitats, we used nested sampling designs in which six and four shores, respectively, were randomly selected. Shores were of ca. 500 m in alongshore length and separated by a few kilometres. In each shore, three sites, separated by few 100s of metres, were randomly located. Within each site, two patches of substratum separated by 10s of metres were randomly located. Within each patch, three plots were randomly located at the low, mid-, and high intertidal, and at shallow (ca. 7 m depth) and deep (ca. 35 m depth) subtidal. On each plot, species percentage covers were estimated in situ and at 1 % resolution by means of 25-field 0.25 m2 quadrats.
-59.1 -58.9
-62.30
-62.25
-62.20
-62.15
-62.10
La
titud
e
Longitude
-59.0 -58.5 -58.0 -57.5
-62.2
-62.0
Figure 1: Study shores in Fildes Peninsula (left), King George Island (right). Filled and open symbol are intertidal and
subtidal study shores, respectively. Figura 1: Costas de estudio en Península Fildes (izquierda), Isla del Rey Jorge (derecha). Los símbolos negros y blancos son
costas intermareales y submareales, respectivamente.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
208
Asymptotic taxon richness was of 24 and 41 for the intertidal and subtidal habitats, respectively. Nested ANOVAs showed significant patterns of taxon richness and diversity (H’) across scales of observations. For the intertidal assemblages, the patterns of taxon richness and diversity along the vertical environmental stress gradient varied across scales of observations, evidenced by significant interactions between elevation (low, mid-, high intertidal) and the finer spatial scale of patch(site(shore)) (i.e. 10s of metres; F36, 216 = 2.49 and 3.1 for richness and diversity, respectively; P < 0.01). The subtidal habitat showed similar results, as vertical patterns of taxon richness and diversity also depended on the spatial scale of horizontal variability (interaction between depth [i.e. shallow vs. deep] and patch(site(shore)) scale; F12, 144 = 3.1 and 2.2 for richness and diversity, respectively; and P < 0.01 and P = 0.14, respectively). According to these results, the largest variance components of species richness and diversity were observed at the finer scale of horizontal variability (i.e. quadrat, decimetres to few metres) in both habitats. Moreover, variance components for the quadrat scale tended to increase toward the sea surface (i.e. from the high to low intertidal and from deep to shallow subtidal; Fig. 2). Canonical analyses of principal coordinates (CAP), conducted on Euclidean distances from standardised data showed that most of the spatial variability in community structure was explained by the filamentous green algae Urospora penicilliformis and Ulothrix sp., the red algae Porphyra endiviifolium, Iridaea cordata, Palmaria decipiens, the brown alga Adenocystis utricularis and the limpet Nacella concinna for the intertidal habitat. In the subtidal, the species better represented in the CAP ordinations were the brown alga Himantothallus grandifolius, a complex of unidentified crustose algae, and the red algae Plocamium cartilagineum and Picconiella plumosa. Excepting for U. penicilliformis, the species with highest representations showed larger variance components at the quadrat scale. The grazer limpet N. concinna showed highest variance components in the mid- and low intertidal heights, while H. grandifolius and the crustose morphs showed a stronger variability at deeper waters (Fig. 3). In addition, the abundance of H. grandifolius was also highly variable in shallow waters (Fig. 3).
Figure 2.Variance components of taxon richness and diversity (Shannon H’) across scales of spatial variability and habitat
(intertidal and subtidal). Figura 2. Componentes de varianza de riqueza y diversidad (H’) de taxa para cada escala especial y hábitat (intermareal y
submareal). These results agree with previous studies pointing to the pervasive nature of fine-scale variability in spatial patterns of species abundances and composition in temperate shores (Terlizzi et al. 2007, Valdivia et al. 2011). At fine resolutions, causes of horizontal variability in community structure include

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
209
physical disturbance (Sousa 1979), frond sweeping by canopy-forming seaweeds (Kiirikki 1996), competition (Benedetti-Cecchi et al. 1999), availability of refugia (Underwood & Chapman 1996), and variations in grazing activity (Díaz & McQuaid 2011). In Antarctic shores, increased fine-scale variability toward the shallow subtidal and low intertidal zone of some algal assemblages could be related to particular photobiological characteristics and environmental stress tolerance (Huovinen and Gómez 2013), and to the occurrence of mechanical disturbances by iceberg scouring (Gutt et al. 2013) and the concomitant re-colonisation process.
Figure 3. Variance components of dominant taxa across scales of spatial variability and habitats (intertidal and subtidal).
Figura 3. Componentes de varianza de taxa dominantes para cada escala especial y hábitat (intermareal y submareal) The larger fine-scale variability observed at the sea surface could also reflect the outcome of biotic interactions. While the spatial variability of opportunistic green alga U. penicilliformis was larger at the high and mid-intertidal height, the variability of N. concinna peaked at the low intertidal zone. Grazing activity of N. concinna has been shown to have significant effects on intertidal re-colonisation by periphyton (Zacher et al. 2007), and could well influence the lower distribution limit of intertidal species. This agrees with the recently described biotic zonation of Antarctic intertidal shores (Waller 2013) and the pervasive vertical zonation described for temperate shores (e.g. Scrosati et al. 2011). In addition, foundation species, such as H. grandifolius, are known to have strong effects on the assembly and maintenance ofcommunity structure (e.g. Bertocci et al. 2010). Our results showed a relatively high variability at the quadrat scale of H. grandifolius, which may have contributed to making a large fine-scale horizontal variation in community structure at the sea-surface level. In summary, these preliminary analyses suggest that fine-scale structuring processes along the coastline may be particularly relevant for coastal community organization in Antarctica. In addition, fine-scale processes arise as being as important in determining community structure as the prevailing physical stresses acting on marine rocky shores across the vertical elevation gradient.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
210
ACKNOWLEDGEMENTS We thank financial support by the Grant CONICYT-AnilloART1101 and MECESUP AUS0805. REFERENCES Benedetti-Cecchi, L., Menconi, M., and Cinelli, F. 1999. Pre-emption of the substratum and the maintenance of spatial pattern on a rocky shore in the northwest Mediterranean. Marine Ecology-Progress Series, 181:13-23. Bertocci, I., Arenas, F., Matias, M.G., Vaselli, S. and others. 2010. Canopy-forming species mediate the effects of disturbance on macroalgal assemblages on Portuguese rocky shores. Marine Ecology-Progress Series, 414:107-116. Díaz, E.R., and McQuaid, C.D. 2011. A spatially explicit approach to trophic interactions and landscape formation: patchiness in small-scale variability of grazing effects along an intertidal stress gradient. Journal of Ecology, 99:416-430. Fraschetti, S., Terlizzi, A., and Benedetti-Cecchi, L. 2005. Patterns of distribution of marine assemblages from rocky shores: evidence of relevant scales of variation. Marine Ecology-Progress Series, 296:13-29. Gutt, J., Cape, M., Dimmler, W., Fillinger, L. and others. 2013. Shifts in Antarctic megabenthic structure after ice-shelf disintegration in the Larsen area east of the Antarctic Peninsula. Polar Biology, 36:895-906. Kiirikki, M. 1996. Experimental evidence that Fucus vesiculosus (Phaeophyta) controls filamentous algae by means of the whiplash effect. European Journal of Phycology, 31:61-66. Scrosati, R.A., van Genne, B., Heaven, C.S., and Watt, C.A. 2011. Species richness and diversity in different functional groups across environmental stress gradients: a model for marine rocky shores. Ecography, 34:151-161. Sousa, W.P. 1979. Disturbance in marine intertidal boulder fields: the nonequilibrium maintenance of species diversity. Ecology, 60:1225-1239. Terlizzi, A., Anderson, M.J., Fraschetti, S., and Benedetti-Cecchi, L. 2007. Scales of spatial variation in Mediterranean subtidal sessile assemblages at different depths. Marine Ecology-Progress Series, 332:25-39. Underwood, A.J., and Chapman, M.G. 1996. Scales of spatial patterns of distribution of intertidal invertebrates. Oecologia, 107:212-224. Valdivia, N., Scrosati, R.A., Molis, M., and Knox, A.S. 2011. Variation in community structure across vertical intertidal stress gradients: how does it compare with horizontal variation at different scales? PLoS ONE, 6:e24062. Waller, C.L. 2013. Zonation in a cryptic Antarctic intertidal macrofaunal community. Antarctic Science, 25:62-68. Wiens, J.A. 1989. Spatial scaling in ecology. Functional Ecology, 3:385-397. Zacher, K., Hanelt, D., Wiencke, C., and Wulff, A. 2007. Grazing and UV radiation effects on an Antarctic intertidal microalgal assemblage: a long-term field study. Polar Biology, 30:1203-1212.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
211
LOS ENSAMBLES MEIOBENTONICOS INTERMAREALES DE LAS ISLAS SHETLAND DEL SUR, COSTA DANCO Y BAHÍA MARGARITA
(The intertidal meiofaunal assemblages of the South Shetland Islands, Danco Coast and Margarita Bay)
Lee, M.R.1, Brante, A.2, Hernandez, C.E.3 and Olivares, B.1
1Centro i~mar, Universidad de Los Lagos, Camino a Chinquihue km.6, Puerto Montt, Chile. E-mail: [email protected]; 2Departamento de Ecología, Facultad de Ciencias, Universidad Católica de la Santísima Concepción, Concepción, Chile; 3Laboratorio de Ecología Molecular y Filoinformática, Departamento de Zoología, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile. INTRODUCCIÓN La meiofauna son invertebrados pequeños que viven en el ambiente bentónico, principalmente asociados con los fondos blandos, pero también en asociación con los sustratos secundarios como algas e invertebrados sésiles (Giere 2009). El estudio de la meiofauna en la Antártica es muy limitado y restringido al mar profundo y los fondos de la plataforma continental. Los estudios de la meiofauna del intermareal son escasos, por lo que se desconoce su diversidad, funcionamiento y estructura comunitaria. Esta carencia de conocimiento se hace más crítica pensando en un contexto de cambio climático global, en donde el periodo en que la zona intermareal permanece libre de hielo va a aumentar (Turner el al. 2009). El objetivo de este trabajo es obtener un primer acercamiento al estudio de los ensambles meiobentónicos, su diversidad, abundancia y distribución, en la zona intermareal de algunas islas de la Antártica, con un foco en los nemátodos, usando técnicas morfológicas y genéticas. Adicionalmente, se comparan los ensambles de meiofauna de la Antártica con los de la región de Magallanes. MATERIALES Y MÉTODOS Muestras cuantitativas (9 sitios) y cualitativas (14 sitios) fueron recolectados de la Antártica, en los siguientes sectores: Islas Shetland del Sur – Isla Rey Jorge (playa ubicada frente a la base Escudero (ESC), Playa Funschloger (FUN), Playa Elefante (ELE)), Isla Decepción (playa ubicada frente a la base Gabriel Castillo (CAS), Bahía Fumerol (FUM), Caleta Péndulo (PEN), Bahía Whaler (WHA)), Isla Livingston – Punta Hannah (HAN); la Costa Danco – Base Gabriel González Videla (GGV) y Port Lockroy (LOC); y Bahía Margarita – base Rothera (ROT), Punta Alexandar (ALX), Isla Jenny (JEN), Isla Herradura (SHO). En cada sitio se tomaron muestras cuantitativas en la zona de retención (o la parte mediana de la playa) donde se observan típicamente las mayores abundancias y diversidad de meiofauna. Las muestras cualitativas fueron tomados en las siguientes zonas: seca (la parte alta de la playa), retención (la parte media de la playa), resurgencia (la parte baja de la playa) y en el nivel del espejo de agua en la zona de retención (más profundo en el sedimento). En algunos sitios se tomaron muestras adicionales: en FUN (Isla Rey Jorge) de algas intermareales (Porphyra sp.), en ELE (Isla Rey Jorge) de algas intermareales (Adenocytis utricularis), en FUM (Isla Decepción) adyacente a la fumarola, en PEN (Isla Decepción) en aguas someras frente a la playa; y en Isla Decepción, Isla Greenwich (Bahía Chile), Port Lockroy, e Isla Herradura, muestras de sedimentos submareales. Las muestras cuantitativas fueron fijadas en terreno con formalina al 5%. Para las muestras cualitativas la meiofauna fue extraída del sedimento por la metodología de decantación, después de 15 minutos de anestesiar los individuos presentes con cloruro de magnesio isotónico. La fauna fue capturada con un tamiz de 45 µm. Las muestras fueron fijadas con DESS o en algunos casos con formalina al 5%. DESS es una solución de EDTA disodio, dimetilsulfoxido (DMSO) saturado en sodio cloruro. Este solución

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
212
tiene la ventaja que fija a los organismos en un estado apto para identificación morfológica pero también no daña el ADN, lo cual permite análisis genéticos posteriores (Yoder 2006). En el laboratorio las muestras cuantitativas de la meiofauna fueron extraídas de los sedimentos por decantación seguido por flotación en LudoxTM. La fauna capturada en un tamiz de 45 µm fue evaporada a glycerol en una desecador y montado dentro de un anillo de cera en portaobjetos grandes. Las muestras fueron examinadas y analizadas bajo microscopio (Olympus BX51), con iluminación DIC conectado a una cámara digital, y un tubo de dibujo. Cada especie fue identificada al nivel taxonómico más bajo posible y se registró su abundancia. En el laboratorio los nemátodos encontrados en las muestras cualitativas fueron montados temporalmente y examinados e identificados bajo el microscopio. En cada caso, los especímenes fueron identificados hasta el nivel taxonómico más bajo posible, dibujado en detalle, fotografiado, registrándose las dimensiones corporales para análisis morfométricos. Después de analizarlos, cada espécimen fue recuperado del portaobjeto y colocado en un micro-tubo de 0.2ml con DESS y enviado para análisis genético. Datos de muestras cuantitativas recolectados durante un proyecto previo (FONDECYT 1080033) en seis sitos del Estrecho Magallanes fueron usados para comparar la meiofauna de la Antártica con la meiofauna del continente. RESULTADOS Las siguientes grupos meiobentónicos fueron identificados en las muestras de sedimento de la Antártica (Tabla 1): Foraminiferos (FOR), Turbelarios (TUR), Nemátodos (NEM), Copépodos harpacticoideos (HAR), Tardigrados (TAR), Ciliados (CIL), Rotíferos (ROT), Ácaros (ACA), Halacaroidea (HAL), Colembolos (COL), Oligoquetos (OLI), Poliquetos (POL), Bivalvos (BIV) y Ostracodos (OST), Amfipoda (AMP), Isopodos, (ISO), Tanaidaceos (TAN), Cumaceos (CUM), Kinorhynchos (KIN), Gastrotrichos (GAS), Priapulidos (PRI), Gastropodos (GAT), y Briozoos (BRI). Una total de 23 grupos fueron encontrados en Antártica, con la riqueza de grupos (Sgrupo) variado entre 17 (LOC) y 0 (SHO). Una menor cantidad de grupos fueron encontrados en las muestras cuantitativas, dado que se representan muestras de un solo sub-hábitat (la parte media de la playa). Las abundancias para todos los grupos fue mayor en la región de Magallanes comparado con la Antártica. Las abundancias en el sector de Bahía Margarita fue la más baja en la Antártica, con una ausencia de nemátodos. Los valores de Sgroup fueron más altos en Isla Rey Jorge comparado con Isla Decepción, aunque los sitios con el Sgrupo más altos fueron del sector Costa Danco; en ambos casos el sedimento fue mal ordenado y enriquecido con materia orgánica derivada de las colonias de pingüinos presentes en sector. Nueve especies de nemátodos fueron encontrados en las muestras cuantitativas de la Antártica (muestras de 2012, las muestras de 2013 están en análisis); siete especies adicionales fueron encontrados en las muestras cualitativas. De esta forma el Snem total para las Islas Decepción y Rey Jorge fue de 16 especies. La Snem para Isla Rey Jorge (5.67) fue más alto que la Snem para Isla Decepción (2.67). La abundancia promedio de nemátodos en la Antártica fue de 13.84 ind/50ml. Las abundancias promedio de Isla Rey Jorge (25.65 ind/50ml) fueron más altas que las abundancias para Isla Decepción (8.25 ind/50ml) y Isla Livingston (2.80 ind/50ml). Las tres órdenes de nemátodos de vida libre fueron encontrados (Enoplida, Chromadorida y Monhysterida). La especies más abundante fue Halomonhystera sp. (Monhysterida: Monhysteridae), seguido por Chromadorita sp. (Chromadorida: Chromadoridae). La relación entre la diversidad de nemátodos y latitud publicada por Lee y Riveros (2012) predice una diversidad de nemátodos en la Antártica (Islas Decepción y Rey Jorge, latitud 62 °S) de entre 0 y 4 especies, siendo el promedio de la Snem observado para las muestras cuantitativas de Antártica de 4.17. La abundancia promedio de nemátodos en el estrecho Magallanes fue 275.93 ind/50ml con una diversidad (Snem) de 13.33 especies. Análisis MDS de los grupos de meiofauna indica que no hay distinción clara en las ensambles de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
213
meiofauna de las islas Antárticas (estrés 0.13). Un análisis que incluye los datos de todos los sitios del Estrecho Magallanes indica que los ensambles de Magallanes y Antártica son distintos (estrés 0.09). El análisis de ANOSIM indica que hay diferencias significativas en los niveles de sitios (p<0.001), islas (p<0.001) (Continente, Tierra del Fuego, Rey Jorge y Decepción) y regiones (p<0.001) (Magallanes y Antártica).
Taxa
Isla Rey Jorge Isla Decepción Isla
Livingston
Costa Danco Bahía Margarita
ESC ELE FUN CAS FUM PEN WHA HAN GGV LOC ROT ALX JEN SHO
NEM X X X X X X X X X X
TAR X
HAR X X X X X X X X X
TUR X X X X X X X X X
OLI X X X X X X X X
HAL X X X
ACA X X X X X
COL X X X X X X
ROT X X X X
OST X X X X
AMP X X
GAT X X
POL X
CUM X
KIN X
ISO X
TAN X
CIL X X
BRY X
BIV X
PRI X
GAS X X
FOR X X
Sgrupo 6 11 6 4 5 1 5 9 8 17 1 2 2 0 Para una clave de los códigos usados para los sitios ver Metodología. Clave a las códigos de taxa usados: NEM = Nematoda, TAR = Tardigrada, HAR = Harpacticoidea, TUR = Turbellaria, OLI = Oligochaeta, HAL = Halacarida, ACA = Otra Acari, COL = Collembola, ROT = Rotifera, OST = Ostracoda, AMP = Amphipoda, GAT = Gastrapoda, POL = Polychaeta, CUM = Cumacea, KIN = Kinorhyncha, ISO = Isopoda, TAN = Tanaidacea, CIL = Cilophora, BRY = Bryozoa, BIV = Bivalva, PRI = Priapulida, GAS = Gastrotricha, FOR = Foraminifera.
Tabla 1. Grupos de meiofauna presentes en la intermareal basados en muestras cuantitativas y cualitativas 2012/13. Table 1. Meiofauna groups present in the intertidal based on both quantitative and qualititative samples (2012/13).
DISCUSIÓN Los resultados de este primer estudio de la meiofauna intermareal de las islas Antárticas indican que los ensambles son distintos en comparación a los encontrados más al norte en Chile continental. Los ensambles intermareales de meiofauna en la Antártica presentan diversidades y densidades bajas. Existe

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
214
evidencia que el enriquecimiento de los sedimentos con fecas de pingüinos puede tener un efecto positivo sobre la diversidad de meiofauna presente, y es necesario poner a prueba este patrón en la siguiente campaña de terreno con muestras cuantitativas. En 2012 existía evidencia observacional que la distribución de la meiofauna es diferencial a lo largo de la playa. Los ensambles de meiofauna en las playas Antárticas fueron dominados por nemátodos con escaso harpacticoideos. Los turbellarios fueron más abundantes en Isla Decepción caracterizada por la presencia de actividad volcánica, y en donde hay observaciones previas de turbellarios con tolerancias a temperaturas altas (Bright et al. 2003). En Isla Rey Jorge los oligoquetos fueron comunes y los rotíferos estuvieron presentes en ambas islas, siendo más frecuentes en Isla Decepción. En Isla Decepción las abundancias fueron bajas, no así la diversidad. La posible causa de este patrón es la presencia de aguas más cálidas encontradas en Isla Decepción. Para una fauna Antártica adaptada a condiciones frías, las temperaturas más altas pueden ser estresantes reduciendo la colonización de estas zonas de algunas especies. Durante la campaña de 2013 un cuarto sitio (WHA) fue maestreado en Isla Decepción, donde las temperaturas son más bajas que en otras partes de la isla, siendo las abundancias de la meiofauna más altas, proporcionando apoyo a la hipótesis previa. Otra hipótesis que se puede sugerir es que la dispersión de la meiofauna hacia las playas del interior de Isla Decepción puede ser menos frecuente debido a que la entrada a la bahía es muy estrecha. La abundancia y diversidad de la meiofauna en las playas del sector de Bahía Margarita fueron muy bajas, con la presencia sólo de oligoquetos (ROT y ALX), Turbelarios (ALX y JEN) y Ciliados (JEN), estando ausente completamente el grupo de los nemátodos. Esto podría ser explicado por la falta de hábitat apropiado: las playas de arena son muy escasas en este sector, siendo las playas de adoquines las más frecuentes. AGRADECIMIENTOS Financiamiento Proyecto INACH T_13-10. REFERENCIAS BIBLIOGRÁFICAS Bright, M. Arndt, C. Keckeis, H. y Felbeck, H. 2003. A temperature-tolerant interstitial worm with associated epibiotic bacteria from the shallow water fumaroles of Deception Island, Antarctica. Deep-Sea Research II, 50:1859- 1871. Giere, O. 2009. Meiobenthology. The Microscopic Motile Fauna of Aquatic Sediments Springer-Verlag, 527pp. Lee, M. y Riveros, M. 2012 Latitudinal trends in the species richness of free-living marine nematode assemblages from exposed sandy beaches along the coast of Chile (18–42 °S). Marine Ecology, 33:317-325. Turner J, et al., eds 2009. Antarctic Climate Change and the Environment (Scientific Committee on Antarctic Research, Cambridge, UK). Yoder, M., Tandingan De Ley, I., King, I., Mundo-Ocampo, M., Mann, J., Blaxter, M., Poiras, L. y De Ley, P. 2006. DESS: a versatile solution for preserving morphology and extractable DNA of nematodes. Nematology, 8:367-376.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
215
DIETA DEL PEZ HARPAGIFER ANTARCTICUS NIBELYN, 1947 (NOTOTHENIOIDEI: HARPAGIFERIDAE), DE POZAS INTERMAREALES DE ISLA REY JORGE: POSIBLE
PERSISTENCIA TEMPORAL EN UN OCÉANO AUSTRAL CALENTÁNDOSE
(Diet of the fish Harpagifer antarcticus Nibelyn, 1947 (Notothenioidei: Harpagiferidae), from interdidal pools of King George Island: possible temporal persistence in a warming Southern Ocean)
Gutiérrez, D.N.1
1Facultad de Ciencias del Mar y Recursos Naturales, Universidad de Valparaíso, Avda. Borgoño 16344, Valparaíso, Chile. [email protected]
INTRODUCCIÓN La actual aceleración del cambio climático a nivel global, en especial en las zonas polares, tiene a la Península Antártica como una de las tres áreas con mayor aumento de temperatura (Vaughan et al. 2003). El océano también ha sido alterado en esta zona, aumentando en 1°C la temperatura del mar en el Mar de Bellingshausen en sólo 50 años (Meredith & King 2005). Esto último ya ha comenzado a tener consecuencias en las redes tróficas, alterando su estructura y dinámica, a través de la abundancia y distribución de poblaciones de organismos (Ducklow et al. 2007). La tolerancia térmica en los organismos que habitan el Océano Austral, se ha generalizado como estenotermal (Clark et al. 2008), con pequeños rangos térmicos, menores a los de animales de latitudes menores, relacionándolos con su historia evolutiva (Bilyk & DeVries 2011). Sin embargo, se ha demostrado una mayor plasticidad térmica fisiológica generalizada entre la ictiofauna antártica (Bilyk & DeVries 2011). Los miembros de la familia Harpagiferidae (Notothenioidei) están representados por un único género, siendo Harpagifer antarcticus Nybelin, 1947 la única especie presente en la Península Antártica y las islas del Arco de Escocia (Hureau 1990). Habita aguas costeras poco profundas sobre fondos rocosos, bajo piedras y grietas (Hureau 1990), alimentándose principalmente de anfípodos gamáridos (Gammaridea: Gammarida) (Duarte & Moreno 1981, Casaux 1998). En base a los antecedentes entregados, el presente estudio tiene como objetivo analizar el contenido estomacal de H. antarcticus, considerando que las descripciones de la dieta en peces permiten inferir sobre los cambios ambientales que influyen en la abundancia y distribución de organismos, tales como alteraciones del hábitat de origen natural o antropogénico (Chipps & Garvey 2007). De esta forma, se evaluará la persistencia o cambio que pudiera existir en la dieta de H. antarcticus, con comentarios sobre los posibles efectos inducidos por el cambio climático que afecta fuertemente el Océano Austral. La obtención de muestras se realizó en Bahía Fildes en Isla Rey Jorge, Shetlands del Sur, durante enero de 2012, durante la Expedición Científica Antártica N° 48, realizada por el Instituto Antártico Chileno. Fueron capturados a mano 45 ejemplares de Harpagifer antarcticus, durante la marea baja, del intermareal rocoso de una playa ubicada en el sector oeste de Bahía Fildes (62º12'S, 58º57'O). En la Facultad de Ciencias del Mar y Recursos Naturales, de la Universidad de Valparaíso, se extrajo el contenido estomacal, se separó y se identificó cada presa hasta el nivel taxonómico más bajo posible, con ayuda de claves especializadas y el empleo de una lupa estereoscópica. Se realizó una curva de diversidad trófica para evaluar el número de estómagos necesarios que representen la dieta de manera adecuada, siguiendo a Ferry & Calliet (1996) y Cortés (1997). La cuantificación dietaria se realizó a través del método frecuencia de ocurrencia, numérico y gravimétrico (Hyslop 1980). Para identificar las principales presas, se empleó el Índice de Importancia Relativa (Pinkas et al. 1971, Cortés 1997). Se evaluó la amplitud de nicho a través del Índice

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
216
Estandarizado de Levin (BA), en donde valores menores a 0,6 indican un estrecho nicho con hábitos especialistas (Krebs 1998). Complementariamente se empleó la medida de amplitud de nicho de Shannon-Wiener (H`) y el de equitatividad (J`) (Krebs 1998), tanto para el presente estudio, como para años anteriores (Duarte & Moreno 1981, Casaux 1998). Para determinar la similaridad de las dietas, en los distintos años, se emplearon el Índice de Similaridad Porcentual (ISP) (Whittaker 1952) y el Índice de Morisita-Horn (CH) (Horn 1966), empleando el Coeficiente Alimentario Gravimétrico (Qp) y Numérico (Qn) (Hureau 1970) para items mayores y anfípodos gamáridos respectivamente. La curva de diversidad trófica alcanzó la asíntota en 35 estómagos, por lo que el tamaño muestral (45 estómagos) fue el adecuado para representar la dieta de Harpagifer antarcticus del actual estudio (2012). De los 5 grupos de presas identificados, siendo los anfípodos gamáridos el ítem más importante (IIR = %88.95), quedando el resto de categorías con porcentajes marginales (Tabla 1). Dentro de los gamáridos en los estómagos de H. antarcticus, Gondogeneia antárctica fue la presa más importante (IIR = %76.71), mientras que Orchomenella sp. quedó en segundo lugar de importancia (IIR = %19.39), presente en casi el %60 de los estómagos, cubriendo entre ambas especies más del %96 del IIR (Tabla 2).
Tabla 1: Izquierda: Porcentaje en frecuencia de ocurrencia (%FO), número (%N), peso (%P) e Índice de Importancia Relativa
(%IIR) de los items presa en los estómagos de Harpagifer antarcticus en Bahía Fildes. (n=44 estómagos con contenido). Table 1: Left: Percentage by frequency of occurrence (%F), number (%N), weight (%W) and index of relative importance
(%IRI) of prey items in the stomachs of Harpagifer antarcticus in Fildes Bay (n=44 stomachs with content)
Tabla 2: Derecha: Anfípodos gamáridos representados en la dieta de Harpagifer antarcticus, Bahía Fildes, como porcentaje en frecuencia de ocurrencia (%FO), número (%N), peso (%P) e Índice de Importancia Relativa (%IIR) (n=44 estómagos con
contenido). Table 2: Right: Gammarid amphipods represented in the diet of Harpagifer antarcticus, Fildes Bay, as percentage by
frequency of occurrence (%F), number (%N), weight (%W) and index of relative importance (%IRI) (n=44 stomachs with content)
Se encontraron hábitos alimentarios especialistas, con un estrecho nicho, tanto para grupos mayores como para anfípodos gamáridos (BA=0.005, H`=0.075, J`=0.042 y BA=0.170, H`=0.689, J`=0.428 respectivamente), en H. antarcticus. Los resultados obtenidos en este estudio (2012), son ampliamente respaldados con lo reportado en otras investigaciones (Moreno 1971, Target 1981, Duarte & Moreno 1981, Daniels 1982, Casaux 1998),
ITEM %FO %N %P %IIR
Crustacea
Gammarida 100,00 70,06 92,82 88,95
Restos de Crustacea
63,64 14,50 4,16 6,49
Mollusca Bivalvia 38,64 7,02 0,87 1,66
Gastropoda 56,82 7,13 1,94 2,82
Polychaeta 6,82 0,35 0,19 0,02
Algas 11,36 0,94 0,01 0,06
Piedra 4,55 - - -
TOTAL 100 100 100
ITEM %FO %N %P %IIR
Gondogeneia antarctica
90,91 60,55 59,98 76,71
Eurymera monticulosa
13,64 3,90 21,62 2,44
Paramoera sp. 31,82 5,28 1,26 1,46
Orchomenella sp.
59,09 30,05 16,84 19,39
Austropleustes cuspidatus?
2,27 0,23 0,29 0,01
TOTAL 100 100 100

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
217
encontrando como grupo principal en aguas someras, a los gamáridos. Sin embargo, esta especialización dietaria, ha sido señalada como una consecuencia de la estrategia alimentaria (acecho, mirar Daniels (1982)) y la disponibilidad de la presa (Arnaud 1977 fide Duarte & Moreno 1981), siendo apoyada esta hipótesis por Casaux (1998), quien encontró krill como ítem principal, en especímenes colectados entre probablemente 46-110 m, probablemente cuando el krill abunda cerca del fondo.
Figura 1. Anfípodos gamáridos presentes en la dieta de Harpagifer antarcticus, como Coeficiente Alimentario Numérico
(%Qn), para el estudio actual, Duarte & Moreno (1981) y Casaux (1998). Figure 1. Gammarid amphipods present in the diet of Harpagifer antarcticus, as Alimentary numerical Coefficient (%Qn),
for the current study, Duarte & Moreno (1981) and Casaux (1998).
Estos patrones de similaridad se observan claramente al comparar los coeficientes alimentarios de los estudios en Isla rey Jorge (Fig.1). Para items mayores, los gamáridos tuvieron la mayor importancia, con %Qp de 98.78, 99.85 y 99.73 para el actual estudio, 1998 (Casaux 1998) y 1981 (Duarte & Moreno 1981) respectivamente. Además, todos tuvieron como principal anfípodo gamárido a Gondogeneia antárctica (Fig. 1). Los índices de similaridad también fueron muy altos, con valores de ISP y CH de %98.8 y 1 respectivamente, entre 2012 y 1998 para items mayores. Así mismo, para los items mayores de años 2012 y 1981, hubo valores de ISP y CH de %99 y 1 respectivamente. Por otra parte, para los anfípodos gamáridos, también hubo altos valores entre los años. Entre 2012 y 1981, hubo valores de ISP y CH de %76.2 y 0.96 respectivamente, mientras que entre 2012 y 1998, hubo valores de ISP y CH de %78.9 y 0.94 respectivamente. Producto del fuerte efecto del calentamiento global en el Océano Austral, se han registrado alteraciones en la abundancia y disponibilidad de presas de diversos depredadores (Ducklow et al. 2007). Esto resulta importante, considerando la baja capacidad de aclimatación en diversos invertebrados antárticos (Peck et al. 2010). Sin embargo, existe la posibilidad de que la dieta entre años (1981, 1998 y 2012) se mantenga sin diferencias, a pesar del aumento de temperatura registrado (Meredith & King 2005), debido a la posible aclimatización que pudieran presentar invertebrados intermareales Antárticos (en particular Gondogeneia antárctica como alimento principal de H. antarcticus en Isla Rey Jorge), estando sujetos a fluctuaciones de temperatura diarias, que pueden alcanzar 18°C (Waller et al. 2006). Esto podría indicar una posible ventaja en el ambiente intermareal austral, respecto otro ambientes térmicamente más estables (por ejemplo, submareal), no obstante, son necesarios más estudios de tolerancia termal en distintos niveles de organización ecológica. AGRADECIMIENTO Gracias al proyecto M_10-11 financiado por INACH, a la Universidad de Valparaíso y a Matthew Lee por la identificación de anfípodos. REFERENCIAS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
218
Amundsen, P., Gabler, H. y Staldvik, F. 1996. A new approach to graphical analysis of feeding strategy from stomach contents data –modification of the Costello method. Journal of Fish Biology, 48:607-614. Bilyk, K. y DeVries, A. 2011. Heat tolerance and its plasticity in Antarctic Fishes. Comparative Biochemistry and Physiology, Part A, 158:382-390. Casaux, R. 1998. The contrasting diet of Harpagifer antarcticus (Notothenioidei, Harpagiferidae) at two localities of the South Shetland Islands, Antarctica. Polar Biology, 19:283-85. Chipps, S. y Garvey, J. 2007. Assessment of Food Habits and Feeding Patterns. En: Guy C & ML Brown (eds). Analysis and interpretation of freshwater fisheries data, pp. 473-514. American Fisheries Society, Bethesda, Maryland. Clark, M., Fraser, K., Burns, G. y Peck, L. 2008. The HSP70 heat shock response in the Antarctic fish Harpagifer antarcticus. Polar Biology, 31:171-180. Cortés, E. 1997. A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Canadian Journal of Fisheries and Aquatic Science, 54:726-738. Daniels, R. 1982. Feeding ecology of some fishes of the Antarctic Peninsula. Fishery Bulletin, 80:575-588. Doyle, S., Momo, F., Brêthes, J. y Ferreyra, G. 2012. Metabolic rate and food availability of the Antarctic amphipod Gondogeneia antarctica (Chevreux 1906): seasonal variation in allometric scaling and temperature dependence. Polar Biology, 35:413-424. Duarte, W. y Moreno, C. 1981. The specialized diet of Harpagifer bispinis: its effect on the diversity of Antarctic intertidal amphipods. Hydrobiologia, 80(3):241-250. Ducklow, H., Baker, K., Fraser, W., Martinson, D., Quetin, L., Ross, R., Smith, R., Stammerjohn, S. y Vernet, M. 2007. Marine pelagic ecosystems: the west Antarctic Peninsula. Philosophical Transaction of the Royal Society of London, B362:67-94. Ferry, L. y Caillet, G. 1996. Sample size and data analysis: are we characterizing and comparing diet properly? En: MacKinlay, D. y Shearer, K. (eds). Feeding, ecology and nutrition in fish, pp. 71-80. American Fisheries Society, San Francisco. Horn, H. 1966. Measurement of overlap in comparative ecological studies. The American Naturalist, 100:419-424. Hureau, J. 1970. Comparative biology of some Antarctic fish (Nototheniidae). Oceanographic Institute Bulletin of Monaco, 68:1-244 Hureau, J. 1990. Harpagiferidae. En: Gon, O. y Heemstra, P. (eds). Fishes of the Southern Ocean, pp. 357-363. Smith Institute of Ichthyology, Grahamstown, South Africa. Hyslop, J. 1980. Stomach contents analysis. A review of methods and their application. Journal of Fish Biology, 17:411-429. Krebs, C. 1998. Ecological methodology, 620 pp. Addison Wesley/Longman, Menlo Park. Meredith, M. y King, J. 2005. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophysical Research Letters, 32:L19604. Morley, S., Martin, S., Bates, A., Clark, M., Ericson, J., Lamare, M. y Peck, L. 2012. Spatial and temporal variation in the heat tolerance limits of two abundant Southern Ocean invertebrates. Marine Ecology Progress Series, 450:81-92. Peck, L., Morley, S. y Clark, M. 2010. Poor acclimation capacities in Antarctic marine ectotherms. Marine Biology, 157:2051-2059. Pinkas, L., Oliphant, S. y Iverson, I. 1971. Food habits of albacore, bluefin tuna, and bonito in Californian waters. California Fish and Game, 152:1-105. Target, T. 1981. Trophic Ecology and Structure of Coastal Antarctic Fish Communities. Marine Ecology Progress Series, 4:243-263. Vaughan, D., Marshall, J., Connolley, W., Parkinson, C., Mulvaney, R., Hodgson, D., King, J., Pudsey, C. y Turner, J. 2003. Recent rapid regional climate warming on the Antarctic Peninsula. Climatic Change, 60:243-274. Waller, C., Barnes, D. y Convey, P. 2006. Ecological contrasts across an Antarctic land–sea interface. Austral Ecology, 31:656-666. Whittaker, R. 1952. A study of summer foliage insect communities in the Great Smokey Mountains. Ecological Monographs, 22:1-44.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
219
ESTRUCTURA DE LA COMUNIDAD BACTERIANA DE SUELOS ANTÁRTICOS CONTAMINADOS CON HIDROCARBUROS DURANTE LA BIOESTIMULACIÓN CON
DIFERENTES NUTRIENTES Y UN SURFACTANTE
(Bacterial community structure of an Antarctic hydrocarbon-contaminated soil during the biostimulation with different nutrients and a surfactant)
Dias, R.L.1,2, Lo Balbo, A.3, Del Panno, M.T.1, Mac Cormack, W.P.3,4 1Centro de Investigación y Desarrollo en Fermentaciones Industriales (CINDEFI), Calles 50 y 115 (1900), La Plata, Argentina. [email protected]; 2Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Av. Rivadavia 1917 (1033), Buenos Aires (Bs. As.), Argentina. [email protected]; 3Cátedra de Biotecnología, FFyB, Universidad de Buenos Aires, Junín 956 6ºP (1113) Bs. As., Argentina; [email protected] 4Instituto Antártico Argentino. Cerrito 1248 (1010) Bs. As., Argentina. [email protected]. La bioestimulación es una estrategia de biorremediación que tiene como objetivo lograr una mayor velocidad de biodegradación de los contaminantes mediante el balance del contenido de nutrientes (especialmente N y P). Si bien se han obtenido resultados positivos al combinar esta estrategia con el agregado de compuestos tensioactivos, éstos no siempre han mostrado ser efectivos y muchas veces no son extrapolables a los ambientes de clima frío. En el caso del continente antártico, las temperaturas bajas constantes limitan fuertemente el metabolismo microbiano. Además, los sistemas y estrategias de biorremediación son, con mucha frecuencia, de naturaleza “sitio específica” por lo cual la optimización de la biorremediación de suelos antárticos debe ser realizada mediante ensayos on site (Yang Si-Zhong y col, 2009). Se ha observado que cuando un suelo es contaminado con hidrocarburos, la comunidad bacteriana autóctona sufre cambios en su composición y su dinámica. El estudio de estos cambios representa una herramienta clave para entender el efecto de cada tipo de contaminante sobre la estructura de las comunidades bacterianas naturales y como éstas evolucionan durante un proceso de biorremediación. El objetivo de este trabajo fue analizar on site, bajo las estrictas condiciones climáticas antárticas, el efecto que tienen distintas fuentes de nutrientes y la combinación de los mismos con un surfactante, sobre las comunidades bacterianas presentes y la eliminación de hidrocarburos en suelos crónicamente contaminados con gasoil. Para ello se utilizaron biopilas como diseño experimental, lascuales han sido reportadas previamente como eficientes en el saneamiento de suelos contaminados en ambientes fríos (Delille y col, 2008; Sanscartier y col, 2009). El estudio se realizó durante el verano antártico, y las biopilas consistieron en montículos de 30 kg de suelo crónicamente contaminado con gasoil antártico, dispuestos en bateas de plástico de 20 cm de altura para así aminorar los efectos de erosión causados por los fuertes vientos. Cada sistema fue bioestimulado con uno de los siguientes nutrientes: i) harina de pescado (HP), utilizada exitosamente por otros autores para estimular el crecimiento bacteriano y la biodegradación de hidrocarburos (Delille y col., 2008); ii) el fertilizante NPK, reportado previamente en estudios de biorremediación (Rosa y Triguis, 2007; Chorom y col., 2010) debido a su ventaja de liberación más lenta de los macronutrientes nitrógeno, fósforo y potasio y iii) una solución de sales inorgánicas de N y P (SI). Las sales se agregaron al inicio del estudio y luego se re-fertilizó con las mismas a los 10 y 30 días de tratamiento para reducir el efecto del lavado generado por las lluvias y el derretimiento de la nieve. Todos los nutrientes fueron combinados con el surfactante Brij700® debido a que, según bibliografía previa (Das y col., 2008),

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
220
resultó efectivo para aumentar la eficiencia de remoción de hidrocarburos en suelos contaminados. Se plantearon los siguientes microcosmos: CC (control no tratado), CCB (control con el agregado de Brij700®), SIB (bioestimulado con sales inorgánicas y con Brij700®), HPB (bioestimulado con harina de pescado y con Brij700®) y NPKB (bioestimulado con el fertilizante agronómico NPK y con Brij700®). El ensayo fue muestreado por triplicado a los 0, 5, 10 y 39 días de iniciado para cuantificar hidrocarburos totales (GC-FID) y estudiar la comunidad bacteriana. La estructura de la comunidad bacteriana se analizó mediante DGGE (electroforesis en gel con gradiente desnaturalizante), una de las técnicas moleculares más utilizadas para identificar la diversidad microbiana en muestras ambientales (Hendrichx y col., 2006). Los resultados mostraron que el sistema control (CC) exhibió al final del ensayo (t39) un porcentaje de eliminación de hidrocarburos totales (HT) del 19,3 %. Todas las biopilas bioestimuladas (SIB, HPB y NPKB) mostraron un descenso significativo en los valores de HT respecto a CC a t39 (p<0,05). Los valores obtenidos fueron de 11,8%, 16,8% y 16,0% respectivamente. La presencia de Brij700® en el suelo control (CCB) no causó diferencias significativas respecto a CC. El agregado de nutrientes produjo cambios considerables en las comunidades bacterianas presentes en el suelo en estudio. El dendrograma obtenido (Figura 1) a partir del gel de DGGE agrupó las muestras de las biopilas control (independientemente de la presencia o no del surfactante) en un cluster distinto al de las biopilas bioestimuladas. Los cambios causados por la presencia de sales y de harina de pescado determinaron que SIB y HPB se agruparan en un mismo sub-cluster. NPKB seleccionó comunidades bacterianas que formaron un sub-cluster separado del resto de los microcosmos.
Figura 1. Dendrograma UPGMA obtenido del análisis de cluster basado enla presencia/ausencia y abundancia de las bandas
del gel de DGGE en las diferentes biopilas a los días 0, 5, 10 y 39 de ensayo. Figure 1. UPGMA dendrogram resulting from cluster analysis of the data reporting the presence/absence and abundance of
DGGE bands in the different biopiles at days 0, 5, 10 and 39 of assay. La mayoría de las bandas obtenidas y secuenciadas a partir del gel de DGGE mostraron un alto porcentaje de similitud (94-100%) respecto de alguna secuencia disponible en la base de datos Genbank. Muchas de las secuencias relacionadas se aislaron de suelos y aguas contaminadas y de zonas de clima frío. Del análisis de las afiliaciones taxonómicas de las bandas secuenciadas puede resaltarse la

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
221
presencia, en todos los sistemas y en todos los tiempos, de miembros del Filum Proteobacteria (especialmente de familia Sphingomonadaceae). Se observó una gran proporción de miembros del Filum Actinobacteria (50%) pertenecientes a las familias Cellulomonadaceae, Propionibacteriaceae; Microbacteriaceae, Nocardioidaceae y Mycobacteriaceae. Cabe destacar que algunos géneros fueron encontrados sólo en los sistemas control (Agromyces) y otros sólo en los sistemas bioestimulados (Dokdonella). La baja eficiencia de remoción de la flora autóctona observada en el sistema control, notoriamente menor a la observada en ensayos previos, podría deberse en parte a la menor disponibilidad de oxígeno en comparación con la de otros sistemas experimentales, como las parcelas. Se debe tener en cuenta que las biopilas sufrieron anegamiento provocado por las constantes precipitaciones, evento que pudo haber disminuido también la disponibilidad del oxígeno, debido a la saturación de los microporos del suelo. La baja pero estadísticamente significativa remoción de HT mostrada por el sistema HPB podría relacionarse con su conocida capacidad para favorecer la porosidad y por ende la entrada de oxígeno al suelo, efecto que no fue significativo en ensayos previos realizados con el mismo suelo utilizando el diseño de parcelas (Dias y col., 2012). Por su parte, el agregado en etapas de las sales inorgánicas y la liberación lenta de nutrientes que proporciona el NPK pueden haber contribuido a la leve eliminación observada en esos sistemas. Respecto al análisis del dendrograma, la agrupación de CC y CCB a t0 en un cluster distinto al del resto de los t0 sugiere un rápido efecto del agregado de los nutrientes sobre las comunidades bacterianas. Este rápido efecto ya había sido observado anteriormente por el grupo de trabajo en un estudio en parcelas conteniendo suelos crónicamente contaminados (Vázquez y col., 2009). Rölling y col. (2004), trabajando con suelos agudamente contaminados, también observaron cambios significativos en la estructura de las comunidades bacterianas un día después de su fertilización con nutrientes inorgánicos. Todos los sistemas bioestimulados, se ubicaron y mantuvieron, a lo largo del ensayo, en un cluster distinto al de los sistemas CC. Estos resultados muestran la estricta selección de comunidades bacterianas diferentes generadas por el agregado de nutrientes, resultado también observado por Ros y col., (2010). La constante presencia de la familia Sphingomonadaceae en todos los microcosmos y en todos los tiempos refleja la capacidad de los miembros de esta familia para tolerar y proliferar en ambientes con alto grado de contaminación por hidrocarburos, así como su gran versatilidad y adaptación a los cambios ambientales. Esta capacidad fue previamente mencionada, tanto en suelos de clima frío (Bell y col., 2011) como de clima templado (Coppotelli y col., 2008). La gran proporción de miembros del Filum Actinobacteria es otro punto a resaltar, ya que su dominancia en suelos antárticos superficiales afectados por hidrocarburos no ha sido observada con frecuencia previamente, aunque sí ha sido mencionada en suelos antárticos prístinos (Saul y col., 2005). Es importante tener en cuenta que este suelo permaneció almacenado, luego de ser contaminado, durante al menos 2 años, muy compactado en barriles metálicos, con poco contacto con el aire atmosférico. Esto podría haber generado una condición de baja tensión de oxígeno y de transferencia de masa muy limitada. Se ha reportado que, bajo estas condiciones las poblaciones bacterianas se encuentran principalmente en un estado de muy baja tasa de crecimiento (Björklöf y col., 2008). Esas condiciones claramente favorecen el establecimiento de microorganismos con bajas tasas de crecimiento y alta capacidad de supervivencia a largo plazo, caracterizándose por una baja demanda energética, la cual es utilizada fundamentalmente como energía de mantenimiento (estrategas k). Este podría ser la causa de la importante proporción de actinobacterias observada en este estudio. El hallazgo de géneros en los microcosmos bioestimulados que no estaban presentes en los microcosmos control sugiere que miembros de estos géneros estaban presentes en la tierra sin bioestimular por debajo

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
222
del límite de detección y que luego aumentaron su número con el agregado de nutrientes. Los resultados observados mostraron que la estrategia de bioestimulación representa una herramienta válida para mejorar la remoción de hidrocarburos en suelos antárticos contaminados así como la falta de efecto de los tensioactivos. También confirman los notorios cambios que el agregado de nutrientes causa sobre la estructura de la comunidad bacteriana presente en el suelo bajo tratamiento. AGRADECIMIENTOS ANPCyT (PICTO N°0124) y UBA (UBACyT 20020100100378). REFERENCIAS BIBLIOGRÁFICAS Bell, T. H., Yergeau, E., Martineau, C., Juck, D., Whyte, L. G. y Greer, C. W. 2011. Identification of Nitrogen-Incorporating Bacteria in Petroleum-Contaminated Arctic Soils by Using [15 N] DNA-Based Stable Isotope Probing and Pyrosequencing. Applied and Environmental Microbiology, 4163-4171. Björklöf, K., Salminen, J., Sainio, P. y Jørgensen, K. 2008. Degradation rates of aged petroleum hydrocarbons are likely to be mass transfer dependent in the field. Environ Geochem Health, 30: 101-7. Chorom, M., Sharifi, H.S. y Motamedi, H. 2010. Bioremediation of a crude oil - polluted soil by application of fertilizers. Iran. J. Environ. Health. Sci. Eng, 7(4):319-326. Coppotelli, B.M., Ibarrolaza, A, Del Panno, M.T. y Morelli, I.S. 2008. Effects of the inoculant strain Sphingomonas paucimobilis 20006FA on soil bacterial community biodegradation in phenanthrene-contaminated soil. Microbial Ecology, 55:173-183. Delille, D., Duval, A. y Pelletier, E. 2008. Highly efficient pilot biopiles for on-site fertilization treatment of diesel oil-contaminated sub-Antarctic soil. Cold Regions Science and Technology, 54: 7–18. Dias, R. L., Ruberto, L.A.M., Vázquez, S.C., Hernández, E., Lo Balbo, A., Del Panno, M. T. y Mac Cormack, W.P. 2012. Bioremediation of an aged diesel oil-contaminated Antarctic soil: evaluation of the "on site" biostimulation strategy using different nutrient sources. Int Biodet Biodeg, 75: 96-103. Hendrichx, B., Dejonghe, W., Faber, F., Boënne W., Bastiaens, L., Verstraete, W., Top, E. M. y Springael, D. 2006. PCR-DGGE method to assess the diversity of BTEX monooxygenase genes at contaminated sites. FEMS Microbiol.Ecol, 55: 262-273. Röling, W. F., Milner, M. G., Jones, M., Lee K., Daniel, F., Swannell, R. J. P. y Head, I. M. 2002. Robust hydrocarbon degradation and dynamics of bacterial communities during nutrient-enhanced oil spill bioremediation. Appl. Environ. Microbiol, 68: 5537-5548. Ros, M., Rodríguez, I., García, C. y Hernández, T. 2010. Microbial communities involved in the bioremediation of an aged recalcitrant hydrocarbon polluted soil by using organic Amendments. Bioresource Technology, 101: 6916–6923. Rosa, A.P. y Triguis, J.A. 2007. Bioremediation process on Brazil shoreline. Laboratory experiments. Environ Sci Pollut Res Int, 14(7):470-6. Sanscartier, D., Zeeb, B., Koch, I. y Reimer, K. 2009. Bioremediation of diesel-contaminated soil by heated and humidified biopile system in cold climates. Cold Regions Science and Technology, 55:167-173. Saul, D.J., Aislabie, J., Brown, C.E., Harris, L. y Foght J.M. 2005. Hydrocarbon contamination changes the bacterial diversity of soil from around Scott Base, Antarctica. FEMs Microbiol Ecol, 53:141-155. Vázquez, B. Nogales, Ruberto L., Hernández E., Christie-Oleza J., Lo Balbo A., Bosch R., Lalucat J. y Mac Cormack W. 2009. Bacterial Community Dynamics during Bioremediation of Diesel Oil-Contaminated Antarctic Soil. S. Microbial Ecology, 57 (4): 598-610. Yang, S., Jin, H., Wei, Z., He, R., Ji, Y., Li, X. y Yu, S. 2009. Bioremediation of Oil Spills in Cold Environments: A Review. Pedosphere, 19(3): 371–381.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
223
EVIDENCIA DE ACTIVIDAD SURFACTANTE EN MICROORGANISMOS PSICRÓFILOS: UNA HERRAMIENTA BIOTECNOLÓGICA PARA LA BIORREMEDIACIÓN DE
HIDROCARBUROS
(Biosurfactant activity evidence in Psychrophiles Microorganisms: A Biotechnology Tool for Hydrocarbon Biorremediation)
Vera N3., Pradel P1,3., Loperena M2., y González A3.
1Programa de Doctorado en Ciencias de Recursos Naturales, Departamento de Química, Universidad de la Frontera., Temuco, Chile. 2Departamento de Bioingeniería, Universidad de la Republica, Montevideo, Uruguay. 3Laboratorio de Microbiología Ambiental y Extremófilos. Departamento de Recursos Naturales. Universidad de Los Lagos, Puerto Montt, Chile. E-mail: [email protected] RESUMEN Los hidrocarburos (HC) son un problema ambiental debido a que sus efectos son persistentes en el tiempo. Actualmente estos contaminantes tienen una amplia distribución geográfica, encontrándose frecuentemente cuerpos de agua como lagos, ríos, océanos, llegando incluso hasta el continente antártico. La prevalencia de los HC en la Antártica se debe principalmente por el mal manejo y transporte de estos compuestos. La estructura química de los HC tiene como resultante una lenta degradación y persistencia, lo que trae como consecuencia efectos negativos al medioambiente. Este problema ambiental ha generado la búsqueda de nuevas alternativas tecnológicas que ayuden a disminuir este tipo de contaminación, una de estas opciones es el uso de los biosurfactantes. Estas biomoléculas extracelulares producidas por microorganismos extremófilos debido a que presentan la capacidad de interactuar con compuestos hidrofóbicos e hidrofílicos como los HC, provocando un cambio en la tensión superficial. La ventaja de la producción de moléculas surfactantes en microorganismos antárticos permite una serie de adaptaciones biológicas para la vida en condiciones climáticas extremas. La presencia de este tipo de actividad en estos microorganismos aislados desde sectores antárticos contaminados sugiere una potencial aplicación en biorremediación a bajas temperaturas. Para ello, en este trabajo se aislaron bacterias antárticas desde sectores contaminados en la bahía Maxwell (S 62º 11' 47'', W 59º 56' 4,3'') medio mínimo (M9) con HC (aceite automotriz nuevo y usado) al 1% como fuente de carbono. Los cultivos se incubaron por 5 días a 4 ºC y 180 rpm. Los microorganismos se traspasaron a medio M9 sólido con HC al 1% a 4º C. El análisis morfológico de las colonias obtenidas reveló que corresponden a bacterias bacilares Gram negativas. Posteriormente se determinó y evaluó la actividad surfactante de cada cepa, utilizando la metodología del colapso de gotas, difusión de hidrocarburos, actividad emulsificante (E24) y concentración micelar crítica (CMC) propuestos por Youseff y col.. 2003, y Aparna ey col., 2012. Las perspectivas de esta investigación vislumbran la caracterización bioquímica de las moléculas surfactantes producidas por estas bacterias antárticas y su potencial aplicación en biorremediación a bajas temperaturas. INTRODUCCIÓN La contaminación por hidrocarburos (HC) es un problema que genera preocupación mundial debido a los daños que ocasionan a la salud y al medio ambiente. Estos compuestos presentan una amplia distribución geográfica debido al transporte de los HC en lagos, ríos, mares, océanos, etc. migrando hasta la zona Antártica (Bao y col., 2012). La presencia de estos contaminantes en estos sitios son ocasionados por el uso y manejo de combustibles derivados del petróleo y explotación de recursos petroleros en alta mar, los cuales frecuentemente producen derrames, ocasionando impactos negativos a

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
224
los ecosistemas marinos (Bao y col., 2012). Con el objetivo de contrarrestar los efectos nocivos causados por la presencia HC, se han desarrollado y aplicado técnicas físicas, químicas y biológicas que buscan remover el mayor porcentaje de este tipo de contaminante y disminuir su impacto. En los últimos años, las tecnologías aplicadas a la remoción de los contaminantes han sido utilizadas, donde la biorremediación ha logrado demostrar altos índices de eficacia (Yang y col., 2009). Actualmente dentro de estos procesos la utilización de moléculas biosurfactantes ha sido exitosa (Pacwa-Plociniczak y col., 2011). Los biosurfactantes son moléculas extracelulares complejas producidas por microorganismos que presentan la capacidad de interactuar con compuestos hidrofóbicos e hidrofílicos que generan un cambio en la tensión superficial que facilitan la degradación biológica y posterior mineralización de HC. Estos biomoléculas se clasifican según su estructura molecular, dentro de estos se encuentran, los glicolípidos (ramnolípidos), lipopéptidos, fosfolípidos, biosurfactantes poliméricos y ácidos grasos (Aparna y col., 2012). Actualmente estas biomoleculas presentan gran interés en los procesos de remediación de sitios contaminados con HC. Principalmente por su efectividad, los biosurfactantes ofrecen ventajas en su aplicación tales como, menor toxicidad, alta biodegradabilidad y presentar mayor estabilidad en condiciones extremas de temperatura, pH y salinidad, en contraste a los surfactantes químicos (Aparna y col., 2012). Los microorganismos antárticos presentan una serie de adaptaciones para vivir a bajas temperaturas y disponibilidad de fuentes de carbono limitada. Esta condición sugiere que las bacterias presentes en sitios contaminados con hidrocarburos presentan la capacidad de utilizar a los HC como fuente de carbono. METODOLOGÍA Las muestras fueron extraídas en la Bahía Maxwell (S 62° 11´, W 59° 56´4,3´) de las Islas Rey Jorge, Antártica. Los aislamientos de las cepas bacterianas se realizaron aeróbicamente a 20 ° C utilizando medio nutritivo mínimo (M9) suplementado con aceite automotriz usado al 1 % como única fuente de carbono. Además se adicionó como agente de selección nistatina 45 µg/ml y ácido nalidíxico 10 µg/ml. Posteriormente se realizó un análisis de morfología a través de la tinción de Gram. La evidencia de la actividad surfactante en los aislados antárticos fue demostrada a través de las pruebas clásicas para la determinación: Parafilm M y Colapso de gotas (Morikawa y col., 1993). La capacidad emulsificante fue determinada a las 24 h (E24) según metodología propuesta por Aparna y col., 2012. Finalmente, se determinó la adhesión bacteriana a los hidrocarburos utilizados (diésel y bencina) (Rosenberg y col., 1980). Para los todos ensayos cualitativos realizados a las bacterias aisladas, se utilizó un control positivo (C+) SDS 5% y blanco (BLC) agua destilada para ratificar los resultados obtenidos. RESULTADOS Aislamiento y caracterización morfológica de microorganismos antárticos, se aislaron 3 cepas bacterianas, denominadas A1R; A2R y A2RN. La tinción Gram de estas cepas antárticas reveló que corresponden a bacilos Gram negativos. En la Tabla 1 se muestra el ensayo de actividad surfactantes basado en el ensayo de parafilm M que mide la extensión del HC en presencia de agentes tensoactivos. En estas cepas se puede observar que todas tienen la misma capacidad de generar extensión del HC en un periodo de 2 min. Para la prueba del colapso de gotas se observa que la cepa A1R y A2RN son las que muestran mayor actividad surfactante lo que sugiere que la cantidad de agentes tensoactivos producidos por estas cepas es mayor y tiene alta actividad.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
225
Tabla 1. Ensayo Parafilm M (arriba) en tiempo 0 y 2 min., los resultados muestran al diámetro de dispersión del HC (mm) en
presencia de los cultivos de cada cepa. Ensayo colapso de gotas (abajo) en tiempo 0 y 2 min., corresponde al grado de actividad de la biosurfactaciòn. C+ (SDS 5%); BLC (Blanco, agua destilada) y las cepas aisladas (A1R; A2R; A2RN).
Table 1. Parafilm M assay (above) at 0 and 2 min. The results show that HC dispersion diameter (mm) in the presence of cultures of each strain. Drop collapse assay (bottom) at time 0 and 2 min., Corresponds to the degree biosurfactation activity .
C + (SDS 5%), BLC (White, distilled water) and the isolates (A1R, A2R, A2RN). En el análisis de la actividad surfactante observado en la Figura 1, muestra que el mayor porcentaje de emulsificación (E24) fue obtenido en la cepa A1R. Este índice sugiere que los agentes tensoactivos producidos por esta bacteria podrían facilitar la adhesión al hidrocarburo para su degradación y eventualmente mineralización. Sin embargo al caracterizar su capacidad de Adhesión Bacteriana a Hidrocarburos (BATH) se logró determinar que esta muestra una diferencia en función del HC con el cual se enfrente, en este caso es posible observar en la Fig 1 B, que el diesel tiene un alto porcentaje de adhesión con respecto a la gasolina (bencina). Esta adhesión podría estar influenciada por el alto porcentaje de alcanos saturados (parafinas y sus derivados) presentes en el diesel, no así en la bencina que su formulación comercial es un destilado de mayor pureza enriquecidos en alcanos de cadenas cortas. Además es posible que los estabilizantes utilizados en la gasolina tengan un rol no determinado y afecten este índice de adhesión en esta bacteria.
Figura 1: A) Índice de emulsión (E24) (en porcentaje) a las 24 h utilizando n-hexano, C+ (SDS 5%.). B) Porcentaje de
adherencia de bacterias antárticas A1R, A2R y A2RN a diesel y bencina. La cepa A1R y A2RN muestran adherencia para los dos hidrocarburos, mientras que, A2R presenta adherencia sólo para diesel. A1R, A2R y A2RN corresponden a las cepas
aisladas. Figure 1: A) Index Emulsion (E24) (percentage) after 24 h using n-hexane, C + (SDS 5%). B) Antarctic bacterial adherence (percentage) A1R, A2R and A2RN diesel and gas. A1R and A2RN strains showing adhesion to the hydrocarbons, whereas
only A2R diesel presents adhesion. A1R, A2R and A2RN correspond to isolates.
CONCLUSIÓN Los ensayos realizados en este trabajo demuestran que las tres cepas antárticas A1R, A2R y A2RN poseen actividad surfactante mediante la técnica de colapso de gotas. Se determinó que A1R posee un alto índice de surfactación y adherencia en presencia de hidrocarburos. Los resultados obtenidos sugieren que A1R, A2R y A2RN presentan capacidad surfactante utilizando como única fuente de carbono HC, por lo que estos microorganismos podrían ser una nueva fuente de estas biomoléculas

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
226
(surfactantes). Estos antecedentes sugieren su utilización como una herramienta biotecnológica de biorremediación para su aplicación en sitios contaminados con HC, derivados del petróleo, especialmente en zonas de baja temperatura. REFERENCIAS Aparna, A., Srinikethan, G. y Smitha, H. 2012. Production and characterization of biosurfactant produced by a novel Pseudomonas sp. 2B. Journal Colloids and Surfaces B: Biointerfaces 95, 23– 29. Bao, M., Wang L., Sun P., Cao, L., Zou J., Li, Y. 2012. Biodegradation of crude oil using an efficient microbial consortium in a simulated marine environment. Journal Marine Pollution Bulletin 64, 1177–1185. Morikawa, M., Daido, H., Takao, T., Murata, S., Shimonishi, Y. y Imanaka, T. 1993. A new lipopeptide biosurfactant produced by Arthrobacter sp. Strain Mis 38. Journal Bacteriology 175, 6459-6466 Pacwa-Płociniczak, M., Płaza, G. A., Piotrowska-Seget, Z., Cameotra, S. S. 2012. Environmental Applications of Biosurfactants: Recent Advances. International Journal of Molecular Sciences 12, 633-654. Rosenberg, M., Gutnick, D. y Rosenberg, E. 1980. Adherence of bacteria to hydrocarbons: a simple method for measuring cell-surface hydrophobicity. FEMS Microbiology Letters 9, 29-33 Yang, Z., Jin, J., Wei, Z., He, R., Ji, J., Li, M. y Yu, S. 2009. Bioremediation of Oil Spills in Cold Environments: A Review. Journal Pedosphere 19(3), 371–381. Youssef, N. H., Duncan, K. E., Nagle, D. P., Savage, K. N., Knapp, R. M., McInerney, M. J. 2003. Comparison of methods to detect biosurfactant production by diverse microorganisms. Journal of Microbiological Methods 56, 339– 347.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
227
CAMBIO CLIMÁTICO O CONDICIONES AMBIENTALES EXTREMAS: ADAPTACIÓN DEL EPILITON EN LAS ISLAS GREENWICH Y REY JORGE, ISLAS SHETLAND DEL SUR,
ANTÁRTICA
(Climate change or extreme environmental conditions: epilithon adaptation in the Greenwich and King George Islands, South Shetland islands – Antarctica)
Guerra C., M.L.1
1Universidad Internacional del Ecuador, Av. Simón Bolívar y Calle Jorge Fernández, Quito, Ecuador [email protected] RESUMEN Durante los veranos australes de los años 2009 y 2013 en las islas Rey Jorge y Greenwich respectivamente, se estudiaron especies de microfitobentos que son consideradas halotolerantes. En la isla Rey Jorge se consideraron cinco playas en Bahía Almirantazgo y en la isla Greenwich, cinco playas en Punta Fort William. Tanto en Rey Jorge como en Greenwich el deshielo de las elevaciones crea cursos de agua con salida al mar, estableciendo zonas estuarinas en el área de confluencia, donde se ha observado mayor crecimiento de fitobentos. El objetivo del estudio es conocer como algunas especies de microalgas son capaces de adaptarse a diferentes hábitats a pesar de que las condiciones ambientales son extremas. El epiliton en cada una de las playas fue colectado utilizando el método NAGISA. En cada playa, durante la marea baja, se midió el largo y el ancho. Se colocaron cuatro transectos de cinco estaciones; en cada uno y se colectó aleatoriamente el epiliton presente. La composición béntica en la zona estuarina fue de 35 taxones, compuestos por diatomeas y cianofíceas, mientras que en la zona intermareal sin influencia de agua dulce se encontraron 45 taxones, compuestos de diatomeas, cianofíceas fitobentónicas y algunas microalgas planctónicas. El género dominante, en la zona intermareal, en ambas islas fue Fragilariopsis, cuyas especies antárticas -entre otras adaptaciones- son halotolerantes. Las diatomeas en la región antártica son poco estudiadas aún, y su comportamiento es apenas conocido. Una ventaja de las especies endémicas antárticas es que tienen un amplio poder de adaptación a diferentes condiciones de temperatura, salinidad, nutrientes y, sobre todo, a la luz. Algunas algas sobreviven en ambientes extremos, protegiendo las paredes celulares de los agentes externos. Por otro lado, con una variabilidad climática, los organismos dominantes -en ambientes extremos como la Antártida- necesitan estar bien adaptados a una dinámica de regímenes diferentes que oscilan entre el agua dulce y la marina. La biomasa de organismos autótrofos como el microfitobentos, en ecosistemas acuáticos, está controlada, entre otros, por dos factores importantes: los nutrientes y la disponibilidad de luz solar. En la Antártica, los organismos que dependen de la luz han tenido que adaptarse bioquímicamente a condiciones extremas, sean estos períodos de luz solar permanente o períodos de oscuridad absoluta. Durante el verano austral, entre los meses de diciembre y abril, los organismos que viven de la fotosíntesis, como las algas, deben aprovechar el corto período de luz para poder realizarla; durante este período tienen un crecimiento acelerado por la gran cantidad de luz y la disponibilidad de nutrientes. Algunos desarrollan eficaces colectores de luz y otros emplean grandes cantidades de proteínas. Durante los seis meses de extrema oscuridad desactivan la fotosíntesis y se alimentan de los nutrientes que llegan con el agua. Durante el verano austral del 2009, entre los meses de enero y febrero, se estudió el crecimiento del microfitobentos (epiliton) en la zona intermareal de las playas Yellow Point, Refugio 1, Ipanema, Henniken, Punta Plaza, así como también aquellas ubicadas frente al Laboratorio de Química y a la Estación de Radio, del área administrada por Brasil, en Bahía Almirantazgo y en la isla Rey Jorge

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
228
(Antártica). Entre los meses de febrero y marzo del 2013 se estudió la biodiversidad de la comunidad microfitobentónica, específicamente de epiliton, presente en la zona intermareal de las playas que conforman Punta Fort William: Caletas Jambelí y Galápagos, Punta Hermosilla y Punta Troncoso e isla Greenwich (Antártica). En las dos islas funcionan bases antárticas: en Greenwich, sólo en verano y en Bahía Almirantazgo, hasta el 2011 durante todo el año. La zona intermareal de Yellow Point recibe el agua procedente del deshielo de la elevación aledaña; por otro lado, la zona intermareal entre Punta Hermosilla y Punta Troncoso recibe las aguas procedentes del río Culebra, que se forma por el deshielo del Glaciar Quito. Para realizar la colecta de las muestras, se utilizó la metodología NAGISA adaptada para el medio antártico. En la zona intermareal, durante la marea baja se midió la playa, se zonificó y se ubicaron los transectos; posteriormente, se establecieron las estaciones. Para la colecta se utilizó un cuadrante de 0,25 m2, a lo largo de los transectos; en cada estación se tomaron las muestras y réplicas. Para realizar una estandarización de todos los puntos de colecta, el substrato que contenía el epiliton tuvo aproximadamente las mismas dimensiones. En el laboratorio se extrajo el epiliton a través del método Peterson (et. Al, 1993). Posteriormente, se las fijó con lugol, para su posterior identificación. Se realizaron mediciones de factores físico-químicos y de radiación UV-B y UV-A. De manera general, en el análisis de las muestras de microfitobentos, obtenidas tanto en la isla Greenwich como en la isla Rey Jorge, presentan una mayor densidad poblacional las diatomeas (Bacillariophytas). En sitios donde hay mayor influencia antropogénica y colonias de pingüinos, los grupos dominantes fueron las algas azul-verdes (Cyanophytas) y verdes (Chlorophytas). La composición béntica en la zona estuarina fue de 35 taxones compuestos por diatomeas y cianofíceas mientras que en la zona intermareal sin influencia de agua dulce se encontraron 45 taxones, compuestos de diatomeas, cianofíceas y algas planctónicas. Por otro lado, el género dominante, en la zona intermareal, en ambas islas fue Fragilariopsis, diatomea cuyas especies antárticas son halotolerantes. En el caso de las Cianoficeas algunos géneros fueron abundantes, entre los que se encuentran Nostoc y Nodularia. Las diatomeas bentónicas son excelentes indicadores biológicos, sobre todo en la Antártica, en primer lugar porque es un componente común de las alfombras microbianas que se encuentran adheridas a las rocas; en segundo lugar, porque claramente reflejan los factores ambientales, como por ejemplo su estrecha relación con la conductividad eléctrica; y finalmente, porque sirve como indicadores ambientales no solo del presente, sino también el pasado, al permanecer sus frústulas un largo período depositadas (Ohtsuka, Taisuke et al. 2006). Las cianofíceas son algas que son conocidas por su característica de colonizar un rango diverso de ambientes, que incluyen rocas, glaciares, diferentes estratos de hielo, riachuelos, lagos y lagunas, y en las regiones polares aún es posible encontrarlas en suelo polares. Estudios realizados en los ambientes antártico y ártico sugieren que por la lenta especiación, en combinación con eficiencia de las cianofíceas, para dispersarse, el endemismo es raro. La mayoría de las formas identificadas aparecen como taxones cosmopolitas. Sin embargo, la simplicidad morfológica de la cianofíceas enmascara un alto nivel de variabilidad genética, un ejemplo de ello es Phormidium autumnale es una especie común en las regiones polares; sin embargo, recientes estudios de este taxón demuestra una diferencia en su pigmento y características de crecimiento (Vincent, W.F., 2000). Las cianofíceas son raras en ambientes marinos; sin embargo, en la Antártida el agua del deshielo es un soporte para el crecimiento de cianofíceas. Las formas más comunes incluyen Nostoc commune y miembros de las oscilatorias dominados por Nodularia, Calothrix, Scytonema y Tolypothrix. La disponibilidad de agua y la salinidad aparecen como el mayor determinante que influencia la distribución de estas cianobacterias (Vincent, W.F. 2000).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
229
Con relación a la radiación ultravioleta (UV) en la Antártica muchas especies, principalmente las cianofíceas, forman oscuras incrustaciones. Estas ocurren en hábitats que son periódicamente suministrados por nieve derretida, comúnmente dominado por Gloeocapsa spp. La coloración oscura se debe a la película UV del pigmento scytonemin. Aunque esta comunidad puede resistir la radiación solar y disecación, esta parece ser intolerante a las condiciones de salinidad (Vincent et al. 1993; Vincent and Quesada, 1994, Quesada y Vincent, 1997). Para llegar a conclusiones sobre qué mecanismos llevan a ciertas especies a ser halotolerantes, es necesario profundizar en estudios sobre la fisiología y el comportamiento de ciertas especies; no obstante, de manera general se conoce que las diatomeas (Basillariophytas) son dominantes en el ecosistema Antártico. En el caso de las Cianofíceas presentan una gran capacidad de colonización de los diferentes ambientes, lo que las ubica como organismos de fácil adaptación. AGRADECIMIENTOS: INAE, Estación Comandante Ferraz y todas las personas que colaboraron durante esta investigación. REFERENCIAS BIBLIOGRÁFICAS Ohtsuka, T., Kudoh, S., Imura, S. y Shuji, O. 2006. “Diatoms composing benthic microbial mats in freshwater lakes of Skarvsnes ice-free area, East Antartica”. In Polar Bioscience, 20, 113-130. Roberts, D. y McMinn, A. 1999. “Diatoms of the saline lakes of the Vestfold Hills, Antarctica” in Biblioteca Diatomológica, Band 44. Sabbe, K., Verleyen, E., Hodgson, D., Vanhoutte, K. y Vyverman, W. 2003. “Benthic diatom flora of freshwater and saline lakes in the Larsemann Hills and Rauer islands, East Antarctica” in Antarctic Science 15 (2) 227-248. Vincent, W.F. 1988. Microbial ecosystems of Antartica. Cambridge University Press, U.K. Vincent, W.F. et al. 1993. “Antarctic cyanobacteria: light, nutrients and photosynthesis in their microbial mat environment” in J. Phycol 29: 745-755. Vincent, W.F. y Quesada, A. 1994. “Ultraviolet radiation effects on cyanobacteria: implica-tions for Antarctic microbial ecosystems”. Antarctic Res Series, 62:111-124. Vincent, W.F. y Quesada, A. 1997. “Microbial niches in the polar environment and the escape from UV radiation in non-marine habitats” in: Battaglia B, Valencia J. and Walton D. (eds), Antarctic Communities: Species, Structure and Survival, pp. 388-395. Cambridge University Press, U.K. Vincent, W.F. 2000. “Cyanobacterial Dominance in the Polar Regions” in Whitton, B.A. and Potts, M. (eds) The Ecology of Cyanobacteria, pp.321-340. Kluwer Academic Publishers.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
230
DISTRIBUCIÓN DE COMPUESTOS TRAZA CON EFECTO CLIMÁTICO Y ACTIVIDAD MICROBIANA EN EL OCÉANO ÁRTICO DURANTE EL VERANO BOREAL,
RESULTADOS PRELIMINARES DEL CRUCERO LOMROG III
(Distribution of trace compound with climate effects and the microbial activity in the Arctic Ocean during boreal summer, preliminary results from LOMROG III Cruise)
Verdugo J.1, Farías L1, Diez B.2, Damn E.3, Sylvander P.4 y P. Snoeijs Leijonmalm4
1Depto. de Oceanografía, Laboratorio de Procesos Oceanográficos y Clima (PROFC) y Centro de Cambio Climático y Resilencia (CR)2, Universidad de Concepción. Concepción, Chile. E-mail: [email protected] 2Depto. Genética Molecular y Microbiología, Fac. de Cs. Biológicas. P. Universidad Católica de Chile, Stgo. Chile. 3Alfred Wegener Institute for Polar and Marine Research, P.O. Box 12061, 27515, Bremerhaven, Alemania. 4Depto. de Ecología. Stockholm University. Estocolmo, Suecia.
Las bajas temperaturas encontradas en aguas de los océanos polares como el Ártico y el Antártico, los cuales experimentan estacionalmente grandes avances y retrocesos de hielos, no limitan o excluyen la presencia y la actividad microbiana (Arrigo y col., 2012). Por el contrario, el hielo marino y el agua oceánica subyacente representan unos de los biomas más activos y dinámicos del planeta. Estos ecosistemas tienen significativos efectos sobre la transferencia de energía y materia, es decir sobre ciclos biogeoquímicos y en particular, sobre gases traza con efecto climático que modifican el balance radiativo de la atmósfera, y se constituyen como los principales forzantes del clima de la Tierra. Estos son los casos de gases de origen biogénico como el metano (CH4), óxido nitroso (N2O), ambos gases de efecto invernadero, y el Dimetil Sulfuro (DMS), precursor de los aerosoles y con un efecto enfriante en la atmósfera. La concentración de gases biogénicos depende de la actividad metabólica de los microbios, a la que se le debe sumar factores físicos como las bajas temperaturas, que facilitan la solubilidad de los gases atmosféricos, y al efecto de la formación y derretimiento de hielos (Randall y col., 2012). Durante décadas se pensó que el hielo marino actuaba como una barrera inerte para las interacciones océano-atmósfera-biosfera. Sin embargo, se conoce que durante el otoño-invierno o periodo de formación de hielo marino, los gases son expulsados del hielo y transportados a las profundidades del océano o ventilado a la atmósfera, mientras que durante en primavera-verano, el derretimiento de hielo sin gases diluye el agua de mar, generando una subsaturación de gases, condición que pudiera actuar como sumidero de gases en la atmósfera (Randall y col., 2012). Es así que estos ecosistemas se pueden comportar como fuentes o sumideros de gases, aunque el rol y la real magnitud de este intercambio aún no se conocen con certeza. El objetivo de este estudio fue caracterizar la distribución de CH4, N2O, DMSP (precursor del DMS) y otras variables oceanográficas y su variabilidad espacial, además de relacionarlos con procesos oceanográficos y biogeoquímicos en transectas a lo largo y a través del Ridge Lomonosov; este último ubicado sobre las cuenca de Makarov, Nansen y Amundsen, las cuencas más profundas del Océano Ártico, cercanas al Polo Norte, en un área totalmente cubierta de hielo. Dicha investigación fue realizada a bordo del rompehielos ODEN en el crucero LOMROG desde el 31 de Julio al 14 de Septiembre 2012, (Snoeijs-Leijonmalm y col., 2012). En dicho crucero fueron muestreadas estaciones de hielo (por medio de helicópteros) y de agua (usando CTD acoplado a una roseta) (Fig. 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
231
Figura 1: a) Distribución de las estaciones de muestreos (puntos negros); b) Roseta con botellas Niskin para extracción de
agua de mar; c) perforaciones por donde se extrajo el agua del “brine” y d) testigos de hielo. Figure 1: a) Spatial distribution of sampling station (black points); b) Rosette with Niskin Bottles to collect water samples; c)
brine water; d) ice cores. A partir de la distribución de salinidad y temperatura (Fig. 2a y 2b), en la banda latitudinal entre los 85-90°N y zonal entre los 70°W y 70°E, se observa una capa superficial más fría y menos salina que caracteriza la presencia de Agua de Mezcla Polar con densidades entre 24,78 a 26,70 kg m-3, siendo menos densa a medida que nos acercamos al Polo Norte. Por debajo de esta capa, se observa una haloclina y una inversión térmica (1°C) que es demarcada por el Agua del Atlántico y que se localiza entre los 500-1000 m. Por debajo de los 1500 m, las cuencas del Océano Ártico están ocupadas por Agua Profunda del Ártico con salinidades y temperaturas de 34,90 y ~ -0,9°C respectivamente, las que conducen a una densidad promedio de 28,10 kg m-3. Respecto al contenido de N2O, las aguas superficiales desde los 10 m de profundidad (cubiertas permanentemente por una capa de hielo de ca. 1,5 metros) pasan por una transición entre supersaturadas, con niveles de hasta 174% en la parte este de la cuenca adyacente al Mar de Kara, a subsaturada en su parte central cercana al Polo Norte y ligeramente saturada o cercana a su equilibrio en la parte oeste del Océano Ártico (ver Fig. 2a). La sobresaturación de N2O (>100%) podría deberse a la actividad nitrificadora, tal como ha sido reportado en ecosistemas Antárticos (Priscu y col., 1990). Por debajo de la capa superficial, las concentraciones de N2O disminuyen con la profundidad y las cuencas de Makarov, Nansen y Amundsen se encuentran ocupadas por agua subsaturadas en este gas. Aunque existen registros de subsaturación en el océano, dada la alta capacidad que tienen estas aguas para acumular gases, los mecanismos que pudieran explicar estos niveles sería una mezcla de masas de agua con bajos niveles de N2O desde su sitio de formación o procesos biológicos que lo consuman; relativo a esto, la presencia de organismos fijadores podrían dar cuanta de un sumidero de este gas (Farías y col., 2013).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
232
Figura 2: a) Distribución espacial de salinidad y b) temperatura durante el crucero LOMROG III en el verano boreal de 2012.
Figure 2: a) Salinity distribution and b) temperature distribution, during LOMROG III Cruise, during a boreal summer of 2012.
Figura 3: Distribución espacial de N2O (nM y % saturación con respecto a la concentración atmosférica), durante el crucero
LOMROG III, en el verano boreal de 2012. Figure 3: Spatial N2O distribution (nM and saturation % respect to the atmosphere concentration), during LOMROG III
Cruise, in a boreal summer of 2012. El contenido de Dimetilsulfopropionato (DMSP), se observa en la Figura 3b. El DMSP es una molécula orgánica, producida por el fitoplancton marino y que da origen al DMS, un compuesto sulfurado volátil precursor de aerosoles en la atmósfera. En la Fig. 3b se observa un núcleo de concentración con niveles de hasta 59 nM en la zona cercana al Polo Norte sobre la cuenca bañada con aguas de origen Atlántico.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
233
La concentración de estos elementos trazas con influencia climática se relaciona con la actividad microbiana medida en el agua de poro del hielo (brine) y en agua de mar por debajo de la cubierta de hielo; resultados preliminares indican que la actividad biológica en términos de fijación de carbono (carbono inorgánico como bicarbonato y orgánico como glucosa) y nitrógeno (tanto de amonio y nitrato) es mayor en aguas de la cuenca Este del Océano Ártico, respecto a la cuenca Oeste; y un orden de magnitud mayor en el agua de mar por debajo del hielo, que en los poros intersticiales del hielo. Por último, cabe señalar que un florecimiento de Phaeocystis sp. fue encontrado en una estación cercana al Polo Norte, esta especie de microalga, aunque cosmopolita y con un habilidad fisiológica sorprendente, ha sido reportado con mayor frecuencia en océanos marginales de la Antártica y del Ártico (Schoemann y col., 2005). No obstante, este es el primer reporte de su presencia en charcas (pond) cercano al Polo Norte. Por lo tanto, es primordial estudiar la dinámica de gases traza biogénicos con efecto directo sobre el balance radiativo del planeta y los procesos microbiológicos que median el reciclaje de estos gases. Esto es especialmente importante, en ecosistemas polares que han estado experimentando cambios ambientales notablemente visibles en las últimas décadas, debido principalmente al Calentamiento Global. AGRADECIMIENTOS Swedish Research Council VR 2012-04-12, PI Pauline Snoeijs Leijonmalm, Centro de Cambio Climático y Resiliencia (CR)2; FONDECYT # 1120719 REFERENCIAS BIBLIOGRÁFICAS Arrigo, K., Perovich, D., Pickart, R., Brown, Z., Van Dijken, G., Lowry, K., Mills, M., Palmer, M., Balch, W., Bahr, F., Bates, N., Benítez-Nelson, C., Bowler, B., Brownlee, E., Ehn, J., Frey, K., Garley, R., Laney, S., Lubelczyk, L., Mathis, J., Matsuoka, A., Mitchell, B., Moore, G., Ortega-Retuerta, E., Pal, S., Polashenski, C., Reynolds R., Schieber, B., M. Sosik, H., Stephens, M. y Swift, J. 2012. Massive Phytoplankton Blooms Under Arctic Sea Ice. Science, 336 (6087); 1408. Farías, L., Faúndez, J., Fernández, C., Cornejo, M., Sanhueza, S. y Carrasco, C. 2012. Biological N2O fixation in the eastern South Pacific Ocean. PloS One 8 (5): e63956. Priscu, J., Downes, M., Priscu, L., Palmisano, A. y Sullivan, C. 1990. Dynamics of ammonium oxidizer activity and nitrous oxide (N2O) within and beneath Antarctic sea ice. Marine Ecology - Progress Series, 62 (97): 37-46. Randall, K., Scarratt, M., Levasseur, M., Michaud, S., Xie, H., y Gosselin, M. 2012. Arctic sea ice: source or sink for nitrous oxide. Journal of Geophysical Research: Oceans, 117, Schoemann, V., Becquevort, S., Stefels, J., Rouseau, V. y Lancelot, C. 2005. Phaeocystis blooms in the global ocean and their controlling mechanisms: a review. Journal of Sea Research, 45 (1-2); 43-66. Snoeijs, P., Sylvander, P., Díez, B. y Farías, L. 2012. Microbial communities in the Arctic Ocean and their contribution to global nitrogen cycling. In: Marcussen C and the Lomrog III Scientific Party. Lomonosov Ridge off Greenland 2012 (Lomrog III) - Cruise Report. Danmarks og Grønlands Geologiske Undersøgelse Rapport 2012/119: 99-115.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
234
LA BÚSQUEDA DEL MEJOR LUGAR PARA OBSERVACIONES ASTRONÓMICAS EN EL ANTÁRTICA OESTE
(The Search for the Best Astronomical Observations Site in Western Antarctica) Mark Falvey1 and Patricio Rojo2
1Departamento de Geofísica, Facultad de Ciencias Físicas y Matemáticas, Universidad de Chile 2Departamento de Astronomía, Facultad de Ciencias Físicas y Matemáticas, Universidad de Chile INTRODUCTION There is ever growing interest in astronomical research in Antarctica. Despite its reputation for having an inhospitable climate, it has been realized that sites within the Antarctic interior may actually have the driest, calmest, longest, and clearest nights in our planet. Indeed, logistical difficulties aside, the Antarctic continent is widely regarded as the world's best site for most forms of southern-hemisphere astronomy. Thus far, most international attention has focused on the high altitude Ice Domes of Eastern Antarctica (EA), where observing campaigns, robotic telescopes (dome A) and fully functioning observatories (dome C, South Pole) have already been successfully installed. In contrast, Western Antarctic (WA), where Chile's bases and territorial claim are located, has been largely unstudied, even though the available data suggest that the area may share many of the favorable characteristics of the EA sites. In this study we examine the potential of the WA for astronomy, based on the analysis of the meteorological characteristics of the region using data from high resolution atmospheric simulations with the Weather Research and Forecasting (WRF) mesoscale model. In the next section the study area and methodology are described. In section 3 we present the key results obtained thus far. Our conclusions are presented in section 4. METHODOLOGY Our study examines the astronomical potential over the spatial region claimed by Chile (Territorio Antártico Chileno) in western Antarctica, ranges from 53W to 90W and from the South Pole to 60S, overlapping the Argentine and British Antarctic claims. The major landform of the area is the Antarctic Peninsula, extending from its northernmost tip at 62S to the Antarctic mainland at around 75S. While this mountainous peninsula may offer some interesting prospects for astronomy, the most promising locations are likely to be found further southwards, on the Antarctic mainland. For example, along the eastern edge of the Ronne Ice Shelf the runs the small but extremely high Ellsworth mountain range. The Vinson Massif (summit of the Ellsworth Mountains) at 4892m is in fact the highest peak in the Antarctica continent and this fact alone makes it an interesting candidate for a hypothetical observatory. To the southwest of the Ellsworth range towards the South Pole the topography rises again to the summit of the WA Ice Sheet, over 3000 m high and situated just to the west of Chiles territorial claim. Given the near total absence of metrological observations in the relevant parts of our study area, we must resort to indirect methods to obtain initial estimates of site suitability. The principal tool employed in this research project is a high resolution mesoscale atmospheric model to simulate the detailed characteristics of WA's atmosphere. Mesoscale models are a mature technology widely used in both atmospheric research and operational forecasting. Such models provide a complete representation of atmospheric phenomenon over a spatial scales ranging from a few hundreds of meters to thousands of kilometers. Mesoscale models represent the state of the atmosphere on a 3D grid with intervals of a few

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
235
kilometers in the horizontal and a few tens of meters in the vertical near to the surface. The model resolution plays a pivotal role in determining the ability of the model to represent small scale meteorological features (particularly important in the boundary layer). Higher resolutions are generally better, but the computational cost increases dramatically. For a given spatial region, a model of 1 km resolution requires nearly 1000 times more processing power to complete a simulation in the same time as a 10 km resolution run. The atmospheric state is defined in terms of a set of 10 or so variables that include wind velocities, temperature, pressure, humidity, clouds, ice and rain. Based on an initial condition (usually derived from large scale gridded metrological analysis) the mesoscale model integrates the equations that govern the time evolution of these variables. During the course of the model integration, the atmospheric state is periodically saved to disk (usually at hourly intervals) for later analysis. On modern computer systems it is feasible to run simulations of the atmosphere for months or even years, and in this way, build up digital climatologies that characterize the spatio-temporal variability of variables of interest.
Figure 1. Study area and comutational domains for the numerical atmosferic simulations. Topography is based on the 1km
RAMP database. Figura 1. Área de estudio y dominios comutacionales para las simulaciones numéricas atmosfericas . Topografía se basa en la
base de datos RAMP 1 km. In this work we applied the Polar version of the Weather Research and Forecasting Advanced Research WRF (WRF-ARW) model (Klemp et al., 2007) version 3.3 (www.wrf-model.org) developed by the Ohio State University (Hines and Bromwich, 2008) . This version of the model includes several modifications to the standard model code, including an optimal surface energy balance and heat transfer scheme for sea ice and permanent ice surfaces. Numerical experiments were conducted for two regions: The first was a large scale simulation at 3km resolution using a modeling domain of approximately 1000 km by 1000 km. This domain extends from the southern part of the Antarctic Peninsula to about 83S, and covers the WA ice dome and the Ellsworth Mountains. The simulation was run for the entire winter period of the year 2011 and provides a useful description of the general low level circulation

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
236
characteristics of the region. Based on the results of this simulation, a second high resolution (1km) simulation was run for the Ellsworth Mountains and surrounding icefields (figure 1), also for the entire winter period of 2011. RESULTS We present results for three key atmospheric variables known to be of primary importance in determining astronomical observing site suitability. These variables are summarized in the table below:
Variable Definition Comments Boundary layer wind speed
Wind speed in the lowest layers (0-100m) of the atmosphere.
A shallow boundary layer and low wind speeds are especially important in for high quality astronomical work, as strong winds will cause the telescopes to vibrate. Many potential sites over the Antarctic continent are discounted due the presence of very strong winds in the boundary layer (the so called Katabatic winds) that develop due to the extreme cooling of the near surface air over the Antarctic ice fields.
Integrated water vapor
Total water vapor content of the atmosphere, usually expressed in mm of water
Dry conditions are ideal for most forms of astronomy and the extreme aridity of the Antarctic atmosphere is one of its most attractive attributes.
Cloudiness Frequency of cloud occurrence. Usually expressed as a percentage of total possible observing time
Few astronomical observations are possible when clouds are present. A very low level of cloudiness is necessary to justify an Antarctic observing site.
Table 1: Variables used to examine site suitability. Tabla 1: Variables utilizadas para examinar la idoneidad de los sitios.
This is not an exhaustive list. Other variables, such as free atmosphere turbulence, temperature and aerosols are also of great importance. Furthermore, non-meteorological factors such as distance to Antarctic bases, accessibility and the presence of exposed bedrock may also be decisive in observatory site selection. The figure 2 shows spatial maps of the variables listed in table 1 both for the focus region centered on the Sentinel Range of the Ellsworth Mountains that was simulated with the high resolution model domain. Water vapor levels are very low over of the study area and not surprisingly show a strong inverse relationship with terrain height. As such lowest water vapor levels are simulated over the high peaks of the Sentinel Range. The maps of cloudiness indicate that winter clouds are frequent over northern and western parts of the study area. However, to the west of the the simulated cloud frequency drops dramatically and reaches levels that could be interesting for high quality astronomy. The maps of mean boundary layer wind speed indicate that fairly strong winds prevail over much of the study area. Winds are particularly strong over the Ellsworth Mountains, reaching mean values of over 10 m /s which effectively eliminate this region as a prospect for astronomy. However, to the west of these mountains, several plateaus of relatively low mean wind speed can be discerned.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
237
Figure 2: Maps of simulated water vapor (a), cloudiness (b) and wind speed (c) over the focus area of our study (Sentinel
Range in the Ellsworth Mountains). Brighter (whiter) areas are more favorable for astronomical observing. Figura 2: Mapas de simulación de vapor de agua (a), nubosidad (b) y velocidad del viento (c) en el área foco de nuestro
estudio (Sentinel Range en las montañas Ellsworth). Las áreas más brillantes (blancos) son más favorables para la observación astronómica.
CONCLUSIONS Our results (although still somewhat preliminary) indicate that there may well be promising sites for astronomical observatories within the Chilean Antarctic claim. Contrary to our initial expectations, the high peaks of the Ellsworth Mountain range do not offer the most favorable sites due to the very strong wind speeds that are simulated over the mountains. On the other hand, the ice rises to the west of the Ellsworth Mountains, which reach over 2600m in some places, appear to offer good conditions for high quality astronomy, characterized by very low mean water vapor, low cloudiness and low to moderate wind speeds. It must be stressed that our results thus far are based entirely on the output of numerical simulations, and there is a need to validate results against available observations. ACKNOWLEDGEMENTS This research was funded by the Instituto Antartico Chileno project INACH G19_11

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
238
GEOQUÍMICA Y MINERALOGÍA DE LOS SEDIMENTOS DE PENÍNSULA Y CALETA POTTER, ISLA 25 DE MAYO (KGI): SUS IMPLICANCIAS EN LOS AMBIENTES
SEDIMENTARIOS ACTUALES DE ANTÁRTIDA
(Geochemistry and mineralogy of Potter Cove and Potter Peninsula sediments, 25 de Mayo (KGI) Island: their implications on current sedimentary environments of Antarctica)
Manograsso Czalbowski, N. T.1, Monien, P.2 , Moretti, L.3 , Chaparro, M.A.E.4, Böhnel, H.N.5, Morrás,
H.3, Paladino, I.R.3, Brumsack, H. J.2 and B. Schnetger2 1Instituto Antártico Argentino-Dirección Nacional del Antártico (IAA-DNA). Balcarce 290, C.P. 1064, CABA, Argentina [email protected] 2Institut für Chemie und Biologie des Meeres (ICBM) Carl-von-Ossietzky Str. 9-11,26129 Oldenburg,Germany [email protected], [email protected], [email protected]
3Instituto Nacional de Tecnología Agropecuaria (INTA) CIRN, Instituto de Suelos. N. Repetto y Los Reseros S/Nº (1712), Castelar, Buenos Aires, Argentina [email protected], [email protected], [email protected] 4IFAS-CONICET (Instituto de Física Arroyo Seco UNCPBA, Argentina). Pinto 399, Tandil (7000), Argentina [email protected] 5Centro de Geociencias (UNAM, México). Blvd. Juriquilla 3001, Querétaro (76230), México [email protected]
RESUMEN La influencia del material particulado en suspensión (MPS) en las comunidades bentónicas marinas ha sido ampliamente estudiada. Dentro del mismo, los óxidos e hidróxidos de hierro son muy importantes ya que son fácilmente reducidos en la descomposición de la materia orgánica y posteriormente metabolizados por los organismos. Este trabajo constituye un aporte a las investigaciones oceanográficas y biológicas de caleta Potter porque genera información referida a la composición y procedencia de sus sedimentos, centrándose mayormente en los minerales portadores de hierro. OBJETIVOS Y METODOLOGÍA A fin de establecer correlación entre los sedimentos de la península y la caleta Potter, se reconocieron las distintas cuencas de drenaje asociadas y se recolectaron 45 muestras de sedimento superficial correspondientes a distintas unidades geomorfológicas. En el sector marino se obtuvieron 53 muestras de sedimento superficial y se extrajeron dos testigos (P06 y P08), de los cuales se obtuvo muestras con un intervalo de 1 cm para análisis geoquímicos y 2 cm para los mineralógicos (Figura 1). Las muestras fueron liofilizadas y tamizadas para obtener la fracción granulométrica menor a 2 mm. En todas las muestras se determinaron elementos mayoritarios y minoritarios a través de análisis cuantitativo por fluorescencia de rayos X (XRF). En muestras selectas se analizaron tierras raras (REE) a través de espectrometría ICP-MS. El estudio mineralógico se centró en los sedimentos fluviales y en los testigos marinos (35 muestras). La mineralogía se determinó por difracción de rayos X (DRX) en polvo total (fracción <2 mm) y en muestras de arcilla (<2 mµ) orientadas naturales (N), glicoladas (EG) y calentadas (C) a 520ºC durante dos horas. A fin de caracterizar la mineralogía magnética, se realizaron estudios de: susceptibilidad magnética, histéresis magnética, magnetización remanente isotérmica (MRI) y mediciones termomagnéticas. RESULTADOS Y DISCUSIÓN

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
239
El estudio de la geología del sector se ve limitada sólo a las zonas libres de hielo. La geología subyacente a los glaciares debe deducirse por las islas rocosas rodeadas por el campo de hielo (Nunataks) y los afloramientos al pie del mismo. La estratigrafía y geoquímica de península Potter (Kraus, 2000) corresponde a una secuencia volcánica terciaria que se inicia con basaltos toleíticos aflorantes en la zona costera al oeste de la península (GPS). Suprayaciendo se encuentran las basandesitas de las elevaciones centrales de la península (CPP). Le suceden basandesitas del Nunatak Yamana o Florence (NY) al este de la caleta y por último las andesitas del Cerro Tres Hermanos (CTH) (Figuras 1 y 2 A). Además de la evolución magmática evidente, existe una diferenciación en la señal del Eu denotada en un enriquecimiento de dicho elemento en GPS y PP, y en un empobrecimiento en NY y CTH (Figura 2 B).
Los sedimentos en su totalidad presentan una señal geoquímica andesítica a basandesítica, y una anomalía negativa de Eu (Figura 2). Esto sugiere que todos los sedimentos de la caleta y península provienen de la erosión de las rocas más jóvenes (NY y CTH), ubicadas en las posiciones más altas del relieve. Los afloramientos del centro de la península (CPP), aunque son de una extensión considerable y se encuentran en sectores topográficos relativamente elevados, no presentan un aporte significativo a la cuenca (Figura 1). A fines comparativos, en la Figura 3 se muestran los difractogramas de una muestra representativa de sedimento fluvial (MS9) y de un testigo marino (P06). En las muestras de polvo total de ambos ambientes se reconocieron plagioclasa, cuarzo, laumontita (mineral del grupo de las ceolitas) y hematita. Por su parte, la goethita es un constituyente importante de los sedimentos fluviales, mientras que en los sedimentos marinos su presencia es minoritaria. De manera opuesta, el clinocloro es importante en la muestra de polvo total de la caleta, mientras que en las del chorrillo es menor. En la fracción arcilla de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
240
ambos ambientes se reconocieron clinocloro, esmectita e illita. No obstante se encontraron las siguientes diferencias: sólo en la muestra fluvial se observan laumontita y goethita, mientras que únicamente en la caleta hay plagioclasa y existe más proporción de esmectita e illita.
Figura 3: Difractogramas de muestra fluvial (MS 9): A) polvo total B) arcillas. Difractogramas de la caleta (P06 17-19 cm):
C) polvo total D) arcillas. N: natural, EG: glicolada, C: calcinada. Clc: clinocloro, Sme: esmectita, Plg: plagioclasa, Qtz: cuarzo, Hem: hematita, Lmt: laumontita, Gt: goethita, Ill: Illita, Kln: caolinita
Adicionalmente, los estudios de remanencia y termomagnéticos realizados en muestras fluviales revelan la presencia de óxidos de hierro (ferrimagnéticos y antiferromagnéticos). En la Figura 4 se presentan resultados representativos de dos muestras fluviales: MS9 y MS50. Aunque estas muestras presentan altas coercitividades de remanencia (Hcr de hasta 96.7 mT), a partir de la separación de fases de la curva de MRI se observan tres componentes, una de baja coercitividad: (titano)magnetita, y dos de alta coercitividad: hematita y goethita. En forma independiente los resultados de la curva M(T) revelan temperaturas de Curie y de Néel correspondientes a los (hidr)óxidos de hierro mencionados. Estas fases de alta coercitividad, también son observadas a partir de los estudios de RX (Figura 3). La susceptibilidad magnética (κ) es una medida de la concentración de los minerales (dia, para, ferri y antiferro)magnéticos. En la Figura 5 se observa una disminución de κ en la parte superior de ambos testigos de la caleta. Esto puede deberse a una destrucción parcial de los minerales magnéticos por reducción del hierro, o a una mayor concentración de minerales antiferromagnéticos (hematita y goethita) sobre minerales ferrimagnéticos del tipo magnetita. Tampoco debe descartarse el efecto de “dilución” de la señal magnética debido al mayor contenido de MO. La primera hipótesis es validada por un aumento de Fe2+ del agua poral (por reducción del Fe3+ de los óxido e hidróxidos de hierro) en los primeros 15 cm de ambos testigos, principalmente en el testigo P08 (Figura 5). Refuerzan la segunda hipótesis un aumento relativo de goethita y hematita en la superficie del testigo P06 observado en los difractogramas y a que un comportamiento similar fue observado en la columna de sedimentos de lagos próximos. En tales sedimentos, la zona de disminución de la susceptibilidad magnética está dominada por fases de alta coercitividad (minerales antiferromagnéticos).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
241
CONCLUSIONES El elevado contenido de materia orgánica en los sedimentos marinos es suficiente para remover por completo la goethita en un período cercano a un año, teniendo en cuenta la vida media de este mineral por efectos de reducción bacteriana (Raiswell. and Canfield, 2012). La composición geoquímica del agua poral refuerza esta hipótesis. En tal sentido la goethita no podría utilizarse como indicador de procedencia de sedimentos de la caleta. Se asume que los restantes minerales de la fracción arcilla son totalmente detríticos y pueden dar información del área de aporte. Además, el comparar sólo esta fracción minimiza el control de la granulometría sobre la mineralogía. Las diferencias en las asociaciones mineralógicas de ambos ambientes indican que los arroyos de península Potter no son la única fuente de sedimentos de la caleta. Según el modelo propuesto por Monien (2013, com. pers.), tendría una mayor importancia (aproximadamente 90%) el aporte del agua de fusión subglacial del campo Fourcade al norte y este de la caleta. La corriente marina procedente del noroeste que circula en sentido horario en la caleta también exportaría el material de los chorrillos de Potter hacia afuera de la misma, impidiendo su sedimentación. REFERENCIAS Kraus S.,2000. Geologie, Vulkanologie und Geochemie tertiärer Vulkanite auf der Potter Halbinsel, King George Island, West-Antarktis (Geology, volcanology and geochemistry of Tertiary volcanics on the Potter Peninsula, King George Island, West Antarctica). Diploma (MSc) Thesis, LMU Munich: Faculty of Geosciences: 77 p. Raiswell, R. and Canfield, D.E., 2012. The Iron Biogeochemical Cycle Past and Present. Geochemical Perspectives 1(1), p. 220.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
242
MINERALOGÍA DE LOS SUELOS DE LA PENINUSLA BYERS, ISLA LIVINGSTON
(Mineralogy of soils peninsula Byers, Livingston Island)
Moura, P.A.1; Francelino, M. R.2; Fernandes Filho, E.I.3 e Schaefer, C.E.R.G.3
1Adnet Florestal, Rua Jorge José Pedro, Nova Friburgo, RJ, Brasil. 2Instituto de Floresta, Universidade Federal Rural do Rio de Janeiro, BR 465 km 07 Seropédica, Rio de Janeiro, Brasil, [email protected]. 3Departamento de Solos, Universidade Federal de Viçosa, MG, Brasil. [email protected] INTRODUCCIÓN A formação de solos na Antártica está restrita às áreas livres de neve e gelo permanente, correspondente a 0,35 % do seu território (Campbel e Claridrige 1987, Ugolini e Bockheim 2007). A maioria dos estudos pedológicos da Antártica foi conduzida na sua parte continental, em especial na região dos vales secos das montanhas Transantárticas. Já os solos formados nas regiões de clima mais ameno e úmido da Antártica Marítima e ilhas Sub-antárticas receberam menor atenção (Bockheim e Tarnocai, 1998, Michel et al. 2006, Simas et al. 2007). Na Antártica marítima, apesar de condições extremas de clima, existem coberturas microfíticas de liquens, algas, briófitas e raras plantas superiores, as quais são capazes de fixar o C atmosférico e incorporá-lo ao solo, sendo sua conservação auxiliada pelas baixas temperaturas que retardam a mineralização (Simas et al. 2006). As áreas livres de gelo da península Byers, na Ilha de Livingston, estão sujeitas aos processos de intemperismo físico e químico de forma mais intensa que nas áreas expostas da Antártica continental. Em Byers os processos químicos são inibidos pelo frio e os processos físicos são favorecidos pelos ciclos de congelamento e descongelamento da água no solo, condições também verificadas por Francelino (2004) e Simas, et al (2007) nas áreas livres de gelo da Baía do Almirantado. Ainda assim, as temperaturas mais amenas e maior disponibilidade de água líquida na península propiciam um intemperismo químico mais acentuado e consequentemente uma pedogênese mais acelerada, sobretudo nas áreas sob influência de atividades ornitogênicas. Porém, o intemperismo físico é mais acentuado e o principal fator responsável pela formação de solos na Antártica em geral. Nestas condições, a mineralogia dos solos deve refletir fortemente às características do material de origem, o qual varia fortemente na península Byers. O objetivo deste trabalho foi caracterizar a mineralogia dos solos presentes na porção norte da península Byers, Ilha de Livingston. MATERIALES Y MÉTODOS O estudo foi realizado na península Byers, localizada na parte oeste da Ilha Livingston, Arquipélago das Shetlands do Sul (62°37' S, 61°06'W) e representa a maior e mais antiga área livre de gelo do arquipélago, ocupando uma área de aproximadamente 60,6 km2, (Lopez-Martinez, 1996). Esse estudo foi realizado numa área de aproximadamente 3.416 hectares (34,16 km2) na parte norte dessa península (Figura 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
243
Figura 1: Localización del área de estúdio.
Figure 1: Location of the study area. As amostras de solos foram coletadas em 23 perfis, distribuídos de forma a cobrir as variações na geologia e geomorfologia da área de estudo. Para o estudo da mineralogia da fração argila foram extraídas amostras deste material por sifonamento em água deionizada, sem o uso de substâncias dispersantes. Posteriormente foram montadas lâminas e analisadas por difratometria de raio-X como o uso de um equipamento PANalytical X´Pert pro, utilizando-se radiação CoKα, com ângulo de varredura de 4 a 50 º2θ. Os minerais foram identificados conforme Chen (1977). Para identificação dos minerais 2:1 foram realizados tratamentos de saturação por Mg, saturação por Mg com Glicerol, saturação com K e saturação com K e aquecimento à 550 oC.
RESULTADOS Y DISCUSIÓN Os resultados serão apresentados em função do material de origem presente no local de amostragem. Solos derivados de tufos vulcânicos A mineralogia da fração argila indicou a presença de argilominerais 2:1 como clorita, e ilita. A clorita foi identificada por espaçamento de 14,9 Å, não apresentando expansão quando solvatadas com glicerol, com espaçamento de 14,05. Não apresentou também alteração significativa com a saturação por K nem com o aquecimento a 550 oC. A presença de rochas com colorações esverdeadas pode ser atribuída à presença de clorita. A análise DRX também indicou a presença de quatzo (4,26 Å e 3,34 Å), plagioclásio (3,79 Å) mirabolita (3,252 Å e 3,20 Å), titanita (2,998 Å ) e hematita (2,51 Å ). A presença do quartzo na fração argila indica forte ação do intemperismo físico nesse local. Solos derivados de basalto As análises DRX revelaram a presença dos argilominerais Clorita, Esmectita e Ilita. Foram também detectados traços de Quartzo, Plagioclásio, Hematita e Chamosita, além do mineral Titanita. A presença de quartzo na fração argila sugere que o intemperismo físico é tão expressivo que é capaz de fragmentar o quartzo até a granulometria de argila. Por outro lado sugere que estes solos

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
244
recebem influência de outro material de origem diferente do basalto. Isso pode ocorrer através do transporte de sedimentos por solifluxão em área de contato entre materiais de origem diferente. Solos derivados de tufo vulcânico misto com brecha vulcânica e basalto Esse grupo apresentou os maiores teores de Ca em média, sendo encontrado valor médio de 43 cmolc/dm3, em função principalmente da presença de calcita (CaCO3), comprovada pelas análises mineralógicas (DRX), que também indicou presença de esmectita e plagioclásio, minerais ricos em sódio (Na+2). A esmectita foi identificada pela reflexão (001) a 14,968 Å nas amostras saturadas por Mg2+ e quando solvatadas com Glicerol observou-se uma expansão da camada para o espaçamento 18,194 Å. Com a saturação por K ocorre colapso da estrutura com deslocamento do pico para 12,578 Å. Com o aquecimento a 550ºC, ocorre novo colapso da estrutura, com deslocamento do pico para 11,910 Å. Na argila natural foram observados picos de segunda ordem da esmectita em 7,513 Å. Com reflexão em 5,079 Å fica evidente a presença de ilita e o pico em 4,495 Å indica a presença de Ilita-Esmectita-regular 1:1, indicando a ocorrência de intemperismo químico. A presença de hematita foi identificada pelos picos em 2,698 Å e 2,519 Å. A presença de plagioclásio, mineral de fácil intemperismo, na fração argila indica o baixo grau de intemperismo químico do material de origem. Solos derivados de argilitos marinhos, arenitos e conglomerados Os solos desse grupo apresentaram difratogramas muito similares, com a presença de Esmectita, Ilita, Quartzo, Plagioclásio e Chamosita, além de Clorita e Vermiculita com hidroxi entrecamadas (VHE). Dentre os solos estudados, os desse grupo foram os mais intemperizados, com pedogênese mais acentuada e com evidências de intemperismo químico mais intenso quando comparado com os solos dos demais grupos. Em parte isso ocorre pelo fato de tratar-se de material de origem sedimentar. Solos ornitogênicos A mineralogia dos solos com influência de atividade da avifauna foi distinta dos demais grupos, com exceção dos solos derivados de argilitos marinhos. Os difratogramas apresentaram picos pouco definidos e com muito ruído. Isso se deve, provavelmente, às interferências da matéria orgânica, que não foi eliminada antes das análises e a presença de minerais amorfos de baixa cristalinidade. Desta forma a análise DRX ficou comprometida nesses perfis, tendo sido, identificados, entretanto a presença de plagioclásio, rhodochrosita e hematita. CONCLUSIONES Os solos presentes em Byers são jovens e pouco desenvolvidos, muito pobres em matéria orgânica e variam significativamente em função do material de origem. Os solos desenvolvidos sobre influência ornitogênica são muito distintos dos demais, apresentando teores mais elevados de matéria orgânica. Nessas áreas os processos de intemperismo químico são muito mais expressivos que nos demais solos, ainda assim, o intemperismo físico predomina. A ocorrência de argilominerais na fração argila e minerais facilmente intemperizáveis indicam o pouco desenvolvimento dos solos e o predomínio do intemperismo físico sobre o químico. AGRADECIMIENTOS Ao CNPq pela bolsa de doutorado e a Marinha Brasileira pelo apoio logístico.
REFERENCIAS Bockheim, J.G., Tarnocai, C. 1998. Recognition of cryoturbation for classifying permafrostaffected soils. Geoderma 81:281-293. Campbell, I.B., Claridge, G.G.C., 1987. Antarctica: Soils,Weathering Processes and Environment. Elsevier, Amsterdam. 368 pp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
245
Francelino, M. R. 2004. Geoprocessamento aplicado ao monitoramento ambiental da Antártica Marítima: solos, geomorfologia e cobertura vegetal da península Keller. Tese de Doutorado, Viçosa, UFV. 102p. López-Martínez, J., Serrano, E. e Martínez DE Pisón, E. 1996. Geomorphological features of the drainage system. In López-Martínez, J., Thomson, J.R.A. and Thomson, J.W. (Eds.) Geomorphological map of Byers Peninsula, Livingston Island. BAS GEOMAP Series, Sheet 5-A, 15-19. Cambridge, British Antarctic Survey. Michel, R.F.M., Schaefer, C.E.G.R., Dias, L., Simas, F.N.B., Benites, V., Mendonça, E.S., 2006. Ornithogenic Gelisols (Cryosols) from Maritime Antarctica: pedogenesis, vegetation and carbon studies. Soil Sci. Soc. Am. J. 70, 1370–1376. Simas, F.N.B., C.E., Shaefer, G.R., Melo, V.F., Guerra, M.B.B., Saunders, M., Gilkes, R.J., 2006. Clay-sized minerals in permafrost-affected soils (Cryosols) from King George Island, Antarctica. Clays Clay Miner. 54, 721–736. Simas, F.N.B., Schaefer, C.E.G.R. Melo, V. F.. Albuquerque-Filho, M.R. , Michel, R.F.M., Pereira, V.V. Gomes, M.R.M.; da Costa L.M. 2007. Ornithogenic cryosols from Maritime Antarctica: Phosphatization as a soil forming process. Geoderma, 138 (3-4), p 191-203. Ugolini, F.C., Bockheim J. G., 2007. Antarctic soils and soil formation in a changing environment: A review, Geoderma, 144 ( 1-2) p 1-8.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
246
REGISTRO POLÍNICO DE MUESTRAS DE SUELO SUPERFICIAL EN ANTÁRTICA MARÍTIMA
(Pollen record of superficial soil samples in the maritime Antarctic)
Rondanelli-Reyes, M. J. (1, 2), Troncoso-Castro, J.M. (1) y Casanova-Katny, A. (3)
1Laboratorio de Palinología. Departamento de Ciencia y Tecnología Vegetal. Escuela de Ciencias y Tecnologías, Universidad de Concepción, Campus Los Ángeles. 2Pasante Sabático, Departamento de Proyectos, Instituto Antártico Chileno (INACH). 3Laboratorio de Microbiología. Facultad de Ciencias Biológicas, Universidad de Concepción, Concepción. E-Mail correspondencia: [email protected] RESUMEN En el contexto general, existe poca información disponible sobre el transporte aéreo de microorganismos desde territorios septentrionales hacia la Antártica y sus ecosistemas insulares asociados, esto incluye, no solo información taxonómica de lo que llega sino también aspectos relacionados sobre su viabilidad, duración en suspensión (distancia recorrida efectiva) y establecimiento gravitacional (Bera y Khandelwal, 2003). No obstante, existen algunos registros bien documentados del transporte de microorganismos al ambiente antártico, ecosistema en donde cobra importancia poder establecer además, si estos microorganismos detectados son elementos de paso o residentes, especialmente considerando la presión selectiva que el ambiente polar antártico ejerce per se. Las fuentes potenciales de colonos aerotransportados son amplias, pudiendo estos provenir de cubiertas vegetales, de animales, corrientes marinas, suelo y también, del hombre (Harmata y Olech, 1991; Pearce et al., 2009). En relación a micropartículas de origen vegetal existen algunos estudios relacionados con el transporte de polenesporas desde regiones subantárticas a Antártica, a través de la circulación de las masas aéreas que circundan la región y en donde existe evidencia de que el continente helado actuaría como un excelente captador del polen transportado a través del viento, el cual provendría de las islas subantárticas o de la vegetación del extremo sur de Sudamérica, incluyendo especies nativas y no nativas (Linskens et al., 1993). Lewis Smith (1991), reporta uno de los escasos registros relacionados con la presencia de palinomorfos en Antártica marítima, con la determinación de taxa que incluyen angiospermas, gimnospermas y hongos, estudiados en muestras provenientes tanto de suelo antártico, como del análisis de musgos. Todos los taxa no nativos determinados están representados en la flora existente en la región del extremo sur de Chile y Argentina. Considerando que la vegetación de las Islas Malvinas guarda estrecha relación con la vegetación patagónica de la región de Magallanes y de Tierra del Fuego, toda esta región debiera ser considerada fuente potencial de emigración de estos propágulos hacia Antártica. El tema no está suficientemente acotado, toda vez que además de necesitar más información taxonómica relacionada con el tipo de especie vegetal que está llegando a Antártica marítima, se hace necesario conocer el patrón de conducta de desplazamiento de los propágulos relacionado con las variables climáticas y también, con el grado de influencia que pudiese tener la entrada cada vez más recurrente del hombre en este ecosistema el cual, con la ayuda del calentamiento global, está recibiendo periódicamente menos nieve y hielo, permitiendo que el flujo de científicos (con sus equipos y maquinarias) y de turistas en tierra firme aumente, lo que indirectamente permitiría la entrada de nuevos inmigrantes no nativos a la Antártica marítima (Cuba et al., 2013). La pregunta es: ¿inmigrantes de paso? o ¿ya establecidos?

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
247
El objetivo general de la presente investigación es conocer la taxonomía de lo que está llegando, como propágulo inmigrante, hacia la Antártica marítima. MATERIALES Y MÉTODOS Durante el verano austral de 2013, a través de la Expedición Científica Antártica (ECA) No 49, organizada por el Instituto Chileno Antártico (INACH), y al alero del Proyecto Fondecyt 1120895, se tomaron muestras de suelo superficial en un transecto longitudinal de aproximadamente 800 km a través de las Islas Shetland del Sur y la Península Antártica, cruzando el límite del círculo polar antártico, con el objetivo de caracterizar las propiedades químicas de los suelos en que crecen las plantas vasculares nativas de la región. Cien gramos de suelo fueron colectados en cada punto muestreado, que de norte a sur del transecto, corresponden a las localidades de Juan Carlos Point (Península Fildes), Collins Harbour (Bahía Fildes), Arctowski (Bahía del Almirantazgo, Isla Rey Jorge), Península Byers (Isla Livingstone), Isla Galindez (Islas Argentinas) e Islas Anchorage, Jenny y Lagotellerie, en Bahía Margarita. Las muestras se mantuvieron congeladas a -20ºC hasta trasladarlas a los laboratorios respectivos en Concepción. Submuestras de estos suelos fueron enviadas al Laboratorio de Palinología de la Universidad de Concepción, Campus Los Ángeles, con el fin de determinar, si lo hubiere, el contenido polínico de las muestras y la taxonomía de las especies encontradas. El tratamiento aplicado a las muestras de suelo fue el clásico para palinología (Faegri e Iversen, 1989), con adición de ácido fluorhídrico y uso de acetólisis. Para realizar el análisis microscópico se montó una alícuota del concentrado polínico, de volumen conocido, en solución de Hoyer glicerinado (Anderson, 1954). Las muestras fueron almacenadas en la palinoteca del laboratorio. Los palinomorfos presentes fueron observados con un microscopio trinonocular planacromático Olympus CX31RTSF adosado a una cámara CCD Microimagi 5.1 megapixeles; luego fueron digitalizados utilizando el software Micrometrics SE Premium 4. RESULTADOS Los resultados preliminares de la determinación palinológica de las muestras analizadas evidencian una interesante flora polínica presente en los suelos superficiales de la Antártica marítima. Todas las muestras hasta ahora analizadas son fértiles polínicamente, por lo que nuestra primera conclusión es que existe influjo polínico a lo largo de todo el transecto muestreado, incluso más allá del límite del círculo polar antártico. El resultado taxonómico de las especies determinadas indica la presencia de esporas de hongo, helechos y granos de polen de gimno y angiospermas (Figura 1). El grupo dominante corresponde al de las angiospermas con representantes de las Familias Poaceae, Caryophyllaceae, Juncaceae, Cyperaceae, Urticaceae, Nothofagaceae, Rosaceae, Fabaceae, Asteraceae. Las gimnospermas presentes corresponden a las Familias Ephedraceae y Podocarpaceae. Los helechos determinados corresponden a los Géneros Hymenophyllum y Polypodium. Existen además, ciertos pólenes encontrados que precisan aún más estudio para su determinación. Las especies polínicas hasta ahora determinadas nos informan de una fuente de origen vegetal característica de ambientes de estepa patagónica y bosques subantárticos, lo que se corrobora con el estudio previo de Lewis Smith (1991). Para poder establecer el desplazamiento que permite que estos propágulos alcancen estas latitudes se hace necesario implementar mecanismos de captura polínica en la Península Antártica, también idealmente, en la Isla del Cabo de Hornos y cotejar los patrones actuales de circulación atmosférica de la región. No debe dejar de considerarse si la presencia de alguno de estos pólenes es producto de una antesis local, resultado de un potencial establecimiento de alguna planta no nativa, como ya ha sido documentado en la literatura científica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
248
Figura 1: A, Asteraceae; B, Fabaceae; C, Juncaceae; D, Cyperaceae; E, Nothofagaceae; F, Podocarpaceae. Figure 1: A, Asteraceae; B, Fabaceae; C, Juncaceae; D, Cyperaceae; E, Nothofagaceae; F, Podocarpaceae.
AGRADECIMIENTOS Se agradece al Proyecto Fondecyt No 1120895 y al Instituto Antártico Chileno (INACH).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
249
REFERENCIAS BIBLIOGRÁFICAS Anderson, L. E. 1954. Hoyer's solution as a rapid mounting medium for bryophytes. Bryologist 57:242-247. Bera, S. K. and khandelwal, A. 2003. Aerospora over Southern Ocean and Schirmacher oasis, East Antarctica. Current Science, 85(2):137-140. Cuba-Díaz, M., Troncoso, J. M., Cordero, C., Finot, V. L. and Rondanelli-Reyes, M. 2013. Juncus bufonius, a new non-native vascular plant in King George Island, South Shetland Islands. Antarctic Science 25(3):385-386. Faegri, K. and Iversen, J. 1989. Textbook of pollen analysis. Chichester, New Jersey: Wiley, The Blacburn Press. Harmata, K. and Olech, M. 1991. Transect for aerobiological studies from Antarctica to Poland. Grana 30:458-463. Linskens, H. F., Bargagli, R., Cresti, M. and Focardi, S. 1993. Entrapment of long-distance transported pollen grains by various moss species in coastal Victoria Land Antarctica. Polar Biol 13: 81–87. Pearce, D. A., Bridge, P. D., Hughes, K. A., Sattler, B., Psenner, R. and Rusell, N. 2009. Microorganisms in the atmosphere over Antarctica. FEMS Microbiol Ecol 69:143-157. Lewis Smith, R. I. 1991. Exotic sporomorpha as indicators of potential immigrant colonists in Antarctica. Grana 30:313-324.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
250
DESARROLLO DE UN SISTEMA DE AREAS MARINAS PROTEGIDAS EN EL SECTOR OCCIDENTAL DE LA PENINSULA ANTARTICA Y ARCO DEL SCOTIA
(Development of a system of Marine Protected Areas in the West Antarctic Peninsula and Scotia Arc)
Arata, J.A.1, Marschoff, E.2, Gaymer, C.3 y F. Squeo4
1Instituto Antártico Chileno. Plaza Muñoz Gamero 1055. Punta Arenas, Chile. Email: [email protected] 2Instituto Antártico Argentino. 3Universidad Católica del Norte. 4Universidad de La Serena. INTRODUCCIÓN La CCRVMA es una organización internacional fundada en 1982, amparada bajo el Sistema del Tratado Antártico, encargada de la conservación de los ecosistemas marinos antárticos. Esto no excluye la explotación de los recursos, siempre que sea hecha de manera sostenible y tenga en cuenta los efectos de la pesca (ej., pesca de kril y bacalao de profundidad) en otros componentes del ecosistema. En concordancia con los acuerdos alcanzados en el Convenio de Diversidad Biológica (CDB), en cuya 10a Conferencia (Aichi, Japón) acordó para el 2020 que los países deben proteger un 10% de sus ecosistemas marinos más relevantes, la CCRVMA decidió en su XXIV reunión (2004) desarrollar un sistema de Áreas Marinas Protegidas en el área de la Convención; actualmente el trabajo se ha dividido en 9 dominios, 3 de los cuales tienen AMPs aprobadas o propuestas (Fig. 1).
Figura 1. Área de la Convención (CCRVMA) (línea negra, gruesa), indicando la distribución de los 9 dominios identificados
para desarrollar propuestas de AMPs. Se indica además la única AMP aprobada (área en gris) a la fecha (junio 2013). Propuestas para los dominios 7 y 8 están bajo análisis por la CCRVMA. Coordenadas: Latitud cada 10°S y Longitud cada
45°E/W. Figure 1. Convention Area (CCAMLR) (black bold line) indicating the distribution of the 9 domains identified for
developing AMP proposals. The only MPA approved to date (June 2013) is also shown (grey area). Proposals for domains 7 and 8 are under consideration by CCAMLR, Coordinates: Latitude every 10°S and Longitude every 45°E/W.
Chile y Argentina han decidido liderar el desarrollo de un sistema de AMPs para el Dominio 1, que incluye la costa occidental de la península Antártica, las islas Shetland del Sur y Orcadas del Sur (Fig. 1). El progreso realizado a la fecha se presenta en este trabajo. Esta presentación busca informar a la comunidad científica Latinoamericana sobre el desarrollo de esta importante tarea, la cual se espera tenga beneficios a nivel de todo el Océano Austral, así como invitar a quienes cuenten con información y/o expertise relevante al proceso a sumarse a este esfuerzo. DATOS Y MÉTODOS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
251
Un primer Taller de Trabajo fue coordinado por Chile y realizado en Valparaíso, 28 mayo a 1 junio 2012, al cual asistieron delegados de 6 países miembros de CCRVMA (EEUU, Reino Unido, Noruega, Japón, Argentina y Chile); la reunión fue financiada parcialmente con fondos de la CCRVMA.
Objetivos Específicos Capa Realizada 1. Ejemplos representativos de hábitats bentónicos Bioregionalización bentónica (Douglass et al. 2011) SI
2. Ejemplos representativos de hábitats pelágicos Bioregionalización pelágica (Raymond et al. 2011) SI 3. Important benthic ecosystem processes a) Large-scale canyons SI
b) Smaller-scale canyons SI c) Benthic areas under ice-shelves SI d) Up/down-welling and mixing areas NO
4. Large-scale pelagic ecosystem processes a) Áreas con alta productividad primaria (clorofila a superficial) SI b) Predictable highly productive areas – water column NO c) Up/down-welling and mixing areas NO d) Frentes oceanográficos (ACCF) SI e) Zona de borde hielo marino (verano) SI f) Polinias SI g) Other dynamic / important areas NO
5. Important (spatially constrained/predictable) areas for mammal and bird life-histories
a) Áreas de forrajeo de especies que se reproducen en tierra (pingüinos, lobos marinos)
SI
b) Distribución de especies presa NO c) Áreas de forrajeo durante el invierno: -borde del hielo marino en Julio -distribución de pingüinos, lobos marinos, focas y ballenas en invierno
SI
6. Important (spatially constrained/predictable) areas for fish life cycles
a) Áreas de reclutamiento de: Notothenia rossii, Gobionotothen gibberifrons
SI
7. Important (spatially constrained/predictable) areas for zooplankton life cycles
NO
8. Rare or unique habitats/features Montes Submarinos y Shackleton Ridge SI 9. Ecosistemas Marinos Vulnerables Según lineamientos de CCRVMA SI 10. Áreas Referencia para estudios científicos a) Sitios actuales de investigación (ej., CEMP) SI
b) Áreas históricas sin pesquería SI c) Áreas históricas con pesquerías SI d) ZAEPs y ZAEA SI e) AMP de South Orkney SI
Tabla 1. Listado de objetivos de conservación y objetos de conservación específicos (capas) identificados en el Taller de Valparaíso (2012).
Table 1. List of conservation objectives and specific conservation objects (layers) identified at the Valparaiso Workshop (2012).
Este primer taller identificó los objetivos específicos de conservación a considerar, así como la información asociada a cada uno de éstos (Tabla 1), en base a los principios delineados por CCRVMA para designar AMPs (Medida de Conservación 91-04) y que son:
I. La protección de ejemplos representativos de ecosistemas, biodiversidad y hábitats marinos en una escala apropiada para la conservación de su viabilidad e integridad a largo plazo;
II. La protección de procesos ecosistémicos, hábitats y especies importantes, incluidas las poblaciones y etapas de los ciclos de vida;
III. El establecimiento de áreas de referencia científica para el seguimiento de la variabilidad natural y de cambios a largo plazo, así como para el seguimiento de los efectos de la explotación y otras actividades humanas sobre los recursos vivos marinos antárticos y sobre los ecosistemas de los cuales forman parte;
IV. La protección de áreas vulnerables al impacto de las actividades del hombre, incluidos los hábitats excepcionales, raros o de gran biodiversidad y múltiples atributos;
V. La protección de atributos esenciales para el funcionamiento de los ecosistemas regionales;

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
252
VI. La protección de áreas para mantener la capacidad de recuperación o de adaptación a los efectos del cambio climático.
Asimismo, se acordaron 3 fuentes de impacto antrópico, vinculados a pesca y turismo (Tabla 2). Una vez reunidas las capas de información geográfica correspondientes a cada objetivo específico, se procederá a realizar un análisis de identificación de las áreas de mayor valor para la conservación, contrastadas con las áreas de impacto antrópico. Este análisis se realizará mediante el software Marxan (Ball et al. 2009). Un análisis preliminar de esta información será realizada en Septiembre 2013, cuyos resultados serán presentados en el Simposio 7: Océano Austral.
Actividad Capa Realizado Distribución pesquería de kril Esfuerzo de pesca (lances) SI Distribución barcos de turismo Tracks de barcos de turismo de la IAATO NO Sitios visitados por turistas Frecuencia de uso de los diferentes sitios SI Tabla 2. Identificación de las fuentes de impacto antrópico utilizados como capas de ‘costo’ en los análisis.
Table 2. Identification of the main anthropogenic impacts used as ‘cost’ layers for the analyses. RESULTADOS PRELIMINARES Un total de 21 objetos de conservación, agrupados en 10 objetivos y 3 fuentes de perturbación antrópica han sido colectados, descritos y transformados en capas de información geográfica (shapefiles) (Tablas 1 y 2). La información colectada a la fecha incluye una extensa colección y análisis sobre bioregionalización, áreas de productividad primaria y aspectos físicos que inciden en la estructuración del Océano Austral (ej., cobertura del hielo marino). Colección de información relativa a distribución y abundancia de zooplancton, peces, comunidades bentónicas y depredadores tope presenta aún vacíos significativos. AGRADECIMIENTOS Este trabajo es fruto de la colaboración de diversos investigadores de países miembros de la CCRVMA. REFERENCIAS BIBIOGRÁFICAS Ball, I.R., Possingham, H.P. and Watts, M. 2009. Marxan and relatives: Software for spatial conservation prioritisation. Chapter 14: Pages 185-195 in Spatial conservation prioritisation: Quantitative methods and computational tools. Eds Moilanen, A., Wilson, K.A. and Possingham, H.P. Oxford University Press, Oxford, UK. Douglass, L.L., Turner, J., Grantham, H.S., Kaiser, S., Constable, A., Nicoll, R., Raymond, B., Post, A., Brandt, A. and Beaver, D. 2011. A hierarchical classification of benthic biodiversity and assessment of protected areas in the Southern Ocean. Submitted to the CCAMLR Marine Protected Area workshop held in Brest, France in 2011. WS-MPA-11/23. Raymond, B. 2011. A circumpolar pelagic regionalisation of the Southern Ocean. WS-MPA-11/6.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
253
SOOS- SOUTHERN OCEAN OBSERVING SYSTEM: MISSION AND OBJECTIVES (SOOS - Sistema de Observaciones del Océano Austral: misión y objetivos)
Costa, D.P.1
1SOOS Science Steering Committee 1Dept of Ecology & Evolutionary Biology, University of California Santa Cruz, USA. Email: [email protected] The Southern Ocean is fundamentally important to the Earth system, influencing climate, biogeochemical and ecological cycles over very large scales. Many of the most difficult and pressing issues faced by society – mitigation and adaptation to climate change and sea-level rise; managing the effects of ocean acidification; conservation of marine resources and biodiversity – cannot be addressed without an improved understanding of Southern Ocean processes, feedbacks and their sensitivity to change. Limited observations suggest the Southern Ocean is changing, yet chronic under-sampling makes the causes and consequences of such changes difficult to assess, and limits the effectiveness of any response. The Southern Ocean Observing System (SOOS) has thus been created, to facilitate integration of resources, to enhance data collection and access, and to guide the sustained development of strategic, multidisciplinary science in the Southern Ocean. SOOS is an initiative of SCAR and SCOR, and is currently endorsed by POGO, CliC and CLIVAR. Opening officially at the end of 2011, SOOS is coordinated from an International Project Office (IPO), which is hosted by the Institute for Marine and Antarctic Studies (IMAS) at the University of Tasmania (Hobart, Australia). Running costs of the IPO are covered by voluntary national/Institutional sponsorship, which is open to all nations. Currently, the IPO is sponsored by the Australian Antarctic Division, Antarctica New Zealand, IMAS, and the New Zealand Antarctic Research Institute. The mission of SOOS is to coordinate and expand the efforts of all nations that gather data from the Southern Ocean. SOOS is developing a coherent and efficient observing system to deliver the observations required to address key scientific and societal challenges. The objectives of SOOS are to:
- Design and implement a comprehensive and multi-disciplinary observing system for the Southern Ocean
- Advocate and guide the development of new observation technologies - Unify current observation efforts and leverage further resources - Effectively integrate and coordinate national and international projects and programmes, across
traditional disciplinary boundaries and between nations - Facilitate and develop a data system that provides seamless access to essential data products for
the Southern Ocean SOOS will focus efforts on collecting data that will address the following 6 most compelling scientific and societal challenges that related to the Southern Ocean:
1) The role of the Southern Ocean in the planet’s heat and freshwater balance 2) The stability of the Southern Ocean overturning circulation 3) The role of the ocean in the stability of the Antarctic Ice Sheet and its future contribution to sea-
level rise 4) The future and consequences of Southern Ocean carbon uptake 5) The future of Antarctic sea ice 6) Impacts of global change on Southern Ocean ecosystems

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
254
SOOS Field Priorities In 2012, SOOS asked the Southern Ocean scientific community to develop a number of workplans along the 6 scientific Themes. These workplans were structured to identify the Essential Ocean Variables (EOVs; e.g., temperature, salinity, CO2 etc) and ecosystem EOVs (eEOVs; e.g., chlorophyll, nutrients, krill) that are required to address each Theme, the status of the technology used to measure these EOVs (and eEOVs), and a priority listing of key gaps in current observations. A number of key gaps were identified. These SOOS Priority Gaps could be addressed with relatively small increments in effort if the international community works together. SOOS Priority Gaps 1) Increased observations offshore from all Antarctic bases (year-round where possible):
- Time series measurements of T, S, O2 and carbon parameters - Sea-ice thickness, extent, concentration and drift - Standardised sampling of marine biota - Seal tagging
2) Increased underway measurements from all Southern Ocean ships (year-round where possible): - Freshwater tracer measurements (del18O, noble gases) - Carbon measurements across the Southern Ocean - Measurements of TSG, pCO2, O2 and if possible nutrients, TCO2, alkalinity, Chlorophyll-a,
Argon/O2, bioptics (using calibrated equipment) - Continuous Plankton Recorder, acoustics, predator tracking/diets - Sea-Ice thickness measurements (from both ship and field data) - Sea-ice extent and concentration measurements
3) Under-ice measurements, particularly from continental shelf regions, under and near ice shelves and polynas (year round) 4) Deep ocean (>2000 m) T, S and O2 measurements (year round) 5) Bathymetry under and near ice shelves 6) Sustained T, S and O2 measurements under and near ice shelves (year round) 7) Sea-ice drift measurements (e.g., from meteorology, buoy arrays etc). For more information go to the SOOS website at: http://www.soos.aq/

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
255
RESISTENCIA VERSUS ELASTICIDAD ECOLÓGICA DE LA COMUNIDAD BENTÓNICA ANTÁRTICA: UNA SIMULACIÓN EXPERIMENTAL EN LA PLATAFORMA
CONTINENTAL DEL MAR DE WEDDELL
(Ecological resistance versus resilience of the Antarctic benthic community: a simulation experiment in the Weddell Sea Continental Shelf)
Montiel, A.1,2, Quiroga, E.3, Sepúlveda, N. 1, Aldea, C.2, Esquete, P.4 y D. Gerdes5
1Instituto de la Patagonia, Universidad de Magallanes, Av. Bulnes #01855, Casilla113-D, 14 Punta Arenas, Chile. Email: [email protected] 2Programa GAIA Antártica, Universidad de Magallanes, Av. Bulnes #01855, Casilla113-D, 14 Punta Arenas, Chile 3Pontificia Universidad Católica de Valparaíso (PUCV), Escuela de Ciencias del Mar, Casilla 1020, Valparaíso, Chile. 4Universidad de Vigo, España. 5Alfred Wegener Institute for Polar and Marine Research, Columbusstrasse, D-27568, Bremerhaven, Alemania. INTRODUCCIÓN La resistencia es un término empleado en ecología de sistemas para indicar cuanto cambiará una variable en la comunidad (e.g. abundancia o diversidad) frente a la variación en el estado de otros parámetros del sistema (e.g. productividad o temperatura). Mientras que la elasticidad o resiliencia se define como la tasa de retorno de una variable luego de la perturbación (Jaksic & Marone 2001; Rodríguez 2001). Desde que en 1972 Dayton & Hessler describieron el rol de las disturbios en las comunidades bentónicas del mar profundo, numerosas investigaciones se han realizado en torno a este tema y las investigaciones sobre fauna bentónica antártica no han sido una excepción. Esto no sólo debido a sus similitudes naturales con el mar profundo (Thiel et al. 1996) sino también a la disturbios producida por icebergs (Smale & Barnes 2008). Sumado a la alta diversidad (Brey et al. 1994) y compleja estructura trófica (Jacobs et al. 2011) hacen a las comunidades bentónicas del Mar de Weddell un ecosistema ideal para investigar cómo éstas responden a distintos grados de perturbaciones. En este contexto, el objetivo de este trabajo fue determinar la respuesta de la comunidad macrobentónica posterior a un evento de perturbación artificial producida mediante red de arrastre y, de este modo, analizar comparativamente el nivel de resistencia y elasticidad que presenta la comunidad bentónica en el Mar de Weddell (Antártica). MATERIAL Y MÉTODOS El experimento de perturbación artificial fue realizado durante la expedición antártica ANTXXI/2 (2003) a bordo del R/V Polarstern (Arntz & Brey 2005). La perturbación experimental fue realizada mediante una red Otter modificada frente al banco de Austasen (Mar de Weddell) entre los 255 y los 310 m de profundidad. La evaluación del impacto sobre las comunidades macrobentónicas posterior al arrastre de la red sobre el fondo marinos se realizó mediante un multibox corer. Para determinar el impacto del experimento se seleccionaron ocho estaciones obtenidas en la misma área; cuatro estaciones obtenidas previo a arrastre “pre-perturbada” (N= 27) y cuatro estaciones “post-perturbada” (N= 22) obtenidas seis días después del arrastre de la red. Los organismos fueron separados del sedimento e identificados al nivel taxonómico más bajo posible. Posteriormente, se estimaron las abundancias (ind. m-2) y biomasas (g.p.h. m-2). Se calculó la diversidad (índice de Shannon-Wiener) y equidad (índice de Pielou) para cada estado de la comunidad. Seguidamente, se determinó el grado de perturbación por medio del índice de Warwick (curvas ABC) y por medio del análisis de similitud (ANOSIM). Además,

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
256
se construyeron espectros normalizados de tamaño - biomasa (NBSS) para evaluar cambios en la estructura de tamaños del macrobentos pre y post perturbación. RESULTADOS Se encontraron un total de 98 taxa, de las cuales se identificaron 88 taxa previo a la perturbación y 72 taxa posterior al arrastre. La abundancia promedio por estación previa a la perturbación fue de 3.460 ind. m-2, mientras que posterior a la perturbación, la abundancia promedio fue de 3.363 ind. m-2. El valor de biomasa promedio previa a la perturbación fue de 216,74 g.p.h. m-2 y el valor post perturbación fue de 123,70 g.p.h. m-2. En el caso del índice de diversidad Shannon-Wiener (H'), éste mostró un leve aumento posterior a la perturbación de 2,6 a 2,8. Mientras que el índice de Pielou (J') no mostró una variación entre el grupo de estaciones pre y post perturbación con 0,861 y 0,893, respectivamente. Al contrastar los valores de abundancia entre el grupo de estaciones de pre y post perturbación, el ANOSIM no mostró diferencias significativas (valores R = 0,011; p = 0,541). En el caso de las curvas ABC, se observó que todas las estaciones mostraron un índice W positivo, indicando que ninguna de ellas presenta un estado perturbado. No se observaron diferencias significativas entre los valores del índice W entre el grupo de estaciones previo y posterior al arrastre de la red (Fig. 1).
Figura 1. Efectos de la red de arrastre en las características de la comunidad macrobentónica en el banco de Austasen (Mar
del Weddell) pre y post perturbación. Figure 1. Effects of Otter trawl on the characteristics of macrobenthic community from Austasen bank (Weddell Sea) pre and
post perturbation.
La estructura de tamaños del macrobentos muestra una pendiente más negativa para las estaciones en post perturbación (Log2Y = Log2-1.09 - 0.52SE=0.13X), mientras que en los sitios previo a la perturbación, la pendiente fue menos negativa (Log2Y = Log2-0.1 - 0.44SE=0.15X) (Fig. 2). Sin embargo, un análisis de comparación de pendientes (test t), indica que no existen diferencias significativas en la estructura de tamaños del macrobentos previo y posterior a la perturbación.
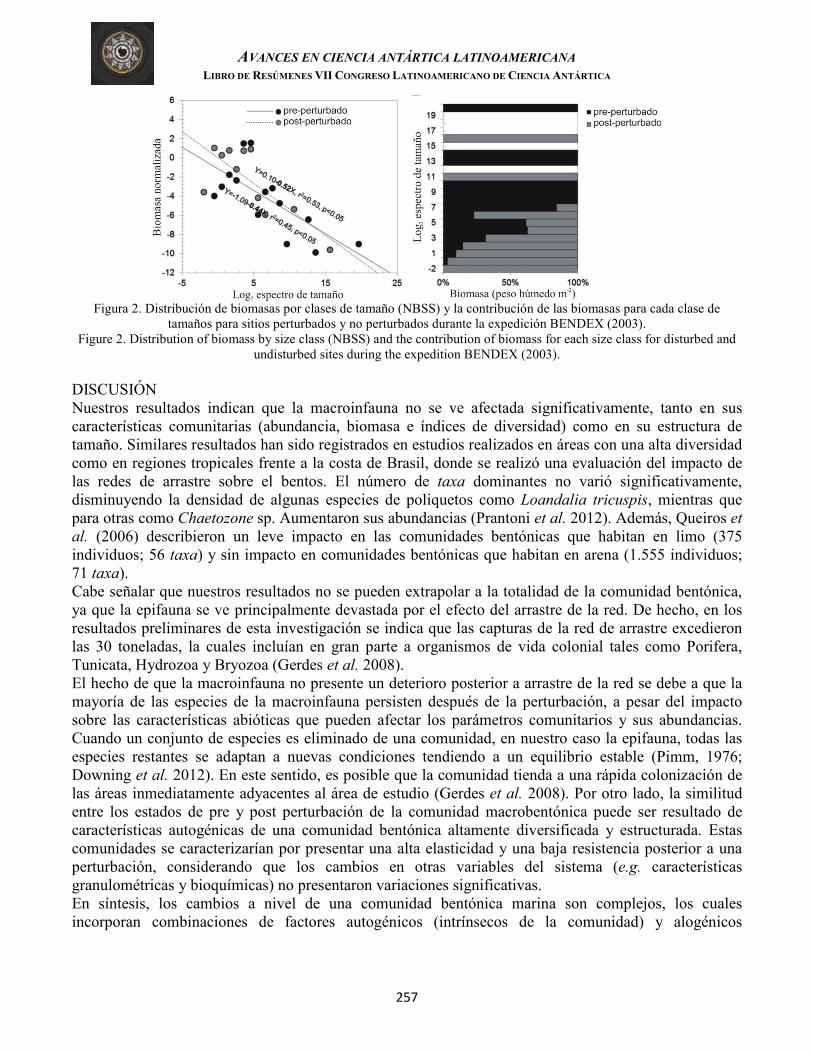
AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
257
Figura 2. Distribución de biomasas por clases de tamaño (NBSS) y la contribución de las biomasas para cada clase de
tamaños para sitios perturbados y no perturbados durante la expedición BENDEX (2003). Figure 2. Distribution of biomass by size class (NBSS) and the contribution of biomass for each size class for disturbed and
undisturbed sites during the expedition BENDEX (2003).
DISCUSIÓN Nuestros resultados indican que la macroinfauna no se ve afectada significativamente, tanto en sus características comunitarias (abundancia, biomasa e índices de diversidad) como en su estructura de tamaño. Similares resultados han sido registrados en estudios realizados en áreas con una alta diversidad como en regiones tropicales frente a la costa de Brasil, donde se realizó una evaluación del impacto de las redes de arrastre sobre el bentos. El número de taxa dominantes no varió significativamente, disminuyendo la densidad de algunas especies de poliquetos como Loandalia tricuspis, mientras que para otras como Chaetozone sp. Aumentaron sus abundancias (Prantoni et al. 2012). Además, Queiros et al. (2006) describieron un leve impacto en las comunidades bentónicas que habitan en limo (375 individuos; 56 taxa) y sin impacto en comunidades bentónicas que habitan en arena (1.555 individuos; 71 taxa). Cabe señalar que nuestros resultados no se pueden extrapolar a la totalidad de la comunidad bentónica, ya que la epifauna se ve principalmente devastada por el efecto del arrastre de la red. De hecho, en los resultados preliminares de esta investigación se indica que las capturas de la red de arrastre excedieron las 30 toneladas, la cuales incluían en gran parte a organismos de vida colonial tales como Porifera, Tunicata, Hydrozoa y Bryozoa (Gerdes et al. 2008). El hecho de que la macroinfauna no presente un deterioro posterior a arrastre de la red se debe a que la mayoría de las especies de la macroinfauna persisten después de la perturbación, a pesar del impacto sobre las características abióticas que pueden afectar los parámetros comunitarios y sus abundancias. Cuando un conjunto de especies es eliminado de una comunidad, en nuestro caso la epifauna, todas las especies restantes se adaptan a nuevas condiciones tendiendo a un equilibrio estable (Pimm, 1976; Downing et al. 2012). En este sentido, es posible que la comunidad tienda a una rápida colonización de las áreas inmediatamente adyacentes al área de estudio (Gerdes et al. 2008). Por otro lado, la similitud entre los estados de pre y post perturbación de la comunidad macrobentónica puede ser resultado de características autogénicas de una comunidad bentónica altamente diversificada y estructurada. Estas comunidades se caracterizarían por presentar una alta elasticidad y una baja resistencia posterior a una perturbación, considerando que los cambios en otras variables del sistema (e.g. características granulométricas y bioquímicas) no presentaron variaciones significativas. En síntesis, los cambios a nivel de una comunidad bentónica marina son complejos, los cuales incorporan combinaciones de factores autogénicos (intrínsecos de la comunidad) y alogénicos

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
258
(externos). Por lo que, es transcendental comprenderlos en forma atingente para predecir las tendencias que tendrán las comunidades bentónicas en el escenario del cambio global actual y futuro. AGRADECIMIENTOS Este investigación fue financiada por Instituto Antárctico Chileno (INACH proyecto N°T25-10, 2010). REFERENCIAS Arntz, W. y Brey T. 2005. The expedition ANTARKTIS XXI/2 (BENDEX) of RV "Polarstern" in 2003/2004. Rep Polar Mar Res, 503:1-149. Brey, T., Klages, M., Dahm, C., Gorny, M., Gutt J., Hain S., Stiller, M. y Arntz, W. 1994. Antarctic benthic diversity. Nature 368. Dayton, P. y Hessler, R. 1972. Rol of biological disturbance in maintaining diversity in the deep sea. Deep sea res. 19: 199-208. Downing, A., van Nes, E., Mooij, W. y Scheffer M. 2012.The Resilience and Resistance of an Ecosystem to a Collapse of Diversity. PLOS ONE 7(9): e46135 Gerdes, D., Isla E., Knust R., Mintenbeck K. y Rossi, S. 2008. Response of Antarctic benthic communities to disturbance: first results from the artificial benthic disturbance experiment on the eastern Weddell Sea Shelf, Antarctica. Polar Biol 31:1469–1480 Jacob, U., Thierry, A., Brose, U., Arntz, W., Berg, S., Brey, T., Fetzer, I., Jonsson, T., Mintenbeck, K., Möllmann, C., Petchey, O., Riede, J., Dunne, J. 2011. The role of body size in complex food webs: a cold case. In Belgrano y Reiss (eds). Adv Ecol Res 45: 181-223. Jaksic F. y Morone L. 2001. Ecología de comunidades. 2ª edición. Ediciones Universidad Católica de Chile. 336 Pp. Pimm, S. L. 1979. Complexity and stability: another look at MacArthur's original hypothesis.Oikos 33: 351-357. Prantoni, A., da Cunha, P., Sandrini, L., Negrello O. y de Oliveira, V. 2012. An experimental evaluation of the short-term effects of trawling on infaunal assemblages of the coast off southern Brazil. JMBA1: 1-8. Rodríguez, J. 2001. Ecología. Ediciones Pirámide Madrid, España. 411 Pp Queirós, A., Hiddink, J., Kaiser M., Hinz H. 2006. Effects of chronic bottom trawling disturbance on benthic biomass, production and size spectra in different habitats. JEMBE 335:91-103 Smale, D. y Barnes, D. 2008. Likely responses of the Antarctic benthos to climate-related changes in physical disturbance during the 21st century, based primarily on evidence from the West Antarctic Peninsula region. Ecography 31: 289-305. Thiel, H., Pörtner, H. y Arntz, W. (1996) Marine life at low temperatures - a comparison of polar and deep-sea characteristics. In: Uiblein F. Ott J, Stachowitsch M. (Eds). Deep Sea and extreme shallow waters habitat: Affinity and Adaptations. Vienna, Austria Academy of Science, Biosystematic and Ecology Serie 11:183-219

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
259
ANÁLISIS DEL COLAPSO Y FALTA DE RECUPERACIÓN EN DOS NOTOTÉNIDOS ANTÁRTICOS USANDO UNA PERSPECTIVA ECOSISTÉMICA
(Analyzing the collapse and lack of recovery in two Antarctic nototheniid stocks using an ecosystem approach)
Arriagada, A.1 y S. Neira1,2
1Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Barrio Universitario s/n, Concepción, Chile. E-mail: [email protected] 2Programa COPAS Sur Austral, Universidad de Concepción, Chile. E-mail: [email protected] INTRODUCCIÓN La explotación pesquera en el continente Antártico se inició el siglo XX a fines de la década de los 60s (Kock 1992). En el sector del Océano Atlántico las pesquerías se desarrollaron alrededor de las islas South Georgia (sub área FAO 48.3) en 1969/70, South Orkney (sub área FAO 48.2) en 1977/78, y South Shetland y la península Antártica (sub área FAO 48.1) en 1978/79. Las principales especies objetivos al inicio de la explotación fueron Notothenia rossii, Champsocephalus gunnari y Gobionotothen gibberifrons. La mayor parte de las capturas se alcanzó en los dos primeros años de la pesquería (fig. 1.a) reduciendo seriamente los stocks (Barrera-Oro et al. 2000, Kock 1992, Kock 1998). En 1985 se clausuró la pesca dirigida a N. rossi y en 1989/90 se declaró una veda indefinida para peces en la región de la Península Antártica. Sin embargo, prospecciones recientes indican que la biomasa de los stocks de N. rossii, C. gunnari y G. gibberifrons habría continuado en descenso en ausencia de pesca (fig. 1.b) sin evidencia de recuperación y con abundancia actual menor que la existente a principios de la pesquería (Barrera-Oro et al. 2003, Jones et al. 2000, Kock 1998, Marschoff et al. 2012).
Figura 1. a) Biomasas estimadas de Notothenia rossii (círculos blancos) y Gobionotothen gibberifrons (círculos negros) alrededor de isla Elefante (área FAO 48.1). b) Capturas de N. rossii (línea continua) y G. gibberifrons (línea punteada)
informadas por CCAMLR en área 48.1 Figure 1. a) Biomass estimates for Notothenia rossii (white circles) and Gobionotothen gibberifrons (black circles) around
Elefant Island (FAO area 48.1). b) Catches for N. rossii (solid line) and G. gibberifrons (dashed line) reported by CCAMLR in area 48.1
En la trama trófica Antártica, en la zona oceánica, el krill es el principal vínculo entre la producción primaria y los depredadores topes del sistema. Sin embargo, en las zonas costeras los peces demersales serían más importantes que el krill al consumir del bentos y también del zooplancton, siendo el vínculo entre los niveles inferiores y superiores de la cadena alimentaria. En efecto, son presas importantes de depredadores como aves y mamíferos asociados a estas zonas (Barrera-Oro 2002, Casaux y Barrera-Oro

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
260
2006, Casaux y Barrera-Oro 2012). Entonces, el objetivo de este estudio es representar la trama trófica que se desarrolla en la península antártica (fig. 2) y evaluar las posibles causas y efectos a nivel de la trama trófica del colapso y la falta de recuperación de los stocks de N. rossii y G. gibberifrons.
Figura 2. Área de estudio, sub área FAO 48.1 Figure 2. Study area, FAO subarea 48.1
MÉTODOS Se construyó un modelo que representa la trama trófica de la sub área FAO 48.1 (fig. 2), que abarca 672000 km2. El modelo representa el período inicial de la pesquería en la zona (i.e., año 1979). El modelo considera 21 grupos funcionales, incluyendo todos los niveles tróficos desde productores primarios, zooplancton, invertebrados, peces, aves y mamíferos marinos, y detritus. Los parámetros de entrada al modelo (biomasa, producción, consumo, dieta y desembarques) fueron calculados a partir de información bibliográfica. Se utilizó el modelo y software Ecopath con Ecosim (EwE), diseñado para la construcción, parametrización y análisis de modelos tróficos con balance de masa (Christensen & Pauly 1992, Walters et al. 1997). Una vez balanceado el modelo se simuló distintos escenarios de pesca para evaluar el colapso y la recuperación de stocks de nototénidos. RESULTADOS El diagrama que representa la trama se presenta en la fig. 3. A partir de las simulaciones se observó que la mortalidad por pesca no causa colapso de la biomasa de los peces nototénidos, a menos que el desembarque haya estado severamente subestimado en los primeros años de la pesquería. Cambios (disminución) en la biomasa de las principales presas (zooplancton) explican la disminución de la biomasa y falta de recuperación de los peces nototénidos en el largo plazo. Sin embargo, la magnitud de la disminución de la biomasa de nototénidos en este escenario parece ser menor que la observada. No se simuló efecto de aumento de depredación como factor explicativo debido a que no existe evidencia que los principales predadores de nototénidos (aves y ballenas) hayan aumentado en las últimas décadas. Por lo tanto, se concluye que el colapso de peces nototénidos en el área de estudio podría explicarse por una sumatoria del efecto de pesca intensa y limitación de presas. Se discuten algunas hipótesis respecto del efecto del ambiente (e.g., indice de oscilación antártico) en los cambios observados.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
261
Figura 3. Diagrama cuantitativo que representa los flujos en la trama trófica en estudio, área FAO 48.1
Figure 3. Flow diagram representing the food web in FAO area 48.1
Figura. 4. Tendencias de la biomasa de distintos grupos funcionales bajo el efecto combinado de disminución de zooplancton y aumento en la mortalidad por pesca en Notothenia rossii y Gobionotothen gibberifrons
Figure 4. Biomass trends for several functional groups under combined effects of zooplankton decline and increasing fishing mortality in Notothenia rossii y Gobionotothen gibberifrons
REFERENCIAS Barrera-Oro, E. 2002. The role of fish in the Antarctic marine food web: differences between inshore and offshore waters in the southern Scotia Arc and west Antarctic Peninsula. Antarctic Science, 14: 293-309. Barrera-Oro, E. Marschoff, E. y Casaux, R. 2000. Trends in relative abundance of fjord Notothenia rossii, Gobionotothen gibberifrons and Notothenia coriiceps at potter cove, South Shetland Islands, after commercial fishing in the area. CCAMLR Science, 7: 43-52. Barrera-Oro, E. Marschoff, E. Casaux, R. y Gonzalez, B. 2003. Monitoring of relative abundance of fjord Notothenia rossii, Gobionotothen gibberifrons and Notothenia coriiceps at Potter Cove, South Shetland Sslands, in years 2000 to 2003. Document WG-FSA-03/89 CCAMLR. Hobart, Australia.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
262
Casaux, R. y Barrera-Oro, E. 2006. Shags in Antarctica: their feeding behaviour and ecological role in the marine food web. Antarctic Science 18(1): 3–14 Casaux, R. y Barrera-Oro, E. 2012. Linking fish and shags population trends. Document WG-EMM-12/36 CCAMLR. Hobart, Australia. Jones, C. Kock, K. y Balguerias, E. 2000. Changes in biomass of eight species of finfish around the South Orkney Islands (subarea 48.2) from three bottom trawl surveys. CCAMLR Science, Vol. 7: 53-74. Kock, K. 1992. Antarctic fish and fisheries. Cambridge, New York: Cambridge University Press. 359 pp. Kock, K. 1998. Changes in the fish biomass around Elephant Island (subarea 48.1) from 1976 to 1996. CCAMLR Science, 5: 165-189. Marschoff, E. Barrera-Oro, E. Alescio, N y Ainley, D. 2012. Slow recovery of previously depleted demersal fish at the South Shetland Islands, 1983–2010. Fisheries Research, 125–126: 206–213. Walters, C.Christensen, V. y Pauly, F. 1997. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments. Reviews in Fish Biology and Fisheries, 7: 139-172.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
263
DETERMINANTES HISTÓRICO-EVOLUTIVOS DEL ENANISMO EN BIVALVOS ANTÁRTICOS MODERNOS
(Historic-evolutionary determinants of the dwarfism in modern Antarctic bivalves)
Rivadeneira, M.M.1, Villafaña, J.A.1, Gordillo, S.2 y S.N. Nielsen3
1Laboratorio de Paleobiología, Centro de Estudios Avanzados en Zonas Áridas (CEAZA), Av. Ossandón 877, CP 1781681, Coquimbo, Chile. Email: [email protected] 2Centro de Investigaciones en Ciencias de la Tierra, Consejo Nacional de Investigaciones Científicas y Técnicas (CICTERRA, CONICET-UNC) & Centro de Investigaciones Paleobiológicas (CIPAL), Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba, Argentina 3Instituto de Ciencias Ambientales y Evolutivas, Facultad de Ciencias, Universidad Austral de Chile, Valdivia, Chile INTRODUCCIÓN En años recientes, el tamaño corporal se ha vuelto una piedra angular para la ecología moderna, debido a su importancia como regulador de procesos desde escala de organismos a ecosistemas (Brown et al. 2004, Marquet et al. 2005). Por esto, el estudio de las variaciones geográficas y evolutivas en el tamaño corporal de las especies puede entregar luces acerca de la forma en que se organizan las poblaciones, comunidades y ecosistemas. La Antártida ofrece una excelente arena para el estudio de patrones y procesos del tamaño corporal de las especies debidas al inusual gigantismo y enanismo reportado en diferentes taxa (<biblio>). En el caso de los moluscos, particularmente gasterópodos y bivalvos, las especies son inusualmente pequeñas comparado a otras regiones (Nicol 1970, Arnaud 1977, Valentine et al. 2006). Por ejemplo, más del 60% de las especies de bivalvos y gasterópodos alcanzan una talla máxima de ≤ 10 mm, dos a tres veces menos que lo observado en la región Ártica (Nicol 1970, Arnaud 1977). No obstante, análisis globales recientes en bivalvos, cuestionan la validez, o al menos la intensidad, del fenómeno, toda vez que no reconocen un gradiente latitudinal de reducción del tamaño hacia la Antártica (Berke et al. 2013). Las causas últimas del fenómeno del enanismo antártico en moluscos permanecen mucho más oscuras. Se ha planteado que una limitación en la precipitación de carbonato de calcio en las conchas, inducida por las bajas temperaturas sería la causante (Vermeij 1978), pero la ausencia de señal de enanismo en el Ártico sugiere que el enanismo no sería consecuencia de una adaptación al frío. Sin embargo, la evidencia paleontológica indica que el tamaño corporal de las especies durante el Eoceno fue mucho más grande que en la actualidad, siendo más similar al resto de al biota global moderna (Krug et al. 2010). Esto sugiere que algún tipo de forzamiento histórico y/o evolutivo sería responsable de la miniaturización de los bivalvos. La existencia de una estructura comunitaria ‘retrograda’ en la fauna somera actual de la Antártica ha sido ampliamente descrita (Dell 1972, Aronson et al. 1997, Aronson et al. 2009). Un análisis reciente (Krug et al. 2010) demostró que el carácter Paleozoico reminiscente de la fauna de bivalvos resultó de la pérdida selectiva de familias derivadas durante el Cenozoico, en contraste con el Ártico donde la extinción no fue selectiva. Debido a que el tamaño corporal exhibe una señal filogenética marcada (Blomberg et al. 2003), y las especies más grandes tienden a habitar zonas costeras (Rex et al. 1999), los avances de los casquetes de hielo sobre la plataforma Antártica gatillados durante el enfriamiento del Neógeno (Poulin et al. 2002) podrían haber inducido la pérdida diferencial de familias de mayor tamaño, y moldeado el enanismo actual en bivalvos. Este trabajo tiene como objetivos: a) re-evaluar la validez del enanismo en bivalvos antárticos a escala global, y b) evaluar la importancia de procesos histórico-evolutivos como moldeadores del enanismo actual.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
264
MATERIALES Y MÉTODOS Se obtuvo información de la talla (largo máximo reportado) para 5117 especies de bivalvos marinos recientes en 17 zonas biogeográficas del mundo, basados en el compendio de Huber (2010). La misma información fue compilada para 107 especies descritas en ensambles fósiles del Paleoceno/Eoceno de Antártica (Stilwell and Zinsmeister 1992, Stilwell 2000). Para las especies modernas, además, se obtuvo información del rango batimétrico de las especies Las asignaciones taxonómicas fueron corregidas de acuerdo al World Register of Marine Species (www.worms.org, consultado en diciembre de 2012). La distribución acumulada de frecuencias de talla actual y fósil de bivalvos antárticos fue comparada con las predicciones de 2 modelos nulos, que consideran un re-muestreo (5000 corridas) desde el pool global de especies presentes en la plataforma continental y en el talud-abismo, empleando la cobertura estadística de los valores de la prueba de Kolmogorov-Smirnov (i.e. proporción de corridas no significativas), donde la similitud de distribuciones de frecuencia puede ser inferida con valores de cobertura superiores a 0.95. Posibles efectos filogenéticos fueron controlados empleando un re-muestreo estratificado por familias, donde solo familias presentes en la Antártica fueron incluidas. Todos los análisis fueron llevados a cabo empleando rutinas ad-hoc en R (Team 2013). RESULTADOS Y DISCUSIÓN La talla de 115 especies antárticas actuales muestra notables diferencias respecto de todas las otras 16 provincias biogeográficas a nivel global (Kolmogorov-Smirnov, p< 0.0001 en todos los casos), con una mediana de solo 8 mm. El enanismo no parece emerger como un artefacto de un muestreo talla-dependiente en Antártica, ya que desde ~1900 esta mediana se ha mantenido notablemente estable e independiente de las nuevas especies reportadas. Comparativamente la fauna ártica exhibe una mediana de ~ 20 mm, notablemente similar a la fauna del Caribe (Fig. 1A). Esta parece ser la primera verificación estadística global del fenómeno de enanismo en bivalvos antárticos, planteado hace más de 40 años (Nicol 1970, Arnaud 1977, Valentine et al. 2006)
Figura 1: Distribución acumulada de frecuencias de tallas de especies de bivalvos antárticos. A) Comparación con biotas de
17 provincias biogeográficas a nivel global. B) Comparación con predicciones de 2 modelos nulos (5000 corridas), usando un re-muestreo desde el pool global de especies de plataforma y talud-abismo. C) Talla en ensambles fósiles del
Paleoceno/Eoceno, comparado con predicciones de 2 modelos nulos. Figure 1: Cumulative frequency distribution of the body size of Antarctic bivalves. A) Comparison with the biotas of 17
biogeographic provinces at a global scale. B) Comparison with the predictions of 2 null models (5000 runs), using a resampling from the global pool of species from inhabiting the shelf or slope-abyss. C) Size in fossil assemblages of the
Paleocene/Eocene, comparing with the predictions of the 2 null models.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
265
Estrategias de adaptación eco-fisiológica al frío, o respuestas a la baja precipitación de carbonato de calcio, o alta concentración de oxígeno suelen ser invocadas para entender los fenómenos de enanismo y gigantismo antártico. Sin embargo la ausencia de una simetría inter-hemisférica en la señal de enanismo en la talla refuerza la idea de que procesos histórico-evolutivos, únicos a la región Antártica - no necesariamente adaptativos -serían responsables de primer orden del fenómeno. El aislamiento geográfico del continente Antártico desde finales del Cretácico tardío (Crame 1999), sumado a la formación y avance de los casquetes de hielo durante el Neógeno sobre la plataforma continental (Poulin et al. 2002) habrían decimado principalmente a la fauna de plataforma, típicamente de mayor tamaño corporal, siendo posible generar dos predicciones: i) la talla de bivalvos antárticos sería mucho más similar a la de zonas de talud-abismo de otras provincias del océano global; ii) la talla de bivalvos previo a la glaciación del continente sería mucho más similar a la fauna actual de zonas de plataforma en otras provincias del océano global. Los resultados apoyan ambas predicciones (Figs. 1B y C). Por un lado, la talla actual de bivalvos no es diferente de lo esperado de un re-muestreo del pool combinado de especies presentes en otras provincias y que habitan en zonas profundas (Fig. 1B, cobertura = 0.98). Por otro lado, las especies en ensambles fósiles del Paleógeno exhiben tallas notablemente mayores (mediana 38 mm), mucho mas similares a la del pool combinado de especies presentes en otras provincias y que habitan en zonas de plataforma (Fig. 1C, cobertura = 0.95). Así, el recambio filogenético inducido por las glaciaciones en la biota de bivalvos antárticos (Krug et al. 2010) habría traído aparejado la desaparición de formas someras y de mayor tamaño (Fig. 2A), y la recolonización por parte de formas profundas típicamente pequeñas (Fig. 2B). Las especies pertenecientes a familias presentes en el Paleógeno y aún representadas en la actualidad también muestran una reducción evolutiva en el tamaño, pero éste nuevamente se asocia a la presencia de formas de zonas profundas (Fig. 2C). Estos resultados validan la hipótesis de que el enanismo de los bivalvos antárticos emergería como un proceso no-adaptativo, subproducto de un proceso batimétrico de selección de especies.
Fig. 2. Recambio evolutivo de bivalvos antárticos desde el Paleoceno/Eoceno al reciente. Distribución acumulada de
frecuencias de talla de especies pertenecientes a familias extintas (A), familias invasoras (B) y familias sobrevivientes (C) en comparación con las predicciones de 2 modelos nulos (5000 corridas), usando un re-muestreo desde el pool global de
especies de plataforma y talud-abismo. En el caso de las familias sobrevivientes se compara además la distribución acumulada de frecuencias de talla de especies fósiles versus recientes.
Fig. 2. Evolutionary turnover of Antarctic bivalves from the Paleocene/Eocene to the recent. Cumulative frequency distribution of the body size of species belonging to extinct families (A), invading families (B) and surviving families (C) in
comparison with the predictions of 2 null models (5000 runs), using a resampling from the global pool of species from inhabiting the shelf or slope-abyss. In the case of surviving families it is also compared the cumulative frequency distribution
of the body size of fossil versus recent species.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
266
AGRADECIMIENTOS Estudio financiado íntegramente por proyecto INACH G_05-11. REFERENCIAS BIBLIOGRÁFICAS Arnaud, P. M. 1977. Adaptations within the Antarctic marine benthic ecosystem. En: Adaptations within the Antarctic ecosystems. G.A. Llano (Ed), The Smithsonian Institution, Washington, D.C., pp. 135-157. Aronson, R. B., D. B. Blake, and T. Oji. 1997. Retrograde community structure in the late Eocene of Antarctica. Geology 25:903-906. Aronson, R. B., R. M. Moody, L. C. Ivany, D. B. Blake, J. E. Werner, and A. Glass. 2009. Climate Change and Trophic Response of the Antarctic Bottom Fauna. PLoS ONE 4:1-6. Berke, S. K., D. Jablonski, A. Z. Krug, K. Roy, and A. Tomasovych. 2013. Beyond Bergmann's rule: size-latitude relationships in marine Bivalvia world-wide. Global Ecology and Biogeography 22:173-183. Blomberg, S. P., T. Garland, and A. R. Ives. 2003. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 57:717-745. Brandt, A., A. J. Gooday, S. N. Brandao, S. Brix, W. Brokeland, T. Cedhagen, M. Choudhury, N. Cornelius, B. Danis, I. De Mesel, R. J. Diaz, D. C. Gillan, B. Ebbe, J. A. Howe, D. Janussen, S. Kaiser, K. Linse, M. Malyutina, J. Pawlowski, M. Raupach, and A. Vanreusel. 2007. First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature 447:307-311. Brown, J. H., J. F. Gillooly, A. P. Allen, V. M. Savage, and G. B. West. 2004. Toward a metabolic theory of ecology. Ecology 85:1771-1789. Chapelle, G. and L. S. Peck. 1999. Polar gigantism dictated by oxygen availability. Nature 399:114-115. Crame, J. A. 1999. An evolutionary perspective on marine faunal connections between southernmost South America and Antarctica. Scientia Marina 63:1-14. Dell, D. K. 1972. Antarctic benthos. Advances in Marine Biology 19:1-216. Huber, M. 2010. Compendium of bivalves. ConchBooks: Hackenhei. 901 pp. Krug, A. Z., D. Jablonski, K. Roy, and A. G. Beu. 2010. Differential Extinction and the Contrasting Structure of Polar Marine Faunas. PLoS ONE 5: e15362. Marquet, P. A., R. A. Quinones, S. Abades, F. Labra, M. Tognelli, M. Arim, and M. Rivadeneira. 2005. Scaling and power-laws in ecological systems. Journal of Experimental Biology 208:1749-1769. Nicol, D. 1970. Antarctic Pelecypod Faunal Peculiarities. Science 168:1248-1249. Poulin, E., A. T. Palma, and J. P. Feral. 2002. Evolutionary versus ecological success in Antarctic benthic invertebrates. Trends in Ecology & Evolution 17:218-222. Rex, M. A., R. J. Etter, A. J. Clain, and M. S. Hill. 1999. Bathymetric patterns of body size in deep-sea gastropods. Evolution 53:1298-1301. Stilwell, J. 2000. Eocene mollusca (Bivalvia, Gastropoda and Scaphopoda) from McMurdo Sound: systematics and paleoecologic significance. Antarctic Research Series 76:261-320. Stilwell, J. D. and W. J. Zinsmeister. 1992. Molluscan systematics and biostratigraphy: Lower Tertiary La Meseta Formation, Seymour Island, Antarctic Peninsula. American Geophysical Union. Team, R. D. C. 2013. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Valentine, J. W., D. Jablonski, S. Kidwell, and K. Roy. 2006. Assessing the fidelity of the fossil record by using marine bivalves. Proceedings of the National Academy of Sciences of the United States of America 103:6599-6604. Vermeij, G. J. 1978. Biogeography and adaptation: patterns of marine life. Harvard University Press. 332 pp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
267
EL “ATAJO ANTÁRTICO” – UNA POSIBLE EXPLICACIÓN PARA EL INCREMENTO DEL INTERCAMBIO FAUNÍSTICO EN EL CENOZÓICO MEDIO ENTRE NUEVA ZELANDA Y
SUDAMÉRICA
(The Antarctic Shortcut – a possible explanation for increased mid-Cenozoic faunal exchange between New Zealand and South America)
Nielsen, S.N.1
1Instituto de Ciencias Ambientales y Evolutivas, Universidad Austral de Chile, Valdivia, Chile. [email protected]. RESUMEN Las estrechas conecciones entre faunas cenozóicas marinas de Nueva Zelanda y el sur de Suramérica (Chile y Argentina), especialmente en moluscos, se conocen ya desde el principio del siglo 20. Entre Nueva Zelanda y Argentina se identificaron 46 géneros de moluscos para el Cenozóico, con un pico de dispersión en el Oligoceno tardío a Mioceno temprano (Beu et al. 1997). Comparando esta base de datos con la de Kiel y Nielsen (2010), se ve que por lo menos 18 de ellos también aparecen en Chile pero ciertos géneros están compartidos con uno de los otros dos paises pero no con ambos. Chile y Argentina comparten por ejemplo Struthiochenopus, Adelomelon y Olivancillaria, mientras que Chile y Nueva Zelanda comparten por ejemplo Lamprodomina, Bedeva y Austrotoma. Los tres comparten géneros como Fusitriton, Austrocominella y Zeacuminia. Aparte de moluscos hay evidencia adicional para conecciones paleobiogeográficas de briozoos cinctopóridos, equinoideos y braquiópodos. Cinctopóridos son una familia de briozoos de que se pensaba que era exclusivo de Nueva Zelanda, sin embargo hace poco fue reportado también del Oligo-Mioceno de la Patagonia (Casadío et al. 2010). Se propone que la Cordillera de los Andes aún baja y el aliniamiento de Nueva Zelanda, Antártica y Suramérica propiciaron varios caminos de migración tal vez hasta tan tarde como el Mioceno tardío. Estas áreas de plataforma o aguas someras proveyeron posibilidades de dispersión 1) entre Nueva Zelanda y Chile por la plataforma antártica, 2) entre Nueva Zelanda y Argentina por el sistema de rift de Antártica Occidental, y 3) entre Chile y Argentina cruzando los Andes por estrechos. El rafting en macroalgas o madera flotante incrementó las posibilidades de dispersión para muchas taxa sin larvas de larga vida. AGRADECIMIENTOS Se agradece financiamiento por proyecto INACH G05-11. REFERENCIAS Beu, A.G., Griffin, M. and Maxwell, P.A. 1997. Opening of Drake Passage gateway and Late Miocene to Pleistocene cooling reflected in Southern Ocean molluscan dispersal: evidence from New Zealand and Argentina. Tectonophysics 281: 83-97. Casadío, S., Nelson, C., Taylor, P., Griffin, M. and Gordon, D. 2010. West Antarctic Rift system: a possible New Zealand-Patagonia Oligocene paleobiogeographic link. Ameghiniana 47: 129-132. Kiel, S. and Nielsen, S.N. 2010. Quaternary origin of the inverse latitudinal diversity gradient among southern Chilean mollusks. Geology 38: 955-958.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
268
PATTERNS OF GENETIC DIVERSITY AND STRUCTURE, AS WELL AS DNA-BASED MODELLING ANALYSES SUPPORTS THE EXISTENCE OF GLACIAL REFUGIA IN
SOUTH GEORGIA ISLAND FOR THE ANTARCTIC LIMPET DURING THE LAST GLACIAL MAXIMA.
(Patrones de diversidad y estructura genética, así como análisis de modelamiento basados en secuencias de DNA, apoyan la existencia de refugio glacial en Isla Georgia del Sur para la lapa Antártica durante el
Último Máximo Glacial)
González-Wevar C.A.1*, Morley S.2, Chown S.L.3, Poulin E.1
1Laboratorio de Ecología Molecular, Instituto de Ecología y Biodiversidad (IEB), Departamento de Ciencias Ecológicas, Universidad de Chile. Las Palmeras # 3425, Ñuñoa, Santiago, Chile. [email protected]. 2British Antarctic Survey, Madingley Road, High Cross Cambridge, Cambridgeshire CB3 0ET, UK. 3School of Biological Sciences, Monash University, Victoria 3800, Australia INTRODUCTION The evolution of benthic invertebrates of the Southern Ocean, and especially in Antarctica, is of considerable interest given the complex biogeographical patterns described for this fauna in space and time. The prevalence of increasingly cold conditions shaped the evolution of the Antarctic marine life and glacial processes radically changed the distribution of intraspecific genetic variation. Several hypotheses have been proposed to explain how the Antarctic benthos survived the glacial events of the Pleistocene. In general, cool-temperate species survived the Last Glacial Maximum (LGM) contracting their distribution to low latitude less ice-impacted refugia, and then re-colonized higher latitudes through range expansion after ice retreat. In Antarctica, three models can be proposed to account for the benthos during the Pleistocene. (1) A “deep-sea refugia” model, where Antarctic shelf species responded to ice advances expanding their bathymetric range toward deep-sea less ice-disturbed areas. However, this model fails to explain how the Antarctic shallow benthos survived the LGM. (2) A “shelf in situ refugia” model suggests that some groups may have survived in situ, in one or several refugia on the Antarctic shelf. (3) Finally, an “island refugia model” where shallow marine species survived the LGM out of the Antarctic continental shelf, at adjacent Antarctic islands or in geographically distant islands of the Scotia Arc. Patterns of population genetic diversity and structure can be used to infer historical and contemporary demographic processes and to identify re-colonization routes. The Antarctic limpet Nacella concinna constitutes a good model to examine the effects of past climate events on Antarctic shallow benthos and to contrast among the competing scenarios. In this study, new molecular mtDNA genetic analyses of N. concinna were performed in populations collected from different areas along the species distribution including the Antarctic Peninsula (West and East), the South Shetlands Islands, South Orkney Island and South Georgia islands to examine these alternative hypotheses. MATERIAL AND METHODS Nacella concinna specimens were collected in the intertidal zone at nine localities distributed in four main areas of interest: the Antarctic Peninsula (AP), the South Shetland Islands (SSI), South Orkney Island (SOI) and South Georgia (SGI). A partial fragment of the mitochondrial gene COI was amplified and sequenced. Sequences were edited and we estimated levels of genetic polymorphism using standard diversity indices (k, S, H, Π, and π) per locality, area, and for the species. We estimated the number of private alleles and we performed neutrality statistical tests (Tajima’s D and Fu’s FS). Genetic

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
269
differentiation was estimated through mean pairwise differences (NST) and through haplotype frequencies (GST). We used two different clustering methods to infer the spatial genetic structure of N. concinna. First, we estimated the number and the composition of panmictic groups, as well as the spatial boundaries among them using a Bayesian model computed with the Geneland program. Second, we estimated the number and composition of groups that were the most differentiated based on sequence data with SAMOVA. We constructed genealogical relationships in the species by median-joining haplotype networks. To assess the patterns of demographic history, we plotted the distribution of pairwise differences between haplotypes (mismatch distributions) for each area. Then, we compared the deviation of mismatch distributions from a model of sudden expansion, using a non-linear least squares method. We estimated past population dynamics through time in N. concinna using a Bayesian skyline plot method using a ten-fold evolutionary rate for nacellids at population level. Three independent runs were made for 250 x 106 generations (sampled every 1,000 iterations), discarding 10% of the trees as burn-in. To assess the influence of glacial cycles on the demographic history and genetic structure of N. concinna, we used the Approximate Bayesian Computation (ABC) method. We compared the posterior probabilities of two competing scenarios of genetic differentiation that are characterized by contrasting population divergence times and demographic histories. Posterior probabilities were estimated with DIY ABC using historical, demographic and mutational parameters drawn from the prior distributions. For each scenario, 1 x 106 data sets were simulated and the relative likelihoods of both scenarios were compared using a logistic regression on 1 % of simulated data closest to the observed data set. RESULTS AND DISCUSSION 269 individuals were analyzed with a fragment of 663 bp of the mtDNA COI gene. We detected low levels of genetic polymorphism in the species. The number of private haplotypes was much higher in SGI than in any of the other analyzed localities. Similarly, the proportion of private haplotypes (8/11) found in SGI was twice that detected in the other areas. Mean general values of differentiation, as measured with GST and NST, were low (GST = 0.039 and NST = 0.045), but highly significant. However, when depicting pairwise comparisons among samples, only those between SGI and the rest of the localities were statistically significant after Bonferroni correction. Two main clusters (K=2) were recognized with Geneland and SAMOVA. The first one includes maritime Antarctic sites from Rothera Station to South Orkney Island and the second one South Georgia Island (Figure 1).
Figure 1. Recognized clusters in Nacella
concinna with Geneland. Black circles indicate the relative position of the sampled
populations. Darker and lighter shading are proportional to posterior probabilities of
membership in clusters, with lighter areas showing the highest probabilities of clusters.
Figura 1. Clusters reconocidos en Nacella concinna con Geneland. Los círculos negros
indican la posición relativa de las poblaciones muestreadas. Áreas sombreadas oscuras y
claras son proporcionales a las probabilidades posteriores de membresía a cada cluster, con las áreas claras mostrando las mayores probabilidades de cluster.
Median-joining haplotype network in the species comprised 28 different haplotypes with a typical star-like topology and a short genealogy (Figure 2). The central haplotype (H1) was the most frequent one (52,4%) and distributed at all localities. In contrast to most localities and areas that are characterized by

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
270
a single dominant haplotype, in South Georgia we found four haplotypes with intermediate frequency values (H1, H21, H22, and H23) and three of them (H21, H22, and H23) were endemic to this peri-Antarctic island (Figure 2).
Figure 2. General median joining haplotype network including 269 Nacella concinna mtDNA COI sequences. Each
haplotype is represented by a circle indicating the main area where it was collected; the size of the circle is proportional to its frequency in the whole sampling effort.
Figura 2. Red de haplotipos de median-joining general que incluye 269 secuencias mtDNA COI de Nacella concinna. Cada haplotipo es representado por un círculo que indica el area donde fue colectado; el tamaño de cada círculo es proporcional a
su frecuencia en todo el esfuerzo de muestreo. Haplotype networks from maritime Antarctic areas showed similarities according to their star-like topologies, short genealogies, and haplotype structure (Figure 3). In contrast, the network of SGI showed a more expanded genealogy and the presence of several private haplotypes of intermediate and low frequency values. SGI also differed from the three other investigated areas in the distribution of pairwise differences between haplotypes. As expected for star-like networks, mismatch distributions are positively skewed and unimodal for AP, SSI, SOI, while the distribution of pairwise differences for SGI was almost unskewed (Figure 3). Figure 3. Median joining haplotype networks and mismatch distribution analyses for each area included in the analyses. Figura 3. Redes de haplotipos de median-joining y análisis
de distribución mismatch para cada área incluida en los análisis.
Bayesian Skyline plot analysis recognized South Georgia Island as older populations than maritime Antarctica ones. The onset of the population expansion in SGI is much more older datedthan for maritime Antarctica. The ABC approach discriminated between the two competing scenarios and the

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
271
posterior probability did not support the ‘in situ shelf refugia’ scenario of N. concinna during the LGM. On the contrary, the ‘island refugia’ scenario received the highest probability value (P = 0.993), which supports the persistence of the species in peri-Antarctic areas and the subsequent colonization of maritime Antarctic through a strong founder effect followed by a rapid expansion. Climatic processes of the Quaternary, together with life-history traits are the major underpinning factors explaining the phylogeographic patterns observed in N. concinna. Supported by the dispersal potential of the species and the observed patterns of genetic diversity and structure in the Antarctic limpet we propose a scenario of rapid post-glacial re-colonization of maritime Antarctic near-shore areas from less ice-impacted peri-Antarctic areas (including SGI), associated with a strong founder effect. This scenario could also prevail for other marine groups so that the following model of Quaternary biogeography could be proposed for shallow-water invertebrates with narrow depth ranges and dispersal capabilities. The model includes: (1) the eradication of near-shore Antarctic populations during glacial maxima of the Pleistocene, (2) the persistence of populations in peri-Antarctic refugia during that time, and (3) the re-colonization of Antarctic near-shore areas after ice retreat, associated with rapid population expansion through larval dispersal. At the same time, this scenario permits us to further understand why one of the most common and dominant Antarctic marine invertebrate is currently restricted to recently deglaciated areas of maritime Antarctica and do not exhibit a circumpolar distribution like other Antarctic taxa. ACKNOWLEDGEMENTS INACH Office project G_04_11, Ph.D thesis projects INACH B_01-07 & Conicyt 24090009 (C.G-W), projects INACH 02-02 & 13-05 and Ecos C06B02 (E.P). Institute of Ecology and Biodiversity (IEB, Universidad de Chile) ICM-P05-002 & PFB-023- CONICYT. Census of Antarctic Marine Life (CAML), Ant-ECO Initiative, EBA-SCAR and PROSUL-Brazil for encouraging and supporting Antarctic research in Evolution.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
272
ESTRUCTURA GENÉTICA DEL ERIZO INCUBANTE ABATUS AGASSIZII EN LA BAHÍA FILDES, ISLA REY JORGE
(Fine-scale genetic structure in the Antarctic brooding sea urchin Abatus agassizii in Fildes Bay, King George Islands)
Poulin E.1, Díaz A.1,2, Maturana C.S.1,3 , González-Wevar C.A.1, Gérard K.1.
1Laboratorio de Ecología Molecular, Instituto de Ecología y Biodiversidad (IEB), Departamento de Ciencias Ecológicas, Universidad de Chile. Las Palmeras # 3425, Ñuñoa, Santiago, Chile. [email protected]. 2Departamento de Ciencias y Recursos Naturales Facultad de Ciencias Universidad de Magallanes,
Punta Arenas, Chile. 3British Antarctic Survey, Madingley Road, High Cross Cambridge, Cambridgeshire CB3 0ET, UK. INTRODUCIÓN Los invertebrados marinos bentónicos de la Antártica se caracterizan por una alta proporción de especies incubantes. La incubación corresponde a la retención de huevos por la hembra, limitando la dispersión de las crías. El género de erizo Abatus es endémico de las regiones antárticas y sub-antárticas e incluye 11 especies, todas incubantes. En estas especies, la baja dispersión de gametos masculinos, así como la limitada movilidad de los adultos, deberían limitar drásticamente la conexión entre parches aislados de individuos y por ende el flujo de genes entre y dentro de las poblaciones. En este estudio, caracterizamos los límites de la población de A. agassizii en la Bahía Fildes (Isla Rey Jorge) con el fin de analizar la diversidad genética intra-poblacional y determinar la escala geográfica a la cual ocurre la diferenciación genética usando marcadores microsatélites. MATERIAL Y METODOS Especímenes de Abatus agassizii fueron colectados en la zona submareal somera de 10 sitios en la Bahía Fíldes. Ninguna otra población de esta especie fue encontrada afuera de esta Bahía a pesar de un importante esfuerzo de muestreo en otras bahías de la isla Rey Jorge, en otras islas de las Shetlands del Sur y a lo largo de la Península Antártica (Figura 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
273
Figura 1. Mapa de distribución de A. agassizii en la Península Antártica y las Shetlands del Sur. A : Bahía Almirantazgo (Isla
Rey Jorge); B: sitios de muestreo de Abatus agassizii en la Bahía Fíldes (Isla Rey Jorge). Figure 1. Distribution of sites of A. agassizii along Antarctic Peninsula. A: Admiralty Bay (King George Island). B: Fildes
Bay (King George Island). Esos sitios se caracterizan por la presencia de parches de individuos agregados en alta densidad y a veces a muy baja profundidad. Todos los ejemplares (n=365) fueron fijados en etanol (95%) y a partir de los cuales se preparó DNA total mediante el método salino. Cada individuo fue genotipificado con 6 marcadores microsatélites polimórficos y se calcularon los índices de diversidad en las 10 poblaciones fue caracterizada por los índices de diversidad clásicos que son la heterozigosidad esperada y el número promedio de alelos por locus. La estructura genética se evaluó a partir de los índices Fst y Rst con el programa Genetix, así como en base a análisis de asignamientos individuales con los programas Structure y Genclass2. RESULTADOS Y DISCUSION Los diferentes sitios mostraron valores similares de heterocigosidad (entre 0,58 y 0,64) así como del número promedio de alelos (entre 10,5 y 13,7). Los análisis de estructuración evidenciaron la existencia de 4 grupos genéricamente diferenciados.
Figura 2. Identificación de las 4 unidades genéticas en la Bahía Fíldes Figure 2. Identification of four genetic units in Fildes Bay Estas unidades genéticas agrupan sitios de muestreo geográficamente cercanos y muestran por lo tanto una coherencia espacial entre sitios continuos, separados generalmente por áreas que no albergan erizos. Aunque claramente significativos, los índices de diferenciación fueron bajos (Tabla 1), indicando solo una leve estructura genética entre estas zonas de la Bahía Fíldes.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
274
Tabla 1: Valores de Fst pareados entre las poblaciones (Weir & Cockerham 1984). Significancia a nivel de 5% (*), 1% (**).
Table 1: paired Fst values between populations (Weir and Cockerham 1984). Significance level of 5% (*), 1% (**). Por otra parte, el análisis de Mantel mostró que la diferenciación genética aumenta con la distancia geográfica, apoyando un modelo de diferenciación de tipo aislamiento por distancia en esta especie.
Figure 3. Relación entre índice de diferenciación y distancia geográfica.
Figure 3. Relationship between differentiation index (FST) and geographic distances.
A pesar de sus limitadas capacidades de dispersión, Abatus agassizii en la Bahía Fíldes demuestra un nivel muy bajo de estructura genética y una homogeneidad genética dentro de parches de más de 1 kilómetro de largo. Una posible explicación a una cierta homogeneidad genética dentro de la población como entre poblaciones, podría ser ligada al comportamiento de los juveniles tardíos luego de la liberación de los bolsillos maternos. Estos juveniles tienen un tamaño aproximado entre 4-5mm, por lo podrían ser desplazado por corrientes de marea u oleajes, permitiendo un flujo genético. AGRADECIMIENTOS Corporación IEB (P05-002ICM, PFB-23), FONDECYT 3100139, INACH F_01-09

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
275
CARACTERIZACIÓN Y COMPARACIÓN DE COMUNIDADES DESNITRICANTES EN TAPETES MICROBIANOS DE LA ISLA DEL REY JORGE (ANTÁRTIDA MARÍTIMA).
(Characterization and comparison of denitrifying bacterial communities in microbial mats from King George Island, maritime Antarctica)
Alcántara-Hernández, RJ1, Centeno, CM1, Ponce-Mendoza, A2, Batista S3, Merino-Ibarra, M4, Campo,
JH5 y Falcón, LI1.
1Laboratorio de Ecología Bacteriana y Epigenética, Instituto de Ecología, Universidad Nacional Autónoma de México, D.F., México, E-mail: [email protected], [email protected], [email protected] 2Centro Nacional de Investigación Disciplinaria en Conservación y Mejoramiento de Ecosistemas Forestales, INIFAP, México. E-mail: [email protected] 3Unidad de Microbiología Molecular, Instituto de Investigaciones Biológicas Clemente Estable (MEC), Montevideo, Uruguay, E-mail: [email protected] 4Unidad Académica de Ecología Marina, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, D.F., México, E-mail: [email protected] 5Laboratorio de Biogeoquímica Terrestre y Clima, Instituto de Ecología, Universidad Nacional Autónoma de México, D.F., México, E-mail: [email protected] Los tapetes microbianos representan una de las principales comunidades bénticas de los lagos y de las zonas encharcables de la Antártida, y que por su actividad, afectan la dinámica de los nutrientes de aquellos sitios donde se establecen (McKnight et al. 2004; Camacho y Fernández-Valiente 2005). Estas comunidades no sólo reflejan una gran diversidad taxonómica, sino también una gran diversidad metabólica, interviniendo en varios ciclos biogeoquímicos (Stal 2012). La desnitrificación es un proceso en el cual óxidos de nitrógeno sirven como aceptores electrónicos para la generación de energía (Zumft 1997). Este proceso es de importancia porque puede ser considerado como una forma de perder nitrógeno (N) de un sistema, ya que formas solubles y asimilables de nitrógeno, como nitratos (NO3
-) y nitritos (NO2-), son reducidos a formas gaseosas como el óxido nítrico
(NO), el óxido nitroso (N2O) y nitrógeno molecular (N2), las cuales regresan a la atmósfera. Las comunidades desnitrificantes pueden ser estudiadas mediante técnicas independientes de cultivo empleando oligonucleótidos para amplificar secciones parciales de genes que codifican para péptidos de nitrito reductasas, como nirK y nirS, y para un péptido de la óxido nitroso reductasa, nosZ (Throbäck et al. 2004). Estas enzimas son clave en el proceso de desnitrificación y han sido ampliamente usadas como marcadores de desnitrificación en diversos ambientes. Se han realizado varios trabajos caracterizando a los microorganismos fijadores de nitrógeno en tapices antárticos. Sin embargo, existen pocos estudios enfocados al proceso de desnitrificación y aún menos a la caracterización de las comunidades potenciales que llevan a cabo el proceso (Joye y Paerl 1994; Taton et al. 2003; Gooseff et al. 2004). Los objetivos de este trabajo fueron i) caracterizar la comunidad potencial desnitrificante en tapetes distribuidos en la Península de Fildes en la Isla de Rey Jorge (Antártida Marítima) por técnicas independientes de cultivo, empleando los marcadores moleculares nirK, nirS y nosZ y ii) comparar el perfil de la comunidad en dos años consecutivos de muestreo. Cinco tapetes microbianos fueron muestreados en diferentes puntos de la Península de Fildes, en la Isla de Rey Jorge (Antártida marítima). El muestreo se llevó a cabo durante los veranos del 2011 y del 2012. Los tapetes muestreados contaban con diferentes características y parámetros como temperatura y pH fueron determinados in situ (Tabla 1). El DNA metagenómico fue extraído de la biomasa de los tapetes empleando el método reportado en Centeno et al, (2012).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
276
Tabla 1 Localización de los sitios de muestreo
Table 1 Location of sampling sites
Id Referencia Localización Temp (ºC) pH Características
S1 Base Artigas (Uruguay)
S 62º 11' 03.0 W 58º 54' 23.4
4.7 a 8.5 b 8.0 a 7.0 b
Tapete verde filamentoso
S2 Base Gran Muralla (China)
S 62º 13' 29.7 W 58º 57' 09.2
2.5 7.0 7.9 8.1 Tapete color rojizo
S3 Caleta Norma S 62º 11' 16.2 W 58º 55' 03.0
5.8 6.6 8.0 8.0 Tapete color naranja y negro
S4 Pasaje de Drake S 62º 09' 37.8 W 58º 55' 32.8
3.0 7.8 8.3 8.3 Tapete color naranja y rojo
S5 Pasaje de Drake S 62º 09' 44.8 W 58º 55' 28.6
2.2 7.6 8.4 8.0 Tapete color naranja
a) Parámetros determinados en el agua circundante en Febrero 2011, b) en Enero 2012.
Los fragmentos nirK, nirS y nosZ fueron amplificados por PCR y los amplicones fueron separados por su composición mediante electroforesis en geles de poliacrilamida con gradiente desnaturalizante (DGGE). Las bandas representativas en el patrón de la comunidad fueron escindidas y eluidas para después ser reamplificadas, secuenciadas y analizadas. Los geles de DGGE no sólo reflejaron los patrones de la comunidad, sino que al realizarse con concentraciones conocidas de DNA, reflejaron la abundancia de cada filotipo en la comunidad, por lo que matrices de abundancia fueron realizadas para las comunidades desnitrificantes por cada gen. Estas matrices fueron concatenadas en una sola matriz que refleja el total de la comunidad desnitrificante, y se calcularon las distancias para establecer relaciones entre las diferentes comunidades. Los resultados mostraron que a) todos los tapetes contaban con una población potencial desnitrificante dominante que oscilaba entre 5 a 18 filotipos, semejantes a lo que se encuentran mediante esta técnica en suelos y lodos activados (Throbäck et al. 2004); b) los marcadores nirK y nosZ cuentan con más resolución para estas comunidades y por ende, es donde se encontraron el mayor número de filotipos desnitrificantes; c) las secuencias encontradas pertenecieron principalmente a Protebacteria y tuvieron entre el 80 y 98 % de identidad con secuencias ambientales reportadas en las bases de datos (Fig. 1); d) los filotipos encontrados fueron asociados principalmente a Alfaproteobacteria y d) la mayoría de las comunidades desnitrificantes en los tapetes microbianos son más parecidas entre sí independientemente del año de muestreo. Los tapetes microbianos antárticos parecen mostrar poca o nula actividad desnitrificante in situ, principalmente por una limitación en NO3
- (Joye and Pearl 1994; Gooseff et al. 2004). Sin embargo, cuentan con una gran comunidad potencial desnitrificante compuesta de filotipos principalmente pertenecientes a Proteobacteria altamente influenciados por el hábitat. Los patrones de la comunidad mostraron mayor similitud entre los mismos sitios de muestreo que entre el año de muestreo. Sin embargo, más estudios deben realizarse para conocer los factores fisicoquímicos que moldean las comunidades funcionales en los tapetes microbianos antárticos. El N2O es un potente gas con efecto invernadero y promueve la destrucción de la capa de ozono estratosférico (Codispoti 2010), por lo cual conocer el potencial desnitrificante de sistemas con relevancia a escala planetaria, como los sistemas Antárticos resulta de gran relevancia.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
277
0.1
nosZ_phylo32, KC951349
Cupriavidus metallidurans CH34, NC_007974
Soil clone DUNnos001, AY912909
nosZ_phylo29, KC951341
nosZ_phylo05, KC951331
Earthworm alimentary canal clone GutLTDNA126, FN859762
nosZ_phylo39, KC951320
Thioalkalivibrio nitratireducens DSM 14787, CP003989
nosZ_phylo28, KC951352
nosZ_phylo49, KC951321
nosZ_phylo60, KC951346
Soil clone 19z35, DQ010842
nosZ_phylo31.1, KC951325
nosZ_phylo12, KC951343
nosZ_phylo47, KC951353
nosZ_phylo14, KC951323
Rice paddy soil clone SNZ1, AB608704
Bradyrhizobiaceae bacterium D195a, AB480505
Soil clone J33 155, JF310476
nosZ_phylo43, KC951328
nosZ_phylo53, KC951334
Sinorhizobium fredii USDA 257, CP003563
nosZ_phylo74, KC951347
Soil clone B42 78, JF310471
Pseudomonas lini PD 15, DQ377787
nosZ_phylo62, KC951329
Pseudomonas sp. MT-1, AB054991
nosZ_phylo36, KC951326nosZ_phylo38, KC951327
Soil clone J49 85, JF310475
Herbaspirillum sp. TSO45-3, AB608728
Mesorhizobium sp. TSA38, AB542283
Azospirillum lipoferum 4B plasmid, FQ311870
nosZ_phylo61, KC951355
Bradyrhizobium japonicum, AJ002531
Pseudomonas brassicacearum, CP002585
Pseudomonas grimontii PD 10, DQ377782
Uncultured soil clone J25 43, JF310470
Soil clone A22 85, JF310667Soil clone B19 205, JF310668
nosZ_phylo23, KC951340
Rice paddy soil clone SNZ14, AB608706
nosZ_phylo45, KC951319
nosZ_phylo55, KC951350
nosZ_phylo51, KC951322
Rhodopseudomonas palustris HaA2, CP000250
nosZ_phylo31, KC951342
Azoarcus sp. KH32C, AP012304
Soil clone B42 81, JF310657
nosZ_phylo01, KC951354
nosZ_phylo52, KC951344
nosZ_phylo69, KC951356
Soil clone US cDNA 023, FN859938
nosZ_phylo30, KC951336
Achromobacter sp. PD 25, DQ377797
Pseudomonas stutzeri CCUG 29243, CP003677
Marinobacter hydrocarbonoclasticus ATCC 49840, FO203363
nosZ_phylo25, KC951333
nosZ_phylo19, KC951318
nosZ_phylo22, KC951335
nosZ_phylo10, KC951332
nosZ_phylo59, KC951345
nosZ_phylo04, KC951330
Soil clone US cDNA 059, FN859918
nosZ_phylo58, KC951338
Soil clone Soil LT DNA 028, FN859951
nosZ_phylo30.1, KC951324
Rhodoferax ferrireducens T118, CP000267
nosZ_phylo15, KC951339
Rice paddy soil clone SNZ10, AB608705
nosZ_phylo76, KC951348
nosZ_phylo63, KC951351
Soil clone J25 110, JF310494
nosZ_phylo42, KC951337
Pseudomonas sp. N48, AB480492
Gamma!
proteobacte
ria!
Betaproteobacte
ria!
Alphapro
teobacte
ria!
14! 12!
1!
4!5!
10!11!
25!19!
15!
32!30!
31!29!23! 28!
27!22!
45!42!
39!
49!51!
53!55!
59!58!60!
68!
61!
65!63!
76!74! 73!75!
A! B!
Figura 1. Comunidades dominantes desnitrificantes nosZ en tapetes microbianos de la Península Fildes. A, patrón de bandeo por DGGE en cinco sitios de muestreo (S1, S2, S3, S4 y S5) en dos años de muestreo: 2011 (-11) y 2012 (-12), los filotipos
secuenciados se indican con un círculo negro. B, árbol filogenético de máxima verosimilitud de secuencias nosZ de los filotipos secuenciados (550 nucleótidos). Los círculos blancos denotan las ramas soportadas en más de un 50% por un solo
método de inferencia (máxima verosimilitud, parsimonia o neighbour-joining, 1000 remuestreos), en gris por dos métodos y en negro por tres. La barra inferior representa 10% de divergencia.
Figure 1. Dominating nosZ communities in microbial mats from Fildes Peninsula. A, DGGE fingerprints from five sites (S1, S2, S3, S5 and S5) in the Fildes Peninsula, in two consecutive years: 2011 (-11)
and 2012 (-12). Sequenced phylotypes are indicated with a mark. B, phylogenetic relationships of the sequenced phylotypes based on the analysis of nosZ sequences (550 nucleotides). Clusters denoted by black, grey and white circles indicate
bootstrap values > 50% supported by three, two or one different approach, respectively (parsimony, neighbour-joining and/or maximum likelihood; 1000 bootstraping). The scale bars represent 10% sequence divergence.
AGRADECEMIENTOS Instituto Antártico Uruguayo y al personal de la Base Científica “Artigas” por el apoyo técnico y logístico para las expediciones de colecta. A la M. en C. Osiris Gaona y al M. en C. Antonio Cruz-Peralta por su gran apoyo técnico en los procesos experimentales y analíticos. Al apoyo económico de UNAM-PAPIIT (IN225709, IT100212) y SEP-CONACyT (056045, 0171596)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
278
REFERENCIAS Camacho González, A. y Fernández-Valiente, E. 2005. Un mundo dominado por los microorganismos: ecología microbiana de los lagos antárticos. Ecosistemas14 (2): 66-78 Centeno, CM. Legendre, P. Beltrán, Y. Alcántara-Hernández, RJ. Lidström, UE. Ashby, MN. Y Falcón, LI. 2012. Microbialite genetic diversity and composition relate to environmental variables. FEMS Microbiol Ecol 82:724-735 Codispoti, LA. 2010. Interesting Times for Marine N2O. Science 327:1339-1340 Gooseff, MN. McKnight, DM. Runkel, RL. y Duff, JH. 2004. Denitrification and hydrologic transient storage in a glacial meltwater stream, McMurdo Dry Valleys, Antarctica. Limnol Oceanogr 49:1884-1895 Joye, S. y Paerl, H. 1994. Nitrogen cycling in microbial mats: rates and patterns of denitrification and nitrogen fixation. Mar Biol 119:285-295 McKnight, DM. Runkel, RL. Tate, CM. Duff, JH. y Moorhead, DL. 2004. Inorganic N and P dynamics of Antarctic glacial meltwater streams as controlled by hyporheic exchange and benthic autotrophic communities. J N Am Benthol Soc 23:171-188 Stal, LJ. 2012. Cyanobacterial Mats and Stromatolites. En: Whitton BA (ed) Ecology of Cyanobacteria II: Their Diversity in Space and Time, vol XV. Springer Netherlands, Dordrecht, pp 65-125. Taton, A. Grubisic, S. Brambilla, E. De Witt, R. y Wilmotte, A. 2003. Cyanobacterial diversity in natural and artificial microbial mats of Lake Fryxell (McMurdo Dry Valleys, Antarctica): a morphological and molecular approach. Appl Environ Microbiol 69: 5157-5169. Throbäck, IN. Enwall, K. Jarvis, Å. y Hallin, S. 2004. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiol Ecol 49:401-417 Zumft, WG. 1997. Cell biology and molecular basis of denitrification. Microbiol Mol Biol R 61:533-616

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
279
DETECCIÓN MOLECULAR DE CRYPTOSPORIDIUM PARVUM EN PYGOSCELIS PAPUA EN BASE PRESIDENTE GABRIEL GONZÁLEZ VIDELA, ANTÁRTICA (64°49’S; 62°51’O)
(Molecular Detection of Cryptosporidium parvum from Pygoscelis papua from Base Presidente Gabriel González Videla, Antártica (64°49’ S; 62°51’W))
Arredondo, C.1,2, Pino, N.1, Dougnac, C.1,3, Díaz-Lee, A1,3, González-Acuña, D.4, Fredes, F.1
1 Departamento de Medicina Preventiva, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile. Av. Santa Rosa 11735, La Pintana, Santiago, Chile. E-mail: [email protected]; [email protected] 2 Programa de Magister en Ciencias Animales y Veterinarias, Facultad de Ciencias Veterinarias y Pecuarias, Universidad de Chile 3 Doctorado en Ciencias Silvoagropecuarias y Veterinarias, Campus Sur, Universidad de Chile 4 Departamento de Ciencias Pecuarias, Facultad de Ciencias Veterinarias, Universidad de Concepción
INTRODUCCIÓN
Actualmente más del 50% las enfermedades infecciosas emergentes (EIE) en seres humanos son de origen zoonótico (Greger, 2007; Jones et al., 2008) y de estas, cerca del 70% tiene su origen en fauna silvestre (Jones et al., 2008), a su vez, la introducción humana de patógenos es la causa más frecuentemente citada de EIE en fauna silvestre (Greger et al., 2007; Gilbert et al., 2008). Esta situación se debe al crecimiento de la población humana y a los procesos de urbanización que, junto a otros factores como el creciente interés en actividades de ecoturismo, aumenta las oportunidades de contacto entre personas, animales domésticos y fauna silvestre (Epstein et al., 2004). Los pingüinos son aves acuáticas que nidifican en el suelo, forman grandes colonias reproductivas y alimentan a sus crías por regurgitación, factores que facilitan la transmisión horizontal de agentes biológicos (Potti et al., 2002). Pygoscelis papua es una especie residente circumpolar subantártica y antártica (Couve y Vidal, 2005), considerada “Casi Amenazada” por la Unión Internacional para la Conservación de la Naturaleza (BirdLife International, 2012). Cryptosporidium spp. es un protozoo zoonótico emergente, especie específico no estricto, capaz de infectar una amplia gama de hospederos (Ng et al., 2006). Es reconocido como un agente prioritario que debe ser monitoreado en fauna silvestre (Tavernier et al., 2011; Humblet et al., 2012). Existen 20 especies de Cryptosporidium reconocidas como válidas, pero se describe la existencia de al menos otros 44 genotipos no pertenecientes a estas (Smith y Nichols, 2010). Dentro de las especies válidas, C. parvum es la de mayor rango de hospederos (Appelbee et al., 2005; Smith y Nichols, 2010). En aves como hospedero principal, la cryptosporidiosis es producida por C. baileyi, C. meleagridis y C. galli, una de las infecciones más prevalentes en individuos domésticos, enjaulados y silvestres (Ng et al., 2006). C. meleagridis es la tercera especie más frecuentemente aislada desde seres humanos inmunocompetentes, con registros de C. baileyi inmunocomprometidos (Smith y Nichols, 2010). Por su parte, C. hominis que supuestamente se encuentra restringido a seres humanos, ha sido encontrado en sirénidos, ovinos y aves silvestres (Ng et al., 2006; Xiao, 2010). En pingüinos se ha detectado la presencia de Cryptosporidium spp. mediante microscopía en P. papua y P. adeliae en Isla Ardley (62º13’S, 58º54’O) (Fredes et al., 2007a; Fredes et al., 2008), mientras que en muestreos realizados el año 2005 en P. papua procedentes de colonias cercanas a la Base Presidente Gabriel González Videla (PGGV) (64°49’S; 62°51’O), arrojaron resultados negativos (Fredes et al., 2007b). Por otro lado, se ha intentado la detección molecular en diferentes zonas antárticas en las tres especies de pygoscélidos con resultados negativos (Palacios et al., 2010).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
280
En este estudio se realizó detección microscópica y confirmación molecular de Cryptosporidium spp. en P. papua de una colonia cercana a la base PGGV en la Antártica.
MATERIALES Y MÉTODOS Se recolectaron 100 muestras de heces de P. papua desde el ambiente de una colonia cercana a la Base PGGV durante la temporada estival del año 2011, conservadas en etanol 70%. Posteriormente tamizadas, homogeneizadas y centrifugadas en el laboratorio, realizando extendidos a partir del sedimento, los cuales fueron teñidos mediante Ziehl-Neelsen modificado (Henricksen y Pohlenz, 1981) y observados bajo microscopio óptico. Se eligieron 12 muestras positivas por microscopía para confirmación mediante PCR. La extracción de DNA siguió un protocolo de disrupción mecánica con perlas de Zirconia-Silica (0.1 y 1.0 mm) y un equipo Mini-Beadbeater-1TM, en 2 ciclos de 4.600 rpm por 80 segundos. Se realizó la purificación con High Pure PCR Template Preparation KitTM (Roche). La reacción de PCR consiste en un ensayo anidado, con una amplificación primaria de un segmento de ~820pb del gen 18S rDNA utilizando los partidores SSU-F3 (5’-GAA GGG TTG TAT TTA TTA GAT AAA G-3’) y SSU-R3 (5’-AAG GAG TAA GGA ACA ACC TCC A-3’). La segunda reacción amplificó una región de ~520 pb con los partidores internos CR18S3543F (5’-GTT AAG TAT AAA CCC CTT TAC AAG TAT C-3’) y CR18S31078R (5’-CCT CCA ATC TCT AGT TGG C-3’). Cada reacción de PCR se desarrolló de acuerdo a lo previamente descrito (Fayer y Xiao, 2008; Muñoz, et al., 2011). Los productos amplificados fueron enviados a secuenciación. RESULTADOS
Detección microscópica De las muestras analizadas por microscopía, se encontraron un total de 59 muestras positivas a
Cryptosporidium spp. (59%) (Figura 1). Figura 1: Muestras positivas a ooquistes de
Cryptosporidium spp. mediante Ziehl-Neelsen Modificado(A: 3,5um; B: 4.9um).
Figure 1: Positive samples to Cryptosporidium spp. oocyst by modified Ziehl-Neelsen (A: 3,5um; B:
4.9um).
Confirmación molecular De las 12 muestras seleccionadas para confirmación molecular, solamente una resultó positiva (Figura 2).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
281
Figura 2: PCR anidada género específica para Cryptosporidium spp. L: estándar de peso molecular, 1–5: muestras, C(-):
control negativo, C(+): control positivo. Figure 2: Genus specific nested PCR for Cryptosporidium spp. L: ladder, 1-5: samples, C(-): negative control, C(+): positive
control.
Secuenciación Las secuencias obtenidas mostraron más de 92% de identidad nucleotídica con Cryptosporidium parvum, de acuerdo a la información disponible en GenBank. DISCUSIÓN Y CONCLUSIONES La búsqueda de Cryptosporidium y la identificación de su especie en pingüinos tiene un doble propósito, en primer lugar para conocer si son portadores de una especie propia de este parásito, y así a futuro estudiar si estas poseen algún rol patógeno que pueda alterar el estado de conservación de estos. O bien, para identificar la posible fuente de transmisión, que aunque no implique patogenicidad, podría ser un indicador de, por ejemplo, efecto antropogénico o cambio climático. Cryptosporidium parvum ha sido previamente descrito en aves acuáticas (Graczyk et al., 1998; Kuhn et al., 2002; Zhou et al., 2004), sin embargo, hasta la fecha nunca en pingüinos. Considerando que las aves no se describen como hospederos primarios de esta especie de Cryptosporidium, y que previamente se había buscado la presencia de este género protozoario en la misma zona con resultados negativos (Fredes et al., 2007b; Palacios et al., 2010), los resultados positivos en P. papua de la Base PGVV puede ser considerado un indicador del efecto antropogénico en la Antártica. El hecho de que sólo una de las 12 muestras positivas mediante microscopía óptica haya sido positiva mediante PCR puede deberse principalmente a la dificultad de ruptura de la pared de los ooquistes, característica de resistencia que ha sido ampliamente descrita (Fayer y Xiao, 2008). Por lo tanto, esta es la primera confirmación molecular de la presencia de Cryptosporidium spp. en pingüinos de la Antártica, y la primera descripción de C. parvum en pingüinos. Así también este es el primer registro microscópico y molecular de Cryptosporidium spp. en P. papua de la Base PGVV. AGRADECIMIENTOS Financiamiento parcial: Proyecto FONDECYT 1110255 e INACH T-27-10 REFERENCIAS BIBLIOGRÁFICAS Appelbee, A.J., Thompson, R.C.A. y Olson, M.E. 2005. Giardia and Cryptosporidium in mammalian wildlife—current status and future needs. Trends Parasitol. 21: 370–376.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
282
BirdLife International. 2012. Pygoscelis papua. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2013.1. <www.iucnredlist.org>. Downloaded on 7 July 2013. Couve, E. y Vidal, C. 2005. Pingüinos. Fantástico Sur. Punta Arenas, Chile. 100 p. Daszak, P., Cunningham, A. y Hyatt, A. 2000. Emerging Infectious Diseases of Wildlife – Threats to Biodiversity and Human Health. Science. 287: 443-449. Epstein, J. y Field, H. 2004. Complexities of seeking agents in wildlife reservoirs. Report: WHO/FAO/OIE; 2004 3-5 May 2004. Fayer, R. 2004. Cryptosporidium: a water-borne zoonotic parasite. Veterinary Parasitology 126: 37–56 Fayer, R. y Xiao, L. 2008. Cryptosporidium and Cryptosporidiosis. Boca Raton, USA, CRC Press. 560pp. Fowler, M. y Cubas, Z. 2001. Biology, Medicine and Surgery of South American Wild Animals. Iowa State University Press. 455pp. Fredes, F., Raffo, E. y Muñoz, P. 2007a. First report of Cryptosporidium spp. Oocysts in Stool of Adélie Penguin From the Antarctic Using Acid-fast Stain. Antarctic Science. 19: 437-438. Fredes, F., Madariaga, C., Raffo, E., Valencia, J., Herrera, M., Godoy, C. y Alcaino, H. 2007b. Gastrointestinal Parasite Fauna of Gentoo Penguins (Pygoscelis papua) from the Península Munita, Bahía Paraíso, Antarctica. Antarctic Science. 19 (1): 93-94. Fredes, F., Díaz, A., Raffo, E. y Muñoz, P. 2008. Cryptosporidium spp. Oocysts Detected Using Acid-fast Stain in Faeces of Gentoo Penguins (Pygoscelis papua) in Antarctica. Antarctic Science. 20: 495-496. Gilbert, M., Slingenbergh, J., Xiao, X.. 2008. Climate change and avian influenza. Rev Sci Tech;27(2):459-466. Graczyk, T., Fayer, R., Trout, J. M., Lewis, E. J., Farley, A., Sulaiman, I. y Lal, A. A. 1998. Giardia sp. and infectious Cryptosporidium parvum oocysts in the feces of migratory Canada geese (Branta canadensis). Appl. Environ. Microbiol. 64: 2736–2738 Greger, M. 2007. The Human/Animal Interface: Emergence and Resurgence of Zoonotic Infectious Diseases. Critical Reviews in Microbiology33(4):243-299. Henricksen, S. y Pohlenz, J. 1981. Staining of cryptosporidia by a modified Ziehl-Neelsen technique. Acta Vet Scand. 22:594-6. Humblet, M., Vandeputte, S., Albert, A., Gosset, C., Kirschvink, N., Haubruge, E., Fecher-Bourgeois, F., Pastoret, P. y Saegerman, C. 2012. Multidisciplinary and Evidence-based Method for Prioritizing Diseases of Food producing Animals and Zoonoses. ISSN:1080-6050 CDC Centers for Disease Control and Prevention. Online Report Jones, K., Patel, N., Levy, M., Storeygards, A., Balk, D., Gittleman, J. y Daszak, P. 2008. Global trends in emerging infectious diseases. Nature. 451: 990-994 Kuhn, R., Rock, C. y Oshima, K. 2002. Occurrence of Cryptosporidium and Giardia in wild ducks along the Rio Grande river valley in Southern New Mexico. Appl. Environ. Microbiol. 68: 161–165 Muñoz, P., Fredes, F., Díaz-Lee, A., Mercado y R., Ozaki, L. 2011.Detección de Cryptosporidium spp. en terneras de lecherías de la Región Metropolitana mediante Ziehl Neelsen y confirmada por inmunocromatografía y ensayo molecular. Arch Med Vet 43, 111-116. Ng, J., Pavlasek, I. y Ryan, U. 2006. Identification of Novel Cryptosporidium Genotypes from Avian Hosts. Applied and environmental Microbiology: 7548-7553 Palacios, M., Barbosa, A., Pedraza-Diaz, S., Ortega-Mora, L., Valera, F., Cuervo, J., Benzal, J. y De la Cruz, C. 2010. Apparent absence of Cryptosporidium, Giardia and Toxoplasma gondii in three species of penguins along the Antarctic Peninsula. Antarctic Science. 22(3), 265–270 Potti, J., Moreno, J., Yorio, P., Briones, V., Garcia-Borboroglu, P., Villar, S. y Ballesteros, C. 2002. Bacteria divert resources from growth for magellanic penguin chicks. Ecol Lett 5:709–714. Smith, H.V., Caccio, S.M., Cook, N., Nichols, R.A.B. y Tait, A. 2007. Cryptosporidium and Giardia as foodborne zoonoses. Veterinary Parasitology 149: 29–40 Smith, H. y Nichols, R. 2010. Cryptosporidium: Detection in water and food. Experimental Parasitolpgy 124: 61 – 79. Tavernier, P., Dewulf, J. y Roelandt, S. 2011. Wildtool, a flexible, first-line risk assessment system for wildlife-borne pathogens. Eur J Wildl Res 57:1065-1075. Xiao, L. 2010. Molecular epidemiology of cryptosporidiosis: An update. Experimental Parasitology 124: 80-89. Zhou, L., Kassa H., Tischler, M. L. y Xiao, L. 2004. Host-adapted Cryptosporidium spp. in Canada geese (Branta canadensis). Appl. Environ. Microbiol. 70:4211–4215

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
283
EVALUACIÓN DE UN MÉTODO ANALÍTICO PARA SU IMPLEMENTACIÓN EN LA BÚSQUEDA DE MICROORGANISMOS ANTÁRTICOS PRODUCTORES DE ÁCIDOS GRASOS
POLIINSATURADOS
(Evaluation of an analytical method for use in detecting polyunsaturated fatty acids in Antarctic microorganisms)
Bruzzone J. 1, Olazábal L.2, Loperena L.1
1Departamento de Bioingeniería, Inst. Ing. Química, Facultad de Ingeniería, Universidad de la República, Calle Julio H. y Reissig 565, 11300 Montevideo, Uruguay, email: [email protected] 2Departamento de Desarrollo de Métodos Analíticos, Laboratorio Tecnológico del Uruguay, Avenida Italia 6201, Montevideo, 11500, Uruguay, email: [email protected]
INTRODUCCIÓN Existe un interés creciente por la producción de compuestos con propiedades benéficas para la salud humana, entre los cuales se destacan los ácidos grasos poliinsaturados (AGPI), particularmente los del tipo omega-3 (ω3), como el ácido eicosapentaenoico (EPA) y docosahexaenoico (DHA). Los efectos positivos de dichos ácidos sobre la salud han sido comprobados por numerosos estudios científicos (Simopoulos, 1991; Lagarde, 1999 Baticic et al. 2011; Kobayashi et al. 2011; Sani et al. 2010). La ingesta de AGPI ω3 se produce principalmente a través del consumo de pescado y mariscos, que se caracterizan por ser ricos en ellos. La pesca sola no puede satisfacer la creciente demanda en AGPI ω-3, la acuicultura tampoco, ya que los peces por si mismos no los producen, al igual que en los seres humanos, los acumulan de su dieta al ingerir microorganismos marinos (productores primarios de EPA y DHA). En la actualidad, más de 106 toneladas de aceite de pescado son producidas anualmente, de las cuales, un 90% se utilizan para la acuicultura (Petrie y Singh 2011; Wijesundera et al. 2011). Fuentes alternativas de producción de AGPI ω-3 pueden ser las bacterias, hongos, microalgas y thraustochytridos (Bergé et al. 2005; Hinzpeter et al. 2006; Kelly et al. 2012; Kobayashi et al. 2011; Shene et al. 2010). Estudios desarrollados en bacterias psicrofílicas de origen marino provenientes de la Antártida han demostrado que una alta proporción de sus ácidos grasos son AGPI y que estas bacterias poseen la propiedad de síntesis de EPA y/o DHA como estrategia de adaptación a las condiciones extremas de dicho ecosistema (Nichols et al. 1993; Nichols et al. 1999; Nogi 2011; Russell & Nichols 1999). Las células tratan de mantener la fluidez de las membranas mediante cambios en el grado de saturación de los ácidos grasos en los fosfolípidos a través de un mecanismo denominado adaptación homeoviscosa. Esta adaptación puede implicar, isomerización de ácidos grasos cis a ácidos grasos trans, cambio en la relación de saturación/insaturación de los ácidos grasos y en la relación ácido graso de cadena larga/ácido graso de cadena corta. Por lo tanto, la producción de AGPI por parte de estos microorganismos representa una ventaja fisiológica adaptativa al frío, la cual se comprueba al comparar la existencia de estos ácidos grasos en especies de Shewanella psicrófilas y su ausencia en especies del mismo género, pero nativas de ambientes con temperaturas mesófilas (Bergé et al. 2005; Nogi 2011). De este modo, algunos de estos microorganismos provenientes de la Antártida pueden resultar muy atractivos para su utilización en la producción industrial de AGPI ω-3. La selección de muchos de los aún desconocidos microorganismos marinos, puede dar lugar a cepas que sean mejores productores de AGPI que los que se conocen actualmente. Algunos investigadores en la búsqueda de encontrar un método sencillo y rápido para detectar a las bacterias productoras de AGPI ω-3 han reportado métodos indirectos para su identificación, por ejemplo Ryan y colaboradores (2010) reportan que en bacterias marinas Gram (-) con solo cultivarlas en un medio conteniendo 2,3,5-triphenyltetrazolium chloride

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
284
(TTC), su reducción a triphenyl formazan (TF) y la consiguiente aparición de un color rojo intenso debelaría la presencia de EPA en las mismas. Dicho método es rápido y sencillo pero no es específico y en nuestras experiencias se obtuvieron falsos positivos, corroborados por GC-MS, método de referencia, con el cual se obtiene en forma directa el espectro de masa de los ácidos grasos extraídos de las bacterias. En el presente trabajo se evalúa una técnica de extracción y metilación de los ácidos grasos con posterior identificación por GC-MS que permite su utilización a partir de la biomasa presente en un cultivo bacteriano en placa y se simplifican las etapas de extracción y metilación lo que valida su utilización en el screening de nuevas cepas. MATERIALES Y MÉTODOS Se utilizo una cepa de Shewanella livingstonensis, la cual fue cultivada en medio marino modificado (MMM) líquido a una temperatura de 13°C durante 24 horas, y también se cultivo en placa en el mismo medio sólido a 20°C durante 48 horas. Para la determinación del perfil de ácidos grasos en la biomasa se utilizaron dos métodos: el método de extracción de lípidos (Bligh y Dyer 1959), con algunas modificaciones por tratarse de una muestra bacteriana con posterior transesterificación, y la técnica propuesta. En el método de Bligh & Dyer se utilizó una mezcla de solvente, cloroformo: metanol en una relación 1:2, para la extracción del material graso, además de una solución de K2HPO4 0,05M, junto con homogenización y tratamiento con ultrasonido, para facilitar la ruptura celular. Una vez extraídos los lípidos, se continuó con el proceso de transesterificación, para la obtención de sus derivados volátiles; que consistió en la saponificación del material graso con NaOH en metanol, a 100ºC bajo atmósfera de nitrógeno y BF3 en metanol 14 % como catalizador de la esterificación. Los ésteres de ácidos grasos se extrajeron con hexano. La técnica propuesta consiste en la transesterificación directa, sin previa extracción de los lípidos de la muestra. Se toma directamente la biomasa de un cultivo líquido o de un medio sólido, y se procede con los pasos de la transesterificación, antes mencionados. Para la determinación de la composición de las extracciones, se utilizó un cromatógrafo de gases Agilent 6890N acoplado a un Detector Selectivo de Masa Agilent 5973 Inert, con columna capilar J&W Scientific HP-88 de 100m * 0.25mm – 0.20 um, utilizando como referencia un estándar de composición conocida SUPELCO de 37 Componentes de una mezcla de Ácidos Grasos Metilados. RESULTADOS Y DISCUSIÓN Los resultados obtenidos por el método de Bligh y Dyer y el propuesto, utilizando biomasa de un cultivo en medio líquido y en medio sólido, se muestran en la figura 1. La diversidad de ácidos grasos determinados coincide en las tres muestras, encontrándose entre estos, ácidos poliinsaturados como el EPA, el cual es de gran interés. Si se compara la proporción de cada ácido graso de los cultivos líquidos desarrollados a la misma temperatura (13°C), se puede ver que hay mayor recuperación de ácidos grasos de menor longitud (≤ 15 C) con la metilación directa y mayor recuperación de ácidos grasos de mayor longitud (>15 C) con la técnica de Bligh y Dyer. La proporción de cada ácido graso en el perfil obtenido por metilación directa del cultivo en medio sólido, difiere de los anteriores, no pudiéndose comparar ya que la temperatura de cultivo varió ( 20°C) (Wang et al, 2009).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
285
Figura 1 Perfil de ácidos grasos de Shewanella livingtonesis. %: respecto al contenido total de ácidos grasos determinados en
la muestra. A: Método de Bligh & Dyer, cultivo liquido a 13°C. B: Método de transesterificación directa, cultivo líquido a 13°C. C: Método de transesterificación directa, cultivo solido a 20°C.
Figure 1 Fatty acid profile of Shewanella livingtonesis. %: Relative to the total fatty acid content in the sample. A: Method of Bligh & Dyer, liquid culture at 13 ° C. B: Direct transesterification method, liquid culture at 13 ° C. C: Direct
transesterification method, solid culture at 20 ° C.
Si se analizan las muestras obtenidas por transterificación directa además de los ésteres de ácidos grasos se determinan otros compuestos, que están ausentes en el método de Bligh & Dyer. Estos compuestos podrían dificultar la identificación de los ácidos grasos de la muestra, por ejemplo el indicado en la Figura 2 (*) que no es un ácido graso y tiene igual tiempo de retención que el C 20:2cis. Por lo cual es conveniente utilizar para la determinación del perfil de ácidos grasos de microorganismos no identificados un cromatógrafo GC-MS.
C 3
1:9C 2
0:5
C 1
8:3
n3
C 1
9:1
C 1
8:1
cis
C 1
9:1
cis
C 1
8:0
C 1
7:1
C 1
7:0
C 1
6:1
cis
C 1
6:0
C 1
5:1C 1
5:0
C 1
5:0
C 1
4:1
C 1
4:0
C 1
4:0
C 1
3:0
C 1
3:0
C 1
2:0
C 1
1:0
( *)
C 3
1:9C 2
0:5
C 1
8:3
n3
C 1
9:1
C 1
8:1
cis
C 1
9:1
cis
C 1
8:0
C 1
7:1
C 1
7:0
C 1
6:1
cis
C 1
6:0
C 1
5:1C 1
5:0
C 1
5:0
C 1
4:1
C 1
4:0
C 1
4:0
C 1
3:0
C 1
3:0
C 1
2:0
C 1
1:0
( *)
Figura 2 Cromatograma de muestra obtenida de biomasa en medio sólido, cultivo 20°C, por transesterificación directa.
Figure 2 Cromatogram obtained by direct transesterification method, biomass from solid culture at 20 ° C. CONCLUSIONES El método de transesterificación directa propuesto y el análisis de los ácidos grasos obtenidos por GC-MS determinan con certeza los ácidos grasos presentes en la muestra. La disminución en el tiempo de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
286
ensayo y de materiales requerido del método propuesto con respecto al método convencional lo validan como método seguro, rápido y sencillo de uso en el screening de nuevas bacterias potenciales productoras de AGPI. AGRADECIMIENTOS Al Instituto Antártico Uruguayo (IAU) y a la Agencia Nacional de Innovación e Investigación (ANII). REFERENCIAS Batičić L., Varljen N. y Varljen J. 2011. Fish lipids as a source of healthy components: fatty acids from Mediterranean fish. In Biomedical Engineering, Trends, Research and Technologies. Malgorzata Komorowska, Anna; Olsztynska-Janus, Sylwia, editores. India: InTech, cap. 16: 383-406. Bergé J.P., Barnathan G. 2005. Fatty Acids from Lipids of Marine Organisms: Molecular Biodiversity, Roles as Biomarkers, Biologically Active Compounds, and Economical Aspects. Adv Biochem Engin/Biotechnol, 96: 49-25. Bligh,E.G. y Dyer,W.J. 1959. A rapid method for total lipid extraction and purification. Can.J.Biochem.Physiol. 37:911-917 Hinzpeter I., Shene C. y Masson L. 2006. Alternativas biotecnológicas para la producción de ácidos grasos poliinsaturados omega-3. Grasas y Aceites, 57 (3):336-342. Kelly J., Scheibling R. 2012. Fatty acids as dietary tracers in benthic food webs. Marine ecology progress series, 446:1-22. Kobayashi T., Sakaguchi K., Matsuda T., Abe E., Hama Y., Hayashi M., Honda D., Okita Y., Sugimoto S., Okino N. y Ito M. 2011. Increase of Eicosapentaenoic Acid in Thraustochytrids through Thraustochytrid Ubiquitin Promoter-Driven Expression of a Fatty Acid _5 Desaturase Gene. Applied and Environmental Microbiology, 77 (11):3870-3876. Lagarde M., Spector A.A, Galli C., Hamazaki T. y Knapp H.R. 1999. Fatty Acids and Lipids from Cell Biology to Human Disease. Lipids, 34: S1-S350. Nichols D.S, Sanderson K., Bowman J., Lewis T., Mancuso C.A, Mc Meekin T.A y Nichols P.D. 1999. Developments with Antarctic microorganisms: PUFA, culture collections, bioactivity screening and cold adapted enzymes. Current Opinions in Biotechnology, 10:240-246. Nichols D.S., Nichols P.D.y Mc Meekin T.A. 1993. Polyunsaturated fatty acids in antartic bacteria. Antarct Sci., 5:149-160. Nogi Y. 2011. Taxonomy of Psychrophiles. Koki Horikoshi (ed.), Extremophiles Handbook. Japan Agency for Marine-Earth Science and Technology (JAMSTEC), Yokosuka, Japan Petrie J.R. y Singh S.P. 2011. Expanding the docosahexaenoic acid food web for sustainable production: engineering lower plant pathways into higher plants. AoB PLANTS 2011 plr011. http://aobplants.oxfordjournals.org/ Ryan J., Farr H., Visnovsky S. y Visnovsky G. 2010. A rapid method for the isolation of eicosapentaenoic acid-producing marine bacteria, J. Microbiol, Methods, 82(1):49-53. Russell N.J. y Nichols D.S. 1999. Polyunsaturated fatty acids in marine bacteria-a dogma rewritten. Microbiology, 145:767-779. Sani Y.M., Afegbua S.L., Glassey J., Ward A.C. y Rajarajan N. 2010. Effect of growth media modifications on cell biomass and polyunsaturated fatty acids (PUFAs) production from Shewanella frigidimarina. African Journal of Biotechnology, 9 (52):8928-8933. Shene C., Leyton A., Esparza Y., Flores L., Quilodrán B., Hinzpeter I. y Rubilar M. 2010. Microbial oils and fatty acids: effect of carbon source on docosahexaenoic acid (c22:6 n-3, DHA) production by thraustochytrid strains. J. soil sci. plant nutr., 10 (3):207 – 216. Simopoulos A.P. 1991. Omega-3 fatty acids in health and disease and in growth and development. Am J Clin Nutr., 54:438-463. Wang F, Xiao X, Ou HY, Gai Y, Wang F. 2009.Role and regulation of fatty acid biosynthesis in the response of Shewanella piezotolerans WP3 to different temperatures and pressures. J Bacteriol. Apr;191(8):2574-84. Wijesundera C., Kitessa S., Abeywardena M., Bignell W. y Nichols P.D. 2011. Long-chain omega-3 oils: Current and future supplies, food and feed applications, and stability. Lipid Technology March, 23 (3): 55-58.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
287
PYROCOCCUS: UN NUEVO MICROORGANISMO ANTÁRTICO CON ACTIVIDAD NITRILASA
(Pyrococcus: A novel antarctic microorganism with nitrilase activity)
Cabrera, M. A.1,2 y Blamey, J. M.1,2 1Fundación Científica y Cultural Biociencia. José Domingo Cañas 2280. Ñuñoa, Santiago. E-mail: [email protected] 2Facultad de Química y Biología, Universidad de Santiago, Santiago. INTRODUCCIÓN Las nitrilasas (EC 3.5.5.1) son enzimas que convierten los nitrilos a su correspondiente ácido carboxílico y amonio. Éstas son atractivos biocatalizadores en la industria de la química fina y farmacéutica, debido a su regio-, quimio- y enantioselectividad. Los compuestos farmacéuticos se sintetizan a altas temperaturas y por este motivo es importante que las nitrilasas mantengan su actividad en estas condiciones. A la fecha, la mejor nitrilasa reportada tiene una vida media de 6 h a 90°C y proviene de la arquea hipertermófila anaeróbica Pyrococcus abyssi. Debido a que esta nitrilasa proviene de un microorganismo hipertermófilo, decidimos buscar otra nitrilasa en microorganismos hipertermófilos antárticos. Esto porque la Antártica es un continente extremo compuesto no solo de medioambientes fríos sino también de varias zonas geotérmicas, tales como fuentes termales, sistemas hidrotermales y playas con fumarolas, las que proveen uno de los medioambientes más extremos. Por lo tanto, nuestro objetivo fue aislar y caracterizar un microorganismo hipertermófilo antártico que posea actividad nitrilasa. MATERIALES Y MÉTODOS Las muestras se tomaron en Bahía Fumarola, Antártica. Los enrequecimientos se realizaron en cultivos líquidos de una muestra de suelo y el aislamiento de un microorganismo hipertermófilo anaeróbico se llevó a cabo por diluciones seriadas. Se extrajo el DNA genómico de dicho microorganismo y se utilizó para amplificar el gen 16S rRNA mediante PCR, usando primers específicos para bacterias y arqueas. La actividad nitrilasa se detectó por ensayo colorimétrico y, posteriormente, se diseñaron primers degenerados para amplificar el gen de esta enzima. RESULTADOS Aquí presentamos la caracterización de un microorganismo hipertermófilo anaeróbico aislado de la Antártica. Los análisis filogenéticos del gen 16S rRNA mostraron que dicho microorganismo pertenece al género Pyrococcus. Se detectó actividad nitrilasa en el extracto crudo de este microorganismo, la cual fue similar a la de Pyrococcus abyssi. Por otro lado, una vez realizada la amplificación del gen de la nitrilasa por PCR, se observaron dos bandas de 850 pb y 950 pb, las que corresponden al tamaño esperadoro de un gen de nitrilasa procarionte. CONCLUSIONES Hasta la fecha se ha descrito sólo un microorganismo hipertermófilo en la Antártica. Aquí reportamos el aislamiento de un nuevo microorganismo hipertermófilo, el que adicionalmente posee actividad nitrilasa y también poseería el gen de esta enzima. Actualmente, estamos en el proceso de clonamiento y expresión de esta nueva nitrilasa hipertermofílica. AGRADECIMIENTOS Becas CONICYT de capital humano avanzado

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
288
REFERENCIAS Afkhami, A. y Norooz-Asl, R. 2008. Micelle-mediated extraction and spectrophotometric determination of ammonia in water samples utilizing indophenol dye formation. J. Braz. Chem. Soc. 19(8): 1546-1552. Amenábar, M. et al. 2013. Archaeal diversity from hydrothermal systems of Deception Island, Antarctica. Polar Biol 36: 373-380. Fawcett, J. K. y Scott, J. E. 1960. A rapid and precise method for the determination of urea. J. clin. Path. 13: 156-159. Mueller, P. et al. 2006. Cloning, overexpression, and characterization of a thermoactive nitrilase from the hyperthermophilic archaeon Pyrococcus abyssi. Protein Expr Purif. 47(2): 672-681. Thuku, R. N. et al. 2008. Microbial nitrilases: versatile, spiral forming, industrial enzymes. J. Appl. Microbiol. 106: 703-727.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
289
AISLAMIENTO Y CARACTERIZACIÓN DE ACTINOBACTERIAS ANTÁRTICAS
(Isolation and Characterization of Antarctic Actinobacterias)
Cid, F.1,3, Lagos, L.1,3, Lamilla, C.2,3, Jorquera, M.1,3, Salazar, L.2,3, Barrientos, L. 2,3
1Centro de Interacción Suelo, Planta y Biotecnología de Recursos Naturales. 2Centro de Biología Molecular y Farmacogenética. [email protected] 3Scientific and Technological Bioresource Nucleus Universidad de La Frontera, Casilla 54-D, Temuco, Chile. INTRODUCCIÓN Las Actinobacterias se describen como bacterias filamentosos Gram-positivas que representan una de las mayores unidades taxonómicas entre los 18 linajes principales reconocidos actualmente dentro del dominio Bacteria (Ventura et al., 2007). Estos microorganismos se encuentran ampliamente distribuidos en los ecosistemas terrestres y acuáticos, sobre todo en el suelo, donde juegan un papel crucial en la biodegradación de éste y en la formación de humus por el reciclado de los nutrientes asociados con polímeros recalcitrantes tales como la queratina, lignocelulosas y quitina (Stach & Bull, 2005). Alrededor del 45% de todos los metabolitos bioactivos microbianos descubiertos, son producidos por Actinobacterias y muchos de estos metabolitos secundarios son potentes antibióticos, principalmente obtenidos desde el género Streptomyces, que son los principales organismos productores de antibióticos explotados por la industria farmacéutica (Berdy, 2005; Wong et al., 2011). La industria farmacéutica, en el transcurso de varias décadas, ha aislado y analizado millones de cepas microbianas de ambientes terrestres. Por consiguiente, las posibilidades de aislar nuevas cepas microbianas, y concomitantemente el hallazgo de compuestos novedosos, se ha reducido sustancialmente (Lam, 2007; Gesheva & Negoita, 2011). Los medicamentos, especialmente nuevos antibióticos, se necesitan con urgencia para contrarrestar y revertir la propagación de agentes patógenos resistentes a los antibióticos (Payne et al., 2007) y para detener las enfermedades potencialmente mortales, como el cáncer (Olano et al., 2009). La Antártica ofrece una oportunidad única para llevar a cabo la investigación científica en uno de los entornos menos caracterizados y prístinos del planeta con condiciones climáticas adversas que delimitan así la diversidad de los seres vivos. Por estas razones, las áreas sin explotar de la Antártica podrían ser unos recursos valiosos para el descubrimiento de microorganismos biotecnológicamente explotables y para obtener nuevos productos secundarios del metabolismo, como resultado de la influencia del medio ambiente local. Dado lo anteriormente expuesto, el objetivo de esta investigación es aislar y caracterizar Actinobacterias provenientes de suelos y sedimentos marinos obtenidos en la Antártica. MATERIALES Y MÉTODOS Las muestras de suelo y sedimento marino, fueron obtenidas durante el muestreo realizado en la Expedición Científica Antártica 2012-2013 (ECA 49). Tanto las muestras de suelo como de sedimento marino se desagregaron por sonicación de 1 y 5 g de muestra, respectivamente, a 100 kHz durante 30 s. Las muestras de suelo se re-suspendieron en 45 ml de agua destilada mientras que los sedimentos marinos se re-suspendieron en 10 ml de solución salina (0,85%). Se prepararon diluciones seriadas, las que fueron plaqueadas sobre el medio NM1, que contienen agua destilada, 0,5 g de extracto de levadura, 0,5 g de peptona, 0,5 g de glucosa, 0,5 g de glutamato de sodio, 0,44 g de KH2PO4, 0,1 g de (NH4) SO4, 0,1 g de Mg SO4 * 7H2O, 1ml vitaminas (Zhang, 2003) y el medio ISP2 que contiene 10,0 g de extracto de malta, extracto de levadura 4,0 g y 4,0 g de glucosa, pH 7,2 (Cruz et al., 2013). Todos los medios fueron suplementados con 18 g de agar L-1 para aislar bacterias generales (medio NM1) y Actinobacterias (medio ISP2). Las muestras de sedimento marino fueron sembradas en medios de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
290
cultivo adicionados con 33 g L-1 de sal marina. La cantidad de Unidades Formadoras de Colonias (UFC) se determinó en los mismos medios descritos anteriormente. Las placas (en triplicado) se incubaron a 4° C y 15° C, durante 3 a 4 semanas. Las colonias obtenidas de esta manera se transfirieron a medio fresco, se purificaron en agar y después de obtener los respectivos cultivos puros, éstos se almacenaron en agar Luria Bertani (LB) adicionado de glicerol a -20° C. También, se determinó la morfología macroscópica y microscópica de cada una de las colonias obtenidas, así como la caracterización bioquímica de los aislados. RESULTADOS Y DISCUSIÓN Los recuentos más altos de UFC fueron encontrados en muestras de suelo, específicamente en las obtenidas en la playa del Glaciar Collins, alcanzando cantidades de 1x107 UFC g de suelo-1 a 15° C en medio de cultivo NM1. La cantidad más baja de UFC g suelo-1 se encontró en las muestras provenientes de la Bahía Fildes crecido en medio NM1 a 4° C, con valores cercanos a 1x105 UFC g suelo-1. Estos resultados coinciden con otros resultados reportados en la literatura con recuentos máximos en el intervalo de 1,17 x 106 a 4,2 × 106 UFC suelo g-1 (Lee & Hwang, 2002).
Figura 1. Recuento de Unidades Formadoras de Colonias (UFC) en medios de cultivo NM1 e ISP2, incubados a 15 y 4° C, en
muestras de suelo provenientes del glaciar Collins y de la Bahía Fildes. Figure 1. Colony Forming Units (CFU) in NM1 and ISP2 culture media incubated at 15 and 4°C, from Collins Glacier and
Fildes Bay soil samples. En las muestras de sedimentos marinos, el recuento más alto obtenido como UFC g de sedimento-1 se registró en la muestra proveniente de la Base O'Higgins, con 1x104 CFU en el medio ISP2, mientras que el área de muestreo con el menor recuento de UFC g de sedimento-1 se encontró en la muestra procedente de la Base Prat, donde no fue posible obtener ningún aislado con el medio de cultivo ISP2 y menos de 1x101 UFC g de sedimento-1 en el medio de cultivo NM1. Estos resultados coinciden con otras investigaciones sobre recuentos de Actinobacterias, cuyos resultados oscilaron entre 1,4x105 a 1,8x102 UFC g sedimentos-1 (Takizawa et al, 1993).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
291
Figura 2. Recuento de Unidades Formadoras de Colonias (UFC) en medios de cultivo NM1 e ISP2, en muestras de sedimento marino obtenidas desde: A: Isla Decepción , B: Base O´Higgins, C: Isla Hanna, D: Isla Dee, F: Base Prat, G: Isla Robert, H:
Isla Nelson, I: Glaciar Collins, J: Bahía Fildes. Figure 2. Colony Forming Units (CFU) obtained in marine sediments samples from: A: Decepción Island , B: O´Higgins Base, C: Hanna Island, D: Dee Island, F: Prat Base, G: Robert Island, H: Nelson Island, I: Collins Glacier, J: Fildes Bay.
Con respecto a la morfología macroscópica de las colonias obtenidas, destaca la abundante presencia de cepas coloreadas, otras blancas, de aspecto rugoso, y mucoso. Las observaciones microscópicas, mediante tinción de Gram, permitió observar la presencia mayoritariamente de bacilos Gram+. Con respecto a la caracterización bioquímica, los aislados muestran una amplia diversidad. CONCLUSIONES Los datos obtenidos a la fecha coinciden con la literatura revisada y nos entregan promisorios resultados, que permitieron aislar diversos microorganismos, tanto para la caracterización de Actinobacterias antárticas como para el posterior estudio de sus aplicaciones biotecnológicas. AGRADECIMIENTOS Proyecto INACH RT_14-12 REFERENCIAS BIBLIOGRÁFICAS Bérdy, J. (2005). Bioactive Microbial Metabolites. The journal of antibiotics, 58(1), 1-26. Cruz, R. B. D., Galdino, P. M., Penna, K. G. B. D., Hoffmann, K., Costa, E. a, & Bataus, L. a M. (2013). Acetone extract from Streptoverticillium sp., a bacterium isolated from Brazilian Cerrado soil, induces anti-inflammatory activity in mice. Anais da Academia Brasileira de Ciencias, 85(2), 595-603. doi:10.1590/S0001-37652013000200009 Gesheva, V., & Negoita, T. (2011). Psychrotrophic microorganism communities in soils of Haswell Island, Antarctica, and their biosynthetic potential. Polar Biology, 35(2), 291–297. doi:10.1007/s00300-011-1052-8 Lam, K. S. (2007). New aspects of natural products in drug discovery. Trends in microbiology, 15(6), 279–89. doi:10.1016/j.tim.2007.04.001 Lee, J. Y., & Hwang, B. K. (2002). Diversity of antifungal actinomycetes in various vegetative soils of Korea, 417, 407–417. doi:10.1139/W02-025 Olano, C., Méndez, C., & Salas, J. a. (2009). Antitumor compounds from marine actinomycetes. Marine drugs, 7(2), 210–48. doi:10.3390/md7020210 Payne, D. J., Gwynn, M. N., Holmes, D. J., & Pompliano, D. L. (2007). Drugs for bad bugs: confronting the challenges of antibacterial discovery. Nature reviews. Drug discovery, 6(1), 29–40. doi:10.1038/nrd2201 Stach, J. E. M., & Bull, A. T. (2005). Estimating and comparing the diversity of marine actinobacteria. Antonie van Leeuwenhoek, 87(1), 3–9. doi:10.1007/s10482-004-6524-1

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
292
Takizawa, M., Colwell, R. R., & Hill, R. T. (1993). Isolation and Diversity of Actinomycetes in the Chesapeake Bayt. Applied and Environmental Microbiology, 59(4), 997–1002. Ventura, M., Canchaya, C., Tauch, A., Chandra, G., Fitzgerald, G. F., Chater, K. F., & van Sinderen, D. (2007). Genomics of Actinobacteria: tracing the evolutionary history of an ancient phylum. Microbiology and molecular biology reviews : MMBR, 71(3), 495–548. doi:10.1128/MMBR.00005-07 Wong, C., Tam, H., Alias, S., González, M., González-Rocha, G., & Domínguez, M. (2011). Pseudomonas and Pedobacter isolates from King George Island inhibited the growth of foodborne pathogens. Polish polar research, 32(1), 3–14. doi:10.2478/v10183 Zhang, H. (2003). Gemmatimonas aurantiaca gen. nov., sp. nov., a Gram-negative, aerobic, polyphosphate-accumulating micro-organism, the first cultured representative of the new bacterial phylum Gemmatimonadetes phyl. nov. International Journal of Systematic and Evolutionary Microbiology, 53(4), 1155–1163. doi:10.1099/ijs.0.02520-0

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
293
PURIFICACIÓN PARCIAL DE LA ACTIVIDAD TELURITO REDUCTASA DEL AISLADO ANTÁRTICO PSEUDOMONAS SP. BNF22
(Partial purification of the tellurite reductase activity from the Antarctic isolate Pseudomonas sp. BNF22)
Cornejo, F.1, Pugin B.1, García J.1, Díaz-Vásquez W.A.1, 2,3, Vásquez C.C.1 y Arenas F.A.1
1 Laboratorio de Microbiología Molecular, Universidad de Santiago de Chile. [email protected] 2 Laboratorio de Microbiología Molecular y Biotecnología Ambiental, Universidad Técnica Federico Santa María. 3 Facultad de Ciencias de la Salud, Universidad San Sebastián. INTRODUCCIÓN El teluro es un metaloide que integra el grupo 16 de la Tabla Periódica, al que también pertenecen elementos esenciales como oxígeno (O), azufre (S) y selenio (Se). La abundancia del teluro en la corteza terrestre es bastante baja, alrededor de 0,027 ppm (Turner y cols., 2011) y se encuentra mayormente asociado a otros metales tales como oro, plata, cobre y azufre (Chasteen y cols., 2009). Aunque la toxicidad del estado elemental no está completamente confirmada, los derivados del teluro telurato (Te+6) y telurito (Te+4) son extremadamente nocivos para la mayoría de los organismos. Las bases moleculares que explican la toxicidad del telurito aún no han sido completamente dilucidadas. Sin embargo, se ha observado que la exposición a TeO3
2- de bacterias como Escherichia coli (Tantaleán y cols., 2003), Pseudomonas pseudoalcaligenes (Tremaroli y cols., 2007, 2009) y Rhodobacter capsulatus (Borsetti y cols., 2005) genera un estado de estrés oxidativo, debido a un aumento en los niveles de especies reactivas de oxígeno (ROS), especialmente de anión superóxido (Tremaroli y cols., 2007, 2009; Pérez y cols., 2007). Desde el punto de vista de los mecanismos de resistencia bacteriano a telurito se ha observado que bacterias resistentes como sensibles (en presencia de concentraciones sub-letales de este tóxico), acumulan depósitos negros intracelulares (Tucker y cols., 1962), identificados como teluro metálico (Tucker y cols., 1966). Esta capacidad de reducción de telurito a teluro ha sido descrita para una serie de enzimas como nitrato reductasas (Avazéri y cols., 1997; Sabaty y cols., 2001), oxidasas terminales de la cadena respiratoria de diversas bacterias Gram negativo (Trutko y cols., 2000), catalasa (Calderón y cols., 2006) y dihidrolipoamida deshidrogenasa (Castro y cols., 2008), entre otras. La reducción de telurito está acompañada de la formación de superóxido (Calderón y cols., 2006). Desde un punto de vista biotecnológico, la reducción microbiana, enzimática de telurito a teluro elemental parece muy valiosa. De hecho, este proceso convierte telurito, una sal sin valor comercial, en materiales con valor agregado como nanoestructuras de teluro y nuevos materiales con aplicaciones aún no descritas. Actualmente, la generación de materiales de teluro se lleva a cabo casi exclusivamente usando procedimientos químicos. El desarrollo de métodos enzimáticos/microbianos, amigables con el medio ambiente, parece ser una buena alternativa. Dado el interés de nuestro laboratorio por encontrar microorganismos que permitan estudiar nuevos mecanismos de resistencia a telurito, principalmente el fenómeno de reducción del oxianión, nos focalizamos en el continente Antártico. La Antártica exhibe una combinación de condiciones extremas, como bajas temperaturas, sequía y alta radiación UV que generan un estrés de carácter oxidativo (He y Häder, 2002). Teniendo en cuenta que el telurito produce estrés oxidativo en la célula, además de la gran variedad de nichos ecológicos y la conservación de microorganismos antiguos y únicos, lleva a la hipótesis que microorganismos aislados de este continente son capaces de resistir telurito. En efecto, nuestro grupo de investigación aisló e identificó nuevos microorganismos provenientes de muestras

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
294
ambientales obtenidas del territorio Antártico chileno que resisten (MIC 200 µg/ml) y reducen grandes cantidades de telurito (E. coli es considerada una bacteria muy sensible, MIC ~1 µg/ml). Debido a esto, en este trabajo se caracterizó un aislado antártico, Pseudomonas sp. BNF22, altamente resistente y reductor de telurito, con la finalidad de purificar y caracterizar nuevas proteínas con actividad telurito reductasa y así ampliar el conocimiento en el fenómeno de reducción enzimática de este tóxico. MATERIALES Y MÉTODOS Cepas bacterianas: La cepa BNF22, que fue identificada por secuenciación de su RNA 16S y pruebas bioquímicas (API), corresponde a Pseudomonas sp. proveniente de muestras antárticas recolectadas durante la expedición INACH ECA-48 (2012). Actividad telurito reductasa (TR): la cepa se creció a 25 °C en medio LB o en LB diluido 1:2 con y sin telurito a una concentración final de 10 µg/ml. Las bacterias se cosecharon en diferentes fases de crecimiento (exponencial, exponencial tardía y estacionaria) a 8.000 x g por 10 min a 4 °C. Los sedimentos obtenidos fueron suspendidos en tampón Tris 50 mM pH 7,4 suplementado con PMSF 20 µM y sonicados durante 25 min en hielo. Tras la ruptura se centrifugó a 12.000 x g por 10 min a 4 °C y el sobrenadante fue considerado como el extracto crudo. La reacción de reducción de telurito fue llevada a cabo con 100 µg de proteínas a temperatura ambiente durante 1 h en tampón telurito reductasa que contuvo Tris 50 mM pH 7,4, TeO3
2- 1 mM, β-mercaptoetanol 1 mM y NAD(P)H 0,2 mM. Purificación de proteínas con actividad telurito reductasa: 9 g (peso húmedo) de la cepa BNF22 (cosechadas en fase estacionaria) fueron suspendidos en 36 ml de tampón A (fosfato de sodio 10 mM pH 6,5, NaCl 50 mM y β-mercaptoetanol 0,5 mM) y sonicados en hielo. Los restos celulares fueron descartados por centrifugación a 12.000 x g por 10 min a 4 °C. El sobrenadante fue tratado con sulfato de estreptomicina (1% p/v final) por 30 min en hielo y el precipitado fue descartado a 12.000 x g por 20 min a 4 °C. El sobrenadante fue tratado con DNasa I y RNasa I (15,4 µg/ml c/u) durante la diálisis contra tampón A por 1 h con reiterados cambios de tampón. EL resultado fue considerado el extracto crudo, que fue cargado en una columna de fosfocelulosa (Sigma) (2,4 x 7,5 cm) equilibrada en tampón A. La actividad telurito reductasa no se unió a la resina por lo que el eluido fue inmediatamente cargado en una columna DEAE Sepharose (Pharmacia) (2,4 x 6,5 cm) equilibrada en tampón A. Luego de lavar con 10 volúmenes de columna, las proteínas adsorbidas fueron eluidas con un gradiente lineal (500 ml) de NaCl 0 a 0,5 M en tampón A. La actividad telurito reductasa de las fracciones fue detectada como se describió más arriba. RESULTADOS Las condiciones óptimas de crecimiento para una mayor reducción se resumen en la Tabla 1. El crecimiento en presencia de K2TeO3 no conlleva un aumento de esta actividad (inducción).
Cepa Medio de
cultivo Fase de
crecimiento Dador de electrones
Tratamiento con SDS
Tratamiento con proteinasa K
BNF22 LB Estacionaria NADPH NR NR SDS 1% final en el ensayo; proteinasa K 20 µg/ml por 10 min. NR: No reducción.
Tabla 1: Condiciones óptimas de crecimiento para mayor reducción del tóxico. Table 1: Optimal condition of growth to obtain the best tellurite reduction rate.
La purificación de la actividad TR se llevó a cabo por diferentes pasos cromatográficos, en los que la mayor parte de la actividad fue adsorbida en la columna de DEAE Sepharosa, resolviéndose en un solo componente (Fig. 1). La Tabla 2 resume la purificación de la actividad TR. Análisis de actividad TR en geles nativos mostraron que estas fracciones poseen gran actividad reductora. Sin embargo, la

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
295
purificación ha sido solamente parcial (no mostrado). Así, nuevos pasos de purificación y optimización están siendo llevados a cabo para obtener la mayor pureza posible.
Figura 1: Perfil de elución de la actividad TR de la cepa BNF22 en DEAE Sepharose. Las condiciones de la cromatografía se
describieron en materiales y métodos. Figure 1: Elution profile of TR activity from the BNF22 strain on DEAE Sepharose. Chromatography conditions are
described in materials and methods.
Paso Proteína total (mg)
[ Proteínas] (µg/µl) Actividad TR (U) Actividad específica (U/mg)
Factor de purificación
Lisado celular 421,74 10,54 2,66 5,05 1 Extracto crudo 396,86 11,34 5,69 10,04 1,99
DEAE Sepharose
63,16 1,40 20,31 289,36 57,27
Tabla 2: Purificación de la actividad telurito reductasa de BNF22. Table 2: Purification of tellurite-reducing activity from BNF22.
Figura 2: Comparación de actividad TR entre el componente E3 de Aeromonas caviae ST purificado (A) y la actividad
parcialmente purificada de BNF22 (B). La reacción contuvo 20 g de proteína en cada caso. Figure 2: Tellurite reductase activity comparison between the purified E3 component of Aeromonas caviae ST and the
partially purified activity from BNF22. The reaction contained 20 µg of protein in each case.
DISCUSIÓN Se purificó parcialmente una actividad TR del aislado antártico Pseudomonas sp. BNF22. La fracción que contiene la enzima responsable presenta características que difieren respecto a telurito reductasas purificadas previamente, que usan preferentemente NADH como dador de electrones en circunstancia que la TR purificada en este trabajo usa preferentemente NADPH como cofactor. Esto podría representar diferencias en el mecanismo de reacción o en el sitio activo. Por otro lado, esta enzima (a pesar de su baja pureza) posee mayor actividad que el componente E3 del complejo piruvato deshidrogenasa de Aeromonas caviae ST (Fig. 2), la que hasta el momento es la que posee mayor actividad entre las descritas a la fecha (Castro et al., 2008). Esto permitirá, luego de su purificación e identificación, utilizarla para generar nuevas nanoestructuras de teluro y caracterizar el mecanismo molecular de reducción.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
296
CONCLUSIÓN Se aisló proteínas con actividad TR a partir del aislado antártico hiperresistente y reductor de telurito Pseudomonas sp. BNF22. La actividad usa preferentemente NADPH como cofactor y presenta mayor actividad que la observada en telurito reductasas descritas previamente.
AGRADECIMIENTOS Este trabajo fue financiado por el proyecto Fondecyt # 1130362 (CCV) y el proyecto Fondecyt de Postdoctorado # 1130362 (FAA). También se agradece a la Expedición INACH ECA-48.
REFERENCIAS Avazéri, C., Turner, R.J., Pommier, J., Weiner, J.H., Giordano, G. y Verméglio, A. 1997. Tellurite reductase activity of nitrate reductase is responsible for the basal resistance of Escherichia coli to tellurite. Microbiology 143: 1181–1189. Borsetti, F., Tremaroli, V., Michelacci, F., Borghese, R., Winterstein, C., Daldal, F. y Zannoni, D. 2005. Tellurite effects on Rhodobacter capsulatus cell viability and superoxide dismutase activity under oxidative stress conditions. Res. Microbiol. 156: 807–813. Calderón, I.L., Arenas, F.A., Pérez, J.M., Fuentes, D.E., Araya, M.A., Saavedra, C.P., Tantaleán J.C., Pichuantes S.E., Youderian P.A. y Vásquez C.C. (2006) Catalases are NAD(P)H-dependent tellurite reductases. PLoS ONE 1:e70. Castro, M.E., Molina, R., Díaz, W., Pichuantes, S.E. y Vásquez, C.C. 2008 The dihydrolipoamide dehydrogenase of Aeromonas caviae ST exhibits NADH-dependent tellurite reductase activity. Biochem. Biophys. Res. Commun. 375: 91–94. Chasteen, T.G., Fuentes, D.E., Tantaleán, J.C. y Vásquez, C.C. 2009. Tellurite: history, oxidative stress, and molecular mechanisms of resistance. FEMS Microbiol. Rev. 33: 1–13. He, Y.-Y. y Häder, D.P. 2002. UV-B-induced formation of reactive oxygen species and oxidative damage of the cyanobacterium Anabaena sp.: protective effects of ascorbic acid and N-acetyl-L-cysteine. J. Photochem. Photobiol. B 66: 115–124. Pérez, J.M., Calderón, I.L., Arenas, F.A., Fuentes, D.E., Pradenas, G.A., Fuentes, E.L., Sandoval J.M., Castro M.E., Elías A.O., y Vásquez C.C. 2007. Bacterial toxicity of potassium tellurite: unveiling an ancient enigma. PLoS ONE 2: e211. Sabaty, M., Avazeri, C., Pignol, D. y Verméglio, A. 2001. Characterization of the reduction of selenate and tellurite by nitrate reductases. Appl. Environ. Microbiol. 67: 5122–5126. Tantaleán, J.C., Araya, M.A., Saavedra, C.P., Fuentes, D.E., Youderian, P. y Vásquez C.C. 2003 The Geobacillus stearothermophilus V iscS Gene, enconding cysteine desulfurase, confers resistance to potassium tellurite in Escherichia coli K-12. J. Bacteriol. 185: 5831–5837. Tremaroli, V., Fedi, S. y Zannoni, D. 2007. Evidence for a tellurite-dependent generation of reactive oxygen species and absence of a tellurite-mediated adaptive response to oxidative stress in cells of Pseudomonas pseudoalcaligenes KF707. Arch. Microbiol. 187: 127–135. Tremaroli, V., Workentine, M.L., Weljie, A.M., Vogel, H.J., Ceri, H., Viti, C., Tatti E., Zhang, P., Hynes, A., Turner, R. y Zannoni, D. 2009. Metabolomic investigation of the bacterial response to a metal challenge. Appl. Environ. Microbiol. 75: 719–728. Trutko, S.M., Akimenko, V.K., Suzina, N.E., Anisimova, L.A., Shlyapnikov, M.G., Baskunov, B.P., Duda, V. y Boronin, A. 2000. Involvement of the respiratory chain of gram-negative bacteria in the reduction of tellurite. Arch. Microbiol. 173: 178–186. Tucker, F.L., Thomas, J.W., Appleman, M.D., Goodman, S.H. y Donohue, J. 1966. X-ray diffraction studies on metal deposition in group D streptococci. J. Bacteriol. 92: 1311–1314. Tucker, F.L., Walper, J.F., Appleman, M.D. y Donohue, J. 1962. Complete reduction of tellurite to pure tellurium metal by microorganisms. J. Bacteriol. 83: 1313–1314. Turner, R.J., Borghese, R. y Zannoni, D. 2011. Microbial processing of tellurium as a tool in biotechnology. Biotechnol. Adv. 30: 954–963.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
297
DETECCIÓN DEL GÉNERO HELICOBACTER EN HECES Y TEJIDOS DE ANIMALES ANTÁRTICOS
(Detection of Helicobacter genus in feces and tissues of Antarctic animals).
Cunachi1,2, M. Contreras1, M. Fernández-Delgado1, D. Sánchez.1, M. A. García-Amado1
1 COEA, Instituto Venezolano de Investigaciones Científicas, Miranda, Venezuela. [email protected] 2 Senescyt-Instituto Antártico Ecuatoriano (INAE). Quito, Ecuador. INTRODUCCIÓN La Antártida constituye un ámbito geográfico de características peculiares y sus glaciares son las acumulaciones de hielo de mayor desarrollo sobre la Tierra. No menos eminente es su megafauna representada principalmente por especies depredadoras de aves y mamíferos de longevidad elevada y reproducción lenta. Aves como skúas y pingüinos y mamíferos como lobos y elefantes marinos, focas y ballenas, se alimentan y viven en comunidad para sobrevivir en este ambiente extremo (Sánchez, 2007). En la Antártida, la base de la cadena alimenticia son los crustáceos llamados “krilles”. Los bancos de krill antártico (Euphausia superba) son estacionales y su abundancia, distribución y ciclo de vida son afectados negativamente por alteraciones ambientales (Tiwari et al. 2008; Flores et al. 2012). El incremento de la actividad antropogénica ha afectado los ecosistemas antárticos a todos los niveles. Uno de estos impactos negativos es la importación de microorganismos que pueden afectar a la fauna silvestre (Nogales et al. 2011). Bacterias entéricas patógenas de humanos como Campylobacter jejuni, Salmonella spp. Yersinia spp. y E coli. ya han sido reportadas en aves y mamíferos marinos (Bonnedahl et al. 2005; Leotta et al. 2006; Grimaldi et al. 2011). El género Helicobacter comprende bacterias que colonizan el tracto gastrointestinal de humanos y animales. La especie gástrica más conocida es H. pylori y su tasa de infección es de alrededor del 50% de la población humana (Cava et al. 2003). Además, algunos estudios sugieren que el agua juega un rol importante en la transmisión de H. pylori y numerosas evidencias han demostrado su presencia en diversas fuentes de agua como microorganismo de vida libre o asociado al plancton (Fernández-Delgado et al. 2008; Artois et al. 2011). Por otra parte, la fauna silvestre antártica es netamente marina y no está exenta de albergar patógenos infecciosos que pueden colonizar o infectar. Helicobacter ya ha sido reportado en heces de aves y mamíferos marinos (Félix et al. 2008; Leotta et al. 2009). En la Antártida, sólo se han reportado bacterias de la familia Helicobactereaceae en pingüinos (Dewar et al. 2013) pero no en otros animales antárticos, por lo que el objetivo de este trabajo fue determinar la presencia del género Helicobacter en la heces de elefantes marinos, focas, skúas y palomas antárticas y en tejidos de anfípodos. METODOLOGÍA Recolección de muestras de heces: Las heces de aves y mamíferos fueron tomadas del suelo y colocadas en recolectores plásticos estériles, las cuales fueron envueltas con parafilm y congeladas a -10°C para su posterior procesamiento. Mientras que los anfípodos, fueron recolectados utilizando una malla sumergida en el mar durante los días de marea baja, luego los anfipodos fueron colocados en un tubo de 50 ml y congelados a -10°C. Extracción de ADN de heces y anfípodos (krill): El ADN de las heces de los mamíferos y aves y del tejido de los anfípodos fue extraído con el kit Power soil DNA isolation (MO BIO Laboratories) utilizando 250 mg de cada muestra siguiendo la metodología sugerida por el fabricante.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
298
Reacción en Cadena de la Polimerasa (PCR): Para comprobar la presencia de ADN bacteriano y descartar posibles inhibiciones se realizó una PCR utilizando cebadores (8F y 1525R) que amplifican el gen 16S ARNr de Eubacteria (Contreras et al. 2007). Para la detección del género Helicobacter se usaron cebadores (HeliF y HeliR) que amplifican una región interna del gen 16S ARNr específica de este género (Germani et al. 1997) y para detectar la especie H. pylori se usaron cebadores específicos (glmMF y glmMR) que amplifican el gen glmM (Kansau et al. 1996; De Reuse et al. 1997). Las PCR se realizaron en un termociclador modelo Gen AMP 9700 utilizando el kit Ready-To-Go PureTaq PCR (Amersham Biosciences). Como control positivo de las reacciones se utilizó ADN de un aislado clínico de H. pylori y como control negativo se utilizó agua estéril. Los amplicones se visualizaron mediante corrida en gel de agarosa-TBE al 2.0% y tinción con bromuro de etidio, observándose por transiluminación a través de radiación UV. RESULTADOS Y DISCUSIÓN Durante el verano austral del 2013, se recolectaron 3 muestras de heces de mamíferos, 3 muestras de heces de aves y 3 muestras de anfípodos de diferentes localidades descritas en la Tabla 1.
Animal N° de muestras
Localidad
Mamíferos 2
Isla Dee Elefante marino (Mirounga leonina)
Foca de weddell (Leptonychotes weddelli)
1 Isla Dee
Aves
Skúa (Stercorarius chilensis) 2 Isla Greenwich
Paloma antártica ( Chionis alba) 1 Isla Barrientos Anfípodos 3 Isla Dee
Tabla 1. Número de muestras de heces y de anfípodos diferentes localidades. Table 1. Number of stool samples and amphipods different locations.
En todas las muestras analizadas de la Antártida se observó la banda esperada para el gen 16S ARNr de Eubacterias confirmando la presencia de ADN bacteriano y ausencia de inhibidores. El género Helicobacter fue detectado sólo en dos muestras, las cuales corresponden a las heces de una skúa y una paloma antártica pero ninguna de éstas presentó la banda esperada para H. pylori (Tabla 2). Estos resultados confirman por primera vez la presencia del género Helicobacter en otras aves además de los pingüinos (Dewar et al. 2013). Sin embargo, la especie H. pylori no fue detectada, por lo que se recomienda hacer una PCR utilizando cebadores específicos para otras especies de este género o secuenciar los 2 productos de PCR para identificar las posibles especies no-H. pylori presentes en estas muestras de heces. En contraste, en los anfípodos y en las heces de los mamíferos no fue detectado el género Helicobacter, a pesar que se ha reportado la presencia de otros patógenos antropogénicos principalmente E. coli entero-patogénica y Salmonella en lobos y elefantes marinos (Palmgren et al. 2000; Hernandez et al. 2007). Se recomienda aumentar el número de muestras de heces en estos mamíferos para confirmar la ausencia de bacterias del género Helicobacter.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
299
Muestras (Samples) Eubacterias Helicobacter spp. H. pylori EM1 (elefante marino) X o EM2 (elefante marino) X o F1 (foca de weddell) X o S1 (skúa Zona C), X S2 (skúa Zona A), X o P1 (Paloma antártica-Barrientos) X anfípodo 1 (krill negro) X o anfípodo 2 (krill anaranjado) X o anfípodo 3 (krill anaranjado) X o
Tabla 2. Detección de Eubacterias, Helicobacter spp. y H. pylori por PCR en las muestras de heces de mamíferos y aves y de tejidos de anfípodos. PCR con resultado positivo √, PCR con resultado negativo X, PCR no aplicable [o].
Table 2. Eubacteria detection, Helicobacter spp. and H. pylori by PCR in stool samples of mammals and birds and amphipod tissues. PCR positive [√], PCR with negative results [X], not applicable PCR [o].
CONCLUSIÓN Este trabajo reporta por primera vez la presencia del género Helicobacter en heces de dos especies de aves antárticas, skúa y paloma antártica, y contribuye al conocimiento de las posibles vías de transmisión de Helicobacter spp. a través de la fauna silvestre en los ambientes marinos. Debido a que H. pylori no fue detectada en el estudio, se considera necesario profundizar en la identificación de otras especies de este género que pudiesen estar presentes en las muestras analizadas utilizando cebadores específicos y/o análisis de secuencias. Se recomienda ampliar el número de muestras de heces de éstas y otras especies de aves para una mejor estimación de la prevalencia de Helicobacter spp. en las aves antárticas. AGRADECIMIENTOS Al COEA. IVIC. Venezuela. INAE (Ecuador). Lab. de Fisiología Gastrointestinal (IVIC). REFERENCIAS Artois, M. Blacou, J. Dupeyroux, O. y Gilot-Fromont, E. 2011. Sustainable control of zoonotic pathogens in wildlife: how to be fair to wild animals? Rev sci. tech.Off. int. Epiz. (30): 733-743. Bonnedahl, J. Broman, T. Waldestróm, J. Palmgren, H. Niskanen, T. y Olsen, B. 2005. In Search of Human-associated bacterial Pathogens in Antarctic Wildlife: Report from Six Penguin Colonies Regularly Visited by Tourists. Ambio. (34):430-432. Cava, F. y Cobas, G. 2003. Dos décadas de Helicobacter pylori. VacciMonitor.1-10. Contreras, M. Morales, A. García-Amado, M. A. De Vera, M. Bermu, y V. Gueneau, P. 2007. Detection of Helicobacter-like DNA in the gastric mucosa of Throughbred horses Letters in Applied Microbiology ISSN 0266-8254. De Reuse, H. Labigne, A. y Mengin-Lecreulx, D. 1997. The Helicobcater pylori ureC Gene Codes for a Phosphoglucosamine Mutase. Journal of Bacteriology. (179): 3488-3493. Dewar, M. Arnould, J.P. Dann, P. Trathan, P. Groscolas, R y Smith, S. 2013. Interspecific variations in the gastrointestinal microbiota in penguins. MicrobiologyOpen. 2(1):195-204. Félix, C. Reyes, E. Orellana, C. Bravo, A. y Abt, Gonzalo. 2008. Actualizaciones en el Patógeno Emergente Helicobacter Pollorum Asociado a Industria Avícola. Ciencia & Trabajo. (28): 47-49. Fernández-Delgado, M. Contreras, M. García-Amado. M.A. Michelangeli, F. Y Suárez, P. 2008. Evidencias de la Transmisión Acuática de Helicobacter pylori. Interciencia. (33): 412-417. Flores, H., Atkinson, A., Kawaguchi, S., Krafft, B., Milinevsky, G., Nicol, S., Reiss, C., Tarling, G.A., Werner, R., Bravo Rebolledo, E., Cirelli, V., Cuzin-Roudy, J., Fielding, S., van Franeker, J.A. Groeneveld, J.J., Haraldsson, M., Lombana, A., Marschoff, E., Meyer, B., Pakhomov, E.A., Van de Putte, A.P., Rombolá, E., Schmidt, K., Siegel, V., Teschke, M., Tonkes, H., Toullec, J.-Y., Trathan, P.N., Tremblay, N., y Werner, T. 2012. The Response of Antarctic Krill to climate change: Implications for management and research priorities. Mar Ecol Prog Ser. (458):1-19.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
300
Germani, Y. Dauga, C. Duval, P. Huerre, M. Levy, M. Pialoux, G. Sansoneti, P. grimont. P. A. 1997. Strategy for the detection of Helicobacter species by amplification of 16S rRNA genes and identification of H. felus in a human gastric biopsy. Research in Microbiology. (148): 315-326. Grimaldi, W. y Jabour, J. 2011. Considerations for minimising the spread of infectious disease in Antarctic seabirds and seals. Polar Record. (47): 56-66. Hernandez, J. Prado, V. Torres, D. Waldenström, J. Haemig, P. y Olsen, B. 2007. Enteropathogenic Escherichia coli (EPEC) in Antártic fur seals Arctocephalus gazella. Polar Biol. (30):1227-1229. Kansau, I. Raymond, J. Bingen, E. Courcoux, P. Kalach, N. Bergeret, M. Braimi, N. Dupon, C. y Labigne, A. 1996. Genotyping of Helicobacter pylori isolates by sequencing of PCR products and comparison with the RAPD technique. Research in Microbiology. (147): 661-669. Leotta, G. Vigo, G. y Giacoboni, G. 2006. Isolation of Campylobacter lari from sea birds in Hope Bay, Antarctica. Polish Polar Research. (27):303-308. Leotta, G. A. Piñeyron, P. Serena, S. y Vigo, G.B. 2009. Prevalence of Edwardsiella tarda in Antarctic wildlife. Polar Biol. (32):809-812. Nogales. B. Lanfranconi, M. P. Piña-Villalonga, J.M. Bosch, R. 2011. Anthropogenic perturbations inmarine microbial communities.FEMS Microbiol Rev. (35):275-298. Palmgren, H. McCafferty, D. Aspán, A. Broman, T. Sellin, M. Wollin, R. Bergström, S y Olsen, B. 2000. Salmonella in sub.Antarctica: low heterogeneity in salmonella serotypes in South Georgian seals and birds. Epidemiol. Infect. (125):257-262. Sánchez, R. A. 2007. Antártida. Introducción a un continente remoto. Editorial Albatros. Buenos Aires. 3-10. Tiwari, A. Krishnan, K.P. Ravindra, R. 2008. The Story of Antarctica. National Center for Antarctic & Ocean Research and Geological Society of India. 64.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
301
UNA NUEVA NITRILASA PROVENIENTE DE MICROORGANISMOS HIPERTERMOFILOS ANTARTICOS
(Novel nitrilase from Antarctic hyperthermophilic microorganisms)
Dennett, G.1,2, Blamey, J.M.1,2 1Departamento de biología, Facultad de Química y Biología, Universidad de Santiago de Chile 2Fundación científica y cultural Biociencia, Santiago, Chile. Email: [email protected] INTRODUCCIÓN La industria farmacéutica y química utiliza distintos compuestos nitrilados para la manufacturación de una variedad de compuestos como ácido nicotínico, ácido mandélico, ibuprofeno, pesticidas, antibióticos, entre otros. Sin embargo, la transformación de estos nitrilos se ve dificultada debido a las duras condiciones de reacción que requiere la hidrólisis de estos compuestos. La hidrólisis química se lleva acabo a elevadas temperaturas, en condiciones de pH altamente alcalinas ó muy acidas. Además la mayor parte de los nitrilos generados como subproducto en las diferentes industrias que utilizan nitrilos son altamente tóxicos. La biotransformación de los nitrilos se ha considerado como una eficiente vía alternativa a la transformación química. La degradación microbiana de nitrilos ocurre a través de procesos enzimáticos que involucran nitrilasas y nitrilo-hidratasas. Las nitrilasas son enzimas tiólicas que catalizan la conversión directa de los nitrilos a su correspondiente ácido carboxílico y amonio. Estas se caracterizan por catalizar la transformación de una amplia variedad de sustratos y por no utilizar cofactores, mientras que la nitrilo-hidratasa cataliza la formación de amidas a partir de nitrilos con la utilización de cofactores. En la actualidad, existen una gran variedad de nitrilasas descritas las cuales poseen temperatura óptima entre 30-65°C y un pH óptimo de entre 6 a 9. Sin embargo la industria continúa necesitando nuevas nitrilasas capaces de trabajar en altas temperaturas, pHs extremos y en presencia de solventes orgánicos. Esto se debe a que la producción de diferentes intermediarios para la generación de ciertos compuestos finales, se realiza principalmente a través de síntesis químicas a elevadas temperaturas. Por lo cual, encontrar una enzima estable a altas temperaturas se hace muy relevante, ya que poco se conoce de nitrilasas activas en estas condiciones. Además, la versatilidad biocatalítica (enantioselectividad y amplio rango de sustratos) y las aplicaciones de esta enzima degradadora de nitrilos, son razones por las cuales su utilización es inmensamente requerida para la producción de diversos compuestos en las diferentes industrias, y para el tratamiento de desechos industriales. En la Antártica debido a su composición geológica y sus características geofísicas, es posible encontrar toda clase de nuevos microorganismos extremos (incluyendo hipertermófilos y termófilos), en los diferentes ambientes extremos que posee este continente. Los hipertermofilos son capaces de vivir óptimamente a 80⁰C y más. Entonces, si estos microorganismos poseen actividad nitrilasa, probablemente estas enzimas sean muy estables a altas temperaturas. A lo largo del presente trabajo, diversas muestras ambientales de la Antártica fueron cultivadas en condiciones anaeróbicas y aeróbicas, en busca de actividad nitrilasa. Dentro de las comunidades de microorganismos hipertermofilos se encontró una nitrilasa termoestable capaz de catalizar la conversión de nitrilos aromáticos como el benzonitrilo a una temperatura de 85°C, siendo una de las nitrilasas más termoestables descritas hasta la fecha. Mediante métodos de purificación enzimática se logro purificar y caracterizar esta nueva nitrilasa. En el presente trabajo se muestra los resultados obtenidos para la caracterización de esta enzima.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
302
MATERIALES Y MÉTODOS: Cultivo de microorganismos Se utilizaron 13 muestras ambientales recolectadas en la Isla Decepción durante la expedición ECA 46, para las cuales se registraron temperatura y pH del sitio original desde donde fueron tomadas. Estas muestras fueron cultivadas en condiciones aeróbicas y anaeróbicas para la obtención de microorganismos. Aislamiento de microorganismos Mediante la realización de reiteradas diluciones seriadas de los cultivos mixtos que poseen una actividad nitrilasa y manteniendo las condiciones de selección se logro separar estos microorganismos. Inducción Para inducir la expresión de nitrilasas las muestras fueron incubadas con distintas concentraciones de nitrilos como la principal fuente de nitrógeno. Preparación de extractos crudos A partir de 6 lt de medio de cultivo las células fueron recoletadas y lavadas con buffer Tris-HCl pH 8.0 50mM conteniendo EDTA 2mM. Las células fueron destruidas por sonicación en un sonicador de baño Branson 1510 durante 12 horas y luego centrifugadas a 17.000 xg por 20 a 4°C. Detección de actividad nitrilasa La detección de la actividad enzimática se realiza a través de un ensayo colorimétrico descrito por Fawcett y Scott en 1960 con diferentes sustratos. Purificación enzimática La purificación de la enzima se realizo de acuerdo a los protocolos descritos por Almatawah y col., 1999 y Kobayashi y col., 1990. RESULTADOS: En el presente trabajo se muestran los resultados obtenidos en la búsqueda de una nueva nitrilasa y su posterior caracterización. Esta enzima fue encontrada en comunidades de microorganismos hipertermófilos antárticos que son capaces de vivir en la presencia de benzonitrilo a una concentración de 30mM. Hasta la fecha esta enzima ha sido purificada a partir de un microorganismo hipertermofilo antártico aislado de muestras ambientales. Su actividad máxima fue obtenida a 85°C y a un pH de 5.5 indicando que esta enzima es capaz de hidrolizar benzonitrilo a altas temperaturas y pH acido entre otros nitrilos aromáticos utilizados. Además, esta enzima es capaz de mantener más del 50% de su actividad (57.14%) luego de ser incubada a 85°C durante 8 horas, en comparación con otras enzimas termoestables descritas, como la de P. abbyssi, y la de B. pallidus las cuales solo permanecen activas durante 6 horas a 90°C y 13 minutos a 70°C respectivamente. CONCLUSIONES: En conclusión en este trabajo se ha logrado encontrar una nueva nitrilasa termoestable a altas temperaturas, con una tendencia a realizar la catálisis enzimática en condiciones acidas, y proveniente de comunidades de microorganismos hipertermofilos antárticos capaces de vivir en la presencia de altas concentraciones de benzonitrilo (30mM). Esto nos lleva a concluir que nuestra enzima es una de las mas termoestables descritas hasta la fecha y que posee las características que la industria requiere para su utilización en la producción de diversos compuestos químicos. AGRADECIMIENTOS: Nuestros agradecimientos a Corfo Innova # 07CN13PXT264 (2008-2012) y a la beca otorgada por el Instituto Antártico Chileno (INACH) código D_06-11 por el financiamiento de este trabajo. REFERENCIAS: Almatawah, Q.A., Cramp, R. y Cowan, D. 1999. Characterization of an inducible nitrilase from a thermophilic bacillus. Extremophiles. 3:283–291

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
303
Fawcett J.K. y Scott J.E. 1960. A rapid and precise method for the determination of urea. J.Clin.Path 13:156-159 Harper, D. 1977. Enzymology of C-N cleavage by Nocardia sp. (Rhodochrous group) NCIB. 11216. Biochem. J. 165:309-319 Kobayashi, M., Yanaka, N., Nagasawa, T. y Yamada, H. 1990. Purification and characterization of a novel nitrilase of Rhodococcus rhodochrous K22 that acts on aliphatic nitriles. J Bacteriol 172: 4807–4815

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
304
BÚSQUEDA, IDENTIFICACIÓN Y CARACTERIZACIÓN DE DIVERSAS ACTIVIDADES ENZIMÁTICAS A PARTIR DE UNA COLECCIÓN DE BACTERIAS CULTIVABLES
AISLADAS DE LA ANTÁRTIDA
(Search, identification and characterization of enzymatic activities in a collection of culturable bacteria isolated from Antarctic samples)
Ferrés, I.1, Amarelle, V., Noya, F. y Fabiano, E.
1Departamento de Bioquímica y Genómica Microbianas, Instituto de Investigaciones Biológicas Clemente Estable, Av. Italia 3318, Montevideo, Uruguay. E-mail: [email protected]
INTRODUCCIÓN La bioprospección se define como la búsqueda dirigida de organismos o funciones de utilidad para diferentes fines tales como la producción de nuevos fármacos, enzimas, nutrientes, etc. Las muestras ambientales son habitualmente las elegidas para realizar bioprospección debido principalmente a la diversidad de organismos presentes dentro de un mismo ecosistema (de Pascale et al. 2012). Los ambientes fríos representan un interesante recurso aún poco explorado. Se estima que un 75% de la biosfera de la Tierra se encuentra en entornos perennemente fríos (menores a 5 ºC) (Margesin, y Haggblom, 2007). Es así que estos ambientes representan un recurso atractivo para el descubrimiento de nuevas especies, nuevos procesos y nuevos genes. A pesar del potencial que tienen los ambientes fríos nuestro conocimiento de la ecología, fisiología, metabolismo, enzimología y genética de los organismos adaptados al frío es muy limitado. OBJETIVOS En este trabajo nos planteamos la búsqueda de diversas enzimas en una colección de bacterias cultivables generada a partir de muestras provenientes de la Antártida. Los objetivos específicos fueron: a) Generar una colección de bacterias cultivables aisladas de muestras provenientes de diferentes ambientes de la Antártida, b) identificar los aislamientos bacterianos de la colección que expresen funciones enzimáticas de interés: celulasas, hemicelulasas, lipasas/esterasas, enzimas ligninolíticas y producción de compuestos antimicrobianos. MATERIALES Y MÉTODOS A partir de un total de 20 muestras ambientales distintas tomadas en la campaña antártica ANTARKOS XXIX en los meses de enero y marzo de 2013, se generó una colección de bacterias cultivables psicrófilas/psicrotolerantes. La ubicación de las muestras fue georeferenciada y la colecta se realizó a partir de diferentes ambientes como ser matas microbianas, sedimento, suelo con combustible, agua de mar, agua de deshielo, témpanos, sedimento con líquenes, entre otros. Todas las muestras fueron tomadas en condiciones asépticas y conservadas a 4ºC hasta su procesamiento en el laboratorio. Para la generación de la colección, se realizaron diluciones seriadas de las muestras en NaCl 0,9% estéril y se sembraron las alícuotas en dos medios de cultivo sólido distintos: R2A, reportado para el cultivo de bacterias de crecimiento lento (Reasoner y Geldreich 1985), y medio Marino (DifcoTM) reportado para el cultivo de heterótrofos marinos. Las placas se incubaron a 4ºC durante 15 días y se seleccionaron un total de 32 aislamientos por muestra en base al fenotipo presentado por las colonias, de modo de obtener una colección con diversidad de especies bacterianas. Se ordenó la colección en placas de 96 pocillos conteniendo el medio líquido adecuado y los aislamientos se incubaron a 4ºC durante 15 días.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
305
Para detectar los aislamientos portadores de las funciones de interés se realizaron ensayos cualitativos donde las bacterias se crecieron en medios que contenían diferentes indicadores específicos: carboximetil celulosa y celulosa microcristalina (avicel) para la detección de celulasas (Kim, et al 2008.), xilano para la detección de xilanasas (Kim, et al. 2008), ABTS para peroxidasas (Hofrichter, et al. 1997), ABTS y MnCl2 para manganeso peroxidasas (Hofrichter, et al. 1997), guaiacol y CuSO4 para lacasas (Kiiskinen, et al. 2004) y tributiril para la detección de actividad lipolítica (Cherry y Fidantsef, 2003). También se evaluó la producción de compuestos antimicrobianos y la capacidad de utilización de avicel como única fuente de carbono. Se incubaron los aislamientos de la colección en los medios específicos a 4ºC y se evaluó la presencia de las distintas actividades enzimáticas a lo largo de los días. Para aquellos ensayos donde se necesitó un paso adicional para la visualización de la actividad de interés (revelado), los aislamientos fueron incubados por un total de 20 días antes de ser sujetos al ensayo de revelado. Los clones de la colección se almacenaron en placas multipocillo con glicerol 25% a -80ºC. RESULTADOS A partir de 20 muestras ambientales recolectadas en distintos sitios de la Antártida, se creó una colección de bacterias cultivables psicrófilas/psicrotolerantes. Esta colección cuenta con un total de aproximadamente 1000 aislamientos y se encuentra almacenada en glicerol a -80ºC. Mediante análisis funcional se identificaron hasta el momento 340 aislamientos con actividad lipolítica, 41 con actividad lacasa, 53 con actividad celulasa, 7 con actividad xilanasa y 21 aislamientos capaces de utilizar avicel como única fuente de carbono. No se identificaron aislamientos con actividad detectable de peroxidasa o manganeso peroxidasa. Cuatro aislamientos fueron capaces de producir compuestos antibacterianos, 3 de ellos efectivos contra bacterias Gram positivas y uno de ellos efectivo en Gram negativas. Los aislamientos que presentaron más actividades positivas, fueron aquellos recuperados de una muestra proveniente de un charco de agua de deshielo con contenido de algas y musgo. En esta muestra, de un total de 63 aislamientos, 19 presentaron actividad lipolítica, 10 actividad lacasa, 9 actividad celulasa y 2 con actividad xilanasa. Interesantemente se detectó la producción de más de una actividad en algunos aislamientos. DISCUSIÓN Y CONCLUSIONES En este trabajo se generó una amplia colección de bacterias cultivables psicrófilas/psicrotolerantes a partir de muestras ambientales tomadas en la Antártida. Esta colección demostró ser un nicho muy interesante desde el punto de vista funcional, ya que se obtuvieron varios aislamientos con funciones enzimáticas de interés académico y biotecnológico. En una etapa posterior, se caracterizarán detalladamente aquellas actividades enzimáticas más promisorias y se determinará el microorganismo encargado de su producción mediante secuenciación de la subunidad 16S del ADNr. Los climas fríos pueden albergar una alta diversidad de bacterias cultivables, capaces de mostrar una gran cantidad de actividades enzimáticas. La microbiología clásica funcional representa una metodología sencilla y rápida para detectar las actividades enzimáticas en aquellos microorganismos que son capaces de crecer en los medios de cultivo conocidos. Se estima que una fracción muy pequeña (menor al 1%) de los microorganismos presentes en un determinado ambiente pueden ser recuperados mediante las técnicas tradicionales de cultivo (Giovannoni, et al. 1990). En este trabajo mediante una aproximación microbiológica clásica y utilizando solamente dos medios de cultivo distintos se construyó una amplia colección de bacterias cultivables psicrófilas/psicrotolerantes, las cuales presentaron varias

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
306
actividades enzimáticas de interés biotecnológico. Podemos concluir entonces que si bien la información obtenida mediante esta aproximación sub-representa la información total disponible en un determinado ambiente, es una aproximación sencilla mediante la cual se obtienen muy buenos resultados. Agradecimientos. Instituto Antártico Uruguayo (IAU), PEDECIBA Biología-Química REFERENCIAS Cherry, J. R. y Fidantsef, A. L. 2003. Directed evolution of industrial enzymes: an update. Curr Opin Biotechnol 14(4): 438-443. de Pascale, D., De Santi, C., Fu, J. y Landfald, B. 2012. The microbial diversity of Polar environments is a fertile ground for bioprospecting. Mar Genomics 8: 15-22. Giovannoni, S. J., Britschgi, T. B., Moyer, C. L. y Field, K. G. 1990. Genetic diversity in Sargasso Sea bacterioplankton. Nature 345(6270): 60-63. Hofrichter, M., Bublitz, F. y Fritsche, W. 1997. Fungal attack on coal: Solubilization of low-rank coal by filamentous fungi. Fuel Process Technol. 52: 55-64. Kiiskinen, L. L., Ratto, M., Kruus, K.. 2004. Screening for novel laccase-producing microbes. J Appl Microbiol 97(3): 640-646. Kim, S. J., Lee, C. M., Han, B. R., Kim, M. Y., Yeo, Y. S., Yoon, S. H., Koo, B. S. y Jun, H. K. 2008. Characterization of a gene encoding cellulase from uncultured soil bacteria. FEMS Microbiol Lett 282(1): 44-51. Margesin, R. y Haggblom, M. 2007. Thematic issue: Microorganisms in cold environments. Reasoner, D. J. y Geldreich, E. E. 1985. A new medium for the enumeration and subculture of bacteria from potable water. Appl Environ Microbiol 49(1): 1-7.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
307
VARIABILIDAD INTERANUAL DE LA ABUNDANCIA DE PICOCIANOBACTERIAS EN EL PASAJE DRAKE E INMEDIACIONES DE LA PENINSULA ANTARTICA
(Internannual variability of picocyanobacteria abundance in the Drake Passage and waters surrounding the Antarctic Peninsula)
Franzosi, C.A. 1 y Alder, V. A. 1, 2
1 Departamento de Ecosistemas y Recursos Marinos. Coordinación de Ciencias de la Vida. Instituto Antártico Argentino, Balcarce 290, Buenos Aires, Argentina. [email protected], [email protected] 2 Departamento de Ecología, Genética y Evolución e IEGEBA (CONICET-UBA). Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, C1428EHA Buenos Aires, Argentina RESUMEN Las picocianobacterias, por sus elevadas abundancias, desempeñan un rol clave en la productividad primaria y el ciclo del carbono y nitrógeno de la gran mayoría de los océanos y ambientes continentales. Sin embargo, en los ecosistemas marinos antárticos sus niveles de abundancia han sido reportados como bajos o nulos, y la posibilidad de que se trate de una comunidad de origen estrictamente marino es aún incierta. Los estudios disponibles indican que, en la columna de agua, las picocianobacterias están prácticamente ausentes en aguas antárticas, al sur del Frente Polar (Vincent et al., 2000; Doolittle et al., 2008), representando un componente minoritario del fitoplancton atribuido a probables fenómenos de resuspensión de organismos y/o detritos bentónicos (Walker y Marchant, 1987). En el hielo marino, por su parte, su frecuencia de aparición es muy rara y su origen también es dudoso (Koh et al., 2012), pudiendo su presencia responder a un proceso de retención durante la formación del campo de hielo durante el periodo invernal. La importancia de analizar la contribución en abundancia y biomasa de los grupos de organismos que conforman el picofitoplancton en áreas submuestradas (Buitenhuis et al., 2012) o vulnerables, tal como es el caso de los ecosistemas marinos antárticos, se sostiene con los resultados de los modelos predictivos, los cuales proyectan que el incremento de la temperatura global generará una expansión en la distribución geográfica y abundancia de Synechococcus (Flombaun et al., 2013), picocianobacteria de relevancia numérica y funcional en los ambientes pelágicos templados. En este contexto, el objetivo del estudio fue conocer la magnitud de la densidad y biomasa de las picocianobacterias y su variabilidad espacio-temporal tanto en aguas oceánicas del Pasaje Drake, las cuales representan una transición entre ambientes templados y polares, como en inmediaciones de la Península Antártica, uno de los ambientes de mayor impacto vinculado con el incremento de la temperatura global. Las muestras y mediciones del presente estudio, provienen de cuatro Campañas Antárticas de Verano realizadas a bordo del rompehielos Almirante Irizar desde el 2002 al 2005. Se efectuaron 66 estaciones oceanográficas en aguas subsuperficiales (9 metros de profundidad), según se detalla en la Figura 1. Mediante una bomba centrífuga, en cada una de las estaciones se colectaron muestras de agua de mar para estimar la concentración de picocianobacterias y clorofila, y se realizaron mediciones de temperatura y salinidad, entre otras variables. Para el análisis cuantitativo de picocianobacterias, se fijaron y filtraron entre 3 y 6 mL de agua de mar por filtros negros de policarbonato de 0,2 µm de poro, que fueron montados sobre portaobjetos y mantenidos en freezer. El recuento celular se realizó en toda el área del filtro con un aumento total de 1000x, bajo excitación con luz verde (480-550 nm) y considerando a los organismos con emisión fluorescente color naranja y tamaño ca. de 1µm, como cianobacterias tipo Synechococcus. Las

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
308
mediciones celulares se realizaron sobre imágenes digitales mediante el uso del software Image Pro Plus. La biomasa se estimó según el modelo de relación constante de BjØrnsen (1986).
Figura 1. Ubicación geográfica de las estaciones realizadas durante el verano 2002-2005 en aguas del Pasaje Drake e
inmediaciones de la Península Antártica. Figure 1. Geographic location of Summer 2002-2005 stations in waters of the Drake Passage and off the Antarctic Peninsula
Las picocianobacterias fueron registradas en el 44% del total de estaciones realizadas en el área de estudio. En el Pasaje Drake, la frecuencia de aparición fue notablemente más elevada que en inmediaciones de la Península Antártica (75% y 15%, respectivamente). Dentro del Pasaje Drake, los promedios de abundancia al norte y al sur del Frente Polar fueron del mismo orden de magnitud (103), exceptuando el mes de marzo del 2005, donde el promedio en aguas antárticas disminuyó en un orden de magnitud. Este último caso sugiere cierta influencia de las propiedades de las masas de agua antárticas o del descenso de la temperatura debido al incremento de la latitud, tal como fuera reportado previamente en una transecta similar (Letelier y Karl, 1987). La abundancia y biomasa alcanzaron, sobre los 56°S y a 6,44°C, sendos valores máximos de 5 x 104 cel. mL-1 y 1,46 µgCL-1. No obstante, las abundancias subsuperficiales no fueron significativamente diferentes en los distintos años investigados (Kruskal-Wallis, p=0,24) o entre aguas antárticas y subantárticas del Pasaje (t-test, p=0,08). La continuidad y la homogeneidad de la distribución de los valores de abundancia confirman una clara presencia de cianobacterias tipo Synechococcus en toda la extensión del Pasaje Drake, desde el Frente Subantártico hasta el Frente Sur de la Corriente Circumpolar Antártica (aprox. 56-62°S). Futuros estudios deberán analizar si estos niveles de abundancia en aguas antárticas oceánicas responden a un mecanismo de advección o al transporte hacia el sur por eddies de aguas subantárticas, y/o si el género Synechococcus incluye a una o más especies planctónicas típicas del área. En inmediaciones de la Península Antártica, las picocianobacterias se detectaron en proximidades de islas, dentro de la interfase plataforma-talud (profundidad promedio: 450 m; intervalo: 259-708 m). Los valores más altos de abundancia (5 x 103 y 1 x 103 cel. mL-1) se registraron en febrero de 2003 al norte y sur de la isla 25 de Mayo/Rey Jorge (Shetland del Sur), donde la temperatura, la salinidad y la profundidad del fondo alcanzaron 2,8ºC, 33 y 435 metros, y 2,20ºC, 33,87 y 708 metros, respectivamente. En las estaciones restantes, los incrementos fueron geográficamente aislados, dentro de un intervalo de temperatura de -0,68 a 0,90ºC. En todos los casos se detectaron valores del orden de 102 cél. mL-1, cifras superiores a las reportadas en aguas costeras próximas a la Base McMurdo (Rivkin et al., 1991:

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
309
103 y 104 cél. L-1). Tanto en marzo de 2002 como en marzo de 2005, entre ca. 65 y 68°S, se registró una lengua de baja salinidad (promedio: 32,96) que indicaría el aporte de aguas de derretimiento de origen continental (Dierssen et.al., 2002). Este proceso favoreció la estratificación de la columna de agua y el desarrollo fitoplanctónico, tal como lo revelan los elevados valores de clorofila registrados. El hecho de que en este sector se haya realizado un único hallazgo en 2005 y ninguno en 2002, genera incertidumbre sobre el hábitat de origen de las picocianobacterias. Por otra parte, las magnitudes que presenta el único sitio con picocianobacterias caen en el orden comúnmente señalado para sistemas marinos de latitudes subpolares, como por ejemplo en Bahía Bedford (Li, 1998). Esto lleva a considerar que su origen es planctónico y que varios otros factores actúan sinérgicamente al nivel de microescala, promoviendo su desarrollo o descenso en la columna de agua. Si bien a nivel interanual, no se detectaron diferencias significativas en la abundancia, tanto en aguas estrictamente oceánicas del Pasaje Drake como al norte de la Península Antártica, las picocianobacterias alcanzaron su máxima abundancia durante febrero (2003), reflejando la influencia del aumento estival de temperatura. AGRADECIMIENTOS Se agradece la colaboración de la tripulación del rompehielos Almirante Irizar durante la ejecución del trabajo de campo. El presente estudio fue realizado con el apoyo de subsidios otorgados a VAA por el Instituto Antártico Argentino-DNA y ANPCYT (PICT 2000- 9108) y CONICET (PIP 02406) REFERENCIAS Buitenhuis, E.T., Li,,W.K.W., Vaulot, D., Lomas, M.W., Landry, M., Partensky, F., Karl, D.M., Ulloa, O., Campbell, L. Jacquet, S., Lantoine, F., Chavez, F., Macias, D., Gosselin, M. y McManus, G.B. 2012. Picophytoplankton biomass distribution in the global ocean. Earth System Science Data, 4(1): 37-46. Bjørnsen, P.K. 1986. Automatic Determination of Bacterioplankton Biomass by Image Analysis. Applied and Environmental Microbiology, 51(6): 1199-1204. Dierssen, H.M., Raymond, C., Smith, R.C. y Vernet, M. 2002. Glacial meltwater dynamics in coastal waters west of the Antarctic peninsula, Proceedings of the National Academy of Sciences of the United States of America, 19: 99(4): 1790–1795. Doolittle D.F., Li W.K.W. y Wood A.M. 2008.Wintertime abundance of picoplankton in the Atlantic sector of the Southern Ocean. Nova Hedwigia, 133:147–160 Flombaum, P., Gallegos, J.L., Gordillo, R.A., Rincón, J., Zabala, L.L., Nianzhi Jiao, N., Karl, D.M., Li, W.K., Lomas, M. W., Veneziano, D., Vera, C.S., Vrugt, J.A. y Martiny, C.A. 2013. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proceedings of the National Academy of Sciences of the United States of America, 110: (24): 9824-9829. Letelier, R.M. y Karl, D.M. 1989. Phycoerythrin-containing Cyanobacteria in surface waters of the Drake Passage during February 1987.Antarctic Journal of the US, 24(5):185-188 Li, W.K. 1998. Annual average abundance of heterotrophic bacteria and Synechococcus in surface waters. Limnology and Oceanography, 43:1745–1753. Koh, E.Y., Cowie, R.O.M., Simpson, A.M., O'Toole, R. y Ryan, K.G. 2012. The origin of cyanobacteria in Antarctic sea ice: marine or freshwater?. Environmental Microbiology Reports, 4:479–483. Rivkin, R,B., Anderson, M.R. y Gustafson D.E.1991. Dynamics of bacterioplankton growth in McMurdo Sound, Antarctica: evidence for substrate sufficient growth. Antarctic Journal of the US, 26: 145-146 Vincent, W.F., Bowman, J.P., Rankin, L.M. y McMeekin, T.A. 2000. Phylogenetic diversity of picocyanobacteria in Arctic and Antarctic ecosystems. En: Bell, C., Brylinsky, M., Johnson-Green, M. (Eds). Microbial biosystems: new frontiers. 8th International Symposium on Microbial Ecology. Atlantic Canada Society for Microbial Ecology, Halifax, 317–322 Walker, T.D. y Marchant H.J. 1989. The seasonal occurrence of chroococcoid cyanobacteria at an Antarctic Coastal site. Polar Biology, 9 :193–196.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
310
BACTERIAS RIZOSFÉRICAS DE Deschampsia antárctica CON ACTIVIDAD ANTIBACTERIANA
(Rhizosferic bacteria of Deschampsia antarctica with antibacterial activity)
Gallardo-Cerda J.1, Lavin P.1, Zurita, A.2
1Laboratorio de Biorrecursos Antárticos, Instituto Antártico Chileno, INACH, Punta Arenas, Chile. [email protected], 2Instituto de Investigaciones Agropecuarias, INIA Intihuasi.
INTRODUCCIÓN Algunas cepas del género Pseudomonas, son conocidas por promover el crecimiento de plantas y ayudar a evitar potenciales consecuencias negativas producto del bajo contenido de nutrientes o ataque de patógenos. Un amplio rango de características de este género son responsables estos efectos benéficos. Muchas Pseudomonas son capaces de solubilizar minerales insolubles como fosfatos, dejándolos disponibles para las plantas promoviendo su crecimiento (Kucey et al., 1989; Rodríguez & Fraga, 1999). Las Pseudomonas colonizadoras de raíces son capaces de promover indirectamente el crecimiento de las plantas otorgando protección en contra de microorganismos patógenos. El mecanismo más importante en contra del ataque de microorganismos patógenos es la inducción de la resistencia sistémica adquirida (Bakker et al., 2007) y la supresión directa de patógenos de suelo por medio de la producción de metabolitos antimicrobianos (Haas & Keel, 2003). Deschampsia antarctica es una de las dos plantas vasculares que han colonizado la Antártica marítima. Esta especie ha aumentado el tamaño de su población en la Antártica con mayor velocidad que los briófitos en respuesta al calentamiento global (Lewis-Smith, 1994) lo que sugiere una gran capacidad de adaptación. Muchos estudios han encontrado que D. antarctica posee mecanismos conservados que le permiten tolerar muchos factores abióticos estresantes, entre los que se encuentran la adaptación del sistema fotosintético a baja temperatura (Edward & Lewis-Smith, 1988; Xiong et al., 2000), acumulación de carbohidratos no estructurales y prolina (Zuñiga et al., 1996; Bravo et al., 2001). A pesar del extensivo estudio sobre las adaptaciones fisiológicas de esta especie, son pocos los estudios sobre el bacterioma asociado a D. antárctica y su relación con la capacidad de esta planta de tolerar condiciones abióticas estresantes. MATERIALES Y MÉTODOS Muestra vegetal. Las muestras de D. antárctica fueron obtenidas desde Isla Edward (Antártica). Las plantas fueron sacadas con su pan radicular, puestas en bolsas estériles en un cooler a 4°C para su traslado a laboratorio. Aislamiento e identificación de bacterias cultivables. Para obtener muestras representativas del rizoplano se tomó cada individuo y se limpió su raíz sacudiéndola hasta dejarla con la menor cantidad de suelo posible. Entonces las raíces fueron lavadas con 10 ml de buffer PBS (8g NaCl, 0.2g KCl, 1.44g Na2HPO4, 0.24g KH2PO4 ). El lavado de raíces de diluyó en escala logarítmica y se sembraron en medio LB y R2A las diluciones 10 -3 y 10 -5 Las colonias crecidas en esto medios fueron repicadas hasta obtener colonias individuales. Posteriormente, cada colonia aislada se cultivó en medio LB líquido y su actividad antibacteriana fue ensayada. Las bacterias purificadas fueron identificadas por medio de API test kit (BIOMERIEUX. USA) y test gram (tinción y viscosidad) Pruebas antimicrobianas.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
311
Las pruebas antimicrobianas fueron realizadas en microplacas de 96 pocillos agregando 50 µl de sobrenadante libre de células (SLC) sin concentrar a 100 µl del cultivo de bacteria patógena con un OD600 = 0.001. Se mide la absorbancia del cultivo después de 24 h de incubación a 35 °C. Como control se utilizó agua en lugar de SLC. RESULTADOS Y DISCUSIÓN. De las diluciones sembradas en los dos medios de cultivo (LB y R2A) se obtuvieron 4 cepas con actividad antibacteriana. Estas fueron identificadas preliminarmente por medio de API test kit como 4 cepas diferentes con grados de homología para cada una superior al 80% (tabla 1). Una de estas cepas coincide en un 99,9% con Pseudomonas fluorescens, especie anteriormente caracterizada como perteneciente a la rizosfera de D. antárctica por Berrios et al., (2013) y con actividad promotora de crecimiento. En este trabajo se logró determinar que además posee actividad antibiótica contra E. coli, una bacteria gram negativa y contra Staphylococcus aureus, una bacteria gram positiva (figura 1).
Cepa aislada de rizoplano
de D. antarctica
Taxón con mayor similitud
ID (%) Gram
D1 Chryseomonas 86,4 - D2 Pseudomonas 81,9 - D3 Burkhorlderia 99,8 - D4 Chrysomonas/Aeromonas 86,3/ ? -
Tabla 1. Resumen de los resultados de asignación taxonómica a partir del test API. Table 1. Summary of taxonomic assignation based in API test results.
Cabe mencionar que la cepa D3 fue capaz de inhibir el crecimiento de 5 de las 6 bacterias utilizadas para las pruebas de actividad antibiótica. En este mismo sentido 3 de las 4 cepas D1, D3 y D4 mostraron la capacidad de reducir nitratos a nitritos (datos no mostrados) lo que sugiere la participación de estas bacterias en más de una vía para favorecer el desempeño de D. antárctica en ambientes extremos.
Figura 1. Actividad antimicrobiana de los sobrenadantes libres de células (SLC) de bacterias aisladas desde el rizoplano de
D. antárctica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
312
Figure 1. Antimicrobial activity of cell free supernatant (CFS) of bacteria isolated from D. antarctica rhizoplane AGRADECIMIENTOS. Este trabajo contó con el apoyo técnico de Geraldine Asencio, asistente de investigación y profesional del Laboratorio de Biorrecursos Antarticos, INACH. REFERENCIAS Bakker, P. A. H. M., Pieterse C. M. J. & van Loon L. C. 2007. Induced systemic resistance by fluorescent Pseudomonasspp. Phytopathology 97:239–243. Berrios G., Cabrera G., Gidekel M., Gutierrez-Moraga A. 2013. Characterization of a novel antarctic plant growth-promoting bacterial strain and its interaction with Antarctic hair grass (Deschampsia antarctica Desv). Polar Biology 36: 349-362. Edward, J. A., Lewis-Smith, R. I. 1988. Photosynthesis and respiration of Colobanthus quitensis and Deschampsia antarctica from the maritime Antarctic. BAS Bull 81: 43-63. Haas, D. & Keel C. 2003. Regulation of antibiotic production in rootcolonizing Pseudomonas spp. and relevance for biological control of plant disease. Annu. Rev. Phytopathol. 41:117–153. Kucey, R. M. N., Jenzen H. H. & Leggett M. E. 1989. Microbially mediated increases in plant available phosphorus. Adv. Agron. 42:199–228. Lewis-Smith R. I. 1994. Vascular plants as bioindicators of regional warming in Antarctica. Oecologia 99: 322-328. Rodríguez, H. & Fraga R. 1999. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 17:319–339. Xiong F. S., Mueller E. C., Day T. A. 2000. Photosynthetic and respiratory acclimation and growth response of Antartic vascular plants to contrasting temperature regimes. Am. J. Bot. 87: 700-710. Zuñiga G. E., Alberdi M., Corcuera, L. J. 1996. Non-structural carbohydrates in Deschampsia antartica Desv. From South Shetland Islands. Maritime Antarctic. Environ Exp Bot. 36: 393-399

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
313
ESTUDIO DE LA BIOMASA DE MICROORGANISMOS FOTOSINTÉTICOS EN DIFERENTES AMBIENTES EN LAS ISLAS GREENWICH, DEE Y BARRIENTOS,
ANTÁRTIDA
(Study of photosynthetic microorganisms in different environments in Greenwich, Dee and Barrientos Islands, Antarctica).
González, N.1, 2, Silva, S.J.1, González, L.1
1Centro de Oceanología y Estudios Antárticos, Instituto Venezolano de Investigaciones Científicas (IVIC) 1020A, Caracas, Venezuela. Email: [email protected], [email protected]. 2Instituto Antártico Ecuatoriano, Blvd. 9 de Octubre 416 y Chile. Ed. Citibank. Guayaquil, Ecuador. E-mail: [email protected].
INTRODUCCIÓN Los cuerpos de agua dulce de la Antártida son considerados los ecosistemas más productivos en este continente debido a que los microorganismos bénticos forman finos tapetes microbianos ya que reciben contribución alóctona de material orgánico y nutrientes de heces de aves y focas, y los deshielos aportan elementos trazas nutricionales (Laybourn-Parry y Marchant, 1992; Tanabe et al., 2010) contribuyendo así al crecimiento de microorganismos fotosintéticos. La mayoría de los estudios sobre microorganismos fotosintéticos y sus pigmentos en la Antártida se han limitado a ciertas regiones como Victoria Land, Larsemann Hills, valles de Mc Murdo e Isla Rey Jorge (Hodgson et al., 2006; Squier et al., 2002), y poco se conoce de las islas Greenwich, Dee y Barrientos, donde los trabajos que se han realizado están centrados a grupos taxonómicos específicos como las diatomeas. El propósito de este trabajo fue estudiar la abundancia y variación espacial de microorganismos fotosintéticos presentes en aguas, sedimentos y biofilms (tapetes microbianos) de las islas Greenwich, Dee y Barrientos a través del análisis de la clorofila a y feopigmentos y su relación con parámetros fisicoquímicos. METODOLOGÍA (MATERIALES Y MÉTODOS): Sitio de muestreo: Durante el desarrollo de la XVII Expedición Ecuatoriana y VI Expedición Científica de Venezuela al Continente Antártico en el verano austral 2013 se colectaron muestras de agua, sedimento superficial y biofilms en un total de 30 sitios en las Islas Greenwich, Barrientos y Dee, Islas Shetland del Sur (Figura 1).
Figura 1. Área de estudio y sitios de muestreo Figure 1. Study area and sampling sites. Medición de Parámetros Fisicoquímicos: Se midieron in situ los parámetros fisicoquímicos, temperatura, pH,

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
314
oxígeno disuelto (OD), sólidos totales disueltos (TDS), potencial reducción oxígeno (ORP), salinidad y conductividad utilizando la sonda multiparamétrica HORIBA, Modelo UX22. Procesamiento de muestras de agua: Las muestras de agua se procesaron siguiendo la metodología de Borghini et al., 2011. Se midieron las absorbancias a 630, 647, 664, 750, 663,645 y 665 nm sin acidificar y acidificadas con 100 µl 0.1N de HCl para detectar la presencia de feopigmentos de acuerdo al método modificado de Jeffrey & Humphrey, 1975. Las absorbancias fueron estimadas por medio del espectrofótometro UV/VIS Spectrophotometer, Optizen POP y el programa de análisis OptizenView™. Procesamiento de muestras de sedimento y biofilms: Aproximadamente 2 g de sedimento superficial y biofilms se liofilizaron y se extrajeron con 6 ml de 100% acetona grado HPLC y se sometió al mismo procedimiento aplicado a los filtros con el agua (Borghini et al., 2011). Análisis Microscópicos: Se realizaron observaciones microscópicas a través de un Microscopio óptico Leica 2500 para identificar los microorganismos fotosintéticos presentes mediante el sistema de clasificación de Anagnostidis y Komarek (1988) y Komarek y Anagnostidis (1989). Análisis Estadísticos: Se aplicó el análisis no paramétrico de Kruskal-Wallis a través del programa estadístico SPSS versión 19.0, para determinar las diferencias en la distribución espacial de la biomasa. Igualmente se aplicó la Correlación de Spearman para determinar la correlación de la biomasa con los parámetros fisicoquímicos de los ambientes muestreados RESULTADOS La concentración de clorofila a varió entre 0,46 µg l-1 y 129,05 µg g-1 (Tabla 1), siendo el biofilm de la Zona A de Isla Greenwich el sustrato con mayor biomasa (129,05 µg g-1) (p=0,013). Otro ambiente con niveles significativos de clorofila a fue la laguna de deshielo en la Isla Barrientos con una media de 77,52 µg l-1 (p= 0,041), y el sedimento de la zona A con una concentración de 18,67 µg g-1 (p= 0,027).
Sitio Agua Sedimento Biofilm Chl a
(µg l-1) Feofitina (µg l-1)
Chl a (µg g-1)
Feofitina (µg g-1)
Chl a (µg g-1)
Feofitina (µg g-1)
Zona A 3,73±4,1 2,48±5,6 18,67±18,4 9,64±8,3 129,05±118,2 23,51±37,8 Zona B 0,46±0,4 0±0,5 9,19±14,3 3,98±5,4 * * Pta. Ambato 0,62±0,7 0,19±0,5 0,54±0,5 0,42±0,3 * * I. Dee 1,37±1,1 0±3,1 0,68±0,4 0,62±0,4 * * I. Barrientos 77,5±47,7 0,9±11,3 6,96±8,2 4,57±4,4 * *
Tabla 1: Valores medios de las concentraciones de Clorofila a (Chl a) y feofitina a de agua, sedimento y biofilm de los sitios muestreados. * Ausencia de biofilm. Table 1: Mean values of the concentrations of chlorophyll a (Chl a) and water pheophytin a, sediment and biofilm of sampled sites. * Biofilm absence. Los valores más bajos de biomasa de acuerdo al tipo de ambiente, se registraron en la laguna de la Zona B (0,46 µg l-1) sin diferencias significativas y en el sedimento de Punta Ambato (0,54 µg g-1) sin diferencias significativas. La concentración de feofitina estuvo entre 0 y 23,51 µg g-1, este valor más alto se registró en las muestras de Biofilm de la Zona A de I. Greenwich con diferencias altamente significativas (p= 0,003). En esta misma zona, este feopigmento del sedimento mostró también una cantidad relativamente alta (9,64 µg g-1) y el agua de deshielo fue el ambiente que mostró menor concentración de feofitina.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
315
Los tapetes microbianos estuvieron presentes en la Zona A, los cuales están compuestos de una serie de cianobacterias y diatomeas. Los principales constituyentes de estos biofilms fueron cianobacterias filamentosas no heterocísticas del orden Oscillatoriales del género Phormidium, Oscillatoria, Lyngbya y Leptolyngbya. Los valores medios de las variables fisicoquímicas se muestran en la tabla 2. El pH de los cuerpos de agua varió entre 3,9 y 6,5 y la temperatura entre 4,5 y 6,2 °C. La conductividad estuvo en un rango de 135,2 a 426,6 µS cm-1, el oxígeno disuelto registró valores entre 6,8 y 8 mg l-1. La concentración de los sólidos totales disueltos estuvo entre 0,08 y 0,3 g l-1 y el Potencial de óxido-reducción entre 243 y 311 mV. El OD y temperatura tuvieron una correlación altamente significativa con la cantidad de clorofila a de la Zona A (r=0,886, p=0,01). En la zona B, el pH tuvo una correlación significativa (r=0,579, p=0,05). En Punta Ambato, el TDS es el parámetro con una correlación altamente significativa, mientras que en Isla Dee existió una correlación inversa altamente significativa del OD (r=-0,843, p=0,01) y en Barrientos la correlación de la conductividad con clorofila-a fue altamente significativa (r=0,777, p=0,01).
Sitio pH Conductividad (µS cm-1)
OD (mg l-1)
Temperatura (°C) Salinidad (%)
TDS (g l-1)
ORP (mV)
Zona A 5,2 ± 0,6 168,1 ± 88,7 7,8 ± 0,2 5,9 ± 1,8 0 0,1 ± 0,08 299 ± 13,1
Zona B 5,3 ± 0,5 135,2 ± 54,3 6,8 ± 2,1 6,1 ± 1,1 0 0,08 ± 0,04 243 ± 55,7
Pta. Ambato 5,6 ± 0,1 159,7 ± 39,7 8 ± 0,9 4,6 ± 1,1 0 0,1 ± 0,03 243 ± 4,2
I. Dee 3,9 ± 0,5 196,8 ± 22,1 7,7 ± 0,3 4,5 ± 1,8 0 0,1 ± 0,01 311 ± 25,3 I. Barientos 6,5 ± 0,5 426,6 ± 158,7 8 ± 0,1 6,2 ± 0,3 0 0,3 ± 0,1 206 ± 23,6
Tabla 2. Valores medios de los parámetros fisicoquímicos de los cuerpos de agua. Table 2. Values of physico-chemical parameters of the water bodies
DISCUSIÓN Y CONCLUSIONES La mayor concentración de clorofila-a se encontró en los tapetes microbianos de la zona A de Isla Greenwich (129,05 µg g-1) debido a que estos microecosistemas están en un ambiente con luz y los factores que posiblemente contribuyen a su desarrollo son la baja temperatura y ausencia de artrópodos y detrívoros (Howard-Williams et al., 1989; Toro et al., 2007). Valores similares se han registrado en biofilms de lagos de Antártida Este (106 – 320 µg g-1) (Tanabe et al., 2010). Mientras que la concentración en los sedimentos de lagos de Victoria Land fue de 0,017-36,17 µg g-1 (Borghini et al., 2011), rango en el cual se encuentran los sedimentos de este trabajo, con un máximo de 18,67 µg g-1 en la Zona A, lo que pudiera estar relacionado a que el sedimento sirve como sustrato para el crecimiento de comunidades del microfitobentos, representadas principalmente por diatomeas y cianobacterias, con un predominio de los géneros Phormidium, Oscillatoria, Lyngbya y Leptolyngbya. En estudios realizados en cuerpos de agua en la Isla Rey Jorge las concentraciones de clorofila a están en un rango de 0,3 a 158,7 µg l-1 (Vinocur y Unrein, 2000) y en lagos de la Antártida Este de 0,1 a 0,4 µg l-1 (Tanabe et al., 2010), y en el conjunto de islas analizadas en este estudio los valores de clorofila a están dentro de este rango, obteniendo el mayor contenido en la laguna de Isla Barrientos (77,52 µg l-1) donde además se registró una elevada conductividad y sólidos totales disueltos debido a su proximidad al mar y guano de aves marinas. La coloración de esta laguna era de un verde intenso, y las clorofitas fue el grupo algal predominante, mientras que el valor más bajo de clorofila a se registró en la Zona B (0,46 µg l-1). Estos valores bajos de Chl a de los cuerpos de agua concuerdan con reportes anteriores en los que se describe que la mayoría de estos ambientes en la Antártida son oligotróficos (Tanabe et al., 2010).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
316
Los resultados obtenidos de este estudio parecen indicar que estos ecosistemas son importantes para el desarrollo y crecimiento de microfitobentos y tapetes microbianos, por ende están contribuyendo en la cadena trófica de estos ambientes antárticos. AGRADECIMIENTOS A la SENESCYT, Instituto Antártico Ecuatoriano, Programa Antártico Venezolano-COEA. REFERENCIAS Anagnostidis, K. y Komárek J. 1988. Modern approach to the classification system of cyanophytes 3 – Oscillatoriales. Archiv für Hydrobiologie/Algological Studies, 50–53: 327–472. Borghini, F., Colacevich, A., Caruso, T. y Bargagli, R. 2011. An Update on Sedimentary Pigments in Victoria Land Lakes (East Antarctica). Arctic, Antarctic, and Alpine Research, 43 (1): 22–34. Hodgson, D., Verleyen, E., Squier, A., Sabbe, K., Keely, B., Saunders, K. y Vyverman, W. 2006. Interglacial environments of coastal east Antarctica: comparison of MIS 1 (Holocene) and MIS 5e (Last Interglacial) lake-sediment records. Quaternary Science Reviews, 25:179–197. Howard-Williams, C., Pridmore, R., Downes, M. y Vincent, W. 1989. Microbial biomass, photosynthesis and chlorophyll a related pigments in the ponds of the McMurdo Ice Shelf, Antarctica. Antarctic Science, 1 (2): 125-13. Komarek J. y Anagnostidis, K 1989. Modern approach to the classification system of Cyanophytes. 4. Nostocales. Archiv fur Hydrobiologie, (Algological Studies), 56 (suppl. 82): 247-345. Laybourn-Parry, J. y Marchant, H. 1992. The microbial plankton of freshwater lakes in the Vestfold Hills, Antarctica. Polar Biology, 12:405-410. Squier, A., Hodgson,D. y Keely, B. 2002. Sedimentary pigments as markers for environmental change in an Antarctic lake. Organic Geochemistry, 33:1655–1665. Tanabe, S., Ohtani, S., Kasamatsu, N., Fukuchi, M. y Kudoh, S. 2010. Photophysiological responses of phytobenthic communities to the strong light and UV in Antarctic shallow lakes. Polar Biology, 33:85–100. Toro, M., Camacho, A., Rochera, C., Rico, E., Bañón, M., Fernández-Valiente, E., Marco, E., Justel, A., Avendaño, M., Ariosa, Y., Vincent, W. y Quesada, A. 2007. Limnological characteristics of the freshwater ecosystems of Byers Peninsula, Livingston Island, in maritime Antarctica. Polar Biology, 30:635-649. Vinocur, A. y Unrein, F. 2000. Typology of lentic water bodies at Potter Peninsula (King George Island, Antarctica) based on physical-chemical characteristics and phytoplankton communities. Polar Biology, 23:858-870.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
317
ESTRUCTURA Y DINÁMICA ESTACIONAL DEL BACTERIOPLANCTON EN AGUAS ANTÁRTICAS CERCANAS A LA CALETA POTTER, ISLA 25 DE MAYO, ISLAS
SHETLAND DEL SUR
(Structure and seasonal dynamics of the bacterioplankton in Antarctic’s sea water near Potter cove, Isla 25 de Mayo, South Shetlands Islands)
Hernández, E.A1,4, Lopez, J.L3, Piquet, A.M-T2, Buma, A.G.J.2, Mac Cormack, W.P.1,4
1Instituto Antártico Argentino. Departamento de Microbiología Ambiental y Ecofisiología. Balcarce 290, Buenos Aires, Argentina. Email: [email protected] 2University of Gröningen. Ocean Ecosystems, Energy and Sustainability Research Institute, Groningen, the Netherlands 3Universidad de Buenos Aires. Facultad de Farmacia y Bioquímica, Cátedra de Virología. Argentina. 4Universidad de Buenos Aires. Facultad de Farmacia y Bioquímica, Cátedra de Microbiología Industrial y Biotecnología. Argentina. INTRODUCCIÓN Se sabe que las bacterias marinas juegan un papel clave en el ciclo de nutrientes y constituyen un factor importante en el flujo de carbono de los ecosistemas acuáticos (Ducklow, 2000). Las áreas abiertas del mar antártico se caracterizan por su baja biomasa fitoplanctónica y una pobre productividad. Lo mismo ocurre en algunas aguas costeras antárticas, como las de Caleta Potter, que a pesar de tener un alto nivel de nutrientes no evidencian florecimientos apreciables del fitoplancton en los meses de verano (Schloss y Ferreyra, 2002). Bajo esta condición, el bacterioplancton parece ser el principal responsable de la incorporación de los nutrientes en su biomasa, permitiendo que estén disponibles para los niveles tróficos superiores. A pesar del papel relevante del bacterioplancton marino antártico, existe un escaso conocimiento sobre la composición de esta comunidad. En este trabajo se analizó, mediante la construcción de bibliotecas de clones del ADNr 16 S, la estructura del bacterioplancton de Caleta Potter, así como su dinámica interestacional (entre verano e invierno), la cual se encuentra muy influenciada por la profunda oscilación anual de las condiciones ambientales. MÉTODOS Las muestras de agua superficial (10 litros) se tomaron en julio del 2008 y enero del 2009 desde botes Zodiac utilizando botellas Niskin. El sitio de muestreo (E2: 62º 14,011’ S, 58º 41,443’ O) se localizó en la entrada de la Caleta Potter, Isla 25 de Mayo, Shetlands del Sur, Antártida. El agua se filtró a través de membranas de acetato de celulosa (0.22 µm) y los filtros fueron congelados a -80ºC hasta su posterior procesamiento. Luego del aislamiento del ADN genómico, el clonado y secuenciación nucleotídica del ADNr16S (382 secuencias) se realizó en el Departamento de Ecosistemas Oceánicos de la Universidad de Groningen, Países Bajos. Las curvas de rarefacción fueron construidas utilizando el programa Analytical Rarefaction (Holland, 2003). Las secuencias fueron alineadas usando la versión en línea de Clustal Omega-Multiple Sequence Alignment (http://www.ebi.ac.uk/Tools/services/ web_clustalo/). Los alineamientos fueron editados manualmente utilizando el programa BioEdit (Hall, 1999). La reconstrucción filogenética se realizó utilizando Mega 5.1 (Tamura y col., 2011) y el método de Neighbour Joining con un bootstrap de 1000. La comparación de las bibliotecas se realizó utilizando Lib Compare a partir del sitio RDP (Cole y col., 2007). RESULTADOS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
318
La curva de rarefacción de todos los clones estudiados mostró que el número de OTUs (unidades taxonómicas operacionales) esperados y los detectados en la biblioteca son congruentes. Utilizando los agrupamientos observados en el árbol filogenético de la figura 1 se describieron 88 OTUs. Figura 1. Afiliación taxonómica de las 382 secuencias de ADNr 16S.
Figure 1. Taxonomic afiliation of the 382 16 S rDNA sequences.
La estructura a nivel de Phyla de la comunidad del bacterioplancton presente en Caleta Potter puede observarse en la tabla 1. Resalta la alta prevalencia de Proteobacteria y Bacteroidetes como la muy escasa presencia de Cyanobac-teria. Además, un 5,5 % del total de las secuencias no pudieron ser clasificadas dentro de ningún Phylum. Dentro del phylum Proteobacteria se observó la presencia de las clases Gamma-proteobacteria (55,1%), Alphaproteobacteria (37,7 %), y Betaproteobacteria (1,6 %). Además se registró un 5,7 % de Proteobacteria no clasificadas. En la clase Gamaproteobacteria se observó un alto porcentaje (57,35 %) de secuencias no clasificadas, que a su vez se repartieron en tres diferentes grupos (Figura 1). En la clase Alphaproteobacteria un 52,69 % de las secuencias pertenecieron al orden Rhodobacterales. Finalmente dentro del phylum Bacteroidetes se encontró un 91,96 % de Flavobacteria, 4,70 % de Sphingobacteria y 3,53 % de Bacteroidetes no clasificados.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
319
Nº de clones Porcentaje
Phylum Cyanobacteria/Cloroplast 3 0,79%
Phylum Firmicutes 1 0,26% Phylum Actinobacteria 22 5,76% Phylum Planctomycetes 1 0,26% Phylum Bacteroidetes 85 22,25% Phylum Proteobacteria 247 64,66% Bacteria no clasificadas 21 5,50% Secuencias no clasificadas 2 0,52% 382 100,00%
Tabla 1. Estructura de la comunidad del bacterioplancton a nivel de Phyla en Caleta Potter. Table 1. Structure, at Phyla level, of the bacterioplankton community in Potter Cove.
La comparación de la estructura del bacterioplancton de invierno y verano mostró diferencias significativas en la distribución de algunos ribotipos (Tabla 2).
Alphaproteobacteria Gammaproteobacteria Verano Invierno Signific. Verano Invierno Signific. Rhodobacterales 37 12 1.2 10-4 Salinisphaerales 5 0 3.03 10-2 Sphingomonadales 3 15 4.74 10-3 Oceanospirillales 20 9 3.16 10-2
Tabla 2. Componentes de la comunidad del bacterioplancton que mostraron diferencias significativas entre invierno y verano.
Table 2. Components of the bacterioplankton community showing significant differences between winter and summer. DISCUSIÓN Desde el punto de vista metodológico, la curva de rarefacción indicó que las 382 secuencias de ADNr 16S fueron suficientes para describir adecuadamente la estructura de la comunidad. De esta manera, el aumento de la cantidad de secuencias clonadas prácticamente no agregaría nuevos OTUs a la descripción (Gotelli y Colwell, 2001). Recientes estudios en aguas costeras de la península antártica (Straza y col. 2010, Grzymski y col. 2012) muestran, al igual que nuestros resultados, que los grupos dominantes del bacterioplacton son miembros de las clases Gammaproteobacteria y Alphaproteobacteria, seguidos por miembros del phylum Bacteroidetes, aunque estos autores encontraron una baja presencia de Deltaproteobacteria, que no fueron detectadas en nuestro trabajo. En síntesis, la estructura de las comunidades bacterianas aquí descriptas mostró que relativamente pocos ribotipos (Rhodobacterales y Gammaproteobacteria no clasificadas) dominan ambas muestras, pero decenas de subpoblaciones poco abundantes le aportan una gran diversidad phylogenética (Sogin y col., 2006). El elevado número de ribotipos no clasificables según el RDP (Cole y col., 2007), implica un aporte original de secuencias a las bases de datos. Dado que este trabajo es el primero en describir la estructura del bacterioplancton en un sitio cercano a caleta Potter, tales ribotipos no clasificables podrían ser endémicos de Potter o simplemente no haber sido descriptos aun en otros sitios del mundo. El estudio de la dinámica temporal mostró que las Rodobacterias incrementaron su dominancia en el verano respecto al invierno, fenómeno éste previamente observado en aguas del Mar del Norte

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
320
(Pernthaler y col., 2002). Esta observación, sumada a la baja prevalencia de cianobacterias, estaría sugiriendo que las Rodobacterias podrían jugar un rol importante como productores primarios en este ecosistema marino. Además, dado que una proporción importante de secuencias de Rhodobacterales no pudieron ser clasificadas a nivel de género, las mismas podrían estar involucrando múltiples fenotipos con capacidades fisiológicas y ecológicas diversas (Buchan y col., 2005). Como perspectiva, seria pertinente profundizar el estudio del rol ecológico de las Rodobacterias como productores primarios en el ecosistema de caleta Potter. BIBLIOGRAFIA Buchan, A., Gonzalez, J.M., y Moran, M.A. 2005. Overview of the Marine Roseobacter Lineage. Appl. Environ. Microbiol. 71: 5665–5677. Cole, J.R., Chai, B., Farris, R.J., Wang, Q., Kulam-SyedMohideen, A.S., McGarrell, D.M., Bandela, A.M., Cardenas, E., Garrity, G.M., Tiedje, J.M. 2007. The ribosomal database project (RDP-II): introducing myRDP space and quality controlled public data. Nucleic Acids Res., 35: D169–D172. (doi: 10.1093/nar/gkl889). Ducklow, H.W. 2000. Bacterioplankton production and biomass in the oceans. In Microbial ecology of the oceans Kirchman D.L pp. 85–120. Eds. New York, NY:Wiley. Gotelli, N.J. y Colwell, R.K. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 4: 379-391. Grzymski, J.J., Riesenfeld, C.S., Williams, T.J., Dussaq, A.M., Ducklow, H., Erickson, M., Cavicchioli, R. y Murray, A.E. 2012. A metagenomic assessment of winter and summer bacterioplankton from Antarctica Peninsula coastal surface waters. The ISME Journal 6: 1901–1915. Hall, T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 41: 95-98. Holland, S. M. 2003. Analytic rarefaction. See http:// www.uga.edu/strata/software/ Pernthaler, A., Pernthaler, J., Schattenhofer, M., y Amann, R. 2002. Identification of DNA-synthesizing bacterial cells in coastal North Sea plankton. Appl. Environ. Microbiol. 68: 5728–5736. Schloss, I.R. y Ferreyra, G.A., 2002. Primary production, light and vertical mixing in Potter Cove, a shallow bay in the maritime Antarctic. Polar Biology, 25: 51-48. Sogin, M.L., Morrison, H.G., Huber, J.A., Welch, D.M., Huse, S.M., Neal, P.R., Arrieta, J.M., y Herndl, G.J. 2006. Microbial diversity in the deep sea and the underexplored “rare biosphere”. PNAS, 103: 12115–12120. Straza, T.R., Ducklow, H.W., Murray, A.E., Kirchman, D.L. 2010. Abundance and single-cell activity of bacterial groups in Antarctic coastal waters. Limnol Oceanogr 55: 2526–2536. Tamura, K. et al. 2011. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods, Mol Biol Evol, 28: 2731-2739.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
321
HONGOS FILAMENTOSOS DE LA ANTÁRTIDA Y SU POTENCIAL EN LA PRODUCCIÓN DE METABOLITOS BIOACTIVOS
(Antarctic fungi and their potential in the production of bioactive metabolites)
Jorcín, G.1, González, A. 1, Vero, S. 1, Garmendia, G. 1 y Alborés, S. 1
1Cátedra de Microbiología, Departamento de Biociencias, Facultad de Química, Universidad de la República, Montevideo, Uruguay E-mail: [email protected] INTRODUCCIÓN Entre las numerosas aplicaciones de los hongos filamentosos en la biotecnología tradicional y moderna se encuentra la producción de metabolitos bioactivos. En particular es muy promisoria la búsqueda de microorganismos provenientes de ambientes no explorados que sean capaces de producir nuevos agentes antimicrobianos para su aplicación en medicina y agricultura (Gloer, 2007; Montermartini et al., 2000). Aunque se han descrito nuevos grupos microbianos provenientes de la Antártida, éstos representan un muy bajo porcentaje de la gran diversidad microbiana presente en esta región. Si bien se han llevado a cabo un gran número de estudios sobre microorganismos procariotas, el número de investigaciones realizadas con eucariotas, en particular hongos, ha sido mucho menor (Cantrell et al., 2011). En este sentido es interesante la identificación de hongos filamentosos provenientes de muestras de esta región para estudios de biodiversidad así como de diversas aplicaciones. La taxonomía de hongos ha estado soportada en técnicas convencionales, basadas en la descripción morfológica y fisiológica de especies y géneros, pero éstas dependen de las condiciones de cultivo de las cepas, por lo que han introducido errores en la ubicación taxonómica. Estas dificultades se han solucionado con la aplicación de las técnicas moleculares, basadas en el análisis de fragmentos de las moléculas de ácidos nucleicos (Orberá Ratón, 2004). La conservación de los hongos filamentosos varía en dependencia del tipo y el grado de esporulación y existen muchos factores que pueden afectar la viabilidad y estabilidad de los cultivos por lo tanto es necesario el mantenimiento y conservación de cultivos fúngicos para garantizar las características originales de las cepas con su estabilidad genética y fenotípica establecidas (Gato Cárdenas, 2010). En el presente trabajo se realizaron aislamientos de hongos filamentosos a partir de muestras colectadas en la Antártida. Se trabajó en la identificación, conservación y evaluación de actividad antimicrobiana de estos aislamientos. METODOLOGÍA Las muestras de suelo y agua (continental y oceánica) de diversos lugares de la isla Rey Jorge, colectadas en 2012 y 2013, se sembraron en placas de PDA con cloranfenicol (a diferentes diluciones) y en algunos casos en caldo de enriquecimiento Sabouraud Dextrosa y posterior aislamiento en PDA. A partir de las colonias aisladas de hongos filamentosos se realizaron cultivos monospóricos y se obtuvieron 26 aislamientos. Identificación Las cepas se cultivaron en medio YES a 25ºC durante 5 días. Luego del tiempo de incubación se separó el micelio y se realizó la extracción de ADN. Se amplificó la región ITS1-5.8SrADN-ITS2. Los amplicones se secuenciaron y las secuencias se compararon con la base de datos del GenBank.. Métodos de conservación

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
322
Tubos de PDA: Los hongos se sembraron en tubos de Potato Dextrosa Agar (PDA) y se incubaron a temperatura ambiente hasta observar crecimiento de micelio. Luego del tiempo de incubación se conservaron en la heladera y a temperatura ambiente con vaselina. Agua estéril: Los hongos se sembraron en tubos conteniendo PDA inclinado. Se incubaron a temperatura ambiente hasta observar crecimiento de micelio y esporulación. A partir de dichos tubos se hizo una suspensión de esporas en tubos con agua estéril. Los tubos se conservaron a temperatura ambiente. Papel de filtro: Cada cepa se sembró en forma puntual, en el centro de una placa de PDA. Alrededor de la siembra, se colocaron trozos pequeños de papel de filtro estéril. Se incubó a temperatura ambiente hasta observar crecimiento de micelio. Los trozos de papel se retiraron con pinza y se guardaron en sobres de papel estériles. Los sobres se conservaron a temperatura ambiente. Para todos los métodos de conservación se estudió la viabilidad de los hongos filamentosos resembrando periódicamente cada cepa en PDA en placa. Actividad antimicrobiana frente a patógenos Se realizaron cultivos duales para detectar inhibición del crecimiento producida por los hongos aislados de la Antártida frente a microorganismos representativos de distintos grupos: bacterias Gram-positivas y Gram-negativas, levaduras y hongos filamentosos. Las pruebas de enfrentamiento con hongos patógenos se realizaron en placas de PDA sembrando en un extremo un disco de agar con micelio del patógeno y en el extremo opuesto otro disco con micelio del hongo de la Antártida a 5 cm aproximadamente entre ellos. Además se sembró como control en otra placa un disco de agar con micelio del hongo patógeno. Todas las placas se incubaron a temperatura ambiente. Se evaluó el porcentaje de inhibición de crecimiento radial (PICR), empleando la fórmula PICR = (R1 – R2)/R1 x 100, donde R1 es el radio del patógeno testigo y R2 es el radio del patógeno en el cultivo dual. Los ensayos con bacterias patógenas se realizaron en placas de TSA, mediante hisopado de una suspensión correspondiente a McFarland 1 de las bacterias patógenas y la siembra de un disco de micelio del hongo de la Antártida en el centro de la placa. La inhibición del crecimiento o lisis producida por el antagonista se observa como un halo alrededor del crecimiento del hongo. RESULTADOS Mediante la amplificación de la región ITS y la secuenciación se logró la identificación molecular de los aislamientos obtenidos del muestreo 2012, complementando dicha identificación con características fenotípicas (observación macro y microscópica). Se han identificado hongos pertenecientes a las especies Aspergillus tubingensis, Aspergillus flavus, Penicillium expansum y al género Geomyces. Para todos los métodos de conservación evaluados durante 12 meses se obtuvo la recuperación de las cepas. En los cultivos duales se observó que varias cepas de hongos filamentosos produjeron inhibición del crecimiento de diferentes microorganismos. Dentro de las bacterias evaluadas se observaron halos de inhibición del crecimiento en los cultivos de Escherichia coli y Staphylococcus aureus, A su vez se observó inhibición del crecimiento de los hongos Aspergillus niger, Fusarium graminearum, Alternaria alternata, Botrytis cinerea, Penicillium expansum y Rhizopus stolonifer. DISCUSIÓN Y CONCLUSIONES Los resultados obtenidos de identificación y conservación de las cepas aisladas a partir del primer muestreo son un gran aporte para la realización de trabajos ya en marcha como ser la búsqueda de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
323
nuevas fuentes de metabolitos bioactivos, así como para el desarrollo de futuros proyectos que exploren nuevas aplicaciones biotecnológicas. Es por ello que estos estudios se extenderán a los aislamientos obtenidos de las muestras colectadas en 2013. AGRADECIMIENTOS Instituto Antártico Uruguayo, PEDECIBA Química, ANII. REFERENCIAS Cantrell, S.A. 2011. Unusual fungal niches. Mycologia. (6): 1161-1174. Gato Cardenas, Y. 2010. Métodos de conservación y formulación de Trichoderma harzianum rifai. Fitosanidad (14): 189-195. Gloer, J.B. (2007). Applications of fungal ecology in the search for new bioactive natural products. In The Mycota vol IV, 2nd edn. Springer-Verlag Berlin Heidelberg. 257–275. Montermartini, A., Liotta , M., Venturi, C. y Calegari, L. 2000. Antibacterial activity of Penicillium spp. strains isolated in extreme environments. Polar Biology. (4): 294-296. Orberá Ratón, T. (2004). Métodos moleculares de identificación de levaduras de interés biotecnológico. Revista Iberoamericana de Micología. (21): 15-19.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
324
DIVERSIDAD DE LAS COMUNIDADES DE ACTINOBACTERIAS EN ECOSISTEMAS ANTÁRTICOS
(Actinobacteria community diversity in Antarctic ecosystems)
Lagos, L.1, 3, Cid, F.1, 3, Marileo, L.1, 3, Jorquera, M.1, 3, Salazar, L.2, 3, Barrientos, L.2, 3 1 Centro de Interacción Suelo, Planta y Biotecnología de Recursos Naturales. 2 Centro de Biología Molecular y Farmacogenética. 3 Scientific and Technological Bioresources Nucleus Universidad de La Frontera, Temuco, Chile. E-mail: [email protected] INTRODUCCIÓN La Antártica ofrece oportunidades únicas para la investigación de la diversidad microbiana y podría representar ventajas para el descubrimiento de microorganismos biotecnológicamente atractivos, dado que es uno de los entornos naturales menos intervenidos (O'Brien et al., 2004, Nichols et al., 1999). Dentro de los procariontes, las Actinobacterias colonizan una gran variedad de nichos, además, es uno de los grupos bacterianos con mayor interés comercial debido a la producción de compuestos metabólicos tales como antibióticos, vitaminas, alcaloides, factores de crecimiento, enzimas, entre otros. Estudiar la comunidad microbiana de Actinobacterias en ecosistemas antárticos puede ser relevante para el aislamiento, caracterización e identificación de ellos. Los métodos independientes de cultivo, tales como el PCR-DGGE (Denaturing Gradient Gel Electrophoresis) han sido usados ampliamente para conocer la diversidad y la función ecológica de las poblaciones microbianas en diferentes ecosistemas. El objetivo principal de nuestro estudio fue estudiar la diversidad de Actinobacterias en ecosistemas antárticos, a partir de muestras de suelo y sedimento marino mediante el empleo de PCR-DGGE. MATERIALES Y MÉTODOS Muestreo de sedimento y de suelo Las muestras de suelos y sedimentos marinos fueron colectadas en distintos sitios de las Islas Shetland del Sur, Península Antártica. Los sitios muestreados fueron Glaciar Collins (suelo 5 cm profundidad), Isla Decepción (sedimento marino, tomado a 4 m de profundidad) y Bahía Fildes (suelo 5 cm profundidad y sedimento marino tomado a 10 m de profundidad), durante la Expedición Científica Antártica 2012-2013 (ECA 49). Análisis de las comunidades de Actinobacterias La composición de las comunidades de Actinobacterias de suelo y sedimento marino fueron evaluadas a través de PCR-DGGE. El DNA fue extraído con Ultraclean Soil DNA Isolation Kit (Mo-Bio Laboratories) de acuerdo a las instrucciones del fabricante. Los genes rARN 16S fueron amplificados a través de PCR anidado con los partidores F27 (5’AGAGTTTGATC(A/C)TGGCTCAG3’), R1492 (5’CGCCCGGGGCGCGCCCC GGGCGGGG CGG GGG CACGGGGG 3’), F984GC (CGC CCG GGG CGC GCC CCG GGC GGG GCG GGG GCA CGG GGG GAA CGC GAA GAA CCT TAC) y R1378 (CGG TGT GTA CAA GGC CCG GGA ACG) (Heuer et al., 1997, Das et al., 2007). Las reacciones de PCR se llevaron a cabo con los reactivos GoTaq Flexi DNA Polymerase (Promega Co.). Para la reacción de amplificación del gen ARNr 16S a partir del ADN extraído de las muestras de suelo y sedimento se utilizaron los partidores universales F27 y R1492. La amplificación del gen ARNr 16S consistió en una desnaturalización inicial de 5 min a 94°C, seguido de 25 ciclos de amplificación (desnaturalización a 94°C por 1 min, alineamiento a 52°C por 1 min y extensión a 72°C por 1 min), seguido por una extensión final a 72°C por 2 min. Para la reacción de amplificación del gen V6 se

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
325
utilizaron los partidores F984GC y R1378. La amplificación de la región variable V6 consistió en una desnaturalización inicial de 5 min a 94°C, seguido de 20 ciclos de amplificación (desnaturalización a 95°C por 1 min, alineamiento a 60°C por 1 min y extensión a 72°C por 1 min), seguido por una extensión final a 72°C por 10 min. Los productos de PCR de la región V6 del gen ARNr 16S se separaron en un gel de poliacrilamida al 6%, con un gradiente denaturante de 40 - 70%. La electroforesis se realizó durante 12 h a 100V en tampón TAE 1X, en un DCode System (Bio-Rad Laboratories, Inc.). El gel fue teñido con SYBR Gold (Molecular Probes, Invitrogen Co.) durante 20 min y fotografiado sobre un Transiluminador UV. El análisis del patrón de bandas de DGGE se realizó mediante el empleo del software de análisis de imagen Phoretix 1D software (TotalLab Ltd.). Basados en los datos entregados por Phoretix 1D, los cambios en la presencia o ausencia de las grupos bacterianos fueron analizados con escalamiento multivariado no métrico (MDS) usando PAST freeware (http://folk.uio.no/ ohammer/past/), índice de similaridad de Bray–Curtis y Shannon–Wiener (Yang et al., 2003). RESULTADOS Y DISCUSIÓN Con el ADN genómico obtenido de los suelos muestreados en el Glaciar Collins y Bahía Fildes y de sedimentos marinos muestreado en Isla Decepción y Bahía Fildes, se logró amplificar el gen ARNr 16S de las comunidades de Actinobacterias en estos suelos y sedimentos. Luego, estos productos de PCR se utilizaron como templado para la amplificación de la región variable V6 del gen ARNr 16S, obteniéndose amplicones de~ 473 pb (Fig. 1). Los perfiles de DGGE analizados mediante un análisis de escalamiento multivariado no métrico (MSD) (Fig.2A), muestran que la estructura de las comunidades de Actinobacterias no presenta similitudes, lo que también se observa en el análisis del dendograma de UPGMA, que muestra que las comunidades de Actinobacterias de suelo (Glaciar Collins y Bahía Fildes) no presentan una similitud en su estructura. Lo mismo ocurre con las comunidades de Actinobacterias de sedimento marino de los sitios de Isla Decepción y Bahía Fildes. Al analizar la diversidad en base a los índices de diversidad de Shannon (H´) de las comunidades de Actinobacterias de suelos y de sedimentos marinos, y determinar la riqueza y el índice de Shannon en base al análisis de los perfiles de DGGE de los genes ARNr 16S de las comunidades bacterianas, observamos que los valores de Shannon–Wiener indican que la diversidad de Actinobacterias es más alta (pero no estadísticamente significativa ≤ 0,05) en las muestras de suelos del Glaciar Collins y Bahía Fildes (1,04 y 1,0, respectivamente) comparado con las muestras de sedimentos marinos de las Islas Decepción y Bahía Fildes (0,76 y 0,77, respectivamente).
Figura 1. Detección de la región variable V6 del gen ARNr 16S del ADN bacteriano de los suelos y sedimentos marinos.
Electroforesis en gel de agarosa (1%) de productos de PCR de región V6. Carril: MM, marcador de peso molecular 100 pb; CN, control negativo; 1-3, suelo Glaciar Collins; 4-6 sedimento Bahía Fildes; 7-9, suelo Bahía Fildes; 10-12, sedimento Isla
Decepción. Figure 1. Detection of V6 variable region of the 16S rRNA bacterial gene from soils and marine sediments. PCR products in
agarose gel electrophoresis (1%) of V6 region. Lane: MM, molecular weight marker 100 bp, CN, negative control; 1-3 Collins Glacier soil; 4-6, Bahía Fildes marine sediment; 7-9, Bahía Fildes soil; 10-12, Deception Island marine sediment.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
326
Figura 2. A. Análisis de escalamiento multivariado no métrico (MSD) y B. Dendograma de UPGMA de perfiles DGGE del gen ARNr 16S de las comunidades bacterianas de los suelos de Bahía Fildes (HSo) y Glaciar Collins (CSo) y sedimentos
marinos de Isla Decepción (DSe) y Bahía Fildes (HSe). Figure 2. A. Non-metric multidimensional scaling (MDS) analysis and B. Dendrogram of DGGE profiles 16S rRNA gene from bacterial community of Bahía Fildes (HSO) and Glacier Collins (CSo) soils and Deception Island (DSe) and Bahía
Fildes (HSe) marine sediments. CONCLUSIONES Basado en el análisis de PCR-DGGE nuestro estudio evidenció que la estructura de las comunidades de Actinobacterias tanto de suelo como sedimento marino no presentó similitud entre los sitios muestreados. Además, a través del índice de diversidad de Shanon-Wiener se evidenció que las comunidades de Actinobacterias en suelos es más diversa comparada con las comunidades de Actinobacterias de sedimentos marinos. La distribución y los roles que cumplen los microorganismos en los diferentes ambientes debe ser estudiada con una combinación de técnicas convencionales y aproximaciones modernas para ser mejor entendida. AGRADECIMIENTOS Proyecto INACH RT_14-12 REFERENCIAS Das, M., Royer, T.V. Y Leff, L.G. 2007. Diversity of Fungi, Bacteria, and Actinomycetes on leaves decomposing on a stream. Applied and Environmental Microbiology, 73: 756-767. Heuer, H., Krsek, M., Baker, P., Smalla, K., y Wellington, E. M. H.1997. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Applied and Environmental Microbiology, 63:3233–3241. Nichols, D., Bowman, J., Sanderson, K., Nichols, C.M., Lewis, T., Mcmeekin, T. y Nichols, P.D. 1999. Development with Antarctic microorganisms: culture collections, bioactivity screening, taxonomy, PUFA production and cold-adapted enzymes. Current Opinion in Biotechnology, 10: 240-246. O’Brien, A., Sharp, R., Russell, N. y Roller, S. 2004. Antarctic bacteria inhibit growth of food-borne microorganisms at low temperatures. FEMS Microbiology Ecology, 48: 157-167. Yang, Y.J., Dungan, R., Ibekwe, A.M., Valenzuela-Solano, C., Crohn, D. y Crowley, D. 2003. Effect of organic mulches on soil bacterial communities one year after application. Biology and Fertility of Soils, 38:273–281
A B

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
327
ACTIVIDAD ANTIMICROBIANA DE COMPUESTOS PROTEICOS Y NO PROTEICOS PROVENIENTES DE DOS CEPAS DE STREPTOMYCES SP. AISLADA DESDE ANTÁRTICA
(Antimicrobial activity of proteinaceous and nonproteinaceous compounds from two strains of Streptomyces sp. isolated from Antarctic)
Lavín, P1., Gallardo-Cerda, J1., Estrada, C2
1Laboratorio de Biorrecursos Antárticos, Instituto Antártico Chileno, Punta Arenas, Chile. E-mail: [email protected] 2Universidad Andrés Bello, Viña del Mar
INTRODUCCIÓN Los Actinomicetos, especialmente las especies del genero Streptomyces, son prolíficos productores de moléculas bioactivas como enzimas, antitumorales, elicitores de crecimiento y bacteriocinas, compuestos con actividad antibiótica, entre otros. Las bacteriocinas (péptidos antimicrobianos producidos por bacterias que inhiben otras bacterias estrechamente relacionadas) han sido los únicos antimicrobianos utilizados en alimentos (Nettles y Barefoot 1993). En general, estos péptidos son estables al calor, además son hipoalergénicos y rápidamente degradados por enzimas proteolíticas en el tracto intestinal humano. El uso de compuestos bioactivos como las bacteriocinas para la conservación de alimentos representa una alternativa para eliminar los peligros a la salud asociados a bacterias patógenas, o al deterioro de los alimentos permitiendo asi extender la vida útil de estos (Diop et al., 2007). Las bacterias antárticas psicrotróficas y psicrofilicas pueden ser una fuente potencial de nuevos compuestos debido a la existencia de nuevas vías metabólicas, como resultado de la selección evolutiva para sobrevivir y crecer en las condiciones extremas de ese ambiente (Van den Burg 2003). El presente trabajo se centró en la caracterización de compuestos antimicrobianos aislados a partir de dos cepas antárticas de Streptomyces sp. MATERIAL Y MÉTODOS A) Aislamiento, cultivo y selección de cepas: Las cepas bacteriana fueron aisladas desde 11 muestras de suelo de la Península Antárctica utilizando medio específico para Actinomycetes (Difco). La selección de cepas se realizó en función de su capacidad antagónica frente a Staphylococcus aureus y/o Escherichia coli, utilizando el test de doble gel con agar molten sobre 4 medios de cultivos (Actinomicete, LB, R2A y medio Avena). Posteriormente las cepas de Streptomycete seleccionadas fueron crecidas en frascos de 500ml utilizando medio avena durante 7 días a temperatura ambiente, con la finalidad de realizar la extracción de metabolitos del sobrenadante libre de células (SLC). B) Extracción de Metabolitos: Las células de los cultivos fueron separadas por centrifugación y los compuestos antagónicos separados del SLC por el método de extracción con solventes, cromatografía y separación por gel de poliacrilamida (SDS-PAGE). La extracción por solvente se realizó con acetato de etilo adicionado al sobrenadante en proporción de 1:1 (V/V) y agitado vigorsamente por una hora. C) Estabilidad de los compuestos: se evaluó la estabilidad de la actividad del SLC y de las fracciones activas después de ser sometidos a un rango de temperaturas entre 20-95 °C durante 15 minutos. RESULTADOS Y DISCUSIÓN Se recuperaron 30 cepas con actividad, caracterizadas parcialmente, que pertenecen al género

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
328
Streptomyces, según morfología, y que poseen actividad antimicrobiana contra bacterias asociadas infecciones alimentarias como Staphylococcus aureus, Escherichia coli y Listeria monocytogenes. De estas dos cepas, denominadas como cepa 3013 y 22E, el uso directo de sus sobrenadantes libres de células resultaron en interesante actividad antimicrobiana contra bacterias multiresistentes a los antibióticos (Figura 1). El proceso de caracterización del compuesto producido por la cepa 3013, utilizando métodos de extracción con solventes, cromatografía y separación por gel de poliacrilamida (SDS-PAGE), permitió obtener dos compuestos semi-purificados apolares, siendo uno de origen peptídico y otro no peptídico. Mientras que el resultado obtenido para la cepa 22E, utilizando la misma metodología, resulto en la obtención de un solo compuesto semi-purificado con carácter polar. Todas las moléculas obtenidas presentaron estabilidad a 95 °C por 15 minutos lo que podría permitir el uso de estos compuestos en empaques de alimentos una vez que se compruebe la ausencia de citotoxicidad de los mismos.
Figura 1. Ensayo de actividad antimicrobiana contra cepas multirresistentes a partir del SLC.
Figure 1: Antimicrobial assay using supernatant from cell-free culture against multidrug resistant strains.
AGRADECIMIENTOS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
329
Proyecto de inserción de capital humano Conicyt 78111301 REFERENCIAS Diop M.B., Dubois-Dauphin R., Tine R., Ngom A., Destain J. y Thonart P. (2007) Bacteriocin producers from traditional food products Biotechnol. Agron Soc Environ 11 (4): 275–281. Nettles, C. G. y Barefoot, S. F. (1993) Biochemical and genetic characteristics of bacteriocins of food-associated lactic acid bacteria. J. Food Prot. 56, 338-356. Van den Burg B. (2003) Extremophiles as a source for novel enzymes. Curr. Opin. Microbiol. 6: 213–218.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
330
CEPA ANTÁRTICA DE BACILLUS SP. CON ACTIVIDAD EXTRACELULAR DE TIPO AGAROLÍTICA Y ALGINATOLIASA.
(Antarctic strain of Bacillus sp. with extracellular agarolitic and alginate-lyase activities)
Lavín, P.1, Gallardo-Cerda, J.1, Torres-Díaz , C2 y Gonzalez, M1.
1Laboratorio de Biorrecursos Antárticos, Instituto Antártico Chileno, Punta Arenas, Chile. [email protected]
2Laboratorio de Genómica y Biodiversidad (LGB), Departamento de Ciencias Básicas, Facultad de Ciencias, Universidad del Bío-Bío, Chillán, Chile.
INTRODUCCIÓN Alginato-liasa y Agarasa son enzimas claves que permiten a ciertas bacterias degradar los azucares estructurales de algas pardas y rojas. Estas enzimas son biocatalizadores interesantes debido a su aplicación en la sacarificación del alginato y agarosa, liberando azucares menos complejos, para una variedad de aplicaciones como la producción de biocombustibles y de compuestos bioquímicos con aplicaciones en el área de salud, cosmética y agricultura. (Giordano, et al. 2006, Kobayashi, et al. 1997, Sulfahri, et al 2011, Wu, et al 2005). Las bacterias de la Antártica, poseen características fisiológicas adaptativas que habrían evolucionado para permitir su supervivencia y funcionamiento dentro de las condiciones adversas de ese ecosistema. Por lo tanto, bacterias aisladas desde algas antárticas podrían tener la capacidad de degradar enzimáticamente azúcares complejos a temperaturas inferiores que aquellas aisladas desde zonas más cálidas, lo que potencialmente podría tener aplicaciones en la mejora de procesos industriales que utilizan enzimas. El objetivo de este estudio fue describir la capacidad de metabolizar agarosa y alginato por una nueva bacteria, Bacillus sp. INACH001, aislada desde algas varadas en la Isla Rey Jorge (Antártica). MATERIAL Y MÉTODOS La cepa bacteriana fue aislada desde algas varadas en la Isla Rey Jorge utilizando un medio de cultivo mínimo R2A sólido, cuya principal fuente de carbono es el agar. Para la identificación de la bacteria se amplificó y sequenció el gen ribosomal 16S. La asignación taxonómica fue realizada mediante la herramienta de libre acceso BLAST (Basic Local Alignment Search Tool) disponible en el sitio web http://blast.ncbi.nlm.nih.gov/Blast.cgi. El análisis filogenético se realizó utilizando el software Bosque (Ramírez-Flandes y Ulloa, 2008) utilizando secuencias del gen ribosomal 16S disponibles en la base de datos Genebank (http://www.ncbi.nlm.nih.gov/genbank/) mediante el software MUSCLE 3,6 (Edgar, 2004). El árbol filogenético de concenso se obtuvo mediante el método de máxima verosimilitud (maximum likelihood) sobre la base del modelo HKY85 (Hasegawa et al., 1985), utilizando inferencia filogenética basada en PhyML (Guindon y Gascuel, 2003) y mediante el algoritmo de agrupamiento de neighbor-joining con el programa Phylip NJ (Felsenstein, 2005). La evaluación estadística de las topologías de los árboles se realizó mediante análisis de bootstrap con 100 repeticiones. La obtención de moléculas activas desde el sobrenadante libre de células se realizó a partir de cultivos incubados por dos semanas a 20°C, en medio mínimo R2A conteniendo 0,15% de agar. Las células fueron separadas del cultivo mediante centrifugación. Posteriormente, el sobrenadante fue filtrado (filtro con un tamaño de poro de 0,22 µm) y liofilizado. El extracto liofilizado, fue resuspendido en tampón fosfato salino (PBS) obteniendo una solución concentrada 40 veces (40x) en relación a la concentración presente en el cultivo original. La actividad catabólica (capacidad de producir azúcares reductores) fue estimada indirectamente a través de la producción azúcares reductores mediante el método de Miller (1959) utilizando como substrato soluciones de agarosa o alginato al 1% (p/v). La reacción se realizó con tres

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
331
réplicas independientes agregando 50 µl del extracto concentrado 40X a 500 µl de substrato y incubanado a tres temperaturas distintas (4°, 10° C y 30° C) durante 30, 120 y 240 min. Los valores de concentración de azúcares reductores, -expresada como mg de Galactosa por ml- fueron estimados a partir de una curva estándar realizada con galactosa. Para evaluar el efecto de la Temperatura sobre la actividad enzimatica se compararon las pendientes de las rectas de regresión lineal mediante un ANOVA de una vía, utilizando la prueba de Bonferroni para evaluar diferencias a posteriori. Los análisis de regresión lineal y el ANOVA fueron realizados por medio del programa estadístico GraphPad Prism version 4.0 para windows, GraphPad Software, San Diego California USA, www.graphpazd.com. RESULTADOS Y DISCUSIÓN El análisis del gen ribosomal 16S indicó que la cepa bacteriana aislada desde algas varadas en la Isla Rey Jorge (Antártica), pertenece al género Bacillus. La cepa descrita en este trabajo es capaz de producir azúcares reductores a partir de agarosa a 4 °C (Fig. 1a). Esta capacidad de degradar agarosa a azúcares más simples ha sido previamente descrita para diversas especies del género Bacillus (e.g., Bacillus megaterium, Bacillus cereus y Bacillus subtilis) aisladas desde sedimento o algas rojas de zonas tropicales (Kim et al., 1999; Khambhaty et al., 2008; Saraswathi et al., 2011). No obstante, ninguna de ellas presenta un máximo de actividad a la temperatura encontrada para la cepa de Bacillus reportada en este estudio. Adicionalmente, es importante mencionar que en ninguna cepa de Bacillus, (sean estas provenientes de la Antártica u otros continentes) descritas anteriormente tienen la capacidad conjunta de degradar agar y alginato (Fig.1). En la mayoría de los estudios, la actividad agarasa se observa cuando los microorganismos son crecidos en un medio con baja concentración de nutrientes y agar como fuente mayoritaria de carbono, fenómeno observado también en la cepa INACH-0001 y que sustenta la hipótesis que plantea que las enzimas agarolíticas, en todos los microorganismos estudiados, están bajo el control de un regulador común (Shin et al., 2010). Un fenómeno similar ocurre con la expresión de la alginatoliasa, la cual puede ser inducida cuando son cultivadas en presencia de alginato, como en Alginovibrio aquatilis (Stevens y Levin, 1977) y otras bacterias marinas como algunas asociadas con Sargassum (Romeo y Preston, 1986; Brown y Preston, 1991). Sin embargo, en algunas especies, como por ejemplo Pseudomonas alginovora, la producción de alginatoliasa es constitutiva (Boyen et al., 1990). Aun cuando la cepa aislada fue crecida en un medio donde la única fuente de carbono correspondía a agar, el sobrenadante libre de células fue capaz de hidrolizar alginato, sugiriendo una posible expresión constitutiva de enzimas degradadoras de otros ficocoloides. Las enzimas de microorganismos marinos, especialmente aquellas obtenidas de microorganismos de ambientes extremos, se han hecho cada vez más importantes por el potencial de sus aplicaciones. Es así como, la capacidad de degradar azúcares complejos mediante enzimas específicas se ha reportado como una característica presente en algunas bacterias epifitas de algas (Hosoda et al., 2003; Fu y Kim, 2010; Kam et al., 2011; Li et al., 2011). En la literatura se pueden encontrar muchos ejemplos como la bacteria marina Saccharophagfus degradans que fue aislada desde Spartina alterniflora en descomposición encontrada en zonas estuarinas (Ekborg et al., 2005). Este microorganismo tiene la capacidad de descomponer el material vegetal completamente debido a un sistema multienzimático que le permite degradar al menos 10 diferentes polisacáridos, entre ellos agar, a sus azúcares elementales o derivados (Shin et al., 2009). La mayoría de las enzimas alginatoliasa aisladas desde microorganismos presentan su actividad máxima a temperaturas que van desde 25 ° C a 50 ° C (Wong et al., 2000). Por otro lado, la mayor parte de las enzimas aisladas y caracterizadas como agarolíticas, tienen un máximo de actividad a temperaturas superiores a 30°C (Hosoda et al., 2003; Fu y Kim, 2010; Kam et al., 2011; Li et al., 2011). Pseudoalteromonas antarctica, una especie marina con actividad agarolítica, fue descrita por Vera et al. (1998) para algas en descomposición presentes en las costas de Valdivia. Estos autores

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
332
encontraron que las colonias de esta especie tienen una alta capacidad agarolítica, siendo capaces de degradar agar 48 h después de comenzado el cultivo. La capacidad agarolítica de P. antarctica resultó ser mayor a la encontrada por Hosoda et al. (2003) en Paenibacillus spp. la cual comienza a degradar agar después de 10 días comenzado el cultivo. En este caso, ambas bacterias presentan la máxima actividad agarolítica a temperaturas que superan los 20 °C. Esta información, además de poseer valor biotecnológico, podría ser extrapolada en un contexto ecológico permitiendo un mayor entendimiento de los procesos de reciclaje del carbono de la biomasa algal varada en la Antártica. Agradecimientos: Proyecto de inserción de capital humano Conicyt 78111301 y Corfo 11IDL2-10651.
Figura 1: Producción de azúcares reductoras de extractos de Bacillus sp. (medida como producción de Galactosa) a través del tiempo a tres temperaturas (4, 10 y 30 ºC) en dos diferentes substratos: Agarosa (a) y Alginato (c). Efecto de la temperatura en la pendiente de producción de Galactosa a través del tiempo en dos diferentes substratos: Agarosa (b) y Alginato (d). Los
valores mostrados corresponden a las medias ± 1 DE (a y c) y ± 2 DE (b y d). Las diferentes letras griegas (, ß) indican diferencias significativas (P < 0.05).
Figure 1: Reducing sugar production of Bacillus sp.(measured as Galactosa production) through time across three temperatures (4, 10 and 30 ºC) in two different substrates: Agarose (a) and Alginate (c). Effect of temperature on the slope of Galactose production through time in two different substrates: Agarose (b) and Alginate (d). The values are averages ± 1 SD
(a and c) and ± 1 SD (b and d). Different Greek letters (, ß) indicate significant differences (P < 0.05). REFERENCIAS Boyen C., Bertheau Y., Barbeyron T. y Kloareg B. 1990. Preparation of guluronate lyase from Pseudomonas alginovora for protoplast isolation in Laminaria. Enzyme and Microbial Technology 12: 885-90 Brown B.J. y Preston JFIII. 1991. L-Guluronan-specific alginate lyase from a marine bacterium associated with Sargassum. Carbohydrate Research 211: 91-102 Edgar R.C. 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research 32: 1792-1797. Ekborg, N., Gonzalez, J., Howard, M.; Taylor, E., Hutcheson, S. y Weiner, R. 2005. Saccharophagus degradans gen nov., sp. nov., a versatile marine degrader of complex polysaccharides. International Journal of Systematic and Evolutionary Microbiology 55: 1545-1549. Felsenstein, J. 2005. PHYLIP (Phylogeny Inference Package) Version 3.6. Seattle, WA, USA: Department of Genetics, University of Washington. Distributed by the author.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
333
Fu, X. y Kim, S. 2010. Agarase: Review of Major Sources, Categories, Purification Method, Enzyme Characteristics and Applications. Marine Drugs 8: 200-218. Giordano, A., G. Andreotti, A. Tramice, y A. Trincone. 2006. Marine glycosyl hydrolases in the hydrolysis and synthesis of oligosaccharides. Biotechnol. J. 1:511–530 Guindon, S. y Gascuel, O. 2003. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Systematic Biology 52: 696-704. Hasegawa M., Kishino H. y Yano T. 1985. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. Journal of Molecular Biology 22: 160-174. Hosoda, A., Sakai, M. y Kanazawa, S. 2003. Isolation and Characterization of Agar-degrading Paenibacillus spp. Associated with the Rhizosphere of Spinach. Bioscience Biotechnology and Biochemistry 67: 1048-1055. Kam N., Park Y.J., Lee E.Y. y Kim HS. 2011. Molecular identification of a polyM-specific alginate lyase from Pseudomonas sp. strain KS-408 for degradation of glycosidic linkages between two mannuronates or mannuronate and guluronate in alginate. Canadian Journal of Microbiology, 57: 1032-1041 Khambhaty Y., Mody K. y Jha B. 2008. Purification, characterization and application of a novel extracellular agarase from a marine Bacillus megaterium. Biotechnology and Bioprocess Engineering 13: 584-591 Kim B.Jo., Kim H.Ju., Duck H.a S., Hee Hwang S., Seok Byun D., Ho Lee T. y Yul Kong Ja. 1999. Purification and characterization of b-agarase from marine bacterium Bacillus cereus ASK202. Biotechnology Letters 21: 1011-1015 Kobayashi, R., M. Takisada, T. Suzuki, K. Kirimura, and S. Usami. 1997. Neoagarobiose as a novel moisturizer with whitening effect. Biosci. Biotechnol. Biochem. 61:162–163. Li J.W., Dong S., Song J., Li C.B., Chen X.L., Xie B.B. y Zhang Y.Z. 2011. Purification and Characterization of a Bifunctional Alginate Lyase from Pseudoalteromonas sp. SM0524 Marine Drugs. 9: 109-123. Miller, G.L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry 31: 426-428. Ramírez-Flandes, S. y Ulloa, O. 2008. Bosque: integrated phylogenetic analysis software. Bioinformatics 24: 2539-2541. Romeo, T. y Preston, JFIII. 1986. Purification and structural properties of an extracellular (1-4)-β-D-mannuronan-specific alginate lyase from a marine bacterium. Biochemistry 25: 8385-8391. Saraswathi, S. Vasanthabharathi, V., Kalaiselvi ,V. y S. Jayalakshmi 2011. Characterization and optimization of agarase from an estuarine Bacillus subtilis. African Journal of Microbiology Research 5: 2960-2968. Shin, M., Lee, D., Skogerson, K., Wohlgemuth, G., Choi, I.G., Fiehn, O. y Kim, K. 2009. Global metabolic profiling of plant cell wall plysacharide degradation by Saccharophagus degradans. Biotechnology and Bioengineering 105: 477-488. Shin, M., Lee, D., Wohlgemuth, G., Choi, I.G., Fiehn, O. y Kim, K. 2010. Global metabolite profiling of agarose degradation by Saccharophagus degradans 2-40. New Biotechnology 27: 156-168. Stevens, R.A. y Levin RE. 1977. Purification and characteristics of an alginase from Alginovibrio aquatilis. Applied and Environmental Microbiology 33: 1156-1161. Sulfahri, Siti Mushlihah, Eko Sunarto, M.Yusuf Irvansyah, Renia Setyo Utami and Sarwoko Mangkoedihardjo 2011 Ethanol Production from Algae Spirogyra with Fermentation by Zymomonas mobilis and Saccharomyces cerevisiae J. Basic Appl. Sci. Res. 1(7): 589-593 Vera, J., Alvarez, R., Murano, E., Slebe, J.C., y Leon, O. 1998. Identification of a marine agarolytic Pseudoalteromonas isolate and characterization of its extracellular agarase. Applied and Environmental Microbiology 64: 4378-4383. Wong, T.Y., Preston, L.A. y Schiller, N.L. 2000. Alginate lyase: review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annual Review of Microbiology 54: 289-340. Wu, S.C. T.-N. Wen, and C. L. Pan, 2005, Algal-oligosaccharide-lysates Prepared by Two Bacterial Agarases Stepwise Hydrolyzed and their Anti-oxidative Properties. Fisheries Science, 71: 1147–1157

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
334
ACTIVIDAD INHIBITORIA DE LA BACTERIA ANTÁRTICA PSEUDOMONAS FRAGI EN LA FORMACIÓN DE BIOPELÍCULAS DE FLAVOBACTERIUM PSYCHROPHILUM
(Inhibitory activity of the antarctic bacterium Pseudomonas fragi on biofilm formation of Flavobacterium psychrophilum)
Leyton, A.1, Urrutia, H1, Vidal, M.1, De la Fuente, M.1, Alarcón, M.1, Aroca, G.3, González-Rocha, G.4,
Sossa, K.1,2 1Laboratorio de Biopelículas y Microbiología Ambiental, Centro de Biotecnología, Universidad de Concepción, Barrio Universitario s/n, Concepción, Chile. E-mail: [email protected]
2Facultad de Ciencias Forestales, Universidad de Concepción, Victoria 631, Barrio Universitario, Concepción, Chile. 3Facultad de Ingeniería Bioquímica, Universidad de Valparaíso, Av. Brasil Nº 2147, Valparaíso, Chile. 4Laboratorio de Investigación en Agentes Antibacterianos, Dpto. de Microbiología, Facultad de Cs. Biológicas, Universidad de Concepción, Concepción, Chile RESUMEN El patógeno de peces Flavobacterium psychrophilum es una bacteria psicrófila Gram negativa cuya temperatura óptima de crecimiento es de 15 °C, es altamente proteolítico e incapaz de degradar los carbohidratos (Bernardet y Kerouault, 1989; Sundell y Wiklund, 2011). Es el agente etiológico de la enfermedad del agua fría y del síndrome del alevín de la trucha arcoíris que afecta a salmones y truchas a nivel mundial (Starliper, 2010). En nuestro país, segundo productor mundial de salmones, las enfermedades causadas por este patógeno causan significativas pérdidas económicas producto de las altas mortalidades que causa en los estados iniciales de desarrollo de las principales especies de cultivo: Salmo salar, Oncorhynchus mykiss y Oncorhynchus kisutch (Henríquez-Núñez et al., 2012). El control de las enfermedades causadas por F. psychrophilum se lleva a cabo mediante quimioterapia. En Chile, los principales antibióticos usados para el tratamiento de las enfermedades provocadas por el patógeno son florfenicol, oxitetraciclina y ácido oxolínico (Henríquez-Núñez et al., 2012). Sin embargo, actualmente se han reportado elevados porcentajes de resistencia de cepas de F. psychrophilum a los principales antibióticos usados en acuicultura (Henríquez-Núñez et al., 2012). Por otro lado, la capacidad de este patógeno de formar biopelículas (Álvarez et al., 2006; Sundell y Wiklund, 2011) es un factor que disminuye la efectividad de la terapia antimicrobiana ya que las células que la constituyen presentan una mayor resistencia a los antibióticos en comparación con las células planctónicas (Costerton et al., 1999). Considerando la diversa bioactividad de bacterias antárticas, se evaluó el efecto inhibitorio en la formación de biopelículas de F. psychrophilum de compuestos secretados por cepas antárticas. Se trabajó con los sobrenadantes libres de células de 67 cepas antárticas, aisladas de distintas muestras recolectadas en la península Fildes, isla Rey Jorge, Territorio Antártico Chileno durante el mes de enero del año 2010. La cuantificación de la biopelícula in vitro se realizó mediante tinción con cristal violeta (Christensen et al., 1985) y la selección del sobrenadante de la cepa que presentó el mayor efecto inhibitorio de la formación de la biopelícula de F. psychrophilum se realizó mediante el cálculo del índice específico de la formación de biopelículas, Specific Biofilm Formation, SBF. (Niu y Gilbert, 2004; Naves et al., 2008). La cepa seleccionada fue identificada mediante el análisis de la secuencia del gen ADNr 16S y se evaluó el efecto antibiopelículas de extractos de la cepa en distintos solventes orgánicos (hexano, diclorometano y acetato de etilo) así como también de las proteínas secretadas por dicha cepa.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
335
Los resultados obtenidos mostraron que un 26,9 % de los sobrenadantes de las cepas antárticas estudiadas presentaron un efecto antibiopelículas fuerte, siendo la cepa M19B la que presentó un mayor efecto inhibitorio. Esta cepa fue identificada como Pseudomonas fragi de acuerdo al análisis de la secuencia del ADNr 16S. Los compuestos extraídos con diferentes solventes orgánicos no mostraron un efecto inhibitorio de la formación de la biopelícula del patógeno. Sin embargo, las proteínas obtenidas, en un rango de concentraciones entre 0,5 y 1,5 mg ml-1 inhibieron la formación de biopelículas de F. psychrophilum. Análisis SDS-PAGE permitieron detectar la presencia de proteínas de entre 48 y 56 kDa en el sobrenadante de la cepa. Estos resultados permiten establecer que estas proteínas fueron las responsables de la actividad inhibitoria de la formación de biopelículas de F. psychrophilum. Este estudio sugiere el potencial uso de las proteínas secretadas por la cepa antártica P. fragi M19B como compuestos inhibidores de la formación de biopelículas de F. psychrophilum. AGRADECIMIENTOS Proyecto INACH M_09-11 de Apoyo a la Realización de Tesis de Postgrado, Proyecto INACH-17-08 y Proyecto Fondecyt 1101009. BIBLIOGRAFÍA Álvarez, B., Secades, P., Prieto, M., McBride, M. y Guijarro, J. 2006. A mutation in Flavobacterium psychrophilum tlpB inhibits gliding motility and induces biofilm formation. Applied and Environmental Microbiology, 72(6): 4044-4053. Bernardet, J. y Kerouault, B. 1989. Phenotypic and genomic studies of "Cytophaga psychrophila" isolated from diseased rainbow trout (Oncorhynchus mykiss) in France. Applied and Environmental Microbiology, 55(7): 1796-1800. Christensen, G., Simpson, W., Younger, J., Baddour, L., Barrett, F., Melton, D. y Beachey, E. 1985. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: a quantitative model for the adherence of staphylococci to medical devices. Journal of Clinical Microbiology, 22(6): 996-1006. Henríquez-Núñez, H., Evrard, O., Kronvall, G. y Avendaño-Herrera, R. 2012. Antimicrobial susceptibility and plasmid profiles of Flavobacterium psychrophilum strains isolated in Chile. Aquaculture, 354: 38–44. Naves, P., Del Prado, G., Huelves, L., Gracia, M., Ruiz, V., Blanco, J., Dahbi, G., Blanco, M., Ponte, M. y Soriano, F. 2008. Correlation between virulence factors and in vitro biofilm formation by Escherichia coli strains. Microbial Pathogenesis, 45(2): 86–91. Niu, C. y Gilbert, E. S. 2004. Colorimetric method for identifying plant essential oil components that affect biofilm formation and structure. Applied and Environmental Microbiology, 70(12): 6951-6956. Starliper, C. 2010. Bacterial coldwater disease of fishes caused by Flavobacterium psychrophilum. Journal of Advanced Research, 2(2): 97-108. Sundell, K. y Wiklund, T. 2011. Effect of biofilm formation on antimicrobial tolerance of Flavobacterium psychrophilum. Journal of Fish Diseases, 34(5): 373-383.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
336
IDENTIFICACIÓN Y BIOPROSPECCIÓN DE LEVADURAS ANTÁRTICAS
(Identification and bioprospecting of antarctic yeasts)
Monserrate-Maggi,L.1, Zambano, K.1, Tacuri, L.1, Cañarte, O.1, Cevallos-Cevallos, J.1 y Peralta, E.1
1Centro de Investigaciones biotecnológicas del Ecuador (CIBE), Escuela Superior Politécnica del Litoral (ESPOL), Campus Gustavo Galindo Km 30.5 vía Perimetral, Ciudad de Guayaquil, Ecuador, [email protected] INTRODUCCIÓN El continente antártico representa un enorme reservorio de microorganismos que producen pigmentos, metabolitos y enzimas frío-activas que presentan altas actividades a bajas temperaturas, lo cual representa un potencial biotecnológico con aplicaciones comerciales. Entre este grupo de extremófilos se encuentran las levaduras, cuya bioprospección puede resultar en el descubrimiento de enzimas frio-activas aplicables en la industria alimenticia. El Centro de Investigaciones Biotecnológicas del Ecuador (CIBE-ESPOL) desde el año 2010 posee una colección de microorganismos de las Islas Shetland del Sur del continente Antártico, entre estas Rey Jorge, Barrientos, Dee y Greenwhich. Uno de los objetivos de este centro es la identificación bioquímica, molecular y bioprospección de aislados de levaduras y su posterior evaluación en la industria de alimentos. MATERIALES Y MÉTODOS Se recolectaron manualmente muestras de suelo en zonas de alta diversidad de especies vegetales, anidación de aves y deshielos en las Islas Torres y Greenwich. Las muestras de suelo se sembraron en los medios de cultivo: Trypcase Soya Agar (TSA), Potato Dextrosa Agar (PDA), Saboraud Dextrosa Agar (SAB) y Yeast Extract Agar. Los aislados de levaduras se caracterizaron en primera instancia por su macro y micromorfología. Además de prueba de catalasa. La extracción de ADN genómico se realizó mediante el protocolo Dellaporta et al. (1983) y Aljanabi y Martinez (1997) y J.L. Cenis (1992) o utilizando PureLink® Genomic DNA Mini Kit (Invitrogen™). Regiones ITS1, 5.8S, y ITS2 fueron amplificadas por PCR y secuenciadas en un laboratorio externo. La secuencia de nucleótidos fue alineada usando BLAST del National Center for Biotechnology Information (NCBI). La identificación se basó en un nivel de identidad de mínimo 98%. La bioprospección se llevó a cabo mediante la incubación de cada una de las levaduras 20 ° C en medio mineral enriquecido con urea, caseína / gluten, almidón de maíz o yuca, y marba (grasa) para determinación de enzimas ureasas, peptidasas, amilasas o lipasas respectivamente; y la producción de alcohol se evaluó por ensayos de fermentación utilizando tubos de Duncan y diversos sustratos incluyendo sacarosa, fructosa, glucosa, maltosa, lactosa, y glicerol a 5, 10, 15, 25 y 30 ° C usando como control positivo Saccharomyces cerevisiae. RESULTADOS Se han obtenido 26 aislados de levaduras de la Isla Greenwich y dos de la Isla Torres, de los cuales se identificó a: Cryptoccocus gastricus, Cryptococcus sp, Candida sp., Candida sake como taxón más cercano, y Rhodotohurla glacialis. Una posible nueva especie de levadura (CIBE 1b.1) se encontró con sólo un 76% de similitud con género Rhodotohurla, además la observación microscópica mostró diferencias significativas en la distribución de los organelos internos en comparación con otras levaduras de dicho genero.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
337
En el perfil bioquímico todas las levaduras dieron positivas para catalasa. Los aislados Candida sake y CIBE12, (ambos de la Isla Torres) y Candida sp. dieron positivos para ureasa, y únicamente la última fue positiva para peptidasa usando gluten como sustrato; las especies del género Candida dieron positivas para amilasa usando como sustrato almidón de yuca o maíz. Ninguno de los aislados mostró actividad de lipasas. Candida sake fue la única que fermentó los azucares: sacarosa, fructosa, glucosa y maltosa con una temperatura óptima de fermentación entre 16 y 25 ° C. Se alcanzó el mayor rendimiento de fermentación cuando se usó glucosa y fructosa como sustrato. DISCUSIÓN Y CONCLUSIONES Las levaduras antárticas presentan un gran potencial como fermentadores e hidrolizadores de almidón y gluten. Como segunda etapa del proyecto determinará el potencial de hidrólisis del almidón o fermentativo para lo cual se determinará la producción y calidad de alcohol lo que determinarán si las capacidades metabólicas de las cepas de levaduras antárticas puedan ser útiles en la industria alimentaria o bebidas alcohólicas AGRADECIMIENTOS Al Instituto Nacional Antártico Ecuatoriano y al Centro de Investigaciones Biotecnológicas del Ecuador . REFERENCIAS BIBLIOGRÁFICAS Baeza, M., Véliz, David., Barahona, S. y Cifuentes, V. 2010. Levaduras antárticas: potencial como herramienta ecológica y fuente de enzimas y metabolitos de interés biotecnológico. Boletín Antártico Chileno. Especial Biodiversidad Antártica.. Volumen 29 Nº 1: 7-8. Akbar Ali Khan Pathan, Bhaskar Bhadra, Zareena Begum, Sisinthy Shivaji. 2010. Diversity of Yeasts from Puddles in the Vicinity of Midre Love´nbreen Glacier, Arctic and Bioprospecting for Enzymes and Fatty Acids. Curr Microbiol 60:307–314. Guamán-Burneo, C., Carvajal-Barriga, J. 2009. Caracterización e identificación de aislados de levaduras carotenogénicas de varias zonas naturales del Ecuador. Universitas. SCIENTIARUM. Vol. 14 N° 2-3: 187-197. Turchetti, B., Buzzini, P.,1, Goretti, M., Branda, E., Diolaiuti, G., D’Agata, C., Smiraglia, C., and Vaughan-Martini, A. 2007. Psychrophilic yeasts in glacial environments of Alpine glaciers. Federation of European Microbiological Societies. Microbiol Ecol 63: 73-83.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
338
EVALUACIÓN DE LEVADURAS MARINAS PROVENIENTES DE LA ANTARTIDA COMO PRODUCTORAS DE ENZIMAS DE INTERÉS INDUSTRIAL Y ÁCIDOS GRASOS POLI-
INSATURADOS
(Evaluation of marine yeast from Antarctica as producers of enzymes and polyunsaturated fatty acids)
Morales D.1, Bruzzone J.1, Lupo S.2, Loperena L.1
1Departamento de Bioingeniería, Inst. Ing. Química, Facultad de Ingeniería, Universidad de la República, Calle Julio H. y Reissig 565, 11300 Montevideo, Uruguay, email: [email protected]
2Laboratorio de Micología, Facultad de Ciencias, Universidad de la República, Calle Iguá 4225, 11400 Montevideo, Uruguay, email: [email protected]
INTRODUCCIÓN La bioprospección puede ser definida como la búsqueda de los productos del metabolismo o de los genomas de los seres vivos para su utilización en procesos industriales o biotecnológicos. En el amplio sentido de la palabra es una actividad que ha sido desarrollada por el hombre desde hace décadas. En los últimos años, el enorme avance logrado por las técnicas de Biología Molecular y el desarrollo de la Biotecnología han incrementado el interés en los programas de bioprospección, los cuales no se limitan actualmente al “screening” de los productos generados por los seres vivos, sino también a la búsqueda de los genes responsables de alguna actividad de potencial interés industrial (Davis 2011). Uno de los focos principales de los actuales programas de bioprospección son los ambientes extremos. Ambientes que presentan muy alta o muy baja temperatura, alta salinidad, valores extremos de pH, son poblados por organismos “extremófilos” (principalmente microorganismos) con adaptaciones que les permiten subsistir bajo esas condiciones extremas que resultan letales para la mayor parte de los seres vivos (Carvalho y Fernandes 2010). El continente Antártico, debido a sus condiciones ambientales específicas, es uno de los hábitats menos poblados de la tierra. Sin embargo, el ambiente marino antártico es muy productivo y esta habitado por organismos procariotas y eucariotas adaptados al frío que poseen características que les permite a ellos sobrevivir en agua de mar a temperaturas permanentemente bajas, cercanas a -1.8ºC (Davis 2011). Los organismos que allí habitan (psicrófilos y psicrotolerantes) presentan adaptaciones en sus sistemas enzimáticos, en sus membranas y por ende en sus genes que tienen un enorme potencial biotecnológico, por lo cual pueden ser objetos de importantes programas de bioprospección (Davis 2011; Hughes y Bridge 2010). Para ser metabolicamente activos en tales condiciones, los microorganismos adaptados al frío deben ajustar sus enzimas y su membrana celular. El cambio más común observado es el incremento de ácidos grasos mono y poliinsaturados en la membrana (Gentile et al. 2003). Dicho ecosistema ha sido objeto de numerosas investigaciones sobre fuentes microbianas de interés tecnológico, como degradadores de compuestos orgánicos en biorremediación, biomineralización, producción enzimática, y fuente de ácidos grasos poliinsaturados, entre otras aplicaciones (Nichols et al. 2002). En el presente trabajo se evalúan levaduras aisladas de agua de mar de las costas de la Isla Rey Jorge como fuente de ácidos grasos esenciales y productoras de enzimas de interés industrial. MATERIALES Y MÉTODOS Las 12 cepas de levaduras en estudio proceden de muestras de algas marinas tomadas en las costas del Mar de Drake y de la Bahía Maxwell (Isla Rey Jorge) e incubadas en medio marino 2216 con antibióticos (penicilina-estreptomicina, 250 mg/l).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
339
La identificación de las cepas se realizó por secuenciación de la región del ADNr 18S e ITS. Las extracciones de ADN genómico se realizaron con el kit AxyPrep Multisource Genomic DNA miniprep (Axygen Biociencies; NY, USA). La amplificación del ADN extraído fue realizada por PCR utilizando los primers NS1, NS (White et al.1990) y un termociclador Palm-CyclerTM (Corbett Research, UK Ltd). La secuenciación fue realizada por Macrogen Inc. (Seúl, Corea). Las secuencias fueron cotejadas y alineadas mediante ClustalW (Thompson et al 1994). Y comparadas con secuencias almacenadas en el banco de datos de nucleótidos del NCBI, mediante el algoritmo BLAST (Altschul et al. 1990). Una relación filogenética fue generada utilizando el modelo de Neighbor-Joining (Saitou y Nei 1987) y el método de Boostrap (evaluación de la significancia estadística) con 1000 iteraciones (Felsestein 1985). Todos los cálculos y análisis se realizaron usando el software Molecular Evolutionary Genetics Análisis, MEGA 5.05, (Tamura et al. 2011). Se determinó la capacidad amilolítica, celulolítica, xilanolítica, ligninolítica, proteolítica (caseinasa y gelatinasa) y lipolítica (degradacion de Tween 80 y tributirina), de 5 cepas pertenecientes a diferentes géneros. Los ensayos se realizaron en placa a 5, 10, 15, 20 y 25ºC. Se midió el halo de hidrólisis del sustrato en estudio. Para la determinación del perfil de ácidos grasos, se utilizó una modificación del método de extracción de lípidos de Bligh y Dyer (1959), posterior transesterificación (saponificación del material graso con NaOH en metanol, a 100ºC bajo atmósfera de nitrógeno y BF3 en metanol al 14% como catalizador). Los ésteres de ácidos grasos se extrajeron con hexano y se determinó su composición utilizando un GC-MS-QP 2010 Ultra con inyector automático AOC-20i. Shimadzu Corporation y columna DB-Wax (30 m largo x 0.25 mm diámetro interno x 0.25 µm de espesor de fase) Agilent J & W.
RESULTADOS Y DISCUSIÓN Mediante el análisis filogenético se identificaron 4 especies: Debaryomyces sp., Metschnikowia sp. , Cryptococcus sp., Rhodotorula sp., figura 1.
Figura 1: Árboles filogenéticos consensos obtenidos por la comparación de las secuencias SSU de los aislamientos del
presente estudio (*) con secuencias del GenBank. Boostrap 1000. Figure 1: Rooted phylogenetic trees showing SSU sequences of the isolates (*) with the highest similarity from the
GenBank database of the NCBI. Boostrap 1000.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
340
Lipolitica (sustrato:tween 80)
Amilolítica Xilanolítica
Temp. de ensayo,°C
5 10 15 20 25 5 10 15 20 25 5 10 15 20 25
Cepa D4 9,0 13,0 12,0 15,0 16,7 - - 2,7 3,2 3,5 - - - 18,3 21,3
Cepa D5 - - - - - - - - - - - - - - -
Cepa D7 - - - - - - - - - - - - - - -
Cepa D9 8,7 8,3 7,7 11,3 9,3 - - - - 6,0 - - - - -
Cepa D14 9,3 16,7 15,3 19,3 18,0 6,0 12,7 12,7 11,7 13,3 - - - 18,7 21,3 El valor reportado es el promedio de tres ensayos, con una desviación estándar de 0.1mm.
Tabla 1: Actividades enzimáticas encontradas, se indican los radios de los halos medidos en mm. Table 1: Enzyme activities, radius of the halos in mm.
Las actividades enzimáticas encontradas se presentan en la tabla 1. Solo en las cepas D5 y D7 se observó actividad lipolítica con tributitina como sustrato (5 y 4 mm de halo respectivamente) y a una única temperatura, a 5°C, no se reporta en tabla. Ninguna de las levaduras en estudio presento actividad proteolítica, celulolítica ni lignolítica independientemente de la temperatura ensayada. Los perfiles de ácidos grasos de las cepas en estudio contenían una proporción de ácidos grasos insaturados de 70-80 %, principalmente ácido oleico (C 18:1), linoleico (18:2 n-6) y linolénico(18:3 n-3), estos dos últimos de gran interés por ser esenciales. El contenido de ácido linoleico y ácido linolénico en las cepas fue de: 28 y 6 % (cepa 5), 18 y 1,6 % (cepa 9), 11 y 3,22 % (cepa D7) respectivamente. Las cepas D4 y D14 solo presentaron ácido linoleico, en orden 10 % y 19%. En la figura 2 se muestra el perfil de ácidos grasos que corresponde a la cepa D5, el mejor perfil obtenido si se busca una fuente de ácidos grasos de valor nutraceutico, por su contenido de ácidos grasos esenciales (18:2 n-6) y (18:3 n-3).
C
18:
3
C 1
8:2C
18:
1
C 1
7:2
C 1
7:1
C 1
6:1
C 1
6:0
C 1
4:0
C 1
8:0
C 1
8:3
C 1
8:2C
18:
1
C 1
7:2
C 1
7:1
C 1
6:1
C 1
6:0
C 1
4:0
C 1
8:0
≈
C 1
8:3
C 1
8:2C
18:
1
C 1
7:2
C 1
7:1
C 1
6:1
C 1
6:0
C 1
4:0
C 1
8:0
C 1
8:3
C 1
8:2C
18:
1
C 1
7:2
C 1
7:1
C 1
6:1
C 1
6:0
C 1
4:0
C 1
8:0
≈ Figura 3: Perfil de ácidos grasos de la cepa 5, cultivada a 15°C, obtenido por GC-MS.
Figure 1: Fatty acid profile of strain 5, cultured at 15 °C, obtained by GC-MS.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
341
CONCLUSIONES Se encontraron solo cuatro actividades enzimáticas de las ocho ensayadas, destacándose la cepa D14 por ser la que presentó las mayores actividades en tres de las cuatro actividades halladas (lipolítica- Tween 80, amilolítica y xilanolítica). Solo se encontró mayor actividad enzimática a baja temperatura (5°C) en las cepas D5 y D7 cuando crecieron sobre agar-tributirina. Todas las cepas contienen un alto porcentaje de ácidos grasos insaturados y se destaca la cepa D5 por su contenido de ácidos grasos esenciales. Confirmándose que las levaduras presentes en Antartida son potenciales fuentes de AGPI de interés biotecnológico. AGRADECIMIENTOS Al Quim. Lic. Manuel Minteguiaga Carvajal, al Instituto Antártico Uruguayo (IAU), a la Agencia Nacional de Innovación e Investigación (ANII). REFERENCIAS BIBLIOGRÁFICAS Altschul, S.F., Gish, W., Miller, W., Myers, E.W. y Lipman, D.J. 1990. Basic local alignment search tool. J. Mol. Biol., 215:403-410. Bligh, E.G. y Dyer W.J. 1959. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol., 37:911-917. Davis, J.M. 2011. Reconsidering Antarctic Bioprospecting through Territorialitis of Science, Property, and Governance. Disertation, pages:1-228, Ohio State University. de Carvalho, C.C.C.R. y Fernandes, P. 2010. Production of Metabolites as Bacterial Responses to the Marine Environment. Mar. Drugs, 8: 5-727. Felsenstein, J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783-791. Gentile, G., Bonasera, V., Amico, C., Giuliano, L. y Yakimov, M.M. 2003. Shewanella sp. GA-22, a psychrophilic hydrocarbonoclastic antarctic bacterium producing polyunsaturated fatty acids. Journal of Applied Microbiology, 95:1124-1133. Hughes, K. y Bridge, P. 2010. Potential impacts of Antarctic bioprospecting and associated commercial activities upon Antarctic science and scientists. Ethics in science and environmental politics, 10:3-18. Nichols, D.S., Sanderson, K., Buia, A., Van de Kamp, J., Holloway, P., Bowman, J.P., Nichols, D., Mc Meekin, T.A., Smith, M., Cancuso, C. y Nichols, P. 2002. Jabour-Green, J. y Haward, M. (Eds). En “Bioprospecting and Biotechnology in Antarctica” in The Antarctic: Past,Present and Future. Antarctic CRC Research Report #28. Hobart, pages:85-103. Saitou, N y Nei, M. 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution 4:406-425. Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S. 2011. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Thompson, J.D., Higgins, D.G. y Gibson, T.J. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22(22):4673-4680. White, T.J., Bruns, T., Lee, S. y Taylor, J.W. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Pages 315-322 In: PCR Protocols: A Guide to Methods and Applications, eds. Innis, M. A., D. H. Gelfand, J. J. Sninsky, and T. J. White. Academic Press, Inc., New York.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
342
BIOPROSPECCIÓN DE BACTERIAS ANTÁRTICAS PARA EL DESCUBRIMIENTO DE NUEVOS COMPUESTOS PEPTÍDICOS CON ACTIVIDAD ANTIMICROBIANA.
(Bioprospection of Antarctic bacteria for finding new peptidic compounds with antimicrobial activity)
Pavlov, M.1, Gallardo, J.2, Lavín, P.2, Luna O 1, Cárdenas, C 3, Guzmán F 3., González, M.2 & Marshall S1,3.
1 Laboratorio de Genética e Inmunología Molecular, Instituto de Biología, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile. E-mail: [email protected] 2 Laboratorio de Biorrecursos Antárticos, Instituto Antártico Chileno, Punta Arenas, Chile. E-mail: [email protected] 3 Núcleo Biotecnología Curauma (NBC), PUCV, Valparaíso, Chile INTRODUCCIÓN El continente antártico y sus mares adyacentes resultan un interesante nicho de descubrimiento de biorrecursos, no sólo por representar una de las áreas más remotas e inexploradas del planeta sino también por la infrecuente combinación de presiones selectivas que dirigen la evolución de los organismos que habitan en este continente (i.e. bajas temperaturas, baja actividad de agua [aw], ciclos de congelamiento/descongelamiento de los suelos, estrés osmótico), tendientes a generar estrategias adaptativas únicas de sobrevivencia (Wong et al., 2011; Marx et al., 2007). Recientemente O´Brien et al. (2004) han sugerido que las bacterias antárticas, no sólo han desarrollado modificaciones celulares/moleculares adaptativas para enfrentar dichas condiciones adversas (i.e. producción de vesículas de membrana, flexibilidad estructural de enzimas, síntesis/degradación macromoléculas atípicas, entre otras), sino que también se han valido de la producción de efectores de antagonismo para reducir la presencia de microorganismos competidores y así lograr una dominancia de los recursos que suelen ser restringidos en estos ambientes extremos. Dentro de los efectores de antagonismo clásicos de bacterias se encuentran los antibióticos, agentes líticos, lisozimas y bacteriocinas, siendo estas últimas compuestos proteicos con actividad antimicrobiana, descritas como “the microbial weapon of choice” debido a la abundancia y diversidad existente entre las bacterias productoras (Riley & Wertz, 2002). Si bien microorganismos pertenecientes a nuevos hábitat están siendo continuamente explorados en la búsqueda de compuestos antimicrobianos, los estudios enfocados en la caracterización intensiva de microorganismos antárticos productores de actividades antimicrobianas/antagónicas son limitados y en la mayor parte de ellos no se caracterizan los compuestos efectores de dichas actividades. [Wong et al., 2011; Rojas et al., 2009; Biondi et al., 2008; Lo Giudice et al., 2007a-b]. Por lo anterior el objetivo del presente trabajo es seleccionar cepas antárticas capaces de inhibir el crecimiento de bacterias indicadoras y aislar compuestos proteicos antimicrobianos responsables de dicha actividad antimicrobiana. MATERIALES Y MÉTODOS Cultivos, condiciones de crecimiento y evaluación de actividad antimicrobiana Se evaluaron 1079 morfotipos bacterianos aislados desde muestras de suelo y agua de la Antártica Marítima (Campaña estival 2010), en cuanto a su capacidad de antagonizar el crecimiento de Staphylococcus aureus ATCC 25923 (cepa indicadora) y/o Escherichia coli ATCC 25922. Cinco de las cepas que presentaron actividad antagonista fueron seleccionadas para las siguientes evaluaciones. Dichas cepas fueron inoculadas en duplicado en medio LB (Lysogeny Broth) e incubadas a 20°C hasta alcanzar la fase exponencial y estacionaria de crecimiento, respectivamente. Los sobrenadantes libres de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
343
células (SLC) obtenidos de estos cultivos fueron concentrado 40 veces y evaluados en cuanto a su capacidad de inhibir el crecimiento de las cepas antes mencionadas, pre-inoculadas en una placa de LA (Lysogeny Agar) 0,7% Agar. Identificación de las bacterias antárticas. Se realizó extracción de DNA genómico de las 5 cepas seleccionadas, utilizando el kit AxyPrep Bacterial Genomic DNA Miniprep (Axygen). Éste gDNA fue utilizado como templado para amplificar por PCR el rDNA 16S. El producto específico resultante fue clonado utilizando TOPO TA Cloning kit (Invitrogen) y secuenciado para su posterior análisis a través del software BLASTn. Fraccionamiento en fase sólida de los sobrenadantes de cultivo. Cada SLC que presentó actividad antimicrobiana fue fraccionado a través de una columna de extracción en fase sólida C18 (1g resina; Sep-Pak, Waters) con un gradiente discontinuo de acetonitrilo (ACN). El volumen de cada fracción recuperada fue ajustado de manera que la concentración final fuera 40 veces mayor respecto al volumen inicial. Dichas fracciones fueron evaluadas en cuanto a su actividad antimicrobiana contra la cepa de indicadora y aquellas que presentaron actividad fueron re-fraccionadas como se señala a continuación. Electroforesis SDS-Tris-Tricina. La separación de compuestos proteicos desde las fracciones activas de los SLC se realizó siguiendo la metodología de electroforesis SDS-Tris-Tricina descrita por Schaegger & Von Jagow [1987] con las siguientes modificaciones. El gel concentrador (15%) y el gel separador (5%) fueron preparados en tampón Tris pH 8,5 - 0,3% SDS. 200 mM Tris pH 8,9 y 100 mM Tris pH 8,5 con 0,1% SDS fueron utilizados como tampones para el compartimento del ánodo y cátodo, respectivamente. Actividad antimicrobiana de productos proteicos Se seleccionaron y cortaron las bandas más prominentes de los geles preparativos Tris-Tricina, las que fueron evaluadas en cuanto a su actividad antimicrobiana siguiendo la metodología señalada previamente. Las proteínas retenidas en ellas fueron electroeluidas por ~16 horas en tampón volátil 50 mM NH4HCO3 - 0.01 % SDS. El producto eluido fue liofilizado, resuspendido en tampón fosfato (10 mM - pH 7,4) y almacenado a -20° C. RESULTADOS Cuarenta y cinco (4,7%) de los aislados de bacterias antárticas evaluados fueron capaces de inhibir el crecimiento de S. aureus y/o E. coli en ensayos de antagonismo. Cinco de estos aislados (Tabla 1) fueron seleccionados, basándose en las dimensiones del área de inhibición producida, para caracterizar parcialmente los compuestos responsables de la actividad antimicrobiana.
Cepa Antártica
Taxón con mayor similitud de secuencia/ Número de acceso GenBank
Identidad (pb)
Similitud (%)
IB20 Pseudomonas fluorescens SBW25 / AM181176.4 1503/1511 99
6A1 Pseudomonas mandelii Asd MV-11 / FM955880.1 1523/1530 99
S6.3 Pseudomonas fluorescens / CCM2115 1492/1500 99
3013 Streptomyces sp. / JX262288.1 490/558 88
CP10 Microbacterium sp. 1294/1300 99 Tabla 1. Identidad más probable de las cepas antárticas productoras de compuestos antimicrobianos, basándose en el análisis
por homología de la secuencia completa/parcial del rADN 16S.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
344
Table 1. Most likely identity Antarctic strains producing antimicrobial compounds based on the analysis complete sequence homology / partial 16S rDNA.
Sobrenadantes libres de células (SLC) obtenidos desde cultivos en fase exponencial (cepas CP10, 3013 e IB20) o en fase estacionaria (cepas 6A1 y S6.3) de las cepas seleccionadas, presentaron actividad antimicrobiana contra S. aureus y/o E. coli. Luego del fraccionamiento de dichos SLC por cromatografía en fase sólida C18, la actividad antimicrobiana asociada a cada cepa quedó restringida a las fracciones eluidas entre 40-60% ACN (CP10), 55-60% ACN (3013), 60-80% ACN (IB20), 0-30% ACN (6A1) y 40-60% ACN (S6.3). Estas fracciones cromatográficas se analizaron en geles SDS-Tris-Tricina para determinar posibles efectores proteicos responsables de la actividad antimicrobiana. La fracción 40-60% ACN de la cepa CP10 presentó 5 bandas proteicas mayoritarias (~10 kDa, 17-27 kDa, ~34 kDa, 43-55 kDa y >55 kDa) que luego de ser electroeluidas presentaron actividad contra la cepa indicadora. La misma actividad se verificó para el producto proteico contenido en las siguientes bandas, provenientes de la fracción activa de cada cepa: banda única de la cepa 3013 (~10 kDa) (Figura 1), 2 bandas proteicas de IB20 (~31 kDa y ~55 kDa), 6 bandas de la cepa 6A1 (<10 kDa, 10kDa, ~ 10 kDa, 10-17 kDa, ~ 17 kDa, 26-34 kDa, 43 kDa) y tres bandas para S6.3 (10-34 kDa, 43 kDa y 55 kDa). Figura 1: (A) Perfil proteico de la fracción cromatográfica activa obtenida del sobrenadante libre de células de la cepa 3013. Panel A carril 1: fracción 55-60% ACN, carril 2: Estándar de peso molecular. La cabeza de flecha indica la banda (~10 kDa)
cuyo producto presentó actividad antimicrobiana contra S. aureus (B). Panel B Arriba: halo inhibición producido por la banda de 10 kDa, Panel B Abajo: control negativo de actividad antimicrobiana.
Figure 1: (A) protein profile of the active chromatographic fraction obtained from cell free supernatant of strain 3013. Panel A lane 1: 55-60% ACN fraction, lane 2: molecular weight standard. The arrowhead indicates the band (~ 10 kDa), whose
product exhibited antimicrobial activity against S. aureus (B). Top Panel B: inhibition halo produced by the band of 10 kDa, Panel B Bottom: negative control of antimicrobial activity.
DISCUSIÓN Y CONCLUSIONES. La frecuencia de aislamiento de bacterias antárticas con actividad antimicrobiana obtenida en la presente prospección (4,7%) supera los valores documentados a partir de evaluaciones masivas de microorganismos heterótrofos antárticos [Wong et al., 2011; Rojas et al., 2009; Lo Giudice et al., 2007b] y se asemeja a las frecuencias obtenidas en evaluaciones rutinarias de hongos filamentosos y actinomicetes, definidos como los productores por excelencia de antibióticos. Lo anterior permite confirmar que las bacterias antárticas representan un nicho interesante de exploración en la búsqueda de nuevos compuestos antimicrobianos. La actividad antimicrobiana verificada en los sobrenadantes de cultivo de las 5 cepas antárticas seleccionadas, al menos en parte, se asocia a compuestos proteicos secretables, con distintos grado de hidrofobicidad, cuyos pesos moleculares varían entre ~10 kDa y ~55 kDa. Cabe destacar que estos compuestos mantienen su actividad antimicrobiana pese a ser purificados a través de SDS-PAGE, cuyas
A
10 kDa
17 kDa
1 2 B

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
345
condiciones denaturantes suelen resultar en la pérdida de funcionalidad para la mayor parte de los compuestos proteicos. Debido a las características antes mencionadas podrían ser incluidos preliminarmente como compuestos tipo bacteriocinas, efectores antimicrobianos que en los últimos años han sido caracterizados en numerosas Pseudomonas spp. y Actinomicetes de áreas templadas , y que se ha demostrado presentan propiedades tales como alta actividad específica y baja toxicidad. En la actualidad se están realizando los análisis de espectrometría de masas de cada una de las bandas purificadas para asociarlas con algún tipo de bacteriocina ya descrita. Estas moléculas se posicionan como alternativas viables para la generación de futuros antimicrobianos de uso profiláctico y terapéutico, tanto en la industria alimentaria como en ámbitos clínicos. AGRADECIMIENTOS Esta investigación es financiada por INNOVA-CORFO. Maria Soledad Pavlov es financiada por CONICYT. LITERATURA CITADA Lo Giudice, A., Bruni, V. y Michaud L. 2007a. Characterization of Antarctic psychrotrophic bacteria with antibacterial activities against terrestrial microorganisms. Journal of Basic Microbiology 47 (6): 496-505. Lo Giudice, A., Brilli, M., Bruni, V., De Domenico, M., Fani, R. y Michaud L. 2007b. Bacterium-bacterium inhibitory interactions among psychrotrophic bacteria isolated from Antarctic seawater (Terra Nova Bay, Ross Sea). FEMS Microbiology Ecology 60 (3): 383-396. O'Brien, A., Sharp, R., Russell, N. y Roller S. 2004. Antarctic bacteria inhibit growth of food-borne microorganisms at low temperatures. FEMS Microbiology Ecology 48 (2): 157-167. Riley, M. y Wertz J. 2002. Bacteriocins: Evolution, Ecology, and Application. Annual Review Microbiology 56: 117-137. Rojas, J., Martin, J., Tormo, J., Vicente, F., Brunati, M., Ciciliato, I., Losi, D., Van Trappen, S., Mergaert, J., Swings, J., Marinelli, F.y Genilloud O. 2009. Bacterial diversity from benthic mats of Antarctic lakes as a source of new bioactive metabolite. Marine Genomics 2 (1): 33-34. Schaegger, H. y Von Jagow, G. 1987. Tricine-Sodium dodecyl sulfate-Polyacrylamide Gel Electrophoresis for the Separation of Proteins in the Range from 1 to 100 kDa. Anal. Biochem. 166: 368-379. Wong, C., Tam, H., Alias, S., González, M., González-Rocha, G. & Yevenes M. 2011. Pseudomonas and Pedobacter isolates from King George Island (Antarctica) inhibited the growth of foodborne pathogens. Polish Polar Research 32: 3-14. Marx, J., Collins, T., D'Amico, S., Feller G. y Gerday C. 2007. Cold-adapted enzymes from marine Antarctic microorganisms. Marine Biotechnology 9 (3):293-304.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
346
PURIFICATION OF AN ALKALINE PHOSPHATASE FROM AN ANTARCTIC BACTERIUM
(Purificación de una fosfatasa alcalina desde una bacteria Antártica).
Pereira, J.1,2, Peralta, R.1,3 , Blamey, L.1 and Blamey J.M.1
1 Fundación Científica y Cultural Biociencia, José Domingo Cañas ·2280, Ñuñoa. Santiago of Chile. [email protected] 2 Universidad Tecnólogica de Chile INACAP, Av. Vicuña Mackenna ·3864, Macul. Santiago of Chile. 3 Department of Biology, Universidad de Santiago de Chile. Av. Lib. Bernardo O'Higgins 3363, Est. Central, Santiago of Chile INTRODUCTION The Antarctic extremophilic microorganisms represent new species and have broad phylogenetic diversity with representatives from Bacteria and Archaea domains. The spectrum of microorganisms includes hyperthermophiles, halophiles, alkalophiles or acidophiles. Cold extreme environments are populated by pyschrophiles and psychrotrophs, which produce cold adapted enzymes, allowing them a high flexibility in their adaptation at these conditions. Some of these microorganisms are alkaliphiles, defined as organisms that have their optimum growth rate at least two pH units above neutrality. The alkalitolerants are capable of growing at values of pH higher than 9 or 10, but their optimum growth rate is around neutrality or less, as reported by Krulwich y Guffanti (1989). These microorganisms have been described as producing a variety of interesting extracellular enzymes, such as alkaline phosphatase. This enzyme belongs to the category of hydrolases, responsible for removing phosphate groups from many types of molecules, including nucleotides, proteins, and alkaloids. This process of removing the phosphate group is called dephosphorylation. MATERIALS AND METHODS 1. Growth conditions and enzymatic activity from alkalitolerant REF3: Samples were collected from Valentino Cape Island and Deception Island during a scientific expedition. Samples were inoculated in sterile medium containing peptone and sodium chloride at the temperature and pH at which samples were collected. The pellet was re-suspended on an isotonic medium, prior the detection of enzymatic activities. Cultures were grown at 4ºC in a medium that contains peptone (15 g/l) and sodium chloride (30 g/l) at different pH values for 10 days. Optical density was measured at defined time intervals to determine the maximum growth rate.Screening for 19 enzymatic activities were analyzed by a semi-quantitative APYZYM method on non-purified extracts of the isolate REF3. A sample culture was prepared and centrifuged at 9000 rpm. The isolate REF3 genomic DNA was extracted using a special cellular disruption method followed by a standard phenol/chloroform protocol. The extracted DNA was used as template for the PCR amplification of the 16S rRNA gene using specific primers for Archaea (751F and 1406R) and Bacteria (334F and 939R). Verification of DNA extraction and PCR amplification were carried out by running the sample on a 1% agarose gel in TAE buffer. 2. Purification of enzyme: The cells were grown in bottles of 15 L flasks with the designed medium. The cells were disrupted by sonication. The insoluble fraction containing the membranes was removed by ultracentrifugation at 9000 rpm at 4ºC for 15 minutes obtaining the supernatant containing APase. This crude extract was loaded in two anionic exchange columns; first a DEAE Sepharose and then a Q HiTrap column. Both columns were previously equilibrated using a 10 mM Tris-HCl containing 0.5 mM CaCl2 buffer at pH 8.0. The proteins in both cases were eluted with a linear gradient of NaCl in buffer 10 mM Tris-HCl, 0.5 mM CaCl2 at pH 8.0. Alkaline phosphatas activity was measured spectrophotometrically by monitoring the release of p-nitrophenol from p–nitrophenyl phosphates

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
347
(pNNP) at 410nm as reported by de Prada et al., 1996. The culture broth was centrifuged at 9000 rpm for 15 minutes at 4ºC.The crude extract was used for the assay. 100 ul of cell free supernatant (crude extract) was mixed with 167 µl of substrate. The mixture was incubated at 37ºC for 10 minutes. The reaction was stopped at 15 minutes with 50 ul of NaOH 4M in order to measure the activity at this time. One unit of enzyme activity was defined as the amount of enzyme required to release 1umol of p-nitrophenol from pNPP in 1min. 3. Genes encoding for alkaline phosphatase in REF3 isolate: The published genes that codify for the alkaline phosphatase enzyme were obtained from NCBI database to design specific the primers. An alignment was performed with nucleotide sequences of the genes coding for alkaline phosphatase found in several species of the genus Shewanella. The alignment was performed by Clustal W2 online program. Primers were designed to amplify the end regions of the gene encoding alkaline phosphatase, and for an internal region of the gene. 4. PCR Detection of the alkaline phosphatase gen in Shewanella: The genetic material from Shewanella was obtained with a standard phenol/chloroform/isoamyl alcohol (25:24:1) protocol and the result of this extraction was checked by agarose gel electrophoresis. The amplification of the alkaline phosphatase gene from Shewanella was performed by PCR in a Corbett thermocycler CG 1-96, according to standard protocol for PCR amplification. RESULTS The psychrophile REF3 isolate is an alkalitolerant bacterium, which has an optimal growth pH of 8 and a maximum growth rate of 0,068 h-1. Molecular analysis of the 16S rRNA gene shows that REF3 belongs to the genus Shewanella. It changes its bacilar morphology with pHs from small rods at acidic pH to long one at alkaline pH. This isolate produces two kind of phosphatases one alkaline and the other acidic based on APYZYM assay. The phylogenetic relationship between REF3 and other species of the genus Shewanella based in the 16S rRNA gene sequence analysis allowed us to construct a phylogenetic tree. In the phylogenetic analysis REF3 showed a 99% of identity with Shewanella putrefaciens. Amplification of the alkaline phosphatase gene: In order to visualize the size of the amplified fragment obtained for the alkaline phosphatase gene, a gel electrophoresis was performed showing that the best condition for PCR was 55ºC using the pair of primers IPALK2F and FPALK3R. The amplified fragments obtained had sizes of 1500 bp and 700 bp. respectively. Each amplified fragment was cut from the gel and cloned using TOPO TA cloning kit DISCUSION AND CONCLUSION REF3 isolate belongs to the genus Shewanella has different cellular morphology depending of the pH media. It changes its cellular morphology with pHs from small rods at acidic pH to long one at alkaline pH. REF 3 is an alkalitolerant bacterium which has an optimal growth pH of 8 and a maximum growth rate of 0,068 h-1. In this study alkaline phosphatase activity was detected through APYZYM system and by monitoring the release of p-nitrophenol from p–nitrophenyl phosphate. Alkaline phosphatase was purified from isolate REF3 using two anionic exchange columns. In different purifications systematic loss of activity was observed along the purification steps. Probably the addition of the divalent cations Zn or Mg into the buffer could increase the stability of the protein along the purification, as reported by Muginova et al, 2007. We have detected the alkaline phosphatase gene in the REF3 genome using degenerated primers IPALK2F/FPALK3R. The amplified genes had a size of 1500 and 700 bp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
348
ACKNOWLEDGEMENTS Our thanks to Corfo Innova # 07CN13PXT264 (2008-2012) and to the Instituto Antartico Chileno (INACH) for financing this work. REFERENCES Krulwich T.A. y Guffanti A.A.1989. Alkalophilic bacteria. Annual Review of Microbiology 43: 435–463. De Prada, P., J. Loveland-Curtze y Brenchley J. 1996. Production of two extracelullar alkaline phosphatases by a psychrophilic Arthrobacter strain. Appl.Environ.Microbiol. 62:3732-8. Muginova S., Havoronkova A.,. Poliakov A. y Shekhovtsova T. 2007. Application of Alkaline Phosphatases from Different Sources in Pharmaceutical and Clinical Analysis for the Determination of Their Cofactors; Zinc and Magnesium Ions. Analytical sciences 23: 357-363.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
349
EL EFECTO DEL GUANO SOBRE LA FIJACIÓN BIOLÓGICA DEL NITRÓGENO EN LAS COSTRAS BIOLÓGICAS EN LA PENÍNSULA DE ARDLEY, ISLA REY JORGE.
(The effect of guano on N fixation of biological soil crusts in Ardley Peninsula, King George Island)
Pérez, C.A.1, Aravena, J. C2., Ivanovich, C2, Mansilla, C.3, McCulloch, R3.
1Instituto de Ecología y Biodiversidad, Las Palmeras 3425, Ñuñoa, Santiago, Chile [email protected] 2Centro de Estudios Cuaternarios de Fuego Patagonia y Antártica (CEQUA), 21 de Mayo 1690, Punta Arenas,Chile. 3Universidad de Stirling, Stirling FK9 4LA, Escocia INTRODUCCIÓN La única fuente de nitrógeno para los líquenes, musgos y hepáticas que forman las costras biológicas del suelo (CBS) en la Península de Ardley es de tipo atmosférico, ya sea por volatilización de amoniaco desde el guano de los pingüinos (Lindeboom 1984), o por fijación simbiótica de nitrógeno (FSN) a través de asociaciones con bacterias diazotrofas (Lindsay y col., 1978, Fernandez y col., 2001). Nuestra principal hipótesis es que el ingreso indirecto de amonio a través de las precipitaciones o directo a través de la adición experimental de guano a las costras biológicas inhibe el proceso de FSN, debido al mecanismo de retroalimentación negativa que controla la actividad de la enzima nitrogenasa a través del nitrógeno mineral (Paul y Clark 1988). El objetivo principal de este estudio fue evaluar el efecto en la FSN, ya sea en forma indirecta a través del amonio disuelto en las precipitaciones, o en forma directa a través de la adición de guano a las costras biológicas. MATERIALES Y MÉTODOS La Península Ardley ofrece un laboratorio natural que permite poner a prueba esta hipótesis debido a que el paisaje está representado por un gradiente de vegetación consistente en un tipo ornitófilo que se ubica muy cercanamente a la colonia de pingüinos ubicada al lado norte de la Península (Pin), sitios intermedios, ubicados entre la pingüinera y el lado sur de la Península (Div, Re) y finalmente un sector al lado sur-oeste de la Península compuesto por una serie de paleoplayas (PP7, PP5, PP3, PP2) distantes de la pingüinera (Fig. 1).
Figura 1: Mapa de los sitios de estudio Figure 1: Map of the study sites
Pin
DivRe
PP7PP5
PP3PP2
Pin
DivRe
PP7PP5
PP3PP2

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
350
Durante el mes de Febrero los años 2012 y 2013 se estimó la tasa de FBN a través del ensayo de reducción de acetileno por la enzima nitrogenasa en ensayos in situ de CBS, conformadas por comunidades de líquenes y briófitas en la Península de Ardley, en muestras a las que se le aplicó guano y muestras control. Las diferencias entre sitios y tratamientos (con y sin guano) se evaluaron mediante un Análisis de Varianza (Andeva) de dos factores con los datos normalizados y a posteriori Tukey tests, con el programa Statistica. Con Andeva de un factor y a posterior Tukey test se evaluaron las diferencias en las tasas de reducción de acetileno entre las especies. Las diferencias fueron consideradas estadísticamente significativas con un nivel de p<0.05). RESULTADOS Los resultados muestran que la concentración de amonio en las precipitaciones es sólo significativa en las precipitaciones de los sitios de correspondientes a la pingüinera (PIN a y b) y el sitio más cercano a ella (Pin Ara a y b) (Fig. 2).
0 .0 0
0 .2 0
0 .4 0
0 .6 0
0 .8 0
1 .0 0
1 .2 0
1 .4 0
1 .6 0
P P 7 P P 5 P P 3 P P 2 P inA ra a
P inA ra b
P in a P in b
si ti o s
mg/L
N it ra t o
A m o n io
Figura 2: Concentración de amonio y nitrato en las precipitaciones en la Península Ardley
Figure 2: Ammonium and nitrate concentration in precipitation in Ardley Peninsula.
Encontramos una completa ausencia de la actividad reductora de acetileno en las CBS del sitio de estudio más cercano a la pingüinera (PIN), pero tasas significativas de la enzima nitrogenas se encontró en las CBS de las paleo-playas y en los sitios intermedios (Fig. 3). La adición de guano (g en la Fig. 3) a las costras biológicas redujo significativamente la actividad de la nitrogenasa en la paleo-playa más antigua y en los sitios intermedios (Fig. 3).
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
P P 7 P P 7 g P P 5 P P 5 g P P 2 P P 2 g R E R E g D IV D IV g P IN
S it io s
nmol etileno/Peso seco/día
*
*
*
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
P P 7 P P 7 g P P 5 P P 5 g P P 2 P P 2 g R E R E g D IV D IV g P IN
S it io s
nmol etileno/Peso seco/día
*
*
*
Figura 3: Actividad reductora de acetileno de la enzima nitrogenasa en costras biológicas del suelo conformadas por líquenes
y briófitas de la Península Ardley Figure 3: Acetylene reduction activity of nitrogenase enzyme in biological soil crusts conformed by lichens and bryophytes
in Ardley Peninsula. Las especies con la actividad más alta de la enzima nitrogenasa fueron los cianolíquenes Placopsis contortuplicata (360-602 nmol etileno/g peso seco/día) y Sterocaulon alpinum (190 nmol etileno/g peso seco/día) y en un menor grado Psoroma hypnorum (30 nmol etileno/g peso seco/día ) (Fig. 4).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
351
0
1
2
3
4
5
Ste
rocu
alon
gla
brum
Bue
llia
coni
ops
Sph
aero
phor
us g
lobo
sus
Riz
ocar
pon
geog
raph
icum
Him
anto
rmia
lugu
bris
Usn
ea fa
scia
ta
Her
zogo
bryu
m te
res
Pol
ytric
hast
rum
alp
inum
Loph
ozia
exc
isa
Cla
doni
a gr
acili
s
San
ioni
a sp
And
reae
reg
ular
is
Leci
dea
crem
onic
olor
Pso
rom
a hy
pnor
um
Pla
cops
is c
onto
rtup
licat
a
Ste
roca
ulon
alp
inum
nmol etileno/peso seco/día (exp.)
Figura 4. Actividad reductora de acetileno de la enzima nitrogenasa en distintas especies líquenes y briófitasde que
conforman las costras biológicas del suelo en la Península Ardley Figure 4. Acetylene reduction activity of nitrogenase enzyme in different species of lichens and bryophytes that conform the
biological soil crusts in Ardley Peninsula. DISCUSIÓN Y CONCLUSIONES Los resultados de nuestros ensayos in situ indican que ingresos significativos de nitrógeno a los ecosistemas de la Isla Ardley estarían ocurriendo efectivamente a través de dos vías atmosféricas importantes: en nitrógeno disuelto en las precipitaciones y la FSN. Nuestros resultados de adición de guano a las CBS indican que existe un significativo mecanismo de retroalimentación negativa en donde el producto de la FBN, el N disponible en forma de amonio, inhibe la actividad de la enzima nitrogenasa. Nuestros resultados entregan el mecanismo que explica la distribución de la vegetación criptogámica nitrofílica en suelo ornitogénicos versus la nitrófoba en suelos no-ornitogénicos, que está determinada por el mecanismo de regulación de las bacterias diazotrofas. AGRADECIMIENTOS Proyecto INACH T01-11 REFERENCIAS Lindeboom, H.1984. The nitrogen pathway in a pinguin rookery. Ecology 65(1): 269-277. Lindsay, D. C. 1978. The role of lichens in Antarctic ecosystems. The Bryologist 81(2): 266-276. Fernandez, E., Quesada, A., Howard, C. y Hawes, I. 2001. N2 Fixation in cyanobacterial mats from ponds on the McMurdo Ice Shelf, Antarctica. Microbial Ecology 42:338-349. Paul, E. y Clark, F.1988. Soil Microbiology and Soil Biochemistry. Academic Press, Nueva York.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
352
RESPUESTA A DAÑO OXIDATIVO DE CEPAS HIPER RESISTENTES A TELURITO PROVENIENTES DEL TERRITORIO ANTÁRTICO CHILENO.
(Response to oxidative damage of tellurite hiper-resistant strains from the Chilean Antarctic territory)
Pozo M.F.1, Pugin B.1, Muñoz C.1, Pinto C. 1, Díaz W.A.1, 2,3, Arenas F.A. 1 y Vásquez C.C.1
1 Laboratorio de Microbiología Molecular, Universidad de Santiago de Chile. Avenida Libertador Bernardo O´Higgins n° 3363. Estación Central. Santiago. Chile. [email protected]. 2 Laboratorio de Microbiología Molecular y Biotecnología Ambiental, Universidad Técnica Federico Santa María. 3 Facultad de Ciencias de la Salud, Universidad San Sebastián. El estrés oxidativo es una condición adversa para la célula que se caracteriza por la génesis elevada especies reactivas de oxígeno (ROS) como anión superoxido (O-2), peróxido de hidrogeno (H2O2) y radical hidroxilo (OH•). Estos subproductos tóxicos del oxígeno reaccionan y dañan biomoléculas como proteínas, lípidos y ácidos nucleicos. Para defenderse de los efectos tóxicos de las ROS, los organismos poseen una variedad de sistemas antioxidantes, enzimáticos y no enzimáticos. Entre los componentes enzimáticos se encuentra la enzima superóxido dismutasa que dismuta superóxido a peróxido de hidrógeno, que puede ser descompuesto por catalasas a oxígeno y agua. Por otra parte, entre los metabolitos antioxidantes en bacterias encontramos NADH, NADPH y tioles reducidos, principalmente glutatión (GSH). Este último se encuentra en grandes concentraciones en bacterias Gram negativo, lo que ayuda a mantener un ambiente reductor en el citoplasma. Las ROS pueden ser generadas en forma endógena durante la respiración celular, o también, por agentes externos como condiciones ambientales extremas y agentes oxidantes, entre otros (Adam-Vizi y Chinopoulos, 2006). Entre los agentes oxidantes que generan estrés oxidativo se encuentra el telurito (TeO3
-2). El telurito es un oxianión de teluro (Te) que presenta un alto nivel de toxicidad para organismos eucariotas y procariotas, siendo específicamente más tóxico para bacterias Gram negativo a concentraciones tan bajas como ~4 µM en Escherichia coli (Summers y Silver, 1978). Sin embargo se han detectado microorganismos naturalmente resistentes a este oxianión de teluro. A pesar de esto, aun no es posible dilucidar los mecanismos de resistencia frente a este tóxico. En base a estudios realizados en E. coli se ha sugerido que el TeO3
-2 ingresaría a la célula utilizando el sistema de transporte de fosfatos. Posteriormente, el TeO3
-2 puede ser reducido por enzimas como la nitrato reductasa y/o por glutatión (GSH) u otros tioles celulares. Si bien la oxidación de GSH permitiría contrarrestar el efecto tóxico del telurito, la disminución del tripéptido comprometería seriamente el balance redox celular. Esta reducción de telurito genera la producción de anión superóxido lo que provoca daños a nivel oxidativo, como aumento del contenido de grupos carbonilo en proteínas celulares, generación de sustancias reactivas a ácido tiobarbitúrico (TBARS), inactivación de centros [Fe-S] de enzimas sensibles al estrés oxidativo como aconitasa y aumento de enzimas antioxidantes como superóxido dismutasa (SOD), entre otros (Pérez y col., 2007). El continente Antártico es un lugar cuyas combinaciones de tensiones ambientales extremas son únicas. En este lugar se dan ambientes áridos y las temperaturas más frías de la tierra donde se desarrolla la vida en condiciones singulares. Los microorganismos que sobreviven a estas condiciones están constantemente expuestos a factores ambientales críticos como bajas temperaturas, alta radiación ultravioleta (ambos factores generadores de estrés oxidativo), salinidad alta de los suelos y limitación de nutrientes y agua. Debido a esto, las bacterias han tenido que desarrollar mecanismos de adaptación eficientes para sobrellevar o aminorar los distintos tipos de estrés que enfrentan constantemente.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
353
Es por esto que el territorio Antártico representa un lugar ideal para la búsqueda y aislamiento de bacterias resistentes a telurito y en este contexto es interesante caracterizar las defensas antioxidantes presentes en estos microorganismos, frente a estrés oxidativo generado por telurito y por otros agentes generadores de ROS. En el este trabajo se propone el uso de bacterias aisladas del continente Antártico para buscar en ellas defensas antioxidantes altamente eficientes contra el estrés oxidativo generado por este tóxico. MATERIALES Y METODOS Cepas bacterianas y condiciones de crecimiento. Las cepas utilizadas en este trabajo fueron Stphylococcus haemolyticus (BNF01), Stphylococcus sciuri (BNF05), Psychrobacter immobilis (BNF08), Acinetobacter haemolyticus (BNF15), Psychrobacter immobilis (BNF20), Pseudomonas lini (BNF22) y E. coli BW25113. Las células se crecieron rutinariamente en medio Luria-Bertani (LB) a 25 ºC (BNF08, BNF15, BNF20 y BNF22) y 37 °C (BNF01, BNF05 y E. coli), con agitación constante. El crecimiento se inició inoculando medio LB fresco con diluciones 1:100 de cultivos saturados. En el caso de medio sólido se agregó agar (2 % p/v) y se incubó las placas durante la noche a la temperatura correspondiente. Determinación del área de inhibición del crecimiento. Cultivos saturados de las cepas Antárticas y E. coli se diluyeron en medio LB y se incubaron con agitación a 25 y 37 °C, respectivamente, hasta una DO600 ~0,4. Posteriormente, se sembró homogéneamente 100 µl de cada cultivo en placas de Petri apropiadas. Luego de secar brevemente al aire, se colocó en el centro de cada placa un disco de papel filtro estéril sobre el que se depositó 10 µl del tóxico a ensayar: H2O2 (1 M), K2CrO4 (2 M), CdCl2 (0,4 M) o K2TeO3 (0,4 M). El área de inhibición del crecimiento (cm2) se determinó luego de incubar las placas a la temperatura correspondiente durante 24 h. Determinación de la concentración mínima inhibitoria (MIC). A partir de soluciones stock estériles de K2TeO3, H2O2, CdCl2 o K2CrO4 se realizaron diluciones seriadas en 200 µl de medio LB en placas de cultivo de 96 pocillos. Posteriormente, se agregaron 10 µl de cultivo crecido a temperatura óptima en medio LB hasta un DO600 ~0,4 a cada pocillo y se incubó con agitación a la temperatura correspondiente. La MIC se determinó luego de 24 h de incubación a las temperaturas óptimas correspondientes (25 o 37 °C). Determinación de ROS. Se detectó los niveles intracelulares de ROS total en las cepas antárticas luego de 75 min de exposición a un cuarto de la MIC a telurito, utilizando la sonda fluorescente 2',7'-diclorofluoresceína diacetato (H2DCFDA), según lo descrito por Pérez y cols., (2007). Los análisis fueron llevados a cabo por citometría de flujo. Actividad catalasa (CAT). Se midió la descomposición de peróxido de hidrógeno a 240 nm. La actividad de la enzima se calculó utilizando un coeficiente de extinción molar de 43,6 mM-1 cm-1 (Chen y col., 2003). RESULTADOS Con el fin de evaluar la sensibilidad de estas cepas a telurito, se determinó el área de inhibición de crecimiento (Fig. 1) y la concentración mínima inhibitoria (Tabla 1) a telurito y otros tóxicos generadores de ROS. Utilizando como referencia la cepa sensible de E. coli BW25113. Los resultados de la Fig. 1 muestran que las cepas antárticas son más resistente que E. coli al tratamiento con telurito y cromato. Estos resultados fueron apoyados por los datos de MIC, donde todas las cepas antárticas mostraron niveles altos de resistencia, llegando a ser ~578 veces más resistente en el caso de BNF20 y ~38 veces más resistente en el caso BNF22 a telurito, respecto a E. coli.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
354
MIC (mM)
Cepa K2TeO3 H2O2 CdCl2 K2CrO4
BNF 01 0,8 0,8 0,08 50
BNF 05 1,6 1 0,1 180
BNF 08 0,8 0,175 0,09 15
BNF 15 0,52 0,1 0,02 5
BNF 20 2,32 0,1 0,02 10
BNF 22 0,16 0,3 0,5 10
E. coli 0,008 0,5 0,5 1
Fig. 1. Área de inhibición del crecimiento de las cepas antárticas y E. coli. Se determinó en células expuestas a K2TeO
3
(A) y
K2CrO4 (B). Los datos representan el promedio de 3 determinaciones independientes. Fig. 1. Zone of growth inhibition of the antarctic strains and E. coli exposed to K2TeO
3
(A) and K2CrO4 (B). Data correspond
to the average of 3 independent experiments.
ROS
E. coli
BNF01
BNF05
BNF08
BNF15
BNF20
BNF22
0
20
40
60
80
100 ControlTelurito (1/4 MIC)
% Fluorescencia
Fig. 2. Determinación de ROS totales y actividad catalasa de cepas Antárticas y E. coli. A) determinación de ROS por citometría de flujo en cepas incubadas con telurito (barra negra) y sin telurito (barra blanca). B) Actividad catalasa presente en extractos crudos de células crecidas con telurito (barra negra) y sin telurito (barra blanca). (Fig. 2. Total ROS levels and catalase activity in the antarctic strains and E. coli. A) ROS determination via flow cytometry in cells incubated with and
without tellurite (black and white bars, respectively). B) Catalase activity from crude extracts of cells grown with and without tellurite (black and white bars, respectively).
Los alta resistencia de cepas antárticas al tratamiento con telurito se correlaciono con la baja presencia de ROS que mostraron estas cepas respecto a E. coli (Fig. 2A), a excepción de BNF15. Los niveles de ROS en BNF15 se correlacionan con la extensa fase de latencia que presentaron las curvas de crecimiento de esta cepa al ser tratadas con un cuarto del MIC (no mostrado). Se evaluó uno de los
Tabla 1. MIC a telurito y otros tóxicos para las cepas antárticas y E. coli: Los
datos representan el promedio de 3 determinaciones independientes. No se
observó variación en los valores de MIC obtenidos.
Table 1. MIC of tellurite and other toxicants for the antarctic strains and E. coli. Data correspond to the average of 3
E. coli 2-
3
E. coli + TeO
BNF01 2-
3
BNF01 + TeOBNF05 2-
3
BNF05 + TeOBNF15 2-
3
BNF15 + TeOBNF08 2-
3
BNF08 + TeOBNF20 2-
3
BNF20+ TeO
BNF22 2-
3
BNF22+ TeO
0
20
40
60
140
160
180
200
220
240
B
U/m
g proteins

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
355
sistemas de protección frente a estrés oxidativo en cepas antárticas, mediante la determinación de la actividad catalasa presente en extractos crudos de estas (Fig. 2B). Los resultaron indican que la cepa BNF01, BNF05 y BNF15 presentaron una mayor actividad catalasa respecto E. coli. Las otras cepas no presentaron diferencias significativas respecto a la cepa control. DISCUSION Las condiciones medioambientales extremas que se presentan en el territorio Antártico condiciona el desarrollo de microorganismos con mecanismos antioxidantes eficientes. Esto se evidencia al evaluar la alta resistencia de cepas antárticas expuestas a tóxicos que generan ROS, como telurito (O-2), peróxido de hidrógeno y cromato (OH*) (Tabla 1). De esta manera, se encontró que las cepas BNF01 y BNF05 son altamente resistentes a especies reactivas de oxígeno generadas por telurito, peróxido de hidrógeno y cromato, mientras que no así para cadmio (control de depleción de tioles) (Tabla 1). Estos resultados son apoyados por los bajos niveles de ROS obtenidos y la alta actividades catalasa de los extractos crudos. Por otra parte, las cepas BNF08, BNF15, BNF20 y BNF22 presentaron una baja resistencia a peróxido de hidrógeno con respecto a E. coli, lo que se relaciona con los bajos niveles de actividad catalasa en los extractos crudos. Sin embargo estas cepas presentaron alta resistencia a telurito, lo cual probablemente se deba a mecanismos eficientes de dismutación del anión superóxido. CONCLUSIÓN Las cepas antárticas utilizadas en este trabajo presentaron una alta resistencia a estrés oxidativo generado por telurito. Lo cual probablemente se deba a los mecanismos antioxidantes eficientes presentes en estos microorganismos, que en condiciones naturales están sometidos constantemente a condiciones extremas. AGRADECIMIENTOS. Fondecyt # 1130362 (CCV), FONDECYT Postdoctorado # 1130362 (FAA), INACH ECA48 (2012). REFERENCIAS Adam-Vizi, V. y Chinopoulos, C. 2006. Bioenergetics and the formation of mitochondrial reactive oxygen species. Trends Pharmacology Science 27: 639-645 Chen, G., Patten, C. y Schellhorn, H. 2003. Controlled expression of an rpoS antisense RNA can inhibit RpoS function in Escherichia coli. Antimicrobial Agents Chemotherapy 47: 3485-3493. Pérez, J., Calderón, I., Arenas, F., Fuentes, D., Pradenas, G., Fuentes, E., Sandoval, J., Castro, M., Elías, A. y Vásquez, C. 2007. Bacterial toxicity of potassium tellurite: unveiling an ancient enigma. PLoS ONE 2: e211. Summers, A. O. y Silver, S. 1978. Microbial transformations of metals. Annual Review Microbiology 32: 637-672.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
356
EFECTO DEL MERCURIO EN LA POTENCIACIÓN DE LA RESISTENCIA A TELURITO EN BACTERIAS AISLADAS DEL TERRITORIO ANTÁRTICO CHILENO
(Effect of mercury on the potentiation of tellurite resistance in bacteria isolated from the Chilean Antarctic territory)
Rodríguez F.1, Muñoz P.1, Díaz-Vásquez W.1, 2, 3, Arenas F.1 y C. Vásquez 1
1 Laboratorio de Microbiología Molecular, Facultad de Química y Biología, Universidad de Santiago de Chile, Av. Libertador Bernardo O’Higgins 2206, Santiago, Chile. 2 Laboratorio de Microbiología Molecular y Biotecnología Ambiental, Universidad Técnica Federico Santa María, Av. España 1680, Valparaíso, Chile. 3 Facultad de Ciencias de la Salud, Universidad San Sebastián, Bellavista 7, Santiago, Chile. E-mail: [email protected]
INTRODUCCIÓN El mercurio (Hg) y el telurito (TeO3
-2) son extremadamente tóxicos para la mayoría de los seres vivos y las Concentraciones Mínimas Inhibitorias (MIC) de ambas especies se ubican muy por debajo de los valores publicados para otros metales en Escherichia coli (Nies, 1999). La toxicidad del Hg se asocia a su interacción con tioles celulares, sulfidrilos, tioéteres e imidazol. La resistencia bacteriana a mercurio se relaciona con: i) la adsorción del metal a la membrana externa, ii) la síntesis de proteínas quelantes de metales, iii) procesos de reducción/oxidación enzimáticos y no enzimáticos, iv) mecanismos de precipitación de sulfuros, fosfatos y bicarbonatos metálicos, y v) actividades de metilación y desmetilación (Valls y col., 2002), que han sido muy estudiadas y se relacionan con la transcripción del operón merRTPGCABDE que permite la reducción del Hg+2 (mediada por MerA y MerB) otorgando resistencia a compuestos organomercuriales (Silver y Hobman, 2007). El teluro no presenta elevada toxicidad para los seres vivos. Sin embargo, sus oxianiones telurito (TeO3
-2) y telurato (TeO4-2) son
extremadamente tóxicos para la mayoría de los microorganismos, especialmente bacterias Gram negativo (Taylor, 1999). Parte de la toxicidad del telurito se relaciona con sus características como oxidante provocando la oxidación de tioles celulares como glutatión y desmantelamiento de centros de hierro-azufre [Fe-S]. E. coli y otras bacterias expuestas a telurito exhiben estrés oxidativo, incrementando el contenido de superóxido (O2
.-), evidenciado cuando enzimas lo reducen a teluro (Te0) en presencia de oxígeno y NADH(P)H in vitro (Calderón y col., 2006). La resistencia a telurito ha sido asociada a la actividad de metilasas y enzimas telurito reductasas (Avazeri y col., 1997) y otras enzimas dependientes de NAD(P)H como la catalasa que reducen el telurito a teluro. Se ha observado que la exposición a bajas concentraciones de un tóxico puede incrementar la resistencia al mismo (respuesta adaptativa) o a otro diferente (respuesta cruzada). Un ejemplo de esto es el sinergismo observado entre cadmio/H2O2 en Xanthomonas campestris (Banjerdkij y col. 2005), y paraquat/TeO3
-2, donde E. coli expuesta a bajas concentraciones de paraquat incrementa la resistencia a telurito explicado en parte por la inducción de los operones oxyRS y soxRS de respuesta a estrés oxidativo. En este sentido, una respuesta sinérgica es plausible de encontrarse entre la asociación Hg/TeO3
-2. Con el objetivo principal de obtener una cepa multiresistente a mercurio y telurito, se realizó un aislamiento de bacterias provenientes del territorio antártico chileno proyectos (ECA47 y ECA48) las que fueron expuestas a mercurio y telurito para la búsqueda de un fenotipo potenciador de la resistencia.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
357
MATERIALES Y MÉTODOS Curvas de crecimiento de los tóxicos por separado y en conjunto: Las curvas de crecimiento fueron realizadas en el lector multimodal Tecan Infinite® m200 PRO a 25°C y 150 rpm por 14 h. El crecimiento fue seguido como el cambio en la densidad óptica a 600 nm cada 30 min. Las condiciones fueron LB, LB + TeO3
-2, LB + HgCl2 o LB + TeO3-2 + HgCl2, y los aislados bacterianos, ATH-43 o
ATH-62, o E. coli BW25113. Las concentraciones utilizadas de HgCl2 fueron 8 μM y 40 μM para E. coli y 40 μM para ATH-43 y ATH-62, respectivamente. Potenciación de resistencia en medio líquido: En placas de 48 pocillos se aplicó una concentración de HgCl2 inferior al MIC para los aislados bacterianos ATH-43, ATH-62 y para la cepa de E. coli BW25113 y 8 concentraciones próximas y/o superiores al MIC de telurito, en un volumen final de 1 ml en medio LB. Las concentraciones de telurito utilizadas para los aislados bacterianos fueron 32, 48, 56, 64, 72, 88, 100 y 120 μM y para E. coli fueron 4, 8, 12, 16, 20, 24, 40 y 48 μM. Halos de inhibición de crecimiento bacteriano: Las bacterias fueron crecidas en medio LB suplementado o no con K2TeO3 32 μM o HgCl2 40 μM según el caso hasta fase exponencial (OD600 ~0,3-0,4) a 25 °C. Luego fueron sembradas en placas con medio LB agar suplementada o no con las mismas concentraciones de los tóxicos. Se utilizó sensidiscos a los que se agregó 10 μl de HgCl2 200 mM o 10 μl de K2TeO3 4mM/400mM, según el experimento. Las placas se incubaron a 25 °C por 16 h. El radio de los halos fue medido y las áreas calculadas con un N total de 6 halos por condición. Como test estadístico se utilizó la prueba t (p < 0,01). RESULTADOS Curvas de crecimiento y potenciación de la resistencia en medio líquido: Como se observa en la Fig. 1, el crecimiento de cada cepa y de E. coli en telurito 40 μM es nulo, debido a que esta concentración es superior al MIC. Sin embargo, en presencia de una concentración sub-letal de HgCl2 se observa crecimiento al cabo de aproximadamente 7 h en el caso de los aislados antárticos y 3 h en la curva de E. coli. Estos datos son corroborados por los resultados observados en potenciación de resistencia en medio líquido, en los que se observa crecimiento aun a concentraciones de telurito 120 μM para ATH-62 y -43, y 48 μM para E. coli. Potenciación a la resistencia de telurito mediada por mercurio: Los resultados de la potenciación de la resistencia a telurito mediada por mercurio muestran que en el caso de la cepa ATH-43 hay un claro efecto potenciador de la resistencia a telurito cuando las bacterias son crecidas en placas LB suplementadas con mercurio. Por otro lado, cuando no crecen en un medio con mercurio, no hay efecto potenciador, ya que las áreas de los halos disminuyen de manera drástica (Fig. 2A). De esta manera, sería necesaria la presencia de mercurio de manera constante en el medio de cultivo sólido para exista efecto en la potenciación de la resistencia a telurito (Fig. 2A). En el caso del experimento utilizando el sensidisco con telurito a una concentración de 4 mM se observa que en las placas de LB agar suplementadas con mercurio, prácticamente casi no hay formación de halo de inhibición de crecimiento (Fig. 2A), no así cuando se usa uno de 400 mM (Fig. 2B). Potenciación de la resistencia a mercurio mediada por telurito: Para comprobar si la presencia de telurito en el medio era capaz de potenciar la resistencia a mercurio en las cepas bacterianas aisladas, se procedió a analizar halos de inhibición en distintas condiciones (Fig. 3). Para ambas cepas se ve que no hay un aumento a la resistencia de mercurio mediada por telurito. DISCUSIÓN Los experimentos de potenciación de resistencia a telurito y mercurio de las cepas utilizadas en este estudio muestran que en ambas cepas hay un aumento a la resistencia de telurito cuando las bacterias

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
358
son crecidas en conjunto con mercurio (curvas de crecimiento, MICs y halos). De manera interesante, para las curvas de crecimiento y los MIC, el control con E. coli muestra que también ocurre un fenómeno de potenciación a la resistencia a telurito en presencia de mercurio cuando se usan valores proporcionales a los MIC para mercurio y telurito. Esto indica que el mercurio podría estar gatillando mecanismos de defensa que son comunes con las cepas antárticas los cuales podría incluir aumento del poder reductor de la célula, moléculas antioxidantes, glutatión, tioredoxinas y moléculas con grupos tioles en general que son los blancos celulares tanto del telurito como del mercurio. Además, podría haber un aumento de enzimas con actividad reductasa que podrían reducir telurito, como se ha visto en cepas que son naturalmente resistentes a este tóxico. Una posibilidad interesante es que la enzima reductora de mercurio, MerA, podría estar participando en la reducción de telurito como lo han indicado análisis bioinformáticos de nuestro laboratorio (Morales y cols., no publicado).
Crecimiento ATH-43
0 5 10 150.0
0.5
1.0
1.5ControlHgCl2 40 uM
TeO3-2 40 uMTeO3-2 40 uM + HgCl2 40um
Tiempo (h)
Densidad Optica (600 nm)
Crecimiento E. coli BW25113
0 5 10 15 200.0
0.5
1.0
1.5
2.0ControlHgCl2 8uMTeO3-2 8uMTeO3-2 8 uM + HgCl2 8um
Tiempo (h)
Densidad Optica (600 nm)Crecimiento ATH-62
5 10 15
-0.5
0.0
0.5
1.0
1.5
2.0Control
HgCl2 40 uMTeO3-2 40 uMTeO3-2 40 uM + HgCl2 40um
Tiempo (h)Densidad Optica (600 nm)
a
b
c
Figura 1. Curvas de crecimiento de las cepas ATH-62 (a), ATH-43 (b) y E. coli (c). En LB, en LB + HgCl2, en LB + TeO3
-2 y en LB + TeO3
-2 + HgCl2. Para las 3 cepas se observa crecimiento en telurito a una concentración superior al MIC en presencia de una concentración sub-letal de mercurio. Fig. Growth curves for bacterial isolates ATH-62 (a), ATH-43 (b) and E. coli (c). In LB , LB + HgCl2, LB + TeO3
-2 and LB + TeO3-2 + HgCl2. For the three strains, growth is observed at a tellurite
concentration above the MIC in presence of sublethal mercury concentrations.
0
1
2
3
4
5
6
7ATH-43
Placa LB LB/Hg LB LB/Hg LB LB/Hg
Cultivo LB LB LB/Hg LB/Hg LB LB
Áre
a de
hal
o de
inhi
bici
ón (c
m
)2
Halos de inhibición
* *0
5
10
15
20
ATH-43
*
A B
Figura 2. Cuantificación de áreas del halo de inhibición de crecimiento con telurito 4 mM (A) y 400 mM (B) para ATH-43. Entre el gráfico y las figuras se muestra el contenido de las placas (Placa) y las condiciones de crecimiento en medio líquido
(Cultivo). Hg: mercurio a 40 μM. Abajo, se muestra una placa representativa de cada experimento (n=6, *p<0,01). Figure 2. Quantification of growth inhibition areas with tellurite 4 mM (A) and 400 mM (B) for ATH-43. Between the plot
and the figures the content of the plates (Placa) and the growing conditions in liquid media (Cultivo) are shown. Hg: mercury 40 μM. Below, a representative plate of each experiment is shown (n=6, *p<0,01).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
359
0
2
4
6
8
10
12ATH-62
0
1
2
3
4
5
6
7
8
9ATH-43
Áre
a d
e h
alo
de
inh
ibic
ión
(cm
)
Placa LB LB LB/Te LB/Te LB LB LB/Te LB/Te
Cultivo LB LB/Te LB LB/Te LB LB/Te LB LB/Te
A B
*
*
*
2
Figura 3. Cuantificación de áreas del halo de inhibición de crecimiento con mercurio 200 mM para ATH-43 y ATH-62. Bajo de los gráficos se muestra el contenido de las placas (Placa) y las condiciones de crecimiento en medio líquido (Cultivo). Te:
telurito 32 μM (n=6, *p<0,01). Figure 3. Quantification of growth inhibition areas with mercury 200 mM for ATH-43 and ATH-62. Below the plots the
content of the plates (Placa) and the growing conditions in liquid media (Cultivo) is shown. Te: tellurite 32 μM (n=6, *p<0,01).
Cepa MIC de Telurito (µM) MIC de Telurito (µM) + Mercurio 40 (µM)
ATH-43 32 120
ATH-62 32 120
E. Coli BW25113 4 >48
Tabla 1. Concentraciones mínimas inhibitorias (MICs) de telurito en presencia de mercurio para las cepas ATH-43, ATH-62 y E. coli .
Table 1. Minimal inhibitory concentrations (MICs) of tellurite in the presence of mercury for the strains ATH-43, ATH-62 and E. coli.
CONCLUSIONES A partir de los resultados se concluye que, tanto en las cepas antárticas como en E. coli, existe un aumento en la resistencia a telurito mediada por la presencia de mercurio en el medio de cultivo. AGRADECIMIENTOS. Fondecyt # 1130362, Fondecyt Postdoctorado # 1130362, Beca Apoyo a la realización de Tesis Doctoral 24121087. REFERENCIAS BIBLIOGRÁFICAS Avazeri, C., Turner, R. J., Pommier, J., Weiner, J. H., Giordano, G. y Vermeglio A. 1997. Tellurite reductase activity of nitrate reductase is responsible for the basal resistance of Escherichia coli to tellurite. Microbiology, 143: 1181-1189 Banjerdkij, P., Vattanaviboon, P. y Mongkolsuk, S. 2005. Exposure to cadmium elevates expression of genes in the oxyR and ohrR regulons and induces cross-resistance to peroxide killing treatment in Xanthomonas campestris. Applied and Environmental Microbiology, 71: 1843-1849 Calderón, I. L., Arenas, F. A., Perez, J. M., Fuentes, D. E., Araya, M. A. y Vásquez C.C. 2006. Catalases are NAD(P)H-dependent tellurite reductases. PLoS ONE, 1: e70 Nies, D. H. 1999. Microbial heavy-metal resistance. Applied Microbiology and Biotechnology, 51: 730-750 Silver, S. y Hobman, J. 2007. Mercury Microbiology: Resistance Systems, Environmental Aspects, Methylation, and Human Health. In Molecular Microbiology of Heavy Metals, 6: 357-370. Taylor, D. E. 1999. Bacterial tellurite resistance. Trends Microbiology, 7: 111-115 Valls, M., Fau - de Lorenzo, V. y de Lorenzo, V. 2002. Exploiting the genetic and biochemical capacities of bacteria for the remediation of heavy metal pollution. FEMS Microbiology Reviews, 26: 327-338

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
360
CARACTERIZACIÓN METABÓLICA Y DE ESTRUCTURA DE MEMBRANA DE AISLADOS BACTERIANOS ANTÁRTICOS CAPACES DE SINTETIZAR QUANTUM DOTS. (Metabolic and Membrane Structure Characterization of Antarctic Bacterial Strain Able to Synthesize
Quantum Dots)
Soto, C.1, Farfán, J.1, Salinas, D.1, Venegas, D.1, Hoare, A.1, Bravo, D.1 y Pérez-Donoso, J.2
1 Laboratorio de Microbiología Oral, Facultad de Odontología, Universidad de Chile, Sergio Livingstone Pohlhammer 943, Independencia, Santiago de Chile. E-mail: [email protected] 2 Laboratorio de Bionanotecnología y Microbiología, Facultad de Ciencias Biológicas, Universidad Andrés Bello, Avenida República 239, Santiago Centro, Santiago de Chile. Los Quantum Dots (QD) son nanopartículas fluorescentes, compuestos principalmente de metales pesados como Cadmio, Selenio o Telurio, utilizadas en las industrias de la tecnología mediante la generación de celdas fotovoltaicas y medicina en el área de la imagenología. Sus diversas cualidades, principalmente la de absorber luz y emitirla en forma de fluorescencia, las han hecho populares en los últimos años. El aumento en la necesidad de generar QD ha logrado que se generen nuevas formas de síntesis que reemplacen las técnicas químicas actuales por otras menos tóxicas, más sencillas y menos costosas (Monrás y col., 2012). En esta búsqueda, se sabe que ciertas bacterias son capaces de sintetizar QD naturalmente al ser expuestas a condiciones de crecimientos específicas. Estas bacterias estas caracterizadas normalmente por poseer mecanismos que las hacen resistentes a altas concentraciones de metales pesados y a condiciones adversas del medio ambiente (alta radiación UV, pH, etc…).
Prueba Resultados
Citrato + : La bacteria metaboliza citrato - : La bacteria metaboliza citrato
Urea + : La bacteria posee ureasas
- : La bacteria no posee ureasas
MIO
Movilidad + : La bacteria es mótil
- : La bacteria no es mótil
Indol + : La bacteria genera indol
- : La bacteria no genera indol
Ornitina + : La bacteria metaboliza ornitina
- : La bacteria no metaboliza ornitina
TSI Solo glucosa: La bacteria sólo metaboliza glucosa
Fermentador Total : La bacteria metaboliza glucosa, lactosa y sacarosa - : La bacteria no metaboliza los azúcares
LIA
Descarboxilación de Lisina + : La bacteria descarboxila la lisina
- : La bacteria no descarboxila la lisina
Desaminación de Lisina + : La bacteria desamina la lisina
- : La bacteria no desamina la lisina Producción Ácido
Sulfhídrico + : La bacteria produce ac. sulfhídrico
- : La bacteria no produce ac. sulfhídrico
Hidrólisis de Gelatina + : La bacteria hidroliza gelatina
- : La bacteria no hidroliza gelatina
Glucosa Campana + : La bacteria fermenta glucosa
- : La bacteria no fermenta glucosa
Catalasa + : La bacteria posee catalasas
- : La bacteria no posee catalasas
Tabla 1: Resumen de los resultados posibles para cada prueba bioquímica realizada. Las convenciones aquí utilizadas de cada resultado serán utilizados en las tablas 2 y 3. (Table 1: Summary of possible results for each biochemical test performed.
Conventions used here will be used for each result in table 2 and 3).
Es por esto que se ha postulado que las bacterias capaces de sobrevivir en medio antárticos son buenos candidatos para la producción de QD. Se tienen diversas cepas bacterianas obtenidas en la Antártida bajo diferentes condiciones las cuales se crecieron y aislaron en nuestro laboratorio. Estos distintos

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
361
aislados necesitan primero ser caracterizados para poder ser utilizados en la producción de QD y lograr hacer de aquel proceso lo más eficaz posible. Con este fin, se realizaron diversas pruebas bioquímicas para conocer sus metabolismos y poder generar condiciones de crecimiento específicas y eficientes para generar QD. En este punto, se realizaron las siguientes pruebas bioquímicas (Tabla 1) con el fin de caracterizar el metabolismo de los 10 aislados bacterianos utilizados en el estudio de la forma más completa posible: LIA (Lisine Iron Agar), Simmons Citrato, TSI (Triple Sugar-Iron Agar), MIO (movilidad, indol y ornitina), ureasa, catalasa, glucosa campana y hidrólisis de gelatina. RESULTADOS Los resultados nos indicaron que las distintas bacterias poseían metabolismos muy diferentes entre ellas (Tablas 2 y 3), incluso las que al ser sometidas a identificación por secuenciación del RNAr 16s resultaron pertenecer a un mismo género (Tabla 4).
Cepa Citrato Urea MIO (Movilidad) MIO (Indol) MIO (Ornitina) TSI
6 - + + + - Fermentador Total 10 + + - - + Solo glucosa 12 - + + - + Solo glucosa 23 + + - - + Solo glucosa 37 - + + - - Solo glucosa 38 + - - - + - 83 + - - - - - 197 + + - - + Solo glucosa 198 + + + + + Fermentador Total 212 - - - - - -
Tablas 2 y 3:
Resumen de los resultados de las distintas pruebas bioquímicas de los aislados bacterianos. El detalle de cada resultado se encuentra descrito en la tabla 1.
Table 2 & 3: Summary of the results of Biochemical Test for all bacterial isolates. The detail for each result is described in table 1.
Cepa Resultado Secuenciación RNAr 16s 6 Yersinia (kristensenii)
10 Pseudomonas (fragi)
12 Psychrobacter (nimivaris)
Cepa LIA (Descarboxilación
de la lisina) LIA (Desaminación
de la lisina) LIA (Producción de ácido sulfhídrico)
Hidrólisis de Gelatina
Glucosa campana
Catalasa
6 - - - - + + 10 + - - - - + 12 + - - - + + 23 - - - - + + 37 + - - - + + 38 + - - - - + 83 + - - - - + 197 - - - - + + 198 - - - - + + 212 + - - - - +

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
362
23 Stenothrophomonas (rizofila)
37 Arthrobacter (psychrophenolycus)
38 Pseudomonas (fragi)
83 Pseudomonas (fragi)
197 Pseudomonas (fragi)
198 Pseudomonas (lini)
212 Pseudomonas (fragi)
Tabla 4: Resultados de la secuenciación del RNAr 16s de los distintos aislados bacterianos. Table 4: Sequencing results of the 16S rRNA of the bacterial isolates.
Además, caracterizamos un componente importante de membrana en bacterias Gram negativo, el lipopolisacárido (LPS), para ver si esta estructura confiere algún tipo de resistencia diferencial frente a diversos componentes antimicrobianos con efecto en la membrana bacteriana. El LPS se caracteriza por ser un factor de virulencia importante en muchas bacterias y un componente importante de la membrana. Está compuesto de 3 regiones: El lípido A anclado a la membrana, un Core de azúcares, y el antígeno O el cual es la repetición sucesiva de una unidad de azucares. Para estos efectos se purificó el LPS de las distintas cepas bacterianas y se visualizó en geles de poliacrilamida; además, relacionamos los distintos perfiles obtenidos con la resistencia diferencial a distintos antimicrobianos mediante ensayos de sensidiscos (antibiograma). El gel de poliacrilamida entregó que los aislados poseían perfiles de LPS distintos entre sí, incluso alguno careciendo de la porción más externa de la estructura, el antígeno O (Bandeo Superior) (Figura 1).Estos perfiles fueron comparados con los resultados del antibiograma realizado con los compuestos EDTA y SDS (desestabilizadores de membrana), Novobiocina (inhibidor de la DNA girasa que aprovecha la desestabilización de la membrana para entrar al espacio intracelular) y Polimixina B (Péptido catiónico capaz de unirse a regiones cargadas de la membrana como el LPS para desestabilizar la membrana bacteriana) (Tabla 5).
Figura 1: Perfiles electroforéticos de LPS para los distintos aislados. La primera banda de cada perfil corresponde al lípido A, la segunda al lípido A + la región Core, la tercera al lípido A + la región Core + la primera porción de antígeno O, y así
sucesivamente agregando porciones de antígeno O. (Figure 1: LPS electrophoretic profiles for the bacterial isolates. First band of each profiles corresponds to lipid A, the second to lipid A + core region, the third to lipid A + core region + first
portion of O antigen and so on adding portions of O antigen.).
Diámetro halo de inhibición (mm)
Cepa EDTA 0,5M Novobiocina
25ug/ml Polimixina B 275 ug/ml
SDS 10%
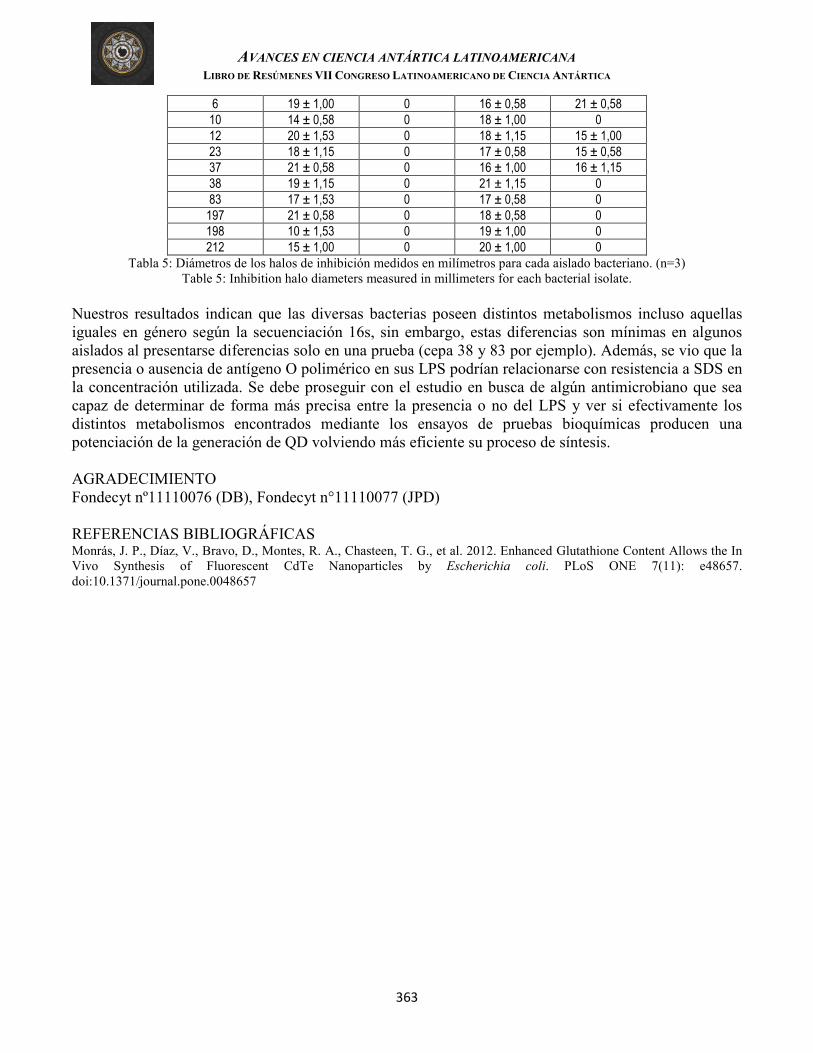
AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
363
6 19 ± 1,00 0 16 ± 0,58 21 ± 0,58 10 14 ± 0,58 0 18 ± 1,00 0 12 20 ± 1,53 0 18 ± 1,15 15 ± 1,00 23 18 ± 1,15 0 17 ± 0,58 15 ± 0,58 37 21 ± 0,58 0 16 ± 1,00 16 ± 1,15 38 19 ± 1,15 0 21 ± 1,15 0 83 17 ± 1,53 0 17 ± 0,58 0
197 21 ± 0,58 0 18 ± 0,58 0 198 10 ± 1,53 0 19 ± 1,00 0 212 15 ± 1,00 0 20 ± 1,00 0
Tabla 5: Diámetros de los halos de inhibición medidos en milímetros para cada aislado bacteriano. (n=3) Table 5: Inhibition halo diameters measured in millimeters for each bacterial isolate.
Nuestros resultados indican que las diversas bacterias poseen distintos metabolismos incluso aquellas iguales en género según la secuenciación 16s, sin embargo, estas diferencias son mínimas en algunos aislados al presentarse diferencias solo en una prueba (cepa 38 y 83 por ejemplo). Además, se vio que la presencia o ausencia de antígeno O polimérico en sus LPS podrían relacionarse con resistencia a SDS en la concentración utilizada. Se debe proseguir con el estudio en busca de algún antimicrobiano que sea capaz de determinar de forma más precisa entre la presencia o no del LPS y ver si efectivamente los distintos metabolismos encontrados mediante los ensayos de pruebas bioquímicas producen una potenciación de la generación de QD volviendo más eficiente su proceso de síntesis. AGRADECIMIENTO Fondecyt nº11110076 (DB), Fondecyt n°11110077 (JPD) REFERENCIAS BIBLIOGRÁFICAS Monrás, J. P., Díaz, V., Bravo, D., Montes, R. A., Chasteen, T. G., et al. 2012. Enhanced Glutathione Content Allows the In Vivo Synthesis of Fluorescent CdTe Nanoparticles by Escherichia coli. PLoS ONE 7(11): e48657. doi:10.1371/journal.pone.0048657

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
364
EFECTO DE EXTRACTO DE LÍQUENES EN LA FORMACIÓN DE BIOPELÍCULAS DEL PATÓGENO DE PECES VIBRIO ANGUILLARUM
(Lichens extract effect in the biofilms formation of fish pathogen Vibrio anguillarum)
Torres, C.1, Jopia, P.1, Urrutia, H.1, Casanova-Katny, A2
1Laboratorio de Biopelículas y Microbiología Ambiental, Centro de Biotecnología, U. de Concepción, Concepción. CHILE. [email protected] 2Laboratorio de Investigación en Agentes Antibacterianos, Facultad de Ciencias Biológicas, U. de Concepción, Concepción. INTRODUCCIÓN Vibrio anguillarum es una importante bacteria patógena de peces, responsable de la vibriosis, enfermedad grave que afecta a una gran variedad de peces y causa grandes pérdidas económicas en la acuicultura Chilena (Austin y Austin, 1993). Biopelículas formadas por cepas patógenas de Vibrio representan serios problemas para la acuicultura (Wang y col., 1998). Para proporcionar una estrategia para la prevención, control y erradicación de las biopelículas, un enfoque prometedor para combatir infecciones, independientes del control convencional con antibióticos, es el mecanismo de comunicación usado por las bacterias para establecer una biopelícula, el llamado quorum sensing (QS) (Jayaraman y Word, 2008), utilizado por muchas especies bacterianas para coordinar su expresión de genes en respuesta a la densidad celular, permitiendo a las células individuales comportarse como una comunidad (Miller y Bassler, 2001). Esto lo logra mediante la producción, liberación y la detección de pequeñas moléculas de señal (Nealson y Hastings, 1979). La detección del QS puede afectar en la ecología y fisiología de la bacteria, así como su patogenicidad (Buch y col., 2003). La alteración de este sistema se ha reconocido como una nueva estrategia anti-infecciosa (Finch y col., 1998). Además, se ha demostrado que la inactivación del quórum en el patógeno puede resultar en una disminución significativa de la expresión de factores de virulencia (Hentzer y col., 2003), donde varios estudios han sugerido que la biopelícula puede ser importante para la supervivencia, virulencia y resistencia en las especies de Vibrio (Faruque y col., 2006). Líquenes y metabolitos secundarios de líquenes se han utilizado en la medicina tradicional hace siglos. Metabolitos liquénicos presentan una amplia variedad de actividades biológicas incluyendo actividad antibiótica, antibacteriana, antitumoral, antimutagénica, anti-viral, entre otras (Huneck y Yoshimura 1996; Johnson y col., 2011). Aunque estas múltiples actividades de metabolitos de líquenes han sido reconocidas, su potencial todavía no se ha explorado completamente. Líquenes Antárticos han permanecido inexplorados, por lo que esta investigación se ha realizado con el objetivo de develar el efectos de sus extractos frente el sistema de comunicación bacteriano, dependiente de la densidad celular (quórum sensing) y formación de biopelículas del patógeno de peces Vibrio anguillarum. MATERIALES Y MÉTODOS Se trabajó con ocho especies de líquenes recolectadas desde la Antártica. Se realizo un protocolo simple de recubrimiento con agar soft, basado en la inhibición de la pigmentación violeta y producción de pigmentación azul de dos cepas reporteras Chromobacterium violaceum 12472 y Agrobacterium tumefaciens NTL4, con el fin de obtener una detección de la presencia potencial de moléculas con capacidad de inhibir el sistemas quórum sensing y detección de AHL (acil homoserina lactona, molécula señal) respectivamente de cada una de las especies de líquenes y de sus extractos acétonicos, metanólico, etanólicos y acuosos. Los extractos que presentaron actividad inhibitoria QS y detección de AHL fueron utilizados en el en ensayos de formación específica de biopelículas (SBF) para evaluar su

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
365
efecto en un la inhibición de la formación de biopelículas de V. anguillarum 3276, los datos proporcionados del índice de formación específica de biopelículas fueron analizados estadísticamente mediante el análisis estadístico de varianza (ANOVA). RESULTADOS En los ensayos para la detección de AHL frente a la cepa reportera Agrobacterium tumefaciens NTL4 (pZLR4) los líquenes Sterocaulon alpinum, Sphaerophus globosus, Usnea aurantiaco atra, Himantormia lugubris, Caloplaca sp. podrían producir en su superficie metabolitos con estructura similar a AHL detectado mediante la aparición de un halo de coloración azul alrededor se los cortes de líquenes. Para la determinación de la actividad inhibitoria de quórum sensing (IQS), Usnea antarctica, Sphaerophus globosus, Himantormia lugubri, Sterocaulon alpinum, Usnea aurantiaco-atra, presentaban en su superficie metabolitos con actividad inhibidora de QS, debido a que fueron capaces de inhibir el sistema de quórum sensing de C. violaceum, detectado en todos los casos mediante la pérdida de coloración violeta alrededor de los cortes de líquenes. Se midió el efecto IQS de extractos de líquenes mediante la medición del halo de inhibición de pigmentación violeta. Un total de 11 extractos de líquenes fueron capaces de inhibir el sistema de quórum sensing del reportero C. violaceum, detectado en todos los casos mediante la pérdida de coloración violeta alrededor de los discos que contenían los extractos, los resultados se presentan en la Tabla 1.
Muestras de Líquenes Antárticos
Halos de Inhibición (mm) Extracto Metanólico Extracto Etanólico Extracto Acuoso Extracto Acetónico
Sterocaulon alpinum 10 - - 15 Caloplaca sp - - - -
Sphaerophus globosus 16 18 - 20 Usnea aurantiaco atra - - - 15
Usnea antartica - 12 - 20 Himantormia lugubris 13 15 - 25
Tabla 1. Inhibición QS de distintos extractos de líquenes Antárticos (0,03g/ml) frente a Chromobacterium violaceum 12472. Table 1. QS inhibition different Antarctic lichens extracts (0.03 g / ml) against Chromobacterium violaceum 12472
En la formación de biopelículas de V. anguillarum se probaron de los distintos extractos de las especies de líquenes que presentaron actividad IQS. Mediante este ensayo SBF se observó, que todos los extractos a excepción del extracto acetónico de Sterocaulon alpinum, fueron capaces de disminuir la formación de biopelícula del patógeno V. anguillarum. Los extractos de líquenes que mostraron una mayor disminución de la biopelícula de patógeno de peces V. anguillarum con respecto al control fueron Himantormia lugubris (Figura 1) y Usnea antarctica (Figura 2).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
366
Figura1: Método SBF, formación de biopelícula del patógeno V. anguillarum 3276 frente a distintos extractos de Himantormia lugubris. *Diferencia significativa con respecto al control (P<0.05 ANOVA). SBF method, biofilm formation of pathogenic V. anguillarum 3276 against different extracts Himantormia lugubris. * Significant difference from control (P <0.05 ANOVA).
Figura 2 Método SBF, formación de biopelícula del patógeno V. anguillarum 3276 frente a distintos extractos de Usnea antarctica. *Diferencia significativa con respecto al control (P<0.05 ANOVA). SBF method, biofilm formation of pathogenic V. anguillarum 3276 against different extracts Usnea antarctica. * Significant difference from control (P <0.05 ANOVA).
DISCUSIÓN Y CONCLUSIÓN Mediante el estudio de producción de AHL se detectó que 5 de 8 especies de líquenes Antárticos podrían producir en su superficie metabolitos con estructura similar a AHL, debido a la respuesta del ensayo con la cepa reportera, sin embargo se debe dejar claro que pueden existir fallas en la detección de estas moléculas, pues pueden ser producidas, pero no alcanzar los umbrales de detección de las bacterias reporteras utilizadas (Cha y col., 1998). El hallazgo de estas cinco especies de líquenes productores de AHL es de importancia biológica, ya que se ha demostrado que la detección de AHL puede representar una estrategia de competencia al actuar como moléculas antagonistas, que impiden la comunicación celular y por lo tanto la generación del quórum (Stickler y col., 1998). Dado que las actividades nocivas de un número de plantas y patógenos de animales están regulados por sistemas asociados a AHL (Fuqua y col., 1994), la capacidad de líquenes para inhibir los sistemas de AHL también puede tener importantes aplicaciones biotecnológicas. En la búsqueda de líquenes con actividad inhibitoria del sistema QS, se detectaron 11 extractos fueron capaces de inhibir el sistema de quórum sensing del biosensor, teniendo fuertes efectos inhibitorios, por lo tanto estos compuestos pueden desempeñar un papel importante en la interacción microorganismos-liquen (Selbmann y col., 2009). Por otra parte, la evidencia hasta la fecha sugiere que el crecimiento lento y las difícil condiciones ambiéntale a las cuales se ven sometido los líquenes en su ambiente natural, hacen que la producción de metabolitos sea una necesidad para ellos, donde muchos componentes secundarios presentan diversas actividad biológicas en protección a ataque de microorganismos (Rankovic y col., 2008). De los extractos acuoso no se observó actividad inhibitoria del sistema QS (IQS), esto es debido a que los componentes activos producidos por los líquenes son insolubles o poco solubles en agua (Baral y Maharjan, 2011). A partir de los extractos de líquenes que presentaron actividad IQS se evaluó el efecto de éstos, sobre en la formación de biopelículas de V. anguillarum, se observó que todos los extractos de líquenes fueron capases de disminuir en menor o mayor grado la formación de biopelícula de V. anguillarum. Esta inhibición puede ser explicada debido a que, la formación de biopelículas de éste patógeno está regulada por el sistema QS (Weber y col., 2008). Por lo que lo extractos podrían estar actuando mediante la inhibición de la comunicación

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
367
celular, de este modo la actividad IQS representa un potencial biocontrol de biopelículas (Steinberg y col., 1997). Mediante este estudio se observó una relación entre la actividad inhibitoria del sistema quórum sensing y la inhibición de la formación de la biopelícula del patógeno de peces V. anguillarum desde extractos de líquenes, resultados que probablemente están asociados a la amplia variedad de actividad biológica descrita en ellos, la cual esta generalmente correlacionado a sus circunstancias ecológicas especiales. AGRADECIMIENTOS Proyectos P_06-11(2012); Apoyo de tesis de pregrados en temática Antártica INACH. REFERENCIAS BIBLIOGRÁFICAS Austin, B. y Austin, D. 1993. Bacterial Fish Pathogens: Disease on Farmed and Wild Fish, 2nd edn. Ellis Horwood, Ltd., 111–171 pp. Baral, B. y Maharjan, B. L. 2011. Assessment of antimicrobial and phytochemical potentials of high altitudinal Nepalese lichens. Journal of Mycrobiology, Biotechnoloy and Food Sciences 1: 98-112 Buch, C., Sigh, J., Nielsen, J. y Gram, L. 2003. Production of acylated homoserine lactones by different serotypes of Vibrio anguillarum both in culture and during infection of rainbow trout. Systematic and Applied Microbiology, 26: 338–349. Cha, C. P. G. O, Chen, Y., Shaw, P. y Farrand, S. 1998. Production of acyl-hommoserine lactone quorum-sensing signals by Gram-negative plant-associated bacteria. MPMI 1: 1110-1129. Faruque, S., Biswas, K., Uddern, S., Ahmad, Q., Sack, D., Nair, G. y Mekalanos. J. 2006. Transmissibility of cholera: in vivo-formed biofilms and their relationship to infectivity and persistence in the environment. Proceedings of the National Academy of Sciences 103: 6350–6355. Finch, R., Pritchard, D., Bycroft, B., Williams, P. y Stewart, G. 1998. Quorum sensing: a novel target for anti-infective therapy. Journal of Antimicrobial Chemotherapy, 42: 569–571. Fuqua, W. C., Winans, S. C. y Greenberg, E. P. 1994. Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. Journal of Bacteriology. 176:269–275. Hentzer, M., Andersen, J., Riedel, K., Rasmussen, T., Bagger, N., Kumar, N., Schembri, M., Song, Z., Kristoffersen, P., Manefield, M., Costerton, J., Molin, S., Hoiby, N. y Givskow, M. 2003. Attenuation of Pseudomonas aeruginosa virulence by quórum sensing inhibitors. The EMBO Journal, 22: 3803–3815. Huneck, S. y Yoshimura, I.. 1996. Identificación of lichen subtances. Springer, Berlin Heidelberg New York. 493 pp. Jayaraman, A. y Wood, T. 2008. Bacterial Quorum Sensing: Signals, Circuits, and Implications for Biofilms and Disease. Annual Review of Biomedical Engineering, 10: 145-167. Jhonson, C. J., Bennett, J. P., Biro, S. M. Duque-Velasquez, J. C. Rodríguez, C. M., Bessen, R. A. y Rockel, T. E. 2011. Degradation of the disease- associated prion protein by a serine protease from lichens. PloS One., 6: 9836- l9836. Miller, M. B. y Bassler, B. L. 2001. Quorum sensing in bacteria. Annual Review of Microbiology, 55:165-199. Nealson, K. y Hastings, J. 1979. Bacterial bioluminescence: its control and ecological significance. Microbiology Reviews, 43: 496-518. Rankovic, B., Misjic, M. y Sukdolak, S. 2008. Antimicrobial activity of substances derived from the lichens Physcia aipolia, Umbilicaria polyphylla, Parmelia caperata and Hypogymnia physodes. World Journal Microbiology Biotechnology, vol. 24: 1239-1242. Selbmann, L., Zucconi, L., Ruisi, S., Grube, M., Cardinale, M. y Onofroi, S. 2009. Culturable bacteria associated whit Antarctic lichens: affiliation and psychrotolerance. Polar Biology. 33:71-83 Steinberg, P., Schneider, R. y Kjelleberg, S. 1997. Chemical defenses of seaweeds against microbial colonization. Biodegradation, 8: 211-220. Stickler, D. J., Morris, N. S. Mclean, R. J. y Fuqua, C. 1998. Biofilms on indwelling urinary catheters produce quorum-sensing signal molecules in situ and in vitro. Applied and Environmental Microbiology 64, 3486– 3490. Wang, X. H., Oon, H. L. Ho, G. W. P. Wong, W. S. F. Lim, T. M. y Leung, K. Y.1998. Internalization and cytotoxicity are important virulence mechanisms in vibrio–fish epithelial cell interactions. Microbiology, 144: 2987-300.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
368
MICROFLORA BACTERIANA CULTIVABLE ASOCIADA AL TRACTO DIGESTIVO DEL BACALAO DE PROFUNDIDAD
(Culturable microbiota along the digestive tract of Patagonian toothfish)
Urtubia, R.1, 2, Gallardo, P.3, y M.2 González.
1Magíster en Ciencias, Facultad de Ciencias, Universidad de Magallanes. Punta Arenas, Chile. [email protected]; 2Laboratorio de Biorrecursos Antárticos, Departamento Científico, Instituto Antártico Chileno. Punta Arenas, Chile; 3Centro de Cultivos Marinos Bahía Laredo, Facultad de Ciencias, Universidad de Magallanes. Punta Arenas, Chile.
INTRODUCCIÓN El bacalao de profundidad (Dissostichus eleginoides) es un pez que se distribuye en aguas del océano Austral como también en la zona subantártica (Belchier y Collins, 2008). Debido a su importancia económica y a los intentos de producción en cautiverio se hace relevante conocer las características microbiológicas en su tracto digestivo, para evaluar el posible impacto del cambio de dieta. El tracto digestivo de peces es colonizado por un ensamble complejo de microorganismos que favorecen al desarrollo del individuo. Se distinguen tres funciones principales de la microflora intestinal: a) funciones de nutrición y metabolismo, como resultado de la actividad bioquímica de la flora; b) funciones de protección, previniendo la invasión de microorganismos patógenos y c) funciones tróficas sobre la proliferación y diferenciación del epitelio intestinal, y sobre el desarrollo y modulación del sistema inmunológico (Guarner y Malagelada 2003). En la actualidad no existe información respecto a la flora normal presente en este organismo y son escasos los reportes de las comunidades bacterianas cultivables asociadas al tracto digestivo de peces antárticos y subantárticos. El objetivo de este trabajo es realizar una primera caracterización de la microflora cultivable, propia de las tres zonas del tracto digestivo del bacalao de profundidad: estómago, intestino medio e intestino posterior. MATERIALES Y MÉTODOS Obtención de muestras Las muestras fueron obtenidas del centro de cultivo de Bahía Laredo, en donde se extrajo el tracto digestivo del bacalao de profundidad. Este se dividió en 3 secciones: estomago, intestino medio e intestino posterior. Con estas secciones se realizaron los cultivos bacteriológicos en distintos medios selectivos. Cultivo de la microflora bacteriana Cada sección de tejido se frotó en 3 medios de cultivo sólido: R2A, AM (Agar Marino) y TCBS (Tiosulfato Citrato Bilis Sucrosa), siendo este último medio selectivo para el aislamiento y cultivo de bacterias del género Vibrio. Estas placas fueron incubadas a 10°C hasta que se observó crecimiento de colonias donde luego fueron purificadas hasta obtener cultivos puros. Aislamiento de cepas bacterianas A partir de las distintas morfologías identificadas, se aislaron colonias representativas desde los medios nutritivos sólidos. Extracción de material genético y RFLP A las cepas seleccionadas se les realizó extracción de ADN genómico mediante el método de fenol-cloroformo. Se amplificó el gen 16S ADNr, mediante PCR con partidores universales 16S Fw (5`-AGAGTTTGATCCTGGCTCAG-3´) y 16S Rv (5´-CGGTTACCTTGTTACG ACTT-3). Con el objeto de identificar cepas bacterianas distintas, se realizó el protocolo de RFLP (Restriction Fragment Length Polymorphisms) sobre los productos de PCR del gen 16S ADNr, utilizando tres enzimas de restricción:

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
369
Rsa I, Alu I y BsuR I (Invitrogen), según las instrucciones del fabricante. De cada grupo de cepas que presentó un mismo patrón de corte, se escogió una o varias colonias para secuenciar el gen ribosomal 16S. Identificación de las cepas Se realizó una búsqueda de homologías a través de la base de datos BLAST en la página del NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi) para realizar la asignación taxonómica a nivel de género. RESULTADOS Y DISCUSIÓN Se aislaron desde las tres zonas del tracto digestivo 113 cepas, 34 correspondieron a la zona del estómago, 28 al intestino medio y 51 al intestino posterior. Del total de cepas aisladas se secuenciaron 70, de estas, 21 pertenecen a la zona del estómago, 22 a la zona del intestino medio y 27 a la zona del intestino posterior. En la zona del estómago se encontraron bacterias pertenecientes a cuatro familias: Micrococcaceae (5%), Pseudomonadaceae (5%), siendo Vibrionaceae (52%) y Moraxellaceae (38%) las más abundantes. En la zona del intestino medio se observaron sólo dos tipos de familias: Moraxellaceae (55%) y Vibrionaceae (45%); finalmente en la zona del intestino posterior se encontró el mayor número de especies diferentes, encontrando cepas pertenecientes a 6 familias distintas que correspondieron a: Vibrionaceae (37%), Microbacteriaceae (19%), Moraxellaceae (19%), Sphingomonadaceae (15%), Xanthomonadaceae (4%) y Planococcaceae (4%) (Figura 1). Estos resultados nos indican que en la sección del intestino posterior existe una mayor diversidad filogenética. Según distintos estudios sobre la microflora asociada al tracto digestivo de especies marinas, es frecuente encontrar bacterias de la clase Gamaproteobacteria (Ward y col., 2009); en este estudio, la clase Gamaprotebacteria presenta una alta dominancia en todo el tracto digestivo del bacalao. En peces nototenoideos antárticos, Ward y colaboradores (2009) han observado una mayor prevalencia en bacterias asociadas al género Photobacterium y Vibrio. Las bacterias aisladas en D. eleginoides se diferenciarían de nototénidos antárticos por una mayor presencia de Psychrobacter, Vibrio y Aliivibrio. Consistente con lo anterior, en el estómago de bacalao Vibrio y Psychrobacter representaron un 52 y 33%, respectivamente. En la zona posterior del intestino, se encontró la mayor variedad, identificando 9 géneros, donde la mayor predominancia correspondió a los géneros de Aliivibrio (18%), Psychrobacter (18%) y Vibrio (18%) (Figura 2). Cepas pertenecientes a los géneros Pseudomonas, Aliivibrio y Vibrio han sido aisladas desde dos peces nototénidos antárticos Notothenia coriiceps y Chaenocephalus aceratus (MacCormack y Fraile, 1990; Ward y col., 2009), lo que concuerda con la presencia de estas cepas en el tracto digestivo del bacalao. Sin embargo, no se observó la presencia de los géneros Photobacterium, Ehrlichia y Neorickettsia que serían característicos de peces aguas frías. Los géneros Sphingomonas y Rhodoglobus han sido aislados en glaciares antárticos, lo que confirma que esta especie es de ambientes fríos. CONCLUSIONES La mayor diversidad filogenética de bacterias asociadas al tracto digestivo se encuentra en la sección posterior del intestino del bacalao de profundidad. Las cepas bacterianas que dominaron el tracto digestivo son del género Vibrio y Psychrobacter, y estas corresponden a una microflora intestinal normal.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
370
Figura 1: Microflora cultivable aislada en tres secciones del tracto digestivo: estómago, intestino medio e intestino final;
según la clase y familia de las bacterias secuenciadas. Figure 1: Cultivable microflora isolated in three sections of the digestive tract: stomach, small intestine middle and end,
according to the family and class sequenced bacteria.
Figura 2: Microflora cultivable aislada en tres secciones del tracto digestivo, según su género. (A) estómago, (B) intestino
medio e (C) intestino posterior. Figure 2: Cultivable microflora isolated in three sections of the digestive tract, according to their kind. (A) stomach, (B)
midgut, and (C) hindgut.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
371
AGRADECIMIENTOS FONDEF DA09I1002 y beca CONICYT - Gobierno Regional de Magallanes y La Antártica Chilena. REFERENCIAS Belchier, M y Collins, M. A. 2008. Recruitment and body size in relation to temperature in juvenile Patagonian toothfish (Dissostichus eleginoides) at South Georgia. Marine Biology, 155: 493-503. Guarner, F. y Malagelada, J. R. 2003. La flora bacteriana del tracto digestivo. Gastroenterology & Hepatology, 26 (1): 1-5. MacCormack, W. P. M. y Fraile, E. R. 1990. Bacterial flora of newly caught Antarctic fish Notothenia neglecta. Polar Biology, 10:413-417. Ward, N., Steven, B., Penn, K., Methé, B. y Detrich III, W. 2009. Characterization of the intestinal microbiota of two Antarctic notothenioid fish species. Extremophiles, 13: 679-685.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
372
BIOPROSPECCIÓN DE MICROORGANISMOS ANTÁRTICOS Y SUS ENZIMAS EXTREMÓFILAS.
(Bioprospecting of Antarctic microorganisms and their extremophilic enzymes)
Vargas, J.1, Cevallos-Cevallos, J.1, Peralta, E.1
1Centro de Investigaciones Biotecnológicas del Ecuador (CIBE), Escuela Superior Politécnica del Litoral (ESPOL), Guayaquil, Ecuador, email: [email protected], [email protected], [email protected], [email protected] RESUMEN La bioprospección de microorganismos en zonas antárticas ha crecido en los últimos años, por el interés de encontrar enzimas activas a bajas temperaturas. El objetivo de este estudio fue de evaluar la presencia de enzimas con alta actividad a bajas temperaturas aisladas de microorganismos antárticos. MATERIALES Y METODOS Zona de muestreo y recolección de muestras Las muestras de suelo fueron colectadas en varios puntos de la estación antártica Pedro Vicente Maldonado en las expediciones del 2010 al 2012. Las Muestras colectadas provinieron de plantas vasculares), torres deshieladas, y de suelos contaminados con hidrocarburos. Aislamiento y Cultivo Bacteriano Se pesó 1g de suelo, y se colocó en tubos con 10 ml de Agua de Peptona para bacterias y 10 ml de agua estéril para hongos. Los tubos inoculados se los dejó incubar durante 2 horas a 4ºC. Se sembraron diluciones hasta 10-3 en medios de cultivo como: TSA (Trypticase Soy Agar) y AN (Agar Nutritivo) para bacterias; YEC (Yeast Extract Cloramphenicol), SAB (Sabouraud Dextrose Agar), y PDA (Potato dextrose agar) para hongos y levaduras. Las muestras sembradas en los diferentes medios se las mantuvo en refrigeradoras reguladas a 4ºC y en una incubadora refrigerada ajustada a 10ºC. Se trabajaron con 137 colonias aisladas para cada prueba. Identificación y Secuenciación de aislados La extracción de ADN se hizo mediante kits comerciales. Se amplificaron fragmentos con primers universales de la región 16s. La secuenciación fue mediante el método de Sanger. Se comprobó la secuencia comparando con la base de datos del NCBI. Bioprospección enzimática Los medios empleados para efectuar las pruebas de actividad enzimática de cada colonia fue efectuada basándose en la publicación (Khan, 2010). Para determinar la actividad enzimática de los diferentes aislados se prepararon medios con sustratos específicos; La prueba de amilasas fue evaluado en medio con 20.1 g/l de NB (Nutrient Broth), 2 g/l Almidón Soluble, 18 g/l agar; para prueba de prueba de proteasa se uso 20.1 g/l de NB, 20 g/l, 20 g/l de Caseína, 18 g/l agar; para prueba de gelatinasa 20.1 g/l de NB, 20g/l glucosa, 8 g/l Gelatina, , 18 g/l agar; para prueba de pectinasa 20.1 g/l de NB, 10 g/l pectina, 18 g/l agar; para prueba de celulasa 20.1 g/l de NB, 20 g/l glucosa, 5 g/l Carboxymethylcellulose, 18 g/l agar; para la prueba de urea 20.1 g/l de NB, 20 g/l glucosa, 1 g/l Urea, 18 g/l agar, 5 g/l de ClNa, 0.018 g/l, Rojo phenol y Agar MacConkey Agar para la prueba de lactasa. Cada microorganismo fue evaluado para cada sustrato para determinar la presencia de las enzimas de interés. La temperatura en la que se efectuó el ensayo fue de 16ºC y la formación de halos con cambios de color indicaba la producción de la enzima de interés.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
373
RESULTADOS Actividad enzimática positiva. Se evaluaron un total de 74 aislados bacterianos y 30 asilados de hongos. La secuenciación de los aislados evaluadas dieron como resultado que la muestra contenía individuos del genero Pseudomonas sp., Arthrobacter sp., Rhodococcus sp. y Pantoea sp.(Gráfico 1.)
Pseudomonas mandelii, es un microorganismo que se aísla con frecuencia en suelos antárticos (Puram, 2009). La especie Pseudomonas presentó en su mayoría positivo para el ensayo de gelatinasa a la temperatura establecida, seguida de lactasa positiva. Actividad amilasa y proteasa y ureasa se obtuvieron entre 5 y 6 aislados positivos. Para celulasas y lactasas no tuvieron resultados positivos en las Pseudomonas evaluadas. (Tabla 1).
Pseudomonas Rhodococus Pantoea Arthrobacter
Amilasa 17 (6) 0 (0) 100 (1) 55 (6)
Proteasa 14(5) 0 (0) 0 (0) 9 (1)
Gelatinasa 86 (30) 25 (1) 100 (1) 45 (5)
Pectinasa 0 (0) 0 (0) 0 (0) 0 (0)
Celulasa 0 (0) 0 (0) 0 (0) 0 (0)
Lactasa 63 (22) 50 (2) 0 (0) 9 (1)
Ureasa 20 (7) 50 (2) 0 (0) 45 (5) 1Los valores en paréntesis indica el numero de aislados positivos (The values in brackets indicate the number of positive isolates)
Tabla 1. Porcentaje de aislados con actividad enzimática por género de microorganismos1
Table 1. Percentages of isolates with enzymatic activity by genus of microorganism El género Arthrobacter presentó mayor actividad enzimática para amilasas, seguido de gelatinas y ureasa, presentando un solo aislado positivo para Proteasas y Lactasas. Se han reportado actividad enzimática para amilasa, lipasa y proteasa en rangos de temperaturas de 5 a 20°C para Arthrobacter citreus (Srinivas, 2009). Los aislados del género Rhodococcus presentaron actividad en las pruebas de gelatinasa, lactasa y ureasa, mientras que el género Pantoea presento resultados positivos para amilasa y gelatinasa (Tabla 1). Las colonias identificadas como CIBE27, CIBE14 y CIBE55 presentaron un mayor halo de actividad de amilasas con 0.9 cm. El halo era notorio sin aplicarle la solución de iodo usada como revelador. Estos aislados corresponden al género Arthrobacter. Este género presento muy pocos resultados positivos en otros sustratos y algunos aislados presentaron resultados positivos hasta tres sustratos, pero la actividad enzimática en ellos fue baja. Las colonia CIBE101 fue la que presento la mayor actividad gelatinasa (1.3 cm halo). Esta colonia fue identificada como Pseudomona sp. Los aislados CIBE68 y CIBE67 presentaron la actividad enzimática en la prueba de urea (1.2 cm de halo). Casi todos los aislados presentaron la una actividad alta durante los 8 días de prueba.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
374
En los gráficos 2 y 3 se muestra una distribución de resultados positivos para los diferentes sustratos evaluados tanto en hongos y en bacterias.
Los sustratos que presentan mayor resultado positivo son Gelatinasas, Lactasas para bacterias, mientras que en hongos se obtuvieron mas resultados positivos en amilasas, ureasas y pectinasas. Del total de enzimas y aislados evaluados, las bacterias no presentaron actividad positiva para celulosa y pectina. Los hongos no presentaron actividad en proteasas y lipasas. La identificacion de los hongos con actividad enzimatica de interes y la caracterizacion de cada enzima se encuentra en proceso. REFERENCIAS Choo, D., Kurihara, T., Suzuki, T., Soda, K. y Esaki, N., 1998, A Cold-Adapted Lipase of an Alaskan Psychrotroph, Pseudomonas sp. Strain B11-1: Gene Cloning and Enzyme Purification and Characterization, Applied and Environmental. Microbiology, 64(2), 486-491 Gerday, C., Aittaleb, M., Bentahir, M. et al. (2000) Cold-adapted enzymes: from fundamentals to Biotechnology. Trends in Biotechnology, 18, 103–107. Khan, A., Bhadra, B., Begum, Z. y Shivaji, S.Khan, A. Bhadra, B., Begum, Z. y Shivaji, S. 2010, Diversity of Yeasts from Puddles in the Vicinity of Midre Love´nbreen Glacier, Arctic and Bioprospecting for Enzymes and Fatty Acids, Current Microbiology, 60, 307–314. Novototskaya, K., Petrovskaya, L., Yakimov, S. y Gilichinsky, D. 2011, Cloning, purification, and characterization of a cold-adapted esterase produced by Psychrobacter cryohalolentis K5T from Siberian cryopeg, FEMS Microbiology Ecology, 82, 367–375. Puram, V., Singireesu, S. Nageswara, R., Mambatta, P., Buddhi, S., Bakka, K., Ravoori, M., Shiv, S., Tanuku, R., y Sisinthy, S. 2009. Bacterial diversity and bioprospecting for cold-active enzymes from culturable bacteria associated with sediment from a melt water stream of Midtre Lovenbreen glacier, an Arctic glacier, ELSEVIER, Research in Microbiology, 160, 538-546. Srinivas, T., Nageswara, R., Puram, V., Pratibha, S., Sailaja, Kavya B., Hara, K., Begum, Z., M. Singh, S., y Shivaji, S., 2009, Bacterial Diversity and Bioprospecting for Cold-Active Lipases, Amylases and Proteases, from Culturable Bacteria of Kongsfjorden and Ny- Alesund, Svalbard, Arctic, Current Microbiology, 59, 537-547 Yong, Y., Hui-Rong, L., Yin-Xin, Z. y Bo, C. 2011. Bacterial Diversity and Bioprospecting for Cold-Active Hydrolytic Enzymes from Culturable Bacteria Associated with Sediment from Nella Fjord, Eastern Antarctica, Marine Drugs, v. 9, p. 184-195.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
375
BACTERIAS ANTARTICAS EXHIBEN ACTIVIDAD ANTIBACTERIANA CONTRA MICROORGANISMOS PATOGENOS
(Antarctic bacteria exhibits antibacterial activity against pathogens)
Viteri, R.1, Quintero, A.1 y J. Cevallos2
1Centro de Química, Instituto Venezolano de Investigaciones Científicas (IVIC), Apartado 20632, Caracas 1020ª, Venezuela. E-mail: [email protected], [email protected] 2Centro de Investigaciones Biotecnológicas del Ecuador (CIBE), Guayaquil-Ecuador. E-mail: [email protected] INTRODUCCIÓN Las bacterias fitopatógenas son capaces de afectar diferentes cultivos, causando grandes pérdidas económicas a nivel mundial (Escalona et al, 2011). Dentro de este grupo se encuentra la batería Xanthomonas albilineans causante de la escaldadura foliar (EF), una de las enfermedades de mayor importancia en el cultivo de la caña de azúcar (Saccharum sp.) ocasionando pérdidas estimadas entre 90-100% de la producción (Alvez, 2011). Otra bacteria patógena es la Xanthomonas phaseoli causante del “tizón común” que afecta a las plantaciones de caraotas (Phaseolus vulgaris L.), ocasionando pérdidas en el rendimiento de 15-30%, (Navarrete, 2000). Pantoea aglomerans es una bacteria que afecta a las plantaciones de cebolla (Allium cepa) causando necrosis foliar (Renaud et al., 2000), pero que también causa enfermedades en seres humanos (personas inmunodeprimidas) (Sanchez, 2006). En las últimas décadas la cura de estas enfermedades se ha basado en el uso de antimicrobianos de origen sintético, cuyo uso indiscriminado ha traído problemas de contaminación, intoxicación y efectos crónicos indeseables en humanos (González, 2005). Una forma de reducir el uso de agentes químicos en la agricultura es mediante el empleo de cepas bacterianas (Gerhardson, 2002). En el escenario actual, los microorganismos como Pseudomonas y Bacillus (Whipps, 2011) son una fuente principal de compuestos antibacterianos. Una vez, demostrado el efecto biológico de una fuente natural (bacteria) es necesario conocer las estructuras de los compuestos responsables de dicha actividad (Romanenko, 2008). En la actualidad, el descubrimiento de nuevos metabolitos a partir microorganismos terrestres ha disminuido, por lo que se ha hecho necesario la búsqueda de compuestos en nuevos entornos (Pathak et al, 2011). Fuentes inusuales, tales como microorganismos de ambientes extremos han llamado la atención de los científicos por la posibilidad de encontrar nuevos productos naturales (Lo Giudice et al, 2007). La región Antártica es de particular interés para la química de productos naturales, ya que sigue siendo relativamente inexplorada y al mismo tiempo agentes microbianos que allí habitan, desarrollan diferentes defensas químicas que les permiten vivir en condiciones extremas de frío, presión y radiación UV (Araujo et al). Por lo antes expuesto, el objetivo de este trabajo fue (I) evaluar la actividad antifitopatógena de cepas antárticas productoras de antimicrobianos, (II) e identificar la bacteria Antártica que exhibe la actividad antimicrobiana. MATERIALES Y MÉTODOS Microorganismos antárticos Las 50 cepas antárticas analizadas pertenecen a la colección de microorganismos antárticos del Centro de Investigaciones Biotecnológicas del Ecuador (CIBE); las mismas fueron aisladas de suelos provenientes de Punta Fort Williams-Isla Greenwich en la Antártida en el laboratorio de Microbiología y Fitopatología del CIBE. Estas fueron conservadas en medio de cultivo Luria-Bertani (LB) a -80ºC y codificadas del CIBE001 al CIBE050.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
376
Evaluación de la actividad antibacteriana La producción de compuestos antibacterianos fue determinada usando una modificación procedimiento de antagonismo de Kekessy y Piguet (1970). Las bacterias fitopatógenas empleadas para este ensayo fueron Xanthomonas phaseoli, Xanthomonas albilineans y Pantoea agglomerans, facilitadas por el Laboratorio de Bacteriología del postgrado de Agronomía de la Universidad Centrooccidental Lisandro Alvarado, Venezuela. Todos los patógenos fueron aislado de cultivos de caraotas (Phaseolus vulgaris L.), caña de azúcar (Saccharum sp.) y cebolla (Allium cepa) respectivamente. Secuenciación de la bacteria El ADN de los microorganismos antárticos que exhibieron potencial antimicorbiano fue extraido usando un kit comercial (Invitrogen). Una parte de la región 16s fue amplificada utilizando cebadores F26 y R574. La secuenciación Sanger fue tercerizada en el Laboratorio de genómica de la Universidad de Florida. RESULTADOS De las 50 cepas antárticas evaluadas, 2 (4%) inhibieron el crecimiento de la bacteria fitopatógena X. phaseoli. Las mismas corresponden a la codificación CIBE015, el cual obtuvo un halo de inhibición con un promedio de 24 mm de diámetro y la cepa CIBE017, tuvo el mayor halo de inhibición, con un promedio de 40 mm de diámetro (Fig. 1). Ninguna de la bacterias probadas inhibió a X. albilineans o a P. aglomerans.
Figure 1: Inhibición del crecimiento de X. phaseoli producido por la actividad antibacteriana de la cepa CIBE017.
Figure 1: Growth inhibition of X. phaseoli due to the antibacterial activity of CIBE017 strain. Las bacterias Antárticas que produjeron actividad antibacteriana fueron identificadas por secuenciación y análisis de su 16S rDNA parcial. Ellas fueron Pseudomonas sp. y Arthrobacter sp. con un 99% de identidad. DISCUSIÓN Las cepas antárticas que produjeron la mayor actividad antibacteriana fueron identificadas como Pseudomonas sp. (CIBE017) y Arthrobacter sp. (CIBE015). Las bacterias del género Pseudomonas sp. son muy utilizadas como agentes para el control biológico de fitopatogenos en la agricultura debido a la excreción de los metabolitos sideróforos y metabolitos antimicrobianos (Hernández et al. 1999). Por otro lado, el género Arthrobacter sp. ha demostrado que produce compuestos orgánicos volátiles que inhiben el crecimiento de hongos fitopatógenos. Es importante destacar que no existen estudios acerca de este género como agente de control biológico contra bacterias fitopatógenas, por lo que este resultado representa un interesante hallazgo. Resultados similares se han informado por otros autores, quienes utilizando metodologías semejantes a la descrita en este trabajo, lograron la inhibición del crecimiento

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
377
de algunas bacterias fitopatógenas utilizando cepas del género Pseudomonas. Reynaldo De La Cruz-Quiroz demostró la capacidad antagónica de nuevas cepas aisladas de P. fluorescens contra 3 bacterias fitopatógenas de gran importancia como: Clavibacter michiganensis, Xanthomonas axonopodis y Erwinia carotovora. Mientras que Romanenko et al., obtuvieron resultados semejantes al nuestro detectando la actividad inhibitoria de 12 cepas de P. fluorescens (aislados de hielo marino) contra Xanthomonas sp. pv. badrii. En este trabajo pudimos confirmar que microorganismos que habitan en condiciones extremas exhiben actividad inhibitoria contra X. phaseoli y pueden producir compuestos antimicrobianos, por lo que representan una fuente potencial para obtener nuevos metabolitos secundarios antibacterianos. CONCLUSIONES Cepas antárticas de Pseudomonas y Arthrobacter producen metabolitos que inhiben a P. phaseoli. Ninguna de las cepas antárticas fue capaz de inhibir el crecimiento de las bacterias X. albilineans y P. aglomerans. En estudios futuros los compuestos antibacterianos serán purificados y caracterizados. AGRADECIMIENTOS AL INAE, SENESCYT, IVIC y el CIBE, instituciones importantes para el desarrollo de este trabajo. REFERENCIAS BIBLIOGRÁFICAS Alvez, B. et al. 2011. Diagnostico molecular de Xanthomonas albilineans (Ashby) Dowson en Venezuela, agente cuasal de la escaldadura foliar de la caña de azúcar. Agronomía Trop. 61(1):27-36. Araújo, L. Kagohara, E. Garcia, T. Pellizari, V. y Andrade, L. 2011. Screening of microorganisms producing cold-active oxidoreductases to be applied in enantioselective alcohol oxidation. An Antarctic survey. Marine drugs. 9:889–905. De La Cruz-Quiroz, R. et al. 2011. Antagonist capacity of Newly Isolated Strains of Pseudomonas Fluorescens against Three Important Phytopathogenic Bacteria. 6(2):267-272. Escalona, S. y Rincón, N. 2011. Enfermedades bacterianas diagnosticadas en el Laboratorio de Bacteriología-UCLA durante el 2010. XXII Congreso Venezolano de Fitopatología, 24(2):60. Gerhardson, B. 2002. Biological substitutes for pesticides. Trends in Biotechnology. 20(8):338-343 González, M., Silva, H. y Mascorro, J. 2005. Ensayo In vitro del péptido Antimicrobiano Melitina contra diferentes Bacterias Fitopatógenas. Revista Mexicana de Fitopatología. 23(2):176-182. Hernández, A., Fernández, A., Pérez, J., Miranda, S., Fons C., Hernández, A. N. y Santander, J. 1999. Producción, purificación y diagnostico de sideróforos a partir de la cepa de Pseudomonas fluorescens J-1443. Cultivos Tropicales. 20(1):21-25 Kekessy, A. y Piguet, D. 1970. New method for detecting bacteriocin production. Applied Microbiology. 20:282-283. Lo Giudice, A. Bruni, V. Michaud, L. 2007. Characterization of Antarctic psychrotrophic bacteria with antibacterial activities against terrestrial microorganisms. Journal of Basic Microbiology. 47:496-505 Navarrete, R. y Acosta J. 2000. Genotipos de frijol (Phaseolus vulgaris L.) resistentes a Xanthomonas campestris pv. phaseoli DE MEXICO. 11(1):17-23 Pathak et al. 2011. Assessment of antibacterial and free radical scavenging activity in psychrophilic Arthrobacter sp. Pharmacologyonline. 1: 344-355 Renaud, D. et al. 2000. Aislamiento e Identificación de Erwinia herbicola pv. Herbicola en cebolla (Allium cepa L) en el Valle de Quibor estado Lara. Resumen VIII Congreso Venezolano de Hortalizas. Barquisimeto estado Lara. Romanenko et al. 2008. Diversity and Antagonistic Activity of Sea Ice Bacteria Isolated from the Sea Japan. Microbes Environ. 23(3):209-214. Sanchez, F. Ruiz, A. Barranco, M. Alonso, A. 2006. Bacteriemia por Pantoea agglomerans. An. Med. Interna (Madrid). 23(5):250-251 Whipps, J. 2001: Microbial interaction and biocontrol in the rhizosphere. J. Exp. Bot. 52: 487-511.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
378
ESPECIFICIDAD Y SELECTIVIDAD DE LOS MICOBIONTES DE LÍQUENES DEL GÉNERO Peltigera DE LAS ISLAS DE TIERRA DEL FUEGO, NAVARINO Y DECEPCIÓN
(Mycobiont Specificity and Selectivity of Peltigera lichens from Tierra del Fuego, Navarino and Deception Islands)
Zúñiga, C. 1, Leiva, D., Frías, F., Carú, M., Orlando, J. 1
1Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile. [email protected]
INTRODUCCIÓN Los líquenes son asociaciones simbióticas estables formadas entre un hongo (micobionte) y al menos un componente foto-autotrófico (fotobionte), que puede ser una micro-alga y/o una cianobacteria. En este último caso se denominan “cianolíquenes”, los cuales tienen la capacidad de fijar nitrógeno y son colonizadores primarios en áreas recientemente expuestas (Chapman y Margulis, 1998). Si bien los líquenes se encuentran entre los grupos de organismos más diversos en la Antártida (Pérez-Ortega y col., 2012), aún existen debates sobre el total de especies actualmente descrito (Kappen, 2000; Robinson y col., 2003). Por otro lado, la correcta clasificación taxonómica de los líquenes basada en caracteres morfológicos representa un paradigma debido a la existencia de especies crípticas (Lumbsch y Leavitt, 2011), y ha sido descrito como particularmente difícil en la zona antártica (Hertel, 1988). El modelo ecológico de liquenización (Yahr y col., 2006) propone que el establecimiento de la interacción depende tanto de la compatibilidad genética de los socios (especificidad), como de la capacidad del hongo para seleccionar (selectividad) entre los fotobiontes disponibles en un determinado ambiente (disponibilidad). Sin embargo, poco se conoce acerca de la formación de la simbiosis liquénica, la especificidad de la interacción, la selectividad de los componentes, o la manera en que éstos se re-liquenizan, y para responder a estas incógnitas resulta indispensable conocer la identidad de ambos simbiontes. El objetivo principal de este estudio fue determinar la especificidad y la selectividad de cianolíquenes del género Peltigera (micobionte: Peltigera sp; cianobionte: Nostoc sp.) a partir de muestras antárticas y compararlas con muestras provenientes de las islas de Tierra del Fuego y Navarino previamente analizadas (Zúñiga y col., 2012).
MATERIALES Y MÉTODOS El sitio de muestreo correspondió a la Bahía Balleneros, en la Isla Decepción (D), Antártida, desde donde se recolectaron 15 muestras de líquenes del género Peltigera (D1-D15). La flora de esta isla es singular ya que en ella se encuentra el mayor número de especies raras y sumamente raras de la Antártida, dentro de las cuáles 14 de los aproximadamente 75 líquenes se consideran raros o sumamente raros (ZAEP 140). El DNA de las muestras se extrajo utilizando el “Power Soil DNA Isolation Kit” (Mobio) y, en el caso de los micobiontes, se amplificó y secuenció el gen que codifica para el rRNA 28S; mientras que para los cianobiontes, el gen para el rRNA 16S. Los amplicones se secuenciaron en un sentido con el partidor forward correspondiente utilizando un Analizador Genético 3730 (Applied Biosystems; Macrogen Inc.). Las secuencias se revisaron y editaron manualmente en el programa Mega v5.2. (Tamura y col., 2011) y se compararon con la base de datos GenBank del NCBI mediante la herramienta bioinformática BLASTn (Altschul y col., 1997), para su identificación. Asimismo, se utilizaron en este trabajo los resultados obtenidos previamente desde muestras de la Isla de Tierra del Fuego (K) y la Isla Navarino (N) (Zúñiga y col., 2012).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
379
Posteriormente, las secuencias nucleotídicas de los simbiontes de las tres localidades se analizaron en el programa Mega v5.2. mediante un análisis filogenético de Máxima Verosimilitud, con el fin de definir operacionalmente los tipos de micobiontes y cianobiontes (OTUs), y determinar si las muestras de la Isla Decepción corresponden a un nuevo clúster de secuencias o si pertenecen a alguna de las OTUs descritas anteriormente. Una vez definidas las OTUs para ambos simbiontes, se calculó la especificidad y la selectividad de cada micobionte. La especificidad (Ep) se definió como el número de cianobiontes con los cuales es capaz de asociarse cada micobionte y fue ponderada por el promedio de la distancia genética entre ellos (distancia-p). Así: Ep = 1- (N° cianobiontes x distancia-p promedio entre los cianobiontes). Por otra parte, la selectividad (Sp) se definió teóricamente como la asociación preferencial de cada micobionte con alguno de sus cianobiontes específicos y se consideró la frecuencia de cada pareja. Se calculó utilizando el Índice de Diversidad de Shannon Ponderado (Poulin y col., 2011), de tal manera que: Sp = 1-(-∑ wi pi log pi), donde pi es la frecuencia de cada pareja y wi es un factor que incorpora la similitud taxonómica de los cianobiontes que forman parte de las parejas. De acuerdo a la Ep y a la Sp de cada micobionte, se definió un rango de categorías que permitió clasificarlos en distintos niveles de especificidad y selectividad. RESULTADOS Y DISCUSIÓN Los análisis permitieron definir un total de 8 OTUs de micobiontes (Fig.1A, OM1-OM8) y 14 de cianobiontes (Fig. 1B, OC1-OC14). En ambos casos se observa la presencia de OTUs frecuentes, con más de 10 representantes, y otras menos frecuentes, llegando algunas a estar formadas por una única muestra. Con respecto a los micobiontes, la mayoría de las OTUs se detectaron en más de una localidad. Sin embargo, ninguno de ellos se encontró presente en las tres localidades estudiadas (K-N-D). Se observa también que algunas OTUs que se encuentran pobremente representadas en uno de los ambientes son comunes en otra localidad; como ocurre en el caso de OC13, que posee un único representante en K, pero 37 en N; o en el de OM3, con 14 representantes en K y sólo uno en N. En el caso de las muestras provenientes de la Isla Decepción, todas (D1-D15) resultaron ser idénticas para ambos simbiontes. Sus micobiontes forman parte del clúster OM4, identificado como P. extenuata, los cuales se reproducen principalmente de manera asexual mediante la producción de soredios. Cabe destacar que, a pesar de formar parte de una misma OTU, las muestras provenientes de K y D no poseen un 100% de identidad nucleotídica, diferenciándose según su origen geográfico. En cuanto a los cianobiontes, la mayoría de ellos se encuentra presente en más de un sitio de muestreo, llegando incluso a estar presentes en los tres sitios, como es el caso de OC10, que fue el único cianobionte detectado en la Isla Decepción. A partir de las OTUs definidas y considerando la distancia genética existente entre los diferentes cianobiontes, se calcularon los índices de especificidad y selectividad para cada micobionte y se definieron 7 categorías, clasificando los valores desde muy bajos hasta muy altos (Tabla 1). Los resultados muestran que los micobiontes más específicos y selectivos fueron OM2 y OM7, los cuales sólo se unen a una única OTU de cianobiontes. Sin embargo, es importante mencionar que la relación entre OM2 y su respectivo cianobionte (OC3) es del tipo “uno es a uno”, a diferencia de OM7 que, si bien sólo se une a OC10, este último es uno de los cianobiontes menos específicos. El micobionte menos específico fue OM1, capaz de asociarse a 5 OTUs de cianobiontes, seguido por OM6, capaz de asociarse con 4. En cuanto a su selectividad, la relación es inversa, siendo OM6 más selectivo que OM1. La OTU que comprende aquellas muestras obtenidas desde la Isla Decepción presentó un grado de especificidad medio-alto (S=3), siendo capaz de asociarse a 3 OTUs de cianobiontes. En cuanto a su selectividad general, esta OTU presenta un grado medio-bajo (SeD=4), formando 1 pareja con OC6, 7

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
380
con OC7 y 15 con OC10. Sin embargo, al analizar su selectividad por ambiente, se observa que es alta en K, ya que solo se une a OC6 y OC7, aun estando OC10 presente (asociado con OM5). Sin embargo, dado que se trata de una especie que se reproduce asexualmente, es menos probable que cambie su fotobionte en comparación a aquellas especies sexuales que necesariamente desacoplan la interacción simbiótica al momento de reproducirse.
Figura 1. Relaciones filogenéticas entre micobiontes (A) y cianobiontes (B) basadas en el análisis de Máxima Verosimilitud. Los números en los nodos indican un soporte estadístico de bootstrap > 75% (1000 repeticiones). K1-K60: Isla de Tierra del
Fuego; N1-N60: Isla Navarino; D1-D15: Isla Decepción. Figure 1. Phylogenetic relationships between the mycobionts (A) and the cyanobionts (B) based in a Maximum Likelihood
analysis. The numbers on the nodes indicate a statistical bootstrap support >75% (1000 repetitions). K1-K60: Tierra del Fuego Island; N1-N60: Navarino Island; D1-D15: Deception Island.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
381
OTU Identidad Especificidad (Ep) Categoría Ep* Selectividad (Sp) Categoría Sp*
OM2 Peltigera sp. 1,0000 MA 1,0000 MA
OM3 P. fuscopraetextata 0,9573 MdB 0,9930 MdA
OM4 P. didactyla 0,9687 MdA 0,9918 MdB
OM5 P. frigida 0,9801 A 0,9937 A
OM6 P. ponojensis 0,9317 B 0,9779 MB
OM7 P. neckeri 1,0000 MA 1,0000 MA
OM8 P. hymenina 0,9633 Md 0,9879 MB
(*) MB: muy baja / very low; B: baja / low; MdB: media baja / moderate low; Md: media / moderate; MdA: media alta / moderate high; A: alta / high; MA: muy alta / very high.
Tabla 1. Especificidad y selectividad de los distintos micobiontes. Table 1. Specificity and selectivity of the different mycobionts.
En conclusión, no se observó variación molecular entre las 15 muestras antárticas estudiadas para ninguno de los simbiontes, lo cual podría ser esperable debido a su estrategia reproductiva. Además, tanto los micobiontes como los cianobiontes fueron clasificados dentro de alguna de las OTUs descritas en el trabajo anterior (OM4 y OC10, respectivamente), aunque en el caso de los micobiontes su secuencia correspondió a un haplotipo diferente al observado en la Isla de Tierra del Fuego. Finalmente, OM4 mostró una selectividad diferente en la Antártida y en la Isla de Tierra del Fuego. Sin embargo, es necesario descartar que esto de deba a la disponibilidad de los cianobiontes en el ambiente, para lo cual es necesario realizar estudios directamente enfocados en esta materia. AGRADECIMIENTOS INACH - ECA 49 (Proyecto F_02-10); Fondecyt 11100381. REFERENCIAS BIBLIOGRÁFICAS Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W. y Lipman, D.J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research 25(17):3389-3402. Chapman, M. J. y Margulis, L. 1998. Morphogenesis by symbiogenesis. International Microbiology. 1(4):319-326. Hertel, H. 1988. Problems in monographing Antarctic crustose lichens. Polarforschung 58(2/3):65-76. Kappen, L. 2000. Some aspects of the great success of lichens in Antarctica. Antarctic Science 12(3):314‐324. Lumbsch, H. T. y Leavitt, S. D. 2011: Goodbye morphology? A paradigm shift in the delimitation of species in lichenized fungi. Fungal Diversity 50:59-72. Pérez-Ortega, S., Ortiz-Álvarez, R., Allan Green, T. G. y De Los Ríos, A. 2012. Lichen myco- and photobiont diversity and their relationships at the edge of life (McMurdo Dry Valleys, Antarctica). FEMS Microbiology Ecology 82(2):429-448. Poulin, R., Krasnov, B. y Mouillot, D. 2011. Host specificity in phylogenetic and geographic space. Trends in Parasitology 27(8):355-361. Robinson, S., Wasley, J., y Tobin, A. 2003. Living on the Edge ‐ plants and global change in continental and maritime Antarctica. Global Change Biology 9(12):1681‐1717. Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. y Kumar, S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution 28(10): 2731-2739. Yahr, R., Vilgalys, R. y Depriest, P. T. 2006. Geographic variation in algal partners of Cladonia subtenuis highlights the dynamic nature of a lichen symbiosis. New Phytologist 171(4):847-860. ZAEP 140. 2005. Plan de gestión de la Zona Antártica Especialmente Protegida N°140: Partes de la Isla Decepción. Islas Shetland del Sur. XXVIII Reunión Consultiva del Tratado Antártico. Zúñiga, C., Ramírez, L., Méndez, M., Carú, M., y Orlando, J. 2012. Diversidad y especificidad de los simbiontes de cianolíquenes del genero Peltigera de las islas de Tierra del Fuego y Navarino (Chile). XXXIV Congreso Chileno de Microbiología. Valdivia.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
382
BUSQUEDA DE MICROORGANISMOS CON POTENCIAL BIOTECNOLOGICO A PARTIR DE SUELOS ANTARTICOS CONTAMINADOS CON HIDROCARBUROS
(Looking for microorganisms with biotechnological potential from an Antarctic hydrocarbon-contaminated soil)
Martinez Alvarez L1,2, Ruberto L1,2,3, Vazquez S2,3, Coria Silvia1, Hernandez Edgardo1, Dias Romina3,
Mac Cormack WP1,2
1Instituto Antártico Argentino, Balcarce 290, CABA, Argentina
2Cátedra de Microbiología Industrial y Biotecnología, FFyB, UBA, Junín 956 6to piso CABA, Arg 3Conicet, Av. Rivadavia 1917, CABA, Argentina INTRODUCTION An Antarctic hydrocarbon-contaminated soil offers a combination of factors which turns it in an interesting source of stress adapted microorganisms. On the one hand, Antarctic ecosystems provide to researchers with the opportunity to find yet undescribed microorganisms. On the other hand, the low temperatures force the microorganisms to the development of strategies to survive and grow under such conditions. Hydrocarbon contamination represents an additional challenge for microorganisms, exposing them to a potentially toxic substance that could be used as carbon and energy sources. All these factors could result in adaptation mechanisms (enzymes and other bioactive molecules) with potential biotechnological applications. METHODS Twenty microbial strains were isolated from hydrocarbon-contaminated soil from Carlini station. Enrichment cultures using a saline basal medium with gasoil (1%) as sole carbon and energy source were incubated during 15 d at 15 °C and 250 rpm in a rotatory shaker. After the enrichment procedure, subsamples of the cultures were plated in the same culture medium supplemented with bacteriological agar (2%) and incubated at 15°C for 10 d. Colonies were picked out from these plates and subcultured for purification. Genomic DNA from each isolated strain was extracted using a commercial kit (Axygen) and used as template for PCR amplification of the 16S rRNA gene (bacteria) and the D1/D2 region of the 26S rRNA gene and the ITS (fungi). Primers sequences are shown in Table 1 and PCR conditions, in Table 2. Sequences were compared against related sequences included in the e-Taxon database (type strains) from EzBioCloud for bacteria and Mycobank for yeast/fungi. The taxonomic affiliation of the isolates and the percentage of similarity between the LSU/ITS sequences obtained from each isolated bacteria, yeast or filamentous fungus and their closest relatives present in databases are shown in the Results section.
Bacteria Primer sequence 27f 5' AGAGTTTGATCMTGGCTCAG 3' 1492r 5' TACGGYTACCTTGTTACGACTT 3´ Fungi ITS4 5' TCCTCCGCTTATTGATATGC 3' ITS5 5' GGAAGTAAAAGTCGTAACAAGG 3´ NL1 5´GCATATCAATAAGCGGAGGAAAAG 3´ NL4 5´GGTCCGTGTTTCAAGACGG 3´
Table 1. Sequences of the primers used for PCR amplification Tabla 1: Secuencias de los partidores usados para la amplificación por PCR.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
383
Temperature Time Number of cycles
Fungal LSU/ITS 16S rRNA 94°C 5 min 2 min 1 94°C 45 seg 1 min
X 35 55°C 45 seg 1 min 72°C 1 min 30 seg 2 min 72°C 10 min 10 min 1
Table 2: PCR conditions. Tabla 2: Condiciones del PCR.
RESULTS Bacteria Pseudomonas frederikbergensis Pseudomonas frederikbergensis JAJ28(T) 98.95% Pseudomonas brenneri Pseudomonas brenneri CFML97-391(T) 99.57% Pseudomonas amygdali Pseudomonas amygdali LMG 2123T (T) (Z76654) 98.3% Caulobacter henricii Caulobacter henricii ATCC 15253(T) 99.13% Variovorax ginsengisoli Variovorax ginsengisoli Gsoil3165 (T) (AB245358) 99.3% Sphingomonas aerolata Sphingomonas aerolata NW12(T) 99.35% Yeast Glaciozyma martinii Glaciozyma martinii strain CBS 8929 (AY040664) 99% Filamentous fungi Cadophora malorum Cadophora malorum (JQ796752) 100% Aspergillus sp Aspergillus versicolor strain UASWS0828 (JX139733) 100% Crytococcus laurentii Cryptococcus laurentii strain LANTA64 99% JN172923.1 DISCUSSION Detection in the studied soil of a number of strains ascribed to the genus Pseudomonas can be considered as a common feature. Members of the Pseudomonadaceae family were reported as frequent inhabitants of hydrocarbon contaminated soils and hence, have been considered as appropriate candidates to be used in bioaugmentation process, mainly considering that they are typical r strategist, tending to quickly consume any abundant energy resource available. The isolation under low temperature conditions suggest their ability to grow as psycrotolerant microorganisms. Caulobacter is refered as a prosthecated bacterial genus well adapted to oligothrophic conditions (Wolf-Rainer et al, 1999). Its member inhabits mainly aquatic habitatss. This bacterial group and the previously mentioned Pseudomonadaceae family are thought to be responsible for mineralization of an important faction of dissolved organic material in aquatic environments. Caulobacter seems to play a relevant role in the organic matter cycling when nutrient concentration and temperature are low (Stanley et al 1987). Variovorax has been reported as a prevalent genus in Antarctic hydrocarbon contaminated soils, along with Pseudomonas, Sphingomonas and Rhodococcus (Saul et al 2005). The isolate obtained in this study, Variovorax ginsengisoli has been recently described as a novel species with denitrifying activity (Wan-Taek et al 2010). If this activity is also confirmed in the isolated variovorax strain, it could be interesting for the development of low-temperature bioremediation processes under anaerobic conditions. Sphingomonas aerolata has been reported for Antarctic soil (Aislabie et al 2000) and also in 4200 years-old ice (Chistner et al 2000, 2001). Members of the Sphingomonas genus are commonly reported as inhabitants of hydrocarbon contaminated soils from cold regions (Saul et al 2005).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
384
Glaciozyma yeast were isolated from cold environments all around the world, such as Alpine and Apenine glaciers and Antarctic soil and sea water. The two species belonging to the new genus were recently described (Turchetti et al 2011). A few months ago Hashin et al (2013) characterized an antifreeze protein from Galciozyma antarctica PI12, highlighting the biotechnological potential of this microorganism. A strain of Cadophora malorum which was isolated from green algae was reported as a producer of sclerosporins and hydroxylated sclerosporin derivatives. One of these compounds showed a weak fat-accumulation inhibitory activity against 3T3-L1 murine adypocites (Almeida et al 2010). A strain of Cadophora malorum was previously isolated by us (Ruberto et al 2011). The strain showed the production of an exudate exhibiting certain inhibitory activity on several other fungi, such us Diploidia mais. Fungi belonging to the genus Aspergillus are well known because its wide distribution and application in fermented food production, heterologous protein expression and toxins production. A psychrotolerant strain of this fungus could be an interesting model for the production of cold enzymes and other secondary metabolites. Finally, Crytococcus laurentii is considered a psychrophilic fungus who poorly growths above 30°C and shows an optimal growth temperature of 15°C. It is frequently found in Antarctic soil (Tosi et al, 2002). Strains of this fungus have been reported as atrazine-degrading microorganisms and have been proposed as bioremediation tools (Evy et al 2012) CONCLUSION Hydrocarbon contaminated soil from Antarctica is, because the stress factors that it represents, an interesting source of microorganisms with potential biotechnological applications. All the microorganisms identified in this work are related to others which have been reported as biotechnological tools. The finding of several members of the Pseudomonas genus in addition to the Caoulobacter, Sphingomonas and the Variovorax strains suggested that an association of these microorganisms prepeared in the laboratory could be an adequate option for the development of a biological tool for bioremediation of cold soils. Its natural occurrence as associated microorganisms could provide to the artificial microbial association some advantages at the moment of its reintroduction into the soil, overcoming the challenges that this procedure implies. The existence of synergic relationships among these strains must be studied in a next step. Because of its recalcitrant-organic-compounds degradation ability, the strain of Crytococcus laurentii could be added to the above mentioned bacterial association, reinforcing its degradation potential and providing some features typical of fungi, such as exoenzymes production, ability to degrade polymers and activity al low water activity. New screenings are necessary using different conditions in order to isolate other microorganisms that allow the complementation of the proposed bioremediation tool. In this sense, bacteria being k strategist (with the ability for a long term utilization of a scarce nutrient), such as Rhodococcus or Arthrobacter, seems to be appropriate partners for this association heading to a more complete biodegradation process. The isolated strains are going to be studied from different approaches (beyond their use as bioremediation tools) tending to identify their biotechnological potential. Cold active enzymes as well as novel or described secondary metabolites production, antibacterial and antifungal activity, antifreeze proteins or compounds will be investigated. This research should be conducted under stress conditions, such us low temperature, pollutant presence and coculture situations, heading to the potential expression of silent genes clusters.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
385
REFERENCES Aislabie, J., Foght, J. & Saul, D. 2000. Aromatic hydrocarbondegrading bacteria from oil near Scott Base, Antarctica. Polar Biol 23,183–188. Celso Almeida, Ekaterina Eguereva, Stefan Kehraus, Carsten Siering, Gabriele M. König. 2010. Hydroxylated Sclerosporin Derivatives from the Marine-derived Fungus Cadophora malorum. J Nat Prod. 73(3): 476–478. doi:10.1021/np900608d. Christner, B. C., Mosley-Thompson, E., Thompson, L. G. & Reeve, J. N. 2001. Isolation of bacteria and 16S rDNAs from Lake Vostok accretion ice. Environ Microbiol 3, 570–577. Christner, B. C., Mosley-Thompson, E., Thompson, L. G., Zagorodnov, V., Sandman, K. & Reeve. J. N. 2000. Recovery and identification of viable bacteria immured in glacial ice. Icarus 144, 479–485. Evy A, Lakshmi V, Nilanjana D. 2012. Biodegradation of atrazine by Cryptococcus laurentii isolated from contaminated agricultural soil. J. Microbiol. Biotech. Res. 2:450-457. Hashim NHF, Bharudin I, Nguong DLS, Higa S, Bakar FDA, Nathan S, Rabu A, Kawahara H,Illias RM, Najimudin N, Mahadi NM, Murad AMA. 2013. Characterization of Afp1, an antifreeze protein from the psychrophilic yeast Glaciozyma Antarctica PI12. Extremophiles 17:63–73. Ruberto L, Levin G, Hernandez E, Stchigel A, Blanco M, Mac Cormack W. 2011. Isolation of a fungus from a hydrocarbon contaminated Antarctic soil and characterization of it exudate. XLVII Reunión Anual de la SAIB, Potrero de los Funes, San Luis, Argentina. October 30- November 2. Saul DJ, Aislabie JM, Brown CE, Harris L, Foght JM. 2005. Hydrocarbon contamination changes the bacterial diversity of soil from around Scott Base, Antarctica. FEMS Microbiology Ecology 53:141–155 Staley T, Konopka AE, Dalmasso P. 1987. Spatial and temporal distribution of Caulobacter spp. in two mesotrophic lakes. FEMS Microbiol Ecol45, 1-6. Tosi S, Casado B, Gerdol R, Caretta G. 2002. Fungi isolated from Antarctic mosses. Polar Biol 25: 262–268. Turchetti B, Thomas Hall SR, Connell LB, Branda E, Buzzini P, Theelen B, Muller WH, Boekhout T. 2011. Psychrophilic yeasts from Antarctica and European glaciers: description of Glaciozyma gen. nov.,Glaciozyma martini sp. nov. and Glaciozyma watsoniisp. nov. Extremophiles, 15:573–586. Wan-Taek I, Qing-Mei L, Kang-Jin L, Se-Young K, Sung-Taik L, Tae-Hoo Y. 2010. Variovorax ginsengisoli sp. nov., a denitrifying bacterium isolated from soil of a ginseng field. International Journal of Systematic and Evolutionary Microbiology, 60, 1565–1569. Wolf-Rainer Abraham, Carsten StrOmpl, Holger Meyer, Sabine Lindholst, Edward R. B. Moore, Ruprecht Christ, Marc Vancanneyt, B. J. Tindali, Antonio Bennasar, John Smit, Michael Tesar. 1999. Phylogeny and polyphasic taxonomy of Caulobacter species. Proposal of Maricaulis gen. nov. with Maricaulis maris (Poindexter) comb. nov. as the type species, and emended description of the genera Brevundirnonas and Caulobacter. International Journal of Systematic Bacteriology, 49, 1053-1073.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
386
LEVADURAS OLEAGINOSAS ANTARTICAS COMO FUENTE DE TRIGLICERIDOS PARA LA PRODUCCIÓN DE BIODISEL
(Oleaginous yeasts as raw material for biodiesel production)
Martínez, A.1, Pereyra, V. 1, González, A. 1, Alborés, S. 1, Garmendia, G. 1,Rufo, C. 2, Vero, S. 1 1 Departemento de Biociencias, Facultad de Química, Universidad de la República, Gral. Flores 2124, Montevideo, Uurguay 2 Polo Tecnológico de Pando, Facultad de Química, Universidad de la República. E-mail: [email protected]
INTRODUCCIÓN La búsqueda de fuentes de energía renovables propulsada por la potencial desaparición de las fuentes de combustible fósil y las consecuencias ambientales de su explotación, ha determinado actualmente un creciente interés en los biocombustibles. En particular, el biodiesel se presenta como una prometedora alternativa. El biodiesel es producido por esterificación de ácidos grasos provenientes de triglicéridos de diferentes orígenes, con alcoholes de cadena corta. Para la producción de biodiesel se han utilizado aceites animales y vegetales tales como el aceite de soja, girasol o canola. Sin embargo, el alto costo asociado a la generación de dichas materias primas puede comprometer la viabilidad económica de la producción de biodiesel. A su vez, la implementación de cultivos comestibles extensivos para la posterior producción de combustibles ha sido cuestionada desde un punto de vista ambiental, social y cultural (Sharma y Sing, 2008). Por lo tanto, se han buscado fuentes alternativas de triglicéridos entre las cuales se encuentran los aceites producidos por microorganismos tales como bacterias, levaduras, microalgas u hongos filamentosos. Las levaduras oleaginosas se definen por la capacidad de acumular lípidos en su interior celular en cantidades que representan al menos un 20% de su peso seco, bajo determinadas condiciones de cultivo. Sin embargo, en varias especies este porcentaje llega a ser de más del 60%. Esta acumulación se produce ante la escasez de algún nutriente, en especial nitrógeno, sólo si la fuente de carbono y energía continúa en exceso (Li et al., 2008, Meng et al., 2009). Este trabajo tiene como propósito el aislamiento y selección de levaduras de la Antártida para ser utilizadas como fuente de triglicéridos para la producción de biodiesel. En particular, se espera obtener cepas de levaduras a partir de muestras de sedimentos y agua de la Antártida con el objetivo de explorar la biodiversidad microbiana local y ampliar la posibilidad de detectar levaduras oleaginosas eventualmente adaptadas a bajas temperaturas. La adaptación a dichas condiciones posiblemente determine en microorganismos oleaginosos una composición de triglicéridos diferente a la que puede encontrarse en muestras de suelo de zonas templadas (Amaretti et al., 2010, Rossi et al., 2009)
METODOLOGÍA Muestras Durante enero de 2012, se tomaron en forma aséptica, 38 muestras de suelo y de aguas continentales y oceánicas de diferentes puntos de la isla Rey Jorge, las cuales se mantuvieron en refrigeración hasta su análisis. Aislamiento de levaduras Para todas las muestras se realizó un aislamiento directo en medio Dicloran Rosa Bengala Agar (DRBA), con agregado de cloranfenicol. Las muestras líquidas fueron estriadas en placa directamente, mientras que para las muestras sólidas se realizó previamente una suspensión 1/10 en agua estéril. Las placas fueron incubadas a 15ºC. Luego de observar crecimiento se realizaron reaislamientos de las colonias de levaduras aisladas en placas con medio PDA, para obtener cepas puras. En todos los casos se identificaron previamente los aislamientos como levaduras, a través de frotis y observación al

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
387
microscopio óptico. Para las muestras en las que no se evidenció crecimiento de levaduras a través del procedimiento mencionado anteriormente se realizaron enriquecimientos en medio Yeast Nitrogen Base con glucosa como fuente de carbono, y agregado de neomicina y cloranfenicol para inhibir el crecimiento de bacterias. Los cultivos fueron incubados a 20ºC en condiciones aerobias (agitación a 100 rpm). Luego de 4 días de incubación se estrió el cultivo en placas PDA con agregado de cloranfenicol y neomicina. Las placas fueron incubadas 9 días a 20ºC y posteriormente se realizaron reaislamientos en medio PDA para la obtención de cepas puras. Todos los aislamientos puros obtenidos fueron almacenados en tubos inclinados con medio PDA a 4ºC, en medio PDA con vaselina líquida a temperatura ambiente y en presencia de glicerol al 20% por duplicado en freezer a -80°C. Identificación La identificación se realizó por secuenciación de la regiones ITS1-ITS2 y D1/D2 y posterior comparación de secuencias con la base de datos GenBank Selección de levaduras oleaginosas Para cada uno de los aislamientos se evaluó de forma cualitativa la capacidad de acumular lípidos en su citoplasma mediante tinción con Sudan Black B en condiciones especiales de cultivo (Thakur et al., 1989). Para dicho análisis, los aislamientos se incubaron a 20ºC con agitación en condiciones de acumulación de lípidos según Thakur et al. (1989). A los 2 y 5 días de cultivo se observaron bajo microscopio óptico frotis de las levaduras teñidos con Sudan Black B. Esta técnica permitió diferenciar rápidamente las cepas oleaginosas de las no oleaginosas, utilizando como control negativo una cepa de Saccharomyces cerevisiae, especie reconocida como no oleaginosa (Burja et al., 2007). Los aislamientos caracterizados como oleaginosos fueron analizados de forma de determinar cuantitativamente la capacidad de acumulación intracelular de lípidos mediante duna técnica fluorimétrica utilizando rojo de Nilo como indicador (Pereyra, 2013). Análisis de lípidos intracelulares Se realizó el análisis de lípidos intracelulares de una de las cepas con mayor capacidad de acumulación y se comparó con el perfil obtenido para una cepa mesófila seleccionada en un trabajo previo. Para ello se cultivaron ambas cepas en condiciones de acumulación de lípidos durante 5 días y luego se realizó la extracción de los lípidos intracelulares por saponificación directa según Burja et al. (2007). Luego se realizó la esterificación de los mismos para el análisis de los metilésteres correspondientes por cromatografía gaseosa. Los metilésteres obtenidos fueron identificados por cromatografía gaseosa en un cromatógrafo de gases Shimadzu GC 2014 equipado con un detector de ionización de llama y una columna Carbowax 20 (30 m, 0.25 mm ID, 0.25 µm), utilizando como estándar la mezcla de esteres metílicos GLC 10 FAME mix, Sigma-Aldrich 1891 (Pereyra, 2013)
RESULTADOS Se aislaron un total de 34 levaduras, de las cuales 26 fueron obtenidas luego de enriquecimiento, lo cual indicó la baja concentración en que estos microorganismos se encontraban en las muestras analizadas. En el caso de algunas muestras líquidas fue posible establecer que la concentración de levaduras era menor a 100 ufc por mililitro. Todas las levaduras aisladas pudieron ser identificadas a nivel de especie. Las especies más abundantes resultaron Cryptococcus victoriae y Candida glaebosa, identificándose en total 12 especies diferentes. Un tercio de los aislamientos fue caracterizado como oleaginoso de acuerdo al contenido de lípidos intracelulares, según el ensayo con Sudan Black y rojo de Nilo, llegando en algunos casos a porcentajes de acumulación cercanos al 50% de su biomasa. En la figura 1 se puede observar con la tinción de Sudan Black la cepa 17 (Leucosporidium scottii) que es capaz de acumular lípidos (glóbulos azules). En

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
388
las figuras 2 y 3 se observa la cepa oleaginosa 8E (Guehomyces pullulans) con tinción de rojo de Nilo en microscopio de fluorescencia (glóbulos fluorescentes intracelulares).
Figura 1 Figura 2 Figura 3
Figura 1: Cepa 17 teñida con Sudan Black en microscopio óptico 1000X. Figura 2: Cepa 8E con filtro visible en microscopio
de fluorescencia 1000X. Figura 3: Cepa 8E teñida con Rojo de Nilo (0.47 µg/ml ) y con filtro de fluorescencia. Figure 1: Strain 17 stained with Sudan Black in optical microscope 1000X. Figure 2: Strain 8E in visible filter fluorescence
microscope 1000X. Figure 3: Strain 8E stained with Nile red (0.47 mg / ml) with fluorescence filter.
El análisis de lípidos intracelulares de uno de los aislamientos demostró una composición de ácidos grasos similar a la obtenida a partir de aceites vegetales, con un mayor porcentaje de ácido oleico y palmítico, sin presentar ácidos grasos poliinsaturados y un contenido de ácido linolénico cercano al 2%, mucho menor de máximo admitido para la fabricación de biodiesel (12%). El porcentaje de acumulación y el perfil de ácidos grasos obtenido fue similar al encontrado para la cepa mesófila Rhodotorula graminis S1/2R, seleccionada anteriormente por nuestro grupo como materia prima potencial para la producción de biodiesel.
DISCUSIÓN Y CONCLUSIONES Este trabajo mostró la diversidad de especies y la baja concentración de levaduras psicrótrofas encontradas en las muestras provenientes de la Isla Rey Jorge. Este resultado lleva a considerar el uso de nuevas estrategias de aislamiento para detectar su presencia en un próximo muestreo. La existencia de levaduras oleaginosas fue confirmada en territorio antártico, describiéndose especies no caracterizadas previamente como oleaginosas. La cantidad de lípidos acumulados y el perfil de ácidos grasos contenidos en las levaduras seleccionadas permite pronosticar su uso como materia prima para la producción de biodiesel (Ramos et al., 2009). El hallazgo de levaduras psicrótofras capaces de acumular lípidos en su citoplasma es importante ya que se ha demostrado que cultivando a temperaturas bajas, el perfil de ácidos grasos podría cambiar (Amaretti et al., 2010), lográndose un perfil de ácidos grasos que permitan producir un biodiesel de características fluidas a bajas temperaturas, de gran potencial de uso en regiones frías (Park et al., 2008). AGRADECIMIENTOS Al Instituto Antártico Uruguayo, OPCW, ANII, Pedeciba. REFERENCIAS Amaretti, A.; Raimondi, S.; Sala, M.; Roncaglia, L.; De Lucía, M.; Leonardi, A.; Rossi, M. 2010. Single cell oils of the cold-adapted oleaginous yeast Rhodotorula glacialis DBVPG 4785. Microbial Cell Factories. 9: 73-79. Burja, A.M.; Armenta, R.E.; Radianingtyas, H.; Barrow, C. 2007.Evaluation of Fatty Acid Extraction Methods for Thraustochytrium sp.ONC-T18. J. Agric. Food Chem. 55, 4795-4801.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
389
Li, Q.; Du, W.; Liu, D. 2008. Perspectives of microbial oils for biodiesel production. Applied Microbiology and Biotechnology. 80: 749–756 Meng, X., Yang, J., Xu, X., Zhang, L., Nie, Q., Xian, M. 2009. Biodiesel production from oleaginous microorganisms. Renewable Energy. 34: 1-5 Pagnoni, U.M. y Matteuzzi, D. 2009. Growth, lipid accumulation, and fatty acids composition in obligate psychrophilic, facultative psychrophilic and mesophilic yeasts. FEMS Microbiol Ecol. 69: 363-372. Park, J., Kim, D., Lee, J., Park, S., Kim, Y., Lee, J. 2008. Blending effects of biodiesels on oxidation stability and low temperature flow properties. Bioresource Technology 99: 1196–1203 Pereyra, V. 2013. Producción de levaduras oleaginosas a partir de glicerol para la obtención de triglicéridos utilizables como sustrato en la producción de biodiesel. Tesis Maestría en Biotecnología. Facultad de Ciencias. Universidad de la República. Uruguay Ramos, M.J., Fernández, C.M., Casas, A., Rodríguez, L., Pérez, A. 2009. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresource Technology 100: 261-268. Rossi, M., Buzzini, P., Cordisco, L., Amaretti, A., Sala, M., Raimondi, S., Ponzoni, C.,Pagnoni, U.M., Matteuzzi, D. 2009. Growth, lipid accumulation, and fatty acids composition in obligate psychrophilic, facultative psychrophilic and mesophilic yeasts. FEMS Microbiol Ecol. 69: 363-372. Sharma, Y.C. y Singh, B. 2008. Development of biodiesel: Current scenario. Renewable and Sustainable Energy Reviews. 13: 1646-1651 Thakur, M.S., Prapulla,S.G., Karanth, N.G. 1989. Estimation of intracelular lipids by the measurement of absorbance of yeast cells stained with Sudan Black B. Enzyme Microbial Technology. 11, 252-254.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
390
ESTUDIOS PRELIMINARES SOBRE METEORIZACION BIOGENICA EN CRONOSECUENCIAS DE SUELOS EN PENINSULA FILDES, ANTARCTICA.
(Preliminary studies about the biogenic weathering of soil chronosequences in Fildes Península, Antarctica)
Godoy, R.1 & Boy, J.2, Mikutta, R.2, Shibistova, O.2, McCulloch, R.3 &Guggenberger, G.2
1Instituto de Ciencias Ambientales y Evolutivas, Universidad Austral de Chile,Valdivia ([email protected]) 2Institut of Soil Sciences, Leibniz Universität Hannover, Germany ([email protected]) 3Biological and Environmental Sciences, University of Stirling, Scotland INTRODUCTION Mineral weathering is of the utmost importance for the support of life on earth, as it turns bedrock into soil and delivers nutrients to organisms. Recent evidence points towards biogenic weathering, the weathering induced by biota, as the major contributor to weathering rates rather than physico-chemical weathering, thus turning weathering into an active process fueled by photosynthesis. This project aims to determine the contribution of biota to weathering processes during soil formation and organism succession along a chronosequence in maritime Antarctica established by glacier retreat at the Fildes Península and along paleobeaches. Selected unweathered minerals were introduced in situ at each developmental stage of the biogeosystem. After an exposure time of 1 and 2 years weathering rates of biota will be analyzed by confocal laser scanning microscopy and scanning electron microscopy and related to functional groups of organisms present, as well as biomass, chlorophyll and Corg in soil. This will clarify whether biogenic weathering is an active directional process, including nutrient selectiveness by weathering organisms in later successional stages. Furthermore, we will identify whether biogenic weathering is linked to photosynthetic active organisms, or can depend on carbon sources from outside the ecosystem alone (e.g. aerosol deposition). As such this project will deliver a significant contribution to the understanding of if and how life alters its own habitat in extreme and early-stage ecosystems. This is a preliminary report of two chronosequences formed by glacier retreat at Fildes Peninsula, King George Island, regarding the C and N accumulation alongside soil development, including a first inventory of lichens and mosses. PRELIMINARY RESULTS AND DISCUSSION Chronosequence identification In order to investigate all possible chronosequences we used all known deglaciation-schemes proposed for the Fildes Peninsula: i) A uniform retreat of the glacier from the south-western tip of Fildes Peninsula towards the actual glacier front (Mäusbacher et al., 1989), ii) an altitudinal retreat from the shoreline towards higher grounds, and iii) the sequences of raised stormbeaches (“palaeobeaches”, Watcham et al., 2011) which were built by increasing uplift of the Fildes Peninsula following deglaciation in progress (locations indicated in Figure 1). In case of the uniform glacier retreat we established one chronosequence of dated lakes close to the shore (Mäusbacher et al., 1989; Watcham et al., 2011; “L”-plots, Fig. 1), and another chronosequence at the Meseta, an inland-altiplano at an altitude of around 200m asl and assumed to be little affected by sea bird feces and anthropogenic activity (“M”-plots, Fig. 1, panel b). The third chronosequence was established along an altitudinal glacier retreat on a pillar at Collins Bay (Fig. 1, panel c). The site was selected because this pillar is the only deglaciated outcrop in this larger glacier area and surrounded and crested by the Collins Ice cap, thus ensuring a consecutive deglaciation without the microclimatic help of adjacent ice-free areas (“C”-plots, Fig. 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
391
Palaeobeach chronosequences dating back to 7200 years were best developed at Ardley island and Loberia Klotz (Watcham et al., 2011; “P”- green plots, Fig. 1, panel a).
Figure 1: left: location of soil chronosequences caused by glacier retreat at the Fildes Peninsula, King George Island; right: a) Ardley palaeobeach sequence (P1-P6,); b) typical plot at the altitudinal chronosequence ‘Meseta’ (“M”), here M6, with soil having a loamy texture; ‘Meseta’ chronosequence consists of plots of comparable altitude (~ 220 masl), almost no impact of bird feces and located as far as possible from direct sea spray, c) chronosequence at the pillar at ‘Collins’, C1-C4. ‘Collins’ represents altitudinal glacier retreat with Deschampsia antarctica as the climax species). Another chronosequence ‘Lake’ (“L”, left) consists of plots close to the dated lakes of Mäusbacher et al. 1989. Other palaeobeach plots: P7 serves as a blank for the palaeobeaches, as it is the same palaeobeach-level (7200 yrs bpd) as P6, only that soil life was consecutively poisoned by Penguin feces, therefore, biogenic weathering had not started (unlike P6, P7 still consists of the original storm-beach pebbles, of diameters around 10cm). P12 is another storm-beach “poisoned” by penguins corresponding to P2 (1200 yrs bpd). P8-P11 is a series of palaeobeaches from the age 2600-7200 yrs bpd, receiving, unlike Ardley, high eolian dust deposits (from nearby Valle Klotz). Figura 1: Izquierda: Localización de cronosecuencias de suelo causadas por el retroceso del Glaciar en la Península Fildes, Isla Rey Jorge. Derecha: a) cronosecuencias Paleoplayas en Ardley (P1-P6); Derecha b) Sitio típico de la cronosecuencia altitudinal “Meseta” (M1-M8,), donde M6, se presenta con suelo de textura limosa; la cronosecuencia de ‘Meseta’ consiste de sitios de altitud comparable (~ 220 masl), siempre libre de impacto por fecas de aves y localizado lejos como posible, de los impactos directos de aerosoles marinos, Derecha c) Cronosecuencia junto al Pilar del Collins (C1-C4) . De esta forma Collins representa el retroceso altitudinal del Glaciar con Deschampsia antarctica como la especie Climax. Otra cronosecuencia ‘Lagos’ (L1-L3) consiste en áreas ubicadas junto a los lagos con cronología según Mäusbacher et al. 1989. Otros sitios Palaeoplayas (P7) sirve como blanco, ubicada al mismo nivel de P6 (7200 yrs bpd), solo que el suelo de este sitio ha sido consecutivamente impactado por fecas de pinguinos. El sitio P12 es otra área con perturbación por fecas de pinguinos equivalente a P2 de Ardley (1200 yrs bpd). P8-P11 corresponde a una serie de Paleoplayas de edades que fluctuantes (2600-7200 yrs bpd), que reciben un aporte de depósito de polvo eólico (Valle Klotz).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
392
SOIL C AND N STOCKS Analyses of C and N content of two chronosequences revealed that the accumulation of organic material was significant and still did not reach the plateau (Fig. 2). It is surprising that quantities of 50 tonnes per hectar can be realized at these climatic conditions. We interpret the accumulation of C and N as a viable proof of that our identification of soil chronosequences was successful.
Figure 2: C and N quantities at the Ardley Palaeobeach chronosequence (top) and the Meseta glacier retreat chronosequence (bottom). Figura 2: Contenido de C y N total, en sitios cronosecuencias de las Paleoplayas de Ardley (arriba) y de sitios de la cronosecuencias del retroceso del Glaciar en Meseta (abajo), Península Fildes, Isla Rey Jorge. Inventory of lichens and mosses We found a relatively high biodiversity in lichens and mosses along our chronosequences. It became apparent, that although high emphasis was put on finding sites with a favourable microclimate, for most species clear successional patterns along a soil age gradient had to be rejected. This is best explained by the finding that even lichen-moss communities which appeared to be directly soil-connected are actually linked to surface rocks only. Further CO2 efflux and isotope-based turnover measurements will be performed to determine the extent of lower plant interference with the mineral soil in the narrow sense.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
393
LICHENS LICHENS BRYOPHYTES
1 Acarospora macrocyclos 13 Ochrolechia frigida 1 Andreaea gainii
2 Caloplaca sp. 14 Placopsis contortuplicata 2 Andreaea sp.
3 Cladonia boralis 15 Pseudephebe pubescens 3 Chorisodontium sp.
4 Cladonia metacollarifera 16 Psora sp 1 4 Polytrichum comune
5 Cladonia sp. 17 Psoroma hypnorum 5 Polytrichum juniperinum
6 Cornicularia aculeata 18 Stereocaulon alpinum 6 Polytrichum piliferum
7 Himantormia lugubris 19 Rinodina cf. turfacea 7 Sanionia uncinata
8 Lecanora sp. 20 Rhyzocarpon geographicum 8 Warnstorfia sp.
9 Lecania brialmotii 21 Rhyzocarpon sp
10 Lecidea sciatrapha 22 Usnea antarctica
11 Lecidea sp 23 Usnea aurantiaco-atra
12 Mastodia tesselata 24 Xanthoria elegans Table 1: Lichens and Bryophytes Flora of Fildes Peninsula, Antarctica. Tabla 1: Flora de Líquenes y Briófitas en Península Fildes, Antárctica.
ACKNOWLEDGEMENTS Project INACH T 28-11 and F. Osorio, Y. Villagra in the identification of collections. REFERENCES Mäusbacher R Muller, J. and Schmidt, R. 1989. Evolution of postglacial sedimentation in Antarctic lakes (King George Island). Zeitschrift fur Geomorphologie, 33, 219-234. Watcham, E.P., Bentley, M.J., Hodgson, D.A., Roberts, S.J., Fretwell, P.T., Lloyd, J.M., Larter, R.D., Whitehouse, P.L., Leng, M.J., Monien, P., and Moreton, S.G., 2011. A new Holocene relative sea level curve for the South Shetland Islands, Antarctica. Quaternary Science Reviews, 2011. 30(21-22): p. 3152-3170.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
394
RELACIÓN FLEXIBILIDAD TÉRMICA DE ESTRUCTURA SECUNDARIA Y ACTIVIDAD ANTICONGELANTE DE HOMOPÉPTIDOS DE PROLINA, LISINA Y ARGININA
(Relationship between thermal flexibility of secondary structure and antifreeze activity of proline, lysine and arginine homopeptides)
Rojas, R.E.1, Carvajal-Rondanelli, P.2, Marshall, S.H.3, Guzmán, F.4 Arostica, M. 1
1Instituto de Química Pontificia Universidad Católica de Valparaíso (PUCV). [email protected] 2Escuela de Alimentos, PUCV 3Instituto de Biología PUCV 4Núcleo de Biotecnología Curauma (NBC), PUCV RESUMEN Generar péptidos anticongelantes es importante para la criopreservación de tejidos de productos cárnicos. Homopéptidos de prolina(Pro), lisina (Lys) y arginina (Arg) presentan actividad anticongelante de forma no coligativa, de forma similar a las glicoproteínas anticongelantes (AFGPs) (Carvajal-Rondanelli P,et al.2011), denominada histéresis térmica, la cual será cuantificada a través de la inhibición del crecimiento de cristales de hielo(Bang, 2013), por medio de la calorimetría diferencial de barrido (DSC). La interacción de péptidos en productos alimenticios es un área de investigación i en la industria actualmente, estas interacciones podemos verlas en los cambios estructurales de estos , cuando se analiza por dicroísmo circular (CD) de homopéptidos de prolina (Pro), arginina (Arg) y lisina (Lys) con estructura secundaria tipo poliprolina II(PPII) y poliprolina I (PPI), esta estructura se caracteriza por tener ángulos diehedrales φ= -75 y ψ= +145 para PPII y φ = -75 y ψ +160 para PPI. A fin de comparar la estabilidad térmica de estos péptidos y su flexibilidad ante los cambios en el medio. MATERIALES Y MÉTODOS Homopéptidos de Pro, Lys y Arg entre 7 y 14 residuos fueron sintetizados con metodología Fmoc en fase sólida (Houghten RA., 1985), los péptidos fueron purificados(Guzmán F.et al. 2007), analizados por RP-HPLC y su masa molecular confirmada por HPLC-MS y MALDI-TOF (Jofré et al., 2011). El análisis de DSC fue con un equipo DSC 1 con sensor HSS8 de sensibilidad ±1µW y un enfriador interno Huber TC45. Se utilizó un crisol de aluminio vacío de 40 µL sin pin para la calibración de la línea base; como control de calibración se utilizó indio y zinc antes de las mediciones de histéresis térmica (THA). Se empleó muestras de 10 µL de solución de péptido a distintas concentraciones (2, 10, 15 y 25mM) en crisoles de aluminio. La masa de las muestras fue registrada usando balanza analítica de precisión. El análisis de CD fue con un equipo J-815 equipado con Peltier. La velocidad de escaneo 50 nm/min, tiempo de respuesta 4 s, ancho de banda 1 nm y 3 acumulaciones. Se utilizaron 250 µL de muestra con concentración 1,1 mM en cubeta de cuarzo con paso óptico de 1 mm; tomando espectros sucesivos cada 5°C entre los 5°C y los 50°C, con tiempo de estabilización de 1min. RESULTADOS Y DISCUSIÓN Los homopéptidos presentan actividad anticongelante entre 0,2°C y 0,7°C, esta actividad anticongelante se hace mayor al aumentar la concentración de homopéptido. Al disminuir los puntos de nucleación aumenta la THA, homopéptidos de prolina presentan la mayor THA seguido de arginina, lisina presenta la menor THA entre estos péptidos(figura2). Los homopéptidos presentan un punto isodicroico, así como mayor definición de estructura PPII a temperaturas bajas(figura1), mientras temperaturas altas

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
395
favorecen estructura PPI. Tanto la magnitud del máximo de elipticidad como el mínimo están aumentados cuando disminuye la temperatura.
Figura 1: Influencia de la temperatura en el máximo de estructura PPII para homopéptidos de prolina, arginina y lisina.
Figure 1: Influence of temperature on the maximum PPII structure for homopéptidos proline, arginine and lysine.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
396
Figura2: Efecto de homopéptidos de Arginina, lisina y prolina entre 7 y 14 residuos en la actividad anticongelante. Figure 2: Antifreeze activity of arginine, lysine and proline homopéptidos between 7 to 14 residues.
CONCLUSIONES La asociación del homopéptido con el cristal de hielo inhibe el crecimiento de este, esto se favorece al disminuir los puntos de nucleación en la muestra. A igual concentración de homopéptido prolina presenta la mayor THA seguido de Arg y Lys. Al aumentar la concentración se aumenta la actividad anticongelante. : Al aumentar el número de residuos estos definen mejor una estructura secundaria. La

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
397
tasa de cambio para este proceso (deg *cm2 /dmol*°C) es menor para homopéptidos de prolina corroborando la rigidez de esta estructura debido a que Pro es un amino acido cíclico, en comparación con Arg y Lys tanto al relacionar los máximos como los mínimos de elipcidad. AGRADECIMIENTOS Proyecto Fondecyt 1100556 REFERENCIAS Bang J., Lee J., Murugan R., Lee S., Do H., Koh H. y Kim H. 2013. Antifreeze Peptides and Glycopeptides, and Their Derivatives: Potential Uses in Biotechnology. Drugs, 11(6), 2013-2041. Carvajal-Rondanelli P, Marshall S. and Guzman F. 2011 Antifreeze glycoprotein agents: structural requirements for activity Journal of the Science of Food and Agriculture.91(14): 2507–2510 Guzmán F., Barberis S., Illanes A. 2007 Peptide Synthesis: Chemical or Enzymatic. Electronic Journal of Biotechnology, 10(2). Houghten RA. 1985. General method for the rapid solid-phase synthesis of large numbers of peptides: specificity of antigen-antibody interaction at the level of individual amino acids. Proceedings of the National Academy of Sciences of the United States of America, 82: 5131-5135. Jofré C., Guzmán F., Cárdenas C., Albericio F. y Marshall S.H.2011. A natural peptide and its variants derived from the processing of infectious pancreatic necrosis virus (IPNV) displaying enhanced antimicrobial activity: A novel alternative for the control of bacterial diseases. Peptides, 328:852-858.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
398
HISTORIA DE LA ARQUITECTURA EN LA ANTÁRTIDA: UNA PROPUESTA DE CLASIFICACIÓN
(Architectural History in Antarctica: a proposal for classification)
Alvarez, C. E.1, Salvador, R. M.2; y Vargas, P. S. de P.3 1LPP/UFES - Laboratório de Planejamento e Projetos da Universidade Federal do Espírito Santo. Av. Fernando Ferrari n° 514, CEMUNI I sala 7. Vitória, ES – Brasil. CEP 29075-910. E-mail: [email protected] 2LPP/UFES. E-mail: [email protected] 3LPP/UFES. Email: [email protected] RESUMO O continente antártico é palco da ocupação humana a mais de um século (Friend, 2009). Neste período, a humanidade buscou, de diferentes formas, soluções construtivas ao desafio de edificar em um ambiente tão desfavorável. Conhecida por seu clima hostil com fortes ventos e baixas temperaturas, a Antártica possui também um ambiente extremamente frágil que se apresenta ainda hoje como um dos locais mais inóspitos e preservados da terra. Estas características afetam diretamente o ato de construir na Antártica, seja pela necessidade das edificações resistirem às condições ambientais, pela necessidade de garantir sobrevivência e conforto aos ocupantes ou pelas dificuldades logística de levar material e mão-de-obra, considerando que lá não existem materiais locais nem tampouco população nativa. Por não haver população nativa, não há o que se possa considerar arquitetura tradicional do continente. As edificações construídas na Antártica são resultado do conhecimento tecnológico e da bagagem cultural dos países que ali edificam (Alvarez, 1995). Aliado a isto, o fato de o meio ambiente antártico ser completamente diferente do que existe em qualquer outra parte do globo faz com que as edificações antárticas tenham como base o contexto histórico, social e cultural do país de origem. Portanto, analisar o que se edifica na Antártica é também analisar o contexto sócio-político das nações envolvidas com a ocupação do continente. A metodologia adotada nesta pesquisa consistiu em ampla revisão de bibliografia referente às edificações antárticas e ao histórico de ocupação humana na região, bem como de investigação junto a órgãos gestores de programas antárticos de diversos países. A partir da caracterização histórica, foram identificados três períodos que reúnem um conjunto de características comuns entre si e já propostos por Alvarez (1995) e denominado tempos históricos, tempos tecnológicos e tempos ambientais. Buscou-se estabelecer o contexto histórico de cada período, identificando fatores que corroboram para sua formação, bem como avaliar suas características e peculiaridades. Uma vez identificados os fatores históricos, foi estabelecido um comparativo entre as edificações produzidas em cada período segundo critérios de análise previamente estabelecidos. Esta pesquisa teve como objetivo caracterizar os três períodos propostos por Alvarez (1995), de acordo com critérios tecnológicos, culturais e antrópicos, bem como exemplificar e demonstrar as principais diferenças que caracterizam cada fase. Como principal resultado, foi possível observar, dentro do contexto da ocupação humana na Antártica, a confirmação dos três diferentes períodos – histórico, tecnológico e ambiental – propostos por Alvarez (1995), ressaltando que a evolução das edificações após 1995 enfatizam ainda mais essa divisão. As primeiras edificações construídas no ambiente antártico, ainda no fim do século XIX, serviam, basicamente, como estrutura de apoio às expedições de reconhecimento, lideradas em sua maioria por desbravadores europeus; ou como estações de apoio à atividade de pesca. Eram em sua maioria

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
399
litorâneas, caracterizadas por terem ocupação temporária, servindo a uma expedição ou atividade pesqueira por um curto período de tempo, não ofertando condições de conforto e segurança mínimas para operar durante todo o ano (Alvarez, 1995). Anterior a esse período, os vestígios de antigas edificações sugerem terem sido construídas com restos de navios e embarcações, provavelmente fruto de naufrágios. Observa-se que os tempos históricos -se por estarem inseridos numa época de desbravamentos, com um grande desconhecimento dos condicionantes da região, fazendo com que a arquitetura deste período fosse marcada pela experimentação. A princípio, adotou-se a madeira como material de construção predominante por já ser amplamente utilizada em situações semelhantes ou nos países de origem dos construtores das edificações. A madeira, enquanto constituinte dos sistemas de vedações externos, permitiu índices aceitáveis de conforto e foi capaz de assegurar a sobrevivência humana por períodos não muito longos de tempo. Além disso, permitiu a pré-fabricação dos componentes das edificações, o que foi fundamental, já que as condições ambientais são altamente adversas para a construção e outros métodos tradicionais de construção mostraram-se inviáveis (Instituto dos Arquitetos do Brasil, 2012). Um exemplo de edificação característica deste período é a cabana, construída por Robert Falcon Scott, como base de apoio à expedição de conquista do Polo Sul, empreendida em 1911 (Figura 1). A construção, que tem seus principais componentes em madeira, foi fabricada na Inglaterra e montada na Antártica e encontra-se preservada ainda hoje, sendo considerada patrimônio arquitetônico da humanidade (Friend, 2009). Posteriormente, sob a tutela do Tratado Antártico (Child, 1988), desenvolveu-se o período denominado “tempos tecnológicos”. Pode-se considerar que a partir dos anos 50 do século passado, com o aumento crescente do interesse de diversas nações pelo continente antártico e com os indícios, a época, de existência de recursos naturais em abundância, criou-se um cenário político internacional favorável a ocupação do continente. Aliado a isso, o desenvolvimento tecnológico e industrial possibilitou novas tipologias de edificação no ambiente. Surge a figura da estação de pesquisa científica, com finalidade civil e um programa muito mais complexo. Essas edificações eram, em sua maioria, desenvolvidas a partir de tecnologia de ponta para a época, com materiais de grande apelo tecnológico, inclusive com ênfase para o emprego do aço (Instituto dos Arquitetos do Brasil, 2012). Observa-se, ainda, a busca por um funcionamento mais autônomo, ou seja, com maior independência do país de origem, alicerçado nas potencialidades que as novas tecnologias construtivas induziam. Caracterizado pelo emprego de alta tecnologia disponível, as edificações dessa época possibilitaram uma ocupação de forma mais sistêmica e permanente na região, caracterizando um passo importante para a pesquisa científica desenvolvida no continente. A utilização de alta tecnologia em larga escala permitiu que as edificações se adequassem às adversidades do ambiente atuando, principalmente no sentido de oferecer isolamento térmico adequado, segurança estrutural e ao fogo, e viabilidade logística com o menor custo (Alvarez, 1995). Entre as várias edificações deste período pode-se destacar a estação Amundsen-Scot, também conhecida como "a Estação do pólo sul" (Figura 2), visto sua localização representar um marco para a ocupação humana no continente, empregando diferentes técnicas construtivas para superar as dificuldades impostas pelo ambiente. Com o domínio das técnicas construtivas necessárias à edificação na Antártica e com o aumento progressivo da segurança e do conforto nas edificações, as atenções se voltaram para o impacto ambiental que a ocupação humana e, consequentemente, as edificações provocam. Paralelamente, o protocolo de Madrid (Ministério do Meio Ambiente, 1995) estabeleceu restrições à presença humana na Antártica e elevou a discussão sobre a responsabilidade ambiental. A partir da

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
400
homologação do acordo, todos os países signatários do tratado devem cumprir requisitos ambientais para manter suas instalações de pesquisa no continente.
Figura 1: Cabaña de Robert Scott, Figura 2: Bóveda antigua de La Estación Amundsen-Scott, Figura 3: Princess Elisabeth,
Figura 4: Bharati, Figura 5: Amundsen-Scott, Figura 6: Proyecto arquitectónico ganador del concurso para la reconstrucción de la Estación Antártica Comandante Ferraz.
Figure 1: Robert Scott's hut, Figure 2: Old dome of the Amundsen-Scott Station, Figure 3: Princess Elisabeth, Figure 4: Bharati, Figure 5: Amundsen-Scott, Figure 6: Architectural design contest winner for the reconstruction of Comandante
Ferraz Antarctic Station. Fontes: Dan Satterfield/American Geophysical Union, Ann Hawthorne/ArticPhoto, Internacional Polar Foundation, BOFArchitecknen, National Science Foundation, Studio 41.
O foco, neste período, deixa de ser apenas a adoção de solução construtiva adequada às adversidades apresentadas e passa a incluir, também, a redução dos impactos ambientais e a adaptação da edificação ao meio. Questões ligadas à eficiência energética, vida útil das edificações, manejo adequado de dejetos, impacto ambiental dos sistemas construtivos adotados e desmonte e destinação adequada após o fim da vida útil tornam-se o foco dos projetos de novos edifícios no continente. Os projetos de novas edificações não apresentam mais uma solução arquitetônica similar, mas passam a utilizar a solução mais apropriada para cada caso. Surge o emprego de novos materiais industrializados,

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
401
com destaque para os derivados, principalmente, de novos plásticos. Destaca-se o uso de diversos materiais em um mesmo projeto, cada um com a finalidade de adequar-se as diferentes necessidades. Cada vez mais diferentes países têm incentivado o desenvolvimento de pesquisas no ambiente antártico e, em paralelo, surge o desafio de conciliar as preocupações ambientais com as necessidades produzidas por esta maior demanda. Observam-se, atualmente, soluções cada vez mais criativas e inovadoras na produção da arquitetura no continente antártico, e como exemplos desta diversidade de soluções podem ser citadas as estações Princess Elizabeth, da Bélgica (Figura 3), Bharati, da Índia (Figura 4), a ampliação da Amundsen-Scott, dos Estados Unidos (Figura 5), e a nova estação Comandante Ferraz, do Brasil (Figura 6), em processo de conclusão do projeto. Como conclusão e proposta de discussão, verifica-se que, vencida essa nova etapa – que os investimentos acelerados no continente indicam que será breve – e com as edificações atuando de forma harmônica com o ambiente, uma questão surge naturalmente: qual será a tônica do novo período? AGRADECIMENTOS À FAPES, ao INCT/APA e à SECIRM pelo apoio à pesquisa. REFERENCIAS Alvarez, C. E. 1995. Arquitetura na Antártica: ênfase nas edificações brasileiras em madeira. Dissertação (Mestrado). Faculdade de Arquitetura e Urbanismo, Universidade de São Paulo, São Paulo, Brasil. American Geopysical Union. 2010. Robert Scott's hut. Recuperado em 08 de julho de 2013, de http://blogs.agu.or g/wildwildscience/2010/06/16/historical-preservation-in-antarctica Arctic Photo. Dome of the Amundsen-Scott South Pole Station. (2013) Recuperado em 08 de julho de 2013, de http://www.arcticphoto.co.uk/supergal/ah/ah00/ah0063-00.htm Bof Architekten. Indische Polarstation. 2013. Recuperado em 08 de julho de 2013, de http://bof-architekten.de/de/projekte/indische-polarstation/ Child, J. 1988. Antarctica and South American Geopolitics. New York: Greenwood Press. Friend, S. H. 2009. Sustainability and habitation in Antarctica. Dissertação (Mestrado). Montana State University, Montana, Estados Unidos. Instituto dos Arquitetos do Brasil. 2012. Termo de Referência: Anexo 1 do concurso da Estação Antártica Comandante Ferraz. Recuperado em 25 de fevereiro de 2013, de http://concursoestacaoantartica.iab.org.br/bases/TERMO_ DE_REFERENCIA_EACF.pdf International Polar Foundation. 2013. Princess Elizabeth Station. Recuperado em 05 de julho de 2013, de http://www.antarcticstation.org/station. Ministério do Meio Ambiente - Secretaria de Biodiversidade e Florestas. (1995). Protocolo ao Tratado da Antártida sobre proteção do Meio Ambiente (Protocolo de Madri). Brasilia, DF National Science Foundation. 2013. Amundsen-Scott South Pole Station. Recuperado em 05 de julho de 2013, de http//www.nsf.gov/geo/plr/support/southp.jsp. Studio 41. 2013. Estação Antártica Comandante Ferraz. Recuperado em 05 de julho de 2013, de http//www.estudio41.com.br/?conteudo=projeto&id=54

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
402
NUEVOS EDIFICIOS EN ANTÁRTIDA: LA BUSQUEDA POR SISTEMAS CONSTRUCTIVOS MÁS EFICIENTES
(New buildings in Antarctica: in search of building systems efficient)
Alvarez, C. E.1, Vargas, P. S. P.2, Fukai, F. M.3 y Salvador, R. M.2
1LPP/UFES - Laboratório de Planejamento e Projetos da Universidade Federal do Espírito Santo. Av. Fernando Ferrari n° 514, CEMUNI I sala 7. Vitória, ES – Brasil. [email protected] 2LPP/UFES. Email: [email protected] 3LPP/UFES. Email: [email protected] 4LPP/UFES. E-mail: [email protected] RESUMO As limitações das ações antrópicas e os cuidados ambientais impostos pelo Protocolo de Madrid (Ministério do Meio Ambiente, 1995) impõem novos princípios para construção e operação das estações de pesquisa no Continente Antártico. Por outro lado, a crescente preocupação em torno da sustentabilidade e da economia de energia nas edificações faz com que as novas propostas de instalações voltadas para a pesquisa na Antártica tendam a considerar os aspectos ambientais como condicionantes fundamentais. Esta pesquisa teve por objetivo identificar a atual tendência das edificações antárticas produzidas no período mais recente – com ênfase nos materiais e técnicas construtivas, conforto ambiental, energia e tratamento das águas residuárias – sendo parte de seus resultados utilizados, também, nos documentos referenciais que nortearam o concurso para a escolha de projeto para as novas edificações brasileiras na Antártica. A metodologia adotada considerou a necessidade de realização de uma revisão da literatura e pesquisa junto aos Programas Antárticos de maior relevância visando à obtenção das informações essenciais para proceder a seleção das edificações consideradas referenciais. Como recorte temporal foi estabelecido o período máximo de 10 (dez) anos de construção, considerando ainda como critério adicional a disponibilidade das informações confiáveis e elencadas como fundamentais para os objetivos da pesquisa. Destaca-se que as informações, principalmente referentes aos valores das obras e áreas de construção, embora oriundos de sites considerados confiáveis, não foram confirmados pelos programas oficiais dos países de origem. A partir do primeiro recorte, foram previamente selecionadas seis edificações referenciais, analisadas sobre os seguintes aspectos: as tecnologias empregadas e os respectivos materiais construtivos básicos, o design formal da edificação, o tratamento das águas residuárias, e o sistema de energia. As informações, após confirmação da confiabilidade da fonte, foram selecionadas e sistematizadas na forma de quadros síntese, sendo mantidas no banco de dados da pesquisa as informações completas, incluindo imagens, relatórios técnicos, contatos e fontes. Como resultado, os dados pesquisados foram sistematizados de forma temática e sintetizados nas tabelas I (dados básicos das estações selecionadas), II (arquitetura), e III (sistemas construtivos, águas residuais e energia). Fonte: Broughton (2013); Berte (2013); International Polar Foundation (2013); Alfred Wegener Institute (2013); British Antartic Survey (2013) e Bof Architekten (2013).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
403
Tabla I: Informaciones generales de los edificios seleccionados.
Table I: General information selected buildings.
Tabla II: Características arquitectónicas.
Table II: Architectural features.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
404
Tabla III: Sistemas constructivos, alcantarillado y energía.
Table III: Building systems, wastewater and energy
Fonte: A3 Water Solutions GMBH (2013); Alfred Wegener Institute (2013); Berte (2013); Bof Architekten (2013); British Antartic Survey (2013); Brooks (2013); Broughton (2013); Dias (2013); International Polar Foundation (2013); Japanese Inspection Report 2010 (2013); Nitschke (2013); Tarasenko (2013);
United States Antarctic Program (2009)
CONCLUSÃO E DISCUSSÃO DOS RESULTADOS É perceptível o uso recorrente de sistemas construtivos baseados em estruturas metálicas (com exceção da estação Juan Carlos I, em monocoque, e Princess Elisabeth, em estrutura de madeira), concebidas de maneira modular e protegidas por revestimentos que as isolam da ação direta das intempéries. A concepção do sistema construtivo envolve especial preocupação com o aspecto logístico da construção buscando soluções ágeis para o transporte e montagem das edificações no menor tempo possível. Nos aspectos relacionados às tecnologias empregadas para o isolamento do ambiente interno da edificação em relação ao meio externo, verificam-se diferentes técnicas e materiais, buscando tanto a eficiência na conservação do calor interno como a minimização dos efeitos decorrentes da condensação de vapor. Outro aspecto que chama atenção refere-se ao desenho aerodinâmico das edificações mais recentes e sua elevação do solo, buscando atenuar o efeito dos fortes ventos e aproveitar o seu efeito para minimizar os problemas decorrentes do acúmulo de neve.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
405
Os sistemas de tratamento de esgoto são variados, mas normalmente baseados em sistemas de biorreatores acoplados a módulos de filtro por membrana, podendo ainda contemplar a utilização de esterilização dos dejetos por raios UV. Eventualmente também se faz o tratamento em separado das águas cinzas visando a sua reutilização de maneira secundária na estação. Os sistemas de energia, de modo geral, empregam geradores a diesel e buscam combinar outras fontes de obtenção de energia e calor tais como painéis fotovoltaicos e turbinas eólicas associados com sistemas conversores de energia e de acumulação, principalmente em tanques d’água. Concluise que há uma notória preocupação com o emprego de tecnologias comprovadamente eficientes e já testadas sem abrir mão da inovação, estabelecendo assim novos patamares de eficiência e durabilidade nas construções antárticas. AGRADECIMENTOS Ao INCT/APA e à SECIRM pelo apoio à pesquisa. REFERÊNCIAS A3 Water Solutions GMBH. Container System for wastewater treatment and drinking water purification. Disponível em: http://www.a3- gmbh.com/ NewsBASE/content_a3/frame_english.php? main_src_framereload= webpages_english/ water_treatment~001.php. Acesso em: 5 de abril de 2013. Alfred Wegener Institute. The new centre of german research in Antarctica – Neumayer Station III. Disponível em: <http://www.awi.de/en/infrastructure/stations/neumayer_station/>. Acesso em: 10 de abril de 2013. Berte, J. COMNAP Energy Management Workshop. Disponível em: <https://www.comnap.aq/Publications/Comnap%20Publications/The_Princess_Elisabeth_Station_(Berte).pdf Bof Architekten. Indische Polarstation. Disponível em: <http://bof-architekten.de/de/projekte/indische-polarstation/>. Acesso em: 10 de abril de 2013. British Antartic Survey. Halley Research Station. Disponível em: < http://www.antarctica.ac.uk/living_and_working/research_stations/halley/>. Acesso em: 10 de abril de 2013. Brooks, W. and Ferraro, J.. Sustainable Design Strategies for the Modernization of the Amundsen-Scott South Pole Station. Disponível em: <http://www.ferrarochoi.com/Publications/SUST%20DESIGN%20STRATEGIES/SUSTDESIGN%2007% 20Energy%20Efficiency.html>. Acesso em 30 de março de 2013.Broughton, K. Halley VI Antarctic Research Station. Disponível em: < http://www.hbarchitects.co.uk/projects.php?project-list=extreme&id=0 >. Acesso em: 10 de abril de 2013. International Polar Foundation. Building the station. Disponível em: <http://www.antarcticstation.org/station/construction>. Acesso em: 20 de março de 2013. Japanese Inspection Report 2010. In: XXXIV Antarctic Treaty Consultative Meeting, 2011, Buenos Aires. Anais da XXXIV ATCM, p 1-89. Disponível em: <http://www.mofa.go.jp/mofaj/gaiko/kankyo/jyoyaku/pdfs/atcm11cep10.pdf>. Acesso em: 15 de março de 2013. Ministério do Meio Ambiente. 1995. Protocolo ao Tratado da Antártida sobre proteção do Meio Ambiente (Protocolo de Madri). Secretaria de Biodiversidade e Florestas. Nitschke, A. Bharati - The Indian Research Station at Larsemann Hills Antarctica. Disponível em: <https://www.comnap.aq/Publications/Comnap%20Publications/COMNAP%20Symposium%202010%20ProceedingsA5.pdf>. Acesso em 28 de março de 2013. Tarasenko, S.. Wastewater Treatment in Antarctica. Disponível em: <http://www.anta.canterbury.ac.nz/documents/2008- 09%20projects%20GCAS/Tarasenko.pdf>. Acesso em: 16 de março de 2013. United States Antarctic Program. Master permit application. Disponível em: <http://www.nsf.gov/about/contracting/rfqs/support_ant/docs/permit_application_text_t agged.pdf>. Acesso em: 15 de março de 2013.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
406
DUAS SITUAÇÕES CONTRASTANTES DE INÍCIO DE VERÃO NO ESTREITO DE BRANSFIELD
(Two contrasting situations at the beginning of summer in the Bransfield Strait region)
Braga, E.S.1 e Castro, C.G.2 1Departamento de Oceanografia Física, Química e Geológica, Instituto Oceanográfico, Universidade de São Paulo, Pça do Oceanográfico, 191, São Paulo, Brazil. E-mail:[email protected] 2Departamento de Oceanografía, Instituto de Investigaciones Marinas, Consejo Superior de Investigaciones Científicas, C/Eduardo Cabello, 6, Vigo, Spain. Email:[email protected] INTRODUÇÃO Informações oceanográficas obtidas na região antártica por dois grupos de pesquisa (espanhol e brasileiro) que trabalharam no Estreito de Bransfield em dezembro de 1995 e em dezembro de 2007 foram comparadas com a finalidade de por em evidência as variações temporais e também, observar diferenças climáticas inter-anuais. O Estreito de Bransfield é um mar antártico semifechado localizado entre as Ilhas Shetlands do Sul e a costa da Península Antártica. O Estreito de Bransfield pode ser definido como a zona de transição entre o Mar de Bellingshausen e o Mar de Weddell. É ocupado majoritariamente por massas de água cujas propriedades são influenciadas pelos fluxos vindos dos mares adjacentes como do Mar de Bellinghausen, com águas relativamente mais quentes e menos salinas, e do Mar de Weddell, com águas mais frias e salinas (García et al., 2002). Nesta região, as principais massas de água encontradas e descritas por García et al. (2002) incluem Águas Transicionais Zonais com influência do Mar de Bellinghausen (ATB), Águas Transicionais Zonais com influência do Mar de Weddell (ATW), Água Profunda Circumpolar modificada (APCM) e Água Profunda de Bransfield (APB). Dada a sua característica de zona de transição entre dois domínios oceanográficos claramente distintos constitui uma zona altamente sensível à variações temporais. Neste trabalho, analisou-se se as variações observadas em duas situações de começo de verão austral podem corresponder à variações climáticas. MATERIAL E MÉTODOS Uma amostragem intensiva do Estreito de Bransfield foi realizada de 12 a 19 de Dezembro de 1995 no âmbito do projeto espanhol FRUELA. Em cada estação foi lançado um CTD GO MkIIIC acoplado em uma rosette General Oceanic® equipada com garrafas Niskin. Submostras para determinação de oxigênio, nutrientes inorgânicos e clorofila-a foram coletadas em profundidades padrões entre a superficie e o fundo. Oxigênio foi analisado pelo método de Winkler. Os nutrientes foram determinados seguindo a metodologia de Hansen & Grasshoff (1983) usando um autoanlizador Technicon AAII®. A clorofila–a foi medida mediante extração com 90% acetona em um fluorômetro Turner 10000R (Yentsch & Menzel, 1963). Como parte do Programa Antártico Brasileiro (PROANTAR) no âmbito do Projeto RADIOANTAR (CNPq 550048/2007-3) foram amostradas 10 estações na região do Estreito de Bransfield em dezembro de 2007. As amostras de água foram obtidas com o uso de rosette contendo garrafas tipo Niskin (General Oceanic®) e CTD (Seabird®) para obtenção dos dados de temperatura e salinidade. Recolheram-se amostras de água para análise de oxigênio, nutrientes inorgânicos e clorofila-a em profundidades escolhidas entre a superfície e o fundo. O oxigênio dissolvido foi analisado segundo o método de Winkler conforme descrito por Grasshoff et al. (1983). Os nutrientes foram determinados por

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
407
colorimetria seguindo-se a metodologia descrita por Grasshoff et al. (op.cit.) utilizando um espectrofotômetro Genesis2 da Bauch & Lomb® para fosfato e silicato e, utilizando um AutoAnalyzer II Bran-Luebbe® para nitrato. A clorofila-a foi estimada por espectrofotometria na faixa de luz visível, segundo a metodologia descrita por Strickland & Parsons (1968), utilizando espectrofotômetro Genesis 2 da Bauch & Lomb®.
Figura 1. Localização das estações realizadas em dezembro de 1995 (esquerda) e as realizadas em dezembro de 2007
(direita) na região do Estreito do Bransfield, Antártica. Figura 1. Ubicación de las estaciones realizadas en diciembre de 1995 (izquierda) y en diciembre de 2007 (derecha) en la
región del Estrecho de Bransfield, Antártica.
Salinidade (psu)
33.6 33.8 34.0 34.2 34.4 34.6
Tem
pera
tura
(C
)
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5 TBW em Dez 1995TWW e APB em Dez 1995TWW e APB em Dez 2007
Figura 2: Gráfico de Temperatura – Salinidade considerando todas as amostras de água coletadas em Dezembro 1995 e em
dezembro 2007 no Estreito de Bransfiled, Antártica. Figura 2: Gráfico de temperatura - salinidad considerando todas las muestras de agua recogidas en diciembre de 1995 y
diciembre de 2007 en el Estrecho de Bransfiled, en la Antártica. RESULTADOS E DISCUSSÃO Em dezembro 1995, a situação hidrográfica no Estreito de Bransfield estava definida pela advecção de ATB e de ATW. ATB dominaram o sector NW do Estreito de Bransfield, caracterizadas por

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
408
Temperaturas > 0°C e valores de silicato entre 72 e 82 µmol kg-1. No sector SE do Estreito de Bransfield encontraram-se ATW com temperaturas <0°C, e valores de silicato acima de 82 µmol kg-1. Os níveis de clorofila-a superficial estiveram, de um modo geral, relativamente altos com valores máximos na região de confluência de ATB e ATW (> 3 mg m-3). A situação encontrada em dezembro de 2007 foi claramente distinta. As temperaturas superficiais em nenhum ponto estiveram acima de 0°C, e além disso, as águas mais frias (< -1°C) foram observadas perto das Ilhas Shetland do Sul, onde em dezembro de 1995 registraram-se temperaturas acima de 0°C. Tanto a temperatura como a salinidade apontaram a presença de ATW em todo o Estreito de Bransfield. No entanto, os níveis de silicato foram muito mais baixos do que o esperado para estas águas. Nas camadas profundas, as situações também foram contrastantemente distintas. Em dezembro de 1995, observou-se de forma menos evidente, os núcleos de APCM e APB tanto nas propriedades termohalinas como nas biogeoquímicas. O core de APCM caracterizou-se por temperaturas e salinidades maiores de 0°C e 34,5 psu, respectivamente e por altas concentrações de silicato (> 90 µmol kg-1) e oxigênio < 280 µmol kg-1. O core de APB encontrou-se na parte SE do Estreito de Bransfield, caracterizado por baixas temperaturas (< 1 °C) e salinidades (<34,50 psu), menores níveis de silicato (<85 µmol kg-1) e altos de oxigênio (>280 µmol kg-1). Por outro lado, em dezembro de 2007, tanto as condições termohalinas como as biogeoquímicas apontaram somente a presença de APB. As condições oceanográficas estudadas demonstraram que dezembro de 2007 foi muito mais frio que a situação observada em dezembro 1995. De fato, o diagrama TS, para todas as profundidades, mostra uma água com um mínimo de temperatura (-1,5°C), que não foi observada em dezembro de 1995. Por sua vez, em dezembro de 2007, todo o Estreito de Bransfield estava dominado pela presença de APB, sem registrar, em nenhum ponto, águas com características de Águas Circumpolares. CONCLUSÃO Ainda que não tenhamos elementos suficientes para comentar sobre a influência de uma mudança climática, a análise comparativa das duas situações observadas no início do verão, mostrou que os cenários termohalinos e biogeoquímicos podem ser muito distintos na região do Estreito de Bransfield. Isto evidencia que se trata de uma região altamente sensível e que seria necessário acompanhá-la mais frequentemente, ou seja, com certa regularidade em virtude dos aspectos apontados neste estudo e diante das mudanças climáticas que estão ocorrendo. AGRADECIMENTOS Registramos nossos agradecimentos às equipes de pesquisa das campanhas FRUELA 95-96 e OPERANTAR XXVI, à tripulação do B/O Hespérides e do NApOc Ary Rongel, bem como ao CNPq, processo N. 550048/2007-3, que fincnanciou o Projeto RADIOANTAR e a equipe LABNUT-IOUSP, em especial ao Vitor G. Chiozzini.
REFERÊNCIAS García, M.A., Castro, C.G., Ríos, A.F., Doval, M.D., Rosón, G., Gomis, D. e López, O. 2002. Water masses and distribution of physico-chemical properties in the Western Bransfield Strait and Gerlache Strait during Austral summer 1995 96. Deep Sea Research II. v. 49. p. 585 – 602. Grasshoff, K.; Ehrhardt, M. e Kremling, K. 1983. Methods of seawater analysis. 2nd ed. Weinhein, Verlag Chemie. 419p. Hansen, H.P. e Grasshoff, K. 1983. Automated chemical analysis.In: Grasshoff, K., Ehrhardt, M., Kremling, K. (Eds.), Methods of Seawater Analysis. Verlag Chemie, Weinheim,pp. 347–395. Strickland, J.D.H. e Parsons, T.R. 1968. A practical handbook of seawater analyses. Ottawa: Fisheries Research Board of Canada. 311p. (Bulletin: Fisheries Research Board of Canada, n167).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
409
A ASSOCIAÇÃO DE PESQUISADORES POLARES EM INÍCIO DE CARREIRA NO BRASIL (APECS-BRASIL): AVANÇANDO E INOVANDO PELO PROGRESSO DA CIÊNCIA
(The Association of Polar Early Career Scientists in Brazil (APECS-Brazil): advancing and innovating to the science progress)
Costa, E.S.; Ivar do Sul, J.A.; Alves, E.; Caramello, N.; Almeida, M.H.; Quaglio, F.; Kerr, R.; Silva, M.
APECS-Brazil. [email protected]; www.apecsbrasil.com INTRODUCCIÓN Durante o 4o Ano Polar Internacional (International Polar Year, IPY, 2007-2008) surgiu a Associação de Pesquisadores Polares em Início de Carreira - APECS (do inglês, Association of Polar Early Carreer Scientists), de âmbito internacional e interdisciplinar com o intuito de apoiar e incentivar a formação das novas lideranças em pesquisa e educação ligadas aos ambientes polares. Mais especificamente, a APECS tem os objetivos de (1) promover o desenvolvimento de redes de pesquisa agregando efetivamente pesquisadores em início de carreira, (2) oferecer oportunidades de desenvolvimento profissional e (3) estimular atividades de educação, comunicação e difusão da ciência polar para estimular as futuras gerações de pesquisadores polares. Pelo trabalho que vem sendo desenvolvido em âmbito internacional a APECS vem ganhando cada vez mais espaço com a formação de novos Comitês Nacionais em diversas as regiões do planeta. Em 2008 foi estabelecido o Comitê Nacional da APECS no Brasil (veja mais em Ivar do Sul et al. 2011). Em cinco anos de existência, a APECS-Brasil desenvolveu com sucesso dezenas de atividades e está fortemente inserida no contexto nacional da pesquisa polar nacional. Membros da APECS-Brasil tem participado de importantes eventos nacionais e internacionais como a Reunião do Tratado da Antártica (ATCM) (maio de 2013, Bélgica). A APECS-Brasil tem ainda colaborado ativamente com a revisão de documentos de interesse nacional e internacional, e para as futuras gerações de pesquisadores e educadores polares, como por exemplo o Plano de Ação para a Ciência Antártica. Com mais de 300 membros que representam as mais diversas áreas da ciência, a APECS-Brasil é considerada um dos Comitês mais ativos dentre os comitês nacionais da APECS. Atualmente, o processo de formalização da Associação está sendo legalizado de acordo com a legislação brasileira para que as atividades possam se expandir ainda mais. Esse é um importante passo para a conquista do apoio formal de órgãos e instituições que buscam a formação das novas gerações de pesquisadores e educadores. A APECS-Brasil tem desprendido grandes esforços para estimular o desenvolvimento de atividade de educação, comunicação e difusão da ciência. Em um país com dimensões continentais como o Brasil, assuntos relacionados aos ambientes polares já chegaram à áreas distantes dos grandes centros (por exemplo Rio de Janeiro e São Paulo), como Rolim de Moura, no Estado de Rondônia, Norte do Brasil. Lá, será realizado o I Seminário Polar Internacional da Amazônia, que vai reunir professores, alunos e a comunidade em geral. É a Amazônia brasileira engajada no diálogo internacional proporcionado pelas Semanas Polares Internacionais da APECS! Este evento será ainda um evento preparatório para a X Semana Polar Internacional (XSPI), que irá ocorrer em setembro de 2013, com participação de convidados internacionais (Bélgica, Espanha, Inglaterra e Portugal) e de todo o Brasil, desde o Rio Grande do Sul até a Amazônia. Escolas de todo o Brasil podem se cadastrar gratuitamente para este evento (http://www.apecsbrasil.com/) que com certeza estará mais uma vez divulgando a ciência polar em nível nacional e internacional. Concomitantemente a XSPI, irá ocorrer o I Workshop de Desenvolvimento de Carreira, onde serão oferecidos mais de 15 minicursos. Este Workshop será aberto a todos os interessados, pesquisadores

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
410
polares ou não, pois os temas discutidos são de interesse para a formação de pesquisadores e educadores de todas as áreas do conhecimento. Logo após a XSPI, irá ocorrer o Simpósio Brasileiro de Pesquisas Antárticas (SBPA), organizado anualmente pela Universidade de São Paulo (USP). Pela primeira vez a APECS-Brasil irá coordenar as atividades do primeiro dia do evento, que será composto por apresentações orais e de painéis realizadas pelos pesquisadores em início de carreira. Está sendo organizado o III Workshop da APECS-Brasil, em Arraial do Cabo, estado do Rio de Janeiro, previsto para Setembro de 2014. Depois de duas edições bem sucedidas (I Workshop APECS-Brasil - 2010 no Rio de Janeiro, RJ; e II Workshop - 2012, em Rio Grande, RS), muitas novidades são aguardadas para o III Workshop que irá reunir pesquisadores, professores, estudantes secundaristas, militares, comunidade em geral, ou seja, todos os que se interessam pelas questões polares, incluindo o Ártico e a Antártica. O tema do III Workshop APECS-Brasil, “integrando pesquisadores de polo a pólo” tem como principal objetivo incentivar a participação de representantes de todos os comitês nacionais da APECS, reunindo no Brasil as futuras lideranças em pesquisa e em educação em nível global. É um grande passo para APECS-Brasil e ainda há muito trabalho pela frente! Para organizar e registrar todas as (muitas) atividades que os membros da Diretoria, Conselho e Participativos da APECS-Brasil desenvolvem em todo o Brasil um Informativo é produzido semestralmente (http://www.apecsbrasil.com/informativo/). Acesse e confira as novidades! Convidamos todos os que se sentirem motivados a se juntar a APECS-Brasil, participando ativamente da formação do futuro da ciência polar no Brasil, integrando a comunidade científica à comunidade geral, promovendo a ciência e motivando estudantes dos vários níveis de educação a serem “nossos futuros pesquisadores e educadores”. Vivemos um momento grandioso para a ciência polar no Brasil, exemplificado pela nova estação científica na Antártica e pelo fortalecimento da pesquisa polar de forma geral, e a APECS-Brasil está sempre presente. Nós assumimos a responsabilidade neste importante momento para ajudar os tomadores de decisão no Brasil a delinear um futuro de excelência para a pesquisa polar no Brasil. Para isso contamos sempre com o apoio dos líderes em pesquisa polar de cada país para que incentivem as iniciativas de educação, comunicação de difusão da ciência de seus pesquisadores em início de carreira por intermédio da APECS.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
411
DISEÑO DE EDIFICACIONES CON ALTA EFICIENCIA ENERGÉTICA Y UTILIZACIÓN DE ENERGÍAS RENOVABLES PARA LA ESTACIÓN PEDRO VICENTE MALDONADO
DEL INSTITUTO NACIONAL ANTÁRTICO ECUATORIANO.
(Design of buildings with high energy efficiency and utilization of renewable energy for the Pedro Vicente Maldonado Station from the National Ecuadorian Institute.)
Peña, P., Montero, A., Labus, J.
Instituto Nacional de Eficiencia Energética y Energías Renovables, Avenida 6 de Diciembre N34-32 e Ignacio Bossano, Quito, Ecuador. Email: [email protected] / [email protected] RESUMEN El Instituto Antártico Ecuatoriano (INAE), en calidad de ente encargado de la investigación en esta región natural del Ecuador, realizó un acercamiento con el Instituto Nacional de Eficiencia Energética y Energías Renovables (INER) para incluir aspectos relevantes de eficiencia energética y energías renovables en los módulos construidos de su estación, así como también, en los que se planifican construir para la estación permanente. En este artículo se presenta la primera parte del estudio que hace referencia únicamente a la inspección energética de la estación. La Estación “Pedro Vicente Maldonado” se encuentra en Punta Fort Williams de la Isla Greenwich (62°26′96″S 59°44′54″O), de acuerdo al portal web del INAE (4). Se encuentra a 400 m. de la playa y aproximadamente a 12 m. sobre el nivel medio del mar. Los módulos estudiados fueron los 1 y 2 para habitabilidad, que se encuentran unidos por una pre cámara de 5,9 m. de longitud y 3,5 de ancho, y el módulo 4 donde se ubican los laboratorios. Además, la estación cuenta con módulos para los generadores, el sistema de calefacción y para la unidad de tratamiento de aguas residuales. Las fachadas posterior y lateral Este de la Estación están protegidas por formaciones montañosas, éstas disminuyen el efecto del viento…” (5). Toda la estructura está elevada 1,5 m., es decir existe un espacio libre entre el piso de la estación y la superficie de la tierra. Existe en el interior un cielo raso de madera en el módulo de vivienda, y de asbesto en el módulo de los laboratorios y túnel. El techo presenta pendientes desde la línea central que divide la estación hacia las paredes anterior y posterior de la fachada principal de la misma. Su altura mayor es a 6,78 m. y la menor en ambas paredes, de 2,5 m.
Laboratorio. Módulo de habitabilidad. Laboratory. Habitability modules.
La investigación en cuanto a energías renovables y eficiencia energética en el Círculo Polar Antártico tiene pocos antecedentes. Algunos países pertenecientes al Tratado Antártico cuentan con estaciones científicas a lo largo de este continente, muchas de ellas con energía renovable supliendo parte de las necesidades energéticas tanto para investigación como para habitabilidad (2). Las publicaciones efectuadas hasta la fecha, muestran un gran potencial del recurso eólico en la zona, alcanzando factores de planta estimados en valores superiores al 60%, mientras que, el recurso solar no

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
412
muestra una aplicación promisoria en esta zona ya que el solamente se cuenta con energía solar durante el verano austral, teniendo factores de planta máximos de 30%, llegando a valores de 0% durante el invierno (1). En el aspecto de la energía renovable, se puede mencionar el estudio de diseño de un sistema de calefacción en base a energía eólica, realizado en base a conductividades referenciales, datos meteorológicos de la Base Arturo Prat (Chile), a 5 km de la estación ecuatoriana y requerimientos energéticos de la estación (3). El alcance planteado del estudio no considera mejorías con respecto a la orientación de los edificios, modificaciones de la edificación para incrementar su eficiencia, ni adición de otro tipo de aislantes térmicos. Por otra parte, la Universidad Católica Santiago de Guayaquil llevó a cabo otra investigación enfocada a la eficiencia energética de las edificaciones que componen la estación científica del INAE haciendo referencia al comportamiento térmico de los materiales que componen las edificaciones de la Estación Pedro Vicente Maldonado (4). Aunque dicha investigación permite tener datos más realistas acerca de la conductividad de los materiales, no se ha considerado los coeficientes convectivos a los que están sometidas las estructuras, que son dependientes de la velocidad del viento y que pueden llegar a ser más influyentes en la transmisión de calor al exterior que los componentes de las paredes. Respecto a los materiales y métodos, en la estación se realizó la monitorización de parámetros para medir el rendimiento energético de los módulos y, a la vez, se realizó también una inspección. Los equipos utilizados fueron:
• Pirómetro. Para medir la temperatura de forma puntual sobre superficies. • Luxómetro. Para medir el nivel de iluminación. • Equipo de adquisición de medidas de temperatura interior. El equipo consta de un data logger,
seis sondas de temperatura, software y una computadora. • Data logger para climas extremos. Para monitorizar parámetros en el ambiente exterior. • Data logger para adquisición de medidas de temperatura y humedad relativa en el interior de los
módulos. • Data logger para medir la ocupación de los módulos. • Higrotermómetro. Para comparar los datos obtenidos en diferentes estancias.
La adquisición de datos se realizó, en su mayor parte, de forma automática. Se instalaron equipos en los diferentes módulos, con diferentes propósitos y el resto de datos se obtuvieron mediante inspecciones in situ.
Instrumentación. Instrumentation.
Los resultados más destacados de la monitorización e inspección revelaron que el principal problema de los módulos de la estación es la rotura de puentes térmicos. De acuerdo a la monitorización de los sensores se obtuvieron las siguientes comparaciones entre la temperatura exterior y la interior de la columna metálica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
413
Temperatura ambiente exterior. Outside temperature.
Temperatura interior, en la estructura metálica del módulo 4. Indoor temperature in the metal structure of module 4.
Se puede apreciar que la diferencia de temperatura entre el exterior y el interior es muy reducida. Esto indica claramente que hay un puente térmico y que provoca pérdidas importantes de energía. Se encontraron infiltraciones de aire en los diferentes módulos de la estación. La cubierta del módulo 1 tiene infiltraciones importantes de aire. Todos los módulos tienen problemas con la rotura de puentes térmicos. Se identificaron puentes térmicos en los marcos de las ventanas, en el sellado, en la entrada de ductos a los módulos y, en especial, en la estructura metálica de los módulos.
Infiltraciones de aire. Air infiltrations.
Las principales conclusiones son que las ventanas con marco de aluminio y cristal simple no son apropiadas para climas extremos. Lo módulos, en general, deben contar con buen aislamiento en la cubierta y en la parte inferior, piso, para evitar pérdidas importantes de energía. La iluminación en los puestos de trabajo es óptima excepto en un puesto del módulo 1. El módulo que cuenta con mejores características constructivas es el módulo 4. Se tiene un solo termómetro de control de temperatura por módulo, esto no permite tener un control de temperatura por espacios y, por tanto, la eficiencia es baja. Se desperdicia energía térmica, en todos los módulos, enviándola al ambiente exterior.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
414
La energía desperdiciada puede ser aprovechada para mantener la temperatura de confort y ahorrar en consumo de energía, en este caso, de combustible. La mayor parte de la energía es demandada para calefacción de los módulos, como se indica en la gráfica siguiente. Se considera de vital importancia mejorar la eficiencia energética de los módulos, como primer paso, antes de implementar sistemas de generación de energía por medio de energías renovables.
Consumo de energía. Energy consumption.
AGRADECIMIENTOS Al Cmdt. José Olmedo y el personal de Logística de la XVII Expedición a la Antártida. REFERENCIAS Brown, C, Guichard, A. and D. Lyons. 1996. Analysis of the potential for wind and solar energy systems in Antarctica. WIND ENERGY CONVERSION. 1 (1), (1): p111-124.1. Flor Fernando (2006). Utilización de la energía eólica en un diseño preliminar de un sistema calefacción para la estación antártica ecuatoriana. Guayaquil: ESPOL. (3): p20-30. INAE. (2012). Instituto Antártico Ecuatoriano. Available: http://www.inae.gob.ec. Last accessed 1st Oct 2012. (5). Jü rgen R. Oliviera,, Thomas M. Harms, Daniël J. Esterhuyse (2007). Technical and economic evaluation of the utilization of solar energy at South Africa’s SANAE IV base in Antarctica. Renewable Energy 33, (7): p 1073–1084. Tina Tin, Benjamin K. Sovacool, David Blake, Peter Magill, Saad El Naggar, Sven Lidstrom, Kenji Ishizawa, Johan Berte. (2009). Energy efficiency and renewable energy under extreme conditions: Case studies from Antarctica. Renewable Energy. XXX (1), (6): p1-9. Universidad Católica Santiago De Guayaquil (2009). estudio de la eficiencia sobre transferencia calórica de los materiales utilizados para la construcción del módulo de laboratorios en la estación ecuatoriana “Pedro Vicente Maldonado” en la Antártida. Guayaquil: INAE. (4): p2-7. XX Reunión de Administradores de Proyectos Antárticos Latinoamericanos (2009). Tecnologías de energías renovables en la Antártida. Montevideo: RAPAL. (2): p2-5.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
415
EVIDENCIA DE LA PRESENÇA DE PAPUACEDRUS (CUPRESSACEAE, CALLITROIDEA) EN EL CRETÁCICO TARDIO DE LA PENÍNSULA ANTÁRTICA
(Fossil evidence to the presence of Papuacedrus – Cupressaceae, Callitroidea - in the Upper Cretaceous of northern Antarctic Peninsula)
Bastos, B.L. 1 Dutra, T.L. 1 Wilberger, T. 1 & C. Trevisan1
1Programa de Pós Graduação em Geologia, PPGeo. Universidade do Vale do Rio dos Sinos, Unisinos. Av. Unisinos, 950, Bairro Cristo Rei, CEP 93.022-000, São Leopoldo, RS, Brasil., [email protected], [email protected] The current geographical position of Antarctica and the lithologies identified in its ice-free areas have corroborated its past connection with other southern continents, especially between the late Cretaceous and Paleogene (Drinnan & Crane, 1990, Lawver & Gahagan, 2003). The macro-and microflora are an important tool in this respect, by providing information derived from subaerial environments, and its generally difficulty in crossing ocean barriers. Thanks to this the paleofloras are also useful in the discrimination of the distinct models suitable for the past dispersion of floras and the understanding of its modern distribution in the Southern Hemisphere (Veblen et al., 1996, Hill, 1994; Dutra, 2001). In a volcanic succession (andesitic/basaltic lavas, tuffs and lapilli) identified at Rip Point, north of Nelson Island (South Shetland Islands, Figure 1), two levels with flora (impressions of twigs and leaves and palynomorphs) allowed to reconstruct a palaeovegetation dominated by Nothofagus, and with rare conifers and ferns, characterized by the predominance of the genus Coniopteris (Dutra et al., 1998; Trevisan, 2011, Bastos, 2012). Although macrofossils are represented by fragmentary and partial preservations and attesting a weak diversity, the presence of “ancestral” types of Nothofagidites pollen grains (Dettmann et al., 1990), and the correlation with similar levels in the nearby King George Island (KGI , Zhaonai Li et al,. 1989) lithostratigraphic units, allowed to assign an age at the end of Campanian to the deposition of paleofloristic assemblage.
Figure 1. Map of King George and Nelson islands detaching the studied area at Rip Point (star). Circles and numbers point to
the previous known fossil occurrences at KGI. Numbers 1-3 and 11, indicates levels considered to represents an Upper Cretaceous (Campanian- Early Maastrictian) age.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
416
The studied material comes from the collections made in the PROANTAR (Brazil) field works and provided 45 impressions with sufficient morphological appeals to be described. In there, a single rest refers to Cupressaceae, although in the microflora pollen grains of Araucariaceae and Podocarpaceae are present. Its taxonomic description was based in Eckenwalder (2009). The Cupressaceae related sample shows strongly dimorphic foliage that allows to propose representing part of a transitional branch (Wilf et al., 2009), with two bilaterally flattened lateral leaves (5 mm wide x 20 mm long), opposite inserted and fused by the margin (informed by a sharp longitudinal scar), and three bifacially flattened and reduced facial leaves (2-1 mm x 3 mm), conspicuously keeled and basally covered by subtending lateral leaf pairs. Both facial and lateral leaves exhibit short or weakly developed acuminate apices (Fig. 2). The laterals leaf surfaces is covered by discontinuous, subparalel and longitudinal rows (stomatal bands?) and, in some sectors shows a more prominent ridge, probably corresponding to the unique main vein or to the resin canal.
Figure 2. Papuacedrus sp., part of transitional-form foliage, with large lateral leaves and keeled and short facial ones (scale bar: 5mm, sample ANTF 34-036)
These morphological characters, despite the scarce preservation of a single sector of the branch, confirm the relation with the Cupressaceae (Farjon, 2005) and the Group E from Eckenwalder (2009). This group includes the endemic modern forms Austrocedrus Florin & Boutelje (Chile), Libocedrus Endl. (New Zealand- New Caledonia), and Papuacedrus H.-L.Li. (Papua New Guinea- Moluccas). Among these, Papuacedrus distinguished by the presence of a resin canal, longitudinal and irregular bands of stomata and the apical fusion of the lateral leaves to each other, and with the basal part of the facial ones (Wilf et al. 2009, fig. 23, and 70). The presence of elongated lateral leaves, with weak developed acuminate apex and without lateral "spreading" confirm that the few preserved leaves was a part of a transitional branch and distinguished the lateral leaves here described from that of Wilf et al. (2009). The size of both lateral and facial leaves is comparable with those found in modern Papuacedrus (Wilf et al., 2009, Table 2) and distinct from the other two genera. Although its sparse and partial representation, this particular morphology allows to suggest that Papuacedrus-related forms and Cupressaceae, Section Callitroidea, were present in the areas of the Antarctic Peninsula before the end of the Cretaceous.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
417
In Antarctic Peninsula, Papuacedrus remains (P. shenii Zhou) had been described for levels of the lower Eocene of KGI by Zhou Zhiyan Haomin & Li (1994), also exhibiting comparable morphologies. Among the numerous and varied material there are young, transitional and mature branches which encourages comparisons and were related by the authors with the modern form, P. papuana (F. Mueller) Li. P. prechilensis, the diversified and complete materials, with associated cones that allow the new combination proposed by Wilf et al. (2009) comes from the Early and Middle Eocene levels of southern South America basins (Laguna del Hunco and Rio Pichileufú). In the new diagnosis they includes the holotipe of Berry (1938) to Libocedrus chilensis from similar levels, and suggested that part of Austrocedrus related materials studied by Paull & Hill (2008) to the Oligocene of Tasmania, could be linked to Papuacedrus (Wilf et al., 2009). Until the identification of the new form here described the ancientest record of the genus come from this eastern side of Southern Hemisphere, been identified in levels from the early Paleocene. In New Zealand they were registered only in the beginning of Miocene, by vegetative branchs and cuticles (Pole, 2007). The phylogenetic work of Mao et al. (2012) led to propose an origin to the Cupressacea in the Triassic, like part of the taxodioid stem, and thatthe first primitive forms of the family appears between Jurassic and Cretaceous. Escapa et al. (2008), when discuss the Jurassic fossil record of Patagonia, are in agree with this and propose that many forms before included in the Taxodiaceae to this levels, must be more certainly be related with Cupressaceae senso latu. The molecular analysis from Gadek et al. (2000) and Farjon (2005) are also in agreed with this proposition. Other comparable features indicative of primitive forms were identified in levels of the Jurassic and Lower Cretaceous of Alexander Island and Larsen Basin, in Antarctic Peninsula (Jefferson, 1982; Hathway et al., 1998), and Patagonia (Spegazzini, 1924; Del Fueyo et al., 2008). To Mao et al. (2012) yet, in the middle part of the Cretaceous, the Northern Hemisphere cupressoides clade diverges from the Gondwanan caliptróides. Quinn & Price (2003) support this proposal and suggest Papuacedrus as the first diverging lineage in the Callitroidea. Given the review of the fossil record and recent molecular analysis, the small piece of branch with Papuacedrus related morphology in the succession of Rip Point (Nelson Island) constitutes the earliest known record to the genus. And his stay until early Eocene in areas of the Antarctic Peninsula and South America, supports the maintenance of continuous landmasses between America and Australasia until this Eocene. Similar rainforests to that from the Cretaceous and early Paleogene from Antarctic Peninsula and austral South America, although currently without Papuacedrus but with other Cupressaceae components (Fitzroya cupressoides, Pilgerodendron, Austrocedus and Libocedrus spp.), still grows today in South America and New Zealand, under temperate to subtropical oceanic climates (Veblen et al., 1996; Cantrill & Poole, 2012). AGRADECIMENTOS PROANTAR-CNPq; CNPq 401780/29010-4; FAPERGS 101012-2 REFERÊNCIAS Bastos, B.L 2012. Flora Fóssil do Pontal Rip, ilha Nelson, Península Antártica: taxonomia e contexto geológico. Dissertação de Mestrado, Programa de Pós-Graduação em Geociências, UNISINOS, São Leopoldo, 99 p. Berry, E.W. 1938. Tertiary flora from the Rio Pichileufú, Argentina. Geological Society of America Special Papers, 12: 1-149. Birkenmajer, K., Delitala, M.C., Narebski, W., Nicoletti, M. & Petrucciani, C. 1986. Geochronology and migration of Cretaceous through Tertiary plutonic centers, South Shetland Islands (West Antarctica): subduction and hot spot magmatism. Bulletin of the Polish Academy of Sciences, Earth Sciences, 34:243- 255. Cantrill, D.J. & Poole, I. 2012. The vegetation of Antarctica through geological time. Cambridge University Press, New York, 480p.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
418
Del Fueyo, G.M., Archangelsky, S., Llorens, M. & Cúneo, R. 2008. Coniferous ovulate cones from the lower Cretaceous of Santa Cruz Province, Argentina. International Journal of Plant Sciences, 169(6):799-813 Dettmann, M.E., Pocknall, D.T., Romero, E.J. & Zamaloa, M.C. 1990. Nothofagidites Erdtman ex Potonie, 1960; a catalogue of species with notes on the palaeogeographic distribution of Nothofagus Bl. (Southern Beech). Paleontological Bulletin of the New Zealand Geological Survey, 60:1-79. Drinnan, A.N. & Crane, P.R., 1990. Cretaceous paleobotany and its bearing on the biogeography of austral angiosperms. In: Taylor, T.N. & Taylor, E.L. (eds.) Antarctic Paleobilogy: its role in the reconstruction of Gondwana. Springer-Verlag, pp.192- 219. Dutra, T.L. 2001. Paleoflora da ilha 25 de Mayo, Península Antártica: contribuição à paleogeografia, paleoclima e para a evolução de Nothofagus. Asociación Paleontologica Argentina, Publicación Especial, 8: 29-37. Dutra, T.L., Hansen, M.A.F. & Fleck, A. 1998. New evidences of wet and mild climates in Northern Antarctic Peninsula at the end of Cretaceous. In: 3rd Annual Conference of IGCP Project 381, Argentina, pp. 10-13. Eckenwalder, J.E. 2009. Conifers of the world: the complete reference. Timber Press, Portland, 720 p. Escapa, I., Cúneo, R. & Axsmith, B. 2008. A new genus of the Cupressaceae (sensu lato) from the Jurassic of Patagonia: Implications for conifer megasporangiate cone homologies. Review of Palaeobotany and Palynology, 151:110–122 . Farjon, A. 2005. A monograph of Cupressaceae and Sciadopitys. Royal Botanic Gardens, Kew, UK. Gadek, P.A., Alpers, D.L., Heslewood, M.M. & Quinn, C.J 2004. Relationships within Cupressaceae sensu lato: a combined morphological and molecular approach. American Journal of Botany, 87(7):1044–1057 Hathway, B., Macdonald, D.I.M., Riding, J.B., Cantrill, D.J., 1998. Table Nunatak: a key outcrop of Upper Cretaceous shallowmarine strata in the southern Larsen Basin, Antarctic Peninsula. Geological Magazine, 135:519–535. Hill, R.S. 1994. The history of selected Australian taxa. In: Hill, R.S. (ed.), History of the Australian Vegetation:Cretaceous to Recent. Cambridge Univ. Press, Cambridge, pp.390-420. Jefferson, T.H. 1982. Fossil forests from the Lower Cretaceous of Alexander Island, Antarctica. Palaeontology, 25(4):681-708. Lawver, L.A. & Gahagan, L.M.. 2003. Evolution of Cenozoic seaways in the circum-Antarctic region. Palaeogeography, Palaeoclimatology, Palaeoecology, 198:11–37. Zhao-Nai, Li, Liu Xiaohan, Zheng Xiangshen, Jin Qingmin & Li Gou 1989. Tertiary volcanism and formation of volcanic rocks in the Fildes Peninsula, King George Island, Antartic. In: GUO KUN (ed.). Proceedings of the International Symposium on Antarctic Research. Beijing, China, Ocean Press, 114-118. Mao, K., Milne, R.I.; Libing Zhangd; Yanling Penga; Jianquan Liua; Thomas, P.; Mill, R.R. & Renner, S.S. 2012. Distribution of living Cupressaceae reflects the breakup of Pangea. PNAS Early Edition, pp. 1-6. Paull, R. & Hill, R.S. 2008. Oligocene Austrocedrus from Tasmania (Australia): Comparisons with Austrocedrus chilensis. International Journal of Plant Sciences, 169:315-330. Pole, M.S. 2007. Conifer and cycad distribution in the Miocene of southern New Zealand. Australian Journal of Botany, 55:143-164. Quinn, C.J. & Price, R.A. 2003. Phylogeny of the Southern Hemisphere Conifers. In: Mill, R.R. (ed.) Proceedings of the 4th IS on Conifer 2003. ISHS Acta Horticulturae, 615:129-136. C. Spegazzini, 1924. Coniferales fósiles Patagónicas. Anuales de La Sociedad Científica Argentina, 98:125–139. Trevisan, C. 2011. Fósseis de pteridófitas das ilhas King George e Nelson. Dissertação de Mestrado, Programa de Pós-Graduação em Geologia, Universidade do Vale do Rio dos Sinos, 44 p. Veblen, T.T., Hill, R.S. & Read, J. 1996. The Ecology and Biogeography of Nothofagus Forests. Yale University Press, New Haven, USA, 403p. Wilf, P., Little, S.A., Iglesias A., Zamaloa, M.C., Gandolfo, M.A., Cúneo, N.R. & Johnson, K.R. 2009. Papuacedrus (Cupressaceae) in Eocene Patagonia, a new fossil link to Australasian rainforests. American Journal of Botany, 96:2031-2047. Zhou Zhiyan & Li Haomin. 1994. Early tertiary gimnosperms from Fildes Peninsula, King George island, Antarctica. In: Shen Yanbin (ed.) Stratigraphy and palaeontology of Fildes Peninsula, King George Island, Antarctica, Monograph 3, pp. 208-219.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
419
ESTUDIO DE LA MADERA DE Araucaria araucana (Molina) K. Koch, VARIABILIDAD Y COMPARACIÓN CON MADERAS FOSILES AFINES
(wood study of araucaria araucana (molina) k. koch, variability and comparison with related fossil woods)
Bozo, A.1, Orell, M.1, Mundaca, T.1, Torres, T.2 y M. Rallo1
1Departamento de Ingeniería en Maderas y sus Biomateriales, Facultad de Ciencias Forestales y Conservación de la Naturaleza, Universidad de Chile. [email protected] 2Departamento de Producción Agrícola, Facultad de Ciencias Agronómicas, Universidad de Chile. RESUMEN Las coníferas de la familia Araucariaceae, alcanzaron su apogeo durante los períodos Jurasico y Cretácico. Actualmente esta familia tiene tres géneros: Agathis con 13 especies, Wollemia con una especie y Araucaria, definida por Jussieu en 1789, con 19 especies restrictas al Hemisferio Sur distribuidas en Nueva Caledonia, Nueva Guinea, oeste de Australia, y Sudamérica. En este último continente existen dos especies: Araucaria angustifolia Kuntze, con una distribución acotada en el sur de Brasil y norte de Argentina, y Araucaria araucana (Mol) K. Koch., en el sur de Argentina y Chile, (Enrigth et al., 1995). Ambas especies han sido consideradas como relictas de bosques que en épocas pretéritas, tenían amplia distribución en Sudamérica y Antártica como lo evidencian los registros fósiles de maderas, polen y hojas de edades cretácicas y terciarias halladas en Chile, Argentina e islas adyacentes a la península Antártica (Cantrill & Falcon-Lang, 2001; Nishida et al., 1990, 2006; Pramparo, 1988; Stephens et al., 2005; Torres y Rallo, 1981, Torres et al., 1982, Torres y Lemoigne, 1988, 1989; Torres, 1990; Torres et al, 1994; Troncoso, 1986, 1991; Troncoso y Romero, 1993; Zamuner y Falachi, 2005). Durante el Cretácico (140-65 millones de años) y especialmente en el Paleógeno (65 a 23,5 millones de años) se encuentran algunas maderas con estructuras anatómicas similares al género Araucaria y en especial a la especie A. araucana, demostrando que la especie no ha sufrido cambios importantes en el xilema, a través de los tiempos geológicos. Esta característica genética ha permitido conocer la historia de la familia Araucariaceae, aportando al conocimiento de la paleogeografía, paleolatitudes, climas y composición de paisajes vegetales pretéritos (Florin, 1940; Krassilov, 1978). En esta investigación se estima de interés abordar el estudio anatómico desde el punto de vista de la variabilidad, considerando que la madera constituye un complejo vivo esencialmente variable, no solamente de un árbol a otro, sino dentro de un mismo individuo. Muchos de los caracteres anatómicos de la madera son estables y otros son fluctuantes y dependen de factores ambientales. Es importante entonces, registrar estas variaciones para determinar si las especies fósiles que han sido descritas en Chile y Antártica, caen en el rango de variabilidad de la especie Araucaria araucana. La importancia principal de este estudio radica en obtener por primera vez una descripción que considere la variabilidad del xilema de Araucaria araucana para establecer relaciones entre la estructura anatómica de la madera y sus símiles fósiles encontrados en el Cretácico y Paleógeno, profusamente señalados en la literatura paleobotánica de Chile y Antártica, contribuyendo al conocimiento evolutivo de uno de los árboles más importantes de los bosques chilenos. El objetivo general es estudiar la variabilidad de la estructura anatómica en la madera de Araucaria araucana (Molina) K. Koch, comparándola con maderas de especies fósiles afines. Este estudio analiza el xilema secundario de Araucaria araucana (Mol) K. Koch considerando muestras de tres árboles obtenidas a diferentes diámetros y alturas del fuste, incorporando madera de ramas y raíces, para establecer si existe variabilidad significativa dentro de la estructura anatómica de la madera y evaluar si

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
420
las descripciones de las especies fósiles afines al género Araucarioxylon Kraus, señaladas en Chile y Antártica durante el Cretácico y Paleógeno, se encuentran dentro de este rango de variabilidad. Las muestras provienen del Parque Nacional Villarrica (39º 38’ y 39º 21’ Latitud Sur y 72º 03’ y 71º 27’ Longitud Oeste). El material paleoxilológico utilizado en los análisis comparativos corresponden a 12 maderas silicificadas en los períodos geológicos Cretácico y Paleógeno, correspondientes a los holotipos o paratipos bien conservados asignadas al morfogénero Araucarioxylon Kraus (1870), eventualmente relacionados con Araucaria araucana. Las maderas fósiles provienen de Chile y Antártica y se encuentran depositadas en la colección Peleoxilológica T. Torres del Laboratorio de Paleobotánica de la Facultad de Ciencias Agronómicas de la Universidad de Chile. Previa autorización del Departamento de Áreas Protegidas de CONAF, Región de la Araucanía, se realizaron las actividades de terreno en el Parque Nacional Villarrica, con el propósito de seleccionar los árboles y recolectar las muestras de madera. Se eligieron tres árboles muertos y caídos, de los cuales se obtuvieron secciones transversales o discos de aproximadamente 3 cm de espesor, en los siguientes niveles de altura: 1,3 m (altura de pecho), 6 m y cada 6 m hasta el ápice, obteniendo un total de cinco discos por árbol y 15 rodelas en total (Figura 2). Adicionalmente, se recolectaron muestras de ramas y raíces correspondientes a cada uno de los árboles seleccionados.
Figura 1: Esquema para la extracción de discos en sentido longitudinal.
Figure 1: Scheme for the disc extraction in longitudinal sense. En cada disco o sección transversal se determinó el radio mayor de la rodela, para luego obtener una faja de 1 cm de ancho por 1 cm de espesor, tomada en dirección de la médula hacia la corteza. De cada faja se tomaron cinco cubos de 1 cm3 (o probetas), formando las unidades básicas de estudio. Los cubos fueron obtenidos en las siguientes ubicaciones; el primero adyacente a la médula, el segundo corresponde a ¼ de la longitud del radio, el tercero ½ de éste, el cuarto ¾ del radio y la última muestra adyacente a la corteza (Figura 3). Para la madera de ramas y raíces se extrajo sólo una muestra.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
421
Figura 2: Esquema de la extracción de muestras en sentido transversal.
Figure 2: Scheme of the sample extraction in transversal sense. Una vez extraídas las muestras, se procedió a la preparación de láminas microscópicas para el estudio xilológico, basado en métodos estándar (Johansen, 1940; Normand, 1972). Para la comparación de Araucaria araucana con maderas fósiles de edades Cretácicas y Paléogenas, se consideraron ocho variables consideradas relevantes en la paleoxilología para la identificación y determinación de nuevas especies, estos parámetros pueden ser cuantificados tanto en maderas actuales como fósiles, facilitando la comparación. Además las maderas fósiles se agruparon por morfogénero y por época geológica. Posteriormente estas características fueron analizadas estadísticamente. Los resultados indican variabilidad significativa del xilema secundario de Araucaria araucana. En sentido longitudinal, los parámetros que presentan variaciones son: grosor y notoriedad de anillos, largo y diámetro de traqueidas, densidad de traqueidas, diámetro de punteaduras, altura y densidad de radios leñoso y número de punteaduras por campo de cruce. En la sección transversal, la variabilidad se observa en notoriedad y grosor de anillos (mm), largo de traqueidas (mm), diámetro de las traqueidas, altura y densidad de radios. Entre las distintas secciones del árbol (fuste, raíz y ramas), el largo de traqueidas, altura de radios y número de punteaduras por campo de cruce, presentan variabilidad significativa. En cuanto a su anatomía, la especie presenta características similares a las descritas por otros autores, exceptuando la presencia ocasional de punteaduras triseriadas con disposición alterna en traqueidas, característica que no ha sido descrita en madera de Araucaria araucana. Este estudio contribuye a la paleoxiloligía, poniendo en evidencia la variabilidad que existe en algunos caracteres anatómicos usualmente utilizados para describir nuevas especies fósiles. Esto acentúa la importancia de realizar estudios de este tipo antes de describir nuevas especies. Se concluye existencia de variabilidad cuantificable en A. araucana y afinidad con 11 especies de Araucarioxylon Kraus. Las mayores similitudes entre los caracteres anatómicos de A. araucana y las maderas fósiles se encontraron con maderas del Paleógeno.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
422
Un hallazgo importante en la descripción anatómica de las punteaduras radiales de las traqueidas, es que muestran punteaduras uni, bi y triseriadas, estas últimas aunque en menor proporción no habían sido señaladas en la literatura para A. araucana. La variabilidad anatómica intra-árbol encontrada en este estudio tiene varias implicancias importantes para los estudios de paleoxilología: Se infiere que es de gran utilidad conocer la variabilidad de las especies actuales, para tener un conocimiento más amplio de las características cualitativas y cuantitativas, útiles en la identificación y comparación con especies fósiles. AGRADECIMIENTOS Al proyecto Anillo Antártico (ART-04) REFERENCIAS Barría M. A., Torres, T. 2008. Estudio Xilotómico comparativo entre Araucaria araucana (Mol) K. Koch y maderas fósiles terciarias de Patagonia y Antártica. En: IV Simposio Latinoamericano sobre Investigaciones Antárticas y VII Reunión chilena de Investigación Antártica, Libro de Resúmenes, Valparaíso, Chile. Septiembre. Libro de Resúmenes: pp. 404-408. Cantrill, D.J. y Falcong-Lang H.J. 2001. Cretaceous (Late Albian coniferales of Alexander Island Antarctica. Leaves, reproductives structuras and roots. Review of Paleobotany and Palynology 115. 119-145. Falcon-Lang, H. 2005. Intra-tree variability in wood anatomy and its implications for fossil wood systematics and paleoclimatic studies. Palaeontology. 48(1): 171-183(13). Jan, 2005. IAWA Committee, 2004. IAWA List of microscopic features for softwood identification. IAWA Journal 25(1): 1-70. Ottone, E. Y Medina, F. 1998. A Wood from Early Cretaceous of James Ross Island, Antartica. Asociación Paleotológica Argentina. (3): 291-298. Panshin A. J., y Zeeuw, C. 1980. Texbook of Wood technology. McGraw-Hill, New York 722 p. Philippe, M, Quiroz, D., Torres, T. 2000. Early Cretaceous fossil woods from Aysen area Patagonia, Chile and their bearing on the role of Araucariaceae in the Andean forest at this time. Simposio Magmatismo y Sedimentación durante el Mesozoico y Cenozoico en Patagonia. 9° Congreso Geológico Chileno, 30 Julio- 5 Agosto, Puerto Varas Chile. Torres, T. 2003. Antártica un mundo oculto bajo el hielo. Ed. Instituto Antártico Chileno. 95 p. Torres, T. G., Marensi, S. y Santillana, S. 1994. Maderas fósiles de la Isla Seymour, Formación La meseta, Antártida. Serie Científica Instituto Antártico Chileno 44: 17-38. Torres, T. y Biro, L. 1986. Xilotomía de coníferas fósiles de la isla Quiriquina. Revista Científica Comunicaciones 37:65-80. Torres, T. y Lemoigne, Y. 1988. Maderas fósiles terciarias de la formación Caleta Arctowsky, isla Rey Jorge, Antártica. Serie Científica INACH. 37: 69-107. Torres, T. y Lemoigne, Y. 1989. Hallazgos de maderas fósiles del Cretácico Superior en Punta Williams, isla Livingstone, islas Shetland del Sur, Antártica. Serie Científica INACH, 39: 9-26. Zamuner A. y Falachi, P. 2005. Agathoxylon matildense n. sp., leño araucariaceo del Bosque Petrificado del cerro Madre e Hija, Formación La Matilde (Jurasico medio), provincia de Santa Cruz, Argentina. Buenos Aires. 42 (2).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
423
ANÁLISIS DE CARBÓN VEGETAL FÓSIL EN DEPÓSITOS CRETÁCICOS DE LA PENÍNSULA ANTÁRTICA
(Análise de carvão vegetal fóssil em depósitos cretáceos da península Antártica)
Manfroi, J.¹, Dutra, T.¹ & Jasper, A.² ¹Programa de Pós-Graduação em Geologia – Universidade do Vale do Rio dos Sinos. Av. Unisinos, 950 - 93022-000, Brasil. [email protected], [email protected] ²Programa de Pós-Graduação em Ambiente e Desenvolvimento – Centro Universitário Univates. Av. Avelino Tallini nº 171 CEP 95900-000, Brasil. [email protected] INTRODUÇÃO De acordo com Belcher & McElwain (2008), as mudanças ambientais globais estão entre os maiores desafios da humanidade para o século XXI. Desta forma, a construção de cenários futuros, que demonstrem quais suas tendências globais, são de suma importância para a tomada de decisões relativas à gestão ambiental. Todavia, a atenção para estes estudos não pode estar voltada somente nos “sinais atuais”, os quais representam uma escala temporal muito restrita para a compreensão de eventos de amplo alcance. Sorokhtin et al. (2007) defendem que a deposições de charcoal podem ser extremamente úteis na caracterização e compreensão dos ambientes pretéritos e na construção de cenários futuros. Conforme Gastaldo et al. (1996) e Sorokhtin et al. (2007), a discussão atual concentra-se em definir a natureza dessas mudanças, e sua vinculação com os processos deposicionais ocorridos em áreas vegetadas do passado. De acordo com Royer et al. (2001), para este estudo são associadas diferentes áreas do conhecimento, que irão contribuir para a distinção dos diferentes processos associados (clima, vulcanismo, paleogeografia) na ocorrência dos níveis de carbonização. Assim, o estudo de processos (paleo)ambientais específicos, são a base para a reconstrução de modelos adequados e como ferramenta preditiva de futuras ocorrências similares, sejam ou não induzidas pelo homem. O fogo é um elemento modelador muito comum em ecossistemas atuais e pretéritos (Scott & Stea, 2002; Preston & Schidt, 2006; Scott, 2010; Scott & Damblon, 2010), podendo ser comparado à herbivoria, como fator de modificação dos biomas ao longo do tempo (Bond & Keeley, 2005). Evidências de incêndios na vegetação são atestadas, entre outras formas, pela presença de carvão vegetal fóssil (charcoal - sensu Jones & Chaloner, 1991; Sott & Glasspool, 2007; Scott, 2010), e identificadas no registro fóssil desde o Siluriano (Edwards & Axe, 2004; Glasspool et al., 2004), até o Quaternário (Macdonald et al., 1991). A análise de carvão vegetal permite inferências paleoambientais que vão desde a caracterização da vegetação queimada (Manfroi et al., 2011; Jasper et al., 2011), até a determinação da origem dos processos e a temperatura aproximada de suas causas (Scott & Damblom, 2010). De acordo com Scott (2000; 2010), os fatores que influenciam a frequência e a intensidade dos incêndios vegetacionais nos diferentes ecossistemas incluem a sazonalidade climática e a disponibilidade de matéria vegetal (combustível) e seus teores de umidade. Permitem avaliar ainda a relação com episódios de vulcanismo e seus tipos (Dutra, 2004; Dutra & Jasper, 2010; Lockwood & Hazlett, 2010). Jones & Challoner (1991), Scott (2000, 2010) e Scott & Damblon (2010) ainda destacam a potencialidade dos depósitos de charcoal para avaliar a variação dos níveis de carbono e oxigênio atmosférico ao longo do tempo geológico e como uma das causas mais importantes para a ocorrência de níveis de charcoal.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
424
Com base nestes argumentos, confirma-se a relevância dos estudos vinculados aos paleoincêndios, no estabelecimento de parâmetros paleoambientais para diferentes localidades, contribuindo para a reconstrução de parâmetros paleoambientais capazes de ser estendidos para construção de modelos globais.
MATERIAIS E MÉTODOS O presente trabalho analisa carvão vegetal fóssil (charcoal) proveniente das ilhas King George e Nelson, arquipélago das Shetland do Sul, Península Antártica, armazenados no NIT-Geo, do Programa de Pós-Graduação em Geologia da UNISINOS e cedidas pelo Instituto Antártico Chileno (INACH). As amostras coletadas foram analisadas em laboratório a olho nu e sob estereomicroscópio (aumentos entre 10 e 40 vezes). Aqueles fragmentos que apresentaram características de carvão vegetal [sensu Scott, 2010, (coloração preto-listrada e brilho levemente lustroso/sedoso)], foram retiradas mecanicamente do sedimento com auxílio de pinça, espátula e agulha histológica, e depositados em placas de Petry devidamente identificadas e numeradas. A análise em andamento e o exame das amostras em Microscópio eletrônico de Varredura (MEV) permitirá a definição de suas características morfo-anatômicas, assim como inferências paleoambientais, que servirão de base para a construção das hipóteses relativas às características dos paleoincêndios responsáveis pela formação do material em estudo. RESULTADOS E CONCLUÇÕES PARCIAIS Até o momento foi possível verificar a ocorrência de fragmentos de carvão vegetal fóssil (charcoal) em duas localidades, uma delas no sul da ilha King George (Pontal Price) e a outra para a área norte da ilha Nelson (Pontal Rip), o que já previamente permite inferir a ocorrência de incêndios vegetacionais nestas localidades. Para ambos os locais, os trabalhos prévios tem proposto uma idade para o final do Cretáceo (Cao Liu, 1994; Dutra, 2004). Além disso, as análises demostraram que os fragmentos vegetais carbonizados apresentam estruturas celulares bem preservadas e, pela presença de raios transversais simples, são relacionados à formas gimnospérmicas. AGRADECIMENTOS Gostaríamos de agradecer ao CNPq pela bolsa de Mestrado de Manfroi, J. e ao Instituto Antártico Chileno (INACH) pela cessão das amostras. REFERÊNCIAS Belcher, C.M.; McElwain, J.C. 2008. Limits for combustion in low O2 redefine paleoatmospheric predictions for the Mesozoic. Science 321: 1197-1200. Cao Liu. 1994. Late Cretaceous palynoflora in King George Island of Antarctic, with references to its paleoclimatic significance. In: SHEN YANBIN (ed.) Stratigraphy and Palaeontology of Fildes Peninsula, King George Island, Antarctic. Beijing, Science Press, Monograph, v. 3, p. 51-83. Dutra, T. L. 2004. Paleofloras da Antártica e sua relação com os eventos tectônicos e Paleoclimáticos nas altas latitudes do Sul. Revista Brasileira de Geociências, 34(3): 401-410. Dutra, T.L. & JASPER, A. 2010. Paleontologia da Antártica. In CARVALHO, I. (org.) Paleontologia (3rd ed.), Interciência, Rio de Janeiro, vol. 1, p. 537-572. Edwards, D.; Axe, L. 2004. Anatomical evidence in the detection of the earliest wildfire. Palaios, 19:113-128. Glasspool, I.J.; Edwards, D.; Axe, L. 2004. Charcoal in the Silurian as evidence for the earliest wildfire. Geology, 32:381-383. Jasper, A., Uhl, D., Guerra-Sommer, M., Bernardes-de-Oliveira, M.E.C., Machado, N.T.G., 2011. The Late Paleozoic charcoal remains from South America: multiple evidence of fire events in the coal bearing strata of the Paraná Basin, Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology 306, 205-218.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
425
Jones, T. P.; Chaloner, W.G. 1991. Fossil charcoal, its recognition and palaeoatmospheric significance. Palaeogeography, Palaeoclimatology, Palaeoecology, 97:39-50 Lockwood, J.P.; Hazlett, R.W. 2010. Volcanoes: Global Perspectives. Wiley-Blackwell, 552p. Manfroi, J.; Jasper.; Secchi, M, I.; Uhl, D.; Hamad, A M.B. 2011 Registro de paleoincêndios vegetacionais no afloramento Morro do Papaléo, Rio Grande do Sul, Brasil. In: Congresso Brasileiro de Paleontologia, 22, Natal, RN, Atas, pp.359-362 MacDonald, G.M.; Larsen, C.P.S.; Szeics, J.M.; Moser, K.A. 1991. The reconstruction of boreal forest fire history from lake sediments-a comparison of coal. Palaeogeography, Palaeoclimatology, Palaeoecology, 291:11-39. Preston, C.M.; Schmidt M. W.I. 2006. Black (pyrogenic) carbon: a synthesis of current knowledge and uncertainties with special consideration of boreal regions. Biogeosciences, Scott, A.C. 2000. The pre-quaternary history of fire. Palaeogeography, Palaeoclimatology, Palaeoecology, 164:281-329. Scott, A.C. 2010. Charcoal recognition, taphonomy and uses in palaeoenvironmental analysis. Palaeogeography Palaeoclimatology, Palaeoecology. 291:11-39. Scott, A.C.; Damblon, F. 2010. Charcoal: taphonomy and significance in geology, botany and archaeology. Palaeogeography, Palaeoclimatology, Palaeoecology. 291:1-10. Scott, A.C.; Glasspool, I. 2007. Observations and experiments on the origin and formation of inertinite group macerals. International Journal of Coal Geology, 70:53-66. Scott, A.C.; Stea, R. 2002. Fire sweep across the Mid-Cretaceous landscape of Nova Scotia. The Geoscientist, 12:3-6. Sorokhtin, O.G., Chilingar, G.V.; Khilyur, L.F. 2007. Global warming and global cooling: evolution of climate on Earth. In: Developments in Earth & Environmental Sciences 5. Elsevier, 313 p.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
426
LABORATORIO DE PALEOBIOLOGÍA DE ANTÁRTICA Y PATAGONIA, INSTITUTO ANTÁRTICO CHILENO, PUNTA ARENAS, CHILE.
(Antarctic and Patagonian Paleobiology Laboratory at the Chilean Antarctic Institute, Punta Arenas, Chile)
Mansilla-Vera, H.1, Levihuan, J.2, Ortiz, H.3, Jujihara, T.4 & Leppe, M.1
1Laboratorio de Paleobiología de Antártica y Patagonia, Instituto Antártico Chileno, Lautaro Navarro 1145, Punta Arenas, Chile. [email protected] 2Laboratorios Antárticos Jorge Berguño Barnes. 3Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción. 4Consejo de Monumentos Nacionales. INTRODUCCION Las investigaciones paleobiológicas entre Antártica y Patagonia desarrolladas por el Instituto Antártico Chileno (INACH) han producido, en los últimos años, un valioso incremento del registro fósil que explica parte de la historia natural pasada entre Antártica y Patagonia. Para hacer frente a este acrecentamiento el INACH ha creado durante el año 2012 un lugar dedicado a la investigación, resguardo y curación de este registro fósil. Las principales líneas de investigación que se desarrollan en el Laboratorio de Paleobiología de Antártica y Patagonia (LPAP) son paleopalinología, paleovertebrados, paleobotánica e icnología del Meso-Cenozoico de la Península Antártica y del extremo sur de Sudamérica. Para el desarrollo de estas disciplinas el LPAP ha establecido redes de cooperación nacional y extranjera, literatura especializada, espacios acondicionados y destinados a la exhibición, almacenamiento y conservación, trabajo de limpieza y consolidación, separación de materia orgánica presente en rocas sedimentarias, fotografiado macro y microscópico y dibujos en cámara clara. Así el laboratorio cuenta con modernos equipos técnicos y con personal especializado en la extracción, preparación y estudio de restos fósiles. Dentro de los espacios especializados destacan una sala de preparación de muestras para trabajo palinológico, manga extractora de polvo para el trabajo con lápices percutores para la preparación de megafósiles, una sala de colección con temperatura controlada y bandejas antideslizante, modernos microscopios ópticos y petrográficos, lupas fijas y de brazo articulado, espacio para recepción de alumnos tesistas y pasantes, y en construcción un espacio para la producción de cortes delgados de todo tipo de muestras paleontológicas (troncos, huesos), proceso que conllevara el aserrado, pulido, pegado y desbaste, para diferentes análisis. Las condiciones y colección de este laboratorio se encuentran de acuerdo a las normas establecidas por el Consejo de Monumentos Nacionales de Chile, organismo técnico del Estado que vela por el patrimonio declarado monumento nacional de acuerdo a la Ley 17.288. FÓSILES ANTÁRTICO-PATAGONICOS El desarrollo de proyectos vinculados a la conexión biológica y evolución de Península Antártica y Patagonia durante el Meso-Cenozoico ha dado como resultado la obtención de exclusivas piezas fósiles que se encuentran almacenadas en el laboratorio. Es importante mencionar que gran parte del material (Tabla 1.) es del final del Cretácico y comienzo de Paleógeno (límite que destaca por la extinción de los grandes reptiles del Mesozoico, dinosaurios, reptiles marinos y reptiles voladores, dejando nichos vacíos para el establecimiento, dispersión y diversificación de las aves y mamíferos) lo que engrandece nuestra colección e información obtenida para este lapso.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
427
Estas muestras se encuentran bajo la codificación CPAP–N° (Colección Paleontológica Antártica y Patagonia + número), descripción de la localidad y edad. Por ejemplo, la primera pluma fósil de Antártica se encuentra almacenada con el código CPAP–0006, hallada en Cerro Fósil (Fossil Hill), isla Rey Jorge, Shetland del Sur, Antártica, con una edad tentativa 47 millones de años (Eoceno Medio). Actualmente la colección cuenta con más de 3.350 fósiles, que se agrupan de acuerdo a las localidades estudiadas (tabla 1).
SITIO DESCRIPCIÓN PIEZAS EDAD FORMACIÓN EXCLUSIVIDAD
Glaciar Tyndall (Tyndall Glacier) Vertebrados 4Tithoniano-Berriasiano-Aptiano
Zapata Ictiosaurios, Peces, Pterosaurio
Punta Hanna (Hanna Point) Improntas foliares 60 Coniaciano-Santoniano Byers Angiospermas, pteridófitasCabeza de Presidente (President Head) Improntas foliares 343 Aptiano Cerro Negro Pteridófitas
Cerro Guido Improntas foliares 399Campaniano-Mastrichtiano-Daniano Dorotea Angioespermas
Las Chinas Improntas foliares, Invertebrados, Vertebrados 1460Campaniano-Mastrichtiano-Daniano
DoroteaDinosaurio, reptiles Marinos, mamiferos
Dumestre Invetebrados, Vertebrados 469 Campaniano-Mastrichtiano Dorotea Plesiosaurios, amonitesCerro Fósil (Fossil Hill) Improntas foliares, Icnofósiles 427 Eoceno Medio Cerro Fósil Pluma, aves, mamífero, equisetumTOTAL 3162 Tabla 1. Colección Paleonlógica Antártica Patagonica, Piezas fósiles que forman parte de la colección con los sitios de recuperación, formación portadora, descripción, edad y exclusividad de las piezas. Table 1. Paleontological Collection of Antartic and Patagonia. Fossil samples that form part of this collecction whit the site of collets, age, descriptions and exclusivity.
Figura 1. Tabla geológica indica el tipo de fósil que forma parte de la colección, para detalles observar tabla 1. Figure 1. Geologic time scale shows the fossil that form part of this colecctions, details in table 1.
Esta colección exhibe una diversa lista de organismos en excelente estado de preservación que incluye, entre lo más destacado, trazas vertebrados antárticos, ejemplares patagónicos de dinosaurios y reptiles marinos con preservación de partes blandas, y un abanico de restos vegetales troncos e improntas foliares de pteridofitas, gimnospermas y angiospermas, junto con un completo set muestras palinológicas los sitios investigados. En los últimos años el constante desarrollo del grupo paleontológico que involucra también la cooperación con especialistas nacionales y extranjeros, ha logrado que profesionales, personal técnico,

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
428
pasantes y estudiantes que desarrollan trabajos con material del laboratorio respondan algunas preguntas relacionadas a la evolución, distribución y establecimiento de biotas australes. Se pueden destacar la evolución de la flora austral-antártica durante el Cretácico (Leppe et al. 2012), los primeros ictiosaurios mas australes de edad Cretácica (Pardo et al. 2012), registros indirecto de vertebrados como pisadas y plumas del Eoceno en excelente estado de conservación (Mansilla et al. 2012), restos de plesiosaurios finicretacicos de la patagonia (Ortiz et al. 2012, Vogt 2012), entre otros.
Figura 1. De izquierda a derecha. Grupo paleontológico durante campaña paleontológica 2012 Patagonia, aleta ictiosaurio CPAP-0019 del Cretático Inferior (Pardo et al. 2012), sala colecciones, preparación aleta plesiosaurio, campaña Antártica
isla Snow 2011, preparación muestras paleopalinologicas. Figure 1. From left to right. Paleontological group during fieldtrip in Patagonia 2012, ichthyosaurian forefin from the Lower Cretaceous (Pardo et al. 2012), collections room, preparation of plesiosaurian forefin, Antartic fieldtrip Snow island 2011,
paleopalinological preparation room.
CONCLUSIONES El Laboratorio de Paleobiología del Instituto Antártico Chileno alberga en su colección (CPAP) fosilífera vestigios únicos que representan un parpadeo de la historia biológica entre Antártica y Suramérica durante finales del Mesozoico y comienzos del Cenozoico (este límite revolucionaría la historia de las composiciones bióticas a nivel global debido a la extinción de los grandes reptiles del Mesozoico), esta colección plasma y crea acontecimientos geológicos-paleontológicos de la estrecha y finita relación que tuvieron las biotas y de los sucesos climáticos entre estas dos masas de tierra. Hoy parte de este relato se encuentra almacenado en el Laboratorio y con este material podremos extender las discusiones con respecto a las relaciones biológicas, ecológicas y climáticas, y de los eventos de fragmentación Antártica-Suramérica que modelarían la pasada y actual distribución de algunos taxas. Los ejemplares yacen en óptimas condiciones de almacenamiento y cuidado, su curación e investigación ha dado como resultado un aumento en el interés por parte de la comunidad regional, nacional e internacional, generando alianzas y cooperación con instituciones como el Museo Nacional de Historia Natural de Chile, La Universidad de Tokio y Chuo de Japón, Universidade do Vale do Rio dos Sinos Brasil, La Universidad de Heidelberg y el Museo de Historia Natural de Karlsruhe, Alemania. La colaboración, establecimiento, visualización y trabajo del Laboratorio de Paleontología Antártica y Patagonia del INACH durante el 2012, tuvo como uno de sus resultado la organización del III Simposio Paleontológico de Chile durante el 2012, con 50 trabajos presentados durante 2 días de exposición (Leppe et al. 2012). La apertura del laboratorio a jóvenes estudiantes ha contribuido de manera significativa al desarrollo de capital humano y al avance de nuevas ideas, teorías y discusiones que enriquecen el debate sobre la relación entre Antártica y Patagonia. REFERENCIAS Leppe, M., Mihoc, M., Varela, N., Stinnesbeck, W., Mansilla, H., Bierma, H., Cisterna, K., Frey, E., Jujihara, T. 2012. Evolution of the Austral-Antarctic flora during the Cretaceous: New insights from a paleobiogeographic perspective, Revista Chilena de Historia Natural, 85, 4, 369-392. Leppe, M., Michea, W., Muñoz, C., Palma-Heldt, S. & Fernandoy F. 2012. Paleobotany of Livingston Island, Vascular Plants and Paleobotany, ISBN: 978-1-926692-98-2, Canada, 1-4.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
429
Leppe, M., Aravena, J.C. & Villa, R. 2012. Abriendo Ventanas al pasado, libro de resúmenes, III Simposio-Paleontología en Chile, Punta Arenas-Chile. Publicación del Instituto Antártico Chileno-Centro de Estudios del Cuaternario y Antártica. Mansilla, H., De Valais, S., Stinnesbeck, W., Varela, N. & Leppe, M. 2012. New Avian tracks from the Lower to Middle Eocene at Fossil Hill, King George Island, Antarctica. Antarctic Science, 24, 5, 500-506. Ortiz, H., Mansilla, H., Jujihara, T. & Leppe, M. 2012. Primer reporte de Plesiosauria (Reptilia: Sauropterygia) en estratos del Cretácico Superior en la Reserva Nacional Laguna Parrillar, Península de Brunswick, Región de Magallanes y Antártica Chilena. Libro de resúmenes II Simposio de Paleontología en Chile: 135-138. Pardo, J., Stinnesbeck, W., Frey, E., Fernández, M., Rivas, L., Salazar, C. & Leppe, M. 2012. An ichthyosaurian forefin from the Lower Cretaceous Zapata Formation of southern Chile: implications for morphological variability within Platypterygius, Palaeobiodiversity and Palaeoenvironments, 92, 2, ISSN: 1867-1594, Editorial: SpringerVerlag. Vogt, M. 2012. Sedimente und fossilien aus der obersten keide (Campanium-Maastrichtium) in der umgebung von Puerto Natales, Chile. ö ruprecht karls universität heidelberg institut für geowissenschaften, Bachelorarbeit, 61 pp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
430
EFECTOS DE EXTRACTOS ACUOSOS DEL LIQUEN U. aurantiaco Y MUSGO W. sarmentosa PROVENIENTES DE LA ANTARTICA SOBRE ERITROCITOS HUMANOS Y
MODELOS MOLECULARES DE SUS MEMBRANAS
(Effects of the aqueous extracts of the Antarctic species U. aurantiaco (lichen) and W. sarmentosa (moss) on human erythrocytes and molecular models of their membranes)
Suwalsky, M.1, Osorio, P.2, Avello, M.2, Villena, F.,3, Strzalka, K.4
1Facultad de Ciencias Químicas, Universidad de Concepción, Concepción, Chile. E-mail: [email protected] 2Facultad de Farmacia, Universidad de Concepción, Concepción, Chile. 3Facultad de Ciencias Biológicas, Universidad de Concepción, Concepción, Chile. 4Faculty of Biochemistry, Biophysics and Biotechnology, Jagiellonian University, Krakow, Poland. INTRODUCCIÓN Hoy en día se han identificado en la Antártica más de 200 especies de líquenes y 50 de briofitas; y sólo se conocen dos especies de plantas florales. El crecimiento de las plantas esta inhibido por las bajas temperaturas, pobre calidad solar, sequías prolongadas, altas radiaciones solares, en especial de rayos UV-B, y por la oscuridad del invierno (Øvstedal y Lewis, 2001). Estas condiciones extremas incrementan la formación de especies reactivas del oxígeno (ROS), por lo que las plantas y líquenes deben contener importantes cantidades de antioxidantes para protegerse del daño causado por la oxidación (Wilson y Brimble, 2008). Para poder sobrevivir los organismos desarrollan varios mecanismos de defensa, algunos de ellos involucran biosíntesis de metabolitos secundarios. Se ha reportado que los musgos y liquenes en la antártica son ricos en antioxidantes lipofilícos (Strzalka et al. 2011).Por estas razones pareció de interés estudiar la interacción de compuestos presentes en el liquenU. aurantiaco y en el musgoW. sarmentosa con membranas celulares como posibles mecanismos moleculares de sus propiedades antioxidantes. MATERIALES Y MÉTODOS U. aurantiaco y W. sarmentosa fueron recolectadas en las islas antárticas Rey Jorge y Ardley durante el verano de 2010. El musgo fue identificado por el profesor RyszardOchyra del Instituto de Botánica WladyslawSzafer de la Academia Polaca de Ciencias (Cracovia, Polonia), y el liquen por la profesora María Olech del Instituto de Botánica de la Universidad Jagiellonian (Cracovia, Polonia). Sus contenidos de tocoferoles α, γ, δ, las formas oxidadas y reducidas de plastoquinona, clorofila y carotenoides fueron determinadas mediante la técnica HPLC (cromatografía líquida de alto rendimiento). Muchas de éstas especies constituyen poderosos antioxidantes y neutralizadores de especies reactivas de oxígeno generadas por varios tipos de estrés oxidativo. A partir de estas especies se obtuvo extractos acuosos que posteriormente fueron liofilizadas. Estos extractos fueron estandarizados mediante la determinación de polifenoles totales por el método de Folin-Ciocalteu (Velioglu et al. 1998). Además, se realizaron reacciones fitoquímicas para conocer la naturaleza de algunos de los compuestos presentes en los extractos. Se determinó el efecto de los extractos sobre la morfología de eritrocitos humanos incubando distintas concentraciones de U. aurantiaco y W. sarmentosa con una suspensión de eritrocitos obtenidos de un donante sano sin tratamiento farmacológico. Las muestras se examinaron mediante un microscopio electrónico de barrido(SEM) (EtecAutoscan, Etec. Corp., Hayward, CA, USA y JEOLJSM-6380LV, Japón).También se realizó un análisis de los efectos que producen los extractos acuosos en bicapas de dimiristoilfosfatidilcolina (DMPC) y dimiristoilfosfatidiletanolamina (DMPE) (Avanti Polar Lipids ALA, USA), clases de fosfolípidos presentes respectivamente en monocapas externas e internas de membranas celulares. Para ello se incubaron ambos fosfolípidos con distintas concentraciones de los respectivos extractos a 30 ºC

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
431
y 60 ºC, tras lo cual se sometieron a difracción de rayos X en capilares especiales de vidrio (Glas-Technik&Konstruktion, Berlin, Alemania) durante 25 minutos a 19 ± 1 ºC, utilizando radiación CuKα (λ=1.52 Å) con filtro de Ni proveniente del generador de rayos X BrukerKristalloflex 760 (Alemania). Las intensidades de las reflexiones fueron obtenidas mediante un detector lineal sensible a posición (MBraun PSD 50 M, Alemania) y procesadas por el software ASA. Estas técnicas han sido utilizadas para el estudio de extractos acuosos de plantas nativas usadas con fines medicinales (Suwalsky et al, 2007, 2008, 2009). RESULTADOS Los resultados obtenidos por HPLC en muestras de U. aurantiacoindicaron la presencia de 2.19 ± 0.8 µg/g seco de α-tocoferol y de0.98 ± 0.04 µg/g seco deplastocromanol. EnW. tormentosaestas concentraciones fueron 88.4 ±5.8 µg/g seco y 3.8 ± 0.8 µg/g seco respectivamente (Strzalka et al. 2011).. El análisiscualitativo de sus extractos acuosos realizado mediante reacciones fitoquímicas reveló la presencia de alcaloides, presentes en mayor cantidad en U.aurantiaco. Por otra parte, resultó negativo para la presencia de taninos, cumarinas, flavonoides y saponinas en ambos extractos. Además,W. sarmentosa contiene gomas y mucilagos mientras que esta reacción resultó negativa para U. aurantiaco. El contenido total de polifenoles determinado por el método de Folin-Ciocalteu fue de 0.09 mM EAG (equivalentes de ácido gálico) en U.aurantiaco y de 1.56 mM EAG en W.sarmentosa. El estudio por microscopía electrónica de barrido (SEM) de eritrocitos humanos reveló cambios morfológicosinducidos por los extractos acuosos de U. aurantiacoy W. sarmentosa, en que predominó la formación de estomatocitos, caracterizados por adquirir una forma de taza. El estudio por difracción de rayos X mostró una disminución en las intensidades de las reflexiones de bicapas de DMPC tanto para U. aurantiaco como para W. sarmentosa,siendo mayores los efectos inducidos por U. aurantiaco; sin embargo, ninguno de estos extractos indujo cambios significativos en DMPE. DISCUSIÓN Los polifenoles son extensamente estudiados por sus conocidas propiedades antioxidantes.La cantidad de polifenoles totales encontrada en U. aurantiaco y W. sarmentosa fue bastante menor que las que se encuentran en plantas mayores (Strzalka et al. 2011). El estudio por microscopía electrónica de barrido mostró en el caso de U. aurantiaco una fuerte interacción con la membrana de los eritrocitos induciendo principalmente la formación de estomatocitos. Estos se originan por la inserción de moléculas presentes en el extracto con la monocapa interna de la membrana. A medida que aumenta la concentración se observa un mayor efecto, el que finalmente provoca la lisis celular. Por otra parte, W. sarmentosa induce la formación mayoritariamente de estomatocitos con crenaciones, lo cual indicaría una interacción del fitocomplejo tanto con la monocapa interna como externa de la membrana celular. Sin embargo, sus efectos fueron de menor intensidad que los inducidos porU. aurantiaco. El estudio por difracción de rayos X mostró disminuciones en las intensidades de las reflexiones de DMPC para ambos extractos, siendo estos efectos estructurales mayores con U. aurantiaco,y prácticamente no se observó ningún efecto sobre DMPE. DMPC es una clase de fosfolípido que se encuentra preferentemente en la monocapa externa de la membrana de los glóbulos rojos humanos. Sin embargo, los resultados obtenidos por SEM indicaron la formación preferente de estomatocitos. De acuerdo a la hipótesis del par-bicapa (Sheetz y Singer), estoimplicaría que los componentes moleculares se ubicaron en la monocapa interna de la membrana de los eritrocitos. Cabe entonces concluir que los compuestos presentes en los extractos acuosos de ambas especies interaccionan preferentemente con proteínas ubicadas en la monocapa interna de la membrana de eritrocitos.La

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
432
localización de estos compuestos en las bicapas de membranas celulares y la consecuente restricción en la fluidez podríanexplicar sus capacidades para neutralizar los efectos oxidativos de radicales libres.Sin embargo, se hace necesaria la identificación de estos compuestos y la realización de pruebas específicas de las capacidades antioxidantes para determinar el verdadero potencial de las especies estudiadas. CONCLUSIONES El líquenU. aurantiaco y el mugo W. tormentosa provenientes de la Antártica contienen antioxidantes lipofílicos constituidos principalmente por tocoferoles y plastocromanol. Extractos acuosos de las especies en estudio contienen polifenoles y alcaloides. Los fitocomplejos inducen cambios en la morfología de eritrocitos humanos, los que se explican por su inserción en la monocapa interna de sus membranas. Los resultados obtenidos por difracción de rayos X, indican que los fitocomplejos interaccionan con DMPC, clase de lípido que se ubica preferentemente en la monocapa externa de eritrocitos humanos. Ambos resultados permiten concluir que los componentes de los extractos acuosos interaccionan preferentemente con proteínas localizadas en la monocapa interna de las membranas de glóbulos rojos humanos. AGRADECIMIENTOS Proyecto FONDECYT 1130043. REFERENCIAS D.O. Øvstedal,y R.I. Lewis Smith.2001. Lichens of Antarctica and South Georgia. Cambridge University Press, Cambridge. M. P. Sheetz, S. J. Singer 1974.Biological membranes as bilayer couples.A molecular mechanism of drug-erythrocytes interactions, Proceedings of the National. Academy of Sciences U. S. A. 71: 4457-4461. K. Strzalka, R. Szymanska, y M. Suwalsky. 2011. Prenyllipids and pigments content in selected Antarctic lichens and mosses.Journal of the Chilean Chemical Society, 56:808-811. M. Suwasky, P. Orellana, M. Avello, F. Villena. 2007. Protective Effect of Ugnimolina eTurcz against Oxidative Damage in Human Erythrocytes. Food and Chemical.Toxicology45: 130-135. M. Suwalsky,P. Vargas, M. Avello, F. Villena, C.P. Sotomayor. 2008. Human erythrocytes are affected in vitro by extracts of Aristoteliachilensis (Maqui) Leaves, International Journal of Pharmaceutics,363: 85-90. M. Suwalsky, K. Oyarce, M. Avello, F. Villena, C.P. Sotomayor. 2009. Human erythrocytes and molecular models of cell membranes are affected in vitro by Balbisiapeduncularis (Amancay) extracts. Chemico- Biological Interactions 179: 413-418. Y. Velioglu, G. Mazza, L. Gao, yB. Oomah. 1998. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products.Journal of Agricultural and Food Chemistry, 46: 4113-4117. Z.E. Wilson y M.A. Brimble. 2008. Molecules derived from the extremes of life. Natural Product Reports, 26:44-71.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
433
EFECTO DEL GRADIENTE DE TEMPERATURA EN LA TASA FOTOSINTETICA DE SPHAGNUM Y PROYECCIONES COMO COLONIZADOR DE LA PENÍNSULA
ANTÁRTICA E ISLAS.
(Effect of a temperature gradient on the photosynthesis rate of Sphagnum and its projections as a colonizer in the Antarctic Peninsula and islands)
Arancibia, P1., Heredia R.1,Torres, C1, Vera, J1
1Depto. de Ciencias Básicas, Universidad del Bío-Bío, Avda. Andrés Bello s/n, Chillán, Chile. E-mail: [email protected]
INTRODUCCIÓN La península Antártica y sus islas (Lat. 61ºS hasta el polo Sur y Long 74º W y 54º E) han experimentado un aumento de temperatura en medio grado Celsius cada década durante los últimos 50 años, debido al calentamiento climático. Es así como esta zona es uno de los lugares de la Tierra con la tasa de calentamiento global más acelerado del planeta (Clarke et al, 2007). Además, la Península Antártica posee la posición geográfica más septentrional del continente, rodeada de una extensa área marítima lo que permite que ésta tenga un clima menos riguroso así como una estación de crecimiento vegetal más larga que los sitios más cercanos al polo, que favorece el establecimiento de una biota terrestre relativamente diversa (Chown, 2012). Por otro lado, su cercanía al continente americano, nos permite inferir que éste último sea considerado una fuente potencial de propágulos vegetales inmigrantes con claras posibilidades de llegar a ser viables en la Antártica. En consecuencia, tanto el clima favorable y porciones de terreno desprovistos de hielo, hace factible que las especies nativas estén expandiendo su rango de distribución y abundancia en nuevas áreas (Chown, 2012) y que el cambio climático está desplazando especies cuyo crecimiento y desarrollo depende de la temperatura como factor principal (Du et al., 2011). Sphagnum es un musgo cuya distribución latitudinal, desde Valdivia hasta el Cabo de Hornos, hace probable que la temperatura sea un factor que influya en su desarrollo. Es por ello que consideramos que Sphagnum podría colonizar áreas de latitudes más altas buscando nichos favorables para su desarrollo. Se comparó la tasa fotosintética de Sphagnum magellanicum y Sphagnum subnitens como factor respuesta al cambio de temperatura, bajo condiciones controladas de laboratorio. MATERIALES Y MÉTODOS Las muestras fueron colectadas en Quellón, Chiloé (Lat. S 43° 0.5801, Long W 73° 30.350). Se trasladaron en bolsas estériles a 15-18°C, en condiciones de temperatura similar a las encontradas en el lugar de muestreo. En el laboratorio las muestras fueron almacenadas en un incubador (Precision Model 818) iluminado a 300 µE*m-2*s-1 y a 15°C temperatura. El pH de las muestras al momento de colectar fue de 3.9. Las determinaciones taxonómicas y ecología general de ambas especies fueron determinadas utilizando la literatura de Clymo (1982) y Crum (2004). Previo a medir las tasas de fotosíntesis se aclimataron las plantas de Sphagnum a la temperatura a la cual serían medidas (1°, 4°, 8° y 15°C), durante una semana. Para medir las tasas de fotosíntesis se utilizó el diseño experimental de Arancibia (2008). Las mediciones de fotosíntesis y de respiración fueron realizadas en condiciones controladas de laboratorio a una irradiación constante de 300 µE*m-
2*s-1 lograda con luz fría mediante un iluminador KL 1500 LCD de Zeiss, y medida con un quantómetro LiCOR 250. La temperatura fue constantemente monitoreada y la variación mantenida a ± 1°C. Se midió la evolución y el consumo de oxígeno de Sphagnum con un oxigenómetro OxiCal-SL WTW durante 30 minutos cada uno. Los filamentos de Sphagnum previamente pesados, eran sumergidos en agua destilada, cuidando de extraer todas las burbujas, en una cámara de volumen conocido. Al terminar de medir respiración los filamentos eran trasladados a un contenedor para su

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
434
congelación y liofilización inmediata. Para medir fotosíntesis, se realizaron lecturas cada cinco minutos por al menos 30 minutos con el fin de obtener una línea de regresión con una pendiente cuyo valor era mg de O2*L -1. La tasa de respiración era medida de la misma forma pero el conjunto de Sphagnum sumergido dentro de la cámara era cubierto con una malla que impedía la llegada de luz. RESULTADOS Por observaciones de terreno, las dos especies de Sphagnum cohabitan en el sector de los pomponales de Quellón. Sphagnum magellanicum se encuentra en lugares más inundados. Sphagnum subnitens se encuentra en lugares más altos y secos. Los óptimos de temperatura para ambas especies se encuentran alrededor de 1°C y ambas especies poseen mayores tasas de respiración a medida que la temperatura aumenta. Sin embargo, las mayores tasas de fotosíntesis neta las registra S. subnitens a cualquier temperatura (Fig. 1 y 2)
Fig.1 Tasa fotosíntesis y de respiración de Sphagnum magellanicum, bajo condiciones controladas de laboratorio, irradiación de 300 µE*m-2*s-1, pH 3,9. Cada barra representa la media aritmética de tres replicados. Los errores estándar se representan por una pequeña barra vertical. La ANOVA muestra valores significativos al p<0,05 en cada una de las temperaturas medidas. Fig.1. Photosynthesis and respiration rates of Sphagnum magellanicum, under laboratory controlled conditions, irradiance of 300 µE*m-2*s-1, pH 3,9. Each bar represents the average of three replicates. Standard errors are represented by a little vertical line above each bar. The ANOVA shows significant values to the level of p<0,05 at each temperature.)
Fig.2 Tasa fotosíntesis y de respiración de Sphagnum subnitens, bajo condiciones controladas de laboratorio, irradiación de 300 µE*m-2*s-1, pH 3,9. Cada barra representa la media aritmética de tres replicados. Los errores estándar se representan
por una pequeña barra vertical. La ANOVA muestra valores significativos al p<0,05 en cada una de las temperaturas medidas.
Fig.2. Photosynthesis and respiration rates of Sphagnum subnitens, under laboratory controlled conditions, irradiance of 300 µE*m-2*s-1, pH 3,9. Each bar represents the average of three replicates. Standard errors are represented by a little vertical
line above each bar. The ANOVA shows significant values to the level of p<0,05 at each temperature.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
435
DISCUSIÓN La tasa fotosintética de ambas especies se ve afectada con la temperatura, hallándose el punto de compensación de ambas alrededor de 8°C por lo que sobre esa temperatura la viabilidad de éstas no es posible. S. magellanicum es más sensible al cambio de temperatura que S. subnitens, lo cual puede explicarse debido a que éste último retiene mayor cantidad de agua en sus tejidos. CONCLUSIONES La temperatura óptima de estas especies en condiciones de laboratorio es posible alcanzarla en la Península Antártica en las estaciones de verano y en el continente, ambas especies de Sphagnum se someten en el invierno a temperaturas bajo cero al igual que en la Península Antártica. Es por ello que la temperatura no sería un factor que podría limitar el desplazamiento de esta especie hacia lugares como la Península Antártica. Por otro lado, es posible que la luz pueda constituir un factor limitante y también el fotoperiodo debido a que ambos factores son extremos en la Antártica y difieren de las encontradas en el continente. AGRADECIMIENTOS Fondecyt 1120619, C. Concepción-Chillán: R .Ávila, W. Chaperón y R. Enríquez REFERENCIAS Arancibia, P. 1994. Physiological ecology of Mougeotia (Zygnemataceae) from an experimentally acidified lake. Tesis de la Universidad de Wisconsin digitalizado en 2008. Chown, S., Huiskesb, H.L., Niek J., Gremmenb M., Leea, J.E., Teraudsa, A., Crosbied, K., Frenote, Y., Hughesf, K.A., Imurag, S., Kieferc, K., Lebouvierh, M., Raymonde, B., Tsujimotoi, M., Warec, C., Van de Vijverk, B. , y Bergstromc, D.M. 2012. Continent-wide risk assessment for the establishment of nonindigenous species in Antarctica. PNAS 109 (13): 4938–4943 Clarke,A., Murphy, E., Meredith, M., King, J., Peck, L., Barnes, D. y Smith, R. 2007. Climate change and the marine ecosystem of the western Antartic Peninsula. Phil.Trans.R.Soc 362:149-166. Clymo, R.S., and Hayward, P.M. 1982. The ecology of Sphagnum. In Bryophyte ecology. Edited by A.E.J. Smith. Chapman & Hall, New York. pp. 229–289 Crum, H. 2004. Mosses of the Great Lakes Forest. 4th ed. The University of Michigan Herbarium, Ann Arbor. 592 pp. Du, H., Wu, Z., & Li, M. 2011. Interdecadal changes of vegetation transition zones and their responses to climate in Northeast China. Theor Appl Climatol 106:179–188

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
436
Colobanthus quitensis DE LA MARISMA, UNA NUEVA POBLACIÓN PARA LA COLECCIÓN GENÉTICA DE LA ESPECIE
(Colobanthus quitensis from Marisma, a new population for specie’s genetic collection)
Cuba-Díaz, M.1, Acuña, D.1, Klagges, M.1, Dollenz, O.2, Cordero, C.3
1Laboratorio de Biotecnología y Estudios Ambientales, Depto. Cs. y Tecnología Veg., Universidad de Concepción, Campus Los Ángeles. 2Depto. Cs. Y Recursos Nat., Universidad de Magallanes, Chile. 3Laboratorio de Genética Molecular Vegetal, INTA, Santiago, Chile.e-mail: [email protected]. INTRODUCCIÓN Uno de los sistemas modelo más interesante para el estudio de la especiación y la evolución de plantas proviene de grupos que se distribuyen ampliamente y en condiciones adversas, como es el caso de Colobanthus quitensis (Kunth) Bartl., con su amplia distribución geográfica y poblaciones nunca antes estudiadas. C. quitensis es la única dicotiledónea que crece de forma natural en la Antártica y algunas de sus propiedades para sobrevivir en las condiciones inhóspitas del ecosistema terrestre Antártico han sido descritas por varios estudios. Recientemente, ha sido objeto de investigaciones centradas en la comparación de poblaciones antárticas con otras a lo largo de su amplia distribución geográfica, que va desde el sur de México hasta la península Antártica, y desde los 0 m. s.n.m. al sur hasta los 4200 m. n.s.m. al norte (Moore 1970, Lewis-Smith 2003). Al comparar la morfología y la variabilidad genética inter-poblacional, se ha observado un patrón de diferenciación entre poblaciones insulares Antárticas y de Chile continental, con dos grupos completamente separados (Cordero 2012, Klagges et al. 2012). La variabilidad genética se relacionó con un bajo flujo génico entre las poblaciones y la fragmentación geográfica entre estas (Cordero 2012). Estas diferencias entre poblaciones, pueden ser el resultado de las interacciones de varios mecanismos, los cuales están directamente relacionados con el historial de colonización, el flujo de genes, la deriva genética y la selección bajo condiciones ambientales que pueden llevar a la adaptación local (Jolivet y Bernasconi 2007), esto puede además, dar origen al desarrollo de geoecotipos si existen barreras geográficas que aíslan a las poblaciones. Considerando las relaciones entre la Antártica y el continente Sub Americano, contar con un banco de germoplasma de individuos provenientes de diferentes regiones sudamericanas, permite la conservación de la variabilidad génica y fenotípica representativa de la especie. El objetivo de este trabajo es describir una nueva población incluida recientemente en varias investigaciones comparativas entre poblaciones de C. quitensis. La población es identificada como la Marisma o pPA; se muestra una breve descripción del sitio de origen, algunos aspectos de la morfología externa en condiciones naturales y jardín común, y su respuesta al cultivo de tejidos como método de propagación y de introducción al banco de germoplasma del Laboratorio de Biotecnología y Estudios Ambientales, Campus Los Ángeles, Universidad de Concepción. MATERIALES Y MÉTODOS Área de estudio y descripción del sitio: El área de estudio se ubica aproximadamente a 28 km al sur de Punta Arenas. El sitio se georreferenció y se colectaron imágenes digitales y material vegetal para realizar una breve descripción del área y especies asociadas a C. quitensis. Se tomaron muestras de suelos asociados a las raíces para su análisis químico en un laboratorio especializado. Morfología y propagación: En el laboratorio las plantas se separaron cuidadosamente del suelo para los respectivos análisis. La propagación vegetativa se realizó en recipientes plásticos en una mezcla de suelo:turba:vermiculita (3:2:1) en cámara de crecimiento a 15±2 ºC, HR de 75-85%, fotoperiodo 16/8 e intensidad lumínica de 100±20 µmol m-2 s-1, fueron fertilizadas con Phostrogen (0,2 g L-1 cada 10 días).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
437
Las mediciones morfológicas se realizaron en plantas recién colectadas (natural) y pasados tres meses después de la propagación, según Klagges et al. 2012. Los datos normalizados fueron analizados mediante un ANOVA de una vía y un test de Tukey para detectar diferencias significativas o mediante estadística no paramétrica detectando diferencias mediante el Mann-Whitney U Test. Germinación y cultivo in vitro: Semillas colectadas desde terreno o en laboratorio fueron mantenidas a 4ºC por una semana, posteriormente se realizaron varios tratamientos de estratificación en frío (Acuña et al. 2012), o mediante escarificación ácida (H2SO4 al 1%) y sembradas en medio MS (Murashige & Skoog 1962). Para la propagación in vitro se probaron diferentes combinaciones hormonales según Zúñiga et al. (2009) y Cuba-Díaz et al. (2013). Los frascos de cultivo se colocaron en cámara de crecimiento con un fotoperiodo 16/8 y a 21±2ºC. Se evaluó: amarillamiento de plántulas o brotes, Nº. de brotes nuevos, Nº. de raíces nuevas y síntomas de estrés a los 42 días de cultivo. El análisis estadístico se realizó con GraphPadPrism 6.0 para Windows (GraphPad Software, La Jolla California USA, www.graphpad.com). RESULTADOS Y DISCUSIÓN C. quitensis se encuentra en ambientes húmedos tanto de agua dulce como salobres (Pisano, 1976). Esta población descrita por primera vez se ubica en el Sector Agua Fresca, a 28 Km al sur de Punta Arenas (52º22’S; 70º58ºW), en un estuario inundable por altas mareas (Fig. 1A) con un estrecha marisma de sedimentos limo arcillosos cubierta por una vegetación herbácea (Fig. 1B) donde además podemos encontrar: Plantago barbata, P. maritima, Sarcocornia fruticosa, Triglochinconcinna, Puccinellia biflora, Armeria marítima ssp. Andina.
Figura 1. Sitio Agua Fresca, población la Marisma (pPA). A: asociación de C. quitensis y P. barbata sobre marisma con
sedimentos limo arcillosos, P. biflora gramínea indicadora de ambiente salino. B: Acercamiento a la formación vegetal con C. quitensis al centro.
Figure 1. Agua Fresca site, Marisma (pPA) population. A: C. quitensis and P. barbata association on clayey silt marshlands sediment, P. biflora, grass saline environment indicator. B. Approach to plant formation with C. quitensis at the center.
C. quitensis es muy frecuente en pastizales costeros de las islas occidentales patagónicas sobre depósitos fluviales inundables por las altas mareas y menos frecuente sobre rocas litorales de las islas (Dollenz, 1982), se ha descrito desde la laguna San Rafael (Pisano, 1988) por el norte hasta las islas Diego Ramírez en el extremo sur archipielágico (Pisano & Schlatter, 1981). La abundante presencia de P. biflora (Fig. 1A) es un indicador de salinidad del suelo, lo que se corrobora por el alto contenido de bases intercambiables y de la suma de bases observados en el análisis químico del suelo. Aunque tanto el pH (6,73) como el contenido de materia orgánica (2%) se consideran dentro de los niveles medios ambos están en el límite de lo que podría considerarse un suelo salino donde predomina el tipo arcilloso, no obstante, más estudios son requeridos. La comparación de parámetros morfológicos evidenció diferencias significativas en los caracteres evaluados (Tabla 1), observándose un mayor desarrollo radicular en condiciones naturales, quizás debido a la baja disponibilidad de nutrientes en las capas más superficiales del suelo, en cambio, en condiciones controladas con mayor disponibilidad de nutrientes y agua, las plantas presenta mayor desarrollo foliar.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
438
Parámetros Morfológicos pPA Natural pPA Jardín común
Longitud de área foliar 2,00 ± 0,20a
3,17 ± 0,69b
Longitud de raíz principal 9,17 ± 3,26a
5,14 ± 0,90b
Diámetro de cuello 0,22 ± 0,19a
0,12 ± 0,03b
Largo de hoja 0,71 ± 0,06a
1,88 ± 0,30b
Ancho de hoja 0,12 ± 0,03a
0,10 ± 0,02a
Tabla 1. Características morfológicas (media ± SD) de la población pPA de C. quitensis en condiciones naturales y en
condiciones controladas (jardín común). Letras diferentes significa p < 0,05. Table 1. Morphology characteristics (mean ± SD) of pPA C. quitensis population from natural and common garden
conditions. Different letters corresponds to p< 0,05.
No tenemos evidencias de trabajos previos con comparación entre plantas en condiciones naturales y propagadas en condiciones controladas. Moore (1970) describe a C. quitensis en condiciones naturales con hojas de 0.5 a 2 cm de largo y 0.8- 1.5 mm de ancho. Gianoli et al. (2004) observaron que en jardín común plantas provenientes de Antártica podían alcanzar 9,9 mm de largo y 1,32 mm de ancho en sus hojas. Al comparar poblaciones de C. quitensis, Klagges et al. (2012) observaron que plantas provenientes de pPA alcanzaban una longitud de hoja de 21,2 mm y un ancho de 1,11 mm. En C. quitensis se ha reportado un bajo porcentaje de germinación, 5,5% en bancos de semillas naturales (Ruhland y Day, 2001), en este estudio se logró un 44% de germinación con la estratificación, sin encontrar diferencias significativas entre los tratamientos usados, este porcentaje se incrementó a un 87% cuando las semillas fueron escarificadas con ácido sulfúrico. La escarificación ha demostrado su efectividad para disminuir la dureza y acelerar el proceso de germinación. Estos resultados concuerdan con los publicados por Orozco et al. (2010) y Pietrosemoli y Mendiri (1997), quienes reportan como mejor método de escarificación el H2SO4 en Hymenaea courbaril L. y Clitorea ternatea L. obteniendo valores de germinación 90% y 59%, respectivamente.
En la Fig. 2 se muestra la respuesta a las combinaciones hormonales usadas, en BAP 0,5 mg L-1, se estimula el desarrollo de nuevos brotes y se evita la aparición de síntomas de estrés, con 50% menos de amarillamiento que los medios propuestos por Zúñiga et al. (2009), en este medio se alcanza una inducción de nuevos brotes en un 75% con un notable 25% de inducción de sistema radicular (Fig. 2). La combinación y concentración de hormonas para la inducción de la regeneración de tejidos es un aspecto relevante, cuando se requiere optimizar un método de propagación para una especie determinada. La respuesta fisiológica del explante, depende de la especie, encontrándose en numerosos casos diferencias entre cultivares, subespecies y ecotipos. Zúñiga et al. 2009, usaron C. quitensis provenientes de una población antártica, lo que podría explicar las diferencias encontradas. En observaciones previas, hemos comprobado que poblaciones de C. quitensis no responden igual al cultivo de tejidos, e incluso la germinación de semillas in vitro presenta porcentajes y tiempos de germinación diferenciales entre poblaciones (Acuña et al., 2012). CONCLUSIONES Se describe una nueva población de C. quitensis, con una alta capacidad para la propagación en condiciones controladas e in vitro, que enriquece el germoplasma representativo de la variabilidad genética y morfológica mostrado por la especie.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
439
AGRADECIMIENTOS Investigación financiada por proyecto INACH T_03-09. REFERENCIAS BIBLIOGRÁFICAS Acuña, D., Matamala, F., Cuba-Díaz, M. 2012. Differential seed germination capacity of Colobanthus quitensis (Kunth) Bartl. (Caryophyllaceae) populartions. VII Reunión Biología Vegetal, Programa y Resúmenes p26. Cordero, C. 2012. Caracterización y análisis de variabilidad morfológica y genética en poblaciones de Colobanthus quitensis (Kunth) Bartl. (Caryophyllaceae). Memoria para optar el título de Ingeniero en Biotecnología Vegetal. Universidad de Concepción, Los Ángeles, Chile 52 pp. Cuba-Díaz, M., Acuña, D., Cordero, C., Klagges, M. 2013. Optimización de parámetros para la propagación in vitro de Colobanthus quitensis (Kunth) Bartl. Gayana Botánica (en prensa). Dollenz, O. 1982. Estudios fitosociológicos en el archipiélago Cabo de Hornos. III. Relevamientos en la isla Deceit. Ans. Ins. Pat. (Punta Arenas, Chile), 13: 145-151. Gianoli, E., Zuñiga-Feest, A., Reyes-Diaz, M., Bravo, L., Corcuera, L. 2004. Ecotypic differentiation in morphology and cold resistance in populations of Colobanthus quitensis from the Andes of central Chile and the maritime Antarctic. Arctic, Antarctic and Alpine Research 36: 470-475. Jolivet, C., Bernasconi, G. 2007. Molecular and quantitative genetic differentiation in European populations of Silena latifolia (Caryophyllaceae). Annals Bot. 100:109-107. Klagges, M.; Matamala, F., Cordero, C. Sepúlveda, J., Cuba-Díaz, M. 2012. Colobanthus quitensis populations have morphological differentiations which might evidence a habitat ecotype formation. VII Reunión Biología Vegetal, Programa y Resúmenes p59. Lewis-Smith, RI. (2003). The enigma of Colobanthus quitensis and Deschampsia antartica. In Huiskes, A . H. L., Gieskes, W. W. C., Rozema, J., Schoro, R. M. L., van der Vies, S. M., and Wolff, W. J. (eds.), Antarctic Biology in a Global Context. Leiden: Backhuys Publishers 2: 34-239. Moore, DM. 1970. Studies in Colobanthus quitensis (Kunth) Bartl. and Deschampsia antarctica Desv. II. Taxonomy, distribution and relationships. Br Br Antarct Surv Bull 23: 63-80. Murashige, T., Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 15: 473-497. Orozco, A., Franco, N., Taborda, L. 2010. Evaluation of scarification methods for seeds of algarrobo (Hymenaea Courbaril L.). Rev. Invest. Univ. Quindío 20: 36 - 41. Pietrosemoli, S., Mendiri, J. 1997. Respuesta a la escarificación de semillas de Clitorea ternatea L. Archivos Latinoamericanos de Producción Animal. 5: 28-29. Pisano, E. 1976. Cormófitos acuáticos de Magallanes. Ans. Ins. Pat. 7: 115-136. Pisano, E., Schlatter, R. 1981. Vegetación y flora de las islas Diego Ramírez. I. Características y relaciones de la flora vascular. Ans. Ins. Pat. 12 : 183-194 Pisano, E. 1988. Sectorización fitogeográfica del archipiélago sudpatagónico-fueguino. II. Vegetación y flora vascular del área del Parque Nacional “Laguna San Rafael”, Aysen, Chile. Ans. Ins. Pat. 18: 5-34. Ruhland, CT., Day, TA. 2001. Size and longevity of seed banks in Antarctica and the influence of ultraviolet-B radiation on survivorship, growth and pigment concentrations of Colobanthus quitensis seedlings. Environmental and Experimental Botany 45:143-154. Zúñiga, GE., Zamora, P., Ortega, P., Obrecht, A. 2009. Micropropagation of Anctarctic Colobanthus quitensis. Antarctic Science 21:149-150.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
440
CARACTERIZACIÓN MORFOLÓGICA Y FENOLOGÍA DE Juncus bufonius L CULTIVADO EN CONDICIONES CONTROLADAS.
(Morphological characterization and phenology of Juncus bufonius L. growing in controlled conditions)
Fuentes. E1,2, Cuba-Díaz M1,2
1Laboratorio de Biotecnología y Estudios Ambientales, 2Departamento de Ciencias y Tecnología Vegetal, Campus Los Ángeles, Universidad de Concepción, Juan Antonio Coloma 0201Casilla 341, Los Ángeles, Chile, e-mail: [email protected]
INTRODUCCIÓN Juncus bufonius L., conocida comúnmente como Junco o Junquillo de sapo, es una planta angiosperma, monocotiledónea, perteneciente a la familia de las Juncaceae, es reconocida por el departamento de agricultura de los E.E. U.U como maleza o planta invasora (PLANTS.USDA.GOV). El género Juncus junto a Luzula, representan los únicos de esta familia que poseen un ciclo de vida anual (Hutchinson 1926; Heywood et al. 1978). La familia Juncacea es considerada como parecida a los pastos pero se diferencian de estos porque las flores poseen tépalos obvios y tienen los frutos en forma de cápsula. Las juncaceas han colonizado todos los ambientes en especial los de las zonas templadas, y se polinizan por el viento (Simpson 2005). En Europa occidental han sido reconocidos cinco especies pertenecientes a un mismo complejo de J. bufonius, donde J. bufonius L. es la única poliploide (Cope & Stace 1978, 1983, 1985). Diferentes estudios fundamentan que dicha especie proviene de la hibridación de las especies diploides pertenecientes al complejo (Cope & Stace 1985), lo que podría hacerla acreedora de una mayor variabilidad genética y fisiológica que le permita una mayor adaptabilidad a condiciones abióticas variables. J. bufonius L. es una planta cespitosa, con follaje fino, tallos erectos o ascendentes, brotes que derivan en forma horizontal, con hojas caulinares y ramificadas (Linneo 1753) que salen desde la base y son largas con un ancho promedio de 1mm. Inflorescencias que puede ocupar el 50% o más de la parte aérea de la planta (Botanicals notes 2000). Las flores son actinomorfas, bisexuales, y la semilla posee una forma ovoide a elipsoidal, sin dificultades para la germinación la cual ocurre generalmente en lugares húmedos de preferencia anegados y con alta intensidad lumínica ya que generalmente no toleran la sombra y ni la competencia con su misma u otras especies (Cope & Stace 1978). Según SMMFLOWERS (2013), son plantas pequeñas en relación a otras perennes, logrando crecer entre 5-20 cm de altura. El color de la planta es típicamente verde, tomando eventualmente tonos rojizos en las bases que pueden extenderse a toda la estructura de esta. La distribución de esta especie es casi cosmopolita (Brooks & Clements 2000), habitando principalmente zonas arenosas, litorales, con climas fríos o de regiones montañosas, con ambientes temporalmente húmedos o mojados (Heywood et al. 1978); asimismo es capaz de vivir en condiciones ocasionalmente salinas. Se puede hallar formando agrupaciones de plantas de la misma especie, o con otras especies. Según Afonin et al. (2008) la distribución de J. bufonius alcanza lugares del planeta como Escandinavia, Europa Occidental, Mediterráneo, Asia Menor, América del Norte, África del Norte, América del Sur, Australia, la parte europea de la antigua URSS, excepto la zona del Ártico, el Cáucaso, Siberia occidental, el este de Siberia, el Lejano Oriente, Asia Central. Recientemente, se ha reportado la existencia de J. bufonius L. var. Bufonius en la Antártica, en las cercanías de la Estación Polaca de Arctowski, encontrándose asociada a plantas vasculares nativas del lugar; D. antártica y C. quitensis (Cuba-Díaz et al. 2013). El objetivo de este trabajo es estudiar las características morfológicas y la fenología de plantas Juncus bufonius L. cultivadas condiciones controladas, a partir de plantas y semillas procedentes del territorio Antártico, y establecer comparaciones con datos ya publicados de Juncus bufonius L que difieren en distribución.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
441
MATERIALES Y MÉTODOS Material vegetal: Se utilizaron plantas y semillas de Juncus bufonius L. provenientes de las cercanías de la Base polaca H. Arctowski, Is. Rey Jorge (Cuba-Díaz et al. 2013), que se encontraban disponibles en el Laboratorio de Biotecnología y Estudios Ambientales de la Universidad de Concepción, Campus Los Ángeles. Las plantas y semillas fueron cultivadas en un sustrato 3:2:1 de tierra: turba: perlita, en cámara de crecimiento de plantas con fotoperiodo de 16/8 horas luz/oscuridad, temperatura de 15±2°C, con una densidad de flujo de fotones fotosintéticos (PPDF) 75±20 µmol fotones m-2 s-1 , y un porcentaje de humedad de 80±5 %. Mediciones morfológicas: Las mediciones morfológicas se realizaron con ayuda de un Vernier o pie de metro digital (MITUTOYO, USA) en 10 plántulas de 20 semanas de edad y se midió: largo total de la planta, largo de la parte aérea, largo de hoja, número de hojas por planta, ancho de la hoja, largo de la raíz, ancho del cuello, número de inflorescencia por plantas. Fenología de Juncus bufonius L.: Se colocaron 200 semillas en el sustrato anteriormente descrito y se evaluaron parámetros como porcentaje de germinación, tiempo de germinación, tiempo entre antesis y formación de semillas. Análisis estadísticos: Los valores promedio de las variables morfológicas comparativas, se realizaron con análisis de varianza de una vía (ANOVA), con un nivel de significancia de p<0,05, utilizando el programa Statistics para Windows, versión 6.0. Los gráficos se construyeron mediante el programa para Windows Origin versión 8.0.
RESULTADOS Y DISCUSIÓN Análisis morfológicos en Juncus bufonius L. En la Tabla 1 se muestra las características morfológicas para J. bufonius procedentes de Antártica. En general los parámetros medidos, muestran que las plantas de J. bufonius cultivadas en las condiciones de crecimiento controladas, establecidas en el laboratorio para las plantas antárticas, son más pequeñas que las descripciones para la especie en condiciones naturales.
Tabla 1: Características morfológicas de Juncus bufonius L. antártico cultivado en condiciones controladas.
Table 1: Morphological characteristic of antarctic Juncus bufonius L. growth in controlled conditions.
Las descripciones de la morfología para Juncus bufonius L, establecen varias medidas, así por ejemplo, SMMFLOWERS (2013) menciona que las plantas pueden alcanzar un largo de entre 5-20 cm, mientras que Wilson & Johnson (2001) mencionan que las plantas pueden medir entre 2-50 cm, las plantas antárticas crecidas en condiciones controladas alcanzan en promedio 10 cm, lo cual es medio respecto a lo informado en la literatura. De la misma forma, otros parámetros medidos tales como el largo y anchos de hojas fueron menores a los informados en la literatura de 15 cm y entre 0,5-5mm, respectivamente (Wilson & Johnson 2001). Ahmadpour & Hossinzadeh (2002) han establecido que las características morfológicas de la especie pueden variar de acuerdo a la distribución y características edafoclimáticas donde estas se desarrollen. De acuerdo a esta observación, se tomaron datos de la morfología de la especie procedente de diferentes regiones geográficas. En la fig. 1, se comparan la altura de plantas, el ancho de hojas y el número de inflorescencia por plantas (únicos datos en común

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
442
encontrados) de las plantas en este estudio (A), plantas creciendo en condiciones naturales en el Este de Europa (Cope & Stoce 1978) (B) y plantas en condiciones naturales en Irán, Asia (Snogerup 1971) (C). Estos análisis comparativos corroboraron que las plantas provenientes de la Antártica y crecidas en las condiciones de laboratorio, son significativamente más pequeñas que plantas creciendo en condiciones naturales, lo que podría ser un efecto de la temperatura, aunque se ha descrito como una planta que es capaz de vivir en condiciones de climas templados, se ha informado que tanto las temperaturas bajas como altas pueden inhibir su crecimiento (Cope & Stace 1978). Excepto para al número de inflorescencia, donde se pudo observar que no existe diferencias significativas con las plantas creciendo en Europa del Este (Fig. 1 Der.).
Figura 1. Características morfológicas comparadas de Juncus bufonius L. antártico (A) en condiciones de crecimiento
controlado, de Europa del Este (B) e Irán (C), ambas en condiciones naturales. Se muestra largo de planta (Izq.), Ancho de hoja (Centro) y No. de inflorescencia (Der.). Letras diferentes corresponden a diferencias de p <0,05.
Figure 1. Comparative morphological characteristics of J. bufonius L. from: Antarctic (A), en controlled growing conditions, Eastern Europe (B) and Iran (C), both in natural conditions. Is shown: plant length (left), leaves width (middle)
and inflorescence number (right). Different letters given for differences in p<0,05. Fenología de Juncus bufonius: En las condiciones del estudio, el porcentaje de germinación obtenido fue bajo (22,5%), el tiempo de germinación fue de 3 semanas, considerándose hasta la aparición de 4-5 brotes o hojas, y el tiempo transcurrido entre la antesis y la formación de semillas fue de aproximadamente 7 semanas, considerándose desde la aparición de los primeros botones florales (fig. 2).
Figura 2. Diferentes estadios del desarrollo fenológico de J. bufonius L. antártico creciendo en condiciones controladas. A:
3 semanas, B-F: 1, 2, 4, 6, 7 meses, respectivamente. Figure 2. Different stages of phenological development of Antarctic J. bufonius L. growing in controlled conditions. A: 3
weeks, B-F: 1, 2, 4, 6, 7 months, respectively.
Estos resultados son similares a los obtenidos para la familia Juncaceae donde se estableció que el tiempo entre antesis y formación de semillas es de aproximadamente 5 a 6 semanas desde la aparición de los primeros botones florales (Michalski & Durka 2007). Se ha informado que en condiciones naturales, el periodo de floración ocurren de marzo a septiembre (Kirschner 2002). El bajo porcentaje de germinación obtenido pudo estar relacionado con disponibilidad hídrica, debido a que por el sistema de la cámara de crecimiento, las temperaturas bajas se logran por inyección de aire frío lo que hace que el sustrato se seque. Aunque el régimen de riego mantenido hace que la HR no sea inferior a 75%, se ha descrito en la literatura que J. bufonius L es poco tolerante a ambientes secos, y una vez iniciado un estrés por sequía para estas plantas la recuperación es muy baja (Cope & Stace 1978). Estos resultados

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
443
muestran por primera vez el comportamiento de J. bufonius L. creciendo en condiciones controladas (bajas temperaturas, fotoperiodo medio y HR media), además en un sustrato más compacto que los sitios arenosos donde ha sido descrito antes (Brooks & Clements, 2000), por lo que más y nuevos estudios son necesarios para conocer el posible comportamiento de esta especie recién arribada al territorio insular antártico.
CONCLUSIONES Estos resultados preliminares muestran una clara inhibición de crecimiento de J. bufonius L al crecer en condiciones controladas, en comparación a datos morfológicos que corresponden a la especie creciendo en condiciones naturales. La fenología de J. bufonius L. en las condiciones de estudio no difiere mayormente con resultados obtenidos para otras juncáceas.
AGRADECIMIENTOS Investigación financiada por proyecto INACH T_03-09.
REFERENCIAS BIBLIOGRÁFICAS Afonin, AN., Greene, SL., Dzyubenko. NI. & Frolov, AN. (eds.). 2008. Interactive Agricultural Ecological Atlas of Russia and Neighboring Countries. Economic Plants and their Diseases, Pests and Weeds[Online]. Available at: http://www.agroatlas.ru Ahmadpour, R. y Hossinzadeh, SR. 2012. Seed coat morphology of the genus Juncus L. (Juncaceae) and its systematic significance in Northeast of Iran. Journal of Biology.1:29-38. Botanicals notes.(2000) Available online athttp://www.woodlotalt.com/publications/publications.htm Number 3, 22 September 2000. Brooks, RE. & Clements, SE. 2000. Juncaceae in Flora of North America Editorial Committee. Flora of North America, Vol. 22. Oxford University Press, NY. Cope, TA. & Stace, CA. 1978 The Juncus bufonius L. aggregate in western Europe. Watsonia 12: 113–128. Cope, TA. & Stace, CA. 1983. Variation in the Juncus bufonius L.aggregate in western Europe. Watsonia 14: 263–272. Cope, TA. & Stace, CA. 1985. Cytology and hybridization in the Juncus bufonius L. aggregate in western Europe. Watsonia 15: 309–320. Cuba-Díaz, M., Troncoso JM., Cordero, C., Finot, VL. & Rondanelli-Reyes M. 2013. Juncus bufonius, a new non-native vascular plant in King George Island, South Shetland Islands. Antarctic Science 25:285-286. Hutchinson, J. 1926. The families of flowering plants, arranged cording to new systems basedon their probable phylogeny. Published by Macmillan & Co. Ltda. Oxforfd University. London. Heywood, VH., Brummintt, RK., Culhan, A. & Seberg, O. 1978 Flowering plant families of the world. Kirschner, J. 2002. Juncaceae 3: Juncus subg.Agathryon. En: Species Plantarum: Flora of the World (Orchard, AE. & Wilson JG., eds. Part 8: 1:192. Canberra, Australia. Linneo C. 1753. Species Plantarum 1: 328. Michalski, SG. & Durka, W. 2007. Synchronous Pulsed Flowering: Analysis of the Flowering Phenology in Juncus (Juncaceae). Annals of Botany 100:1271–1285. Simpson, MG. 200). Juncaceae. Plant Systematics. Elsevier Inc. pp. 206. SMMFLOWERS. 2013. National park service U.S department of interior, and Santa Monica Mountains National Recreation Area. http://www.smmflowers.org/bloom/bloom.htm Snogerup, S. 1971. Juncaceae, En: Rechinger, KH., ed. Flora Iranica, 75: 15-18. Wilson, KL. & Johnson, LAS. 2001. The genus Juncus (Juncaceae) in Malesia and allied septate-leaved species in adjoining Regions. Telopea 9:357-397.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
444
IDENTIFICACION Y ANALISIS DE EXPRESION DE G3PD, FtsH Y SOD, TRES GENES REGULADOS POR FRIO EN Deschampsia antarctica Desv.
(Identification and expression analysis of G3PD, FtsH and SOD, three genes regulated by low temperature in Deschampsia antarctica Desv.)
Velásquez, R1. Rabert, C1-2. Gutiérrez, A1-2. Sandoval, A1-2*.
1Laboratorio de Fisiología y Biología Molecular Vegetal, Facultad de Ciencias Agropecuarias y Forestales, [email protected] 2Center of Plant, Soil Interaction and Natural Resources Biotechnology, Scientific and Technological Bioresource Nucleus, Universidad de La Frontera, Casilla 54-D, Temuco, Chile. INTRODUCCIÓN Las bajas temperaturas son un factor limitante para la productividad y distribución geográfica de muchas especies vegetales (Allen and Ort 2001). Estas afectan el equilibrio osmótico, alteran la fluidez de las membranas, la conformación de las proteínas e impactan el metabolismo celular, reduciendo drásticamente la velocidad de reacciones bioquímicas y provocan una reprogramación de la expresión génica. Muchas plantas incrementan su tolerancia al frío en respuesta a estímulos medioambientales como las bajas temperaturas, fenómeno conocido como aclimatación al frío (Thomashow 1999). Este proceso está asociado a la expresión de novo de genes, síntesis de proteínas y varios cambios fisiológicos. Las plantas son capaces de aclimatarse rápidamente en respuesta a bajas temperaturas. Pero como la aclimatación al frío contrarresta el crecimiento, las plantas son capaces a su vez, de des-aclimatarse rápidamente una vez que las temperaturas comienzan a subir. Los mecanismos que contribuyen a la tolerancia incluyen la prevención de la desnaturalización de proteínas inducida por bajas temperaturas, la prevención de la precipitación de moléculas y la atenuación de las consecuencias de la formación de hielo intercelular (Guy 1985). Deschampsia antarctica Desv. (Poaceae), es una planta gramínea que ha despertado el interés de muchos especialistas, dada su capacidad para sobrevivir bajo las hostiles condiciones ambientales del territorio Antártico. Su singular respuesta se debería a la expresión coordinada de genes especializados, interesantes de ser identificados y caracterizados, para su uso en el mejoramiento de cultivos o la industria farmacéutica. Nuestro grupo realiza desde hace algunos años estudios a nivel de campo y laboratorio orientados a identificar genes responsables de la tolerancia de esta planta al ambiente Antártico. Para obtener la información contenida en el transcriptoma de Deschampsia nuestra metodología contempló el uso de Hibridación Sustractiva por Supresión (SSH) para identificar genes diferencialmente expresados durante condiciones de frio. Nosotros encontramos un total de 112 genes a partir de una biblioteca de cDNA de Deschampsia. Análisis de PCR en Tiempo Real confirmaron las diferencias de expresión. Mediante una búsqueda por similaridad identificamos tres transcritos que codifican para proteínas que presentan homología con G3PD, FtsH y una SOD. Actualmente estudiamos estos genes y también sus respectivas proteínas, siendo candidatos para regular la actividad en respuesta a situaciones de estrés provocados por las bajas temperaturas. Esto puede ser de relevante ayuda para comprender el o los mecanismos por los cuales Deschampsia sobrevive bajo el extremo ambiente Antártico. MATERIALES Y MÉTODOS Material vegetal: El material vegetal necesario para los análisis, fue obtenido de plantas de Deschampsia antarctica DESV. (Poaceae), colectadas durante el periodo estival en la bahía Glaciar Collins (62° 10’ 05,6’’ S; 58° 51’ 06,2’’ W) de la Isla Rey Jorge (islas Shetland del Sur, Antártica). Para ensayos de laboratorio, una fracción del material fue propagado vegetativamente in vitro. Las

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
445
condiciones de crecimiento fueron 13 ± 1.5°C en una cámara de crecimiento (Forma Scientific, Inc., Marieta, OH, USA), con fotoperíodo de 21/3horas luz / oscuridad y 100-120 µmol m-2s-1 de densidad de flujo de fotones fotosintéticos (PPFD). Tratamientos: Pasado el proceso de aclimatación, se aplicaron los tratamientos de frío en cámara de cultivo a una temperatura de 4°C y se tomaron muestras a distintos tiempos de aclimatación (0 y 72 hrs). Las plantas aclimatadas en el laboratorio fueron comparadas con las muestras crecidas a 13°C y con muestras tomadas en territorio antártico. Extracción de ARN y síntesis de cDNA: Para la extracción del RNAtotal se utilizó el reactivo Trizol (Invitrogen, USA), siguiendo las recomendaciones del fabricante. Posteriormente cada muestra es tratada con DNAsa I (Ambion Inc, USA), y purificado mediante clean up usando columnas RNeasy mini spin (Qiagen, Germany). La concentración y pureza del RNA extraído se determina en un espectrofotómetro UV NanoDrop ND-1000; la síntesis de cDNA fue realizada con el kit comercial AffinityScript qRT-PCR de acuerdo a las instrucciones del fabricante. PCR tiempo Real: Los partidores FtsH, G3PH y SOD fueron diseñados utilizando el software AmplifX 1.5 (http://ifrjr.nord.univ-mrs.fr/AmplifX-Home-page). El gen alfa-tubulina fue utilizado para normalizar los datos. Actualmente, se prueban tres genes normalizadores para incorporarlos al estudio. Para la PCR se utilizó el kit cDNABrilliant II SYBR Green (Stratagene, USA). El procesamiento de los datos de la PCR es realizado con el software MxPro (Stratagene, USA), siguiendo las instrucciones del fabricante, y posteriormente con el programa Gene Expression Analysis for iCycler iQ Real-Time PCR Detection System (Laboratories, BioRad, Hercules, CA). El análisis estadístico de los datos es determinado con el Software GraphPad Prism version 5.00 (GraphPad Software Inc., USA), utilizando análisis de tipo T-test de Students y los valores de p-value < 0.05 son considerados estadísticamente significativos. RESULTADOS Los ensayos para estudio de genes inducidos por condiciones de frío presentes en la condición Antártica establecidos con anterioridad indicaron la expresión diferencial de 5 genes los cuales dentro de la búsqueda de similitud arrojaron coincidencia con G3PD, Rubisco, FtsH, desconocido, SOD (Figura 1). Para el ensayo que compara las condiciones antárticas con condiciones de frío en laboratorio (4°C) se trabajó solo con los genes G3PD, FtsH y SOD; esto debido a que estos genes y sus proteínas respectivas presentan un mayor atractivo en la industria.
Figura 1. Validación de genes sobre-expresados a través de PCR en tiempo real. Cantidad relativa de mRNA para Gen
normalizador a-tubulina; Gen Da5F11 (G3PD); Gen Da9G8 (RubisCO); Da3D11 (UnKnown) y DaSOD (SOD). Figure 1. Gene validation expresed through real time PCR. Relative quantity of mRNA normalized by a.tubuline, Gen
Da5F11 (G3PD); Gen Da9G8 (RubisCO); Da3D11 (UnKnown) y DaSOD (SOD). Se realizó una nueva validación de genes seleccionados a través de PCR en tiempo real esta vez bajo condiciones de frío en laboratorio (4°C) contrastado con la condición de crecimiento óptima a 13°C comparando los niveles basales de expresión a tiempo 0 y luego de 72 horas de estar sometidos a la

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
446
condición de frío. Para todos los genes analizados que en primera instancia presentaron una expresión diferencial entre las condición Antártica versus laboratorio (13°C) se ratificó que la expresión de los transcrito fue mayor en la condición Antártica, siendo los niveles basales para todos los genes analizados muy similares entre las condiciones 13 y 4°C. DISCUSIÓN Si bien todos los organismos vivos soportan numerosos factores endógenos y exógenos de estrés, al mismo tiempo poseen numerosos sistemas de defensas. De acuerdo con los resultados obtenidos en el presente trabajo podemos concluir que mediante la metodología de SSH se logró identificar genes regulados por frío, los cuales participan activamente como mecanismos de defensa y tolerancia en Deschampsia antarctica. La expresión diferencial de un grupo de genes fue corroborada mediante PCR Tiempo Real. Tras la secuenciación, alineamientos en BLASTN y BLASTX, encontraron similitud principalmente con otras especies gramíneas. Uno de los problemas con los que nos encontramos durante la interpretación fue la existencia de un alto porcentaje de secuencias similares a proteínas hipotéticas, de las cuales podemos conocer su secuencia de aminoácidos pero nada directamente sobre su bioquímica, su estructura o su función. Podremos deducir indirectamente algunos de estos rasgos mediante la búsqueda de dominios conservados de otras proteínas conocidas en otros organismos diferentes, con las que la secuencia de la proteína en cuestión presente similitud. La respuesta de Deschampsia frente al estrés por frío fue la activación transcripcional de genes específicos. La regulación espacial y temporal de patrones de expresión de genes de estrés específicos es una parte importante de la adaptación de la planta al estrés abiótico. La expresión de G3PD juega un rol importante en la vía glucolítica y se ha reportado su incremento bajo condiciones de estrés en Zea mays (Russell and Sachs 1989) y Arabidopsis thaliana (Yang et al., 1993) ensayos que incluyeron exposición a radiación UV y aclimatación a frío. Esto coincide con lo obtenido experimentalmente en nuestro trabajo; en donde los niveles de expresión de la enzima G3PD fueron incrementados en condiciones de crecimiento de frió de laboratorio, observando un nivel basal de expresión en el tiempo 0 muy similar entre las condiciones evaluadas. La proteína FstH es conocida por su rol como componente del sistema de reparación del complejo del fotosistema II (FSII) que opera para que la planta sea capaz de sobreponerse a fenómenos de fotoinhibición (Lindahl et al., 2000). De acuerdo a los resultados de expresión diferencial obtenidos en nuestro ensayo podemos detectar que en condiciones de frío de 4°C se aprecia una mayor abundancia de transcrito y por lo tanto esta proteína también se establecería en un gen con respuesta positiva a la aclimatación por frío. Las SODs juegan un papel primordial en la detoxificación de productos derivados del estrés. Se sabe que SOD contribuye a la protección frente al estrés oxidativo, tanto mediante la inactivación directa de peróxidos (de los lípidos o del DNA) como mediante la detoxificación de xenobióticos, los cuales son conocidos cofactores para la formación de radicales libres (Dixon et al. 2002). Entre los genes sobre-expresados en la muestra Antártica, encontramos Superóxido Dismutasa (SOD), la cual dentro de su rol de protección contra la producción de radicales libres tendría un mayor expresión lo cual fue corroborado con el ensayo de frío en laboratorio donde también se encontró una mayor cantidad de transcrito que en su condición de crecimiento óptimo de laboratorio13°C. Para todos los genes analizados se visualizó que su nivel de transcrito fue mayor en la condición Antártica versus frio de laboratorio (4°C); esto viene a confirmar que D. antarctica se encuentra expuesto a extremas condiciones de crecimiento en la península Antártica por lo cual su nivel de transcritos es mayor. AGRADECIMIENTOS Proyectos FONDEF D03I-1079, INACH 01-03, CONICYT CTE DIUFRO DI13-0048.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
447
REFERENCIAS BIBLIOGRÁFICAS Allen DJ, Ort DR (2001) Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci 6:36–42. Dixon, D.P., Lapthorn, A. & Edwards, R. 2002. Plant glutathione transferases. Genome Biology, 3, REVIEWS3004. Guy CL, Niemi KJ, Brambl R (1985) Altered gene expression duringcold acclimation of spinach. Proc Natl Acad Sci USA82:3673–3677. Lindahl M, Spetea C, Hundal T, Oppenheim AB, Adam Z, Andersson B (2000) The thylakoid FtsH protease plays a role in the lightinduced turnover of the photosystem II D1 protein. Plant cell 12:419–431 Russell DA, Sachs MM (1989) Differential expression and sequence analysis of the maize glyceraldehyde-3-phosphate dehydrogenase gene family. Plant cell 1:793–803. Thomashow MF (1999) Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annu Rev Plant Physiol Plant Mol Biol 50:571–599. Yang Y, Kwon HB, Peng HP, Shih MC (1993) Stress responses and metabolic regulation of glyceraldehyde-3-phosphate dehydrogenase genes in Arabidopsis. Plant Physiol 101:209–216

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
448
VARIACIÓN DEL ALBEDO ESPECTRAL DE LA NIEVE EN PUNTA FORT WILLIAMS, ISLA GREENWICH, ANTÁRTIDA MARINA
(Variation of the snow spectral albedo in Fort William Point, Greenwich Island, marine Antartic)
Alfonso, J.A.1, Carrera, J.M. 1, Espinoza, J.2, Silva, S1. 1Centro de Oceanología y Estudios Antárticos, Instituto Venezolano de Investigaciones Científicas, Apartado 20632, Caracas 1020A, Venezuela 2Servicio de Hidrografía y Navegación, Armada Bolivariana de Venezuela, Caracas, Venezuela. La nieve superficial es un importante componente del sistema climático (Flanner y otros, 2011). El albedo (fracción de luz solar reflejada) es una de las variables mas criticas que afectan el clima de la tierra y la característica fundamental de las áreas cubiertas de nieve es su alto albedo. Así, una reducción en la cobertura de nieve provoca un incremento en la absorción de la energía que calienta el planeta. En este contexto, son de crucial importancia los esfuerzos que hoy se realizan para ayudar a comprender el balance energético de las superficies cubiertas de nieve (Carmagnola y otros, 2012). El albedo de la nieve depende fundamentalmente de la densidad y tamaño de grano de la nieve, presencia de impurezas (Flanner y otros, 2012) y de las distribuciones espectrales y angulares de la radiación solar incidente (Warren y Wiscombe, 1980). El varia fuertemente a través del espectro solar (Warren, 1982), de acuerdo fundamentalmente a las variaciones espectrales del índice refractivo del hielo. En la región visible (λ= 0.35-0.75 µm), la absorción del hielo es débil y la radiación puede ser retrodispersada hacia la atmosfera o puede penetrar dentro de la nieve varias decenas de centímetros. Aquí, debido precisamente a esta débil absorción del hielo, aun pequeñas cantidades de impurezas absorbentes, tales como hollín, polvo y ceniza volcánica, pueden reducir significativamente el albedo de la nieve (Doherty y otros, 2010). Ha sido probado que el hollín (Black carbon) es el absorbente más eficiente en nieve (Flanner y otros, 2012), así tanto que el efecto de 1 ppm de hollín es tan grande como el efecto de 100 ppm de polvo (Warren y Wiscombe, 1980). Este trabajo reporta resultados preliminares de un proyecto de investigación del COEA-IVIC, financiado por el Fondo Nacional de Ciencia, Tecnología e Innovación (FONACIT) de Venezuela, que pretende estudiar la variación espacial y temporal del albedo espectral de nieve antártica y su interrelación con la distribución de impurezas superficiales. Los resultados aquí reportados están limitados al estudio de la variación del albedo espectral de la nieve en la parte visible del espectro. Las mediciones de reflectancia espectral de la radiación solar fueron realizadas sobre 7 sitios seleccionados cerca y sobre el glacial Quito, ver Fig. 1. Los sitios S1, S2, S3 y S4 fueron medidos en febrero del 2010 y los sitios 5, 6 y 7 en febrero del 2012, en el marco de la III y la V expediciones científicas venezolanas al continente antártico. Los espectros de reflectancia se midieron en función del Factor de Reflectancia Hemisférica Direccional (HDRF) (G. Schaepman-Strub y otros, 2006) en dirección nadir, utilizando un Spectralon de 30 cm x 30 cm, como superficie de referencia y un espectroradiómetro portátil modelo PR-650 (380-780 nm) manufacturado por Photho Researh Inc., posicionado sobre un trípode a una altura aproximada de 1 metro sobre la superficie de la nieve. Las mediciones fueron realizadas utilizando un campo de visión (FOV) de 1º, entre las 10 am y las 3 pm con poca o nula cobertura de nubes. En las Figuras 2 y 3 se presentan los albedos espectrales medidos en los sitios de estudiados. El albedo espectral de S1, una superficie de nieve visualmente blanca cercana a la costa, es parecido al albedo reportado para nieve pura, ver Fig. 4 (Warren y Wiscombe, 1980). El pequeño pico de reflectancia cercano a 760 nm, que aparece generalmente en todos los sitios estudiados, no pudo ser identificado. El sitio S2, en el momento de la medición se encontraba bastante cubierto de hollín proveniente de un incinerador muy cercano, y su espectro parece reflejar fundamentalmente la absorción del hollín: una significativa reducción del albedo en toda la región visible del espectro (Carmagnola y otros 2012). El sitio S4 fue

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
449
seleccionado muy cerca de un nido de petreles y su nieve superficial estaba cubierta de una coloración rojiza y el albedo medido en el coincide con la respuesta espectral de altas cantidades de impurezas de materia orgánicas (Takeuchi, 2009).
Figura 1. Sitios de estudio.
Figure 1: Study sites.
Figura 2: Reflectancia espectral medida en los sitios S1, S2, S3 y S4. Figure 2: Measured spectral reflectance in the sites S1, S2, S3 and S4.
El albedo medido en S3, parece indicarnos una combinación de efectos de impurezas de hollín y materia orgánica, esto era de esperar debido a su ubicación entre los sitios S2 y S4. Los sitios S5, S6 y S7 fueron seleccionados sobre el glacial Quito, a alturas media sobre el nivel del mar de 35, 80 y 106 m, respectivamente. En la Fig. 3 se observa, que el albedo de la nieve del glacial Quito, en toda la región visible del espectro, aumenta con la altitud y disminuye al acercarnos a la Estación Científica Ecuatoriana, en los sitios estudiados. Warren y Wiscombe (1980), estudiaron el efecto de la concentración de hollín sobre el albedo, ver Fig. 4., y reportaron grandes cambios en la intensidad y

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
450
ligeros cambios en la forma del espectro, al aumentar la concentración de hollín en la nieve entre 0.1 y 10 ppm.
Figura 3: Reflectancia espectral medida en los sitios S5, S6 y S7. Figure 3: Measured spectral reflectance in the sites S5, S6 and S7.
Figura 4: Efecto de la concentración de hollín sobre el albedo
Figure 4: Effect of soot concentration on albedo. (Warren y Wiscombe, 1980).
El tipo de nieve (fresca, vieja) y el área superficial especifica de nieve (SSA, esta variable caracteriza el tamaño ópticamente relevante de los granos de nieve), también afectan notablemente la intensidad del albedo en toda la región visible del espectro (Carmagnola y otros 2012). Nosotros sugerimos, que el contenido de impurezas inorgánicas, posiblemente hollín, es la variable que más afecta a la variación del albedo entre los sitios S5 y S6. El albedo en S7 parece ser consecuencia de una mezcla más compleja de factores. Nuestros resultados sugieren que la variación en la distribución de impurezas de hollín sobre la nieve superficial, es uno de los más importantes factores que influencian la variación espacial del albedo espectral de la nieve en Punta Fort Williams, Isla Greenwich, Antártida marina. Que se conozca, estas

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
451
son las primeras mediciones “in situ” que se toman del albedo espectral de la nieve en la Antártida marina. Considerando la importante influencia del albedo de la nieve sobre el cambio climático, se sugiere desarrollar un monitoreo espacial y temporal de la reflectancia espectral (espectro completo) en la Antártida a través de sensores remotos, para lo cual es imprescindible realizar previamente extensivas mediciones “in situ”, para una adecuada calibración e interpretación de los resultados. AGRADECIMIENTOS Al FONACIT de Venezuela y al INAE de Ecuador. REFERENCIAS Carmagnola, C.M., Domine, F., Dumont, M., Wright, P., Strellis, B., Bergin, M., Dibb, J., Picard, G., Morin, S. 2012. Snow spectral albedo at Summit, Greenland: comparison between in situ measurements and numerical simulations using measured physical and chemical properties of the snowpack. The Cryosphere Discuss., 6: 5119-5167. Doherty, S.J., Warren, S.G., Grenfell, T.C., Clarke, A.D. y Brandt, R.E. 2010. Light-absorbing impurities in Arctic snow. Atmos. Chem. Phys., 10: 11647-11680. Flanner, M., Shell, K., Barlage, M., Perovich, D. y Tschudi, M. 2011. Radiative forcing and albedo feedback from the Northern Hemisphere cryosphere between 1979 and 2008, Nat. Geosci. 4: 151-155. Flanner, M., Liu, X., Zhou, C., Penner, J. y Jiao, C. 2012. Enhanced solar energy absorption by internally-mixed black carbon in snow grains. Atmos. Chem. Phys., 12: 4699-4721. Schaepman-Strub, G., Schaepman, M.E., Painter, T.H., S. Dangel, S. y Martonchik, J.V. 2006. Reflectance quantities in optical remote sensing – definitions and case studies. Remote Sens. Environ., 103(1): 27-42. Takeuchi, N. 2009. Temporal and spatial variations in spectral reflectance and characteristics of surface dust on Gulkana Glacier, Alaska Range. Journal of Glaciology, 55: 701-709 Warren, S. y Wiscombe, W. 1980. A model for the spectral albedo of snow. II: Snow containing atmospheric aerosols. J. Atmos. Sci., 37: 2734-2745. Warren, S. 1982. Optical properties of snow. Rev. Geophys., 20: 67-89.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
452
THE POTENTIAL OF TERRASAR-X INTERFEROMETRY ON UNION GLACIER
(El potencial de la interferometría Terrasar X en glaciar Unión)
E. Johnson1, C. Cárdenas1, D. Floricioiu2
1Universidad de Magallanes, Dirección de Programas Antárticos y Sub-Antárticos (erling.johnson, carlos.cardenas)@umag.cl 2German Aerospace Center (DLR), Remote Sensing Technology Institute (IMF) [email protected]
ABSTRACT The Union Glacier (79°45′0″S, 82°30′0″W) has a great importance for our country because a new Chilean base station will be established in that area. This will increase the scientific research and logistics on the surrounding area generating the need to study the sector by SAR techniques. Therefore, in this work we will investigate the potential of SAR interferometry applications with TerraSAR-X satellite data in the area of the Union Glacier. The information will be compared with data already obtained of the sector, and also could be checked by future expeditions from a group of “Universidad de Magallanes” researchers. TerraSAR-X provides high resolution SAR data at X-band (9.65 GHz) with a repeat cycle of 11 days, which for areas in the interior of the Antarctic ice sheet is short enough to preserve the InSAR phase coherence. We analyze TerraSAR-X single look slant range (SSC) products over Union glacier in respect to glacier motion. Two techniques will be applied on these data and compared: InSAR and speckle tracking (Floricioiu et al, 2009). Also the TerraSAR-X velocities will be verified to previous velocities based on DGPS measurements of Rivera (2010), as well as with the recent published velocity map of Antarctica by Rignot (2011). The glacier velocity analysis improve the understanding of ice derives and the mechanics on flux transportation. Is possible to obtain the tributaries and main streams, proving that in Antarctica all the glacier masses are connected between them (Rignot, E., Mouginot, J., & Scheuchl, B., 2011). We expect a detailed view of the velocity pattern of the Union glacier, which gives an insight on the flow mechanisms of this glacier. A comparison with the older GPS data will also reveal possible changes in the surface velocity. This analysis would improve the existing data that our scientific community got from that area, presenting a complementary technique to the field measurements. Also the results can be used by the Chilean government as information for logistics and knowledge, to develop practical activities on the area surrounding this new Antarctic base station. ACKNOWLEDGEMENTS We want to thank the Deutschen Zentrums für Luft- und Raumfahrt (DLR), that provide the TSX data for this work. REFERENCES Floricioiu, Dana, et al. "TerraSAR-X interferometric observations of the recovery glacier system." FRINGE (2009). Rignot, E., J. Mouginot, and B. Scheuchl. "MEaSUREs InSAR-Based Antarctica Velocity Map." NASA EOSDIS DAAC at NSIDC: Boulder, CO, USA(2011). Rignot, E., Mouginot, J., & Scheuchl, B. (2011). Ice flow of the Antarctic ice sheet. Science, 333(6048), 1427-1430. Rivera, Andres, et al. "Glaciological investigations on Union Glacier, Ellsworth Mountains, West Antarctica." Annals of Glaciology 51.55 (2010): 91-96.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
453
ANÁLISIS DE MODELOS DIGITALES DE ELEVACIÓN PARA APLICACIÓN EN LA PENÍNSULA ANTÁRTICA.
(Analysis of digital elevation models for application in the Antarctic Peninsula) Petsch, C.1, Simões, J. C.2
1, 2 Universidade do Rio Grande do Sul - UFRGS/Centro Polar Climático (CPC), Av. Bento Gonçalves 9500 – 91501-970 – Porto Alegre - RS, Brasil. email: [email protected]; [email protected]
INTRODUÇÃO Modelos digitais de elevação (MDE), apesar de apresentarem vantagens como a obtenção de dados topográficos de forma remota, exibem elevações com erros significativos, os quais dependem da limitação inerente dos instrumentos de observação, da rugosidade e declividade da superfície; também carecem de pontos de controle localizados precisamente, para minimizar o erro sistemático, o que nem sempre é fácil de ser obtido, principalmente em regiões remotas (DIXON, 1995). É importante considerar que erros são inevitáveis, pois os MDEs são representações simplificadas da realidade, submetidas a um processo de generalização (FELICÍSIMO, 1994). Portanto, é necessário realizar uma avaliação prévia sobre os MDEs disponíveis para cada área de estudo e optar por aquele que mais se aproxima da realidade. Diante do exposto, é objetivo desse trabalho testar a aplicabilidade de 3 modelos digitais de elevação: (1) o do Radarsat Antarctic Mapping Project Digital Elevation Model (RAMP DEM) versão 2 (resolução espacial de 200 m), (2) do ASTER Global Digital Elevation Model (GDEM) versão 1 e (3) de sua versão 2 (resolução espacial de 30 m), pelos dados de altitude máxima, altitude mínima, orientação predominante das geleiras e declividade. MATERIAIS E MÉTODOS Foram selecionadas 9 bacias de drenagem glacial da ilha Joinville (localizada entre as coordenadas 63°00’ – 63°30’S e 56°45’ – 55°00’W, Figura 01), que foram vetorizadas com base nas curvas de nível da carta topográfica da área e na banda 8 da imagem de satélite Landsat 7 (29/01/2000). O DEM do RAMP DEM é disponível no site do National Snow and Ice Data Center (NSIDC) (http://nsidc.org/data/nsidc-0082.html); e as duas versões do ASTER GDEM podem ser obtidos no site do National Aeronautics and Space Administration (NASA) e Ministério da Economia, Comércio e Indústria do Japão (METI) (http://asterweb.jpl.nasa.gov/gdem.asp). O banco de dados geográficos para elaboração desse estudo foi convertido para a projeção Universal Transversa de Mercator (Zona 21S) e Datum WGS 84.
Figura 01: Cuencas seleccionados para el estudio.
Figure 01: Glacial drainage basins selected for this study.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
454
Os MDEs utilizados nesta investigação apresentam algumas discrepâncias nos valores encontrados, no entanto não foi aplicado nenhum filtro para suavização desses dados pois de acordo com Andrada et al. (2013) mesmo a aplicação dupla do filtro mediana 9 x 9, não altera grandemente esses valores espúrios. Os dados analisados para comparação entre os MDEs foram: altitude máxima, altitude mínima, orientação predominante e a declividade da geleira, todos obtidos no Sistema de Informação Geográfica ENVI 4.3. Para orientação predominante da bacia glacial não foi considerada a média, mas a classe que ocupou maior área. Para a declividade foi utilizada a mesma paleta de cores e intervalos de classes para permitir comparação entre os mapas obtidos.
RESULTADOS E DISCUSSÕES O MDE do ASTER GDEM 2 foi o que mostrou as maiores altitudes, sendo que sua diferença em relação ao RAMP DEM chegou a ser de 238, 153, 322 e 202 metros para as geleiras 3, 5, 6 e 9, respectivamente. Esses valores acentuados de altitude se configuram em picos na imagem, resultam de mais ruídos no ASTER GDEM 2 em relação aos outros MDEs. No caso das altitudes mínimas, o ASTER GDEM 2 também apresentou ruídos, com valores negativos (que foram considerados como 0), exibindo a maior variação em relação ao RAMP DEM nas geleiras 6 e 9 com diferença de 23 e 22 metros, respectivamente. As altitude máximas do RADARSAT e ASTER GDEM 1 foram similares em 6 geleiras, enquanto que para as altitudes mínimas a discrepância foi acentuada. O RAMP DEM mostrou altitudes mínimas variando entre 21 e 26 metros enquanto que o ASTER GDEM 1 variou entre 0 e 8 metros (Figura 02).
Figura 2: Valores de máxima y mínima altitud según los modelos digitales de elevación examinados.
Figure 2: Values of maximum and minimum altitude according to the digital elevation models examined.
Os dados de declividade também mostram diferenças significativas entre as bacias glaciais analisadas. De maneira geral, os mapas obtidos com o RAMP DEM e ASTER GDEM 1 possuem resultados similares, ocorrendo declividades mais altas (acima de 20%) nos mapas do RAMP DEM. Nas bacias glaciais 8 e 9 essa situação se inverte, enquanto que no RAMP DEM algumas faixas foram classificadas como de declividade entre 0-3% e 3-6%, no ASTER GDEM 1 estas áreas são de declividade entre 10-20% e 20-30%. Todavia, os dois MDEs apresentam a faixa de declividade predominante como sendo a de 3-6%. O MDE do ASTER GDEM 2 mostra resultados diferenciados, em todas as bacias glaciais a faixa predominante de declividade foi a de 10-20%, havendo poucas áreas na faixa de 3-6%, que era a predominante segundo os outros MDEs. Outro problema observado é a presença de declividade acima de 30% em áreas que são consideradas planas (0-3%) segundo o ASTER GDEM 1 e RAMP DEM (Figura 03).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
455
Figura 3: Declividad de las cuencas estudiadas.
Figure 3: Slope of the glacial drainage basins studied.
A orientação predominante das bacias glaciais apontou grande variação, não sendo possível obter esse parâmetro para o MDE do ASTER GDEM 2 devido à confusão de direção dos pixels. Comparando os dados do RAMP DEM e ASTER GDEM 1, somente as geleiras 1, 2 e 3 foram agrupadas com a mesma orientação (NO). Nas demais bacias glaciais houve diferenças discrepantes quanto à orientação: as bacias 4 e 5 foram denominadas de orientação Leste para o RAMP DEM enquanto que para o ASTER GDEM 1 foram classificadas como NO; a bacia 8 foi identificada como NO e S, e a bacia 9 é de orientação SE e NE para RAMP DEM e ASTER GDEM 1, respectivamente (Figura 04). Após o exame

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
456
visual, pode-se considerar como mais condizente com a realidade o resultado obtido com o RAMP DEM.
Figura 4: Orientación predominantes de las cuencas estudiadas.
Figure 4: Orientation predominant of the glacial drainage basins studied.
Considerações finais Avaliando os resultados obtidos com os dados de altitude, orientação predominante e declividade das bacias glaciais, recomenda-se o uso do RAMP DEM, apesar da sua escala espacial ser de menor detalhe em relação à do ASTER GDEM 1 e 2. O ASTER GDEM 1 apresentou informações semelhantes ao RAMP DEM nos critérios de altitude máxima, e declividade, mas quanto às altitudes mínimas exibiu valores negativos, provenientes de ruídos nas imagens, e na orientação predominante das vertentes foram observados resultados divergentes da realidade. O ASTER GDEM 2 demonstrou resultados insatisfatórios em todos os casos analisados, com valores de altitude máxima na ordem de 320 metros maior que o encontrado nos demais MDEs. Quanto à declividade, houve superestimação dos valores nos mapas obtidos a partir do ASTER GDEM 2, além da presença de áreas com declividade acima de 30% onde o ASTER GDEM 1 e RAMP DEM classificaram como plana (0-3%).
AGRADECIMENTOS C. Petsch agradece à CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) pela concessão de sua bolsa de Mestrado.
REFERÊNCIAS ANDRADA, R. R.; OLIVEIRA, J. A.; BICCA, C. E.; MENDES JUNIOR, C. W.; ARIGONY NETO, J. Análise de parâmetros morfométricos de geleiras derivados de diferentes modelos digitais de elevação da Península Antártica. In: XVI Simpósio Brasileiro de Sensoriamento Remoto - SBSR, 2013, Foz do Iguaçu, PR. Anais... Foz do Iguaçu: 2013, p. 8782 - 8789. DIXON, T. H. SAR Interferometry and surface change detection. Miami: University of Miami, Rosenstiel School of Marine and Atmospheric Sciences, RASMAS TR 95-003, 1995. 97p. FELICÍSIMO, A. M. Modelos digitales del terreno: Introducción aplicaciones en las ciencias ambientales. Oviedo: Universidad de Oviedo, 1994.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
457
PLUMAS DE SEDIMENTOS GLACIARES EN PEQUEÑAS BAHÍAS Y FIORDOS
DE LA COSTA DE DANCO, PENÍNSULA ANTÁRTICA (Glacial sediment plumes in small bays and fjords of the Danco Coast, Antarctic Peninsula)
Rodrigo, C.1, Varas, A.2
1Escuela de Ciencias de la Tierra, Universidad Andrés Bello, Quillota 910, Viña del Mar, Chile. E-mail: [email protected]
2Escuela de Ciencias del Mar, Pontificia Universidad Católica de Valparaíso, Av. Altamirano 1480, Valparaíso, Chile. E-mail: [email protected]
INTRODUCCIÓN La Antártica está sometida a varios cambios de su ambiente (e.g. Masson y Stammerjohn, 2010; Gutt et al., 2011; Rignot et al., 2004; Mayewski et al., 2009; Turner et al., 2005). Es importante conocer y evaluar dichos cambios debido a la conexión natural de la Antártica con el resto del mundo, en términos del clima, la circulación marina y el consumo de carbono. La península Antártica es una de las área más dinámicas y sensibles de la Antártica, por lo cual, se espera que las evidencias de los cambios sean más notorios en esa región. Algunos de estos cambios pueden ser registrados en las secuencias sedimentarias marinas. Los ambientes glaciomarinos y los procesos sedimentarios asociados, pueden entregar pistas para entender la evolución climática y oceanográfica de la región. Uno de los elementos de los procesos sedimentarios es la generación de plumas de sedimentos que provienen del glaciar, en la cual un aumento de la concentración de partículas podría indicar un mayor derretimiento de los glaciares. La costa de Danco representa una región de transición subpolar-polar, por lo cual, se espera que los cambios recientes sean más notorios en esta localidad. Griffith y Anderson (1989) sostuvieron que la costa de Danco se caracteriza por una escasez de plumas de sedimentos y posee aguas superficiales muy claras. Debido al cambio climático, nosotros esperamos un aumento de la presencia y concentración de las plumas de sedimentos, en especial para aquellos glaciares más pequeños, comparados a aquellos estudiados por Griffith y Anderson (1989). METODOLOGÍA En febrero de 2013 se realizó la Expedición Científica Antártica N°49 a bordo del buque de la Armada de Chile AP “Aquiles”. En dicha oportunidad se visitaron 4 bahías o fiordos relativamente pequeños y fueron las siguientes (carta de referencia SHOA N°15.300): bahía Guesalaga, bahía Frei, bahía Briand y una bahía pequeña más al sur de bahía Paraíso (Fig. 1). A bordo de un bote de goma, se aproximó lo más cerca posible del glaciar principal y se realizaron alrededor de 6 estaciones oceanográficas perpendiculares al glaciar y equidistantes, aproximadamente, en 1 Mn. Se realizó batimetría exploratoria con un ecosonda Garmin portátil de 50 KHz y mediciones con CTD SBE 19 Plus V2 con turbidímetro (T), fluorómetro, sensores de pH y oxígeno, hasta 100 m de profundidad. El procesamiento de los datos de CTD consistió simplemente en la remoción manual de los datos erróneos. Las secciones oceanográficas se prepararon utilizando el programa Ocean Data View (Schlitzer, 2010).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
458
Figura 1. Área de estudio. Los pines muestran las bahías estudiadas. Figure 1. Study area. The pins show the bays studied.
RESULTADOS Y DISCUSIONES Cada bahía presentó plumas de sedimentos provenientes del glaciar, pero con concentraciones bajas de partículas (< 1 NTU). Predominaron los flujos sub-superficiales y se asociaron a aguas más frías (< 0°C) y a salinidades intermedias (34 psu). La distribución de las plumas en la vertical se asemejaron a aquellas mostradas por Domack e Ishman (1993) en su modelo de los procesos depositacionales activos de la costa de Danco. Estas plumas también fueron asociadas a lenguas sub-superficiales de agua más fría. De lo anterior se deprende que, comparando con los sistemas de fiordos mayores del área, las localidades más pequeñas también pueden ser una fuente importante de sedimentos para todo el sistema de depositación de la región. Dado que, en genaral, las mediciones sólo se realizaron en superficie (a 1/3 de la profundidad total), no se sabe cómo son las características de las plumas a mayor profundidad, pero se espera que el flujo de sedimentos sea mayor cerca del fondo (Domack e Ishman, 1993). Las bahías que presentaron un sólo glaciar que desembocaba en la localidad, presentaron una lengua o pluma mejor definida, donde los valores de concentración de partículas fueron disminuyendo paulatinamente a medida de su alejamiento del glaciar. Estas bahías fueron las de Frei y Guesalaga. En cambio, en las bahías Beaupré y Paraíso, la distribución de plumas en la vertical fue algo más irregular debido a la presencia de otros glaciares en sus cercanías que se unieron al flujo principal de las bahías. Los valores de concentración de partículas de estos flujos se mantuvieron relativamente constantes a medida que se desplazaban más hacia afuera del glaciar. La figura 2 muestra un ejemplo de los resultados de las mediciones oceanográficas obtenidos en en la bahía Frei. El origen de la distancia (0 km) está en la estación oceanográfica más alejada del frente del glaciar. Se observa una pluma de sedimentos de baja concentración que proviene del glaciar y desaparece a casi 4 km de su frente (Fig. 2a). Esta pluma se asocia a una masa de agua más fría de alrededor de 0°C (Fig. 2b) y con una salinidad > 34.2 psu (Fig. 2c). Lo anterior es congruente con el modelo de Domack e Ishman (1993), y apoya el hecho de que los sedimentos terrígenos se ubicarán mayormente en las proximidades del glaciar, dado el corto alcance horizontal de la pluma.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
459
(a) (b) (c) Figura 2. Secciones de variables oceanográficas para la bahía Frei, costa de Danco. (a) turbidez, NTU; (b) temperatura, °C
(C) salinidad, psu. El origen de la distancia corresponde a la estación más alejada del frente del glaciar. Figure 2. Oceanographic variable sections for the Frei Bay, Danco Coast. (a) tubidity, NTU; (b) temperature, °C (b) salinity,
psu. The origin of the distance corresponds to the farthest station from the glacier front. CONCLUSIONES Se encontró que en cada bahía estudiada de la costa de Danco existieron una o más plumas de sedimentos provenientes del glaciar, aunque de bajas concentraciones de partículas. Para establecer si estas plumas son efímeras o no, es necesario realizar mediciones repetidas en el tiempo de las variables oceanográficas. Los resultados fueron consistentes con el modelo de Domack e Ishman (1993), y se espera además que el flujo de sedimentos sea mayor en el fondo. Por otro lado, los resultados concuerdan con las observaciones de Griffith y Anderson (1989) en lo que respecta a que hubo una predominancia de plumas subsuperficiales más que superficiales, encontrándose aguas claras en superficie. De todo lo anterior se desprende que la contribución de sedimentos de los sistemas de bahías y fiordos menores, pueden jugar un rol importante en los procesos sedimentarios regionales y, en especial, considerando su mayor vulnerabilidad al cambio climático. AGRADECIMIENTOS Al proyecto FONDECYT N°11121522, al Instituto Antártico Chileno y a la Armada de Chile. REFERENCIAS

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
460
Domack, E y S. Ishman. 1993. Oceanographic and physiographic controls on modern sedimentation within Antarctic fjords. Geological Society of America Bulletin, 105: 1175-1189. Griffith, T. y J. Anderson. 1988. Climatic control of sedimentation in bays and fjords of the Northern Antarctic Peninsula. Marine Geology, 85: 181-204. Gutt J., I. Barratt, E. Domack, C. d'Udekem d'Acoz, W. Dimmler, A. Grémare, O. Heilmayer, E. Isla, D. Janussen, E. Jorgensen, K.H. Kock, L.S. Lehnert, P. López-Gonzáles, S. Langner, K. Linse, M.E. Manjón-Cabeza, M. Meissner, A. Montiel, M. Raes, H. Robert, H. Robert, A. Rose, E.S. Sené Schepisi, T. Saucéde, M. Scheidat, H.W. Schenke, J.Seiler y C. Smith. 2011. Biodiversity change after climate-induced ice-shelf collapse in the Antarctic. Deep-Sea Research Part II: Topical Studies in Oceanography, 58(1-2): 74-83. Massom R.A. y S.E. Stammerjohn. 2010. Antarctic sea ice change and variability - Physical and ecological implications. Polar Science, 4(2): 149-186. Mayewski, P.A., M.P. Meredith, C.P. Summerhayes, J. Turner, A. Worby, P.J. Barrett, G. Casassa, N. A.N. Bertler, T. Bracegirdle, A.C. Naveira Garabato, D. Bromwich, H. Campbell, G.S. Hamilton, W.B. Lyons, K.A. Maasch, S. Aoki, C. Xiao y T. van Ommen. 2009. State of the Antarctic and Southern Ocean Climate System. Reviews of Geophysics, 47. Rignot, E., G. Casassa, P. Gogineni, W. Krabill, A. Rivera y R. Thomas. 2004. Accelerated ice discharge from the Antarctic Peninsula following the collapse of Larsen B ice shelf, Geophysical Research Letters, 31, L18401, doi:10.1029/2004GL020697. Schlitzer, R. 2010. Ocean Data View, http://odv.awi.de. Turner, J., S.R. Colwell, G.J. Marshall, T.A. Lachlan-Cope, A.M. Carleton, P.D. Jones, V. Lagun, P.A. Reid y S. Iagovkina. 2005. Antarctic climate change during the last 50 years. Int. J. Climatol., 25: 279–294. doi: 10.1002/joc.1130.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
461
QUÍMICA SANGUÍNEA DE DOS ESPECIES DE PINGÜINOS ANTÁRTICOS EN ÁREAS CON ALTO Y BAJO IMPACTO AMBIENTAL
(Plasma and serum biochemistry from two different species of Antarctic penguins in areas with low and high environmental impact)
Di Fonzo,C.I.1, Cebuhar J.D.1, Libertelli, M.1, Zappala, C.1y M. Ansaldo1,2
1Instituto Antártico Argentino, Dirección Nacional del Antártico. Cerrito 1248, (C1010AAZ) Ciudad Autónoma de Buenos Aires, Argentina. [email protected], [email protected] 2Laboratorio de Ecotoxicología. Universidad Argentina JF Kennedy. Sarmiento 4564, (C1197AAR) Ciudad Autónoma de Buenos Aires, Argentina. INTRODUCCIÓN Los predadores tope de la península Antártica, como focas y pingüinos, son indicadores muy sensibles a los cambios en el ecosistema. Sus vidas integran los efectos de la variabilidad del ambiente marino a través de largos períodos y vastas áreas dado que suelen ser longevos y su distribución es amplia (Mc Clintock, 2008). En bahía Esperanza habitan poblaciones de dos especies de pingüinos antárticos Pygoscelis adeliae (Adelia) y Pygoscelis papua (Papúa), que se encuentran distribuidas en parches cercanos y lejanos de la Base Antártica Esperanza. En esta zona Acero y col. (1991) verificaron sitios de alto impacto, donde existe un alto el ruido de helicópteros, residuos sólidos, derrames de combustibles, área de desembarco de turistas y circulación de vehículos y/o personas (Mazón, 2013); y otros de bajo impacto, o sea áreas alejadas de la actividad antrópica. El aumento de dichas actividades en Antártida ha dado lugar a interacciones muy frecuentes entre personas y fauna antártica, pudiéndose generar efectos adversos en la fisiología y comportamiento de los pingüinos (Culik y col., 1990; Giesse, 1998). Las acciones perturbadoras del ambiente aumentan durante los meses de verano (mayor actividad científica y logística), coincidiendo con el momento en el que las aves se encuentran en su etapa reproductiva (puesta de huevos y cría). Este hecho determina que en la práctica, las aves constituyen el elemento ambiental más afectado, siendo necesario por lo tanto de un análisis más detallado. Es importante comprender las respuestas fisiológicas de los organismos a las perturbaciones humanas, para así proporcionar datos sobre las causas subyacentes a los problemas de conservación, tales como la disminución del tamaño de las poblaciones (Carlini y col., 2009). Con el objetivo de determinar el estado fisiológico de las poblaciones de dos especies de pingüinos que habitan zonas de alto y bajo impacto ambiental de Bahía Esperanza, se tomaron muestras de sangre de pichones y adultos, sobre las que se analizaron distintos parámetros bioquímicos sanguíneos. MATERIALES Y MÉTODOS El presente trabajo fue realizado en los alrededores de la Base Antártica Esperanza situada en Bahía Esperanza, Península Antártica (63°23′54″S 56°59′46″O) durante el verano 2012-2013. El muestreo fue realizado en dos áreas diferenciadas según el impacto ambiental producido por las actividades de la base: una zona de alto impacto (ZAI) y otra zona de bajo impacto (ZBI) (Carlini y col., 2007). En dichas zonas se encuentran las áreas de nidificación de las dos especies de pingüinos antárticos estudiados, Adelia y Papúa. Se tomaron muestras de sangre en 15 pichones y 15 adultos de cada especie de cada zona. En el laboratorio de la Base Antártica Esperanzase centrifugaron las muestras (1500g x 10 min) para separar los eritrocitos de suero o plasma. Todas las muestras se conservaron en frío hasta su análisis en el laboratorio. Mediante técnicas espectrofotométricas se determinaron: las proteínas totales (Lowry y col., 1951), los niveles de ácido úrico, triglicéridos, calcio, fósforo inorgánico, albúmina

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
462
glicosilada y glucosa; y en suero, los niveles de hierro (kits Wiener Lab, ref.: 1840101, 1780111, 1152002, 1382321, 1400050, 1400106 y 1492001 respectivamente). En el análisis estadístico se empleó el modelo lineal general, teniendo en cuenta los factores: tipo de impacto y edad. El mejor modelo se obtuvo por comparación de los IC de Akaike y Schwartz (InfoStat, 2012). Se utilizó como post test para las comparaciones entre los grupos la prueba DGC (Di Rienzo y col., 2002). RESULTADOS
BAJO ALTO BAJO ALTO0
2
4
6
8a
bb
c
d)
Tipo de impacto
PR
OT
.TO
T.
PLA
SM
A(m
g/m
l)
Figura 1. Concentración de glucosa, albúmina glicosilada, fósforo inorgánico y proteínas totales vs tipo de impacto (BAJO y ALTO) en pichones y adultos de P. adeliae. Los valores están expresados como media ± error estándar. Diferentes letras indican diferencias significativas (p< 0,05). Figure 1. Glucose, glycosylated albumin, inorganic phosphorus and plasma protein concentration vs type of impact (LOW and HIGH) in P. adeliae chicks and adults. Values are expressed as mean ± standard error. Different letters indicate significant differences (p <0.05).
Los pichones de Papúa presentaron valores de triglicéridos, calcio y proteínas totales significativamente (p<0.005) más bajos que los adultos. A su vez los adultos presentaron niveles de albúmina glicosilada en plasma sanguíneo significativamente menores (p<0.001) que los pichones. En Adelia solo hubo diferencias en los niveles de ácido úrico relacionadas a la edad, siendo significativamente mayor en pichones que en adultos.
0
100
200
300
AdultoPichón
a
bbb
GLU
CO
SA
(m
g/dl
)
0
150
300
450
600
a
bb
b
ALB
UM
INA
GLI
CO
SIL
AD
A(u
mol
/l)
BAJO ALTO BAJO ALTO0
3
6
9
12
aa
b
c
Tipo de impacto
FÓ
SF
OR
O I
NO
RG
ÁN
ICO
(mg/
dl)
a)
b)
c)
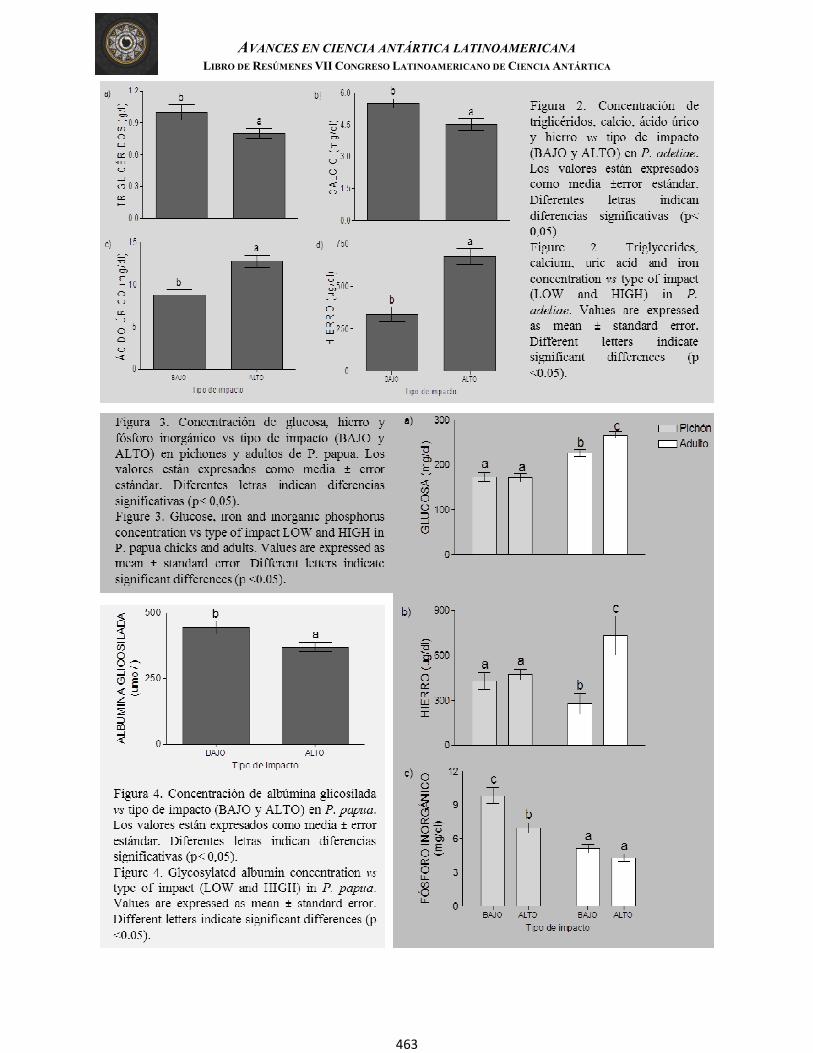
AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
463

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
464
Por otro lado, adiferencia de lo observado para Adelia, no se registraron diferencias en los niveles de triglicéridos, calcio y ácido úrico entre los pingüinos Papúa que habitan zonas de alto y bajo impacto. En el caso de Adelia, todos los parámetros bioquímicos sanguíneos estudiados se ven afectados por la actividad antrópica (Fig.1 y 2). En Papúa, en cambio, se observa una menor influencia de la actividad humana ya que en la ZAI únicamente se encuentran: aumentados los niveles de hierro y glucosa (adultos), disminuidos los niveles fósforo inorgánico (pichones) (Fig.3), y disminuidos los niveles de albúmina glicosilada (ambos estadíos) (Fig.4). CONCLUSIONES En el presente trabajo pudimos observar que, de las dos especies estudiadas, P. adeliae se encuentra más afectada por la contaminación asociada a las actividades antrópicas. Esta situación podría relacionarse a la ontogenia de cada especie y sus preferencias de hábitats, pese a que comparten el mismo género (Mc Clintock, 2008). Además, la determinación de valores basales de parámetros bioquímicos sanguíneos, proporciona información sobre el estado metabólico-sanitario presente de estas poblaciones de pingüinos y un punto de referencia para evaluaciones futuras o comparaciones con otras poblaciones. Por último, en virtud de nuestras observaciones y en desacuerdo con otros estudios realizados como el de Carney y col. (1999), donde se sugiere que las actividades humanas no afectan las poblaciones de pingüinos, consideramos que debe prestarse mayor atención a la clara determinación de las alteraciones fisiológicas de los pingüinos, ya que allí subyace la respuesta real al fenómeno de impacto antrópico. AGRADECIMIENTOS Este proyecto fue realizado con el subsidio PICTO 0091 (FONCyT e IAA-DNA) y la Universidad Argentina JF Kennedy. Se agradece la colaboración de la dotación de la Base Antártica Esperanza. REFERENCIAS Acero, J. y col. 1996. Revisión ambiental de las actividades antárticas en Bahía Esperanza, Península Antártica. Instituto Antártico Argentino. Publicación especial N°26. Carney, K. y William S. 1999. A Review of Human Disturbance Effects on Nesting Colonial Waterbirds. Waterbirds 22(1): 68-79. Carlini, A. y col. 2007. Breeding succes and population trends in Adélie penguins in areas with low and high levels of human disturbance. Polar Biol. 30:917-924 Carlini, A. y col. 2009. Responses of P. adeliae and P. papua populations to environmental changes at Isla 25 de Mayo (King George Island). Polar Biol (2009) 32:1427–1433. Culik, B. y col. 1990. The Effect of Disturbance on the Heart Rate and Behaviour of Adélie Penguins (Pygoscelis adeliae) During the Breeding Season. Antarctic Ecosystems, pp 177-182 Di Rienzo, J. Guzman, A. y Casanoves, F. 2002. A Multiple Comparisons Method based On the Distribution of the Root Node Distance of a Binary Tree Obtained by Average Linkage of the Matrix of Euclidean Distances between Treatment Means. JABES 7(2), 129-142. Giesse, M. 1998. Guidelines for people approaching breeding groups of Adélie penguins (Pygoscelis adeliae). Polar Record, 34, pp 287-292. InsfoStat 2012. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Di Rienzo J.A., Casanoves F., Balzarini M.G., Gonzalez L., Tablada M., Robledo C.W. Lowry, O. y col. 1951.Protein measurement with the phenol reagent.J. Biol. Chem. 193: 265-275. Mazón, H. 2013. GAT 04 “gestión de bases antárticas” informe CAV 2012-2013 “Base Esperanza”, Programa de gestión Ambiental y Turismo, DNA Mcclintock, J. y col. 2008. Ecological responses to climate change on the Antarctic Peninsula. Am. Scientist 96:302-310.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
465
EFECTO DE LA RADIACIÓN ULTRAVIOLETA SOBRE LA RAZÓN RNA: DNA DE COPÉPODOS ANTÁRTICOS Y DE LA ZONA CENTRAL DE CHILE
(Effects of the ultraviolet radiation on the RNA: DNA ratio of Antarctic and central Chile zone copepods)
Lagos, P. F.1 y K. Manríquez1
1Universidad Andrés Bello, Facultado de Ecología y Recursos Naturales, Escuela de Ciencias del mar. Republica 440, Santiago, Chile. Email: [email protected] RESUMEN El zooplancton se caracteriza por desempeñar un rol primordial en los ecosistemas marinos en relación al traspaso de energía entre el fitoplancton y los niveles tróficos superiores. Pese a su importancia, poco es lo que se conoce sobre la influencia del forzamiento ambiental sobre la dinámica del zooplancton, la cual se encuentra lejos de ser apropiada para fines de diagnóstico y de modelación. Se ha señalado que el zooplancton es sensible a las perturbaciones ambientales externas (Harris et al., 2000; Irigoien et.al, 2000) y es por esto que puede actuar como un indicador sensible de la variabilidad ambiental a escalas espacio–temporales diversas, incluso a aquellas asociadas a efectos globales (ejemplo: calentamiento global: Beaugrand et.al., 2002). Si consideramos la importancia ya mencionada que tiene el zooplancton dentro de un ecosistema pelágico, es necesario entender, que como todo organismo que habita en nuestro planeta, el zooplancton está inmerso dentro de un sistema tridimensional, donde las variables físicas, como la radiación ultravioleta, juegan un papel crucial al momento de definir estrategias de supervivencia que adopta un grupo de organismos en particular y en modificar los caracteres de historia de vida de éste. Se ha señalado que la disminución del ozono estratosférico es un problema global, siendo en el hemisferio sur, particularmente en Chile, en donde se ha documentado una disminución de hasta el 50% del ozono que cubre la Antártica durante la primavera y un aumento a una razón de 14% por década de la radiación ultravioleta de 305 nm., en la zona central de Chile, lo que permite inferir que el ozono que cubre esta región está disminuyendo (Cabrera et.al. 2000). Desde una perspectiva oceanográfica, este tipo de radiación puede considerarse un factor de importancia ya que un 95% de la radiación UV penetra en el agua y hasta un 50% llega hasta una profundidad de 3 mt. (Delgado, 2003). No obstante, se ha prestado escasa importancia a los efectos que la radiación ultravioleta pudiese tener sobre especies zooplanctónicas de latitudes medias, como la zona central de Chile, especialmente teniendo en cuenta las alzas que este tipo de radiación ha tenido en estas áreas en las últimas décadas y menos aún a cuáles son los efectos que este tipo de radiación pudiese tener en grupos zooplanctónicos Antárticos claves en la trama trófica, donde el derretimiento de la capa de hielo inducida por el calentamiento global a generado disminuciones en las áreas oceánicas cubiertas por hielos, generando un incremento de las zonas marinas expuestas a la radiación UV, la cual es letal para el zooplancton que no es capaz de ajustar su protección contra la radiación UV, radiación que es considerada un factor de estrés importante para el zooplancton pues procesos de reparación enzimáticos y destoxificación de elementos reactivos a oxígeno (ROS) son menores a bajas temperaturas (Rautio & Tartarotti, 2010). Considerando los antecedentes antes señalados es que nos proponemos como hipótesis de trabajo, que la radiación UV ejerce un efecto diferencial en la relación ARN:ADN influyendo a escala fisiológica sobre grupos funcionales claves como los copépodos, pero a diferentes magnitudes según la localidad donde habitan y el grado de exposición a la radiación UV a la que están expuestos. Nuestro objetivo general fue evaluar en forma experimental los daños fisiológicos y como varia la razón ARN: ADN de copépodos en dos localidades distintas (1) Bahía Fíldes (62°11’S; 58°54’O) y (2) Bahía Quintay (33°11’S; 71°42’O).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
466
Para poder responder nuestra hipótesis planteada, nuestro diseño experimental consistió en evaluar los efectos de la radiación UV en copépodos Antárticos y de la zona central de Chile, de donde se obtuvieron muestras mediante redes de zooplancton de 200 µm de tamaño de poro, a dos profundidades, superficial y 15 metros. Las muestras extraídas fueron trasladas al laboratorio de INACH (Bahía Fíldes) y CIMARQ (Universidad Andrés Bellos) de forma inmediata para ser utilizadas en ensayos experimentales (y no sufrieran daños posteriores). Para esta experiencia solo se utilizaron copépodos, los cuales fueron experimentalmente irradiadas por periodos de tiempo de 4 y 8 horas bajo distintos tratamientos lumínicos los cuales correspondieron a luz ultravioleta UV como grupo experimental, luz blanca como control positivo (C+) y ningún tipo de luz como control negativo (C-). Luego de los cuales se cuantificó diferencias significativas en la razón ARN: ADN de copépodos capturados e irradiados en ambas localidades. Utilizando el kit de extracción (E.Z.N.A. RNA: DNA isolation Kit, Omega Bio-Tek).
Figura 1: Modelo general lineal (ANDEVA) para la razón ARN: ADN de copépodos Antárticos (Izquierda) y copépodos de Quintay (Derecha) expuestos a 4 y 8 horas de radiación ultravioleta. C (-) Control negativo, C (+) Control positivo, UV grupo experimental expuesto a radiación UV. Grafico muestra interacción F (2, 24) = 24,102; P = 0,000. Datos ± Desviación estándar. Figure 1: General lineal model (ANOVA) for the RNA:DNA ratio of Antarctica (Left) and Quintay (Right) exposed to 4 and 8 Hours of ultraviolet radiation. (C-) Negative control, (C+) Positive control, UV experimental group expose to radiation. Graph shows interaction F (2, 24) = 24,102; P = 0,000. Data ± Estándar deviación. Los resultados indican que existen diferencias significativas en la razón ARN:ADN principalmente asociadas a los tiempos de exposición a radiación ultravioleta y a las diferentes localidades de estudio, observándose que copépodos Antárticos responden de forma más lenta a la radiación UV, mostrando una razón ADN:ARN más elevada luego de 8 horas de exposición a radiación UV mientras que copépodos de la zona central de Chile son más sensibles a este tipo de radiación en las primeras horas de exposición mostrando una razón ARN:ADN más elevada en las primeras 4 horas de exposición a luz UV no observándose diferencias significativas en los valores de la razón ARN:ADN en los grupos control positivo y negativo de ambas localidades de estudio (Fig.1). Los resultados fueron analizados mediante ANDEVA de dos vías. Los resultados reflejaron cómo distintos ensambles de copépodos presentan estrategias diferenciales de supervivencia frente a eventos de radiación ultravioleta. Se tienen antecedentes de que este tipo de estudio ha sido muy utilizado en organismos intermareales (Wieters, 2005), pero aun es escasa la información que hay en relación a organismos planctónicos, siendo los modelos de estudio más analizados el bacterioplancton (Hewson et al. 2006). Al ser este un trabajo uno de los pocos trabajos existentes hasta el momento en como la radiación ultravioleta es un factor que modifica la fisiología de copépodos, las complicaciones sobre las implicancias de este tipo de trabajo pueden ser insospechadas y sólo atribuibles a un factor filogenético que al parecer es un factor

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
467
que sólo puede ser calculado y explicado en función de la claridad de los resultados obtenidos, los cuales indican que existen efectos asociados a la radiación UV tanto en copépodos Antárticos como en copépodos de la zona central de Chile. Como conclusión es de esperar que las respuestas fisiológicas de los copépodos de ambas localidades de estudio estén relacionadas con la escala de tiempo a la que organismos de la zona central de Chile han sido expuestos a dosis elevadas de radiación ultravioleta, donde alzas sólo se han reportado en los últimos años, en contraste con la zona Antártica que exhibe índices elevados de radiación UV desde los años 80 (Gies et al. 2009; Laat et al. 2010; Purich & Son, 2012). Finalmente se espera que en un escenario de aumento de radiación UV los efectos sobre distintas comunidades de copépodos puedan presentar respuestas acopladas en donde sin importar la distribución o historias de vida de los organismos zooplanctónicos la radiación UV será un factor que module y cambie la forma en que se estructuran la comunidades de copépodos. Es por esto que la relevancia de este trabajo es el impulso a generar nuevas interrogantes sobre el tema y a seguir desarrollando temáticas que ayuden a entender como las comunidades zooplanctonicas, en general, están siendo afectadas de una manera directa al aumento en la radiación Ultravioleta. AGRADECIMIENTOS Proyecto P01-11 Instituto Antártico Chileno (INACH). REFERENCIAS Cabrera, S. 2000. Radiación ultravioleta en Chile : variaciones con la altura y la latitud. Boletín Cientifíco, Asociación Chilena de Seguridad 66–71. Delgado, L. 2003. Índice ultravioleta. Departamento de Física, Universidad de Antofagasta. Gies, P., Watzl R., Javorniczky J., Roy C., Henderson S., Ayton J. and Kingston, M. 2009. Measurement of the UVR Exposures of Expeditioners on Antarctic Resupply Voyages. Photochemistry and photobiology 85, 1485–1490. Harris, R. P., Irigoien, X., Head, R. N., Rey, C., Hygum, B. H., Hansen, B. W., Niehoff, B., Meyer-Harms, B., and Carlotti, F. 2000. Feeding, growth, and reproduction in the genus Calanus. ICES Journal of Marine Science, 57: 1708–1726. Hewson, I., Steele, J. A., Capone, D. G. & Fuhrman, J. A. 2006. Temporal and spatial scales of variation in bacterioplankton assemblages of oligotrophic surface waters. Marine Ecology Progress Series 311, 67–77. Irigoien, X., Harris, R. P. and Head, R.N. 2000. Does turbulence play a role in feeding and reproduction of Calanus finmarchicus ?. Journal of Plankton Research 22, 399–407. Laat, A. T. J, van der R.J.A., Allaart M.A.F., Van Weele M., Benitez C.C., Leme N.M.P., Quel E., Salvador J. and Wolfram, E. 2010. Extreme sunbathing : Three weeks of small total O3 columns and high UV radiation over the southern tip of South America during the 2009 Antarctic O3 hole season. Geophysical research letters 37, 1–6. Purich A. and Son, S. 2012. Impact of Antarctic Ozone Depletion and Recovery on Southern Hemisphere Precipitation, Evaporation and Extreme Changes. Journal of Climate 25, 3145–3154. Rautio, M. and Tartarotti, B. 2010. UV radiation and freshwater zooplankton : damage, protection and recovery. Freshwater Reviews 3, 105–131. Williams R., Conway D.V.P. and Hunt, H.G. 1994. The role of copepods in the planktonic ecosystems of mixed and stratified waters of the Europeans shelf seas. Hydrobiologia 292/293: 521-530. Wieters, E. A. 2005. Upwelling control of positive interactions over mesoscales: a new link between bottom-up and top-down processes on rocky shores. Marine Ecology Progress Series 301, 43–54.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
468
METABOLITOS SECUNDARIOS BIO-ACTIVOS DE ORGANISMOS MARINOS DE LA ANTÁRTICA
(Bio-active secondary metabolites from Antarctic marine organisms)
Rovirosa J1. y San-Martín A1,2. 1Universidad de Chile, Facultad de Ciencias, Depto. de Química, Casilla 653, Santiago, Chile. 2Facultad de Ciencias, Universidad de Magallanes. Email: [email protected] INTRODUCCIÓN. En contraposición a lo que ocurre con plantas terrestres, el medio marino es menos estudiado desde el punto de vista del metabolismo secundario, no obstante que la fuente de recursos naturales más importantes de nuestro planeta es el mar. Además, existen numerosos estudios que demuestran que las interacciones químicas entre organismos e individuos, tanto de la misma especie como de especies diferentes, también se encuentran en el medio marino. Asimismo se ha comprobado que la mayoría de los metabolitos secundarios aislados desde organismos marinos (esponjas, algas, moluscos, etc.), son tóxicos para algunos peces, inhibidores de la fijación y/o crecimiento de larvas, antimicrobianos y/o antifúngicos, lo que permite suponer que dichas substancias constituyen un verdadero sistema de defensa químico. Los organismos marinos han desarrollado adaptaciones químicas y biológicas propias como respuesta a su medio ambiente, siendo probablemente una de ellas la producción de metabolitos secundarios para la supervivencia. La síntesis y acumulación de moléculas complejas le demanda al ente productor una enorme cantidad de energía. Si esta función se ha conservado durante la evolución, podría significar que la biosíntesis de tales compuestos ha ayudado a la sobrevivencia (a través de la adaptación) de dicho organismo. Las diferencias estructurales de los compuestos de origen marino con respecto a los terrestres y el gran número de ellos informados en la literatura, explican la esperanza puesta en encontrar sustancias de interés biomédico en los océanos. Sin embargo pocos de ellas han sido sometidas a estudios farmacológicos detallados y menos aún alcanzado el mercado de las drogas comerciales, aunque en numerosos casos se cita, basándose en ensayos simples, un amplio espectro de actividades biológicas, entre las que se puede mencionar: antialimentaria, repelente, feromonas, antimicrobiana, antivirales, citotóxicas, etc. Un medicamento que está en uso para el tratamiento de sarcomas del tejido blando contra el cáncer de ovario es Yondelis (Le Cesne, 2013), cuyo principio activo es trabectedina que fue aislada desde la ascidia Ecteinascidia turbinata, que habita en el mar Caribe (Gajdos y Elias 2011). Uno de los métodos mas utilizados por los agricultores en el control de pestes, es el uso de compuestos químicos, por su considerable éxito en la protección de los vegetales. Sin embargo, el uso continuo de un pesticida induce una selección y un aumento de resistencia en los insectos, lo que obliga a aumentar no solo las concentraciones de los compuestos en cada aplicación, sino también la frecuencia de su uso, aumentando la polución y los costos del cultivo. La búsqueda de nuevos compuestos químicos en el control de plagas ha cobrado importancia en los últimos tiempos, ya que representa una buena posibilidad de reemplazar a los pesticidas tradicionales. Los monoterpenos halogenados son compuestos típicos de algas de la familia Plocamiaceae y Rhizophyllidaceae (Orden Gigartinales). A diferencia de los monoterpenos de origen terrestre, que comúnmente presentan funciones oxigenadas, los de origen marino presentan un alto grado de sustituciones con átomos de halógeno. De los más de 160 monoterpenos halogenados marinos descritos, muy pocos han incorporado oxígeno en la molécula. Muchos compuestos contienen 5 o aún 6 halógenos en una estructura con 10 átomos de carbono. La familia Plocamiaceae, distribuida en todo el mundo, es una fuente importante de monoterpenos polihalogenados, los cuáles han sido estudiados

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
469
desde 1974, en diferentes especies del género Plocamium, tales como P. cartilagineum, P. violaceum, P. mertensi, por nombrar algunos. En el transcurso de nuestro programa de investigación de la química de los productos naturales de los organismos marinos de las costas de Chile y de la Antártica, ahora hemos aislado tres monoterpenos polihalogenados de P. cartilagineum conseguidas en Bahía Fildes, Península Antártica. En el presente estudio, dos de esos compuestos se sometieron a ensayos plaguicidas. MATERIALES Y MÉTODOS Recolección y extracción Plocamium cartilagineum (Dixon) se recogió por medio de buceo autónomo (-15m) en Bahía Fildes, Península Antártica. Un ejemplar de muestra (N º 13032) es depositado en el herbario de la Facultad de Ciencias, Universidad de Chile. El alga (820 g) se secó al aire, se molió y luego se extrajo con n-hexano exhaustivamente. El disolvente se eliminó entonces a presión reducida por medio de un evaporador rotatorio, obteniéndose un extracto oleoso. El extracto crudo se sometió a cromatografía en columna a presión normal y media presión sobre gel de sílice, utilizando n-hexano y cantidades crecientes de acetato de etilo como eluyente. A partir de las fracciones eluídas con n-hexano, fueron purificados aún más por HPLC en fase normal. Este procedimiento permitió aislar los compuestos 1, 2 y 3 que fueron identificados por métodos espectroscópicos habituales. Actividades biológicas En los ensayos de actividad insecticida / acaricida, los controles no tratados y el disolvente se replicaron x 10. Los insecticidas estándar se aplicaron en tres concentraciones y se replican tres veces. Se probaron ocho especies insectos / ácaros. Todos estos bioensayos fueron diseñados para detectar la actividad de contacto y de ingestión. También se realizó un bioensayo antialimentario con Spodoptera frugiperda. La edad / fases de insectos probados fueron tercer estadio para el gusano cogollero y el gusano de la raíz, áfidos inmaduros y adultos y araña de dos manchas blancas adultas. El cuarto estadio del gusano cogollero fueron utilizados en el bioensayo antialimentario. Todos los compuestos experimentados se formularon en un disolvente de acetona-agua (3:1) a una concentración de 1000 ppm. Los compuestos formulados se aplicaron mediante un pulverizador de correa hidráulico RESULTADOS Plocamium cartilagineum (L.) Dixon (Plocamiaceae, Plocamiales orden) se recogió en la Península Antártica. A partir del extracto crudo, se aislaron después de cromatografía y filtración en gel y HPLC, el compuesto 1, 2 y 3 (Figura 1). El componente principal aislado fue 2, junto con pequeñas cantidades de 1 y 3, todos ellos identificados por comparación de sus propiedades físicas y espectroscópicas con la reportada en la literatura (Rovirosa, J. 1990; Stierle, D.1979a; Stierle, D.1979b). Se estudiaron los efectos antialimentarios de dos monoterpenos polihalogenados (1 y 2) contra varias especies de insectos. El compuesto 1 mostró la actividad anti-insecto más potente entre los dos compuestos ensayados, principalmente contra Heliothis virescens. El compuesto 2 mostró una actividad moderada hacia Diabrotica undecimpunctata. La pequeña cantidad obtenida a partir del compuesto 3 no permitió realizar las pruebas biológicas.
Cl Cl
Cl
Cl
BrCl
Cl
R
Cl
R2 Cl3 OH
1
Figura 1. Compuestos aislados de P. cartilagineum.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
470
Figure 1. Compounds isolated from P. cartilagineum AGRADECIMIENTOS Se agradece a E. I. Du Pont de Nemours y al Instituto Antártico Chileno (Proyecto T_15-09). REFERENCIAS Gajdos, C. y Elias, A. 2011. Trabectedin: safety and efficacy in the treatment of advanced sarcoma, Clinical Medicine Insights Oncology ,5, 35-43. Le Cesne, A. 2013. 13 years of trabectedin, 5 years of Yondelis: what have we learnt?, Axel Expert Review of Anticancer Therapy, 13(6s), 11-19. Rovirosa, J., Sánchez, I., Palacios, Y., Darias, J. y San-Martín, A. 1990. Antimicrobial activity of a new monoterpene from Plocamiun cartilagineum from Antarctic Peninsula. J. Chil. Chem. Soc. 35,131-133. Stierle, D., Wing, R. y Sims, J. 1979a Marine natural products. XVI. Polyhalogenated cyclic monoterpenes from the red alga Plocamium cartilagineum of Antarctica Tetrahedron, 35, 2855-2859. Stierle, D. y Sims, J. 1979 Marine natural products. XV. Polyhalogenated acyclic monoterpenes from the red alga Plocamium of Antarctica Tetrahedron, 35, 1261-1370.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
471
VIGILANCIA DE VIRUS INFLUENZA EN AVES MARINAS DE ELEPHANT ISLAND, ANTARTIDA, DE 2010 a 2012
(Surveillance of Influenza virus in seabirds from Elephant Island, Antarctica, from 2010 to 2012)
Seixas, M.M.M.1; Araújo, J.1; Ometto, T.1; Hurtado, R.1; Kruger, L.; Petry, M.V.2; Durigon, E.L.1 1.Laboratory of Clinical and Molecular Virology, Biomedical Science Institute, São Paulo University. [email protected] 2 Laboratory of Ornithology and Marine Animals, Vale do Rio do Sinus University
INTRODUCTION Influenza A virus belongs to the Family Orthomyxoviridae and causes acute respiratory illness considered emerging zoonotic disease by the World Health Organization (WHO, 2012). The transmission occurs in humans by inhalation of aerosols containing the virus (NICHOLSON, 1998), while in poultry, the natural reservoir of the virus (REED et al., 2003), the type of transmission is fecal-oral (FEREIDOUNI et al., 2010). Approximately 61 species of seabirds migrate between Australia, New Zealand, South Africa, Antarctica and South America. The Antarctic avifauna consists of 40 different species of seabirds (BRITO, 2009), of which 19 reproduced within the continent, while the rest reproduced in oceanic islands of the sub-Antarctic region. This study aims to analyze the presence of influenza virus in samples of seabirds of the Antarctic environment, characterize the positive samples, and learn about the microbial ecology of Antarctic birds, potential health risks of the Antarctic ecosystem and animal and public health. MATERIAL AND METHODS Elephant Island (61°08'S; 55°07'W) is located in Northeast of the South Shetland Islands (Figure1). The study site was chosen for sampling, as the island is the point of nesting colonies of seabirds, seals and elephant seals breeding during the austral summer.
Figure 1: A. Location map of the South Shetland Islands; B. Map showing the location of Elephant Island in the South Shetland.
Figura 1: A. Mapa de localización de las Islas Shetland del Sur, B. Mapa que muestra la ubicación de la isla Elefante en las Shetland del Sur.
From 2010 to 2012, 493 seabirds from different species were captured using dip net in Elephant Island, Antarctic region. Orotracheal and cloacal sterile swabs were collected from each bird and placed in sterile cryotubes containing 500µL of transport media (glycerol 5% in phosphate buffered saline with

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
472
penicillin and fungizone) according to WALLENSTEN (2007). All samples were immediately transferred to liquid nitrogen in the field and stored at freezer -70ºC until processing. The project is underway, therefore, the samples will be analyzed further. The extraction and detection of viral material will be carried out in the Laboratory Biosafety Level 3+ "Prof. Dr. Klaus Eberhard Stewien", in Biomedical Science Institute of São Paulo University, Brazil. The extraction will be done by "Magmax ™ -96 Total RNA Isolation Kit", according to the manufacturer's instructions, using "Magmax ™ Express" Applied Biosystems® equipment. The detection will be done by Real Time One Step RT-PCR kit using the Taqman ® AIV-M Reagents (Applied Biosystems ®). Samples positive for influenza will have the gene coding for hemagglutinin and neuraminidase amplified. The DNA sequencing will be performed using "BigDye Terminator Cycle Sequencing Ready Reaction Kit - Ampli Taq DNA Polymerase" (Applied Biosystems ®). RESULTS We will analyze 493 samples already collected from tracheal and cloacal swab seabird species Catharacta lonngbergi (Skua), Daption capense (Cape petrel), Pygoscelis antarcticus (Chinstrap penguin) and Pygoscelis Papua (Gentoo penguin) between the years 2010 and 2012 in the Elephant Island, Antarctica (Table1). Samples have not yet been detected once the project is underway.
2010 2011 2012 specie n° samples n° samples n° samples TOTAL
Catharacta lonnbergi 25 24 30 79 Daption capense 53 32 30 115 Pygoscelis antarcticus 22 62 40 124 Pygoscelis papua 28 36 111 175 TOTAL 128 154 211 493
Table 1. Number (n°) of samples collected by species, year and the total. Tabla 1. Número (n °) de muestras extraídas por especie, año y el total.
DISCUSSION In recent times the contact between humans and animals in Antarctica has increased due to the research bases and increased tourism (THOMAZELLI, 2010). This endangers the integrity of Antarctic environment, increasing the possibility of new species input, contamination by pollutants, disease in native species (MEC, 2006). The possibility of exotic diseases in Antarctic fauna and flora is known since the creation of the Antarctic Treaty in 1959, and the risk is of concern since the first meeting of the Scientific Committee on Antarctic Research (SCAR) (KERRY, 1998). CONCLUSION Lack of knowledge about biological and genetic diversity, especially of microbial communities, emphasizes that prevention, and monitoring should be considered in planning any activity in Antarctica. The continent has indisputable scientific importance, so knowledge of its characteristics and natural phenomena may clarify issues of regional and global importance. It is important to monitoring wildlife in Antarctica, especially knowing that influenza A virus spreads so quickly through the planet. ACKNOWLEDGMENT Elisa Petersen, Fernanda Colabuono, Gabriel Vier, Julia Finger, Liana Chesini, Roberta Piuco, and the Brazilian Navy.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
473
REFERENCES Brito, T. 2009. Antártica: Bem comum da humanidade. Brasília. Ministério do Meio Ambiente.69p. Fereidouni, S. R.; Werner, O.; Starick, E.; Beer, M.; Harder, T. C.; Aghakhan, M.; Modirrousta, H.; Amini, H.; Moghaddam, M. K.; Bozorghmehrifard, M. H. et al. 2010. Avian influenza vírus monitoring in wintering waterbirds in Iran, 2003-2007. Virology Journal. 7(43): 1-14. Kerry, K.; Riddle, M.; Clarke, J.1998. Diseases of Antactic wildlife: A report on the ‘Workshop on Diseases of Antactic Wildlife’. Australian Antactic Division. MEC. 2006. Antártica : ensino fundamental e ensino médio / coordenação Maria Cordélia S. Machado, Tânia Brito. – Brasília : Ministério da Educação, Secretaria de Educação Básica, 2006. 167 p. : il. (Coleção explorando o ensino ; v. 9). Nicholson, K. G. 1998. Human influenza. In: NICHOLSON, K. G.; WEBSTER, R. G., HAY, A. J. Textbook of influenza. Blackwell Science, London, England. p. 219-264. Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. 2003. Birds, Migrations and Emerging Zoonoses: West Nile Virus, Lyme Disease, Influenza A and Enteropathogens. Clin.Med & Res., v.1, n1:5-12. Thomazelli L.M., Araujo J., Oliveira D.B., Sanfilippo L., Ferreira C.S., Brentano L., Pelizari V.H., Nakayama C., Duarte R., Hurtado R., Branco J.O., Walker D., Durigon E.L. 2010. Newcastle disease virus in penguins from King George Island on the Antarctic region. Vet. Microbiol. 146: 155–160.. Wallensten, A.T.; Munster, V.J.; Latorre-Margalef, N.; Brytting, J.; Fouchier, R.A.M. et al. 2007. Surveillance of Influenza A virus in migratory waterfowl in Northern Europe. Emerg Infec Dis. v.13, n.3. 404-411. WHO. World Health Organization. 2012. Available in: www.who.int/zoonoses/emerging_zoonoses/en/ [2012 mar 28].

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
474
MENSURANDO OS NÍVEIS DE ESTRESSE EM PINGUIM-ANTÁRTICO (PYGOSCELIS ANTARCTICA): UMA COMPARAÇÃO ENTRE AS FASES DE REPRODUÇÃO E MUDA
(Measuring stress levels in chinstrap penguins (Pygoscelis antarctica): a comparison between nesting and molting stages)
Souza, J.S.1, 2, Costa, E.S.1,2, Convey, P.3, Viau, P.4, Oliveira, C.A.4, Torres, J.P.M.1 y M.A.S. Alves2
1Laboratório de Radioisótopos Eduardo Penna Franca, Universidade Federal do Rio de Janeiro, (UFRJ), Rio de Janeiro, RJ, Brazil. Email: [email protected] 2Laboratório de Ecologia de Aves, Universidade do Estado do Rio de Janeiro (UERJ), Rio de Janeiro, RJ, Brazil. 3British Antarctic Survey. NERC, High Cross, Madingley Road, Cambridge, Cambs, England 4Laboratório de Dosagens Hormonais, Universidade de São Paulo (USP), São Paulo, SP, Brazil INTRODUÇÃO: O estresse é inevitável e indispensável para a vida (Selye, 1976; Greenberg et al., 2002). Diariamente animais são submetidos a uma série de estressores em potenciais como privação de alimento ou água, disputa por territórios, variações das condições ambientais (temperatura, luminosidade, umidade e pressão), presença de predadores, perturbação antrópica, além dos estresses com muda e reprodução (Greenberg et al., 2002, Wingfield, 2003). Organismos submetidos a fatores estressores sofrem uma série de alterações a nível hormonal, como aumento nos níveis de glicorticóides, que são hormônios indicadores dos níveis de estresse (Palme et al., 2005). Estas alterações hormonais permitem que os indivíduos suportem períodos de estresse por meio de alterações fisiológicas e comportamentais que visam garantir a sobrevivência do indivíduo em curto prazo suprimindo qualquer atividade que possa por em risco a vida do animal (Wingfield et al., 1998). Em aves, a corticosterona (CORT) é o principal glicocorticóide indicador dos níveis de estresse (Ringer, 1976; Palme et al., 2005) e pode ser medido nas excretas cloacais, por meio de radioimunoensaio (RIE). O período de reprodução e muda são ocasiões extremamente estressantes. Durante a reprodução o organismo requer muita energia para manutenção corporal e cuidado com a prole. Enquanto no período de muda, pinguins permanecem desprotegidos, ficando impossibilitados de entrar na água ou procurar alimentos por semanas (Ninnes, et al., 2009). A Antártica é considerada um ambiente estressor, embora o estresse mais evidente enfrentado pelos organismos seja o frio e o risco de congelamento, há uma variedade de outros estressores, como por exemplo a escassez de recursos, que são fatores significativos e extremos aos quais os organismos são expostos e precisam suportar para garantir sua sobrevivência (Mayr, 1988; Convey, 1996; Greenberg et al., 2002). Vivendo nos limites da vida, esses organismos podem ser bons modelos para estudar como a vida sobrevive em ambientes extremos (Convey & Stevens, 2007). O objetivo deste trabalho foi determinar a concentração de CORT e comparar variações nos níveis deste GC entre adultos maduros e adultos maduros em fase de muda em excretas cloacais de Pygoscelis antarctica capturados na Baia do Almirantado, Ilha Rei George, Península Antártica. MATERIAL E MÉTODOS As amostras foram coletadas durante o verão austral de 2010/2011, na Baía do Almirantado, Antártica. Os indivíduos de Pygoscelis antactica foram capturados durante dois períodos: reprodução (n=4) e muda (n=11). As amostras espontâneas foram armazenada em potes plásticos identificados e mantidas congeladas a 20 -ºC até o transporte para o Brasil.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
475
A determinação dos níveis de CORT nas excretas cloacais foram realizadas por radioimunoensaio (RIE), segundo protocolo geral de Nakagawa et al. (2003). Utilizando-se o conjunto diagnóstico comercial de duplo anticorpo (MP BIOMEDICALS, INC, CA-USA) no Laboratório de Dosagens Hormonais (LDH) da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo (FMVZ/USP). RESULTADOS E CONCLUSÃO Os resultados indicaram que a concentração média de CORT para Pygoscelis antactica foi de 5,14 ng/ml ± 3,04 . Sendo as médias para muda e reprodução de 5.24 ng/ml (± 3.42) e 4.88 ng/ml (± 1.96), respectivamente. Apesar de aparentemente os indivíduos em muda, demonstrarem valores mais elevados de CORT nas excretas que indivíduos em reprodução, os dois grupos analisados não diferiram significativamente. Consequentemente, podemos considerar que apesar de muda e reprodução representarem períodos estressantes distintos, eles apresentam relação similar quanto aos níveis de CORT liberados pelo organismo. Para melhor entendimento da fisiologia do estresse nesta espécie, serão necessárias mais análises considerando outros fatores como idade e sexo dos indivíduos. AGRADECIMENTOS O presente trabalho contou com apoio financeiro do CNPq (MCT/CNPq: 557049/2009-1) e FAPERJ (E-26/111.505/2010) e logístico do Programa Antártico Brasileiro (PROANTAR). REFERÊNCIAS Convey, P. 1996. The influence of environmental characteristics on life history attributes of Antarctic terrestrial biota. Biological Reviews, 71:191–225. Convey, P. and Stevens, M. I. 2007. Antarctic biodiversity. Science, 317(5846): 1877-1878. Greenberg, N., Carr, J.A. and Summers, C.H. 2002. Causes and consequences of stress. Integrative and Comparative Biology, 42: 508-516. Mayr, E. 1988. Toward a new philosophy of biology. Harvard University Press, Cambridge Mass. Nakagawa, S., Möstl, E. and Waas, J.R. 2003. Validation of an enzyme immunoassay to measure faecal glucocorticoid metabolites from Adelie penguins (Pygoscelis adeliae): a non-invasive tool for estimating stress? Polar Biology, 26:491–493. Ninnes, C. E., Waas, J. R., Ling, N., Nakagawa, S., Banks, J. C., Bell, D. G., Bright, A., Carey, P. W., Chandler, J., Hudson, Q. J., Ingram, J. R., Lyall, K., Morgan, D. K. J., Stevens, M. I., Wallace, J. and Möstl, E. 2009. Comparing plasma and faecal measures of steroid hormones in Adelie penguins Pygoscelis adeliae. Journal of Comparative Physiology Part B, 180:83–94. Palme, R., Rettenbacher, S., Touma, C., El-Bahr, S.M. and Mostl, E. 2005. Stress hormones in mammals and birds: comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Annals of the New York Academy of Sciences, 1040: 162-171. Ringer, R.K. 1976. Adrenals. Pp. 372-382. In: Sturkie, P.D. (ed) Avian physiology. Springer, New York. Selye, H. 1976. The stress of life, rev. ed. McGraw Hill, New York. WINGFIELD, J. C. & SAPOLSKY, R. M. 2003. Reproduction and Resistance to Stress: When and How. Journal of Neuroendocrinology, Vol. 15, 711-724

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
476
CAMBIOS MICROCLIMÁTICOS INDUCIDOS POR OTC Y SUS CONSECUENCIAS EN EL CRECIMIENTO DE PLANTAS ANTÁRTICAS
(Microclimatic changes induced by OTC and their consequences on antarctic growth plants)
Sáez P1, Sanhueza C2, Corcuera LJ3, Cavieres L2, Sierra-Almeida A2, Alvear C4, Bravo LA4*
1Laboratorio Cultivo de Tejidos Vegetales, Centro de Biotecnología, Departamento de Silvicultura, Facultad de Ciencias Forestales, Universidad de Concepción, Casilla 160-C, Concepción, Chile. 2Laboratorio de ECOBIOSIS, Departamento de Botánica, Facultad de Ciencias Naturales y Oceanográficas,Universidad de Concepción.Barrio Universitario s/n, Concepción. Casilla 160-C.Concepción, Chile. 3Laboratorio de Fisiología Vegetal, Facultad de Ciencias Naturales y Oceanográficas,Universidad de Concepción. Barrio Universitario s/n, Concepción. Casilla 160-C.Concepción, Chile. 4Laboratorio de Fisiología y Biología Molecular Vegetal, Instituto de Agroindustria, Departamento de Ciencias Agronómicas y Recursos Naturales, Facultad de Ciencias Agropecuarias y Forestales, Universidad de La Frontera. Center of Plant, Soil Interaction and Natural Resources Biotechnology, Scientific and Technological Bioresource Nucleus. Universidad de La Frontera, Casilla 54-D, Temuco, Chile. [email protected]*. INTRODUCCIÓN En la Península Antártica los factores limitantes para el desarrollo de las plantas, tales como: bajatemperatura estival, estación de crecimiento corta, baja disponibilidad de agua y suelo, están siendo dramáticamente modificados debido al acelerado calentamiento regional. Estecambio ambiental ha sido correlacionado con un aumento significativo del tamaño de las plantas, cobertura y tamaño de laspoblaciones de Deschampsia antarctica y Colobanthus quitensis. Dentro de los métodos utilizados para evaluar el posible impacto del calentamiento sobre estos parámetros, las cámaras de calentamiento pasivo del tipo OTC (open top chamber), permiten examinar la respuesta de la vegetación al calentamiento experimental y comparar esta respuesta en diferentes comunidades (Bokhorst et al. 2007). Estos sistemas, además de modificar la temperatura dentro de ellos, pueden alterar también otros factores bióticos, tales como la humedad, luz (Marion et al. 1997) y disponibilidad de nutrientes (Rustad et al. 2001). Si bien, el nivel de alteración puede variar dependiendo de la estación del año y las condiciones climáticas (Bokhorst et al. 2011), en conjunto afectarán al crecimiento y desarrollo de las plantas. El objetivo de esta investigación fue estudiar los cambios producidos por OTC en las condiciones microclimáticas y su posible efecto sobre el crecimiento de plantas en la Antártida. MATERIALES Y MÉTODOS Cámaras de calentamiento pasivo (OTC) fueron instaladas en Diciembre de 2102 en poblaciones de D. antarctica y C. quitensis creciendo en Isla Rey Jorge, en las cercanías de la base polaca Arctowski (62° 09' S, 58°28' W) y posteriormente evaluadas en Febrero de 2013. Se instaló un sistema de registro microclimático (DL2 Datalogger) con el cual se evaluó dentro y fuera de OTC: temperatura en la superficie del suelo, temperatura foliar (utilizando sensores infrarrojos), radiación fotosintéticamente activa (PAR) y humedad de suelo. Adicionalmente se tomaron muestras de suelo, dentro y fuera de OTC para realizar análisis nutricional. Al comienzo de la estación de crecimiento (Diciembre) se midieron plantas de ambas especies, las que fueron marcadas en terreno. Al final de la estación de crecimiento (Febrero), estas plantas fueron nuevamente medidas y se determinó su porcentaje de crecimiento dentro y fuera de OTC, estas mediciones se realizaron en dos sitios, un sitio con cobertura principalmente de C. quitensis y otro sitio donde ambas especies estaban presentes. RESULTADOS Las OTC generaron cambios en todas las variables microclimáticas evaluadas, los cuales fueron dependientes de la hora del día. La temperatura del aire y PAR fueron incrementadas por la OTC

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
477
principalmente entre las 8:00 y las 16:00. Respecto a la humedad relativa, se redujo dentro de la OTC durante el día, pero fue más alta que en EA durante la noche. El incremento de la temperatura circundante dentro de OTC generó un incremento significativo de la temperatura foliar (evaluada en D. antartica) durante el día, mientras que durante la noche las temperaturas de la hoja no fueron diferentes dentro y fuera de OTC y se compensaron con la temperatura ambiental (Fig. 1). .
07:00:00 11:00:00 15:00:00 19:00:00 23:00:00 03:00:00
2D Graph 18
Horas del día
07:00:00 11:00:00 15:00:00 19:00:00 23:00:00 03:00:00
Hum
edad
Rel
ativ
a (%
)
66
68
70
72
74
76
78
80
82
84
Tem
peratura foliar (°C
)
-10
-5
0
5
10
15
20
Tem
pera
tura
del
aire
(°C
)
-2
0
2
4
6
8
OTC EA P
AR
(µm
ols fotones m-2s -1)
0
500
1000
1500
2000
Fig. 1. Diferencias entre la temperatura del aire, humedad relativa, radiación fotosintéticamente activa (PAR) y temperatura foliar de plantas antárticas creciendo en sistema de calentamiento pasivo (OTC) y espacio abierto (EA). Los registros fueron
tomados cada una hora durante el verano en Isla Rey Jorge. Fig.1. Differences among air temperature, air moisture, photosynthetically active radiation (PAR) and foliar temperature of antarctic plants growth under passive warming chambers (OTC) and open site (EA). Recordings were taken every one-hour
during summer in King George Island.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
478
Nitrogeno disponible Nitratos Amonio
Nitrog
eno (m
g/K
g)
0
10
20
30
40
50
60
Potasio dospinible Fósforo Olsen
Nu
trie
nte
s (m
g/K
g)
0
50
100
150
200
250
% M
ateria orga
nica
0
5
10
15
20
25
OTCEA
C. quitensis
% C
reci
mie
nto
0
20
40
60
80
100
120
140
160
EA OTC
C. quitensis D. antarctica
% C
reci
mie
nto
0
20
40
60
80
100
120
140
160
Fig. 3. Crecimiento (% de incremento del area) de los individuos de C. quitensis y D. antarctica en espacio abierto (EA) y
dentro de OTC (izquierda: sitio con presencia de ambas especies; derecha, sitio con C. quitensis). El área inicial fue determinada en Diciembre de 2012 y el área final en Febrero de 2013.
Fig. 3. Growth (% of area increase) in. quitensis and D. antarctica growing in open site (EA) and within OTC (left: site with both species; right: site with C. quitensis). The initial area was determined in December 2012 and final area in February
2013.
A pesar de no existir diferencias significativas respecto a porcentaje de crecimiento entre plantas dentro de OTC y EA, posiblemente debido a la relativamente corta duración del experimento, existe una influencia positiva de la OTC en el crecimiento de las plantas, principalmente en C. quitensis (Fig.3). Otros estudios han reportado una disminución en el crecimiento de especies dentro de las OTC, atribuido principalmente a una disminución en la humedad relativa del suelo (Bokhorts et al. 2011). A pesar de que nuestros resultados indican que la OTC generó una disminución de la humedad durante las horas de luz, esta tendencia se invierte durante las horas de oscuridad, lo que sugiere que este parámetro no tuvo una influencia negativa sobre el crecimiento. Dado que la asimilación de carbono es altamente dependiente de la temperatura y la luz, un incremento de ambos factores durante el día (Fig.
Fig.2. Cantidad de materia orgánica y concentraciones de nutrientes presentes en suelos dentro de OTC y en EA. Valores medios ± SE. El pH para ambos suelos fue de 4.3. Fig.2.Organic matter amount and nutrient concentrations from soils in OTC and EA. Mean ± SE. The pH for both soils was 4.3.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
479
1), pudo tener un efecto positivo sobre las tasas fotosintéticas, ya que se ha demostrado que la baja temperatura en terreno es un factor limitante para la fotosíntesis. Además, se ha reportado que la temperatura óptima de fotosíntesis para estas especies es mayor a las temperaturas de campo (Xiong et al. 1999). DISCUSIÓN Aumento de la productividad inducido por el calentamiento ha sido previamente reportado (Chapin et al. 1995, Hartley et al. 1999). Un efecto directo del calentamiento sería que incrementos en la temperatura pueden producir un aumento en la tasa de fotosíntesis lo cual conlleva a mayores tasas de crecimiento(Day et al. 1999). Por otra parte, efectos indirectos serían que mayores temperaturas en el suelo contribuyen aun descongelamiento más rápido de éste prolongando los periodos de captura de agua y nutrientes (Rustad et al. 2001). Además, un incremento en la disponibilidad de nutrientes sería el resultado de un incremento en las tasas de descomposición de la hojarasca y la mineralización de N por un aumento en la actividad microbiana (Rustad et al. 2001). Todas estas condiciones en conjunto estimulan la producción de biomasa y la captura de carbono (Strömgren y Linder 2002). En nuestro estudio, incrementos en el crecimiento observados en estas dos especies creciendo dentro de OTC, fueron relacionados con aumento en la concentración de Nitrógeno (nitratos, amonios), Fosforo y Potasio los cuales fueron determinados mediante análisis nutricionales de suelo (Fig. 2). Nuestros resultados indican que las modificaciones ambientales producidas por efecto de la OTC tuvieron un efecto positivo sobre el crecimiento de D. antarctica y C. quitensis, lo que estaría corroborando el efecto positivo del calentamiento sobre el incremento en las poblaciones de plantas antárticas. Sin embargo, es necesario prolongar los períodos de evaluación e incluir nuevasvariables a monitorearpara finalmente predecir el real impacto del cambio climático sobre la ecofisiología de las plantas antárticas. AGRADECIMIENTOS Proyecto PIA ART 1102. REFERENCIAS Bokhorst, S., Huiskes, A., Convey, P., Aerts, R. 2007. The effect of environmental change on vascular plant and cryptogam communities fron Falkland Island and the Maritime Antarctic. BMC Ecology, 7: 15-27. Bokhorst, S., Huiskes, A., Convey, P., Sinclair, B., Lebouvier, M., Van de Vijver, B., Wall, D. 2011. Microclimatic impacts of passive warming methods in Antarctica: implications for climate change studies. Polar Biol, 34: 1421-1435. Day, T., Ruhland, C., Grobe, C., Xiong, F. 1999. Growth and reproduction of Antarctic vascular plants in response to warming and UV-B radiation reductions in the field. Oecologia,119: 24–35. Marion, G., Henry, G., Freckman, D., Johnstone, J., Jones, G., Jones, M., Levesque, E., Molau, U., Molgaard, P., Parsons, A. 1997. Open-top designs for manipulating field temperature in high-latitude ecosystems.Global Change Biology, 3: 20-32. Rustad, L., Campbell, J., Marion, G., Norby, R., Mitchel, M., Hartley, J., Cornelissen, J., Gurevitch, J. 2001. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia, 126: 543-562. Strömgren, M., Linder, S. 2002. Effects of nutrition and soil warming on stemwood production in a boreal Norway spruce stand. Global Change Biology, 8: 1195-1204. Xiong, F., Ruhland, C., Day, T. 1999. Photosynthetic temperature reponse of the Antartic vascular plants Colobanthus quitensis and Deschampsia Antarctica. Physiologia Plantarum,106: 276-286.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
480
CAMBIOS MORFO-FISIOLÓGICOS EN POBLACIONES DE Colobanthus quitensis A LO LARGO DE UN GRADIENTE LATITUDINAL ANTARTICO-PATAGÓNICO
(Morphophysiological changes in Colobanthus quitensis populations along an antarctic-patagonian latitudinal gradient)
Sanhueza C1, Sáez P,2 Venegas R2, Vallejos V2, Corcuera1 LJ, Cavieres L3 , Sierra-Almeida A3, Bravo
LA4*
1Laboratorio de Fisiología Vegetal, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción. Barrio Universitario s/n, Concepción. Casilla 160-C. Concepción, Chile. 2Laboratorio Cultivo de Tejidos Vegetales, Centro de Biotecnología, Departamento de Silvicultura, Facultad de Ciencias Forestales, Universidad de Concepción, Casilla 160-C, Concepción, Chile. 3Laboratorio de ECOBIOSIS, Departamento de Botánica, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción. Barrio Universitario s/n, Concepción. Casilla 160-C. Concepción, Chile. 4Laboratorio de Fisiología y Biología Molecular Vegetal, Instituto de Agroindustria, Departamento de Ciencias Agronómicas y Recursos Naturales, Facultad de Ciencias Agropecuarias y Forestales, Universidad de La Frontera. Center of Plant, Soil Interaction and Natural Resources Biotechnology, Scientific and Technological Bioresource Nucleus. Universidad de La Frontera, Casilla 54-D, Temuco, Chile. [email protected]*. INTRODUCCIÓN Colobanthus quitensis (Kunth) es la única planta dicotiledónea que ha colonizado exitosamente la Antártica Marítima (Bravo et al. 2001). En Chile, esta especie se extiende a través de los Andes, donde habita humedales a gran altura, hasta las latitudes más polares donde se encuentra creciendo al nivel del mar (Hoffmann et al. 1998). Debido a este amplio rango de distribución C. quitensis muestra una considerable variabilidad morfológica (Moore 1972, Gianoli et al. 2004), encontrándose diferenciaciones ecotípicas entre poblaciones antárticas y andinas (Gianoli et al. 2004, Sierra-Almeida et al. 2007). Estas diferenciaciones estarían gatilladas en parte por los factores ambientales más característicos en cada hábitat(e.g., frió, sequía, salinidad o luz). Las poblaciones antárticas de C. quitensis han sido descritas como morfológica y fisiológicamente adaptadas a tener éxito en ambientes extremadamente fríos (Pérez-Torres et al. 2004), presentando una maquinaria fotosintética bien adaptada a bajas temperaturas (Xiong et al. 2000). Sin embargo, no se ha evaluado si poblaciones de C.quitensis distribuidas a lo largo de un gradiente latitudinal hacia el círculo polar, donde además de una disminución de la temperatura se presentan variaciones en la longitud del día, presentan variaciones morfológicas y fisiológicas relacionadas con los cambios en el ambiente. En el presente estudio se evaluaron aspectos morfológicos y fisiológicos de C. quitensis colectado en tres poblaciones diferentes dentro de un gradiente latitudinal antártico-patagónico. Las poblaciones seleccionadas correspondieron al sector Madre de Dios en Punta Arenas (50°16’S; 75°15’W); Isla Rey Jorge, en las cercanía de la base polaca Arctowski (62° 09' S; 58°28' W) e Isla Lagotellerie en Bahía Margarita (67° 52’ S; 68° 42' W). Las condiciones mesoclimáticas preponderantes en cada población se muestran en la Tabla 1. MATERIALES Y MÉTODOS Se evaluó si existen diferencias entre poblaciones respecto a: 1) anatomía foliar; 2) temperatura optima para la fotosíntesis y 3) partición de la energía lumínica a nivel del fotosistema II (PSII).Las plantas fueron colectadas de cada población con su propio sustrato y transferidas a una cámara en donde fueron cultivadas en contendores con tierra de hoja:turba:perlita (3:1:1), a 11/5 °C día/noche de temperatura ambiental, fotoperíodo de 16/8 horas luz/oscuridad y a 60 µmol de fotones m-2 s-1. Las muestras para anatomía foliar fueron fijadas en FAA y posteriormente teñidas con safranina fast green. Los cortes fueron analizados por microscopía óptica y las micrografías fueron evaluadas utilizando el software

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
481
ImageJ. Las curvas de fotosíntesis a diferentes temperaturas (5-35°C) fueron realizadas a 300 µmol de fotones con un IRGA (GFS-3000; Heinz Walz GmbH, Effeltrich, Alemania). La partición de la energía fue analizada utilizando un sistema de fluorescencia modulada (FMS II, Hansatech, R.U.). La tasa de transporte de electrones (ETR) fue calculada de acuerdo a Genty et al. (1989) y el apagamiento fotoquímico (qL)y no fotoquímico (NPQ) fueron determinados de acuerdo a Kramer et al. (2004).
Tabla 1. Temperatura promedio mensual, temperaturas máximas y mínimas absolutas y longitud del día para cada sitio de estudio durante Febrero 2013, correspondiente al último mes de la estación de crecimiento. Fuente: Base de datos de la Utah
State University y Solartopo. Table 1. Monthly average temperature, maximum and minimum absolute air temperatures and length day for each study site during February 2013 corresponding to the last month of the growing season. Source: Utah State University Database and
Solartopo.
Procedencia Ubicación T° Media
(°C) T°Max
(°C) T°Min (°C)
Longitud del día (horas luz/horas oscuridad)
Pta. Arenas 53° 10' S, 70°56' W 10.25 18.6 4.5 14/10
Arctowski 62° 09' S, 58°28' W 0.7 2.9 -4.1 15/9
I. Lagotelerie 67° 52’ S, 68° 42' W -3.5 1 -10.9 17/8
Tabla 2: Anatomía foliar de C. quitensis proveniente de distintas poblaciones dentro del gradiente latitudinal antártico-patagónico. Valores medios ± error estándar (n=20). Letras distintas indican diferencias significativas entre poblaciones
(P<0,05). Derecha: Sección transversal de hoja de C. quitensis proveniente de Punta Arenas (A-B), Arctowski (C-D) e Isla Lagotelerie (E-F). Barras representan 30 µm (izquierda) y 50 µm (derecha).
Table 2: Foliar anatomy of C. quitensis from different population within antarctic-patagonian latitudinal gradient. Media± SE (n=20). Different letters indicates significant differences among population (P<0.05). Right: Tranversal leaf section of
C. quitensis from Punta Arenas (A-B), Arctowski (C-D) and Lagotellerie Island (E-F). Bars represent 30 µm (left) y 50 µm (right).
Procedencia Pta. Arenas Arctowski I. Lagotelerie Área sección transversal (cm2)
0,44±0,02a 0,32±0,02b 0,23±0,02c
Largo superficie adaxial (µm)
1415,09±31,2a 1073,36± 41,9b 749,36±17,4c
Ancho mesófilo (µm)
378,73±7,56a 400,41±20,2ab 440,37±16,0b
Ancho mesófilo de empalizada (µm)
89,67±4,21a 90,94±3,48ab 93,02±3,12a
Ancho epidermis inferior (µm)
19,81±0,53a 17,51±0,57b 13,54±0,83c
Ancho epidermis superior(µm)
18,39±0,71a 18,43±0,70a 12,98±0,67b
Long. cel. epidermis inferior (µm)
21,88±1,03a 16,67±0,57b 14,73±1,58b
Long. cel. epidermis superior (µm)
21,40±1,33ab 23,52±,56a 14,77±2,18b
RESULTADOS Y DISCUSIÓN La anatomía foliar de C. quitensis de distintas procedencias (Tabla 1) indican que desde el extremo austral del continente (Pta. Arenas) hacia las poblaciones más polares, existe una disminución significativa del área de la sección transversal. Esta disminución está dada principalmente por una disminución en la longitud de las hojas, de modo que las hojas de poblaciones más polares presentan un menor ancho de la superficie adaxial. A medida que se avanza hacia el polo las hojas van
A B
C D
E F

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
482
disminuyendo su superficie expuesta (haz), pero mantienen el grosor del mesófilo. De acuerdo a Gianoli et al. (2004), estas diferencias entre distintas procedencias podrían constituir ecotipos morfológicos. Sin embargo, las respuestas similares de la fotosíntesis respecto a la temperatura y de algunos parámetros asociados a fluorescencia podría indicar que existen aspectos funcionales que no responden al gradiente latitudinal. Así, se determinó que individuos provenientes de la población más polar (I. Lagotellerie) presentaron una mayor fotosíntesis mantenida a mayor rango de temperatura (10 y 25 ºC), seguidos por los individuos prevenientes de Pta. Arenas y Arctowski (Fig. 1), quienes despliegan su máxima fotosíntesis en torno a los 15ºC. Destaca el hecho que a baja temperatura (rango de 5 a10 °C) las plantas provenientes de la mayor latitud presentan mayores tasas fotosintéticas,lo que demuestra la capacidad de estas plantas para maximizar en uso de energía en su crecimiento y reproducción en un corto periodo de tiempo, en el cual las condiciones son más favorables (Xiong & Day 2001). El incremento de las tasas fotosintéticas a temperaturas superiores a las de campo, confirman lo planteado por Xiong et al. (2000), quienes indican que las bajas temperaturas en la antártica serían limitantes para la fotosíntesis y por tanto temperaturas superiores a las de campo producen incrementos en las tasas fotosintéticas, esto se aplicaría no sólo para las poblaciones de C. quitensis en la antártica, sino también para las poblaciones más australes en el continente.
Fig. 1. Efecto de la temperatura en la fotosíntesis neta a saturación lumínica en plantas de C. quitensis procedentes de distintas poblaciones dentro del gradiente latitudinal antártico-patagónico. Valores medios ± error estándar (n=6). Fig.1. Photosynthetic response curve of C. quitensis from different population in the antarctic-patagonian latitudinal gradient. Media ± SE (n=6).
La partición de la energía lumínica a nivel del PSII mostró diferencias significativas en cuanto a la tasa de transporte de electrones (Fig. 2), en donde plantas provenientes deArctowski muestran las mayores ETR en comparación con plantas provenientes del sector I. Lagotellerie y Punta Arenas. Respecto a la actividad fotoquímica (qL) se determinó que las plantas de las poblaciones más polares mantienen mayoresqL que las plantas provenientes de Punta Arenas, con lo cual son capaces de mantener una menor presión de excitación sobre el PSII (1-qL) a medida que incrementa la luz. Debido a que en climas polares las bajas temperaturas y episodios de alta luz son condiciones típicas (Pérez-Torres et al. 2004), las plantas deben desarrollar mecanismos que les permitan mantener una baja presión de excitación sobre el PSII y evitar desbalances energéticos que podrían inducir a fotoinhibición (Wilson et al. 2006). Finalmente, es probable que, al igual queDeschampsia antarctica (Pérez-Torres et al. 2004), los individuos de C. quitensis provenientes de la Antártida,dependan principalmente de las reacciones fotoquímicas para la fotoprotección más que de la disipación del exceso de energía como calor (NQP), debido a que no existieron diferencias para este parámetro entre estas poblaciones y la de Punta Arenas.
Temperatura (°C)
0 5 10 15 20 25 30 35 40
Fo
tosí
nte
sis
(µm
ol C
O2 m
-2 s
-1)
0
2
4
6
8
10
12
14ArctowskiI. LagoteleriePta.arenas

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
483
Fig. 2. Tasa de transporte de electrones (ETR), eficiencia máxima del PSII (Fv’/Fm’), apagamiento fotoquímico (qL) y no fotoquímico (NPQ) en plantas de C. quitensis procedentes de distintas poblaciones dentro del gradiente latitudinal antártico-
patagónico. Valores medios ± error estándar (n=6). Fig. 2. Electron transport rate (ETR), maximal efficiency of PSII (Fv’/Fm’), photochemical (qL) and non photochemical
quenching (NPQ) in C. quitensis from different population in anantarctic-patagonian latitudinal gradient. Media ± SE (n=6). AGRADECIMIENTOS Proyecto PIA ART 1102. REFERENCIAS Bravo, L., Ulloa, N., Zuñiga, G., Casanova, A., Corcuera, L., Alberdi, M. 2001. Cold resistance in antarctic angiosperms. Physiologia Plantarum, 111: 55–65. Genty, B., Briantais. J., y Baker, N. 1989. The relationship between the quantum yield of photosynthetic electron transport andquenching of chlorophyll fluorescence. Biochem Biophys Acta, 990: 87-92 Gianoli, E., Hinostroza, P., Zúñiga-Feest, A., Reyes-Díaz, M., Caviares, L., Bravo, LA., Corchera, LJ. 2004. Ecotypic Differentiation in Morphology and Cold Resistance in Populations ofColobanthus quitensis (Caryophyllaceae) from the Andes of Central Chile andthe Maritime Antarctic. Arctic, Antarctic, and Alpine Research, 36(4): 484–489 Hoffmann, A., Arroyo, M., Liberona, F., Muñoz, M., Watson, J. 1998. Plantas Altoandinas en la Flora Silvestre de Chile. Santiago: Ediciones Fundación Claudio Gay. 281 pp. Kramer, D., Johnson, G., Kiirats, O. Edwards, G. 2004. New fluorescence parameters for the determination of QAredox state and excitation energy fluxes. Photosynthesis Research,79: 209–218. Pérez-Torres, E., García, A., Dinamarca, J., Alberdi, M., Gutiérrez, A., Gidekel, M. 2004. The role ofphotochemical quenching and antioxidants in photoprotection of Deschampsia antarctica. Funct. Plant Biol, 31: 731–741. Sierra-Almeida, A., Casanova-Katny, M., Bravo, L., Corcuera, L., Cavieres, L. 2007. Photosynthetic responses totemperature and light of Antarctic and Andean populations of Colobanthus quitensis (Caryophyllaceae). Revista Chilena de Historia Natural, 80: 335-343. Wilson, K., Ivanov, A., Öquist, G., Grodzinki, B., Srhan, F., Huner, N. 2006. Energy balance, organellar redox status and acclimation to environmental stress. Can. J. Bot. 84, 1355-1370. Xiong, F., Mueller, E. y Day, T. 2000. Photosynthetic and respiratory acclimation and growth responses of Antarctic vascular plants to contrasting temperature regimes. Am. J. Bot, 87: 700–710.
ET
R0
30
60
90
120
150
Fv'
/Fm
'
0,0
0,2
0,4
0,6
0,8
0 300 600 900 1200 1500 1800
qL
0,0
0,2
0,4
0,6
0,8
1,0
0 300 600 900 1200 1500 1800
NPQ
0
1
2
3
DFF (µmol fotones m-2 s-1)
Arctowski
I. Lagotelerie
Pta. Arenas

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
484
EL PROGRAMA CIENTÍFICO Y AMBIENTAL ANTÁRTICO URUGUAYO
(The Uruguayan Scientific and environmental program)
Abdala, J.1, Dematteis, S.1, Juri, E.1, Machado, A.1
1 Dirección de Coordinación Científica, Instituto Antártico Uruguayo. Av. 8 de Octubre 2958 CP 11600 Montevideo, Uruguay. [email protected] INTRODUCCIÓN La información global sobre los programas científicos nacionales es manejada en general por los administradores antárticos y en diferentes instancias internacionales (RAPAL, SCAR,RCTA,CPA), sin embargo muchas veces esa información no llega a los investigadores así como los mecanismos de participar en los mismos. El Programa Científico y Ambiental Antártico Uruguayo (PCAAU) es el conjunto de actividades de Investigación que promueve, coordina y supervisa el Instituto Antártico Uruguayo (IAU). Desde 1984 el Uruguay cuenta con una Base Científica Antártica en la isla Rey Jorge / 25 de mayo y desde esa fecha se desarrollan actividades científicas. Desde esa fecha hasta la actualidad los investigadores uruguayos de diferentes instituciones académicas y de servicios han mantenido una actividad ininterrumpida. Abarcando diferentes áreas del conocimiento como meteorología, biología, oceanografía, geología, psicología, medicina y paleontología, entre otras. A partir de 1996, Uruguay cuenta con una estación científica en la península Antártica. En los últimos años y siguiendo las recomendaciones y debates de la Comisión de asuntos científicos y ambientales de la Reunión de Administradores de Programas Antárticos Latinoamericanos (RAPAL), Uruguay ha seguido un proceso de organización de las actividades de investigación antártica. En ese sentido se han clasificado las mismas en Actividades de investigación, Actividades de investigación Institucionales y proyectos Científicos. METODOLOGÍA Para el proceso de selección de los proyectos y actividades de investigación, el IAU realiza un llamado público y abierto a presentar propuestas de investigación. Un Grupo de Trabajo Científico Asesor, externo al IAU, evalúa los contenidos científico-técnicos y selecciona las propuestas que encuentra convenientes. En última instancia el Consejo Directivo del IAU aprueba la ejecución de los proyectos y actividades seleccionadas por el Grupo de Trabajo Científico Asesor. El IAU aporta los traslados y manutención de los investigadores a las bases y estaciones antárticas así como gastos de materiales de los proyectos y actividades de investigación. Para ello cuenta con el apoyo de un avión Hércules C-130 de la Fuerza Aérea Uruguaya que realiza cinco vuelos por año y de un buque logístico de la Armada Nacional. La Dirección de Coordinación Científica del IAU tiene a su cargo la coordinación de todas estas actividades tanto a nivel nacional como internacional. RESULTADOS Actualmente el PCAAU lo integran diez instituciones, Facultad de Ciencias, Facultad de Química y Facultad de Ingeniería e Instituto de Bella Artes de la Universidad de la República, Instituto de Investigaciones Biológicas Clemente Estable, servicio Geográfico Militar, Servicio de Oceanografía, Hidrografía y Meteorología de la Armada, Dirección Nacional de Recursos Acuáticos, Dirección Nacional de Meteorología, Fuerza Aérea Uruguaya, con 11 proyectos, tres actividades de investigación y dos actividades institucionales con la participación de 67 investigadores (48 mujeres y 19 hombres) y la concurrencia a la Base Científica Antártica de 22 investigadores (16 mujeres y seis hombres).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
485
Siguiendo los lineamientos de SCAR en los mismos participan jóvenes investigadores con trabajos de tesis de grado y posgrado. En el mismo sentido se ha atendido a la participación internacional de los proyectos en cooperación con países como México, Chile, Argentina, Brasil, EEUU, Reino Unido y Venezuela entre otros. Las áreas del conocimiento en las cuales se desarrollan investigaciones actualmente son Microbiología, Ecología y Limnología, Ciencias de la Tierra y Ciencias Sociales. Dentro de estas áreas se desarrollan investigaciones de carácter fundamental y de aplicación, así como actividades de colecta de datos y monitoreo de variables ambientales que sirven de apoyo a otras investigaciones y a bases de datos internacionales. En el área Microbiología destacan los estudios de los procesos de desnitrificación de los ecosistemas en la Antártida, Identificación y bioprospección de levaduras de la Antártida, Búsqueda de nuevas enzimas bacterianas, Monitoreo de poblaciones bacterianas, Búsqueda y caracterización de bacterias promotoras del crecimiento vegetal, Aspectos tecnológicos y estructurales de enzimas activas a bajas temperaturas y la Caracterización de microorganismos antárticos como fuente de ácidos grasos poliinsaturados del tipo omega 3 y 6. En el área Ecología y Limnología se destacan estudios sobre la diversidad de crustáceos y estudios sobre las lagunas asociadas a los glaciares antárticos y el Relevamiento de Restos Marinos e identificación de estructuras de poblaciones de mamíferos marinos en KGI 25 de mayo. En el área de Ciencias de la tierra se destacan los estudios en la Meteorología antártica, Infraestructura Geodésica y Mareas Terrestres, Mareas Oceánicas y Monitoreo de la Capa de Ozono. En el área de Ciencias Sociales se destaca el Proyecto Austral (Construcción de archivos que contribuyan desde el campo del arte a difundir una mirada sobre la Antártida). Las perspectivas para el futuro inmediato son de fortalecer las convocatorias a proyectos y actividades de investigación con fondos de la Agencia Nacional de Investigación e Innovación y la remodelación y acondicionamiento de los laboratorios en la Base Científica Antártica Artigas y en la Estación Científica Antártica Ruperto Elichiribehety. También se adaptarán las plataformas marinas móviles en apoyo a la actividad de investigación así como el traslado de un helicóptero de la Fuerza Aérea Uruguaya. Se estima que con estas mejoras el Programa Científico y Ambiental Antártico Uruguayo podrá tener un incremento tanto en calidad como en cantidad de participantes y avanzar hacia las actividades de cooperación internacional con programas de otros países.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
486
MONITOREO AMBIENTAL EN BASES ANTÁRTICAS: UNA HERRAMIENTA PARA LA TOMA DE DECISIONES Y LA MEJORA DEL DESEMPEÑO AMBIENTAL
(Environmental monitoring in Antarctic Stations: a tool for decision-making and improvement environmental performance)
Calisto-Ulloa, N. y Gómez-Fuentes, C.
CIMAA, Departamento de Ingeniería Química, Universidad de Magallanes, Punta Arenas, Chile. [email protected] INTRODUCCIÓN Actualmente se reconoce que los programas de monitoreo ambiental son una herramienta eficaz para la adopción de medidas de mitigación en la Antártica (Deacon, 2003; Kennicutt et al., 2009; Harris, 1998), sin embargo la implementación de programas regulares es compleja debido a las dificultades logísticas y técnicas, siendo usual encontrar sólo medidas esporádicas. Las plantas de tratamiento de aguas residuales en la Antártica en general carecen de control de calidad de sus efluentes, debido a su aislamiento, la ausencia de laboratorios cercanos y falta de personal capacitado. Esta situación no permite saber si los sistemas funcionan adecuadamente y si existe un impacto real en el medio ambiente. En la actualidad existe preocupación a nivel internacional, debido a que se han realizado estudios que muestran que los efluentes estarían afectando la calidad de las aguas cercanas a las descargas (Gröndahl et al., 2009; Deacon, 2003). Un parámetro que se ha identificado en el medio son las bacterias coliformes, las cuales tienen claro origen humano y constituyen un excelente indicador de la dispersión de la contaminación y del impacto en el punto de vertido y sus alrededores (Hernandez et al., 2012; Hughes & Nobbs, 2004; Delille & Gleizon, 2003). Este trabajo presenta los resultados obtenidos por el Centro de Investigación y Monitoreo Ambiental Antártico, CIMAA, en la Base Antártica Chilena O’Higgins. CIMAA, perteneciente al Departamento de Ingeniería Química de la Universidad de Magallanes, trabajando en coordinación con el Instituto Antártico Chileno, INACH, y el Departamento Antártico del Ejército de Chile, ha generado información científica para la toma de decisiones respecto de los sistemas de tratamientos de aguas residuales, representando un modelo de cooperación entre operadores antárticos y Universidad, con la finalidad de proteger el medio ambiente. METODOLOGÍA Durante los años 2012 y 2013 se realizaron monitoreos de la calidad del aguas de mar en las proximidades de la Base O’Higgins, de acuerdo al plan de monitoreo presentado en la Figura 1. Este plan incluye la toma de muestras en la costa y en el mar, utilizando como puntos centrales las dos descargas existentes en el Cabo Legoupil (Base Chilena O’Higgins y Base Alemana GARS-O’Higgins), alrededor de los cuales se trazan semicírculos para identificar la dirección del impacto. En el mes de enero, de ambos años, estos monitoreos se realizaron por personal del CIMAA con el apoyo personal de las dotaciones antárticas de la Base O’Higgins y durante el mes de abril el personal de la dotación replicó este muestreo. Adicionalmente durante ambos años se tomaron muestras mensuales del efluente de la planta de tratamiento de aguas residuales de la base O’Higgins. En las muestras de agua de mar y en las muestras del efluente de la planta de tratamiento se realizó la cuantificación de coliformes totales utilizando la técnica de filtración por membrana y cultivo en medio cromogénico específico, Chromocult Coliform Agar ES de Merck.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
487
Figura 1: Plan de muestreo
Figure 1: Sampling plan RESULTADOS Y DISCUSIÓN La Figura 2 muestra la concentración de coliformes en el borde costero. Se observa que la primera medida realizada en enero de 2012 mostraba que la costa se encontraba afectada por las descargas de los efluentes tratados, este impacto se limitaba exclusivamente a las muestras de costa ya que no se detectaron coliformes en muestras más allá de los 10 metros del borde costero. Estos resultados unidos a los obtenidos para la calidad de los efluentes de las planta de tratamiento, presentados en la Figura 3, indicaban que el sistema no estaba funcionando adecuadamente. La información obtenida durante el muestreo del mes de enero permitió a los responsables de la Base O´Higgins detectar la necesidad de realizar un mantenimiento a la planta de tratamiento de aguas residuales. Las acciones correctivas se realizaron en el mes de marzo del año 2012 y esto se reflejó en los resultados en el medio marino del mes de abril, los cuales fueron significativamente mejores a los obtenidos durante el mes de enero, como se puede observar en la Figura 2. Estos nuevos resultados son coincidentes con la mejora en la calidad de los efluentes de la planta de tratamiento de la Base O’Higgins (Figura 3). Durante el mes de enero del año 2013 no se detectan coliformes en el medio marino y tampoco en los efluentes del sistema de tratamiento de aguas residuales, situación que se replicó para los meses siguientes. Estos resultados indicarían un correcto funcionamiento del sistema de tratamiento de aguas residuales de la Base O’Higgins y una disminución en el impacto generado en el medio ambiente marino por la presencia de esta Base.
EO EO+1 EO-1 EA
EA-1
O4
EA+1
OC10
OC9
O3

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
488
Figura 2: Concentración de coliformes totales en el borde costero para los años 2012 y 2013.
Figure 2. Total coliform concentration in the coastline for 2012 and 2013.
Figura 3: Concentración de coliformes totales en el efluente de la planta de tratamiento de aguas residuales de la Base
O'Higgins para los años 2012 y 2013. Figure 3. Total coliform concentration in the effluent of treatment plant wastewater O'Higgins Station for 2012 and 2013.
CONCLUSIONES Los resultados obtenidos en el medio marino en el año 2013, son consecuencia de la toma de acción inmediata una vez conocidos los problemas de operación del sistema de tratamiento. Se demuestra, por lo tanto, la importancia del monitoreo continuo como herramienta para la toma de decisiones; más aún, el contar con resultados históricos permite a las bases reconocer en forma oportuna posibles problemas en el tratamiento sin necesidad de llegar a situaciones críticas. Este trabajo presenta un ejemplo de distintas instituciones cooperando para el cuidado del medio ambiente antártico. Los resultados presentados muestran que es posible mejorar significativamente el desempeño de las bases antárticas mediante la capacitación de las dotaciones y el monitoreo periódico del medio ambiente y de los sistemas de tratamiento.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
489
AGRADECIMIENTOS Al INACH y al Departamento Antártico del Ejército de Chile, por financiamiento del proyecto CIMAA (MA_01-12), así como a las dotaciones de la Base O’Higgins en el período de ejecución de la actividad. REFERENCIAS Deacon C. 2003. Environmental health on Australian bases in the Antarctic and sub-Antarctic. Journal of Rural and Remote Environmental Health 2(1), 1-5. Delille D. and Gleizon F. 2003. Distribution of enteric bacteria in Antarctic seawater surrounding the Port-aux-Francais permanent station (Kerguelen Island). Mar Pollut Bull 46(9), 1179-83. Gröndahl F., Sidenmark J. and Thomsen A. 2009. Survey of waste water disposal practices at Antarctic research stations. Polar Research 28(2), 298-306. Harris C. M. 1998. Science and environmental management in the McMurdo Dry Valleys, southern Victoria Land, Antarctica. 72. Hernández, J.; Stedt, J.; Bonnedahl, J.; Molin, Y.; Drobni, M.; Calisto-Ulloa, N.; Gomez-Fuentes, C.; Astorga, S.; González-Acuña, D.; Waldenström, J.; Blomqvist, M. y Olsen, B. 2012. Human-Associated Extended-Spectrum beta-lactamase in the Antarctic. Applied and Environmental Microbiology, 78(6):2056-2058. Hughes K. A. and Nobbs S. J. 2004. Long-term survival of human faecal microorganisms on the Antarctic Peninsula. Antarctic science 16(3), 293-7. Kennicutt M., Wolff G. A., Alsup D. and Klein A. G. 1999. Long-term monitoring, McMurdo Station, Antarctica: pilot project design.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
490
BIOACUMULACION DE CONTAMINANTES ORGANICOS PERSISTENTES EN PINGUINOS PYGOSCELIS: UNA MIRADA A LOS PROCESOS DE BIOMAGNIFICACION
EN PENINSULA ANTARTICA
(Bioaccumulation of Persistent Organic Pollutants in Pygoscelis penguins: an insight to biomagnification processes in the Antarctic Peninsula)
Chiang, G.1, Rudolph, I.2, Jara, S.2, Munkittrick, K.1y R. Barra2
1Canadian Rivers Institute, Department of Biology, University of New Brunswick, Saint John NB Canada. [email protected] 2Unidad de Sistemas Acuáticos, Centro EULA-Chile, Universidad de Concepción, Concepción, Chile. The importance of studying Persistent Organic Pollutants (POPs) in Antarctica is the dynamics of these compounds in the environment through global fractionation (Wania & Mackay, 1993, Bengston Nash, 2011). Global fractionation explains why and how the concentrations of certain POPs are much higher in Polar areas (e.g. Arctic/Antarctic Regions) than in areas of warm-temperate climate. Physico-chemical properties of these compounds and surrounding environmental conditions (temperature and precipitation) explain the fractioning through volatilization-condensation-deposition phenomenon. Currently, this hypothesis remains verified by adopting different approaches to study either from an analytical or methodological standpoint (Bengston Nash, 2011). Although there is information that supports this hypothesis, mainly in the northern hemisphere, less is known about the behavior of POPs in the southern hemisphere. Despite the monitoring efforts of many Countries, POP data from the Antarctic remains fragmentary and scarce. The proposed project aims to provide information on the biomagnification of POPs through Antarctic aquatic food chain. The current study focused in the role of Pygoscelis penguins (as the principal Antarctic biomass) in bioaccumulation of POPs. Due to the conservation status of the Antarctic continent biota, Penguin feces is a valid alternative to assess POP concentrations and exposure because, (1) is a non lethal sampling approach that may be an adequate alternative to traditional invasive sampling procedures used in contaminant studies (2) feces from a nest will reflect the food source from chick(s) and mating penguins who forage in the vicinity of the breeding colonies at that time. The influence of the diet on the observed contaminant concentration was further investigated by stable isotopes (δ13C, δ15N) analysis. The study area includes sites near the Chilean Station General Bernardo O'Higgins, one of the main Chilean Antarctic stations, and Kopaitic Island, both located on the Antarctic Peninsula. Sampling was performed with the utmost care to avoid stress on the colonies of penguins (Gentoo, Chinstrap and Adélie penguins) and nests with chicks. Fresh stool samples from 5 nests for each species were collected for each colony. A metal blade used to collect droppings as close to the nest and choosing the freshest available. Samples were stored individually in aluminum foil, labeled, placed in airtight bags and stored at -20° C until processing in the laboratory. Extraction of PCBs from feces samples was based on the laboratory methods developed for other tissues (Montory et al. 2011). Identification and quantification of compounds were done by gas chromatography with electron capture detector (GC-ECD; Perkin Elmer gas chromatograph 9000 series Autosystem), using the external standard method. Standard mixtures consisted of 4 CLB1 (CLB1-A, CLB1-B, CLB1-C and-D) obtained from the National Research Council of Canada representing a total of 51 PCB congeners. Penguin dropping samples for each species were analyzed for stable isotopes of nitrogen (δ15N) and carbon (δ13C) at the Environmental Isotope Laboratory, University of Waterloo. Dried samples were ground and the aliquots (0.25–0.30 mg) weighed in tin cups and analyzed using a Delta Plus

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
491
Continuous Flow Stable Isotope Ratio Mass Spectrometer (Thermo Finnigan, Bremen, Germany) coupled to a Carlo Erba Elemental Analyzer (CHNS-O EA1108, Milan, Italy). The occurrence of 23 congeners in Adélie penguin’s droppings was determined with the maximum of 4.47 ng g-1 (wet weight (ww)) of PCB-121 followed by 3.30 ng g-1 (ww) of PCB-52. The total PCBs concentrations ranged from 10.06 to 16.19 (ng g-1 ww) with an average of 12 individual congeners detected per sample (Fig 1A). Chinstrap penguins (Fig 1B) shows a similar distribution, but lower concentrations compared to the observed PCBs concentration of the Adélie penguins (2.95 to 5.92 ng g-1 ww), which also had fewer congeners detected, averaging 9 congeners per sample. Gentoo penguins (Fig 1) showed the highest concentrations among the three species studied (5.15 to 100.73 ng g-1 ww). The stable isotope analysis indicates differences in δ15N between Gentoo and Chinstrap (> 9‰); Gentoo and Adélie (≈ 8‰) penguins, but minor differences between Adélie and Chinstrap (< 2‰). The isotopic signal of δ13C evidenced minor changes between Adélie and Chinstrap penguin colonies (< 0,5‰), but a greater decrease in δ13C is showed in Gentoo penguins, with differences ≈ 0,8‰ smaller to Adélie and > 1‰ to Chinstrap penguin colonies (Fig. 2A). A positive relationship between the total levels of PCBs and their respective δ15N is observed (r2 = 0.9898), with lower total PCBs concentrations in Chinstrap and higher in Gentoo penguins (Fig 2B).
Figure 1.- PCB concentrations (By Congener) in penguin droppings (Mean±Standard Error). (A) Adélie, (B) Chinstrap and
(C) Gentoo. The results in PCB contents in feces along with stable isotopes showed differences that could be attributed to food web position and carbon source. As differences in the diet of these species has been reported: the diet of the Adélie penguin (Pygoscelis adeliae) is based primarily on krill (Lynnes et al. 2004), although they may have a broader diet that includes fish and zooplankton (Tierney et al. 2009). The highest levels of δ15N in Gentoo penguin feces may be due to a more diverse and higher trophic level diet relative to Adélie and Chinstrap penguins (Fry 2006, Lynnes et al. 2004). Adélie and Chinstrap δ15N signature has a difference of < 2‰, that could result in a higher trophic position for Adélie (explaining higher POPs concentration), but variability is very high with some possible overlap in prey, and there could be a very small difference in diet responsible for the POPs differences. Along with trophic position, stable isotopes helps explain energy source and, since δ13C is similar between Adélie and Chinstrap colonies, but ≈1‰ smaller in Gentoo penguin feces. This is further evidence of the influence in diet on source of POPs showing that Gentoo penguins might get their prey deeper or farther offshore (Fry 2006). This differences in trophic position and penguin diet is consistent with previous data (Lynnes et al. 2004, Tierney et al. 2009), and shows that this approach is critical to identify possible sources of POPs and biomagnification processes in Antarctica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
492
Figure 2.- Correlation between: (A) stable Isotopes (δ13C‰ / δ15N‰), (B) δ15N‰ and total PCB concentration present in penguin droppings. Values are represented by mean±standard error. A: Adélie; C: Chinstrap; G: Gentoo. ACKNOWLEDGEMENTS Cpt. Luis Torres (Carabineros de Chile), for scuba diving sampling support during field work. REFERENCES Bengtson Nash S. 2011. Persistent organic pollutants in Antarctica: current and future research priorities. Journal of Environmental Monitoring, 13, 497-504. Fry, B. 2006. Stable Isotope in Ecology. Springer, New York. 308 p. ISBN 978-0-387-30513-4 Lynnes, A.S., Reid, K., & Croxall, J.P. 2004. Diet and reproductive success of Adélie and Chinstrap penguins: linking response of predators to prey population dynamics. Polar Biology, 27(9), 544-554. Montory, M., Habit, E., Bahamonde, P., Fernandez,P., Grimalt, J.O., Saez,K., Rudolph,I. & Barra, R.,2011. Polychlorinated biphenyls in farmed wild Onchorhynchus kisutch and Oncorhynchus mykiss from the Chilean Patagonia. Environmental Science and Pollution Research, 18(4), 629-637. Tierney, M., Emmerson, L., & Hindell, M. 2009. Temporal variation in Adélie penguin diet at Bechervaise Island, east Antarctica and its relationship to reproductive performance. Marine Biology, 156(8), 1633-1645. Wania, F., & Mackay, D. 1996. Tracking the distribution of persistent organic pollutants. Environmental Science & Technology, 30(9), A390-A396.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
493
CONCENTRACIÓN DE METALES TRAZA EN EXCRETAS DE DOS ESPECIES DE PINGÜINOS EN DIFERENTES LOCALIDADES DE LA ANTÁRTICA.
(Concentration of trace metals in excrements of two species of penguins from different locations of the Antarctica)
Espejo W1, Celis J1, González-Acuña D 1, Jara S2, Barra R2.
1Facultad de Ciencias Veterinarias, Universidad de Concepción, Casilla 537, Chillán, Chile. [email protected] 2Unidad de Sistemas Acuáticos, Centro de Ciencias Ambientales EULA-Chile, Universidad de Concepción, Concepción, Chile. RESUMEN Cierta evidencia indica que los metales pueden ser transportados a grandes distancias y podrían estar afectando a regiones prístinas como las polares (Smichowski et al., 2006). Ciertamente, la Antártica se está convirtiendo en un espacio donde diversas actividades antropogénicas se están manifestando con más fuerza desde la última década (Lohan et al., 2001). El biomonitoreo podría proporcionar información útil sobre los niveles de los metales en la fauna polar, como los pingüinos, ya que son de amplia distribución, abundantes, de larga vida y ocupan los niveles superiores en la cadena trófica (Boersma, 2008). Los metales traza son tóxicos para los mamíferos y las aves, y pueden causar graves daños a los riñones, el hígado y el sistema nervioso central (Kim et al., 1998; Horai et al., 2007). Estudios recientes han relacionado la contaminación con Cd, Pb, As, Cu y Zn con derrames de combustible, procesos de combustión, disposición de residuos, así como productos químicos desechados a través de la red de alcantarillado (Tin et al., 2009). El cadmio es un elemento biotóxico que está considerado como uno de los metales que provoca serios problemas de contaminación en los ecosistemas marinos (Smichowski et al., 2006). Algunos metales tales como Cu y Zn en bajas dosis son esenciales para las funciones fisiológicas y bioquímicas de los organismos vivos (Tahri et al. 2005). Sin embargo, todos los metales pueden ser potencialmente tóxicos e incluso cancerígenos, dependiendo de los niveles a los cuales son detectados y la naturaleza química del metal (Bengtsson et al. 2006) Un estudio concluyó que los excrementos de animales marinos se pueden utilizar como bioindicadores eficaces para la contaminación por metales y los cambios de calidad ambiental (Yin et al. 2008). Recientemente un estudio mostró que el excremento fresco de pingüino Papúa puede ser utilizado para indicar el nivel de contaminación por metales en la Península Antártica (Celis et al., 2012). Paralelamente, las aves marinas son organismos que actúan como biovectores de contaminantes, transfiriendo contaminantes químicos vía excretas desde el mar hacia tierra (Michelutti et al. 2009). En el presente estudio se recolectaron muestras de excretas en diferentes lugares de la Península Antártica durante la temporada 2011/2012: ocho sitios (Base O'Higgins, Yankee Harbour, Mikkelsen Harbour, Isla Danco, González Videla Base, Base Yelcho y Base Brown) en colonias de pingüinos Papúa, y cuatro lugares (Hydrurga Rocks, Cabo Shirreff, Punta Narębski e Isla Kopaitic) en colonias de pingüinos de Barbijo. Las excretas se recolectaron utilizando guantes de látex y mediante el uso de espátulas de plástico. Cada muestra recogida (5 g) fue de un pool de varios individuos, no siendo posible evaluar la contaminación individualmente, pero sí en un grupo de esa especie. Las muestras se guardaron en bolsas de plástico selladas a -20 º C para su transporte hasta sus determinaciones analíticas. En el laboratorio, las muestras se lavaron con agua destilada (Milli-Q), se secaron a temperatura ambiente y luego se molieron (malla 24 dm-2). Después, las muestras fueron digestadas con ácido nítrico, ácido clorhídrico, y ácido perclórico para Pb, Cu y Zn. en los laboratorios del centro EULA-Chile. Las concentraciones de As se determinaron mediante espectrometría de absorción atómica, horno de grafito para Cd y Pb, mientras que para Cu y Zn se usó espectrofotometría de

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
494
absorción atómica. Los límites de detección fueron 0,01 µg g-1 (peso seco) para todos los elementos estudiados. Triplicados de cada muestra se prepararon de forma independiente. Con el fin de asegurar el control de calidad, se utilizó un material de referencia certificado (cabello humano) como estándar interno. Los niveles detectados se expresan en µg g-1 ± desviación estándar. Las diferencias entre las concentraciones de metales traza se evaluaron mediante el uso de análisis no paramétrico de la varianza Kruskal-Wallis y U de Mann-Whitney y Pruebas post hoc se realizaron para los análisis de Kruskal-Wallis, con las diferencias críticas de rango medio. Coeficientes de corrección de Spearman fueron calculados entre los niveles de los metales. Las diferencias estadísticas significativas fueron estimadas a 0,05. Los análisis estadísticos se realizaron con el programa SPSS versión 15.0. Los resultados en excrementos de P. papua mostraron diferencias geográficas significativas entre los lugares estudiados para los metales traza. Los niveles de Pb no mostraron diferencias significativas entre la Base O'Higgins, Mikkelsen Harbour y la Base González Videla. Los niveles de Pb fueron significativamente mayores en la Base O'Higgins y la Base González Videla en comparación con Bahía Yankee e Isla Danco. Los mayores niveles de Zn fueron detectados en Base O'Higgins (379,99 µg g-1), que difieren estadísticamente de los niveles de Zn encontrados en Bahía Yankee, Isla Danco, Base González Videla, Base Yelcho y Base Brown (que van desde 172,92 hasta 246,95 µg g-1). No hubo diferencias significativas entre los niveles de Zn detectados en Mikkelsen Harbour (324,30 µg g-1) y la Base Yelcho (172,92 mg g-1). Con respecto a los niveles de Cu, las excretas de P. papua no mostraron diferencias significativas entre Base O'Higgins, Base González Videla, Mikkelsen Harbour y Base Yelcho. Hubo significativamente más Cu en las excretas colectadas en Base O'Higgins que en Bahía Yankee e Isla Danco, mientras que el Cu fue significativamente mayor en las excretas de Base González Videla que en Bahía Yankee, Isla Danco, Base Yelcho y Base Brown. Los niveles de As y Cd en las heces de P. papua fueron similares en todos los lugares estudiados. Los niveles de metales en los excrementos de P. antarctica también mostraron diferencias geográficas significativas entre los lugares estudiados. Los mayores niveles (p<0,05) de Cd fueron encontrados en Hydrurga Rocks (3,30 µg g-1) y Punta Narebski (3,13 µg g-1), no encontrándose diferencias significativas entre ambos lugares. Ambos sitios mostraron niveles de Cd significativamente más altos que Isla Kopaitic (1.88 mg g-1) y que Cabo Shirreff (1.89 µg g-1). Los mayores niveles Cu (p<0,05) se encontraron en los excrementos recogidos en Isla Kopaitic (286,69 µg g-1), aunque no difieren significativamente de Punta Narębski (259,99 µg g-1). Ambos lugares mostraron niveles de Cu mayores (p <0,05) que los niveles encontrados en Hydrurga Rocks (168,90 µg g-1). Los niveles de As y Zn no mostraron diferencias significativas entre las localidades estudiadas. Hubo correlaciones positivas significativas entre los metales no esenciales (Cd y Pb) y metales esenciales (Cu y Zn) en los excrementos de ambas especies de pingüinos. DISCUSIÓN Al considerar en su conjunto todos los puntos geográficos de muestreo, se observaron las siguientes relaciones entre los metales y elementos traza en las excretas de los pingüinos Papúa: Zn> Cu> Cd> Pb> As. Esta secuencia es similar a la observada por Metcheva et al. (2011) en excrementos de pingüinos Papúa de Isla Livingston (Shetland del Sur). Para los pingüinos de Barbijo, la relación entre los metales fue la siguiente: Cu> Zn> Cd> Pb> As. El mismo orden descendente entre Cu, Zn y Pb fue reportado por Yin et al. (2008) en excretas de pingüinos de Barbijo en la Isla Rey Jorge. Un menor contenido de Pb en las excretas se explicaría porque el Pb es un elemento que tiende a bioacumularse en los huesos (Teodorova et al. 2003). Otros estudios han demostrado que las concentraciones de Pb son más altas en las plumas que en los excrementos de los pingüinos Adélia (Honda et al., 1986; Jerez et al., 2011). Se observaron correlaciones positivas (p <0,05) entre Pb y Zn, Pb y Cu, y Cu y Zn en los excrementos de pingüino, lo que puede indicar la misma fuente de contaminación. La concentración de metales

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
495
puede diferir entre las colonias de la misma especie que viven lejos unos de otros debido a la dieta y la presencia de contaminantes en aguas de la Antártida (Metcheva et al., 2006). Es interesante destacar que los pingüinos pueden actuar como biovectores al depositar los metales en el suelo vía excrementos. Una investigación realizada por Michelutti et al. (2009) demostró que las concentraciones de Cd fueron hasta 16 veces mayores en los sitios terrestres frecuentados por aves marinas que en aquellos sitios no visitados. Debido a que los pingüinos se alimentan en lo alto de la cadena alimentaria marina, biomagnifican los contaminantes y transfieren así los metales bio-acumulados en los sitios de defecación, presentando concentraciones más allá de las originadas por el transporte a través de procesos abióticos (Blais et al. 2007). Nuestros hallazgos sugieren que las colonias de pingüinos estudiadas pueden transferir metales como Cd, As, Cu, Pb y Zn desde el mar hasta los ecosistemas terrestres vía excrementos, que es una vía importante para el transporte de contaminantes en la Antártida, como también lo notó Nie et al. (2012). En conclusión, los niveles de Pb, Cu y Zn en las heces de los pingüinos Papúa y de Cd y Cu en las excretas de los pingüinos de Barbijo fueron mayores en aquellos lugares con mayor actividad antropogénica. Estos resultados señalan la actividad humana como responsable de la concentración de metales en las excretas de los pingüinos estudiados. En general, nuestros niveles de Pb, As, Cd, Cu y Zn fueron similares o levemente superiores a los metales traza encontrados previamente en los excrementos de pingüinos Papúa y de Barbijo en la Antártica. Los niveles de As, Pb y Cd fueron inferiores a los niveles de metales en excrementos de otros animales marinos que viven en la Antártica, que los niveles en heces de aves que viven en otras regiones del mundo, y que en las excretas de los pingüinos Adélia que habitan Terra Nova, respectivamente. Debido a procesos de bioacumulación, los pingüinos Papúa y de Barbijo, como la mayoría de los organismos que actúan como biovectores, pueden transferir metales desde el mar hacia los ecosistemas terrestres a través de los excrementos, lo cual podría estar afectando negativamente a los organismos vivos que viven en ambientes terrestres de algunas zonas de la Península Antártica y las Islas Shetland del Sur. AGRADECIMIENTOS Al Instituto Antártico Chileno y a los proyectos INACH T_18-09 y T_27-10. REFERENCIAS BIBLIOGRÁFICAS Bengtsson H, Alvenas G, Nilsson S, Hultman B, Oborn I. (2006). Cadmium, copper and zinc leaching and surface run-off losses at the Öjebyn farm in Northern Sweden-temporal and spatial variation. Agric Ecosyst Environ 113: 120-138. Blais J, Macdonald R, Mackey D, Webster E, Harvey C, Smol J (2007) Biologically mediated transport of contaminants to aquatic systems. Environ Sci Technol 41: 1075-1084. Boersma PD (2008) Penguins as marine sentinels. Bioscience 58: 597-607. Celis J, Jara S, González-Acuña D, Barra R, Espejo W (2012) A preliminary study of trace metals and porphyrins in excreta of Gentoo penguins (Pygoscelis papua) at two locations of the Antarctic Peninsula. Arch Med Vet 44: 311-316. Honda K, Yamamoto Y, Hidaka H, Tatsukawa R (1986) Heavy metal accumulations in Adélie penguin, Pygoscelis adeliae, and their variations with the reproductive processes. Mem Natl Inst Polar Res 40: 443-453. Horai S, Watanabe I, Takada H, Iwamizu Y, Hayashi T, Tanabe S, Kuno K (2007) Trace element accumulations in 13 avian species collected from the Kanto area, Japan. Sci Total Environ 373: 512–525. Jerez S, Motas M, Palacios M, Valera F, Cuervo J, Barbosa A (2011) Concentration of trace elements in feathers of three Antarctic penguins: Geographical and interspecific differences. Environ Pollut 159: 2412-2419. Kim EY, Goto R, Tanabe S, Tanaka H, Tatsukawa R (1998) Distribution of 14 elements in tissues and organs of Oceanic seabirds. Arch Environ Contam Toxicol 35: 638–645. Lohan MC, Statham PJ, Peck L (2001) Trace metals in the Antarctic soft-shelled clam Laternula elliptica: implications for metal pollution from Antarctic research Bases. Polar Biol 24: 808-817. Metcheva R, Yurukova L, Teodorova S (2011) Biogenic and toxic elements in feathers, eggs, and excreta of Gentoo penguin (Pygoscelis papua ellsworthii) in the Antarctic. Environ Monit Assess 182: 571–585. Metcheva R, Yurukova L, Teodorova S, Nikolova E (2006) The penguin feathers as bioindicator of Antarctica environmental state. Sci Total Environ 362: 259–265.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
496
Michelutti N, Keatley B, Brimble S, Blais, J, Liu H, Douglas M, Mallory M, Macdonald R, Smol J (2009) Seabird-driven shifts in Arctic pond ecosystems Proc. R. Soc. B 276: 591–596. Nie Y, Liu X, Sun L, Emslie SD (2012) Effect of penguin and seal excrement on mercury distribution in sediments from the Ross Sea region, East Antarctica. Sci Total Environ 433: 132-140. Smichowski P, Vodopivez C, Muñoz-Olivas R, Gutierrez A (2006) Monitoring trace elements in selected organs of Antarctic penguin (Pygoscelis adeliae) by plasma-based techniques. Microchem J 82: 1-7. Tahri M, Benyaïch F, Bounakhla M, Bilal E, Gruffat J, Moutte J, García D. (2005). Multivariate analysis of heavy metal contents in soils. Sediments and water in the region of Meknes (central Morocco). Environ Monit Assess 102: 405-417. Teodorova S, Metcheva R, Topashka-Ancheva M (2003). Bioaccumulation and damaging action of polymetal industrial dust on laboratory mice Musmusculus alba I. Analysis of Zn, Cu, Pb and Cd disposition and mathematical model for Zn and Cd bioaccumulations. Environ Res 91: 85–94. Tin T, Fleming ZL, Hughes KA, Ainley DG, Convey P, Moreno CA, Pfeiffer S, Scott J, Snape I (2009) Impacts of local human activities on the Antarctic environment. Antarct Sci 21: 3-33. Yin X, Xia L, Sun L, Luo H, Wang Y (2008) Animal excrement: A potential biomonitor of heavy metal contamination in the marine environment. Sci Total Environ 399: 179-185.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
497
CARACTERIZACIÓN QUÍMICA Y BIOLÓGICA DE SUELOS CONTAMINADOS CON DIESEL PROVENIENTES DE LA ISLA REY JORGE, ANTÁRTICA
(Chemical and biological characterization of diesel contaminated soils obtained from Isla Rey
Jorge, Antártica)
Gran-Scheuch, A.12, Órdenes-Aenishanslins, N.12, Monrás, J.P.13, Coppottelli, B.4, Pérez-Donoso, J.M.1
1Laboratorio de Bionanotecnología y Microbiología, Centro de Bioinformática y Biología Integrativa (CBIB), Universidad Andrés Bello, República 237, Santiago, Chile [email protected] 2Facultad de Ciencias Químicas y Farmacéuticas, Universidad de Chile, Sergio Livingstone Pohlhammer 1007, Santiago, Chile 3Facultad de Química y Biología, Universidad de Santiago de Chile, Alameda 3363, Santiago, Chile 4Coppotelli, Centro de Investigación y Desarrollo en Fermentaciones Industriales (CINDEFI), Universidad Nacional de La Plata, Provincia de La Plata, Argentina. INTRODUCCIÓN La Antártica es el continente más austral de la tierra, presenta condiciones extremas, principalmente bajas temperaturas y alta radiación ultravioleta. El hombre ha sido capaz de sobrevivir en la región y realizar actividades científicas y de explotación durante más de 80 años. Como consecuencia, las distintas zonas habitadas han sufrido contaminación, principalmente asociada a diesel. Este compuesto es almacenado muchas veces en contenedores en mal estado, provocando filtraciones y la consecuente liberación de diesel y metales pesados al suelo Antártico (provenientes del petróleo y de los mismos contenedores). El diesel es nocivo ya que presenta una gran cantidad de moléculas orgánicas como los hidrocarburos policíclicos aromáticos (HPAs), que son compuestos mutágenos, cancerígenos y teratógenos. Además, presenta compuestos inorgánicos como metales pesados (Cd, Cr y Pb) que también han sido asociados con cáncer (Boffetta, 2007; Adeniyi, 2010). La descontaminación de HPAs se puede llevar a cabo mediante procesos físicos, químicos y biológicos. Los procesos biológicos representan una opción ecológica, económica y segura, que involucra una menor alteración de los suelos, constituyendo una excelente alternativa para la descontaminación del territorio Antártico por su carácter ecofriendly. A la fecha, se ha reportado un gran número de bacterias capaces de degradar HPAs, sin embargo debido al Tratado Antártico no está permitido ingresar microorganismos o ADN foráneo al continente, por lo que es vital encontrar bacterias nativas que puedan ser usadas en la biorremediación de diesel. Por su mayor solubilidad, menor toxicidad para el humano y similitud estructural con otros HPAs, el compuesto que se utiliza como modelo para aislar y estudiar microorganismos degradadores de HPAs es el fenantreno (Wick, 2011). Sin embargo, no basta sólo una adecuada selección de la fuente de carbono para obtener una cepa con potencial biotecnológico, es de vital importancia el contexto ambiental en donde las cepas son aisladas. Los microorganismos usados en biorremediación normalmente se desempeñan de mejor manera en condiciones similares a las de su sitio de origen, debido a variables como la temperatura, pH, nutrientes, biodiversidad y presencia de tóxicos como metales pesados. De este modo, la caracterización química y biológica de los suelos contaminados con petróleo, es un paso fundamental que permitirá determinar las condiciones que favorecen la metabolización de contaminantes in situ, fenómeno que en el caso de Antártica resulta de especial interés por las características especiales del continente y debido a lo poco estudiado de la contaminación de estas zonas (Bamforth, 2005). Se plantea que la contaminación antropogénica con petróleo y metales en la Antártica ha favorecido el desarrollo de bacterias degradadoras de fenantreno resistentes a metales. El objetivo general de este trabajo es caracterizar química y biológicamente suelos expuestos a diésel con fines aplicados en la biorremediación.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
498
MATERIALES Y MÉTODOS Muestreo Las muestras de suelo fueron aisladas desde la Isla Rey Jorge en la Antártica durante una expedición, por nuestro grupo, durante Febrero del 2013. Determinación de compuestos apolares La extracción de compuestos apolares se realizó según el protocolo de Ikhtiar ud Din y colaboradores, en laboratorios institucionales de la Universidad Andrés Bello. La muestra de suelo (1 gramo) se mezcló con 5 ml de diclorometano. La mezcla se centrifugó y se separó el sobrenadante. El extracto se cargó en una columna con sílica humedecida con n-hexano. El extracto se eluyó con una mezcla n-hexano/diclorometano (1:1). El eluído se evapora completamente y el residuo sólido es resuspendido en acetonitrilo. La determinación cuantitativa de fenantreno y los cromatogramas de los perfiles de suelo son realizados inyectando 20 µL de extracto en un equipo HPLC con detector UV (KONIK) equipado con una columna RP-C18 y usando una gradiente acetonitrilo: agua (60:40, v/v) como fase móvil a un flujo de 1.25 ml/min. El fenantreno es determinado por absorbancia a 254 nm (ud Din, 2013). Aislamiento de bacterias A las muestras de suelo se les agregó agua destilada y fenantreno a una concentración final de 2.000 ppm. Diferentes diluciones de sobrenadante tras 5 días de incubación fueron sembradas en medio R2A-agar. Curva de Crecimiento Las cepas aisladas se inocularon en medio mínimo con fenantreno 2.000 ppm, como única fuente de carbono, en constante agitación. La acumulación de productos parciales de la oxidación de fenantreno lleva a la coloración del medio en tonos ocres y permite así evidenciar cualitativamente la degradación de dicho HAP. Se tomaron alícuotas a distintos tiempos y se midió absorbancia a 600 nm de longitud de onda. RESULTADOS Extracción de compuestos apolares Para la determinación de fenantreno mediante HPLC-UV se realizó una curva de calibración de fenantreno en acetonitrilo. Además se analizó muestras de suelo Antártico con y sin petróleo. Se detectó fenantreno en los suelos contaminados. En las muestras de suelo contaminado se diferencian mayor cantidad de picos en el cromatograma en comparación a las tierras vírgenes (Fig. 1).
Figura 1.Medición de extracto orgánico de muestras de suelos Antártico a 254 nm. Cromatograma de muestras de tierras contaminadas (1 y 2) y a muestra de suelo sin contaminar (Suelo sin exposición). Tiempo de retención de fenantreno aproximado de 6,8 min. Figure 1. Measuring organic extract Antarctic soil samples at 254 nm. Chromatogram of contaminated and uncontaminated soil samples (1 and 2, respectively). Phenanthrene retention time of approximately 6.8 min.
Curva de crecimiento Tras sembrar diluciones de sobrenadante de suelo con fenantreno 2000 ppm se aislaron 22 cepas en medio R2A-agar. Estas cepas fueron capaces de metabolizar fenantreno, evidenciándose con el cambio de coloración del medio mínimo con el hidrocarburo como única fuente de carbono (transparente a ocre). Junto con esto, se evidenció un aumento de la densidad óptica (OD600) en el tiempo en este

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
499
mismo medio (Fig. 2).
Figura 2. Curva de crecimiento a 600 nm. Curva de 3 cepas aisladas desde suelo contaminado con diesel en medio de cultivo con
fenantreno como única fuente de carbono. Figure 2. Growth curve at 600 nm. Three growth curves of isolated strains from soil contaminated with diesel in phenanthrene culture
medium as a sole carbon source.
DISCUSIÓN Y CONCLUSIONES Las determinaciones por cromatografía líquida de alta resolución indican 2 conclusiones importantes; la primera es que en suelos contaminados existe mayor presencia de componentes apolares que en suelos sin exponer a diésel, representado en la intensidad de los picos de absorbancia en los cromatogramas (Fig. 1). En segundo lugar, al tiempo de retención del estándar de fenantreno aparecen en los cromatogramas de suelos contaminados señales intensas, es decir, efectivamente existe contaminación con fenantreno y otras moléculas apolares en estos suelos. La curva de crecimiento de cepas aisladas (Fig. 2) indica que a los 5 días las bacterias se encuentran en fase estacionaria, evidenciándose que las cepas son capaces de crecer con fenantreno como fuente de carbono y energía. El cambio de coloración del medio de cultivo ocurrió alrededor del tercer día (de transparente a ocre). Este es un método sencillo para determinar cepas capaces de degradar fenantreno, permitiendo realizar un screening con una gran cantidad de potenciales bacterias degradadoras de HPAs. Encontramos una gran número de cepas capaces de metabolizar fenantreno, lo que abre las puertas a seguir estudiándolas para evaluar la potencialidad de las cepas para ser usadas con fines biotecnológicos en la biorremediación de suelos antárticos. Estas cepas capaces de crecer en fenantreno serán estudiadas en mayor profundidad en estudios de cinética de degradación y caracterización microbiológica. Las bacterias serán identificadas mediante secuenciación del gen 16S rDNA. La caracterización química de suelos Antárticos es vital para un potencial proceso de biorremediación. La concentración de fenantreno y de metales pesados da un indicio de la presencia de Hidrocarburos Policíclicos Aromáticos y la toxicidad del suelo, pudiendo establecer una relación directa entre la contaminación de hidrocarburos y metales con la presencia de diesel. Consecuentemente se puede generar prioridad en un posible tratamiento de suelo. Al presente, nuestro grupo está cuantificando la cantidad de metales disponibles y totales de cadmio, cromo y plomo de muestras de suelo antártico por espectroscopía de absorción atómica de llama, debido a que la presencia o ausencia de metales pesados es información trascendental si se va a tratar un suelo en particular, ya que los metales pesados son tóxicos para bacterias, por lo que hay que tratar con cepas resistentes a estos compuestos inorgánicos. Para esto, se debe conocer la concentración en el suelo y relacionarlo según las características de cada bacteria degradadora de fenantreno. Este trabajo es un primer estudio de nuestro grupo para investigar una alternativa de descontaminación de ecosistemas antárticos y a profundizar el conocimiento sobre el efecto de la contaminación antropogénica sobre la biodiversidad de nuestro planeta, específicamente en el continente Antártico. En este punto cabe resaltar cómo las bacterias han sido capaces de adaptarse al medio antártico, encontrándose una abundancia y diversidad de organismos antárticos capaces de tolerar condiciones extremas y los efectos del hombre.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
500
AGRADECIMIENTOS INACH T_19-11, FONDECYT 11110077, ANILLO ACT 1107 y Proyecto de Intercambio del Programa de Cooperación Internacional CONICYT-MINCYT (Argentina), “Estudio de los procesos de Biorremediación de suelos contaminados con metales pesados e hidrocarburos policíclicos aromáticos (PAH)”, 2011-2013 REFERENCIAS Adeniyi, A. A., & Owoade, O. J. (2010). Total petroleum hydrocarbons and trace heavy metals in roadside soils along the Lagos–Badagry expressway, Nigeria. Environmental Monitoring and Assessment, 167(1-4), 625-630. Bamforth, S. M., & Singleton, I. (2005). Bioremediation of polycyclic aromatic hydrocarbons: current knowledge and future directions. Journal of Chemical Technology and Biotechnology, 80(7), 723-736. Boffetta, P., Jourenkova, N., & Gustavsson, P. (1997). Cancer risk from occupational and environmental exposure to polycyclic aromatic hydrocarbons. Cancer Causes & Control, 8(3), 444-472. ud Din, I., Rashid, A., Mahmood, T., & Khalid, A. (2013). Effect of land use activities on PAH contamination in urban soils of Rawalpindi and Islamabad, Pakistan. Environmental Monitoring and Assessment, 1-10. Wick, A. F., Haus, N. W., Sukkariyah, B. F., Haering, K. C., & Daniels, W. L. (2011). Remediation of PAHcontaminated soils and sediments: A literature review. Virginia Polytechnic Institute and State University.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
501
ESTUDIO INMUNO-HEMATOLÓGICO DE EFECTOS ATRIBUIBLES A CONTAMINANTES ORGÁNICOS PERSISTENTES EN PINGUINOS Pygoscelis DE LA
PENÍNSULA ANTÁRTICA
(Immunohaematological study on effects atributable to persistant organic pollutants in Pygoscelis penguins in Antarctic Peninsula)
Mattatal, P.1,2, Jara, S. 1, González, M.2, Barra, R.1
1Laboratorio de Biomarcadores, Centro de Ciencias Ambientales EULA, Universidad de Concepción. 2Departamento de Bioquímica Clínica e Inmunología, Facultad de Farmacia, Universidad de Concepción. [email protected] Los contaminantes orgánicos persistentes (COPs) tienen efectos nocivos a corto y largo plazo sobre la biota, que son incrementados por los procesos de bioacumulación y biomagnificación, siendo los principales afectados los organismos con elevada posición en las tramas tróficas (Van der Oost et al, 2003; Goerke et al, 2004). Estudios en contaminantes indican alteraciones en el sistema inmune, disfunción de la tiroides, alteraciones hormonales, tumores, problemas reproductivos; feminización de machos y masculinización de hembras, alteración en el tamaño de las gónadas, disminución en el número de crías, malformaciones fetales si es que las hembras han estado expuestas durante periodo de gestación, etc. (Fernández et al, 2005; Lopez 2005; AMAP, 2009). El destino y los efectos de los COPs en la biota de zonas con escasa intervención antrópica como la Península Antártica son motivo de gran preocupación por el valor de biodiversidad de este ecosistema. Dentro de esta biota, los pingüinos representan más del 70% de la fauna (en biomasa) y están cercanos al tope de la trama trófica acuática. Los pingüinos poseen, además, en sus compartimentos internos un mayor contenido lipídico (particularmente tejido graso) y al metabolizar este contenido, se libera simultáneamente toda la carga de contaminantes bioacumulada en dichos depósitos (Rajchard, 2010). El objetivo de este estudio fue evaluar alteraciones hematológicas e inmunológicas atribuibles a la exposición a COPs en pingüinos del género Pygocelis (P. papua, P. adeliae, P. antárctica) de la Península Antártica. Frotis sanguíneos teñidos con May-Grünwald Giemsa fueron analizados en microscopio Zeiss. Se contó un total de 200 leucocitos para recuento diferencial leucocitario y se determinó el índice de abundancia relativa de heterófilos y linfocitos (H:L). Para el recuento de eritrocitos inmaduros se consideró un total de 400 eritrocitos. Para cuantificación de IgY, primero se obtuvo un estándar de IgY de pollo a partir de yema de huevo mediante purificación por salting out (Pinto et al., 2005). Se cuantificó las proteínas del estándar por método de Bradford y se comprobó el éxito de la purificación realizando una electroforesis en gel de poliacrilamida no denaturante. La cuantificación se realizó por método ELISA (directo) utilizando un anticuerpo comercial contra IgY de pollo desarrollado en conejo por la empresa Sigma-Aldrich y utilizando como bloqueante leche descremada en PBS. La purificación fue exitosa, lo que se demostró con la existencia de una única banda en el gel de poliacrilamida, alrededor de los 175 kDa. Se observó diferencias significativas entre especies y en relación a los datos existentes de 1987 (ver figura 1), que incluyen alteraciones celulares, células con granulaciones citotóxicas, linfocitos con núcleo segmentado, linfocitos granulados y en diferentes estados de activación, poiquilocitosis, anisocitosis, y monocitos vacuolados. Aunque el número de eritrocitos inmaduros estuvo en rango fisiológico para las tres especies, se observó grados de hipocromía. La relación H:L resultó sin diferencias significativas entre especies; P. antárctica con 2,37±0,20; P. adeliae con 2,71±0,59 y P. papua con 3,28±0,41, si bien los tres valores indican estrés celular.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
502
Fig 1. Recuento diferencial leucocitario por especie en frotis de pingüino Pygoscelis. A. Porcentaje de basófilos. B. Porcentaje de eosinófilos. C. Porcentaje de heterófilos D. Porcentaje de linfocitos. E. Porcentaje de monocitos. papua: P.
papua; adelia: P. adeliae; antarctica: P. antarctica Fig 1. Differential leucocyte count by specie in Pygoscelis penguin smears. A. Basophil percentage. B. Eosinophil
percentage. C. Heterophil percentage. D. Lymphocyte percentage. E. Monocyte percentage. papua: P. papua; adelia: P. adeliae; antarctica: P. antarctica. (*) P<0,05 (**)P<0,001
De la cuantificación de IgY se obtuvo los valores 194,5±35,00; 95,97±12,94 y 109,9±17,04 ug/ml para P. papua, P. adelia y P. antarctica respectivamente, donde P. papua dobla en cantidad a las otras
B
E
A
Heterófilos
papua
adelia
antarctica
0
20
40
60
80
Especie
Tota
l de c
élu
las
(%
)
Linfocitos
papua
adelia
antarctica
0
10
20
30
Especie
Tot
al d
e cé
lula
s (
%)
Monocitos
papua
adelia
antarctica
0
2
4
6
Especie
Total de células
(%)
C D
Basófilos
papua
adelia
antarctica
0
1
2
3
4
5Total de células
(%)
*
*
Especie
Eosinófilo
papua
adelia
antarctica
0
5
10
15
**
Especie
Total de células
(%)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
503
especies. Por otra parte, las tres especies de pingüino fueron encontradas bajo lo documentado para aves, donde el valor establecido es entre 6 y 13 mg/ml. No se encontró relaciones significativas entre niveles de contaminantes comparados y los valores obtenidos de frotis o IgY. Las variaciones en la alimentación, en desplazamiento, y en definitiva, en la exposición a agentes contaminantes pueden ser responsables de las alteraciones en cada especie y aunque la etiología de las alteraciones no es clara, se infiere que la condición ambiental alterada en los últimos años afecta los procesos celulares en pingüinos. AGRADECIMIENTOS Proyecto INACH T 18-09, laboratorio de biomarcadores y departamento de Bqca. Clin. E Inm, Universidad de Concepción. BIBLIOGRAFÍA AMAP Assessment 2009: Human Health in the Arctic. Arctic Monitoring and Assessment Programme (AMAP), Oslo, Norway. Xiv. 256 Pp. Fernández A, Yarto M & Castro J 2005 Las sustancias tóxicas persistentes en México. Secretaría de Medio Ambiente y Recursos Naturales Instituto Nacional de Ecología. Instituto Nacional de Ecología. México. 266. Goerke H, Weber K, Bornemann H, Ramdohr S & Plotz J. 2004 Increasing levels and biomagnification of persistent organic pollutants (pops) in Antarctic biota. Marine pollution Bulletin (48): 295–302. López O. 2005 Influencia de la exposición crónica a plaguicidas sobre diversos marcadores bioquímicos (esterasas y enzimas antioxidantes) en trabajadores de invernadero de la costa oriental de Andalucía. Editorial de la Universidad de Granada Pinto, J., Barco, M, Afanador, M., Merchán, A., Montañez, M., Andrade, F., Torres, O.2005 Obtención de anticuerpos policlonales igy antiparvovires canino a partir de yema de huevo de gallina. Universitas Scientiarum. 10 (1):37-44. Rajchard J. 2010 Biologically active substances of bird skin: a review. Veterinarni Medicina, 55, (9): 413–421 Van Der Oost R, Beyer J., Vermeulen N. 2003 Fish bioaccumulation and biomarkers in environmental risk assessment: a review. Environmental Toxicology and Pharmacology (13): 57-149

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
504
MERCURY CONCENTRATION IN FEATHERS OF PHALACROCORAX ATRICEPS FROM KING GEORGE ISLAND, SOUTH SHETLAND ARCHIPELAGO, ANTARCTICA
(Concentración de mercurio en plumas de Phalacrocorax atriceps de isla Rey Jorge, archipiélago Shetland del Sur, Antártica)
Pereira, L.E.1, Costa, E.S.*1, 2, Pessôa, A.R.L.1, Souza, J.S.1,2, Alves, M.A.S.2, Torres, J.P.1 & Malm,
O.1
1Laboratório de Radioisótopos Eduardo Penna Franca, Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, RJ. Brazil. CEP: 21941-900. [email protected] 2Laboratório de Ecologia de Aves e Comportamental, Departamento de Ecologia, Instituto de Biologia Roberto Alcantara Gomes, Universidade do Estado do Rio de Janeiro, RJ. Brazil. CEP 20550-011. ABSTRACT Antarctica is one of the most isolated regions, and is considered one of the most pristine on the planet. Mercury (Hg) is a trace metal with high dispersive capability in the environment and with a high level of toxicity. Seabirds such as Blue-Eyed-Shag (Phalacrocorax atriceps) are top predators in the food chain, and they are exposed to mercury bioaccumulation through diet. Herewith we present preliminary results on Hg levels in feathers of P. atriceps (Admiralty Bay, King George Island, South Shetland Archipelago). The feathers were collected during summer of 2011/2012 at the breeding colony of P. atriceps in Shag Island in Admiralty Bay. The samples (n=10), around 1 g were analyzed for their Hg content using cold vapor atomic absorption technique according the method proposed by Bastos (1997). The recovery for the certified samples was between 90 and 110%. Our initial results show that the average value of Hg for the population of P. atriceps studied was 4.95 mg/kg (s.d. ± 2.40). The minimum and maximum values were 1.99 and 8.08 mg/g, respectively. Mercury concentrations varied significantly (KW=9.27, p<0.05, n=10) and were higher in three samples. This variation may be due to differences in age and sex of the birds, or differences in feeding, since individuals who consume more fish than krill may have higher levels of Hg bioaccumulation. Saeki et al. (2000) determined the concentrations of Hg in feathers of P. carbo in two places in Japan: Tokyo (1.9 ± 0.6 mg/kg, n=9) and Laka Biwa (2.9 ± 0.8 mg/kg, n=10). The lowest concentrations for the species of Japan may be associated with several factors, such as type of feather used, but this information was not mentioned by the authors. Our study used the wing feathers easily found in breeding areas. Other factors that may influence these values are the physiology of different species, sex and age, differences in dietary habits and migration. There were no publications evaluating the levels of Hg contamination in feathers of P. atriceps. Thus, further analysis is needed, including different types of feathers, in order to achieve a better understanding on the dynamics of Hg contamination in individuals and in populations of P. atriceps in sub-Antarctic islands. ACKNOWLEDGEMENTS CNPq: 557049/2009-1; 48.4002/2011-2, PROANTAR, FAPERJ E-26/111.505/2010. REFERENCIAS W.R. Bastos; Malm, O.; Pfeiffer, W.C. y Cleary, D. 1998. Establishment and analytical quality control of laboratories for Hg determination in biological and geological samples in the Amazon, Brazil. Ciência e Cultura, 50: 255-260. K. Saeki; Okabe, Y.; Kim, E.-Y.; Tanabe, S.; Fukuda, M. y Tatsukawa, R. 2000. Mercury and cadmium in common cormorants (Phalacrocorax carbo). Environmental Pollution, 108: 249-255.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
505
PLOMO EN SUELOS DE COLONIAS DE PINGÜINOS PYGOSCELIDOS EN LA PENINSULA ARDLEY, ANTARTICA
(Lead in colonies of Pygoscelid penguins in Ardley Peninsula, Antarctica)
Perfetti-Bolaño, A.1 y Moreno, L.1 1Departamento de zoología, Universidad de Concepción, Concepción, Barrio Universitario s/n, Casilla 160-C, Concepción, Chile. E-mail: [email protected] INTRODUCCIÓN El plomo se encuentra presente en la corteza terrestre en forma natural y se produce primariamente por fundición del mineral (Sepúlveda et al., 2000). Es un metal pesado que posee la capacidad de bioacumularse en las cadenas tróficas de plantas y animales, por lo tanto, su concentración se magnifica a lo largo de la cadena alimentaria, en la cual no tiene un papel esencial conocido (Halliwell et al., 2000; Sun & Xie, 2001). Varios estudios han reportado la presencia de metales pesados en la Antártica, entre ellos el plomo, el que ha sido detectado en el suelo, nieve, agua y en tejidos y heces de diferentes especies de aves marinas (Lebedeva, 1997; Sun & Xie, 2001; Celis et al., 2012). Las aves marinas son consideradas bioindicadores de la contaminación ambiental, ya que pueden acumular las formas biológicamente disponibles de los contaminantes, pudiendo biomagnificarlos, debido a que se encuentran en una posición alta en las cadenas tróficas (Burger & Gochfeld, 2004). Además, se caracterizan por formar colonias en la época reproductiva, produciendo a lo largo de varios años un aporte importante al suelo de heces y otros restos orgánicos (Otero & Mouriño, 2002), lo que produciría un incremento del contenido total de metales en el suelo (García et al. 2007). Los pingüinos del género Pygoscelis (Pygoscelis adeliae, P. antarctica, P. papua), presentan poblaciones con distribución exclusiva en mares antárticos y sub-antárticos (Martínez & González, 2004), nidifican en colonias, utilizando las mismas áreas geográficas, pero con la existencia de segregación ecológica entre ellas para la nidificación (Forcada, 2007). Estudios recientes han identificado la presencia de plomo y otros metales pesados en plumas, contenido estomacal y heces en estas especies (Ancora et al., 2002; Celis et al., 2012). Del mismo modo, estudios realizados en sedimentos de un lago que contenía excrementos de pingüinos en la Antártica, reportaron un aumento significativo en las concentraciones de plomo durante los últimos 200 años y especialmente durante los últimos 50 años, coincidiendo con la revolución industrial. Esto sugiere que la contaminación del medio ambiente mundial ha afectado el ecosistema antártico (Sun & Xie, 2001). El objetivo principal de este trabajo es evaluar la influencia de las aves en la acumulación de metales pesados en suelos antárticos. Para esto se determinará la concentración de plomo en colonias de pingüinos de las especies P. adeliae, P. antarctica y P. papua en la Península Ardley, la que será comparada con las concentraciones de plomo encontrada en suelos donde transitan los pingüinos y zonas sin pingüinos. MATERIALES Y MÉTODOS El muestreo se realizó en diciembre de 2012 en la Península Ardley, Zona Antártica Especialmente Protegida N° 150, en la cual existe una población estimada de 4.978 pingüinos pygoscelidos, siendo la especie dominante P. papua con 4.635 individuos (Secretaría del Tratado Antártico, 2009). La zona se clasificó en tres sitios de muestreo, los cuales corresponden a lugares sin pingüinos, lugares de tránsito de pingüinos y en colonias de pingüinos de las especies P. adeliae, P. antarctica y P. papua. Se utilizó el muestreo aleatorio estratificado descrito por Mason (1992), identificando en cada zona de muestreo 3 sitios al azar en los que se extrajeron 3 muestras, hasta obtener un total de 27 muestras.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
506
Cada muestra de sedimento de suelo se extrajo a una profundidad de 20 cm, utilizando una cucharilla de plástico, hasta obtener 30 gramos, los cuales se almacenaron en una bolsa plástica individual rotulada. La muestra se secó a 21ºC en una estufa de secado. Luego se realizó la digestión de las 27 muestras de suelo siguiendo el protocolo EPA 3050-B. La lectura de las muestras se realizó en un espectrofotómetro de absorción atómica-llama. Los datos de concentración de plomo fueron transformados a logaritmo natural y se evaluó el supuesto de normalidad de los datos mediante una prueba de bondad de ajuste de Shapiro Wilk (W= 0,96; p=0,51). Se realizó un análisis de varianza (ANOVA) para evaluar la diferencia entre las zonas de estudio. RESULTADOS Las concentraciones de plomo de las 27 muestras recolectadas en suelos de zonas de paso, zonas de ausencia de pingüinos y colonias de pingüinos se muestran en la Tabla 1.
Tabla 1. Concentraciones de plomo (mg/kg) por zonas. Table 1. Lead concentrations (mg/kg) zones.
ZONA CONCENTRACIÓN DE Pb (mg/kg)
Media SD MIN MAX A 3,63 1,11 1,82 5,02 B 5,92 2,07 3,57 9,54 C 4,45 1,44 2,80 6,75
A: Zona de ausencia de pingüinos, B: zona de paso y C: colonia de pingüinos. SD: Desviación estándar, MIN: mínimo, MAX: máximo.
La concentración de plomo en suelos de zona de paso fue significativamente mayor (p=0,0053) que la concentración de plomo en la zona de ausencia de pingüinos. Contrario a lo esperado, la concentración de plomo en suelos de colonias de pingüinos no se diferenció significativamente de las zonas de paso y ausencia de pingüinos (p>0,05; Figura 1).
0,5
1
1,5
2
2,5
Ln c
once
ntra
cion
(mg/
kg)
A B C
Zona
Each Pair
Student's t
0,05 Figura 1. Análisis de varianza de una vía de Ln de concentración (mg/kg) de plomo por zona. A: zona de ausencia de
pingüinos, B: zona de paso y C: colonia de pingüinos. La línea media de los diamantes indican la media y las líneas en los extremos los intervalos de confianza (95%).
Figure 1. One-way analysis Ln concentration (mg/kg) per zone. A: absence of penguin area, B: transit area and C: penguin colony.
DISCUSIÓN Y CONCLUSIONES El kril, alimento básico en la cadena trófica antártica, posee concentraciones de plomo de 1,16 mg/kg (Barbante et al. 2000), las cuales se relacionan con las concentraciones detectadas en plumas de pingüino papúa (1,6 mg/kg) y pingüino barbijo (1,73 mg/kg) en Isla Livingston (Metcheva et al. 2005),

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
507
y pingüino Adelia en Terra Nova (>1,0 mg/kg) (Ancora et al. 2002). En esta última especie también se han encontrado concentraciones similares en músculos (0,1 mg/kg) e hígado (2,0 mg/kg) en Isla Rey Jorge (Smichowski et al. 2005). Estos valores son inferiores a los detectados en este trabajo en suelos ocupados por pingüinos (5,19 mg/kg), los cuales coinciden con aquellos encontrados en suelos de la misma zona (5,68 mg/kg) por Tapia (2011). En condiciones de precipitaciones leves, los excrementos percolan el suelo de colonias, los cuales poseen una mayor eficiencia en el filtrado de plomo debido al alto contenido de arcilla y minerales, éstos últimos derivados de la actividad microbiana, reduciendo así los riesgos de lixiviación y la contaminación de aguas subterráneas (Mendonça et al. 2013; Myrcha & Tatur, 1991) y cuando las precipitaciones son fuertes, los excrementos son desplazados de las colonias pendiente abajo, hasta suelos de zona de paso de pingüinos (media: 5,92 mg/kg). Lo anterior explica la menor concentración de plomo (media: 3,63mg/kg) en suelos de ausencia de pingüinos. Las concentraciones de plomo en suelos utilizados por pingüinos evidencian procesos de bioacumulación, los cuales contribuyen a la polución del ambiente. AGRADECIMIENTOS Al Instituto Antártico Chileno y Correos de Chile, por su apoyo en la realización de este proyecto. REFERENCIAS BIBLIOGRÁFICAS Ancora, S., V. Volpi, S. Olmastroni, S. Focardi & C. Leonzio. 2002. Assumption and elimination of trace elements in Adélie penguins from Antarctica: a preliminary study. Marine Environmental Research, 54: 341-344. Barbante, C., G. Cozzi, G. Capodaglio & P. Cescon. 2000. Trace element determination in a candidate reference material (Antarctic Krill) by ICP-sector field MS. Journal of Analytical Atomic Spectrometry, 15: 377-382. Burger, J. & M. Gochfeld. 2004. Marine birds as sentinels of environmental pollution. EcoHealth, 1: 263-274. Celis, J., S. Jara, D. González-Acuña, R. Barra & W. Espejo. 2012. A preliminary study of trace metals and porphyrins in excreta of Gentoo penguins (Pygoscelis papua) at two locations of the Antarctic Peninsula. Archivos de Medicina Veterinaria, 44: 311-316. Forcada, J. 2007. El cambio climático y sus repercusiones para la megafauna antártica. In: Duarte, C. & Fundación BBVA (eds.). Impactos del calentamiento global sobre los ecosistemas polares. Bilbao, pp. 83-108. García, L.V., E. Gutiérrez, T. Marañón, R. Redondo & L. Clemente. 2007. Acumulación de metales pesados en el suelo mediada por aves marinas: evidencias derivadas del análisis isotópico. Libro de Actas del III Simposio sobre control de la degradación de suelos y la desertificación, pp. 255-256. Halliwell, D., N. Turoczy & F. Stagnitti. 2000. Lead Concentrations in Eucalyptus sp. in a Small Coastal Town. Bulletin of Environmental Contamination and Toxicology, 65: 583-590. Lebedeva, N.V. 1997. Accumulation of Heavy Metals by Birds in the Southwest of Russia. Russian Journal of Ecology, 28(1): 41–46. Martínez, D. & G. González. 2004. Las aves de Chile, nueva guía de campo. Ediciones del Naturalista, Santiago, Chile. 620 pp. Mason, B. 1992. Preparation of soil sampling protocols: sampling techniques and strategies. Environmental Protection Agency, Washington, 169 pp. Mendonça, T., V. Melo, L. Alleoni, C. Schaefer & R. Michel. 2013. Lead adsorption in the clay fraction of two soil profiles from Fildes Peninsula, King George Island. Antarctic Science, 25(3): 389-396. Metcheva, R., L. Yurukova, S. Teodorova & E. Nikolova. 2005. The penguin feathers as bioindicator of Antarctica environmental state. Science of the Total Environment, 362: 259-265. Myrcha, A. & A. Tatur. 1991. Ecological role of the current and abandoned penguin rookeries in the land environment of the maritime Antarctic. Polish polar research, 12 (1): 3-24. Otero, X. L. & J. Mouriño. 2002. Nitrógeno (NH4
+, NO3-), fósforo asimilable y metales traza (Hg, Cd, Pb, Zn, Ni y Cu) en
suelos de la colonia de la gaviota patiamarilla (Larus cachinnans) en el parque natural de las islas Cíes. Actas de la I reunión sistemas agroforestales - I reunión espacios naturales, en Cuadernos de la sociedad española de ciencias forestales, 14: 143-149. Secretaría del Tratado Antártico. 2009. Informe final de la trigésima segunda reunión Consultiva del tratado antártico. Plan de gestión revisado de la Zona Antártica Especialmente Protegida N°150 Isla Ardley, bahía Maxwell, isla Rey Jorge (isla 25 de Mayo), 713 pp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
508
Sepúlveda, V., J. Vega & I. Delgado. 2000. Exposición severa a plomo ambiental en una población infantil de Antofagasta, Chile. Revista Médica de Chile, 128(2): 221-232. Smichowski, P., C. Vodopivez, R. Muñoz-Olivas & A. Gutiérrez. 2005. Monitoring trace elements in selected organs of Antarctic penguin (Pygoscelis adeliae) by plasma-based techniques. Microchemical journal, 82(1): 1-7. Sun, L. & Z. Xie. 2001. Changes in lead concentration in antarctic penguin droppings during the past 3.000 years. Environmental Geology, 40: 1205-1208. Tapia, R. 2011. Evaluación de la calidad del aire en la Península Fildes, Isla Rey Jorge, Antártica. Biomonitoreo de líquenes como herramienta de gestión. Tesis (Magíster en Gestión y Planificación Ambiental), Santiago, Chile, Universidad de Chile, 66 pp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
509
FEATHERS AND SOIL AS INDICATORS OF ENVIRONMENTAL CONTAMINATION
(Plumas y suelo como indicadores de contaminación ambiental)
Silva, D.G.A.1, Costa, E.S.*1, 2, Pereira, L.E. 1, Souza J.S.1, 2, Pessôa, A.R.L.1, Cunha, L.1, Alves, M.A.S.2, Torres1, J.P.M, Malm, O.1
1Laboratório de Radioisótopos Eduardo Penna Franca, Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, RJ. CEP: 214941-900. [email protected] 2Laboratório de Ecologia de Aves, Instituto de Biologia Roberto Alcantara Gomes, Universidade do Estado do Rio de Janeiro, RJ. CEP 20550-011.
The Mercury (Hg) is considered an environmental contaminant of global importance due to its high dispersion capacity, by bioaccumulate and biomagnification in the food chain. Seabirds are good bioindicators of environmental contamination because they are widely spread and they are top predators. Highest levels of contamination by Hg are expected in breeding areas of birds when compared with no breeding areas. Macronectes giganteus is a pelagic seabird with circumpolar distribution in the Southern Hemisphere. The aim of this study was set up Hg contamination levels in samples (feathers and soil) collected in a M. giganteus colony from Vaureal Peak in Admiralty Bay (King George Island, Antarctic Peninsula). The field took place from December 2010 to March 2011. The soil samples were separated in three groups according to the collect area of the colony: inside (n=3); around (n=4) and far away from the colony (n=3; control). The samples were analyzed according to specific methodology to feathers (n=10, Bastos 1997) and soil (n=10, Malm et al. 1989). The Hg levels in feathers ranged between 2.96 and 17.21 mg/kg ( =8.55±3.96 mg/Kg). The Hg concentrations of soil ( =0.19±0.14 mg/kg) showed significant variation (q>3.517; p<0.001) in different areas of the breeding colony. The highest values were found to samples collected inside the colony ( =0.37±0.16 mg/kg) and the lowest values to samples collected far away from the colony ( =0.09±0.02 mg/kg). Thompson et al. (1993), analyzing Hg contamination in feathers of M.
giganteus, presented mean results 11.90 (±6.15) to Gough Island (n=8) and 11.38 (±6.40) to Marion Island (n=30). The comparison between the present study and Thompson (1993) did not result in significant differences. Although the values for soil found in our study were not considered high, we can see from the data that the birds can collaborate to increase the Hg concentration in the soil. Analyzes will be conducted considering other trace elements to understand the dynamics of these contaminants in the environment.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
510
PROPIEDADES FÍSICAS Y COMPOSICIÓN ELEMENTAL DE LOS AEROSOLES EN LA ESTACIÓN ANTÁRTICA PERUANA MACHU PICCHU
(Physical properties and elemental composition of aerosols in Machu Picchu Peruvian Antarctic Station)
Luis Suarez1,2, Ana Contreras1, Julio Angeles2 y Abilia Larrauri2
1Instituto de Investigación, Universidad Continental, Av. San Carlos 1980, Huancayo, Perú 2Escuela de Ingeniería Ambiental, Universidad Alas Peruanas, Av. San Felipe 1109, Jesus Maria, Lima, Perú 3Facultad de Ingeniería Quimica, Universidad Nacional del Centro del Perú, Av. Mariscal Castilla 3909, El Tambo, RESUMEN El presente trabajo describe los resultados del estudio de las propiedad físicas y composición de los aerosoles, obtenidos de la campaña de mediciones y monitoreos realizada durante la XVII Expedición Antártica Peruana desde el mes de Enero 2007, en la Estación Científica Antártica Machu Picchu (ECAMP) a 62° de latitud Sur. El instrumental utilizado para la determinación de las propiedades físicas, fue un fotómetro solar SP02-L, el cual se encuentra dentro del marco instrumental del Año Polar Internacional. Este fotómetro dispone de 4 canales 412, 500, 675 y 862 nm lo que ha permitido medir espectros de irradiancia directa. Las observaciones utilizadas para el cálculo del espesor óptico de aerosol (AOD) fueron las realizadas entre las 10 am. y 2 pm. que corresponden a ángulos cenit solar inferiores a 45° y con cero de cobertura nubosa 30° alrededor del Sol. Además, también se determino el exponente de Ángstrom (α) que permite estudiar el tipo de aerosol presente en la atmósfera. Para la captura de material particulado se utilizaron soportes de filtro tipo cónico (Stacked Filter Unit, SFU), con filtros de teflón y nuclepore de 0.2 µm y 8 µm de diámetro respectivamente y analizados por la técnica de ICP (inductively coupled plasma) para la identificación de la composición elemental. El estudio de los valores y variaciones del AOD y α, son importantes debido a los pocos datos existentes en la región polar antártica y la necesidad de conocer sus características, dado que los aerosoles en esta región son un tema de gran interés, por su influencia en el clima y los efectos adversos en la salud. En este trabajo se presenta un estudio de los valores y variaciones del AOD y α para evidenciar los distintos tipos de partículas presentes en la atmósfera y verificar que la relación entre el AOD y el α brinde información para distinguirlas.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
511
LOS VARAMIENTOS DE LOS CETÁCEOS ¿SON UNA CAUSA DE MUERTE NATURAL?
(The cetaceans strandings, Are they a natural a cause of death?)
Aguayo-Lobo, A.1
1 Departamento Científico, Instituto Antártico Chileno (INACH). Plaza Benjamín Muñoz Gamero 1055, Punta Arenas, Chile. E-mail: [email protected]
Con motivo del varamiento de orcas falsas, Pseudorca crassidens, ocurrido en caleta Susana, estrecho de Magallanes, el 24 de febrero del presente año (Haro et al., en preparación) decidí abordar este interesante tema, que por segunda vez se presenta con esta especie de delfín en Magallanes (Ver Oporto et al., 1994), dónde existen canales estrechos y bahías cerradas, con desniveles grandes de marea, que permiten responder como hipótesis (morfología costera) a la pregunta formulada por los investigadores ¿Por qué se varan los Cetáceos? Los otros lugares del estrecho dónde han ocurrido varamientos son las playas norte y sur de la boca Oriental y la primera y segunda Angostura; así como al sur del canal Beagle, en bahía Windhond , sur de la isla Navarino, en la provincia Antártica. En este trabajo se define por varamiento de cetáceos, cuando uno o más ejemplares vivos arriban a la costa y posteriormente mueren. Por el contrario cuando uno o más ejemplares muertos, flotando en el mar, arriban a la costa, se define como ejemplares que fallecieron y son arrastrados por corrientes marinas que los arrojan en la playa sin vida. Teniendo en mente estas definiciones, intentaremos explicar las diferentes hipótesis que se han planteado para explicar los varamientos de las distintas especies de cetáceos: pero primero mencionaremos las causas de muerte conocidas de estos mamíferos marinos. Las causas de muerte de los ejemplares de cetáceos son diversas, como en los otros Ordenes de la Clase Mammalia: • Inanición, que es frecuente en las crías recién nacidas. • Traumas y Heridas Profundas, comunes en las colisiones con los barcos. • Enfermedades Infecciones como Erisipela, Pasteurilosis, Psedomoniasis, Hepatitis. Micosis
producidas por hongos de los géneros: Nocardia, Cándida, Aspergillus, Mucor, Actinobacillus, Blastomyces, Histoplasma, entre otros (Sweeney, 1986). Los agentes patógenos y virales encontrados en la Orca falsa, Pseudorca crassidens, y en los Calderones (Globicephala spp.) son: Streptococcus, Pseudomonas, Escherichia, Staphilococcus y el virus de la Influenza Humana tipo A. (Higgins et al. 1980).
• Enfermedades Parasitarias Internas causadas por Tremátodos, como Nasitrema spp., Braunina y Campula. Céstodos como Tetrabothrium y Strobilocephalus. Nemátodos como Anisakis spp., Contracaecum, Halocercus, Atenurus y Crassicauda. Acantocéfalos como Balbosoma spp. (Dailey & Brawnell, 1972). También se han descrito Parásitos Externos como especies de piojos de cetáceos, de los géneros Cyamus, Syncyamus e Isocyamus (Bowman, 1955) y algunos Epizoitos como Xenobalanus (Caldwell et al. 1970).
La competencia entre los ejemplares de cetáceos y los seres humanos por aprovechar las diversas poblaciones de peces, moluscos y crustáceos de importancia comercial, que viven en las comunidades del bentos, plancton y necton, han abierto un nuevo campo de investigación entre las interacciones de los ejemplares de mamíferos marinos y algunas pesquerías principales del mundo. Además, dichas interacciones han puesto en primera plana el tema de la Conservación de las poblaciones de mamíferos marinos, especialmente las correspondientes a los cetáceos, por ser el grupo más diverso (Aguayo Lobo, 1999). Otra causa de muerte son los contaminantes, los mas frecuentes registrados en cetáceos son los químicos y físicos. Entre los primeros está el DDT, el mercurio y los hidrocarburos derivados del

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
512
Petróleo (Gaskin, 1962), y entre los físicos el mas importante es la contaminación acústica; la cual en nuestro país ha sido poco estudiada, especialmente la producida por los ejercicios bélicos navales, dónde además del uso de sonares militares se efectúan explosiones submarinas, causando algunos varamientos de cetáceos (Aguayo Lobo et al. 1998). En el estudio de los contaminantes es importante separar las fuentes de la contaminación natural de las fuentes antropogénicas, como las causadas por la minería, agricultura, industrias varias y por los efluentes urbanos. Las hipótesis planteadas sobre los Varamientos se pueden reducir a cuatro: La primera fue sugerida por Slijper (1958; 1962) quién comparó a las poblaciones de cetáceos con las manadas de algunas especies de Arctiodáctilos, las cuales a veces son sobrecogidas por una sensación de pánico y de miedo repentino, como resultado de tormentas eléctricas con rayos y truenos, saliendo los animales disparados en una estampida, corriendo ciegos en varias direcciones en la pradera. Algunos grupos desorientados caen a quebradas, en ríos, otros en acantilados, quedando heridos, moribundos y otros muertos. Según el autor dichas estampidas podrían explicar algunos varamientos masivos de odontocetos, recordando que los cetáceos se orientan emitiendo y recibiendo sonidos, los cuales se reflejan en los objetos externos y vuelven hacia el animal emisor, fenómeno que fue demostrado posteriormente por Dudock van Heel, (1966) como Ecolocation, traducido por nosotros como Eco-ubicación. Las ondas sonoras extrañas de la tormenta: es decir de intensidad diferente al rango normal de los animales, alterarían el sistema receptor de los ejemplares, el nervio acústico y el centro auditivo del encéfalo, desorientándolos para terminar varándose en las costas más cercanas. Esta hipótesis de la estampida, del pánico y miedo, recibió luego otro apoyo con la descripción inequívoca del sistema denominado Sonar Biológico por Norris (1968). Los cetáceos a diferencia de los Primates (Visión) y de los Carnívora (Olfato) perciben la realidad del mundo exterior o del medio ambiente, usando un sistema de comunicación llamado Sonar Biológico; el cual funciona con un sistema emisor de ondas sonoras, situado en el aparato nasal complejo de los cetáceos y con un sistema receptor, situado en el oído del animal, el cual recibe las ondas sonoras reflejadas, vía las ramas mandibulares. El animal se forma así una imagen sonora del medio al captar el eco de las ondas que el mismo emite, ya sea para navegar, comunicarse con sus compañeros o para encontrar alimento. Otro apoyo más reciente a esta hipótesis es la relación encontrada por un grupo de investigadores (Fernández et al. 2005) entre el uso del Sonar Militar y los Varamientos de algunos ejemplares sanos de la Familia Ziphiidae. El sonar usado por los militares Mid Frequency Military Sonar es de intensidad alta y de frecuencia media. La pregunta que se hicieron los investigadores fue ¿Porqué ejemplares sanos de los géneros Ziphius, Hyperoodon y Mesoplodon se varan y mueren en la costa, a las pocas horas que se inician los ejercicios navales antisubmarinos? Estos varamientos singulares se conocieron por primera vez en los años de 1963, y aumentaron en los años de 1985, 1986 y 1989 en las islas Canarias, España, en 1996 en Grecia y en el año 2000 en las Bahamas. En las necrosis realizadas por los investigadores se registraron hemorragias cerebrales y en el oído y en el tejido adiposo del sistema complejo acústico (Frantzis, 1998), así como embolia de gases y de grasa, indicando que los animales varados habían experimentado la Enfermedad del Buzo o Decompression Sickness (Ridgway & Howards, 1979) al salir en estampida desde las profundidades, sin alcanzar a realizar la descompresión habitual en sus sumergidas; o por el contrario, se sumergieron más aún en estampida, más allá de sus profundidades habituales debido al pánico producido por las ondas sonoras intensas del Sonar Militar. La segunda hipótesis considera a los Varamientos como un mecanismo de control del tamaño de la población (Sergeant, 1982). Según este autor los Varamientos de cetáceos sólo se producen cuando el tamaño poblacional aumenta mucho, con alta densidad poblacional y en el centro espacial del área de distribución, no en los bordes; es decir, los varamientos servirían como un control interno del tamaño poblacional. Algunos autores han interpretado esta hipótesis como un acto de suicidio de los cetáceos

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
513
(Perrin & Geraci, 2002), denominándola en inglés como Suicidal Mass Stranding o Suicidio Masivo. El varamiento sería un acto voluntario, un esfuerzo para seguir respirando, sin importar el costo que se pague por ello, costo que sería la vida misma. En estos varamientos si uno intenta devolver al mar al o a los ejemplares varados, éste o éstos vuelven al lugar del varamiento una y otra vez, hasta terminar muriendo en el lugar. La tercera hipótesis corresponde a la denominada Morfología Costera (Gresson, 1968). En algunos lugares la costa forma bahías cerradas, canales estrechos, caletas abrigadas de aguas poco profundas y con amplios desniveles de marea, que al producirse la bajamar, las aguas dejan ver el fondo marino expuesto en grandes extensiones. Las diferencias de mareas producen corrientes rápidas de flujo y de reflujo. Como es fácil imaginar, estos accidentes del borde costero son verdaderas trampas para los ejemplares de cetáceos que viven en alta mar, en aguas profundas y lejos de la costa. Cuando grupos de esas especies de odontocetos se acercan a la costa persiguiendo a sus presas, como peces y calamares, los cuales desovan en aguas tranquilas en bahías con amplias variaciones de mareas, al bajar la marea terminan varándose en dichos lugares, pues quedan entrampados en el fondo somero de dichos lugares. La cuarta hipótesis es la de las Anomalía Magnéticas (Klinoska, 1985). Según algunos autores, entre ellos Gould (1980), todos los animales tienen en el Sistema Nervioso Central una estructura con cristales de magnetita, capaz de detectar las diferentes ondas magnéticas de nuestro planeta En su estudio Klinoska (op. cit.) encontró una relación directa entre los varamientos ocurridos en las islas Británicas y los lugares donde se presentan las anomalías geomagnéticas; es decir dónde existen rocas con propiedades geomagnéticas. En consecuencia para esta investigadora los Varamientos de los Cetáceos se deberían considerar como errores exclusivos cometidos por los animales que intentaron orientarse utilizando la topografía geomagnética. Posteriormente, Klinoska et al. (1986) mostraron esta misma relación en la costa este de los Estados Unidos de América. CONCLUSIONES Las causas de los varamientos aún son objeto de discusiones entre los investigadores, debido a que ninguna de las hipótesis propuestas, por si sola, puede explicarlos todos. 1.Se conoce sólo el final del proceso del varamiento, cuando los animales recién llegan vivos a la costa, pero no se conoce porqué y cuando se inicia el proceso en alta mar. 2.Se debe estudiar el comportamiento de los animales durante el proceso y el estado de salud o la condición médica de los ejemplares varados. 3.De los ejemplares que se logran salvar y son devueltos al mar, se deberían marcar y hacer un seguimiento, para conocer su período de de sobrevivencia, después de haber pasado el stress del varamiento. 4.La mayoría de los animales varados deben ser examinados vivos y luego realizar las necropsias respectivas, con el objeto de poder acumular datos suficientes para conocer la condición médica de los ejemplares. 5.Los animales que llegan muertos a la costa también deben ser examinados y ser sometidos a la necropsia, para poder a futuro diferenciar muy bien, los varamientos masivos de las muertes masivas de los cetáceos. REFERENCIAS Aguayo-Lobo, A., Torres, D., y Acevedo, J. 1998. Los mamíferos marinos de Chile. I. Cetácea. Serie Científica INACH, 48: 19-159. Aristóteles. 1965. History of Animals. Loeb Classical Library Nº 437. Harvard University Press. M A. (Greck with English Translation). Dailey, M. 1986. Parasitology-Basic Considerations. Pp.781-785. In: M.E. Fowler (Ed.). Zoo and Wild Animal Medicine. W.B. Sounder Co.Philadelphia.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
514
Gaskin, D.E. 1982. Social structure and social behaviour. Pp.112-118. In: The ecology Of Whales and Dolphins. Heinemann, London. Klinoska, M. 1985. Cetacean live stranding sites related to geomagnetic topography. Aquatic Mammals, 1: 27-32. Norris, K.S. 1968. The evolution of acoustic mechanisms in odontocete cetacean. Pp.297-324. In: E.T. Drake (Ed.). Evolution and Environment. Yale. University Press. Sergeant, D.E. 1982. Mass Stranding of toothed whales (Odontoceti) as a population phenomenon. Science Report Whales Research Institute, 34: 1-47. Slijper, E.J. 1958. 1962. Behaviour. Pp.179-202. In: Whales. Hutchinson of London.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
515
DIVERSIDAD DE POLIPLACÓFOROS ANTÁRTICOS CON ESPECIAL ÉNFASIS EN NUEVOS REGISTROS DE LAS CAMPAÑAS BENTART
(Diversity of Antarctic Polyplacophorans with special reference to new records from BENTART cruises)
Aldea, C.1, Troncoso, J.S.2 y F.J. García3
1Programa Gaia-Antártica, Universidad de Magallanes, Avenida Bulnes 01855, Punta Arenas, Chile. E-mail: [email protected]. 2Departamento de Ecología y Biología Animal, Facultad de Ciencias del Mar, , Universidad de Vigo, E- 36310, España. E-mail: [email protected]. 3Departamento de Sistemas Físicos, Químicos y Naturales, Universidad Pablo de Olavide, Sevilla, España. INTRODUCCIÓN Desde que comenzó el conocimiento de los moluscos en el Océano Antártico hacia finales del siglo XIX, la Clase Polyplacophora estuvo representada en los reportes generados de las expediciones científicas. El primer trabajo publicado, titulado “Report on the Polyplacophora collected by H.M.S. Challenger during the years 1873-76” (Haddon, 1886), incluyó poliplacóforos de algunas islas subantárticas. Desde esa época, numerosas expediciones fueron registrando especies desde diversas zonas ubicadas alrededor del continente antártico y áreas adyacentes, exclusivamente a través de reportes extensivos (e.g. Dell, 1964) y monografías (Kass & Van Belle, 1985-1990). La primera lista sobre la distribución y diversidad de poliplacóforos antárticos fue publicada por Götting (1989). Posteriormente, se continuaron aportando registros concretos de especies (Dell, 1990; Götting, 1993; Schwabe, 2008). A partir de la literatura publicada, hoy en día se tiene registro de 15 especies de poliplacóforos que habitan en el Océano Antártico. No obstante, el conocimiento sobre la diversidad de poliplacóforos en esta zona aún permanece difuso producto de la dispersión de literatura y registros imprecisos. En este contexto, el objetivo de este trabajo es describir la diversidad de especies de poliplacóforos que habitan en el Océano Antártico, agregando nuevos registros de las especies colectadas en las campañas BENTART 2003 y 2006, del Programa Antártico Español. MATERIALES Y MÉTODOS Diversidad antártica Teniendo como información preliminar la lista de Götting (1989), se revisaron todas las publicaciones que han registrado poliplacóforos en el Océano Antártico. Todos los registros se analizaron sistemáticamente y fueron ingresados a una base de datos. Adicionalmente se revisaron los registros de especies identificadas en el Museo Nacional de Historia Natural, USNM (http://invertebrates.si.edu/antiz/). Campañas BENTART 2003-2006 El área de estudio fue la zona occidental de la Península Antártica, desde las Islas Shetland del Sur hasta el Mar de Bellingshausen frente a la Isla Thurston (Fig. 1). A bordo del B/O Hespérides fueron obtenidas muestras bentónicas entre 5 y 2044 m de profundidad, utilizando diversos artes de muestreo: rastra Agassiz, rastra de roca, box-corer, trineo suprabentónico y buceo SCUBA (ver Aldea & Troncoso, 2008). Todos los especímenes recolectados fueron separados, fijados en etanol (70%) identificados a nivel de especie y registrados en la base de datos. Su clasificación taxonómica se realizó en base a Sirenko (2006) y Gofas (2013). Para efectos de consulta se mantienen en la colección del Departamento de Ecología y Biología Animal de la Universidad de Vigo (España).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
516
Figura 1. Área de estudio y estaciones de muestreo de las Campañas BENTART 2003-2006. Datos de las estaciones en
Aldea & Troncoso (2008). Figure 1. Study area and sampling stations of BENTART 2003-2006 cruises. For further information see Aldea & Troncoso
(2008). RESULTADOS Fueron registradas un total de 17 especies habitando al sur del frente polar, pertenecientes a seis familias y tres subórdenes diferentes (Tabla 1). La familia mejor representada es Ischnochitonidae, con cinco especies de tres géneros distintos. La especie con mayor cantidad de registros dentro del Océano Antártico es Nuttallochiton mirandus, la cual se distribuye alrededor de todo el continente antártico y algunas islas subantárticas.
Tabla 1. Polyplacohora distribuidos en el Océano Antártico. Las especies registradas en las campañas BENTART 2003-2006 se señalan con asteriscos en negrita.
Table 1. Polyplacohora distributed in Antarctic Ocean. Species recorded from BENTART cruises are highlighted with boldface asterisk.
Orden / Suborden / Familia Especie Fuentes actuales Lepidopleurida Lepidopleurina Leptochitonidae Leptochiton alveolus (Sars MS, Lovén, 1846) 2 Leptochiton columnarius (Hedley & May, 1908) 4 Leptochiton kerguelensis Haddon, 1886* 2 Chitonida Chitonina Ischnochitonidae Ischnochiton stramineus (Sowerby in Broderip & Sowerby, 1832) 2 Stenosemus chiversi Ferreira, 1981 4 Stenosemus exaratus (Sars, 1878) 1 Stenosemus simplicissimus (Thiele, 1906) 3 Tonicina zschaui (Pfeffer in Martens & Pfeffer, 1886)* 2, 4 Chaetopleuridae Chaetopleura brucei Iredale in Melvill & Standen, 1912 2 Callochitonidae Callochiton bouveti Thiele, 1906* 2, 4 Callochiton gaussae Thiele, 1908 2, 4 Callochiton steinenii (Pfeffer in Martens & Pfeffer, 1886) 2 Leloupia belgicae (Pelseneer, 1903) 1

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
517
Orden / Suborden / Familia Especie Fuentes actuales Acanthochitonina Mopaliidae Plaxiphora aurata (Spalowsky, 1795) 2 Nuttallochiton hyadesi (Rochebrune, 1889) 2, 4 Nuttallochiton mirandus (Smith MS, Thiele, 1906)* 2, 4 Hemiarthridae Hemiarthrum setulosum Carpenter in Dall, 1876* 2, 4
1: Kass & Van Belle (1985-1990); 2: Götting (1989); 3: Schwabe (2008); 4: USNM (invertebrates.si.edu/antiz/). De las muestras BENTART, fueron examinados un total de 127 poliplacóforos, identificándose cinco especies distintas: Leptochiton kerguelensis, Tonicina zschaui, Callochiton bouveti, Nuttallochiton mirandus y Hemiarthrum setulosum (Tabla 1, Fig. 2). La especie mejor representada fue L. kerguelenensis, correspondiendo al 69% de todos los individuos estudiados. Dos nuevos registros geográficos son dados para el Mar de Bellingshausen: L. kerguelenensis y N. mirandus. Además, una ampliación de rango batimétrico hasta 2044 m de profundidad es dada para L. kerguelensis.
Figura 2. Polyplacophora recolectados en las campañas BENTART 2003-2006. A, Leptochiton kerguelensis; B, Tonicina zschaui; C, Callochiton bouveti; D, Nuttallochiton mirandus; E, Hemiarthrum setulosum. Barra de escala: A, B, C, E (1
mm.); D (10 mm.). Figure 2. Polyplacophora collected in BENTART 2003-2006 cruises. A, Leptochiton kerguelensis; B, Tonicina zschaui; C, Callochiton bouveti; D, Nuttallochiton mirandus; E, Hemiarthrum setulosum. Scale bar: A, B, C, E (1 mm.); D (10 mm.).
DISCUSIÓN Y CONCLUSIONES Este trabajo entrega una sinopsis actual de la diversidad de poliplacóforos que habitan en el Océano Antártico, es decir en aguas ubicadas al sur del frente polar, reconociéndose 17 especies citadas de diversas fuentes. El trabajo anterior de esta temática reconocía nueve especies de distribución Antártica, más dos especies de distribución Sud-Atlántica (50-70ºS) y una de Islas Kerguelen (Götting, 1989). Adicionalmente, dos especies fueron confirmadas por Kass & Van Belle (1985-1990) y un nuevo registro de una especie sudafricana fue entregado por Schwabe (2008). Por consiguiente, a través de la literatura formal solamente se conocían 15 especies en el Océano Antártico. Los dos registros adicionales que se aportan este trabajo (Leptochiton columnarius y Stenosemus chiversi) aparecen ingresadas en el Museo Nacional de Historia Natural (USA), USNM, registradas para la zona de las Islas Shetland del Sur y fueron identificadas por Boris Sirenko. Al respecto, se hace necesario revalidar estos registros, con datos e imágenes de las especies.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
518
A pesar de que algunas especies son comunes y de amplia distribución en la Antártica, aún faltan registros concretos en numerosas áreas. Tal es el caso de los dos nuevos registros BENTART (L. kerguelenensis y N. mirandus), correspondientes a especies comunes que no se conocían para el Mar de Bellingshausen. Con el conocimiento existente actualmente, se puede concluir la baja diversidad existente de poliplacóforos en el Océano Antártico, con pocos y dispersos registros; considerando que gasterópodos y bivalvos suman 707 especies en esta zona (fide Clarke et al., 2007). Por consiguiente, es necesaria la disponibilidad de mayor cantidad de datos, información taxonómica fidedigna y muestras provenientes de distintos puntos al sur del frente polar. AGRADECIMIENTOS Se reconoce el trabajo de todos los participantes de las campañas Antárticas BENTART. REFERENCIAS Aldea, C. y Troncoso, J.S. 2008. Systematics and distribution of shelled mollusks (Gastropoda, Bivalvia and Scaphopoda) from the South Shetland Island to the Bellingshausen Sea, West Antarctica. Iberus, 26(2): 43-117. Clarke, A., Griffiths, H.J., Linse, K., Barnes, D.K.A. y Crame, J.A. 2007. How well do we know the Antarctic marine fauna? A preliminary study of macroecological and biogeographical patterns in Southern Ocean gastropod and bivalve molluscs. Diversity and Distributions, 13: 620-632. Dell, R.K. 1964. Antarctic & sub-Antarctic Mollusca: Amphineura, Scaphopoda & Bivalvia. Discovery Reports, 33: 99-250. Dell, R.K. 1990. Antarctic Mollusca with special reference to the fauna of the Ross Sea. Bulletin of the Royal Society of New Zealand, 27: 1-311. Gofas, S. 2013. Polyplacophora. World Register of Marine Species (http://www.marinespecies.org/aphia.php?p=taxdetails&id=55 on 2013-07-11). Götting, K.J. 1989. Los poliplacóforos (Mollusca) de las regiones antárticas y subantárticas. Medio Ambiente, 10(2): 54-60. Götting, K.J. 1993. Polyplacophora des Weddell-Meeres und von King George Island, Süd-Shetlands, Antarktis. Arch. Moll. Zilch-Festschr., 122: 171-187. Haddon, A.C., 1886. Report on the Polyplacophora collected by H.M.S. Challenger during the years 1873-76. Challenger Report. Zoology, 15: 1-51. Kaas, P. y Van Belle, R.A. 1985-1990. Monograph of Living Chitons (Mollusca: Polyplacophora). Vol. 1 (1985): Order Neoloricata: Lepidopleurina, 240 pp. Vol. 2 (1985): Suborder Ischnochitonina, Ischnochitonidae: Schizoplacinae, Callochitoninae and Lepidochitoninae, 198 pp. Vol. 3 (1987): Suborder Ischnochitonina. Ischnochitonidae: Chaetopleurinae & Ischnochitoninae (pars), 302 pp. Vol. 4 (1990): Suborder Ischnochitonina: Ischnochitonidae: Ischnochitoninae (continued), 298 pp. Vol. 5 (1990): Suborder Ischnochitonina: Ischnochitonidae: Ischnochitoninae (concluded), 402 pp. E. J. Brill/W. Backhuys, Leiden, The Netherlands. Schwabe, E. 2008. Discovery of the South African polyplacophoran Stenosemus simplicissimus (Thiele, 1906) (Mollusca, Polyplacophora, Ischnochitonidae) in the Southern Ocean. American Malacological Bulletin, 24(1/2): 71-77. Sirenko, B. 2006. New outlook on the system of chitons (Mollusca: Polyplacophora). Venus (Japanese Journal of Malacology), 65(1-2): 27-49.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
519
BIOGEOGRAFÍA DE LÍQUENES ANTÁRTICOS: UN ESTUDIO A PEQUEÑA ESCALA
(Biogeography of Antarctic lichens: a small-scale study) Caiafa, M.1, Renderos, L., Zepeda, P., Leiva, D., Farías, F., Carú, M., Orlando, J.1
1Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile. [email protected]
INTRODUCCIÓN Los líquenes, junto a los musgos, son los componentes predominantes de la flora criptogámica de los ecosistemas antárticos terrestres (Smith 1984). Además, son considerados como especies pioneras en la colonización de hábitats terrestres (Kappen 1973), probablemente debido a su gran tolerancia a condiciones extremas de temperatura y foto-estrés (Kappen 2000). De las más de 380 especies de líquenes descritas en la Antártida, cerca del 50% son endémicas (Øvstedal y Lewis Smith, 2001; Convey y col., 2009) y han sufrido procesos de especiación asociados a máximos glaciales (Peat y col., 2007). De acuerdo a Casanovas y col,. (2013) existen diversos factores que pueden influir en la riqueza de especies de líquenes a diferentes escalas espaciales. Entre ellos destaca la temperatura media anual, la cual se correlaciona negativamente con el número de especies en una escala regional (Green y col. 2011). Por otro lado, dentro de una escala local, la riqueza de especies de líquenes se correlaciona positivamente con los tamaños de las colonias de pingüinos menores a 20.000 parejas reproductoras (Casanovas y col., 2013). La mayor parte de las especies de líquenes que se han descrito en el continente antártico se encuentran entre los 60o S y los 62o S, observándose una fuerte disminución de esta riqueza hacia los 70o S (Peat y col., 2007), sin embargo, la predominancia relativa de los líquenes aumenta a mayores latitudes (Kappen 2000). Tradicionalmente, el continente antártico ha sido dividido en cuatro regiones de acuerdo a sus características climáticas y a la biodiversidad que albergan, éstas son: zona antártica templada del sur, zona sub-antártica, zona antártica marítima y zona antártica continental (Holdgate 1970). Recientemente, Terauds y colabores (2012) propusieron una clasificación en áreas biológicamente distintas que requieren una representación en una red de protección, generando 15 Regiones Biogeográficas de Conservación Antártica (ACBR, por sus siglas en inglés). El presente estudio se realizó en las Islas Shetland del Sur, las cuales se encuentran en la región Nor-Este de la península Antártica, de acuerdo a dicha clasificación. La mayoría de los estudios sobre líquenes antárticos se han centrado en los patrones de distribución global y regional. El objetivo del presente estudio es evaluar la existencia de una diferenciación genética del micobionte en tres géneros de líquenes, representantes de tres formas de crecimiento (crustoso, fruticoso y folioso), de acuerdo a un aislamiento geográfico a pequeña escala (menos de 1o de latitud). MATERIALES Y MÉTODOS Los sitios de muestreo antárticos correspondieron a la Isla Ardley (Isla Rey Jorge, Islas Shetland del Sur; ZAEP 150) y la Península Byers (Isla Livingston, Islas Shetland del Sur; ZAEP 126). En total se recolectaron 60 muestras -30 en cada uno de los sitios- de líquenes de los géneros Placopsis (crustoso; P1-P10 y P11-P20 desde Ardley y Byers, respectivamente), Stereocaulon (fruticoso; S1-S10 y S11-S20 desde Ardley y Byers, respectivamente) y Umbilicaria (folioso; U1-U10 y U11-U20 desde Ardley y Byers, respectivamente). El DNA de las muestras se extrajo utilizando el “PowerSoil DNA Isolation Kit” (Mobio) y a partir de éste se amplificaron los genes que codifican para el rRNA18S y el rRNA 28S usando los partidores EF4-EF3 (Smit y col. 1999) y LIC24R-LR7 (Moncalvo y col., 1993; Miadlikowska y Lutzoni 2000), respectivamente. Los amplicones se secuenciaron en un sentido con el partidor forward correspondiente utilizando unAnalizador Genético 3730 (Applied Biosystems; Macrogen Inc.).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
520
Las secuencias se revisaron y editaron manualmente en el programa Mega v5.2 (Tamura y col., 2011) y se compararon con la base de datos GenBank del NCBI mediante la herramienta bioinformática BLASTn (Altschul y col., 1997), para su identificación. Posteriormente, las secuencias nucleotídicas de los tres tipos de líquenes, se analizaron en el programa Mega v5.2 (Tamura y col., 2011) para establecer la distancia neta (distancia-p) entre los grupos de líquenes de cada sitio.Luego, el análisis filogenético de Máxima Verosimilitud se realizó comparando 1682 posiciones. Las secuencias alineadas mediante la herramienta ClustalW (Thompson y col., 1994) se analizaron en el programa GeneticDNASp v.5.10.1 (Librado y Rozas 2009). Las dos poblaciones (Ardley vs Byers) de cada género de liquen (Placopsis, Stereocaulon y Umbilicaria) se analizaron en el programa Arlequin v.3.01 (Excoffier y col., 2005) para calcular el índice de estructuración FST, y su significancia estadística. Finalmente, para determinar si existe algún tipo de correlación entre las distancias geográficas de los puntos de muestreo, calculadas con base en los datos de geo-referenciación transformados en distancia lineal (www.movable-type.co.uk/scripts/latlong.html), con respecto a las distancias genéticas de los diferentes líquenes (distancia-p), se realizó la Prueba de Mantel mediante la herramienta XlStat (Addinsoft; www.xlstat.com). RESULTADOS El análisis filogenético muestra una clara y robusta diferenciación de los tres líquenes estudiados. Sin embargo, sólo para el caso de Placopsissp. se observa una separación de las muestras de acuerdo al sitio de procedencia, aunque sin soporte estadístico (Figura 1). Las muestras de Ardley (P1-P10) forman un grupo separándose de las muestras
de Byers (P11-P20), las cuáles se separan a su vez en dos subgrupos bien definidos.
U-16 U-17 U-15 U-14 U-13 U-7 U-1 U-2 U-19
U-20 U-8
U-9 U-10
U-4 U-5
U-3 U-18
U-11 U-12
U-6 P-17 P-18 P-15 P-13
P-16 P-20 P-12 P-11
P-14 P-19
P-10 P-7 P-8 P-9 P-1 P-2 P-3 P-4 P-5 P-6
S-14 S-15
S-20 S-11
S-16 S-2 S-6 S-9
S-12 S-19
S-7 S-8
S-3 S-1 S-4 S-5 S-10 S-13 S-17 S-18
9987
76
100
100
100
100
86100
0.01 Figura 1. Relaciones filogenéticas entre los líquenes Placopsis(P), Stereocaulon (S) y Umbilicaria (U) basadas en el análisis de Máxima Verosimilitud. Los números en los nodos indican un soporte estadístico de bootstrap> 75% (1000 repeticiones). P1-P10; S1-S10 y U1-U10: Ardley; P11-P20; S11-S20 y U11-U20: Byers. Figure 1. Phylogenetic relationships between the lichens Placopsis(P), Stereocaulon (S) and Umbilicaria (U) (A) based in a Maximum Likelihood analysis. The numbers on the nodes indicate a statistical bootstrap support >75% (1000 repetitions). P1-P10; S1-S10 y U1-U10: Ardley; P11-P20; S11-S20 y U11-U20: Byers.
La distancia genética entre las dos poblaciones (Ardley y Byers) de cada género de liquen indica que las muestras de Stereocaulon y Umbilicaria son muy similares entre sí independientemente de su ubicación geográfica; mientras que la distancia entre los miembros del género Placopsis muestran una mayor distancia genética, probablemente por la diferente agrupación observada en el análisis filogenético dependiente del sitio de procedencia de las muestras (Tabla 1). El análisis de estructuración de las diferentes poblaciones arroja resultados que avalan lo mencionado anteriormente, ya que al calcular el índice FST se puede observar un valor de aproximadamente 0,5 en el caso de Placopsis, mientras que para los otros dos géneros de líquenes el valor es cero. Cabe

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
521
mencionar que los valores de FST oscilan entre 0 y 1, un valor cero implica que las dos poblaciones comparadas se entrecruzan libremente; un valor de uno implica que toda la variación genética se explica por la estructura de la población y que las dos poblaciones no comparten ninguna diversidad genética (Tabla 1). Finalmente, la correlación entre las distancias genéticas y la ubicación de las muestras no dio valores significativos por lo que se podría desestimar que la variación observada sea explicada por la distancia geográfica entre las muestras (Tabla 1).
Tabla 1. Parámetros de distancia genética, estructuración y correlación del test de Mantel. Table 1. Parameters of the genetic distance, structuring and Mantel correlation test.
Ardley vs Byers
Placopsis sp. Stereocaulon sp. Umbilicaria sp. distancia-p 0,00164 0,00024 0,00015 FST (estructuración) 0,45855 0,00000 0,00000 p (estructuración) 0,00000 0,00020 0,00002 R (Mantel) -0,38140 p (Mantel) 0,92700
DISCUSIÓN La diversidad genética que se observa en las distintas poblaciones de líquenes antárticos pueden explicarse por los procesos de colonización que han dado origen a dichas poblaciones, en este sentido Romeike y colaboradores (2002) plantea tres hipótesis sobre los posibles orígenes de los líquenes antárticos: (1) endemismos relictos de paleoclimas antárticos más templados, (2) eventos de colonización ocasionales de ecotipos especializados adaptados al clima antártico seguidos por procesos de especiación y propagación por el continente, y (3) constantes eventos de colonización independiente. Los resultados obtenidos indican que sólo en Placopsis se observó una diferenciación genética entre las poblaciones estudiadas, esto puede deberse en alguna medida a los mecanismos de reproducción utilizados por los líquenes, ya que en general los líquenes crustosos utilizan diasporas vegetativas en menor medida que los líquenes foliosos y fruticosos (Bowler y Rundel 1975). Según Smith (1984), frente a condiciones de estrés ambiental, las criptógamas favorecen estrategias de reproducción vegetativas por sobre la reproducción sexual. Además, se ha descrito que algunas especies de Umbilicaria se reproducen únicamente mediante propágulos vegetativos (Sancho y col., 1998), lo cual podría explicar la baja diferenciación observada en los representantes de dicho género. Sin embargo, también se deben considerar las limitaciones que presenta el uso de marcadores moleculares como los genes ribosomales en estudios de genética de poblaciones de micobiontes, los cuales pueden presentar una baja variabilidad (Zoller y col., 1999, Buschbom 2007), de manera que pueden no ser muy resolutivos en estudios a pequeña escala. CONCLUSIONES A partir del análisis de las secuencias nucleotídicas, se pudo determinar que las poblaciones del género Placopsis recolectadas en Ardley y Byers, muestran una tendencia a diferenciarse genéticamente. Sin embargo, este comportamiento no se observó en las poblaciones de los géneros Stereocaulon y Umbilicaria, probablemente debido a las estrategias reproductivas que presentan estos líquenes en la Antártida marítima. AGRADECIMIENTOS INACH - ECA 49 (Proy. F_02-10); Fondecyt 11100381.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
522
REFERENCIAS Altschul, S. F., Madden, T. L., A. Schäffer, A., Zhang, J., Zhang, Z., W.Miller y Lipman, D. J. 1997 Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research, 25(17):3389-3402. Bowler, P. y Rundel, P. 1975. Reproductive strategies in Lichens. Botanical Journal of the Linnean Society, 70(4): 325-340. Buschbom, J. 2007. Migration between continents: geographical structure and long-distance gene flow in Porpidiaflavicunda (lichen-forming Ascomycota). Molecular Ecology, 16(9):1835-1846 Casanovas, P., Lynch, H. J. y Fagan, W. F. 2013. Multi-scale patterns of moss and lichen richness on the Antarctic Peninsula.Ecography, 36(2):209-219. Convey, P., Stevens, M. I., Hodgson, D. A., Smellie, J. L., Hillenbrand, C. D., Barnes, D. K. A., Clarke, A., Pugh, P. J. A., Linse, K. y Cary, S. C. 2009. Exploring biological constraints on the glacial history of Antarctica.Quaternary Science Reviews, 28(27-28):3035-3048. Excoffier, L., Laval, G. y Schneider, S. 2005. Arlequinver 3.01.An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online, 1(1):47-60. Green, T. G. A., Sancho, L. G., Pintado, A. y Schroeter, B. 2011. Functional and spatial pressures on terrestrial vegetation in Antarctica forced by global warming.Polar Biology 34(11):1643-1656. Holdgate, M.W. (eds.) 1970. Antarctic Ecology. Academic Press, London. Kappen, L. 1973 Response to extreme environments. In:Ahmadiian, L. V. & Hale, M. E. (eds), The lichens. Academic Press, pp. 311-380. Kappen, L. 2000 Some aspects of the great success of lichens in Antarctica. Antarctic Science, 72 (3):314-324 Librado, P. y Rozas, J. 2009. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 25(11):1451-1452. Miadlikowska, J, y Lutzoni, F. 2000. Phylogenetic revision of the genus Peltigera (Lichen forming Ascomycota) based on morphological, chemical and large subunit nuclear ribosomal DNA data. International Journal of Plant Sciences, 161(6):925-958. Moncalvo, J. M., Rehner,S. A., y Vilgalys, R. 1993. Systematics of Lyophyllum section Difformia based on evidence from culture studies and ribosomal DNA sequences. Mycologia, 85(5):788-794. Øvstedal, D. O. y Lewis Smith, R.I. 2001.Lichens of Antarctica and South Georgia.A Guide to Their Identification and Ecology. Cambridge University Press, Cambridge. Peat, H. J., Clarke, A. y Convey, P. 2007. Diversity and biogeography of the Antarctic flora. Journal of Biogeography, 34(1):132-146. Romeike, J., Friedl, T., Helms, G. y Ott, S. 2002. Genetic diversity of algal and fungal partners in four species of Umbilicaria (Lichenized Ascomycetes) along a transect of the Antarctic Peninsula. Molecular Biology and Evolution 19(8):1209–1217. Sancho, L., Shroeter, B. y Valladares, F. 1998. Umbilicaria kappeni (Umbilicariaceae) a new lichen species from Antarctica with multiple mechanisms for the simultaneous dispersal of both bionts. Nova Hedwinia, 67(3-4):279-288. Smit, E., Leeflang, P., Glandorf, B., van Elsas, J. D., y Wernars, K. 1999 Analysis of fungal diversity in the wheat rhizosphere by sequencing of cloned PCR-amplified genes encoding 18S rRNA and temperature gradient gel electrophoresis. Applied and Environmental Microbiology, 65(6):2614-2621. Smith, R. I. L. 1984. Terrestrial plant biology of the sub-Antarctic and Antarctic.In: Laws, R. M. (ed.), Antarctic ecology. Academic Press, pp. 61-162. Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. y Kumar, S. 2011 MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 28(10):2731-2739. Terauds, A., Chown, S. L, Morgan, F., Peat, H. J., Watts, D. J., Keys, H., Convey, P., y Bergstrom, D. M. 2012. Conservation biogeography of the Antarctic. Diversity and Distributions, 18(2):726-741. Thompson, J. D., Higgins, D. G.,y Gibson, T. J. 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22(22):4673-4680. ZAEP 126. 2011. Plan de Gestión revisado de la Zona Antártica Especialmente Protegida Nº 126: Península Byers, Isla Livingston, Islas Shetland del Sur. XXXIV Reunión Consultiva del Tratado Antártico. ZAEP 150. 2009. Plan de gestión revisado de la Zona Antártica Especialmente Protegida Nº 150: Isla Ardley, Bahía Maxwell, Isla Rey Jorge. XXXII Reunión Consultiva del Tratado Antártico. Zoller, S., Lutzoni, F. y Scheidegger, C. 1999.Genetic variation within and among populations of the threatened lichen Lobariapulmonaria in Switzerland and implications for its conservation. Molecular Ecology, 8(2):2049-2059.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
523
BIOLOGÍAY ESTRUCTURA POBLACIONAL DEL KRILL (Euphausia superba) EN EL ESTRECHO DE BRANSFIELD DURANTE EL VERANO DEL 2013 (EXPEDICIÓN
ANTAR XXI - PERÚ)
(Biology and population structure of krill (Euphausia superba) in the Bransfield Strait during summer of 2013(ANTAR XXI Expedition - Perú)
Correa, J.1,2, Quiñones, J.1,3, Díaz, E.1,4 y M. Romero1,5
1Instituto del Mar del Perú – IMARPE. Esquina Gamarra y General Valle s/n, Chucuito, Callao – Perú. 2Área Funcional de Investigaciones en Oceanografía Biológica –IMARPE.E-mail: [email protected] 3Centro Regional de Investigaciones Pesqueras de Pisco- IMARPE.E-mail: [email protected] 4Área Funcional de Dinámica Poblacional y Evaluación de Recursos Pelágicos – IMARPE. E-mail: [email protected] 5Área Funcional de Biodiversidad – IMARPE. E-mail:[email protected] INTRODUCCIÓN Desde 1988, una de las principales investigaciones que realiza el Perú en la Antártida está referida a la determinación de los aspectos biológico-poblacionales del krill (Euphausia superba) en el Estrecho de Bransfield. En el presente trabajo se resumen los principales resultados obtenidos al respecto, realizados a bordo del BIC Humboldt entre el 15 y 22 de febrero durante el verano del 2013. MATERIALES Y MÉTODOS La información fue obtenida a partir de 6 lances positivos de pesca usando una red pelágica tipo Engel 1000/400 de poliamida con copo de 13mm y 5 arrastres de red bongo de 500µm sobre los 200 m de profundidad. A esta última, se le agregó un flujómetro para medir el volumen de agua filtrada. Se analizó la variación circadiana de la intensidad alimentaria ajustando a un Modelo Aditivo Generalizado (GAM). Se determinó los estadios de madurez sexual usando la clave de Kirkwood, 1982. Se determinó la estructura por tallas total y por grado latitudinal, comparándose mediante la prueba no paramétrica de Kolmogorov-Smirnov. Se calculó la abundancia relativa del E. superba provenientes de ambas redes, se estandarizó y expresó las abundancias y la distribución de krill en número de individuos/1000 m3 (Reiss et al, 2008). RESULTADOS Y DISCUSIÓN El análisis de la intensidad alimentaria demostró que el krill se alimentó durante las horas luz (Figura 1). Considerando todos los lances, se estableció que el 45.5% fueron juveniles, el 46.7% fueron sub-adultos y solamente el 8.6% fueron adultos. De éstos últimos, las hembras que pertenecen a las subcategorías con espermatóforos en el thelycum (3DF, fecundadas), no superaron el 3% debido a que existe variación interanual en el inicio del desove (Siegel y Loeb, 1995). La estructura de tallas del krill estimada para el área de estudio presentó rangos que variaron entre 15.0 y 53.0 mm para la captura con red bongo y entre 25.0 y 50.0 mm de longitud total para la red de pesca. Dicha estructura estuvo compuesta por más de dos grupos modales, notándose uno principal con tallas entre 33.0 – 40.0 mm y uno secundario con tallas entre 42.0 – 45.0 mm, además de resaltar aquellos grupos modales que pertenecerían a grupos de edades menores a 1 y hasta 3 años (de la Mare, 1995).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
524
Se notó diferencias significativas en las tallas del krill entre los diferentes grados longitudinales evaluados. Así, se observó una creciente ocurrencia de ejemplares adultos desde el oeste en el estrecho de Bransfield hacia el este en dirección a la Isla Elefante (figura 2).
Figura 1. Variación circadiana de la intensidad alimentaria del krill en el Estrecho de Bransfield.
Figure 1.Circadian variation of food intensity of krill in the Bransfield Strait.
Figura 2. Estructura de tallas del krill estimada en el Estrecho de Bransfield. Figure 2.Size structure of krill estimatedin the Bransfield Strait.
Considerando lo capturado por ambas redes, la abundancia promedio fue de 122.7±198.2 individuos/1000m3 (rango: 0-674.5, N = 30) cuya distribución homogénea estuvo cercana a la

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
525
Península Antártica. De acuerdo con las abundancias relativas registradas por AMLR (United States, Antartic Marine Living Resources) en la misma área geográfica, las abundancias relativas de krill encontradas durante la presente expediciónson comparables a la de los años 1982, 1996, 1998 y 2003 (Reiss et al, 2008). Las altas abundancias reportadas se debieron probablemente a la dominancia de juveniles y sub-adultos en las estructuras por tallas, además de mínimas abundancias de salpas y a una zona fría cercana a la Península Antártica. Así, las abundancias de krill suelen ser mayores en años con bajas temperaturas superficiales, mayor cobertura de hielos y mayores concentraciones de nutrientes, mientras que las salpas muestran el patrón contrario (Lee et al, 2011). CONCLUSIONES Las abundancias relativas del krill registradas durante esta expedición ANTAR XXI-2013 serían consideradas altas al ser comparadas con datos determinados por AMLR (United States, Antartic Marine Living Resources). Las altas abundancias de krill se debieron a la dominancia de estadios juveniles y sub-adultos, mínima abundancia de salpas y una zona fría (-1.0ºC) con influencia del mar de Weddell, estrecho de Gerlache. De los resultados obtenidos se podría inferir que el presente verano austral 2013 favorecería el recurso krill. AGRADECIMIENTOS A todos los científicos y tripulación del BIC Humboldt por su apoyo durante dicha expedición. REFERENCIAS De la Mare, W. 1995. Estimating krill recruitment and its variability.CCAMLR Science, Vol. 1: 55-69. Kirkwood, J.M. 1982. A guide to the Euphausiacea of the Southern Ocean. Australian National Antarctic Research Expeditions. Research Notes 1: 1-45. Lee, H., Pakhomov, E., Atkinson, A. and Siegel, V. 2011. Long-TermRelationships between theMarine Environment, Krill and Salps in the Southern Ocean.Journal of Marine Biology, 2010, 18p. Reiss, Ch., Cossio, A., Loeb, V. and Demer, D. 2008. Variations in the biomass of Antarctic krill (Euphausia superba) around the South Shetland Islands, 1996–2006.ICES Journal of Marine Science, 65: 12pp. Siegel, V., and Loeb, V. 1995.Recruitment of Antarctic krill Euphausia superba and possible causes for its variability. Marine Ecology Progress Series, 123: 45–56.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
526
VARIACIONES DE LAS COMUNIDADES EUCARIONTES PICO, NANO Y MICROFITOPLANCTÓNICOS EN BAHIA FILDES, ISLA REY JORGE, ANTARTICA,
DURANTE EL VERANO 2011 Y 2012.
(Variations of pico-, nano- and micro- phytoplanktonic eukaryotic communities in Fildes Bay, King George Island, Antarctic, during summer 2011 and 2012).
Egas C.1, Ibarra C1,2, Trefault N.2 & De la Iglesia R.1
1Departamento de Genética Molecular y Microbiología, Pontificia Universidad Católica de Chile, Alameda 340; 2Centro de Genómica y Bioinformática, Universidad Mayor, Camino La Pirámide 5750. E-mail: [email protected] INTRODUCCIÓN La Antártica es una zona que presenta grandes variaciones en intensidad y duración de los regímenes de luz. Esto convierte a la luz en un factor ambiental de gran relevancia para los organismos fotosintéticos que habitan en esta región. Sin embargo, poco se sabe de los cambios que estas variaciones generan, tanto en la composición comunitaria como en la estructura del fitoplancton en función del tamaño celular. En esta zona es esperable que las fluctuaciones de luz y en especial las variaciones en la Radiación Fotosintéticamente Activa (PAR), afecten la diversidad y la composición taxonómica del fitoplancton, generando cambios en la estructura de tamaño de éste. El objetivo de este trabajo es determinar la diversidad y la composición taxonómica de las comunidades eucariontes fotosintéticos en las distintas fracciones de tamaño (microfitoplancton (100-20 µm), nanofitoplancton (20-3 µm) y picofitoplancton (3-0,2 µm)) y analizar si estas presentan variaciones frente a distintos valores de PAR.
MATERIAL Y METODOS Para este trabajo se analizaron muestras recolectadas en Bahía Fildes, Isla Rey Jorge, Antártica (62°12’11’’ S; 58°55’15’’ O) durante enero 2011 y febrero 2012. A partir del total de muestras colectadas se seleccionó aquellas correspondientes a los días en que se detectó el mayor y el menor PAR y aquellas correspondientes al PAR intermedio entre los valores elegidos. La diversidad y composición taxonómica de las comunidades de eucariontes fotosintéticos para cada fracción de tamaño fue determinada mediante análisis de T-RFLP y librerías de clones para el gen 16S rRNA cloroplastidial. Las variaciones en su composición fueron determinadas mediante análisis estadísticos multivariados. RESULTADOS Los resultados muestran que las variaciones de PAR en el rango analizado no explican los cambios observados en la composición de las comunidades eucariontes fitoplanctónicas, sino que es el tamaño celular del fitoplancton el factor que mejor explica dicha variación. Interesantemente, la estructura del fitoplancton en función del tamaño celular que se observa en este estudio, no se condice con la estructura de tamaño canónica establecida por la literatura. A diferencia de lo esperado (tres fracciones de tamaño funcionales), se observó dos fracciones en la estructura de tamaño del fitoplancton. La fracción de tamaño de 12-0,2 µm se encontraba dominada por Bacilariofíceas y Criptofíceas y la fracción de tamaño 100-12 µm por Bacilariofíceas y Dinofíceas (Figura 1). Estos análisis contribuyen a mejorar la comprensión de la dinámica de las comunidades de eucariontes fitoplanctónicos en las aguas costeras de la Antártica.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
527
AGRADECIMIENTOS Este trabajo fue financiado por el proyecto INACH T16-10.
Figura 1. Análisis de NMDS de los perfiles de T-RFLPHaeIII de todas las muestras estudiadas. A, clasificación en base a la agrupación de las fracciones de tamaño 12-0,2 y 100-12 µm. B, abundancia relativa de la OTU a 251 nucleótidos en cada muestra. C, abundancia relativa de la OTU a 437 nucleótidos en cada muestra. D, abundancia relativa de la OTU a
834 nucleótidos en cada muestra. Figure 2. Analysis of NMDS T-RFLPHaeIII profiles of all samples studied. A, classification based on the grouping of the size fractions from 12-0,2 and 100-12 µm. B, relative abundance of the 251 nucleotides OTU in each sample. C, relative
abundance of the 437 nucleotides OTU in each sample. D, relative abundance of the 834 nucleotides OUT in each sample.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
528
PRIMERA COMPARACIÓN ISOTÓPICA DE LAS ÁREAS DE ALIMENTACIÓN DE LA BALLENA JOROBADA (Megaptera novaeangliae) EN AGUAS SUBÁNTARTICAS DE
SUDAMÉRICA Y PENÍNSULA ANTÁRTICA (First isotopic comparison of feeding grounds of humpback whale (Megaptera novaeangliae) from
the South American subantarctic waters and Antarctic Peninsula)
Haro, D.1, Riccialdelli, L.2, Aguayo-Lobo, A.3, Olavarría, C.4, 5 y J. Acevedo5
1Departamento de Presupuestos y Costos, Instituto Antártico Chileno (INACH), Plaza Muñoz Gamero 1055, Punta Arenas, Chile. E-mail:[email protected] 2Centro Austral de Investigaciones Científicas (CADIC), Bernardo A. Houssay 200, Ushuaia, Argentina. 3Departamento Científico, Instituto Antártico Chileno (INACH), Plaza Muñoz Gamero 1055, Punta Arenas, Chile. 4Dirección actual: 39 Moncrieff Avenue, Britannia Heights, Nelson 7010, Nueva Zelanda. 5Centro de Estudios del Cuaternario de Fuego-Patagonia y Antártica (CEQUA). 21 de Mayo 1690, Punta Arenas, Chile.
INTRODUCCIÓN Las ballenas jorobadas del Pacífico suroriental se alimentan principalmente en aguas de la costa oeste de la Península Antártica (Mackintosh 1965; Tomilin 1967), y en aguas del estrecho de Magallanes (Acevedo 2005; Acevedo et al. 2006) y golfo Corcovado (Haro 2009; Hucke-Gaete et al. 2013). En la Antártica se conoce que la dieta de esta especie está compuesta exclusivamente de krill (Euphausia superba) (Tomilin 1967). En el estrecho de Magallanes y aguas del golfo Corcovado, los estudios de ballenas jorobadas son más recientes, sin embargo, a través de observación directa de animales alimentándose en superficie, se ha señalado en Magallanes que la presa principal de esta ballena correspondería a la sardina fueguina (Sprattus fueguensis), seguida por el langostino de los canales (Munida gregaria) y el krill (Euphausia lucens) (Acevedo 2005; Acevedo et al. 2011). El análisis de isótopos estables ha demostrado ser una herramienta útil para determinar tanto áreas de alimentación de poblaciones marinas, así como diferentes grupos alimentarios para una misma especie (Gendron et al. 2001; Riccialdelli et al. 2012). Con el propósito de revalidar las áreas de alimentación de la población de ballenas jorobadas del Pacífico suroriental, se examinaron las diferencias isotópicas entre dos zonas de alimentación, costa oeste de la península Antártica y estrecho de Magallanes, medianteanálisis de isótopos estables de carbono y nitrógeno.
MATERIALES Y MÉTODOS Se colectaron muestras de piel de 33 ballenas jorobadas en el estrecho de Magallanes durante febrero y mayo de 2011 y 2012. Se colectaron muestras de las presas indicadas para esta zona, krill (E. lucens), langostino (M. gregaria) y sardina (S. fueguensis). Por otra parte, se realizó el análisis de isótopos estables de carbono y nitrógeno de dos muestras de piel de ballenas jorobadas obtenidas en la costa oeste de la Península Antártica, durante las temporadas de alimentación de 2006 y 2007. Se utilizaron datos isotópicos publicados de eufáusidos antárticos (Schmidt et al. 2003; Valenzuela 2008; Polito & Goebel 2010; Polito et al. 2011), con el fin de comparar sus valores con los de las ballenas. Todas la muestras fueron procesadas (liofilizadas, homogenizadas y con extracción de lípidos) previo al análisis de isótopos estables de carbono y nitrógeno, el cual fue realizado en un espectrómetro de masas en la Universidad de Concepción. Se realizaron comparaciones estadísticas entre los valores isotópicos obtenidos dentro de cada área de alimentación y entre áreas, y se determinó, mediante modelos de mezcla isotópicos (SIAR, Parnell et al. 2010), la contribución de cada presa en la dieta de las ballenas.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
529
RESULTADOS La ballena jorobada en el estrecho de Magallanes, presentó valores promedios de 14,7 (±1,0) ‰ y -16,3 (±0,6) ‰ en δ15N y δ13C respectivamente, no existiendo diferencias significativas entre clases etarias ni entre sexos (p>0,05). Los valores isotópicos fueron significativamente mayores durante el 2011 (p<0,05). Se encontraron promedios de δ15N y δ13C en krill de 11,8 (+0,7) ‰ y -17,7 (+ 0,4) ‰, en langostinos de 11,7 (+1,7) ‰ y -17,9 (+0,5) ‰, y en sardinas de 14,9 (+2,1) ‰ y -16,9 (+0,4) ‰ respectivamente. Entre años se observó una inversión de la contribución de las presas a la dieta de las ballenas analizadas. Durante el 2011, la contribución de la sardina (55 +12%) resultó ser significativamente mayor (p<0,05) que la del langostino (45 + 12%). Mientras que para el 2012 la contribución de la sardina (18 + 9%), fue significativamente menor (p<0,05) respecto a los crustáceos (82 + 9%). En la península Antártica, las ballenas presentaron valores promedios de 8,7 (±0,4) ‰en δ15N y -23,1 (±1,2) ‰ en δ13C. Se encontraron promedios de δ15N y δ13C de 3,5 (+0,5) ‰ y -26,7 (+1,2) ‰ en E. superba, y de 2,3‰ y -29,4 ‰, en Thysanoessa sp. La contribución de E. superba (53+24%) resultó ser significativamente mayor (p<0,05) que la de Thysanoessa sp. (47+ 24%) en la dieta de la ballena en la Antártica. Las proporciones isotópicas encontradas en ballenas y eufáusidos fueron significativamente diferentes en las dos áreas de alimentación (p<0,05) (Fig. 1). Se encontró una diferencia de 6‰ en los valores medios de δ15N, y de 6.8‰ en los valores medios de δ13C siendo, en ambos casos, significativamente mayores para las ballenas que se alimentan en aguas Magallánicas. En los eufáusidos, se encontró una diferencia de 8,9‰ en δ15Ny de -10,4‰ en δ13C, siendo significativamente mayores en el estrecho de Magallanes. DISCUSIÓN Y CONCLUSIONES Las marcadas diferencias isotópicas encontradas tanto entre las ballenas como en sus principales presas entre Magallanes y la Península Antártica (Fig. 1), verifican que ambas zonas son áreas de alimentación diferenciadas para la población del Pacífico suroriental. Los resultados encontrados refuerzan estudios previos, que señalan diferencias entre las ballenas de ambas áreas en relación a la ausencia de intercambio de animales (Acevedo et al. 2008) y a diferencias en la diversidad genética (análisis de ADN mitocondrial) (Olavarría et al. 2006; Olavarría 2007). En esta primera aproximación, se ha demostrado que el análisis isotópico sobre muestras de piel de ballenas jorobadas resultaría un método útil para lograr diferenciar las áreas de alimentación de los individuos que transitan por las aguas del Pacifico suroriental. A pesar de necesitarse mayores estudios sobre la trama trófica así como de la dieta de esta especie en ambas áreas, para determinar posibles diferencias entre los grupos alimentarios, los análisis realizados revalidan la dieta de esta especie en ambos sectores. Para Magallanes, los resultados indican que tanto los crustáceos como los peces son presas importantes, pudiendo variar el consumo anual posiblemente en relación a la disponibilidad de alimento (Haro 2013). Tanto adultos como juveniles, así como ejemplares machos y hembras de ballenas, consumirían presas similares. En la Península Antártica las ballenas consumieron principalmente E. superba respecto a Thysanoessa sp, resultado que es apoyado por el conocimiento de la dieta de esta especie en esa zona (Tomilin 1967).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
530
Figura 1: Valores promedio y desviaciones estándar de carbono (δ13C) y nitrógeno (δ15N) en muestras de ballena jorobaday en muestras presas en aguas Magallánicas (círculo superior derecho), y en muestras de ballena jorobada y de eufáusidos en la Península Antártica (Círculo inferior). Figure 1. Average values and standard deviations of carbon (δ13C) and nitrogen (δ15N) in samples of hump back whale and prey from Magellanic waters (upper right circle) and hump back whale samples and euphausiids in the Antarctic Peninsula (lower circle). REFERENCIAS BIBLIOGRÁFICAS Acevedo, J. 2005. Distribución, filopatría, residencia e identidad poblacional de la ballena jorobada, Megaptera novaeangliae, que se alimentan en las aguas del estrecho de Magallanes, Chile. Tesis de Maestría en Ciencias. Facultad de Ciencias. Universidad de Magallanes, 144 Págs. Acevedo, J., Aguayo-Lobo, A. y Pastene, L. 2006. Filopatría de la ballena jorobada (Megaptera novaeangliae Borowski, 1781), al área de alimentación del estrecho de Magallanes. Revista de Biología Marina y Oceanografía, 41(1):11-19. Acevedo, J., Hucke-Gaete, R., Secchi, E., Allen, J., Aguayo-Lobo, A., Dalla Rosa, L., Haro D. and Pastene, L. 2008. Photo-identification analysis of humpback whales from three high latitude locaties of the Eastern South Pacific Population (Stock G). Comité Científico de la Comisión Ballenera Internacional SC/60/SH27, Santiago, Chile. Acevedo, J., Plana, J., Aguayo-Lobo A. and Pastene, L.2011. Surface feeding behaviors in the Magellan Strait humpback whales. Revista de Biología Marina y Oceanografía, 46(3): 483-490. Gendron, D., Aguiniga, S. and Carriquiry, J.D.2001.δ15N and δ13C in skin biopsy samples: a note on their applicability for examining the relative trophic level in three rorqual species. Journal of Cetacean Research Management, 3:41-44. Haro, D. 2009. Identificación individual de ballenas jorobadas, Megaptera novaeangliae (Borowski, 1781), en el golfo Corcovado, Patagonia Norte, Chile: 2003-2009. Tesis de Biología Marina. Facultad de Ciencias. Universidad Austral de Chile, 92 Págs. Haro, D. 2013. Relaciones tróficas de la ballena jorobada (Megaptera novaeangliae) en el Área Marina Costera Protegida Francisco Coloane, estrecho de Magallanes, Chile. Tesis de Maestría en Ciencias. Facultad de Ciencias. Universidad de Magallanes,139 Págs. Hucke-Gaete, R., Haro, D., Torres-Flórez, J.P., Montecinos, Y., Viddi, F., Bedriñana-Romano, B., Nery, M. and Ruiz, R. 2013. A historical feeding ground for humpback whales in the Eastern South Pacific revisited: the case of northern Patagonia, Chile. Aquatic Conservation: Marine & Freshwater Ecosystems. DOI: 10.1002/aqc.2343 Mackintosh, N.A. 1965.The stocks of whales.Fishing News (Books) Ltd. London, 232 Págs. Olavarría, C., Aguayo-Lobo, A., Acevedo, J., Medrano, L., Thiele, D. and Baker, C.S. 2006. Genetic differentiation between two feeding areas of the Eastern South Pacific humpback whale population: Update on SC/57/SH3. Report SC/A06/HW29 to the Inter-sessional workshop for the Comprehensive Assessment of Southern Hemisphere humpback whales Scientific Committee of the International Whaling Commission. Hobart, Australia. 3 - 7 April.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
531
Olavarría, C. 2007. Population structure of Southern Hemisphere humpback whales.Unpublished thesis, The University of Auckland, Auckland, New Zealand, 268 pp. Parnell, A.C., Inger, R., Bearhop, S. and Jackson, A.L. 2010 Source partitioning using stable isotopes: coping with too much variation. Plos One.5 (3) e9672. Polito, M.J. and M.E. Goebel. 2010. Investigating the use of stable isotope analysis of milk to infer seasonal trends in the diets and foraging habitats of female Antarctic fur seals. Journal Experimental Marine Biology Ecology, 39:1–9. Polito, M.J., Lynch, H.J., Naveen, R. and Emslie, S.D. 2011.Stable isotopes reveal regional heterogeneity in the prebreeding distribution and diets of sympatrically breeding Pygoscelis spp. penguins. Marine Ecology Progress Series, 421:265-277. Riccialdelli, L., Newsome, S.D., Goodall, R.N.P., Fogel, M., Bastida, R. 2012. Insight into niche separation of Risso’s dolphin (Grampus griseus) in the southwestern South Atlantic via d13C and d15N values. Marine Mammal Science, 28:e503-e515 Schmidt, K., Atkinson, A., Stubing, D., McClelland, J.W., Montoya, J.P., Voss, M. 2003. Trophic relationships among southern ocean copepods and krill: some uses and limitations of a stable isotope approach. Limnology Oceanography, 48:277-289. Tomilin, A.G. 1967. Megaptera nodosa. In: Cetacea. Vol. IX. Mammals of the U.S.S.R. and adjacent countries. Academy of Science of the USSR. Moscow. Translated from Russian Israel Program for Scientific Translations. Jerusalen, pp 246-295. Valenzuela, L. 2008. Population structure and foraging ecology of southern right whales (Eubalaena australis): insights from isotopic and genetic analyses. Doctor of Philosophy Thesis. University of Utah, United states, 140 pp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
532
ESPECIFICIDAD Y SELECTIVIDAD EN LÍQUENES TRIPARTITOS DEL GÉNERO Placopsis DESDE DOS SITIOS DE LAS ISLAS SHETLAND DEL SUR
(Specificity and selectivity in tripartite lichens of the genus Placopsis from two sites in the South Shetland Islands)
Leiva, D.1, Zúñiga, C., Farías, F., Carú, M., Orlando, J.1
1Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile. Email: [email protected] INTRODUCCIÓN Los líquenes son asociaciones simbióticas compuestas por dos o tres organismos (bi o tripartitos, respectivamente), entre los cuales está siempre presente un hongo (micobionte) y un fotobionte, el cual puede corresponder a un alga (clorobionte), una cianobacteria (cianobionte), o ambos a la vez. El micobionte aporta con la estructura para alojar al organismo fotosintético, mientras que el fotobionte genera compuestos carbonados simples que pueden ser aprovechados por el hongo; además el cianobionte aporta con la fijación de nitrógeno (cianolíquenes). En los líquenes tripartitos, el cianobionte se aloja en estructuras especializadas llamadas cefalodios. Según su forma de crecimiento, los líquenes pueden clasificarse en tres tipos principales: crustosos (forma de corteza), foliosos (forma de hoja) y fruticosos (forma de fruto) (Nash, 2008). En la formación exitosa de esta asociación es fundamental el tipo de reproducción que origina al organismo simbionte, en el caso de una reproducción asexual (propágulos vegetativos), probablemente se mantendrá la misma relación micobionte-fotobiontes, mientras que en el caso de existir reproducción sexual (esporas fúngicas), debe generarse una asociación de novo con los organismos fotosintéticos disponibles y compatibles (Nash, 2008). Se ha descrito que la disponibilidad de fotobiontes en regiones antárticas es limitada, observándose pocos organismos asociados a distintos hongos liquenizables (Pérez-Ortega y col., 2012; Wirtz y col., 2003). Placopsis (Ascomycota; Lecanoromycetes; Agyriales) es un género de hongos formadores de líquenes crustosos tripartitos. Se le ha observado en zonas de climas extremos (Antárticos y Subantárticos) y con un llamativo crecimiento en lugares de desglaciación reciente (Raggio y col., 2012; Sancho y col., 2010). Crece adherido a la roca como sustrato y por lo tanto su manejo al momento de extraer muestras y luego tratarlas es complejo. La gran velocidad de crecimiento que se ha observado en estos líquenes, así como sus diversas estrategias de reproducción (Galloway y col., 2005), hacen prever una distribución de especies similares incluso en lugares aislados geográficamente, es por esto que se propone un análisis de los marcadores moleculares SSU rDNA a nivel de haplotipos y la determinación de la especificidad y la selectividad de las asociaciones liquénicas en representantes de este género. MATERIALES Y MÉTODOS Las muestras de talo liquénico se recolectaron desde 2 locaciones de las islas Shetland del Sur: Isla Ardley (Isla Rey Jorge; ZAEP 150) y la Península Byers (Isla Livingston; ZAEP 126). Diez muestras se extrajeron desde cada sitio en un radio de 2 metros desde las coordenadas 62º12'49.32"S-58º56'14.64"O y 62º39'6.84"S-61º5'59.28"O, respectivamente, y se almacenaron en bolsas de papel selladas, rotuladas y a temperatura ambiente hasta el momento de su uso. Para la extracción de DNA de cada muestra, primero se procedió a separar mecánicamente el talo del sustrato y luego se aplicó el protocolo de extracción mediante el kit PowerLyzer™ PowerSoil®

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
533
DNA Isolation Kit (MoBio). Luego se amplificaron mediante la técnica de PCR los marcadores moleculares rDNA 18S (Smit y col., 1999) y rDNA 16S (Wilmotte y col., 1993) con partidores específicos para hongos y cianobacterias, respectivamente. Los amplicones se secuenciaron con el partidor forward correspondiente utilizando un Analizador Genético 3730 (AppliedBiosystems; Macrogen Inc.). Las secuencias nucleotídicas se revisaron y editaron manualmente en el programa Mega v5.2 (Tamura y col., 2011) y se compararon con la base de datos GenBank del NCBI mediante la herramienta bioinformática BLASTn (Altschul y col., 1990). Las secuencias se analizaron en el programa Network v4.6, mediante el algoritmo Median Joining (Bandelt y col., 1999) para definir los haplotipos de micobiontes (HMs) y cianobiontes (HCs). La especificidad de cada HM se definió como el número de haplotipos de cianobiontes con los que se asocian, ponderado por la relación taxonómica entre ellos, que se calculó como el promedio de sus distancias genéticas (distancias-p). Por lo tanto: Especificidad ponderada (Ep) = 1 – [(n° de haplotipos de cianobiontes) x (distancia-p promedio entre los haplotipos de cianobiontes)]. La selectividad, por otro lado, se define teóricamente como la asociación preferencial de los micobiontes con alguno de sus cianobiontes específicos. Para su cálculo se consideró la frecuencia de las diferentes parejas simbióticas formadas por cada micobionte. Para ello se usó el índice de diversidad ponderada de Shannon (Poulin y col., 2011). Por lo tanto: Selectividad ponderada (Sp) = 1 - (-∑wi pi log pi), donde pi es la frecuencia de cada pareja simbiótica y wi es un factor que incorpora la relación taxonómica de los haplotipos de cianobiontes que forman las parejas. De acuerdo con la Ep y la Sp de cada micobionte, se estableció una serie de categorías que los clasifica en diferentes grados de especificidad y selectividad. RESULTADOS Y DISCUSIÓN Luego del análisis de las 40 secuencias obtenidas, tanto para el marcador rDNA 18S (965 pb, n=20) como para el rDNA 16S (780 pb, n=20), se obtuvo un total de 4 haplotipos de micobiontes (HM1, HM2, HM3 y HM4), todos relacionados según la base de datos a Placopsis gelida, y 6 de cianobiontes (HC1, HC2, HC3, HC4, HC5 y HC6) asociados al género Nostoc; la distribución de las secuencias de las muestras en haplotipos se detalla en la Tabla 1.
Tabla 1. Distribución de las secuencias de las muestras en haplotipos. Table 1. Distribution of sample’s sequences in haplotypes.
Haplotipo Micobionte Secuencias Incluidas HM1 MA1, MA2, MA3, MA4, MA5, MA6. HM2 MA7, MA8, MA9, MA10, MB1, MB2, MB4, MB6, MB10. HM3 MB3, MB5, MB7, MB8. HM4 MB9.
Haplotipo Cianobionte Secuencias Incluidas HC1 CA1, CA2, CA7, CA9. HC2 CA3, CA4. HC3 CA5. HC4 CA6. HC5 CA8, CA10. HC6 CB1, CB2, CB3, CB4, CB5, CB6, CB7, CB8, CB9, CB10.
Haplotipos de micobiontes (HM), de cianobiontes (HC) y secuencias de micobiontes y cianobiontes de Ardley (MA, CA) o Byers (MB, CB). Haplotypes of mycobionts (HM), cyanobionts (HC), and associated mycobiont and cyanobiont sequences from Ardley (MA, CA) or Byers (MB,
CB).
En cuanto a los cianobiontes, se observa una clara distribución de haplotipos según su ubicación geográfica, ya que los haplotipos HC1-HC5 se encuentran en Ardley y sólo el HC6 se presenta en

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
534
Byers. En el caso de los micobiontes, se observan 2 haplotipos en Ardley (HM1 y HM2) y 3 en Byers (HM2, HM3 y HM4), siendo el HM2 el único presente en ambas locaciones (4 muestras de Ardley y 5 de Byers), lo que se explicaría por la alta capacidad de crecimiento y diversidad de estrategias reproductivas observadas en Placopsis. En la Figura 1 se observa un diagrama con la distribución de las parejas HM-HC. Las distribuciones graficadas en esta figura indican una alta especificidad de los haplotipos de micobionte HM3 y HM4, ya que están asociados a un sólo cianobionte; mientras que para HM1, que se une a 4 HCs, y HM2, asociado con 3 HCs, es necesario considerar las distancias filogenéticas para realizar una estimación más precisa de su especificad (Tabla 2).
Figura 1. Distribución de las parejas HM-HC en ambos sitios de muestreo. Los círculos muestran los haplotipos de micobiontes (HM) con su respectiva frecuencia. Las diferentes texturas indican los diversos haplotipos de cianobiontes (HC), detallados en la leyenda. Los puntos negros indican la base nucleotídica que diferencia a cada haplotipo, el número adyacente da cuenta de la posición de dicha base en la secuencia del rDNA 18S (965 pb en total). Figure 1. Distribution of HM-HC pairs in both sampling sites. Circles show mycobiont haplotypes (HM) with their respective frequency. The different textures indicate the cyanobiont haplotypes (HC), which are detailed in the key. Black dots indicate the nucleic base that differs from each haplotype; the adjacent number shows the position of such base at the 18S rDNA sequence (965 bp in total).
Tabla 2. Especificidad y selectividad de los distintos micobiontes. Table 2. Specificity and selectivity of the different mycobionts.
Haplotipo Micobionte
N° Haplotipos Cianobiontes
Asociados
Distancia-p Promedio
Especificidad Ponderada
Grado de Especificidad*
Selectividad Ponderada
Grado de Selectividad**
HM1 4 0,0045 0,9765 Alto 0,9941 Medio HM2 3 0,0026 0,9924 Muy Alto 0,9975 Alto HM3 1 0,0000 1,0000 Muy Alto 1,0000 Muy Alto HM4 1 0,0000 1,0000 Muy Alto 1,0000 Muy Alto
*Grados de Especificidad / Specificity degrees: Muy Bajo / Very Low = [0.9200, 0.9299]; Bajo / Low = [0.9400, 0.9549]; Medio / Medium = [0.9550, 0.9749]; Alto / High = [0.9750, 0.9899]; Muy Alto / Very High = [0.9900, 1.0000]. **Grados de Selectividad / Selectivity degrees: Muy Bajo / Very Low = [0.9800, 0.9819]; Bajo / Low = [0.9820, 0.9849]; Medio / Medium = [0.9850, 0.9949]; Alto / High = [0.9950, 0.9979]; Muy Alto / Very High = [0.9980, 1.0000].
En esta última tabla, que muestra los resultados de los cálculos de selectividad y especificidad para cada haplotipo de micobionte, se verifica que el grado de selectividad disminuye según aumenta el número de HCs asociados a cada HM, aunque siempre en el rango “medio” o superior. En cuanto a la especificidad, se observa que en el caso de HM2 su grado es “muy alto” debido a la cercanía filogenética de sus HCs asociados; para HM1 en cambio, se obtiene un grado “alto” de especificidad debido a que los cianobiontes con los cuales se asocia no son tan cercanos genéticamente comparado con aquellos HCs que se asocian con HM2. En el caso de HM3 y HM4

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
535
se obtiene un grado “muy alto”, lo cual es esperable debido a su asociación con solo un HC. En general, se observa un grado de especificidad “alto” o “muy alto” en los líquenes estudiados, debido tanto a la asociación con sólo un representante cianobacteriano o a la cercanía filogenética entre los cianobiontes, esto puede deberse a la escasez de especies cianobacterianas en estos ambientes, descrito también por Wirtz y col. (2003). CONCLUSIONES La presencia del haplotipo HM2 en ambas localidades se condice con la alta capacidad de crecimiento y diversas estrategias reproductivas observadas en líquenes del género Placopsis. Debido probablemente a la escasa diversidad de representantes de cianobacterias que se espera en estos ambientes, se observó una alta especificidad en las asociaciones micobionte-cianobionte. Finalmente, el análisis de haplotipos para el cálculo de la especificidad y la selectividad ponderadas por las distancias genéticas entre ellos, supone un buen acercamiento para el estudio ecológico de la simbiosis liquénica, sobre todo para el caso de muestras cercanas geográfica y genéticamente como las estudiadas en esta investigación. AGRADECIMIENTOS INACH - ECA 49 (Proyecto F_02-10); Fondecyt 11100381. REFERENCIAS Altschul, S. F., Gish, W., Miller, W., Myers, E. W., y Lipman, D. J. 1990. Basic local alignment search tool. Journal Of Molecular Biology, 215(3):403-410. Bandelt, H., Forster, P., y Röhl, A. 1999. Median-joining networks for inferring intraspecific phylogenies. Molecular Biology And Evolution, 16(1):37-48. Galloway, D. J., Lewis-Smith, R. I., y Quilhot, W. 2005. A new species of Placopsis (Agyriaceae: Ascomycota) from Antarctica. The Lichenologist, 37(04):321-327. Nash, T. 2008. Lichen Biology. (T. H. Nash, Ed.) (2nd ed.). Cambridge: Cambridge University Press. Pérez-Ortega, S., Ortiz-Álvarez, R., Allan Green, T. G., y De Los Ríos, A. 2012. Lichen myco- and photobiont diversity and their relationships at the edge of life (McMurdo Dry Valleys, Antarctica). FEMS Microbiology Ecology, 82(2):429-448. Poulin, R., Krasnov, B. R., y Mouillot, D. 2011. Host specificity in phylogenetic and geographic space. Trends In Parasitology, 27(8):355-361. Raggio, J., Green, T. G. A., Crittenden, P. D., Pintado, A., Vivas, M., Pérez-Ortega, S., Ríos, A., y Sancho, L. G.. 2012. Comparative ecophysiology of three Placopsis species, pioneer lichens in recently exposed Chilean glacial forelands. Symbiosis, 56(2):55-66. Sancho, L. G., Palacios, D., Green, T. G. A., Vivas, M., y Pintado, A. 2010. Extreme high lichen growth rates detected in recently deglaciated areas in Tierra del Fuego. Polar Biology, 34(6):813-822. Smit, E., Leeflang, P., Glandorf, B., Van Elsas, J. D., y Wernars, K. 1999. Analysis of fungal diversity in the wheat rhizosphere by sequencing of cloned PCR-amplified genes encoding 18S rRNA and temperature gradient gel electrophoresis. Applied And Environmental Microbiology, 65(6):2614-2621. Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., y Kumar, S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology And Evolution, 28(10):2731-2739. Wilmotte, A., Auwera, G. Van der, y Wachter, R. De. 1993. Structure of the 16 S ribosomal RNA of the thermophilic cyanobacterium Chlorogloeopsis HTF (’Mastigocladus laminosus HTF’) strain PCC7518, and phylogenetic analysis. FEBS Letters, 317(1,2):96-100. Wirtz, N., Lumbsch, H. T., Green, T. G. A., Türk, R., Pintado, A., Sancho, L., y Schroeter, B. 2003. Lichen fungi have low cyanobiont selectivity in maritime Antarctica. New Phytologist, 160(1):177-183. ZAEP 126. 2011. Plan de Gestión revisado de la Zona Antártica Especialmente Protegida Nº 126: Península Byers, Isla Livingston, Islas Shetland del Sur. XXXIV Reunión Consultiva del Tratado Antártico. ZAEP 150. 2009. Plan de Gestión revisado de la Zona Antártica Especialmente Protegida Nº 150: Isla Ardley, Bahía Maxwell, Isla Rey Jorge. XXXII Reunión Consultiva del Tratado Antártico.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
536
ESTIMACION DE LA EDAD ABSOLUTA EN FOCAS CANGREJERAS LOBODON CARCINOPHAGA MOMIFICADAS DE LA PENÍNSULA ANTÁRTICA.
(Absolute age determination of mummified crabeater seals Lobodon carcinophaga from Antarctic Peninsula)
Loza, CM.1, del Corro, ME.1, Soibelzon, LH.1, Negrete, J.2,3, Acosta, W.4, Coria, NR.3, Márquez,
M.E.I.3 Carlini, AA.1
1 División Paleontología Vertebrados, Museo de La Plata., Facultad de Ciencias Naturales y Museo, UNLP, Paseo del Bosque s/nº, La Plata, Bs As, Argentina. 2 CONICET, Av. Rivadavia 1917 (C1033AAJ), CABA, Argentina. 3. Departamento de Biología de Predadores Tope, Coordinación de Ciencias de la Vida, Instituto Antártico Argentino, Cerrito 1248, C1010AAZ-CABA, Argentina 4 Facultad de Ciencias Veterinarias. UNLP Av. 60 s/n, La Plata, Argentina Email: [email protected] INTRODUCCIÓN Los agrupamientos de focas momificadas cangrejeras (Lobodon carcinophaga), fueron hallados en el continente antártico desde hace décadas (Pewé et al 1959, Stirling, et al 1968 Barwick y Balham 1967, Nelson et al 2008, Negrete et al, 2011). Sin embargo, aún hoy no está claro por qué estas focas, típicamente habitantes del pack de hielo marino, son halladas en grandes grupos tierra adentro. Algunas investigaciones postulan que estos animales pudieron haber sufrido de enfermedades que llevaran a su desorientación, o bien que la desorientación que los llevó tierra adentro es consecuencia de la corta edad de los ejemplares que se encuentran habitualmente momificados (Nelson et al 2008). No obstante los individuos de foca cangrejera hallados en el Cabo Bodman, Isla Marambio parecerían ser adultos. Es por ello que el objetivo del presente proyecto es realizar la determinación de las edades absolutas de una muestra de ejemplares recolectados durante la precampaña 2012 de los especímenes de focas cangrejeras momificadas que se encuentran en dicho sitio. Esta información será de suma utilidad al momento de establecer los motivos de dichos varamientos.
Figura 1: Dientes utilizados para la determinación de las edades.
Figure 1. Teeth used for the determination of the ages. MATERIALES Y MÉTODOS El muestreo fue realizado durante la precampaña (Octubre-Noviembre) de 2012 en la isla Marambio, en las inmediaciones del accidente geográfico conocido como cabo Bodman (64º15´S, 56º48´O) en el extremo oeste de la bahía López de Bertodano. Los individuos elegidos para la determinación de edades (n=28) fueron, siempre que se pudo, sexados y diseccionados para obtención de distintos tipos de muestras. Para la determinación de edades se colectaron dientes postcaninos (Fig. 1) que fueron procesados en el laboratorio de la Div. Paleontología de Vertebrados del Museo de la Plata UNLP. Las edades de los individuos se

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
537
obtuvieron mediante la extracción y análisis de un diente siguiendo las pautas de trabajo sugeridas por Laws (1953) y Shiller et al (2004) para cada espécimen a partir del siguiente protocolo: Eliminación de materia orgánica de los dientes mediante hervido en solución de agua y peróxido de hidrógeno 100 volúmenes. Los dientes fueron medidos, fotografiados y se realizó un desgaste del mismo en el plano mesio-distal, mediante una pulidora de grano grueso al agua, y luego se lo pulió por una de grano fino para eliminar rayaduras Una vez pulido, el hemidiente obtenido, fue observado mediante lupa, con luz transmitida, donde se contabilizaron las líneas de dentina y de cemento (Fig. 2)
Figura 2: Corte mesio-distal del postcanino, marcando las líneas de crecimiento.
Figure 2. Mesiodistal cut postcanine, marking the growth lines. RESULTADOS En base a la determinación de edades se pudo constatar que el 82% de los individuos superaban la edad de madurez sexual que en el mayor de los casos en la especie sería de 5 años (Adam 2005). Asimismo, mediante la determinación de sexo realizada a campo se observó que el 80% eran hembras, muchas de las cuales se encontraban preñadas con fetos a término. Las edades obtenidas son presentadas en la Tabla 1 y Figura 3.
ID de la Muestra 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
Sexo H H I H M H H H H M H H H H H H H H I H I M H H H H M M
Edad absoluta 0 3 3 4 4 5 5 5 5 5 6 6 6 6 6 7 7 7 7 8 8 8 10 10 12 15 15 19
Tabla 1: Edades y sexo de los individuos de foca cangrejera momificados estudiados. H: hembra; M: macho; I: sexo indeterminado.
Table 1. Age and sex of mummified crabeater individuals studied. H: female, M: male, I: Sex undetermined.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
538
Figura 3: Representación de clases de edades, representadas en cada uno de los sexos.
Figure 3. Representation of age classes represented in each of the sexes. DISCUSIÓN Los resultados así obtenidos fueron contrastados con la técnica de conteo de anillos externos de cemento en la raíz, verificando que esta última técnica subestima la edad como ya fue observado en otros taxones. Los anillos fueron contados en la base de la corona y en el ápice de las raíces y se pudo determinar que en este último sector los anillos son más abundantes. Dada la particular morfología de la corona de los postcaninos de L. carcinophaga se recomienda realizar el corte en un plano que contenga a la cúspide principal de la corona o desgastar la pieza hasta llegar a ese plano. El conjunto de individuos de Cabo Bodman está representado en su mayoría por focas adultas, muchas de ellas presumiblemente con participación en varias temporadas reproductivas. Este hallazgo es contrario a lo observado en otros varamientos de focas cangrejeras (Stirling and Rudolph 1968, Nelson et al 2008) por lo cual muchas de las explicaciones respecto al motivo de los varamientos planteadas en estos trabajos no pueden ser enteramente aplicadas a este grupo en particular. CONCLUSIONES Dado que la mayoría de los individuos el área de estudio son adultos sexualmente maduros y que además predominan las hembras (muchas de ellas preñadas) existiría una migración conjunta por parte de la especie en etapas previas a la reproducción. A su vez los datos de edades obtenidos descartan las hipótesis planteadas por otros investigadores respecto a que los varamientos son consecuencia de la desorientación de individuos jóvenes. AGRADECIMIENTOS Agradecemos al Sr. Jorge Lusky y a todo el personal del IAA que facilito las tareas de campo. REFERENCIAS BIBLIOGRÁFICAS Adam, PJ. 2005. Lobodon carcinophaga Mammalian species, No. 772, pp. 1–14. Barwick, RE. y Balham, RW. 1967. Mummified Seal Carcases in a Deglaciated Region of South Victoria Land, Antarctica Tuatara 15(3): 165-180. Laws, R.M. 1953. A new method of age determination in mammals with special reference to elephant seal (Mirounga leonina L.) Flalkland Is. Dep. Surv. Sci. Rep. 2:1-111. Negrete, J., Soibelzon, E., Tonni, E.P., Carlini, A., Soibelzon, L.H., Poljak, S., Huarte, R.A. y Carbonari, J.E. 2011. Antarctic radiocarbon reservoir. The case of the mummified crabeater seals Lobodon carcinophaga (Mammalia: Pinnipedia) in Bodman Cape, Seymour Island (Antarctica). Radiocarbon 53, (1): 161-166.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
539
Nelson, AE., Smellie, JL., Williams, M. y Moreton, S. 2008. Age, geographical distribution and taphonomy of an unusual occurrence of mummified crabeater seals on James Ross Island, Antarctic Peninsula. Antarctic Science 20(5):485–93. Shiller, D.M. y Pinedo, M.C. 2004. Growth layer Patterns in Artocephalus australis canine teeth: evaluation of techniques for age determination. LAJAM, 3 (2): 107-108. Stirling, I. y Rudolph ED. 1968. Inland record of a live crabeater seal in Antarctica. Journal of Mammalogy 49: 161–2. Péwé, TL., Rivard, NR. y Llano, GA. 1959. Mummified seal carcasses in the McMurdo Sound Region, Antarctica. Science 30 (3377):716.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
540
VALOR CALÓRICO DE LAS PRINCIPALES PRESAS DE UN PREDADOR GENERALISTA, LA FOCA LEOPARDO HYDRURGA LEPTONYX, Y LOS
REQUERIMIENTOS ENERGÉTICOS DE SUS DIFERENTES GRUPOS ETARIOS
(Caloric value of main prey of a generalist predator, the leopard seal Hydrurga leptonyx and the energetic requirements of their different age groups)
Márquez, M.E.I.1,2, Casaux, R.J.1,3, Baroni, A.V.2, Negrete, J.3 y Mennucci, J.1
1 Departamento de Biología de Predadores Tope, Coordinación de Ciencias de la Vida, Instituto Antártico Argentino. E-mail: [email protected]. 2 Cátedra de Bromatología, Facultad de Farmacia y Bioquímica (UBA). 3 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). INTRODUCCION Estudios desarrollados en diferentes localidades antárticas evidenciaron que la dieta de la foca leopardo Hydrurga leptonyx es diversa y que krill, peces, pingüinos y focas son sus presas principales (Casaux et al. 2009, 2011). La foca leopardo se alimenta frecuentemente en aguas superficiales y su alimento varía con la estación y la localidad (Laws 1984). Este comportamiento podría relacionarse con la estrategia alimentaria generalista del leopardo marino. El conocimiento del valor energético de sus presas es de particular importancia para entender el patrón de alimentación de este predador antártico. OBJETIVO: Determinar el aporte calórico de las principales presas de la foca leopardo con el objetivo de analizar su relación con el requerimiento energético de los ejemplares adultos, juveniles y cachorros de ambos sexos.
MATERIALES Y METODOS Se analizaron las siguientes muestras recolectadas en Caleta Potter (62° 14´S, 58° 40´O), Islas Shetland del Sur (ISS), y en inmediaciones de Punta Cierva (64° 09´S,60° 57´O), Península Antártica (PA), durante el verano, que es un período intensivo de alimentación:. 1) Eufaúsidos: krill antártico Euphausia superba (capturado en el Estrecho de Gerlache, PA, en Marzo de 1976). 2) Peces: ejemplares bentónico-demersales pertenecientes a las familias: Nototheniidae o Bacalaos Antárticos (Gobionotothen gibberifrons, Trematomus newnesi, T. bernacchii, T. hansoni) y Channichthyidae o peces de Hielo (Chaenocephalus aceratus) capturados entre Enero y Marzo de 1999 y de 2002 en ISS, y de 2000 en PA, y un ejemplar de la familia meso-pelágica Myctophidae o peces Linterna (Electrona antarctica), obtenido durante una cesión de alimento del Gaviotín Antártico Sterna vittata. Inmediatamente luego de la captura, los ejemplares fueron medidos y pesados, luego fueron congelados a -20 °C hasta su análisis químico en el laboratorio. 3) Pingüinos: vísceras de pichones de pingüinos Papúa Pygoscelis papua encontrados recientemente muertos en ISS durante los meses de Octubre y Noviembre de 2000. 4) Focas: tejido graso de un cachorro destetado de elefante marino del sur Mirounga leonina encontrado recientemente muerto en el mes de Diciembre de 2001 en ISS. El análisis del contenido de agua de los ejemplares frescos se realizó siguiendo el método AOAC (1980) por deshidratación en estufa a 100 ± 2 °C. Para la determinación de la energía bruta, ejemplares enteros de invertebrados y peces, vísceras de pingüinos y tejido graso de foca, fueron

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
541
secados, molidos y posteriormente sometidos a combustión “in vitro” en presencia de oxígeno mediante una bomba calorimétrica (Miller y Payne 1959). Asumimos una eficiencia digestiva (ED) del 93% para la asimilación de la energía bruta aportada por los peces, pingüinos y focas, y del 86% para los crustáceos (Worthy 2001). RESULTADOS En la Tabla 1 se muestran los valores energéticos obtenidos para cada ítem alimentario. Tabla 1. Tallas (largo total en cm.), pesos (en g.) y contenido calórico (kJ g -1 peso húmedo) de los ejemplares analizados. Table 1. Size, total length (in cm.), weight (in g.) and caloric content (kJ g -1 wet weight) of analyzed specimens.
ESPECIE N TALLA PESO VALOR CALORICO (*)
REFERENCIA (#)
Crustáceos Krill Euphausia superba 4 pools
de ej. 4,5 – 6,0
(ejemplares individuales)
0, 6 – 1,0 (ejemplares individuales)
6,3 ±0,3 Casaux et al. 2009, Rogers et al. 2012
Peces Nototheniidae G. gibberifrons 10 ej. 19,9 – 36,0 50,54 – 466,79 4,4 ±0,5 Casaux et al. 2009,
2011 T. newnesi 10 ej. 17,0 – 21,0 66,41 – 117,10 5,3 ±1,4 Casaux et al. 2011 T. bernacchii 10 ej. 18,0 – 28,0 66,79 – 364,20 4,6 ±0,8 Casaux et al. 2011 T. hansoni 3 ej. 24,9 – 34,0 164,25 – 533,96 5,1 ±0,8 Hall-Aspland &
Rogers 2004 Channichthyidae Oritsland 1977 C. aceratus 6 ej. 49,0 – 58,5 724,33 – 1902,11 4,7 ±0,9 Myctophidae E. antarctica 1 ej. 9,5 6,83 5,6 Casaux et al. 2009,
2011 Pingüinos P. papua pichones (vísceras)
3 ej. n.d. n.d. 4,7 ±0,4 Casaux et al. 2009
Focas Mirounga leonina cachorro destetado (grasa)
1 ej. n.d. n.d. 39,0 Forcada et al. 2009
N= número de ejemplares; n.d.= no determinado. (*) Valores expresados como energía bruta, sin considerar la digestibilidad de cada ítem alimentario. Media ± Desvío estándar. (#) Las referencias bibliográficas citan a cada ítem alimentario y especie como componentes de la dieta de Hydrurga leptonyx. La Tabla 2 muestra el consumo estimado según los datos de los contenidos energéticos de los principales ítems alimentarios, después de ser corregidos de acuerdo a su ED, necesarios para cubrir los requerimientos nutricionales informados por Forcada et al. (2009).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
542
Tabla 2.- Masa corporal (MC) promedio de la foca leopardo, requerimiento energético diario (a) y consumo diario de las presas (b).
Table 2. Mean body mass (MC) of leopard seal, daily energy requirement (a) and daily prey intake (b). Foca leopardo
MC (Kg) Energía diaria (MJ)
Krill antártico (Kg) (1)
Peces mictófidos (Kg) (1)
Pingüino papúa (Kg) (2)
Elefante marino del sur (Kg) (3)
Cachorros (0 año) Hembras 103 29 5,4 5,6 Machos 105 30 5,6 5,8 Juveniles (1 a 5 años) Hembras 196 43 8,0 8,3 9,8 1,2 Machos 227 48 8,9 9,2 10,9 1,3 Adultos Hembras 348 64 11,8 12,3 14,5 1,8 Machos 338 67 12,4 12,9 15,2 1,8 (a) Según los datos informados por Forcada et al. 2009. (b) Se asume que cada especie presa es el único alimento ingerido para cubrir el 100% de los requerimientos energéticos de las focas. (1) Ejemplares enteros; (2) Vísceras; (3) Grasa.
DISCUSION El krill, cuyo contenido calórico es elevado (Tabla 1), es la presa más frecuente y abundante en la dieta de ejemplares juveniles y adultos, seguido en importancia por pingüinos y peces (Casaux et al. 2009). Los peces bentónico-demersales presentaron una baja densidad energética (Tabla 1, Márquez et al. 2010). Casaux et al. (2009, 2011) sugieren que cuando las especies meso-pelágicas (mayormente mictófidos) están presentes en abundancia en el área de forrajeo la foca leopardo preda intensamente sobre ellas en lugar de hacerlo sobre las especies bentónico-demersales debido a su alto valor energético (Tabla 1). Forcada et al. (2009) estimó que el consumo de 4-5 pingüinos cubren el requerimiento energético diario de las focas leopardo. Observaciones realizadas sobre el consumo de pingüinos y otras aves, indican que las presas no son comidas enteras sino desmembradas e ingeridas como pequeñas piezas (Edwards et al. 2010), así como sus vísceras (Hall-Aspland y Rogers 2004). El elevado contenido energético de las vísceras de pingüinos, se justifica por ser éstas un gran reservorio de grasa corporal. La foca leopardo frecuentemente consume focas jóvenes, las que representan un gran aporte energético debido a su tejido graso. La falta de huesos o intestinos encontrados en las heces (Hall-Aspland & Rogers 2007) sugieren que la foca leopardo solo consume su carne, grasa y piel (Bonner 1994). El valor calórico estimado para el elefante marino en este trabajo está muy sobreestimado por considerarse solo el aporte energético de su grasa. El consumo de la presa con músculo y grasa estaría bajando su aporte calórico y debería aumentar la cantidad consumida para cubrir los requerimientos. Asumiendo una tasa de alimentación correspondiente al 7% del peso corporal, Oritsland (1977) estimó que un leopardo marino adulto consume diariamente 20 kg de alimento. La diferencia entre esa y nuestras estimaciones de consumo diario se explicaría porque asumimos que cada especie presa es el único alimento ingerido para cubrir el 100% de los requerimientos energéticos de las focas. Si en las estimaciones se hubiera considerado una dieta mas variada, acorde con la estrategia generalista de este predador, que incluyera otras presas: macroalgas, ascidias y cefalópodos

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
543
(Oritsland 1977, Laws 1984) de menor valor energético (Brown 1989, Márquez et al. Datos no publicados) quizás nuestras estimaciones fueran más cercanas a las de Oritsland (1977). CONCLUSIONES Las presas mayormente consumidas por el leopardo marino son invertebrados y peces pelágicos, pingüinos y cachorros de focas, los que parecen ser seleccionadas positivamente debido a su alto valor energético. El comportamiento alimentario generalista de esta foca, la disponibilidad de presas en las áreas de forrajeo, así como su valor energético y su accesibilidad de acuerdo a la habilidad de buceo de este pinnípedo, condicionan el rol de las diferentes presas en la dieta de la foca leopardo. AGRADECIMIENTOS Dedicado a nuestro amigo y colega Alejandro Carlini. REFERENCIAS BIBLIOGRAFICAS A.O.A.C. 1980. Official Methods of Analysis of the AOAC (Association of Official of Analytical Chemist). 13th. Ed. Washington D.C., USA. Bonner, N. 1994. Seals and sea lions of the world. Cassell plc, Blandford, 149 pp. Brown, C.R. 1989. Energy requirements and food consumption of Eudyptes penguins at the Prince Edward Islands. Antarctic Science, 1(1): 15-21. Casaux, R., Baroni, A., Ramón, A., Carlini, A., Bertolin, M. y DiPrinzio, C.Y. 2009. Diet of the Leopard Seal Hydrurga leptonyx at the Danco Coast, Antarctic Peninsula. Polar Biology, 32: 307-310. Casaux, R., Bertolin, M.L. y Carlini, A. 2011. Feeding habits of three seal species at the Danco Coast, Antarctica: a re-assessment. Polar Biology, 34: 1615-1620. Edwards, E. Forcada, J. y Crossin, G.T. 2010. First documentation of leopard seal predation of South Georgia pintail dick. Polar Biology, 33: 403-405. Forcada, J. Malone, D., Royle, J.A. y Staniland, I.J. 2009. Modelling predation by trnasient leopard seals for an ecosistema-based Management of Southern Ocean fisheries. Ecological Modelling, 220: 1513-1521. Hall-Aspland, S.A. y Rogers, T.L. 2004. Summer diet of leopard seals in Prydz Bay, Eastern Antarctica. Polar Biology, 27(12): 729-734. Hall-Aspland, S.A. y Rogers, T. 2007. Identification of hairs found in leopard seal (Hydrurga leptonyx) scats. Polar Biology, 30: 581-585. Laws, R.M. 1984. Seals. En: Laws, R.M., ed. Antarctic ecology, Vol. II, London: Academic Press, pp. 621-715. Márquez, M., Casaux, R.J., Baroni, A.V., Bertolin, M.L. y Carlini, A.R. 2010. Caloric content of fish preys of Antarctic pinnipeds. XXXI SCAR y Open Science Conference. Buenos Aires, Argentina. 3-6 de Agosto de 2010. Resumen, Nro. 482, 1 pp.. Miller, D.S. y Payne, P.R. 1959. A balistic bomb calorimeter. British Journal of Nutrition, 13: 501. Oritsland. T. 1977. Food consumption of seals in the Antarctic pack ice. En: Adaptions within Antarctic Ecosystems, ed. Llano, G.A. Proceedings of the 3rd SCAR Symposium on Antarctic Biology, pp. 749-768, Smithsonian Institution, Washington D.C., USA. Rogers, T., Ciaglia, A.M., O¨Connell, T.C., Slip, D., Meade, J., Carlini, A.R. y Márquez, M.E.I.. 2012. WAP Antarctic top predator behaves differently whiskers reveal WAP leopard seals are krill-feeding specialists. XXXII SCAR (Comité Científico de Investigaciones Antárticas) Open Science Conference, Portland, Oregon, U.S.A. 16-19 de Julio de 2012. Resumen en CD-ROM Worthy, G. 2001. Nutrition and energetics. En: Dierauf, L.A., Gulland, F.M.D. (Eds.), CRC Handbook of Marine Mammal Medicine, 2nd ed. CRC Press, Boca Raton, Florida, USA, pp. 791-827.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
544
VARIACIÓN EN LA DISTRIBUCIÓN DE MEIOFAUNA INTERMAREAL, UNA COMPARACIÓN ENTRE LA ANTÁRTICA Y LA REGIÓN DE MAGALLANES
(Variation in meiofaunal distribution in intertidal beaches, a comparison between Antarctica and the region of Magallanes)
Olivares, B.1 y M.R. Lee1
1Centro i~mar, Universidad de Los Lagos, Puerto Montt, Chile. E-mail: [email protected] INTRODUCCIÓN La meiofauna es un componente importante del bentos marino, involucrados en el reciclaje de material orgánico (Nascimento et al. 2012). Sin embargo, los estudios sobre la meiofauna y la fauna bentónica de pequeños metazoos en la Antártica son escasos (véase Giere, 2009 para una revisión). La poca información de los aspectos biológicos y ecológicos básicos, tales como su diversidad y distribución, han limitado por muchos años el desarrollo del conocimiento de las especies de meiofauna; una razón principal es la falta de taxónomos expertos en los diferentes grupos de la meiofauna. Otros aspectos importantes que están relacionados con la abundancia y distribución de meiofauna son la temperatura, granulometría, salinidad, saturación de la disponibilidad de oxígeno, porosidad del sustrato, y el agua, se cree que estos son los principales factores abióticos que afectan a los patrones de distribución a gran escala de meiofauna (Giere 2009). Por ello se investigó la abundancia, diversidad y ubicación de meiofauna en Antártica y el continente de América del Sur, en relación con la temperatura, materia orgánica, y la productividad bentónica. Es probable que la temperatura influya la riqueza de especies, debido a que cualquier especie puede ser incapaz de habitar en un sitio por su temperatura. Temperaturas bajas retardan el ciclo reproductivo en tal medida que una población determinada se vuelve vulnerable a la extinción local. Esta conocido que nematodos, por ejemplo, crecen más lentos en temperaturas más bajas (Van Voorhies 1996). En este caso, la resistencia fisiológica debe ser un factor determinante importante en la distribución en el sedimento. Johnson (1965) sugiere que los animales que viven en los centímetros superiores de una playa de arena fina están expuestos a una temperatura diaria que es hasta tres veces mayor que la que experimentan los individuos submareales, mientras que las especies que viven a 10 cm de profundidad o más, experimentan temperaturas comparables a las de los ambientes submareales. Hay evidencia de que muchas especies no se restringen a hábitats específicos o rangos de profundidad. Especies intermareales también se pueden encontrar en aguas más profundas y especies que se registran primero en sustratos de algas después de un muestreo adicional también aparecen en las muestras de sedimento (M. Lee, comunicación personal). El presente estudio tiene como objetivo verificar si existen diferencias en la distribución de meiofauna en el continente Sudamericano en comparación con Antártica y si esta respuesta está afectada por la temperatura y/o otras variables ambientales que existen en cada continente. Se tomaron muestras en dos playas de antártica y dos de la región de Magallanes para verificar si existe diferencia en la abundancia y diversidad en las especies de meiofauna y si esta discrepancia es por la diferencia de temperatura que hay entre antártica y el continente sudamericano. MATERIALES Y MÉTODOS Se estudió la abundancia, diversidad y ubicación de meiofauna en 4 playas, dos de Antártica en la localidad de Isla Livingstone, playa Punta Hannah (62°39'0.14"S; 60°36'15.08"W) e Isla Rey Jorge,

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
545
playa Elefantera (62°11'52.23"S; 58°59'40.28"W) y se compararon con dos playas de la región de Magallanes, playa Tres Puentes (53°08'3.7"S;70°51'37.3"O) y la Desembocadura de Rio San Juan (53°38'17.5"S; 70°56'45.9"O); los resultados de este último sitio no están considerados en este estudio, pero serán presentados en el congreso. Se evaluó el intermareal de cada playa y se dividió en tres zonas de muestreo (secado, retención y resurgencia), los muestreos se llevaron a cabo en dos distintas profundidades (superficie y profundidad). Se tomaron 30 muestras por playa con una jeringa de 50 ml modificado para formar una corer de tipo pistón. Se sacaron 5 replicas con una caminata al azar con marea baja en cada zona de muestreo, se colocaron en frascos plásticos de 50 ml con tapa rotatoria, y fueron fijados con formalina al 5 %. En el laboratorio se realizó extracción de meiofauna por el Método de Decantación simple, y para extraer los múltiples componentes del sedimento y obtener solo la muestra original de meiofauna utilizamos extracción de meiofauna con Ludox; para mayor detalle de los métodos véase en (http://meiochile.matthewlee.org). El conteo para determinar la abundancia y diversidad de meiofauna se realizó en un microscopio Olympus BX43.
RESULTADOS
Figura 1. Abundancia de meiofauna total en el intermareal en relación con la temperatura en Punta Arenas (superficie, 30 cm profundidad). PAS = Punta Arenas, Alto-Superficie; PAP = Punta Arenas, Alto-Profundidad; PMS = Punta
Arenas, Medio-Superficie; PMP = Punta Arenas, Medio-Profundidad; PBS = Punta Arenas, Bajo-Superficie; PBP = Punta Arenas, Bajo-Profundidad.
Figure 1. Abundance of intertidal meiofauna in relation to the temperature in Punta Arenas (surface and 30cm deep). PAS = Punta Arenas, High-Surface; PAP = Punta Arenas, High-Deep; PMS= Punta Arenas, Mid-Surface; PMP= Punta
Arenas, Mid-Deep; PBS= Punta Arenas, Low-Surface; PBP= Punta Arenas, Low-Deep.
Se realizó un análisis de regresión para las playas de Antártica y Punta Arenas, entre la abundancia de meiofauna total y la temperatura de cada playa que dio como resultado la ausencia de correlación para ambas playas de Antártica, playa Punta Hannah con un R2=0,0348 y un p=0,8778, playa Elefantera con un R2=0,0209 y un p=0,5292; sin embargo, para la playa de Punta Arenas, existe significancia entre las correlación de las variables con un R2=0,2132 y un p=0,0059, con una reducción en la abundancia y aumentos en temperatura. El análisis MDS da como resultado dos grupos distintos de similaridad, con un 2D Estrés=0,05, que agrupa las playas de Antártica, difiriendo de la playa de Punta Arenas.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
546
Figura 2 Abundancia de meiofauna total en el intermareal en relación con la temperatura en Punta Hannah (superficie, 30 cm profundidad). HAS = Punta Hannah, Alto-Superficie; HAP = Punta Hannah, Alto-Profundidad; HMS = Punta
Hannah, Medio-Superficie; HMP = Punta Hannah, Medio-Profundidad; HBS = Punta Hannah, Bajo-Superficie; HBP = Punta Hannah, Bajo-Profundidad.
Figure 2. Abundance of intertidal meiofauna in relation to the temperature in Punta Hannah (surface and 30cm deep). HAS = Punta Hannah, High-Surface; HAP = Punta Hannah, High-Deep; HMS= Punta Hannah, Mid-Surface; HMP=
Punta Hannah, Mid-Deep; HBS= Punta Hannah, Low-Surface; HBP= Punta Hannah, Low-Deep.
Figura 3. Abundancia de meiofauna total en el intermareal en relación con la temperatura en Playa Elefantera (superficie, 30 cm profundidad). EAS = Elefantera, Alto-Superficie; EAP = Elefantera, Alto-Profundidad; EMS = Elefantera, Medio-Superficie; EMP = Elefantera, Medio-Profundidad; EBS = Elefantera, Bajo-Superficie; EBP =
Elefantera, Bajo-Profundidad. Figure 3. Abundance of intertidal meiofauna in relation to the temperature in Playa Elefantera (surface and 30cm deep).
EAS = Playa Elefantera, High-Surface; EAP = Playa Elefantera, High-Deep; EMS= Playa Elefantera, Mid-Surface; EMP= Playa Elefantera, Mid-Deep; EBS= Playa Elefantera, Low-Surface; EBP= Playa Elefantera, Low-Deep.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
547
CONCLUSIONES Los gráficos (Fig. 1, Fig. 2, Fig. 3) muestran la variable abundancia total de meiofauna y temperatura de los sitios de estudio en las playas. La cantidad de abundancia que encontramos en las playas de Antártica no se relaciona con la temperatura, sin embargo en la playa de Punta Arenas existe una leve correlación, pero esto no indica que la variable temperatura determine la abundancia de meiofauna, estas conductas se pueden explicar por el estrés térmico que se presenta en el intermareal en la parte alta de la playa de Punta Arenas. En el caso de Antártica la distribución de la meiofauna es más homogénea, y sus distribuciones pueden ser relacionadas con otras variables como la productividad bentónica, la sedimentología y también la presencia y cantidad de materia orgánica. La materia orgánica puede ser responsable para el peak en la abundancia de meiofauna en la parte alta de la playa Elefantera (Fig. 3), donde se encontró gran cantidad de algas en el sedimento al momento de realizar el muestreo, pero estas variables son tema de estudio. AGRADECIMIENTOS Financiamiento Proyecto INACH T_13-10. REFERENCIAS Giere, O. 2009. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments. 2nd Edition. Springer-Verlag, Berlin, Germany. 527pp. Hulings, N. y Gray, J. 1971. A Manual For The Study Of Meiofauna. Smithsonian Contributions To Zoology, 78, 1- 104. Johnson, R. 1965. Temperature variation in the infaunal environment of a sand flat. Limnology and Oceanography, 10, 114-120. Lee, M. y Riveros, M. 2012. Latitudinal trends in the species richness of free-living marine nematode assemblages from exposed sandy beaches along the coast of Chile (18–42° S). Marine Ecology, 33, 317-325. Nascimento, F., Naslund, J. y Elmgren, R. 2012. Meiofauna enhances organic matter mineralization in soft sediment ecosystems- Limnology and Oceanography, 57, 338-346. Van Voorhies, W. 1996. Bergmann size clines: A simple explanation of their occurrence in ectotherms. Evolution, 50, 1259-1246.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
548
SOBRE LA NECESIDAD DE ACTUALIZAR LAS DISTRIBUCIONES DE LAS AVES MARINAS PELÁGICAS EN OCÉANO ATLÁNTICO SUR Y ANTÁRTIDA
(The need to update the seabird pelagic distribution in South Atlantic Ocean and Antarctica)
Orgeira, J. L.
Departamento Biología de Predadores Tope, Instituto Antártico Argentino. Cátedra Ecología General, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. Email: [email protected]
INTRODUCCIÓN. Durante más de 30 años diferentes especies de aves marinas pelágicas han sido registradas en áreas geográficas que no corresponden con la distribución dada en la literatura corriente. Estos registros pueden ubicarse en tres clases diferentes: i) apariciones ocasionales debidas a la intervención de fuertes vientos, siendo ésta la explicación más frecuente en la literatura (Lazo y Yánez 1989, Parmelee & Fraser 1989, entre otros); ii) cambios ambientales a nivel regional como fluctuaciones térmicas o prolongadas sequías, presionando a algunas especies a migrar hacia ecosistemas diferentes, como la presencia del Cisne de cuello negro en Antártida (Bennett 1922, Orgeira & Fogliatto 1991, Aguirre 1995); iii) conducta migratoria y colonizadora de la especie, como la Garcita bueyera en el Océano Atlántico Sur y Antártida (Prince & Payne 1979, Prince & Croxall 1983, Lange & Nauman 1990). Otro grupo de especies de aves marinas pelágicas también ha sido registrado en regiones geográficas que no coinciden con las citadas en la literatura. Esto sucede, presumiblemente, porque de igual forma son consideradas como apariciones extralimitales ocasionales. Sin embargo y a diferencia de los ejemplos citados arriba, han sido observadas en forma recurrente en las mismas áreas, en la misma época del año y bajo las mismas condiciones ambientales. El objetivo de este trabajo es determinar si estos registros se tratan de apariciones ocasionales o si corresponden a los verdaderos rangos de distribución de las especies, los cuales no se han actualizado. Parte de la información presentada ya ha sido publicada; se presentan aquí nuevas observaciones, inéditas, obtenidas en la temporada de verano 2013 que completan las precedentes. MATERIALES Y MÉTODOS. Los datos provienen de nueve temporadas de verano tomados a bordo del rompehielos Almirante Irizar (temporadas de verano 1990, 1991, 1993 y 1995) y del buque de investigación oceanográfico Puerto Deseado (2009 a 2013) siguiendo la metodología de censos de aves marinas de 10 minutos a bordo de buques (SCAR 1981). El área de estudio cubierta fue Pasaje de Drake, norte de la península antártica, Mar del Scotia y norte del Mar de Weddell. Diferentes variables ambientales fueron registradas al momento del censo: temperatura superficial del mar (ºC), intensidad y dirección del viento (en escala Beaufort y en grados respectivamente) y cobertura de hielo (%). RESULTADOS. Listado de especies. Petrel Atlántico o Petrel cabeza parda Pterodroma incerta. La distribución dada para esta especie es el Océano Atlántico Sur al norte de los 50ºS (Onley & Scofield 2007). El primer registro en Antártida fue reportado por Brown et al. (1975), quien observó cinco individuos al sur de los 61°S. A partir de entonces lo hemos observado en las

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
549
temporadas de verano 1990, 1991, 1993, 1994, 2009, 2011, 2012 y 2013 en el Pasaje de Drake, Islas Shetland del Sur, Is. Orcadas del Sur y Mar de Weddell (Orgeira et al. 2013, Tabla 1).
Especie Área Fecha Abundancia Petrel Atlántico Mar de Weddell Marzo 1990 5 Mar del Scotia Marzo 1991 5 Pasaje de Drake Marzo 1993 3 Mar del Scotia Diciembre 1994 14 Islas Shetland del Sur Marzo 2009 24 Pasaje de Drake Marzo 2009 3 Pasaje de Drake Enero 2011 4 Pasaje de Drake Febrero 2011 3 Pasaje de Drake Febrero 2012 9 Pasaje de Drake Marzo 2013 4 Petrel de collar gris Pasaje de Drake Marzo 1990 65 Pasaje de Drake Febrero 2010 17 Pasaje de Drake Marzo 2010 7 Pasaje de Drake Febrero 2011 26 Pasaje de Drake Marzo 2011 21 Pasaje de Drake Febrero 2012 4 Petrel de alas grandes Mar de Weddell Enero 1994 59 Mar de Weddell Febrero 1994 6 Mar del Scotia Enero 1996 47 Mar del Scotia Enero 2012 3 Petrel de Kerguelen Pasaje de Drake Febrero 1993 1 Islas Shetland del Sur Marzo 1993 5 Pasaje de Drake Marzo 1993 6 Tierra del Fuego Noviembre 1993 1 Mar de Weddell Diciembre 1994 14 Mar de Weddell Enero 1995 268 Pasaje de Drake Marzo 1995 4 Pasaje de Drake Febrero 1996 1 Islas Shetland del Sur Febrero 2010 1 Islas Shetland del Sur Marzo 2012 2
Tabla1. Abundancias de las cuatro especies tratadas, discriminadas por año y regiones geográficas. Table 1. Abundance of studied species by year and by geographical regions.
Figura 1. Distribución geográfica de las especies tratadas. Los puntos negros indican la posición geográfica de cada
conteo de 10 minutos donde las aves fueron registradas en las temporadas de verano 1990, 1991, 1993, 1995 y 2009 a 2013. Las áreas grises indican la distribución dada en la bibliografía citada.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
550
Figure 1. Geographical distribution of studied species. Black dots indicate geographical position of 10 min counts where birds were recorded in summer seasons 1990, 1991, 1993, 1995 and 2009 to 2013. Grey areas indicate the
distribution given in the literature cited. Petrel de collar gris Pterodroma mollis. Según Rodríguez Mata et al. (2006) y Onley & Scofield (2007) se distribuye en el Océano Atlántico Sur al norte de los 50ºS. Nuestros registros más importantes corresponden a marzo de 1990, cuando fueron registrados 65 individuos en el Pasaje de Drake, incluida una bandada de 15 individuos. La especie fue registrada nuevamente en el Pasaje de Drake en las temporadas de verano de 2010, 2011 y 2012. Petrel de alas grandes P. macroptera. La distribución conocida en el Océano Atlántico Sur es al norte de los 50ºS aproximadamente. En enero y febrero de 1994 registramos 65 individuos en el centro del Mar de Weddell, algunos de ellos al sur del Círculo Polar Antártico, constituyendo los únicos registros conocidos para esta especie en el Mar de Weddell. En enero de 1996 registramos numerosas bandadas al norte del Mar del Scotia. Otros individuos fueron observados en enero de 2012, esta vez en los alrededores de las Islas Shetland del Sur y de las Is. Orcadas del Sur. Petrel de Kerguelen Lugensa brevirostris. La distribución de esta especie es contradictoria en la literatura. Según Harrison (1987), del Hoyo et al (1992) y Onley & Scofield (2007) es circumpolar frecuentando los bordes de campo de hielo hasta los 40ºS, mientras que para Rodríguez Mata et al. (2006) su límite austral en el Océano Atlántico Sur es el paralelo 37ºS. Nuestros registros no sólo confirman su presencia en aguas antárticas y subantárticas sino que lo extienden: entre diciembre de 1994 y marzo de 1995 registramos en el Mar de Weddell 286 individuos (incluyendo numerosas bandadas) y el registro más austral obtenido para la especie, al sur de los 62ºS. La mayoría de estos registros ocurrieron en presencia de hielo. DISCUSIÓN. Los programas plurianuales de monitoreo de aves marinas a bordo de buques cubren extensas áreas geográficas con la obvia ventaja de acumular información de diferentes ecosistemas marinos. Esta información nos permite diferenciar entre las especies cuya presencia es ocasional y las recurrentes. Las cuatro especies citadas en este trabajo han sido registradas reiteradamente desde la década de 1990. Algunas de ellas, como el Petrel Atlántico, están siendo observadas en las mismas regiones antárticas donde fueron registradas por primera vez hace casi 40 años. Lo interesante de estos registros es que han ocurrido independientemente de los cambios en las condiciones ambientales; es decir, de fluctuaciones climáticas (como la gran sequía regional de fines de 1988 o los incrementos térmicos de los años 2008 a 2012). Ello sugiere que su presencia en Antártida y en el extremo sur del Atlántico Sur no está determinada por presiones exógenas y que estas áreas constituyen sus rangos de distribución efectivos. No obstante ello, la distribución geográfica de estas cuatro especies no ha sido actualizada en la literatura. Una distribución incompleta significa desconocer las tolerancias fisiológicas de la especie, la amplitud de su nicho trófico, sus interacciones con otras especies y con el ecosistema que habitan; todos motivos que justifican la necesidad de una inmediata actualización.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
551
AGRADECIMIENTOS. A la Armada Argentina y las tripulaciones de los buques Almirante Irizar y Puerto Deseado. Este trabajo fue realizado con el soporte logístico y financiero del Instituto Antártico Argentino. REFERENCIAS BIBLIOGRAFICAS. Aguirre, C.A. 1995. Distribution and abundance of birds at Potter Peninsula, 25 de Mayo (King George) Island, South Shetland Islands, Antarctica. Mar. Ornithol., 23: 23-31. Bennett, A.G. 1922. Notas sobre aves subantárticas. Hornero, 2: 255-258. Brown, R.G.B., Cooke, F., Kinnear, P.K. and Mills, E.L. 1975. Summer seabird distributions in Drake Passage, the Chilean fjords and off Southern South America. Ibis Vol. 117: 339-356. Harrison, P.T. 1987. A field guide to Seabirds of the world. Lexington: Stephen Green Press. Lange, U. and Naymann, J. 1990. Weitere erstnachweise von Vogelarten im Sudwesten von King George Island (Sudshetland-Inseln, Antarktis) — Beitr. Vogelkd. 36: 165-170. Lazo, I.F. and Yanez, J. 1989. First record of black-necked swan Cygnus melanocoryphus in South Shetland and Antarctica. Polar Record 25: 354. Onley, D. and Scofield, P. 2007. Albatrosses, Petrels & Shearwaters of the world. (Princeton Field Guides. Princeton University Press). Orgeira, J.L. y Fogliatto, O.N. 1991. Presence of the Blacknecked Swan Cygnus melanocoryphus in Antarctica. Mar. Ornithol., 19: 140-143. Orgeira, J.L., Scioscia, G., Torres, M.A. and Dellabianca, N.A. 2013. New at-sea records of pelagic seabirds in the South Atlantic Ocean and Antarctica. Polar Research 32, 18972, http://dx.doi.org/10.3402/polar.v32i0.18972. Parmelee, D.F. and Fraser, W.R. 1989. Multiple sightings of Black-necked Swans in Antarctica. — American Birds, Bird Count., 43: 1231-1232. Prince, P.A. and Payne, M.R. 1979. Current status of birds at South Georgia. Br. Antarct. Surv. Bull., 48: 103-118. Prince P. A. and Croxall J.P. 1983. Birds of South Georgia: new records and re-evaluation of status. Br. Antarct. Surv. Bull., 59: 15-27 Rodríguez Mata, J., Erize, F. and Rumboll, M. 2006. Aves de Sudamérica. Guía de Campo Collins. (Birds of South America. Collins Field Guide). Buenos Aires: Editorial Letemendía, Harper Collins Publisher. S.C.A.R. 1981. BIOMASS Seabird Ecology. Post FIBEX Data Workshop. BIOMASS Report Series, 22.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
552
LIQUENOBIOTA EN LA ESTACIÓN CIENTÍFICA ANTÁRTICA MACHU PICCHU
(Lichen biota at the Antarctic research station Machu Picchu)
Ramírez, A.1, Bello, C.2, Domínguez, N.2 y P. Gallegos3
1Laboratorio de Florística, Departamento de Dicotiledóneas, Museo de Historia Natural, Universidad Nacional Mayor de San Marcos. Email. [email protected] 2Dirección Antártica-Ministerio de Relaciones Exteriores del Perú 3Universidad Científica del Sur-Perú
Los líquenes son organismos simbiontes que soportan condiciones extremas del clima como temperatura y radiación, entre otras; por ende son de interés de estudio para entender las variaciones climáticas. Con la finalidad de conocer la diversidad liquénica para luego correlacionar el comportamiento de la liquenobiota frente a los parámetros ambientales y así poder elaborar modelos predictivos sobre el cambio climático, se decidió estudiar a estos organismos en la Estación Científica Antártica Machu Picchu (ECAMP), la cual esta dentro del South Shetland Island. En tal sentido se realizaron colectas en el ECAMP durante el verano austral del 2011-2012 y del 2012-2013; se llevó a cabo un muestro representativo, colocando una grilla imaginaria en el área de estudio con más de 100 puntos de evaluación, separados entre sí por 50 m; las muestras fueron fotografiadas y colectadas depositándolas en sobres de papel con sus datos respectivos para luego ser trasladas al territorio peruano (Perú); paralelamente, se instalaron cuadrantes de 50 cm x 50 cm sobres las rocas, a las cuales se fotografiaron para estudiar el crecimiento de los líquenes. Las muestras de líquenes fueron analizadas en el laboratorio mediante equipo especializado para conocer sus características; se consultó literatura pertinente y a especialistas en el tema. La diversidad liquénica preliminar en el ECAMP es de 49 especies, conformadas por 16 familias y 22 géneros; entre las familias representativas están: Acarosporaceae, Agyriaceae, Cladoniaceae, Collemataceae, Lecanoraceae, Parmeliaceae, Rhizocarpaceae, Stereocaulaceae, Teloschistaceae, Umbilicariaceae, la especie predominante fue Usnea antarctica (Parmeliaceae) y la familia más

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
553
abundante fue Lecanoraceae con seis especies; entre los líquenes que se sugieren para estudiar el cambio climático se propone a Rhizocarpon geographicum por se un liquen asequible medir su tamaño en área y a Usnea antarctica por poseer bandas negras para protegerse de la radiación y por la producción de acido úsnico frente al UV.
Placopsis antarctica Rhizocarpon geographicum
Rhizoplaca aspidophora Usnea aurantiaco-atra
REFERENCIAS Hooker Y., J. Amanzo, J. Pérez, D. Schories y J. Zavala. 2003. Biodiversidad submarina y terrestre de la ensenada Mackellar (Isla Rey Jorge-Antártida). Antar XIV. Informe del Instituto del Mar de Perú. Olech, M. 2004. Lichens of King George Island Antarctica. The institute of Botany of the Jagiellonian University pp. 391. Kraków. 391 pag. Quilot, W., Fernandez, E., Rubio, C., Goddard, M. and Hidalgo, E. 1998. In M.P. Marcelli & M.R.D. Seaward (Eds). Lichenology in Latin America: history current knowledge and applications. P. 171-179. CETESB. Sao Paulo. Spielmann, A. and Batista, A. 2012. Lichens and the Maritime Antartica. Revista Electrónica del Grupo Latinoamericano de Liquenologos. Vol 4(3):33. Wirtz, N., Printzen, C., Sancho, L. and Lumbsch, T. 2006. The phylogeny and classificaction of Neuropogon and Usnea (Parmeliaceae, Ascomycota) revisited. Taxon 55(2). Mayo 2006:367-376.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
554
DISTRIBUCIÓN Y DENSIDAD INCIDENTAL DE AVES MARINAS EN UN CRUCERO DEL ATLÁNTICO SUR OCÉANO ANTÁRTICO
(Distribution and density of seabirds counting in a cruise from South Atlantic to Antarctic Ocean)
Silva, M.1, Costa, E.S. 2,3
1Universidade Federal Fluminense. Curso de Graduação em Geografia. 2Universidade Federal do Rio de Janeiro (UFRJ), Laboratório de Biofísica, Ilha do Fundão, Rio de Janeiro, RJ, Brasil. CEP: 21941-540. 3Laboratório de Ecologia de Aves, Departamento de Ecologia, Instituto de Biologia Roberto Alcantara Gomes, Universidade do Estado do Rio de Janeiro, RJ. CEP 20550-011. The census of marine birds is important tools to assess distribution and movement patterns of these organisms, help us analyzing and understanding the distribution and abundance of this group. By 03 from 10 of December 2012, during the cruise in the Oceanographic vessel Ary Rongel, between Rio de Janeiro (Brazil, Atlantic Ocean) to South Shetland Islands (Antarctic Ocean) the census was conducted with duration of 10 minutes every two hours while the brightness allowed the identification of seabirds. We performed 51 censuses during the travel and counted 3.119 seabirds, classified in 20 species. Maps of the distribution, abundance and richness were obtained by grouping the data by degree of latitude. Here we present the maps obtained with censuses data and discuss distribution and species richness patterns according to environmental variables.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
555
ANÁLISIS PRELIMINAR DE LA DIVERSIDAD GENETICA Y FILOGEOGRAFÍA DEL CARACOL ENDÉMICO NEOBUCCINUM EATONI EN EL OCÉANO AUSTRAL
(Preliminary analysis of diversity genetics and phylogeography of the endemic snail Neobuccinum eatoni in the southern ocean)
Díaz, A.1, 3, Poulin E.2,3, Mansilla, A.1,3
1 Laboratorio de Macroalgas Antárticas y Subantárticas, Instituto de la Patagonia, Universidad de Magallanes, Avenida Bulnes # 01890, Punta Arenas, Chile. E-mail: [email protected], [email protected] 2 Laboratorio de Ecología Molecular, Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras # 3425, Ñuñoa, Santiago, Chile. E-mail: [email protected] 3 Instituto de Ecología y Biodiversidad (IEB), Facultad de Ciencias, Universidad de Chile, Las Palmeras # 3425, Ñuñoa, Santiago, Chile. RESUMEN El océano Austral que rodea a Antártica es considerado como uno de los ecosistemas marinos más grandes y bien delimitados de la tierra. Al sur por el continente antártico y al norte por la zona del frente polar (ZFP o convergencia antártica). Este ecosistema alberga a una particular macrofauna bentónica que se caracteriza por su alta diversidad, incluyendo a un número de especies que puede exceder las 15000. El origen de estas especies se establece desde el aislamiento que experimentó Antártica hace aproximadamente 30 Ma., junto con el posterior establecimiento de importantes barreras alrededor del continente: Oceanográfica (Corriente circumpolar Antártica), Geográfica y Batimétrica. Estas barreras promovieron la evolución y la diversificación de una rica fauna a lo largo de la plataforma continental y del talud que rodea al continente. Sin embargo, debido a distintos periodos de cambios en la CCA, la fauna presente hoy está compuesta por una mezcla de taxa con diferentes afinidades biogeográficas. Entre las hipótesis que explican el origen de dicha fauna se cuentan: (1) Fauna relicta autóctona; (2) Fauna derivada proveniente de las zonas profundas adyacentes; (3) Fauna que ha dispersado desde Sudamérica por el Arco de Escocia y; (4) Fauna Antártica que ha avanzado hacia zonas norte por el Arco de Escocia. Recientes estudios apoyan la tesis de que la biogeografía de varias especies del OA, en particular de aquellas que presentan una elevada capacidad dispersiva, está influenciada por eventos de dispersión mucho más recientes que la historia tectónica del continente Antártico. En particular, un reciente estudio sobre relaciones evolutivas y procesos de diversificación en las zonas profundas y someras del Océano Austral, realizó un importante aporte para el entendimiento del origen de la fauna de invertebrados marinos someros, al determinar que las diversas poblaciones alrededor del OA del erizo Sterechinus neumayeri, muestran señales genéticas de una expansión poblacional reciente desde un único refugio fuera del margen del continente (probablemente una isla del Arco de Escocia), con posteriores eventos de recolonización. Sin embargo, el escenario biogeográfico histórico sobre el origen de la fauna bentónica sigue incompleto al desconocer lo que ocurre con especies que no presentan un estado larval de vida libre y poseen una amplia distribución batimétrica. Entre los moluscos que forman parte de la fauna bentónica marina de Antártica, la especie monotípica Neobuccinum eatoni (Smith, 1875), presenta características que la hacen un modelo de estudio adecuado para investigar y completar el escenario biogeográfico de cómo los procesos climáticos y oceanográficos históricos pueden haber influenciado la historia evolutiva de la fauna marina bentónica del OA. Su distribución es circumantártica en el OA y en la provincia subantártica, en el Archipiélago de Kerguelen (Figura 1), presenta una importante euribatia y posee

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
556
un desarrollo intracapsular prolongado de aproximadamente 15 meses, del que salen juveniles (Hain y Arnaud 1992).
Figura 1: Distribución de la especie Neobuccinum eatoni (Smith, 1875), en el Océano Austral.
Figure 1: Distribution of the species Neobuccinum eatoni (Smith, 1875), in the Southern Ocean. La obtención del material biológico analizado fue realizada mediante buceo autónomo en dos sitios de la Península Antártica, Bahía Fildes (Isla Rey Jorge) y Base Rothera (Isla Adelaida). La preparación del material genético a partir de las muestras fijadas en etanol (95%) se realizó siguiendo el protocolo del método de sales (Aljanabi & Martínez 1997), mientras que la amplificación parcial por PCR del gen mitocondrial COI se realizó con partidores universales (LCO1490 y HCO2198, Folmer et al. 1994). Para las muestras obtenidas en Fildes se determinaron la diversidad haplotípica (h), el número de sitios polimórficos (S), y el número promedio de diferencia entre pares de secuencias (π), con el programa DnaSP (Rozas et al. 2003). Se estimaron además las relaciones genealógicas entre los distintos haplotipos de distintas regiones biogeográficas mediante el programa Network 4.1.5 (Röhl 2002), incluidas secuencias obtenidas desde GENBANK, que representan a otros dos sitios alrededor del continente (Figura 2). La amplificación parcial del gen citocromo Oxidasa I obtenida fue de 627pb para un total de 6 individuos en Bahía Fildes y 1 para Base Rothera. Los resultados preliminares para Bahía Fildes indican que existe una elevada diversidad de haplotipos (H= 0.867) con elevado número de sitios polimórficos (S= 29), pero con un bajo número de haplotipos (K= 4). La red de haplotipos da cuenta de la alta diversidad, al observar la distancia en pasos mutacionales que separan a dichos haplotipos (Figura 3a).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
557
Figura 2: Distribución de la muestras analizadas. Bahía Fildes (1), Base Rothera (2) y dos localidades obtenidas desde
GENABNK, Mar de Ross (3) y Elizabeth Land (4). Figure 2: Distribution of the analyzed samples. Fildes Bay (1), Rothera Base (2) and two localities obtained from
GENBANK, Ross Sea (3) and Elizabeth Land (4). Es interesante el escenario que se observa al incluir la muestra obtenida en la Base Rothera a dicha red de haplotipos, ya que esta muestra se sitúa al medio sin relación aparente con la distribución geográfica de las muestras, con una distancia aproximada de 730 km entre ambos sitios (Figura 3b).
Figura 3: Red de haplotipos para las muestras obtenidas en Bahía Fildes (a) y Bahía Fildes/Base Rothera (b). Figure 3: Haplotypes network for the obtained samples from Fildes Bay (a) and Fildes Bay/Rothera Base (b). Aún más interesante es observar cómo se sitúan en la red de haplotipos las secuencias obtenidas en GENBANK, provenientes de Mar de Ross y Elizabeth Land (Figura 4), ya que se relacionan con las muestras analizadas pero no es posible establecer un patrón filogeográfico claro.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
558
Figura 4: Red de haplotipos para las muestras de N. eatoni obtenidas de todos los sitios analizados. Bahía Fildes (1), Base Rothera (2), Mar de Ross (3) y Elizabeth Land (4). Figure 4: Haplotypes network for samples of N. eatoni obtained from all the analyzed sites. Fildes Bay (1), Rothera Base (2), Ross sea (3) and Elizabeth Land (4). Los resultados preliminares para la especie en estudio indican que en bahía Fildes existe una mayor diversidad genética que lamentablemente no se puede comparar con ningún otro sitio ya que tanto en Base Rothera y Mar de Ross se tienen solamente un individuo, mientras que las cuatro secuencias analizadas para Elizabeth Land presentan el mismo haplotipo (no hay diversidad genética). No obstante, debido a que son resultados muy recientes con pocas muestras analizadas, es altamente probable que la diversidad genética que al parecer presenta N. eatoni (antes de descartar un importante cuello de botella) aún no es analizada, por lo que faltan individuos por analizar y sitios intermedios por muestrear. AGRADECIMIENTOS FONDECYT 3130677, Instituto de Ecología y Biodiversidad (IEB), proyecto ICM P05-002 y PFB – 23. BIBLIOGRAFÍA Aljanabi, S.M. y I. Martínez. 1997. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Research 25: 4692-4693. Folmer, O., Black, M., Hoeh, W., Lutz, R., Vrijenhoek, R. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology & biotechnology, 3, 294–299 Hain, S. y P.M. Arnaud. 1992. Notes on the reproduction of high-Antarctic molluscs from the Weddell Sea. Polar Biology 12: 303-312. Röhl, A. 2002. Network, netw3111.exe downloaded in the web site: www.fluxusengineering

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
559
PATRONES DE DIVERSIDAD GENÉTICA Y CONECTIVIDAD EN POBLACIONES NATURALES DEL Puye (Galaxias maculatus) EN PATAGONIA
(Patterns of genetic Diversity and connectivity in natural populations of el Puye (Galaxias maculatus) in Patagonia)
González-Wevar C.A.1, Salinas P.1, Hüne M.1,2, Yañez D, Poulin E.1
1Instituto de Ecología y Biodiversidad (IEB), Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Santiago, Chile. 2Departamento de Recursos Naturales, Universidad de Magallanes, Punta Arenas, Chile. [email protected] INTRODUCCIÓN La dinámica historia geológica y climática de Sudamérica durante el Neogeno modelaron la distribución, abundancia y estructura genética de la biota terrestre y especialmente la de especies de agua dulce. Estudios moleculares recientes han demostrado que los patrones de diversidad y estructura genética en especies sudamericanas es el resultado de complejas interacciones entre la biogeografía pasada, el potencial de flujo génico y la realización de éste en función a barreras históricas y contemporáneas. Durante el Último Máximo Glacial (∼ 20 ka), una extensa capa de hielo continental cubrió el margen occidental de la Cordillera de los Andes hasta la altura de los 35º S y los cambios geomorfológicos asociados a los avances y retrocesos del hielo glacial eliminaron casi por completo los hábitats dulceacuícolas. Por lo tanto, las especies que los habitaban se vieron desplazadas hacía sectores menos impactados y/o redujeron sus distribuciones a través del uso de refugios. Posteriormente, los procesos de deglaciación generaron rearreglos de las cuencas hidrográficas mediados por cambios en la dirección de los drenajes y aumento de los caudales favoreciendo el intercambio de fauna íctica entre cuencas. Estudios genético moleculares sobre los patrones de dispersión y re-colonización post-glacial en peces sudamericanos sugieren tanto la sobrevivencia de poblaciones al este de los Andes como también la el uso de refugios glaciales al oeste de los Andes. La familia Galaxiidae incluye cerca de 60 especies distribuidas en el Hemisferio Sur, a excepción de Antártica. La mayoría de las especies son exclusivas de agua dulce pero algunas pasan parte de su ciclo de vida en ambientes marinos someros. Entre éstas, Galaxias maculatus es una especie catadrómica en la que la toda la fase adulta ocurre en sistemas dulceacuícolas a excepción de la reproducción y la fase larval que ocurren en sistemas estuarinos y marinos, respectivamente. Además de estas poblaciones migratorias, se ha descrito la existencia de poblaciones no migratorias o residentes que llevan todo su ciclo de vida en agua dulce. Esta catadromía facultativa es considerada una adaptación clave de la especie para utilizar ambientes marinos adyacentes a los hielos continentales durante las glaciaciones cuaternarias. Estudios en G. maculatus se han centrado en aspectos globales de su diversidad genética entre poblaciones de Australia y Sudamérica, así como también en los mecanismos de dispersión utilizados y los tiempos de divergencia entre poblaciones de ambos continentes. Estudios recientes han determinado el patrón de diversidad genética general y la historia post-glacial de la especie en Patagonia. No obstante, la mayor parte de los estudios han sufrido sesgos en el número de individuos analizados por localidad, el uso de un solo marcador molecular y en el área de estudio abarcada centrándose la mayoría de estos los 39º y 42º S. Actualmente, se requieren estudios a escala más fina sobre la estructura filogeográfica de G. maculatus en distintas áreas a lo largo de su distribución considerando la historia glacial en la zona austral de Sudamérica y rasgos de historia de vida de la especie como el carácter migratorio y residente de sus poblaciones.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
560
La presente investigación buscará determinar los patrones de estructura y diversidad genética en poblaciones migratorias y residentes de G. maculatus a lo largo de un gradiente latitudinal. Se estimará las relaciones genéticas en poblaciones de G. maculatus provenientes de distintas cuencas hidrográficas ubicadas en áreas fuertemente impactadas por los ciclos glaciales del Cuaternario, al sur de los 44º S (XI Región de Aysén). Para efectos comparativos, además se incluirá un sector que se habría visto menos impactado por lo cambios geomorfológicos glaciales (XIV Región de los Ríos). Para evaluar el legado de los ciclos glaciales del Cuaternario sobre la demografía de la especie se realizarán análisis filogeográficos mediante el uso de la región control mitocondrial (D-Loop). La información contenida en las secuencias de DNA permitirá determinar el efecto de los ciclos glaciales sobre la diversidad y estructura genética en distintos sectores de la distribución de la especie.
MATERIALES Y MÉTODOS Se realizaron muestreos en 6 localidades de la Región de Aysén (Isla Concoto, Canal Williams, Canal María Eugenia, Estuario Río Pascua, Estuario Río Baker y lago Quetru) y en 6 localidades de la Región de los Ríos en los ríos Chaihuín, Moncul, Lingue y Queule. Todos los ejemplares (n = 302) fueron fijados en etanol (95%) donde se preparó DNA total mediante el método salino. Se amplificó un fragmento de 950 pares de bases del gen mitocondrial D-loop o región control (RC) los cuales fueron secuenciados en ambas direcciones. Las secuencias fueron editadas y alineadas para posteriores análisis comparativos. Para comparaciones genético poblacionales se incluyeron al menos 24 ejemplares por localidad (tabla 1). Se estimaron los niveles de polimorfismo genético a través de índices estándares de diversidad incluyendo el número de haplotipos (k), sitios polimórficos (S), diversidad haplotípica (H) para cada localidad. Además se estimó el número promedio de diferencias pareadas (Π) y la diversidad nucleotídica (π). Se estimaron los niveles de diferenciación entre poblaciones a través de la frecuencia de los haplotipos (GST). Se realizó un análisis de la Varianza Molecular (AMOVA) que permite estimar asignar las unidades que maximizan la varianza entre grupos y un análisis de paisajes genéticos implementado en el programa Geneland. Se construyeron las relaciones genealógicas entre las poblaciones de N. concinna usando redes de haplotipos de Median Joining. Se estimaron los índices de neutralidad de Tajima (D) y Fu (FS), y se construyó la distribución de desajuste (Mismatch distribution). Finalmente, se realizó un de Skyline Plot para estimar cambios demográficos históricos en la especie.
RESULTADOS (PRELIMINARES) Las secuencias provenientes de Isla Concoto, Canal Williams, Canal María Eugenia y de los ríos Queule, Chaihuín, Moncul y Lingue acaban de llegar a las instalaciones del laboratorio de Ecología Molecular, donde actualmente están siendo analizadas. Contaremos con estos datos y análisis al momento de la presentación. No obstante, actualmente contamos con datos para la cuenca del río Pascua y Baker, los cuales a continuación detallaremos. Se detectaron altos niveles de diversidad genética en la especie generadas por 92 sitios polimórficos presentes en un total de 102 ejemplares (Tabla 1). Los sitios de muestreo ubicados en las desembocaduras de ríos Baker y Pascua mostraron niveles similares de diversidad genética en comparación a los menores índices detectados en el lago Quetru (Tabla 1).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
561
Localidad N S K H π Desembocadura Baker
27 70 26 0.997 ± 0.011
15.467
Desembocadura Pascua
43 74 41 0.998 ± 0.006
12.784
Lago Quetru 32 22 11 0.714 ± 0.085
3.026
Tabla 1. Índices de diversidad molecular en poblaciones de Galaxias maculatus. N = número de individuos, S = Sitios polimórficos, K = número de
haplotipos; H = diversidad haplotípica y π diversidad nucleotídica. Table 1. Molecular diversity indices in populations of Galaxias maculatus. N = number of individuals; S = polymorphic sites; K = number of haplotypes; H =
haplotype diversity and π = nucleotide diversity.
La mayor parte de los haplotipos son únicos, proporción que aumenta en el fiordo donde los haplotipos únicos alcanzan el 97%. Del total de haplotipos del fiordo (66) sólo dos se encuentran compartidos entre muestras del Río Baker y Pascua. A diferencia de lo observado en el Fiordo, sólo se detectaron 11 haplotipos en el lago, 6 de los cuales son únicos. El haplotipo 2 (H2) del lago muestra una frecuencia elevada en comparación al resto de los haplotipos detectados en la especie (Figura 1).
Figura 1. Frecuencia de los 76 haplotipos encontrados en G. maculatus en fiordos (ríos Pascua y Baker) y en el lago
Quetru. Figure 1. Frequency of the 76 haplotypes found in G. maculatus in fjords (Pascua and Baker Rivers) and in the Quetru
lake. La red de haplotipos (Figura 2) muestra haplotipos de ambas desembocaduras (Baker y Pascua) mezclados dentro de una extensa y compleja red. Contrariamente, los haplotipos del lago se agrupan en una genealogía en forma de estrella compacta donde la mayoría de ellos derivan del haplotipo central de mayor frecuencia (H2). Sólo dos haplotipos del lago se mezclan dentro de la red formada por los haplotipos de la desembocadura de ambos ríos. Dichos haplotipos son genéticamente cercanos entre ellos.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
562
•
Figura 2. Red de haplotipos de Median Joining que incluye la relación entre los 77 haplotipos detectados en G. maculatus. Cada haplotipo esta representado como círculos cuyo tamaño es proporcional a su frecuencia y cuyo color
indica su procedencia. Cada línea entre dos haplotipos es proporcional al número de mutaciones que los separa. Figure 2. Median-Joining haplotype Network including 77 haplotypes detected in G. maculatus. Each haplotype is
represented by a circle which size is proportional to its frequency and the color indicates the locality. Las comparaciones pareadas de GST entre poblaciones de G. maculatus sugieren niveles significativos de diferenciación genético poblacional entre el Lago Quetru y las localidades en la desembocadura del río Baker y río Pascua (Tabla. La comparación entre Río Pascua y Río Baker no mostró diferencia significativa entre ambos sitios lo que sugiere la existencia de una única población a nivel del Fiordo.
Localidades GST Valor de p Lago – desembocadura Baker
0.46478 < 0.0001
Lago – desembocadura Pascua
0.46244 < 0.0001
Baker - Pascua 0.00858 0.75
Tabla 2. Pruebas pareadas de GST entre poblaciones de G. maculatus. Table 2. GST Pairwise comparisons between populations of G. maculatus.
Las pruebas de neutralidad de Tajima y Fu mostraron diferencias notorias entre las localidades del Fiordo y lago. Valores negativos y significativos observados en el lago son señal de procesos demográficos muy recientes como expansión poblacional, efecto cuello de botella o selección positiva; a diferencia de lo observado en el fiordo donde las poblaciones se han mantenido más estables en el tiempo. La distribución de desajuste de la población del fiordo es multimodal mientras que la población del lago parece ajustarse a una curva del modelo de expansión.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
563
Figura 3. Distribución del número de diferencias pareadas para la población del fiordo (a) y lago Quetru (b). El eje x
indica el número de diferencias y el eje y la frecuencia. Se muestra la distribución esperada según el modelo de expansión súbita. En los gráficos pequeños se muestra la dinámica del tamaño efectivo esperada para cada población
estimada por el parámetro τ. Figure 3. Distribution of pairwise differences in fjord (a) and (b) lake populations. X-Axis indicate the number of
pairwise differences and y-Axis the frequency. The expected distribution according to a sudden expansion model is also shown. In the small graphics is shown the effective population sizes dynamics estimated through the τ parameter.
Los resultados obtenidos en G. maculatus provenientes de la cuenca de los ríos Pascua y Baker (XI Región) sugieren la existencia de dos poblaciones claramente diferenciadas las que habitan de manera aislada el lago Quetru y el Fiordo. Los bajos niveles de diversidad genética encontrados en el lago, junto a su genealogía en forma de estrella sugieren que esta población habría pasado por un evento de expansión reciente. Por el contrario, las poblaciones del fiordo muestran niveles extramadamente altos de diversidad genética con una extensa y compleja red de haplotipos, lo que da cuenta de poblaciones más antiguas. De esta forma, las poblaciones del fiordo podrían haber sobrevivido durante la última glaciación, permaneciendo probablemente en el paleolitoral Pacífico. Además, los patrones de diversidad genética sugieren que la población actual del lago habría sido fundada posterior a la deglaciación a partir de un número reducido de migrantes provenientes del fiordo. AGRADECIMIENTOS: Proyecto Postdoctorado FNC3120075, Proyecto Fondecyt FNC 1110798, IDEAWILD. Instituto de Ecología y Biodiversidad (IEB) ICM-P05-002 & PFB-023- CONICYT.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
564
NUEVOS MARCADORES NUCLEARES PARA PINGÜINOS PYGOSCELIS (New DNA nuclear markers for Pygoscelis penguins)
Mura-Jornet, I.1*, Valenzuela-Guerra, P.1, González-Acuña, D.2, Vianna, J.A1*.
1Pontificia Universidad Católica de Chile, Departamento de Ecosistemas y Medio Ambiente, Vicuña Mackenna 4860, Macul, Santiago, Chile. *E-mail : [email protected], [email protected] 2Universidad de Concepción, Facultad de Ciencias Veterinarias, Departamento de Ciencias Pecuarias, Chillán, Chile. INTRODUCCIÓN El género Pygoscelis, de la familia Spheniscidae, clase Aves, está compuesto por tres especies: Pygoscelis adeliae, Pygoscelis papua y Pygoscelis antarctica (Ksepka et al. 2006). Pygoscelis adeliae posee una distribución reproductiva entre los 54 y 77ºS; Pygoscelis antarctica entre 54 y 64ºS y, finalmente, Pygoscelis papua con una distribución entre los 46 y 65ºS (Ancel et al. 2013). Actualmente, Pygoscelis adeliae y Pygoscelis papua se encuentran clasificados como casi amenazados (NT), mientras que Pygoscelis antarctica se encuentra con preocupación menor (LC) (IUCN, 2013). A pesar de que Pygoscelis antarctica se encuentra etiquetado como preocupación menor, se han reportado disminuciones de hasta un 84% en poblaciones de este pingüino en la Isla Rey Jorge (Korczak-Abshire et al. 2012). Debido a lo antes mencionado, es importante realizar estudios con marcadores genéticos, puesto que estos nos permitirán resolver temas relacionados con la conservación de los Pygoscelis gracias a la información que nos proporcionen, siendo los marcadores microsatélite una alternativa a los estudios de comportamiento y seguimiento de una especie (Schlosser, J. A et al. 2003). En este estudio, nosotros reportamos tres loci de microsatélite obtenidos desde el genoma de Pygoscelis adeliae, los que podrán ser utilizados en estudio de diversidad genética de las tres especie de Pygoscelis. MATERIALES Y MÉTODOS La obtención del genoma de Pygoscelis adeliae y el posterior análisis de éste por el Centro Nacional de Genómica, Proteómica y Bioinformática de Chile (OMICS), nos permitió identificar aproximadamente 5200 loci de tetranucleótidos simples. De estos, se seleccionaron 32 loci y se mandaron a sintetizar los partidores. Posteriormente, se evaluó en dos muestras de cada especie, cuáles de estos partidores amplificaron en las tres especies de Pygoscelis, pre-seleccionándose nueve de estos (Tabla 1). Se colectaron 80 muestras de sangre de las tres especies de Pygoscelis de colonias reproductivas, de 7 localidades diferentes durante el periodo 2011/2013. Pygoscelis papua (n=30) desde: Isla Ardley-South Shetland Islands (n=10), Bernardo O’Higgins-Península Antártica (n=10) y Gabriel González Videla-Península Antártica (n=10); Pygoscelis antarctica (n=30) desde: Narebski (n=10), Kopaitic (n=10) y Shirreff (n=10); y, finalmente, Pygoscelis adeliae (n=20) desde: Arctowski (n=10) y Bernardo O’Higgins-Península Antártica (n=10). Todo el ADN genómico fue extraído utilizando el protocolo de extracción con sal. Los nueve loci microsatélites preseleccionados fueron evaluados en 9 muestras de Pygoscelis papua, 9 muestras de Pygoscelis antarctica y 8 muestras de Pygoscelis adelie seleccionadas al azar, con el objetivo de evaluar polimorfismo (Tabla 1). La amplificación por PCR fue llevada a cabo utilizando un volumen final de 30uL, conteniendo 2uL de ADN, 2pmol de cada partidor (marcados con fluorescencia HEX, FAM o NED), 1X Buffer de reacción de PCR, 2.5mM de MgCl2, 200µM

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
565
de cada DNTPs y 0,8 unidades de Taq polimersa (Invitrogen®). Los ciclos de PCR fueron realizados siguiendo las siguientes condiciones: denaturación inicial a 95°C por 10min, seguido de una secuencia touchdown a 95°C por 15min, 60-50°C por 30min, 72°C por 45 minutos, con dos ciclos en cada temperatura de alineamiento y 35 ciclos de amplificación a 95°C por 15min, 50°C por 30min, 72°C por 5 minutos, seguido de una extensión final a 72°C por 30min. Los productos PCR fueron visualizados en un gel de agarosa al 3% con GelRedTM y secuenciados por Macrogen, Korea. Los loci que resultaron ser más polimórficos (AP-3, AP-19 y AP-26) fueron seleccionados y evaluados en las 80 muestras totales. Los alelos fueron identificados comparando su tamaño con los estándares de M13 en GeneMarker. Finalmente, se calculó la heterocigosidad observada (Ho) y esperada (He). RESULTADOS Y DISCUSIÓN De los 9 loci pre-seleccionados, 6 loci resultaron polimórficos por lo menos para dos de las tres especies (Tabla 1).
Tabla 1. Detalle de la amplificación de los loci microsatélites. M-monomórfico y P-polimórfico, con el número de alelos en paréntesis. (*)Representa loci donde no amplificaron todas las muestras.
Table 1. Detail of the amplification of microsatellite loci. M = monomorphic and P = polymorphic, with the number of alleles in parenthesis. (*) Represents loci where not all the samples amplified.
En los 9 loci, Pygoscelis adeliae fue el más polimórfico, al igual que en el estudio desarrollado por Trakiiska et al. (2005). Por otra parte, los loci AP-3, AP-19 y AP-26 fueron los más variables (polimórficos en todas las especies, con la excepción de AP-3 en pingüinos Barbijo y AP-19 en pingüinos Papua). Los tres loci más polimórficos (AP-3, AP-19 y AP-26), al ser analizados en 80 individuos de las tres especies de Pygoscelis (Tabla 2), provenientes de diferentes localidades, mostraron ser polimórficos para todas las especies. Esto muestra la importancia del número de individuos al momento de evaluar diversidad genética, ya que entre más individuos se tenga, mejor será la predicción que pueda hacerse. Como ejemplo, en el estudio de Trakiiska et al. (2005) en el que se utilizan 102 individuos.
a)Pygoscelis adeliae b)Pygoscelis papua c)Pygoscelis antarctica Locus Repetición del
alelo Rango de tamaño del alelo
No alelos
Rango de tamaño (en pb)
No alelos
Rango de tamaño (en
pb)
No alelos
AP-3 (ATAA)7 174-190 5 174-238
9
174-182 3
AP-19 (ATAA)7 250-290 5 206-254
5
250-290 7
AP-26 (GATA)7 226-262 8 210-250 8 218-262 10 Tabla 2. Caracterización de los tres loci de microsatélites analizados (número de alelos y su rango de tamaño) en a)
Pygoscelis adeliae, b) Pygoscelis papua y c) Pygoscelis antarctica.
Especie N° de muestras
AP-2 AP-3 AP-4 AP-12 AP-14 AP-19 AP-24 AP-26 AP-29
P. antarctica 9 M* M M P(2)* P(2) P(4)* M* P(5) M P. papua 9 M P(2) M M M M* M P(6) P(2)* P. adeliae 8 M P(3) M P(2) P(2) P(3) M P(8)* P(2)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
566
Table 2. Characterization of the three microsatellite loci analyzed (number of alleles and size range) in a) Pygoscelis adeliae, b) Pygoscelis papua and c) Pygoscelis antarctica.
Los menores valores de Ho fueron encontrados en Pygoscelis adeliae al ser comparados con Pygoscelis papua y Pygoscelis antarctica (Tabla 3). En Pygoscelis papua encontramos un mayor Ho para la localidad de Gabriel González Videla comparado con las otras dos localidades.
Tabla 3. Heterocigosidad esperada, observada y número de alelos para cada población de P. papua, P. adeliae y P.
antarctica utilizando los 3 loci microsatélites polimórficos identificados. Table 3. Expected and observed heterozygosity and number of alleles for each population of P. papua, P. adeliae and P.
antarctica obtained using the three polymorphic microsatellite loci identified. Especie Población N Loci He Ho No Alelos
P. papua Gabriel González Videla 10 3 0,6912 0,7333 7
Bernardo O’Higgins 10 3 0,4298 0,3667 4
Isla Ardley 10 3 0,5632 0,4667 4
P. adeliae Arctowski 10 3 0,4526 0,3667 5
Bernardo O’Higgins 10 3 0,6807 0,4333 4
P. antarctica Narebski 10 3 0,5098 0,5185 4
Kopaitic 10 3 0,5351 0,5000 4
Shirreff 10 3 0,6789 0,6333 5
A pesar de que en los primeros análisis se vio que Pygoscelis adeliae fue el más polimórfico, al evaluar los loci microsatélite, la heterocigosidad observada (Ho) fue la menor en comparación a las otras especies. Esto podría atribuirse a una baja en la población de pingüinos Adelie en casi todas las localidades de Antártica (Lynch et al. 2012). A pesar de nuestros resultados, consideramos que se deben realizar más estudios sobre la estructura genética poblacional de esta especie. En cuanto a los pingüinos Papua, se observó la mayor heterocigosidad observada en Gabriel González Videla. Estos resultados son similares a los encontrados en estudios de variación morfológica en las mismas poblaciones, donde los individuos de esta localidad presentan diferencias significativas morfológicamente con las otras localidades, principalmente con los individuos de Bernardo O’Higgins (datos por publicar). CONCLUSIÓN Dado que muchos estudios se ven limitados por la falta de marcadores genéticos que funcionen correctamente, hemos encontrado nuevos marcadores nucleares microsatélites para Pygoscelis. Lo cual permitirá estudiar con mayor profundidad las relaciones intra e inter poblacionales de las tres especies que conforman este género. A su vez, estos nuevos marcadores permitirán estudiar la diversidad de las especies, siendo una buena herramienta para la conservación de éstos. Finalmente, se sugiere la utilización de estos en estudios poblacionales de otras especies de pingüinos. AGRADECIMIENTOS INACH-G_06-11; INACH-T-27-10; INACH-MG_02-12 BIBLIOGRAFÍA Ancel, A., Beaulieu, M., & Gilbert, C. 2013. The different breeding strategies of penguins: A review. Comptes rendus biologies, 336(1):1-12 IUCN 2013. The IUCN Red List of Threatened Species. Version 2013.1. <http://www.iucnredlist.org>. Downloaded on 09 July 2013. Korczak-Abshire, M., Chwedorzewska, K., Wąsowicz, P., & Bednarek, P. 2012. Genetic structure of declining chinstrap penguin (Pygoscelis antarcticus) populations from South Shetland Islands (Antarctica). Polar

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
567
Biology, 35(11): 1681-1689. Ksepka, D. T., Bertelli, S., & Giannini, N. P. (2006). The phylogeny of the living and fossil Sphenisciformes (penguins). Cladistics, 22(5): 412-441. Lynch, H., Naveen, R., Trathan, P., & Fagan, W. 2012. Spatially integrated assessment reveals widespread changes in penguin populations on the Antarctic Peninsula. Ecology, 93(6): 1367-1377. Schlosser, J. Garner, T. Dubach, J. & McElligott, A. 2003. Characterization of microsatellite loci in Humboldt penguin (Spheniscus humboldti) and cross‐amplification in other penguin species. Molecular Ecology Notes, 3(1): 62-64. Trakiiska, E., Bichev, S., Telegeev, G., Dybkov, M., Bezrukov, V., Jankov, J., & Savov, A. 2005. Study of Three DNA Microsatellite Markers in Pygoscelis papua Penguins (Aves: Sphenisciformes) from the Livingston Island. Acta zoologica bulgarica, 57(3): 333-340.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
568
DISTRIBUCIÓN DE METALES, GRANULOMETRÍA Y COMPOSICIÓN ELEMENTAL E ISOTÓPICA EN SEDIMENTOS SUPERFICIALES DE LA BAHIA DE ALMIRANTADO Y
DEL ESTRECHO DE BRANSFIELD - PENÍNSULA ANTÁRTICA
(Distribution of metals, granulometry and isotopic and elementary composition in surface sediments of Admiralty Bay and Bransfield Strait - Antarctic Peninsula)
Cherene B.V.O.¹, Araujo B.F.¹, Almeida1 M.G.1, Vilaça M.S.1, Bernardes M.C.2, Campos
L.S.3 e Rezende C. E.¹
1Universidade Estadual do Norte Fluminense, Centro de Biociências e Biotecnologia, Laboratório de Ciências Ambientais, Campos dos Goytacazes, RJ, Brasil, 28013-602; [email protected] e [email protected] 2Universidade Federal Fluminense, Instituto de Química, Departamento de Geoquímica, 5º andar, Campus do Valonguinho, Centro, Niterói –RJ, 24029-000. 3Universidade Federal do Rio de Janeiro, Centro de Ciências da Saúde, Instituto de Biologia, – Bloco A – Ilha do Fundão – RJ – CEP 21949-900
RESUMO A Baía do Almirantado, atualmente considerada como Área Antártica Especialmente Gerenciada (AAEG), possui uma área de 362 km². O Estreito de Bransfield é uma bacia extensional, vulcânica e sismicamente ativa. O presente estudo tem como objetivo avaliar a distribuição de metais traço (Pb, Ba, Cd, Cr, Cu, Mn, P, Ni, Zn, Sr, V, Ca, Ti, S, Al, Fe) e composição elementar e isotópica da matéria orgânica nos sedimentos superficiais da Baia do Almirantado e Estreito de Bransfield. Os metais estudados apresentaram resultados que expressam a baixa probabilidade de ocorrência de efeitos adversos a vida aquática, porém Cr, Cu e Ni despertam atenção e devem ser monitorados com mais intensidade e ampliando o entendimento das potenciais fontes para estes elementos. A composição elementar e isotópica mostram a predominância de fontes autóctones, porém vemos com fundamental o aprofundamento do entendimento entre as fontes, as transformações na interface continente - oceano, e finalmente nos seus destinos finais.
INTRODUÇÃO O sedimento é formado pelas combinações de material inorgânico, como argilo-minerais, e matéria orgânica, tanto detrítica como viva. Esta mistura que compõe o sedimento representa o resultado de processos físicos, químicos e biológicos que ocorrem nos ecossistemas aquáticos, podendo interagir com diferentes poluentes. Dessa forma, os sedimentos de fundo são considerados importantes reservatórios de metais, podendo atuar como fontes de contaminação para a coluna d’água e para a biota. Contudo, o impacto que estes elementos causam nas estruturas das comunidades bentônicas e seus efeitos associados nos corpos d’água superficiais, depende diretamente da concentração, da biodisponibilidade e da forma química que estes elementos estão presentes nos sedimentos (Giesy et al., 1989). O Sistema do Tratado Antártico tem gerado mecanismos para promover o gerenciamento ambiental e das atividades humanas na Antártica com o objetivo de evitar e minimizar o risco de impactos acumulativos no meio ambiente, intensificando a assistência e a cooperação entre os países que operam na baía do Almirantado e protegendo importantes características ambientais e históricas (Neto et al, 2004). Em síntese, o objetivo deste estudo foi determinar a distribuição de metais e, composição elementar e isotópica nos sedimentos superficiais compreendido em um gradiente espacial que envolve a Plataforma e Margem Continental.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
569
MATERIAIS E MÉTODOS As amostragens do presente estudo foram feitas na Operação Antártica XXIII e Operação Antártica XXVII que foram realizadas durante os verões de 2004/2005 e de 2008/2009. Os sedimentos foram coletados com amostradores do tipo Mini Box Corer dentro da Baia do Almirantado em áreas rasas, e nas áreas profundas foram coletados com auxílio de “box-core” que estava a bordo do Navio Oceanográfico Ary Rongel. Neste estudo estão sendo apresentados resultados de 29 amostras de sedimentos superficiais (0 a 2cm) coletados em diferentes profundidades entre 24,4 a 1100m dentro e fora da Baia do Almirantado. Inicialmente, as amostras de sedimentos foram devidamente congeladas e armazenadas. Após o descongelamento, uma sub-amostra foi peneirada na fração <2 mm por via úmida e posteriormente levada para a determinação da granulometria através do analisador de distribuição de tamanho de partícula por difração a laser (modelo SALD-3101, Shimadzu). Os sedimentos secos e homogeneizados foram utilizados para determinação de metais traço e a composição elementar e isotópica. A abertura para os metais totais nos sedimentos foi realizada a partir de 0,5 g de sedimento seco foi colocado em bomba de teflon em forno microondas com tubo fechado (CEM modelo X-PRESS) e adicionados 9mL de HNO3, 4mL de HF e 2 mL de HCl as quais foram aquecidos a 180ºC com tempo de rampa de 10 minutos e depois mantidos por 20 minutos nessa temperatura. Após o que foram adicionados 25 mL de solução de ácido bórico 4% (m/v) e o extrato final foi reaquecido a 180ºC com tempo de rampa de 10 minutos e depois mantido por 20 minutos nessa temperatura. Após resfriamento, os extratos foram filtrados e analisados por ICP-OES Varian modelo 720 ES Baseado na U. S. EPA (1996) método 3052. Os testes sobre a exatidão utilizando-se o padrão certificado marinho NIST 2702 apresentaram os seguintes resultados: Al (93%); Fe (91%) Ba (97%); Cd (91%); Cr (92%); Cu (100%); Mn (90%); Ni (96%); Pb (93%); V (103%) e Zn (94%), S (95%) e P (90%). A composição elementar e isotópica foi determinada na amostra bruta (N total e δ15N) e na amostra descarbonatada (Corgânico e δ13C). A massa de material utilizada para as determinações analíticas foi aproximadamente 10 mg. As determinações foram realizadas no Analisador Elementar Flash 2000 acoplado a um espectrometro de massa Thermo Finnigan Delta V Advantage. A quantificação foi realizada utilizando-se curvas de calibração com padrão fornecido pelo National Institute Standards (NIST) Buffalo River. Os valores foram expressos em percentual (%), sendo a precisão obtida com três repetições, realizadas a cada 20 amostras. Nosso nível de reprodutibilidade analítica dentro da mesma amostra foi a partir de 95%. Os testes de exatidão para C e N realizados com Standard OAS/isotope – Low Organic Soil (microelement al microanalysis)foram realizados e revelaram recuperações acima de 94%. O limite de detecção para C e N foram 0,05%, 0,02%, respectivamente (Meyers, 1994). Os resultados são obtidos inicialmente relativos a padrões de trabalho para depois serem expressos em relação ao carbonato da formação Pee Dee (Belemnite) para δ13C e em relação ao N2 atmosférico para δ 15N. A precisão da análise é de 0,2‰ para os isótopos de C e N (Kenenedy et al., 2005; Meyers, 1994). RESULTADOS E DISCUSSÃO A distribuição granulométrica mostra um predomínio da fração silte e um aumento gradual do teor de argila em direção as estações mais profundas. Este padrão é corroborado por estudo anterior que indicou as regiões centrais da Baía do Almirantado como sedimentos lamosos nas regiões mais profundas Gruber (2008). Os valores do presente estudo, apresentaram resultados similares quando comparados a outros trabalhos na região Antártica (Santos et al 2005, Alam and Sadiq 1993, Sammonza et al, 2004, Jae

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
570
et al, 2005). Porém, segundo a literatura, os sedimentos da Baia do Almirantado possuem grandes quantidades de calcopirita (CuFeS2) que estão associadas a litologia da região o que acaba aumentando o nível de base para o Cu e não caracterizando um enriquecimento devido a atividade antrópica (Machado et al, 2001). Para Ni e Cr apenas a região E.B. (Estreito de Bransfield), apresentou os resultados mais elevados, e estes valores também podem estar ligado a grande atividade tectônica existente no estreito, que vem promovendo a abertura do Estreito de Bransfield a uma taxa de 11 ± 3 mm/ano (Dietrich et al, 1998).
Figura 1: Distribución granulométrica superficial de la Bahía de Almirantado y del Estrecho de Bransfield. Figure 1: Surface granulometric distribution in Admiralty Bay and Bransfield Strait.
A correlação não paramétrica de Spearman foi utilizada para se testar as associações entre os metais e, a composição elementar e isotópica. O Fe obteve correlações significativas com Cr, Mn Ni, Ti, Ca e S (P<0,05); o Al com Ba e Zn; o Mn com Sr, Ti, Ca, Fe e Cr. A composição elementar e isotópica não apresentaram correlações significativas com os metais. Estes resultados indicam que os óxidos e hidróxidos de Mn, Fe e Al são importantes suportes geoquímicos para os metais analisado (Salomons e Förstner, 1984). As concentrações da grande maioria dos elementos analisados não apresentaram valores acima das referências estabelecidos pelo NOOA (2008), exceto para Cr, Cu e Ni (Tabela 1). Estes resultados chamam atenção e mesmo que possamos atribuir estes resultados a litologia da região, as concentrações destes elementos devem ser acompanhas nos sedimentos e organismos da região.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
571
Estações Fe (mg.g-¹) Al (mg.g-¹) Ca (mg.g-¹) Ti (mg.g-¹) S (mg.g-¹) P (µg.g-¹) Mn (µg.g-¹) V (µg.g-¹) Sr (µg.g-¹) Ba (µg.g-¹) Zn (µg.g-¹) Cu (µg.g-¹) Cr (µg.g-¹) Ni (µg.g-¹) Pb (µg.g-¹) Cd (µg.g-¹)
Média 41 36,7 11,2 4,09 3,5 970 452 164 110 123 63 63 24 12 5,1 1,48
Intervalo 40 ± 42 28 ± 41 10 ± 13 4,03 ± 4,16 2,9 ± 4 835 ± 1088 409 ± 497 156 ± 171 92 ± 121 100± 140 60 ± 68 60 ± 67 20 ± 30 10 ± 15 4,6 ± 6,1 1,27 ± 1,81
Média 41 32,9 10,0 3,98 3,7 911 416 164 92 116 58 64 18 9 5,2 1,54
Intervalo 40 ± 42 24 ± 43 7 ± 13 3,67 ± 4,15 2,9 ± 5 637± 1072 375 ± 477 158 ± 172 67 ± 110 93 ± 134 55 ± 60 61 ± 68 17 ± 19 8 ± 11 4,3 ± 6,6 1,43 ± 1,72
Média 42 32,7 10,5 3,87 3,1 847 458 168 106 118 61 64 22 11 4,5 1,91Intervalo 39 ±45 27 ± 42 7 ± 13 3,57 ± 4,29 1,7 ± 6 632 ± 1101 393 ± 625 162 ± 177 63 ± 193 87 ± 149 57 ± 64 58 ± 66 16 ± 28 8 ± 14 3,3 ± 5,8 1,74 ± 2,12
Média 42 24,4 9,4 4,05 2,1 778 500 164 88 99 57 63 23 12 4,9 1,97
Intervalo 41 ± 44 13 ± 35 7 ± 12 3,8 ± 4,3 1,7 ± 2,5 667 ± 928 445 ± 562 155 ± 172 64 ± 127 72 ± 122 56 ± 58 59 ± 68 21 ± 27 10 ± 15 3,8 ± 6,2 1,68 ± 2,31
Média 46 23,0 12,8 4,83 4,1 864 698 190 163 120 49 61 49 16 4,1 1,05
Intervalo 44 ± 49 18 ± 29 10 ± 13 4,2 ± 4,99 0,9 ± 7,1 667 ± 963 480 ± 794 169 ± 196 93 ± 206 111 ± 129 45 ± 57 49 ± 69 22 ± 58 12 ± 18 3,8 ± 5 0,88 ± 2,00
Média 44 16,0 12,0 4,86 0,9 823 860 191 185 129 72 48 58 17 4,8 1,19
Intervalo 44 ± 45 6 ± 24 6 ± 13 4,84 ± 4,88 0,9 ± 7,1 762 ± 963 92 ± 926 63 ± 196 164 ± 206 11 ± 129 51, ± 94,81 31 ± 65 57 ± 58 16 ± 18 4,3 ± 5,3 1,17 ± 1,27
Ferraz sediments (AB, KGI) b 65 31,6 5,8 5,8 - - 624 204 63 41 89 92 31 10 10,5 -
Control sediments (AB, KGI) b 69 30,2 6,8 5,8 - - 527 127 121 - 52 67 35 7,9 5,5 -
Sediments (Intertidal area,
KGI) c 24 - - 0,4 - - 280 82 334 28 47 53 2,6 11 121 -
Deception Islanda d - - - - - - 819 207 - - 53 27 15 2.0 - -
Brandsfield Straitd e - - - - - - 1009 207 - - 123 68 98 34 - -NOAA (TEL) - - - - - - - - - - 124 18 52 15 30 -
NOAA (PEL) - - - - - - - - - - 271 108 160 42 112 4,2
E.B
CF1
CF2
BP1
BP2
B.A.
Tabla 1: Tabla comparativa de valores de metales en la región Antártica.
Table 1: Comparative table of values of metals in the Antarctic.
A composição elementar apresentou uma variabilidade abaixo de 35% para o Corgânico e 19% para o Ntotal, enquanto a composição isotópica do Corgânico ficou abaixo de 7% (δ13C -29,9 a -23,3‰), e o Ntotal apresentou uma ampla variabilidade (286%; δ15N -7,34 a 2,77‰). A razão atômica C:N apresentou baixa variabilidade (<12%) sendo o intervalo entre 4,3 e 7,3 típico de bactérias e fitoplancton. O Corgânico apresentou corelação siguinificativa com Ptotal e Ntotal sugerindo a predominância de formas orgânicas dentros das concentrações totais de P e N. Esta abordagem tem sido utilizada por vários autores sob diferentes circunstâncias. O intervalo da composição isotópica do carbono e nitrogênio obtido neste estudo variam de <0,05 a 0,15% e a razão isotópica variou de -29,36 a -23,31‰ para δ13C e de -7 a 3‰ para δ15N está dentro da faixa descrita na literatura, onde o fitoplanctônico da região do Equador varia entre -19 a -22‰ enquanto para as regiões polares está entre -26 a -31‰ para δ13C segundo (Rau et al.1991) e -5,6 a 2,8‰ para δ15N segundo (Bishop et al 2001). CONCLUSÕES O padrão de distribuição granulométrica observado apontou a fração silto-argilosa como predominante, sendo as isóbatas mais profundas com maior presença de argila e nas regiões mais rasas maior porcentagem de silte. Para as concentrações de metais, foi observado valores abaixo dos níveis considerados críticos para efeitos biológos, sendo que o Cu, Cr e Ni possuem níveis que chamam atenção, e devem ser monitorados, porém esses valores podem estar associados à litologia da região. A composição isotópica dos sedimentos podem ser consideradas leves para sedimentos tipicamente marinhos, porém as fontes locais descritas na literatura justificam plenamente estes resultados, mas faz-se necessário um inventário da composição isotópica de forma que possamos abordar de forma mais consistente a biogeoquímica da matéria orgânica.
AGRADECIMENTOS Ao Laboratório de Ciencias Ambientais LCA – UENF pelas facilidades analíticas necessárias para realização de todas as determinações; ao Projeto MABIREH. O Prof. CER é membro do INTC-MCTOcean sobre a Transferência de Materiais na Interface Continente-Oceano e ao INCT-APA.
REFERÊNCIAS Bishop, J.L., Lougear, A., Newton, J., Doran, P.T., Froeschl, H., Trautwein, A.X., Korner, W. and Koeberl, C. Mineralogical and geochemical analyses of Antarctic lake sediments: A study of reflectance and Mo¨ ssbauer

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
572
spectroscopy and C, N, and S isotopes with applications for remote sensing on Mars Geochimica et Cosmochimica Acta, Vol. 65, No. 17, pp. 2875–2897, 2001 Galimov E. M., 1999. Carbon isotope composition of Antarctic plants. Geochimica et Cosmochimica Acta, Vol. 64, No. 10, pp. 1737–1739 Giesy, J. P.; Hoke R.A.1989. Freshwater sediment toxicity bioassessment: rationale for species selection and test design . Journal of Great Lakes Research, 15 (4), p. 539-569 Gruber, N.L.S. 1990. Contribuição ao estudo da Sedimentação glacio-marinha da Baía do Almirantado, Ilha Rei George, Antártica. Dissertação de Mestre, UFRGS, 174p Kennedy, P., Kennedy, H., Papadimitrious, S., 2005. The effect of acidification on the determination of organic carbon, total nitrogen and their stable isotopic composition in algae and marine sediment. Rapid Communication in Mass Spectrometry 19, 1063-1068. Machado, A., Lima, E.F., Chemale Jr., F., Liz, J.D., A ´ vila, J.N., 2001.Quı´mica mineral de rochas vulcânicas da Peninsula Fildes (Ilha ReiGeorge), Anta´rtica. Revista Brasileira de Geocieˆncias 31, 299–306. Meyers, P. (1994). Preservation of elemental and isotopic source identification of sedimentary organic matter. Chemical Geology 114 (1994) 289-302. Rau, G. H., P. N. Froelich, T. Takahashi, and D. J .Des Marias. 1991. Does sedimentary organic delta 13C record variations in Quaternary ocean (CO 2(aq) )? Paleoceanography, 6:335-347.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
573
DISTRIBUCION VERTICAL DEL ZOOPLANCTON ANTARTICO EN LOS ALREDEDORES DE LA ISLA ELEFANTE. VERANO AUSTRAL 2007. PERU ANTAR
XVII (Vertical distribution of Antarctic zooplankton around Elephant Island. Austral summer 2007. Peru
ANTAR XVII) Pinedo, E.1, Nakazaki, C.1, Ayón, P.1, Vásquez, L.1 y J. Ledesma1
1Dirección General de Investigaciones Oceanográficas y Cambio ambiental. Área Funcional en Oceanografía biológica. IMARPE. Email: [email protected]
RESUMEN Durante el verano austral 2007, la distribución vertical del zooplancton fue estudiada en los alrededores de la Isla Elefante. Los promedios más altos de biovolúmenes y abundancias del zooplancton se encontraron en la capa más profunda (200-100 m) en horas de día. En total, se determinaron 12 grupos taxonómicos, siendo los copépodos el grupo más abundante y frecuente, seguido de los eufaúsidos y sifonóforos, los cuales se encontraron distribuidos en toda la columna de agua, presentando sus mayores concentraciones en la capa más profunda en horas del día, y disminuyendo hacia la superficie. Se observó patrones de migración nictimeral solamente en Oithona similis, Calanoides acutus (CV, CII y CI) y Thysanoessa sp. encontrados hacia la superficie en horas de la noche. A pesar de que en la capa superficial se encontraron las mayores concentraciones de clorofila, las abundancias del zooplancton se concentraron en la capa más profunda, lo cual estaría relacionado con la evasión de los depredadores en la capa superficial en horas de luz. MATERIALES Y MÉTODOS Durante la expedición ANTAR XVII realizada por el Perú durante el verano Austral del 2007 entre el 24 y 26 de enero, se realizó un experimento de pequeña escala entre la Isla Elefante y Clarence, en una línea de 10 mn (Fig. 1) colectándose un total de 27 muestras con intervalos de 6 horas, aproximadamente. Las muestras de zooplancton fueron colectadas con una red de bongo de cierre de 60 cm. de diámetro de 300 y 500 µ. Para conocer la cantidad de agua se utilizó un flujómetro marca Hydrobios. La red fue lanzada en tres estratos diferentes: 0-50, 50-100 y 100-200 m de profundidad. Se tomaron datos de temperatura, salinidad y clorofila. Para detectar si hubo migración nictimeral en las especies más abundantes, se determinó Weighted Mean Depth (m): WMD= Σ(ni × zi × di)/ Σ (ni × zi)
55.8ªW 55.4ªW 55ªW 54.6ªW 54.2ªW
55.8ªW 55.4ªW 55ªW 54.6ªW 54.2ªW
61
.6ºS
61
.4ºS
61
.2ºS
61
ºS
61
.6ªS
61
.4ªS
61
.2ªS
61
ªS
I. ELEFANTE
I. GIBBS
I. CLARENCE
PROYECTO COPEPOD IIEXPERIMENTO I. ELEFANTE
24-26 ENERO 2007
Figura 1. Área de estudio.
Figure 1. Study area.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
574
donde di es el centro de cada estrato (m), zi es el espesor del estrato (m) y ni es la biomasa o abundancia de los organismos (Nº ind. m-3).RESULTADOS Y DISCUSIÓN Los biovolúmenes promedio de zooplancton fueron más altos en el estrato más profundo en horas de luz, mientras que en horas de la noche los mayores promedios estuvieron en el estrato más superficial (Fig 2). Los mayores valores obedecen a la presencia de altas densidades de eufaúsidos (Euphausia superba y Thysanoessa spp.), sifonóforos, quetoganatos y copépodos de gran tamaño (Rhincalanus gigas). En cuanto a las abundancias, los promedios más altos se encontraron en la capa más profunda, disminuyendo hacia el estrato superficial tanto de día como de noche, lo cual estaría relacionado con su composición. El zooplancton estuvo conformado por 12 grupos taxonómicos, siendo los más abundantes y frecuentes los copépodos, seguido de los eufaúsidos y sifonóforos. Dentro de los copépodos, Ctenocalanus sp., Calanoides acutus (Fig. 3) y Oithona similis (Fig.4), fueron las especies más abundantes, las cuales se encontraron distribuidas en los tres estratos, presentando una mayor densidad en el estrato más profundo. Los resultados obtenidos del WMD para las especies más abundantes, muestran que sólo C. acutus (CV, CII y CI) (Fig.5), O. similis, y Thysanoessa sp. (Fig. 6) mostraron migración vertical más clara, concentrándose por debajo de los 100 m durante las horas luz y migrando hacia la capa superficial en horas de la noche. C. acutus presentó una migración vertical ontogénica, donde los estadios más jóvenes se encuentran en la capa superficial y los más avanzados en zonas más profundas lo que estaría de acuerdo con descripciones previas (Voronia, 1972 y Shnack-Schiel, 1992). Los otros componentes del zooplancton presentaron una mayor preferencia por el estrato más profundo en horas de luz. La capa superficial presenta una mayor concentración de clorofila y una distribución termo-halina más variable que es más homogénea hacia el estrato más profundo. Sin embargo, no se encontró correlación de las abundancias del zooplancton con ninguno de estos componentes. Las altas concentraciones alcanzadas por los grupos zooplanctónicos en la capa de 200-100 m. y en horas de luz, estaría relacionado con evitar la predación en la capa superficial en horas del día, donde estarían vulnerables a ser capturados.
Figura 2. Biovolúmenes y abundancias promedio del zooplancton por rango de profundidad.
Figure 2. Mean biovolumen and abundance of zooplankton by vertical range)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
575
AGRADECIMIENTOS Este estudio fue realizado dentro del proyecto COPEPOD II financiado por el Instituto Antártico Peruano (INANPE) y el Instituto del Mar del Perú (IMARPE).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
576
REFERENCIAS Atkinson, A., Shreeve, R., Pakhomov, E., Priddle, J., Blight, S. and Ward, P. 1996. Zooplankton response to phytoplankton Bloom near South Georgia, Antartica. Marine Ecoloy Progress Series 144: 195-210. Bonicelli, J., López, D., Ochoa, N. y Shreeve, R. 2008. Estructuta Comunitaria del Zooplancton Asociada con el fitoplancton y las masas de agua del Estrecho de Bransfield y la Isla Elefante durante el verano austral del 2006. EcologìaAplicada, 7(1,2). Schnack-Schiel, S, Hagen, W. and Mizdalski, E. 1991. Seasonal comparison of Calanoidesacutusand Calanus propinquus (Copepoda: Calanoida) in the southeastern Weddell Sea, Antarcticancias. Marine EcologyProgress Series. Vol. 70:17-27. Vásquez, L. 2007. Condiciones Ambientales en la Expedición ANTAR XVII. Proyecto COPEPD II.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
577
FEEDING HABITS AND DIET OVERLAP IN MARINE FISH LARVAE FROM THE PERIANTARCTIC MAGELLAN REGION
(Hábitos alimentarios y sobreposición trófica en larvas de peces marinos de la región periantártica de Magallanes)
Salas-Berrios, F.1, Valdés-Aguilera, J.1, Landaeta, M.F.1,
Bustos, C.A.1,2, Pérez-Vargas, A.3 y F. Balbontín4
1Laboratorio de Ictioplancton (LABITI), Facultad de Ciencias del Mar y de Recursos Naturales, Universidad de Valparaíso, Avenida Borgoña 16344, Reñaca, Viña del Mar, Chile. [email protected] 2Programa Doctorado en Acuicultura, Universidad Católica del Norte campus Guayacán, Larrondo 1281, Coquimbo, Chile. 3Programa Doctorado en Ciencias mención Recursos Naturales Acuáticos, Universidad de Valparaíso, Avenida Borgoño 16344, Reñaca, Viña del Mar, Chile. 4Laboratorio de Peces, Facultad de Ciencias del Mar y de Recursos Naturales, Universidad de Valparaíso, Avenida Borgoño 16344, Reñaca, Viña del Mar, Chile.
INTRODUCTION The Magellan region is a unique periantarctic ecosystem due to its geographical position. However, the knowledge about the distribution and feeding ecology of fish larvae is scarce. Physiographic and bathymetric features such as sills, shallow shelves, and deep basins affect the intrusion of water and plankton. Microbasins delimited by bathymetric features are consistent with the distribution of oceanographic variables, copepods, meso- macrozooplankton, and ichthyoplankton. One of the most recurrent fish species in the region is the black southern cod Patagonotothen tessellata (Richardson, 1844). This is a small Nototheniid fish (up to 22 cm body length, BL) that inhabits shallow subtidal zones (<15 m depth). Embryonic development last around 28 days, during which the male does not ingest food (Rae and Calvo 1995). Larvae hatch with pigmented eyes and a yolk-sac which is absorbed at 5-7 days post hatch (Rae et al. 1999). There is no information about distribution or feeding habits of the larval stages. Another prominent epipelagic marine fish species recurrent along the Patagonian fjords and channels is Bathylagichthys parini Kobyliansky 1990 (Bathylagidae). Although larval B. parini are recurrently collected in the plankton samples from Chilean Patagonia (Bustos et al. 2008, 2011), there is no information about distribution patterns and larval fish feeding of this species. Both marine fish species inhabit different bathymetric zones as adults; however, larval stages coexist in the water column during the austral spring season along southern Patagonia. The main objective of this study is to address the feeding habits of larval stages of Patagonotothen tessellata and Bathylagichthys parini, their niche breadth and overlap in southern Patagonia during the austral spring season. We hypothesize that both larval species predate over different prey items, and this will be expressed as a reduced diet overlap. MATERIALS AND METHODS During October-November 2009 and 2010, two bio-oceanographic cruises were carried out between 50º07’-52º45’ S, and 52º26’-55º23’ S, respectively. At each station the hydrographic features of the water column were measured with vertical casts of a CTD Seabird SB-19 from surface to 500 m depth or to 10 m from the floor. Zooplankton was captured using oblique tows from 200 m depth to surface or from 10 m above the bottom to surface (in shallower stations) with a 1 m2 mouth Tucker trawl (300 µm mesh size) equipped with a General Oceanics flow meter mounted in the frame of the

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
578
net to estimate the filtered seawater. Filtered seawater fluctuated from 22.71 to 897.23 m3 tow-1 (mean ± standard deviation: 221.94 ± 150.94 m3 tow-1) during 2009, and from 32.63 to 631.60 m3 tow-1 (264.04 ± 132.09 m3 tow-1) during 2010. Once onboard, nets were washed and plankton samples were initially fixed in 5% formaline buffered with sodium borate and then preserved in 96% ethanol. All fish larvae from plankton samples were separated, counted and identified to the lowest taxonomic level. Larval Bathylagichthys parini and Patagonotothen tessellata were identified. Abundance was standardized to individuals (ind.) 10 m-2, taking into account the number of larvae captured and volume of the seawater filtered by the net. Body length (BL, notochord length in preflexion larvae; standard length in flexion and postflexion larvae), upper jaw length (UJL, from the tip of the snout to the posterior end of the maxilla) and mouth width (MW, in the ventral view, the width between the posterior edges of the maxillae) of all intact larval B. parini (n = 300) and P. tessellata (n = 62) were measured under an Olympus SZ61 stereomicroscope attached to a camera (Motic moticam 2500, resolution 5.0 MPixel) using Motic Image Plus 2.0 software. The gut of each larva was dissected from the body and opened length-wise with fine needles. Under a Motic BA310 microscope, prey items were counted and identified to the lowest possible taxon, except for indigestible prey remains such as setae. The maximum BL and width of each prey item (maximum prey width, MPW) were measured with the microscope and a Motic moticam 2500 camera (resolution 5.0 MPixel) using the Motic Image Plus 2.0 software. The volume of each prey item was estimated using the three-dimensional shape that most closely resembled the item. Only prosome length of copepodite prey was measured, due to the urosome often being missing. All larvae with identifiable prey in their guts were used for the analysis. The diet was described using the percent frequency of occurrence (%F) of a diet item in larvae with food in their guts, the percent of the total number (%N) of diet items that were examined and the percent of volume (%V) of each item out of the total volume of prey items. An index of relative importance (IRI) was calculated as follows: IRI=(%N+%V)×%F. To readily allow comparisons among prey items, the IRI was the standardized to %IRI for each prey item i (Cortés 1997). Feeding incidence (FI) was calculated as a percentage of the total number of larvae that had gut contents out of the total number of larvae examined for each species and for the day and night samples. The larvae collected between 1 h after sunrise and 1 h before sunset, and between 1 h after sunset and 1 h before sunrise, were considered ‘daytime’ and ‘nighttime’ samples, respectively. Comparisons between day and night were carried out with contingency tables. RESULTS The 11 and 14 different prey items detected in the guts of B. parini and P. tessellata, respectively during 2009-2010 are listed in Table 1. Bathylagichthys parini larvae showed high percentage of largely digested prey items (unidentified and chitinous remains, 54.24 and 13.35% of %IRI, respectively), suggesting that digestion occurs fast. Also, during the larval development, feeding was on the basis of phytoplankton (diatoms of the genus Fragilaria), zooplankton (several life stages of copepods, invertebrates eggs, ostracods) and even fecal pellets (from appendicularians), showing an omnivorous tactic. Numerically, invertebrate eggs and fecal pellets were the most important items in the diet (Table 1). Volumetrically and most frequent prey items were unidentified (highly digested) and chitinous remains, explaining the high percentage of IRI% (67.6%) in the gut contents. Most of the identified prey are static (eggs, 15.35%; fecal pellets, 4.82%; diatoms, 3.12%) or show slow movements in the water column (copepod nauplii, 4.86%). Larval P. tessellata showed a diet based exclusively on zooplankton, being carnivorous in its feeding (Table 1). Numerically, the most important prey taxa were calanoid copepodites (25.11%)

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
579
and Calanus simillimus (23.81%), being also the most frequent prey in the guts (93.55% and 85.48%, respectively). Volumetrically, chitinous remains and C. simillimus were the largest (Table 3). Diet of larval P. tessellata was largely dominated by different life stages of copepods (74.25%IRI), mostly adult copepods (41.36%). The feeding incidence (FI) of all larval B. parini examined was low (FI = 26.67%). It did not differ between locations (north vs south areas, χ2 = 0.338, df = 1, P =0.561), but it was higher in larvae collected during day compared those captured at night (FI = 46.96% and 14.05%, respectively; χ2 = 39.3, df = 1, P < 0.001), suggesting that feeding occurs mostly during daylight. On the other hand, FI of larval P. tessellata was high (FI = 70.97%) and significantly larger than B. parini (χ2 = 44.8, df = 1, P < 0.001). FI of P. tessellata differ between day and night, being larger at night (84.21% at night, 50% at day, χ2 = 8.36, df = 1, P = 0.004). This result suggests that larval P. tessellata are able to feed with low light intensities and/or it is a nocturnal feeder. Table 1. Diet composition of larval Bathylagichthys parini and Patagonotothen tessellata from the Magellan region, austral spring 2009-2010. Tabla 1. Composición de la dieta de larvas de Bathylagichthys parini y Patagonotothen tessellata de la región de Magallanes, primavera de 2009-2010
Bathylagichthys parini Patagonotothen tessellata
Prey taxa %N %V %F %IRI %N %V %F %IRI
Fragilaria sp. 12.06 1.24 10 3.121 - - - -
Bivalvia D larva 1.56 0.90 5 0.288 0.43 0.03 1.61 0.006
Invertebrate egg 17.51 1.02 18.75 8.151 - - - -
Pluteus larva - - - - 0.43 0.10 1.61 0.007
Copepod egg 14.01 1.33 20 7.200 4.76 0.06 17.74 0.722
Copepod nauplius 6.23 6.98 12.5 3.873 12.99 1.35 48.39 5.858
Calanoid copepodite 7.39 11.06 11.25 4.871 25.11 8.19 93.55 26.303
Aetideus arcuatus - - - - 7.79 9.44 29.03 4.225
Calanoides patagoniensis
- - - - 1.30 10.60 4.84 0.486
Calanus simillimus - - - - 23.81 25.57 85.48 35.639
Paracalanus indicus - - - - 3.90 4.03 14.52 0.971
Phaenna spinifera - - - - 0.43 0.01 1.61 0.006
Rhincalanus nasutus - - - - 0.87 0.64 3.23 0.041
Zoea - - - - 1.30 0.19 4.84 0.061
Mysis - - - - 1.30 1.19 4.84 0.102
Ostracoda 0.39 0.02 1.25 0.012 - - - -
Salp 0.39 0.00 1.25 0.012 - - - -
Fecal pellet Appendicularia
16.34 1.94 11.25 4.826 - - - -
Chitinous remains 8.95 21.41 18.75 13.359 15.15 38.48 56.45 25.565
Unidentified 14.40 54.10 33.75 54.242 0.43 0.13 1.61 0.008
Saetas 0.78 - 2.5 0.046 - - - -

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
580
DISCUSSION Marine fish larvae inhabiting the Periantarctic region of Magellan and Fuegian zones of South America showed different tactics in their feeding, during austral spring. Both species were collected along the southern tip of South America in low abundance during 2009 and 2010. Larvae of the bathylagid fish Bathylagichthys parini showed a lower increase in the mouth gape at size, predates during day hours mostly over no motile prey from different origin (phytoplankton, zooplankton, fecal pellets). They show no increase of its feeding success with increasing larval size, and the niche breadth is independent of larval size. On the other hand, larvae of the nototheniid fish Patagonotothen tessellata showed a large mouth gape at size, which may partially explain the predation over motile prey like large calanoid copepods (Calanus simillimus). They are nocturnal feeders whose diet is exclusively carnivorous, feeding over more and large prey during their larval development; nonetheless, niche breadth is also independent of larval size. Finally, diet overlap was important only in individuals with smaller mouth gape, and diminished as larvae (and their jaw) grow. Because of the small size and absence of swimming behaviour of prey items ingested by B. parini, it is plausible that B. parini larvae need sunlight for the detection of prey, in the case of most fish larvae. Differences in the timing when FI increases among larval species from the Magellan zone, potentially may reduce any competence for food during early life stages, when diet overlap is the largest. ACKNOWLEDGEMENTS CIMAR 15-F 09-03, CIMAR 16-F 10-03, Fondecyt 11090020 (MFL). REFERENCES Bustos, C.A., Landaeta, M.F. y Balbontín, F. 2008. Environmental effects on the spatial variability of the ichthyoplankton from southern Chile during November 2005. Revista Chilena de Historia Natural 81:205–219 Bustos, C.A., Landaeta, M.F. y Balbontín, F. 2011. Ichthyoplankton spatial distribution and its relation with water column stratification in fjords of southern Chile (46º48’S-50º09’S) in austral spring 1996 and 2008. Continental Shelf Research 31:293–303 Cortés, E. 1997. A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Canadian Journal of Fish Aquatic Science 54:726–738 Rae, G.A. y Calvo, J. 1995. Fecundity and reproductive habits in Patagonotothen tessellata (Richardson, 1845) from the Beagle Channel, Argentina. Antarctic Science 7:235–240 Rae, G.A., San Roman, N.A. y Spinoglio, D.E. 1999. Age validation and growth of yolked larvae of Patagonotothen tessellata (Richardson, 1845) (Pisces: Nototheniidae) from the rocky littoral of the Beagle Channel, Argentina. Scientia Marina 63(Supplement 1):469–476

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
581
EVALUACIÓN HIDROACÚSTICA DEL KRILL (Euphausia superba) EN EL ESTRECHO DE BRANSFIELD Y ALREDEDORES DE LA ISLA GIBBS DURANTE LA XXI
CAMPAÑA ANTÁRTICA PERUANA 2013
(Hidroacustic survey of krill (Euphausia superba) in Bransfield and Gibbs Island during XXI Peruvian Antartic Campaign 2013)
Salazar, C.M.1, Cornejo, R.M.1 y M. Flores1
1Dirección General de Investigaciones en Hidroacústica, Sensoramiento Remoto y Artes de Pesca, Instituto del Mar del Perú (IMARPE), Perú. Email: [email protected] La evaluación hidroacústica del krill (Euphausia superba) se efectuó a bordo del BIC HUMBOLDT del 15 al 22 de febrero del 2013, el área evaluada fue de 13.607 mn2 localizadas entre el estrecho de Bransfield y las proximidades de la Isla Gibbs, dicha zona fue cubierta por 10 transectos con longitudes que variaron entre 15 y 40 mn, la Unidad Básica de Muestreo (UBM) fue de 1 mn y se rastrearon en total 507 UBM. MÉTODOS Se utilizó un diseño de muestreo de tipo sistemático estratificado (perfiles paralelos y equidistantes), en el cual se asume una aleatoriedad en la distribución del recurso con relación a la localización de los transectos. El sistema de ecointegración estuvo compuesto por la ecosonda científica EK60 SIMRAD con transductores modelos ES120-7 y ES38B de haz dividido, frecuencias de 120 y 38 kHz, respectivamente. Este sistema es calibrado de acuerdo al procedimiento señalado por su fabricante, el cual sigue las recomendaciones efectuadas al respecto por el ICES para el uso cuantitativo de mediciones acústicas. La cobertura del muestreo vertical a 120 kHz fue de 0 a 2 000 metros y de 0 a 2 000 m para 38 kHz. Las calibraciones acústicas se realizaron en la Ensena McKellar, de acuerdo a la metodología descrita en Simmonds and MacLennan (2005). Para el análisis de los datos acústicos se utilizó un programa de post-procesamiento (Echoview 3.00.81.32), que contiene los módulos de detección de cardúmenes y ecogramas virtuales que facilitan la extracción de la información correspondiente a los enjambres de kril y otras especies. A partir del 2000 se viene utilizando un al Algoritmo de identificación a través del cual las señales acústicas (ecoregistros) son separadas según la presencia de krill o zooplancton en base al algoritmo de Green et al (1996), quienes hallaron una diferencia de 2 a 16 dB en las señales retrodispersadas por el krill utilizando frecuencias de 38 y 120 kHz. Esta metodología es ampliamente utilizada en estudios relativos al krill en la Antártica. Para los lances de arrastre pelágico y semipelágica se utilizó la red Pelágica ENGEL 1000/400, con un copo de 13mm. Se monitoreó la red con un net-sonda SIMRAD modelo ITI. Complementariamente se registró información térmica utilizando un sensor marca FURUNO. Para ejecutar los lances de arrastre de fondo, se contó con información de la expedición ANTARKTIS XIV/2 del buque aleman “Polarstern” en 1996-97 y datos de la campaña realizada por Antarctic Marine Living Resources Program de la NOOA. Para la realización de los lances de fondo, previamente se detectó los fondos para identificar áreas arrastrables, teniendo en cuenta la configuración de fondos, estado de mar, corrientes, vientos entre otros parámetros. La red utilizada es de modelo Granton con copo de 90 mm.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
582
RESULTADOS Resultados respecto a la relación de los enjambres krill con respecto a las horas del día y profundidad indicaron que El krill durante el día (05-11 horas y 16-18 horas) se presentó en enjambres con altas densidades, que principalmente durante el amanecer (04-08 horas) tienen mayor altura. Conforme transcurrieron las horas de luz disminuyó la extensión de estas agregaciones que se localizaron entre los 20 y 50 m de profundidad. Durante la noche (22-04 horas) el krill se encuentra disgregado y muy disperso, presentando extensas capas o manchas muy cerca de la superficie hasta los 20 m de profundidad. Las principales características morfológicas (densidad, extensión y altura) de los enjambres variaron según las horas de luz y noche. Durante las horas de luz se incrementó el número y la densidad de los enjambres de krill, se registraron 354 agregaciones principalmente con densidades bajas y medias entre 40 y 5 000 m²/mn² respectivamente, alcanzando densidades altas llegando a medir entre 10 000 y 14 000 m²/mn². En la noche se registraron solo 118 enjambres de krill con densidades entre 80 y 4 000 m²/mn² densidades bajas y medias respectivamente y pocas agregaciones con densidades altas superiores a 5 000 m²/mn². En cuanto a la extensión de los ecotrazos de krill, durante las horas de luz se presentaron más estrechos alcanzando hasta 500 m de longitud, sin embargo, durante la noche prevalecieron los estratos o capas de krill con extensiones de 10 a 1000 m, alcanzando hasta 2000 m longitud . En el caso de la altura, en general los enjambres de krill se presentaron ligeramente más altos durante las horas de oscuridad alcanzando hasta 30 m, durante el día esta altura alcanzó hasta 18 m. Los valores ecointegrados predominantes fueron calificados como de magnitud de tipo ‘disperso’. Densidades más altas de krill se observaron en el extremo oriental del estrecho de Bransfield. Se observó una gran zona de distribución al Este del estrecho entre las Islas Rey Jorge y Gibbs donde predominaron densidades medias (500-1 000 m²/mn²) y altas (> 1 000 m²/mn²), y del lado Oeste del estrecho entre la Isla Decepción y la isla Torre se detectó un núcleo más pequeño con las mismas densidades. Entre estas zonas de gran concentración se presentaron enjambres con densidades muy bajas (< 250 m²/mn²). Verticalmente se observó el patrón de comportamiento típico del krill, durante las horas de luz (de las 05 a 22 horas). Se distribuyó principalmente entre los 20 y 60 m, alcanzando hasta 240 de profundidad; acercándose a la superficie en horas de la noche (22 a 05 horas) donde se localizó entre los 05 y 40 m. El rango de temperatura de los enjambres de krill estuvo entre -1.0 y 1,5 °C pero con mejores relaciones entre valores de -0.8 a 0.9 °C. Con respecto a la salinidad las agregaciones del kril fueron observadas en un amplio rango con valores entre 33.96 y 34.35 ups.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
583
Figura 1. Trayectos prospectados y ubicación de lances de pesca efectuados en Estrecho de Bransfield. Figure 1. Prospected track and location of fishing station in Bransfield Strait.
Figura 2. Distribución de la densidad acústica de krill según categorías de abundancia relativa. Frecuencia 120 kHz (EK60
SIMRAD). Figure 2. Acoustic density distribution of krill according to relative abundance. Frequency 120 kHz (EK60 SIMRAD)
Figura 3. Características físicas de los enjambres de krill en el experimento de comportamiento.
Figure 3. Physical characteristics of krill swarms in behavioral experiment.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
584
REFERENCIAS BIBLIOGRAFICAS Gutiérrez, M. 1999. Mediciones in situ de TS para el krill (Euphausia superba) a 120 kHz. Verano austral 1998. Perú ANTAR IX. Inf. Inst. Mar Perú 144: 7-22. Castillo, R., Gutierrez, M., Ganoza, F. y Aliaga, A. 1999. Biomasa y distribución del krill (Euphausia superba) utilizando 38 y 120 kHz Verano austral 1998. Perú ANTAR IX. Inf. Inst. Mar Perú 144. Ganoza, F. y Peraltilla, S. 1999. Comportamiento de la red de arrastre pelágico Engel 998/400 para la captura de krill. Verano austral 1998. Perú ANTAR IX. Inf. Inst. Mar Perú 144. Hooker, Y., Quiñones, J., Pellón, J. y Bustamante, M. 1999. Aspectos biológicos del krill (Euphausia superba) durante la expedición de evaluación hidroacústica. Verano austral 1998. Perú ANTAR IX. Inf. Inst. Mar Perú 144.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
585
EFECTO DEL VIENTO Y LA MAREA EN LAS PROPIEDADES FISICO-QUIMICAS DE CALETA POTTER
(Wind and tide effects on physical and chemical properties of Potter Cove)
Sierra, M.1,2, Ruiz Barlett, E.1,2 y G. Tosonotto1
1 Dirección Nacional del Antártico - Balcarce 290, C1064AAF Buenos Aires, Argentina. [email protected]; [email protected]; [email protected] 2 Universidad de Buenos Aires - Intendente Güiraldes 2160 - Ciudad Universitaria - C1428EGA Buenos aires, Argentina INTRODUCCIÓN La Caleta Potter se encuentra al Sudoeste de la isla 25 de Mayo (King George Island), Shetlands del Sur. Es un cuerpo de agua semicerrado, con una orientación SW – NE y una superficie de aproximadamente 7 Km2. Su extremo Noreste y la costa Norte están rodeados por el glaciar Fourcade, mientras que la costa Sur es una playa de arena y grava. Su extremo Sudoeste está abierto a la Bahía Guardia Nacional (Maxwell Bay) que a su vez la conecta con el Mar de la Flota (Brandsfield Strait). (Figura 1) La topografía de fondo es irregular, comprendiendo una serie de umbrales y cuencas. Se subdivide a la caleta en una parte interior, con profundidades menores a 60 m, y una exterior, que aumenta abruptamente de 40 a 200 m de profundidad. (Tosonotto et al., 2010).
Figura 1. Península Antártica, Isla 25 de Mayo (King George Island) y Caleta Potter. Los asteriscos muestran la
posición de las estaciones de la transecta y los círculos la posición de las estaciones yo-yo. Figure 1. Antarctic Peninsula, 25 de Mayo Island (King George Island) and Potter Cove. The asterisks show the
position of the transect stations and circles show the position of the yo-yo stations.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
586
El campo de viento es bidireccional, predominantemente del sector Este – Oeste. La velocidad media es de ∼10 m/s , pudiendo llegar a valores de hasta ∼20 m/s. (Klöser et al.; 1994). El régimen de las mareas es mixto predominantemente semidiurno. Las corrientes de marea son de poca intensidad. Mediciones directas de corrientes indican que la circulación es de tipo ciclónica dentro de la Caleta. Por otra parte, existe una celda vertical cuando la intensidad del viento supera los 5 m/s, produciendo en la parte mas interna de la caleta surgencia (upwelling) con viento procedente del Este y hundimiento (downwelling) con viento procedente del Oeste (Roese et al., 1998). En este trabajo se describen los cambios en las propiedades físico-químicas de la columna de agua en relación al régimen de marea reinante en el momento de la medición, como también variaciones debidas a cambios meteorológicos. MATERIALES Y MÉTODOS Durante los veranos australes que van del 2010 al 2013, se realizaron estaciones oceanográficas (ver Figura 1) con un perfilador continuo CTD Sea Bird 19 plus V.2 con sensor de turbidez. Con el fin de analizar la variación de las condiciones hidrográficas influenciadas por diferentes condiciones de viento y mareas, se tomo como referencia una transecta a lo largo de la parte interna de la caleta. Durante el período correspondiente al verano 2011/12 y diciembre del 2012 se realizaron 3 perfiles tipo yo-yo (estaciones a, b, c; mostradas en la Figura 1). La medición se repitió para regímenes de marea diurno y semidiurno, durante las pleamares y bajamares, a lo largo de un día. RESULTADOS Y DISCUSIÓN Durante las pleamares se observan aguas más frías y salinas en la parte exterior de la transecta. La profundización de isotermas e isohalinas hacia la cuenca interior, indican mezcla en la columna de agua y en consecuencia hundimiento. Para las bajamares, en ambos regímenes, se observa que agua mas fría y salina se concentra en la parte profunda de la cuenca interna. El agua proveniente del glaciar se extiende a lo largo de toda la transecta, obedeciendo principalmente a la dirección y velocidad del viento. Para velocidades mayores a 5 m/s, cuando la dirección predominante es del sector Oeste, una lengua de agua cálida ingresa desde la caleta exterior, mezclándose con la pluma de agua dulce y fría del glaciar que se encuentra retraída hacia el extremo NE de la caleta. Cuando el viento es del sector Este, el agua de deshielo proveniente del glaciar se extiende a lo largo de toda la transecta, dejando a las aguas cálidas restringidas a la zona más externa de la caleta. Este comportamiento, que afecta la estructura térmica y salina, también altera la distribución de densidad. La inclinación de las isopicnas indica, en el primer caso, surgencia costera y, en el segundo, hundimiento. (Figura 2). Los datos obtenidos con las estaciones yo-yo revelan, para el régimen semidiurno, un aumento en la profundidad de la haloclina durante la bajamar, en especial para las estaciones b y c. La temperatura en la capa de mezcla muestra aumento y descenso, relacionado al flujo y reflujo de la marea respectivamente, principalmente en zona de la boca de la caleta. Para el régimen diurno se aprecia un aumento de temperatura con el flujo de marea, especialmente en las estaciones b y c, lo que es evidencia de la entrada de agua relativamente más cálida proveniente de la Bahía Guardia Nacional siguiendo la costa norte, en concordancia con la circulación ciclónica descripta por Roese et al., 1998.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
587
(a)
.
(b)
Figura 2. Distribución de densidad potencial a lo largo de la transecta para a) viento del Este y b) viento del Oeste. Figure 2. Distribution of potential density along the transect for a) Eastern winds and b) Western winds.
Los valores de salinidad de 27 a 34 ups, observados en las estaciones yo-yo realizadas en el verano de 2012, cubren un rango más amplio que el observado en Diciembre del 2012, con valores entre 34 y 34.15 ups. También se observan diferencias en los rangos de temperatura, siendo respectivamente de 0.97 a 2.21°C y de -0.44 a 0.32 °C (Figura 3).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
588
Figura 3. Diagrama TS que muestra la distribución de temperatura potencial y salinidad de las estaciones yo-yo. Figure 3. TS diagram showing potential temperature and salinity distribution from yo-yo stations. CONCLUSIÓN La Caleta Potter es un cuerpo de agua dominado por fuertes vientos y afectado por el derretimiento de los glaciares. Por lo tanto, las características físico – químicas de la columna de agua responden a cambios en las corrientes de marea, descarga de agua dulce, temperatura del aire y acción del viento. El análisis de sus variaciones, es de gran utilidad en estudios desarrollados por disciplinas como biología y geología, entre otras. El monitoreo de las propiedades oceanográficas de dicho lugar es fundamental, ya que su variación afecta directamente a la flora y fauna autóctona, alterando su tasa de crecimiento, reproducción y desarrollo. AGRADECIMIENTOS A la Dra. Doris Abele. A Oscar González y Alejandro Ulrich por su cooperación en el trabajo de campo. REFERENCIAS BIBLIOGRÁFICAS Klöser, H., Ferreyra, G.A., Schloss, I.R., Mercuri, G., Laturnus, F., Curtosi, A. 1994. Hydrography of Potter Cove, a Small Fjord – like Inlet on King George Island. Estuarine, Coastal and Shelf Science 38. Pp 523 - 537 Roese, M y Drabble, M. 1998. Wind-Driven Circulation in Potter Cove. Berichte zur Polarforschung, Reports on Polar Research. Pp. 40-46. Tosonotto, G., Sierra, M. y Ruiz Barlett, E. 2010. Oceanographic Characteristics in Potter Cove During Summer 2010. 4th Open Science Conference CD Rom. Abstract number 775, 1pp.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
589
COMPARACIÓN DE LA RESPUESTA FOTOSINTÉTICA A LA VARIACIÓN DE LA INTENSIDAD DE LUZ DE DIATOMEAS BENTÓNICAS ANTÁRTICAS DE LOS
GÉNEROS NAVICULA Y NITZSCHIA.
(Comparison of the photosynthetic response to light intensity variation of benthic Antarctic diatoms from the genus Navicula and Nitzschia)
Uribe, P.1, Petrou, K.2 y M. Zbinden2
1Fundación Ciencia para la Vida e INTA, Universidad de Chile, Santiago, Chile. Email: [email protected] 2 Plant Functional Biology & Climate Change Cluster (C3) University of Technology of Sydney, Australia. INTRODUCCIÓN Las comunidades de diatomeas bentónicas son responsables de un 40% de la producción primaria en zonas costeras ((Falkowski y Raven, 1997; Underwood y Kromkamp, 1999). En la costa Antártica, están expuestas a grandes diferencias de intensidad de luz durante el ciclo anual. En particular, pueden persistir en esas áreas durante el invierno austral bajo la cubierta de hielo marino, en niveles muy bajos de intensidad de luz (Robinson et al, 1995). Por lo tanto, tienen un rol crucial en la red trófica de invierno, como soporte o “forraje de invierno” para una diversidad de organismos incluyendo pequeños crustáceos y krill. Sin embargo, las características de esta comunidad en su composición, y su respuesta a este ambiente lumínico cambiante es poco conocido. En estudios preliminares, se caracterizó la composición de especies de diatomeas de la comunidad bentónica obtenidas en diferentes profundidades de la Bahía Covadonga en la Base Antártica Bernardo O’Higgins y la composición relativa de sus pigmentos fotosintéticos y foto-protectores mediante HPLC. Esta comunidad presenta una gran diversidad y riqueza de especies de diatomeas, con diferentes morfologías, rango de tamaño y formas, algunas presentan movilidad, y producción de sistemas de túbulos. Para examinar si esta diversidad se expresa además en su comportamiento frente a la variación de la intensidad de luz, en este estudio, comparamos la respuesta foto-fisiológica y los rangos de tolerancia de cuatro especies pertenecientes a los géneros Navicula y Nitzschia aisladas a partir de estas comunidades. MATERIALES Y MÉTODOS Las muestras de las comunidades de diatomeas bentónicas se recolectaron en diferentes sitios de la bahía Covadonga (63°19′15″S, 57°53′55″W) y alrededores de la Base O’Higgins, Antártica. Las muestras se obtuvieron a 20 metros de profundidad en la bahía, en la zona intermareal, y en frentes de hielo en las playas (Fig. 1). Las especies Navicula perminutum y Navicula cancellata, Nitzschia sp1 y Nitzschia sp2 se aislaron mediante “fishing” y se cultivaron a 4ºC y 0,5 - 5 µMol Fotones m-2 s-1, en medio f/2 (Guillard y Ryther1962) (Fig. 2). Alícuotas de 3 ml de los cultivos fueron expuestos por 5 horas a diferentes intensidades de luz, y se determinaron los parámetros fotosintéticos, Quantum Yield (φPSII), mediante un fluorómetro Water PAM (Fluorometro de Pulso Modulado), inmediatamente, y luego de 30 min. de recuperación en oscuridad. Además se compararon las variaciones de la composición relativa de diferentes pigmentos fotosintéticos y fotoprotectores en estas especies, medidos por HPLC, a partir de muestras tomadas antes y después del tratamiento (Petrou, et al, 2011).

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
590
Figura 1. Tipos de muestras. A: Bentos submarino, B: Bentos de la zona intermareal. C: Microalgas de frentes de hielo
en playas. Figure 1. Sample types. A: Submarine benthos, B: Intertidal benthos C: Ice fronts microalgae.
Figura 2. Especies de diatomeas bentónicas antárticas en cultivo.
Figure 2. Antarctic benthic diatom species in culture. RESULTADOS Respuesta Fotosintética. Aunque todas las especies muestran foto-inhibición a 200 µMol Fotones m-2 s-1, las especies del género Navicula muestran valores más bajos del Effective Quantum yield (ϕII) que las especies del genero Nitzschia en respuesta a la variación de la intensidad de luz en el rango entre 0,5 y 100 µMol Fotones m-2 (Fig. 3). Después de 30 minutos en oscuridad, las especies del género Navicula tienen una menor capacidad de recuperación que las especies del género Nitzschia (30-40% y 70-80%, respectivamente) en el rango de intensidad de luz estudiado (Fig. 4). Proporción de pigmentos. Normalizados por Chl-a, el contenido Chl c2, es menor en las especies de Navicula que el las del género Nitzschia, y el contenido de Fucoxantina es similar en todas en el rango de intensidad de luz estudiado. En todas las especies no se observa una variación de estos cuocientes asociados a la variación de la intensidad de luz. (Fig 5A y B). Por otra parte, las especies Navicula cancellata y Navicula perminuta mostraron una mayor proporción de pigmentos fotoprotectores (PP: diadinoxantina, diatoxantina y ß caroteno) respecto a los pigmentos fotosintéticos (PS: Clorofilas a, c2 y Fucoxantina) que en las especies de Nitzschia, además en las Navicula esta proporción decae al aumentar la intensidad de luz (Fig. 5C). También las tasa de conversión de diadinoxantinas a Diatoxantinas expresada en la tasa de de-epoxidación

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
591
(DT/DD+DT) fue más alta en las dos especies de Navicula, y aumenta con la intensidad de luz, comparadas con las obtenidas en las especies del genero Nitzschia que muestran valores menores y similares en todo el rango de intensidad de luz estudiado (Fig. 5D).
Figura 3. Respuesta Fotosintética en las diferentes intensidades de luz. Effecive Quantum Yield (ϕII) de Navicula perminuta (círculos negros), Navicula cancellata (triángulos claros) Nitzschia sp1 (cuadrados negros) y Nitzschia sp2 (rombos claros). Figure 3. Photosynthetic response at different light intensities. Effecive Quantum Yield (ϕII) of Navicula perminuta (black dots); Navicula cancellata (clear triangles) Nitzschia sp1 (black squares) y Nitzschia sp2 (clear rhombs).
Figura 4. Porcentaje de recuperación de la respuesta fotosintética en respuesta a diferentes intensidades de luz, después
de 30 min. en oscuridad. Figure 4. Recovery of the photosynthetic response of species at different light intensities, after 30 min of darkness.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
592
Fig. 5. Variación del cuociente de pigmentos fotosintéticos y fotoprotectores en respuesta a diferentes intensidades de luz. Np: Navicula perminuta; Nc: Navicula cancellata; Nsp1 y Nsp2: Nitzschia sp1 y Nitzschia sp2, respectivamente.
Figure 5. Variation of photosynthetic to photoprotective pigments ratios at different light intensities. Np: Navicula perminuta; Nc: Navicula cancellata; Nsp1 y Nsp2: Nitzschia sp1 y Nitzschia sp2, respectively.
DISCUSIÓN Y CONCLUSIONES Las 4 especies de diatomeas bentónicas antárticas estudiadas están aclimatadas a baja intensidad de luz, y muestran foto-inhibición a los 200 µMol Fotones m-2, sin embargo, las especies mostraron una respuesta fisiológica fotosintética diferente y característica frente a la variación de la intensidad de luz. Las especies del género Navicula estarían aclimatadas a ambientes de intensidades de luz más bajas, con una menor tolerancia a la luz alta, y con mayores tasas de de-epoxidación. Por otra parte, las especies del género Nitzschia podrían acomodarse y tolerar ambientes más iluminados, con menor proporción de pigmentos fotoprotectores y con una tasa de de-epoxidación menor. Estos resultados sugieren que las diferencias en la plasticidad de esta respuesta de las especies Navicula y Nitzschia estudiadas, podrían asociarse a diferencias en sus mecanismos de foto-protección mediante síntesis de pigmentos fotoprotectores y la tasa de de-epoxidación de las xantofilas. AGRADECIMIENTOS INACH T11-08. Ernesto Molina, UTS, Sydney Australia. REFERENCIAS BIBLIOGRÁFICAS Falkowski, P., Raven, J. 1997. Aquatic Photosynthesis.Aquatic Photosynthesis, Blackwell, Oxford UK. Guillard, R. and Ryther, J. 1962. Studies on marine plankton diatoms I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve). Canadian Journal of Microbiology 8, 229-239. Petrou, K., Doblin, M. and Ralph, P. 2011. Heterogeneity in the photoprotective capacity of three Antarctic diatoms during short-term changes in salinity and temperature. Marine Biology 158, 1029-1041. Robinson, D. 1995. Microalgal light-harvesting in extreme low-light environments in McMurdo sound, Antarctica. Journal of Phycology 31, 508-520.
Underwood, G. and Kromkamp, J. 1999. Primary Production by Phytoplankton and Microphytobenthos in Estuaries. Advanced Ecology Research 29, 93-153.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
593
DESCRIPCION DEL MITOGENOMA EN Nacella concinna Y COMPARACION CON OTRAS NACELLAS SUBANTARTICAS.
(The mitogenome of Nacella concinna and the comparison with others Subantartic Nacellids)
Cárdenas, L.1, Gaitan-Espitia, J.C.1 y Cea G.1
1Instituto de Ciencias Ambientales y Evolutivas, Facultad de Ciencias, Universidad Austral de Chile. email: [email protected] RESUMEN El análisis del genoma mitocondrial ha permitido avanzar en la comprensión de las relaciones filogenéticas y filogeográficas de numerosas especies (White et al. 2011). Mas aun, solo para Metazoa actualmente existen 3.436 reportes de genomas mitocondriales completos disponibles en la base de datos de NCBI (www.ncbi.nlm.nih.gov); y esta información va creciendo constantemente debido al avance en las tecnologías de secuenciación. Estas tecnologías de ultima generación o next generation sequences (NGS) permiten obtener información genómica de manera intensiva, rápida y de alta confiabilidad a un costo bastante menor al de cualquier técnica de secuenciación tradicional (Metzker 2010). Tradicionalmente, el genoma mitocondrial de cualquier organismo es obtenido mediante la técnica de Long PCR (Hu et al. 2002) pero recientes trabajos han demostrado la utilidad de secuenciación de genomas organelares con NGS sugiriendo que este método es mas viable para la secuenciación de genomas mitocondriales básicamente debido a su alta cobertura, calidad y bajo costo (Jex et al. 2010). Pese a todo lo anterior, existen todavía grupos taxonómicos prácticamente desconocidos, en particular aquellos conformados por especies no modelos que habitan ambientes poco explorados como las zonas Subantárticas y Antárticas. El genero Nacella (Patellogastropoda: Nacellidae) actualmente esta distribuido en diferentes regiones de la Antártica y Sudamérica (González-Wevar et al. 2010). N concinna es la única especie representante de este genero en la Antártica. Dentro de las especies subantárticas, N. clypeater se distribuye en la provincia biogeográfica peruviana habitando desde el sur de Perú hasta alrededor de los 42º S coincidente con el principal quiebre biogeográfico descrito en esta área (Valdovinos & Rüth 2005). En la provincia magallánica, se distinguen al menos cuatro grupos taxonómicos siendo el mas representativo y abundante N. magallanica (Gonzalez-Wever et al. 2010). Recientes trabajos muestran que la separación de estas especies ocurrió durante el mioceno, sin evidencia que apoye la existencia de actual flujo genético entre ellos (González-Wevar et al. 2011, 2012). La mayoría de las inferencias filogeográficas están basadas en el análisis de unos pocos genes como el gen mitocondrial citocromo oxidasa I (cox1) y citocromo b (cyt B), dejando la interrogante de si estos patrones biogeográficos mayores serán también apoyados por la inclusión de nuevos genes o por el genoma mitocondrial completo. A nivel de las relaciones filogenéticas dentro del grupo Patellogastropoda, la información genética disponible también es escasa, existiendo solo un genoma mitocondrial completo hasta la fecha (Simison et al. 2006). En este trabajo determinamos el mitogenoma de N. concinna, caracterizando 12 genes que codifican proteínas (protein coding genes o PCGs) en base al análisis de información obtenida desde una base de datos de marcas de secuencias expresadas o expressed seqenced tag (EST) generadas por pirosecuenciación utilizando una plataforma 454 GS FLX (Roche). Después de la anotación usando blast2go (Götz et al. 2011), se extrajeron los contigs que se anotaron relacionadas a genes mitocondriales. A partir de esta base de datos y utilizando el programa CLC se caracterizaron los

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
594
genes presentes en N. concinna, N. magallánica y N. clypeater. La reconstrucción de los genes se realizo mediante un tBlastx utilizando un e-value de 1x10-5 y utilizando a las especies Lottia digitalis (Patellogastropoda) y Haliotis rubra (Vestigastropoda) como taxa de referencia. Para la comparación entre los genes, se determinaron el numero de sitios polimórficos y la divergencia promedio entre secuencias y especies. En total, 3792 contig asociados al mitogenoma, de los cuales 1485 fueron encontrados en N. concinna, 1478 para N magallánica y 829 para N clypeater. En todas las especies el mayor numero de secuencias fue para el gen citocromo oxidasa I (Cox1) siendo al menos 5 veces mayor que los otros PCGs (Figura 1), sugiriendo un mayor nivel de expresión de este gen probablemente debido a su naturaleza de gen constitutivo (Järving et al. 2004). Interesantemente, se identificaron 12 de los 13 PCGs, no encontrándose en ninguna de las especies analizadas el gen ATP8. Varios moluscos y nematodos carecen de este gen, como por ejemplo Cassotrea Virginica, sugiriéndose que el gen podría haber sido transferido al núcleo o constituye una versión completamente modificada lo que impide su identificación (Milbury & Gaffney 2005). Aunque recientemente Breton et al (2010) logra identificar este gen en Mytilus spp., posterior secuenciación y análisis permitirán confirmar la ausencia de este gen en Nacella. Figura 1: Número de contigs mitocondriales encontrados en Nacella concinna, tomando como referencia 2 especies de
gastrópodos más cercanos. Figure 1: Contigs mitochondrial number found in Nacella concinna, referring two closest gastropod species.
AGRADECIMIENTOS Proyecto INACH T22-10. REFERENCIAS Breton S, Stewart DT & WR Hoeh. 2010. Characterization of a mitochondrial ORF from the gender-associated mtDNAs of Mytilus spp. (Bivalvia: Mytilidae): Identification of the “missing” ATPase 8 gene. Marine Genomics 3: 11-18. González-Wever CA, Nakano T, Cañete JI & E Poulin. 2010. Molecular phylogeny and historical biogeography of Nacella (Patellogastropoda: Nacellidae) in the Southern Ocean. Molecular Phylogenetics and Evolution 56: 115-124. González-Wevar CA, Nakano T, Cañete JI & E Poulin. 2011. Concerted genetic, morphological and ecological diversification in Nacella limpets in the Magellanic Province. Molecular Ecology 20: 1936-1951. Götz S et al. 2011. B2G-FAR, a species centered GO annotation repository. Bioinformatics 27: 919-924. Hu M, Chilton NB & RB Gasser. 2002. Long PCR-based amplification of the entire mitochondrial genome from single parasitic nematodes. Mol. Cell Probes 16: 261-267.
num
ero
de c
ontig
s
Genes que codifican proteinas (PCGs)
0
50
100
150
200
250
300
350
Cox1
Cox2
Cox3
Cytb
Nadh1
Nadh2
Nadh3
Nadh4
Nadh4
Nadh5
Nadh6
Atp6
Atp8
Haliotis rubra
Lottia digitalis

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
595
Jex A, Hall R, Timothy D, Littlewood J & RB Gasser. 2010. An integrated pipeline for next-generation sequencing and annotation of mitochondrial genomes. Nucleic Acids Research 38(2): 522-533. Jarving R, Jarving I, Kurg R, Brash AR & N Samel. 2004. On the Evolutionary Origin of Cyclooxygenase (COX) Isozymes. The Journal of Biological Chemistry 279: 13624-13633. Metzker M. 2010. Sequencing technologies-the next generation. Nature Review Genetic 11:31-46. Milbury CA & PM Gaffney. 2005. Complete Mitochondrial DNA Sequence of the Eastern Oyster Crassostrea virginica. Marine Biotechnology 7: 697-712. Simison WB, Lindberg DR, Boore JL. 2006. Rolling circle amplification of metazoan mitochondrial genomes. Mol Phylogenet Evol 39:562-567. White TR, Conrad MM, Tseng R, Balayan S, Golding R, de Frias Martins AM & BA Dayrat. 2011. Ten new complete mitochondrial genomes of pulmonates (Mollusca: Gastropoda) and their impact on phylogenetic relationships. BMC Evolutionary Biology 11:295.

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
596
Índice de Autores Abdala, et al. ….................................................................................................................................................. 483 Acuña-Rodríguez ............................................................................................................................................... 12 Aguayo-Lobo, .................................................................................................................................................... 510 Alcántara-Hernández, et al. ............................................................................................................................... 274 Aldea, et al. ....................................................................................................................................................... 514 Alfonso, et al. .................................................................................................................................................... 447 Alvarez, et al. .................................................................................................................................................... 397 Alvarez, et al. .................................................................................................................................................... 401 Antelo, et al. ...................................................................................................................................................... 59 Arancibia, et al. .................................................................................................................................................. 432 Arata, et al. ........................................................................................................................................................ 249 Arredondo, et al. ................................................................................................................................................ 278 Arriagada, et al. ................................................................................................................................................ 258 Bastos, et al. ...................................................................................................................................................... 414 Bastos, et al. ...................................................................................................................................................... 190 Bergstrom, et a. .................................................................................................................................................. 21 Bezada, et al. ..................................................................................................................................................... 134 Bozo, et al. ....................................................................................................................................................... 418 Braga, et al. ....................................................................................................................................................... 405 Bruzzone, et al. ................................................................................................................................................. 282 Cabrera, et al. .................................................................................................................................................... 286 Caiafa, et al. ...................................................................................................................................................... 518 Calisto-Ulloa, et al.............................................................................................................................................. 485 Callejas, et al...................................................................................................................................................... 26 Canário, et al. ..................................................................................................................................................... 82 Cárdenas, et al. ................................................................................................................................................ 592 Carmona, et al. .................................................................................................................................................. 140 Castro, et al. ………………………….……………………………………………………………………… 159 Cavieres, et al. ................................................................................................................................................... 22 Cebuhar , et al. .................................................................................................................................................. 67 Chacón, et al. .................................................................................................................................................... 145 Cherene, et al. .................................................................................................................................................. 567 Chiang, et al. .................................................................................................................................................... 489 Chiarandini, et al. .............................................................................................................................................. 5 Cid, et al. ........................................................................................................................................................... 288 Contreras, et al. .................................................................................................................................................. 101 Corcuera, et al. .................................................................................................................................................. 40 Costa………… .................................................................................................................................................. 252 Cornejo, et al. ................................................................................................................................................... 292 Correa, et al. ..................................................................................................................................................... 522 Costa, et al. ...................................................................................................................................................... 408 Costa, et al. ....................................................................................................................................................... 170 Cuba-Díaz, et al. ............................................................................................................................................... 435 Cunachi, et al. .................................................................................................................................................. 296 Dennett, et al. ................................................................................................................................................... 300 Di Fonzo, et al. .................................................................................................................................................. 71 Di Fonzo, et al. .................................................................................................................................................. 460 Dias, et al. .......................................................................................................................................................... 218 Díaz-Vásquez, et al. ........................................................................................................................................... 117 Díaz, et al. .......................................................................................................................................................... 554 E. Johnson, et al. ............................................................................................................................................... 451 Egas, et al. ......................................................................................................................................................... 525 Espejo, et al. ...................................................................................................................................................... 492 Ferrés, et al. ....................................................................................................................................................... 303 Ferrés, et al. ....................................................................................................................................................... 29 Franco, et al. ...................................................................................................................................................... 57 Franzosi, et al. ................................................................................................................................................... 306 Fuentes, et al. .................................................................................................................................................... 439 Gallardo-Cerda, et al. ....................................................................................................................................... 309

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
597
García-Sancho, .................................................................................................................................................. 21 Garcia, et al. ...................................................................................................................................................... 202 Godoy, et al. ...................................................................................................................................................... 389 Gómez-Fuentes, et al. ........................................................................................................................................ 1 González-Wevar, et al. ..................................................................................................................................... 267 González-Wevar, et al. ...................................................................................................................................... 558 González, et al. .................................................................................................................................................. 75 González, et al. .................................................................................................................................................. 85 González, et al. .................................................................................................................................................. 312 Goyanes, et al..................................................................................................................................................... 137 Gran-Scheuch, et al. .......................................................................................................................................... 496 Guerra, ............................................................................................................................................................... 226 Gutiérrez, ........................................................................................................................................................... 214 Gutiérrez, et al. .................................................................................................................................................. 53 Haro, et al. ......................................................................................................................................................... 527 Hernández, et al. ............................................................................................................................................. 316 Jaña, et al. .......................................................................................................................................................... 149 Jara, et al. ......................................................................................................................................................... 78 Jorcín, et al. ..................................................................................................................................................... 320 Klagges, et al. ................................................................................................................................................... 49 Lagos, et al. ...................................................................................................................................................... 323 Lagos, et al........................................................................................................................................................ 464 Lavín, et al. ....................................................................................................................................................... 329 Lavín, et al. ....................................................................................................................................................... 326 Lee, et al. ........................................................................................................................................................... 210 Leiva, et al. ........................................................................................................................................................ 531 Lemmertz, et al. ................................................................................................................................................. 16 Leppe, et al. .................................................................................................................................................... 171 Leyton, et al. .................................................................................................................................................. 333 Loza, et al. ..................................................................................................................................................... 535 MacDonell, et al. ............................................................................................................................................ 130 Manfroi, et al. .................................................................................................................................................. 422 Manograsso, , et al. ......................................................................................................................................... 237 Mansilla , et al. ................................................................................................................................................. 45
Mansilla-Vera, et al. .......................................................................................................................................... 425 Mark, et al. ........................................................................................................................................................ 233 Márquez, et al. .................................................................................................................................................. 539 Martinez Alvarez, et al. .................................................................................................................................... 381 Martínez, et al. .................................................................................................................................................. 385 Mattatal, et al. .................................................................................................................................................. 500 Michel, et al. .................................................................................................................................................... 89 Mónica Puentes, et al. ....................................................................................................................................... 9 Monserrate-Maggi, et al. ................................................................................................................................... 335 Montiel, et al. .................................................................................................................................................. 254 Morales, et al. ................................................................................................................................................... 337 Moreno, et al. .................................................................................................................................................. 194 Moura, et al...................................................................................................................................................... 241 Muñoz-Villagrán, et al. .................................................................................................................................... 121 Mura-Jornet, et al. …........................................................................................................................................ 563 Nielsen, ............................................................................................................................................................ 266 Nishida, et al. ................................................................................................................................................... 175 Olivares, et al. .................................................................................................................................................. 543 Órdenes-Aenishanslins, et al. .......................................................................................................................... 113 Orgeira, ........................................................................................................................................................... 547 Orgeira, et al. ................................................................................................................................................... 162 Ortiz, et al. ....................................................................................................................................................... 182 Pavlov, et al. .................................................................................................................................................... 341 Pellizari, et al. .................................................................................................................................................. 17 Peña, et al. ....................................................................................................................................................... 410 Pereira, et al. .................................................................................................................................................. 345 Pereira, et al. .................................................................................................................................................. 503 Pérez, et al. .................................................................................................................................................... 348

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
598
Perfetti-Bolaño, et al. ….................................................................................................................................. 504 Pętlicki, et al. .................................................................................................................................................. 153 Petsch, et al. .................................................................................................................................................. 452 Pinedo, et al. .................................................................................................................................................. 572 Plaza, et al. ...................................................................................................................................................... 109 Poulin, et al. .................................................................................................................................................... 271 Pozo, et al. ...................................................................................................................................................... 351 Rabbia, et al. .................................................................................................................................................. 105 Ramírez, et al. .................................................................................................................................................. 551 Ramírez, et al. .................................................................................................................................................. 63 Ratti, et al. ....................................................................................................................................................... 37 Reguero, et al. .................................................................................................................................................. 179 Reyes, et al. ...................................................................................................................................................... 41 Rivadeneira, et al. ............................................................................................................................................. 262 Rodrigo, et al. ................................................................................................................................................... 456 Rodríguez-Marconi, et al. ................................................................................................................................. 33 Rodríguez, et al. ............................................................................................................................................... 355 Rojas, et al. ...................................................................................................................................................... 97 Rojas, et al. ...................................................................................................................................................... 393 Rondanelli-Reyes, et al. ................................................................................................................................... 245 Rovirosa, et al. .................................................................................................................................................. 467 Ruiz Barlett, et al. .............................................................................................................................................. 186 Sáez, et al. ....................................................................................................................................................... 475 Salamene, et al. .................................................................................................................................................. 93 Salas-Berrios, et al. …....................................................................................................................................... 576 Salazar, et al. ..................................................................................................................................................... 580 Sanhueza, et al. ................................................................................................................................................. 198 Sanhueza, et al. .................................................................................................................................................. 479 Seixas, et al. ...................................................................................................................................................... 470 Sierra, et al. ...................................................................................................................................................... 584 Silva, et al. ...................................................................................................................................................... 553 Silva, et al. ....................................................................................................................................................... 508 Simões, ............................................................................................................................................................. 129 Soto, et al. ....................................................................................................................................................... 359 Souza, et al. ...................................................................................................................................................... 473 Suarez, et al. ................................................................................................................................................... 509 Suwalsky, et al. .................................................................................................................................................. 429 Torres, et al. ....................................................................................................................................................... 363 Uribe, et al. ........................................................................................................................................................ 588 Urtubia, et al. .................................................................................................................................................... 367 Valdivia, et al..................................................................................................................................................... 125 Valdivia, et al. .................................................................................................................................................. 206 Valenzuela-Guerra, et al. ............................................................................................................................. 166 Vargas, et al. .................................................................................................................................................. 371 Velásquez, et al. .. .......................................................................................................................................... 443 Vera, et al. ...................................................................................................................................................... 222 Verdugo, et al. .................................................................................................................................................. 229 Viteri, et al. ..................................................................................................................................................... 374 Zamora, et al. .................................................................................................................................................. 155 Zúñiga, et al. .................................................................................................................................................. 377

AVANCES EN CIENCIA ANTÁRTICA LATINOAMERICANA LIBRO DE RESÚMENES VII CONGRESO LATINOAMERICANO DE CIENCIA ANTÁRTICA
599




















