
Pretreatment With TCDD Exacerbates Liver Injury From Concanavalin A: Critical Role for NK Cells

ORGAN TOXICITY AND MECHANISMS
Matti Viluksela á Mikko Unkila á Raimo PohjanvirtaJouni T. Tuomisto á Bernhard U. StahlKarl K. Rozman á Jouko Tuomisto
Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)on liver phosphoenolpyruvate carboxykinase (PEPCK) activity,glucose homeostasis and plasma amino acid concentrationsin the most TCDD-susceptible and the most TCDD-resistant rat strains
Received: 29 March 1999 /Accepted: 17 May 1999
Abstract Reduced gluconeogenesis due to decreasedactivity of key gluconeogenic enzymes in liver, togetherwith feed refusal, has been suggested to play an import-ant role in 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-induced lethality in rats. This study was carried out tofurther analyse the toxicological signi®cance of reducedgluconeogenesis by comparing dose-responses and time-courses of e�ects of TCDD on the activity ofphosphoenolpyruvate carboxykinase (PEPCK) in liver,liver glycogen concentration as well as plasma concen-trations of glucose and amino acids in both genders ofTCDD-sensitive Long-Evans (L-E) rats and TCDD-re-sistant Han/Wistar (H/W) rats. A dose-dependent de-crease in PEPCK activity was observed in H/W rats, butin L-E rats the activity was not decreased. However,TCDD impaired the strong increase in liver PEPCKactivity observed in pair-fed controls of the L-E strain.Liver glycogen concentrations were severely decreased inL-E rats and moderately in H/W rats. This e�ect seemsto be secondary to reduced feed intake, since a similardecrease was seen in pair-fed controls. Decreases inplasma glucose concentrations were also more profound
in L-E rats than in H/W rats, but pair-fed controls weregenerally less a�ected. Circulating concentrations ofamino acids were markedly increased in TCDD-treatedL-E rats, which is likely to re¯ect increased mobilizationof amino acids and their decreased metabolism in liver.Reduction of liver PEPCK activity cannot account forthe sensitivity di�erence of these two strains of rats interms of mortality. Nevertheless, the response of bothstrains of TCDD-treated rats regarding gluconeogenesisis di�erent from that seen in pair-fed controls andsuggesting that impairment of this pathway contributesto the development of the wasting syndrome.
Key words 2,3,7,8-Tetrachlorodibenzo-p-dioxin,TCDD á Phosphoenolpyruvate carboxykinase,PEPCK á Glucose á Glycogen á Amino acids
Introduction
Toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD),the most potent congener of polychlorinated dibenzo-p-dioxins, is characterized by a typical wasting syndromein lethally intoxicated animals. Decreased feed intake,body weight loss and exhaustion of energy stores fol-lowed by hypoglycaemia, are well-documented mani-festations of the wasting syndrome, further complicatedby a wide variety of alterations in intermediary metab-olism (Pohjanvirta and Tuomisto 1994). Key lesions andunderlying mechanism(s) of this wasting syndrome arestill incompletely understood.
It has been suggested that the progressive hypo-glycaemia, developing as a consequence of feed refusaland decreased ability to form glucose via the gluconeo-genic pathway, would be at least contributory butpossibly even the ultimate cause of death in TCDD-in-toxicated animals (Gorski and Rozman 1987; Gorskiet al. 1990; Lentnek et al. 1991; Weber et al. 1991a, b).Furthermore, a reduced rate of conversion of 14C-ala-
Arch Toxicol (1999) 73: 323±336 Ó Springer-Verlag 1999
M. Viluksela (&) á R. Pohjanvirta á J.T. Tuomisto á J. TuomistoNational Public Health Institute, Laboratory of Toxicology,P.O. Box 95, FIN-70701 Kuopio, Finlande-mail: matti.viluksela@ktl.®Tel.: +359-17-201329; Fax: +358-17-201265
M. UnkilaDepartment of Pharmacology and Toxicology,University of Kuopio, P.O. Box 1627, FIN-70211 Kuopio,Finland
B.U. Stahl á K.K. RozmanDepartment of Pharmacology, Toxicology and Therapeutics,University of Kansas Medical Center,Kansas City, KS 66160-7417, USA
B.U. Stahl á K.K. RozmanSection of Environmental Toxicology,GSF-Institut fuÈ r Toxikologie,85758 Neuherberg, Germany

nine into 14C-glucose observed in TCDD-treated rats(Gorski et al. 1990) was later concluded to result fromdecreased activity of phosphoenolpyruvate carboxyki-nase (PEPCK), the rate-limiting enzyme of gluconeo-genesis, in liver (Weber et al. 1991a, b).
A peculiar feature in the toxicity of TCDD is hugesensitivity di�erences between species or even strains ofthe same species. The guinea-pig is the most TCDD-sensitive mammalian species with 50% lethal dose(LD50) of �1 lg/kg (McConnell et al. 1978; Schwetzet al. 1973), whereas the hamster is extremely resistantwith an LD50 of >3000 lg/kg (Olson et al. 1980).Studies in our laboratory have discovered an almostsimilar extent of intraspecies di�erence in sensitivitybetween two rat strains (Pohjanvirta et al. 1987, 1993;Unkila et al. 1994). Han/Wistar (Kuopio; H/W) rats are�1000 times more resistant to the acute lethality ofTCDD than Long-Evans (Turku/AB; L-E) rats, havingoral LD50 values of >9600 and 9.8 lg/kg in females,respectively. This di�erence seems to be speci®c forTCDD, since both strains were equally sensitive to theacute toxicity of other types of weight reducing chemi-cals (Unkila et al. 1992).
An interesting di�erence was recently detected byphysicochemical characterization and molecular cloningand sequencing of the hepatic aryl hydrocarbon recep-tors (AHRs) of these strains (Pohjanvirta et al. 1998,1999). The AHR of L-E rat is identical to the previouslyreported Sprague-Dawley rat AHR, while the H/WAHR is structurally deviant. The ®rst invariant nucleo-tide of intron 10 harbours a point mutation, whichresults in an insertion/deletion type alteration at the 3¢end of the coding region of cDNA and a smaller mo-lecular mass for the receptor protein. In spite of thisdi�erence the binding a�nity of TCDD to the AHR isapproximately equal in liver cytosol of both strains. Alsothe H/W type AHR is fully functional in terms of cy-tochrome P450 (CYP) 1A1 induction by TCDD, as theethoxyresoru®n O-deethylase (EROD) activity seems tobe similar in liver and brain of TCDD-treated H/W andL-E rats (Pohjanvirta et al. 1988; Unkila et al. 1993a).Our recent studies suggest that the deviant AHR gene isone of the genes contributing to the resistance of H/Wrats to the lethality of TCDD (Tuomisto et al. 1999).
A conspicuous di�erence between the L-E and H/Wstrains that could be at least partially related to thesensitivity di�erence is their feeding behaviour and bodyweight change. In L-E rats TCDD causes an irreversibleanorexia and body weight loss, while H/W rats generallyresume eating within 1±2 weeks after dosing graduallyreaching a body weight slightly less than that of theircontrols (Pohjanvirta et al. 1987). Correspondingly,increased de novo biosynthesis and turnover of seroto-nin in brain, and elevated concentrations of plasma freetryptophan were detected in lethally intoxicated L-Erats, but not in H/W rats (Tuomisto et al. 1990; Unkilaet al. 1993b, 1994; reviewed by Unkila et al. 1995). Inthis study we have taken advantage of the wide intra-species sensitivity di�erence between H/W and L-E rats
to analyse the role of altered gluconeogenesis and glu-cose homeostasis in the toxicity of TCDD. For thispurpose we have determined dose-response relationshipsof liver PEPCK activity, liver glycogen concentrationsand plasma glucose and amino acid concentrations inboth genders of these rat strains.
Materials and methods
Test chemical
The purity of TCDD was >98% as assessed by gas chromatog-raphy ± mass spectrometry. TCDD was dissolved in corn oil aspreviously described (Pohjanvirta et al. 1987).
Animals
Adult (10- to 16-week-old) male and female inbred Long-Evans(Turku/AB) and outbred Han/Wistar (Kuopio) rats were purchasedfrom the National Laboratory Animal Centre, Kuopio, Finland.Rats were kept individually in plastic metabolism cages (Tecniplast,Buguggiate, Italy) or stainless steel wire bottom cages and given R3feed (Ewos, SoÈ dertaÈ lje, Sweden), and tap water ad libitum. Theroom was arti®cially illuminated from 7.00 a.m. to 7.00 p.m., andthe ambient temperature was 21.5 � 1 °C and relative humidity55 � 10%. The animals were acclimated to the experimentalconditions for at least 2 weeks before dosing.
Experimental design
A detailed description of the experimental design has been givenpreviously (Unkila et al. 1994). Rats were randomly allocated intoexperimental groups of 4±6, corn oil or TCDD was administered byoral gavage, and body weights were recorded every 2 days.
Time-course experiment
Female L-E rats were given a single dose of 50 lg/kg TCDD in thedose volume of 4 ml/kg. Ad libitum -fed and feed-restricted controls(FRC) were given the same volume of the vehicle. FRCs were fedaccording to the daily feed intake pattern recorded in previous ex-periments with L-E rats given 50 lg/kg TCDD, adjusted to theirmetabolic body size. The di�erent groups were scheduled for tissuesampling at 1, 2, 4 and 10 days (FRCs on days 2, 4 and 9) afterdosing. FRCs scheduled for tissue sampling on day 10 wereeuthanized on day 9 because of their debilitated condition (cf.Pohjanvirta et al. 1990). Tissue samples were collected identically tothe other groups. In statistical tests, FRCs of day 9 were comparedwith the ad libitum-fed controls and TCDD-treated group of day 10.
Dose-response experiment
Both genders of L-E andH/W rats were used.Male L-E rats receivedtotal doses of 5, 10, 15, 20 and 50 lg/kg TCDD. Since females aremore sensitive than males (LD50 of 9.8 and 17.7 for females andmales, respectively; Pohjanvirta et al. 1993), the doses for the fe-males were 2.5, 5, 10, 15 and 50 lg/kg. Both genders of H/W ratsreceived 50, 500, 1000 and 9600 lg/kg. Because the highest dosecannot be given as a single dose due to the low solubility of TCDD incorn oil, all the doses were given in a volume of 16 ml/kg divided intofour aliquots of 4 ml/kg each and given over a duration of 48 h.Control groups were given the same volume of vehicle. In addition toad libitum-fed controls, pair-fed control (PFC) L-E rats were addedto control for secondary e�ects due to highly reduced feed intake inlethally intoxicated rats. The PFC were closely matched by bodyweight to the L-E rats of the 50 lg/kg dosage group and given the
324

same amount of feed as their TCDD-treated pairs had consumedvoluntarily on the previous day. No PFCs of the H/W strain wereincluded, because their feed intake and body weight development isonly slightly a�ected by TCDD. Tissue samples were taken 6 daysafter the beginning of dosing.
Tissue sampling
At the end of the treatment period rats were decapitated and trunkblood collected into a dish containing �25 IU heparin/ml blood,centrifuged at 1000 g for 10 min, plasma separated and stored at)80 °C. Livers were quickly removed, weighed, frozen in liquidnitrogen and stored at )80 °C until analyses.
Biochemical analyses
Liver EROD activity
Ethoxyresoru®n O-deethylase activity in liver S9 fractions wasmeasured ¯uorometrically according to Dutton Parkinson (1989)with minor modi®cations as described by Weber et al. (1992). Onlymales of the dose-response experiment were studied.
Liver PEPCK activity
Liver PEPCK (EC 4.1.1.32) activity was measured using the bio-luminescent method of Wimmer (1988) with minor modi®cations asdescribed by Viluksela et al. (1995b). Liver total PEPCK capacitywas calculated by multiplying the speci®c activity with liver S9protein concentration and liver weight.
Liver glycogen content
Liver glycogen concentration was measured by the method ofHultman (1967) with minor modi®cations as described by Unkilaet al. (1995). Liver glycogen content was calculated by multiplyingthe glycogen concentration with liver weight.
Plasma glucose concentration
Plasma glucose concentration was determined with Peridochrom
GlucoseÒ kits (Boehringer Mannheim GmbH, Mannheim, Ger-many).
Plasma amino acid and urea concentrations
Plasma concentrations of amino acids and urea were measuredwith an amino acid analyser (LKB Biochrom 4151 Alpha Plus)using an Ultrapac C8-column and lithium citrate bu�er (PharmaciaBiotech Ltd, Cambridge, England).
Statistics
Datawere tested for homogeneity of group variances using Bartlett'stest. If the data were homogeneous as such or after a logarithmic orsquare root transformation (at the level of P £ 0.01), comparisonsbetween treatment groups were performed using one-way analysis ofvariance (ANOVA) followed by the least signi®cant di�erence(LSD) test. If the data were heterogeneous comparisons were madeusing the Kruskal-Wallis test followed by theMann-WhitneyU-test.
Results
Body weight development
Body weights of L-E rats were dose-dependentlydecreased in all TCDD-treated groups 6 days after
dosing, maximally by 18 (males) and 27% (females) at50 lg/kg TCDD, respectively (data not shown). In pair-fed rats the decrease in body weight was identical totheir TCDD-treated counterparts. Body weights of H/Wrats were only slightly a�ected resulting in a decrease by9 (males) and 7% (females) at 9600 lg/kg TCDD. Thisdose was previously shown to cause mortality of only10% in female H/W rats (Unkila et al. 1994).
Liver EROD activity
TCDD-treatment resulted in a similar maximum in-duction of liver EROD activity in both L-E and H/Wrats (Fig. 1). Dose-responses of EROD activity are dif-®cult to compare, because the lowest doses causedmaximal or nearly maximal induction in L-E but not inH/W rats. In L-E rats the highest EROD activity wasmeasured at the lowest dose of TCDD (5 lg/kg),whereas with increasing doses the activity declined, mostlikely due to overt toxicity and liver atrophy (see below).In H/W rats there was a dose-related increase in ERODactivity from 50 to 1000 lg/kg TCDD, after which theactivity plateauxed (Fig. 1). EROD activity of PFCs didnot di�er from that of ad libitum-fed controls.
Relative liver weights
Relative liver weights of TCDD-treated (50 lg/kg) fe-male L-E rats were signi®cantly (P < 0.001) increasedon days 1±4 after dosing, but signi®cantly (P < 0.001)decreased on day 10 compared to ad libitum-fed controls(Fig. 2A). In FRCs the values decreased progressivelyon days 2±9 (P < 0.01±0.001). When examined 6 daysafter the beginning of dosing, the relative liver weights ofboth male and female L-E rats were increased at lowerdoses and decreased at the highest dose (s) (Fig. 3A, B);female L-E rats were more sensitive. In PFCs of bothgenders the decrease was signi®cantly more pronounced(P < 0.001) than at 50 lg/kg TCDD. In both genders
Fig. 1 Dose-responses of liver 7-ethoxyresoru®n O-deethylase(EROD) activity in male Long-Evans (L-E) and Han/Wistar (H/W)rats 6 days after starting the dosing regimen. (PF Pair-fed controls).Group mean � SE, n = 4±6
325

of H/W rats the relative liver weights showed a patternof dose-dependent increase.
Liver PEPCK activity
Liver PEPCK activity in TCDD-treated (50 lg/kg) fe-male L-E rats was not signi®cantly di�erent from adlibitum-fed controls on days 1±4. The activity subse-
quently increased to a level �2.2 times higher than thecontrol activity (P < 0.01) by day 10 (Fig. 2B). FRCsshowed a parallel time-dependent increase in PEPCKactivity as was seen in the TCDD-treated group, whichwas maximally 3.5 times higher than the activity inad libitum -fed controls. The PEPCK activity of FRCswas consistently �100 nmol/(mg ´ min) above that ofthe TCDD-treated group (P < 0.001 on days 2 and 4).
Basal liver PEPCK activity in both male and femaleL-E controls was only about one-half of the activitymeasured in male and female H/W controls (Fig. 3C,D). In both strains the activity was slightly lower infemales than in males. In TCDD-treated H/W rats thePEPCK activity was dose-dependently decreased start-ing at 50 lg/kg in males (P < 0.001) and 5000 lg/kg infemales, maximally to 45 and 75% of the control ac-tivity, respectively. Male L-E rats showed decreasedPEPCK activity at the two lowest doses only (P < 0.05vs ad libitum -fed controls at 5 and 10 lg/kg), whereas infemale L-E rats the activity was dose-dependently in-creased to 174% of the control activity (P < 0.05 at10 lg/kg and P < 0.001 at 50 lg/kg). Pair-feeding for 6days increased the liver PEPCK activity 2.6- and 2.7-fold compared with ad libitum-fed control males andfemales, respectively (P < 0.001). PEPCK activity inthe pair-fed males and females was signi®cantly(P < 0.001; 2.5- and 1.5-fold, respectively) higher thanin their TCDD-treated counterparts, indicating thatTCDD-treatment suppressed the physiological responseto reduced feed intake.
Because TCDD treatment resulted in strain-speci®cchanges in liver weigths, liver total PEPCK capacitieswere calculated. Due to decreased liver weights in L-Erats at higher doses of TCDD their liver total PEPCKcapacities were rather invariable (Figs. 2C; 3E, F).However, total PEPCK capacities of FRC/PFCs wereclearly higher than those of their TCDD-treated coun-terparts.
Plasma glucose concentration
TCDD treatment resulted in signi®cantly decreasedplasma glucose concentrations in female L-E rats ondays 1±4 after dosing (Fig. 4A). Surprisingly, the levelswere close to those of ad libitum-fed controls on day 10.The reason for this unexpected ®nding for a single datapoint is not known. FRCs were slightly higher than theTCDD-treated group on day 4, but the plasma glucoseconcentration continued to decrease to 58% of that inad libitum-fed controls by day 9 (P < 0.01). In bothgenders of H/W rats plasma glucose concentrations werevery slightly decreased at higher doses of TCDD(P < 0.01 in females only), while L-E rats showed asigni®cant (P < 0.05±0.001) dose-dependent decreasewithin the whole dose-range studied (Fig. 5A, B). Thevalues for PFCs were higher than their TCDD-treatedcounterparts, but signi®cantly (P < 0.01) lower thanthe ad libitum -fed controls.
Fig. 2 Time-course of A relative liver weight, B liverphosphoenolpyruvate carboxykinase (PEPCK) activity and C livertotal PEPCK capacity in female Long-Evans rats after a single oraldose of 50 lg/kg 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). (Ctrad libitum-fed controls á FR feed-restricted controls). Group mean �SE, n=6, except for FR day 9 where n=5. *P < 0.05, **P < 0.01,***P < 0.001. aSigni®cantly di�erent from ad libitum -fed controls,bsigni®cantly di�erent from FR controls
326

Liver glycogen content
Progressively decreasing liver glycogen contents wereobserved in TCDD-treated (50 lg/kg) female L-E ratsfrom day 4 after dosing (Fig. 4B). On day 10 the mean
glycogen content was only 16% of that in ad libitum -fedcontrols (P < 0.001). The decrease was even faster andmore severe in FRCs: the level was statistically signi®-cant already on day 2 (P < 0.01 vs. ad libitum -fedcontrols) and the maximum decrease (on day 9) was13% of that in ad libitum -fed controls (on day 10;P < 0.001).
In male H/W rats, the liver glycogen contents wereslightly but still dose-dependently decreased on day 6after the beginning of TCDD-treatment (Fig. 5C), butthere was no e�ect in female H/W rats (Fig. 5D). Inboth genders of L-E rats the dose-dependent decreasewas very profound with maximum e�ect at 50 lg/kg
Fig. 3 Dose-responses of A, B relative liver weight, C, D liver PEPCKactivity and E, F liver total PEPCK capacity in male (left panel) andfemale (right panel) L-E and H/W rats 6 days after starting the dosingregimen. PF placed for better comparison on x-axis at 50 lg/kgTCDD. Group mean � SE, n=4±6. *P < 0.05, **P < 0.01,***P< 0.001. aSigni®cantly di�erent from ad libitum -fed controls,bsigni®cantly di�erent from pair-fed controls
327

TCDD (16 and 8% of ad libitum -fed controls in malesand females, respectively).
Plasma amino acid concentrations
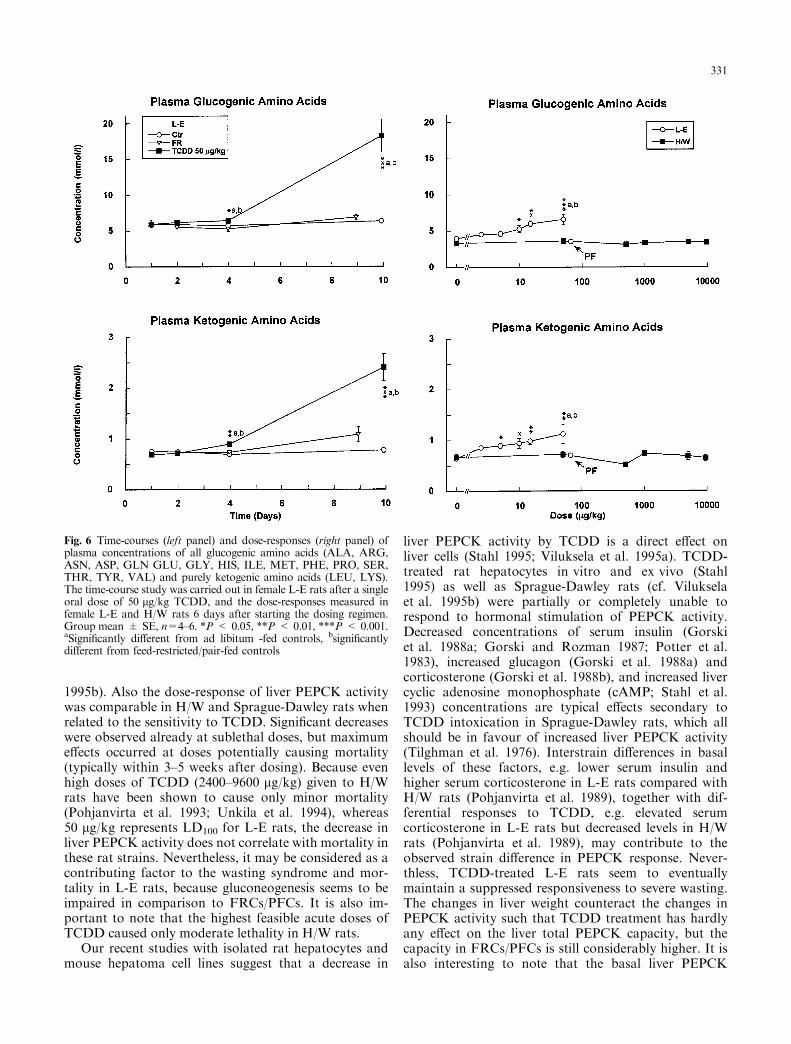
Most remarkable changes in plasma amino acid con-centrations were observed in female L-E rats 10 daysafter a dose of 50 lg/kg TCDD (Table 1, Figs. 6 and 7).At lower doses and at shorter exposure times the chan-ges were de®nitely smaller (Table 2, Figs. 6 and 7). Ingeneral, only minor changes were observed in H/W rats,and females were more a�ected than males at the dosesemployed (data for males not shown). An overall sum-mary of plasma amino acid concentrations is shown inFig. 6, where plasma concentrations of all glucogenicamino acids (amino acids that can be utilized for glu-coneogenesis, i.e. alanine, arginine, asparagine, aspar-tate, glutamine, glutamate, glycine, histidine, isoleucine,methionine, phenylalanine, proline, serine, threonine,tyrosine and valine), as well as plasma concentrations ofpurely ketogenic amino acids (amino acids that cannotbe converted to glucose, i.e. leucine and lysine; Stryer1996) are summarized separately. These groups of amino
acids followed very similar time-courses and dose-re-sponses. Plasma amino acid concentrations were highly(�three-fold; P < 0.001) elevated in TCDD-treated ratson day 10 of the time-course study. There was also adose-dependent increase in TCDD-treated L-E rats,when the samples were taken on day 6 after the begin-ning of dosing.
Nearly all amino acids followed the general trend ofhighly elevated plasma concentrations on day 10 afterTCDD treatment in the time-course study. Only con-centrations of glutamine and isoleucine were not chan-ged, and those of proline, valine and leucine displayedonly minor increases (Table 1). In FRCs, the concen-trations of most amino acids were either not changed orwere slightly and temporarily decreased, except thatthose of arginine were consistently decreased(P < 0.05±0.01). Valine, leucine and isoleucine, how-ever, were increased (on day 9) compared with concen-trations found in TCDD-treated rats, but ornithine,phenylalanine and tyrosine only increased above ad li-bitum -fed controls. Thus, amino acids with a branchedand hydrophobic side-chain (valine, leucine and isoleu-cine) responded di�erently from other amino acids.Their plasma concentrations were more elevated inFRCs than in TCDD-treated rats (increases in isoleucineconcentrations were very small and nonsigni®cant).Also, phenylalanine and tyrosine (amino acids with anaromatic side-chain) responded similarly in that theirplasma concentrations were increased in FRCs on day 9,although less than in TCDD-treated rats. The same wasalso true for ornithine.
A similar trend with less pronounced dose-relatedchanges was observed in L-E rats in the dose-responsestudy (Table 2). In H/W rats statistically signi®cant in-creases were only found in plasma concentrations ofserine and threonine. These hydroxylated amino acids,as well as ornithine and lysine, were the most sensitivealso in L-E rats, as their concentrations were elevatedalready at low doses. Plasma concentrations of tyrosineshowed an unusual pattern in L-E rats in both time-course and dose-response studies. Concentrations weresigni®cantly (P < 0.001) decreased already on day 1,started to recover after day 4, and were signi®cantly(P < 0.05) increased on day 10 (Fig. 7). (In fact, theother hydroxylated amino acids serine and threoninealso exhibited a similar but less clear decrease on days 1±2.) In the dose-response study plasma tyrosine concen-trations were signi®cantly (P < 0.05±0.01) decreased at2.5 and 5 lg/kg, but signi®cantly (P < 0.01) increasedat 50 lg/kg TCDD.
Plasma urea concentrations
In FRC female L-E rats plasma urea concentrationswere signi®cantly (P < 0.001) elevated on day 9(Fig. 7). TCDD treatment, in contrast, resulted in aslight but signi®cant decrease from day 2 (P < 0.05) today 10 (P < 0.01). In the dose-response study plasma
Fig. 4 Time-course of A plasma glucose concentration and B liverglycogen content in female L-E rats after a single oral dose of 50 lg/kg TCDD. Group mean � SE, n=6, except for FR day 9 wheren=5. *P < 0.05, **P < 0.01, ***P < 0.001. aSigni®cantly di�erentfrom ad libitum -fed controls, bsigni®cantly di�erent from feed-restricted controls
328

urea concentrations of pair-fed L-E rats were increasedless (P < 0.05) than in the time-course study. InTCDD-treated L-E rats plasma urea concentrationstended to increase, but in H/W rats there was adecreasing trend.
Discussion
This study was carried out to characterize the role ofglucose homeostasis and reduced gluconeogenesis inlethality and wasting syndrome caused by TCDD. Ingeneral, liver PEPCK activity, liver glycogen concen-tration and plasma concentrations of glucose and aminoacids were less a�ected in H/W rats than in L-E rats.This is consistent with the observations on feed intake,body weight development and mortality in these ratstrains (Pohjanvirta et al. 1987, 1993; Unkila et al.1993b). Due to limited solubility of TCDD comparisonof these rat strains at doses completely equipotent in
terms of mortality is unfortunately not feasible in acutestudies. In spite of the vast di�erence in sensitivity tomortality it is remarkable that the maximum ERODinduction is quite similar in these strains (Pohjanvirtaet al. 1988; Unkila et al. 1993a; this study), although atlethal doses EROD activity seems to be compromised inL-E rats along with liver atrophy.
TCDD treatment has been previously shown todecrease liver PEPCK activity in Sprague-Dawley rats(Stahl et al. 1993; Viluksela et al. 1995b; Weber et al.1991a, b), in a Charles River substrain of the Long-Evansrat (Fan and Rozman 1994, 1995), as well as in mice(Weber et al. 1995) and hamsters (Unkila et al. 1995).Contrary to those ®ndings, TCDD treatment caused nochange or an increase in liver PEPCK activity in L-E rats(except in males at the two lowest doses, i.e. at doses thatdo not cause wasting) as compared to ad libitum -fedcontrols. However, it prevented or suppressed the pro-gressive increase in PEPCK activity caused by restrictedfeed intake, as seen in FRCs/PFCs.At 50 lg/kg of TCDDliver PEPCK activity was only 40 or 65% of that in PFCmales and females, respectively. Thus, TCDD treatmentimpaired the well established physiological response toundernutrition (Tilghman et al. 1976) in L-E rats. InH/Wmales, however, TCDD treatment resulted in a decreaseof PEPCK activity to 45% of the activity measured in adlibitum -fed controls, which is nearly as much as in themore sensitive Sprague-Dawley rats (Viluksela et al.
Fig. 5 Dose-responses of A, B plasma glucose concentrations and C,D liver glycogen content in male (left panel) and female (right panel)L-E and H/W rats 6 days after starting the dosing regimen. Groupmean � SE, n=4±6. *P < 0.05, **P < 0.01, ***P < 0.001.aSigni®cantly di�erent from ad libitum -fed controls, bsigni®cantlydi�erent from pair-fed controls. The results of female rats were partlypublished previously (Pohjanvirta et al. 1995), but are shown here forcomparison
329

Table
1Plasm
aconcentrationsofaminoacidsandurea(m
mol/l;groupmean�
SE,n=
5±6)in
thetime-coursestudyin
femaleLong-Evansrats
after
asingleoraldose
of50
lg/kg
2,3,7,8-tetrachlorodibenzo-p-dioxin
(TCDD)
Group
Tim
e
(Days)
ALA
ARG
ASN
ASP
GLN
GLU
GLY
HIS
ILE
LEU
Control
10.650�
0.024
0.246�
0.009
0.134�
0.005
0.0215�
0.0018
0.243�
0.010
0.268�
0.011
0.331�
0.008
0.0664�
0.0109
0.118�
0.011
0.199�
0.014
Control
20.581�
0.028
0.223�
0.016
0.099�
0.008
0.0475�
0.0096
0.328�
0.017
0.155�
0.007
0.265�
0.013
0.0546�
0.0025
0.139�
0.012
0.200�
0.009
Control
40.580�
0.038
0.221�
0.011
0.098�
0.007
0.0297�
0.0092
0.283�
0.013
0.183�
0.014
0.272�
0.008
0.0539�
0.0025
0.137�
0.009
0.211�
0.004
Control
10
0.658�
0.034
0.242�
0.017
0.127�
0.011
0.0129�
0.0064
0.322�
0.019
0.198�
0.025
0.315�
0.029
0.0612�
0.0049
0.134�
0.009
0.212�
0.012
Feed-restricted
20.461�
0.035
0.182�
0.013
0.105�
0.010
0.0269*�
0.0019
0.214***�
0.009
0.223**�
0.009
0.285�
0.017
0.0461�
0.0033
0.144�
0.013
0.210�
0.012
Feed-restricted
40.389***�
0.027
0.170**�
0.007
0.119�
0.029
0.0340�
0.0179
0.258�
0.022
0.210�
0.030
0.298�
0.024
0.0402***�
0.00240.157�
0.020
0.213�
0.025
Feed-restricted
90.765�
0.116
0.112*�
0.032
0.154�
0.022
0.0205�
0.0020
0.294�
0.045
0.215�
0.029
0.290�
0.034
0.0743�
0.0059
0.170�
0.026
0.347*�
0.047
TCDD
50
lg/kg
10.636�
0.034
0.237�
0.015
0.121�
0.015
0.0309�
0.0060
0.258�
0.032
0.262�
0.015
0.276**�
0.015
0.0502�
0.0047
0.155�
0.012
0.213�
0.009
TCDD
50
lg/kg
20.745*�
0.078
0.202�
0.014
0.077*�
0.007
0.0202**�
0.0034
0.345�
0.012
0.259***�
0.023
0.277�
0.013
0.0546�
0.0034
0.110�
0.005
0.191�
0.004
TCDD
50
lg/kg
40.612�
0.033
0.028***�
0.007
0.108�
0.004
0.0089�
0.0040
0.427***�
0.019
0.197�
0.019
0.277�
0.009
0.0536�
0.0019
0.133�
0.006
0.240�
0.003
TCDD
50
lg/kg
10
2.028***�
0.285
0.676***�
0.108
0.488�
0.077
0.0596***�
0.00571.732***�
0.371
0.220�
0.019
0.702***�
0.078
0.2182***�
0.03780.165�
0.006
0.297*�
0.013
Group
Tim
e
(Days)
LYS
MET
ORN
PHE
PRO
SER
THR
TYR
VAL
UREA
Control
10.551�
0.028
0.0801�
0.0148
0.146�
0.024
0.102�
0.007
0.446�
0.043
0.315�
0.007
0.331�
0.016
0.111�
0.005
0.225�
0.017
7.93�
0.42
Control
20.547�
0.020
0.0964�
0.0109
0.133�
0.009
0.092�
0.008
0.353�
0.041
0.274�
0.015
0.310�
0.011
0.113�
0.004
0.244�
0.015
8.05�
0.54
Control
40.482�
0.030
0.0898�
0.0102
0.147�
0.012
0.103�
0.004
0.363�
0.031
0.252�
0.012
0.248�
0.017
0.125�
0.005
0.254�
0.009
7.65�
0.42
Control
10
0.559�
0.044
0.0835�
0.0078
0.135�
0.010
0.089�
0.008
0.556�
0.102
0.296�
0.020
0.314�
0.024
0.129�
0.008
0.263�
0.022
9.85�
0.65
Feed-restricted
20.514�
0.031
0.0878�
0.0112
0.117�
0.009
0.100�
0.008
0.319�
0.040
0.246�
0.011
0.262�
0.023
0.119�
0.010
0.224�
0.018
7.61�
0.88
Feed-restricted
40.518�
0.018
0.0913�
0.0235
0.105*�
0.006
0.112�
0.014
0.290�
0.090
0.247�
0.030
0.266�
0.036
0.117�
0.020
0.211�
0.026
6.49�
0.75
Feed-restricted
90.737�
0.109
0.0968�
0.0148
0.329*�
0.095
0.147*�
0.015
0.300*�
0.052
0.214�
0.034
0.317�
0.026
0.184�
0.029
0.448*�
0.062
23.08***�
3.34
TCDD
50
lg/kg
10.475�
0.029
0.0974�
0.0104
0.116�
0.009
0.110�
0.014
0.415�
0.030
0.283*�
0.011
0.285�
0.018
0.057***�
0.008
0.224�
0.011
7.99�
0.68
TCDD
50
lg/kg
20.524�
0.023
0.0798�
0.0030
0.230**�
0.029
0.098�
0.007
0.289�
0.034
0.308�
0.018
0.317�
0.019
0.042***�
0.002
0.229�
0.010
5.76�
0.32
TCDD
50
lg/kg
40.655***�
0.031
0.0946�
0.0021
0.444***�
0.020
0.128�
0.008
0.291�
0.030
0.344**�
0.017
0.495***�
0.022
0.070***�
0.005
0.263�
0.011
6.14�
0.45
TCDD
50
lg/kg
10
2.109***�
0.251
0.4120***�
0.0754
0.653***�
0.115
0.200***�
0.022
0.657�
0.055
1.257***�
0.157
1.631***�
0.240
0.214*�
0.021
0.336�
0.031
6.12*�
0.39
*P<
0.05,**P<
0.01,***P<
0.001vsadlibitum
-fed
controls
330
1995b). Also the dose-response of liver PEPCK activitywas comparable in H/W and Sprague-Dawley rats whenrelated to the sensitivity to TCDD. Signi®cant decreaseswere observed already at sublethal doses, but maximume�ects occurred at doses potentially causing mortality(typically within 3±5 weeks after dosing). Because evenhigh doses of TCDD (2400±9600 lg/kg) given to H/Wrats have been shown to cause only minor mortality(Pohjanvirta et al. 1993; Unkila et al. 1994), whereas50 lg/kg represents LD100 for L-E rats, the decrease inliver PEPCK activity does not correlate with mortality inthese rat strains. Nevertheless, it may be considered as acontributing factor to the wasting syndrome and mor-tality in L-E rats, because gluconeogenesis seems to beimpaired in comparison to FRCs/PFCs. It is also im-portant to note that the highest feasible acute doses ofTCDD caused only moderate lethality in H/W rats.
Our recent studies with isolated rat hepatocytes andmouse hepatoma cell lines suggest that a decrease in
liver PEPCK activity by TCDD is a direct e�ect onliver cells (Stahl 1995; Viluksela et al. 1995a). TCDD-treated rat hepatocytes in vitro and ex vivo (Stahl1995) as well as Sprague-Dawley rats (cf. Vilukselaet al. 1995b) were partially or completely unable torespond to hormonal stimulation of PEPCK activity.Decreased concentrations of serum insulin (Gorskiet al. 1988a; Gorski and Rozman 1987; Potter et al.1983), increased glucagon (Gorski et al. 1988a) andcorticosterone (Gorski et al. 1988b), and increased livercyclic adenosine monophosphate (cAMP; Stahl et al.1993) concentrations are typical e�ects secondary toTCDD intoxication in Sprague-Dawley rats, which allshould be in favour of increased liver PEPCK activity(Tilghman et al. 1976). Interstrain di�erences in basallevels of these factors, e.g. lower serum insulin andhigher serum corticosterone in L-E rats compared withH/W rats (Pohjanvirta et al. 1989), together with dif-ferential responses to TCDD, e.g. elevated serumcorticosterone in L-E rats but decreased levels in H/Wrats (Pohjanvirta et al. 1989), may contribute to theobserved strain di�erence in PEPCK response. Never-thless, TCDD-treated L-E rats seem to eventuallymaintain a suppressed responsiveness to severe wasting.The changes in liver weight counteract the changes inPEPCK activity such that TCDD treatment has hardlyany e�ect on the liver total PEPCK capacity, but thecapacity in FRCs/PFCs is still considerably higher. It isalso interesting to note that the basal liver PEPCK
Fig. 6 Time-courses (left panel) and dose-responses (right panel) ofplasma concentrations of all glucogenic amino acids (ALA, ARG,ASN, ASP, GLN GLU, GLY, HIS, ILE, MET, PHE, PRO, SER,THR, TYR, VAL) and purely ketogenic amino acids (LEU, LYS).The time-course study was carried out in female L-E rats after a singleoral dose of 50 lg/kg TCDD, and the dose-responses measured infemale L-E and H/W rats 6 days after starting the dosing regimen.Group mean � SE, n=4±6. *P < 0.05, **P < 0.01, ***P < 0.001.aSigni®cantly di�erent from ad libitum -fed controls, bsigni®cantlydi�erent from feed-restricted/pair-fed controls
331

speci®c activity in control L-E rats is close to themaximally reduced speci®c activity measured in Spra-gue-Dawley rats at doses of TCDD causing mortality(Viluksela et al. 1995b).
Liver glycogen contents were severely decreased inTCDD-treated L-E rats, but because decreases of similarextent (although faster) were also seen in PFCs andFRCs, the decrease is most likely a consequence of re-duced feed intake. In H/W males the decrease wasmodest, following closely the pattern of decrease in liverPEPCK activity. Consistent with our results, previousstudies have demonstrated decreased liver glycogencontents in rats treated with TCDD or other dioxins(Pohjanvirta et al. 1995; Weber et al. 1983), even aftercontinuous total parenteral nutrition (containing nearly20% dextrose) (Gasiewicz et al. 1980).
Also in accordance with previous reports (Pohjan-virta et al. 1989; Potter et al. 1983; Zinkl et al. 1973),serum glucose concentrations decreased in both strains,though more so in L-E rats. Their PFCs were less af-fected, in agreement with the notion that intact gluco-neogenesis alleviates the development of hypoglycaemiain starvation. However, plasma glucose concentrationsof lethally intoxicated female L-E rats returned to nor-mal at day 10 of the time-course study. This ®ndingsuggests that the rate of gluconeogenesis, although de-creased in comparison to FRCs, was still su�cient tomaintain normoglycaemia in the presence of high con-centrations of circulating amino acids.
TCDD intoxication resulted in an overall increase inconcentrations of circulating amino acids, which wasultimately elevated about three-fold on day 10 in L-Erats. This increase was not secondary to reduced feedintake as such, because in FRCs/PFCs the concentra-tions of most amino acids did not change or were onlyslightly increased. Our results are in overall agreementwith the data of Christian et al. (1986b) from schedule-fed mature Sprague-Dawley rats (although this studyfound slightly decreased plasma levels of glutamine,leucine and isoleucine in TCDD-treated rats). The in-creases were generally smaller than in our study withyounger and more sensitive L-E rats. Plasma alanineconcentrations reported by Gorski et al. (1990) forTCDD-treated as well as PFC and ad libitum-fed controlSprague-Dawley rats are also very similar to our data.
The increase in circulating levels of amino acids mayresult from enhanced mobilization of amino acids frommuscle, their decreased metabolism, or both. Previousstudies have reported a severe muscle atrophy (Maxand Silbergeld 1987) and a signi®cant loss of totalcarcass protein (Christian et al. 1986a) in TCDD-treated rats at dose-levels causing lethality. Thesechanges result from reduced feed intake alone, becausePFCs were a�ected to a similar extent (Christian et al.1986a). The fact that plasma urea concentrations wereelevated only in FRCs/PFCs (especially on day 9),suggests that TCDD treatment prevents the starvation-accelerated metabolism of amino acids, which in turnresults in increased levels of circulating amino acids.
Decreased amino acid metabolism in TCDD-treatedrats has been previously suggested based on measure-ments of plasma amino acid concentrations (Christianet al. 1986b) and analysis of respiratory quotients(Muzi et al. 1989). This was con®rmed in experiments,which demonstrated a markedly decreased disappear-ance of 14C-alanine from plasma of TCDD-treated ratscompared to ad libitum -fed controls and PFCs, as wellas highly reduced conversion of 14C-alanine into14C-glucose in TCDD-treated rats (Gorski et al. 1990).These changes were observed 4±16 days after dosing.Reduced utilization of amino acids for gluconeogenesisdue to decreased activity of liver PEPCK (in compar-ison to FRCs/PFCs) is an obvious reason for increasedplasma amino acid levels in TCDD-treated rats;because, however, the levels of purely ketogenic aminoacids were similarly increased, this cannot be the onlymechanism involved. It is therefore likely that TCDD-treatment, in addition to reduced feed intake, results inincreased mobilization of amino acids in L-E rats. InH/W rats plasma amino acid levels were not a�ected byTCDD treatment, although liver PEPCK activity wasdecreased. A likely explanation is that H/W rats werenot dependent on gluconeogenesis, because their feedintake and body weight development were only slightlydecreased.
Amino acids with branched and hydrophobic side-chains, i.e. valine, leucine and isoleucine, di�ered fromthe general trend seen with other amino acids, becausetheir plasma levels in TCDD-treated rats were maxi-mally only 1.2- to 1.4-fold higher than in ad libitum -fedcontrols. Interestingly, branched-chain amino acids areprimarily metabolized in muscles and brain, not in liver.If the metabolism of these amino acids in muscles is nota�ected by TCDD, their circulating concentrationswould not be expected to rise. It is also interesting tonote that liver disease is often characterized by elevatedcirculating concentrations of amino acids that are me-tabolized in liver with a simultaneous reduction in cir-culating levels of branched-chain amino acids andaspartate (cf. Christian et al. 1986b). Other amino acidsexhibiting responses di�erent from the general trendwere aromatic amino acids phenylalanine and tyrosine,which share a common pathway of metabolism (Stryer1996), and the hydroxylated amino acids serine andthreonine, which responded already at low doses ofTCDD.
In conclusion, glucose homeostasis, as indicated bychanges in plasma glucose and liver glycogen levels,was severely a�ected by acute exposure to TCDD inL-E rats, but moderately in H/W rats. Increased levels
Fig. 7 Time-courses (left panel) and dose-responses (right panel) ofplasma concentrations of some amino acids and urea. Selected aminoacids represent di�erent types of responses (see text). See the legend toFig. 6 for experimental details. Group mean � SE, n=4±6.*P < 0.05, **P < 0.01, ***P < 0.001. aSigni®cantly di�erent fromad libitum -fed controls, bsigni®cantly di�erent from feed-restricted/pair-fed controls
c
332

333

Table
2Plasm
aconcentrationsofaminoacidsandurea(m
mol/l;groupmean�
SE,n=
4±6)in
thedose-response
studyin
female
Long-EvansandHan/W
istarrats
6daysafter
startingthedosingregim
en(C
trAdlibitum
-fed
controlsPFpair-fed
controls)
Strain
Dose
(lg/kg)
ALA
ARG
ASN
ASP
GLN
GLU
GLY
HIS
ILE
LEU
Long-Evans
Ctr
0.693�
0.011
0.289�
0.008
0.109�
0.013
0.032�
0.002
0.580�
0.010
0.331�
0.045
0.337�
0.005
0.086�
0.005
0.162�
0.004
0.202�
0.003
30.852�
0.031
0.212�
0.022
0.105�
0.007
0.055�
0.006
0.729�
0.016
0.313�
0.014
0.342�
0.006
0.086�
0.006
0.159�
0.003
0.222�
0.001
50.850�
0.031
0.181*�
0.014
0.169�
0.012
0.062�
0.005
0.662�
0.010
0.412�
0.034
0.386�
0.009
0.073�
0.005
0.158�
0.005
0.224�
0.003
10
0.938�
0.049
0.211�
0.010
0.156�
0.010
0.054�
0.010
0.784**�
0.029
0.322�
0.028
0.424�
0.016
0.071�
0.003
0.144�
0.004
0.224�
0.006
20
1.196**�
0.067
0.218�
0.011
0.160�
0.006
0.027�
0.001
0.928***�
0.0190.240�
0.004
0.485**�
0.013
0.062�
0.001
0.154�
0.003
0.222�
0.004
50
1.153**�
0.106
0.367�
0.018
0.222*�
0.010
0.054�
0.006
1.143***�
0.0320.286�
0.000
0.522**�
0.032
0.086�
0.002
0.174�
0.008
0.258�
0.012
PF
0.487*�
0.008
0.202�
0.022
0.114�
0.008
0.027�
0.001
0.618�
0.025
0.228�
0.025
0.357�
0.016
0.073�
0.005
0.176�
0.007
0.272�
0.012
Han/W
istar
Ctr
0.486�
0.016
0.285�
0.016
0.056�
0.002
0.029�
0.002
0.609�
0.023
0.211�
0.0154
0.314�
0.014
0.086�
0.009
0.144�
0.002
0.177�
0.004
50
0.568�
0.024
0.255�
0.011
0.072�
0.008
0.029�
0.000
0.617�
0.032
0.321�
0.028
0.343�
0.011
0.102�
0.008
0.130�
0.003
0.175�
0.006
500
0.478�
0.014
0.245�
0.009
0.065�
0.003
0.038�
0.004
0.421�
0.029
0.181�
0.005
0.315�
0.027
0.098�
0.040
0.128�
0.004
0.154�
0.003
1000
0.482�
0.016
0.249�
0.007
0.086�
0.003
0.031�
0.002
0.637�
0.030
0.308�
0.014
0.336�
0.010
0.089�
0.007
0.131�
0.004
0.175�
0.005
5000
0.531�
0.010
0.244�
0.023
0.120�
0.015
0.022�
0.002
0.530�
0.024
0.303�
0.043
0.342�
0.012
0.084�
0.008
0.145�
0.005
0.189�
0.006
9600
0.580�
0.023
0.252�
0.007
0.105�
0.011
0.032�
0.003
0.493�
0.013
0.327�
0.031
0.318�
0.017
0.083�
0.008
0.109�
0.004
0.152�
0.005
Strain
Dose
(lg/kg)
LYS
MET
ORN
PHE
PRO
SER
THR
TYR
VAL
UREA
Long-Evans
Ctr
0.439�
0.009
0.069�
0.005
0.095�
0.004
0.074�
0.002
0.323�
0.023
0.374�
0.011
0.368�
0.013
0.092�
0.002
0.217�
0.004
10.04�
0.54
30.630**�
0.013
0.086�
0.003
0.235***�
0.0190.086�
0.002
0.257�
0.013
0.463�
0.008
0.583*�
0.015
0.052**�
0.0020.245�
0.002
12.15�
0.50
50.667**�
0.020
0.085�
0.008
0.244***�
0.0150.094�
0.001
0.331�
0.026
0.489*�
0.014
0.617**�
0.016
0.062*�
0.001
0.231�
0.005
13.88*�
0.49
10
0.718**�
0.035
0.113�
0.005
0.233***�
0.0190.120***�
0.0050.357�
0.026
0.584**�
0.021
0.729***�
0.0290.084�
0.003
0.226�
0.008
11.84�
0.23
20
0.763**�
0.022
0.076�
0.009
0.226***�
0.0060.131***�
0.0030.405�
0.020
0.722***�
0.0130.991***�
0.0260.092�
0.003
0.247�
0.004
10.64�
0.22
50
0.868�
0.083
0.117�
0.008
0.210**�
0.007
0.162***�
0.0060.382�
0.030
0.713***�
0.0531.018***�
0.0580.129**�
0.0040.275�
0.011
14.41*�
0.71
PF
0.434�
0.020
0.070�
0.005
0.097�
0.009
0.106**�
0.002
0.164�
0.009
0.297�
0.017
0.329�
0.014
0.101�
0.009
0.260�
0.015
17.07*�
1.05
Han/W
istar
Ctr
0.486�
0.025
0.050�
0.003
0.084�
0.008
0.066�
0.001
0.271�
0.011
0.290�
0.008
0.298�
0.009
0.080�
0.005
0.203�
0.006
12.99�
0.91
50
0.546�
0.023
0.030�
0.014
0.088�
0.066
0.087�
0.005
0.242�
0.029
0.396*�
0.013
0.364�
0.016
0.076�
0.006
0.167�
0.004
8.32�
0.70
500
0.365�
0.012
0.056�
0.000
0.065�
0.004
0.071�
0.022
0.241�
0.025
0.404*�
0.006
0.376�
0.009
0.075�
0.003
0.177�
0.004
9.84�
0.12
1000
0.567�
0.020
0.041�
0.008
0.109�
0.007
0.083�
0.002
0.215�
0.015
0.342�
0.007
0.401*�
0.012
0.052�
0.005
0.158�
0.005
8.85�
0.54
5000
0.500�
0.028
0.067�
0.015
0.103�
0.005
0.093�
0.002
0.227�
0.006
0.372*�
0.009
0.458*�
0.012
0.066�
0.008
0.182�
0.005
8.08�
0.66
9600
0.494�
0.019
0.025�
0.009
0.107�
0.012
0.073�
0.003
0.296�
0.026
0.387*�
0.014
0.403*�
0.010
0.059�
0.007
0.164�
0.004
7.37�
0.31
*P
<0.05,**P
<0.01,***P
<0.001vsadlibitum
-fed
controls
334

of circulating amino acids, observed primarily inTCDD-treated L-E rats, are likely to re¯ect increasedmobilization of amino acids and their decreased me-tabolism in liver. Reduction of liver PEPCK activitycannot account for the sensitivity di�erence of thesetwo strains of rats in terms of mortality. Nevertheless,the response of both strains of TCDD-treated ratsregarding gluconeogenesis was di�erent from that seenin PFCs suggesting that impairment of this pathwaycontributes to the development of the wastingsyndrome.
Acknowledgements We are grateful for the technical assistance ofCindy Palmer, Margitta Lebofsky, Arja Tamminen and MinnaVoutilainen. This study was supported by the Academy of Finland,Research Council for Environmental Sciences (grant no. 5410/4011/89), European Commission (grant ENV4-CT96-0336), andthe GSF-Forschungszentrum fuÈ r Umwelt und Gesundheit, Neu-herberg, Germany. Dr Bernhard Stahl was supported by a fel-lowship of the Deutsche Forschungsgemeinschaft (Sta 300/3-3).
References
Christian BJ, Inhorn SL, Peterson RE (1986a) Relationship of thewasting syndrome to lethality in rats treated with 2,3,7,8-tet-rachlorodibenzo-p-dioxin. Toxicol Appl Pharmacol 82: 239±255
Christian BJ, Menahan LA, Peterson RE (1986b) Intermediarymetabolism of the mature rat following 2,3,7,8-tetrachlorodi-benzo-p-dioxin treatment. Toxicol Appl Pharmacol 83: 360±378
Dutton DR, Parkinson A (1989) Reduction of 7-alkoxyresoru®nsby NADPH-cytochrome P-450 reductase and its di�erentiale�ects on their O-dealkylation by rat liver microsomal cyto-chrome P-450. Arch Biochem Biophys 268: 605±616
Fan F, Rozman K (1994) Relationship between acute toxicity of2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) and disturbanceof intermediary metabolism in the Long-Evans rat. Arch Tox-icol 69: 73±78
Fan F, Rozman KK (1995) Short- and long-term biochemical ef-fects of TCDD in female Long-Evans rats. Toxicol Lett 75:209±216
Gasiewicz TA, Holscher MA, Neal RA (1980) The e�ect of totalparenteral nutrition on the toxicity of 2,3,7,8-tetrachlorodi-benzo-p-dioxin in the rat. Toxicol Appl Pharmacol 54: 469±488
Gorski JR, Rozman K (1987) Dose-response and time course ofhypothyroxinemia and hypoinsulinemia and characterization ofinsulin hypersensitivity in 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD)-treated rats. Toxicology 44: 297±307
Gorski JR, Muzi G, Weber LWD, Pereira DW, Arceo RJ,Iatropoulos MJ, Rozman K (1988a) Some endocrine andmorphological aspects of the acute toxicity of 2,3,7,8-tetra-chlorodibenzo-p-dioxin (TCDD). Toxicol Pathol 16: 313±320
Gorski JR, Muzi G, Weber LWD, Pereira DW, Iatropoulos MJ,Rozman K (1988b) Elevated plasma corticosterone levels andhistopathology of the adrenals and the thymuses in 2,3,7,8-tetrachlorodibenzo-p-dioxin -treated rats. Toxicology 53: 19±32
Gorski JR, Weber LWD, Rozman K (1990) Reduced gluconeo-genesis in 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-treatedrats. Arch Toxicol 64: 66±71
Hultman E (1967) Muscle glycogen in man determined in needlebiopsy specimens. Methods and normal values. Scand J ClinLab Invest 19: 209±217
Lentnek M, Gri�th OW, Rifkind AB (1991) 2,3,7,8-Tetrachloro-dibenzo-p-dioxin increases reliance on fats as a fuel source in-dependently of diet: evidence that diminished carbohydratesupply contributes to dioxin lethality. Biochem Biophys ResCommun 174: 1267±1271
Max SR, Silbergeld EK (1987) Skeletal muscle glucocorticoid re-ceptor and glutamine synthetase activity in the wasting syn-drome in rats treated with 2,3,7,8-tetrachlorodibenzo-p-dioxin.Toxicol Appl Pharmacol 87: 523±527
McConnell EE, Moore JA, Haseman JK, Harris MW (1978) Thecomparative toxicity of chlorinated dibenzo-p-dioxins in miceand guinea pigs. Toxicol Appl Pharmacol 44: 335±356
Muzi G, Gorski JR, Rozman K (1989) Mode of metabolism isaltered in 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-treatedrats. Toxicol Lett 47: 77±86
Olson JR, Holscher MA, Neal RA (1980) Toxicity of 2,3,7,8-tet-rachlorodibenzo-p-dioxin in the Syrian golden hamster. ToxicolAppl Pharmacol 55: 67±78
Pohjanvirta R, Tuomisto J (1994) Short-term toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin in laboratory animals: e�ects,mechanisms and animal models. Pharmacol Rev 46: 483±549
Pohjanvirta R, Tuomisto J, Vartiainen T, Rozman K (1987) Han/Wistar rats are exceptionally resistant to TCDD I. PharmacolToxicol 60: 145±150
Pohjanvirta R, Juvonen R, KaÈ renlampi S, Raunio H, Tuomisto J(1988) Hepatic Ah-receptor levels and the e�ect of 2,3,7,8-tet-rachlorodibenzo-p-dioxin (TCDD) on hepatic microsomalmonooxygenase activities in a TCDD-susceptible and -resistantrat strain. Toxicol Appl Pharmacol 92: 131±140
Pohjanvirta R, Kulju T, Morselt AFW, Tuominen R, Juvonen R,Rozman K, MaÈ nnistoÈ P, Collan Y, Sainio E-L, Tuomisto J(1989) Target tissue morphology and serum biochemistry fol-lowing 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) exposurein a TCDD-susceptible and a TCDD-resistant rat strain. Fun-dam Appl Toxicol 12: 698±712
Pohjanvirta R, Sankari S, Kulju T, Naukkarinen A, Ylinen M,Tuomisto J (1990) Studies on the role of lipid peroxidation inthe acute toxicity of TCDD in rats. Pharmacol Toxicol 66: 399±408
Pohjanvirta R, Unkila M, Tuomisto J (1993) Comparative acutelethality of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD),1,2,3,7,8-pentachlorodibenzo-p-dioxin and 1,2,3,4,7,8-hex-achlorodibenzo-p-dioxin in the most TCDD-susceptible and themost TCDD-resistant rat strain. Pharmacol Toxicol 73: 52±56
Pohjanvirta R, Unkila M, Linde n J, Tuomisto JT, Tuomisto J(1995) Toxic equivalency factors do not predict the acute tox-icities of dioxins in rats. Eur J Pharmacol 293: 341±353
Pohjanvirta R, Wong JMY, Li W, Harper PA, Tuomisto J, OkeyAB (1998) Point mutation in intron sequence causes alteredC-terminal structure in the AH receptor of the most TCDD-resistant rat strain. Mol Pharmacol 54: 86±93
Pohjanvirta R, Viluksela M, Tuomisto JT, Unkila M, KarasinskaJ, Franc M-A, Holowenko M, Giannone JV, Harper PA,Tuomisto J, Okey AB (1999) Physicochemical di�erences in theAH receptors of the most TCDD-susceptible and the mostTCDD-resistant rat strains. Toxicol Appl Pharmacol 155:82±95
Potter CL, Sipes GL, Haddock RD (1983) Hypothyroxinemia andhypothermia in rats in response to 2,3,7,8-tetrachlorodibenzo-p-dioxin administration. Toxicol Appl Pharmacol 69: 89±95
Schwetz BA, Norris JM, Sparschu GL, Rowe VK, Gehring PJ,Emerson JL, Gebrig CG (1973) Toxicology of chlorinateddibenzo-p-dioxins. Environ Health Perspect 5: 87±99
Stahl BU (1995) 2,3,7,8-Tetrachlorodibenzo-p-dioxin blocks thephysiological regulation of phosphoenolpyruvate carboxyki-nase in primary rat hepatocytes. Toxicology 103: 45±52
Stahl BU, Beer DG, Weber LWD, Rozman K (1993) Reduction ofhepatic phosphoenolpyruvate carboxykinase (PEPCK) activityby 2,3,7,8-tetrachlordibenzo-p-dioxin (TCDD) is due to de-creased mRNA levels. Toxicology 79: 81±95
Stryer L (1996) Biochemistry, 4th edn. Freeman, New York, pp629±652
Tilghman SM, Hanson RW, Ballard FJ (1976) Hormonal regula-tion of phosphoenolpyruvate carboxykinase (GTP) in mam-malian tissues. In: Hanson RW, Mehlman MA (eds)Gluconeogenesis: its regulation in mammalian species. Wiley,New York, pp 49±91
335

Tuomisto J, Pohjanvirta R, MacDonald E, Tuomisto L (1990)Changes in rat brain monoamines, monoamine metabolites andhistamine after a single administration of 2,3,7,8-tetrachloro-dibenzo-p-dioxin (TCDD). Pharmacol Toxicol 67: 260±265
Tuomisto JT, Viluksela M, Pohjanvirta R, Tuomisto J (1999) TheAh receptor and a novel gene determine acute toxic responses toTCDD: segregation of the resistant alleles to di�erent rat lines.Toxicol Appl Pharmacol 155: 71±81
Unkila M, Pohjanvirta R, Tuomisto J (1992) Acute toxicity ofper¯uorodecanoic acid and cobalt protoporphyrin in a TCDD-sensitive and a TCDD-resistant rat strain. Chemosphere 25:1233±1238
Unkila M, Pohjanvirta R, Honkakoski P, ToÈ rroÈ nen R, Tuomisto J(1993a) 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) inducedethoxyresoru®n-O-deethylase (EROD) and aldehyde dehydro-genase (ALDH3) activities in the brain and liver. A comparisonbetween the most TCDD-susceptible and the most TCDD-re-sistant rat strain. Biochem Pharmacol 46: 651±659
Unkila M, Pohjanvirta R, MacDonald E, Tuomisto J (1993b)Di�erential e�ect of TCDD on brain serotonin metabolism in aTCDD-susceptible and a TCDD-resistant rat strain. Chemo-sphere 27: 401±406
Unkila M, Pohjanvirta R, MacDonald E, Tuomisto JT, Tuomisto J(1994) Dose response and time course of alterations in tryp-tophan metabolism by 2,3,7,8-tetrachlorodibenzo-p-dioxin(TCDD) in the most TCDD-susceptible and the most TCDD-resistant rat strain: relationship with TCDD lethality. ToxicolAppl Pharmacol 128: 280±292
Unkila M, Ruotsalainen M, Pohjanvirta R, Viluksela M, Mac-Donald E, Tuomisto JT, Rozman K, Tuomisto J (1995) E�ectof 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on tryptophanand glucose homeostasis in the most TCDD-susceptible and themost TCDD-resistant species, guinea pigs and hamsters. ArchToxicol 69: 677±683
Viluksela M, Stahl BU, Rozman KK (1995a) Lack of correlationbetween Ah receptor mediated induction of cytochrome P450activity and decrease in phosphoenolpyruvate carboxykinase
(PEPCK) activity in genetically altered mouse hepatoma celllines after TCDD treatment. Abstract XIII, ScandinavianWorkshop on In Vitro Toxicology, September 21±24, Tampere,Finland
Viluksela M, Stahl BU, Rozman KK (1995b) Tissue-speci®c e�ectsof 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on the activityof phosphoenolpyruvate carboxykinase (PEPCK) in rats.Toxicol Appl Pharmacol 135: 308±315
Weber G, Luzi P, Resi L, Tandanelli P (1983) Natural history ofTCDD-induced liver lesions in rats as observed by transmissionelectron microscopy during a 32-week period after a single in-traperitoneal injection. J Toxicol Environ Health 12: 533±540
Weber LWD, Lebofsky M, Greim H, Rozman K (1991a) Keyenzymes of gluconeogenesis are dose-dependently reduced in2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-treated rats. ArchToxicol 65: 119±123
Weber LWD, Lebofsky M, Stahl BU, Gorski JR, Muzi G, RozmanK (1991b) Reduced activities of key enzymes of gluconeogenesisas possible cause of acute toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in rats. Toxicology 66: 133±144
Weber LWD, Lebofsky M, Stahl BU, Kettrup A, Rozman K(1992) Comparative toxicity of four chlorinated dibenzo-p-di-oxins (CDDs) and their mixture. Part III: Structure-activityrelationships with increased plasma tryptophan levels, but norelationship to hepatic ethoxyresoru®n O-deethylase activity.Arch Toxicol 66: 484±488
Weber LWD, Lebofsky M, Stahl BU, Smith S, Rozman KK (1995)Correlation between toxicity and e�ects on intermediary me-tabolism in 2,3,7,8-tetrachlorodibenzo-p-dioxin-treated maleC57BL/6 J and DBA/2 J mice. Toxicol Appl Pharmacol 131:155±162
Wimmer M (1988) A bioluminescent assay for the determination ofphosphoenolpyruvate carboxykinase activity in nanogram-sizedtissue samples. Anal Biochem 170: 376±381
Zinkl JG, Vos JG, Moore JA, Gupta BN (1973) Hematologic andclinical chemistry e�ects of 2,3,7,8-tetrachlorodibenzo-p-dioxinin laboratory animals. Environ Health Perspect 5: 111±118
336
Copyright © 2022 FDOKUMEN














