
Assessment by c-Fos Immunostaining of Changes in Brain Neural Activity Induced by...
9
C Basic & Clinical Pharmacology & Toxicology 2006, 98, 363–371. Printed in Denmark . All rights reserved Copyright C ISSN 1742-7835 Assessment by c-Fos Immunostaining of Changes in Brain Neural Activity Induced by 2,3,7,8-Tetrachlorodibenzo-p- Dioxin (TCDD) and Leptin in Rats* Sanna Lensu 1,2 , Riitta Miettinen 3,4 , Raimo Pohjanvirta 5,6 , Jere Linde ´n 6 and Jouko Tuomisto 1 1 National Public Health Institute, Department of Environmental Health, P.O.B. 95, FI-70701 Kuopio, 2 Department of Pharmacology and Toxicology, University of Kuopio, P.O.B. 1611, FI-70211 Kuopio, 3 Department of Neuroscience and Neurology, University of Kuopio, P.O.B. 1611, FI-70211 Kuopio, 4 University Hospital of Kuopio, P.O.B. 1777, FI-70211 Kuopio, 5 National Veterinary and Food Research Institute, Kuopio Department, P.O.B. 92, FI-70701 Kuopio, 6 Department of Food and Environmental Hygiene, Faculty of Veterinary Medicine, University of Helsinki, P.O.B. 66, FI-00014 University of Helsinki, Finland (Received July 9, 2005; Accepted October 11, 2005) Abstract: The environmental toxicant 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) causes multiple effects in laboratory animals. One of these is a wasting syndrome (a dramatic loss of body weight over 2–5 weeks) whose mechanism is still largely unknown. We exploited the over 1000 times difference in TCDD sensitivity between Long-Evans (Turku/AB); (L- E) and Han/Wistar (Kuopio); (H/W) rats to reveal brain areas that might be activated by a single dose of TCDD (50 mg/kg) given 24 hr previously. Leptin (1.3 mg/kg intraperitoneally 2 hr before tissue harvest) was used as a reference compound, as its neural pathway for decreasing food intake in the control of energy homeostasis is fairly well known. Serial sections of the brains were immunostained with an antibody for the activity marker c-Fos, and selected areas – primarily in the hypothalamus – were analysed with a computer-assisted microscope. Given alone, TCDD did not elicit any major alter- ations in c-Fos protein levels in the hypothalamic nuclei at the early time-point studied (24 hr after administration), neither in pooled data nor in individual strains. The control substance leptin proved that the method is valid as it increased the number of c-Fos-immunopositive cells in the hypothalamic ventromedial andarcuate nuclei. Although the present findings are not suggestive of a primary role for the hypothalamus in the wasting syndrome, a time-course study covering also the feeding-active dark hours is warranted for their verification. Chlorinated dibenzo-p-dioxins and dibenzofurans are en- vironmental pollutants, which are commonly called ‘‘diox- ins’’. They are highly persistent in the environment and poorly soluble in water showing a tendency to accumulate in adipose tissue and food chain (Van den Berg et al. 1994; Geyer et al. 2002). Among dioxins, 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) is the most potent congener. Its LD50 for guinea pig is only about 1 mg/kg whereas that for rats varies de- pending on strain and gender from ∂10 to over 9600 mg/kg (Schwetz et al. 1973; Pohjanvirta et al. 1993; Unkila et al. 1994). TCDD and related xenobiotics exert most of their effects via binding to the cytosolic aryl hydrocarbon recep- tor (AHR). Upon ligand-triggered transformation the AHR translocates into the nucleus, heterodimerizes with a related protein (AHR nuclear translocator, ARNT) and causes re-adjustments in transcriptional activities of multiple genes with the best-known effect being the induc- tion of CYP1A1 (Whitlock 1990; Okey et al. 1994). TCDD causes a wide variety of effects in laboratory ani- Author for correspondence: Sanna Lensu, National Public Health Institute, Laboratory of Toxicology, P.O.B. 95, FI-70701 Kuopio, Finland (fax π358 17 201 265, e-mail sanna.lensu/ktl.fi). * Part of this study was presented at the 10th International Con- gress of Toxicology, held in Tampere, Finland July 11–15, 2004. mals (for reviews see (Pohjanvirta & Tuomisto 1994; Birnbaum & Tuomisto 2000; Dragan & Schrenk 2000)). One of the striking effects after TCDD exposure in labora- tory animals is a wasting syndrome: food intake is severely suppressed and body weight gain is reversed. A lethal dose results in a body weight loss of up to 60% before death ensues (Pohjanvirta & Tuomisto 1994). Several studies sup- port the notion that altered central regulation of energy homeostasis and body weight underlies the syndrome (See- feld et al. 1984; Pohjanvirta & Tuomisto 1990; Pohjanvirta et al. 1990a & 1991). Wasting is not due to gross malabsorp- tion or diarrhoea, nor does it seem to be attributable to nausea (Pohjanvirta et al. 1994), but the animals appear to pursue a lower body weight level by adjusting their food intake accordingly (Seefeld et al. 1984; Pohjanvirta & Tuo- misto 1990). Even after a high sublethal dose, treated ani- mals retain and defend a lowered body weight level (Seefeld et al. 1984; Tuomisto et al. 1995 & 1999a). The conspicuous change in food intake behaviour is of specific interest, as other neurobehavioural effects (e.g. anxiety, nociception or motor coordination) are rather in- significant in adult rats (Sirkka et al. 1992). Similarly, the neural effects – such as changes in spatial or visual learn- ing – observed so far in the progeny of TCDD-treated ani- mals are minor (Seo et al. 2000; Widholm et al. 2003). To understand the mechanism of TCDD-elicited anor-
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Assessment by c-Fos Immunostaining of Changes in Brain Neural Activity Induced by...

C Basic & Clinical Pharmacology & Toxicology 2006, 98, 363–371.Printed in Denmark . All rights reserved
Copyright C
ISSN 1742-7835
Assessment by c-Fos Immunostaining of Changes in BrainNeural Activity Induced by 2,3,7,8-Tetrachlorodibenzo-p-
Dioxin (TCDD) and Leptin in Rats*Sanna Lensu1,2, Riitta Miettinen3,4, Raimo Pohjanvirta5,6, Jere Linden6 and Jouko Tuomisto1
1National Public Health Institute, Department of Environmental Health, P.O.B. 95, FI-70701 Kuopio, 2Department ofPharmacology and Toxicology, University of Kuopio, P.O.B. 1611, FI-70211 Kuopio, 3Department of Neuroscienceand Neurology, University of Kuopio, P.O.B. 1611, FI-70211 Kuopio, 4University Hospital of Kuopio, P.O.B. 1777,
FI-70211 Kuopio, 5National Veterinary and Food Research Institute, Kuopio Department, P.O.B. 92, FI-70701 Kuopio,6Department of Food and Environmental Hygiene, Faculty of Veterinary Medicine, University of Helsinki,
P.O.B. 66, FI-00014 University of Helsinki, Finland
(Received July 9, 2005; Accepted October 11, 2005)
Abstract: The environmental toxicant 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) causes multiple effects in laboratoryanimals. One of these is a wasting syndrome (a dramatic loss of body weight over 2–5 weeks) whose mechanism is stilllargely unknown. We exploited the over 1000 times difference in TCDD sensitivity between Long-Evans (Turku/AB); (L-E) and Han/Wistar (Kuopio); (H/W) rats to reveal brain areas that might be activated by a single dose of TCDD (50 mg/kg)given 24 hr previously. Leptin (1.3 mg/kg intraperitoneally 2 hr before tissue harvest) was used as a reference compound, asits neural pathway for decreasing food intake in the control of energy homeostasis is fairly well known. Serial sections ofthe brains were immunostained with an antibody for the activity marker c-Fos, and selected areas – primarily in thehypothalamus – were analysed with a computer-assisted microscope. Given alone, TCDD did not elicit any major alter-ations in c-Fos protein levels in the hypothalamic nuclei at the early time-point studied (24 hr after administration), neitherin pooled data nor in individual strains. The control substance leptin proved that the method is valid as it increased thenumber of c-Fos-immunopositive cells in the hypothalamic ventromedial and arcuate nuclei. Although the present findingsare not suggestive of a primary role for the hypothalamus in the wasting syndrome, a time-course study covering also thefeeding-active dark hours is warranted for their verification.
Chlorinated dibenzo-p-dioxins and dibenzofurans are en-vironmental pollutants, which are commonly called ‘‘diox-ins’’. They are highly persistent in the environment andpoorly soluble in water showing a tendency to accumulatein adipose tissue and food chain (Van den Berg et al. 1994;Geyer et al. 2002).
Among dioxins, 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD) is the most potent congener. Its LD50 for guineapig is only about 1 mg/kg whereas that for rats varies de-pending on strain and gender from ∂10 to over 9600 mg/kg(Schwetz et al. 1973; Pohjanvirta et al. 1993; Unkila et al.
1994). TCDD and related xenobiotics exert most of theireffects via binding to the cytosolic aryl hydrocarbon recep-tor (AHR). Upon ligand-triggered transformation theAHR translocates into the nucleus, heterodimerizes with arelated protein (AHR nuclear translocator, ARNT) andcauses re-adjustments in transcriptional activities ofmultiple genes with the best-known effect being the induc-tion of CYP1A1 (Whitlock 1990; Okey et al. 1994).
TCDD causes a wide variety of effects in laboratory ani-
Author for correspondence: Sanna Lensu, National Public HealthInstitute, Laboratory of Toxicology, P.O.B. 95, FI-70701 Kuopio,Finland (fax π358 17 201 265, e-mail sanna.lensu/ktl.fi).* Part of this study was presented at the 10th International Con-
gress of Toxicology, held in Tampere, Finland July 11–15, 2004.
mals (for reviews see (Pohjanvirta & Tuomisto 1994;Birnbaum & Tuomisto 2000; Dragan & Schrenk 2000)).One of the striking effects after TCDD exposure in labora-tory animals is a wasting syndrome: food intake is severelysuppressed and body weight gain is reversed. A lethal doseresults in a body weight loss of up to 60% before deathensues (Pohjanvirta & Tuomisto 1994). Several studies sup-port the notion that altered central regulation of energyhomeostasis and body weight underlies the syndrome (See-feld et al. 1984; Pohjanvirta & Tuomisto 1990; Pohjanvirtaet al. 1990a & 1991). Wasting is not due to gross malabsorp-tion or diarrhoea, nor does it seem to be attributable tonausea (Pohjanvirta et al. 1994), but the animals appear topursue a lower body weight level by adjusting their foodintake accordingly (Seefeld et al. 1984; Pohjanvirta & Tuo-misto 1990). Even after a high sublethal dose, treated ani-mals retain and defend a lowered body weight level (Seefeldet al. 1984; Tuomisto et al. 1995 & 1999a).
The conspicuous change in food intake behaviour is ofspecific interest, as other neurobehavioural effects (e.g.anxiety, nociception or motor coordination) are rather in-significant in adult rats (Sirkka et al. 1992). Similarly, theneural effects – such as changes in spatial or visual learn-ing – observed so far in the progeny of TCDD-treated ani-mals are minor (Seo et al. 2000; Widholm et al. 2003).
To understand the mechanism of TCDD-elicited anor-

SANNA LENSU ET AL.364
exia it would be important to know whether specific brainareas are involved. We have shown in simultaneous studiesthat at the same dose, TCDD alters mRNA expression ofcertain genes in the hypothalamus (Korkalainen et al. 2005;Linden et al. 2005). The present study was undertaken todetect the exact localization of brain areas activated byTCDD by using c-Fos immunohistochemistry, whereasmRNA alterations were studied using hypothalamic homo-genates. c-Fos immunostaining is a widely used techniquefor screening neuronal responses to various stimuli (Dragu-now & Faull 1989; Herrera & Robertson 1996; Wyss et al.
1999). A rat model of two rat strains with different sensi-tivities to the toxic outcomes of TCDD was used. At lethaldoses, L-E rats show severely diminished food intake andconsequent body weight loss eventually culminating indeath. Although H/W rats do not exhibit the lethal wastingsyndrome, high doses of TCDD (Ø1000 mg/kg) result in areduced or even permanently halted body weight gain(Pohjanvirta & Tuomisto 1994; Tuomisto et al. 2000). As apositive control substance we used leptin, which is a peptidesecreted by the adipose tissue. It is involved in a number ofphysiological functions, but foremost it is known to regulateenergy balance by decreasing food intake (Schwartz et al.
2000; Baskin et al. 2001). Location of leptin receptors inthe brain has been described (Oldfield et al. 2002), andElmquist et al. (1997 & 1998) have demonstrated by c-Fosimmunostaining the areas in the rat brain which are acti-vated by peripheral leptin treatment.
Materials and Methods
Animal studies. Male rats of both TCDD-sensitive, Long-Evans(Turku/AB); (L-E) rats, (nΩ4–5/group), and TCDD-resistant, Han/Wistar (Kuopio); (H/W) rats, (nΩ4/group), strains were obtainedfrom the breeding colony of the National Public Health Institute,Kuopio, Finland. The LD50 value for L-E rats is from 10 to 20 mg/kg, whereas that for H/W rats is above 9600 mg/kg (Pohjanvirta &Tuomisto 1987; Pohjanvirta et al. 1987; Unkila et al. 1994; Tuomistoet al. 1999b). Rats were 14–15 weeks old at the start of the experi-ments (except for two L-E rats, which were 22 weeks old in theTCDD-treated group) and weighed 297∫5 g and 338∫5 g(mean∫S.E.M.), respectively. Rats were housed in groups of fiveanimals in conventional stainless-steel wire mesh cages, but at leastone week before the start of the experiments they were moved intoindividual stainless-steel wire mesh cages. The rats had standardpelleted food (R36, Ewos, Södertälje, Sweden) and water availablead libitum throughout the study. In the animal room, temperaturewas 21∫1 æ, relative humidity 50∫10% and the lighting rhythm was12 hr light, 12 hr dark with lights on at 7 a.m. The experimentswere reviewed and approved by the Committee for the Welfare ofLaboratory Animals of the University of Kuopio and by the Provin-cial Government. The procedures were conducted in accordancewith the Guidelines of the European Community Council directives86/609/EEC.
The exposure time of TCDD was 24 hr, after which tissue distri-bution is considered to be complete and TCDD has been shown toexist in the brain (Pohjanvirta et al. 1990b; Weber et al. 1993). Theearliest changes in feeding and drinking behaviour can be observedalready on the first day, some of them in a few hours after exposure(Tuomisto et al. 2000; Lensu et al. 2005). As an initial screen of thepossible brain targets of TCDD, and to keep the total number ofanimals moderate but not to decrease group sizes we decided tohave only one time point in this study. Twenty-four hr exposure was
also chosen to catch the acute effects of exposure per se in the brain,with an attempt to avoid confounding primary neural effects withpossible secondary effects arising from e.g. liver toxicity. Responsesto leptin have been shown to appear in the brain already within twohours after doses similar to those now used (Elmquist et al. 1997 &1998; Morimoto et al. 2000).
The rats were treated with either TCDD (UFA-Oil institute, Ufa,Russia; 50 mg/kg, intragastric administration) or the vehicle, cornoil (4 ml/kg, intragastrically) 24 hr before perfusion fixation. Thesame rats were further dosed with either leptin (Calbiochem, LaJol-la, CA, USA; mouse recombinant, 1.3 mg/kg) or saline (0.9% Na-trosterilA, Orion, Espoo, Finland, 1.3 ml/kg) intraperitoneally 22hours after TCDD exposure. The experimental setting is shown infig. 1.
TCDD was dissolved in corn oil as previously described (Pohjan-virta et al. 1987). TCDD was �99% pure as determined by gaschromatography – mass spectrometry. For the sensitive L-E rats thedose of TCDD used is 100% lethal, but lethality due to the TCDDexposure and wasting does not emerge until 2–5 weeks after ex-posure (Pohjanvirta & Tuomisto 1994).
The animal experiments were carried out between 8 a.m.–4 p.m.The rats in different groups were randomised for the exposure time.Twenty-four hr after exposure, the animals were anaesthetised intra-peritoneally with 50 mg/kg sodium-pentobarbital (MebunatA,Orion, Espoo, Finland) and perfused via ascending aorta with 0.9%saline, followed by fresh fixative. The fixative contained 4% para-formaldehyde (P001; TAAB Laboratories Equipment, Aldermas-ton, Berks, UK), 0.01% glutaraldehyde (G002, TAAB LaboratoriesEquipment) and 0.26% picric acid (623; Merck, Darmstadt, Ger-many) in 0.1 M phosphate buffer, pH 7.4. Brains were cryoprotect-ed with a solution containing 30% glycerol (R.P.Normapur, Prola-bo, France), 30% ethylene glycol (Riedel-de Haen, Sigma-AldrichGmbH, Germany) and 40% 0.25 M phosphate buffer, pH 7.4. Fi-nally, brains were stored at ª20 æ.
The brains were cut into 30 mm coronal sections with vibratome(Leica VT1000S, Leica Instruments, Wetzlar, Germany). Cuttingwas started from the level of the optic chiasm and the sections werecollected serially into the 0.1 M phosphate buffer in a 24-well cell-culture plate and washed several times with phosphate buffer. Thensections were cryoprotected with the same solution as before andfrozen (ª20 æ) until used for immunohistochemistry.
Immunostaining. c-Fos-immunostaining was used as an indicator ofactivated cells. Free-floating serial sections (every 8th section, thefirst section randomly selected, through the hypothalamic area:from anterior-posterior level ª0.8 mm to ª3.9 mm from bregma)according to a brain atlas (Paxinos & Watson 1998) were stainedusing immunoperoxidase techniques. Following 6 washes for 30min. with 0.1 M phosphate buffer and 2 washes for 20 min. with0.05 M tris-buffered saline, pH 7.4 (TBS), the sections were treatedwith 10% normal goat serum (NGS; CS-0922, Colorado SerumCompany, Denver, CO, USA) for 40 min. and with 1% NGS for 15min. NGS was dissolved in TBS containing 0.5% Triton X-100(TBST). Polyclonal rabbit primary c-Fos antiserum (1:2000, sc-52,Santa Cruz Inc., USA, Lot. K089) was diluted with 1% NGS andincubation with the primary antibody was carried out at 4 æ for 3days.
The primary polyclonal antiserum for c-Fos was raised againstthe synthetic N-terminal fragment (residues 4–17) of human c-Fos
Fig. 1. Outline of the study design.
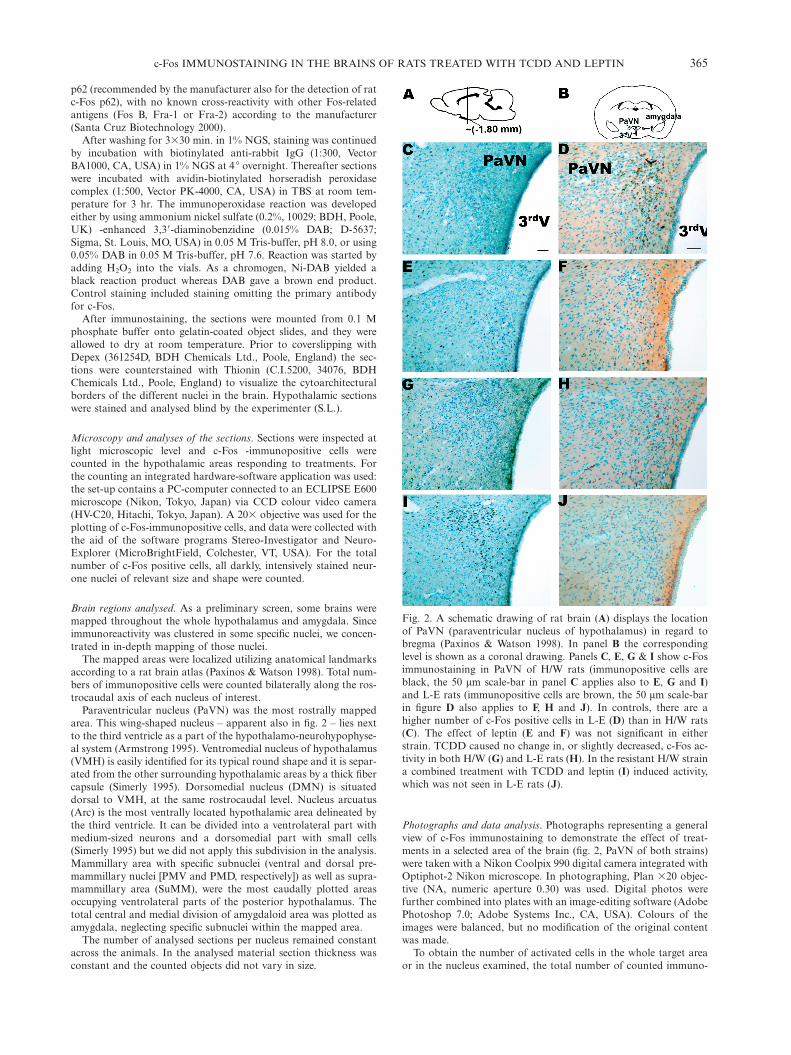
365c-Fos IMMUNOSTAINING IN THE BRAINS OF RATS TREATED WITH TCDD AND LEPTIN
p62 (recommended by the manufacturer also for the detection of ratc-Fos p62), with no known cross-reactivity with other Fos-relatedantigens (Fos B, Fra-1 or Fra-2) according to the manufacturer(Santa Cruz Biotechnology 2000).
After washing for 3¿30 min. in 1% NGS, staining was continuedby incubation with biotinylated anti-rabbit IgG (1:300, VectorBA1000, CA, USA) in 1% NGS at 4 æ overnight. Thereafter sectionswere incubated with avidin-biotinylated horseradish peroxidasecomplex (1:500, Vector PK-4000, CA, USA) in TBS at room tem-perature for 3 hr. The immunoperoxidase reaction was developedeither by using ammonium nickel sulfate (0.2%, 10029; BDH, Poole,UK) -enhanced 3,3ø-diaminobenzidine (0.015% DAB; D-5637;Sigma, St. Louis, MO, USA) in 0.05 M Tris-buffer, pH 8.0, or using0.05% DAB in 0.05 M Tris-buffer, pH 7.6. Reaction was started byadding H2O2 into the vials. As a chromogen, Ni-DAB yielded ablack reaction product whereas DAB gave a brown end product.Control staining included staining omitting the primary antibodyfor c-Fos.
After immunostaining, the sections were mounted from 0.1 Mphosphate buffer onto gelatin-coated object slides, and they wereallowed to dry at room temperature. Prior to coverslipping withDepex (361254D, BDH Chemicals Ltd., Poole, England) the sec-tions were counterstained with Thionin (C.I.5200, 34076, BDHChemicals Ltd., Poole, England) to visualize the cytoarchitecturalborders of the different nuclei in the brain. Hypothalamic sectionswere stained and analysed blind by the experimenter (S.L.).
Microscopy and analyses of the sections. Sections were inspected atlight microscopic level and c-Fos -immunopositive cells werecounted in the hypothalamic areas responding to treatments. Forthe counting an integrated hardware-software application was used:the set-up contains a PC-computer connected to an ECLIPSE E600microscope (Nikon, Tokyo, Japan) via CCD colour video camera(HV-C20, Hitachi, Tokyo, Japan). A 20¿ objective was used for theplotting of c-Fos-immunopositive cells, and data were collected withthe aid of the software programs Stereo-Investigator and Neuro-Explorer (MicroBrightField, Colchester, VT, USA). For the totalnumber of c-Fos positive cells, all darkly, intensively stained neur-one nuclei of relevant size and shape were counted.
Brain regions analysed. As a preliminary screen, some brains weremapped throughout the whole hypothalamus and amygdala. Sinceimmunoreactivity was clustered in some specific nuclei, we concen-trated in in-depth mapping of those nuclei.
The mapped areas were localized utilizing anatomical landmarksaccording to a rat brain atlas (Paxinos & Watson 1998). Total num-bers of immunopositive cells were counted bilaterally along the ros-trocaudal axis of each nucleus of interest.
Paraventricular nucleus (PaVN) was the most rostrally mappedarea. This wing-shaped nucleus – apparent also in fig. 2 – lies nextto the third ventricle as a part of the hypothalamo-neurohypophyse-al system (Armstrong 1995). Ventromedial nucleus of hypothalamus(VMH) is easily identified for its typical round shape and it is separ-ated from the other surrounding hypothalamic areas by a thick fibercapsule (Simerly 1995). Dorsomedial nucleus (DMN) is situateddorsal to VMH, at the same rostrocaudal level. Nucleus arcuatus(Arc) is the most ventrally located hypothalamic area delineated bythe third ventricle. It can be divided into a ventrolateral part withmedium-sized neurons and a dorsomedial part with small cells(Simerly 1995) but we did not apply this subdivision in the analysis.Mammillary area with specific subnuclei (ventral and dorsal pre-mammillary nuclei [PMV and PMD, respectively]) as well as supra-mammillary area (SuMM), were the most caudally plotted areasoccupying ventrolateral parts of the posterior hypothalamus. Thetotal central and medial division of amygdaloid area was plotted asamygdala, neglecting specific subnuclei within the mapped area.
The number of analysed sections per nucleus remained constantacross the animals. In the analysed material section thickness wasconstant and the counted objects did not vary in size.
Fig. 2. A schematic drawing of rat brain (A) displays the locationof PaVN (paraventricular nucleus of hypothalamus) in regard tobregma (Paxinos & Watson 1998). In panel B the correspondinglevel is shown as a coronal drawing. Panels C, E, G & I show c-Fosimmunostaining in PaVN of H/W rats (immunopositive cells areblack, the 50 mm scale-bar in panel C applies also to E, G and I)and L-E rats (immunopositive cells are brown, the 50 mm scale-barin figure D also applies to F, H and J). In controls, there are ahigher number of c-Fos positive cells in L-E (D) than in H/W rats(C). The effect of leptin (E and F) was not significant in eitherstrain. TCDD caused no change in, or slightly decreased, c-Fos ac-tivity in both H/W (G) and L-E rats (H). In the resistant H/W straina combined treatment with TCDD and leptin (I) induced activity,which was not seen in L-E rats (J).
Photographs and data analysis. Photographs representing a generalview of c-Fos immunostaining to demonstrate the effect of treat-ments in a selected area of the brain (fig. 2, PaVN of both strains)were taken with a Nikon Coolpix 990 digital camera integrated withOptiphot-2 Nikon microscope. In photographing, Plan ¿20 objec-tive (NA, numeric aperture 0.30) was used. Digital photos werefurther combined into plates with an image-editing software (AdobePhotoshop 7.0; Adobe Systems Inc., CA, USA). Colours of theimages were balanced, but no modification of the original contentwas made.
To obtain the number of activated cells in the whole target areaor in the nucleus examined, the total number of counted immuno-
SANNA LENSU ET AL.366
positive cells was multiplied by 8 to correct for the sampling fraction(every 8th section was stained). The volume (mm3) of regions wasobtained by multiplying the area of contour (mm2) drawn duringthe plotting around each region by 240 mm (section sampling frac-tion¿section thicknessΩ8¿30 mm). Activated cell density wascounted by dividing the number of c-Fos positive cells with the totalvolume (mm3) of each plotted nucleus.
For statistical analyses, SPSS version 10.0.7 was used. Data wereassessed by three-way analysis of variance (strain, TCDD treatmentand leptin treatment) after verifying homogeneity of variances byLevene’s test. Data of different rat strains were tested by two-wayanalysis of variance. In the case of a statistically significant interac-tive effect with TCDD and leptin, a full-factorial model with con-trast coefficient was calculated to assign the statistical differencesamong the groups. If the interaction was not statistically significant,data were analysed by two-way analysis of variance followed bypost-hoc testing (Dunnett’s 2-sided t test and Dunnett’s T3 test).The level of a statistically significant difference was set at P-valueof 0.05.
Results
In the majority of positively stained cells, c-Fos-immuno-reactivity (c-Fos-ir) was present in the nuclei of individualcells. c-Fos-ir was detectable as a dense, darkly stained,either black (Ni-DAB as a chromogen) or brown (DAB asa chromogen) aggregate. The immunoreaction product andits counterstain (Nissl) could readily be discriminated (fig.2) in both cases.
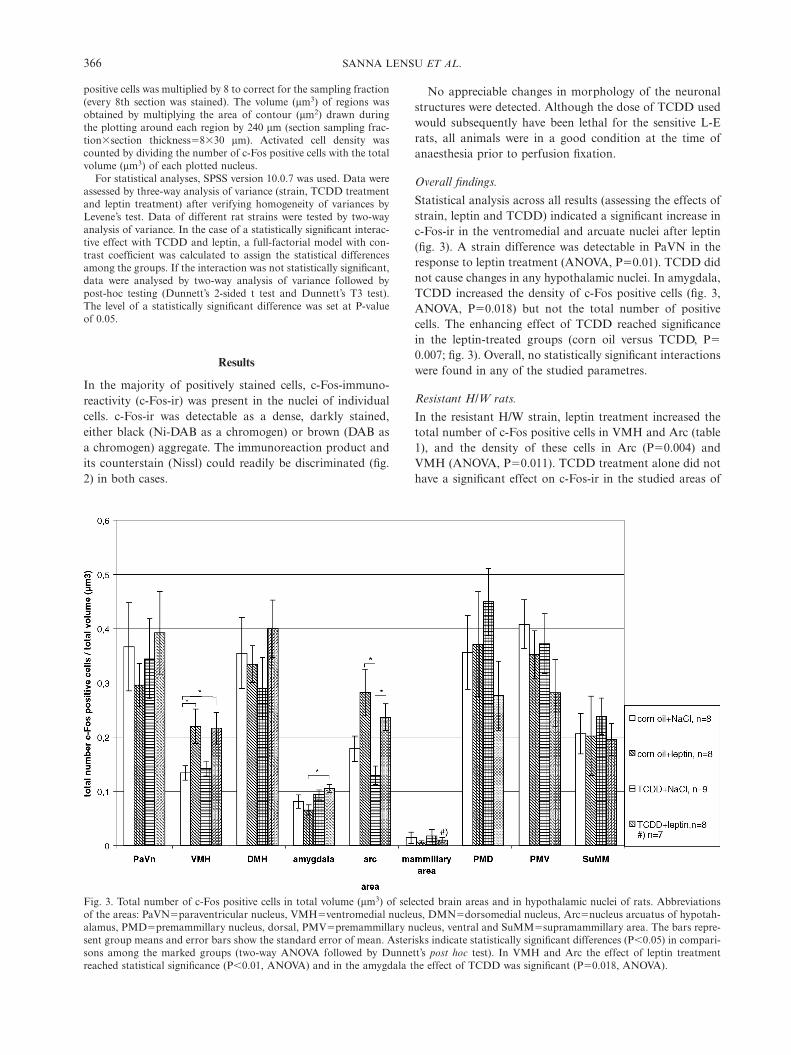
Fig. 3. Total number of c-Fos positive cells in total volume (mm3) of selected brain areas and in hypothalamic nuclei of rats. Abbreviationsof the areas: PaVNΩparaventricular nucleus, VMHΩventromedial nucleus, DMNΩdorsomedial nucleus, ArcΩnucleus arcuatus of hypotah-alamus, PMDΩpremammillary nucleus, dorsal, PMVΩpremammillary nucleus, ventral and SuMMΩsupramammillary area. The bars repre-sent group means and error bars show the standard error of mean. Asterisks indicate statistically significant differences (P�0.05) in compari-sons among the marked groups (two-way ANOVA followed by Dunnett’s post hoc test). In VMH and Arc the effect of leptin treatmentreached statistical significance (P�0.01, ANOVA) and in the amygdala the effect of TCDD was significant (PΩ0.018, ANOVA).
No appreciable changes in morphology of the neuronalstructures were detected. Although the dose of TCDD usedwould subsequently have been lethal for the sensitive L-Erats, all animals were in a good condition at the time ofanaesthesia prior to perfusion fixation.
Overall findings.
Statistical analysis across all results (assessing the effects ofstrain, leptin and TCDD) indicated a significant increase inc-Fos-ir in the ventromedial and arcuate nuclei after leptin(fig. 3). A strain difference was detectable in PaVN in theresponse to leptin treatment (ANOVA, PΩ0.01). TCDD didnot cause changes in any hypothalamic nuclei. In amygdala,TCDD increased the density of c-Fos positive cells (fig. 3,ANOVA, PΩ0.018) but not the total number of positivecells. The enhancing effect of TCDD reached significancein the leptin-treated groups (corn oil versus TCDD, PΩ0.007; fig. 3). Overall, no statistically significant interactionswere found in any of the studied parametres.
Resistant H/W rats.
In the resistant H/W strain, leptin treatment increased thetotal number of c-Fos positive cells in VMH and Arc (table1), and the density of these cells in Arc (PΩ0.004) andVMH (ANOVA, PΩ0.011). TCDD treatment alone did nothave a significant effect on c-Fos-ir in the studied areas of

367c-Fos IMMUNOSTAINING IN THE BRAINS OF RATS TREATED WITH TCDD AND LEPTIN
Ta
ble
1.
Tota
lnu
mbe
rof
c-F
ospo
siti
vece
llsin
sele
cted
hypo
thal
amic
nucl
eian
dam
ygda
la(b
ilate
rally
,bo
thhe
mis
pher
esin
clud
ed).
Abb
revi
atio
nsof
the
area
s:Pa
VN
Ωpa
rave
ntri
cula
rnu
cleu
s,V
MH
Ωve
ntro
med
ial
nucl
eus,
DM
NΩ
dors
omed
ial
nucl
eus
and
Arc
Ωnu
cleu
sar
cuat
usof
hypo
taha
lam
us;
PM
DΩ
prem
amm
illar
ynu
cleu
s,do
rsal
;P
MV
Ωpr
emam
mill
ary
nucl
eus,
vent
ral
and
SuM
MΩ
supr
amam
mill
ary
nucl
eus.
The
data
are
give
nas
mea
n∫S.
E.M
.C
Eis
the
unbi
ased
esti
mat
orof
coef
ficie
ntof
vari
atio
n(S
.E.M
./mea
n).
The
data
wer
est
atis
tica
llyan
alyz
edby
two-
way
AN
OV
Afo
llow
edby
calc
ulat
ion
ofco
ntra
stco
effic
ient
sfo
rdi
ffer
ent
trea
tmen
ts.S
igni
fican
tef
fect
sar
ein
bold
.Val
ues
appr
oach
ing
the
set
sign
ifica
nce
leve
l(0.
05)
are
show
nin
ita
lics
(see
foot
erof
the
tabl
efo
rex
act
P-v
alue
s).
PaV
NV
MH
DM
NA
myg
dala
Arc
Mam
mill
ary
area
PM
DP
MV
SuM
Mm
ean∫
S.E
.M.
mea
n∫S.
E.M
.m
ean∫
S.E
.M.
mea
n∫S.
E.M
.m
ean∫
S.E
.M.
mea
n∫S.
E.M
.m
ean∫
S.E
.M.
mea
n∫S.
E.M
.m
ean∫
S.E
.M.
Han
-Wis
tar
Cor
noi
lπsa
line
1822
∫52
414
76∫
108b
3232
∫10
1352
46∫
1041
2908
∫51
442
∫34
608∫
57g
346∫
4361
2∫14
9n
Ω4
CE
Ω0
.29
CE
Ω0
.07
CE
Ω0
.31
CE
Ω0
.20
CE
Ω0
.19
CE
Ω0
.81
CE
Ω0
.09
CE
Ω0
.12
CE
Ω0
.24
Cor
noi
lπle
ptin
2398
∫63
222
40∫
310c
2574
∫23
544
88∫
1255
5364
∫13
55f
32∫
1769
6∫20
854
4∫16
860
0∫37
1n
Ω4
CE
Ω0
.26
CE
Ω0
.14
CE
Ω0
.09
CE
Ω0
.28
CE
Ω0
.25
CE
Ω0
.53
CE
Ω0
.30
CE
Ω0
.31
CE
Ω0
.62
TC
DD
πsa
line
1576
∫38
0a17
88∫
151d
1672
∫37
7e55
90∫
1044
2054
∫38
312
2∫74
936∫
44g
348∫
100
804∫
219
nΩ
4C
EΩ
0.2
4C
EΩ
0.0
8C
EΩ
0.2
3C
EΩ
0.1
9C
EΩ
0.1
9C
EΩ
0.6
0C
EΩ
0.0
5C
EΩ
0.2
9C
EΩ
0.2
7
TC
DD
πle
ptin
34
28∫
83
5a
3108
∫54
9b,c
,d42
18∫
824e
7414
∫11
4136
28∫
182f
59∫
17*
640∫
269
342∫
128
540∫
95n
Ω4
CE
Ω0
.24
CE
Ω0
.18
CE
Ω0
.20
CE
Ω0
.15
CE
Ω0
.05
CE
Ω0
.30
CE
Ω0
.42
CE
Ω0
.37
CE
Ω0
.18
Lon
g-E
vans
Cor
noi
lπsa
line
3220
∫87
016
10∫
390
3442
∫12
4464
80∫
2362
2116
∫49
830
∫17
990∫
509
374∫
154
1162
∫44
7n
Ω4
CE
Ω0
.27
CE
Ω0
.24
CE
Ω0
.36
CE
Ω0
.36
CE
Ω0
.24
CE
Ω0
.58
CE
Ω0
.51
CE
Ω0
.41
CE
Ω0
.38
Cor
noi
lπle
ptin
1814
∫24
438
78∫
716h
3582
∫42
656
44∫
1192
41
12∫
11
01
i52
∫23
916∫
390
178∫
985
6∫10
4n
Ω4
CE
Ω0
.13
CE
Ω0
.18
CE
Ω0
.19
CE
Ω0
.21
CE
Ω0
.27
CE
Ω0
.44
CE
Ω0
.43
CE
Ω0
.05
CE
Ω0
.12
TC
DD
πsa
line
2941
∫63
914
66∫
183
3346
∫85
467
42∫
938
1550
∫35
113
∫3
856∫
209
230∫
4899
5∫33
9n
Ω5
CE
Ω0
.22
CE
Ω0
.12
CE
Ω0
.26
CE
Ω0
.14
CE
Ω0
.23
CE
Ω0
.25
CE
Ω0
.24
CE
Ω0
.21
CE
Ω0
.34
TC
DD
πle
ptin
2036
∫54
02
77
2∫
62
8h
3230
∫80
382
04∫
957
34
90∫
70
2i
20∫
754
2∫18
410
8∫40
1044
∫36
3n
Ω4
CE
Ω0
.27
CE
Ω0
.23
CE
Ω0
.25
CE
Ω0
.12
CE
Ω0
.20
CE
Ω0
.35
CE
Ω0
.34
CE
Ω0
.37
CE
Ω0
.35
aC
ontr
ast
betw
een
salin
ean
dle
ptin
inT
CD
D-t
reat
edgr
oups
:P
Ω0.
055.
bD
iffe
renc
ebe
twee
nco
rnoi
lπsa
line
vs.
TC
DD
πle
ptin
:P
Ω0.
011
(Dun
nett
’st-
test
).c
Eff
ect
ofle
ptin
:P
Ω0.
008.
dC
ontr
ast
betw
een
salin
ean
dle
ptin
inT
CD
D-t
reat
edgr
oups
:P
Ω0.
015.
eIn
tera
ctio
nof
lept
inan
dT
CD
Dtr
eatm
ents
:P
Ω0.
039,
cont
rast
betw
een
salin
ean
dle
ptin
trea
tmen
tin
TC
DD
-tre
ated
grou
ps:
PΩ
0.02
3.f
Eff
ect
ofle
ptin
:P
Ω0.
02,
cont
rast
betw
een
salin
ean
dle
ptin
trea
tmen
tin
corn
oil-
trea
ted
grou
ps:
PΩ
0.04
.g
Dif
fere
nce
betw
een
corn
oilπ
NaC
lve
rsus
TC
DD
πN
aCl:
PΩ
0.02
2(D
unne
tt’s
t-te
st).
hE
ffec
tof
lept
in:
PΩ
0.00
3,co
ntra
stbe
twee
nsa
line
and
lept
intr
eatm
ent
inco
rnoi
-tre
ated
grou
ps:
PΩ
0.00
8;in
TC
DD
-tre
ated
grou
ps:
PΩ
0.07
7.i
Eff
ect
ofle
ptin
:P
Ω0.
013,
cont
rast
betw
een
salin
ean
dle
ptin
trea
tmen
tin
corn
oil-
trea
ted
grou
ps:
PΩ
0.06
7;in
TC
DD
-tre
ated
grou
ps:
PΩ
0.06
1.*
nΩ3.

SANNA LENSU ET AL.368
the brain as compared with the control treatment except forthe PMD (table 1). A trend to increased number of c-Fospositive cells was detectable in PaVN (ANOVA, PΩ0.055;table 1).
Combined treatment with TCDD and leptin increasedthe total number of c-Fos positive cells in the dorsomedial,ventromedial and (almost significantly, PΩ0.055) paraven-tricular nuclei of hypothalamus as compared with TCDDtreatment alone (table 1).
Sensitive L-E rats.
As was the case in H/W rats, TCDD exposure alone in L-Edid not have a significant effect on c-Fos-ir in the studiedareas of the brain as compared with the control treatment.Treatment with leptin – with or without TCDD exposure –increased the total number of c-Fos positive cells in VMH(ANOVA, PΩ0.003) and in Arc (ANOVA, PΩ0.013; table 1).In PaVN, leptin treatment had a subtle tendency to decreasethe activation (ANOVA, PΩ0.088; table 1), but the baselineactivity in the control group was high and variation amongthe animals was large. Combined treatment with leptin andTCDD did not cause a similar upward trend in c-Fos-ir towhat was seen in PaVN, VMH and DMN in H/W rats.
Discussion
The control of energy homeostasis requires an appropriatebalancing of food intake with energy expenditure. This con-trol is centrally orchestrated with hypothalamic nuclei play-ing an indispensable role as key regulatory sites (Schwartzet al. 2000 & 2003). TCDD is the most potent congener inthe group of dioxins. It is the most potent anorexigenknown, but the mechanism by which it exerts its drasticeffect on food intake is obscure (Pohjanvirta & Tuomisto1990; Tuomisto et al. 1995; & 2000). Therefore, we set outthe present study to examine whether TCDD might activateor deactivate those hypothalamic nuclei previously estab-lished to be involved in the control of feeding behaviour.As a physiological, positive control substance, we employedleptin that is a major satiety signal in the body and actsthrough the hypothalamus (Schwartz et al. 1996; Hakans-son et al. 1998; Shioda et al. 1998).
c-Fos immunoreactivity is a useful index of neuronal acti-vation. Upon stimulation, the first response of neurons inthe CNS involves immediate early gene activation. One ofthe immediate early genes is c-Fos, which may have a gen-eral role in the cellular signal transmission pathway (Hugh-es & Dragunow 1995). c-Fos is the protein product of the c-
fos gene, and staining of neural tissue with a c-Fos antibodyenables mapping of functional pathways (Sagar et al. 1988;Smeyne et al. 1993; Curran & Morgan 1995; Herdegen et
al. 1995), as the c-Fos protein is detectable after stimuli al-ready in 30 min. Maximal levels are not reached until 2 to3 hr later (Dragunow & Faull 1989; Herdegen et al. 1995).Following TCDD exposure, elevated c-Fos levels in thebrain have been found even several weeks afterwards(Cheng et al. 2002).
Induction of c-Fos can be detected after cell membranedepolarization. An increase in intracellular calcium follow-ing opening of voltage-sensitive calcium channels regulatesthe expression of immediate early genes (Morgan & Curran1986 & 1988; Sagar et al. 1988). Induction of c-Fos mayalso follow from activation of protein kinase C (Stumpo &Blackshear 1986). Based on data from cell culture studies,TCDD has been suggested to induce expression of immedi-ate early genes by activation of Ah receptor complex-de-pendent and -independent signal transduction pathways(Hoffer et al. 1996). In vivo, increased c-Fos-ir was detectedin guinea pig liver 1 day after exposure to 1 mg/kg TCDD(Ashida & Matsumura 1998), but brain c-Fos-ir levels atthat time point have not been reported previously. Anyhow,it is known that induction of immediate early genes is notspecifically stimulus-dependent and there is a certain levelof basal neuronal c-Fos-ir in naive rats (Sagar et al. 1988;Smeyne et al. 1993; Curran & Morgan 1995; Herdegen et
al. 1995). The c-Fos-ir levels of control animals found in thepresent study are well in line with those in previous reports.
In the present study, leptin-induced elevations of c-Fosimmunoreactivity were found in VMH and Arc (ANOVA,P�0.01) implying that these two are the most sensitive hy-pothalamic nuclei to leptin action. Cells in the VMH havebeen shown to increase their activity following leptin treat-ment in electrophysiological studies in vivo and in vitro (Shi-raishi et al. 1999 & 2000; Nagamori et al. 2003). In studieswith Sprague-Dawley rats, increased c-Fos-ir following pe-ripheral leptin treatment (intravenously or intraperito-neally) was detected in VMH and Arc, but also in PaVNand DMN (Elmquist et al. 1997 & 1998; Kalra et al. 1999).
In inter-strain comparisons, some differences betweenTCDD-sensitive L-E and TCDD-resistant H/W ratsemerged in neural activity. Surprisingly, more alterations inc-Fos-ir were recorded in H/W rats than in L-E rats. InDMN, VMH and PaVN of H/W rats, the highest levels ofc-Fos-ir were found in the combined treatment group (leptinπ TCDD). This finding may suggest that TCDD exposurecan lower the signalling threshold for the cellular responseto leptin, such that a preceding TCDD treatment may po-tentiate the effect of exogenous leptin in these nuclei. Analternative explanation is that the increase would resultfrom activation of distinct neuronal populations by leptinand TCDD. However, TCDD on its own tended to diminishc-Fos-ir in some hypothalamic nuclei, suggesting thatTCDD, acting alone, might have a preferably inhibitory ef-fect on certain neurones in H/W rats. Observations of tran-scriptionally repressive actions of TCDD have recently beenmade in in vitro studies with renal proximal tubule cells(Han et al. 2005).
In contrast to the situation in H/W rats, in the hypothala-mus of L-E rats c-Fos expression was not induced by a com-bined treatment with TCDD and leptin. Although the bio-logical basis and toxicological significance of these unex-pected findings is hard to envisage at present, it should beborne in mind that these experiments were carried out atthe time of day (during light hours) when rats consume only

369c-Fos IMMUNOSTAINING IN THE BRAINS OF RATS TREATED WITH TCDD AND LEPTIN
a minor proportion (some 10%) of their daily food intake.The rats were thus in a state of satiety at the time of brainsample harvesting. Should TCDD mainly act by dimin-ishing hunger sensation, no major alterations in neural ac-tivity would be expected to be revealed in the present con-ditions. Verification of this possibility requires a similaranalysis to be conducted during dark hours.
Regarding basal c-Fos-ir levels, the greatest differencewas seen in PaVN, where an almost 2 times higher consti-tutive c-Fos expression was recorded in L-E than in H/Wrats. One possible reason for this could be a more pro-nounced stress reaction by L-E rats to intragastric gavagestimulating CRH expression and leading to elevated c-Foslevels. However, the rats were adapted to handling prior tothe experiment to minimize the stress. Interestingly, a recentstudy discovered about 2 times higher basal mRNA levelsof e.g. AhR, ARNT and ARNT2 in the hypothalamus ofL-E rats as compared with H/W rats (Korkalainen et al.
2005). Since in that study hypothalamic blocks were usedas starting material, it is not known from which nuclei thesevarious mRNA levels originated; nor can one infer whetherthe differences in mRNA expression levels would also reflectin c-Fos immunoreactivity. However, it is evident that theserat strains have important physiological differences.
Cheng et al. (2002) showed that the same dose of TCDD(50 mg/kg by gavage) to adult Long-Evans rats elevated thenumber of c-Fos-immunopositive neurones in various nucleiof hypothalamus and amygdala, but their data originatedfrom 3–4 days after exposure. The authors did not reportearlier effects, despite the fact that in their study, samplingwas started 12 hr after exposure. In central nucleus ofamygdala and in bed nucleus of stria terminalis inductionof c-Fos protein was still detectable 1–5 weeks after ex-posure. However, those Long-Evans rats were outbred andtheir LD50 was not defined. In the present study with in-bred, extremely TCDD-sensitive L-E rats we did not see anycomparable increase at 24 hr after TCDD exposure, but wedid not analyse the subnuclei of amygdala distinctly in thisstudy. When we included both strains in the 3-way ANOVAanalysis of the density of c-Fos positive cells, a TCDD-in-duced increase in amygdala immunoreactivity was found.Thus, TCDD may affect central regulatory pathways alsooutside of the hypothalamus. It should be borne in mindthat there are important extrahypothalamic feeding centersthat were not included in the present study such as the soli-tary nucleus in the hindbrain.
The molecular mechanisms of the effects of TCDD in thebrain are unknown, but in order to set up a study for speci-fic molecular mechanisms in vitro, changes in the livingbrain need to be screened in vivo. By using quantitative RT-PCR we have shown that TCDD elevates mRNA levels ofthe AHR repressor, CYP1A1 and CYP1A2 genes (Korka-lainen et al. 2005). We have also studied the mRNA levelsof certain neuropeptides, proteins and their receptors ex-pressed in the rat brain, and found that TCDD affectedmainly expression of orexigenic factors: after an initial sup-pression there was reversal to enhanced expression by 96 hr
after the exposure (Linden et al. 2005). Though thesestudies were undertaken by using hypothalamic blocks, andsome cell-specific responses may have been masked in thehomogenated tissue, they support the hypothesis of a neuralpathway behind the wasting syndrome. Fetissov et al. (2004)have studied by in situ hybridisation the mRNA expressionof some hypothalamic neuropeptides. They found increasedlevels of mRNA expression of orexigenic or anorexigenicpeptides in the Spraque-Dawley rat brain 6 days after asingle dose of TCDD (15 mg/kg), but they did not includeany earlier data of immediate changes after the exposure.
So far, no individual hypothalamic nucleus, other regionin the CNS or neurotransmitter (either peptide or mono-amine) has stood out as a prominent candidate for the pri-mary key target of TCDD. As outlined above, it is conceiv-able that this is due to brain sample harvesting, which inall cases has taken place at light hours and thus missed theperiod of day most relevant for feeding in rats. Although itis equally possible that the primary impact of TCDD oc-curs in a peripheral tissue or organ such as the liver, thefinal effect leading to altered feeding should be also detect-able in the CNS.
In conclusion, in the studied brain areas TCDD (50 mg/kg) did not have any detectable effect on c-Fos protein ex-pression at 24 hr after exposure in TCDD-sensitive L-Erats, but interacted with leptin in PaVN and DMN inTCDD-resistant H/W rats. In both strains, leptin elevatedc-Fos-ir in VMH and in Arc testifying for the high sensi-tivity of these nuclei to leptin. To verify whether TCDDexerts a delayed effect on c-Fos-ir, a detailed time-coursestudy will be necessary. Furthermore, TCDD’s possible im-pact on neural activity should also be analysed at darkhours when the rats are hungry.
AcknowledgementsThe authors want to thank Mrs. Virpi Tiihonen for her
excellent technical help in the animal experiment and Mrs.Anna-Lisa Gidlund among other personnel at the labora-tory for advice on histology. M.Sc. Pekka Tiittanen is ac-knowledged for providing statistical advice. The authorsalso want to express their sincere thanks to Doc. Thomasvan Groen for valuable comments and advice.
The work was supported by grants from the Centre ofExcellence Program of the Academy of Finland to theCentre of Environmental Health Risk Analysis (Grant no.53307) and from the Academy of Finland (Grants no.200980 and no. 211120 [R.P.]). Financial support [S.L.]from the Graduate School in Environmental Health (Minis-try of Education) is acknowledged.
References
Armstrong, W. E.: Hypothalamic supraoptic and paraventricularnuclei. In: The rat nervous system. Ed.: G. Paxinos. AcademicPress, San Diego, California, 1995, pp. 377–390.
Ashida, H. & F. Matsumura: Effect of in vivo administered 2,3,7,8-tetrachlorodibenzo-p-dioxin on DNA-binding activities of nu-

SANNA LENSU ET AL.370
clear transcription factors in liver of guinea pigs. J. Biochem.
Mol. Toxicol. 1998, 12, 191–204.Baskin, D. G., J. E. Blevins & M. W. Schwartz: How the brain
regulates food intake and body weight: the role of leptin. J. Phar-
macol. Exp. Therap 2001, 14, 1417–1429.Birnbaum, L. S. & J. Tuomisto: Non-carcinogenic effects of TCDD
in animals. Food Addit. Cont. 2000, 17, 275–288.Cheng, S. B., S. Kuchiiwa, I. Nagatomo, Y. Akasaki, M. Uchida,
M. Tominaga, W. Hashiguchi, T. Kuchiiwa & S. Nakagawa:2,3,7,8-Tetrachlorodibenzo-p-dioxin treatment induces c-Fos ex-pression in the forebrain of the Long-Evans rat. Brain Res. 2002,931, 176–180.
Curran, T. & J. I. Morgan: Fos: an immediate-early transcriptionfactor in neurons. J. Neurobiol. 1995, 26, 403–412.
Dragan, Y. P. & D. Schrenk: Animal studies addressing the carcino-genicity of TCDD (or related compounds) with an emphasis ontumour promotion. Food Addit. Cont. 2000, 17, 289–302.
Dragunow, M. & R. Faull: The use of c-fos as a metabolic markerin neuronal pathway tracing. J. Neurosci. Meth. 1989, 29, 261–265.
Elmquist, J. K., R. S. Ahima, C. F. Elias, J. S. Flier & C. B. Saper:Leptin activates distinct projections from the dorsomedial andventromedial hypothalamic nuclei. Proc. Natl. Acad. Sci. USA
1998, 95, 741–746.Elmquist, J. K., R. S. Ahima, E. Maratos-Flier, J. S. Flier & C. B.
Saper: Leptin activates neurons in ventrobasal hypothalamus andbrainstem. Endocrinology 1997, 138, 839–842.
Fetissov, S. O., P. Huang, Q. Zhang, J. Mimura, Y. Fujii-Kuriyama,A. Rannug, T. Hokfelt & S. Ceccatelli: Expression of hypothal-amic neuropeptides after acute TCDD treatment and distributionof Ah receptor repressor. Regulatory Peptides 2004, 119, 113–124.
Geyer, H. J., K. W. Schramm, E. A. Feicht, A. Behechti, C.Steinberg, R. Bruggemann, H. Poiger, B. Henkelmann & A.Kettrup: Half-lives of tetra-, penta-, hexa-, hepta-, and octachlor-odibenzo-p-dioxin in rats, monkeys, and humans – a critical re-view. Chemosphere 2002, 48, 631–644.
Hakansson, M. L., H. Brown, N. Ghilardi, R. C. Skoda & B. Meis-ter: Leptin receptor immunoreactivity in chemically defined tar-get neurons of the hypothalamus. J. Neurosci. 1998, 18, 559–572.
Han, H. J., M. J. Lim, Y. J. Lee, E. J. Kim, Y. J. Jeon & J. H. Lee:Effects of TCDD and estradiol-17beta on the proliferation andNa(π)/glucose cotransporter in renal proximal tubule cells. Toxi-
cology in vitro 2005, 19, 21–30.Herdegen, T., K. Kovary, A. Buhl, R. Bravo, M. Zimmermann & P.
Gass: Basal expression of the inducible transcription factors c-Jun, JunB, JunD, c-Fos, FosB, and Krox-24 in the adult rat brain.J. Comp. Neurol. 1995, 354, 39–56.
Herrera, D. G. & H. A. Robertson: Activation of c-fos in the brain.Progr. Neurobiology 1996, 50, 83–107.
Hoffer, A., C. Y. Chang & A. Puga: Dioxin induces transcriptionof fos and jun genes by Ah receptor-dependent and -independentpathways. Toxicol. Appl. Pharmacol. 1996, 141, 238–247.
Hughes, P. & M. Dragunow: Induction of immediate-early genesand the control of neurotransmitter-regulated gene expressionwithin the nervous system. Pharmacol. Rev. 1995, 47, 133–178.
Kalra, S. P., M. G. Dube, S. Pu, B. Xu, T. L. Horvath & P. S. Kalra:Interacting appetite-regulating pathways in the hypothalamicregulation of body weight. Endocrine Rev. 1999, 20, 68–100.
Korkalainen, M., J. Linden, J. Tuomisto & R. Pohjanvirta: Effectof TCDD on mRNA expression of genes encoding bHLH/PASproteins in rat hypothalamus. Toxicology 2005, 208, 1–11.
Lensu, S., J. Linden, J. Tuomisto & R. Pohjanvirta: Continuousfollow-up of feeding and drinking patterns in rats after TCDDexposure. Organohalogen Compounds 2005, 67, 2549–2551.
Linden, J., M. Korkalainen, S. Lensu, J. Tuomisto & R. Pohjanvir-ta: Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) andleptin on hypothalamic mRNA expression of factors partici-pating in food intake regulation in a TCDD-sensitive and a
TCDD-resistant rat strain. J. Biochem. Mol. Toxicol. 2005, 19,139–48.
Morgan, J. I. & T. Curran: Role of ion flux in the control of c-fosexpression. Nature 1986, 322, 552–555.
Morgan, J. I. & T. Curran: Calcium as a modulator of the immedi-ate-early gene cascade in neurons. Cell Calcium 1988, 9, 303–311.
Morimoto, T., Y. Yamamoto & A. Yamatodani: Leptin facilitateshistamine release from the hypothalamus in rats. Brain Res. 2000,868, 367–369.
Nagamori, K., M. Ishibashi, T. Shiraishi, Y. Oomura & K. Sasaki:Effects of leptin on hypothalamic arcuate neurons in Wistar andZucker rats: an in vitro study. Exp. Biol. Med. 2003, 228, 1162–1167.
Okey, A. B., D. S. Riddick & P. A. Harper: The Ah receptor: me-diator of the toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD) and related compounds. Toxicol. Lett. 1994, 70, 1–22.
Oldfield, B. J., M. E. Giles, A. Watson, C. Anderson, L. M. Col-vill & M. J. McKinley: The neurochemical characterisation ofhypothalamic pathways projecting polysynaptically to brown adi-pose tissue in the rat. Neuroscience 2002, 110, 515–526.
Paxinos, G. & C. Watson: The rat brain in stereotaxic coordinates,fourth edition. Academic Press, San Diego, California, USA,1998.
Pohjanvirta, R. & J. Tuomisto: Han/Wistar rats are exceptionallyresistant to TCDD. II. Arch. Toxicol. Suppl. 1987, 11, 344–347.
Pohjanvirta, R. & J. Tuomisto: 2,3,7,8-Tetrachlorodibenzo-p-dioxinenhances responsiveness to post-ingestive satiety signals. Toxi-
cology 1990, 63, 285–299.Pohjanvirta, R. & J. Tuomisto: Short-term toxicity of 2,3,7,8-tetra-
chlorodibenzo-p-dioxin in laboratory animals: effects, mechan-isms, and animal models. Pharmacol. Rev. 1994, 46, 483–549.
Pohjanvirta, R., J. Tuomisto, T. Vartiainen & K. Rozman: Han/Wistar rats are exceptionally resistant to TCDD. I. Pharma-
cology & Toxicology 1987, 60, 145–150.Pohjanvirta, R., M. Unkila & J. Tuomisto: The loss of glucoprivic
feeding is an early-stage alteration in TCDD-treated Han/Wistarrats. Pharmacology & Toxicology 1990a, 67, 441–443.
Pohjanvirta, R., T. Vartiainen, A. Uusi-Rauva, J. Monkkonen & J.Tuomisto: Tissue distribution, metabolism, and excretion of 14C-TCDD in a TCDD-susceptible and a TCDD-resistant rat strain.Pharmacology & Toxicology 1990b, 66, 93–100.
Pohjanvirta, R., M. Unkila & J. Tuomisto: Characterization of theenhanced responsiveness to postingestive satiety signals in2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-treated Han/Wistarrats. Pharmacology & Toxicology 1991, 69, 433–441.
Pohjanvirta, R., M. Unkila & J. Tuomisto: Comparative acute leth-ality of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), 1,2,3,7,8-pentachlorodibenzo-p-dioxin and 1,2,3,4,7,8-hexachlorodibenzo-p-dioxin in the most TCDD-susceptible and the most TCDD-resistant rat strain. Pharmacology & Toxicology 1993, 73, 52–56.
Pohjanvirta, R., M. Unkila & J. Tuomisto: TCDD-induced hypo-phagia is not explained by nausea. Pharmacol. Biochem. Behav.
1994, 47, 273–282.Sagar, S. M., F. R. Sharp & T. Curran: Expression of c-fos protein
in brain: metabolic mapping at the cellular level. Science 1988,240, 1328–1331.
Santa Cruz Biotechnology, Inc: Datasheet, April 2000.Schwartz, M. W., R. J. Seeley, L. A. Campfield, P. Burn & D. G.
Baskin: Identification of targets of leptin action in rat hypothala-mus. J. Clin. Invest. 1996, 98, 1101–1106.
Schwartz, M. W., S. C. Woods, D. Porte, Jr, R. J. Seeley & D. G.Baskin: Central nervous system control of food intake. Nature
2000, 404, 661–671.Schwartz, M. W., S. C. Woods, R. J. Seeley, G. S. Barsh, D. G.
Baskin & R. L. Leibel: Is the energy homeostasis system in-herently biased toward weight gain? Diabetes 2003, 52, 232–238.
Schwetz, B. A., J. M. Norris, G. L. Sparschu, U. K. Rowe, P. J. Gehr-ing, J. L. Emerson & C. G. Gerbig: Toxicology of chlorinated di-benzo-p-dioxins. Environ. Health Perspect. 1973, 5, 87–99.

371c-Fos IMMUNOSTAINING IN THE BRAINS OF RATS TREATED WITH TCDD AND LEPTIN
Seefeld, M. D., S. W. Corbett, R. E. Keesey & R. E. Peterson: Char-acterization of the wasting syndrome in rats treated with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Appl. Pharmacol. 1984, 73,311–322.
Seo, B. W., B. E. Powers, J. J. Widholm & S. L. Schantz: Radial armmaze performance in rats following gestational and lactationalexposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Neuro-
toxicol. Teratol. 2000, 22, 511–519.Shioda, S., H. Funahashi, S. Nakajo, T. Yada, O. Maruta & Y.
Nakai: Immunohistochemical localization of leptin receptor inthe rat brain. Neurosci. Letters 1998, 243, 41–44.
Shiraishi, T., Y. Oomura, K. Sasaki & M. J. Wayner: Effects ofleptin and orexin-A on food intake and feeding related hypothal-amic neurons. Physiol. Behav. 2000, 71, 251–261.
Shiraishi, T., K. Sasaki, A. Niijima & Y. Oomura: Leptin effects onfeeding-related hypothalamic and peripheral neuronal activitiesin normal and obese rats. Nutrition 1999, 15, 576–579.
Simerly, R. B.: Anatomical substrates of hypothalamic integration.In: The rat nervous system. Ed.: G. Paxinos. Academic Press,1995, pp. 353–376.
Sirkka, U., R. Pohjanvirta, S. A. Nieminen, J. Tuomisto & P. Ylita-lo: Acute neurobehavioural effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in Han/Wistar rats. Pharmacology & Toxi-
cology 1992, 71, 284–288.Smeyne, R. J., M. Vendrell, M. Hayward, S. J. Baker, G. G. Miao,
K. Schilling, L. M. Robertson, T. Curran & J. I. Morgan: Con-tinuous c-fos expression precedes programmed cell death in vivo.Nature 1993, 363, 166–169.
Stumpo, D. J. & P. J. Blackshear: Insulin and growth factor effectson c-fos expression in normal and protein kinase C-deficient 3T3-L1 fibroblasts and adipocytes. Proc. Natl. Acad. Sci. USA 1986,83, 9453–9457.
Tuomisto, J. T., R. Pohjanvirta, M. Unkila & J. Tuomisto: 2,3,7,8-Tetrachlorodibenzo-p-dioxin-induced anorexia and wasting syn-drome in rats: aggravation after ventromedial hypothalamiclesion. Eur. J. Pharmacol. 1995, 293, 309–317.
Tuomisto, J. T., R. Pohjanvirta, M. Unkila & J. Tuomisto: TCDD-
induced anorexia and wasting syndrome in rats: effects of diet-induced obesity and nutrition. Pharmacol. Biochem. Behav.
1999a, 62, 735–742.Tuomisto, J. T., M. Viluksela, R. Pohjanvirta & J. Tuomisto: The
AH receptor and a novel gene determine acute toxic responses toTCDD: segregation of the resistant alleles to different rat lines.Toxicol. Appl. Pharmacol. 1999b, 155, 71–81.
Tuomisto, J. T., M. Viluksela, R. Pohjanvirta & J. Tuomisto:Changes in food intake and food selection in rats after 2,3,7, 8-tetrachlorodibenzo-p-dioxin (TCDD) exposure. Pharmacol. Bio-
chem. Behav. 2000, 65, 381–387.Unkila, M., R. Pohjanvirta, E. MacDonald, J. T. Tuomisto & J.
Tuomisto: Dose response and time course of alterations in tryp-tophan metabolism by 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD) in the most TCDD-susceptible and the most TCDD-resistant rat strain: relationship with TCDD lethality. Toxicol.
Appl. Pharmacol. 1994, 128, 280–292.Van den Berg, M., J. De Jongh, H. Poiger & J. R. Olson: The toxic-
okinetics and metabolism of polychlorinated dibenzo-p-dioxins(PCDDs) and dibenzofurans (PCDFs) and their relevance fortoxicity. Crit. Rev. Toxicol. 1994, 24, 1–74.
Weber, L. W., S. W. Ernst, B. U. Stahl & K. Rozman: Tissue distri-bution and toxicokinetics of 2,3,7,8-tetrachlorodibenzo-p-dioxinin rats after intravenous injection. Fundam. Appl. Toxicol. 1993,21, 523–534.
Whitlock, J. P. Jr: Genetic and molecular aspects of 2,3,7,8-tetra-chlorodibenzo-p-dioxin action. Ann. Rev. Pharmacol. Toxicol.
1990, 30, 251–277.Widholm, J. J., B. W. Seo, B. J. Strupp, R. F. Seegal & S. L. Schantz:
Effects of perinatal exposure to 2,3,7,8-tetrachlorodibenzo-p-di-oxin on spatial and visual reversal learning in rats. Neurotoxicol.
Teratol. 2003, 25, 459–471.Wyss, J. M., K. Tilokskulchai, K. King, I. Kadish & T. van Groen:
Distribution of neurons in the anterior hypothalamic nucleus ac-tivated by blood pressure changes in the rat. Brain Res. Bull.
1999, 49, 163–172.












