
Auditory motion affects visual motion perception in a speeded discrimination task
7
Exp Brain Res (2007) 178:415–421 DOI 10.1007/s00221-007-0919-y 123 RESEARCH NOTE Auditory motion aVects visual motion perception in a speeded discrimination task Daniel Sanabria · Juan Lupiáñez · Charles Spence Received: 11 August 2006 / Accepted: 19 February 2007 / Published online: 13 March 2007 © Springer-Verlag 2007 Abstract Transient auditory stimuli have been shown to inXuence the perception of ambiguous 2D visual motion displays (the bouncing-disks eVect; e.g. Sekuler et al. in Nature 385:308, 1997). The question addressed here was whether continuous moving auditory stimuli can also inXu- ence visual motion perception under the same experimental conditions. In Experiment 1, we used a modiWcation of Sanabria et al.’s (Exp Brain Res 157:537–541, 2004) para- digm (involving an indirect behavioural measure of the bouncing-disks eVect), in which the 2D visual display was presented together with either a brief tone, a continuous moving sound, or in the absence of any form of auditory stimulation. Crucially, the results showed that, together with the eVect of the brief tone on bouncing trials, the pres- ence of the continuous moving sound speeded-up partici- pants’ responses on streaming trials as compared to the brief tone or no sound conditions. The results of a second experiment revealed that the eVect of the continuous mov- ing sound reported in Experiment 1 was not caused simply by the presence of continuous auditory stimulation per se. Keywords Crossmodal integration · Multisensory perception · Motion perception Introduction The bouncing-disks eVect (Sekuler et al. 1997), whereby the perception of an ambiguous 2-dimensional (2D) visual motion display is aVected by the presence of a brief audi- tory stimulus, represents an oft-cited example of the multi- sensory inXuences on unimodal (visual) motion perception. In the present study, we utilized this crossmodal phenome- non to investigate whether continuous auditory motion (as opposed to the brief pulse of auditory stimulation typically used in previous studies) can also inXuence the perceived direction of visual motion in a speeded discrimination task. In their inXuential paper, Sekuler et al. (1997) described an experiment in which they presented an ambiguous visual motion display, consisting of two identical visual stimuli moving towards each other. These Wlled black disks coin- cided and then moved apart again. Given that the two objects were identical and were presented using a 2D visual display, the perceptual interpretation of the display was ambiguous: the objects could either be perceived as coin- ciding, and then bouncing oV each other (bouncing trials), or else as coinciding and then passing through each other (streaming trials; see Bertenthal et al. 1993). The crucial result to emerge from Sekuler et al.’s study was that when a brief auditory stimulus (a 2.5 ms duration click) was pre- sented at approximately the same time as the two objects coincided, the probability of participants reporting a bounc- ing percept (as measured by subjective report) increased signiWcantly with respect to the ‘default’ predominant streaming percept (cf. Ramachandran and Anstis 1983). According to Ramachandran and Anstis (1983), the per- ceptual ambiguity produced by the 2D visual motion dis- play at the moment of collision is solved by ‘default’ in line with Newton’s Wrst law of motion, which states that an object moving at a uniform velocity in a given direction D. Sanabria · C. Spence Department of Experimental Psychology, University of Oxford, Oxford, UK D. Sanabria (&) · J. Lupiáñez Departamento de Psicología Experimental, Facultad de Psicología, Universidad de Granada, 18071 Granada, Spain e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Auditory motion affects visual motion perception in a speeded discrimination task

Exp Brain Res (2007) 178:415–421
DOI 10.1007/s00221-007-0919-yRESEARCH NOTE
Auditory motion aVects visual motion perception in a speeded discrimination task
Daniel Sanabria · Juan Lupiáñez · Charles Spence
Received: 11 August 2006 / Accepted: 19 February 2007 / Published online: 13 March 2007© Springer-Verlag 2007
Abstract Transient auditory stimuli have been shown toinXuence the perception of ambiguous 2D visual motiondisplays (the bouncing-disks eVect; e.g. Sekuler et al. inNature 385:308, 1997). The question addressed here waswhether continuous moving auditory stimuli can also inXu-ence visual motion perception under the same experimentalconditions. In Experiment 1, we used a modiWcation ofSanabria et al.’s (Exp Brain Res 157:537–541, 2004) para-digm (involving an indirect behavioural measure of thebouncing-disks eVect), in which the 2D visual display waspresented together with either a brief tone, a continuousmoving sound, or in the absence of any form of auditorystimulation. Crucially, the results showed that, togetherwith the eVect of the brief tone on bouncing trials, the pres-ence of the continuous moving sound speeded-up partici-pants’ responses on streaming trials as compared to thebrief tone or no sound conditions. The results of a secondexperiment revealed that the eVect of the continuous mov-ing sound reported in Experiment 1 was not caused simplyby the presence of continuous auditory stimulation per se.
Keywords Crossmodal integration · Multisensory perception · Motion perception
Introduction
The bouncing-disks eVect (Sekuler et al. 1997), wherebythe perception of an ambiguous 2-dimensional (2D) visualmotion display is aVected by the presence of a brief audi-tory stimulus, represents an oft-cited example of the multi-sensory inXuences on unimodal (visual) motion perception.In the present study, we utilized this crossmodal phenome-non to investigate whether continuous auditory motion (asopposed to the brief pulse of auditory stimulation typicallyused in previous studies) can also inXuence the perceiveddirection of visual motion in a speeded discrimination task.
In their inXuential paper, Sekuler et al. (1997) describedan experiment in which they presented an ambiguous visualmotion display, consisting of two identical visual stimulimoving towards each other. These Wlled black disks coin-cided and then moved apart again. Given that the twoobjects were identical and were presented using a 2D visualdisplay, the perceptual interpretation of the display wasambiguous: the objects could either be perceived as coin-ciding, and then bouncing oV each other (bouncing trials),or else as coinciding and then passing through each other(streaming trials; see Bertenthal et al. 1993). The crucialresult to emerge from Sekuler et al.’s study was that when abrief auditory stimulus (a 2.5 ms duration click) was pre-sented at approximately the same time as the two objectscoincided, the probability of participants reporting a bounc-ing percept (as measured by subjective report) increasedsigniWcantly with respect to the ‘default’ predominantstreaming percept (cf. Ramachandran and Anstis 1983).
According to Ramachandran and Anstis (1983), the per-ceptual ambiguity produced by the 2D visual motion dis-play at the moment of collision is solved by ‘default’ in linewith Newton’s Wrst law of motion, which states that anobject moving at a uniform velocity in a given direction
D. Sanabria · C. SpenceDepartment of Experimental Psychology, University of Oxford, Oxford, UK
D. Sanabria (&) · J. LupiáñezDepartamento de Psicología Experimental, Facultad de Psicología, Universidad de Granada, 18071 Granada, Spaine-mail: [email protected]
123

416 Exp Brain Res (2007) 178:415–421
will preserve its state unless an external force acts tochange that state (Newton 1687; cited in Ramachandranand Anstis 1983). Sekuler et al.’s (1997) results demon-strate that this bias inherent in the visual system toward thestreaming percept can be overcome by the simple presenta-tion of a brief tone (at the moment of coincidence), thusincreasing the subjective report of bouncing.
The bouncing-disks eVect demonstrates that the percep-tion of a visual object in motion can be inXuenced by thepresence of a transient auditory stimulus. The general viewin the literature (e.g. see Shimojo and Shams 2001) is thatonly transient stimuli would inXuence the perception ofvisual motion under conditions of ambiguity. The questionaddressed in the present study is whether continuous audi-tory motion information can also bias visual motion percep-tion in the event of perceptual uncertainty.1 While visionusually dominates motion processing (cf. Soto-Faraco andKingstone 2004), we hypothesized that the presence ofauditory motion information would favour the perceptionof streaming in the bouncing-disks eVect under conditionsof temporary visual uncertainty. We tested the hypothesisof whether, under the presence of (temporary) uncertaintyin the visual modality, auditory motion information wouldinXuence visual motion processing as it provides the mostreliable information at that particular moment (cf. Heronet al. 2004).
Finding that auditory motion information modulatesvisual motion processing in the present study would favourthe idea that crossmodal perception is a Xexible (rather thana ‘winner-takes-all’) process, in which each sensory inputreceives a ‘weight’ depending on its respective reliability ata particular moment and in a particular situation (cf. Batta-glia et al. 2003; Ernst and Banks 2002). This idea makessense from an ecological perspective, as perception will bebiased by the sensory modality that provides the most con-sistent information (i.e. with the least variability) concern-ing a particular (relevant) object or event at a given timeand in a given context. Following on from this rationale, weexpect that auditory motion will inXuence visual motiongiven the presence of temporal uncertainty in the visualmodality (see below). Moreover, any such Wnding woulddirectly challenge the idea that only transient auditory stim-
uli can inXuence the perception of visual motion (see Shim-ojo and Shams 2001).
Researchers have traditionally used subjective reports tomeasure the eVect of brief sounds on the visual perceptionof moving objects. That is, participants have typically beenrequired to explicitly report whether they saw the two mov-ing objects streaming or bouncing oV each other in eachtrial (e.g. Sekuler et al. 1997). Therefore, one might arguethat the eVect reported in previous studies is the conse-quence of the task demands present in the experimental sit-uations, or any other kind of cognitive bias, rather than agenuine crossmodal eVect (that does not depend on the sub-jective interpretation of the experimental situation by theparticipant). In fact, a number of researchers have pointedout (Prinzmetal et al. 2005; Watt 1991) that it is importantto try and isolate any perceptual eVects of interest from thedecisional or cognitive biases that might inXuence one’sdependent measure.
In the present study, rather than using the typical para-digm found in previous studies to test our hypothesis (e.g.Sekuler et al. 1997), we used an indirect measure of thebouncing-disks eVect which circumvented the potentialmethodological limitations associated with the use of sub-jective report measures present in a number of previousstudies. Following Sanabria et al. (2004), we measured par-ticipants’ RTs. In this way, we reduced the inXuence of anycognitive bias, which helps to isolate any perceptual eVectunder study. In Experiment 1, we used modiWcation of San-abria et al.’s task,2 in which, in addition to the typical brieftone condition and a no sound condition, we also includedthe crucial new experimental manipulation: A sound mov-ing horizontally which was presented together with thevisual motion display in order to test our predictions. Inparticular, we expected that in addition to the eVect of thebrief sound on bouncing trials, auditory motion wouldaVect participants’ response latencies to streaming trials (incomparison to the no sound condition and the brief tone
1 It should be noted that the level of ambiguity present in our set-up wasreduced with respect to Sekuler et al.’s (1997) study, since they usedtwo disks of the same colour while we used two disks of diVering col-our. In their study, the presence of visual ambiguity started at the mo-ment of collision between the two disks and lasted until the end of thetrial. In our case, there was ambiguity only during the period in whichthe two disks were occluded. In any case, we still consider that, duringthe occlusion of the visual moving stimuli in our study there was per-ceptual uncertainty. Uncertainty that was suYcient to give rise to theinXuence of the auditory information on the visual judgment measuredby the RT to the reappearance of one of the visual stimuli (see Methodsfor details).
2 The participants in Experiment 1 also completed a replication of San-abria et al.’s (2004) study in which the disks only moved along the hor-izontal axis. The order of the two experiments was counterbalancedacross participants. Crucially, there was a signiWcant interaction be-tween the motion and sound factors, F(1,19) = 9.56, P < 0.01. Pairwisecomparisons revealed that in the brief tone condition, response laten-cies were signiWcantly faster on bouncing trials (384 ms) than onstreaming trials (M = 419 ms), t(19) = 5.05, P < 0.001. This eVect wasnot present in the no sound condition, in which there was no signiWcantdiVerence in mean RTs between the bouncing (M = 401 ms) andstreaming (416 ms) trials, t(19) = 1.84, P = 0.08. These results par-tially replicate Sanabria et al.’s data, since in their study, participantsresponded more rapidly to streaming trials in the no sound conditionthan in the brief tone condition. As noted by the authors (see Sanabriaet al. 2004, Footnote 1) in order to obtain the crossover interaction (i.e.faster RT in the bouncing trials when the sound is present versus fasterRT in the streaming trials when the sound is absent) a paradigm inwhich diVerent motion trajectories are used may be more appropriate.
123

Exp Brain Res (2007) 178:415–421 417
condition). Experiment 2 was conducted in order to test analternative explanation of the results of the moving soundcondition obtained in Experiment 1. Namely, whether theeVect of the moving sound (compared to the no sound con-dition and the brief tone condition) on participants’ RTs tostreaming trials might simply have been caused by the pres-ence of a continuous auditory stimulus throughout themovement of the visual stimulus.
Experiment 1
Methods
Participants
Twenty participants (10 men; age range: 18–33 years, meanage of 25 years), from the University of Oxford took part inthis experiment. All of the participants reported normalhearing and normal or corrected-to-normal vision, andreceived a 5-pound UK sterling gift voucher in exchangefor their participation. The experiment took »20 min tocomplete and was performed in accordance with the ethicalstandards laid down in the 1964 Declaration of Helsinki.All of the participants gave their informed consent prior totheir inclusion in the study.
Apparatus and materials
The participants sat 50 cm from a computer monitor in adark experimental booth. A chin-rest was used to keep theparticipant’s head still and at a constant distance from themonitor. A yellow circle (diameter = 0.3° of visual angle)presented in the middle of the screen was used as a centralWxation point. The visual occluder consisted of a black rect-angle (6° of visual angle horizontally £ 23° vertically) pre-sented from directly behind the Wxation point. Two disks(diameter = 0.8°), one blue and the other red, were pre-sented symmetrically one from the left side of the computerscreen and the other from the right side (15° from centralWxation). The disks moved towards the black occluder at29° per second, and then disappeared behind it. Two loud-speaker cones, one placed directly to either side of the com-puter screen, were used to present the auditory stimuli. A14 ms (i.e. equivalent to one screen refresh) 450-Hz puretone (70 dB[A], as measured from the participant’s ear)was used in the brief tone condition. On post-experimentalquestioning, the participants reported perceiving the soundas emanating from the centre of the screen (i.e. fromapproximately the same location as the two disks wouldhave met behind the occluder). In the crucial moving soundcondition, auditory motion was induced by gradually modi-fying the relative intensity of the sound from each of the
two loudspeakers (e.g. from 100% to the left and 0% to theright to 0% to the left and 100% right loudspeaker; cf. Soto-Faraco et al. 2004) while presenting a 450 Hz (70 dB[A] atparticipants’ ear level) auditory stimulus for a duration of1,026 ms. The presentation of the auditory and visual stim-uli and the timing of responses were controlled by the E-Prime software package.
Procedure
On each trial, the red and blue disks appeared randomly onopposite sides of the screen level with the Wxation point(see Fig. 1). The disks moved toward each other, followinga horizontal trajectory. The disks disappeared behind theoccluder just prior to their coincidence and then reappearedafter a delay of 168 ms. The disks could either reappearfrom behind the opposite side of the occluder moving in thesame direction as they had been travelling before occlusion,or else they could reappear from the same side of theoccluder, moving in the opposite direction to that prior totheir occlusion. The presence of the sound (present versusabsent), and the direction in which the disks moved afterocclusion (same versus diVerent) was randomly intermixedacross trials.
The participants were instructed to attend to either thered or blue disk (counterbalanced across participants), andto respond to the side (left versus right) on which the diskreappeared from behind the occluder (by pressing the “z” or
Fig. 1 Schematic illustration of the 2D visual displays used in Exper-iments 1 and 2. In Experiment 1, the visual movement occurred in thepresence of either a brief tone, a moving sound, or in the absence of anyauditory stimulation. In Experiment 2, the moving sound was replacedby a continuous stationary sound
Bouncing trial
Streaming trial
SOUND
TIME
TIME
SOUND
T1 T2 T3
T1 T2 T3
123
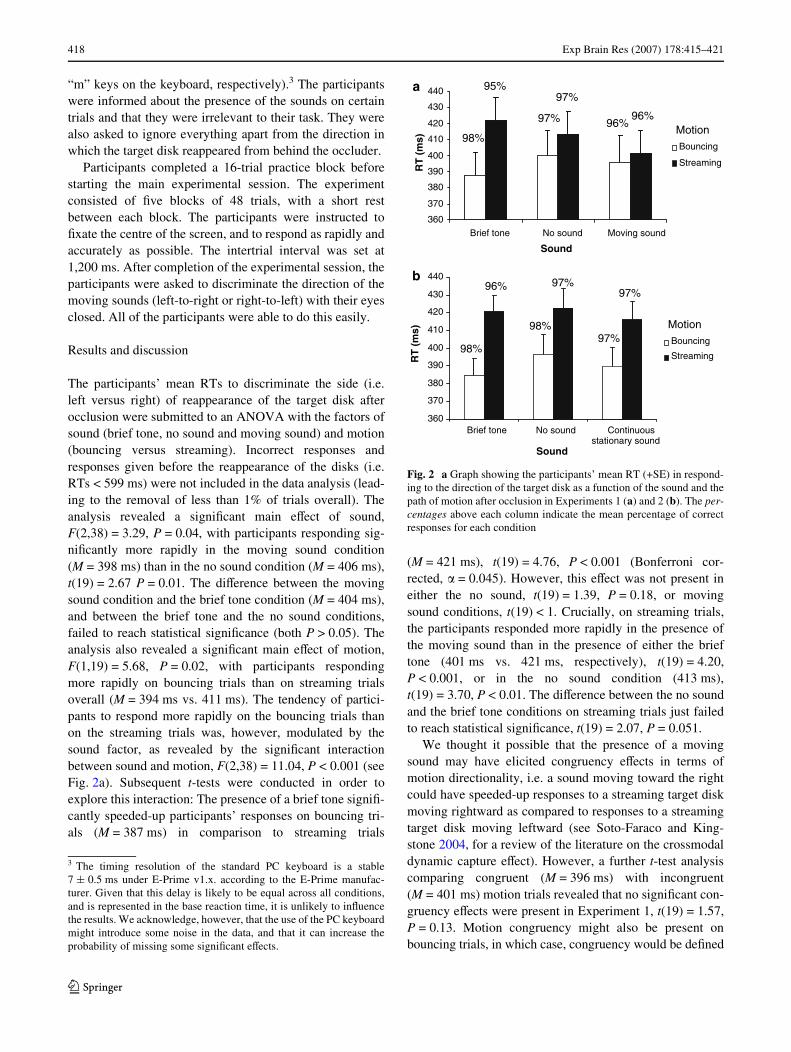
418 Exp Brain Res (2007) 178:415–421
“m” keys on the keyboard, respectively).3 The participantswere informed about the presence of the sounds on certaintrials and that they were irrelevant to their task. They werealso asked to ignore everything apart from the direction inwhich the target disk reappeared from behind the occluder.
Participants completed a 16-trial practice block beforestarting the main experimental session. The experimentconsisted of Wve blocks of 48 trials, with a short restbetween each block. The participants were instructed toWxate the centre of the screen, and to respond as rapidly andaccurately as possible. The intertrial interval was set at1,200 ms. After completion of the experimental session, theparticipants were asked to discriminate the direction of themoving sounds (left-to-right or right-to-left) with their eyesclosed. All of the participants were able to do this easily.
Results and discussion
The participants’ mean RTs to discriminate the side (i.e.left versus right) of reappearance of the target disk afterocclusion were submitted to an ANOVA with the factors ofsound (brief tone, no sound and moving sound) and motion(bouncing versus streaming). Incorrect responses andresponses given before the reappearance of the disks (i.e.RTs < 599 ms) were not included in the data analysis (lead-ing to the removal of less than 1% of trials overall). Theanalysis revealed a signiWcant main eVect of sound,F(2,38) = 3.29, P = 0.04, with participants responding sig-niWcantly more rapidly in the moving sound condition(M = 398 ms) than in the no sound condition (M = 406 ms),t(19) = 2.67 P = 0.01. The diVerence between the movingsound condition and the brief tone condition (M = 404 ms),and between the brief tone and the no sound conditions,failed to reach statistical signiWcance (both P > 0.05). Theanalysis also revealed a signiWcant main eVect of motion,F(1,19) = 5.68, P = 0.02, with participants respondingmore rapidly on bouncing trials than on streaming trialsoverall (M = 394 ms vs. 411 ms). The tendency of partici-pants to respond more rapidly on the bouncing trials thanon the streaming trials was, however, modulated by thesound factor, as revealed by the signiWcant interactionbetween sound and motion, F(2,38) = 11.04, P < 0.001 (seeFig. 2a). Subsequent t-tests were conducted in order toexplore this interaction: The presence of a brief tone signiW-cantly speeded-up participants’ responses on bouncing tri-als (M = 387 ms) in comparison to streaming trials
(M = 421 ms), t(19) = 4.76, P < 0.001 (Bonferroni cor-rected, � = 0.045). However, this eVect was not present ineither the no sound, t(19) = 1.39, P = 0.18, or movingsound conditions, t(19) < 1. Crucially, on streaming trials,the participants responded more rapidly in the presence ofthe moving sound than in the presence of either the brieftone (401 ms vs. 421 ms, respectively), t(19) = 4.20,P < 0.001, or in the no sound condition (413 ms),t(19) = 3.70, P < 0.01. The diVerence between the no soundand the brief tone conditions on streaming trials just failedto reach statistical signiWcance, t(19) = 2.07, P = 0.051.
We thought it possible that the presence of a movingsound may have elicited congruency eVects in terms ofmotion directionality, i.e. a sound moving toward the rightcould have speeded-up responses to a streaming target diskmoving rightward as compared to responses to a streamingtarget disk moving leftward (see Soto-Faraco and King-stone 2004, for a review of the literature on the crossmodaldynamic capture eVect). However, a further t-test analysiscomparing congruent (M = 396 ms) with incongruent(M = 401 ms) motion trials revealed that no signiWcant con-gruency eVects were present in Experiment 1, t(19) = 1.57,P = 0.13. Motion congruency might also be present onbouncing trials, in which case, congruency would be deWned
3 The timing resolution of the standard PC keyboard is a stable7 § 0.5 ms under E-Prime v1.x. according to the E-Prime manufac-turer. Given that this delay is likely to be equal across all conditions,and is represented in the base reaction time, it is unlikely to inXuencethe results. We acknowledge, however, that the use of the PC keyboardmight introduce some noise in the data, and that it can increase theprobability of missing some signiWcant eVects.
Fig. 2 a Graph showing the participants’ mean RT (+SE) in respond-ing to the direction of the target disk as a function of the sound and thepath of motion after occlusion in Experiments 1 (a) and 2 (b). The per-centages above each column indicate the mean percentage of correctresponses for each condition
95%
360
370
380
390
400
410
420
430
440
Brief tone No sound Moving sound
Bouncing
Streaming
98%
97%
97% 96%96%
Motion
Bouncing
Streaming
Motion
Continuous360
370
380
390
400
410
420
430
440
Brief tonestationary sound
Sound
Sound
No sound
RT
(m
s)R
T (
ms)
98%
96%
98%
97%97%
97%
a
b
123

Exp Brain Res (2007) 178:415–421 419
as the relation between the direction of the moving soundand the direction of the target disk after occlusion. How-ever, a t-test comparison between the congruent and incon-gruent trials of this kind (M = 388 ms vs. 396 ms,respectively) revealed no signiWcant diVerence, t(19) =1.66, P = 0.11. None of the terms in an ANOVA conductedon the accuracy data reached signiWcance.
However, before drawing any Wrm conclusions from theresults of Experiment 1, an alternative explanation for theresults obtained in the moving sounds condition needs to beconsidered Wrst. It could be argued that the faster RTsobserved on the streaming trials in the moving sound condi-tion relative to the other two conditions may simply have beencaused by the continuous presence of a sound throughout thetrial (rather than by the presence of auditory motion informa-tion per se). In other words, participants’ responses on trials inwhich the attended disks continued along the same path afterocclusion may have been biased by the presence of continuousauditory stimulation, i.e. the continuity in visual motion (asopposed to the change in the direction that occurred on bounc-ing trials) may have been biased by the continuity in the audi-tory domain. Experiment 2 was conducted in order to evaluatethis possibility, which, if it were to have been the case, wouldargue against an eVect of auditory motion per se on visualmotion perception (cf. Soto-Faraco and Kingstone 2004).
Experiment 2
Methods
Participants
Twenty-six new participants (11 males; age range: 19–33,mean 25 years) took part in Experiment 2.
Apparatus, materials, design and procedure
These were exactly the same as in the previous experimentexcept for the following: In the continuous stationary soundcondition, a 450 Hz auditory stimulus was presentedthroughout the trial for 1,026 ms (70 dB[A] at participant’sear level). Participants completed a 12-trial practice blockbefore starting the main experiment. After completion ofthe experiment, the participants were asked whether theyperceived auditory motion or not in the continuous station-ary sound condition. None of the participants reported per-ceiving any kind of auditory motion.
Results and discussion
The participants’ mean RTs for discriminating the side (leftversus right) of appearance of the target disk after occlusion
were submitted to an ANOVA with the factors of sound(brief tone, no sound and continuous stationary sound) andmotion (bouncing versus streaming). Incorrect responsesand responses given before the reappearance of the disks(i.e. RTs < 599 ms) were not included in the analysis, lead-ing to the removal of less than 1% of trials overall. Therewas a main eVect of sound, F(2,50) = 4.63, P = 0.01, withthe participants responding more rapidly in the brief tonecondition (M = 402 ms) than in the no sound condition(409 ms), t(25) = 2.19, P = 0.03. Participants alsoresponded signiWcantly faster in the continuous stationarysound condition (M = 402 ms) than in the no sound condi-tion, t(25) = 3.45, P < 0.01. The participants also respondedsigniWcantly more rapidly on bouncing trials than onstreaming trials overall (M = 390 ms vs. 419 ms), givingrise to a signiWcant main eVect of motion, F(1,25) = 39.79,P < 0.001. As in Experiment 1, the interaction betweensound and motion was also signiWcant, F(2,50) = 3.76,P = 0.02 (see Fig. 2b). In order to investigate this interac-tion, we conducted pairwise comparisons (Bonferroni cor-rected, � = 0.045). These revealed that in the brief tonecondition, participants responded signiWcantly more rapidlywhen the disks bounced oV one another (M = 384 ms) thanwhen they streamed (M = 420 ms), t(25) = 7.65, P < 0.001.Participants also responded signiWcantly more rapidly inbouncing trials than in streaming trials in both the nosound, t(25) = 5.42, P < 0.001, and continuous stationarysound conditions, t(25) = 5.13, P < 0.001. However, thediVerence in the mean RT between bouncing and streamingtrials was signiWcantly larger in the brief tone condition(mean diVerence of 36 ms) than in either the no sound con-dition (mean diVerence of 26 ms), t(25) = 2.39, P = 0.02, orthe continuous stationary sound condition (mean diVerenceof 26 ms), t(25) = 2.09, P = 0.04.
We conducted a between-experiments analysis to com-pare the performance of participants in Experiments 1 and 2in order to test the hypothesis that the eVect of the movingsound on participants’ responses in Experiment 1 mighthave been caused simply by the presence of a continuoussound being presented throughout the trial, and not by anyperception of auditory movement. If this were to have beenthe case then we should not expect to Wnd any signiWcantdiVerence in performance between the moving sound con-dition of Experiment 1 and the continuous stationary soundcondition of Experiment 2. In contrast, if the presence ofauditory motion (regardless of the presence of a continuoussound) inXuenced participants’ performance, one wouldhave expected to Wnd a signiWcant diVerence in the continu-ous sound conditions between the two experiments. Thebetween-experiment ANOVA conWrmed our hypothesis,revealing a signiWcant interaction between Experiment,motion and sound, F(2,88) = 3.24, P = 0.04. Crucially, thediVerence in participants’ RTs between bouncing and
123

420 Exp Brain Res (2007) 178:415–421
streaming trials in the moving sound condition of Experi-ment 1 was signiWcantly smaller than in the continuous sta-tionary sound condition of Experiment 2 (mean diVerenceof 5 ms vs. 26 ms, respectively), F(1,44) = 5.03, P = 0.02.That is, RTs to streaming trials were aVected to a greaterextent by the presence of a continuous sound in Experiment1 (moving) than in Experiment 2 (stationary). Importantly,however, this between-experiments diVerence was not pres-ent in either the brief tone, F < 1, or no sound conditions,F(1,44) = 1.81, P = 0.18. Therefore, the presence of themoving sound in Experiment 1 aVected participants’ per-formance (i.e. it speeded-up responses on streaming trials)to a greater extent than did the presence of a continuous sta-tionary sound in Experiment 2. The between-experimentsANOVA also highlighted signiWcant main eVects of sound,F(2,88) = 6.48, P < 0.01, and motion, F(1,44) = 31.81,P < 0.001, as well as a signiWcant interaction betweensound and motion, F(2,88) = 15,64, P < 0.001. The interac-tions between experiment and motion and between experi-ment and sound were not signiWcant (both P > 0.05).
A further between-experiments ANOVA comparing par-ticipants’ performance in Experiments 1 and 2 in the brieftone and no sound conditions revealed no interactionbetween experiment, sound and motion, F(1,44) = 2.23,P = 0.14, suggesting that the diVerence in performancebetween the two experiments was speciWcally attributableto the diVerent pattern of results between the moving sound(Experiment 1) and continuous stationary sound conditions(Experiment 2).
An ANOVA on the accuracy data only revealed a sig-niWcant main eVect of motion, F(1,25) = 10.30, P = 0.003,with participants responding more accurately on bouncingtrials than on streaming trials overall (98% vs. 96% correct,respectively).
Conclusions
The results of the two experiments reported here conWrmthe view that information concerning auditory motion caninXuence people’s responses to visual motion stimuli. Inparticular, the presence of a continuous moving sound over-came the participants’ general bias to respond more rapidlyto bouncing trials than to streaming trials. One might arguethat the presence of auditory motion information in thepresent study did not facilitate the perception of streamingwith respect to the perception of bouncing, as there was nodiVerence between the two conditions in terms of partici-pants’ RTs. We note, however, that there was a general ten-dency among our participants to respond signiWcantly morerapidly to bouncing trials than to streaming, even in theabsence of any auditory stimulus being presented. Theimportant result in our study is that the presence of auditory
motion information overcame this tendency, resulting in aslowing of RTs on bouncing trials and a facilitation of RTson streaming trials (with respect to brief tone and no soundconditions in Experiment 1). It remains possible that varia-tions in certain parameters of our study (such as improvingthe quality of the auditory motion information presented toparticipants) would result in faster response to streamingtrials than to bouncing trials in the auditory motion condi-tion. In any case, when compared to the baseline no soundcondition and the brief tone condition, the presentation ofauditory motion information aVected participants’responses to streaming trials. Crucially, this result cannotbe accounted for solely by the presence of continuous audi-tory stimulation, as revealed by the results of the between-experiments comparison.
Our results have both theoretical and methodologicalimplications. First, they extend previous accounts of theinXuence of multisensory integration on unimodal (visual)motion processing, demonstrating that not only can tran-sient auditory stimulation inXuence the resolution of anyperceptual uncertainty regarding the direction of visualmotion, but information consistent with continuous audi-tory motion can also have a signiWcant eVect. This resultagrees with recent accounts suggesting the existence ofneural systems specialized for the processing of auditorymotion information (e.g. GriYths et al. 1998). Second, ourstudy represents a step forward with respect to Sanabriaet al.’s (2004) study in terms of extending their methodol-ogy to the analysis of the eVect of auditory motion informa-tion on visual motion processing using an objectivemeasure (participants’ RTs), in contrast to all previousstudies on this matter that have used subjective reports (e.g.Sekuler et al. 1997).
The results of the present study also add to the extantevidence regarding the multisensory integration of motioninformation. While motion judgments in vision are typi-cally little inXuenced by the presence of motion informa-tion in other sensory modalities (see Soto-Faraco andKingstone 2004, for a review), the present results show thatunder the appropriate conditions (i.e. under conditions ofperceptual uncertainty), visual motion processing can beaVected by auditory motion information. Meyer and Wuer-ger (2001) also reported a study in which auditory motioninformation was shown to aVect visual motion processing.In their study, participants had to discriminate the directionof motion of a random dot kinematogram. The analysis oftheir results showed that the eVect of auditory motion wasonly present under conditions of high uncertainty regardingthe direction of visual motion (i.e. at low levels of motioncoherence), and that auditory motion primarily aVected par-ticipants’ response criterion, and not their perception. Con-trary to Meyer and Wuerger’s study, we used a paradigm inwhich the direction of visual motion was never ambiguous
123

Exp Brain Res (2007) 178:415–421 421
(apart from during the occlusion of the moving disks), andthat was primarily based on participants’ RTs. In fact, ourparticipants were able to discriminate the direction ofmotion in the experiments presented here very accurately(see Fig. 2), and, importantly, the RT eVects were not sim-ply caused by a speed/accuracy trade-oV. Moreover, of the20 participants tested in Experiment 1, only four reportedsubjectively feeling any inXuence of the moving sound ontheir responses to the direction of the attended disk (i.e. thatwhen listening to a moving sound they were expecting thedisks to follow the same path). It therefore appears that inour study the presence of a moving sound aVected partici-pants’ perception of visual motion rather than simply bias-ing their criterion for responding.
Our study shows that auditory motion can inXuencevisual motion perception in a speeded discrimination task.Under conditions of perceptual uncertainty, visual motionprocessing can be aVected by motion information availablein a diVerent sensory modality (cf. James and Blake 2004).This Wts with the view that our perception of the externalworld depends on the relative reliability of the various sen-sory inputs involved in a given situation. Introducinguncertainty, as in the present study (temporarily occludingthe visual stimuli), or reducing the quality of the sensoryinformation in the visual modality (e.g. Ernst and Banks2002; Heron et al. 2004), increases the likely inXuence ofother sensory modalities, with the Wnal purpose of main-taining coherent perception and behaviour (see Bedford2004). In conclusion, our results add further evidence to thehypothesis that the interaction/integration of informationfrom diVerent sensory modalities involves a Xexible, ratherthan a winner-takes-all, mechanism (cf. Welch and Warren1980). Moreover, the present study demonstrated that audi-tory motion information, as compared to auditory tran-sients, can also modulate the perception of 2D visualmotion.
Acknowledgements This research was supported by the SpanishMinisterio de Educación y Ciencia (Research Project - BSO2002-04308-C02-02- to J. L.). Correspondence regarding this article should be direct-ed to: Daniel Sanabria, Departamento de Psicología Experimental y
Fisiología del Comportamiento, Facultad de Psicología, Universidadde Granada, 18071, Granada, Spain. E-mail: [email protected].
References
Battaglia PW, Jacobs RA, Aslin RN (2003) Bayesian integration of vi-sual and auditory signals for spatial localization. J Opt Soc Am A20:1391–1397
Bedford FL (2004) Analysis of a constraint on perception, cognition,and development: one object, one place, one time. J Exp Psychol:Hum Percept Perform 30:907–912
Bertenthal B, Banton T, Bradbury A (1993) Directional bias in the per-ception of translating patterns. Perception 22:193–207
Ernst MO, Banks MS (2002) Humans integrate visual and haptic infor-mation in a statistically optimal fashion. Nature 415:429–433
GriYths TD, Rees G, Rees A, Green GGR, Witton C, Rowe D, et al(1998) Right parietal cortex is involved in the perception of soundmovement in humans. Nat Neurosci 1:74–79
Heron J, Whitaker D, McGraw PV (2004) Sensory uncertainty governsthe extent of audio-visual interaction. Vision Res 44:2875–2884
James TW, Blake R (2004) Perceiving object motion using vision andtouch. Cogn AVect Behav Neurosci 4:201–207
Meyer GF, Wuerger SM (2001) Cross-modal integration of auditoryand visual motion signals. Neuroreport 12:2557–2560
Prinzmetal W, McCool C, Park S (2005) Attention: reaction time andaccuracy reveal diVerent mechanisms. J Exp Psychol Gen134:73–92
Ramachandran VS, Anstis SM (1983) Extrapolation of motion path inhuman visual perception. Vision Res 23:83–85
Sanabria D, Correa A, Lupiáñez J, Spence C (2004) Bouncing orstreaming? Exploring the inXuence of auditory cues on the inter-pretation of ambiguous visual motion. Exp Brain Res 157:537–541
Sekuler R, Sekuler AB, Lau R (1997) Sound alters visual motion per-ception. Nature 385:308
Shimojo S, Shams L (2001) Sensory modalities are not separate modal-ities: plasticity and interactions. Curr Opin Neurobiol 11:505–509
Soto-Faraco S, Kingstone A (2004) Multisensory integration of dy-namic information. In: Calvert GA, Spence C, Stein BE (eds) Thehandbook of multisensory processes. MIT Press, Cambridge,MA, pp 49–68
Soto-Faraco S, Spence C, Kingstone A (2004) Cross-modal dynamiccapture: congruency eVects in the perception of motion acrosssensory modalities. J Exp Psychol: Hum Percept Perform 30:330–345
Watt RJ (1991) Understanding vision. Academic Press, LondonWelch RB, Warren DH (1980) Immediate perceptual response to inter-
sensory discrepancy. Psychol Bull 88:638–667
123




















