
analysis of steam propulsion plants - Deep Blue Repositories
Upload
khangminh22Category
view
10download
0
Acc
epte
d A
rtic
leDeletion of Macrophage Migration Inhibitory Factor Reduces Severity of Osteoarthritis in
Aged Mice
Meredith A. Rowe1, Lindsey R. Harper
2, Margaret A. McNulty
3, Anthony G. Lau
4, Cathy S.
Carlson2, Lin Leng
5, Richard J. Bucala
5, Richard A. Miller
6, Richard F. Loeser
1
Running title: Mif Deletion Reduces Age-related OA
This work was supported by grants from the National Institute on Aging (AG046990) to M.A.R.,
(AG044185) to R.F.L., and (AG024824) to R.A.M. and from the National Institute of Arthritis
and Musculoskeletal and Skin Diseases (AR049610) to R.J.B.
1Meredith A. Rowe, PhD and Richard F. Loeser, MD: Wake Forest School of Medicine,
Winston-Salem, North Carolina and University of North Carolina at Chapel Hill, Chapel Hill,
North Carolina. 2Lindsey R. Harper, BS and Cathy S. Carlson, DVM, PhD: University of
Minnesota, St. Paul, Minnesota. 3Margaret A. McNulty, PhD: Louisiana State University, Baton
Rouge, Louisiana. 4Anthony G. Lau, PhD: University of North Carolina at Chapel Hill, Chapel
Hill, North Carolina. 5Lin Leng, PhD and Richard J. Bucala, MD, PhD: Yale University, New
Haven, Connecticut. 6Richard A. Miller, MD, PhD: University of Michigan, Ann Arbor,
Michigan.
Page 1 of 36 Arthritis & Rheumatology
This is the author manuscript accepted for publication and has undergone full peer review but has not beenthrough the copyediting, typesetting, pagination and proofreading process, which may lead to differencesbetween this version and the Version record. Please cite this article as doi:10.1002/art.39844.
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
2
Address correspondence to: Richard F. Loeser M.D., Thurston Arthritis Research Center,
University of North Carolina, 3300 Thurston Building, CB #7280, Chapel Hill, NC 27599-7280.
E-mail: [email protected]
Page 2 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
3
Abstract
Objective: Macrophage migration inhibitory factor (MIF) is a pro-inflammatory cytokine
that is elevated in the serum and synovial fluid of osteoarthritic (OA) patients. In this study, the
potential role of MIF in OA was studied using human joint tissues and in vivo in mice with age-
related and surgically induced OA.
Methods: MIF in conditioned media from human chondrocytes and meniscal cells and
from cartilage explants was measured by ELISA. Severity of OA was analyzed histologically in
male wild-type and Mif-/- mice at 12- and 22-months of age and following destabilization of the
medial meniscus (DMM) surgery in 12-week old Mif-/- mice as well as in wild-type mice treated
with a neutralizing MIF antibody. Synovial hyperplasia was graded in S100A8 immunostained
histologic sections. Bone morphometric parameters were measured by microCT analysis.
Results: Human OA chondrocytes secreted 3-fold higher levels of MIF than normal
chondrocytes, while normal and OA meniscal cells produced equivalent amounts. Compared to
age- and strain-matched controls, the cartilage, bone, and synovium in older adult mice with Mif
deletion were protected from changes of naturally occurring age-related OA. No protection from
DMM-induced OA was seen in young adult Mif-/- mice or in wild-type mice treated with anti-
MIF. Increased bone density in 8 week-old mice with Mif deletion was not maintained at 12-
months.
Conclusions: These results demonstrate a differential mechanism in the pathogenesis of
naturally occurring age-related OA compared to injury-induced OA. The inhibition of MIF may
represent a novel therapeutic target in the reduction of age-related OA.
Page 3 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
4
Aging has been well-described as a key risk factor for the development of osteoarthritis
(OA) (1). While aging does not directly cause OA, aging changes in joint tissues and possibly
circulating factors that change with age increase the susceptibility for the development of the
disease. Aging changes are reflected in the cartilage matrix including thinning of the articular
cartilage (2) and the accumulation of advanced glycation end products (AGE) which alters the
biomechanical properties of the joint (3). Aging changes are also evident in chondrocytes, where
chondrocytes isolated from aged individuals are more resistant to stimulation with the growth
factors insulin-like growth factor 1 (IGF-1) (4, 5), transforming growth factor-β (TGF-β) (6), and
bone morphogenetic protein-6 (BMP-6) (7) than chondrocytes isolated from younger individuals.
Inflammatory mediators have been found to be up-regulated in aged joint tissues including IL-7
in chondrocytes and synovial fluid in humans and IL-33, CXCL13, CCL8, and CCL5 in the
mouse knee joint (reviewed in Greene and Loeser (8)). The increased expression of these
inflammatory mediators, which is compounded by aging changes in the tissues, may contribute
to the development of OA.
Macrophage migration inhibitory factor (MIF) is an inflammatory cytokine that has been
studied for its role in the immune system. MIF has been shown to function as a cytokine that
signals through the CD74 receptor (9) to increase neutrophil migration to regions of
inflammation and to promote the innate immune response (10, 11). Additionally, MIF promotes
macrophage activation, increasing phagocytosis and destruction of pathogens (11, 12). MIF has
been studied in autoimmune diseases including rheumatoid arthritis (RA) and systemic lupus
erythematosus (SLE). It has been shown to promote MMP-1 and MMP-3 production in synovial
fibroblasts isolated from the knee joints of RA patients (13) and also has been shown to be
significantly elevated in RA synovial fluid (14). In a more recent study, inhibition of MIF
Page 4 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
5
function either by deletion of Mif, or its receptor, Cd74, was found to reduce the severity of
disease in an RA model by reducing both inflammation and bone erosion (15). In SLE, the
amount of MIF in serum is positively correlated with tissue damage (16), and the renal
expression of MIF was shown to be significantly elevated in an SLE-prone mouse model (17).
The level of MIF was noted to be elevated in the serum and synovial fluid of patients with knee
OA compared to healthy controls (14, 18). However, the potential contribution of MIF to the
development of OA has not been previously studied.
Recently, Mif-/- 129Sv/C57Bl/6 mice were reported to have an increased lifespan when
compared to wild-type (WT) controls, suggesting a role for MIF in the aging process (19),
although more recent data (Miller, unpublished) suggest that this beneficial effect may be absent
on other genetic backgrounds. Like OA, atherosclerosis is a common disease of aging and
inhibition of MIF has been found to reduce atherosclerosis in a mouse model of the disease (20)
suggesting that MIF may represent a therapeutic target for age-related disease. Given the
association between aging and OA, we sought to determine the contribution of MIF to the
development of OA. We found that genetic disruption of Mif decreased the severity of naturally-
occurring OA in aged mice but not in a surgically-induced model of OA in younger adult mice.
MATERIALS & METHODS
Chondrocyte and meniscal cell isolation and culture. Normal human donor tissue
(both knee and ankle joints) was obtained from the Gift of Hope Organ and Tissue Donor
Network (Elmhurst IL) through the Rush University Medical Center in Chicago, IL.
Osteoarthritic tissue was obtained as surgical waste from total knee replacement surgeries
performed in the Department of Orthopaedic Surgery at Wake Forest Baptist Health and at the
Page 5 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
6
University of North Carolina Hospitals. The use of human tissue was approved by the
Institutional Review Boards at Rush University, Wake Forest Baptist Health, and the University
of North Carolina at Chapel Hill. All human donor tissue was de-identified before receipt, and
the cartilage and meniscus were processed separately. The cartilage was dissected away from
the subchondral bone in small flakes and rinsed in serum-free DMEM-F12 media (Gibco Life
Technologies) to prevent the tissue from drying out. After the excess fat and ligamentous tissue
were removed, the meniscus was dissected into small pieces to enable more efficient digestion.
Cells were isolated separately from cartilage and meniscus using pronase and collagenase
digestion as previously described (21). The cells were cultured in 10% serum DMEM-F12
media until confluent. Cultures were serum-starved overnight before media collection.
A 4mm biopsy punch was used to harvest human cartilage explants from the cartilage
pieces that had been dissected away from the subchondral bone. Three explants per donor were
pooled in one well and cultured in 10% serum DMEM-F12 media for approximately 72 hours.
The explants were serum-starved for 48 hours before media collection. The explants were
digested in papain, and the DNA content was quantified using the PicoGreen kit (Life
Technologies) as previously described (22).
Experimental animals. The mice used for these studies were housed and maintained
according to the IACUC guidelines at the respective institutions where each study was
performed (The University of Michigan, Wake Forest School of Medicine, and The University of
North Carolina at Chapel Hill). Knee joints from Mif-/- and wild-type (WT) mice for the aging
experiment were from a previously reported study (19). For the surgically-induced OA studies,
the Mif-/- colony was bred on the SV129/C57Bl/6 mixed background. The colony was
maintained by heterozygous breeding, so the Mif-/- and WT mice used in these studies were age-
Page 6 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
7
and strain-matched littermates. The C57Bl/6 mice used in the neutralizing antibody study were
purchased directly from Jackson Labs.
ELISA. MIF content in serum-free conditioned media was quantified by solid-phase
ELISA (R&D Systems) according to the manufacturer’s protocol. Due to the high level of MIF
in the media, the media samples were diluted 1:25 in the assay.
MIF immunostaining. Human normal and OA cartilage sections were a kind gift from
Martin Lotz (Scripps Research Institute, La Jolla, CA). Human cartilage sections and mid-
coronal sections from 12-month old wild-type and Mif-/- mouse knee joints were immunostained
with anti-MIF (Life Technologies). Sections were de-paraffinized and rehydrated in serial
ethanol washes followed by antigen retrieval in citrate buffer. Sections were first blocked with
3% H2O2 (Fisher Scientific) and then with Protein Block (Dako). The primary antibody was
diluted in antibody diluent (Dako) and incubated on the sections overnight at 4°C. The
following day, the sections were incubated with HRP-linked secondary antibody (Dako) and
developed with the DAB chromagen (Dako). The sections were counterstained in Mayer’s
Hematoxylin (Sigma) and then dehydrated in serial ethanol washes.
DMM procedure. The destabilization of the medial meniscus (DMM) procedure was
performed in 12-week old male Mif-/- and WT mice as described previously (23). This
procedure induces OA by transecting the medial meniscotibial ligament (MMTL). For the sham
surgery, the joint was opened and visualized, but the MMTL was not cut. We randomly assigned
14-16 mice to each surgical group and to each of the anti-MIF treatment and control groups
(described below). The number of mice chosen for the present study was based on a power
calculation using data from a previously published DMM and sham control experiment from our
group that included 12-week old male C57BL/6 mice (23) and found that a sample size of 12
Page 7 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
8
mice per group would provide greater than 80% power to detect at least a 50% difference in
articular cartilage structure scores between groups. After the surgical procedure, the mice were
monitored closely for signs of apparent pain or adverse effects of the procedure. The mice were
maintained in normal housing conditions and allowed to exercise through normal activities. The
mice were euthanized 10 weeks post-surgery, and the hind-limbs were collected in 10% formalin
(Fisher Scientific) for histological analysis.
In the anti-MIF study, a neutralizing murine anti-MIF IgG1 (clone NIHIIID.9) and
isotype control antibody were purified from ascites fluid using Protein A/G spin columns
(Thermo Scientific). Following isolation, the antibodies were dialyzed overnight in DPBS
(Lonza) and then filter-concentrated by centrifugation (Millipore). The antibody concentration
was measured by the BCA assay (Thermo Scientific). After DMM surgery, mice were allowed
to recover for five days before antibody treatment began. The antibodies were administered by
IP injection for the duration of the study at a dose (20 mg/kg twice per week) previously shown
to neutralize MIF in vivo (24). The mice were euthanized 10 weeks after surgery, and the legs
were collected in formalin for histological analysis. Because repeated handling and injections
can cause pain or other adverse effects which would be reflected with a decrease in food intake,
the body weight of each mouse was monitored throughout the course of the study. All mice
gained weight consistently over the course of the study, and there was no difference in the
average body weights of each group at any time point during the study (data not shown).
Histology processing and OA grading. The formalin-fixed hind-limbs were transferred
to 70% ethanol, and the excess soft tissue was removed. The samples were decalcified in 19%
EDTA (Fisher Scientific), and the intact joints were embedded in paraffin. The joints were
sectioned along the coronal plane at 4µm. Mid-coronal sections were stained with hematoxylin
Page 8 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
9
and eosin (H&E) and scored using the articular cartilage structure (ACS) score developed by
McNulty et al (25). This system scored the integrity of the articular cartilage on a scale of 0-12,
where 0 represents normal healthy cartilage and 12 represents full-thickness loss of the articular
cartilage across more than two-thirds of the surface scored. Adjacent mid-coronal sections were
also stained with Safranin O and fast green and scored using the Saf-O score developed by
McNulty et al (25). The Saf-O system scored the proteoglycan content of the articular cartilage
on a scale of 0-12, where 0 represents uniform staining of healthy cartilage and 12 represents
complete loss of staining across more than two-thirds of the surface. The Saf-O score is more
sensitive to the mild to moderate lesions that result from the DMM procedure, as it relies on
proteoglycan loss from the matrix vs. fibrillation and loss of cartilage. Therefore, it (rather than
the ACS scoring system) was used to score OA in those studies. Additional joint measurements
were made using the OsteoMeasure histomorphometry system (OsteoMetrics) as described (25).
The additional parameters measured were: articular cartilage area and thickness, subchondral
bone area and thickness, number of viable chondrocytes, area of chondrocyte necrosis, and
calcified cartilage area and thickness.
Osteophyte assessment. Osteophytes were graded using a system that was modified
from a previously published study (26). Osteophytes were graded on a scale of 0-3 as follows: 0
= no osteophyte, 1 = questionable/borderline osteophyte, 2 = small osteophyte, and 3 = large
osteophyte. The summed osteophyte scores for the medial tibial plateau and the medial femoral
condyle are presented.
Synovial assessment. Coronal sections were immunostained with anti-S100A8 (kindly
provided by Johannes Roth) which has been used as a method to detect synovitis with synovial
hyperplasia in mouse joints with OA (27). Immunohistochemistry was performed as above
Page 9 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
10
except that proteinase K was used for antigen retrieval. Synovial hyperplasia was graded on a
single mid-coronal section from each mouse using a scale of 0-3 as follows: 0 = 1-3 cell layers
in synovium, 1 = 4-6 cell layers, 2 = 7-9 cell layers, and 3 = 10 or more cell layers. The medial
and lateral compartments of the joint were scored separately, and the sum of the two scores is
presented.
MicroCT analysis. The knee joints from 8-week old (n=3 each Mif-/- and WT) and 12-
month old (n=4 Mif-/- and n=5 WT) male mice were scanned with 10 µm resolution by microCT
(µct80; Scanco Medical AG, Brüttisellen, Switzerland) as previously described (28). Bone
morphometric parameters were measured in the trabecular bone of the proximal metaphysis of
the tibia using Scanco software (Scanco Medical AG, Brüttisellen, Switzerland). The region of
interest was consistently defined to begin inferior to the growth plate and extended for 1.00mm
distally. Bone image data were rotated as necessary to adjust for any vertical alignments. The
following histomorphometric trabecular bone parameters (29) were analyzed using the same
threshold for each animal: bone volume fraction, tissue mineral density, connectivity density,
trabecular number, trabecular separation, and trabecular thickness.
Statistical Analysis. All statistical analyses were performed using GraphPad Prism 6
with one exception: OA histomorphometry measurements were analyzed using SPSS, as noted.
Data are presented graphically as individual data points with horizontal lines representing the
mean of each group. MIF ELISA samples were collected from monolayer cultures of cells or
explant cultures from unique human tissue donors (n=7-16 donors/group) and analyzed by
unpaired t-test within each tissue type. OA severity (ACS and Saf-O scores) was analyzed from
individual mice (n=13-16 mice/group) by Kruskal-Wallis nonparametric one-way ANOVA
followed by Dunn’s multiple comparisons test. Histomorphometric measurements were
Page 10 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
11
analyzed by heteroscedastic t-test. Synovial hyperplasia was evaluated in a subset of mice from
the 12- and 22-month old groups (n=5-11 mice/group) and analyzed by Kruskal-Wallis
nonparametric one-way ANOVA followed by Dunn’s multiple comparisons test. Bone
parameters were measured in individual mice (n=3-5 mice/group) and analyzed by two-way
ANOVA followed by Tukey’s multiple comparisons post-hoc test.
RESULTS
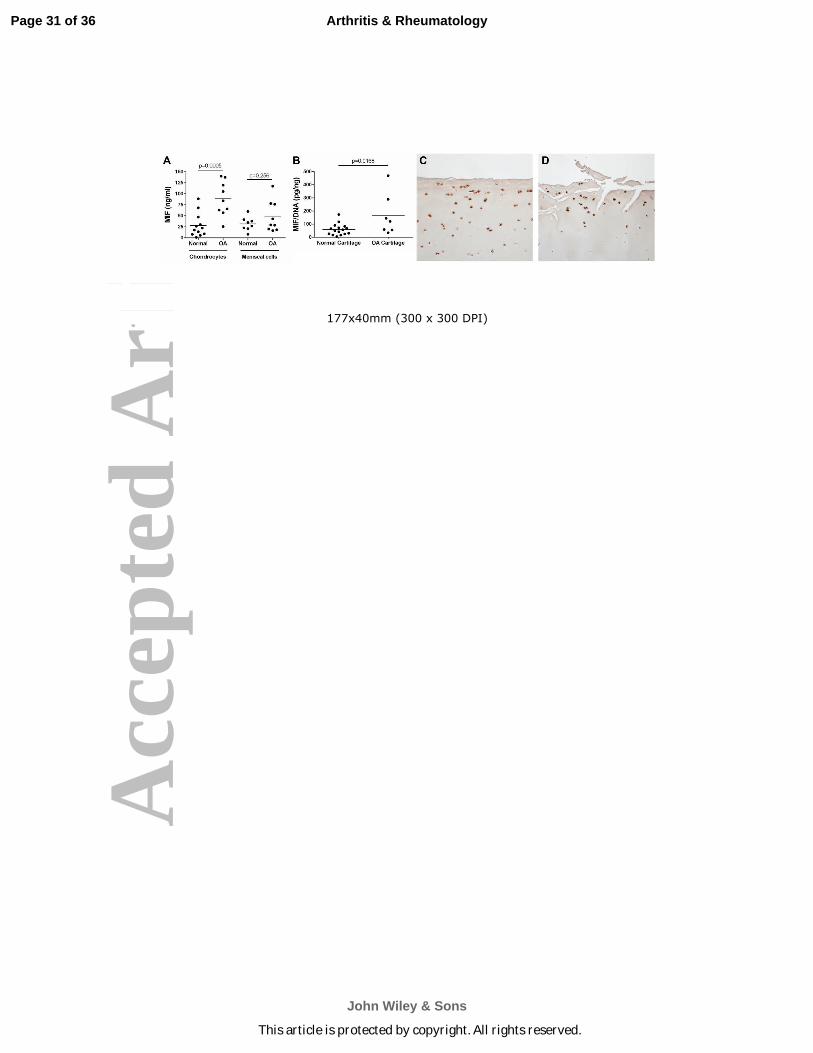
Human OA chondrocytes secrete higher levels of MIF than normal chondrocytes.
Due to previously published results showing that MIF is present in higher levels in OA synovial
fluid than in normal synovial fluid (14, 18), the amount of MIF secreted by joint tissue cells was
measured to determine which tissues could be a source of MIF within the joint. Conditioned
media samples were collected from unstimulated monolayer cultures of chondrocytes and
meniscal cells and from unstimulated cartilage explant cultures, and the MIF protein level in the
media was measured by ELISA (Fig. 1A and 1B). Chondrocytes isolated from OA cartilage
secreted an average of 88.76±13.14ng/mL of MIF over 16 hours, which was significantly higher
(p=0.0005) than that produced by chondrocytes from normal cartilage, which secreted an average
of 28.23±7.78ng/mL of MIF. Cells from OA meniscus secreted an average of
47.62±11.73ng/mL of MIF, which was not significantly different from cells isolated from normal
meniscus (31.65±5.54ng/mL). MIF in conditioned media from OA synovial fibroblasts was not
detectable (data not shown).
OA cartilage explants released an average of 167.9±55.92pg MIF/ng DNA, which was
significantly higher (p=0.0168) than that produced by normal cartilage explants, which released
Page 11 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
12
an average of 59.21±10.87pg MIF/ng DNA. MIF levels in human cartilage were visualized by
immunostaining. MIF was localized to chondrocytes in sections from both normal (Fig. 1C) and
OA donors (Fig. 1D) without significant differences in immunopositivity suggesting that the
differences noted in cultured cells and explants were due to differences in MIF release into the
media.
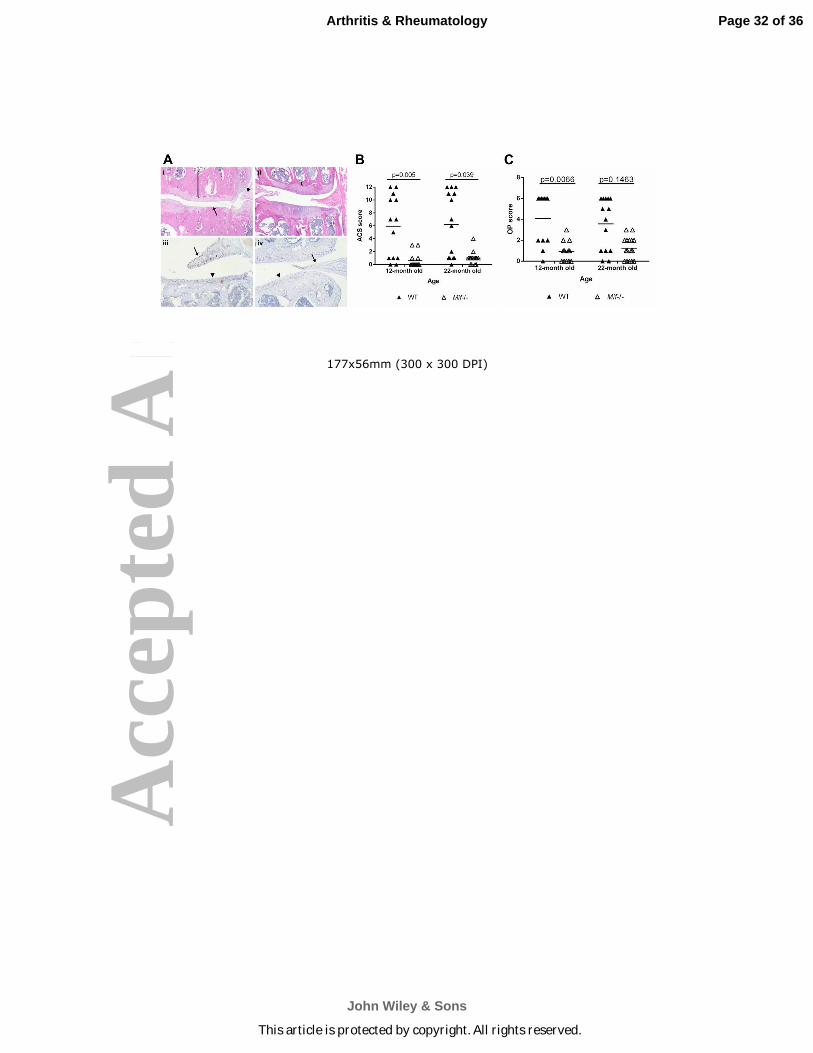
Mif deletion in mice reduces the severity of naturally occurring OA with age. In
order to determine if MIF contributes to the development of OA in vivo, the stifle joints from 12-
and 22-month old male Mif-/- and WT mice were analyzed for OA severity. Histological
evaluation of hematoxylin and eosin (H&E) sections revealed characteristic OA changes in the
joints of the WT mice at both ages, including degradation and loss of the articular cartilage,
thickening of the subchondral bone, and osteophyte formation (Fig. 2A). In contrast, the joints
of the Mif-/- mice displayed more healthy appearing articular cartilage, normal thickness of the
subchondral bone, and no osteophyte formation. Anti-MIF immunostaining in a WT section
showed MIF expression in the meniscus and the articular cartilage (Fig. 2A). As expected, there
was a lack of staining for MIF in the sections from Mif-/- mice.
OA severity in the 12- and 22-month old Mif-/- and WT mice, graded using the articular
cartilage structure (ACS) score, revealed significantly higher ACS scores in the WT relative to
the Mif-/- mice at both 12 and 22 months of age (Fig. 2B). WT mice at 12 and 22 months
developed significantly larger osteophytes than the Mif-/- mice (Fig. 2C). The joints of the 12-
and 22-month old Mif-/- and WT mice were further analyzed using histomorphometric analysis
(Table 1). In both age groups, the Mif-/- mice maintained significantly greater articular cartilage
area and thickness and more viable chondrocytes than the WT mice. Subchondral bone changes
were also evident between the genotypes. The subchondral bone area and thickness were
Page 12 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
13
significantly greater in WT than in Mif-/- mice. The only two parameters which were not
significantly different between Mif-/- and WT were the area of chondrocyte necrosis and the
calcified cartilage area.
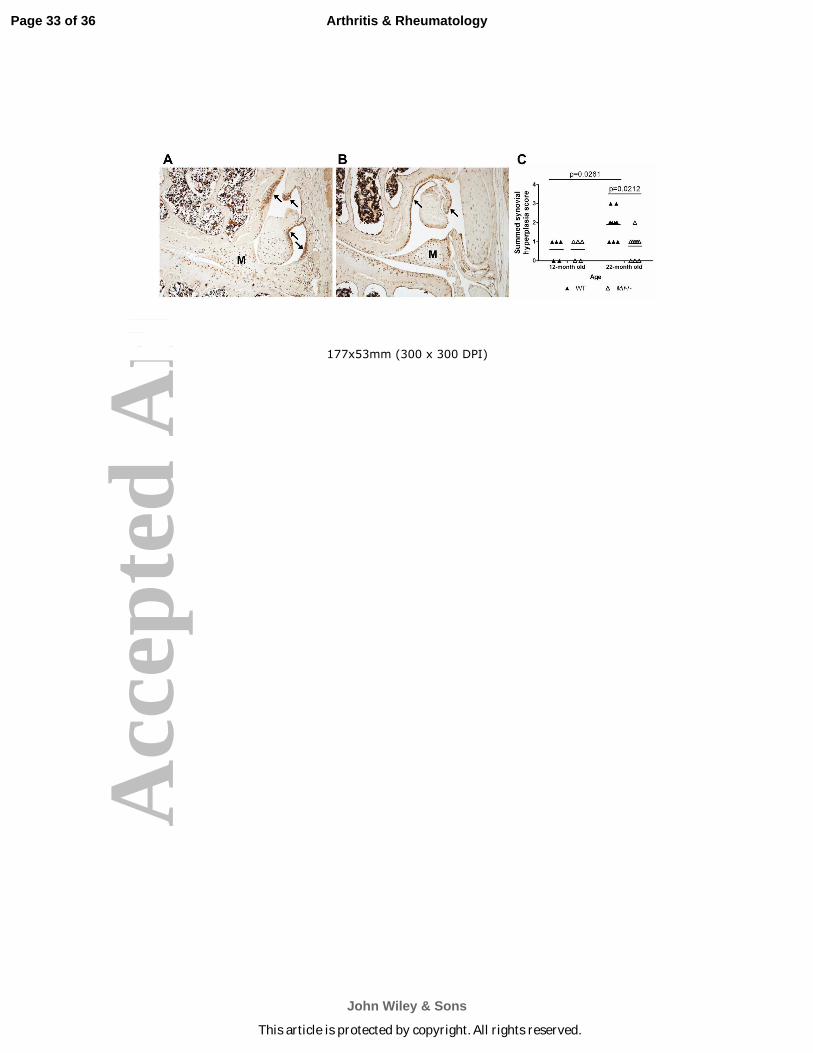
Synovitis with synovial hyperplasia is another characteristic of OA. S100A8 is an
alarmin protein that is produced by synovial cells as well as activated macrophages and has been
shown to correlate with synovitis (27, 30). Here, immunohistochemistry was performed using
anti-S100A8 on coronal sections from 12- and 22-month old WT and Mif-/- mice (representative
images are shown in Fig. 3A and 3B). In both genotypes, immunopositivity was evident
throughout the joint – in the articular cartilage, the meniscus, and the synovium. Specifically in
the WT joints, S100A8 immunopositivity was strong in both the cells and matrix of the
synovium. As shown in Fig. 3A, S100A8 immunopositivity correlated with areas of synovial
hyperplasia and thickening of the synovial lining. In contrast, the S100A8 synovial
immunostaining results in the Mif-/- mice revealed minimal synovial hyperplasia. Based on the
described synovial scoring system, there was no difference in the synovial hyperplasia between
the 12-month old WT and Mif-/- mice, while the 22-month old WT mice exhibited more severe
synovial hyperplasia than the Mif-/- mice (Fig. 3C). Additionally, the severity of synovial
hyperplasia increased between 12 and 22 months in the WT mice.
Early bone differences in mice with Mif deletion are not maintained with age.
Because of conflicting reports of bone density differences in Mif-/- mice compared to WT mice
(31, 32) and the possibility that bone density differences could influence the development of OA
(33), we measured trabecular bone parameters by µCT scans of the proximal metaphysis of the
tibia from 8-week old and 12-month old male Mif-/- and WT mice. The 8-week old male Mif-/-
mice had significantly greater bone volume fraction, tissue mineral density, and connectivity
Page 13 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
14
density than the age-matched WT mice. These differences were not maintained with age, as
there were no significant differences in any bone parameters between 12-month old male Mif-/-
and WT mice. Bone density parameters were also measured in female mice of the same ages,
but there were no differences in these parameters between Mif-/- and WT mice in either age
group (data not shown).
Mif deletion or treatment with a MIF neutralizing antibody does not reduce the
severity of surgically-induced OA in young adult mice. We next wanted to determine if MIF
was required for the development of surgically-induced OA in young adult mice. Here, we used
the destabilization of the medial meniscus (DMM) surgical model to induce OA in 12-week old
male Mif-/- and WT mice and evaluated the severity of cartilage lesions 10 weeks after surgery.
Compared to the sham operated controls, both the WT and Mif-/- mice developed significant
cartilage lesions; however, in contrast to the data on natural age-related OA, there was no
difference in severity between the two groups (Fig. 5A). Both the WT and Mif-/- DMM mice
developed significantly larger osteophytes than the sham mice, but there was no difference
between the two groups (Fig 5B).
We also determined if inhibition of MIF systemically, using a neutralizing antibody,
would alter the severity of surgically-induced OA. Twelve-week old male C57Bl/6 mice
underwent the DMM procedure and were then treated for ten weeks with either a MIF
neutralizing antibody or an IgG control antibody. The dose of anti-MIF used here was sufficient
to inhibit MIF activity in a previous study (24). OA severity was analyzed ten weeks after
surgery (Fig. 5C). Due to limitations in the amount of MIF antibody available and our previous
data indicating that a sham control group did not develop OA changes, the contralateral legs
were scored as the control group in this study. The DMM limbs of the control IgG-treated group
Page 14 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
15
and the anti-MIF-treated group developed significantly more severe OA than the contralateral
limbs, and there was no difference in OA severity between the two treatment groups. The DMM
limbs of both the IgG and MIF treatment groups developed significantly larger osteophytes than
the contralateral limbs (Fig 5D).
DISCUSSION
Although OA is characterized by common pathologic changes within affected joints, the
pathways that lead to OA can vary depending on the inciting factors. Here, we demonstrate that
deletion of the pro-inflammatory cytokine MIF protects mice from developing naturally
occurring age-related OA but not from developing injury-induced OA. These findings are in
stark contrast to previous studies on mice with deletion of the pro-inflammatory cytokine IL-6
where age-related OA was more severe in the IL-6 knockouts (34) while they were protected
from injury-induced OA using the same DMM model used in our MIF study (35). Other studies
have also shown different effects when age-related and injury-induced OA were evaluated in
mice with specific gene deletions including Mmp3 which, similar to our results, had less age-
related OA but no effects on OA severity in an injury model (reviewed in (36)). These studies
emphasize the need to consider more than one model of OA when determining the role of a
particular factor and indicate that the successful treatment of age-related OA and post-traumatic
OA may require different targets.
The aged Mif knockouts not only exhibited less articular cartilage damage compared to
age-matched wild types but also less synovial hyperplasia and fewer OA bone changes, including
Page 15 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
16
osteophytes. Synovial inflammation is evident in over 60% of clinical cases of OA as measured
by MRI (37). Synovial inflammation is a key source of pain in OA due to macrophage
infiltration and increased vascularization and innervation of the synovium (38). We used the
alarmin S100A8 as a marker of synovial changes due to its strong correlation with synovial
hyperplasia and synovitis as well as its association with macrophage infiltration in the synovium
(27, 30). However, while there is a clear correlation between the Mif genotype, synovial
hyperplasia, and OA severity, it is difficult to determine if lack of MIF resulted in less synovial
involvement and this contributed to cartilage protection or if less cartilage damage in the Mif-/-
mice resulted in less synovial hyperplasia. The DMM model effectively induces OA changes in
the cartilage and bone, but minimal synovitis has been observed in this model (39). We also did
not observe significant synovitis or synovial hyperplasia in our DMM mice (data not shown)
suggesting that the differential effect of Mif knockout on age-related OA and DMM-induced OA
may be due to a differential role of the synovium. This would be consistent with studies that
have shown that MIF contributes to synovitis in models of inflammatory arthritis (40). As MIF
is constitutively expressed in the joint, it is possible that it exerts an age-related deleterious
action on joint homeostasis but is not important in the acute setting of joint injury when the
expression of other inflammatory mediators prevail.
The finding that the ACS scores in the wild-type mice did not change significantly
between 12- and 22-months was unexpected. Since this was a cross-sectional study it is not
possible to determine if this was due to a lack of progression of the cartilage changes or just a
chance occurrence in two different sets of mice. However, we did note that there was a
significant decrease in the subchondral bone area between the 12- and 22-month old WT mice,
and there was a trend towards a significant decrease in the articular cartilage area between the
Page 16 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
17
two groups, but we did not note differences in the number of viable chondrocytes or in the area
of chondrocyte necrosis. These aging changes in the mice are similar to aging changes in
humans where there is a thinning of the articular cartilage (2), a decrease in the number of viable
chondrocytes (41), and an overall increase in bone turnover (1).
There is some controversy over the role of MIF in bone. Studies have shown that MIF
inhibits osteoclastogenesis through activation of the tyrosine kinase Lyn (42), and that Mif-/-
mice have significantly less bone volume than WT mice (32). Alternatively, MIF has been
shown to be necessary for osteoclastogenesis in a mouse model of rheumatoid arthritis and Mif-/-
mice were protected from development of bone erosions in this model (15). Because increased
bone density has been shown to be a potential risk factor for incident OA while increased bone
turnover may promote progression (43, 44), we examined bone density in young and older adult
Mif-/- mice. We found that the BV/TV and other bone parameters were higher in young Mif-/-
mice compared to WT mice, but these early bone differences were not maintained with age.
Therefore, it is unlikely that these early bone density differences were responsible for the
reduced OA severity seen in the older adult Mif-/- mice.
It is not clear from the present study if Mif deletion had a direct effect on the articular
chondrocytes that could explain less severe age-related OA. Immunohistochemistry results
revealed that chondrocytes, as well as meniscal cells, were immunopositive for MIF and released
MIF into the media when cultured. Similar intracellular staining and release has been seen with
other cell types such as macrophages (12) and RA synovial fibroblasts (14). Chondrocytes, but
not meniscal cells, from OA joints secreted more MIF than cells from normal joints. However,
we could not detect an effect of extracellular MIF on joint tissue cells. We performed a series of
in vitro studies using recombinant MIF in doses up to 1 µg/mL and could not detect an increase
Page 17 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
18
in ERK, p38, or JNK MAP kinase activation or increase in MMP production in cultures of
human chondrocytes, meniscal cells, or synovial fibroblasts (supplemental Fig.1).
These in vitro results with joint tissue cells are contrary to published studies with other
cell types that reported recombinant MIF-stimulated phosphorylation of the ERK MAP kinase in
Raji cells (a B cell line) (9) and in primary rat osteoblasts (45). Additional published studies
show recombinant MIF-stimulated production of MMP-1 and MMP-3 production in synovial
fibroblasts isolated from OA and RA patients (13) as well as MMP-9 and MMP-13 production in
primary rat osteoblasts (45). However, those studies required doses of MIF up to 10 µg/mL to
detect a response that far exceeds the amount of MIF measured in synovial fluid, which ranges
from 3 to 19 ng/mL (14, 18), or the amount of MIF we measured in conditioned media from
confluent chondrocyte and meniscal cell monolayers, which ranged from 1 to 140 ng/ml. We
examined chondrocytes for the presence of CD74, which is the primary receptor for MIF, but
could not detect significant levels on the cell surface by flow cytometry (unpublished results).
These findings suggest that chondrocytes produce and secrete MIF but that it does not have an
autocrine effect on cartilage. MIF may instead promote OA by promoting macrophage
infiltration into the joint, which would explain the reduced synovial hyperplasia observed in the
aged mice with MIF deleted. MIF also has been described to exert intracellular functions by
intracytoplasmic interaction with the COP9 signalosome that influences cell cycle progression
(46). Accordingly, the distinction we observed between the effects Mif deletion versus
immunoneutralization may be accounted for by a strictly intracellular role for MIF in
chondrocytes.
Taken together, the results presented here indicate that the lack of MIF has a strong
protective effect on naturally occurring age-related OA. As the percentage of adults over the age
Page 18 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
19
of 65 continues to increase, the prevalence of aging-related diseases, including OA, is also
increasing (47). MIF is encoded in a functionally polymorphic locus that occurs commonly in
the population (48), and our data prompt investigation into the potential role of variant MIF
alleles in OA incidence or progression. Therapies that are able to stop or slow the progression of
diseases of aging are necessary in order to improve the quality of life and to extend the healthy
lifespan of the aging population.
ACKNOWLEDGEMENTS
We thank the Gift of Hope Tissue and Organ Donor Network, Dr. Susan Chubinskaya,
and the donor families for providing normal donor tissue. We also thank the Department of
Orthopaedic Surgery at Wake Forest Baptist Health and the University of North Carolina for
providing osteoarthritic tissue. We thank Dr. Tom Smith (Department of Orthopaedic Surgery,
Wake Forest School of Medicine) and Kathryn Kelley (Thurston Arthritis Research Center,
University of North Carolina at Chapel Hill) for their assistance in performing the DMM
surgeries as well as Melissa Roy, Dr. Alexandra Armstrong, and Dr. Laura Pritzker (University
of Minnesota) for their assistance with histological grading/morphometry and Dr. Ted Bateman
and Eric Livingston (University of North Carolina at Chapel Hill) for their assistance with the
microCT. We also thank Dr. Martin Lotz (Scripps Research Institute, La Jolla, CA) for his kind
gift of the human cartilage sections, Dr. Peter van der Kraan (Radboud University Medical
Center, Nijmegen) for his advice regarding our synovial studies, and Dr. Johannes Roth (Institute
of Immunology, University of Münster, Münster) for his kind gift of the S100A8 antibody.
REFERENCES
Page 19 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
20
1. Loeser RF. Age-related changes in the musculoskeletal system and the development of
osteoarthritis. Clin Geriatr Med. 2010;26:371-386.
2. Ding C, Cicuttini F, Scott F, Cooley H, Jones G. Association between age and knee
structural change: a cross sectional MRI based study. Ann Rheum Dis. 2005;64:549-555.
3. Chen AC, Temple MM, Ng DM, Verzijl N, DeGroot J, TeKoppele JM, et al. Induction of
advanced glycation end products and alterations of the tensile properties of articular
cartilage. Arthritis Rheumatol. 2002;46:3212-3217.
4. Loeser RF, Gandhi U, Long DL, Yin W, Chubinskaya S. Aging and oxidative stress
reduce the response of human articular chondrocytes to insulin-like growth factor 1 and
osteogenic protein 1. Arthritis Rheumatol. 2014;66:2201-2209.
5. Messai H, Duchossoy Y, Khatib AM, Panasyuk A, Mitrovic DR. Articular chondrocytes
from aging rats respond poorly to insulin-like growth factor-1: an altered signaling
pathway. Mech Ageing Dev. 2000;115:21-37.
6. Blaney Davidson EN, Scharstuhl A, Vitters EL, van der Kraan PM, van den Berg WB.
Reduced transforming growth factor-beta signaling in cartilage of old mice: role in
impaired repair capacity. Arthritis Res Ther. 2005;7:R1338-1347.
7. Bobacz K, Gruber R, Soleiman A, Erlacher L, Smolen JS, Graninger WB. Expression of
bone morphogenetic protein 6 in healthy and osteoarthritic human articular chondrocytes
and stimulation of matrix synthesis in vitro. Arthritis Rheumatol. 2003;48:2501-2508.
8. Greene MA, Loeser RF. Aging-related inflammation in osteoarthritis. Osteoarthritis
Cartilage. 2015;23:1966-1971.
9. Leng L, Metz CN, Fang Y, Xu J, Donnelly S, Baugh J, et al. MIF signal transduction
initiated by binding to CD74. J Exp Med. 2003;197:1467-1476.
Page 20 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
21
0. Rajasekaran D, Zierow S, Syed M, Bucala R, Bhandari V, Lolis EJ. Targeting distinct
tautomerase sites of D-DT and MIF with a single molecule for inhibition of neutrophil
lung recruitment. FASEB J. 2014;28:4961-4971.
1. Mitchell RA, Liao H, Chesney J, Fingerle-Rowson G, Baugh J, David J, et al.
Macrophage migration inhibitory factor (MIF) sustains macrophage proinflammatory
function by inhibiting p53: regulatory role in the innate immune response. Proc Natl
Acad Sci U S A. 2002;99:345-350.
2. Calandra T, Bernhagen J, Mitchell RA, Bucala R. The macrophage is an important and
previously unrecognized source of macrophage migration inhibitory factor. J Exp Med.
1994;179:1895-1902.
3. Onodera S, Kaneda K, Mizue Y, Koyama Y, Fujinaga M, Nishihira J. Macrophage
migration inhibitory factor up-regulates expression of matrix metalloproteinases in
synovial fibroblasts of rheumatoid arthritis. J Biol Chem. 2000;275:444-450.
4. Onodera S, Tanji H, Suzuki K, Kaneda K, Mizue Y, Sagawa A, et al. High expression of
macrophage migration inhibitory factor in the synovial tissues of rheumatoid joints.
Cytokine. 1999;11:163-167.
5. Gu R, Santos LL, Ngo D, Fan H, Singh PP, Fingerle-Rowson G, et al. Macrophage
migration inhibitory factor is essential for osteoclastogenic mechanisms in vitro and in
vivo mouse model of arthritis. Cytokine. 2015;72:135-145.
6. Foote A, Briganti EM, Kipen Y, Santos L, Leech M, Morand EF. Macrophage migration
inhibitory factor in systemic lupus erythematosus. J Rheumatol. 2004;31:268-273.
Page 21 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
22
7. Hoi AY, Hickey MJ, Hall P, Yamana J, O'Sullivan KM, Santos LL, et al. Macrophage
migration inhibitory factor deficiency attenuates macrophage recruitment,
glomerulonephritis, and lethality in MRL/lpr mice. J Immunol. 2006;177:5687-5696.
8. Liu M, Hu C. Association of MIF in serum and synovial fluid with severity of knee
osteoarthritis. Clin Biochem. 2012;45:737-739.
9. Harper JM, Wilkinson JE, Miller RA. Macrophage migration inhibitory factor-knockout
mice are long lived and respond to caloric restriction. FASEB J. 2010;24:2436-2442.
0. Bernhagen J, Krohn R, Lue H, Gregory JL, Zernecke A, Koenen RR, et al. MIF is a
noncognate ligand of CXC chemokine receptors in inflammatory and atherogenic cell
recruitment. Nat Med. 2007;13:587-596.
1. Loeser RF, Pacione CA, Chubinskaya S. The combination of insulin-like growth factor 1
and osteogenic protein 1 promotes increased survival of and matrix synthesis by normal
and osteoarthritic human articular chondrocytes. Arthritis Rheumatol. 2003;48:2188-
2196.
2. Yin W, Park JI, Loeser RF. Oxidative stress inhibits insulin-like growth factor-I
induction of chondrocyte proteoglycan synthesis through differential regulation of
phosphatidylinositol 3-Kinase-Akt and MEK-ERK MAPK signaling pathways. J Biol
Chem. 2009;284:31972-31981.
3. Loeser RF, Olex AL, McNulty MA, Carlson CS, Callahan MF, Ferguson CM, et al.
Microarray analysis reveals age-related differences in gene expression during the
development of osteoarthritis in mice. Arthritis Rheumatol. 2012;64:705-717.
Page 22 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
23
4. Leng L, Chen L, Fan J, Greven D, Arjona A, Du X, et al. A small-molecule macrophage
migration inhibitory factor antagonist protects against glomerulonephritis in lupus-prone
NZB/NZW F1 and MRL/lpr mice. J Immunol. 2011;186:527-538.
5. McNulty MA, Loeser RF, Davey C, Callahan MF, Ferguson CM, Carlson CS. A
comprehensive histological assessment of osteoarthritis lesions in mice. Cartilage.
2011;2:354-363.
6. Miller RE, Tran PB, Das R, Ghoreishi-Haack N, Ren D, Miller RJ, et al. CCR2
chemokine receptor signaling mediates pain in experimental osteoarthritis. Proc Natl
Acad Sci U S A. 2012;109:20602-20607.
7. Schelbergen RF, de Munter W, van den Bosch MH, Lafeber FP, Sloetjes A, Vogl T, et al.
Alarmins S100A8/S100A9 aggravate osteophyte formation in experimental osteoarthritis
and predict osteophyte progression in early human symptomatic osteoarthritis. Ann
Rheum Dis. 2014;75:218-225.
8. Lau AG, Sun J, Hannah WB, Livingston EW, Heymann D, Bateman TA, et al. Joint
bleeding in factor VIII deficient mice causes an acute loss of trabecular bone and
calcification of joint soft tissues which is prevented with aggressive factor replacement.
Haemophilia. 2014;20:716-722.
9. Muller R, Van Campenhout H, Van Damme B, Van Der Perre G, Dequeker J, Hildebrand
T, et al. Morphometric analysis of human bone biopsies: a quantitative structural
comparison of histological sections and micro-computed tomography. Bone. 1998;23:59-
66.
0. Schelbergen RF, van Dalen S, ter Huurne M, Roth J, Vogl T, Noel D, et al. Treatment
efficacy of adipose-derived stem cells in experimental osteoarthritis is driven by high
Page 23 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
24
synovial activation and reflected by S100A8/A9 serum levels. Osteoarthritis Cartilage.
2014;22:1158-1166.
1. Oshima S, Onodera S, Amizuka N, Li M, Irie K, Watanabe S, et al. Macrophage
migration inhibitory factor-deficient mice are resistant to ovariectomy-induced bone loss.
FEBS Lett. 2006;580:1251-1256.
2. Jacquin C, Koczon-Jaremko B, Aguila HL, Leng L, Bucala R, Kuchel GA, et al.
Macrophage migration inhibitory factor inhibits osteoclastogenesis. Bone. 2009;45:640-
649.
3. Hardcastle SA, Dieppe P, Gregson CL, Davey Smith G, Tobias JH. Osteoarthritis and
bone mineral density: are strong bones bad for joints? Bonekey Rep. 2015;4:624.
4. de Hooge AS, van de Loo FA, Bennink MB, Arntz OJ, de Hooge P, van den Berg WB.
Male IL-6 gene knock out mice developed more advanced osteoarthritis upon aging.
Osteoarthritis Cartilage. 2005;13:66-73.
5. Ryu JH, Yang S, Shin Y, Rhee J, Chun CH, Chun JS. Interleukin-6 plays an essential role
in hypoxia-inducible factor 2alpha-induced experimental osteoarthritic cartilage
destruction in mice. Arthritis Rheumatol. 2011;63:2732-2743.
6. Little CB, Zaki S. What constitutes an "animal model of osteoarthritis"--the need for
consensus? Osteoarthritis Cartilage. 2012;20:261-267.
7. Wang X, Jin X, Han W, Cao Y, Halliday A, Blizzard L, et al. Cross-sectional and
longitudinal associations between knee joint effusion synovitis and knee pain in older
adults. J Rheumatol. 2015;43:121-130.
8. Benito MJ, Veale DJ, FitzGerald O, van den Berg WB, Bresnihan B. Synovial tissue
inflammation in early and late osteoarthritis. Ann Rheum Dis. 2005;64:1263-1267.
Page 24 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
25
9. Jackson MT, Moradi B, Zaki S, Smith MM, McCracken S, Smith SM, et al. Depletion of
protease-activated receptor 2 but not protease-activated receptor 1 may confer protection
against osteoarthritis in mice through extracartilaginous mechanisms. Arthritis
Rheumatol. 2014;66:3337-3348.
0. Morand EF, Bucala R, Leech M. Macrophage migration inhibitory factor: an emerging
therapeutic target in rheumatoid arthritis. Arthritis Rheumatol. 2003;48:291-299.
1. Kuhn K, D'Lima DD, Hashimoto S, Lotz M. Cell death in cartilage. Osteoarthritis
Cartilage. 2004;12:1-16.
2. Mun SH, Oh D, Lee SK. Macrophage migration inhibitory factor down-regulates the
RANKL-RANK signaling pathway by activating Lyn tyrosine kinase in mouse models.
Arthritis Rheumatol. 2014;66:2482-2493.
3. Dequeker J, Aerssens J, Luyten FP. Osteoarthritis and osteoporosis: clinical and research
evidence of inverse relationship. Aging Clin Exp Res. 2003;15:426-439.
4. Zhang Y, Hannan MT, Chaisson CE, McAlindon TE, Evans SR, Aliabadi P, et al. Bone
mineral density and risk of incident and progressive radiographic knee osteoarthritis in
women: the Framingham Study. J Rheumatol. 2000;27:1032-1037.
5. Onodera S, Nishihira J, Iwabuchi K, Koyama Y, Yoshida K, Tanaka S, et al. Macrophage
migration inhibitory factor up-regulates matrix metalloproteinase-9 and -13 in rat
osteoblasts. Relevance to intracellular signaling pathways. J Biol Chem. 2002;277:7865-
7874.
6. Kleemann R, Hausser A, Geiger G, Mischke R, Burger-Kentischer A, Flieger O, et al.
Intracellular action of the cytokine MIF to modulate AP-1 activity and the cell cycle
through Jab1. Nature. 2000;408:211-216.
Page 25 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
26
7. Cross M, Smith E, Hoy D, Nolte S, Ackerman I, Fransen M, et al. The global burden of
hip and knee osteoarthritis: estimates from the global burden of disease 2010 study. Ann
Rheum Dis. 2014;73:1323-1330.
8. Bucala R. MIF, MIF alleles, and prospects for therapeutic intervention in autoimmunity. J
Clin Immunol. 2013;33 Suppl 1:S72-78.
Page 26 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
27
TABLES
Table 1. Histomorphometric analysis of aged mice with Mif deletion and wild-type controls.
12-month old 22-month old
WT
12- vs
22-
month
olds
Parameter Mif-/-
Mean (SD)
WT
Mean (SD) p-value
Mif-/-
Mean (SD)
WT
Mean (SD) p-value p-value
Art Cart Area
(mm2)
0.064
(0.011)
0.040
(0.016) <0.001
0.064
(0.008)
0.029
(0.018) <0.001 .089
Art Cart
Thickness
(mm)
0.072
(0.013)
0.043
(0.017) <0.001
0.078
(0.009)
0.037
(0.019) <0.001 .339
Subchondral
Bone Area
(mm2)
0.080
(0.016)
0.156
(0.069) 0.002
0.070
(0.025)
0.103
(0.038) 0.009 .024
Subchondral
Bone
Thickness
(mm)
0.059
(0.011)
0.104
(0.046) 0.004
0.070
(0.023)
0.100
(0.037) 0.013 .772
Number of
Viable
Chondrocytes
138.39
(16.15)
74.31
(41.38) <0.001
115.50
(21.98)
64.14
(45.24) 0.001 .549
Area of
chondrocyte
necrosis
(mm2)
0.008
(0.006)
0.006
(0.004) 0.396
0.008
(0.011)
0.007
(0.008) 0.859 0.602
Calcified
cartilage area
(mm2)
0.063
(0.012)
0.059
(0.019) 0.504
0.049
(0.005)
0.053
(0.019) 0.368 .421
Histomorphometry measurements of cartilage (Cart) and bone parameters in the medial tibial
compartment of mid-coronal stifle sections were analyzed using the Osteomeasure
Page 27 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
28
Histomorphometry Program (OsteoMetrics®). 12-month old WT: n=13; 12-month old Mif-/-:
n=13; 22-month old wild-type (WT): n=14; 22-month old Mif-/-: n=16. Significance differences
determined by heteroscedastic t-test using SPSS.
FIGURE LEGENDS
Figure 1. MIF levels in conditioned media from normal and OA chondrocyte and meniscal cell
cultures and in normal and OA cartilage sections. A. Serum-free medium was collected after 16
hours of culture from unstimulated confluent monolayers of normal and OA chondrocytes and
meniscal cells. MIF protein in media was measured by human MIF ELISA. Horizontal lines on
the graph represent average MIF level for each tissue type; individual data points are shown.
Normal chondrocytes: n=12 donors; OA chondrocytes: n=9 donors; normal meniscus: n=8
donors; OA meniscus: n=9 donors. Statistical significance was evaluated by unpaired t-test
within tissue types. B. Serum-free conditioned medium was collected after 48 hours of culture
from unstimulated human and OA cartilage explants. MIF protein in media was measured by
human MIF ELISA and was normalized to the DNA content of the explants. Individual data
points are shown. Normal cartilage explants: n=16 donors; OA cartilage explants: n=7 donors.
Statistical significance was evaluated by unpaired t-test, and no outliers were identified by the
Grubbs outlier test. Immunohistochemistry results for MIF in sections of (C) human normal
knee cartilage and (D) human OA knee cartilage.
Figure 2. Mice with Mif deletion are protected from age-related OA. A. Representative
histologic images of knee joint sections in 12-month old Mif-/- and WT mice. (i) WT mouse,
medial tibial plateau (MTP) articular cartilage structure (ACS) score 11. Degradation and loss of
Page 28 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
29
articular cartilage (arrow), osteophyte formation (arrowhead), and thickening of subchondral
bone (bracket). (H&E) (ii) Mif-/- mouse, ACS score 3. Normal subchondral bone thickness
(bracket). (H&E) (iii) WT mouse section immunostained for MIF. Intracellular MIF is present in
meniscus (arrow) and articular cartilage (arrowhead). (iv) Mif-/- mouse. Lack of MIF
immunopositivity in meniscus (arrow) and cartilage (arrowhead). B. Articular cartilage structure
scores in Mif-/- and WT mice. The medial tibial plateau of mid-coronal stifle sections was
scored using the articular cartilage structure (ACS) score. 12-month old WT: n=13; 12-month
old Mif-/-: n=13; 22-month old WT: n=14; 22-month old Mif-/-: n=16. C. Osteophyte scores in
Mif-/- and WT mice from (B). The summed scores from the medial tibial plateau (MTP) and
medial femoral condyle (MFC) are presented. Horizontal lines on graph represent the mean of
each group. Significant differences were determined by Kruskal-Wallis nonparametric one-way
ANOVA followed by Dunn’s multiple comparisons test.
Figure 3. Synovial hyperplasia is less severe in 22-month old mice with Mif deletion.
Immunohistochemistry results for S100A8 in (A) 22-month old WT mouse, ACS score 11, and
(B) 22-month old Mif-/- mouse, ACS score 0. Arrows indicate regions of strong synovial
S100A8 immunopositivity and synovial hyperplasia. Representative histologic images are
shown. M=meniscus. C. Synovial hyperplasia scores of the medial and lateral compartments
were summed from 12-month old WT (n=5) and Mif-/- (n=5) and from 22-month old WT (n=11)
and Mif-/- (n=9) mice. Horizontal lines on graph represent the mean of each group. Significant
differences were determined by Kruskal-Wallis nonparametric one-way ANOVA followed by
Dunn’s multiple comparisons test.
Page 29 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
30
Figure 4. µCT analysis of 8-week old and 12-month old male mice with Mif deletion and wild-
type controls. Bone density parameters of the proximal metaphysis of the tibia were analyzed
using Scanco software in 8-week old WT (n=3) and Mif-/- (n=3) and in 12-month old WT (n=4)
and Mif-/- (n=5) mice. A. Bone volume fraction. (BV/TV=bone volume/total volume) B. Tissue
mineral density. (HA=hydroxyapatite) C. Connectivity density. D. Trabecular number. E.
Trabecular separation. F. Trabecular thickness. Horizontal lines on graphs represent the mean
of each group. Significant differences were determined by two-way ANOVA followed by
Tukey’s multiple comparisons post-hoc test.
Figure 5. Inhibition of MIF activity either by gene deletion or by neutralizing antibody does not
protect young adult mice from injury-induced OA. A. Twelve-week old Mif-/- or WT mice
received either the DMM or sham procedure. OA severity was analyzed 10 weeks after surgery
by the Saf-o score. Mif-/- DMM: n=14; Mif-/- sham: n=16; WT DMM: n=16; WT sham: n=13.
B. Osteophytes were measured in the mice from (A) and summed scores from the MTP and
MFC are presented. C. Twelve-week old WT mice received the DMM procedure and were
treated for 10 weeks with either the MIF neutralizing antibody (Anti-MIF) or IgG control
antibody. The contralateral limb was used as the un-operated control. OA severity was analyzed
10 weeks after surgery by the Saf-O score. Anti-MIF DMM: n=15; Anti-MIF contralateral
control: n=15; IgG DMM: n=14; IgG contralateral control: n=14. D. Osteophytes were
measured in the mice from (C) and summed scores from the MTP and MFC are presented.
Horizontal lines on the graph represent the mean of each group. Significant differences were
determined by Kruskal-Wallis nonparametric one-way ANOVA followed by Dunn’s multiple
comparisons test.
Page 30 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
177x40mm (300 x 300 DPI)
Page 31 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
177x56mm (300 x 300 DPI)
Page 32 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
177x53mm (300 x 300 DPI)
Page 33 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
177x85mm (300 x 300 DPI)
Page 34 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
177x49mm (300 x 300 DPI)
Page 35 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
177x76mm (300 x 300 DPI)
Page 36 of 36
John Wiley & Sons
Arthritis & Rheumatology
This article is protected by copyright. All rights reserved.
Copyright © 2022 FDOKUMEN




















