Assembly of ovarian follicles in the caecilians Ichthyophis tricolor and Gegeneophis ramaswamii:...
15
Zygote 15 (August), pp. 199–213. C 2007 Cambridge University Press doi:10.1017/S0967199407004327 Printed in the United Kingdom Assembly of ovarian follicles in the caecilians Ichthyophis tricolor and Gegeneophis ramaswamii: light and transmission electron microscopic study R.S. Beyo 1 , P. Sreejith 1 , L. Divya 1 , O.V. Oommen 1 and M.A. Akbarsha 2 University of Kerala, Thiruvananthapuram and Bharathidasan University, Thiruchirappalli, India Date submitted: 09.10.2006. Date accepted: 29.01.2007 Summary Though much is known about various aspects of reproductive biology of amphibia, there is little information on the cellular and mechanistic basis of assembly of ovarian follicles in this group. This is especially true of the caecilians. Therefore, taking advantage of the abundant distribution of caecilians in the Western Ghats of India, two species of caecilians, Ichthyophis tricolor and Gegeneophis ramaswamii, were subjected to light and transmission electron microscopic analysis to trace the sequential changes during the assembly of ovarian follicles. The paired ovaries of these caecilians are elongated sac-like structures each including numerous vitellogenic follicles. The follicles are connected by a connective tissue stroma. This stroma contains nests of oogonia, primary oocytes and pregranulosa cells as spatially separated nests. During assembly of follicles the oocytes increase in size and enter the meiotic prophase when the number of nucleoli in the nucleus increases. The mitochondrial cloud or Balbiani vitelline body, initially localized at one pole of the nucleus, disperses through out the cytoplasm subsequently. Synaptonemal complexes are prominent in the pachytene stage oocytes. The pregranulosa cells migrate through the connective tissue fibrils of the stroma and arrive at the vicinity of the meiotic prophase oocytes. On contacting the oocyte, the pregranulosa cells become cuboidal in shape, wrap the diplotene stage oocyte as a discontinuous layer and increase the content of cytoplasmic organelles and inclusions. The oocytes increase in size and are arrested in diplotene when the granulosa cells become flat and form a continuous layer. Soon a perivitelline space appears between the oolemma and granulosa cells, completing the process of assembly of follicles. Thus, the events in the establishment of follicles in the caecilian ovary are described. Keywords: Caecilian, Follicular assembly, Granulosa cells, Mitochondrial cloud, Oocyte Introduction The oocyte associates with somatic granulosa cells in the vertebrate ovary and becomes a structural and functional complex known as the ovarian follicle. At its earliest appearance, the oocyte enters the meiotic prophase and is wrapped by a single layer of All correspondence to: Oommen V. Oommen, Department of Zoology, University of Kerala, Kariavattom 695 581, Thiruvananthapuram, India. Tel: +91 471 2418906. e-mail: [email protected] 1 Department of Zoology, University of Kerala, Kariavattom 695 581, Thiruvananthapuram, India. 2 Department of Animal Science, School of Life Sciences, Bharathidasan University, Thiruchirappalli 620 024, India. flattened granulosa cells with little, if any, membrane associations among themselves or between the oolemma and the membrane of the granulosa cells. In mammals the earliest such fully established follicle, known as the primordial follicle, is established during fetal development or early postnatal period (Skinner, 2005) and in birds this occurs during hatching (Tokarz, 1978). With the onset of reproductive activity, a number of primordial follicles, depending on the species, develop into primary and subsequent follicular stages (Van Voorhis, 1999). However, in fishes, amphibians and reptiles, populations of oogonial stem cells are present in the ovary as discrete nests amidst somatic cells at locations depending upon whether the ovary is an elongated structure or formed into lobes. In the adult ovary number of oogonia produced from the stem
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Assembly of ovarian follicles in the caecilians Ichthyophis tricolor and Gegeneophis ramaswamii:...

Zygote 15 (August), pp. 199–213. C© 2007 Cambridge University Pressdoi:10.1017/S0967199407004327 Printed in the United Kingdom
Assembly of ovarian follicles in the caecilians Ichthyophis tricolorand Gegeneophis ramaswamii: light and transmission electronmicroscopic study
R.S. Beyo1, P. Sreejith
1, L. Divya
1, O.V. Oommen
1and M.A. Akbarsha
2
University of Kerala, Thiruvananthapuram and Bharathidasan University, Thiruchirappalli, India
Date submitted: 09.10.2006. Date accepted: 29.01.2007
Summary
Though much is known about various aspects of reproductive biology of amphibia, there is littleinformation on the cellular and mechanistic basis of assembly of ovarian follicles in this group. Thisis especially true of the caecilians. Therefore, taking advantage of the abundant distribution of caeciliansin the Western Ghats of India, two species of caecilians, Ichthyophis tricolor and Gegeneophis ramaswamii,were subjected to light and transmission electron microscopic analysis to trace the sequential changesduring the assembly of ovarian follicles. The paired ovaries of these caecilians are elongated sac-likestructures each including numerous vitellogenic follicles. The follicles are connected by a connectivetissue stroma. This stroma contains nests of oogonia, primary oocytes and pregranulosa cells as spatiallyseparated nests. During assembly of follicles the oocytes increase in size and enter the meiotic prophasewhen the number of nucleoli in the nucleus increases. The mitochondrial cloud or Balbiani vitellinebody, initially localized at one pole of the nucleus, disperses through out the cytoplasm subsequently.Synaptonemal complexes are prominent in the pachytene stage oocytes. The pregranulosa cells migratethrough the connective tissue fibrils of the stroma and arrive at the vicinity of the meiotic prophaseoocytes. On contacting the oocyte, the pregranulosa cells become cuboidal in shape, wrap the diplotenestage oocyte as a discontinuous layer and increase the content of cytoplasmic organelles and inclusions.The oocytes increase in size and are arrested in diplotene when the granulosa cells become flat andform a continuous layer. Soon a perivitelline space appears between the oolemma and granulosa cells,completing the process of assembly of follicles. Thus, the events in the establishment of follicles in thecaecilian ovary are described.
Keywords: Caecilian, Follicular assembly, Granulosa cells, Mitochondrial cloud, Oocyte
Introduction
The oocyte associates with somatic granulosa cells inthe vertebrate ovary and becomes a structural andfunctional complex known as the ovarian follicle.At its earliest appearance, the oocyte enters themeiotic prophase and is wrapped by a single layer of
All correspondence to: Oommen V. Oommen, Departmentof Zoology, University of Kerala, Kariavattom 695 581,Thiruvananthapuram, India. Tel: +91 471 2418906. e-mail:[email protected] of Zoology, University of Kerala, Kariavattom695 581, Thiruvananthapuram, India.2Department of Animal Science, School of Life Sciences,Bharathidasan University, Thiruchirappalli 620 024, India.
flattened granulosa cells with little, if any, membraneassociations among themselves or between theoolemma and the membrane of the granulosa cells.In mammals the earliest such fully established follicle,known as the primordial follicle, is established duringfetal development or early postnatal period (Skinner,2005) and in birds this occurs during hatching (Tokarz,1978). With the onset of reproductive activity, a numberof primordial follicles, depending on the species,develop into primary and subsequent follicular stages(Van Voorhis, 1999). However, in fishes, amphibiansand reptiles, populations of oogonial stem cells arepresent in the ovary as discrete nests amidst somaticcells at locations depending upon whether the ovaryis an elongated structure or formed into lobes. In theadult ovary number of oogonia produced from the stem

200 R.S. Beyo et al.
cells, as may be required depending upon the pattern ofreproduction and somatic pregranulosa cells are drawnfor the development of follicles. This is referred to asfollicular assembly and brings about soma–germ cellinteractions that play a critical role in both the germline and the physiological functions of the sexuallymature gonad (Gilula et al., 1978). Once assembled,the oogonium, which differentiates into the oocyte,is arrested in the first meiotic division (Sanchez &Villecco, 2003).
Amphibia are an appropriate biological model tostudy the interactions that take place in ovarianfollicles since the follicles pass through differentperiods (previtellogenic, vitellogenic and maturation),when various processes related to oocyte growthand maturation occur (Sharon et al., 1997; Villeccoet al., 2002). Before establishment of follicles theoocytes and pregranulosa cells exist as distinct cellpopulations (Wischnitzer, 1966; Uribe, 2001, 2003;Sanchez & Villecco, 2003). Though the morphologicaland functional relationships between amphibiandeveloping oocytes and their investing layer havebeen the subject of several investigations (Balinsky &Devis, 1963; Wallace & Dumont, 1968; Dumont, 1972;Holland & Dumont, 1975; Brummet & Dumont, 1976;Exbrayat & Collenot, 1983; Exbrayat & Laurent, 1983;Exbrayat, 1986; Villecco et al., 2002) and the outcomeshave been reviewed (Lofts, 1974; Saidapur, 1989;Uribe, 2003; Sanchez & Villecco, 2003; Exbrayat, 2006),the mechanistic details of the process of assemblyof oocytes and pregranulosa cells to establish theprevitellogenic follicle is only poorly understood.According to Uribe (2003), the morphological featuresthat distinguish oogonia, the relationship of oocyteto somatic cells and the initiation of folliculogenesisrequire definition using the electron microscope.The ultrastructural studies of folliculogenesis inamphibians as well as the comparative morphologicalaspects of the process with other vertebrates are of greatinterest for future research.
During assembly of follicle the developing oocyteis progressively surrounded by somatic prefollicularcells (Wischnitzer, 1966). The assembly of follicles iscompleted when the oocyte is surrounded by a layerof pregranulosa cells that become the granulosa cells(Dumont, 1972; Dumont & Brummet, 1978; Wallace,1985; Wallace & Selman, 1990; Sretarugsa et al., 2001;Villecco et al., 2002). Thus, the previtellogenic follicleconsists of an oocyte, arrested in diplotene, surroundedby a single layer of follicle cells (Uribe, 2003; Sanchez &Villecco, 2003). In the salamander three stages havebeen identified during early folliculogenesis: stage 1,in which oogonia divide and form cell nests; stage2 in which oogonia differentiate into oocytes; andstage 3, in which the oocyte cytoplasm increases involume (Sharon et al., 1997). Exbrayat (2006), in his
review, discusses germinal nests, follicle cells and theprevitellogenic follicles as aspects of the first (Stage A)and second (Stage B) stages in the ovary of caeciliansbut there is no description of assembly of follicles.Thus, a detailed description of assembly of folliclesin amphibia in general and caecilians in particular, ispertinent.
Caecilians are unique among the amphibians in sev-eral aspects of reproductive anatomy, physiology andbehavior. All caecilians practice internal fertilizationusing the eversible phallodeum as the intromittentorgan and many are viviparous. Even the oviparousspecies are practically ovo-viviparous in the sensethat the eggs at laying contain embryos. Caeciliansare known to have parental care. Yet, the biology offemale reproduction of caecilians has been studiedfor only a few species due to several reasons (Wake,1968, 1970a, b, 1972, 1977, 1980; Exbrayat & Collenot,1983; Exbrayat & Laurent, 1983; Exbrayat, 1986;Berois & de Sa, 1988; Masood-Parveez & Nadkarni,1993a, b; Anjubault & Exbrayat, 2004). Detailedmorphological and anatomical descriptions of caecilianovary and oogenesis are limited to three species,Chthonerpeton indistinctum (Berois & de Sa, 1988),Typhlonectes compressicuda (Exbrayat & Collenot, 1983;Exbrayat & Laurent, 1983; Exbrayat, 1986; Anjubault &Exbrayat, 2004) and Ichthyophis beddomei (Masood–Parveez & Nadkarni, 1993a, b). These studies haverelied on conventional light microscopic histology andhistochemistry. Ultrastructural descriptions of oocyteand granulosa cells during assembly of follicles andsubsequent development have not yet been attempted.In this study we describe in detail the assembly offollicles in two species of caecilians of the WesternGhats of India, Ichthyophis tricolor and Gegeneophisramaswamii.
Materials and methods
Ichthyophis tricolor (Ichthyophiidae) and Gegeneophisramaswamii (Caeciliidae) were collected from terracedplantations of mixed coconut and rubber fromThekkada (08◦37′N, 76◦57′E) in the Trivandrum districtof Kerala and Maramalai (08◦26′N, 77◦24′E) inthe Kanyakumari district of Tamil Nadu, SouthernIndia, from June 2004 to June 2005. They arerelatively abundant in the Western Ghats of Keralaand Tamil Nadu (Oommen et al., 2000). Monthlysamples (three animals from each species) werecollected, anesthetized with MS-222 (tricaine methanesulphonate) and dissected to expose the femalereproductive system. Slices of ovary were preparedfor light microscopic histological observation (paraffinembedding; hematoxylin–eosin staining). Tissues

Ovarian follicles in the caecilians 201
representing different phases of ovarian structurewere fixed in 2.5% glutaraldehyde prepared incacodylate buffer, postfixed in 1% osmium tetroxideand embedded in thin viscosity resin. Semi-thinsections (1 µm thick) were stained in toluidine blueO (TBO). In addition to making observations of thestatus of the ovary, the diameters of oogonia, oocytes,pregranulosa cells and granulosa cells, including thenuclei, were measured in five randomly selected cellsof each type using a research microscope supportedwith Q-Win software (Leica, Jena) and the data arepresented as the range. Ultrathin sections, obtainedwith a Leica ultra-microtome (Jena), were stainedwith uranyl acetate and lead citrate and subjectedto transmission electron microscopic analysis usinga Philips 201C transmission electron microscope.The images were processed using Adobe Photoshopversion 7.0.
Results
Gross anatomical and light microscopic observations
Ichthyophis tricolor and Gegeneophis ramaswamii havesimilar reproductive patterns, with regression ofovaries during March to June. The paired ovaries ofboth caecilian species are elongated sac-like structureswith previtellogenic, vitellogenic or postvitellogenicfollicles disposed like a string of beads. Theneighbouring follicles are connected by interfollicularstromal tissue (Fig. 1A, B). (The seasonal pattern ofovarian morphology will be described elsewhere.) Thenests of oogonia, premeiotic oocytes and pregranulosacells occur as clusters throughout the year in thegerminal ridges present in the stroma that connectsthe follicles in the string (Fig. 2A–C). The sphericaloogonia are 10.2–12.6 µm diameter and have 6.7–8.3 µmdiameter darkly stained nuclei lying in a slightlyeosinophilic (Fig. 2B)/acidophilic (Fig. 2C) cytoplasm.
The premeiotic oocytes are 40.8–51.6 µm diameterand contain spherical to irregularly shaped nuclei 10.9–31.3 µm diameter lying in a pale-stained cytoplasm. Thelightly staining nuclei contain one or two prominentnucleoli. There are a few patches of slightly denseheterochromatin in the nucleus. A mitochondrial cloud,either as dense aggregate or diffuse, is present atone pole of the nucleus (Fig. 2D). The pregranulosacells occur as a few clusters, separated from the germcells and are highly compact. They are elongatedcells 20.7–28.2 µm long and 12.1–15.6 µm diameter.The dark staining highly heterochromatic elongatednuclei are 15.4–23.2 µm long and 10.5–12.2 µm diameter.Fibroblast cells and capillaries are also present in thestroma (Fig. 2B–D).
Figure 1 The female urinogenital system of Ichthyophis tricolor(A) and Gegeneophis ramaswamii (B). The elongated ovary(OV) on each side is a beaded structure running parallel tothe oviduct (OD), kidney (KD) and fat body (FB). In boththe cases there are yellowish vitellogenic (VO) and whitishprevitellogenic (PF) follicles. In (B) the vitellogenic folliclesare at a more advanced stage than in (A). Scale bar represents2 cm.
TEM observations of oocytes
The premeiotic oocytes are spherical or elongatedcells surrounded by stromal tissue (Fig. 3A, B).Neighbouring oocytes are held together by membranejuxtaposition, with an occasional intercellular spacecontaining a heterogeneous material (Fig. 3B). Thenuclei of the premeiotic oocytes are spherical tohighly irregular in shape and each have a singlenucleolus in a section, but there may be more than onenucleolus in an oocyte. The chromatin is fairly uniformin the premeiotic and leptotene oocytes (Fig. 3A)but forms dense aggregates in the zygotene cells(Fig. 3B). The cytoplasm stains lightly and contains

202 R.S. Beyo et al.
Figure 2 Light micrographs of sections of ovary of Ichthyophis tricolor. (A) Toluidine blue O-stained semithin section showingthe connective tissue strand (CS) connecting vitellogenic follicles (VO). There are oogonial nests (OG) and primary oocytes (PO)in the connective tissue. Scale bar represents 60 µm. (B) Magnified view of a hematoxylin- and eosin-stained section showingoogonia (OG), primary oocytes (PO) and pregranulosa cell nests (PG). Scale bar represents 30 µm. (C) A further magnified viewof a TBO-stained semithin section showing oogonial nests (OG), primary oocytes (PO), fibroblast cells (FB), pregranulosa cellnests (PG) and a capillary (BC). Scale bar represents 20 µm. (D) A highly magnified view of a TBO-stained semithin sectionshowing ovarian epithelium (OE), oogonia (OG), primary oocytes (PO), pregranulosa cell nest (PG) and fibroblast cells (FB).The primary oocyte contains an irregularly shaped nucleus (NU) with one or two prominent nucleoli (NL). The cytoplasmcontains a prominent mitochondrial cloud (MC). Scale bar represents 7.5 µm.
mitochondria, endoplasmic reticulum, ribosomes andGolgi apparatus (not shown). The mitochondrial cloud,in view of its juxtanuclear localization, is seen only inappropriately cut sections (Fig. 3C–E).
A peculiar spherical vesicle is invariably presentin the cytoplasm in the vicinity of the mitochondrialcloud of meiotic oocytes (Fig. 3B, C–E). The contentof these vesicles is either homogenous (Fig. 3B)

Ovarian follicles in the caecilians 203
Figure 3 Early meiotic prophase oocytes of Gegeneophis ramaswamii. TEM of prefollicular oocytes. (A) This leptotene oocytehas a large irregularly shaped nucleus, containing a single nucleolus. There are few mitochondria and scarce endoplasmicreticulum in the ooplasm. Scale bar represents 0.6 µm. (B) This pachytene oocyte is increased size with the nucleus becomingproportionately smaller. The nuclear chromatin exhibits synaptonemal complexes (arrows). There are many nucleoli (only oneis shown) located around the periphery of the nucleus. Mitochondria have become more abundant in the ooplasm and form thejuxtanuclear mitochondrial cloud. There is a dense body in the ooplasm indicating the formation of Balbiani vitelline body. Theendoplasmic reticulum is still scarce. Two oocytes are held together by membrane apposition, but at places the membranes areseparated resulting in a space containing an amorphous material (arrowhead). Scale bar represents 0.6 µm. (C–E) Portions ofoocytes magnified, showing the gradual development of the Balbiani vitelline body. (C) Zygotene oocyte. Scale bar represents0.2 µm. (D) A portion of (C) further magnified showing the dense body in the mitochondrial cloud. Arrowhead points toassociation between mitochondria and dense body. Scale bar represents 0.06 µm. (E) Pachytene oocyte showing abundantmitochondria with a dense body associating with them, forming the Balbiani vitelline body. The rough endoplasmic reticulumis also abundant. The chromatin is formed into synaptonemal complexes (PC). Scale bar represents 0.1 µm. DB, dense body ofBalbiani vitelline body; ER, endoplasmic reticulum; MC, mitochondrial cloud; MT, mitochondria; NL, nucleolus; NU, nucleus;OP, ooplasm.
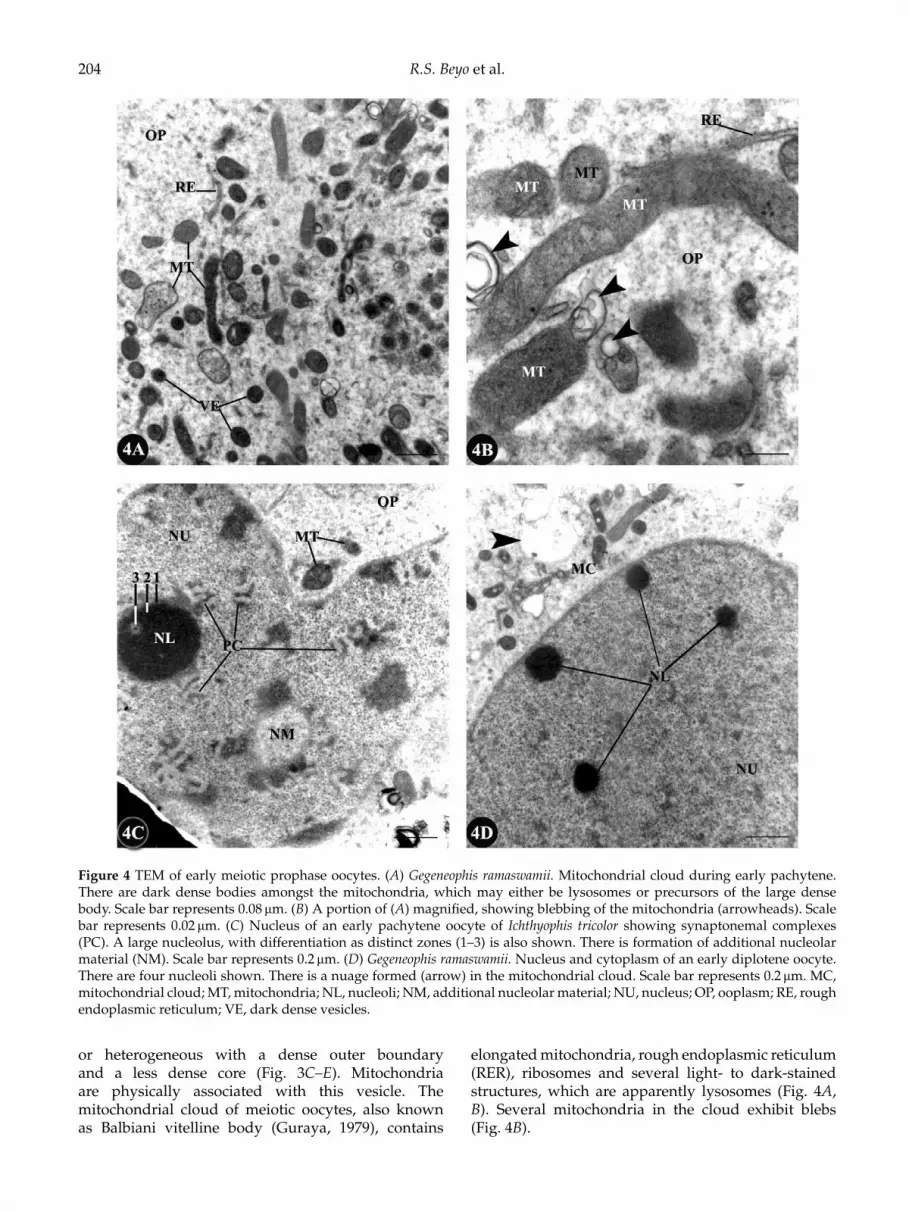
204 R.S. Beyo et al.
Figure 4 TEM of early meiotic prophase oocytes. (A) Gegeneophis ramaswamii. Mitochondrial cloud during early pachytene.There are dark dense bodies amongst the mitochondria, which may either be lysosomes or precursors of the large densebody. Scale bar represents 0.08 µm. (B) A portion of (A) magnified, showing blebbing of the mitochondria (arrowheads). Scalebar represents 0.02 µm. (C) Nucleus of an early pachytene oocyte of Ichthyophis tricolor showing synaptonemal complexes(PC). A large nucleolus, with differentiation as distinct zones (1–3) is also shown. There is formation of additional nucleolarmaterial (NM). Scale bar represents 0.2 µm. (D) Gegeneophis ramaswamii. Nucleus and cytoplasm of an early diplotene oocyte.There are four nucleoli shown. There is a nuage formed (arrow) in the mitochondrial cloud. Scale bar represents 0.2 µm. MC,mitochondrial cloud; MT, mitochondria; NL, nucleoli; NM, additional nucleolar material; NU, nucleus; OP, ooplasm; RE, roughendoplasmic reticulum; VE, dark dense vesicles.
or heterogeneous with a dense outer boundaryand a less dense core (Fig. 3C–E). Mitochondriaare physically associated with this vesicle. Themitochondrial cloud of meiotic oocytes, also knownas Balbiani vitelline body (Guraya, 1979), contains
elongated mitochondria, rough endoplasmic reticulum(RER), ribosomes and several light- to dark-stainedstructures, which are apparently lysosomes (Fig. 4A,B). Several mitochondria in the cloud exhibit blebs(Fig. 4B).

Ovarian follicles in the caecilians 205
The nucleus of pachytene oocyte contains a singlelarge nucleolus (Fig. 4C). The nucleolus has a lessdense diffuse peripheral zone and a dense core.Occasionally, the dense surrounds another less-densezone, resulting in a nucleolus that appears ring-shaped(Fig. 4C). The pairing of homologous chromosomes intosynaptonemal complexes and formation of additionalnucleolar material are also evident (Fig. 4C). Thenucleus of diplotene oocyte contains several smallnucleoli but the synaptonemal complexes are lessprominent (Fig. 4D). A few nuages are evident amongthe mitochondria (Fig. 4C, D).
Ultrastructural organization of pregranulosa cells
In their earliest appearance, the squamous pregranu-losa cells lie embedded in the stroma in the formof discrete nests (Fig. 2B–D) but later the cellsdisaggregate and tend to move away from eachother (Fig. 5A). The nuclei of pregranulosa cellsare oval to elongated and densely heterochromaticwith a continuous patch of dense heterochromatinunderneath the nuclear envelope. The nucleus occupiesalmost the entire cell with little cytoplasm around it.The cytoplasm contains few organelles (Fig. 5A, B).
Assembly of follicles
The pregranulosa cells, which are spatially separatedfrom the premeiotic and leptotene to pachytene stagemeiotic oocytes, migrate through the connective tissuefibrils of the stroma to reach the oocytes when thelatter are in the early diplotene stage and stationary(Fig. 5A–D). During this migration a large numberof dark dense bodies appear in the cytoplasm ofpregranulosa cells (Fig. 5C, D). The migratory pre-granulosa cells, on arrival at the vicinity of the oocytes,establish physical contact with the latter by displacingthe connective tissue fibrils of the stroma (Fig. 6A).A few pregranulosa cells migrate between the oocytesso that the juxtaposed oocytes separate (Fig. 6A).By this time the nucleus of the oocyte has becomespherical. The pregranulosa cells lodge themselvesaround the outer boundary of the oocytes, initially asa discontinuous layer of cuboidal pregranulosa cells(Fig. 6B). At this stage, little intercellular space is pre-sent between the oocyte and the pregranulosa cells(Fig. 6C). In the oocyte cytoplasm the mitochondrialcloud is dispersed and lysosome-like bodies appearamongst the mitochondria. During subsequent deve-lopment, the pregranulosa cells elongate and flatten,with their nuclei also transforming in shape ac-cordingly, to form the granulosa cells and the oocyteis arrested in diplotene of meiotic prophase I (Fig. 6D).The perivitelline space appears between the granulosacells and the oocyte, with occasional junctional com-plexes, either desmosomes or gap junctions, between
the two. A basal lamina appears around the follicleand sparse thecal elements associate with the follicleexternal to the basal lamina of the granulosa cells(Fig. 6D).
The cytoplasmic content of the granulosa cellincreases and dense organelles appear in the cytoplasm(Fig. 7A, B). The organelles include mitochondria,Golgi apparatus and RER. Dense fibrils are alsoseen in the cytoplasm. Later, the perivitelline spacebecomes continuous (Fig. 7C). The granulosa cellsproduce highly tortuous out-pocketing, which invadethe perivitelline space but these are not comparable tomacrovilli that would appear later in preparation tovitellogenesis (Fig. 7D).
Discussion
The period of oogonial proliferation varies from speciesto species and may be continuous or cyclical (Tokarz,1978). In the caecilians it appears to be continuousbecause fully developed ovary with vitellogenic,previtellogenic and early mitotic prophase I oocytes arepresent through out the year in spite of regression of theovary in size during a small part of the year (Masood-Parveez & Nadkarni, 1993a, b; Exbrayat, 2006; thepresent observation) as in the case of anurans (Tokarz,1978; Kanamadai & Saidapur, 1982; Pancharatna& Saidapur, 1985a, b; Pramoda & Saidapur, 1984;Saidapur, 1989; Stretarugsa et al., 2001). Following thisproliferative phase, the oogonial cells must undergoa phenotypic change as they transform to oocytes(Begovac & Wallace, 1988). In the caecilians, theearliest changes are increase in size with greateramounts of cellular organelles that appear as ajuxtanuclear aggregate as it occurs in the toadXenopus laevis (Al-Mukhtar & Webb, 1971) and in thepolyautochronic lizard Calotes versicolor (Shanbhag &Krishna Prasad, 1993). In the latter, once a batchof follicles enters vitellogenesis, no other batch ofprevitellogenic follicles (stage III) is recruited to thevitellogenic stage indicating that follicle recruitmentcould still be continuous. This is inferred from thenumber of primary oocytes and primordial folliclespresent in the ovaries during all months of the year.
In the caecilians once the oocyte has been formed, thenuclear features with respect to synaptonemal complexformation, nucleoli fragmentation and arrest in dip-lotene of prophase I are similar to those in Xenopus laevis(Al-Mukhtar & Webb, 1971; Coggins, 1973). The prom-inent features of the later phases of primary oocytegrowth include intense transcriptional activity, forma-tion and subsequent dispersal of the Balbiani vitellinebody (mitochondrial cloud) concomitant with an in-crease in cytoplasmic organelles and volume (Guraya,1979; Wallace & Selman, 1990). The several nucleoli

206 R.S. Beyo et al.
Figure 5 TEM showing the early events in the assembly of follicles. (A) Ichthyophis tricolor. Pregranulosa cells migrate throughthe stromal connective tissue and arrive at the vicinity of oocytes. Scale bar represents 1.0 µm. (B) One pregranulosa cellin (A) magnified showing the large heterochromatic nucleus and poor cytoplasmic content of the cell. Scale bar represents0.05 µm. (C) Gegeneophis ramaswamii. On arrival closer to the oocyte, the pregranulosa cell increased in organelle content.Scale bar represents 0.02 µm. (D) Ichthyophis tricolor. Accumulation of dark dense bodies, apparently ribosomal material,in the cytoplasm of pregranulosa cell. Scale bar represents 0.02 µm. CT, connective tissue strand; DG, dense granules; MT,mitochondria; NU, nucleus; OC, oocyte; OE, ovarian epithelium; PG, pregranulosa cells.
in the pachytene and early diplotene stage oocytes andtheir ring-shaped structure, as observed in the caecilianearly meiotic prophase oocytes, have been reported in
anuran and urodelan oocytes. This is referred to asamplification of nucleoli, which is meant to producethe enormous rRNA required for protein synthesis

Ovarian follicles in the caecilians 207
Figure 6 TEM showing follicular assembly. (A) Ichthyophis tricolor. Migratory pregranulosa cells are shown to establish contactwith the oocyte. Still, the neighbouring oocytes are attached by juxtaposition of membranes (arrowhead). Scale bar represents2.1 µm. (B) Gegeneophis ramaswamii. The pregranulosa cells lodge themselves around the boundary of the oocyte. Scale barrepresents 2.8 µm. (C) A portion of (B) magnified showing absence of an intercellular space between oocyte and pregranulosacell. A mitochondrial cloud is still evident in the cytoplasm of this early diplotene oocyte. Scale bar represents 1.4 µm.(D) Gegeneophis ramaswamii. The pregranulosa cell has become a flat granulosa cell. Basal lamina is seen outside the granulosacell. The perivitelline space is under formation (asterisks) but the oolemma and granulosa cell membrane are held together bymembrane junctions (arrowheads). An incipient theca is under formation. Scale bar represents 1.2 µm. BL, basal lamina; CT,connective tissue fibrils; GR, granulosa cell; MC, mitochondrial cloud; OC, oocyte; OL, oolemma; PG, pregranulosa cell; TH,theca.

208 R.S. Beyo et al.
Figure 7 Gegeneophis ramaswamii. TEM. (A, B) Increase in the cytoplasmic organelle content of granulosa cell is shown. (B) isa portion of (A) magnified. Scale bar (A) represents 0.8 µm and (B) represents 0.2 µm. (C) The perivitelline space is continuous(asterisk). Scale bar represents 1.2 µm. (D) The membrane of granulosa cell forms into folds in the perivitelline space (OF). GA,Golgi apparatus; GR, granulosa cell; MT, mitochondria; NU, nucleus; OC, oocyte; RE, rough endoplasmic reticulum.
until the mid-blastula stage (Sanchez & Villecco, 2003;Uribe, 2003). Although the amplification process in theanurans begins in premeiotic oogonia, the main periodfor the selective replication of rRNA is the pachyteneof the meiotic prophase, followed by completion in
early diplotene. The amplified nucleoli appear eitheras compact spheroidal bodies 2–10 µm in diameter oras ring-like structures (Sanchez & Villecco, 2003).
The mitochondrial cloud or Balbiani vitelline bodyhas been described for many organisms (Guraya, 1979;

Ovarian follicles in the caecilians 209
Heasman et al., 1984; Wallace & Selman, 1990) but isnot evident in the oocyte of viviparous placentotrophiclizard Mabuya mabouya (Gomez & Ramırez-Pinilla,2004). The Balbiani vitelline body is the most prominentcytoplasmic structure in the premeiotic oocyte ofthe caecilians but it disperses through out theoocyte prior to early meiotic prophase I. DuringXenopus oogenesis, the message transport organizer(METRO) pathway delivers germinal granules andlocalized RNAs to the vegetal cortex of the oocytevia the Balbiani vitelline body. According to thetraditional model, the mitochondrial cloud is thoughtto break up at the onset of vitellogenesis and thegerminal granules and METRO-localized RNAs aretransported within the mitochondrial cloud fragmentsto the vegetal cortex of the oocyte. However, astudy using light and electron microscopy, in situhybridization and three-dimensional reconstructionhas shown that germinal granules and METRO-localized RNAs are delivered to the oocyte cortexbefore the onset of mitochondrial cloud fragmentation.The delivery involves accumulation of localized RNAsand aggregation of germinal granules at the vegetal tipof the mitochondrial cloud. The subsequent internalexpansion of the mitochondrial cloud is betweenits animal (nuclear) and vegetal tips, which drivesthe germinal granules and METRO-localized RNAstowards the vegetal cortex (Wilk et al., 2005). Thesituation prevailing in the caecilians in this connectionis under investigation. Perhaps, the dark dense vesiclesappearing within the mitochondrial cloud in thecaecilian oocytes are accumulations of localized RNAs.
Follicle assembly is a critical aspect of female repro-duction, the mechanistic details of which are poorlyunderstood. Follicle assembly requires development ofoocytes from the oogonial nests and the pregranulosacells from the somatic elements of the ovarian stroma.During follicle assembly each developing oocyte isprogressively surrounded by the pregranulosa cells(Wischnitzer, 1966). At the end of this assembly thefollicle is a well established structural and functionalentity but the underlying process in the amphibianshas not been traced in adequate detail (Guraya, 1979;Uribe, 2003; Sanchez & Villecco, 2003; Exbrayat, 2006).As of now, most descriptions of ovarian cycles inthe amphibians begin with stage I, the previtellogenicfollicles, which consist of oocytes wrapped by granu-losa cells (Lofts, 1974; Rastogi et al., 1983; Pancharatna &Saidapur, 1985a, b; Saidapur, 1989; Cabada et al., 1996;Villecco et al., 1996, 2002; Sanchez & Villecco, 2003;Uribe, 2003; Exbrayat, 2006). Our light microscopic andTEM description of follicle assembly in the two speciesof caecilians fills this gap.
Folliculogenesis in caecilians commences whenoogonia differentiate into oocytes with the initiationof meiosis (Exbrayat, 2006), as in the urodeles and
anurans (Uribe, 2003; Sanchez & Villecco, 2003). Inthe few caecilian species so far studied in detail(Exbrayat & Collenot, 1983; Exbrayat & Laurent, 1983;Exbrayat, 1986; Berois & de Sa, 1988; Masood-Parveez &Nadkarni, 1993a, b; Anjubault & Exbrayat, 2004), theproliferative oocytes occur as several germinal nestsor proliferation areas disposed in a segmental fashionalong the entire length of the ovary. The ovarianfollicles lie between the islets of germinal elements(Masood-Parveez & Nadkarni, 1993b), as is the casein the caecilians in this study. The earliest stageof follicles so far described for a caecilian is theprimary follicle, which contains the oocyte with thecentral nucleus possessing one to several nucleoli,surrounded by a discontinuous single layer of flattenedepithelial cells (Exbrayat, 1986; Exbryat & Collenot,1983; Masood-Perveez & Nadkarni, 1993a, b). Exbrayat(2006) describes isolated oogonia in the caecilian ovaryas the first stage or Stage A and when the oogoniatransform into primary oocytes, which are either nakedor surrounded by a small layer of flattened follicle cells,it is the second stage or Stage B. The sequential lightmicroscopic and ultra structural changes taking placeduring these stages are described in this study.
After the assembly of follicles the granulosa cellseither remain as a simple layer consisting of a singlecell type as in most anuran and urodelan amphi-bians (Dumont & Brummet, 1978; Saidapur, 1989;De Oliviera & Santos, 2004), chelonian and crocodilianreptiles (Sarkar, et al., 1996; Uribe & Guillette, 2000;Calderon et al., 2004) and domestic goose (Kovacset al., 1992), or a simple layer differentiating intodark cells and clear cells as in the anuran Ceratophryscranwelli (Sanchez & Villecco, 2003) and the Japanesequail (Callebault, 1991) or differentiate into a hetero-geneous and multilayered structure consisting of threetypes of cells as in the case of the spotted rays Torpedomarmorata (Marina, et al., 2002, 2004) and Urolophusjamaicensis (Hamlett et al., 1999), the hylid anuranScinax fuscovarius (De Oliviera & Santos, 2004), thedove (Zarnescu, 2004) and the squamate reptiles(Andreuccetti et al., 1978; Filosa et al., 1979;Klosterman, 1987; Sarkar & Shivanandappa, 1989;Ibrahim & Wilson, 1989; Taddei & Andreuccetti, 1990;Andreuccetti, 1992; Gomez & Ramırez-Pinilla, 2004;Ricchiari et al., 2004; Hernandez-Franyutti et al., 2005).Whether caecilian follicular granulosa cells continueto remain as a single-layered homogenous structureor become a single-layered heterogeneous structureor a multilayered heterogeneous structure duringsubsequent development is under investigation.
The structural changes in the granulosa cells duringfolliculogenesis in the caecilians indicate that thegranulosa cells are the primary cell type in the ovarythat provide the physical support and the microen-vironment required for the developing oocytes. In the

210 R.S. Beyo et al.
anurans and urodeles the follicle cells form the interfacebetween the blood capillaries and the oocyte andplay critical roles in the sequestration of vitellogenin,the precursor of yolk protein, from the blood to theoocyte (Wallace & Dumont, 1968; Wallace & Bergink,1974; Wallace & Jared, 1976; Brummet & Dumont,1977; Dumont, 1978; Polzonetti-Magni, 1998; Wallaceand Selman, 1990; Hamlett et al., 1999; Stretarugsaet al., 2001; Villecco et al., 2002; Uribe, 2003; Sanchez &Villecco, 2003). In addition to this passive role, thefollicle cells in other amphibians play an active rolein the formation of vitelline envelope (Dumont &Brummet, 1978), steroid biosynthesis (Redshaw, 1972;Guraya, 1979; Wallace, 1985; Mahmoud et al., 2006),providing cAMP, which would be a regulatorymolecule in the meiotic arrest of the oocyte at diplotene(Villecco et al., 1996, 2000, 2002). The production ofa maturation inducing hormone (MIH) brings aboutoocyte maturational competence (OMC) so as torespond to MIH (Villecco et al., 1996; Patino et al.,2001), synthesis and secretion of the components of thevitelline envelope (Cabada et al., 1996). The possiblerole of the follicle cells in these aspects in the caeciliansis under investigation.
During previtellogenesis in the anuran follicles, thegranulosa cells increase in the content of RER, freeribosomes and glycogen. They become metabolicallyvery active and are involved in the synthesis of nucleicacids. It is conceivable that the follicle cells surroundingthe oocytes of caecilians are concerned with theseprocesses. A possible involvement of granulosa cellsin the transfer of ribosomes to the oocyte in lizardshas been proposed (Taddei, 1972; Andreuccetti et al.,1978; Klosterman, 1987). Cytoplasmic dense-coredgranules accumulate in the granulosa cells in closeassociation with fenestrated cisternae and networksof tubule derived from the RER of domestic goose.These granules consist of spheres and strands of anamorphous substance of unknown origin (Kovacs et al.,1992). The dark dense inclusions that appear in thepregranulosa cells during their migration towards theoocyte and later present in these cells less prominentlyafter the assembly of follicle, in the caecilians arecomparable with these granules. Perhaps, thesegranules are packaged ribosomes for export to theoocyte, an aspect worthy of further investigation.Intermediate filament bundles containing keratin havebeen shown in the follicle cells of the lizard Podarcissicula (Maurizii et al., 2000). The filaments in thepregranulosa cells of the caecilian early follicles maycorrespond to these fibrils and provide a cytoskeletalframework. The junction between granulosa cells andoocyte in the caecilians are either tight or gap junctions.In Xenopus laevis oocytes these junctions were identifiedas gap junctions based on the passage of microinjectedfluorescent dye from oocytes to follicle cells (Browne
et al., 1979). Before beginning of vitellogenesis in theanurans, the membranes of granulosa cells are apposedto the oocyte membrane, without an interface. Soon thegranulosa cell plasma membrane happens to separatefrom the oolemma, forming an interface made up ofsmall spaces with a homogenous material. Only atcertain points are both membranes in close apposition(Sanchez & Villecco, 2003). This matches with theobservation made in the caecilians in this study.
The establishment of primordial follicles in themammalian ovary requires individual oocytes tosegregate and associate with pregranulosa cells. Toprovide for this, nests of associated oocytes undergorandom apoptosis of individual oocytes to deriveisolated oocytes, which then associate with precursorsquamous granulosa cells (Skinner, 2005). During theestablishment of follicles in the caecilian ovary thepregranulosa cells physically separate the juxtaposedoocytes thereby not requiring apoptotic death ofoocytes for purpose of isolating them from one another.On the contrary in the common snook, Centropomusundecimali, cytoplasmic processes of epithelial cells andnot the entire cells, encompass meiotic oocytes andtransform into prefollicle cells, which become folliclecells at the completion of folliculogenesis (Grier, 2000).
Thus, this study describes the sequential changes inthe transformation of premeiotic oocyte into diploteneoocyte. When it is arrested in the meiotic prophase, theassembly of diplotene oocyte and pregranulosa cellsoccur. The latter becomes granulosa cells in the twospecies of caecilians.
Acknowledgements
The TEM facility of Welcome Trust ResearchLaboratory, Christian Medical College and Hospital,Vellore, is heartily acknowledged. The research wassupported with funds from the Kerala State Councilfor Science, Technology and Environment (KSCSTE),through the Science and Research Development(SARD) facility. Support under the FIST scheme ofDepartment of Science and Technology, Governmentof India, New Delhi, to the Department of Zoology,University of Kerala, Thiruvananthapuram andthe Department of Animal Science, BharathidasanUniversity, Tiruchirappalli, is also acknowledged.We thank Dr B. Kadalmani for help in the imageprocessing.
References
Al-Mukhtar, K.A.K. & Webb, A.C. (1971). An ultrastructuralstudy of primordial germ cells, oogonia and early oocytesin Xenopus laevis. J. Embryol. Exp. Morphol. 26, 195–21.
Andreuccetti, P. (1992). An ultra-structural study ofdifferentiation of pyriform cells and their contribution to

Ovarian follicles in the caecilians 211
oocyte growth in representative squamata. J. Morphol. 212,1–11.
Andreuccetti, P., Taddei, C. & Filosa, S. (1978). Intercellularbridges between follicle cells and oocyte during thedifferentiation of follicular epithelium in Lacerta sicula Raf.J. Cell.Sci. 33, 341–50.
Anjubault, E. & Exbrayat, J.M. (2004). Contribution ala connaissance de l’appareil genital de Typhlonectescompressicauda (Dumeril et Bibron, 1841), AmphibienGymnophione. I. Gonadogenese. Bull. Mens. Soc. Linn. Lyon.73, 379–92.
Balinsky, B.L. & Devis, B.J. (1963). Origin and differentiationof cytoplasmic structures in the oocytes of Xenopus laevis.Acta. Embryol. Morphol. Exp. 6, 55–108.
Begovac, P.C. & Wallace, R.A. (1988). Stages of oocytedevelopment in the pipe fish, Syngnathus scovelli. J. Morphol.197, 353–69.
Berois, N. & de Sa, R. (1988). Histology of the ovaries andfat bodies of Chthonerpeton indistinctum. J. Herpetol. 22,146–51.
Browne, C.L., Wiley, H.S. & Dumont, J.N. (1979). Oocytefollicle cell gap junctions in Xenopus laevis and the effectsof gonadotropin on their permeability. Science 203, 182–3.
Brummett, A.R. & Dumont, J.N. (1976). Oogenesis in Xenopuslaevis (Daudin) III. Localization of negative charges on thesurface of developing oocytes. J. Ultrastruct. Res. 55, 4–16.
Brummett, A.R. & Dumont, J.N. (1977). Intracellulartransport of vitellogenin in Xenopus laevis oocytes: anautoradiographic study. Dev. Biol. 60, 482–6.
Cabada, M.O., Sanchez Riera, A.N., Genta, H.D., Sanchez,S.S. & Barisone, G.A. (1996).Vitelline envelope formationduring oogenesis in Bufo arenarum. Biocell 20, 77–86.
Calderon, M.L, De Perez, G.R. & Ramirez-Pinilla, M.P. (2004).Morphology of the ovary of Caiman crocodilus (Crocodylia:Alligatoridae). Ann. Anat. 186, 13–24.
Callebault, M. (1991). Pyriform-like and holding granulosacells in the avian ovarian follicular wall. Eur. Arch. Biol.(Bruxelles) 102, 135–45.
Coggins, L.W. (1973). An ultrastructural and radioauto-graphic study of early oogenesis in the toad Xenopus laevis.J. Cell. Sci. 12, 71–93.
De-Oliveira, C. & Santos, L.R.D.S. (2004). Histologicalcharacterization of cellular types during Scina fuscovariusoogenesis (Lutz)(Anura, Hylidae). Rev. Brazil. de Zool. 21,919–23.
Dumont, J.N. (1972). Oogenesis in Xenopus laevis (Daudin).I. Stages of oocyte development in laboratory maintainedanimals. J. Morphol. 136, 153–79.
Dumont, J.N. (1978). Oogenesis in Xenopus laevis (Daudin).VI. The route of injected tracer transport in the follicle anddeveloping oocyte. J. Exp. Zool. 204, 193–217.
Dumont, J.N. & Brummet, A.R. (1978). Oogenesis in Xenopuslaevis (Daudin). V. Relationships between developingoocytes and their developing follicular tissues. J. Morphol.155, 73–98.
Exbrayat, J.M. (1986). Quelques observations sur lareproduction en elevage de Typhlonectes compressicaudus,amphibien apode vivipare. Possibilite de rythmesendogenes. Bull. Soc. Herp. Fr. 40, 52–62.
Exbrayat, J.M. (2006). Oogenesis and folliculogenesis. In:Reproductive Biology and Phylogeny of Gymnophiona, Vol. 3,
(ed. B.G.M. Jamieson & J.M. Exbrayat), pp. 275–290.Enfield, New Hampshire: Science Publishers Inc.
Exbrayat, J.M. & Collenot, G. (1983). Quelques aspects deI’evolution de I’ ovaire de Typhlonectes compressicaudus(Dumeril & Bibron, 1841), Batracien Apode vivipare. Etudequantitative et histochimique des corps jaunes. Reprod.Nutr. Dev. 23, 889–98.
Exbrayat, J.M. & Laurent, M.T. (1983). Premieres observationssur le cycle annuel de l’ ovxaire de Typhlonectescompressicaudus (Dumeril & Bibron, 1841), Batracien ApodeVivipare. CR. Acad. Sci 296, 493–8.
Filosa, S., Taddei, C. & Andreuccetti, P. (1979). Thedifferentiation and proliferation of follicle cells duringoocyte growth in Lacerta sicula. J. Embryol. Exp. Morphol.54, 5–15.
Gilula, N.B., Epstein, M.L. & Beers, W.H. (1978). Cell-to-cellcommunication and ovulation: a study of the cumulus–oocyte complex. J. Cell Biol. 78, 58–75.
Gomez, D. & Ramırez-Pinilla, M.P. (2004). Ovarian histologyof the placentotrophic Mabuya mabouya (Squamata,Scincidae). J. Morphol. 259, 90–105.
Grier, H. (2000). Ovarian germinal epithelium andfolliculogenesis in the common snook, Centropomusundecimalis (Teleostei: Centropomidae). J. Morphol. 243,265–81.
Guraya, S.S. (1979). Recent advances in the morphology,cytochemistry and function of Balbiani vitelline body inanimal oocytes. Int. Rev. Cytol. 59, 249–321.
Hamlett, W.C., Jezior, M. & Spieler, R. (1999). Ultrastructuralanalysis of folliculogenesis in the ovary of the yellowspotted stingray, Urolophus jamaicensis. Ann. Anat. 181, 159–72.
Heasman, J., Quarmby, J. & Wylie, C.C. (1984). Themitochondrial cloud of Xenopus oocytes: the source ofgerminal material. Dev. Biol. 105, 458–69.
Hernandez-Franyutti, A., Uribe-Aranzabal, M.C. & Guillette,L.J. Jr (2005). Oogenesis in the viviparous matrotrophiclizard Mabuya brachypoda. J. Morphol. 265, 152–64.
Holland, C.A. & Dumont, J.N. (1975). Oogenesis in Xenopuslaevis (Daudin). IV. Effects of gonadotropin, estrogen andstarvation on endocytosis in developing oocytes. Cell TissueRes. 162, 177–84.
Ibrahim, M.M. & Wilson, I.B. (1989). Light and electronmicroscopic studies on ovarian follicles in the lizardChalcides ocelletus. J. Zool. 218, 187–208.
Kanamadai, R.D. & Saidapur, S.K. (1982). Pattern of ovarianactivity in the Indian toad Bufo melanostictus (Schn). Proc.Natl. Sci. Acad. Indian B 48, 307–16.
Klosterman, L.L (1987). Ultrastructural and quantitativedynamics of the granulosa of the ovarian follicles of thelizard Gerrhonotus coeruleus (Family: Anguidae). J. Morphol.192, 125–44.
Kovacs, J., Forgo, V. & Peczely, P. (1992). The fine structure ofthe follicular cells in growing and atretic ovarian folliclesof the domestic goose. Cell Tissue Res. 267, 561–9.
Lofts, B. (1974). Reproduction. In: The Physiology of theAmphibia, Vol. II, (ed. B. Lofts), pp. 53–106. New York:London Academic Press.
Mahmoud, I.Y., Ba-Omar, T. & Alkindi, A. (2006). Partialdevelopment of the steroidogenic ultra structural featuresin degenerative corpora lutea after a single injection of

212 R.S. Beyo et al.
pituitary extract in the Western painted turtle (Chrysemyspicta). Tissue Cell 38, 171–6.
Marina, P., Loredana, R. & Piero, A. (2002). Ultrastructuralstudies on developing follicles of the spotted ray Torpedomarmorata. Mol. Reprod. Dev. 61, 78–86.
Marina, P., Salvatore, V., Maurizio, R., Loredana, R.,Annamaria, L., Vincenza, L., Ermelinda, L. & Piero, A.(2004). Ovarian follicle cells in Torpedo marmorata synthesizevitellogenin. Mol. Reprod. Dev. 67, 424–9.
Masood-Parveez, U. & Nadkarni, V.B. (1993a). The ovariancycle in an oviparous gymnophione amphibian, Ichthyophisbeddomei (Peters). J. Herpetol. 27, 59–63.
Masood-Parveez, U. & Nadkarni, V.B. (1993b). Morpholo-gical, histological and histochemical studies of the ovaryof an oviparous caecilian, Ichthyophis beddomei (Peters).J. Herpetol. 27, 63–9.
Maurizii, M.G., Alibardi, L. & Taddei, C. (2000). Organizationand characterization of the keratin cytoskeleton in theprevitellogenic ovarian follicle of the lizard Podarcis sicularaf. Mol. Reprod. Dev. 57, 159–66.
Oommen, O.V., Measey, G.J., Gower, D.J., Wilkinson, M.(2000). Distribution and abundance of the caecilianGegeneophis ramaswamii (Amphibia: Gymnophiona) insouthern Kerala. Curr. Sci. 79, 1386–9.
Pancharatna, M. & Saidapur, S.K. (1985a). Ovarian cycle inthe frog Rana cyanophlyctis: a quantitative study of follicularkinetics in relation to body mass, oviduct and fat bodycycles. J. Morphol. 186, 135–47.
Pancharatna, M. & Saidapur, S.K. (1985b). Seasonal variationin oocyte recruitment and development in the long-termhypophysectomized frog, Rana cyanophlyctis, in relationto homoplastic pituitary pars distalis homogenate andexogenous hormones. Indian J. Exp. Biol. 23, 183–7.
Patino, R., Yoshizaki, G., Thomas, P. & Kagawa, H. (2001).Gonadotrophic control of ovarian follicular maturation:the two-step concept and its mechanisms. Comp. Biochem.Physiol. 129, 427–39.
Polzonetti-Magni, A.M. (1998). Amphibian ovarian cycles. In:Encyclopedia of Reproduction, Vol. 1 (ed. E. Knobil & J.D. Nill),San Diego: Academic Press. pp. 154–60.
Pramoda, S. & Saidapur, S.K. (1984). Annual changes in thesomatic weight, hypophysial gonadotrophs, ovary, oviductand abdominal fat bodies in the Indian bullfrog Ranatigerina. Proc. Natl. Sci. Acad. Indian B50, 387–98.
Rastogi, R.K., Izzo-Vitiello, I., DiMeglio, M., Di Matte, L.,Franzese, R., Dicostanzo, M.G., Minncci, S, Iela, L. & Chieffi,G. (1983). Ovarian activity and reproduction in the frogRana esculenta. J. Zool. Lond. 200, 223–47.
Redshaw, M.R. (1972).The hormonal control of amphibianovary. Am. Zool. 12, 289–306.
Ricchiari, L., Carmela, V.M., Marina, P., Rosa, C., Annamaria,L. & Piero, A. (2004). Alpha and beta spectrin distributionduring the differentiation of pyriform cells in follicles oflizard Podarcis sicula. Mol. Reprod. Dev. 67, 101–7.
Saidapur, S.K. (1989). Reproductive cycles of amphibians.In: The Reproductive Cycles of Indian Vertebrates. ed. S.K.Saidapur, New Delhi: Allied Publishers. pp. 166–224.
Sanchez, S. & Villecco, E.I. (2003). Oogenesis. In: ReproductiveBiology and Phylogeny of Anura, Vol. 2, ed. B.G.M.Jamieson, Enfield, New Hampshire: Science Publishers Inc.pp. 27–71.
Sarkar, H.B.D. & Shivanandappa, T. (1989). Reproductivecycles of reptiles. In: Reproductive Cycles of Indian Vertebrates.ed. S.K. SAIDAPUR, pp. 225–72. New Delhi: AlliedPublishers.
Sarkar, S., Sarkar, N.K., Das., P. & Maiti, B.R. (1996).Photothermal effects on ovarian growth and function in thesoft-shelled turtle Lissemys punctata punctata. J. Exp. Zool.274, 41–55.
Shanbhag, A. & Krishna Prasad, B.S. (1993). Folliculardynamics and germinal bed activity during the annualovarian cycle in the lizard, Calotes versicolor. J. Morphol.216, 1–7.
Sharon, R., Degani, G. & Warburg, M.R. (1997). Oogenesis andthe ovarian cycle in Salamandra salamandra infraimmaculataMertens (Amphibia; Urodela; Salamandridae) in fringeareas of the taxon’s distribution. J. Morphol. 231, 149–60.
Skinner, M.K. (2005). Regulation of primordial follicleassembly and development. Human Reprod. Update 11, 461–71.
Sretarugsa, P., Weerachatyanukul, W., Chavadej, J.,Kruatrachue, M. & Sobhon, P. (2001). Classification ofdeveloping oocytes, ovarian development and seasonalvariation in Rana tigerina. Science Asia 27, 1–14.
Taddei, C. (1972). Significance of pyriform cells in ovarianfollicles of Lacerta sicula. Exp. Cell Res. 72, 562–6.
Taddei, C. & Andreuccetti, P. (1990). Structural modificationsof the nuclear components during lizard oogenesis inrelation to the differentiation of the follicular epithelium.Cell Differ. Dev. 29, 205–15.
Tokarz, R.R. (1978). Oogonial proliferation, oogenesis andfolliculogenesis in non-mammalian vertebrates. In: TheVertebrate Ovary. ed. R.R. Jones, pp. 145–79. New York:Plenum Press.
Uribe, M.C.A. (2001). Reproductive systems of caudateamphibians. In: Vertebrate Functional Morphology (ed. H.M.Dutta & J.S. Datta Munshi), pp. 267–94. Enfield, NewHampshire: Science Publishers Inc.
Uribe, M.C.A. (2003). Reproductive biology and phylogenyof Urodela. In: Ovary and Oogenesis (ed. B.G.M. Jamieson &D.M. Sever), pp. 135–50. Enfield, New Hampshire: SciencePublishers Inc.
Uribe, M.C.A. & Guillette, L.J. Jr. (2000). Oogenesis andovarian histology of the American alligator Alligatormississippiensis. J. Morphol. 245, 225–40.
Van Voorhis, B.J. (1999). Follicular development. In:Encyclopedia of Reproduction, Vol. 2, (ed. E. Knobil & J.D.Nill), pp. 376–389. San Diego: Academic Press.
Villecco, E.I., Aybar, M.J., Sanchez, S.S. & Sanchez Riera,A.N. (1996). Heterologous gap junctions between oocyteand follicle cells in Bufo arenarum: hormonal effects ontheir permeability and potential role in meiotic arrest.J. Exp. Zool. 276, 76–85.
Villecco, E.I., Aybar, M.J., Genta, S.B., Sanchez, S.S. & SanchezRiera, A.N. (2000). Effect of gap junction uncoupling in fullBufo arenarum ovarian follicles: participation of cAMP inmeiotic arrest. Zygote 8, 171–9.
Villecco, E.I., Genta, S.B., Sanchez Riera, A.N. & Sanchez, S.S.(2002). Ultrastructural characteristics of the follicle cell–oocyte interface in the oogenesis of Ceratophrys cranwelli.Zygote 10, 163–73.

Ovarian follicles in the caecilians 213
Wake, M.H. (1968). Evolutionary morphology of the caecilianurogenital system. Part I: The gonads and fat bodies.J. Morphol. 126, 291–332.
Wake, M.H. (1970a). Evolutionary morphology of thecaecilian urogenital system. Part II: The kidneys andurogenital ducts. Acta. Anat. 75, 321–58.
Wake, M.H. (1970b). Evolutionary morphology of thecaecilian urogenital system. Part III: The bladder.Herpetologica 26, 120–8.
Wake, M.H. (1972). Evolutionary morphology of the caecilianurogenital system. Part IV: The cloaca. J. Morphol. 136, 353–66.
Wake, M.H. (1977). The reproductive biology of caecilians.In: The Reproductive Biology of Amphibians: An EvolutionaryPerspective (ed. D.H. Taylor & S.I Guttman), pp. 73–101.New York: Plenum Press.
Wake, M.H. (1980). Reproduction, growth and populationstructure of the Central American caecilian Dermophismexicanus. J. Herpetol. 36, 244–56.
Wallace, R.A. (1985). Vitellogenesis and oocyte growth innon-mammalian vertebrates. In: Developmental Biology,Vol. I, ed. L.W. Browder, pp. 127–77. New York: PlenumPress.
Wallace, R.A. & Bergink, E.W. (1974). Amphibian vitellogenin:properties, hormonal regulation of hepatic synthesis andovarian uptake and conversion to yolk proteins. Am. Zool.14, 1159–75.
Wallace, R.A. & Dumont, J.N. (1968). The induced synthesisand transport of yolk protein and their accumulation bythe oocyte in Xenopus laevis. J. Cell Comp. Physiol. 72, 73–89.
Wallace, R.A. & Jared, D.W. (1976). Protein incorporation byisolated amphibian oocytes. V. Specificity for vitellogeninincorporation. J. Cell Biol. 69, 345–51.
Wallace, R.A. & Selman, K. (1990). Ultra structural aspectsof oogenesis and oocyte growth in fish and amphibians.J. Electron Microscopy Techniques 16, 175–201.
Wilk, K., Balinski, S., Dougherty, M.T. & Kloc, M. (2005).Delivery of germinal granules and localized RNAs viathe messenger transport organizer pathway to the vegetalcortex of Xenopus oocytes occurs through directionalexpansion of the mitochondrial cloud. Int. J. Dev. Bol. 49,17–21.
Wischnitzer, S. (1966). The ultrastructure of the cytoplasm ofthe developing amphibian egg. Adv. Morphog. 5, 131–79.
Zarnescu, O. (2004). Ultrastructural observations ofprevitellogenic ovarian follicles of dove. Zygote 12, 285–92.




![Microscopic and macroscopic creativity [Comment]](https://static.fdokumen.com/doc/165x107/63222cba63847156ac067f99/microscopic-and-macroscopic-creativity-comment.jpg)