
Mutations in STAT3 and diagnostic guidelines for hyper-IgE syndrome
Upload
independentCategory
view
0download
0
Research Communication
Association of the Glucocorticoid Receptor with STAT3, C/EBPb,and the Hormone-responsive Element Within the Rat HaptoglobinGene Promoter During the Acute Phase Response
Jelena Arambasic, Goran Poznanovic, Svetlana Ivanovic-Matic, Desanka Bogojevic,
Mirjana Mihailovic, Aleksandra Uskokovic and Ilijana GrigorovDepartment of Molecular Biology, Institute for Biological Research, University of Belgrade, Belgrade, Serbia
Summary
Upregulation of haptoglobin (Hp) expression in the rat dur-
ing the acute phase (AP) response is the result of synergistic
effects of IL-6–, IL-1b–, and corticosterone-activated signaling
pathways. IL-6 signaling terminates in cis–trans interactions of
the Hp gene hormone-responsive element (HRE) with tran-
scription factors STAT3 and C/EBPb. The aim of this study
was to examine the unresolved molecular mechanism of gluco-
corticoid action. A 3-fold rise in serum corticosterone at 2 and
4 h of the AP response induced by turpentine administration
preceded a 2.3-fold increase in the rate of Hp gene transcrip-
tion at 12 h that was accompanied by a 4.8-fold increase in
glucocorticoid receptor (GR), the appearance of an 86-kDa
STAT3 isoform and 3.9-, 1.9-, and 1.7-fold increased amounts
of 91-kDa STAT3, 35- and 42-kDa C/EBPb isoforms in the
nucleus. These events resulted in 4.6- and 2.5-fold increased
Hp levels in the liver and serum at 24 h. HRE affinity chro-
matography and immunoblot analysis revealed that maximal
occupancy of the HRE with GR, STAT3, and C/EBPb at 12 h
correlated with increased transcriptional activity of the Hp
gene. Coimmunoprecipitation experiments showed that acti-
vated GR established de novo interaction with STAT3 isoforms
while GR–C/EBPb interactions observed during basal tran-
scription increased during the AP response. Computer analysis
of the HRE disclosed two potential GR-binding sites: one
overlapping STAT3, another adjacent to a C/EBPb-binding
site. This finding and the experimental results suggest that
activated GR through direct interactions with STAT3 and
C/EBPb, participates in Hp gene upregulation as a transcrip-
tional coactivator. � 2010 IUBMB
IUBMB Life, 62(3): 227–236, 2010
Keywords acute phase response; glucocorticoid receptor; haptoglo-
bin; STAT3; C/EBPb; hormone-responsive element.
INTRODUCTION
Haptoglobin (Hp) is a major hemoglobin binding and trans-
port protein that belongs to the evolutionary conserved group of
acute phase (AP) plasma proteins (1, 2). These proteins are gen-
erally viewed as being relevant in providing protective functions
during tissue injury and inflammation. Hp fulfils this role by
binding free hemoglobin that is released from erythrocytes and
thereby restricts its oxidant activity (3). Hp also inhibits cathep-
sin, serves as a chaperone, assists in iron recycling, and
regulates the immune cell response (4, 5). After acute systemic
tissue injury, the concentration of Hp in the plasma increases
three to sixfold (6, 7). This increase has primarily been attrib-
uted to the transcriptional induction of the Hp gene in response
to AP mediators. The regulation of Hp gene transcription is
complex and both tissue and species specific (8). In cultured rat
liver cells, maximal expression of the Hp gene is achieved by
the combined action of glucocorticoids, IL-6, and IL-1b (9).
Sevaljevic et al. (10) revealed that adrenalectomized Wistar rats
exhibit unchanged basal level of Hp expression and its full
expression in response to turpentine-induced inflammation,
which suggests that AP-cytokines are the principal mediators of
Hp gene expression. Dexamethasone treatment alone in different
cell lines has no effect on Hp expression (11); whereas, in rats,
it promoted incomplete expression of Hp (9, 10, 12). This data
implicate that the glucocorticoids acted as factors that in inflam-
mation enhance the AP cytokine-induced stimulation of Hp
gene expression. The glucocorticoid- and IL-6–responsive ele-
ment is localized in the promoter-proximal region at 2170 to
256 bp relative to the transcription start site (6). The HRE that
is also referred to as fragment ABC consists of three separate,
cooperatively interacting cis-acting sequences designated as
Address correspondence to: Ilijana Grigorov; Department of Molecu-
lar Biology, Institute for Biological Research, University of Belgrade,
Bulevar Despota Stefana 142, 11060 Belgrade, Serbia. Tel: 1381-11-
20-78-301. Fax: 1381-11-27-61-433. E-mail: [email protected]
Received 30 October 2009; accepted 13 January 2010
ISSN 1521-6543 print/ISSN 1521-6551 online
DOI: 10.1002/iub.313
IUBMB Life, 62(3): 227–236, March 2010

regions A, B, and C, each of which are capable of binding a
distinct set of transcription factors under basal and inflammatory
states (13). IL-6– and dexamethasone-responsive elements that
could not be physically separated were localized to the pro-
moter-proximal region, 2146 to 256 (6). Sequence analysis of
this region revealed the existence of two types of IL-6–response
elements: type I that is responsible for binding constitutive and
AP-inducible transcription factors that belong to the CCAAT-
enhancer binding protein (C/EBP) family (14, 15), and type II
that binds the signal transducers and activators of transcription
(STAT) 3 and 5b (16, 17) proteins. Dexamethasone acts through
a specific glucocorticoid-response element (GRE); however, no
sequence information is available regarding the functional GRE
in the HRE region of the rat Hp gene (6). Colocalization of glu-
cocorticoid and IL-6 responsiveness in the rat Hp gene suggests
that their synergistic relationship during transcriptional activa-
tion of the Hp gene is due to the presence of common transcrip-
tional regulatory elements and synergistic interactions between
the glucocorticoid receptor (GR), the principal mediator of
the physiological actions of glucocorticoid hormones, and
IL-6–activated, HRE-bound transcription factors.
The GR protein that is not bound to its hormone predomi-
nantly resides in the cytoplasm. Binding of glucocorticoid hor-
mones induces conformational changes in the receptor, dissocia-
tion of chaperone proteins, dimerization of the receptor, its
import into the nucleus, and DNA binding. Depending on the
cell type and promoter context, GR regulates the transcriptional
activity of target genes by two basic modes of action (18, 19).
GRs can be recruited as homodimers to GREs in the promoter
region of target genes where GRE is an inverted repeat of a
hexameric nucleotide sequence separated by three base pairs
(AGAACAnnnTGTTCT) (20). Once localized to the promoter,
the receptor mediates transcriptional activation or inhibition of
the target gene either directly through contacts between the GR
transactivation domains and transcription factors, or indirectly
by recruiting transcriptional coactivators (19, 21, 22). The ab-
sence of a DNA binding site for the GR in many glucocorti-
coid-modulated genes has raised the question of the mechanism
that mediates the responses to glucocorticoids. Experimental
evidence suggests that the effect of the GR can be modulated
through cross-talk with members of other transcription factor
families, without the need for DNA binding (23, 24). Of the
transcription factors that are expressed in liver cells and which
could be major partners for activated GR are AP-1 (25), nuclear
factor-jB (NF-jB) (26), members of the STAT (23, 27, 28) and
C/EBP (29, 30) families of transcription factors. Zhang et al.
(27) showed that the synergistic effect of glucocorticoids and
interleukins primarily relies on the functional interaction
between STAT3 and GR. Subsequent observation of direct
physical interaction of GR with STAT5 proteins supports a
model in which GR acts as a coactivator for STAT5-dependent
transcription following glucocorticoid hormone stimulation of
liver cells (31). This event plays a key role in controlling body
growth.
To obtain a better understanding of the molecular mecha-
nisms that underlie Hp gene transcription, we examined the
mode(s) of action of the GR, which plays a central role in the
upregulation of the rat Hp gene by glucocorticoid hormones.
Using the sterile model of systemic inflammation induction by
turpentine administration (32), which effectively mimics the
in vitro stimulation of liver cells with proinflamatory cytokines
and dexamethasone, we examined dynamic associations of the
GR with major gene regulatory proteins STAT3 and C/EBPband the hormone-responsive element (HRE) of the rat Hp gene
during the AP response.
EXPERIMENTAL PROCEDURES
Experimental Animals and Treatment
The study was carried out on a 2.5-month-old adult male
Wistar rats weighing between 230 and 270 g. The animals were
kept at constant temperature, humidity, and 12-h dark/light
intervals. Tap water and standard laboratory chow were avail-
able ad libitum. The AP response was induced by a subcutane-
ous injection of turpentine oil (1 lL/g body weight) in the
lumbar region of rats (five per group). Control rats were rou-
tinely injected with pyrogen-free saline. The animals were
killed at 2, 4, 12, and 24 h after the injection. All animal proce-
dures were approved by the Committee for Ethical Animal Care
and Use of the Institute for Biological Research, Belgrade,
which acts in accordance with the Guide for the Care and Use
of Laboratory Animals published by the US National Institute
of Health (NIH Publication No. 85/23, revised 1986).
Measurement of Serum Hp Concentrations
The serum levels of Hp were assessed by rocket immunoe-
lectrophoresis with anti-human Hp antibody (Sigma–Aldrich,
Milwaukee, WI), which is cross-reactive with rat Hp (33). The
method is based on the immunoprecipitation reaction between
the Hp present in the serum and the polyspecific anti-Hp anti-
body incorporated in the agarose gel in regions of their optimal
concentration and is characterized by the formation of immuno-
precipitation peaks. The area under the formed immunoprecipi-
tation peak was used to quantify the relative Hp concentration
in the serum. The peaks were drawn out on tracing paper and
their areas were quantified. The values obtained after quantifica-
tion were expressed as the means 6 SEM from three separate
experiments.
Determination of Corticosterone Concentration
The concentration of corticosterone was determined as
described by Hodges and Sadow (34). Corticosterone was
extracted from the plasma of individual rats with chloroform,
and the chloroform was in turn extracted with 1 M NaOH,
followed by water. A fluorescent reagent (H2SO4 in 50% etha-
nol, 2.5:1 v/v) was added to the chloroform. After 2 h,
absorption was measured at 470 and 520 nm with a spectropho-
228 ARAMBASIC ET AL.

tofluorimeter (Amico-Bowman). The concentration of corticos-
terone was expressed in lg/100 mL plasma.
Isolation of Nuclei and In Vitro Nuclear Transcription
Rat liver nuclei were isolated and purified by ultracentrifuga-
tion and the in vitro nuclear transcription assay was performed
as described by Vannice et al. (35), except that the nuclei were
incubated with 18.5 MBq of uridine 50-[a-32P] triphosphate and
0.4 mM each of unlabeled adenosine, cytidine, and guanosine
50-triphosphates. The 32P-labeled RNA was hybridized to nitro-
cellulose filters containing 3 lg of linearized, denatured plasmid
DNA containing an insert complementary to rat Hp mRNA
(obtained from Dr. Heinz Baumann, Roswell Park Memorial
Institute, Buffalo, NY). The filters were hybridized with all of
the in-vitro–labeled RNA isolated from 108 nuclei of control
and treated rats. Prehybridization, hybridization, and washing of
the filters were carried out as described (35). The relative
changes of the transcriptional activities after treatment were
estimated by scanning the autoradiogram using a Phosphor
ImagerTM (Molecular Dynamics) and expressed as the average
percentage of the values obtained from control rats (taken as
100%). The values were corrected for background binding to
nonrecombinant pBR322.
Preparation of Whole Liver Homogenates
Whole liver homogenates were prepared according to Choi
and Waxman (36). Frozen rat liver tissues (200 mg) were
pooled from three animals and homogenized at 4 8C in a
Dounce tissue grinder (10 strokes) in ice-cold homogenization
buffer (10 mM Tris pH 7.6, 1 mM EDTA, and 250 mM su-
crose) containing a mixture of protease and phosphatase inhibi-
tors (5 mg/mL aprotinin, 0.1 mM PMSF, 1 mM NaVO3, and 5
mg/mL leupeptin). Samples were centrifuged at 10,000g for 20
min at 4 8C in an Eppendorf 5415 R centrifuge. The superna-
tants were divided into aliquots that were stored at 280 8C.
Preparation of Liver Nuclear Extracts
Nuclear extracts (NEs) were prepared from a pool of three
livers from control or turpentine-treated rats following the pro-
cedure elaborated by Gorski et al. (37). The tissues were
excised and homogenized in buffer containing 10 mM Hepes
pH 7.6, 25 mM KCl, 5 mM MgCl2, 1 mM spermidine, 1 mM
DTT, 1 mM EDTA, 1 mM PMSF, 2 M sucrose, and 10% glyc-
erol. The nuclei were pelleted by centrifugation of the homoge-
nate through a cushion of the same solution at 72,000g in an
SW 28 rotor (Beckman Instruments, Palo Alto, CA) for 30 min
at 4 8C. The obtained nuclear pellet was resuspended in lysis
buffer (10 mM Hepes pH 7.6, 100 mM KCl, 3 mM MgCl2, 0.1
mM EDTA, 1 mM DTT, 0.1 mM PMSF, and 10% glycerol).
The chromatin was precipitated by slow addition with constant
stirring of (NH4)2SO4, pH 7.9 to a final concentration of
0.36 M. Chromatin was sedimented by centrifugation at
82,000g in a Ti 50 rotor (Beckman) for 60 min at 4 8C. The
nucleoproteins were precipitated from the supernatant by the
addition of solid (NH4)2SO4 to final concentration of 2.6 M, fol-
lowed by sedimentation at 82,000g in a Ti 50 rotor (30 min,
4 8C). NEs were dialyzed overnight against 25 mM Hepes pH
7.6, 40 mM KCl, 0.1 mM EDTA, 1 mM DTT, and 10% glyc-
erol, resuspended in dialysis buffer and frozen in small aliquots
at 280 2C.
Immunoprecipitation
Immunoprecipitation was carried out with NEs (500 lg) thatwere precleared by incubation with control IgG and Protein
A-Sepharose. Supernatants obtained after preclearing were incu-
bated for 2 h on ice with 3 lg of monoclonal anti-GR antibody
(BuGR, Affinity BioReagents) or 1 lg of polyclonal C/EBPb(C-19) antibody (Santa Cruz Biotechnology, Santa Cruz, CA).
Protein A-Sepharose-coupled beads (Santa Cruz Biotechnology,
Santa Cruz, CA) were added for 2 h at 4 8C under constant agi-
tation. The beads were pelleted and washed three times with
lysis buffer (50 mM Tris 7.4, 150 mM NaCl, 1 mM EDTA,
0.1% TritonX-100, 1 mM PMSF, 1 lg/mL aprotinin, and 1 lg/mL leupeptin). The immunoprecipitated proteins were resus-
pended in sample buffer and after boiling were separated by
12% SDS-polyacrylamide gel electrophoresis (SDS-PAGE). The
obtained immunoprecipitates were analyzed by Western immu-
nobloting.
DNA Affinity Chromatography
For DNA affinity, column nucleotide fragment between
2165 and 249 bp from the 50-flanking region of the rat Hp
gene (6) was used as a DNA probe. This fragment, functionally
identified as a HRE, was obtained from Dr. Heinz Baumann
(Roswell Park Memorial Institute, Buffalo, NY). A DNA affin-
ity column with the rat Hp gene HRE was prepared according
to Kadonaga and Tjian (38). The HRE fragment was annealed
and ligated to obtain oligomers and then covalently coupled to
CNBr-activated Sepharose CL-2B (Amersham Pharmacia Bio-
tech, Uppsala, Sweden). The HRE affinity resin was equili-
brated in a Bio-Rad Econo-Column with dialysis buffer
(25 mM Hepes pH 7.6, 0.1 mM EDTA, 1 mM DTT, and 10%
glycerol) containing 0.1 M KCl. Equal amounts of NEs (8 mg)
prepared from the livers of control and turpentine-treated
animals were mixed with competitor salmon sperm DNA and
incubated for 10 min at room temperature. The protein–DNA
mixture was then applied to the HRE affinity column and after
30 min, HRE-bound nucleoproteins were eluted with dialysis
buffer containing 1 M KCl. Total amounts of eluted proteins
were subjected to 12% SDS-PAGE and analyzed by Western
immunobloting.
Western Immunoblot Analysis
Western immunoblot analysis was performed according to
the procedure of Towbin et al. (39). Protein concentrations were
determined according to Lowry et al. (40). Proteins (20 lg)
229ASSOCIATION OF GR WITH STAT3, C/EBP�, AND HRE

were separated by 12% SDS-PAGE and transferred to nitrocel-
lulose (Hybond-C, Amersham Pharmacia Biotech) or PVDF
membrane (Hybond-P, Amersham Pharmacia Biotech, Uppsala,
Sweden) and immunoblotted with polyclonal antibodies to GR
(M-20), STAT3 (H-190), and C/EBPb (C-19) (Santa Cruz Bio-
technology, Santa Cruz, CA). Proteins from the whole liver ho-
mogenate (40 lg) were transferred to a nitrocellulose membrane
and probed with goat anti-human Hp antibody (Sigma–Aldrich,
Milwaukee, WI). After incubation with blocking solution (0.2%
Tween 20, 50 mM Tris-HCl pH 7.6, 150 mM NaCl, and 5%
nonfat condensed milk), the membranes were incubated with
the relevant primary antibodies for 1.5 h at room temperature.
After rinsing, the blots were incubated with horseradish peroxi-
dase-conjugated anti-rabbit or anti-goat immunoglobulin as a
secondary antibody for 1 h. Immunoreactive bands were identi-
fied by an enhanced chemiluminescence (ECL) detection system
according to the manufacturer’s instructions (Santa Cruz Bio-
technology, Santa Cruz, CA). Each PVDF membrane was re-
probed according to the supplier’s protocol for reprobing.
PVDF membranes were incubated in 2% SDS, 100 mM b-mer-
captoethanol, and 62.5 mM Tris-HCl pH 6.8, for 35 min at 50
8C and then rinsed three times in blotto base buffer, blocked,
and probed again with another antibody. The blots were
scanned and the intensities of the signals were quantified using
TotalLab (Phoretix, Newcastle upon Tyne, England) electropho-
resis software (ver. 1.10).
Sequence Analysis of the Hp Gene HRE
Putative GR- and C/EBPb-binding sites in the HRE region
of the rat Hp gene were identified by computer search using
Alggen Promo software (available on the web page: www.alg-
gen.lsi.upc.es/cgi/bin/promo).
Statistical Analysis
Differences between the experimental groups were tested
using nonparametric Mann-Whitney’s U-test (Statistica v.5.0,
StatSoft, Tulsa, OK). P \ 0.05 was considered the threshold
level for significance.
RESULTS
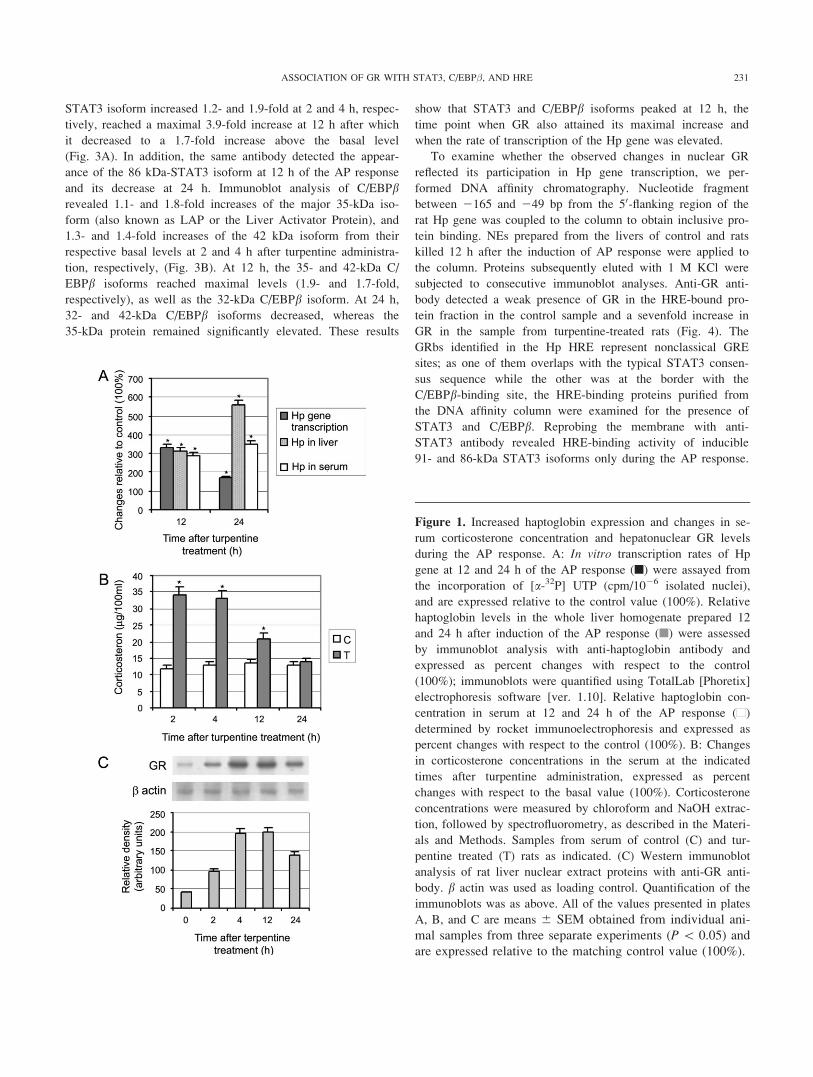
A hallmark of the AP response is the establishment of ele-
vated concentrations of AP proteins in the circulation about 24
h after its inception, and maintenance of raised concentrations
of AP proteins for a further 24 h. We assessed the dynamic
changes in Hp expression during the AP response induced by
turpentine injection by examining the changes of: (i) transcrip-
tional activity of the Hp gene, (ii) Hp protein levels in the liver
where its synthesis almost exclusively occurs (41), and (iii) Hp
serum concentration. Using an in vitro nuclear transcription
run-on assay, we observed that the transcriptional activity of the
Hp gene was significantly higher at 12 h after turpentine admin-
istration when a 232% (P \ 0.05) increase above the basal
level was observed, than at 24 h when the relative rate of Hp
gene transcription was 69% (P \ 0.05) above the basal level
(Fig. 1A, black square n). Increased relative rates of Hp gene
transcription preceded the rise in Hp protein level that was
accordingly higher at 24 h than at 12 h after induction of the
AP response. By immunoblot analysis, 210% (P \ 0.05) and
460% (P \ 0.05) increases of Hp levels in total liver homoge-
nates were observed at 12 and 24 h, respectively (Fig. 1A, gray
square n). An essentially similar profile of increase in serum
Hp concentration was observed. A higher increase (250%; P \0.05) was measured at 24 h than at 12 h after turpentine admin-
istration (190%; P\ 0.05) (Fig. 1A, white square n).
Temporal changes of Hp expression were next compared
with the dynamic changes of the central components of the
stress hormone signaling pathway—serum corticosterone con-
centrations and GR levels in the liver cell nucleus. GRs have
low affinity for glucocorticoids and their activation occurs when
glucocorticoid levels are elevated, such as following exposure
to stress (42). After 2 and 4 h of turpentine injection, 3-fold
(P\ 0.05) and 2.9-fold (P\ 0.05) increases of serum corticos-
terone concentrations above the basal level, respectively, were
measured (Fig. 1B). After 12 h, the corticosterone concentration
was slightly elevated (1.5-fold; P \ 0.05), and by 24 h, it was
at basal level. This result shows that the changes in Hp expres-
sion were preceded by a rapid increase in serum corticosterone
concentration. According to Spiga and Lightman (43), low cor-
ticosterone concentration does not have a significant effect on
nuclear GR, whereas at higher concentrations, corticosterone
increases the nuclear abundance and activity of the GR. Immu-
nobloting revealed a rapid rise in GR levels after turpentine
treatment (Fig. 1C). Four hours into the AP response, nuclear
GR was 4.8-fold higher in treated rats when compared with the
controls. Despite the decrease in corticosterone concentration
after 4 h, high levels of nuclear GR were maintained until 12 h.
The level of nuclear GR was about 15% lower at 24 h. This
result confirms that high nuclear levels of GR correlate with the
high relative rate of transcription of the rat Hp gene at 12 h of
the AP response.
The essential Hp gene cis-element is defined by the HRE
mapped at 2165/249 bp. Computer search of the HRE by All-
gen Promo revealed the presence of two potential GR-binding
sites (GRbs) (Fig. 2). One GRbs was found within the type II
IL-6 RE in subelement B at position 2106/2113 bp that partly
overlaps with the STAT3 consensus-binding site at 2109/2117
bp described by Grigorov et al. (16). The second GRbs was
found within the type I IL-6 RE in fragment C at position
270/277 bp, proximal to the C/EBPb-binding site at 262/269
bp. Previously, we showed that transcriptional induction of the
rat Hp gene during the turpentine-induced AP response is pri-
marily regulated by IL-6 REs and transcription factors C/EBPband STAT3 (14, 16). By immunoblot analysis of NEs, we
obtained the basic nuclear expression profiles of STAT3 and
C/EBPb and compared them to that of the GR during the AP
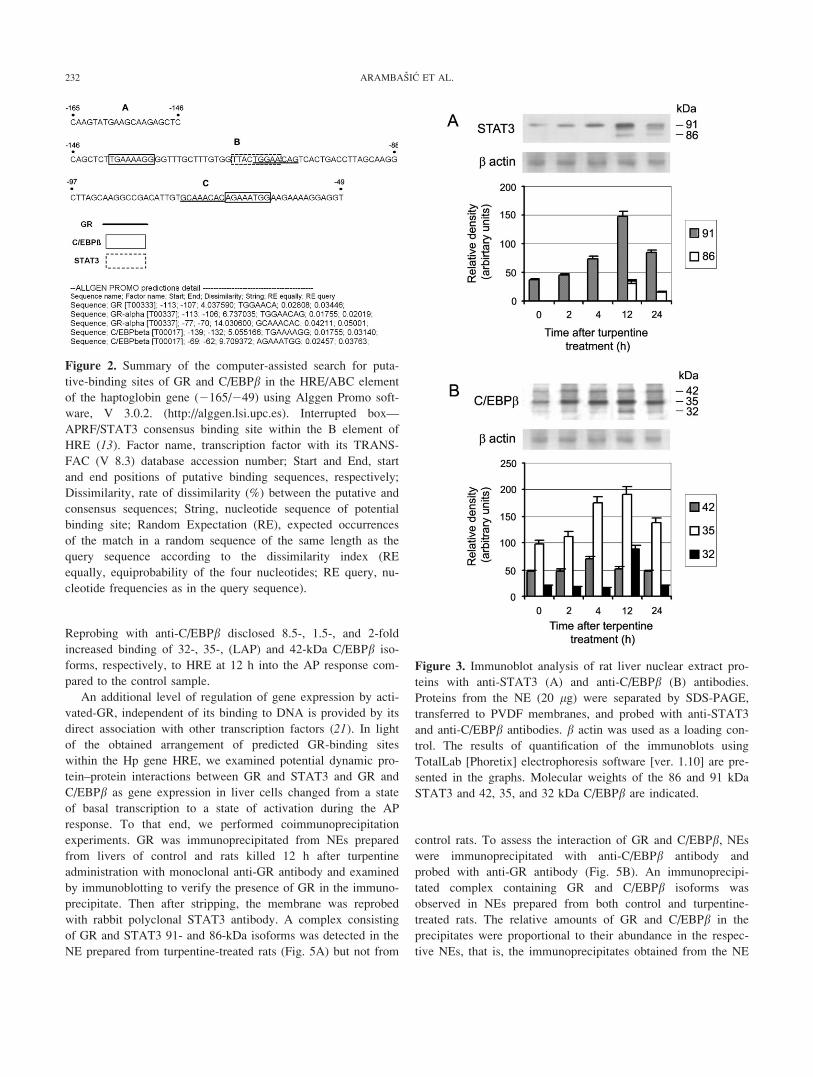
response. We observed that from the low level of constitutive
basal activity in the course of the AP response, the 91 kDa-
230 ARAMBASIC ET AL.
STAT3 isoform increased 1.2- and 1.9-fold at 2 and 4 h, respec-
tively, reached a maximal 3.9-fold increase at 12 h after which
it decreased to a 1.7-fold increase above the basal level
(Fig. 3A). In addition, the same antibody detected the appear-
ance of the 86 kDa-STAT3 isoform at 12 h of the AP response
and its decrease at 24 h. Immunoblot analysis of C/EBPbrevealed 1.1- and 1.8-fold increases of the major 35-kDa iso-
form (also known as LAP or the Liver Activator Protein), and
1.3- and 1.4-fold increases of the 42 kDa isoform from their
respective basal levels at 2 and 4 h after turpentine administra-
tion, respectively, (Fig. 3B). At 12 h, the 35- and 42-kDa C/
EBPb isoforms reached maximal levels (1.9- and 1.7-fold,
respectively), as well as the 32-kDa C/EBPb isoform. At 24 h,
32- and 42-kDa C/EBPb isoforms decreased, whereas the
35-kDa protein remained significantly elevated. These results
show that STAT3 and C/EBPb isoforms peaked at 12 h, the
time point when GR also attained its maximal increase and
when the rate of transcription of the Hp gene was elevated.
To examine whether the observed changes in nuclear GR
reflected its participation in Hp gene transcription, we per-
formed DNA affinity chromatography. Nucleotide fragment
between 2165 and 249 bp from the 50-flanking region of the
rat Hp gene was coupled to the column to obtain inclusive pro-
tein binding. NEs prepared from the livers of control and rats
killed 12 h after the induction of AP response were applied to
the column. Proteins subsequently eluted with 1 M KCl were
subjected to consecutive immunoblot analyses. Anti-GR anti-
body detected a weak presence of GR in the HRE-bound pro-
tein fraction in the control sample and a sevenfold increase in
GR in the sample from turpentine-treated rats (Fig. 4). The
GRbs identified in the Hp HRE represent nonclassical GRE
sites; as one of them overlaps with the typical STAT3 consen-
sus sequence while the other was at the border with the
C/EBPb-binding site, the HRE-binding proteins purified from
the DNA affinity column were examined for the presence of
STAT3 and C/EBPb. Reprobing the membrane with anti-
STAT3 antibody revealed HRE-binding activity of inducible
91- and 86-kDa STAT3 isoforms only during the AP response.
Figure 1. Increased haptoglobin expression and changes in se-
rum corticosterone concentration and hepatonuclear GR levels
during the AP response. A: In vitro transcription rates of Hp
gene at 12 and 24 h of the AP response (n) were assayed from
the incorporation of [a-32P] UTP (cpm/1026 isolated nuclei),
and are expressed relative to the control value (100%). Relative
haptoglobin levels in the whole liver homogenate prepared 12
and 24 h after induction of the AP response (n) were assessed
by immunoblot analysis with anti-haptoglobin antibody and
expressed as percent changes with respect to the control
(100%); immunoblots were quantified using TotalLab [Phoretix]
electrophoresis software [ver. 1.10]. Relative haptoglobin con-
centration in serum at 12 and 24 h of the AP response (n)
determined by rocket immunoelectrophoresis and expressed as
percent changes with respect to the control (100%). B: Changes
in corticosterone concentrations in the serum at the indicated
times after turpentine administration, expressed as percent
changes with respect to the basal value (100%). Corticosterone
concentrations were measured by chloroform and NaOH extrac-
tion, followed by spectrofluorometry, as described in the Materi-
als and Methods. Samples from serum of control (C) and tur-
pentine treated (T) rats as indicated. (C) Western immunoblot
analysis of rat liver nuclear extract proteins with anti-GR anti-
body. b actin was used as loading control. Quantification of the
immunoblots was as above. All of the values presented in plates
A, B, and C are means 6 SEM obtained from individual ani-
mal samples from three separate experiments (P\ 0.05) and
are expressed relative to the matching control value (100%).
231ASSOCIATION OF GR WITH STAT3, C/EBP�, AND HRE
Reprobing with anti-C/EBPb disclosed 8.5-, 1.5-, and 2-fold
increased binding of 32-, 35-, (LAP) and 42-kDa C/EBPb iso-
forms, respectively, to HRE at 12 h into the AP response com-
pared to the control sample.
An additional level of regulation of gene expression by acti-
vated-GR, independent of its binding to DNA is provided by its
direct association with other transcription factors (21). In light
of the obtained arrangement of predicted GR-binding sites
within the Hp gene HRE, we examined potential dynamic pro-
tein–protein interactions between GR and STAT3 and GR and
C/EBPb as gene expression in liver cells changed from a state
of basal transcription to a state of activation during the AP
response. To that end, we performed coimmunoprecipitation
experiments. GR was immunoprecipitated from NEs prepared
from livers of control and rats killed 12 h after turpentine
administration with monoclonal anti-GR antibody and examined
by immunoblotting to verify the presence of GR in the immuno-
precipitate. Then after stripping, the membrane was reprobed
with rabbit polyclonal STAT3 antibody. A complex consisting
of GR and STAT3 91- and 86-kDa isoforms was detected in the
NE prepared from turpentine-treated rats (Fig. 5A) but not from
control rats. To assess the interaction of GR and C/EBPb, NEswere immunoprecipitated with anti-C/EBPb antibody and
probed with anti-GR antibody (Fig. 5B). An immunoprecipi-
tated complex containing GR and C/EBPb isoforms was
observed in NEs prepared from both control and turpentine-
treated rats. The relative amounts of GR and C/EBPb in the
precipitates were proportional to their abundance in the respec-
tive NEs, that is, the immunoprecipitates obtained from the NE
Figure 2. Summary of the computer-assisted search for puta-
tive-binding sites of GR and C/EBPb in the HRE/ABC element
of the haptoglobin gene (2165/249) using Alggen Promo soft-
ware, V 3.0.2. (http://alggen.lsi.upc.es). Interrupted box—
APRF/STAT3 consensus binding site within the B element of
HRE (13). Factor name, transcription factor with its TRANS-
FAC (V 8.3) database accession number; Start and End, start
and end positions of putative binding sequences, respectively;
Dissimilarity, rate of dissimilarity (%) between the putative and
consensus sequences; String, nucleotide sequence of potential
binding site; Random Expectation (RE), expected occurrences
of the match in a random sequence of the same length as the
query sequence according to the dissimilarity index (RE
equally, equiprobability of the four nucleotides; RE query, nu-
cleotide frequencies as in the query sequence).
Figure 3. Immunoblot analysis of rat liver nuclear extract pro-
teins with anti-STAT3 (A) and anti-C/EBPb (B) antibodies.
Proteins from the NE (20 lg) were separated by SDS-PAGE,
transferred to PVDF membranes, and probed with anti-STAT3
and anti-C/EBPb antibodies. b actin was used as a loading con-
trol. The results of quantification of the immunoblots using
TotalLab [Phoretix] electrophoresis software [ver. 1.10] are pre-
sented in the graphs. Molecular weights of the 86 and 91 kDa
STAT3 and 42, 35, and 32 kDa C/EBPb are indicated.
232 ARAMBASIC ET AL.

extract prepared from the AP sample was considerably enriched
in GR and C/EBPb isoforms compared to the control sample.
Neither GR, STAT3, nor C/EBPb was found to immunoprecipi-
tate with control IgG (results not shown). The results of the
coimmunoprecipitation experiments suggest that STAT3 iso-
forms establish de novo contacts with activated GR while
C/EBPb isoforms interact with GR in the basal state and during
the burst of transcriptional activity in the course of the AP
response when a quantitative increase in protein–protein con-
tacts occurs.
DISCUSSION
We used the in vivo sterile model of inflammation induced
by turpentine administration to examine the role of the GR in
Hp gene expression. The turpentine-induced hind limb abscess
is an established model of acute inflammation in rodents (44). It
is associated with very high levels of circulating IL-6 and pro-
longed activation of the hypothalamic–pituitary–adrenal axis
(45) which leads to corticosterone elevation whose effects are
mediated by the GR, that is, whose activity depends on the
binding of the hormone. The accumulation of the GR that was
observed in the rat liver nucleus after the turpentine treatment
was the result of dissociation of the GR from chaperone pro-
teins, and its translocation to the nucleus in response to the
surge of corticosterone that peaked at 4 h post-turpentine
administration. While the serum corticosterone level started to
decrease after 4 h, it remained significantly elevated at 12 h.
This finding is in accordance with the data published by Veni-
haki et al. (46). Elevated corticosterone concentration coincided
with maximal transcriptional activity of the Hp gene. The
decrease in transcriptional activity of the Hp gene by about
50% at 24 h was accompanied by the establishment of basal
corticosterone levels and a decrease of nuclear GR. These
results point to the involvement of the corticosterone-activated
GR in transcriptional activation of the rat Hp gene. Previously,
Baumann et al. (11) described dexamethasone attenuation of
GR expression in long-term steroid-treated cells. The authors
ascribed it to a change in transcriptional activity of genes under
GR control and to post-translational processing and increased
stability of GR mRNA. Similar results were obtained by Karten
et al. (47) who also examined long-term exposure of male Wis-
tar rats to high corticosterone concentration. When the treat-
ment with hormone was limited to 24 h, the reduction of GR
expression was not significant. We observed that the level of
GR was decreased by about 15% at 24 h after the turpentine
treatment, whereas at the same time the transcriptional activity
of the Hp gene remained increased, that is, nearly twofold
Figure 4. Immunoblot analysis of nuclear proteins with anti-
GR, anti-STAT3, and anti-C/EBPb antibodies after DNA-affin-
ity chromatography. Equal quantities (8 mg) of nuclear proteins
were applied to the DNA affinity column and eluted with 1 M
KCl as described in Materials and Methods. Twenty micro-
grams of NE and the total amount of the corresponding eluted
fractions (1 M) were separated by SDS-PAGE, transferred to
PVDF membranes, and probed with an anti-GR, anti-STAT3,
and anti-C/EBPb antibodies. Lane 1, control NE; lane 2, control
NE eluted with 1 M KCl; lane 3, NE obtained 12 h after AP
induction; lane 4, NE obtained 12 h after AP induction eluted
with 1 M KCl.
Figure 5. Coimmunoprecipitation of GR-STAT3 and C/EBPb–GR from nuclear extracts from control and rats 12 h post-
turpentine administration. A: Immunoprecipitation of GR was
performed with precleared nuclear extracts and BuGR antibody.
Precipitates were analyzed with anti-GR and anti-STAT3 anti-
bodies. B: Immunoprecipitation of C/EBPb was performed with
precleared nuclear extracts and anti-C/EBPb antibody. Precipi-
tates were analyzed with anti-C/EBPb and anti-GR antibodies.
Immunoprecipitations were performed according to the proce-
dure described in Materials and Methods. Lane 1, control NE;
lane 2, immunoprecipitates from control NE; lane 3, NE
obtained 12 h after AP induction; lane 4, immunoprecipitates
from NE obtained 12 h after AP induction.
233ASSOCIATION OF GR WITH STAT3, C/EBP�, AND HRE

higher than basal transcription. The result shows the importance
that GR levels, together with other trans-acting factors, have in
maintaining an efficient rate of transcription of the Hp gene.
The importance of GR levels in regulating the promoter ele-
ments of rat a1-acid glycoprotein and b fibrinogen genes was
shown (11).
As a result of increased IL-6 levels after turpentine injection
(results not shown) and initiation of the signal transduction cas-
cade, Hp gene transcription is stimulated by increased activities
of transcription factors STAT3 and C/EBPb (14–16). The accu-
mulation of GR and establishment of elevated levels of nuclear
STAT3, which is sometimes also referred to as the AP response
factor, and of CEBPb/LAP were observed at 12 h after induc-
tion of the AP response (Fig. 4). A strong correlation of the
time course of changes in nuclear abundance of these proteins
was observed until 24 h after turpentine administration. Accord-
ing to DNA affinity chromatography, the occupancy of the Hp
gene HRE by these proteins also correlated with the activation
of the Hp gene transcription.
Increasing evidence demonstrates the overlap of IL-6 and
glucocorticoid signal transduction pathways in the regulation of
AP protein gene activities (48, 49). Although there is no func-
tional GRE within the Hp gene HRE/ABC element, computer
search revealed the presence of two GRbs that could be respon-
sible for AP-related activation of Hp gene transcription. Not
every site where GR specifically binds to the DNA represents a
GRE. The binding sites for transcription factors on the DNA
are regions whose affinity for the given protein are elevated
because of specific nucleotide sequences that distinguish it from
other regions (50). The GRbs found by Allgen Promo program
within the IL-6–responsive element B (2146/288) of the Hp
gene HRE have a typical hexanucleotide motif of a new
variant of the palindromic nucleotide sequence G/AGnA-
CAnnnTGTTCT. The GR can interact with this hexanucleotide
(GGAACA) as a monomer, however, the GR-hexanucleotide
complex is not very stable. Potential GRbs detected in element
B overlap with the STAT3 AP-response element consensus
binding site called APRE, which is responsible for the forma-
tion of the AP-specific DNA-protein complex through interac-
tions of both STAT3 isoforms (16). As dexamethasone alone
does not increase Hp gene transcription in different cell lines
(11), but is capable to induce incomplete expression of Hp in
vivo (10), we assumed that GR controls Hp-gene–induced activ-
ity through interactions with STAT3. STAT3 synergy with the
GR for full transcriptional increase of the a2-macroglobulin (49,
51) and human b fibrinogen gene (52) has been documented. In
both instances, the GR does not increase transcription alone but
through interaction with STAT3 and without direct DNA bind-
ing. Coimmunoprecipitation of both STAT3 isoforms and GR
only during the AP response, together with the results from
HRE affinity chromatography imply that one of the functions of
the GR in activation of the Hp gene is to recruit and maintain
STAT3 bound to the APRE. In this case, the GR acts as a coac-
tivator. Zhang et al. (27) described the association of IL-6–acti-
vated STAT3 with ligand-bound GR in a trans-activating/sig-
naling complex that functions through the IL-6 RE. Similar
results were obtained by Savoldi et al. (29) who showed a glu-
cocorticoid-dependent interaction of the GR and C/EBPb at the
steroid response unit of the a1-acid glycoprotein promoter that
contains several binding sites for C/EBPb, some of which over-
lap with the GRE.
The stability of the GR-hexanucleotide complex can be
increased by heterodimerization of the GR with other transcrip-
tion factors. Thereby, the binding of the GR monomer to the
hexanucleotide motif is stabilized by another protein that pos-
sesses a binding site adjacent to the hexanucleotide (50). GRbs
found within element C of the Hp gene HRE contain a hexanu-
cleotide sequence (CAAACA) that slightly differs from the
sequence found in element B (GGAACA). After comparing the
two hexanucleotide sequences, we observed a substitution of
GG ? CA in the GRbs residing in element C of the Hp gene.
The ACA motif within this sequence is conserved, rendering
this hexanucleotide, according to Merkulov and Merkulova
(50), a possible binding site for the GR monomer. As the
C/EBPb-binding site within the type I IL-6 RE of the Hp gene
HRE is adjacent to the GRbs, and since the 35 kDa-C/EBPbisoform displays strong HRE binding affinity in both basal and
especially under AP conditions, we assume that C/EBPb is
responsible for the binding of the GR monomer to the GRbs.
This assumption is supported by coimmunoprecipitation
experiments which point to cooperativity between the GR and
C/EBPb at both basal and AP conditions. At present, we cannot
distinguish between the contributions of the GR and C/EBPb in
basal regulation of this transcription unit. Nisho et al. (53)
reported that C/EBPb could exert not only a cooperative action
on this site but also an inhibitory effect on the activity of the
GR. However, GR induction of reporter gene expression in a
dose-dependent manner (18) allows us to conclude that the
amount of GR–C/EBPb complexes in the basal state is not suf-
ficient to increase Hp gene activity. This supports our results
(15) that show that the main C/EBP family transcription factor
involved in the regulation of Hp gene activity in noninduced
conditions is C/EBPa which binds to the same binding site as
C/EBPb, and which is more abundant than C/EBPb. No interac-
tion between GR and C/EBPa was detected (results not shown).
We conclude that full transcriptional activity of the Hp gene
requires increased amount of activated GR that can interact
with STAT3 besides C/EBPb.Tissue-specific responses to glucocorticoid hormones play
important roles in homeostasis. It is, therefore, important to elu-
cidate the contribution of GRs that mediate the effects of gluco-
corticoids to homeostasis—how they are induced by extracellu-
lar signals, how their functioning is regulated in a tissue-specific
manner and to identify their target genes. It will be interesting
to compare the Hp promoter with promoters of other AP protein
genes that are responsive to the same inflammatory mediators
and establish whether they share common mechanisms of GR
action.
234 ARAMBASIC ET AL.

ACKNOWLEDGEMENTS
This work was supported by the Ministry of Science and Tech-
nological Development of the Republic of Serbia, Grant
No. 143002.
REFERENCES1. Mackiewicz, A., Kushner, I., and Baumann, H. (1993) Acute Phase Pro-
teins: Molecular Biology, Biochemistry and Clinical Applications. CRCPress, Boca Raton, FL.
2. Baumann, H. and Gauldie, J. (1994) The acute-phase response. Immu-
nol. Today 15, 74–80.
3. Tseng, C. F., Lin, C. C., Huang, H. Y., Liu, H. C., and Mao, S. J.
(2004) Antioxidant role of human haptoglobin. Proteomics 4, 2221–
2228.
4. Huntoon, K. M., Wang, Y., Eppolito, C. A., Barbour, K. W., Berger, F.
G., Shrikant, P. A., and Baumann, H. (2008) The acute phase protein
haptoglobin regulates host immunity. J. Leukoc. Biol. 84, 170–181.
5. Wasell, J. (2000) Haptoglobin: function and polymorphism. Clin. Lab.
16, 547–552.
6. Marinkovic, S. and Baumann, H. (1990) Structure, hormonal regulation,
and identification of the interleukin-6- and dexamethasone-responsive
element of the rat haptoglobin gene. Mol. Cell. Biol. 10, 1573–1583.
7. Gabay, C. and Kushner, I. (1999) Acute-phase proteins and other sys-
temic responses to inflammation. N. Engl. J. Med. 340, 448–454.
8. Xia, L., Xiao, T., Chen, H., Li, P., Wang, Ya., and Wang, H. (2008)
Regulation of haptoglobin expression in a human keratinocyte cell line
HaCaT by inflammatory cytokines and dexamethasone. Chin. Med. J.
121, 730–734.
9. Baumann, H., Prowse, K. R., Marinkovic, S., Won, K.-A., and Jahreis,
G. P. (1989) Stimulation of hepatic acute phase response by cytokines
and glucocorticoids. Ann. NY Acad. Sci. 557, 280–296.
10. Sevaljevic, Lj., Macvanin, M., Zakula, Z., Kanazir, T. D., and Ribarac-
Stepic, N. (1998) Adrenalectomy and dexamethasone treatment alter the
patterns of basal and acute phase response-induced expression of acute
phase protein genes in rat liver. J. Steroid Biochem. Mol. Biol. 66, 347–353.
11. Baumann, H., Jahreis, G. P., and Morella, K. K. (1990) Interaction of
cytokine- and glucocorticoid-response elements of acute-phase plasma
protein genes. J. Biol. Chem. 265, 20390–20399.
12. Fey, G. and Gauldie, J. (1990) The acute phase of liver in inflama-
tion. In Progress in Liver Disease, (Popper, H. and Schaffner, F.,
eds.). pp. 89–116, WB Saunders Co, Philadelphia.
13. Sevaljevic, Lj., Marinkovic-Pajovic, S., Grigorov, I., Bogojevic, D., Iva-
novic-Matic, S., and Petrovic, M. (1995) Acute-phase-dependent
changes in the binding of rat liver nucleoproteins to the cytokine
response element of the rat haptoglobin gene. Int. J. Biochem. Cell Biol.
27, 185–194.
14. Grigorov, I., Milosavljevic, T., Cvetkovic, I., and Petrovic, M. (1998)
Participation of two isoforms of C/EBPb transcription factor in the
acute-phase regulation of the rat haptoglobin gene. Cell Biol. Int. 22,
685–693.
15. Milosavljevic, T., Lazic, T., Uskokovic, A., Petrovic, M., and Grigorov,
I. (2003) Expression of the rat liver haptoglobin gene is mediated bz
isoforms of C/EBPa, -b and -d proteins. Gen. Physiol. Biophys. 22,
181–190.
16. Grigorov, I., Lazic, T., Cvetkovic, I., Milosavljevic, T., and Petrovic,
M. (2000) STAT3 involvement in the acute phase-related expression of
the rat haptoglobin gene. Mol. Biol. Rep. 27, 81–86.
17. Grigorov, I., Lazic, T., Cvetkovic, I., Milosavljevic, T., and Petrovic,
M. (2002) Opposite nuclear level and binding activity of STAT5b and
STAT3 proteins with rat haptoglobin gene under normal and turpentine
induced acute phase conditions. Mol. Biol. Rep. 28, 217–222.
18. Lu, Z. N. and Cidlowski, A. J. (2006) Glucocorticoid receptor isoforms
generate transcription specificity. Trends Cell Biol. 16, 301–307.
19. Beato, M., Herrlich, P., and Schutz, G. (1995) Steroid hormone recep-
tors: many actors in search of a plot. Cell 83, 851–857.
20. Scheidereict, S., Geisse, H., Westpha, M., and Beato, M. (1983) The
glucocorticoid receptor binds to defined nucleotide sequences near the
promoter of mouse mammary tumor virus. Nature 304, 749–752.
21. Truss, M. and Beato, M. (1993) Steroid hormone receptors: interaction
with deoxyribonucleic acid and transcription factors. Endocr. Rev. 14,
459–479.
22. Bastian, L. S. and Nordeen, S. K. (1991) Concerted stimulation of tran-
scription by glucocorticoid receptors and basal transcription factors: lim-
ited transcriptional synergism suggests mediation by coactivators/adap-
tors. Mol. Endocrinol. 5, 619–627.
23. Stocklin, E., Wissler, M., Gouflleux, F., and Gronet, B. (1996) Func-
tional interactions between Stat5 and glucocorticoid receptor. Nature
383, 726–728.
24. Herrlich, P. (2001) Cross-talk between glucocorticoid receptor and
AP-1. Oncogene 20, 2465–2475.
25. Pfahl, M. (1993) Nuclear receptor/AP-1 interaction. Endocr. Rev. 14,
651–658.
26. Ray, A. and Prefontaine, K. E. (1994) Physical association and func-
tional antagonism between the p65 subunit of transcription factor NF-
jB and glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 91, 752–
756.
27. Zhang, Y., Jones, S., Hagood, J. S., Fuentes, N. L., and Fuller, G. M.
(1997) STAT3 acts as a co-activator of glucocorticoid receptor signal-
ing. J. Biol. Chem. 272, 30607–30610.
28. Doppler, W., Windegger, M., Soratroi, C., Tomasi, J., Lechner, J., Rus-
coni, S., Cato, A. C., Almlof, T., Liden, J., Okret, S., Gustafsson, J. A.,
Richard-Foy, H., Starr, D. B., Klocker, H., Edwards, D., and Geymayer,
S. (2001) Expression level dependent contribution of glucocorticoid re-
ceptor domains for functional interaction with STAT5. Mol. Cell. Biol.21, 3266–3279.
29. Savoldi, G., Fenaroli, A., Ferrari, F., Rigaud, G., Albertini, A., and Di
Lorenzo, D. (1997) The glucocorticoid receptor regulates the binding of
C/EBPb on the alpha-1-acid glycoprotein promoter in vivo. DNA Cell
Biol. 16, 1467–1476.
30. Rudiger, J. J., Roth, M., Bihl, P. M., Cornelius, C. B., Johnosn, M., Zie-
sche, R., and Block, L. (2002) Interaction of C/EBPa and the glucocor-
ticoid receptor in vivo and in nontransformed human cells. FASEB J.16, 177–184.
31. Tronche, F., Opherk, C., Moriggl, R., Kellendonk, C., Reimann, A.,
Schwake, L., Reichardt, H. M., Stangl, K., Gau, D., Hoeflich, A., Beug,
H., Schmid, W., and Schutz, G. (2004) Glucocorticoid receptor function
in hepatocytes is essential to promote postnatal body growth. GenesDev. 18, 492–497.
32. Denham, W. and Norman, J. (1998) Inflammatory mediators in sterile
models of inflammation: early evaluation of new mediator-directed
therapies. Sepsis 2, 221–226.
33. Baumann, H. (1988) Electrophoretic analysis of acute-phase proteins.
Methods Enzymol. 163, 566–594.
34. Hodges, J. R. and Sadow, J. (1967). Impairment of pituitary adrenocor-
ticotrophic function by corticosterone in the blood. Br. J. Pharmacol.
Chemother. 30, 385–391.
35. Vannice, J. L., Taylor, J. M., and Ringold, G. M. (1984) Glucocorti-
coid-mediated induction of alpha-1-acid glycoprotein: Evidence for hor-
mone-regulated RNK processing. Proc. Natl. Acad. Sci. USA 81, 4241–
4245.
36. Choi, K. H. and Waxman, J. D. (1999) Growth hormone, but not pro-
lactin, maintains low-level activation of STAT5a and STAT5b in female
rat liver. Endocrinology 140, 5126–5135.
37. Gorski, K., Carneiro, M., and Schibler, U. (1986) Tissue-specific in
vitro transcription from the mouse albumin promoter. Cell 47, 767–776.
235ASSOCIATION OF GR WITH STAT3, C/EBP�, AND HRE

38. Kadonaga, J. T. and Tijan, R. (1986) Affinity purification of sequence-spe-
cific DNA binding proteins. Proc. Natl. Acad. Sci. USA 83, 5889–5893.
39. Towbin, H., Staehelin, T., and Gordon, J. (1979) Electrophoretic trans-
fer of proteins from polyacrilamide gels to nitrocellulose sheets: proce-
dure and some applications. Proc. Natl. Acad. Sci. USA 76, 4350–4354.
40. Lowry, O. H., Rosebrough, W. J., Farr, A. L., and Randall, R. J. (1951)
Protein measurements with the Folin phenol reagent. J. Biol. Chem.
193, 429–432.
41. Kalmovarin, N., Friedrichs, W., O’Brien, H., Linehan, I., Bowman, B.,
and Yang, F. (1991) Extrahepatic expression of plasma protein genes
during inflammation. Inflammation 15, 369–379.
42. Buckingham, C. J. (2006) Glucocorticoids: exemplars of multi-tasking.
Br. J. Pharmacol. 147, 258–268.
43. Spiga, F. and Lightman, L. S. (2009) Dose-dependent effects of corti-
costerone on nuclear glucocorticoid receptors and their binding to DNA
in the brain and pituitary of the rat. Brain Res. 1293, 101–107.
44. Turnbull, A. V., LEE, S., and Rivier, C. (1994) Mechanisms of activa-
tion of the pituitary-adrenal axis by tissue injury in the rat. Psychoneur-
oendocrinology 19, 165–178.
45. Turnbull, A. V. and Rivier, C. (1996) Corticotropin-releasing factor, va-
sopressin, and prostaglandins mediate, and nitric oxide restrains, the
hypothalamic-pituitary-adrenal response to acute local inflammation in
the rat. Endocrinology 137, 455–463.
46. Venihaki, M., Dikkes, P., Carrigan, A., and Karalis, P. K. (2001) Corti-
cotropin-releasing hormone regulates IL-6 expression during inflamma-
tion. J. Clin. Invest. 108, 1159–1166.47. Karten, Y. J. G., Nair, S. M., van Essen, L., Siburg, R., and Joels, M.
(1999) Long-term exposure to high corticosterone levels attenuates sero-
tonin responses in rat hippocampal CA1 neurons. PNAS 96, 13456–
13461.
48. Heinrich, C. P., Behrmann, I., Haan, S., Hermanns, M. H., Muller-
Newen, G., and Schaper, F. (2003) Review article Principles of interleu-
kin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 374,1–20.
49. Lerner, L., Henriksen, M. A., Zhang, X., and Darnell, J. E. Jr. (2003)
STAT3-dependent enhanceosome assembly and disassembly: synergy
with GR for full transcriptional increase of the alpha 2-macroglobulin
gene. Genes Dev. 17, 2564–2577.
50. Merkulov, M. V. and Merkulova, I. T. (2009) Structural variants of glu-
cocorticoid receptor binding sites and different versions of positive glu-
cocorticoid responsive elements: Analysis of GR-TRRD database.
J. Steroid. Biochem. Mol. Biol. 115, 1–8.
51. Takeda, T., Kurachi, H., Yamamoto, T., Nishio, Y., Nakatsuji, Y., Mor-
ishige, K., Miyake, A., and Murata, Y. (1998) Crosstalk between the
interleukin-6 (IL-6)-JAK-STAT and the glucocorticoid-nuclear receptor
pathway: synergistic activation of IL-6 response element by IL-6 and
glucocorticoid. J. Endocrinol. 159, 323–330.
52. Duan, H. O. and Simpson-Haidaris, P. J. (2006) Cell type-specific dif-
ferential induction of the human {gamma}-fibrinogen promoter by inter-
leukin-6. J. Biol. Chem. 281, 12451–12457.
53. Nisho, Y., Isshiki, H., Kishimoto, T., and Akira, S. (1993) A nuclear
factor for IL-6 expression (NF-IL-6) and the glucocorticoid receptor
synergistically activate transcription of the rat alpha-1-acid glycoprotein
gene via direct protein-protein interaction. Mol. Cell. Biol. 13, 1854–
1862.
236 ARAMBASIC ET AL.
Copyright © 2022 FDOKUMEN




















