
Physical properties of ordered mesoporous SBA-15 silica as immunological adjuvant
Upload
independentCategory
view
3download
0
Assessing the immunopotency of Toll-like receptor agonists in anin vitro tissue-engineered immunological model
Yifan Ma,1,2 Louis Poisson,1
Guzman Sanchez-Schmitz,1 Santosh
Pawar,1 Chunfeng Qu,3 Gwendalyn
J. Randolph,3 William L. Warren,1
Eric M. Mishkin1 and Russell G.
Higbee1
1VaxDesign Corporation, Orlando, FL, USA,2Shenzhen Institutes of Advanced Technology,
Chinese Academy of Sciences, Shenzhen,
China, and 3Department of Gene and Cell
Medicine, Icahn Research Institute, Mount
Sinai School of Medicine, New York, NY, USA
doi:10.1111/j.1365-2567.2009.03237.x
Received 22 September 2009; revised 9
December 2009; accepted 15 December 2009.
Correspondence: R. G. Higbee, VaxDesign
Corporation, 12612 Challenger Parkway,
Suite 365, Orlando, FL 32826, USA.
Email: [email protected]
Senior author: Russell G. Higbee
Summary
The in vitro Peripheral Tissue Equivalent (PTE) module is a three-dimen-
sional tissue-engineered endothelial cell/collagen matrix culture system,
which has been reported to reproduce in vivo physiological conditions
and which generates dendritic cells (DC) autonomously. In the present
study, we used the PTE module to investigate the immunopotency of
Toll-like receptor (TLR) agonists, including polyinosine-polycytidylic acid,
Gardiquimod, CpG 2006 and lipopolysaccharide. Application of TLR
agonists in the PTE module induced a wide range of cytokines, including
interleukins 1a/b, 6, 8 and 10 and tumour necrosis factor-a. Compared
with traditional peripheral blood mononuclear cell (PBMC) cultures, the
PTE module produced twofold to 100-fold higher levels of cytokine secre-
tion, indicating that it can be a highly sensitive assay system. This
increased sensitivity is the result of the natural synergy between the leuco-
cytes and the endothelium. Furthermore, the application of TLR agonists,
such as lipopolysaccharide and Gardiquimod, to the PTE module
enhanced DC differentiation and promoted DC maturation, as indicated
by up-regulated expression of CD83, CD86 and CCR7(CD197). In addi-
tion, functional assays indicated PTE-derived DC treated with Gardiqui-
mod, a TLR-7 agonist, significantly augmented anti-tetanus toxoid
antibody production. Interestingly, replacing PBMC with purified myeloid
cells (CD33+) significantly reduced the responsiveness of the PTE module
to TLR stimulation. The reduced sensitivity was partly the result of the
removal of plasmacytoid DC that participated in the response to TLR
stimulation and sensitization of the PTE module. Overall, the in vitro
PTE module clearly demonstrated the effects of TLR agonists on DC gen-
eration, maturation and antigen-presenting capacity, and may serve as a
sensitive and predictive test bed for the evaluation of adjuvant candidates.
Keywords: cytokine; dendritic cell; endothelial cells; human; in vitro tissue
engineered immunological model; three-dimensional; Toll-like receptor
Abbreviations: APC, antigen-presenting cells; DC, dendritic cell; ELISPOT, enzyme-linked immunosorbent spot-forming cellassay; GM-CSF, granulocyte–macrophage colony-stimulating factor; HBSS, Hanks’ buffered salt solution; HUVEC, humanumbilical vein endothelial cell; iDC, immature DC; IFN, interferon; IgG, immunoglobulin G; IL-4, interleukin-4; IP-10,interferon inducible protein-10; LC, Langerhans cells; LPS, lipopolysaccharide; mDC, mature DC; ODN, oligonucleotides;PBMC, peripheral blood mononuclear cell; pDC, plasmacytoid DC; PE, phycoerythrin; Poly I:C, polyinosine-polycytidylic acid;PTE module, in vitro peripheral tissue equivalent module; PTE-DC, PTE-derived DC; RANTES, regulated on activation, normalT-cell expressed and secreted; RT, reverse transmigrated; TLR, Toll-like receptor; TNF, tumour necrosis factor; TT, tetanustoxoid.
374 � 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387
I M M U N O L O G Y O R I G I N A L A R T I C L E

Introduction
Dendritic cells (DC) are the most potent antigen-present-
ing cells (APC), and they play an essential role in both
innate and adaptive immunity. They normally develop
from circulating bone-marrow-derived DC precursors that
distribute into the peripheral tissues and give rise to
immature DC (iDC).1 The tissue-residing iDC capture
antigens from the local environment and release cyto-
kines/chemokines, thereby participating in innate immu-
nity. Moreover, antigen capture also triggers DC
maturation and migration into draining lymph nodes. In
the T-cell region of lymph nodes, mature DC (mDC)
present antigens to naive T cells via major histocompati-
bility complex molecules, triggering the adaptive immune
response.2 Hence, DC are an important link between
innate and adaptive immunity.
In vitro DC production is an important strategy for
generating large numbers of DC. To date, the most com-
monly used method to generate human DC is to culture
blood monocytes with granulocyte–macrophage colony-
stimulating factor (GM-CSF) and interleukin-4 (IL-4) for
5–7 days.3 Although this method can produce a large
population of DC, it remains questionable whether this
method faithfully recapitulates DC development in vivo.4
Besides cytokine-derived DC, Randolph et al.5–8 have
reported a tissue-engineered in vitro immunological
model that allows for autonomous generation of DC. We
termed this system the in vitro ‘Peripheral Tissue Equiva-
lent’ (PTE) module. The PTE module is a three-dimen-
sional culture system consisting of a confluent monolayer
of unstimulated endothelial cells grown on top of an
endotoxin-free collagenous matrix (see Fig. 1). When
peripheral blood mononuclear cells (PBMC) are applied
onto the endothelial monolayer, monocytes preferentially
migrate across the endothelium into the sub-endothelial
collagen. This process probably mimics the constitutive
entry of monocytes into tissues as observed in vivo.9 After
48 hr in culture, approximately half of the migrated
monocytes differentiate into DC with varying degrees of
maturity, and migrate across the intact endothelium in
the abluminal-to-luminal direction (reverse transmigra-
tion),5 which may mimic the in vivo DC migration from
peripheral tissue into the local lymphatics.1 In the
unstimulated PTE module, the percentage of mature DC
in the reverse-transmigrated (RT) cell fraction is < 10%,
with the majority of RT cells resembling immature DC or
monocytes. Incorporation of various stimuli, such as lipo-
polysaccharide (LPS), influenza virus or zymosan, signifi-
cantly increases the percentage of mature DC in the RT
cell fraction.6 Compared with conventional cytokine-
derived DC, PTE-derived DC (PTE-DC) differentiate
more rapidly and do not require application of exogenous
cytokines. Moreover, the composition of PTE-DC is more
heterogeneous than that of cytokine-cultured DC, and
may more closely resemble the composition of the DC
populations developed in vivo.5,10 In addition, the PTE
module contains endothelial cells that can sense ‘danger
signals’ in peripheral tissues and contribute to immune
responses.11,12 Indeed, the presence of endothelial cells
has a critical role in the generation of DC in the system,5
which probably increases the sensitivity of the PTE mod-
ule in response to various stimuli.
The Toll-like receptor (TLR) family is a group of pat-
tern-recognition receptors that play a crucial role in both
innate and adaptive immunity. TLRs can recognize con-
served microbial structures or products of microbial
metabolism called ‘pathogen-associated molecular pat-
tern’, which consequently triggers innate immunity. The
TLR signalling also promotes the activation and matura-
tion of APCs, thereby facilitating adaptive immunity. In
addition, cytokines and chemokines elicited by TLR stim-
ulation further regulate downstream T-cell and B-cell
responses.13 Currently, several TLR agonists are being
evaluated as potential adjuvants for vaccine development
against infectious diseases and cancer. For example,
polyinosine-polycytidylic acid (Poly I:C), a TLR3 agonist,
has been shown to be a potent adjuvant to enhance vac-
cine-induced protective immune responses.14 Agonists of
TLR7/8, such as Imiquimod and Resiquimod, have been
used to treat skin neoplasms and viral infections in
humans.15 CpG oligonucleotides (ODN), TLR9 agonists,
have been reported to significantly enhance the antibody
responses induced by hepatitis B and anthrax vaccines in
human clinical trials.16,17 Previous studies have demon-
strated that conventional and plasmacytoid DC (pDC) are
the major cell types mediating the adjuvant effect of TLR
agonists.18 Stimulation by TLR can promote the differen-
tiation and maturation of freshly isolated DC or cyto-
kine-cultured DC, triggering the release of cytokines.19–21
Moreover, there is evidence that TLR stimulation, such as
stimulation of TLR8 by single-stranded RNA, triggers
PTE-DC maturation.7 However, a generally broader sur-
vey as to how TLR stimulation will affect the phenotype
and function of PTE-DC has not been performed, nor is
PBMC
Remain
Monocyte/DC
48 hr
Monocytes
M φ
(a) (b)
Figure 1. The scheme of the in vitro peripheral tissue equivalent
(PTE) module. The PTE module consists of a quiescent monolayer
of human umbilical vein endothelial cells (HUVEC) grown on a col-
lagen matrix. When peripheral blood mononuclear cells (PBMC) are
applied onto HUVEC, monocytes migrate into the subendothelial
collagen and form a PTE module (a). After an additional 48 hr of
culture, about half of the monocytes reverse transmigrate across the
endothelium and develop into dendritic cells (b).
� 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387 375
Assessing the immunopotency of TLR agonists in a 3D immunological model

it known how the response to TLR agonists in the PTE
module compares with traditional PBMC culture and
cytokine-cultured DC. In the present study, the immuno-
potency of TLR agonists, such as Poly I:C (TLR3), LPS
(TLR4), Gardiquimod (TLR7/8) or CpG 2006 (TLR9),
was tested in the PTE module. These TLR agonists evoked
cytokine and chemokine production, and promoted DC
generation and maturation in the PTE module. When
compared with traditional PBMC culture methods, the
PTE module was more sensitive and so was a promising
predictor of immune response. These studies support the
use of the PTE module as a reliable immunological test
bed for adjuvant and vaccine screening and development.
Materials and methods
Human PBMC preparation
Apheresis blood products were obtained from Florida’s
Blood Centers (Orlando, FL). The study was reviewed
and approved by Chesapeake Research Review Inc.
(Columbia, MD). All donors were in good health and all
blood products were negative for blood-borne pathogens
as detected by standard blood bank assays. The PBMC
were enriched by Ficoll density gradient separation
according to standard laboratory procedures. After wash-
ing, PBMC were either resuspended in culture media or
cryopreserved in dimethylsulphoxide-containing freezing
media for extended storage in liquid nitrogen vapour
phase for future assays.
In vitro PTE module preparation
The PTE module was fabricated as described previously,10
as shown in Fig. 1. Briefly, second-passage human umbili-
cal vein endothelial cells (HUVEC; Lonza, Walkersville,
MD) were cultured on a polymerized type I collagen
matrix (Inamed Biomaterials, Fremont, CA) and main-
tained in M199 media (Lonza) supplemented with 20%
fetal bovine serum until confluent. The PBMC or purified
monocytes were suspended in M199 media containing
0�1% (weight/volume) bovine serum albumin, and
applied onto the HUVEC monolayer at 500 000 cells/well
and 200 000 cells/well, respectively. In some experiments,
pDC were depleted from whole PBMC by using CD304
(BDCA-4/Neuropilin-1) MicroBeads (Miltenyi Biotec
Inc., Auburn, CA). After a 90-min incubation, non-
migrated cells were removed, and PTE modules were
incubated in culture medium (M199 medium with 20%
heat-inactivated human autologous serum) with or with-
out the following TLR agonists: 5 lg/ml CpG 2006,
25 lg/ml Poly I:C, 5 lg/ml Gardiquimod (all from Invivo-
Gen, San Diego, CA), or 10 ng/ml LPS (Sigma, St Louis,
MO). The PTE culture supernatants were collected at
24 hr or 48 hr for cytokine analysis, and the RT cells were
carefully harvested at 48 hr for fluorescence-activated cell
sorting analysis.
PBMC culture
The PBMC were resuspended at 2�5 · 106 cells/ml in
RPMI-1640 supplemented with 10% heat-inactivated
autologous human serum, and stimulated with or without
the following TLR agonists: 5 lg/ml CpG 2006, 25 lg/ml
Poly I:C, 5 lg/ml Gardiquimod (all from InvivoGen), or
10 ng/ml LPS (Sigma). PTE culture supernatants were
collected at 24 hr for cytokine analysis.
Cell isolation
The T and B cells were negatively selected using pan-
T-cell isolation kit II or B-cell isolation kit II from Miltenyi
Biotec, Inc. For isolation of CD33+ myeloid cells, PBMC
were stained with phycoerythrin (PE) -labelled anti-
human CD33 antibody (BD Biosciences, San Jose, CA).
Labelled cell populations were then enriched to > 99%
purity by selection over LS separation columns after label-
ling with anti-PE microbeads (Miltenyi Biotec Inc). For
pDC reconstitution experiments, the CD33/pDC fraction
was positively selected using CD33 and BDCA-4 micro-
beads, and the percentage of pDC in the CD33/pDC-
enriched fraction was 1�4–2%.
Cytokine-cultured DC preparation and stimulation
CD14+ monocytes were positively selected using CD14+
microbeads (Miltenyi Biotec), and cultured with GM-CSF
(100 ng/ml) and IL-4 (25 ng/ml) for 5 days to generate
iDC. The iDC were then stimulated with equivalent doses
of TLR agonists as previously described. After 48 hr cul-
ture, cells were collected for DC phenotypic analysis.
Flow cytometry
The phenotype of DC was determined using seven-colour
staining. The RT cells or iDC were first incubated with
mouse immunoglobulin G1 (IgG1; 1 lg/sample; Sigma)
at room temperature to block Fc receptor along with the
vital dye Annexin V (2�5 ll/sample; Invitrogen Corpora-
tion, Carlsbad, CA) to identify apoptotic cells. After
15 min, the cells were incubated for 40 min on ice with
the cocktail of fluorochrome-labelled monoclonal anti-
bodies. The antibody cocktail contained non-myeloid
lineage markers, allophycocyanin-conjugated CD3, CD19
and CD56, and the relevant DC surface markers, allo-
phycocyanin-Cychrome 7-conjugated CD14, peridinin
chlorophyll protein-Cychrome 5.5-conjugated human leu-
cocyte antigen (HLA) -DR, CCR7-PE-Cy7, fluorescein
isothiocyanate-conjugated CD83 and CD86-PE (BD Bio-
sciences). After incubation, the cells were washed twice
376 � 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387
Y. Ma et al.

with Hanks’ buffered salt solution (HBSS, +Ca/+Mg) fol-
lowed by fixation in 2% (volume/volume) paraformalde-
hyde, also in HBSS. Viable cellular events were accrued
on a BD LSRII flow cytometer equipped with BD FACS-
DIVA software (BD Biosciences). Data analysis was with
FLOWJO software (Tree Star Inc., Ashland, OR).
Cytokine and chemokine analysis
Production of cytokines and chemokines was analysed
using a Beadlyte human 22-plex multi-cytokine detection
system (Millipore, Billerica, MA). Briefly, 50 ll appropri-
ately diluted cytokine standard or culture supernatants
was incubated with 25 ll of multi-cytokine beads in a 96-
well filter plate at room temperature for 2 hr. The beads
were then washed and incubated with 25 ll of multi-
cytokine biotin for 90 min, followed by mixing with 25 ll
of diluted Beadlyte strepavidin-PE. After 30 min, liquid
was removed, and the beads were resuspended in 125 ll
of assay buffer and analysed using the Bio-Plex 200 sys-
tem (Bio-Rad, Hercules, CA). Cytokine and chemokine
concentrations were calculated based on relevant standard
curves using the BIO-PLEX manager software (Bio-Rad).
Antigen presentation
The PTE modules were prepared as previously described.
After removing the non-migrated cells, the PTE modules
were incubated at 37� with either culture medium alone,
tetanus toxoid (TT, 1 lg/ml; Calbiochem, La Jolla, CA),
Gardiquimod (5 lg/ml), or TT plus Gardiquimod. The
RT cells were carefully collected after 48 hr and co-cul-
tured with 2�5 · 105 autologous T cells and 2�5 · 105
autologous B cells in a 24-well plate at an APC : lympho-
cyte ratio of 1 : 30. The anti-TT IgG response was
evaluated on day 7 using enzyme-linked immunosorbent
spot-forming cell assay (ELISPOT) detection.
Enumeration of anti-TT antibody-secreting cells
Numbers of TT-specific antibody-secreting cells were
assessed by ELISPOT as previously described, with minor
modifications.22 Briefly, a 96-well Multiscreen HA filtration
plate (Millipore Corporation) was coated with 1 lg/ml TT
at 4� overnight, followed by blocking with RPMI-1640 con-
taining 1% bovine serum albumin at 37� for 2 hr. Serially
diluted lymphocyte suspensions were added to the plate
and incubated at 37� for 4–5 hr. After incubation, cells
were discarded and the plate was incubated with 1 lg/ml
goat anti-human IgG-biotin (Jackson ImmunoLab, West
Grove, PA) at 4�, overnight. On the next day, the plate was
washed and incubated with 5 lg/ml horseradish peroxi-
dase-conjugated avidin (Vector Laboratories, Burlingame,
CA) for 1 hr at room temperature. The plate was washed
and incubated with 3-amino-9-ethylcarbazole substrate
solution until spots were well developed. The plate was
rinsed with deionized water to stop the reaction and spots
were counted using the AID ELISPOT reader system (Cell
Technology, Columbia, MD).
Results
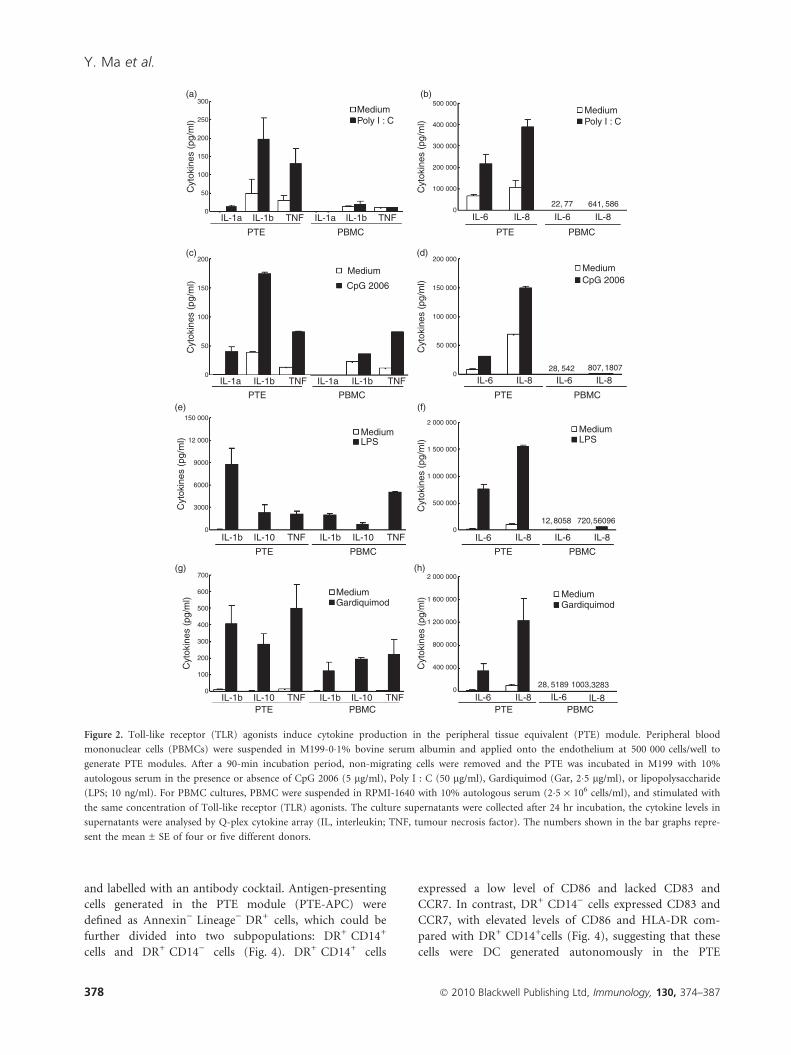
TLR agonists induce more cytokines in the PTEmodule than in PBMC culture
The TLR-induced cytokine production in the PTE mod-
ule was measured first to evaluate the immunopotency of
the TLR agonists. Overall, TLR agonists induced higher
levels of cytokines in the PTE module than in conven-
tional PBMC cultures (Fig. 2). For example, both Poly
I:C and CpG 2006 induced the pro-inflammatory cyto-
kines IL-1a/b in the PTE module, which were not
observed in PBMC cultures (Fig. 2a,c). Poly I:C also trig-
gered tumour necrosis factor-a (TNF-a) production solely
in the PTE culture (Fig. 2a). Moreover, the production of
IL-6 and IL-8 induced by Poly I:C and CpG 2006 treat-
ments in the PTE module was significantly higher than in
PBMC culture (Fig. 2b,d). Although Gardiquimod and
LPS dramatically induced IL-1b, IL-6, IL-8, IL-10 and
TNF-a, both in the PTE module and in the PBMC cul-
ture, the PTE module produced approximately three- to
sixfold more IL-1b and IL-10, and 10- to 50-fold more
IL-6 and IL-8, than PBMC culture (Fig. 2e–h). Conse-
quently, the PTE module was more sensitive than conven-
tional PBMC culture in response to TLR stimulation.
The PTE module contains a confluent HUVEC mono-
layer, so it was important to determine if the higher sen-
sitivity of the PTE module to TLR agonists was
attributable to the presence of HUVEC. There was a basal
production of IL-6 (286 ± 26 pg/ml) and IL-8
(3822 ± 347 pg/ml) in unstimulated HUVEC cultures.
Although treatment with Poly I:C or LPS significantly
increased the production of IL-6 and IL-8 by about two-
fold to fourfold, Gardiquimod and CpG 2006 did not
trigger cytokine production in the HUVEC culture
(Fig. 3). These data suggest that enhanced TLR-induced
cytokine production in the PTE module is attributed to a
synergistic interaction between migrated cells and the
endothelium.
TLR agonists promote differentiation and maturationof DC in the PTE module
Previous studies have shown that TLR stimulation can
induce DC maturation and enhance antigen presenta-
tion.23 The PTE module has been reported as a novel tis-
sue-engineered system to generate DC autonomously, so
we determined if TLR stimulation could also regulate the
phenotype of the PTE-DC. For this purpose, RT cells
were carefully harvested from unstimulated PTE modules
� 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387 377
Assessing the immunopotency of TLR agonists in a 3D immunological model
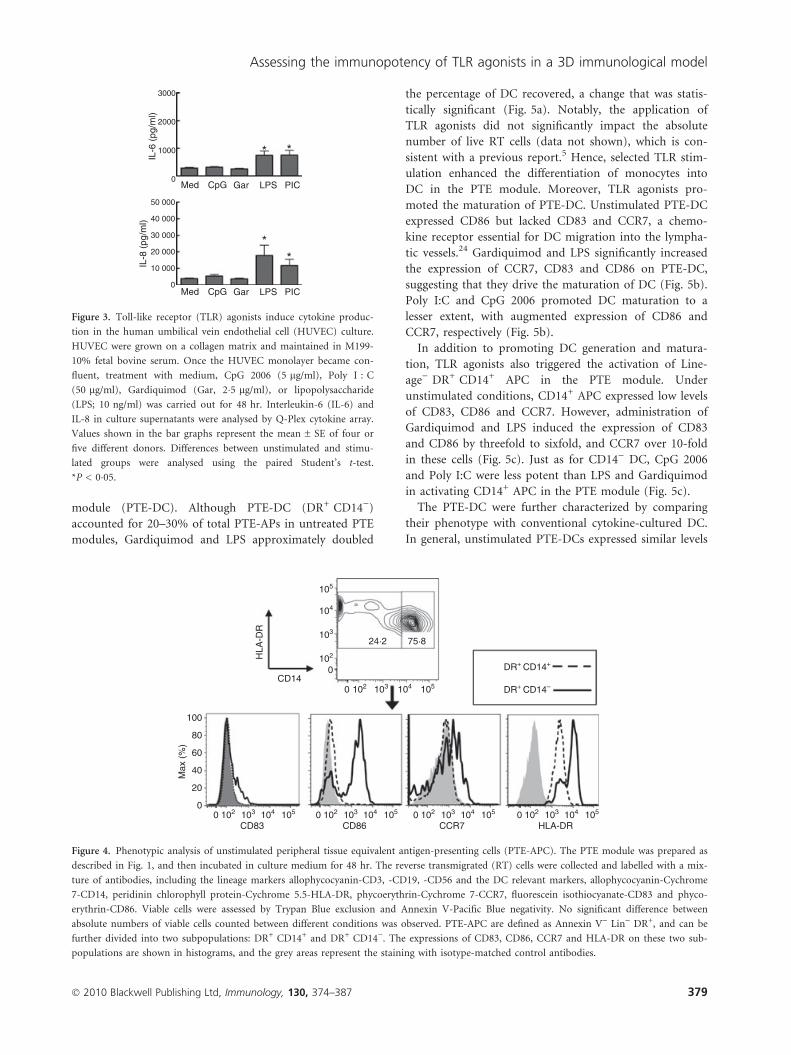
and labelled with an antibody cocktail. Antigen-presenting
cells generated in the PTE module (PTE-APC) were
defined as Annexin) Lineage) DR+ cells, which could be
further divided into two subpopulations: DR+ CD14+
cells and DR+ CD14) cells (Fig. 4). DR+ CD14+ cells
expressed a low level of CD86 and lacked CD83 and
CCR7. In contrast, DR+ CD14) cells expressed CD83 and
CCR7, with elevated levels of CD86 and HLA-DR com-
pared with DR+ CD14+cells (Fig. 4), suggesting that these
cells were DC generated autonomously in the PTE
300
250
200
150
100
Cyt
okin
es (
pg/m
l)
Cyt
okin
es (
pg/m
l)
50
IL-1a IL-1b TNF IL-1a
PBMCPTE PBMCPTE
PBMCPTE
IL-1b IL-6 IL-8 IL-8IL-6
IL-6 IL-8 IL-8IL-6
IL-6 IL-8 IL-8IL-6
PBMCPTEIL-6 IL-8 IL-8IL-6
22,
807, 180754228,
5189 328328, 1003,
12,8058 56096720,
641,77 586
TNF
IL-1a IL-1b TNF IL-1a
PBMCPTE PBMCPTE
IL-1b TNF
IL-10IL-1b TNF IL-10
PBMCPTE
IL-1b TNF
IL-10IL-1b TNF IL-10PBMCPTE
IL-1b TNF
MediumPoly I : C
(a) (b)
(c) (d)
(e) (f)
(g) (h)
MediumPoly I : C
Medium
Medium Medium
CpG 2006
Cyt
okin
es (
pg/m
l)C
ytok
ines
(pg
/ml)
Cyt
okin
es (
pg/m
l)
Cyt
okin
es (
pg/m
l)C
ytok
ines
(pg
/ml)
Cyt
okin
es (
pg/m
l) CpG 2006
MediumGardiquimod
MediumGardiquimod
200 000
500 000
400 000
300 000
200 000
100 000
0
150 000
100 000
50 000
0
2 000 000
2 000 000
1 600 000
1 200 000
800 000
400 000
0
1 000 000
1 500 000
500 000
0
LPSMediumLPS
0
200
150
100
50
0
150 000
12 000
9000
6000
3000
0
700
600
500
400
300
200
100
0
Figure 2. Toll-like receptor (TLR) agonists induce cytokine production in the peripheral tissue equivalent (PTE) module. Peripheral blood
mononuclear cells (PBMCs) were suspended in M199-0�1% bovine serum albumin and applied onto the endothelium at 500 000 cells/well to
generate PTE modules. After a 90-min incubation period, non-migrating cells were removed and the PTE was incubated in M199 with 10%
autologous serum in the presence or absence of CpG 2006 (5 lg/ml), Poly I : C (50 lg/ml), Gardiquimod (Gar, 2�5 lg/ml), or lipopolysaccharide
(LPS; 10 ng/ml). For PBMC cultures, PBMC were suspended in RPMI-1640 with 10% autologous serum (2�5 · 106 cells/ml), and stimulated with
the same concentration of Toll-like receptor (TLR) agonists. The culture supernatants were collected after 24 hr incubation, the cytokine levels in
supernatants were analysed by Q-plex cytokine array (IL, interleukin; TNF, tumour necrosis factor). The numbers shown in the bar graphs repre-
sent the mean ± SE of four or five different donors.
378 � 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387
Y. Ma et al.
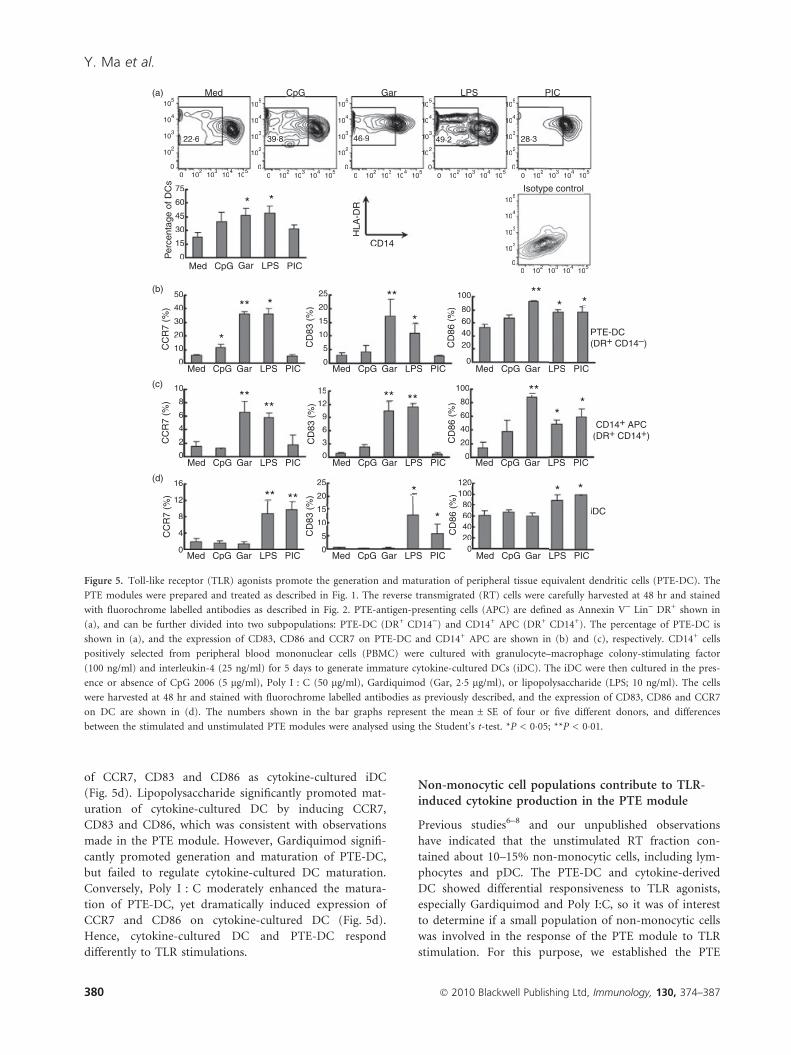
module (PTE-DC). Although PTE-DC (DR+ CD14))
accounted for 20–30% of total PTE-APs in untreated PTE
modules, Gardiquimod and LPS approximately doubled
the percentage of DC recovered, a change that was statis-
tically significant (Fig. 5a). Notably, the application of
TLR agonists did not significantly impact the absolute
number of live RT cells (data not shown), which is con-
sistent with a previous report.5 Hence, selected TLR stim-
ulation enhanced the differentiation of monocytes into
DC in the PTE module. Moreover, TLR agonists pro-
moted the maturation of PTE-DC. Unstimulated PTE-DC
expressed CD86 but lacked CD83 and CCR7, a chemo-
kine receptor essential for DC migration into the lympha-
tic vessels.24 Gardiquimod and LPS significantly increased
the expression of CCR7, CD83 and CD86 on PTE-DC,
suggesting that they drive the maturation of DC (Fig. 5b).
Poly I:C and CpG 2006 promoted DC maturation to a
lesser extent, with augmented expression of CD86 and
CCR7, respectively (Fig. 5b).
In addition to promoting DC generation and matura-
tion, TLR agonists also triggered the activation of Line-
age) DR+ CD14+ APC in the PTE module. Under
unstimulated conditions, CD14+ APC expressed low levels
of CD83, CD86 and CCR7. However, administration of
Gardiquimod and LPS induced the expression of CD83
and CD86 by threefold to sixfold, and CCR7 over 10-fold
in these cells (Fig. 5c). Just as for CD14) DC, CpG 2006
and Poly I:C were less potent than LPS and Gardiquimod
in activating CD14+ APC in the PTE module (Fig. 5c).
The PTE-DC were further characterized by comparing
their phenotype with conventional cytokine-cultured DC.
In general, unstimulated PTE-DCs expressed similar levels
3000
50 000
40 000
30 000
20 000
10 000
0
2000
1000
0Med
IL-6
(pg
/ml)
IL-8
(pg
/ml)
CpG Gar LPS PIC
PICMed CpG Gar LPS
*
**
*
Figure 3. Toll-like receptor (TLR) agonists induce cytokine produc-
tion in the human umbilical vein endothelial cell (HUVEC) culture.
HUVEC were grown on a collagen matrix and maintained in M199-
10% fetal bovine serum. Once the HUVEC monolayer became con-
fluent, treatment with medium, CpG 2006 (5 lg/ml), Poly I : C
(50 lg/ml), Gardiquimod (Gar, 2�5 lg/ml), or lipopolysaccharide
(LPS; 10 ng/ml) was carried out for 48 hr. Interleukin-6 (IL-6) and
IL-8 in culture supernatants were analysed by Q-Plex cytokine array.
Values shown in the bar graphs represent the mean ± SE of four or
five different donors. Differences between unstimulated and stimu-
lated groups were analysed using the paired Student’s t-test.
*P < 0�05.
CD14
HLA
-DR
00
CD83 CD86 CCR7 HLA-DR102 103 104 105
0
0
102
102
24·2 75·8
DR+ CD14+
DR+ CD14–103
103
104
104
105
105
0 102 103 104 105 0 102 103 104 105 0 102 103 104 105
20
40
60
80
100
Max
(%
)
Figure 4. Phenotypic analysis of unstimulated peripheral tissue equivalent antigen-presenting cells (PTE-APC). The PTE module was prepared as
described in Fig. 1, and then incubated in culture medium for 48 hr. The reverse transmigrated (RT) cells were collected and labelled with a mix-
ture of antibodies, including the lineage markers allophycocyanin-CD3, -CD19, -CD56 and the DC relevant markers, allophycocyanin-Cychrome
7-CD14, peridinin chlorophyll protein-Cychrome 5.5-HLA-DR, phycoerythrin-Cychrome 7-CCR7, fluorescein isothiocyanate-CD83 and phyco-
erythrin-CD86. Viable cells were assessed by Trypan Blue exclusion and Annexin V-Pacific Blue negativity. No significant difference between
absolute numbers of viable cells counted between different conditions was observed. PTE-APC are defined as Annexin V) Lin) DR+, and can be
further divided into two subpopulations: DR+ CD14+ and DR+ CD14). The expressions of CD83, CD86, CCR7 and HLA-DR on these two sub-
populations are shown in histograms, and the grey areas represent the staining with isotype-matched control antibodies.
� 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387 379
Assessing the immunopotency of TLR agonists in a 3D immunological model
of CCR7, CD83 and CD86 as cytokine-cultured iDC
(Fig. 5d). Lipopolysaccharide significantly promoted mat-
uration of cytokine-cultured DC by inducing CCR7,
CD83 and CD86, which was consistent with observations
made in the PTE module. However, Gardiquimod signifi-
cantly promoted generation and maturation of PTE-DC,
but failed to regulate cytokine-cultured DC maturation.
Conversely, Poly I : C moderately enhanced the matura-
tion of PTE-DC, yet dramatically induced expression of
CCR7 and CD86 on cytokine-cultured DC (Fig. 5d).
Hence, cytokine-cultured DC and PTE-DC respond
differently to TLR stimulations.
Non-monocytic cell populations contribute to TLR-induced cytokine production in the PTE module
Previous studies6–8 and our unpublished observations
have indicated that the unstimulated RT fraction con-
tained about 10–15% non-monocytic cells, including lym-
phocytes and pDC. The PTE-DC and cytokine-derived
DC showed differential responsiveness to TLR agonists,
especially Gardiquimod and Poly I:C, so it was of interest
to determine if a small population of non-monocytic cells
was involved in the response of the PTE module to TLR
stimulation. For this purpose, we established the PTE
Med (a)
(b)
(c)
(d)
22·6
75
50 25 100
80 60 40 20
0
100
80
60
40
20
0
100 120
80 60 40 20
0
20
15
10
5
0
25
15
12
9
6
3
0
20
15
10
5
0
40
30
20
10
0
10
16
12
8
4
0
8
6
4
2
0
60
45
30
15
0
39·8 46·9 49·2 28·3
105
104
104
105
103
103
102
102
0 0
105
104
104
105
103
103
102
102
0 0
105
104
104
105
103
103
102
102
0 0
105
104
104
105
103
103
102
102
0 0
105
104
104
105
103
103
102
102
0 0
105
104
104
105
103
103
102
102
0 0
CpG Gar LPS PIC
Isotype control
CD14
Med
Per
cent
age
of D
Cs
CC
R7
(%)
CD
83 (
%)
CD
83 (
%)
CD
83 (
%)
CD
86 (
%)
CD
86 (
%)
CD
86 (
%)
CC
R7
(%)
CC
R7
(%)
CpG Gar
*
*
* *
*
*
*
*
* *
*
* ** ** **
** ** ** ** **
** **
*
LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS
iDC
CD14+ APC (DR+ CD14+)
PTE-DC(DR+ CD14–)
PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
HLA
-DR
ed
22·6
75
50 25 100
80604020
0
100
80
60
40
20
0
100 120
80 60 40 20
0
20
15
10
5
0
25
15
12
9
6
3
0
20
15
10
5
0
40
30
20
10
0
10
16
12
8
4
0
8
6
4
2
0
60
45
30
15
0
39·8 46·9 49·2 28·3
105
104
104
105
103
103
102
102
0 0 10
410
510
3 10
20
105
104
104
105
103
103
102
102
0 0
105
104
104
105
103
103
102
102
0 0
105
104
104
105
103
103
102
102
00
105
104
104
105
103
103
102
102
00
CpG Ga S C
Isotype control
CD14
Med
CD
83 (
%)
CD
83 (
%)
CD
83 (
%)
CD
86 (
%)
CD
86 (
%)
CD
86 (
%)
CpG Gar
*
*
**
*
*
*
*
* *
*
* **** **
** ** ******
** **
*
LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
Med CpG Gar LPS PIC Med CpG Gar LPS
iD
(
P(D
PIC
Med CpG Gar LPS PIC
HLA
-DR
Figure 5. Toll-like receptor (TLR) agonists promote the generation and maturation of peripheral tissue equivalent dendritic cells (PTE-DC). The
PTE modules were prepared and treated as described in Fig. 1. The reverse transmigrated (RT) cells were carefully harvested at 48 hr and stained
with fluorochrome labelled antibodies as described in Fig. 2. PTE-antigen-presenting cells (APC) are defined as Annexin V) Lin) DR+ shown in
(a), and can be further divided into two subpopulations: PTE-DC (DR+ CD14)) and CD14+ APC (DR+ CD14+). The percentage of PTE-DC is
shown in (a), and the expression of CD83, CD86 and CCR7 on PTE-DC and CD14+ APC are shown in (b) and (c), respectively. CD14+ cells
positively selected from peripheral blood mononuclear cells (PBMC) were cultured with granulocyte–macrophage colony-stimulating factor
(100 ng/ml) and interleukin-4 (25 ng/ml) for 5 days to generate immature cytokine-cultured DCs (iDC). The iDC were then cultured in the pres-
ence or absence of CpG 2006 (5 lg/ml), Poly I : C (50 lg/ml), Gardiquimod (Gar, 2�5 lg/ml), or lipopolysaccharide (LPS; 10 ng/ml). The cells
were harvested at 48 hr and stained with fluorochrome labelled antibodies as previously described, and the expression of CD83, CD86 and CCR7
on DC are shown in (d). The numbers shown in the bar graphs represent the mean ± SE of four or five different donors, and differences
between the stimulated and unstimulated PTE modules were analysed using the Student’s t-test. *P < 0�05; **P < 0�01.
380 � 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387
Y. Ma et al.
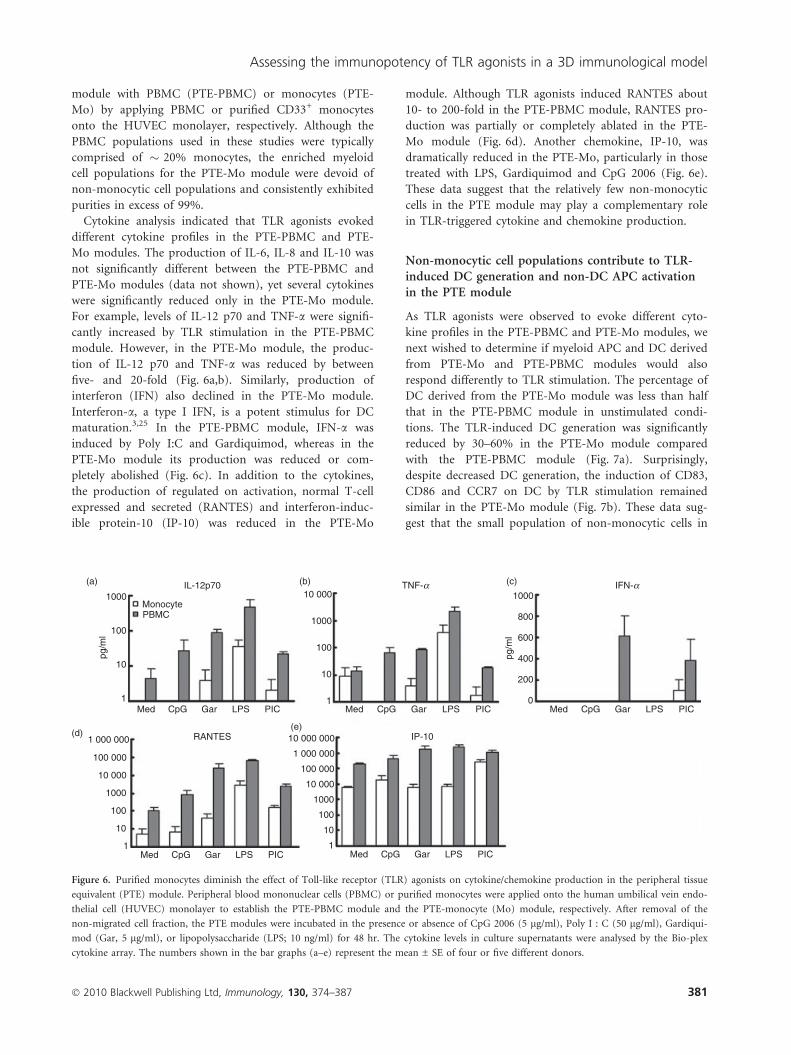
module with PBMC (PTE-PBMC) or monocytes (PTE-
Mo) by applying PBMC or purified CD33+ monocytes
onto the HUVEC monolayer, respectively. Although the
PBMC populations used in these studies were typically
comprised of � 20% monocytes, the enriched myeloid
cell populations for the PTE-Mo module were devoid of
non-monocytic cell populations and consistently exhibited
purities in excess of 99%.
Cytokine analysis indicated that TLR agonists evoked
different cytokine profiles in the PTE-PBMC and PTE-
Mo modules. The production of IL-6, IL-8 and IL-10 was
not significantly different between the PTE-PBMC and
PTE-Mo modules (data not shown), yet several cytokines
were significantly reduced only in the PTE-Mo module.
For example, levels of IL-12 p70 and TNF-a were signifi-
cantly increased by TLR stimulation in the PTE-PBMC
module. However, in the PTE-Mo module, the produc-
tion of IL-12 p70 and TNF-a was reduced by between
five- and 20-fold (Fig. 6a,b). Similarly, production of
interferon (IFN) also declined in the PTE-Mo module.
Interferon-a, a type I IFN, is a potent stimulus for DC
maturation.3,25 In the PTE-PBMC module, IFN-a was
induced by Poly I:C and Gardiquimod, whereas in the
PTE-Mo module its production was reduced or com-
pletely abolished (Fig. 6c). In addition to the cytokines,
the production of regulated on activation, normal T-cell
expressed and secreted (RANTES) and interferon-induc-
ible protein-10 (IP-10) was reduced in the PTE-Mo
module. Although TLR agonists induced RANTES about
10- to 200-fold in the PTE-PBMC module, RANTES pro-
duction was partially or completely ablated in the PTE-
Mo module (Fig. 6d). Another chemokine, IP-10, was
dramatically reduced in the PTE-Mo, particularly in those
treated with LPS, Gardiquimod and CpG 2006 (Fig. 6e).
These data suggest that the relatively few non-monocytic
cells in the PTE module may play a complementary role
in TLR-triggered cytokine and chemokine production.
Non-monocytic cell populations contribute to TLR-induced DC generation and non-DC APC activationin the PTE module
As TLR agonists were observed to evoke different cyto-
kine profiles in the PTE-PBMC and PTE-Mo modules, we
next wished to determine if myeloid APC and DC derived
from PTE-Mo and PTE-PBMC modules would also
respond differently to TLR stimulation. The percentage of
DC derived from the PTE-Mo module was less than half
that in the PTE-PBMC module in unstimulated condi-
tions. The TLR-induced DC generation was significantly
reduced by 30–60% in the PTE-Mo module compared
with the PTE-PBMC module (Fig. 7a). Surprisingly,
despite decreased DC generation, the induction of CD83,
CD86 and CCR7 on DC by TLR stimulation remained
similar in the PTE-Mo module (Fig. 7b). These data sug-
gest that the small population of non-monocytic cells in
1000Monocyte
IL-12p70 TNF-a IFN-a
PBMC
1 000 000 RANTES IP-10
100 000
10 000
1000
100
10
1Med CpG Gar LPS PIC Med CpG Gar LPS PIC
Med CpG Gar LPS PIC Med CpG Gar LPS PICMed CpG Gar LPS PIC
10 000 000
1 000 000
100 000
10 000
10 000
1000
1000
800
600
400
200
0
100
10
1
1000
100
10
1
100
10
pg/m
l
pg/m
l
1
(a)
(d)(e)
(b) (c)
Figure 6. Purified monocytes diminish the effect of Toll-like receptor (TLR) agonists on cytokine/chemokine production in the peripheral tissue
equivalent (PTE) module. Peripheral blood mononuclear cells (PBMC) or purified monocytes were applied onto the human umbilical vein endo-
thelial cell (HUVEC) monolayer to establish the PTE-PBMC module and the PTE-monocyte (Mo) module, respectively. After removal of the
non-migrated cell fraction, the PTE modules were incubated in the presence or absence of CpG 2006 (5 lg/ml), Poly I : C (50 lg/ml), Gardiqui-
mod (Gar, 5 lg/ml), or lipopolysaccharide (LPS; 10 ng/ml) for 48 hr. The cytokine levels in culture supernatants were analysed by the Bio-plex
cytokine array. The numbers shown in the bar graphs (a–e) represent the mean ± SE of four or five different donors.
� 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387 381
Assessing the immunopotency of TLR agonists in a 3D immunological model
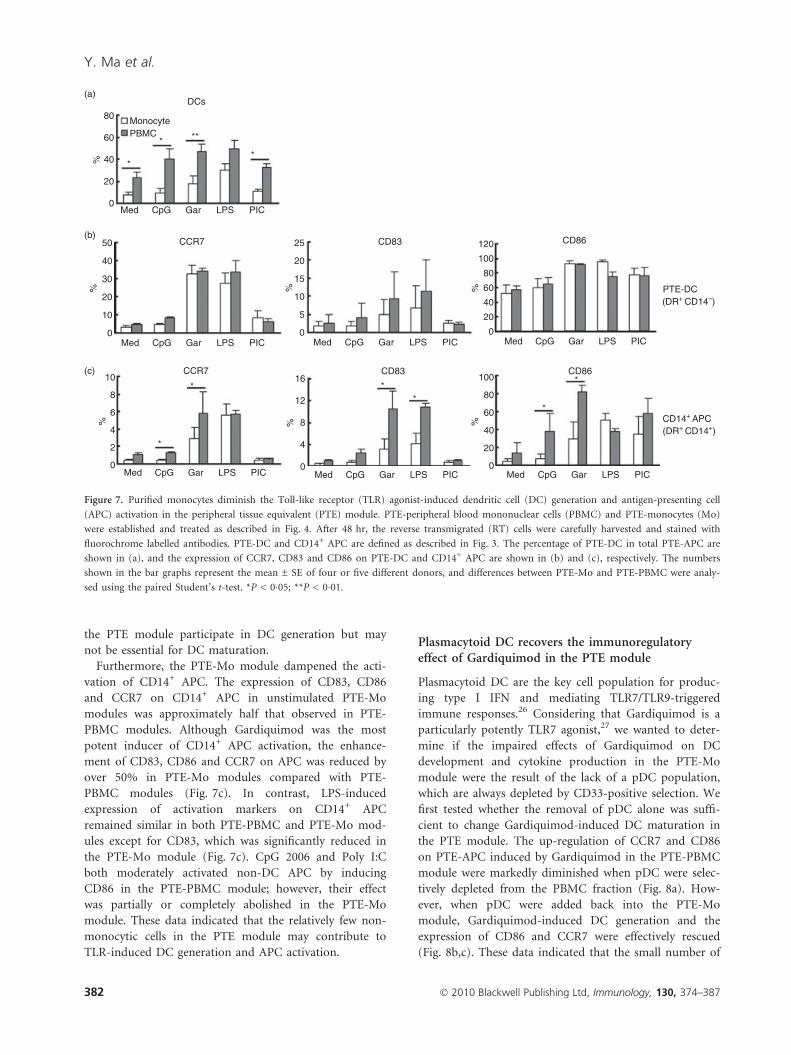
the PTE module participate in DC generation but may
not be essential for DC maturation.
Furthermore, the PTE-Mo module dampened the acti-
vation of CD14+ APC. The expression of CD83, CD86
and CCR7 on CD14+ APC in unstimulated PTE-Mo
modules was approximately half that observed in PTE-
PBMC modules. Although Gardiquimod was the most
potent inducer of CD14+ APC activation, the enhance-
ment of CD83, CD86 and CCR7 on APC was reduced by
over 50% in PTE-Mo modules compared with PTE-
PBMC modules (Fig. 7c). In contrast, LPS-induced
expression of activation markers on CD14+ APC
remained similar in both PTE-PBMC and PTE-Mo mod-
ules except for CD83, which was significantly reduced in
the PTE-Mo module (Fig. 7c). CpG 2006 and Poly I:C
both moderately activated non-DC APC by inducing
CD86 in the PTE-PBMC module; however, their effect
was partially or completely abolished in the PTE-Mo
module. These data indicated that the relatively few non-
monocytic cells in the PTE module may contribute to
TLR-induced DC generation and APC activation.
Plasmacytoid DC recovers the immunoregulatoryeffect of Gardiquimod in the PTE module
Plasmacytoid DC are the key cell population for produc-
ing type I IFN and mediating TLR7/TLR9-triggered
immune responses.26 Considering that Gardiquimod is a
particularly potently TLR7 agonist,27 we wanted to deter-
mine if the impaired effects of Gardiquimod on DC
development and cytokine production in the PTE-Mo
module were the result of the lack of a pDC population,
which are always depleted by CD33-positive selection. We
first tested whether the removal of pDC alone was suffi-
cient to change Gardiquimod-induced DC maturation in
the PTE module. The up-regulation of CCR7 and CD86
on PTE-APC induced by Gardiquimod in the PTE-PBMC
module were markedly diminished when pDC were selec-
tively depleted from the PBMC fraction (Fig. 8a). How-
ever, when pDC were added back into the PTE-Mo
module, Gardiquimod-induced DC generation and the
expression of CD86 and CCR7 were effectively rescued
(Fig. 8b,c). These data indicated that the small number of
80
60
40
20
%%
0
40
20
0
% %%
%%
0Med CpG Gar LPS PIC Med CpG Gar LPS PIC Med CpG Gar LPS PIC
Med CpG Gar LPS PIC
PTE-DC
CD14+ APC(DR+ CD14+)
(DR+ CD14–)
Med CpG Gar LPS PICMed CpG Gar LPS PIC
Med
Monocyte
DCs
CCR7
CCR7 CD83
CD83 CD86
CD86
PBMC
*
*
*
*
**
*
*
*
**
CpG Gar LPS PIC
2
4
6
8
8
4
0
12
1610
10
30
50 25 120
100
80
60
40
20
0
100
80
60
40
20
0
20
15
10
5
0
(a)
(b)
(c)
Figure 7. Purified monocytes diminish the Toll-like receptor (TLR) agonist-induced dendritic cell (DC) generation and antigen-presenting cell
(APC) activation in the peripheral tissue equivalent (PTE) module. PTE-peripheral blood mononuclear cells (PBMC) and PTE-monocytes (Mo)
were established and treated as described in Fig. 4. After 48 hr, the reverse transmigrated (RT) cells were carefully harvested and stained with
fluorochrome labelled antibodies. PTE-DC and CD14+ APC are defined as described in Fig. 3. The percentage of PTE-DC in total PTE-APC are
shown in (a), and the expression of CCR7, CD83 and CD86 on PTE-DC and CD14+ APC are shown in (b) and (c), respectively. The numbers
shown in the bar graphs represent the mean ± SE of four or five different donors, and differences between PTE-Mo and PTE-PBMC were analy-
sed using the paired Student’s t-test. *P < 0�05; **P < 0�01.
382 � 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387
Y. Ma et al.

pDC remaining in the PTE module crucially contribute
to Gardiquimod-induced PTE-DC differentiation and
APC activation.
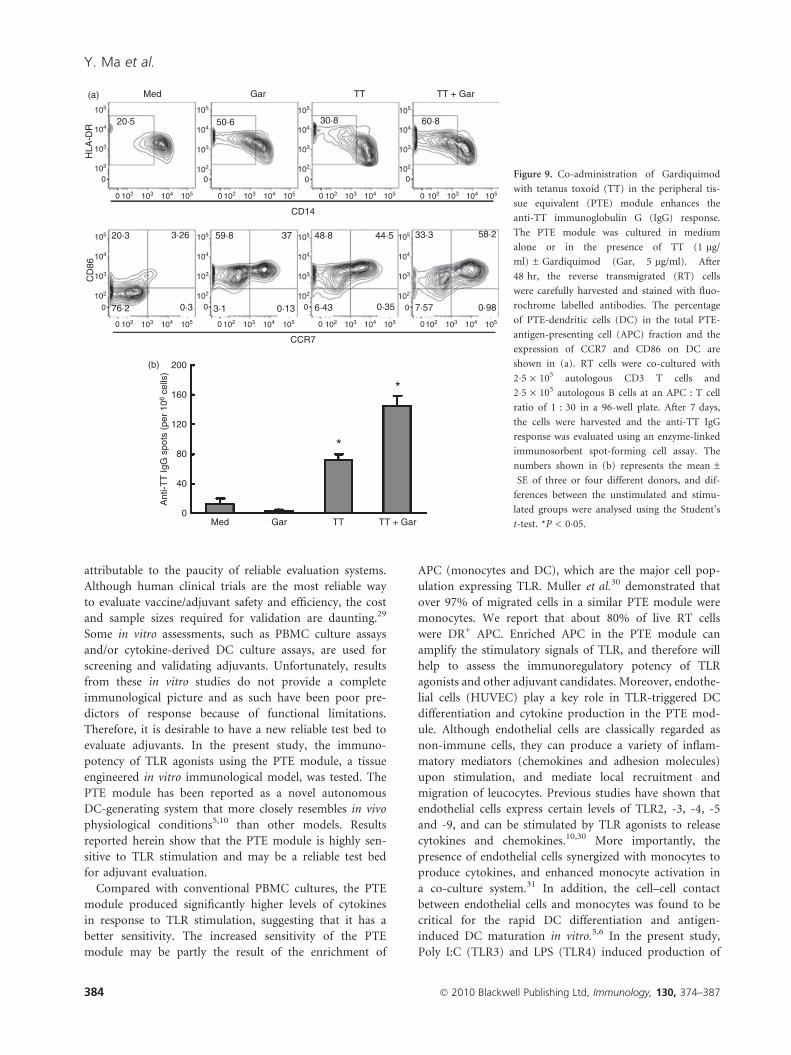
Gardiquimod enhances TT-specific IgG responsesin vitro
The major function of APC is to present antigen to T
cells and initiate adaptive immunity. As TLR agonists
could enhance the generation and/or maturation of APC
and DC, we next investigated if adjuvant-treated PTE-
APC could facilitate antigen-specific immune responses.
For these studies, we used Gardiquimod because it is a
novel TLR7 ligand and has been shown to potently
enhance the maturation of PTE-APC. The RT cells were
collected from the PTE module treated with either culture
medium alone or with TT ± Gardiquimod for 48 hr.
Consistent with previous results, Gardiquimod alone sig-
nificantly enhanced DC generation and maturation; TT
alone also markedly increased the DC generation and
promoted the expression of CD86 and CCR7 on DC. The
combination of TT and Gardiquimod resulted in the
highest percentage of CD86+ CCR7+ DC (Fig. 9a),
indicating that TT synergized with Gardiquimod in the
generation and maturation of PTE-DC. When co-cultured
with autologous T cells and B cells, PTE-DC treated with
Gardiquimod and/or TT did not impact TT-specific
T-cell and B-cell proliferation (data not shown). However,
TT-primed PTE-DC significantly induced the anti-TT IgG
response, suggesting that these DC were competent APC.
Notably, the PTE-DC treated with TT + Gardiquimod
elicited about a twofold increase in the number of anti-TT
IgG-secreting cells (Fig. 9b). Clearly, administration of
TLR agonists in the PTE module not only promoted the
generation and maturation of PTE-DCs, but also facilitated
a more pronounced antigen-specific B-cell response.
Discussion
Vaccination is currently the most cost-effective and suc-
cessful approach to prevent infectious disease in the
world. Adjuvants are an important part of vaccines and
are frequently used to enhance vaccine immunogenicity.
Over the past few decades, numerous adjuvants have been
discovered, but only a few are licensed for human use.28
The limited application of new vaccine adjuvants can be
0 102 103 104 105
0102 103 104 105 0 102 103 104 105
0 102 103 104 105 102 103 104 101 100 102 103 104 101 100
102 103 104101100 102 103 104101100
102 103 104101100 102 103 104101100
Gardiquimod
Gardiquimod
Medium
Isotype
Medium
Isotype
Cel
l num
ber
Cel
l num
ber
Who
le P
BM
Cs
Cel
l num
ber
pDC
dep
lete
d C
ell n
umbe
r
Unstained control Medium
Isotype control Gardiquimod
PBMCs CD33 CD33/pDC
Who
le P
BM
Cs
Cel
l num
ber
Pur
ified
CD
33
Cel
l num
ber
CD
33+
pD
Cs
Cel
l num
ber
CCR7 CD86
Med Gar 0
10203040506070
DC
(%
)
Unstained
Unstained
CCR7 CD86
(a) (b) (c)
Figure 8. Plasmacytoid dendritic cells (pDC) are involved in Gariquimod-induced DC development and antigen-presenting cell (APC) activation
in peripheral tissue equivalent (PTE) modules. Different PTE modules were established either using peripheral blood mononuclear cells (PBMC)
versus PTE. PBMC depleted with pDC (a), or using purified monocytes (CD33+ cells) versus purified monocytes with pDC (CD33/pDC) (b, c),
as described in the Materials and methods. The PTE modules were treated with medium only or Gardiquimod for 48 hr. The reverse transmi-
grated (RT) cells were carefully harvested and labelled with a mixture of antibodies, including the lineage markers allophycocyanin-CD3, -CD19,
-CD56 and the DC relevant markers, allophycocyanin-Cychrome 7-CD14, peridinin chlorophyll protein-Cychrome 5.5-HLA-DR, phycoerythrin-
Cychrome 7-CCR7, fluorescein isothiocyanate-CD83 and phycoerythrin-CD86. The expression of CCR7, CD83 and CD86 on PTE-APC are
shown in (a) and (c) (a representative of three independent experiments, using different donors) and the percentage of PTE-DC is shown in (b).
Filled area: unstained control; dash line: isotype control; thin line: medium-treated group; thick line: Gardiquimod-treated group.
� 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387 383
Assessing the immunopotency of TLR agonists in a 3D immunological model
attributable to the paucity of reliable evaluation systems.
Although human clinical trials are the most reliable way
to evaluate vaccine/adjuvant safety and efficiency, the cost
and sample sizes required for validation are daunting.29
Some in vitro assessments, such as PBMC culture assays
and/or cytokine-derived DC culture assays, are used for
screening and validating adjuvants. Unfortunately, results
from these in vitro studies do not provide a complete
immunological picture and as such have been poor pre-
dictors of response because of functional limitations.
Therefore, it is desirable to have a new reliable test bed to
evaluate adjuvants. In the present study, the immuno-
potency of TLR agonists using the PTE module, a tissue
engineered in vitro immunological model, was tested. The
PTE module has been reported as a novel autonomous
DC-generating system that more closely resembles in vivo
physiological conditions5,10 than other models. Results
reported herein show that the PTE module is highly sen-
sitive to TLR stimulation and may be a reliable test bed
for adjuvant evaluation.
Compared with conventional PBMC cultures, the PTE
module produced significantly higher levels of cytokines
in response to TLR stimulation, suggesting that it has a
better sensitivity. The increased sensitivity of the PTE
module may be partly the result of the enrichment of
APC (monocytes and DC), which are the major cell pop-
ulation expressing TLR. Muller et al.30 demonstrated that
over 97% of migrated cells in a similar PTE module were
monocytes. We report that about 80% of live RT cells
were DR+ APC. Enriched APC in the PTE module can
amplify the stimulatory signals of TLR, and therefore will
help to assess the immunoregulatory potency of TLR
agonists and other adjuvant candidates. Moreover, endothe-
lial cells (HUVEC) play a key role in TLR-triggered DC
differentiation and cytokine production in the PTE mod-
ule. Although endothelial cells are classically regarded as
non-immune cells, they can produce a variety of inflam-
matory mediators (chemokines and adhesion molecules)
upon stimulation, and mediate local recruitment and
migration of leucocytes. Previous studies have shown that
endothelial cells express certain levels of TLR2, -3, -4, -5
and -9, and can be stimulated by TLR agonists to release
cytokines and chemokines.10,30 More importantly, the
presence of endothelial cells synergized with monocytes to
produce cytokines, and enhanced monocyte activation in
a co-culture system.31 In addition, the cell–cell contact
between endothelial cells and monocytes was found to be
critical for the rapid DC differentiation and antigen-
induced DC maturation in vitro.5,6 In the present study,
Poly I:C (TLR3) and LPS (TLR4) induced production of
0
0102
103
104
105
102 103 104 105 0
0102
103
104
105
102 103 104 105 0
0102
103
104
105
102 103 104 105 0
0102
103
104
105
102 103 104 105
0
0102
103
104
105
102 103 104 105 0
0102
103
104
105
102 103 104 105 0
0102
103
104
105
102 103 104 105 0
0102
103
104
105
102 103 104 105
200
CCR7
CD14
Med(a)
(b)
20·5
20·3 3·26 59·8 37
3·1 0·13
48·8 44·5
6·43 0·35
33·3 58·2
7·57 0·9876·2 0·3
50·6 30·8 60·8
HLA
-DR
CD
86Gar TT TT + Gar
160
120
80
Ant
i-TT
IgG
spo
ts (
per
106
cells
)
40
0Med
*
*
Gar TT TT + Gar
Figure 9. Co-administration of Gardiquimod
with tetanus toxoid (TT) in the peripheral tis-
sue equivalent (PTE) module enhances the
anti-TT immunoglobulin G (IgG) response.
The PTE module was cultured in medium
alone or in the presence of TT (1 lg/
ml) ± Gardiquimod (Gar, 5 lg/ml). After
48 hr, the reverse transmigrated (RT) cells
were carefully harvested and stained with fluo-
rochrome labelled antibodies. The percentage
of PTE-dendritic cells (DC) in the total PTE-
antigen-presenting cell (APC) fraction and the
expression of CCR7 and CD86 on DC are
shown in (a). RT cells were co-cultured with
2�5 · 105 autologous CD3 T cells and
2�5 · 105 autologous B cells at an APC : T cell
ratio of 1 : 30 in a 96-well plate. After 7 days,
the cells were harvested and the anti-TT IgG
response was evaluated using an enzyme-linked
immunosorbent spot-forming cell assay. The
numbers shown in (b) represents the mean ±
SE of three or four different donors, and dif-
ferences between the unstimulated and stimu-
lated groups were analysed using the Student’s
t-test. *P < 0�05.
384 � 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387
Y. Ma et al.

IL-6 and IL-8 about two- to threefold in the HUVEC cul-
tures, indicating a direct stimulation of HUVEC by TLR
agonists. More importantly, compared with the conven-
tional PBMC culture, the PTE module produces signifi-
cantly higher levels of cytokines after TLR stimulation,
confirming that HUVEC and migrated monocytic cells
synergistically contributed to TLR-induced cytokine pro-
duction in the PTE module.
Furthermore, the PTE module is capable of elucidating
the effect of TLR agonists on both DC differentiation and
maturation. Both LPS and Poly I : C have been shown to
fully induce the maturation and the activation of cyto-
kine-cultured DC.32,33 However, their effect on DC gener-
ation remains unclear. The present study shows that LPS
up-regulates the expression of CD83, CD86 and CCR7 on
PTE-DC, consistent with observations in cytokine-
cultured DC. Moreover, LPS significantly increases the
percentage of DC in the PTE module. Hence, LPS pro-
motes both DC generation and maturation in the PTE
module. On the other hand, although Poly I : C induces
CD86 expression on PTE-DC, it does not significantly
regulate the percentage of DC produced. These data sug-
gest that Poly I:C can moderately induce PTE-DC matu-
ration without impacting DC generation. Surprisingly,
our results showed a weaker effect of Poly I : C on
PTE-DC maturation in comparison with that shown in
cytokine-cultured DC, which may be to the result of the
heterogeneity of the PTE-APC population, consisting of
� 20% PTE-DC and 80% monocyte-like APC. As human
TLR3 has been shown to be restricted to freshly isolated
mature DC and cytokine-derived DC,34 the presence of
monocyte-like APCs may diminish the stimulatory effect
of Poly I:C in the PTE module. In fact, in vivo DC have
been shown as a heterogeneous population. Morelli
et al.10 reported that migrating DC from skin tissue were
heterogeneous and contained both dermal DC and Lan-
gerhans cells (LC) as well as a subpopulation (� 20%) of
CD14+ LC precursors. Additionally, cells in healthy
human superficial lymph are known to contain three- to
fourfold more monocytes than DC.10 Therefore, although
the heterogeneity of PTE-APC diminished the extent of
Poly I:C-induced DC maturation, it may more closely
resemble in vivo physiological conditions.
The CpG ODNs are synthetic oligonucleotides contain-
ing unmethylated CpG motifs, which can mimic bacterial
DNA and stimulate both innate and adaptive immunity.16
Latest clinical studies also indicated that co-administra-
tion of CpG ODN with vaccine was safe and effectively
increased vaccine-specific immune responses in humans.35
The immunostimulatory effects of CpG ODN are medi-
ated by intracellular TLR9 expression predominantly in
pDC and B cells.36 The recognition of CpG ODN by
TLR9 can directly activate B cells and pDC, up-regulating
type I IFN production.37 Moreover, CpG ODN are
known to induce the differentiation of monocytes into
DC via pDC-derived type I IFN.38 Interestingly, although
cytokine-cultured DC did not respond to CpG ODN,
CpG significantly induces differentiation and maturation
of freshly isolated peripheral blood DC.37 The different
responses of cytokine-cultured DC compared with in vivo
DC implies that there are also functional differences. In
the present study, the application of CpG 2006 increased
PTE-DC generation by about 50%, and moderately
enhanced PTE-DC maturation by inducing CCR7 expres-
sion. These data are consistent with observations made in
fresh-blood DC.37 On the other hand, CpG 2006 failed to
augment DC generation in the PTE-Mo modules, suggest-
ing that CpG-induced PTE-DC generation should require
the involvement of pDC, which account for 1–2% of live
migrated cells in the PTE-PBMC module but were absent
in the PTE-Mo module. Compared with conventional
PBMC cultures and cytokine-derived DC, the PTE mod-
ule shows better sensitivity in response to CpG 2006, and
therefore could serve as a promising test bed for evaluat-
ing the immunopotency of different CpG motifs.
Gardiquimod, as a novel TLR7 ligand, has been shown
to enhance vaccine-induced immune responses in animal
models.39 However, the role of TLR7 in DC differentia-
tion and maturation remains controversial. Ito et al.40
reported that TLR7 is expressed on freshly isolated mDC
and pDC, and the treatments with TLR7 ligands, such as
Imiquimod and Resiquimod, significantly enhances
expression of CD80 and CD86 on freshly isolated DC. In
contrast, cytokine-induced iDC neither express TLR7 nor
respond to TLR7 agonists.41,42 Furthermore, co-adminis-
tration of TLR7 agonists with GM-CSF suppressed differ-
entiation and maturation of cytokine-derived DC.43 These
conflicting observations between cytokine-cultured DC
and freshly isolated DC further supports the theory that
functional differences exist between cytokine-derived and
in vivo DC. Interestingly, although cytokine-induced iDC
did not respond to TLR7 agonist, priming iDC with type
I IFN induces TLR7 expression and sensitizes iDC to
TLR7 stimulation.42
In the present study, Gardiquimod did not affect the
maturation of traditional cytokine-cultured DC; how-
ever, it did enhance PTE-DC generation and matura-
tion as well as cytokine/chemokine production. Notably,
the immunoregulatory effects of Gardiquimod were sig-
nificantly reduced when PBMC were replaced with
purified monocytes in the PTE module. Analysis of
whole PBMC selectively depleted of pDC indicates that
reduced sensitivity of the PTE module in response to
Gardiquimod could be the result of the absence of
pDC in the PTE module. Furthermore, the reconstitu-
tion of pDC effectively rescued the immunoregulatory
effect of Gardiquimod in the PTE module. Taken
together, our studies indicate that pDC are indispens-
able for Gardiquimod-triggered immune responses in
the PTE module.
� 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387 385
Assessing the immunopotency of TLR agonists in a 3D immunological model

In addition to promoting PTE-DC development and
inducing cytokine production, Gardiquimod co-adminis-
tered with TT in the PTE module also significantly
increases the production of anti-TT IgG antibodies, which
is consistent with the results in animal models.39 The cor-
relation between the PTE-DC development and antibody
production suggests that adjuvant-triggered immune
responses in the PTE module could be used to predict
their modulatory effects on the downstream antigen-
specific immune response.
Overall, the PTE module shows marked sensitivity to
TLR stimulation. Application of TLR agonists into the
PTE module not only induces cytokine and chemokine
production, but also promotes DC differentiation and
maturation. Moreover, the PTE module mimics the gen-
eration of DC that functionally resembles in vivo DC.
Hence, the tissue-engineered PTE module may serve as a
sensitive and reliable in vitro immunological model for
adjuvant assessment.
Disclosures
G.J.R. has received stock options from VaxDesign. G.J.R.
and Mount Sinai School of Medicine (MSSM) have
applied for a patent with VaxDesign for technology in
which vascular and connective tissue is reconstructed
from human cells for the purposes of vaccine testing and
selection. If this technology were licensed to a commercial
entity, then G.J.R. and MSSM would benefit financially.
References
1 Shortman K, Liu YJ. Mouse and human dendritic cell subtypes. Nat Rev Immunol 2002;
2:151–61.
2 Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K.
Immunobiology of dendritic cells. Annu Rev Immunol 2000; 18:767–811.
3 Santini SM, Di Pucchio T, Lapenta C, Parlato S, Logozzi M, Belardelli F. A new type I
IFN-mediated pathway for the rapid differentiation of monocytes into highly active
dendritic cells. Stem Cells 2003; 21:357–62.
4 HC ON, Wilson HL. Limitations with in vitro production of dendritic cells using cyto-
kines. J Leukoc Biol 2004; 75:600–3.
5 Randolph GJ, Beaulieu S, Lebecque S, Steinman RM, Muller WA. Differentiation of
monocytes into dendritic cells in a model of transendothelial trafficking. Science 1998;
282:480–3.
6 Qu C, Moran TM, Randolph GJ. Autocrine type I IFN and contact with endothelium
promote the presentation of influenza A virus by monocyte-derived APC. J Immunol
2003; 170:1010–8.
7 Qu C, Nguyen VA, Merad M, Randolph GJ. MHC class I/peptide transfer between den-
dritic cells overcomes poor cross-presentation by monocyte-derived APCs that engulf
dying cells. J Immunol 2009; 182:3650–9.
8 Randolph GJ, Sanchez-Schmitz G, Liebman RM, Schakel K. The CD16+ (FccRIII+) sub-
set of human monocytes preferentially becomes migratory dendritic cells in a model tis-
sue setting. J Exp Med 2002; 196:517–27.
9 Muller WA, Randolph GJ. Migration of leukocytes across endothelium and beyond:
molecules involved in the transmigration and fate of monocytes. J Leukoc Biol 1999;
66:698–704.
10 Morelli AE, Rubin JP, Erdos G et al. CD4+ T cell responses elicited by different subsets
of human skin migratory dendritic cells. J Immunol 2005; 175:7905–15.
11 Krishnaswamy G, Smith JK, Mukkamala R, Hall K, Joyner W, Yerra L, Chi DS. Multi-
functional cytokine expression by human coronary endothelium and regulation by
monokines and glucocorticoids. Microvasc Res 1998; 55:189–200.
12 Pober JS, Kluger MS, Schechner JS. Human endothelial cell presentation of antigen and
the homing of memory/effector T cells to skin. Ann N Y Acad Sci 2001; 941:12–25.
13 Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses.
Nat Immunol 2004; 5:987–95.
14 Ma Y, Chen Q, Ross AC. Retinoic acid and polyriboinosinic:polyribocytidylic acid stim-
ulate robust anti-tetanus antibody production while differentially regulating type 1/type
2 cytokines and lymphocyte populations. J Immunol 2005; 174:7961–9.
15 Dockrell DH, Kinghorn GR. Imiquimod and Resiquimod as novel immunomodulators.
J Antimicrob Chemother 2001; 48:751–5.
16 Verthelyi D, Klinman DM. Immunoregulatory activity of CpG oligonucleotides in
humans and nonhuman primates. Clin Immunol 2003; 109:64–71.
17 Klinman DM. CpG oligonucleotides accelerate and boost the immune response elicited
by AVA, the licensed anthrax vaccine. Expert Rev Vaccines 2006; 5:365–9.
18 Seya T, Akazawa T, Tsujita T, Matsumoto M. Role of Toll-like receptors in adjuvant-
augmented immune therapies. Evid Based Complement Alternat Med 2006; 3:31–8; dis-
cussion 133–7.
19 Pasare C, Medzhitov R. Toll-like receptors and acquired immunity. Semin Immunol
2004; 16:23–6.
20 Napolitani G, Rinaldi A, Bertoni F, Sallusto F, Lanzavecchia A. Selected Toll-like recep-
tor agonist combinations synergistically trigger a T helper type 1-polarizing program in
dendritic cells. Nat Immunol 2005; 6:769–76.
21 Walker J, Tough DF. Modification of TLR-induced activation of human dendritic cells
by type I IFN: synergistic interaction with TLR4 but not TLR3 agonists. Eur J Immunol
2006; 36:1827–36.
22 Pihlgren M, Tougne C, Schallert N, Bozzotti P, Lambert PH, Siegrist CA. CpG-motifs
enhance initial and sustained primary tetanus-specific antibody secreting cell responses
in spleen and bone marrow, but are more effective in adult than in neonatal mice. Vac-
cine 2003; 21:2492–9.
23 Reis e Sousa C. Toll-like receptors and dendritic cells: for whom the bug tolls. Semin
Immunol 2004; 16:27–34.
24 MartIn-Fontecha A, Sebastiani S, Hopken UE, Uguccioni M, Lipp M, Lanzavecchia A,
Sallusto F. Regulation of dendritic cell migration to the draining lymph node: impact
on T lymphocyte traffic and priming. J Exp Med 2003; 198:615–21.
25 Longhi MP, Trumpfheller C, Idoyaga J et al. Dendritic cells require a systemic type I
interferon response to mature and induce CD4+ Th1 immunity with poly IC as adju-
vant. J Exp Med 2009; 206:1589–602.
26 Smits EL, Ponsaerts P, Berneman ZN, Van Tendeloo VF. The use of TLR7 and TLR8
ligands for the enhancement of cancer immunotherapy. Oncologist 2008; 13:859–75.
27 Zhu J, Lai K, Brownile R, Babiuk LA, Mutwiri GK. Porcine TLR8 and TLR7 are both
activated by a selective TLR7 ligand, imiquimod. Mol Immunol 2008; 45:3238–43.
28 Pashine A, Valiante NM, Ulmer JB. Targeting the innate immune response with
improved vaccine adjuvants. Nat Med 2005; 11:S63–8.
29 Aguilar JC, Rodriguez EG. Vaccine adjuvants revisited. Vaccine 2007; 25:3752–62.
30 Muller WA, Weigl SA. Monocyte-selective transendothelial migration: dissection of
the binding and transmigration phases by an in vitro assay. J Exp Med 1992;
176:819–28.
31 Ward JR, Francis SE, Marsden L, Suddason T, Lord GM, Dower SK, Crossman DC,
Sabroe I. A central role for monocytes in Toll-like receptor-mediated activation of the
vasculature. Immunology 2009; 128:58–68.
32 Lutz MB, Kukutsch N, Ogilvie AL, Rossner S, Koch F, Romani N, Schuler G. An
advanced culture method for generating large quantities of highly pure dendritic cells
from mouse bone marrow. J Immunol Methods 1999; 223:77–92.
33 Verdijk RM, Mutis T, Esendam B, Kamp J, Melief CJ, Brand A, Goulmy E. Polyriboi-
nosinic polyribocytidylic acid (poly(I:C)) induces stable maturation of functionally
active human dendritic cells. J Immunol 1999; 163:57–61.
34 Kadowaki N, Ho S, Antonenko S, Malefyt RW, Kastelein RA, Bazan F, Liu YJ. Subsets
of human dendritic cell precursors express different toll-like receptors and respond to
different microbial antigens. J Exp Med 2001; 194:863–9.
35 Sagara I, Ellis RD, Dicko A et al. A randomized and controlled Phase 1 study of the
safety and immunogenicity of the AMA1-C1/Alhydrogel((R))+CPG 7909 vaccine for
Plasmodium falciparum malaria in semi-immune Malian adults. Vaccine 2009; 27:7292–
8.
36 Watts C, Zaru R, Prescott AR, Wallin RP, West MA. Proximal effects of Toll-like recep-
tor activation in dendritic cells. Curr Opin Immunol 2007; 19:73–8.
37 Hartmann G, Weiner GJ, Krieg AM. CpG DNA: a potent signal for growth, activation,
and maturation of human dendritic cells. Proc Natl Acad Sci U S A 1999; 96:9305–10.
38 Krug A, Rothenfusser S, Selinger S et al. CpG-A oligonucleotides induce a monocyte-
derived dendritic cell-like phenotype that preferentially activates CD8 T cells. J Immunol
2003; 170:3468–77.
39 Baldwin SL, Bertholet S, Kahn M, Zharkikh I, Ireton GC, Vedvick TS, Reed SG, Coler
RN. Intradermal immunization improves protective efficacy of a novel TB vaccine can-
didate. Vaccine 2009; 27:3063–71.
386 � 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387
Y. Ma et al.
40 Ito T, Amakawa R, Kaisho T et al. Interferon-a and interleukin-12 are induced differen-
tially by Toll-like receptor 7 ligands in human blood dendritic cell subsets. J Exp Med
2002; 195:1507–12.
41 Gorden KB, Gorski KS, Gibson SJ et al. Synthetic TLR agonists reveal functional differ-
ences between human TLR7 and TLR8. J Immunol 2005; 174:1259–68.
42 Severa M, Remoli ME, Giacomini E et al. Sensitization to TLR7 agonist in IFN-b-pre-
activated dendritic cells. J Immunol 2007; 178:6208–16.
43 Assier E, Marin-Esteban V, Haziot A, Maggi E, Charron D, Mooney N. TLR7/8 agonists
impair monocyte-derived dendritic cell differentiation and maturation. J Leukoc Biol 2007;
81:221–8.
� 2010 Blackwell Publishing Ltd, Immunology, 130, 374–387 387
Assessing the immunopotency of TLR agonists in a 3D immunological model
Copyright © 2022 FDOKUMEN




















